Embed Size (px)
Citation preview

Journal of Experimental Botany, Vol. 49, Special Issue, pp. 387–398, March 1998
Stomatal control of photosynthesis and transpiration
Hamlyn G. Jones1
Department of Biological Sciences, University of Dundee, Dundee DD1 4HN, UK
Received 21 July 1997; Accepted 14 October 1997
Abstract has been little speculation as to the precise ‘goal’ ofstomatal movements; for example, Meidner and
The stomata occupy a central position in the pathways Mansfield (1968) in their classic text simply point outfor both the loss of water from plants and the exchange
that ‘… both (transpiration and photosynthesis) are to aof CO
2. It is commonly assumed that they therefore considerable extent controlled by stomata’.
provide the main short-term control of both transpir- Perhaps the first rigorous attempt to consider explicitlyation and photosynthesis, though the detailed control the ‘goal’ of stomatal movements was that by Cowancriteria on which their movements are based are not (Cowan, 1977; Cowan and Farquhar, 1977) who crystal-well understood and are likely to depend on the par- lized the hypothesis put forward by Parkhurst and Loucksticular ecological situation. This paper first reviews (1972) that stomata operate in such a way as to minimizethe main methods available for quantifying the control water loss relative to the amount of CO2 uptake. Cowanexerted by stomata over transpiration and photosyn- expressed this concept in the following way: ‘stomatalthesis in the absence of feedbacks between gas- aperture would vary so that the average rate of evapora-exchange and stomatal function. The discussion is tion is a minimum for a particular average rate ofthen extended by using very simple models to investi- assimilation’. Subsequent analyses have extended thisgate the role of stomata in the control of gas exchange approach to the longer term where soil moisture may bein the presence of hydraulic feedbacks and to clarify declining with time and future rainfall is uncertainthe nature of causality in such systems. Comparison (Cowan, 1982, 1986). One weakness of this approach hasof a limited number of different mechanistic models been that it does not provide an explicit solution for theof stomatal function is used to investigate likely mech- optimal transpiration rate; another crucial problem isanisms underlying stomatal responses to environment. that it takes no direct account of the effects of plant
water deficits on physiological processes such as growthKey words: Feedback photosynthesis, stomatal function, or water transport. Attempts to respond to these objec-stomatal limitation, transpiration. tions have been made by a number of authors who have
incorporated a sensitivity to water status into the photo-synthetic model (Givnish, 1986; Friend, 1991).Introduction
On the basis of a quite different analysis, based on anIt is a general assumption amongst plant physiologists analysis of the potential role of stomata in the avoidanceand ecologists that stomata have evolved to provide a of xylem cavitations and possible consequent runawaymeans for controlling water loss from plants while embolism (Tyree and Sperry, 1988), Jones and Sutherlandallowing photosynthesis. Notwithstanding the enormous (1991) have proposed that the prime role of stomataamount of research, especially in recent years, into the might be to avoid damaging plant water deficits. Theremechanism of stomatal operation, there has, however, is substantial circumstantial evidence in favour of thisbeen relatively little rigorous consideration of their precise hypothesis, not least the consistently good control of leaffunction in terms of the physiological or ecological pro- water status in so-called ‘isohydric’ plants (Stocker, 1956)cesses that are regulated or optimized by the observed such as cowpea (Bates and Hall, 1981) or maize (Tardieustomatal movements in various environments. Although et al., 1993) or the evidence that stomata close to avoidthere is general agreement that stomata evolved in some cavitation in oak (Cochard et al., 1996). Yet another
possibility is that stomatal control of transpiration has ageneral way as a means of controlling water loss, there
1 Fax: +44 1382 344275. E-mail: [email protected]
© Oxford University Press 1998
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

388 Jones
role in maintaining leaf temperature within an optimal approach to estimation of the importance of the stomatain controlling processes such as photosynthesis and trans-range (Burke et al., 1988; Mahan and Upchurch, 1988).
Of course these different functions are not necessarily piration was provided by the introduction of resistanceanalogues (Maskell, 1928), and their subsequent develop-exclusive.
Although it can be relatively straightforward to deter- ment (Jones, 1992). An assumption implicit in the use ofresistance analogues is that the relative magnitudes of themine the effect of a given change in stomatal conductance
on transpiration or assimilation, analysis of the role of resistances of each component in series (e.g. the intracellu-lar resistance (ri, whether defined as the initial slope ofstomata in the control of these exchanges is complicated
by the existence of several feedback loops (Fig. 1) where an A/ci curve or as a residual resistance), stomatal resist-ance (rs) and the boundary layer resistance (ra)) providechanges in assimilation or transpiration rates resulting
from changes in stomatal conductance can themselves a measure of their relative importance in limiting theoverall rate of the process according to:affect conductance (Cowan, 1972; Farquhar, 1973;
Raschke, 1975; Jones, 1992; Jarvis and Davies, 1998).ls=rs/(rs+ra+ri) (1)
The first part of this paper reviews approaches for thequantification of stomatal limitations to both photosyn- where ls is the relative stomatal limitation. Unfortunately,
as pointed out by Jones (1973, 1985a), the non-linearitythesis and transpiration in the absence of feedbacks; thesecond part then investigates the use of simple models to of the A/ci curve means that this calculation can often be
seriously misleading, potentially leading to a large overes-define the role of stomata in the control of gas exchangein the presence of feedbacks and attempts to clarify the timation of the importance of the stomata in situations
where photosynthetic saturation is approached. Innature of causality in such systems.extreme cases where assimilation is on the horizontalportion of a standard A/ci curve, the use of resistanceQuantification of stomatal control of transpirationanalogues leads to the estimation of significant stomataland assimilationlimitations even though changes in the stomatal conduct-ance (gs) may have no effect on assimilation rate underQuantification of the stomatal control of assimilationsuch circumstances.
Physiologists, breeders and ecologists often wish to quant-A recent example where large changes in gs have been
ify the control exercised by stomata over gas-exchangeshown to have no effect on assimilation is illustrated
processes. A number of approaches to such quantificationin Fig. 2, which shows the dynamics of changes in assim-
have been used.ilation rate and stomatal conductance in response to
Resistance analogues: The first useful quantitativechanges in irradiance in Phaseolus leaves. In these studies,assimilation by Phaseolus leaves was found to increaserapidly after irradiance was increased from 50 to350 mmol m−2 s−1, stabilizing after about 10 min; in con-trast, stomatal conductance continued to rise for about20 min (only reaching about half the final value after10 min). It follows that the large changes in stomatalconductance in the second 10 min had no influence onassimilation rate so that the stomatal limitation shouldhave been negligible over this period, though the valueof ls calculated from equation (1) was approximately 0.3.This conclusion is supported by analysis of the time-course of changes in intercellular space CO2 concentration(ci) which also continued to change over the same periodwhen assimilation was constant. Similar conclusions canbe drawn from studies of circadian rhythms in tomato(J.E. Corlett, S. Wilkinson and A.J. Thompson, unpub-lished data) who showed that after the first 12 h incontinuous light, stomatal conductance declined to aboutone-third of its normal daytime value, yet assimilationcontinued undiminished. Again this implied no substantialFig. 1. Some of the interactions involved in control of stomatal
conductance (gs). Not only does gs affect both assimilation rate (A) stomatal control of assimilation in their experiment.
and transpiration rate (E ), but these have a range of feedback effectson gs, either direct or indirect as illustrated. In addition, environmental
Sensitivity analysis: Partly as a response to the recognitionfactors may also affect any of these variables and the gain of thefeedback loops. of the errors arising from the resistance analogue
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

Photosynthesis and transpiration control 389
step in the photosynthetic pathway with the stomatalcontrol coefficient (Cs) being equivalent to ls from equa-tions (2) and (3). An important feature of metaboliccontrol analysis (MCA) is that it provides the necessarytools for one to calculate the appropriate control coeffi-cients from a knowledge of the kinetics of all the compon-ent biochemical and biophysical processes involved inphotosynthesis (for an example of the approach seeWoodrow, 1994). Another experimentally based approachto determining control coefficients for photosyntheticcomponents such as ribulose-1,5-bisphosphate carb-oxylase (Rubisco) is to make use of transgenic plantswhose activity of certain photosynthetic enzymes has beenmanipulated by use of ‘antisense’ technology (Rodermelet al., 1988). By use of the MCA connectivity theorem itis possible to estimate the stomatal control coefficient(Stitt et al., 1991). The approximate approach used inthis particular case, however, tends to overestimate thetrue stomatal control coefficient because it does not fullytake account of the feedback effect of ci on stomatalconductance. Another problem is that a range of otherfeedbacks may operate in the ‘antisense’ plants modulat-ing other enzymes than the target, in fact there is evensome indication that stomatal conductance itself maydecline in the low Rubisco plants (Stitt et al., 1991;Lauerer et al., 1993).
Application of any of equations (2), or (3) or MCAto the data in Fig. 2 gives an ls close to zero as expected.
Limitation analysis: A number of other approaches to theFig. 2. Responses of CO2 assimilation rate (a), leaf conductance (b)estimation of stomatal limitations have been proposedand intercellular CO2 partial pressure (c) to changing irradiance from
50 to 350 mmol m−2 s−1 and back. The times of changes are indicated which are based on the extrapolation to what might beby the arrows (from Barradas and Jones, 1996). considered to be unrealistic conditions. One of the most
popular of these approaches is to define the stomatalapproach, a more rigorous approach to the quantification limitation as the relative change in assimilation that wouldof the role of stomata in controlling photosynthesis was occur if all stomatal restriction were eliminatedproposed by Jones (1973). In this the relative stomatal (Bjorkman et al., 1972; Farquhar and Sharkey, 1982). Inlimitation to photosynthesis (ls) was defined as the relative this, ls is defined assensitivity of assimilation to an infinitesimal change instomatal conductance ls=(Ao−A)/Ao (4)
ls=(∂A/A)/(∂gs/gs) (2) where Ao is the assimilation rate that would occur withan infinite stomatal conductance. Alternatively, andThis was shown (Jones, 1973) to be equivalent toequally logically perhaps, one might define ls on eliminat-
ls=rs/(rs+ra+r*) (3) ing all the biochemical limitation (Jones, 1985a) but thisgives a very different answer.where r* is the slope of the tangent to the A/ci curve at
the operating point.MCA extension: A major limitation of MCA is that itonly applies to infinitesimally small changes in gs. ItMetabolic control analysis: The sensitivity approach
described above is essentially equivalent to the approach therefore cannot reliably be used to predict the effect ofthe larger changes in gs that might occur with transgenicbased on what is now known as Metabolic Control
Analysis (Kacser and Burns, 1973; Jones, 1995), where plants. An attempt to overcome this restriction has beenmade by the introduction of the so-called Deviation Indexthe relative flux control by different components of a
pathway is given by the relative magnitude of their flux (D) by Small and Kacser (1993) though this strictly onlyapplies to ‘linear systems’, which may limit the practicalcontrol coefficients (C ). These can be calculated for each
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

390 Jones
application of this technique to photosynthesis (Jones, 1985a). It is also worth noting that standard gas-exchangecalculations of ci may also be inaccurate if significant1995).
It will be apparent that the different methods can give stomatal heterogeneity or patchy stomatal closure occurs(see Jones, 1992).very different answers in different situations. Some
examples are summarized in Table 1; in each case thelimitation calculated by the resistance analogue approach Quantification of stomatal control of transpirationis greater than or equal to that calculated either by the
In a similar way to the stomatal control of photosynthesis,sensitivity approaches, including those based on MCA,the role of stomata in controlling transpiration may beor on Farquhar and Sharkey’s (1982) elimination method.defined analogously as the relative change of transpirationAs a generalization, the stomatal limitation calculated byrate for a given relative change in stomatal conductance.these latter, perhaps more realistic, approaches tends toThe role of stomata in the control of transpiration hasbe a rather small fraction ( less than about 20%) of thebeen the subject of debate for many years, not leasttotal photosynthetic limitation as long as the data arebecause Brown and Escombe (1900) in their classicalobtained for well-adapted plants growing at high lightwork omitted consideration of the boundary layer resist-(Stitt et al., 1991; Lauerer et al., 1993; Woodrow, 1994).ance, which was rather unfortunate in that it took manyUnfortunately, the choice of method is somewhat subject-years for this omission to be corrected. Many workersive, depending on one’s particular objectives in attemptinghave concurred with Lloyd’s (1908) conclusion thatthe quantification. For example, a breeder, who may bechanges in stomatal aperture are of greatest significanceconcerned only with rather small changes in stomatalto transpiration at small stomatal apertures with stomataconductance could probably use a sensitivity method,having relatively little regulatory effect when more open.while the use of elimination methods may give a more
The situation for single leaves was clarified well bywidely applicable answer.Bange (1953), who showed that the sensitivity of transpir-An even more difficult problem is to define the contribu-ation from single leaves to changes in stomatal aperturetion that stomata make to determining a change inwas dependent on windspeed (and hence the boundaryassimilation between two different conditions (caused bylayer resistance). In general, in still air transpiration iseither environmental or physiological changes). In prin-only responsive to stomatal aperture when the stomataciple, the most informative approach would be to takeare nearly closed, but as air movement increases, breakingaccount of the precise path of the changes that occur,down the boundary layer resistance, transpirationbut in most cases the requisite information to allow fullbecomes responsive to changes in aperture over a widertracing of the changes in stomatal limitation during therange.change is usually not available so simplified ‘state-
Although the special features of stomata and theirfunction’ approaches have been adopted (Jones, 1973,obvious role in regulation of water loss have been recog-1985a; Assmann, 1988; Peisker and Vaclavık, 1987). Innized for many years, the contrasting views of physiolo-contrast to the conclusion reached above, much of thegists who considered that ‘… stomata must be the primarychange between conditions is often attributable tocontrol of transpiration …’ (Bange, 1953) and meteor-stomatal changes. One common assumption is that anologists who argued that ‘… transpiration from plantincrease of ci implies an increase in the relative limitationcanopies was in general independent of plant water statusdue to intracellular processes (Farquhar and Sharkey,and plant type …’ were only properly reconciled when1982), but this conclusion can be shown to be misleadingMcNaughton and Jarvis (1983) reformulated the classicalwhere the actual sequence of changes is known (Jones,Penman–Monteith combination evaporation equation toincorporate the degree to which leaves are ‘coupled’ toenvironmental conditions. In particular, they proposed aTable 1. Some examples where stomatal limitations have beendecoupling coefficient (V=(e+1)/(e+1+ga/gs), where ecomputed using different approachesis the increase of latent heat content of air per increase
Resistance Sensitivity Elimination of sensible heat content of saturated air). This can beused to give a direct estimate of the degree of stomatalPhaseolus vulgaris 50% 0% 0% (1)a
Phaseolus vulgaris 44% 28% 23% (2) control of transpiration (in terms of the relative sensitiv-Cotton: well watered 47% 25% 6% (3) ity) asCotton: stressed 59% 51% 36% (3)Tidestromia oblongifolia 75% 10% 3% (4)
(∂E/E )/(∂gs/gs)=1−V (5)Sunflower 10%b (5)Tobacco 17%b (6)
By analogy with equation (2) it is apparent that (1−V )aReferences: (1) Barradas and Jones, 1996; (2) Farquhar and von is the stomatal control coefficient for the control of
Caemmerer, 1981; (3) Jones, 1973; (4) Bjorkman et al., 1975; (5)transpiration (Cs) while V represents the ‘control’ exertedWoodrow et al., 1990; (6) Lauerer et al., 1993.
bMetabolic control analysis. by all other factors. An important consideration in the
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

Photosynthesis and transpiration control 391
use of this equation is the need for estimation of the tions can be compared with experimental data. For con-venience, it is assumed that conductance depends on yleafrelevant value of the boundary layer conductance. The
appropriate value is the transfer resistance from the plant according tocanopy to the unmodified air. For a single leaf this may
gs=gm(1+kyleaf ) (6)be a distance of a few millimetres, while for an extensivearea of homogeneous crop it may be hundreds or even where k=0.4 MPa−1, and subject to the restriction
that gs=0 if gm(1+kyleaf ) ≤0 (Jones, 1992). This equa-thousands of metres above the surface. Plant physiolo-gists, in general, have tended to overestimate the control tion gives a positive relationship between these two vari-
ables as illustrated in Fig. 3a (which also shows aexerted by stomata as a result of ignoring the canopy andregional boundary layers, thus they have tended to under- potentially more realistic continuous function; Fisher
et al., 1981). This positive relationship is what one wouldestimate V.expect where yleaf is the independent variable whichdetermines gs. In practice, however, equation (6) is onlypart of the complete control system, because gs itselfThe use of models to investigate hydraulic
feedbacks in the control of stomata affects the transpiration rate (according to the Penman–Monteith equation), and this in turn affects yleaf (Jones,
Thus far our discussion of the stomatal control of gas1992).
exchange has only explicitly taken account of the directThe other part of the control system can be modelled
effects of stomata on transpiration or assimilation. Inby treating gs as the independent variable. In this case,
reality, the situation is much more complex with feedbackfor well-coupled canopies such as isolated plants (where
control and interactions with a wide range of environ-V approaches 0), the vapour pressure at the leaf surface,
mental conditions (Fig. 1). The feedbacks have beenD, is nearly independent of gs and the rate of water loss
separated into CO2 feedback, possibly operating throughis approximately proportional to gs, so one can write
either the internal CO2 concentration (ci) or throughassimilation rate (Wong et al., 1985), and hydraulic E=Dgs (7)feedbacks dependent on aspects of stomatal or plant
The effect of increasing transpiration rate on leaf waterwater relations (Raschke, 1975; Jarvis and Davies, 1998).
potential as a result of frictional losses attributable to theAlthough the usual models of the hydraulic feedback
resistance (Rsoil–plant) in the conducting pathway isloop in the control of stomatal action are based on an
described by the Van den Honert equation asassumption that shoot water status determines stomatalaperture, there is increasing evidence from split-root yleaf=ysoil−ERsoil–plant (8)experiments and from root pressure-chamber studies that
Combining equations (7) and (8) givessoil water status may have a controlling effect on stomata(for reviews see Davies and Zhang, 1991; Jones and yleaf=ysoil−DgsRsoil–plant (9)Tardieu, 1998; but compare Fuchs and Livingston, 1996).
This equation describes a negative relationship betweenIn either case, however, it is assumed that some aspect of
yleaf and gs, which is illustrated for a range of soil waterplant water status is a critical variable determining
potentials but otherwise constant environmental condi-stomatal conductance.
tions in Fig. 3b (Rsoil–plant=2.0 MPa m2 s mol−1). TheIn what follows, the expected consequences for the
slope of this line is opposite to that in Fig. 3a whichrelationships between stomatal conductance, leaf water
suggests that a relationship with this sense would implypotential and transpiration rate of some of the main
stomatal control of yleaf, rather than vice versa.hydraulic signalling mechanisms that have been proposed
Although the solid line in Fig. 3a gives the locus forfor the control of stomatal aperture will be investigated
all possible combinations of gs and yleaf, the actual posi-and compared using simple models of plant water rela-
tion on the relationship in Fig. 3a at any time is con-tions with arbitrary parameter values chosen for conveni-
strained by the hydraulic feedback shown in the centralence (e.g. the maximum stomatal conductance, gm, is set
right-hand portion of Fig. 1 (Jarvis and Davies, 1998).at 1.0 mol m−2 s−1). The results of these predictions will
The range of possible values for any particular set ofbe compared with published data on such relationships.
conditions is a restricted subset determined by simultan-eous solution (see equation A.1 in Appendix I ) of equa-
Simple models of stomatal control: (a) response to leaftions (6) and (9). It now becomes clear that the true
water status alonedriving variables in this model are D and ysoil, even thoughthe direct mechanistic link is through yleaf. As an example,The traditional assumption is that gs depends on leaf
water status alone. Even though, as shown later, this Fig. 4 shows the possible combinations of gs and yleaf,
and of gs and ysoil, for D=2.0 mmol mol−1 as ysoil varies;assumption has been widely invalidated, analysis of theconsequences of this assumption is instructive, and predic- only values of gs below 0.4 mol m−2 s−1 are possible.
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from
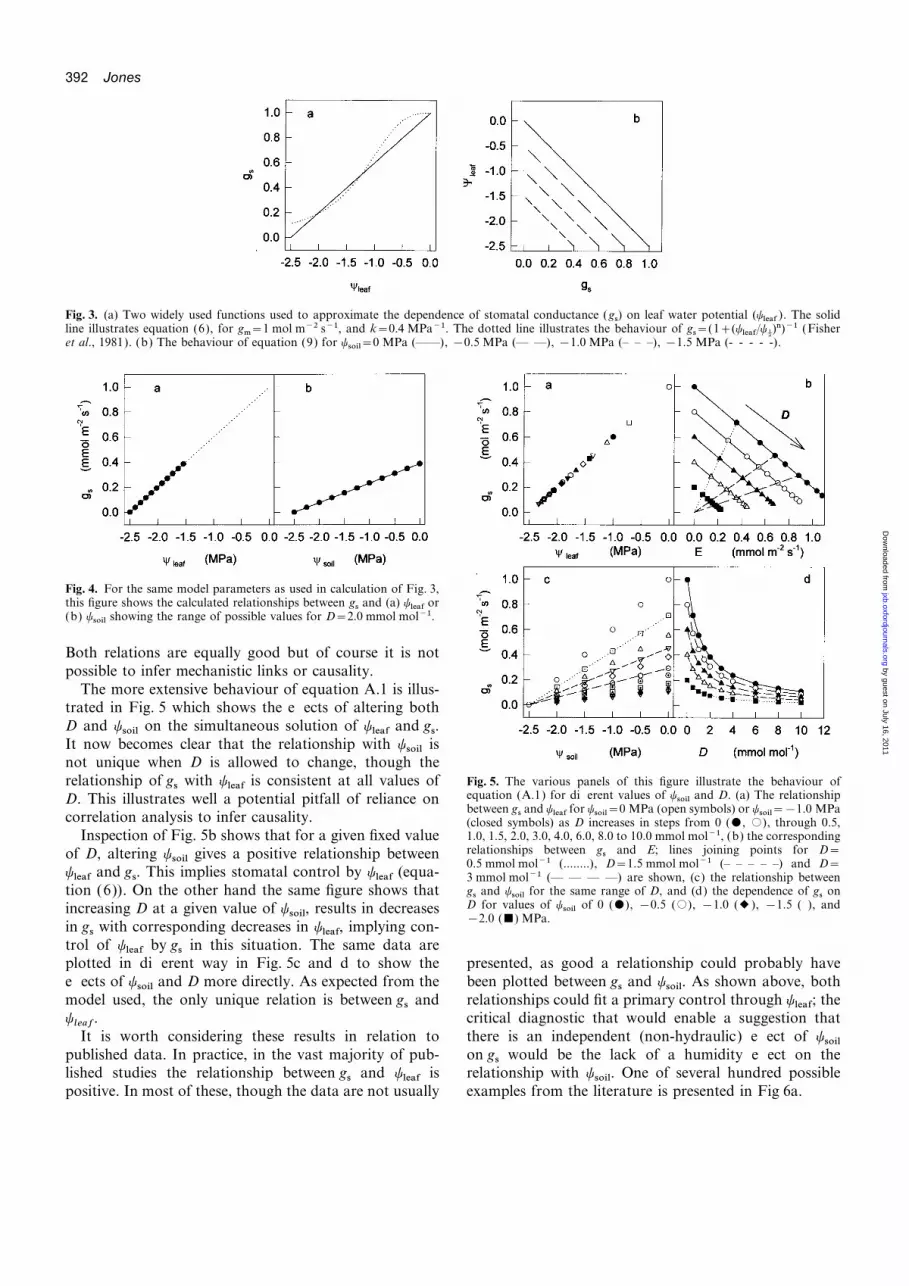
392 Jones
Fig. 3. (a) Two widely used functions used to approximate the dependence of stomatal conductance (gs) on leaf water potential (yleaf ). The solidline illustrates equation (6), for gm=1 mol m−2 s−1, and k=0.4 MPa−1. The dotted line illustrates the behaviour of gs=(1+(yleaf/yD)n)−1 (Fisheret al., 1981). (b) The behaviour of equation (9) for ysoil=0 MPa (——), −0.5 MPa (— —), −1.0 MPa (– – –), −1.5 MPa (- - - - -).
Fig. 4. For the same model parameters as used in calculation of Fig. 3,this figure shows the calculated relationships between gs and (a) yleaf or(b) ysoil showing the range of possible values for D=2.0 mmol mol−1.
Both relations are equally good but of course it is notpossible to infer mechanistic links or causality.
The more extensive behaviour of equation A.1 is illus-trated in Fig. 5 which shows the effects of altering bothD and ysoil on the simultaneous solution of yleaf and gs.It now becomes clear that the relationship with ysoil isnot unique when D is allowed to change, though the
Fig. 5. The various panels of this figure illustrate the behaviour ofrelationship of gs with yleaf is consistent at all values ofequation (A.1) for different values of ysoil and D. (a) The relationshipD. This illustrates well a potential pitfall of reliance onbetween gs and yleaf for ysoil=0 MPa (open symbols) or ysoil=−1.0 MPa
correlation analysis to infer causality. (closed symbols) as D increases in steps from 0 ($, #), through 0.5,1.0, 1.5, 2.0, 3.0, 4.0, 6.0, 8.0 to 10.0 mmol mol−1, (b) the correspondingInspection of Fig. 5b shows that for a given fixed valuerelationships between gs and E; lines joining points for D=of D, altering ysoil gives a positive relationship between0.5 mmol mol−1 (........), D=1.5 mmol mol−1 (– – – – –) and D=
yleaf and gs. This implies stomatal control by yleaf (equa- 3 mmol mol−1 (— — — —) are shown, (c) the relationship betweengs and ysoil for the same range of D, and (d) the dependence of gs ontion (6)). On the other hand the same figure shows thatD for values of ysoil of 0 ($), −0.5 (#), −1.0 (2), −1.5 ( ), andincreasing D at a given value of ysoil, results in decreases−2.0 (&) MPa.
in gs with corresponding decreases in yleaf, implying con-trol of yleaf by gs in this situation. The same data areplotted in different way in Fig. 5c and d to show the presented, as good a relationship could probably have
been plotted between gs and ysoil. As shown above, botheffects of ysoil and D more directly. As expected from themodel used, the only unique relation is between gs and relationships could fit a primary control through yleaf; the
critical diagnostic that would enable a suggestion thatyleaf
.It is worth considering these results in relation to there is an independent (non-hydraulic) effect of ysoil
on gs would be the lack of a humidity effect on thepublished data. In practice, in the vast majority of pub-lished studies the relationship between gs and yleaf is relationship with ysoil. One of several hundred possible
examples from the literature is presented in Fig 6a.positive. In most of these, though the data are not usually
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

Photosynthesis and transpiration control 393
Fig. 6. (a) A typical set of experimental data obtained from a field experiment on sorghum at a range of soil moisture contents showing a positiverelationship between gs and yleaf (redrawn from Henzell et al., 1976), (b) an example for apple where a negative relationship between gs and yleafhas been observed; open symbols refer to well irrigated plants and closed symbols to droughted plants (from Jones, 1985b), (c) the relationshipbetween gs and yleaf. for field- ($) and greenhouse-grown (l ) sugarcane in response to manipulations of leaf area (Meinzer and Grantz, 1990), and(d) the relationship between gs and yleaf. for field-grown maize over a growing season in non-compacted (open symbols) and compacted (closedsymbols) soil (replotted from data in Tardieu, 1993).
Less frequently, but importantly, a number of examples yleaf. For example, one might hypothesize a direct stomatalresponse to either humidity deficit (Raschke, 1975;of negative relationships between gs and yleaf have been
observed (Fig. 6b). It is interesting to note that the Grantz, 1990) or to evaporation rate (e.g. the feedforwardresponse of Cowan, 1977) that does not involve feedbackpositive relationship tends to occur where the variable
being altered is the soil water status, as in standard soil through bulk leaf water status. If one assumes a linearstomatal response to E, as indrying experiments. Where, however, the evaporation rate
is manipulated, for example, by changes in air humiditygs=gm(1−aE ) (11)or other environmental conditions, the opposite slope is
often observed (Morison and Gifford, 1983). Cases of (again subject to a minimum gs=0), one can substitutesuch negative relationships are probably much more from equation (7) to get the equivalent response to Dcommon than is usually recognized. Not only do they
gs=gm/(1+agmD) (12)probably underlie much of the apparent stomatal responseto humidity (Monteith, 1995), but they probably underlie These two primary assumptions are indistinguishablemuch of the variation between leaves or plants within because of their linkage through equation (7) (thoughany drought treatment. These negative relationships imply see Mott and Parkhurst, 1991, who concluded fromthat stomatal conductance may have a greater role in measurements in Helox that stomata respond to thecontrolling plant water status than is often implied by ‘evaporative potential of the air’, i.e. the product ofthe analysis of standard drought experiments. diffusion coefficient and the water vapour concentration
difference). Either assumption gives the responses shownSimple models of stomatal control: (b) environmental
in Fig. 7, where the relationships between gs and both Eresponses without feedback
and D are unique. Although a majority of studies(Monteith, 1995) have shown that gs tends to decreaseThe models used so far have included hydraulic feedback.
It is instructive to compare these results with those approximately linearly with E when D changes (with aninverse hyperbolic relationship to D as in equation (12))expected where the environmental response of stomata is
assumed not to involve a hydraulic feedback through as shown in Fig. 7, this is not always apparent. For
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from
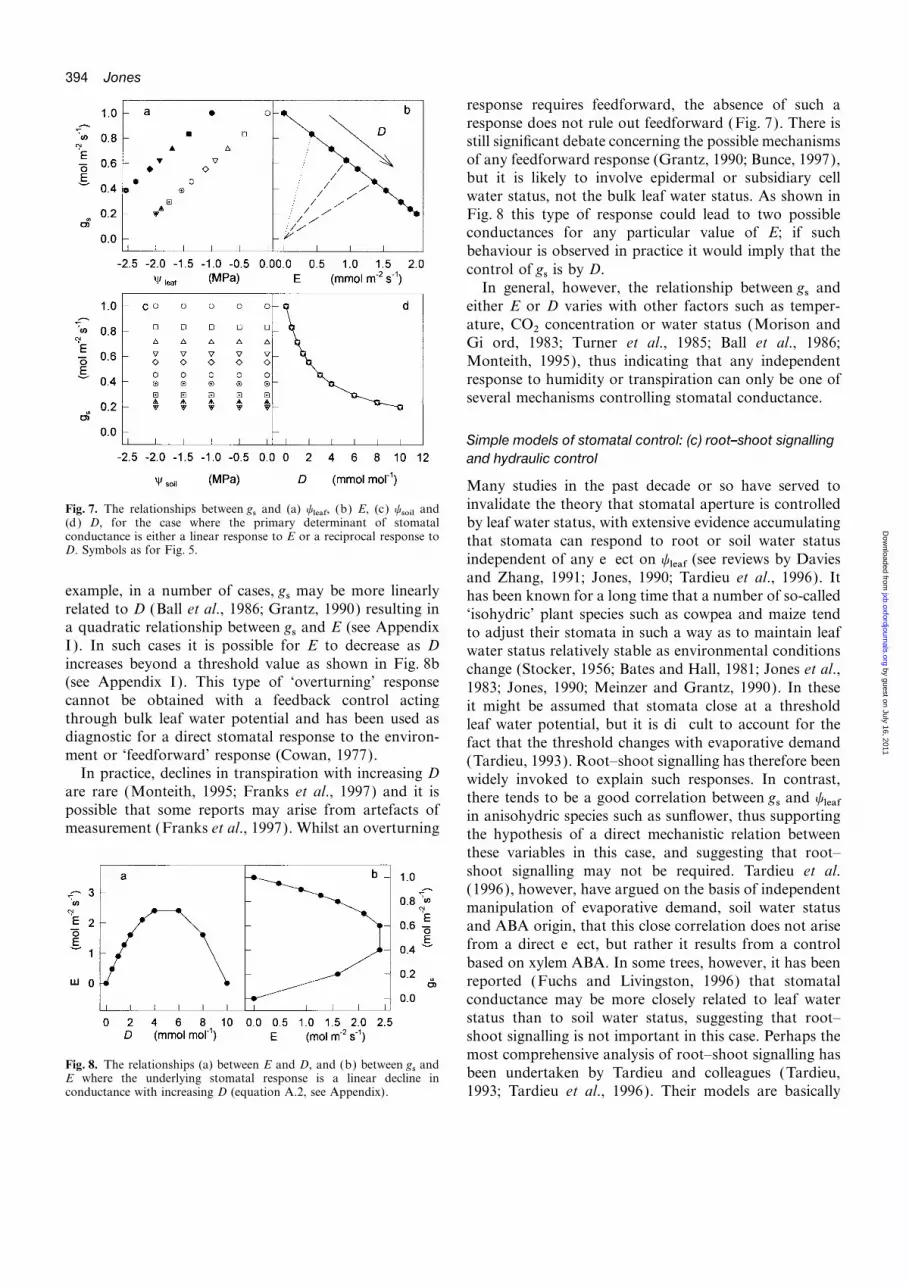
394 Jones
response requires feedforward, the absence of such aresponse does not rule out feedforward (Fig. 7). There isstill significant debate concerning the possible mechanismsof any feedforward response (Grantz, 1990; Bunce, 1997),but it is likely to involve epidermal or subsidiary cellwater status, not the bulk leaf water status. As shown inFig. 8 this type of response could lead to two possibleconductances for any particular value of E; if suchbehaviour is observed in practice it would imply that thecontrol of gs is by D.
In general, however, the relationship between gs andeither E or D varies with other factors such as temper-ature, CO2 concentration or water status (Morison andGifford, 1983; Turner et al., 1985; Ball et al., 1986;Monteith, 1995), thus indicating that any independentresponse to humidity or transpiration can only be one ofseveral mechanisms controlling stomatal conductance.
Simple models of stomatal control: (c) root–shoot signallingand hydraulic control
Many studies in the past decade or so have served toinvalidate the theory that stomatal aperture is controlledFig. 7. The relationships between gs and (a) yleaf, (b) E, (c) ysoil and
(d) D, for the case where the primary determinant of stomatal by leaf water status, with extensive evidence accumulatingconductance is either a linear response to E or a reciprocal response to that stomata can respond to root or soil water statusD. Symbols as for Fig. 5.
independent of any effect on yleaf (see reviews by Daviesand Zhang, 1991; Jones, 1990; Tardieu et al., 1996). It
example, in a number of cases, gs may be more linearly has been known for a long time that a number of so-calledrelated to D (Ball et al., 1986; Grantz, 1990) resulting in ‘isohydric’ plant species such as cowpea and maize tenda quadratic relationship between gs and E (see Appendix to adjust their stomata in such a way as to maintain leafI ). In such cases it is possible for E to decrease as D water status relatively stable as environmental conditionsincreases beyond a threshold value as shown in Fig. 8b change (Stocker, 1956; Bates and Hall, 1981; Jones et al.,(see Appendix I). This type of ‘overturning’ response 1983; Jones, 1990; Meinzer and Grantz, 1990). In thesecannot be obtained with a feedback control acting it might be assumed that stomata close at a thresholdthrough bulk leaf water potential and has been used as leaf water potential, but it is difficult to account for thediagnostic for a direct stomatal response to the environ- fact that the threshold changes with evaporative demandment or ‘feedforward’ response (Cowan, 1977). (Tardieu, 1993). Root–shoot signalling has therefore been
In practice, declines in transpiration with increasing D widely invoked to explain such responses. In contrast,are rare (Monteith, 1995; Franks et al., 1997) and it is there tends to be a good correlation between gs and yleafpossible that some reports may arise from artefacts of in anisohydric species such as sunflower, thus supportingmeasurement (Franks et al., 1997). Whilst an overturning the hypothesis of a direct mechanistic relation between
these variables in this case, and suggesting that root–shoot signalling may not be required. Tardieu et al.(1996), however, have argued on the basis of independentmanipulation of evaporative demand, soil water statusand ABA origin, that this close correlation does not arisefrom a direct effect, but rather it results from a controlbased on xylem ABA. In some trees, however, it has beenreported (Fuchs and Livingston, 1996) that stomatalconductance may be more closely related to leaf waterstatus than to soil water status, suggesting that root–shoot signalling is not important in this case. Perhaps themost comprehensive analysis of root–shoot signalling has
Fig. 8. The relationships (a) between E and D, and (b) between gs andbeen undertaken by Tardieu and colleagues (Tardieu,E where the underlying stomatal response is a linear decline in
conductance with increasing D (equation A.2, see Appendix). 1993; Tardieu et al., 1996). Their models are basically
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

Photosynthesis and transpiration control 395
extensions of those described in this article and cover thesituations for a number of anisohydric and isohydricspecies.
A strongly favoured hypothesis to explain root–shootsignalling has been that abscisic acid (ABA) or someother signalling compound is synthesized by roots inresponse to soil drying (Davies and Zhang, 1991). Thesignal compound is then transported in the xylem to theleaves, though there is still some uncertainty as to whetherstomatal conductance is better related to the concentrationof ABA in the xylem sap or to the rate of arrival of ABA(Gowing et al., 1993). [As an aside it is somewhat difficultto envisage a stomatal regulation mechanism that dependsdirectly on the rate of supply of a signal compound;rather it seems likely that an apparent response to arrivalrate results from the balance between arrival rate andany removal mechanism such as metabolism affectingconcentration at a receptor.]
On the basis of this model, the rate of ABA transportto the shoots in a steady-state should depend only on therate of synthesis of ABA in the roots, even though theABA concentration in the xylem sap would depend also
Fig. 9. The relationships between gs and (a) yleaf, (b) E, (c) ysoil andon the rate of water flow that effectively dilutes the signal(d) D, expected for the model where gs depends linearly on the flux ofconcentration. If one assumes as a simplification that theABA from the roots (symbols as for Fig. 5).
rate of synthesis of ABA (JABA) is linearly related to rootwater potential (i.e. JABA=−ayroot; Tardieu, 1993), andthat yroot=ysoil, changes in the stomatal conductance found that it was also necessary to modulate the stomatal
sensitivity to [ABA] as a function of yleaf to fit their datawould have no direct feedback effect on the rate of ABAsupply, so that gs would be uniquely related to ysoil adequately.(Appendix I; Fig. 9). The assumption of a sequence ofsteady-states, however, is likely to be an oversimplification Statistical assessment of stomatal effectiveness(Tardieu, 1993). in control
In spite of the indications of the importance of ysoil incontrolling gs (Turner et al., 1985; Davies and Zhang, It has been known for a long time that in isohydric plants
stomata tend to adjust in such a way as to maintain leaf1991) a model based on a response to the rate of ABAsupply does not fit the widespread observations that gs is water status relatively stable (Stocker, 1956). It is there-
fore common in field experiments (Fig. 6c, d) to observesensitive to D (Grantz, 1990), or the fact that otherstudies (Ferreira and Katerji, 1992; Fuchs and Livingston, a wide range of stomatal conductances for a rather limited
range of leaf water potentials (Bates and Hall, 1981;1996) have shown that stomatal conductance may notalways be well related to soil water potential. As an Jones et al., 1983; Jones, 1990; Meinzer and Grantz,
1990). This behaviour is what has been predicted byalternative it is more commonly assumed that stomatarespond to the ABA concentration in the xylem ([ABA]), Jones and Sutherland (1991) if stomata were to operate
to maximize productivity by avoiding xylem cavitation.which would be proportional to (JABA/E). This systemdoes not in general have a stable solution; stomatal It has even been suggested (Jones, 1974) that information
on the variability of stomatal conductance relative to theclosure tends to decrease E, and hence increase theconcentration of ABA, which in turn leads to further variability of yleaf can be a useful indicator of the plant’s
efficiency at controlling leaf water status in response toclosure, and so on. It is, of course possible that suchunstable feedbacks are advantageous in a rapidly chan- developing stress. A stomatal control index, I, was defined
by Jones (1974) asging environment where steady-states are not achieved(Farquhar, 1973).
I=s2( lngs)/s2( lnyleaf ) (10)Tardieu (1993) and others (Johnson et al., 1991) have
got round this instability problem by including the where s2( lngs) is the short-term variance of ( lngs), ands2( lnyleaf ) is the short-term variance of ( lnyleaf ). I is ahydraulic flow resistance between the soil and the root,
so that yroot becomes dependent on flow rate in the same measure of the degree to which a particular variety orspecies controls leaf water potential by variation inway as yleaf does in equation (9). Indeed Tardieu (1993)
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

396 Jones
stomatal conductance. Log transformed data are used to stomatal conductance, and so on, together with transportof possible signalling molecules in the xylem. Thereensure that variances are approximately independent of
the mean. Values of I as great as 20 (indicating a high therefore remains a need for more complete data setsfrom a range of situations to clarify the balance betweendegree of stability in yleaf ) were found on occasions for
wheat growing in the UK, while similar stomatally con- the different possible controls.trolled homeostasis of yleaf has been found in many othersituations (Fig. 6; Stocker, 1956; Kanemasu and Tanner,
Appendix I1969; Bates and Hall, 1981; Tardieu et al., 1992).Solution of hydraulic feedback
Concluding discussion Simultaneous solution of equations (6) and (9) gives the valueof yleaf asAlthough techniques are available for describing the
yleaf=(ysoil−bDRsoil–plantgm)/(1−bDRsoil–plantgmk) (A.1)importance of stomata in controlling photosynthesis, thedegree to which the choice of method is subjective is not with the corresponding value of gs being given by equation (6).widely recognized. There is, however, general agreement
Conductance directly related to Dthat at least in well-adapted plants the stomata play arelatively small part in determining the rate of photosyn-
When stomatal conductance is linearly related to D, bythesis, comprising less than about 20% of the totalphotosynthetic limitation. Notwithstanding this, stomata gs=gm(1−aD); for D≤1/a
gs=0; for all other values of D(A.2)
may play a major part in determining the difference inassimilation rate between two plants or treatments (Jones, substituting from equation (7) and rearranging, gives (for1985a), but there still remains a need to develop a rigorous D≤1/a)yet widely acceptable approach to defining the role of gs2−gmgs+agmE=0 (A.3)stomata in causing such changes in assimilation rate
which can be solved by the usual method for a quadratic.between two plants or two treatments.Similarly the relationship between E and D is quadratic
The complex feedbacks involved in stomatal operation (Fig. 8b).mean that is often difficult to decide whether stomata arecontrolling gas-exchange or vice versa. Although it is Root–shoot signallingapparent from other articles in this issue that a lot is
If one assumes that stomatal conductance depends on the rateknown about the detailed molecular processes involvedof supply of ABA (JABA) according to
in stomatal movement, significant uncertainty about thegs=gm(1−kJABA) (A.4)physiological controls and their interactions remains. The
implication of the diversity of observed environmental and thatresponses of stomata is that a single response mechanism JABA=−ayroot$−aysoil (A.5)cannot be expected to explain all features of stomatal
one getsbehaviour. This paper has summarized the types ofresponse expected for a limited number of different gs=gm(1+akysoil ) (A.6)stomatal control mechanisms (with discussion beingrestricted to a few possible aspects of the hydrauliccontrols). Comparison of these can be valuable in elimin-ating possible mechanisms, but agreement between the Referencespredictions from any proposed mechanism and data
Assmann SM. 1988. Stomatal and non-stomatal limitations tocannot be taken as confirmation that the mechanism is carbon assimilation: an evaluation of the path-dependentrelevant. method. Plant, Cell and Environment 11, 577–82.
Ball JT, Woodrow IE, Berry JA. 1986. A Model predictingAlthough there are data-sets in the literature which canstomatal conductance and its contribution to the control ofbe used to support each of the proposed mechanisms,photosynthesis under different environmental conditions. In:there has been particular emphasis on root–shoot signal-Biggins J, ed. Proceedings of VII International Photosynthesis
ling in recent years. It should not be concluded, however, Congress. Dordrecht: Martinus Nijhoff, 221–34.that such signalling is the only mechanism that occurs, Bange GGJ. 1953. On the quantitative explanation of stomatal
transpiration. Amsterdam: North-Holland Publishingas it may often occur in conjunction with the ‘classical’Company.control by leaf water status and other environmental
Barradas V, Jones HG. 1996. Responses of CO2 assimilation tocontrols. Unfortunately, it is rather rare that experimentschanges in irradiance: laboratory and field data and a model
measure all the relevant data including water status in for beans (Phaseolus vulgaris L.). Journal of Experimentaldifferent parts of the soil–plant system, environmental Botany 47, 639–45.
Bates LM, Hall AE. 1981. Stomatal closure with soil moistureconditions, assimilation rate, evaporation rate, and
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

Photosynthesis and transpiration control 397
depletion not associated with changes in bulk water status. ures with two rainforest trees: short communication. Plant,Cell and Environment 20, 142–5.Oecologia 50, 62–5.
Friend AD. 1991. Use of a model of photosynthesis and leafBjorkman O, Boardman NK, Anderson JM, Thorne SW,microenvironment to predict optimal stomatal conductanceGoodchild DJ, Pyliotis NA. 1972. The effect of light intensityand leaf nitrogen partitioning. Plant, Cell and Environmentduring growth of Atriplex patula on the capacity of photosyn-14, 895–905.thetic reactions, chloroplast components and structure.
Fuchs EE, Livingston NJ. 1996. Hydraulic control of stomatalCarnegie Institution Year Book 71, 115–35.conductance in Douglas fir [Pseudotsuga menziesii (Mirb.)Bjorkman O, Mooney HA, Ehleringer JR. 1975. PhotosyntheticFranco] and alder [Alnus rubra (Bong)] seedlings. Plant, Cellresponses of plants from habitats with contrasting thermaland Environment 19, 1091–8.environments. Carnegie Institution Year Book 74, 743–8.
Givnish TJ. 1986. Optimal stomatal conductance, allocation ofBrown H, Escombe F. 1900. Static diffusion of gases and liquidsenergy between leaves and roots, and the marginal cost ofin relation to the assimilation of carbon and translocation intranspiration. In: Givnish TJ, ed. On the economy of plantplants. Philosophical Transactions of the Royal Society ofform and function. Cambridge University Press, 171–213.London, Series B 193, 223.
Gowing DJG, Jones HG, Davies WJ. 1993. Xylem transportedBunce JA. 1997. Does transpiration control stomatal responsesabscisic acid: the relative importance of its mass and itsto water vapour pressure deficit? Plant, Cell and Environmentconcentration in the control of stomatal aperture. Plant, Cell20, 131–5.and Environment 16, 453–9.Burke JJ, Mahan JR, Hatfield JL. 1988. Crop-specific thermal
Grantz DA. 1990. Plant response to humidity. Plant, Cell andkinetic windows in relation to wheat and cotton biomassEnvironment 13, 667–79.production. Agronomy Journal 80, 553–6.
Guehl JM, Aussenac G. 1987. Photosynthesis decrease andCochard H, Breda N, Granier A. 1996. Whole tree hydraulicstomatal control of gas exchange in Abies alba Mill. inconductance and water loss regulation in Quercus duringresponse to vapour pressure difference. Plant Physiologydrought—evidence for stomatal control of embolism. Annales83, 316–22.des Sciences Forestieres 53, 17–206.
Henzell RG, McCree KJ, van Bavel CHM, Schertz KF. 1976.Cowan IR. 1972. Oscillations in stomatal conductance and plantSorghum variation in stomatal sensitivity to leaf water deficit.functioning associated with stomatal conductance:Crop Science 16, 660–2.Observations and a model. Planta 106, 185–219.
Jarvis AJ, Davies WJ. 1998. Modelling stomatal responses toCowan IR. 1977. Stomatal behaviour and environment. Advancessoil and atmospheric drought. Journal of Experimental Botanyin Botanical Research 4, 117–228.49, Special Issue, 399–406.Cowan IR. 1982. Water-use and optimization of carbon
Johnson IR, Melkonian JJ, Thornley JHM, Riha SJ. 1991. Aassimilation. In: Lange OL, Nobel PS, Osmond CB, Zieglermodel of water flow through plants incorporating shoot/rootH, eds. Encyclopedia of plant physiology, New series, Vol. 12B.‘message’ control of stomatal conductance. Plant, Cell andBerlin, Heidelberg, New York: Springer-Verlag, 589–613.Environment 14, 531–44.Cowan IR. 1986. Economics of carbon fixation in higher plants. Jones HG. 1973. Limiting factors in photosynthesis. NewIn: Givnish TJ, ed. On the economy of plant form and function. Phytologist 72, 1089–94.Cambridge University Press, 133–70. Jones HG. 1974. Assessment of stomatal control of plant water
Cowan IR, Farquhar GD. 1977. Stomatal function in relation status. New Phytologist 73, 851–9.to leaf metabolism and environment. Symposium of Society Jones HG. 1985a. Partitioning stomatal and non-stomatalfor Experimental Biology 31, 471–505. limitations to photosynthesis. Plant, Cell and Environment
Davies WJ, Zhang J. 1991. Root signals and the regulation of 8, 95–104.growth and development of plants in drying soil. Annual Jones HG. 1985b. Physiological mechanisms involved in theReview of Plant Physiology and Plant Molecular Biology control of leaf water status: implications for the estimation42, 55–76. of tree water status. Acta Horticulturae 171, 291–6.
Farquhar GD. 1973. A study of the responses of stomata to Jones HG. 1990. Physiological aspects of the control of waterperturbations of the environment. PhD thesis, Australian status in horticultural crops. HortScience 25, 19–26National University, Canberra. Jones HG. 1992. Plants and microclimate: a quantitative
Farquhar GD. 1978. Feedforward responses of stomata to approach to environmental plant physiology, 2nd edn.humidity. Australian Jounal of Plant Physiology 5, 769–72. Cambridge University Press.
Farquhar GD, Sharkey TD. 1982. Stomatal conductance and Jones HG. 1995. Photosynthetic limitations: use in guidingphotosynthesis. Annual Review of Plant Physiology 33, 317–45. effort in crop improvement. Journal of Experimental Botany
Ferreira MI, Katerji N. 1992. Is stomatal conductance in a 46, 1415–22.tomato crop controlled by soil or atmosphere? Oecologia Jones HG, Luton MT, Higgs, KH, Hamer PJC. 1983.92, 104–7. Experimental control of water status in an apple orchard.
Finnigan JJ, Raupach MR. 1987. Transfer processes in plant Journal of Horticultural Science 58, 301–16.canopies in relation to stomatal characteristics. In: Zeiger E, Jones HG, Sutherland RA. 1991. Stomatal control of xylemFarquhar GD, Cowan IR, eds. Stomatal function. Stanford embolism. Plant, Cell and Environment 6, 607–12.University Press, 385–429. Jones HG, Tardieu F. 1998. Modelling water relations of
Fisher MJ, Charles-Edwards DA, Ludlow MM. 1981. An horticultural crops: a review. Scientia Horticulturae (in press).analysis of the effects of repeated short-term water deficits Kacser H, Burns JA. 1973. The control of flux. Symposium ofon stomatal conductance to carbon dioxide and leaf photosyn- the Society for Experimental Biology 27, 65–107.thesis by the legume Macroptilium atropupureum cv. Siratro. Kanemasu ET, Tanner CB. 1969. Stomatal diffusion resistanceAustralian Journal of Plant Physiology 8, 347–57. of snap beans. I. Influence of leaf water potential. Plant
Franks PJ, Cowan IR, Farquhar GD. 1997. The apparent Physiology 44, 1547–52.feedforward responses of stomata to air vapour pressure Lauerer M, Saftic D, Quick WP, Labate C, Fichtner K, Schulze
E-D, Rodermel SR, Bogorad L, Stitt M. 1993. Decreaseddeficit: information revealed by different experimental proced-
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from

398 Jones
ribulose-1,5-bisphosphate carboxylase-oxygenase in trans- Stitt M, Quick MP, Schurr U, Schulze E-D, Rodermel SR,Bogorad L. 1991. Decreased ribulose-1,5-bisphosphate carb-genic tobacco transformed with ‘antisense’ rbcS VI. Effect on
photosynthesis in plants grown at high irradiance. Planta oxylase-oxygenase in transgenic tobacco transformed with‘antisense’ rbcS. II. Flux-control coefficients for photosyn-190, 332–45.
Lloyd FE. 1908. The physiology of stomata. Carnegie Institution thesis in varying light, CO2, and air humidity. Planta183, 555–66.of Washington Bulletin 82.
Mahan JR, Upchurch DR. 1988. Maintenance of constant leaf Stocker O. 1956. Die Abhangigkeit des transpiration von denunweltfaktoren. In: Ruhland W. ed. Encyclopedia of planttemperature by plants. I. Hypothesis—limited homeothermy.
Environmental and Experimental Botany 28, 351–7. physiology, Vol. 3. Springer-Verlag.Tardieu F. 1993. Will increases in our understanding of soil-Maskell EJ. 1928. Experimental researches on vegetable assim-
ilation and respiration. XVIII. The relation between stomata root relations and root signalling substantially alter waterflux models? Philosophical Transactions of the Royal Societyopening and assimilation—a critical study of assimilation
rates and porometer rates of cherry laurel. Proceedings of the of London, B 341, 57–66.Tardieu F, Zhang J, Katerji N, Bethenod O, Palmer S, DaviesRoyal Society, Series B 102, 488–533.
McNaughton KG, Jarvis PG. 1983. Predicting the effects of WJ. 1992. Xylem ABA controls the stomatal conductance offield-grown maize subjected to soil compaction or drying.vegetation changes on transpiration and evaporation. In:
Kozlowski TT, ed. Water deficits and plant growth 7. New Plant, Cell and Environment 15, 193–7.Tardieu F, Zhang J, Gowing DJC. 1993. Stomatal control byYork: Academic Press, 1–47.
Meidner H, Mansfield TA. 1968. Physiology of stomata. both [ABA] in the xylem sap and leaf water status: test of amodel and alternative hypotheses for droughted or ABA-fedMaidenhead: McGraw-Hill.
Meinzer FC, Grantz DA. 1990. Stomatal conductance in growing field-grown maize. Plant, Cell and Environment 16, 413–20.Tardieu F, Lafarge T, Simonneau Th. 1996. Stomatal control bysugarcane: stomatal adjustment to water transport capacity.
Plant, Cell and Environment 13, 383–8. fed or endogenous xylem ABA in sunflower: interpretationof correlations between leaf water potential and stomatalMonteith JL. 1995. A reinterpretation of stomatal responses to
humidity. Plant, Cell and Environment 18, 357–64. conductance in anisohydric species. Plant, Cell andEnvironment 19, 75–84.Morison JIL, Gifford RM. 1983. Stomatal sensitivity to carbon
dioxide and humidity. Plant Physiology 71, 789–96. Turner NC, Schulze E-D, Gollan T. 1985. The response ofstomata and leaf gas exchange to vapour pressure deficitsMott KA, Parkhurst DF. 1991. Stomata response to humidity
in air and helox. Plant, Cell and Environment 14, 509–15. and soil water content. II. In the mesophytic herbaceousspecies. Helianthus annuus. Oecologia 65, 348–55.Parkhurst DF, Loucks OL. 1972. Optimal leaf size in relation
to environment Journal of Ecology 60, 505–37. Tyree MT, Sperry JS. 1988. Do woody plants operate near thepoint of catastrophic xylem dysfunction caused by dynamicPeisker M, Vaclavık J. 1987. A simple path-independent method
for apportioning nonstomatal and stomatal contributions to water stress? Plant Physiology 88, 574–80.Woodrow IE, Ball JT, Berry JA. 1990. Control of photosyntheticdiminutions in net CO2 uptake rate. Biochemie. Physiologie
Pflanzen 182, 31–40. carbon dioxide fication by the boundary layer, stomata andribulose-1,5-bisphosphate carboxylase-oxygenase. Plant, CellRaschke K. 1975. Stomatal action. Annual Review of Plant
Physiology 26, 309–40. and Environment 13, 339–47.Woodrow IE. 1994. Control of steady-state photosynthesis inRodermel SR, Abbott MS, Bogorad L. 1988. Nuclear-organelle
interactions: nuclear antisense gene inhibits ribulose- sunflowers growing in enhanced CO2. Plant, Cell andEnvironment 17, 277–86.1,5-bisphosphate carboxylase enzyme levels in transformed
tobacco plants. Cell 56, 673–81. Wong SC, Cowan IR, Farquhar GD. 1985. Leaf conductance inrelation to rate of CO2 assimilation. I. Influence of nitrogenSmall JR, Kacser H. 1993. Responses of metabolic systems to
large changes in enzyme activities and effectors. I. The linear nutrition, phosphorous nutrition, photon flux density andambient partial pressure of CO2 during ontogeny. Planttreatment of unbranched chains. European Journal of
Biochemistry 213, 613–24. Physiology 78, 821–5.
by guest on July 16, 2011jxb.oxfordjournals.org
Dow
nloaded from





























