Embed Size (px)
Citation preview

Helicobacter pylori single-stranded DNA bindingprotein – functional characterization and modulationof H. pylori DnaB helicase activityAtul Sharma1,*, Ram G. Nitharwal1,*, Bhupender Singh2, Ashraf Dar1, Santanu Dasgupta2
and Suman K. Dhar1
1 Special Centre for Molecular Medicine, Jawaharlal Nehru University, New Delhi, India
2 Department of Cell and Molecular Biology, Uppsala University, Biomedical Center, Sweden
Helicobacter pylori causes gastric ulcer and gastric
adenocarcinoma related diseases in humans [1,2].
Although there are effective therapies against these
bacteria, an increasing incidence of antibiotic
resistance and recurrent infection following treatment
complicates the situation [3,4]. Considerable research
has been conducted on the clinical aspects of H. pylori
infection but the fundamental aspects of cell cycle and
DNA replication are poorly understood.
H. pylori can transform from the active helical bacil-
lary form into the dormant coccoid form, which is the
manifestation of a bacterial response towards anti-
biotics, stress, aging and unfavorable conditions [5–8].
Almost nothing is known regarding the molecular
mechanisms involved in vegetative to coccoid transi-
tion and the biology of the dormant coccoid form.
DNA replication requires the timely interplay of
various proteins that co-ordinate initiation, elongation
and termination. Although H. pylori fall into the king-
dom of Gram negative bacteria, the sequence analysis
of Helicobacter genome reveals interesting features that
include the location of the dnaA gene, approximately
600 kb away from the dnaN-gyrB gene cluster and the
absence of important genes such as recF and the
Keywords
DNA replication; helicase; Helicobacter
pylori; replication foci; single-stranded
DNA binding protein
Correspondence
S. K. Dhar, Special Centre for Molecular
Medicine, JNU, New Delhi 110067, India
Fax: +91 11 26741781
Tel: +91 11 26742572
E-mail: [email protected]
*These authors contributed equally to this
work
(Received 7 October 2008, revised 10
November 2008, accepted 13 November
2008)
doi:10.1111/j.1742-4658.2008.06799.x
Helicobacter pylori, an important bacterial pathogen, causes gastric ulcer
and gastric adenocarcinoma in humans. The fundamentals of basic biology
such as DNA replication are poorly understood in this pathogen. In the
present study, we report the cloning and functional characterization of the
single-stranded DNA (ssDNA) binding protein from H. pylori. The N-ter-
minal DNA binding domain shows significant homology with E. coli
single-stranded DNA binding protein (SSB), whereas the C-terminal
domain shows less homology. The overall DNA-binding activity and tetra-
merization properties, however, remain unaffected. In in vitro experiments
with purified proteins, H. pylori (Hp) SSB bound specifically to ssDNA
and modulated the enzymatic ATPase and helicase activity of HpDnaB
helicase. HpSSB and HpDnaB proteins were co-localized in sharp, distinct
foci in exponentially growing H. pylori cells, whereas both were spread
over large areas in its dormant coccoid form, suggesting the absence of
active replication forks in the latter. These results confirm the multiple
roles of SSB during DNA replication and provide evidence for altered
replicative metabolism in the spiral and coccoid forms that may be central
to the bacterial physiology and pathogenesis.
Abbreviations
dsDNA, double-stranded DNA; Ec, E. coli; FITC, fluoroscein isothicyanate; GST, glutathione S-transferase; Hp, Helicobacter pylori; IPTG,
isopropyl thio-b-D-galactoside; Pi, inorganic phosphate; SSB, single-stranded DNA binding protein; ssDNA, single-stranded DNA.
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 519

helicase loader dnaC [9,10]. We have shown recently
that the H. pylori (Hp) DnaB helicase can bypass
E. coli (Ec) DnaC function in vivo that may explain
the absence of the dnaC gene in H. pylori [11,12]. The
C-terminal region of HpDnaB is unique, with a 34
amino acid residue insertion region that is essential for
its function [13]. Recently, a protein HobA, the struc-
tural homolog of E. coli protein DiaA has been shown
to interact with the initiator protein DnaA and this
interaction is essential for DNA replication in Helicob-
acter [14,15].
One protein that is central to the DNA replication,
repair and recombination is single-stranded DNA
binding protein (SSB) [16,17]. The N-terminal domain
of SSB is highly conserved and forms an oligonucleo-
tide binding fold, and this region is also responsible
for oligomerization, typically homotetramerization in
eubacteria. The C-terminal region is less conserved and
is responsible for protein–protein interaction [18,19].
The proteins that may interact with SSB include DNA
polymerase, RNA polymerase and DNA helicases [20–
22]. Although, no direct interaction has been shown
between SSB and DnaB replicative helicase, the physi-
cal interaction between SSB and PriA helicase, the
major DNA replication restart protein, has been dem-
onstrated recently [23]. The extreme C-terminal ten
residues are essential for the interaction of EcSSB with
PriA helicase and the deletion of these residues affects
the stimulation of helicase activity of PriA mediated
through SSB [23]. Interestingly, deletion of 10 amino
acid residues from the extreme C-terminus affects
in vivo function of EcSSB [24]. Taken together, these
results suggest that the extreme C-terminal residues of
SSB are important for protein–protein interaction.
To understand the basic DNA replication machinery
of H. pylori in detail, we have cloned, over-expressed
and characterized the functional properties of HpSSB
both in vitro and in vivo. We found that HpSSB is a
true homolog of SSB in vivo because it can comple-
ment the Ecssb mutant strain and is localized in the
replisome assembly of E. coli. Furthermore, we show
that HpSSB can modulate the enzymatic activities of
HpDnaB significantly. Finally, we report that both
HpSSB and HpDnaB are co-localized in distinct foci
in replicating H. pylori but not in the dormant coccoid
form, indicating an important difference between the
two forms regarding bacterial physiology and growth.
These results further enhance our knowledge on SSB
proteins from a slow growing pathogenic bacteria and
offer great potential to study DNA–protein and pro-
tein–protein interaction that is central to the DNA
replication machinery in prokaryotes. To the best of
our knowledge, this is the first probe into the coccoid
stage demonstrating its distinction from the vegetative,
spiral stage in DNA replication activity.
Results and Discussion
Cloning, expression, purification and biochemical
activity of HpSSB
The coding region of the ORF, HP1245 (annotated as
the putative HpSSB homlog) was amplified using spe-
cific primers (as shown in the Experimental proce-
dures) and genomic DNA from H. pylori strain 26695.
The amplified PCR product was subsequently cloned
in the expression vector pET28a and was sequenced
completely. The deduced amino acid sequence was
aligned with E. coli and Bacillus subtilis SSB sequences
using the multiple sequence alignment program
clustalw (Fig. 1A). Overall, HpSSB shows 30% iden-
tity and 45% homology with EcSSB. The analysis
shows more homology at the N-terminal DNA-binding
domain (� 67%) compared to the C-terminal domain
(� 34%), which is assumed to be the region responsi-
ble for protein–protein interaction. Sequence compari-
son reveals many interesting features that include the
absence of some important residues in HpSSB com-
pared to that of EcSSB. The tryptophan residues
(Trp40 and Trp54 in EcSSB) have been replaced by
phenyl alanine residues [25]. In vitro, mutations in
these residues in EcSSB show a moderate effect on
DNA binding. His55, a residue important for oligo-
merization, is replaced by Ile in HpSSB. However,
mutation of His55 to Ile does not affect in vitro
oligomerization of EcSSB [26].
To purify recombinant HpSSB for biochemical char-
acterization, the E. coli BL21 codon plus strain was
transformed with pET28a-HpSSB construct, as
described in the Experimental procedures, and the
His-tagged fusion protein was purified using Ni-NTA
agarose beads (Fig. 1B). The purified His6-HpSSB
protein shows an apparent molecular mass of approxi-
mately 25 kDa, which is very close to the deduced
molecular mass of untagged HpSSB (� 20 kDa).
Polyclonal antibodies were raised in mice using the
purified His-HpSSB as antigens. These antibodies
effectively recognized the purified HpSSB antigen
(Fig. 1C). To prove that HpSSB is truly expressed in
H. pylori, a western blot experiment was performed
using the same antibodies against H. pylori bacterial
lysate. A single band was detected in the lane contain-
ing bacterial lysate confirming the expression of
HpSSB in H. pylori (Fig. 1C). There is a difference in
the migration of the recombinant protein and the
endogenous protein because the recombinant protein is
H. pylori SSB protein A. Sharma et al.
520 FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS
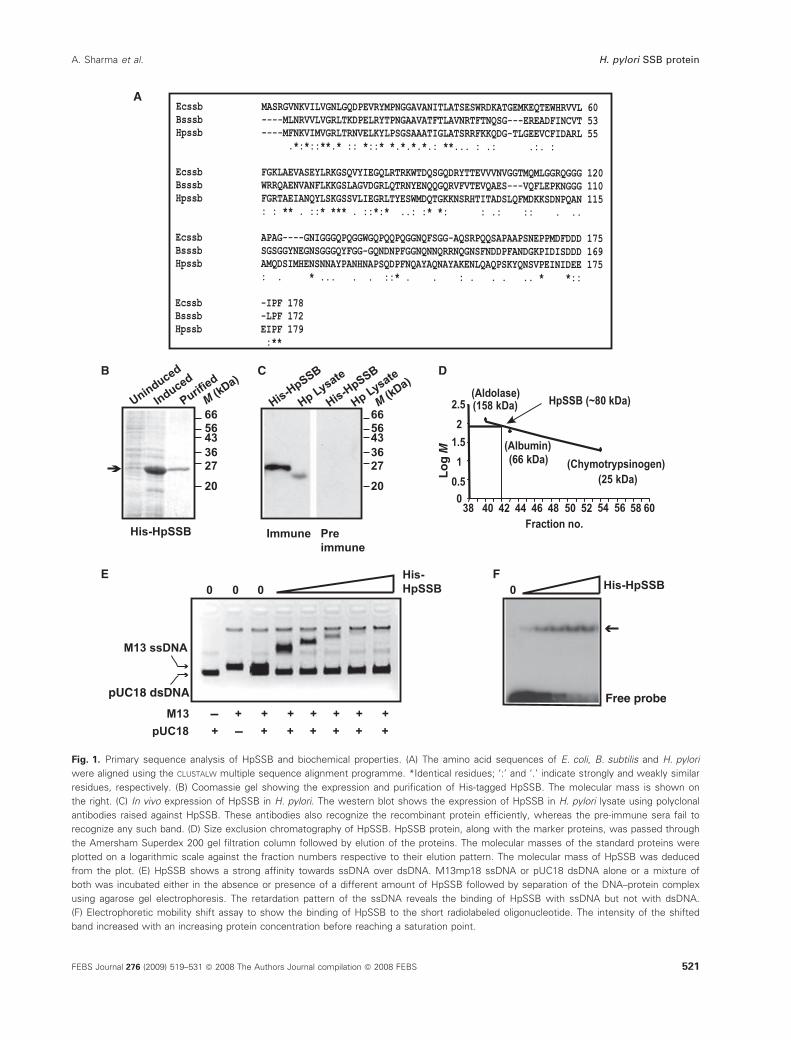
Fig. 1. Primary sequence analysis of HpSSB and biochemical properties. (A) The amino acid sequences of E. coli, B. subtilis and H. pylori
were aligned using the CLUSTALW multiple sequence alignment programme. *Identical residues; ‘:’ and ‘.’ indicate strongly and weakly similar
residues, respectively. (B) Coomassie gel showing the expression and purification of His-tagged HpSSB. The molecular mass is shown on
the right. (C) In vivo expression of HpSSB in H. pylori. The western blot shows the expression of HpSSB in H. pylori lysate using polyclonal
antibodies raised against HpSSB. These antibodies also recognize the recombinant protein efficiently, whereas the pre-immune sera fail to
recognize any such band. (D) Size exclusion chromatography of HpSSB. HpSSB protein, along with the marker proteins, was passed through
the Amersham Superdex 200 gel filtration column followed by elution of the proteins. The molecular masses of the standard proteins were
plotted on a logarithmic scale against the fraction numbers respective to their elution pattern. The molecular mass of HpSSB was deduced
from the plot. (E) HpSSB shows a strong affinity towards ssDNA over dsDNA. M13mp18 ssDNA or pUC18 dsDNA alone or a mixture of
both was incubated either in the absence or presence of a different amount of HpSSB followed by separation of the DNA–protein complex
using agarose gel electrophoresis. The retardation pattern of the ssDNA reveals the binding of HpSSB with ssDNA but not with dsDNA.
(F) Electrophoretic mobility shift assay to show the binding of HpSSB to the short radiolabeled oligonucleotide. The intensity of the shifted
band increased with an increasing protein concentration before reaching a saturation point.
A. Sharma et al. H. pylori SSB protein
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 521

His-tagged. The pre-immune sera under the same
experimental conditions fail to recognize any band,
suggesting the specificity of these antibodies (Fig. 1C).
EcSSB forms homotetramers in solution. The critical
residue for EcSSB homotetramer formation (His55) is
not conserved in HpSSB [26]. To investigate whether
HpSSB forms homotetramers in solution, gel filtration
analysis was performed using different marker proteins
as standards followed by HpSSB. A standard curve
was plotted using the log molecular mass values of
various standard proteins against the fraction numbers
of the proteins at which they are eluted (Fig. 1D).
From this standard curve, the molecular mass of
HpSSB was calculated to be approximately 80 kDa.
These results suggest that HpSSB forms a tetramer in
solution because the molecular mass of monomeric
HpSSB is approximately 20 kDa.
Finally, we investigated the DNA-binding property
of HpSSB. For this purpose, single-stranded M13mp18
DNA or double-stranded pUC18 DNA, or a mixture
of both, was incubated in the absence or presence of
different quantities of HpSSB followed by resolving
the DNA–protein complexes using agarose gel electro-
phoresis (Fig. 1E). The band corresponding to the
M13mp18 ssDNA is retarded significantly with an
increasing amount of HpSSB protein, whereas pUC18
double-stranded DNA (dsDNA) band is not retarded
at all under the same experimental conditions, indicat-
ing that HpSSB shows a strong affinity towards
ssDNA compared to dsDNA. Furthermore, the affin-
ity of HpSSB towards ssDNA was documented by per-
forming gel retardation assay using a small
oligonucleotide single-stranded radiolabeled probe. No
shift was observed in the absence of HpSSB, whereas
an increasing amount of HpSSB resulted in a more
intense shifted band, finally reaching a saturation point
due to the exhaustion of the free probe (Fig. 1F). The
above results indicate that, although HpSSB shows
some differences with EcSSB at the amino acid level,
overall, HpSSB shows oligomeric properties and
ssDNA binding activities similar to that of EcSSB.
Complementation of E. coli Dssb strain with
HpSSB and in vivo localization of HpSSB
in E. coli
Although HpSSB showed oligomerisation and ssDNA
binding activity in vitro typical of SSB related proteins,
we further analyzed its function as a true SSB homo-
log in vivo. For this purpose, we performed plasmid
bumping experiments where we tried to replace an
Ecssb containing plasmid (pRPZ150, ColE1 ori, TcR)
in E. coli RDP317 (Dssb::kan) (a kind gift from
U. Varshney, IISC, Bangalore, India) with plasmids
(AmpR) containing either Ecssb or HpssbWt or
HpssbDC20 (deletion of 20 amino acid residues from
the C-terminus) or pTRC vector (ColE1 ori, AmpR)
alone where the above genes have been cloned [27].
The details of the bacterial strains and plasmid con-
structs are shown in Table 1. It is important to note
that SSB is an essential protein. Therefore, if the
incoming AmpR plasmids containing test SSB coding
Table 1. Bacterial strains and plasmids.
Strain ⁄ plasmid Genotype ⁄ relevant characteristics Reference
DH10b F-mcrAD(mrr-hsdRMS-mcrBC)u80lacZDM15 DlacX74
recA1 endA1 araD139 D(ara, leu)7697
galU galK k- rpsL nupG
Invitrogen
pET28a T7, his, kanR Novagen
pET28a
HpSSBWt and HpSSB DC 20
pET28 a derivative containing 540 bp and 480 bp of
H. pylori SSB full length and C-terminal
deletion mutant
This study
pET28a HpDnaB pET28a derivative containing 1.5 kb of H. pylori dnaB Soni et al. [11]
pGEX-2T HpDnaB pGEX derivative containing 1.5 kb of H. pylori dnaB Soni et al. [12]
BL21 (DE3) F ± ompT hsdSB (rB ± mB) gal dcm (DE3) Novagen
E. coli RDP317 strain Carries a deletion in its chromosomal ssb gene (ssb::Kan)
and a wild-type copy of the ssb gene on a support plasmid,
pRPZ150 (ColE1 ori, TcR)
Gift from U. Varshney
(IISC, Bangalore, India)
pTRC E. coli SSB Wt Plasmid expressing E. coli SSB ColE1 ori, AmpR Gift from U. Varshney
pTRC HpSSBWt and
HpSSB DC 20.
Plasmid expressing HpSSBWt and DC20. ColE1 ori, AmpR This study
pET28a HpSSB-mCherry pET28a derivative expressing fusion protein of Wt HpSSB and mCherry This study
H. pylori 26695 Gift from A. Mukhopadhyay
(NICED, Kolkata, India)
H. pylori SSB protein A. Sharma et al.
522 FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS

genes are capable of complementing the Dssb E. coli
strain, TcR plasmids will show the TcS, AmpR pheno-
type. Using this strategy and Ecssb as a positive con-
trol, we found that continuous subculture of the
bacteria in the media containing Amp and Kan but
lacking Tc helps to replace the TcR plasmid with the
incoming AmpR plasmid with greater than 90% effi-
ciency. Similarly HpssbWt shows very high efficiency
(� 87%) compared to that of HpssbDC20 or vector
alone control. The results are summarized in Table 2
and clearly indicate that Hpssb can complement E. coli
Dssb strain in vivo. Moreover, the data suggest that the
last twenty amino acid residues are important for
in vivo function of HpSSB because HpssbDC20 cannot
complement the E. coli mutant strain. It is important
to note that the amino acid residues at the extreme
C-terminal residues of EcSSB have been reported to be
essential because they may be involved in protein–pro-
tein interaction [24]. EcSSB, HpSSBWt and
HpSSBDC20 were expressed efficiently in the E. coli
mutant strain as shown by SDS ⁄PAGE and Coomassie
staining of the bacterial lysate from the transformed
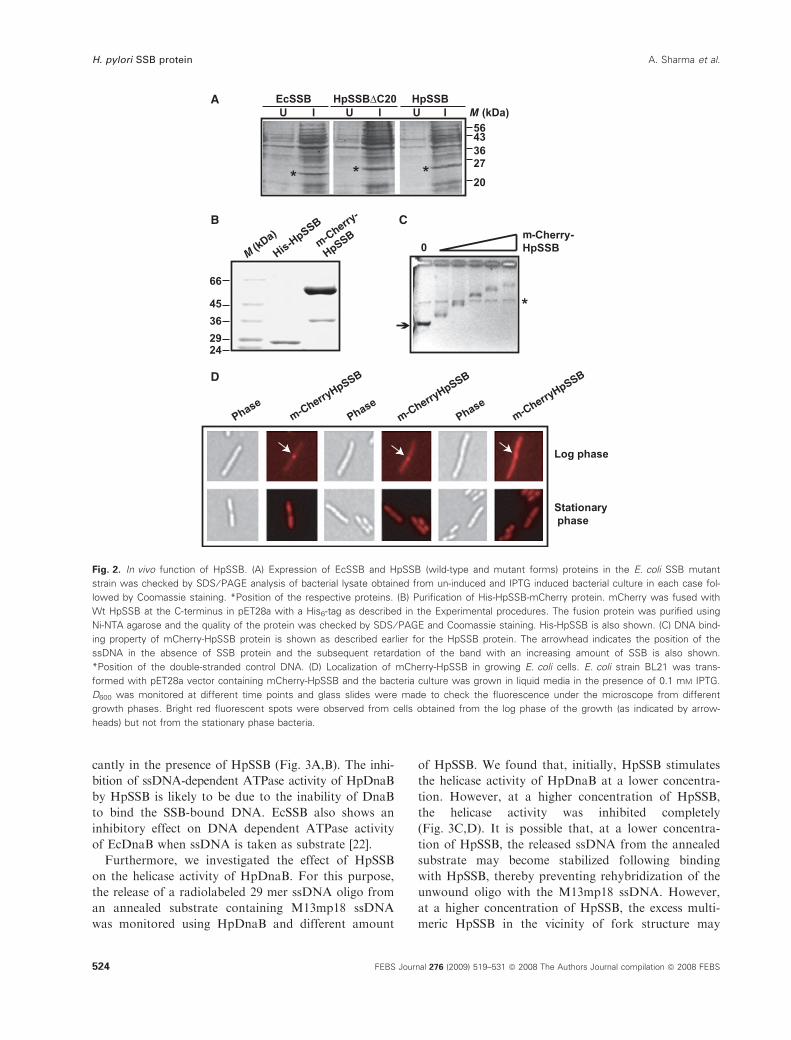
cells (Fig. 2A).
Complementation of E. coli Dssb strain using
HpSSB ensures that it can take over EcSSB function
in vivo. It has been shown recently using green fluores-
cent protein-SSB that replisome machinery containing
replication proteins assemble at the replication origin
[28]. To investigate whether HpSSB can take part in
replisome machinery, we made a His-HpSSB-mCherry
fusion construct where His-tagged HpSSB is fused at
the N-terminus of fluorescent mCherry protein. The
protein was expressed and purified from E. coli BL21
strain and the purified protein was used for DNA
binding activity. mCherry-HpSSB shows DNA-binding
activity that is similar to the Wt HpSSB, suggesting
that the fusion of mCherry does not affect the DNA
binding property of HpSSB (Fig. 2B,C). Subsequently,
we performed in vivo localization experiments using
either lag phase or log phase or stationary phase
E. coli BL21 cells transformed with mCherry-HpSSB
where the expression of mCherry-HpSSB could be
induced using isopropyl thio-b-d-galactoside (IPTG) if
required. The in vivo localization experiments to local-
ize fluorescent mCherry proteins indicate that the
expression of mCherry-HpSSB is poor in the majority
of the lag phase cells, with somewhat diffused staining
pattern (data not shown). Interestingly, the majority of
the cells from the logarithmic phase show moderate
expression of mCherry-HpSSB with distinct foci
(Fig. 2D, upper panel). At the stationary phase of
growth, these cells show expression of mCherry all
over the cell without foci formation (Fig. 2D, upper
panel). Green fluorescent protein-SSB fusion has
recently been used to label replication forks in E. coli
in time-lapse microscopy to demonstrate the dynamics
of replication fork movement during a round of repli-
cation of the bacterial chromosome [28]. We strongly
believe that these distinct foci are the replisome foci
because the foci are not present in the bacteria from
the control stationary phase. These results clearly indi-
cate that HpSSB can take part in the replisome foci in
E. coli, which is consistent with the complementation
of EcSSB mutant strain with HpSSB.
Effect of HpSSB on HpDnaB enzymatic activity
We have shown that HpSSB is a true homolog of SSB
both in vitro and in vivo. SSB interacts with many pro-
teins at the replication fork and modulates their activi-
ties. One of these proteins is DnaB helicase whose
activity can be modulated by SSB [22]. We have
recently cloned, characterized, purified and performed
structure–function analysis of the major replicative
helicase DnaB from H. pylori [13]. We were interested
to see whether HpSSB would modulate the enzymatic
activities of HpDnaB.
One of the hallmarks of the replicative helicases is
its DNA-dependent ATPase activity, which is central
to the helicase activity because it provides energy for
the DNA unwinding and forward translocation on the
replication fork. We have recently shown that the
ATPase activity of HpDnaB can be stimulated many
times in the presence of ssDNA [13]. It has been
reported that DNA-dependent ATPase activity of
DnaB helicase can be inhibited in the presence of SSB
protein [22]. We also found that the ssDNA-dependent
ATPase activity of HpDnaB can be inhibited signifi-
Table 2. Complementation analysis of HpSSB. E. coli RDP 317
Dssb strain was transformed with various AmpR plasmids (as indi-
cated) and, subsequently, they were grown in continuous subcul-
tures in liquid media in the presence of Amp and Kan. Samples
after four subcultures were streaked on agar plates and the resul-
tant single colonies were further patched on agar plates containing
Amp or Amp and Tc. The ability of the patches to grow on the
different plates was monitored and the efficiency of plasmid
replacement was counted. Amp, ampicillin; Tc, tetracyclin.
Test
SSB
genes
No. of
colonies (Amp
resistant)
No. of colonies
(Amp, Tc
resistant)
Efficiency of plasmid
replacement Tc(R)
to Tc(S) (%)
EcSSB 60 5 91
HpSSB 60 8 86.6
HpSSBDC20 52 50 3.8
pTRC vector 50 50 0
A. Sharma et al. H. pylori SSB protein
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 523
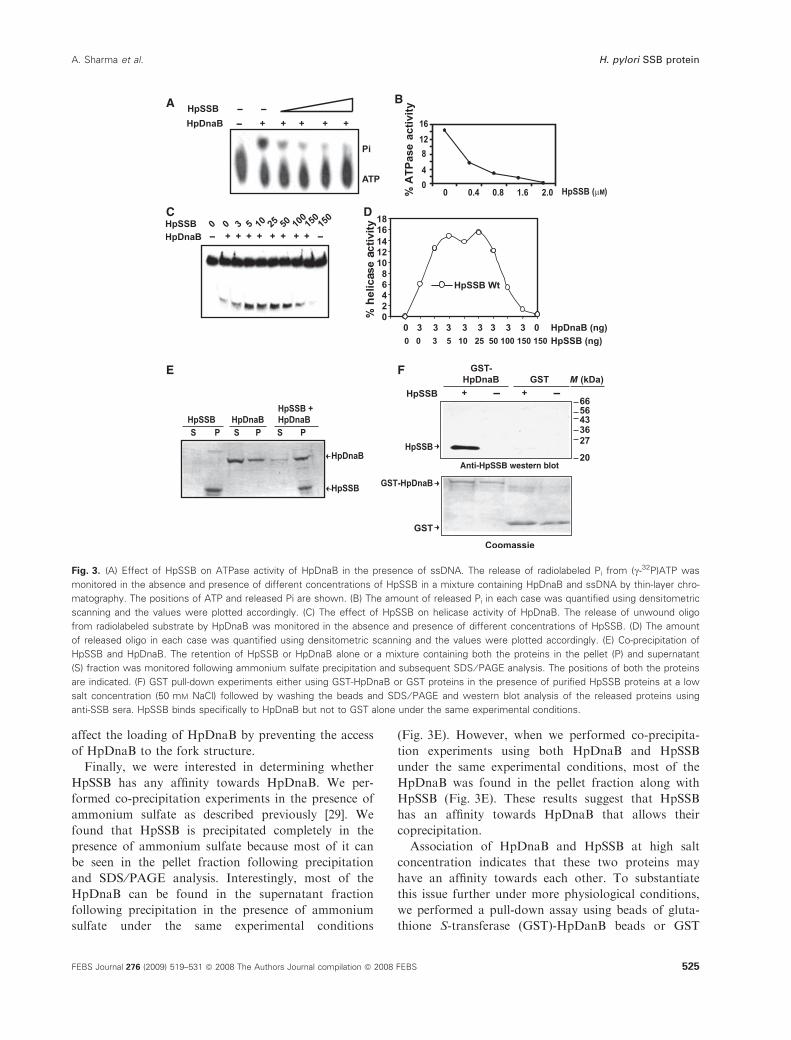
cantly in the presence of HpSSB (Fig. 3A,B). The inhi-
bition of ssDNA-dependent ATPase activity of HpDnaB
by HpSSB is likely to be due to the inability of DnaB
to bind the SSB-bound DNA. EcSSB also shows an
inhibitory effect on DNA dependent ATPase activity
of EcDnaB when ssDNA is taken as substrate [22].
Furthermore, we investigated the effect of HpSSB
on the helicase activity of HpDnaB. For this purpose,
the release of a radiolabeled 29 mer ssDNA oligo from
an annealed substrate containing M13mp18 ssDNA
was monitored using HpDnaB and different amount
of HpSSB. We found that, initially, HpSSB stimulates
the helicase activity of HpDnaB at a lower concentra-
tion. However, at a higher concentration of HpSSB,
the helicase activity was inhibited completely
(Fig. 3C,D). It is possible that, at a lower concentra-
tion of HpSSB, the released ssDNA from the annealed
substrate may become stabilized following binding
with HpSSB, thereby preventing rehybridization of the
unwound oligo with the M13mp18 ssDNA. However,
at a higher concentration of HpSSB, the excess multi-
meric HpSSB in the vicinity of fork structure may
Fig. 2. In vivo function of HpSSB. (A) Expression of EcSSB and HpSSB (wild-type and mutant forms) proteins in the E. coli SSB mutant
strain was checked by SDS ⁄ PAGE analysis of bacterial lysate obtained from un-induced and IPTG induced bacterial culture in each case fol-
lowed by Coomassie staining. *Position of the respective proteins. (B) Purification of His-HpSSB-mCherry protein. mCherry was fused with
Wt HpSSB at the C-terminus in pET28a with a His6-tag as described in the Experimental procedures. The fusion protein was purified using
Ni-NTA agarose and the quality of the protein was checked by SDS ⁄ PAGE and Coomassie staining. His-HpSSB is also shown. (C) DNA bind-
ing property of mCherry-HpSSB protein is shown as described earlier for the HpSSB protein. The arrowhead indicates the position of the
ssDNA in the absence of SSB protein and the subsequent retardation of the band with an increasing amount of SSB is also shown.
*Position of the double-stranded control DNA. (D) Localization of mCherry-HpSSB in growing E. coli cells. E. coli strain BL21 was trans-
formed with pET28a vector containing mCherry-HpSSB and the bacteria culture was grown in liquid media in the presence of 0.1 mM IPTG.
D600 was monitored at different time points and glass slides were made to check the fluorescence under the microscope from different
growth phases. Bright red fluorescent spots were observed from cells obtained from the log phase of the growth (as indicated by arrow-
heads) but not from the stationary phase bacteria.
H. pylori SSB protein A. Sharma et al.
524 FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS
affect the loading of HpDnaB by preventing the access
of HpDnaB to the fork structure.
Finally, we were interested in determining whether
HpSSB has any affinity towards HpDnaB. We per-
formed co-precipitation experiments in the presence of
ammonium sulfate as described previously [29]. We
found that HpSSB is precipitated completely in the
presence of ammonium sulfate because most of it can
be seen in the pellet fraction following precipitation
and SDS ⁄PAGE analysis. Interestingly, most of the
HpDnaB can be found in the supernatant fraction
following precipitation in the presence of ammonium
sulfate under the same experimental conditions
(Fig. 3E). However, when we performed co-precipita-
tion experiments using both HpDnaB and HpSSB
under the same experimental conditions, most of the
HpDnaB was found in the pellet fraction along with
HpSSB (Fig. 3E). These results suggest that HpSSB
has an affinity towards HpDnaB that allows their
coprecipitation.
Association of HpDnaB and HpSSB at high salt
concentration indicates that these two proteins may
have an affinity towards each other. To substantiate
this issue further under more physiological conditions,
we performed a pull-down assay using beads of gluta-
thione S-transferase (GST)-HpDanB beads or GST
A
C D
B
E F
Fig. 3. (A) Effect of HpSSB on ATPase activity of HpDnaB in the presence of ssDNA. The release of radiolabeled Pi from (c-32P)ATP was
monitored in the absence and presence of different concentrations of HpSSB in a mixture containing HpDnaB and ssDNA by thin-layer chro-
matography. The positions of ATP and released Pi are shown. (B) The amount of released Pi in each case was quantified using densitometric
scanning and the values were plotted accordingly. (C) The effect of HpSSB on helicase activity of HpDnaB. The release of unwound oligo
from radiolabeled substrate by HpDnaB was monitored in the absence and presence of different concentrations of HpSSB. (D) The amount
of released oligo in each case was quantified using densitometric scanning and the values were plotted accordingly. (E) Co-precipitation of
HpSSB and HpDnaB. The retention of HpSSB or HpDnaB alone or a mixture containing both the proteins in the pellet (P) and supernatant
(S) fraction was monitored following ammonium sulfate precipitation and subsequent SDS ⁄ PAGE analysis. The positions of both the proteins
are indicated. (F) GST pull-down experiments either using GST-HpDnaB or GST proteins in the presence of purified HpSSB proteins at a low
salt concentration (50 mM NaCl) followed by washing the beads and SDS ⁄ PAGE and western blot analysis of the released proteins using
anti-SSB sera. HpSSB binds specifically to HpDnaB but not to GST alone under the same experimental conditions.
A. Sharma et al. H. pylori SSB protein
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 525

alone in the presence of HpSSB protein. The pull-
down experiments were carried out at a low salt
concentration (50 mm NaCl) followed by washing the
beads first using the binding buffer and, finally, at high
stringency (300–500 mm salt concentration). The pull-
down experiments indicate that HpSSB interact specifi-
cally with GST-HpDnaB but not with control GST
protein under the same experimental conditions
(Fig. 3F). Thus, association of HpDnaB and HpSSB
both at the low and high salt concentrations suggests
that these proteins may physically interact with each
other. A similar interaction has been reported between
replication restart helicase PriA and SSB protein in
E. coli [23].
The interaction of SSB with DnaB helicase appears
to be biologically relevant because the loading of the
HpDnaB helicase may be facilitated by SSB bound to
single-stranded moiety at the fork structure. In chro-
mosomal DNA replication, initiation of Okazaki frag-
ments requires SSB coating of the lagging strand;
similar coating plays a critical function in the restart
of paused replication forks where SSB–DnaB interac-
tions might play critical, although yet undefined roles
[22]. Unlike EcDnaB, HpDnaB does not require a heli-
case loader (EcDnaC) [12]. Hence, HpSSB might have
a closer and more specific interaction with the HpSSB
C-terminal that shows poor homology compared to
that of the EcSSB C-terminal region.
Comparison of localization of replication proteins
between the active helical bacillary form and the
dormant coccoid form of H. pylori
As discussed earlier, H. pylori undergoes morphologi-
cal transition from the spiral shape to the coccoid
form under physiologically unfavorable conditions. It
is reported that the coccoid form is the degenerate
form of the bacteria leading to cell death [30]. There
are also reports indicating the presence of bacterial
enzymatic activities in the dormant form, suggesting
the continuation of metabolic activity at this stage [31–
33]. We compared the DNA replication machinery in
H. pylori in the helical bacillary form and in the coc-
coid stage by attempting to detect and localize active
replication forks. For this purpose, we used two inde-
pendent markers of active growing replication forks
(HpSSB and HpDnaB, respectively) and followed their
localization pattern in the above two forms by immu-
nofluorescence microscopy using specific antibodies
against these markers. We obtained striking results,
where the majority of the active bacillary forms show
clear distinct foci of HpDnaB and HpSSB (wherever
A
C
B
Fig. 4. Immunolocalization of HpDnaB and SSB proteins in H. pylori bacillary and coccoid forms. (A) Glass slides containing either the bacil-
lary form (upper panel) or coccoid form (lower panel) were treated for immunofluorescence (as described in the Experimental procedures)
using HpSSB antibodies (mice, 1 : 500 dilution) followed by FITC conjugated anti-mice sera as secondary antibodies. Green fluorescence
was detected using a fluorescence microscope. (B) Localization of HpDnaB in the above two stages of H. pylori. HpDnaB antibodies (rabbit,
1 : 500) were used as primary antibodies and Alexafluor594 conjugated anti-rabbit sera were used as secondary antibodies. (C) Co-localiza-
tion of HpDnaB and HpSSB proteins in the bacillary form. Both the HpDnaB and HpSSB antibodies were used in combination as primary
antibodies. Alexafluor594 conjugated anti-rabbit and FITC conjugated anti-mice secondary sera were used in combination.
H. pylori SSB protein A. Sharma et al.
526 FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS

staining was obtained) that are the manifestation of
active replication forks in these bacteria (Fig. 4A,B,
upper panels). HpDnaB and HpSSB foci also co-local-
ized completely with each other, confirming the pres-
ence of active replication forks in the bacillary form
(Fig. 4C). These results also suggest that these proteins
are the components of the replisome complex in vivo
and validate our in vitro co-precipitation and pull-
down results (Fig. 3E,F). Interestingly, the coccoid
forms showed diffused staining pattern for both the
proteins (Fig. 4A,B, lower panels). The absence of dis-
tinct replication foci in the coccoid forms clearly sug-
gests that these forms are physiologically different
from the bacillary form. It has been reported previ-
ously that the DNA content of the coccoid forms is
very low compared to the bacillary forms [30]. Taken
together, these results suggest that either very low or
no DNA replication takes place in the coccoid forms.
In summary, we have reported the functional char-
acterization of the SSB protein from an important
pathogen H. pylori. Although it shows divergence from
the EcSSB at the key residues involved in DNA bind-
ing and oligomerization for EcSSB, surprisingly, it can
complement an Ecssb mutant strain and is localized at
the replisome containing growing replication fork in
E. coli and also in H. pylori. Moreover, both DNA-
dependent ATPase and helicase activity of HpDnaB
can be modulated by HpSSB. Whether the modulation
effect is due to the titration of ssDNA in the presence
of HpSSB, or due to the possible interaction between
the two proteins, remains to be elucidated. However,
co-precipitation of HpSSB and HpDnaB, in vitro pull-
down experiments and in vivo co-localization of these
proteins in the bacillary form raise the possibility that
these two proteins may have an affinity with each
other. Finally, the absence of distinct replication foci
in the coccoid form clearly indicates a physiological
difference from the active bacillary form.
Experimental procedures
Bacterial strains
The bacterial strains and plasmids used in the present study
are listed in Table 1. E. coli strains were grown in LB
media (supplemented with 100 mgÆmL)1 ampicillin or
50 mgÆmL)1 kanamycin wherever needed) either at 37 or
22 �C, as required.
H. pylori culture
H. pylori strain 26695 was grown on brain heart infusion
agar (Difco, Sparks, MD, USA) supplemented with 7%
horse blood serum, 0.4% IsoVitaleX and the antibiotics
amphotericin B (8 mgÆmL)1), trimethoprim (5 mgÆmL)1)
and vancomycin (6 mgÆmL)1). The plates were incubated at
37 �C under microaerobic conditions (5% O2, 10% CO2)
for 36 h.
The coccoid form of H. pylori cells was obtained from
the culture plates kept for prolonged periods of 10–14 days,
as described previously [34,35], at 37 �C under the same
conditions. The morphology of bacteria was observed
under the microscope and cells from both the bacillary and
coccoid form cultures were harvested and used for the
immunofluorescence assay.
DNA preparation methods
E. coli plasmids DNA were prepared by the alkaline lysis
method [36]. Bacteriophage M13mp18 single-stranded
circular DNA was prepared as per the protocol described
previously [37]. H. pylori genomic DNA was isolated from
confluent culture grown on BHI agar using the cetyl
trimethyl ammonium bromide-phenol method [38].
DNA manipulation
In the H. pylori genomic database, an ORF (HP1245) was
annotated as the putative HpSSB homolog. The 540 bp
long DNA fragment representing the ORF was amplified
by PCR using H. pylori strain 26695 genomic DNA as tem-
plate with forward and reverse primers having BamHI
restriction sites using Pfu DNA polymerase. Similarly, a
fragment with a deletion of 60 bp representing the last 20
amino acids at the C-terminus of the Hpssb gene was
amplified by PCR.
The PCR-amplified HpssbWt (540 bp) and HpssbDC20(480 bp) DNA fragments were cloned in the expression
vector pET28a (Novagen, Madison, WI, USA) at the
BamHI site and subsequently sequenced. For the
complementation assay, wild-type and HpssbDC20 genes
were subcloned from the respective pET28a recombinant
clones into pTRC vector at the NcoI–HindIII restriction
sites. For pET28a-HpSSB-mCherry constructs, the Hpssb
gene was amplified using the same forward primer, but a
reverse primer without a stop codon and with a SacI site,
and cloned into pET28a at the BamHI–SacI site followed
by cloning of PCR amplified mCherry gene from PRSET-B-
mCherry [39] at the SacI–XhoI site [HpSSB full length for-
ward BamHI, 5¢-CG GGATCCATGTTTAATAAAGTGA
TTATGG-3¢; HpSSB full length reverse BamHI,5¢CG GG
ATCCCTTCATCAATATTGATTTCAGG-3¢; HpSSBDC20reverse BamHI, 5¢-CGGGATCCTCACTGTGCTTGTAA
ATTCTC-3¢; SSB reverse SacI (without stop codon), 5¢-CGAGCTC AAA GGG GAT TTC TTC TTC-3¢; mCherry
forward SacI, 5¢-CGAGCTC ATG GTG AGC AAG GGC
GAG-3¢; mCherry reverse XhoI, 5¢-CCGCTCGAG TTA
CTT GTA CAG CTC GTC C-3¢].
A. Sharma et al. H. pylori SSB protein
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 527

Purification of His-tagged Wt and DC20 HpSSB
protein
E. coli strain BL21 (DE3) (Novagen) harboring pET28a
HpSSB (Wt), DC20 SSB and SSB-mCherry constructs was
grown at 37 �C in LB media containing 50 mgÆmL)1 kana-
mycin. The bacterial cultures were induced for the expres-
sion of the recombinant proteins using 0.25 mm IPTG at
22 �C for 4 h. His-tagged proteins were purified using
Ni-NTA agarose beads (Qiagen, Hilden, Germany) in
accordance with the manufacturer’s instructions. The eluted
proteins were dialyzed against dialysis buffer containing
50 mm Tris–Cl (pH 7.5), 1 mm EDTA, 100 mm NaCl,
100 mm phenylmethanesulfonyl fluoride and 10% glycerol.
For helicase and ATPase assays, HpSSB (Wt) and
DC20SSB were dialysed against MonoQ and MonoS buf-
fers and subjected to ion exchange chromatography using
MonoQ and MonoS ion-exchange columns (GE Health-
care, Uppsala, Sweden) in accordance with the manu-
facturer’s instructions. The fractions of ion exchange
chromatography were then checked on 10% SDS ⁄PAGE
and pooled and dialysed against dialysis buffer.
Protein concentrations were determined by the Brad-
ford method (Bio-Rad, Hercules, CA, USA) in accor-
dance with the manufacturer’s instructions with BSA as
standard. Western blot analysis was carried out following
standard procedures to check the proteins.
Agarose gel retardation assay
ssDNA binding activity of Wt HpSSB, DC20 and HpSSB-
mCherry was checked by incubating the Wt and DC20SSB protein in varying concentrations (0, 0.45, 0.9, 1.8,
2.7 and 3.6 lg, respectively) with 300 ng of M13mp18
single-stranded circular DNA and ⁄ or 300 ng of pUC18
double-stranded circular DNA in binding buffer (20 mm
Tris–HCl, pH 8.0, 1 mm MgCl2, 100 mm KCl, 8 mm
dithiothreitol, 4% sucrose and 80 lgÆmL)1 BSA) in a
20 lL reaction mixture. After 30 min of incubation on ice,
reaction mixtures were resolved in 0.7% agarose gel along
with M13mp18 ssDNA alone, as a control. The increas-
ing retardation of the nucleoprotein complex with increas-
ing concentrations of SSB indicates the ssDNA binding
activity of test proteins. BSA was taken as a negative
control.
Electrophoretic mobility shift assay
Thirty-two nucleotide ssDNA oligo (CGGGA CCATGCG
CCAAAAAATGCCTAAAGAC) from Microsynth (Balgach,
Switzerland) was radiolabeled using (32P)ATP(cP) with the
help of polynucleotide kinase enzyme and the purified
labeled oligos were incubated in the absence or presence of
HpSSB (20, 60, 100, 140 and 180 ng) in binding buffer
(20 mm Tris–HCl, pH 8.0, 1 mm MgCl2, 100 mm KCl,
8 mm dithiothreitol, 4% sucrose, 80 lgÆmL)1 BSA) for
30 min at room temperature (25 �C) and separated on a
6% native PAGE. The native gel was run at 150 V for 2 h
in 1 · TBE buffer (Tris 89 mm, pH 8, boric acid 89 mm,
EDTA 2 mm). The complex and the free DNA were visual-
ized by autoradiography.
Oligomerization status
Wt HpSSB (500 lg) was subjected to size-exclusion
chromatography on a Pharmacia Superdex 200 gel filtration
column (Amersham Biosciences, Uppsala, Sweden) in a
buffer containing 50 mm Tris–HCl (pH 7.4), 1 mm EDTA,
100 mm phenylmethanesulfonyl fluoride, 10% glycerol,
10 mm b-mercaptoethanol and 100 mm NaCl. The column
was previously calibrated using Pharmacia low- and high-
molecular weight standards as indicated. Fractions
(0.3 mL) were collected and checked for the presence of
proteins by SDS ⁄PAGE.
ATP hydrolysis assay
The ATPase activity of HpDnaB with and without SSB
was measured in a reaction mixture (20 lL) containing
20 mm Tris–HCl (pH 8.0), 1 mm MgCl2, 100 mm KCl,
8 mm dithiothreitol, 4% sucrose, 80 lgÆmL)1 BSA, 1 mm
ATP, 3.4 fmol of (c-32P)ATP and the required amount of
DnaB (50 ng), along with 1 pmol of M13mp18 ssDNA and
various concentrations of SSB. The reaction mixtures were
incubated at 37 �C for 30 min and the reactions were
stopped by putting the tubes on ice. Released inorganic
phosphate (Pi) was separated by thin-layer chromatography
on a poly ethylenemine cellulose strip (Sigma-Aldrich,
St Louis, MO, USA) in 0.5 m LiCl and 1 m formic acid at
room temperature for 1 h. The thin-layer chromatography
plate was dried, autoradiographed and analyzed by a phos-
phorimager (Fuji�lm-BAS-1800; Fuji, Tokyo, Japan) for
quantitation.
Helicase assay
The substrate for helicase assay was prepared by annealing
a 29 mer oligo (5¢-CCAAAACCCAGTCACGACGTTGT
AAAACG-3¢) to M13mp18 single-stranded circular DNA.
This annealed substrate has a six bases long 5¢ tail. Helicase
assay was carried out in a 20 lL reaction mixture contain-
ing 20 mm Tris–Cl (pH 8.0), 8 mm dithiothreitol, 2.5 mm
MgCl2, 2 mm ATP, 80 lgÆmL)1 BSA, 10 mm KCl, 4%
sucrose and 10 fmol of helicase substrate and the indicated
amount of HpDnaB and HpSSB. HpDnaB protein (3.0 ng)
was incubated in above buffer for 15 min (on ice) and then
the indicated amount of HpSSB was added to the reaction.
This mixture was incubated at 37 �C in a water bath for
H. pylori SSB protein A. Sharma et al.
528 FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS

30 min. The reaction was stopped by the addition of 5 lLof 5 · stop buffer (1.25% SDS, 75 mm EDTA, 25% glyc-
erol) and the products were separated in 10% native gel
(run in 0.5 · TBE). The gel was dried and exposed to
X-ray film and the quantification was performed using a
phosphorimager.
Complementation assay – plasmid bumping
assay
E. coli RDP317 strain, in which the chromosomal ssb gene
is replaced by a kanamycin resistance (ssb::Kan) marker
and harboring a support plasmid pRPZ150 (ColEl ori,
TcR) coding wild-type SSB protein was transformed with
the pTRC HpSSB Wt or pTRC HpSSBDC20 or pTRC
vector alone. Transformants were grown in the presence
of ampicillin (100 lgÆmL)1) and kanamycin (25 lgÆmL)1)
in four consecutive subcultures in 5 mL of LB media, and
then streaked on plain LB agar plates. The isolated single
colony from the streaked plate was patched on LB agar
plates containing ampicillin alone (100 lgÆmL)1) and on
plates containing both tetracycline (25 lgÆmL)1) and ampi-
cillin (100 lgÆmL)1). The number of patches growing in
both the plates was recorded and the plasmid replacement
efficiency was calculated as a percentage. Because SSB is
an essential protein, the original TcR plasmid can be
replaced by the incoming AmpR plasmid only when it
complements the function of Ecssb in vivo. The conversion
of TcR strain into TcS AmpR phenotype shows that the
HpSSB complements the Dssb strain of E. coli. The com-
plementation ability is analysed in terms of the plasmid
replacement efficacy of the E. coli mutant strain trans-
formed with plasmid containing test genes [27]. The
pTRCEcssb was used as a positive control whereas pTRC
vector alone was used as a negative control to assess the
efficacy of the experiment.
Replication foci study
E. coli BL21 (DE3) strain was transformed with pET28a
HpSSB-mCherry construct and the transformants were
grown in 100 mL of LB media containing Kanamycin
(50 lgÆmL)1) in the absence and presence of various con-
centration of IPTG (0.1–2 mm). At various time intervals,
the cell growth was estimated by taking the value at D600
using a spectrophotometer. A 1 mL sample for each growth
stages was collected and the cell pellets were resuspended in
100 lL of growth media. Some 10 lL of this cell suspen-
sion were placed on agarose gel slab (1% agarose gel slab
in 0.9% NaCl or growth media) and covered by a coverslip.
The cells were then observed under a Nikon fluorescent
microscope (Nikon, Tokyo, Japan). The excitation and
emission spectra for mCherry are 587 and 610 nm, respec-
tively. Images were analysed for red fluorescence signals for
a number of bacterial cells.
Co-precipitation experiments
HpSSB and HpDnaB (alone or a mixture containing both
proteins) were precipitated in the presence of ammonium
sulfate using a previously described protocol [29]. In brief,
HpSSB or HpDnaB or both the proteins (10 lm each) were
mixed in 20 lL of co-precipitation buffer (50 mm Tris–
HCl, pH 7.5, 100 mm NaCl and 10% glycerol) and incu-
bated on ice for 15 min. A solution of 20 lL of ammonium
sulfate (250 gÆL)1, final concentration 125 gÆL)1) was added
to the protein mixture and incubated on ice for another
15 min followed by centrifugation at 10 000 g for 1 min.
Supernatant was collected separately and the pellet was
washed three times with 50 lL of wash buffer (co-precipita-
tion buffer plus 125 gÆL)1 ammonium sulfate). Pellet and
soup fractions were suspended in 2 · SDS ⁄PAGE loading
buffer and 20 lL of each was loaded on 10% polyacryl-
amide gel and stained with Coomassie brilliant blue.
GST pull-down assay
GST pull-down assays were performed by incubating
purified His6-HpSSB (6 lg) in the presence of either GST-
HpDnaB or GST proteins bound on glutathione beads in
the binding buffer (20 mm Tris–HCl, pH 7.5, 1 mm
dithiothreitol, 0.1% NP40, 5 mm MgCl2, 50 mm NaCl,
100 lm phenylmethanesulfonyl fluoride) at room tempera-
ture (25 �C) with gentle rotation. The beads were then
washed three times with binding buffer containing 300 mm
NaCl and the bound proteins were analysed by
SDS ⁄PAGE and western blotting.
Generation of HpSSB antibody
Polyclonal antibodies against His-HpSSB protein were gen-
erated in mice essentially following the protocol described
previously [40]. The antibodies were tested for specificity by
western blot analysis using standard protocols.
Immunofluorescence assays
H. pylori cells were harvested on poly-lysine coated glass
slides and fixed with 4% paraformaldehyde in NaCl ⁄Pi for
15 min at room temperature. Cells were then washed with
NaCl ⁄Pi and treated with 50 mm glucose, 20 mm Tris–HCl,
pH 8.0, 10 mm EDTA, 2 mgÆmL)1 lysozyme and 0.1% Tri-
ton X-100 for 1 h at 37 �C. Cells were further washed with
1 · NaCl ⁄Pi and blocked with 2% BSA in 1 · NaCl ⁄Pi.
Cells were then incubated with primary antibodies (1 : 500
for anti-HpDnaB in rabbit and 1 : 500 for anti-HpSSB in
mice) with 2% BSA in NaCl ⁄Pi at 4 �C overnight. After
washing with NaCl ⁄Pi, cells were incubated with secondary
antibodies [1 : 20 dilution for Alexafluor594 conjugated
IgG antibodies raised in goat against rabbit IgG molecules
A. Sharma et al. H. pylori SSB protein
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 529

and 1 : 20 dilution for fluoroscein isothicyanate (FITC)
conjugated IgG antibodies raised in goat against mouse
IgG molecules; Molecular Probes (Carlsbad, CA, USA)
and Invitrogen (Carlsbad, CA, USA), respectively]. Cells
were further washed with NaCl ⁄Pi and 20% glycerol was
used as the mounting medium. An Axioplan2 fluorescence
microscope (Nikon) was used to capture images. For detec-
tion, a FITC filter was used (exciter 480 ⁄ 40 nm and emitter
535 ⁄ 50 nm). For detection of Alexafluor594, a suitable filter
with an exciter range of 540–590 nm and an emission range
of 600–650 nm was used. axiovision, release 4.6 (Nikon)
software was used for analysis of the images.
Acknowledgements
This work was partially supported by an Indo-Swiss
link grant (SIDA-VR No. 348-2006-6709) provided
to S. D. and S. K. D. We acknowledge Dr Umesh
Varshney (Indian Institute of Science, Bangalore, India)
for providing the E. coli SSB mutant strain RDP317
and Dr Ashish Mukhopadhyay (National Institute of
Cholera and Enteric Diseases, Kolkata, India) for his
help in establishing H. pylori culture. A. S. acknowl-
edges the University Grant Commission (UGC) as well
as the Council of Scientific and Industrial Research
(CSIR). R. G. and A. D. acknowledge UGC for fellow-
ships. Dr Rahma Wehelie (Uppsala University,
Sweden) is greatly acknowledged for providing the
H. pylori culture for the immunofluorescence assay.
References
1 Atherton JC (2006) The pathogenesis of Helicobacter
pylori-induced gastro-duodenal diseases. Annu Rev
Pathol 1, 63–96.
2 Dhar SK, Soni RK, Das BK & Mukhopadhyay G
(2003) Molecular mechanism of action of major Heli-
cobacter pylori virulence factors. Mol Cell Biochem 253,
207–215.
3 Dore MP, Piana A, Carta M, Atzei A, Are BM, Mura I,
Massarelli G, Maida A, Sepulveda AR, Graham DY
et al. (1998) Amoxycillin resistance is one reason for fail-
ure of amoxycillin–omeprazole treatment of Helicobacter
pylori infection. Aliment Pharmacol Ther 12, 635–639.
4 Niv Y (2008) H. pylori recurrence after successful eradi-
cation. World J Gastroenterol 14, 1477–1478.
5 Catrenich CE & Makin KM (1991) Characterization of
the morphologic conversion of Helicobacter pylori from
bacillary to coccoid forms. Scand J Gastroenterol
26(Suppl. 181), 58–64.
6 Cellini L, Allocati N, Angelucci D, Lezzi T, Di Campli
E, Marzio L & Dainelli B (1994) Coccoid Helicobacter
pylori not culturable in vitro reverts in mice. Microbiol
Immunol 38, 843–850.
7 Cellini L (1996) Coccoid forms of Helicobacter pylori.
J Infect Dis 173, 1288.
8 Chan WY, Hui PK, Leung K, Chow J, Kwok F & Ng
CS (1994) Coccoid forms of Helicobacter pylori in the
human stomach. Am J Clin Pathol 102, 503–507.
9 Tomb JF, White O, Kerlavage AR, Clayton RA, Sutton
GG, Fleischmann RD, Ketchum KA, Klenk HP, Gill
S, Dougherty BA et al. (1997) The complete genome
sequence of the gastric pathogen Helicobacter pylori.
Nature 388, 539–547.
10 Alm RA, Ling LS, Moir DT, King BL, Brown ED,
Doig PC, Smith DR, Noonan B, Guild BC, deJonge
BL et al. (1999) Genomic-sequence comparison of
two unrelated isolates of the human gastric pathogen
Helicobacter pylori. Nature 397, 176–180.
11 Soni RK, Mehra P, Choudhury NR, Mukhopadhyay G
& Dhar SK (2003) Functional characterization of Heli-
cobacter pylori DnaB helicase. Nucleic Acids Res 31,
6828–6840.
12 Soni RK, Mehra P, Mukhopadhyay G & Dhar SK
(2005) Helicobacter pylori DnaB helicase can bypass
Escherichia coli DnaC function in vivo. Biochem J 389,
541–548.
13 Nitharwal RG, Paul S, Dar A, Choudhury NR, Soni
RK, Prusty D, Sinha S, Kashav T, Mukhopadhyay G,
Chaudhuri TK et al. (2007) The domain structure of
Helicobacter pylori DnaB helicase: the N-terminal
domain can be dispensable for helicase activity whereas
the extreme C-terminal region is essential for its func-
tion. Nucleic Acids Res 35, 2861–2874.
14 Zawilak-Pawlik A, Kois A, Stingl K, Boneca IG, Skro-
buk P, Piotr J, Lurz R, Zakrzewska-Czerwinska J &
Labigne A (2007) HobA – a novel protein involved in
initiation of chromosomal replication in Helicobacter
pylori. Mol Microbiol 65, 979–994.
15 Natrajan G, Hall DR, Thompson AC, Gutsche I &
Terradot L (2007) Structural similarity between the
DnaA-binding proteins HobA (HP1230) from Helicob-
acter pylori and DiaA from Escherichia coli. Mol Micro-
biol 65, 995–1005.
16 Shamoo Y, Friedman AM, Parsons MR, Konigsberg
WH & Steitz TA (1995) Crystal structure of a replica-
tion fork single-stranded DNA binding protein (T4
gp32) complexed to DNA. Nature 376, 362–366.
17 Webster G, Genschel J, Curth U, Urbanke C, Kang
C & Hilgenfeld R (1997) A common core for binding
single-stranded DNA: structural comparison of the
single-stranded DNA-binding proteins (SSB) from
E. coli and human mitochondria. FEBS Lett 411,
313–316.
18 Murzin AG (1993) OB(oligonucleotide ⁄oligosaccharidebinding)-fold: common structural and functional
solution for non-homologous sequences. EMBO J 12,
861–867.
H. pylori SSB protein A. Sharma et al.
530 FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS

19 Kinebuchi T, Shindo H, Nagai H, Shimamoto N &
Shimizu M (1997) Functional domains of Escherichia coli
single-stranded DNA binding protein as assessed by
analyses of the deletion mutants. Biochemistry 36, 6732–
6738.
20 Kelman Z, Yuzhakov A, Andjelkovic J & O’Donnell M
(1998) Devoted to the lagging strand-the subunit of
DNA polymerase III holoenzyme contacts SSB to pro-
mote processive elongation and sliding clamp assembly.
EMBO J 17, 2436–2449.
21 Richard DJ, Bell SD & White MF (2004) Physical and
functional interaction of the archaeal single-stranded
DNA-binding protein SSB with RNA polymerase.
Nucleic Acids Res 32, 1065–1074.
22 Biswas EE, Chen PH & Biswas SB (2002) Modulation
of enzymatic activities of Escherichia coli DnaB helicase
by single-stranded DNA-binding proteins. Nucleic Acids
Res 30, 2809–2816.
23 Cadman CJ & McGlynn P (2004) PriA helicase and
SSB interact physically and functionally. Nucleic Acids
Res 32, 6378–6387.
24 Curth U, Genschel J, Urbanke C & Greipel J (1996)
In vitro and in vivo function of the C-terminus of
Escherichia coli single-stranded DNA binding protein.
Nucleic Acids Res 24, 2706–2711.
25 Khamis MI, Casas-Finet JR, Maki AH, Murphy JB &
Chase JW (1987) Investigation of the role of individual
tryptophan residues in the binding of Escherichia coli
single-stranded DNA binding protein to single-stranded
polynucleotides. A study by optical detection of mag-
netic resonance and site-selected mutagenesis. J Biol
Chem 262, 10938–10945.
26 Curth U, Bayer I, Greipel J, Mayer F, Urbanke C &
Maass G (1991) Amino acid 55 plays a central role in
tetramerization and function of Escherichia coli single-
stranded DNA binding protein. Eur J Biochem 196,
87–93.
27 Handa P, Acharya N, Thanedar S, Purnapatre K &
Varshney U (2000) Distinct properties of Mycobacte-
rium tuberculosis single-stranded DNA binding protein
and its functional characterization in Escherichia coli.
Nucleic Acids Res 28, 3823–3829.
28 Reyes-Lamothe R, Possoz C, Danilova O & Sherratt
DJ (2008) Independent positioning and action of
Escherichia coli replisomes in live cells. Cell 133,
90–102.
29 Genschel J, Curth U & Urbanke C (2000) Interaction
of E. coli single-stranded DNA binding protein (SSB)
with exonuclease I. The carboxy-terminus of SSB is the
recognition site for the nuclease. Biol Chem 381, 183–
192.
30 Kusters JG, Gerrits MM, Van Strip JAG & Van-
debroucke-grauls MJE (1997) Coccoid forms of Heli-
cobacter pylori are the morphologic manifestation of
cell death. Infect Immun 65, 3672–3679.
31 Bode G, Mauch F & Malfertheiner P (1993) The
coccoid forms of Helicobacter pylori. Criteria for their
viability. Epidemiol Infect 111, 483–490.
32 Cole SP, Cirillo D, Kagnoff MF, Guiney DG &
Eckmann L (1997) Coccoid and spiral Helicobacter
pylori differ in their abilities to adhere to gastric
epithelial cells and induce interleukin-8 secretion. Infect
Immun 65, 843–846.
33 Sorenberg M, Nilsson M, Hanberger H & Nilsson LE
(1996) Morphologic conversion of Helicobacter pylori
from bacillary to coccoid form. Eur J Clin Microbiol
Infect Dis 15, 216–219.
34 Mizoguchi H, Fujioka T, Kishi K, Nishizono A,
Kodama R & Nasu M (1998) Diversity in protein
synthesis and viability of Helicobacter pylori coccoid
forms in response to various stimuli. Infect Immun 66,
5555–5560.
35 Enroth H, Wreiber K, Rigo R, Risberg D, Uribe A &
Engstrand L (1999) In vitro aging of Helicobacter pylori:
changes in morphology, intracellular composition and
surface properties. Helicobacter 4, 7–16.
36 Ausubel FM, Brent R, Kingston RE, Moore DD,
Seidman JG, Smith JA & Struhl K (eds) (1999) Short
Protocols in Molecular Biology, 4th edn, pp. 1–22.
John Wiley & Sons, New York, NY.
37 Sambrook J, Fritsch EF & Maniatis T (ed.) (1989)
Molecular Cloning: A Laboratory Manual, 2nd edn, pp.
4.21–4.32. Cold Spring Harbor Press, Cold Spring
Harbor, NY.
38 Ausubel FM, Brent R, Kingston RE, Moore DD,
Seidman JG, Smith JA & Struhl K (eds) (1998) Current
Protocols in Molecular Biology. Vol. 1, pp. 1.1.1.
John Wiley & Sons, New York, NY.
39 Shaner NC, Campbell RE, Steinbach PA, Giepmans
BN, Palmer AE & Tsien RY (2004) Improved mono-
meric red, orange and yellow fluorescent proteins
derived from Discosoma sp. red fluorescent protein.
Nat Biotechnol 22, 1567–1572.
40 Harlow E & Lane D (1988) Antibodies. Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, NY.
A. Sharma et al. H. pylori SSB protein
FEBS Journal 276 (2009) 519–531 ª 2008 The Authors Journal compilation ª 2008 FEBS 531





























