Embed Size (px)
Citation preview

Free Radical Biology & Medicine, Vol. 36, No. 9, pp. 1126 –1133, 2004Copyright D 2004 Elsevier Inc.
Printed in the USA. All rights reserved0891-5849/$-see front matter
doi:10.1016/j.freeradbiomed.2004.02.001
Original Contribution
OXYGEN TENSION REGULATES REACTIVE OXYGEN GENERATION
AND MUTATION OF Helicobacter pylori
AH-MEE PARK,* QUAN LI,* KUMIKO NAGATA,y TOSHIHIDE TAMURA,
y KUNIO SHIMONO,z
EISUKE F. SATO,* and MASAYASU INOUE*,b
*Department of Biochemistry and Molecular Pathology, Osaka City University Medical School, 1-4-3 Asahimachi,Abeno, Osaka 545-8585, Japan; yDepartment of Bacteriology, Hyogo College of Medicine, 1-1 Mukogawa,
Nishinomiya, Hyogo 663-8501, Japan; and zHinode-En Elder Health Center, Ohishi-Minamimachi,Nada, Kobe, Hyogo 657-0852, Japan
(Received 3 October 2003; Revised 21 January 2004; Accepted 2 February 2004)
Ad
Bioche
School
6645-3
Abstract—Although both bacillary and coccoid forms of Helicobacter pylori reside in human stomach, the
pathophysiological significance of the two forms remains obscure. The present work describes the effect of oxygen
tension on the transformation and reactive oxygen species (ROS) metabolism of this pathogen. Most H. pylori cultured
under an optimum O2 concentration (7%) were the bacillary form, whereas about 80% of cells cultured under aerobic or
anaerobic conditions were the coccoid form. The colony-forming unit of H. pylori decreased significantly under both
aerobic and anaerobic culture conditions. The bacillary form of H. pylori generated predominantly superoxide radical,
whereas the coccoid form generated preferentially hydroxyl radical. Specific activities of cellular respiration, urease, and
superoxide dismatase decreased markedly after transformation of the bacillary form to the coccoid form, with
concomitant generation of protein carbonyls and 8-hydroxyguanine. The frequency of mutation of cells increased
significantly during culture under nonoptimum O2 conditions. These results indicate that ROS generated by H. pylori
catalyze the oxidative modification of cellular DNA, thereby enhancing the transformation from the bacillary to the
coccoid form. The enhanced generation of mutagenic hydroxyl radicals in the coccoid form might accelerate mutation
and increase the genetic diversity of H. pylori. D 2004 Elsevier Inc. All rights reserved.
Keywords—Helicobacter pylori, Oxidative stress, Superoxide, Mutation, Gastric cancer, Free radicals
INTRODUCTION
Helicobacter pylori is a gram-negative and microaerobic
bacterium that plays important roles in the pathogenesis
of gastritis, peptic ulcer, and gastric cancer [1,2]. Two
types of H. pylori, bacillary and coccoid forms, are seen
in human gastric mucosa; the bacillary form is the
predominant form found in human stomach, and the
coccoid form is the major form found in extragastric
environments [3]. Both oral-to-oral and fecal-to-oral
routes have been postulated to be involved in the
dress correspondence to: Dr. Masayasu Inoue, Department of
mistry and Molecular Pathology, Osaka City University Medical
, 1-4-3 Asahimachi, Abeno, Osaka 545-8585, Japan; Fax: +81-6-
721; E-mail: [email protected].
1126
mechanism of infection of human subjects [4–6]. We
previously reported that the oxygen tension in gastric
juice changes significantly depending on the injected
solutions [7]. Gastric mucosal oxygen tension has been
reported to decrease significantly in animals exposed to
restraint stress [8]. When the bacillary form of H. pylori
is exposed to various stresses, such as unfavorable O2
tension and antibiotics, it undergoes transformation to the
coccoid form [9]. The coccoid form of H. pylori has been
postulated to be viable but difficult to grow under culture
conditions [10]. The mechanism and pathophysiological
significance of the transformation of H. pylori remain
unknown. It has been postulated that the transformation
of H. pylori from the bacillary to the coccoid form might
occur irreversibly, and, hence, the latter form might
represent a dormant stage in the life cycle [11,12].
However, the coccoid form of H. pylori has been known

Transformation of H. pylori 1127
to exhibit activities to synthesize DNA and proteins [13].
Moreover, both forms of H. pylori are known to adhere
to epithelial cells and activated cellular metabolism,
including tyrosine phosphorylation of specific proteins
[14]. Thus, not only the bacillary form but also the
coccoid form of H. pylori has been postulated to con-
tribute to infection [15]. We previously showed that the
bacillary form of H. pylori generates the superoxide
radical, and the coccoid form generates preferentially
the hydroxyl radical [16]. These results suggested that
the transformation of H. pylori might affect its biological
properties, including infectivity and toxicity to gastric
mucosal cells. To understand the roles of the transfor-
mation of H. pylori in the mechanism of infection and in
the pathogenesis of gastric mucosal injury, we studied the
effects of O2 tension on its activities urease, SOD, and
cytochrome c oxidase activities; generation of reactive
oxygen species, oxidative modification of proteins and
DNA, and growth.
EXPERIMENTAL PROCEDURES
Materials
The reagents used in the present experiments, such as
2-methyl-6-[ p-methoxyphenyl]-3,7-dihydroimidazol
[1,2- a]pyrazin-3-one (MCLA), 8-amino-5-chloro-7-phe-
nyl-pyridopyridazine (L012), Brucella broth and agar,
and ascorbic acid, were purchased from Tokyo Kasei
(Tokyo, Japan), Wako Company (Osaka, Japan), Becton
Dickinson (Cockeysville, MD, USA) and Sigma Chem-
ical Company. (St Louis, MO, USA). All other reagents
used were of the highest grade commercially available
and were obtained from Wako Company.
H. pylori NCTC-11637 was cultured in Brucella broth
containing 5% horse serum under optimal microaerobic
conditions (7% O2) with gentle shaking at 37jC [13]. H.
pylori was also cultured under anaerobic (<1% O2) and
aerobic (21% O2) conditions without using CO2. Anae-
roPack Campylo and AnaeroPack Keep (Mitsubishi Gas
Chemical, Tokyo, Japan) were used to obtain micro-
aerobic and anaerobic atmospheres, respectively. The
number of suspended cells was determined optically;
an optical density of 0.2 at 550 nm corresponds to
approximately 108 cells/ml. The number of living cells
was determined by a colony-forming unit (CFU). Cul-
tured cells were washed with 10 mM Hepes buffer (pH
7.4) containing 0.9% NaCl and used for experiments.
Assay of cellular respiration and enzyme activities
Respiration and cytochrome c oxidase activity of H.
pylori were polarographically monitored using a Clark-
type oxygen electrode (Rank Brothers, Ltd., Cambridge,
UK) in a closed cell containing 10 mM Hepes buffer (pH
7.4), 0.9% NaCl, and 108 to 109 cells/ml at 37jC as
described previously [17]. Pyruvate 5 mM and ascorbic
acid 5 mM with N,N,NV,NV-tetramethyl-p-phenylenedi-
amine (TMPD) 0.5 mM were used as substrates for
respiration and cytochrome c oxidase, respectively. Urease
and SOD activities were assayed essentially by the me-
thod’s of Ferraro et al. [18] and Crapo et al. [19],
respectively, after solubilization with 1% Triton X-100
containing 10 mM potassium phosphate (pH 7.4) at 4jCfor 10 min, followed by centrifugation at 15,000g for 10
min. Enzyme activities in the soluble fractions were
determined.
Analysis of reactive oxygen species
Superoxide and hydroxyl radicals generated by H.
pylori (5 � 108 cells/ml) were assayed in 10 mM Hepes
buffer (pH 7.4) containing 0.9% NaCl and 0.5% Triton
X-100 in the presence of either 1 AM MCLA [20] or 0.4
mM L012 [21] using a luminescence reader BLR-201
(Aloka Co., Tokyo, Japan). To determine the specificity
of MCLA and L012 for reactive oxygen species (ROS),
chemiluminescence intensity was analyzed in the pres-
ence and absence of either deferoxamine (50 AM) or Cu/
Zn-SOD (200 U/ml).
Analysis of protein carbonyls
Oxidatively modified proteins in H. pylori were
analyzed by an immunoblotting method as described
previously [22]. Briefly, cells were disrupted in 10 mM
sodium phosphate buffer (pH 7.4) containing 0.5 mM
phenylmethylsulfonyl fluoride and 0.5 mM deferox-
amine in the presence of either 0.5% Triton X-100 or
2% SDS at 4jC for 10 min and centrifuged at 15,000g
for 10 min. The supernatant fractions were used for the
assay of protein carbonyls. Carbonyl groups in cellular
proteins were reacted with 8 mM 2,4-dinitrophenyl-
hydrazine at 15jC for 60 min. Dinitrophenol (DNP)-
conjugated proteins were determined by SDS-polyacryl-
amide gel electrophoresis followed by Western blotting
analysis using specific antibodies to DNP (Intergen Co.,
Manhattanville, NY, USA) and an ECL kit (Amersham,
Buckinghamshire, England).
Analysis of oxidative modification and degradation of
DNA
DNA was purified from H. pylori cultured under the
three different conditions with the use of a DNA extrac-
tion WB kit (Wako Co.). The amount of 8-OHdG was
determined by the HPLC method as previously described
[16]. The integrity of DNA from the H. pylori specimens
was evaluated by electrophoresis on 0.7% agarose gels
containing SYBR Green I (FMC BioProducts, Rockland,
ME, USA).
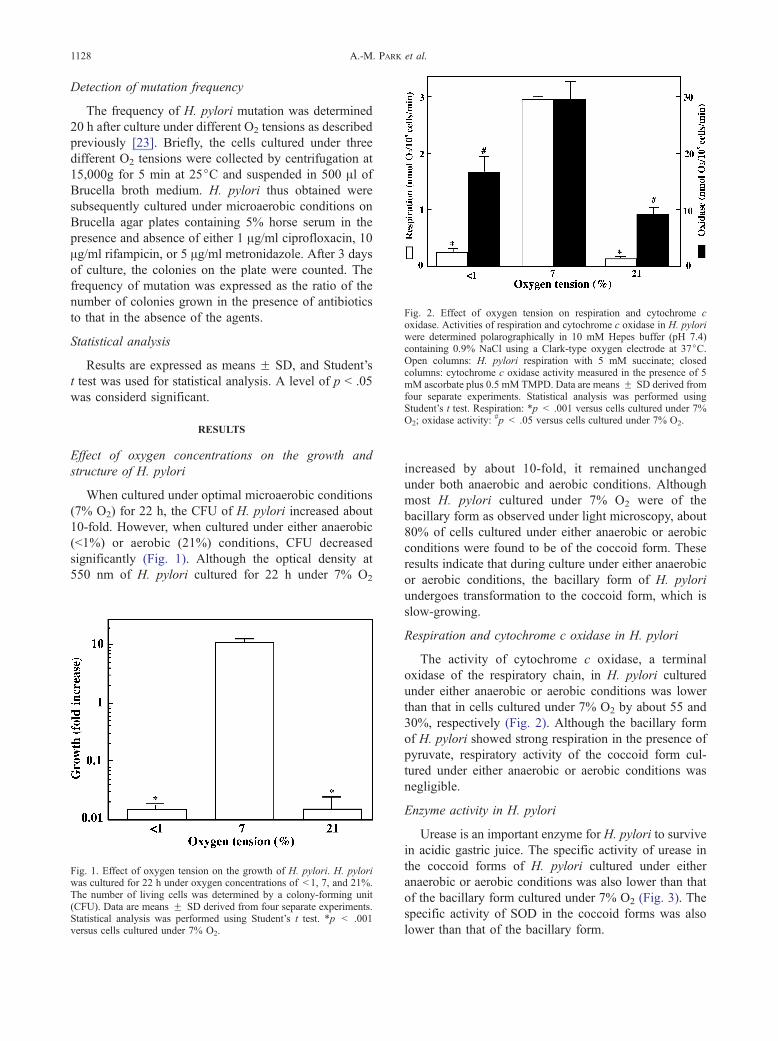
Fig. 2. Effect of oxygen tension on respiration and cytochrome coxidase. Activities of respiration and cytochrome c oxidase in H. pyloriwere determined polarographically in 10 mM Hepes buffer (pH 7.4)containing 0.9% NaCl using a Clark-type oxygen electrode at 37jC.Open columns: H. pylori respiration with 5 mM succinate; closedcolumns: cytochrome c oxidase activity measured in the presence of 5mM ascorbate plus 0.5 mM TMPD. Data are means F SD derived fromfour separate experiments. Statistical analysis was performed usingStudent’s t test. Respiration: *p < .001 versus cells cultured under 7%O2; oxidase activity:
#p < .05 versus cells cultured under 7% O2.
A.-M. PARK et al.1128
Detection of mutation frequency
The frequency of H. pylori mutation was determined
20 h after culture under different O2 tensions as described
previously [23]. Briefly, the cells cultured under three
different O2 tensions were collected by centrifugation at
15,000g for 5 min at 25jC and suspended in 500 Al ofBrucella broth medium. H. pylori thus obtained were
subsequently cultured under microaerobic conditions on
Brucella agar plates containing 5% horse serum in the
presence and absence of either 1 Ag/ml ciprofloxacin, 10
Ag/ml rifampicin, or 5 Ag/ml metronidazole. After 3 days
of culture, the colonies on the plate were counted. The
frequency of mutation was expressed as the ratio of the
number of colonies grown in the presence of antibiotics
to that in the absence of the agents.
Statistical analysis
Results are expressed as means F SD, and Student’s
t test was used for statistical analysis. A level of p < .05
was considerd significant.
RESULTS
Effect of oxygen concentrations on the growth and
structure of H. pylori
When cultured under optimal microaerobic conditions
(7% O2) for 22 h, the CFU of H. pylori increased about
10-fold. However, when cultured under either anaerobic
(<1%) or aerobic (21%) conditions, CFU decreased
significantly (Fig. 1). Although the optical density at
550 nm of H. pylori cultured for 22 h under 7% O2
Fig. 1. Effect of oxygen tension on the growth of H. pylori. H. pyloriwas cultured for 22 h under oxygen concentrations of < 1, 7, and 21%.The number of living cells was determined by a colony-forming unit(CFU). Data are means F SD derived from four separate experiments.Statistical analysis was performed using Student’s t test. *p < .001versus cells cultured under 7% O2.
increased by about 10-fold, it remained unchanged
under both anaerobic and aerobic conditions. Although
most H. pylori cultured under 7% O2 were of the
bacillary form as observed under light microscopy, about
80% of cells cultured under either anaerobic or aerobic
conditions were found to be of the coccoid form. These
results indicate that during culture under either anaerobic
or aerobic conditions, the bacillary form of H. pylori
undergoes transformation to the coccoid form, which is
slow-growing.
Respiration and cytochrome c oxidase in H. pylori
The activity of cytochrome c oxidase, a terminal
oxidase of the respiratory chain, in H. pylori cultured
under either anaerobic or aerobic conditions was lower
than that in cells cultured under 7% O2 by about 55 and
30%, respectively (Fig. 2). Although the bacillary form
of H. pylori showed strong respiration in the presence of
pyruvate, respiratory activity of the coccoid form cul-
tured under either anaerobic or aerobic conditions was
negligible.
Enzyme activity in H. pylori
Urease is an important enzyme for H. pylori to survive
in acidic gastric juice. The specific activity of urease in
the coccoid forms of H. pylori cultured under either
anaerobic or aerobic conditions was also lower than that
of the bacillary form cultured under 7% O2 (Fig. 3). The
specific activity of SOD in the coccoid forms was also
lower than that of the bacillary form.
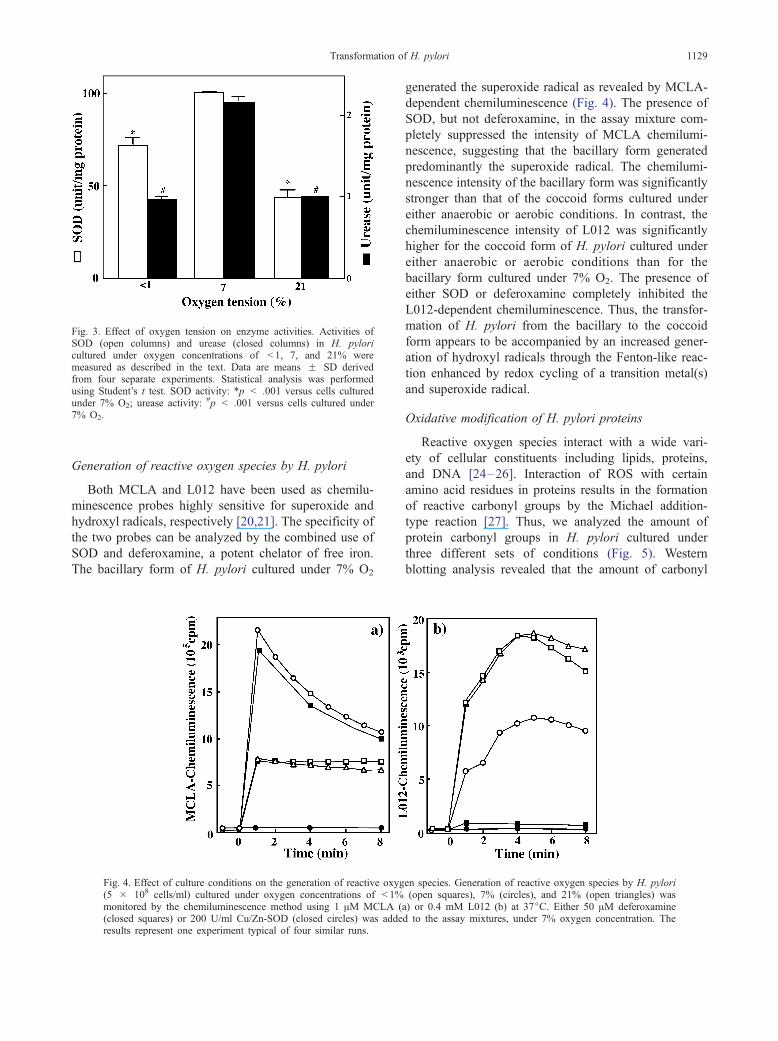
Fig. 3. Effect of oxygen tension on enzyme activities. Activities ofSOD (open columns) and urease (closed columns) in H. pyloricultured under oxygen concentrations of < 1, 7, and 21% weremeasured as described in the text. Data are means F SD derivedfrom four separate experiments. Statistical analysis was performedusing Student’s t test. SOD activity: *p < .001 versus cells culturedunder 7% O2; urease activity: #p < .001 versus cells cultured under7% O2.
Transformation of H. pylori 1129
Generation of reactive oxygen species by H. pylori
Both MCLA and L012 have been used as chemilu-
minescence probes highly sensitive for superoxide and
hydroxyl radicals, respectively [20,21]. The specificity of
the two probes can be analyzed by the combined use of
SOD and deferoxamine, a potent chelator of free iron.
The bacillary form of H. pylori cultured under 7% O2
Fig. 4. Effect of culture conditions on the generation of reactive oxyg(5 � 108 cells/ml) cultured under oxygen concentrations of < 1%monitored by the chemiluminescence method using 1 AM MCLA ((closed squares) or 200 U/ml Cu/Zn-SOD (closed circles) was adderesults represent one experiment typical of four similar runs.
generated the superoxide radical as revealed by MCLA-
dependent chemiluminescence (Fig. 4). The presence of
SOD, but not deferoxamine, in the assay mixture com-
pletely suppressed the intensity of MCLA chemilumi-
nescence, suggesting that the bacillary form generated
predominantly the superoxide radical. The chemilumi-
nescence intensity of the bacillary form was significantly
stronger than that of the coccoid forms cultured under
either anaerobic or aerobic conditions. In contrast, the
chemiluminescence intensity of L012 was significantly
higher for the coccoid form of H. pylori cultured under
either anaerobic or aerobic conditions than for the
bacillary form cultured under 7% O2. The presence of
either SOD or deferoxamine completely inhibited the
L012-dependent chemiluminescence. Thus, the transfor-
mation of H. pylori from the bacillary to the coccoid
form appears to be accompanied by an increased gener-
ation of hydroxyl radicals through the Fenton-like reac-
tion enhanced by redox cycling of a transition metal(s)
and superoxide radical.
Oxidative modification of H. pylori proteins
Reactive oxygen species interact with a wide vari-
ety of cellular constituents including lipids, proteins,
and DNA [24–26]. Interaction of ROS with certain
amino acid residues in proteins results in the formation
of reactive carbonyl groups by the Michael addition-
type reaction [27]. Thus, we analyzed the amount of
protein carbonyl groups in H. pylori cultured under
three different sets of conditions (Fig. 5). Western
blotting analysis revealed that the amount of carbonyl
en species. Generation of reactive oxygen species by H. pylori(open squares), 7% (circles), and 21% (open triangles) was
a) or 0.4 mM L012 (b) at 37jC. Either 50 AM deferoxamined to the assay mixtures, under 7% oxygen concentration. The
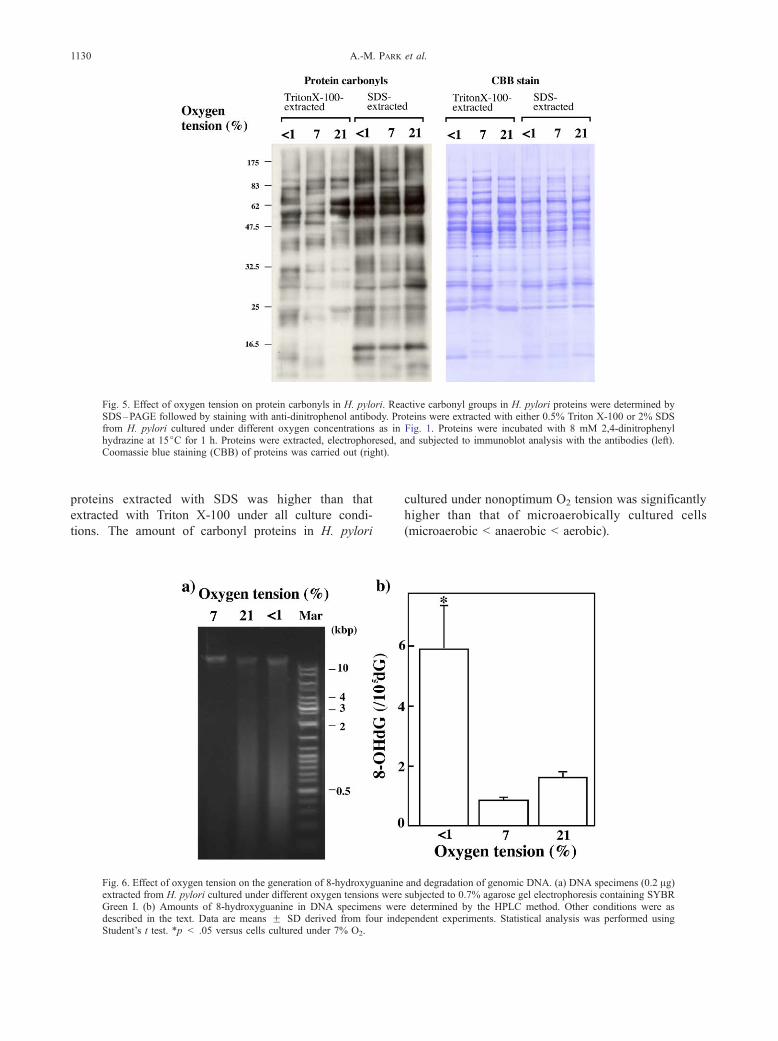
Fig. 5. Effect of oxygen tension on protein carbonyls in H. pylori. Reactive carbonyl groups in H. pylori proteins were determined bySDS–PAGE followed by staining with anti-dinitrophenol antibody. Proteins were extracted with either 0.5% Triton X-100 or 2% SDSfrom H. pylori cultured under different oxygen concentrations as in Fig. 1. Proteins were incubated with 8 mM 2,4-dinitrophenylhydrazine at 15jC for 1 h. Proteins were extracted, electrophoresed, and subjected to immunoblot analysis with the antibodies (left).Coomassie blue staining (CBB) of proteins was carried out (right).
A.-M. PARK et al.1130
proteins extracted with SDS was higher than that
extracted with Triton X-100 under all culture condi-
tions. The amount of carbonyl proteins in H. pylori
Fig. 6. Effect of oxygen tension on the generation of 8-hydroxyguanineextracted from H. pylori cultured under different oxygen tensions wereGreen I. (b) Amounts of 8-hydroxyguanine in DNA specimens werdescribed in the text. Data are means F SD derived from four indeStudent’s t test. *p < .05 versus cells cultured under 7% O2.
cultured under nonoptimum O2 tension was significantly
higher than that of microaerobically cultured cells
(microaerobic < anaerobic < aerobic).
and degradation of genomic DNA. (a) DNA specimens (0.2 Ag)subjected to 0.7% agarose gel electrophoresis containing SYBRe determined by the HPLC method. Other conditions were aspendent experiments. Statistical analysis was performed using
Transformation of H. pylori 1131
Changes in DNA after transformation of cells
We analyzed the amounts of 8-OHdG in DNA of H.
pylori cultured under three different O2 tensions (Fig.
6). The DNA sample from the coccoid form was
significantly larger than that from the bacillary form.
Interestingly, the amount of 8-OHdG in the coccoid
form cultured under anaerobic conditions was signifi-
cantly larger than that in the coccoid form cultured
under aerobic conditions. Agarose gel electrophoresis
revealed that, although most DNA from the bacillary
form cultured under 7% O2 was intact, significant
fractions of DNA from the coccoid forms cultured
under either anaerobic or aerobic conditions had under-
gone degradation.
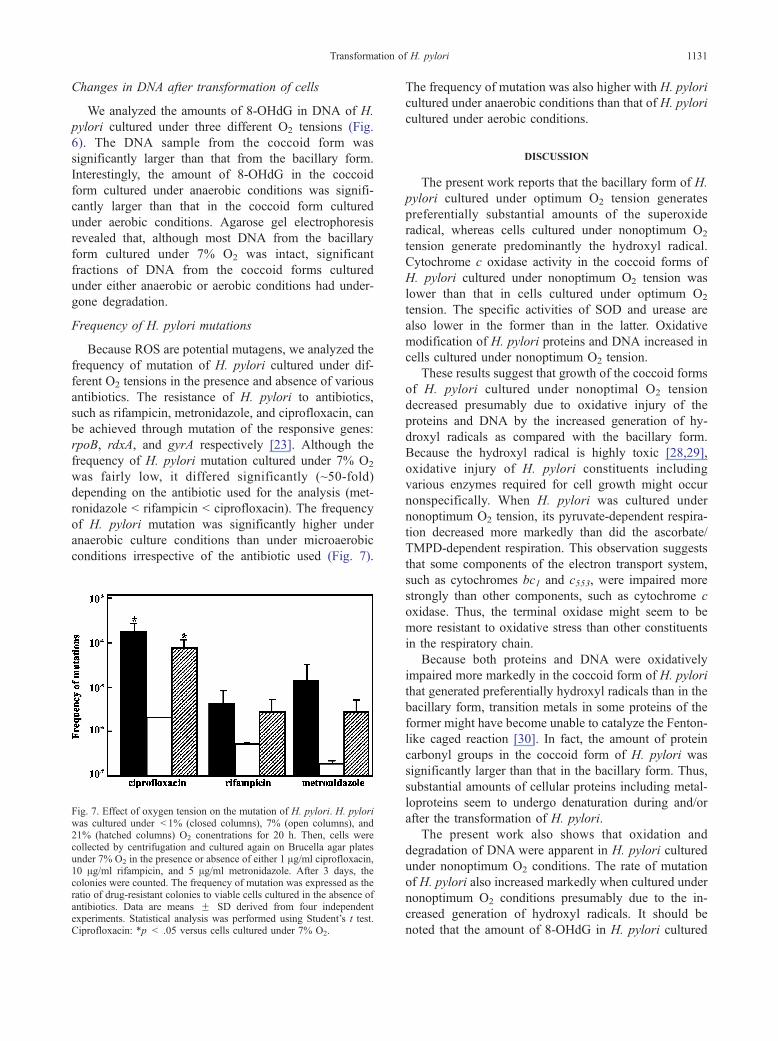
Frequency of H. pylori mutations
Because ROS are potential mutagens, we analyzed the
frequency of mutation of H. pylori cultured under dif-
ferent O2 tensions in the presence and absence of various
antibiotics. The resistance of H. pylori to antibiotics,
such as rifampicin, metronidazole, and ciprofloxacin, can
be achieved through mutation of the responsive genes:
rpoB, rdxA, and gyrA respectively [23]. Although the
frequency of H. pylori mutation cultured under 7% O2
was fairly low, it differed significantly (~50-fold)
depending on the antibiotic used for the analysis (met-
ronidazole < rifampicin < ciprofloxacin). The frequency
of H. pylori mutation was significantly higher under
anaerobic culture conditions than under microaerobic
conditions irrespective of the antibiotic used (Fig. 7).
Fig. 7. Effect of oxygen tension on the mutation of H. pylori. H. pyloriwas cultured under < 1% (closed columns), 7% (open columns), and21% (hatched columns) O2 conentrations for 20 h. Then, cells werecollected by centrifugation and cultured again on Brucella agar platesunder 7% O2 in the presence or absence of either 1 Ag/ml ciprofloxacin,10 Ag/ml rifampicin, and 5 Ag/ml metronidazole. After 3 days, thecolonies were counted. The frequency of mutation was expressed as theratio of drug-resistant colonies to viable cells cultured in the absence ofantibiotics. Data are means F SD derived from four independentexperiments. Statistical analysis was performed using Student’s t test.Ciprofloxacin: *p < .05 versus cells cultured under 7% O2.
The frequency of mutation was also higher with H. pylori
cultured under anaerobic conditions than that of H. pylori
cultured under aerobic conditions.
DISCUSSION
The present work reports that the bacillary form of H.
pylori cultured under optimum O2 tension generates
preferentially substantial amounts of the superoxide
radical, whereas cells cultured under nonoptimum O2
tension generate predominantly the hydroxyl radical.
Cytochrome c oxidase activity in the coccoid forms of
H. pylori cultured under nonoptimum O2 tension was
lower than that in cells cultured under optimum O2
tension. The specific activities of SOD and urease are
also lower in the former than in the latter. Oxidative
modification of H. pylori proteins and DNA increased in
cells cultured under nonoptimum O2 tension.
These results suggest that growth of the coccoid forms
of H. pylori cultured under nonoptimal O2 tension
decreased presumably due to oxidative injury of the
proteins and DNA by the increased generation of hy-
droxyl radicals as compared with the bacillary form.
Because the hydroxyl radical is highly toxic [28,29],
oxidative injury of H. pylori constituents including
various enzymes required for cell growth might occur
nonspecifically. When H. pylori was cultured under
nonoptimum O2 tension, its pyruvate-dependent respira-
tion decreased more markedly than did the ascorbate/
TMPD-dependent respiration. This observation suggests
that some components of the electron transport system,
such as cytochromes bc1 and c553, were impaired more
strongly than other components, such as cytochrome c
oxidase. Thus, the terminal oxidase might seem to be
more resistant to oxidative stress than other constituents
in the respiratory chain.
Because both proteins and DNA were oxidatively
impaired more markedly in the coccoid form of H. pylori
that generated preferentially hydroxyl radicals than in the
bacillary form, transition metals in some proteins of the
former might have become unable to catalyze the Fenton-
like caged reaction [30]. In fact, the amount of protein
carbonyl groups in the coccoid form of H. pylori was
significantly larger than that in the bacillary form. Thus,
substantial amounts of cellular proteins including metal-
loproteins seem to undergo denaturation during and/or
after the transformation of H. pylori.
The present work also shows that oxidation and
degradation of DNA were apparent in H. pylori cultured
under nonoptimum O2 conditions. The rate of mutation
of H. pylori also increased markedly when cultured under
nonoptimum O2 conditions presumably due to the in-
creased generation of hydroxyl radicals. It should be
noted that the amount of 8-OHdG in H. pylori cultured

A.-M. PARK et al.1132
under anaerobic conditions was higher than that of H.
pylori cultured under 21 and 7% O2 by about 3.7- and 9-
fold, respectively. Furthermore, the mutation rate was
also higher in H. pylori cultured under anaerobic con-
ditions than in H. pylori cultured under aerobic condi-
tions by about 1.5- to 5-fold. It is known that DNA-
repairing enzymes are often induced in various cells
exposed to oxidative stress. In fact, Escherichia coli
shows adaptive response to hydrogen peroxide [31] and
superoxide [32] by upregulating OxyR, SoxR, and SoxS
[33–36]. Because H. pylori lacks these stress-induced
genes [37,38], degradation of oxidized DNA would have
been enhanced more markedly under nonoptimum O2
tension. Furthermore, ATP generation would not occur
under anaerobic conditions so that DNA-repairing
enzymes functioned minimally. Although the amounts
of hydroxyl radicals generated were similarly high in H.
pylori cultured under anaerobic and aerobic conditions,
gene mutation caused by DNA oxidation occurred more
markedly under anaerobic conditions than under aerobic
conditions. H. pylori is known to have an unusually large
genetic diversity; hence, the genomic DNAs of cells
obtained from one patient differ significantly from those
of another [39,40]. The types of mutation found in H.
pylori are point mutation, recombination, insertion, sub-
stitution, and deletion. It is well documented that ROS
accelerate the mutation of DNA [41,42]. Hence, the
property of H. pylori to generate ROS seems to be an
important factor that enhances the genetic diversity of
this bacterium.
Because H. pylori upregulates the activities of CO2-
fixation enzymes, particularly under aerobic conditions,
it grows preferentially in the presence of 5–10% CO2
[43]. However, the transformation of H. pylori from
bacillary to coccoid form occurs preferentially in the
absence of CO2. Thus, we cultured H. pylori without
exogenous use of CO2. Preliminary experiments in this
laboratory showed that H. pylori cultured under aerobic
or microaerobic conditions with 5% CO2 generated
predominantly superoxide radicals (data not shown).
Thus, the presence of CO2 seems to affect the ability
of H. pylori to generate either superoxide or hydroxyl
radicals irrespective of the morphological transformation.
Although the coccoid form of H. pylori does not
grow under culture conditions, this form has been
postulated to retain virulence and induce gastric inflam-
mation in BALB/cA mice [44]. In fact, the coccoid
form of H. pylori is frequently found in human stomach
and extragastric environments [4–6,45,46]. Chan et al.
showed that 94 and 50% of H. pylori-positive patients
with gastric adenocarcinoma and peptic ulcer have the
coccoid form of cells, respectively [45]. Thus, the ROS
generated by the bacillary and/or coccoid forms of H.
pylori may have cytotoxic effects not only on H. pylori
itself but also on gastric mucosal cells in its hosts. The
possible involvement of ROS generated by the two forms
of H. pylori in the mechanism of gastric carcinogenesis
should be studied further.
Acknowledgments—This work was supported by Special CoordinationFunds for Promoting Science and Technology from the Ministry ofEducation, Culture, Sports, Science and Technology, the JapaneseGovernment (11877031 to M.I.).
REFERENCES
[1] Parsonnet, J.; Hansen, S.; Rodriguez, L.; Gelb, A. B.; Warnke,R. A.; Jellum, E.; Orentrich, N.; Vogelman, J. H.; Friedman, G.D. Helicobacter pylori infection and gastric lymphoma.N. Engl. J.Med. 330:1267–1271; 1994.
[2] Hahm, K. B.; Lee, K. J.; Kim, J. H.; Cho, S. W.; Chung, M. H.Helicobacter pylori infection, oxidative DNA damage, gastriccarcinogensis, and reversibility by rebamipide. Dig. Dis. Sci. 43(Suppl.):72S–77S; 1998.
[3] Kabir, S. Detection ofHelicobacter pylori in faeces by culture, PCRand enzyme immunoassay, J. Med. Microbiol., 50:1021–1029;2001.
[4] Klein, P. D.; Graham, D. Y.; Gaillour, A.; Opekun, A. R.; Smith,E. O. Water source as a risk factor for Helicobacter pylori infec-tion in Peruvian children. Lancet 337:1503–1506; 1991.
[5] Mendall, M. A.; Northfield, T. C. Transmission of Helicobacterpylori infection. Gut 37:1–3; 1995.
[6] Sarker, S. A.; Mahalanabis, D.; Hildebrand, P.; Rahaman, M. M.;Bardhan, P. K.; Fuchs, G.; Beglinger, C.; Gyr, K. Helicobacterpylori: prevalence, transmission, and serum pepsinogen II concen-trations in children of a poor periurban community in Bangladesh.Clin. Infect. Dis. 25:990–995; 1997.
[7] Inoue, M.; Sato, E. F.; Park, A. M.; Nishikawa, M.; Kasahara, E.;Miyoshi, M.; Ochi, A.; Utsumi, K. Cross-talk between NO andoxyradicals, a supersystem that regulates energy metabolism andsurvival of animals. Free Radic. Res. 33:757–770; 2000.
[8] Schwille, P. O.; Schellerer, W.; Steiner, H.; Reitzenstein, M. Ratgastric mucosal oxygen tension, ulcer index, plasma gastrin andglucagon following restraint stress. Influence of vagotomy,splanchnicotomy and exogenous secretin. Res. Exp. Med. (Berlin)167:149–158; 1976.
[9] Sorberg, M.; Nilsson, M.; Hanberger, H.; Nilsson, L. E. Morpho-logic conversion of Helicobacter pylori from bacillary to coccoidform. Eur. J. Clin. Microbiol. Infect. Dis. 15:216–219; 1996.
[10] Gribbon, L. T.; Barer, M. R. Oxidative metabolism in noncultu-rable Helicobacter pylori and Vibrio vulnificus cells studied bysubstrate-enhanced tetrazolium reduction and digital image pro-cessing. Appl. Environ. Microbiol. 61:3379–3384; 1995.
[11] Cellini, L.; Robuffo, I.; Di Campli, E.; Di Bartolomeo, S.; Tarabo-relli, T.; Dainelli, B. Recovery of Helicobacter pylori ATCC43504from a viable but not culturable state: regrowth or resuscitation?APMIS 106:571–579; 1998.
[12] Kusters, J. G.; Gerrits, M. M.; Van Strijp, J. A.; Vandenbroucke-Grauls, C. M., Coccoid forms of Helicobacter pylori are the mor-phologicmanifestationof cell death. Infect. Immun.65:3672–3679;1997.
[13] Bode, G.; Mauch, F.; Malfertheiner, P. The coccoid forms ofHelicobacter pylori: criteria for their viability. Epidemiol. Infect.111:483–490; 1993.
[14] Segal, E. D.; Falkow, S.; Tompkins, L. S. Helicobacter pyloriattachment to gastric cells induces cytoskeletal rearrangementsand tyrosine phosphorylation of host cell proteins. Proc. Natl.Acad. Sci. USA 93:1259–1264; 1996.
[15] Wang, X.; Sturegard, E.; Rupar, R.; Nilsson, H. O.; Aleljung,P. A.; Carlen, B.; Willen, R.; Wadstrom, T. Infection of BALB/c A mice by spiral and coccoid forms of Helicobacter pylori.J. Med. Microbiol. 46:657–663; 1997.
[16] Nakamura, A.; Park, A. M.; Nagata, K.; Sato, E. F.; Kashiba, M.;

Transformation of H. pylori 1133
Ogata, T.; Inoue, M. Oxidative cellular damage associated withtransformation of Helicobacter pylori from a bacillary to a coc-coid form. Free Radic. Biol. Med. 28:1611–1618; 2000.
[17] Park, A. M.; Nagata, K.; Sato, E. F.; Tamura, T.; Shimono, K.;Inoue, M. Mechanism of strong resistance of Helicobacter pylorirespiration to nitric oxide. Arch. Biochem. Biophys. 411:129–135;2003.
[18] Ferraro, R. L.; Hazell, S. L.; Lee, A. The urease enzymes ofCampylobacter pylori and a related bacterium. J. Med. Microbiol.27:33–40; 1988.
[19] Crapo, J. D.; McCord, J. M.; Fridovich, I. Preparation and assayof superoxide dismutases. Methods Enzymol. 53:382–393; 1978.
[20] Sugioka, K.; Nakano, M.; Kurashige, S.; Akuzawa, Y.; Goto, T. Achemiluminescent probe with a Cypridina luciferin analog, 2-methyl-6-phenyl-3,7-dihydroimidazo[1,2-a]pyrazin-3-one, speci-fic and sensitive for O2
� production in phagocytizing macro-phages. FEBS Lett. 197:27–30; 1986.
[21] Imada, I.; Sato, F. E.; Miyamoto, M.; Ichimori, Y.; Minamiyama,Y.; Konaka, T.; Inoue, M. Analysis of reactive oxygen speciesgenerated by neutrophils using a chemiluminescence probe L-012. Anal. Biochem. 271:53–58; 1999.
[22] Levine, R. L.; Williams, J. A.; Stadtman, E. R.; Shacter, E. Car-bonyl assays for determination of oxidatively modified proteins.Methods Enzymol. 233:346–357; 1994.
[23] Wang, G.; Wilson, T. J.; Jiang, Q.; Taylor, D. E. Spontaneousmutations that confer antibiotic resistance in Helicobacter pylori.Antimicrob. Agents Chemother. 45:727–733; 2001.
[24] Choi, J. H.; Yu, B. P. Brain synaptosomal aging: free radicalsand membrane fluidity, Free Radic. Biol. Med., 18:133–139;1995.
[25] Berlett, B. S.; Stadtman, E. R. Protein oxidation in aging, disease,and oxidative stress. J. Biol. Chem. 272:20313–20316; 1997.
[26] Beckman, K. B.; Ames, B. N. Oxidative decay of DNA. J. Biol.Chem. 272:19633–19666; 1997.
[27] Kato, Y.; Uchida, K.; Kawakishi, S. Oxidative fragmentation ofcollagen and prolyl peptide by Cu(II)/H2O2: conversion of prolineresidue to 2-pyrrolidone. J. Biol. Chem. 267:23646–23651; 1992.
[28] Davies, J. A. Protein damege and degradation by oxygen radicals.I. general aspects. J. Biol. Chem. 262:9895–9901; 1987.
[29] Davies, J. A. Protein damage and degradation by oxygen radicals.II. Modification of amino acid. J. Biol. Chem. 262:9902–9907;1987.
[30] Yim, M. B.; Berlett, B. S.; Chock, P. B.; Stadtman, E. R. Man-ganese(II)-bicarbonate-mediated catalytic activity for hydrogenperoxide dismutation and amino acid oxidation: detection of freeradical intermediates. Proc. Natl. Acad. Sci. USA 87:394–398;1990.
[31] Demple, B.; Halbrook, J. Inducible repair of oxidative DNAdamage in Escherichia coli. Nature 304:466–468; 1983.
[32] Farr, S. B.; Natvig, D. O.; Kogoma, T. Toxicity and mutagenicityof plumbagin and the induction of a possible new DNA repairpathway in Escherichia coli. J. Bacteriol. 164:1309–1316; 1985.
[33] Christman, M. F.; Morgan, R. W.; Jacobson, F. S.; Ames, B. N.Positive control of a regulon for defenses against oxidative stressand some heat-shock proteins in Salmonella typhimurium. Cell41:753–762; 1985.
[34] Greenberg, J. T.; Monach, P.; Chou, J. H.; Josephy, P. D.; Demple,B. Positive control of a global antioxidant defense regulon acti-vated by superoxide-generating agents in Escherichia coli. Proc.Natl. Acad. Sci. USA 87:6181–6185; 1990.
[35] Tsaneva, I. R.; Weiss, B. soxR, a locus governing a superoxideresponse regulon in Escherichia coli K-12. J. Bacteriol.172:4197–4205; 1990.
[36] Wu, J.; Weiss, B. Two divergently transcribed genes, soxR andsoxS, control a superoxide response regulon of Escherichia coli.J. Bacteriol. 173:2864–2871; 1991.
[37] Tomb, J. F.; White, O.; Kerlavage, A. R.; Clayton, R. A.; Sutton,G. G.; Fleischmann, R. D.; Ketchum, K. A.; Klenk, H. P.; Gill, S.;Dougherty, B. A.; Nelson, K.; Quackenbush, J.; Zhou, L.; Kirk-ness, E. F.; Peterson, S.; Loftus, B.; Richardson, D.; Dodson, R.;Khalak, H. G.; Glodek, A.; McKenney, K.; Fitzegerald, L. M.;Lee, N.; Adams, M. D.; Venter, J. C. The complete genome se-quence of the gastric pathogen Helicobacter pylori. Nature388:539–547; 1997.
[38] Alm, R. A.; Ling, L. S.; Moir, D. T.; King, B. L.; Brown, E. D.;Doig, P. C.; Smith, D. R.; Noonan, B.; Guild, B. C.; deJonge,B. L.; Carmel, G.; Tummino, P. J.; Caruso, A.; Uria-Nickelsen,M.; Mills, D. M.; Ives, C.; Gibson, R.; Merberg, D.; Mills, S.D.; Jiang, Q.; Taylor, D. E.; Vovis, G. F.; Trust, T. J. Genomic-sequence comparison of two unrelated isolates of the humangastric pathogen Helicobacter pylori. Nature 397:176–180;1999.
[39] Marshall, D. G.; Dundon, W. G.; Beesley, S. M.; Smyth, C. J.Helicobacter pylori: -a conundrum of genetic diversity. Microbi-ology 144:2925–2939; 1998.
[40] Blaser, M. J.; Berg, D. E. Helicobacter pylori genetic diversityand risk of human disease. J. Clin. Invest. 107:767–773; 2001.
[41] Oya, Y.; Yamamoto, K.; Tonomura, A. The biological activity ofhydrogen peroxide: I. Induction of chromosome-type aberrationssusceptible to inhibition by scavengers of hydroxyl radicals inhuman embryonic fibroblasts. Mutat. Res. 172:245–253; 1986.
[42] Farr, S. B.; Touati, D.; Kogoma, T. Effects of oxygen stress onmembrane functions in Escherichia coli: role of HPI catalase. J.Bacteriol. 170:1837–1842; 1988.
[43] Donelli, G.; Matarrese, P.; Fiorentini, C.; Dainelli, B.; Taraborelli,T.; Di Campli, E.; Di Bartolomeo, S.; Cellini, L. The effect ofoxygen on the growth and cell morphology of Helicobacter py-lori. FEMS Microbiol. Lett. 168:9–15; 1998.
[44] Wang, X.; Sturegard, E.; Rupar, R.; Nilsson, H. O.; Aleljung, P.A.; Carlen, B.; Willen, R.; Wadstrom, T. Infection of BALB/c Amice by spiral and coccoid forms of Helicobacter pylori. J. Med.Microbiol. 46:657–663; 1997.
[45] Chan, W. Y.; Hui, P. K.; Leung, K. M.; Chow, J.; Kwok, F.; Ng, C.S. Coccoid forms of Helicobacter pylori in the human stomach.Am. J. Clin. Pathol. 102:503–507; 1994.
[46] Ogata, M.; Araki, K.; Ogata, T. An electron microscopic study ofHelicobacter pylori in the surface mucous gel layer. Histol. His-topathol. 13:347–358; 1998.
ABBREVIATIONS
DNP—dinitrophenol
LO12—8-amino-5-chloro-7-phenyl-pyridopyridazine
MCLA—2-methyl-6-[f-methoxyphenyl]-3,7-dihydroi-
midazol[1,2-a]pyrazin-3-one
ROS—reactive oxygen species
SOD—superoxide dismutase
TMPD—N,N,NV,NV-tetramethyl-p-phenylenediamine





























