Embed Size (px)
Citation preview

Ž .Developmental Brain Research 109 1998 281–292
Interactive report
Ontogeny of cation–Cly cotransporter expression in rat neocortex 1
Gerald H. Clayton a,c, Geoffrey C. Owens b,c, Jason S. Wolff a, Roderic L. Smith a,c,)
a Neurology and Pediatrics, Department of Neurology, B-182, UniÕersity of Colorado Health Sciences Center, 4200 E. Ninth AÕe., DenÕer, CO 80262,USA
b Biochemistry and Molecular Genetics, UniÕersity of Colorado Health Sciences Center, DenÕer, CO 80262, USAc Neuroscience Program, UniÕersity of Colorado Health Sciences Center, DenÕer, CO 80262, USA
Accepted 9 May 1998
Abstract
Neuronal precursors and immature cortical neurons actively accumulate Cly and as a consequence depolarize in response to GABAA
receptor activation. With maturity, intracellular Cly decreases resulting in a shift towards GABA inhibition. These observations suggestA
that changes in expression of cation–Cly cotransporters may have a significant role in the ontogeny of neuronal Cly homeostasis. Usingribonuclease protection analysis and in situ hybridization we examined the developmental expression of all presently known members ofthe cation–Cly cotransporter gene family in rat brain. Of the inwardly directed cotransporters, NKCC-1, NKCC-2, and NCC-1, onlyNKCC-1 was detected at significant levels in brain. NKCC-1 was expressed in neurons, appearing first in cortical plate but not inventricular or subventricular zone. Expression levels peaked by the third postnatal week and were maintained into adulthood. Theoutwardly directed cotransporters, KCC-1 and KCC-2, demonstrated significantly different levels and time courses of expression. KCC-1was expressed prenatally at very low levels which increased little over the course of development. In contrast, KCC-2 expressionappeared perinatally and increased dramatically after the first week of postnatal life. Differential changes in expression of this gene familyoccurred during periods of critical shifts in chloride homeostasis and GABA response suggestive of a role in these processes. Furthermorethe absence of expression of known inwardly directed cotransporters in Cly accumulating neuroepithelia and lack of evidence for glialexpression suggests that as yet unidentified members of this gene family may be involved in chloride homeostasis in immature neuronalprecursors and neuroglia. q 1998 Elsevier Science B.V. All rights reserved.
Keywords: GABA; Chloride; Development; Inhibition; Chloride homeostasis; GABA function; RNase protection analysis; In situ hybridizationA
histochemistry
1. Introduction
The functional components of the gamma-amino butyricŽ .acid GABA neurotransmitter system are present during
early development of the mammalian central nervous sys-Ž . w xtem CNS 22,26,32,50 . In the adult brain GABA is the
principal neurotransmitter responsible for inhibition andelicits a hyperpolarizing response by activation of GABAA
w xreceptorrchloride channel complexes 28 . In contrast,GABA activation in the embryonic brain mediates aA
striking depolarization that is sufficient to activate2q w xvoltage-gated Ca channels 24,30 . The embryonic depo-
larizing response to GABA activation is the result of highA
) Corresponding author. Tel.: q1-303-315-5220; Fax: q1-303-315-7583; E-mail: [email protected]
1 Published on the World Wide Web on 27 May 1998.
intra-neuronal concentrations of Cly, which is achieved byw xactive accumulation via furosemide-sensitive transport 24 .
During cortical development, neuronal intracellular Cly
decreases significantly from embryonic to early postnatalw xlife 31 , resulting in a shift from an excitatory to in-
hibitory GABA response. During the first week of post-A
natal life GABA -mediated excitation is observed in hip-A
pocampal pyramidal neurons of CA3 before disappearingw xduring the second postnatal week 3,4,6 . Changes in the
expression of Cly homeostatic mechanisms has been sug-gested as a significant factor in maturation of the GABAA
w xresponse in both neocortex and hippocampus 24,25,28,31 .Developmental control of neuronal Cly homeostasis is
important for understanding the ontogeny of GABA inhi-A
bition and age-related changes in neuronal homeostasis.Recently, increasing insight into the complexity of Cly
homeostasis has been provided by molecular identification
0165-3806r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0165-3806 98 00078-9

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292282
of the components of this process. Mechanisms involvingvoltage-gated Cly channels, cation–Cly cotransport, andanion-exchange are currently receiving intense scrutiny.Members of the voltage-gated Cly channel family that are
w xexpressed at significant levels in brain include ClC-2 49 ,w x w x w x w xClC-3 17 , ClC-4 51 , ClC-5 42,48 , ClC-6 5 , and
w x yClC-7 5 . The voltage-gated Cl channel, ClC-2 has beenshown to stabilize Cly concentrations in adult neuronsw x45–47 . The expression of ClC-2 is also regulated duringdevelopment and both molecular and functional studiessuggest that it contributes to the maturation of neuronal
y w xCl homeostasis 7 . Anion exchange mechanisms mayalso significantly contribute to the developmental changesin Cly homeostasis within the nervous system. Theseproteins have been identified in neuronal and non-neuronalcells, have been shown to be developmentally regulated insome populations, and have been suggested to play a role
w xin neural induction 20,21,23,39,44 .Although voltage gated Cly channels, such as ClC-2,
are capable of preventing Cly accumulation and can func-tion to prevent neuronal Cly accumulation other mecha-nisms are required for the cell to actively accumulate orextrude this ion. The finding that some vertebrate neuronsare capable of active accumulation of Cly by cation–Cly
cotransporters was an important step in elucidating aw xmechanism for GABA -mediated depolarization 1,2 . TheA
cation–Cly cotransporter gene family now consists of 5members that are designated by their dependence on cation
Ž . qgradients as KCC KCC-1 and KCC-2 for K dependenty q ŽCl cotransporters, NCC for Na dependent NCC-
. Ž1rTSC-1 , or NKCC NKCC-1rBSC-2 and NKCC-
.2rBSC-1 for cotransporters that depend on the transmem-q q w xbrane gradients of Na and K 8,9,12,33,34,52 . The
only thiazide sensitive cotransporter identified to date,NCC-1, has been localized only the renal cortex in adult
w xanimals 9 . Those cotransporters inhibitable by furosemideare NKCC-1, NKCC-2, KCC-1, and KCC-2 and to datethree members of this gene family, NKCC-1, KCC-1, andKCC-2, have been found to be expressed in brainw x8,12,35,36 . The finding that the initially depolarizingactions of GABA agonists on neuronal precursors areA
dependent on active accumulation of Cly against the con-centration gradient by a furosemide-inhibitable mechanismfurther suggests an important role for inwardly directed
y w xcation–Cl cotransport 24,31 . Postnatally, the intraneu-ronal concentration of Cly decreases until an adult patternis attained with Cly maintained at concentrations less than
w xpredicted by E 31 suggesting a role for outwardlyCl
directed transport mechanisms such as the recently de-q y w xscribed K dependent cation–Cl cotransporters 12,35 .
These observations collectively provide strong support forthe concept that changes in expression of cation–Cly
cotransporters are important for ontogeny of mature neu-ronal Cly homeostasis and GABA function.
The experiments presented here describe an analysis ofthe expression of cation–Cly cotransporters in developing
rat neocortex using in situ hybridization histochemistryŽ . Ž .ISHH and ribonuclease protection analysis RPA . Thecombination of these molecular and histologic techniquesprovides the sensitivity and the cellular resolution neces-sary for an accurate analysis. These data suggest a role inthe maturation of the GABA response for a limitedA
subset of the cation–Cly cotransporter gene family.
2. Materials and methods
All tissues for these experiments were obtained fromŽSprague Dawley Rats Harlan Sprague Dawley, Inc.; Indi-
.anapolis, IN . Prenatal embryos and early postnatal pupswere of undetermined sex and late postnatal and adultanimals were male.
2.1. Cloning of a partial rat NKCC-1rBSC-2 cDNA andinitial characterization of expression
Ž .Murine BSC-2 mBSC-2 has been cloned and thew xcomplete sequence is available 8 . In the course of PCR
screening for cation–Cly cotransporters in rat brain usingthe degenerate prim ers 5X-RTNM GNTGYAT -GYTNAAYATHTGG-3X and 5X-NMMNSWRAARTT-DATNARNGCRTA-3X, partial clones for rat BSC-2Ž .rBSC-2 were obtained corresponding to transmembranedomains 1–10, which is a 1.2 kb region from bp 985–2187
w xof the published mouse sequence 8 . These primers arepotentially capable of amplification of all known membersof both the thiazide and furosemide sensitive cotrans-porters because of the conserved nature of transmembranedomains 1 and 10, although intervening sequences diverge
Ž .widely. Multiple independent clones 6 were sequencedyielding identical results for the 5X and 3X ends of this 1.2kb clone. These data were highly homologous to equiva-lent regions of the mouse BSC-2 sequence described above.A truncated subclone corresponding to the 5X end of the
Ž .larger clone bp 955–1177 referenced to the mouse clone ,was prepared by XbaI restriction digest and was used togenerate probes for both in situ hybridization and RPA
Ž .analysis. This partial clone accession no. AF061084 is92% homologous to bp 985–1177 of mBSC-2 at thenucleic acid level and 97% homologous at the amino acid
w xlevel 8 which spans membrane domain regions 1 and 2.Both the shortened and 1.2 kb probes have been used for
Ž .in situ hybridization ISHH with identical results whereasthe short clone only was utilized for ribonuclease protec-
Ž .tion analysis RPA . Details of probe labeling and subse-quent ISHH and RPA follow below.
2.2. Cloning of rat NKCC-2rBSC-1
A clone homologous to 1–575 of the published ratŽ .sequence accession a U10096 was generously provided
w xby Dr. S.C. Hebert 9 . This clone, which has no signifi-cant homology with other co-transporter genes, was used

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292 283
as template for the production of riboprobes for ISHHstudies and RPA.
2.3. Cloning of rat NCC-1rTSC-1
A 285 bp clone from the 3X UTR of the publishedŽsequence for rat NCC-1rTSC-1 Genebank accession a
w x.U10097, bp 3705-3989 9 was obtained via standardŽRT-PCR methods cDNA cycle kitrTA ‘TOPO’ cloning
.kit; Invitrogen according to the manufacturers’ instruc-tions from adult rat kidney using 5X-CAGAGTCCT-TATBTBBTCCAGGCTG-3X and 5X-TTTTCAAAGCACG-GGTCACTG-3X as forward and reverse primers, respec-tively. This 3X UTR region lacks any significant homologywith other members of the cation–Cly cotransporter genefamily. This clone was appropriately linearized by restric-tion digest and used as template for the generation of senseand anti-sense digoxigenin labeled riboprobes per methodsdescribed below. Riboprobes were subsequently used forin situ hybridization and ribonuclease protection analysisof cotransporter expression in developing brain.
2.4. Cloning of rat KCC-1
A 233 bp clone from the 3X UTR region of the pub-Žlished rat sequence accession aU55815, bp 3278–3510
w x. X35 was obtained from adult rat brain using 5 -CCTG-GAGTTGGGTTGTCTAAGA-3X and 5X-CATCAGCC-CTCACCAGTCATCTC-3X as forward and reverse primers,
Žrespectively, via standard RT-PCR methods cDNA cycle.kitrTA cloning kit; Invitrogen according to the manufac-
turers’ instructions. This clone, lacking significant homol-ogy to other co-transporter genes, was subsequently usedas template for the digoxigenin and 32 P labeled riboprobesused for ISHH and RPA, respectively.
2.5. Cloning of rat KCC-2
A 399 bp clone from the amino terminal end of theŽ w x.published sequence accession a U55816, bp 4–402 35
was obtained from adult rat brain using 5X-CTCAACAAC-CTGACGGACTG-3X and 5X-GCAGAAGGACTCCAT-GATGCCTGCG-3X as forward and reverse primers, re-
Žspectively, via standard RT-PCR methods see KCC-1.above according to the manufacturers’ instructions. This
clone was shortened to 168 bp by restriction digest withEcoRVrNcoI such that the resulting product correspondedto bp 119–286 of the published rat sequence. The short-ened clone, which uniquely identifies rKCC-2, was subse-quently used as template for the digoxigenin and 32 Plabeled riboprobes used for ISHH and RPA, respectively.
2.6. Tissue preparation
Embryonic brains were immersion fixed in molecularŽ .biology grade HC fixative Amresco, Solon, OH for 24 h
at room temperature, dehydrated and paraffin embedded bystandard methods. Postnatal and adult rat brain was pre-pared by perfusion with molecular biology grade HCfixative followed by immersion fixation for 2 h. The brainwas then cut into 0.5 cm slices and fixed overnight byimmersion in fresh fixative prior to dehydration and paraf-fin embedding. Ten mm sections were prepared from
Žparaffin blocks and mounted on pretreated slides Super-.frost plus, Fisher , and stored desiccated at y2008C until
use. Immediately prior to use, the tissue sections werecleared of paraffin with xylene washes and rehydrated.
( )2.7. In situ hybridization histochemistry ISHH
In situ hybridization probes were prepared from thetemplate DNAs described above. Digoxigenin labeledcRNA was generated from template DNA linearized withthe appropriate restriction enzyme using either SP6, T3 orT7 DNA dependent RNA polymerase in the presence of alabeling mixture containing digoxigenin-UTP and UTP at
Ž . w xa ratio of 0.54 Boehringer-Mannheim 45 . Only thecRNA prepared from the 1.2 kb region of rat NKCC-1 wassubjected to partial alkaline hydrolysis prior to use. In situhybridization for detection of all species of mRNA wasperformed at a final probe concentration of 5 mgrml witheither an anti-sense or sense-strand specific probe tran-scribed from a plasmid linearized with the appropriaterestriction enzyme. The conditions of hybridization, RNase
w xtreatment and washing were as previously described 45 .To control for non-specific signal, Levamisol was added tothe chromagen reaction. In addition, sense and anti-sensestrand probes were reacted simultaneously and the chroma-gen reaction was stopped before any signal was apparentin tissue hybridized with sense strand probes. For all agesof rat brain multiple tissue sections prepared from at leasttwo animals were utilized.
2.8. Imaging
Images of representative sections were captured using aŽCOHU 4900 series CCD camera Cohu, Inc.; San Diego,
. ŽCA fitted to a Nikon Optiphot microscope Nikon Inc.;. ŽMelville, NY equipped with Hoffman optics Modulation
.Optics, Inc.; Greenvale, NY . Digitized images were cap-Žtured with a PowerMac 7100 computer Apple, Inc., Cu-
.pertino, CA equipped with a video framegrabber cardŽ .LG-3, Scion Corp.; Frederick, MD and the public domainimage analysis software ‘NIH Image’ 2. Images were thenprocessed using the software package Photoshop 3.0.5Ž .Adobe Systems Inc., Mountainview, CA and printed on a
ŽTektronics Phaser IISDX dye-sub printer Tektronix, Inc.,.Wilsonville, OR .
2 http:rrrsb.info.nih.govrnih-image
( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292284
( )2.9. Ribonuclease protection analysis RPA
Ž .Ribonuclease protection analysis RPA was performedŽusing total RNA obtained by extraction with Trizol Life
.Technologies; Gaithersburg, MD from the homogenizedŽ . Ž .brains of embryonic E15 , postnatal P0, P7, P21 and
adult animals and in some experiments from astrocyticŽ . Ž . w xA7 and Schwann 1.17 cell lines 10,38 . At least 6brains were pooled at embryonic time points and withincreasing developmental mass fewer brains were pooled,
Ž .however no fewer than 2 brains were combined adult .For use as positive controls in some experiments totalRNA from adult rat kidney was prepared as above. Foranalysis 10 mg of total RNA from each time point washybridized with 32 P labeled derived from DNA templates
Ž32 .described above. Labeling P and subsequent RNasedigestion were performed using the Maxiscript and RPA II
Ž .kits Ambion Inc.; Austin, TX per the manufacturers’instructions. These riboprobes were designed to hybridizeand protect fragments of 198, 466, 285, 233, and 168 bpsfor NKCC-1, NKCC-2, NCC-1, KCC-1, and KCC-2, re-spectively. RNA was simultaneously hybridized with a 32 Plabeled probe which protected a 104 bp fragment of theconstitutively expressed housekeeping gene cylclophilinŽpTRI-Cyclophilin DNA template, Ambion Inc.; Austin,
.TX . This was done to provide a normalizing reference forsubsequent analysis. Both probes were also hybridizedwith 10 mg of total yeast RNA as a control. Precipitatedhybrids were then electrophoresed on a sequencing gelŽ .6% Long Ranger; J.T. Baker Inc., Phillipsburg, NJ under
w xdenaturing conditions using standard methods 43 andscanned and analyzed using a Molecular Dynamics Phos-
Žphoimager or via routine autoradiographic methods Sun-.nyvale, CA .
3. Results
3.1. Expression of inwardly directed cotransporter genesin deÕeloping brain
The ontogeny of neuronal chloride homeostasis in-volves active accumulation of Cly in the most immatureneurons and active removal in many mature neurons; bothprocesses believed to involve cation–Cly cotransportersand which result in changing actions of GABA stimula-A
tion. The molecular expression of known members of thecation–Cly cotransporter gene family was examined in ratbrain cortex at different stages of development using RPA
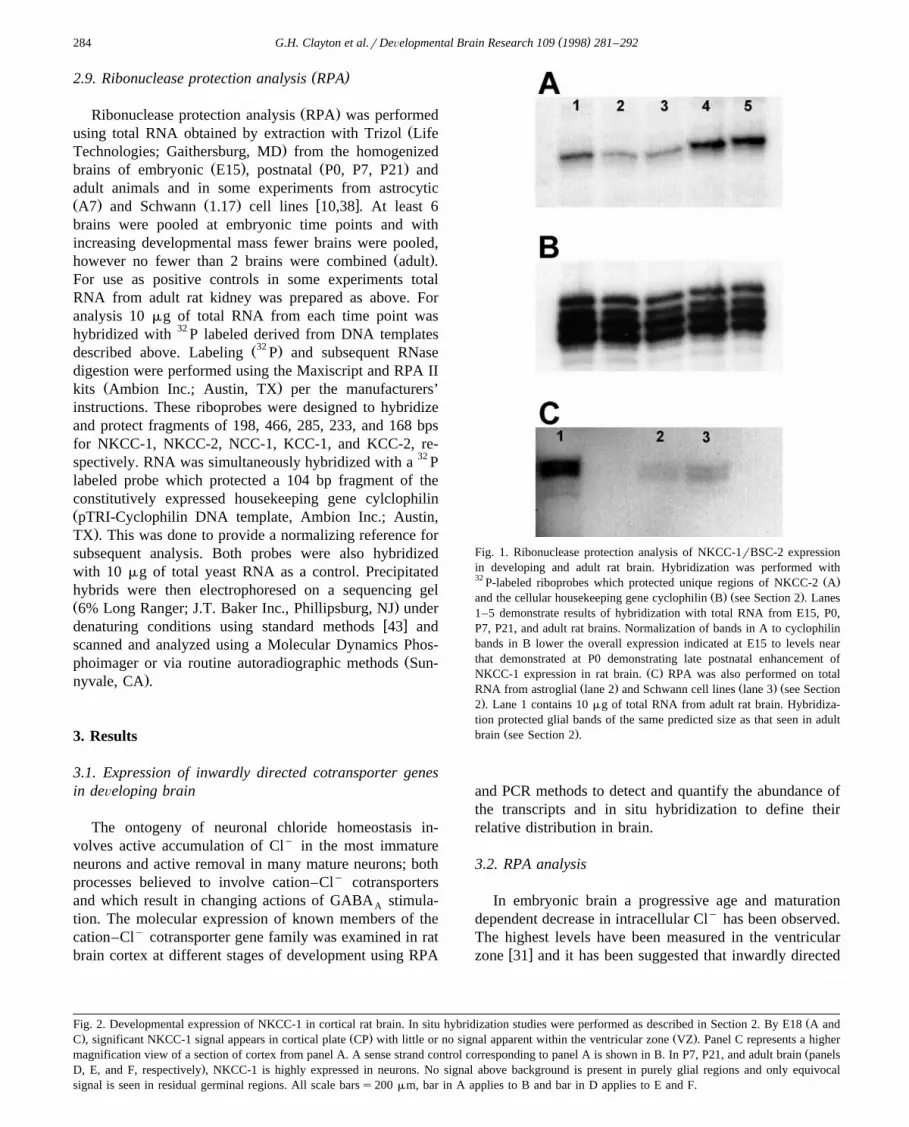
Fig. 1. Ribonuclease protection analysis of NKCC-1rBSC-2 expressionin developing and adult rat brain. Hybridization was performed with32 Ž .P-labeled riboprobes which protected unique regions of NKCC-2 A
Ž . Ž .and the cellular housekeeping gene cyclophilin B see Section 2 . Lanes1–5 demonstrate results of hybridization with total RNA from E15, P0,P7, P21, and adult rat brains. Normalization of bands in A to cyclophilinbands in B lower the overall expression indicated at E15 to levels nearthat demonstrated at P0 demonstrating late postnatal enhancement of
Ž .NKCC-1 expression in rat brain. C RPA was also performed on totalŽ . Ž . ŽRNA from astroglial lane 2 and Schwann cell lines lane 3 see Section
.2 . Lane 1 contains 10 mg of total RNA from adult rat brain. Hybridiza-tion protected glial bands of the same predicted size as that seen in adult
Ž .brain see Section 2 .
and PCR methods to detect and quantify the abundance ofthe transcripts and in situ hybridization to define theirrelative distribution in brain.
3.2. RPA analysis
In embryonic brain a progressive age and maturationdependent decrease in intracellular Cly has been observed.The highest levels have been measured in the ventricular
w xzone 31 and it has been suggested that inwardly directed
ŽFig. 2. Developmental expression of NKCC-1 in cortical rat brain. In situ hybridization studies were performed as described in Section 2. By E18 A and. Ž . Ž .C , significant NKCC-1 signal appears in cortical plate CP with little or no signal apparent within the ventricular zone VZ . Panel C represents a higher
Žmagnification view of a section of cortex from panel A. A sense strand control corresponding to panel A is shown in B. In P7, P21, and adult brain panels.D, E, and F, respectively , NKCC-1 is highly expressed in neurons. No signal above background is present in purely glial regions and only equivocal
signal is seen in residual germinal regions. All scale barss200 mm, bar in A applies to B and bar in D applies to E and F.

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292 285

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292286
Fig. 3. Neither NKCC-2rBSC-1 or NCC-1rTSC-1 are detected in brainby ribonuclease protection analysis. No bands of predicted size were
Ž . Ž . Ž .detected for NKCC-2 A or NCC-1 B in either embryonic E15 oradult brain but were present in kidney. In ‘A’, lane 1 contains undigested
Ž . Ž . Ž .NKCC-2 probe 484 bp while lane 2 E15 brain and lane 3 adult braincontained no protected bands. Lane 4 demonstrates a protected band of
Ž .the correct size 466 bp obtained from kidney RNA. In ‘B’, lane 1Ž . Žcontains undigested NCC-1 probe 433 bp whereas lanes 3 and 4 E15
.and adult brain, respectively exhibit no protected bands. Lane 2 demon-Ž .strates a protected band of appropriate size 285 bp obtained from kidney
RNA. All lanes contained 10 mg of total RNA from the indicated source.Ž .Cyclophilin controls not shown indicated no loading variability.
Cly cotransport is the mechanism by which this occurs.Because of their dependence on Naq the cotransportersNKCC-1, NKCC-2, and NCC-1 promote accumulation ofCly to concentrations higher than predicted by passiveequilibrium distribution. NKCC-1 is known to be ex-
w xpressed in early postnatal life 36 while neither NCC-1nor NKCC-2 are detected in adult brain; however, expres-sion at earlier stages of development have not been exam-ined. To examine the abundance of NKCC-1 and the renal
Fig. 4. Ribonuclease protection analysis of KCC-2 in developing ratbrain. RPA was performed by hybridizing 10 mg of total RNA with32 Ž .P-labeled riboprobes which protected unique regions of KCC-2 A and
Ž .the cellular housekeeping gene cyclophilin B . In both A and B, lanes1–5 contain total RNA from E15, P0, P7, P21, and adult rat brains,respectively. Normalization of bands in ‘A’ to cyclophilin did not alterthe general impression of the results presented.
specific cotransporters NCC-1 and NKCC-2, RPA analysiswas used to measure the total amount of these transcriptspresent in total brain RNA prepared at different develop-mental stages. Protected fragments of the appropriate sizeŽ .see Section 2 were identified in controls which includedrenal RNA samples. As shown in Fig. 1a, NKCC-1 wasdetected as early as E15 and steadily increased about 2–3fold until adult levels of abundance were observed by P21.Expression of NKCC-1 has been previously reported in rat
Ž . w xglioma C6 8 , and was found by us to be present in RNAŽ .samples prepared from transformed astrocytic A7 and
Ž . Ž . w xSchwann cell 1.17 cultures see Fig. 1c 10,38 . Incontrast, neither NCC-1 nor NKCC-2 were detected by
Ž .RPA in any of the developmental stages examined Fig. 3Ž .or in Schwann or astrocytic cultures data not shown but
were present in control RNA prepared from adult ratŽ .kidney Fig. 3 . These results differ from previous studies
which detected a decrease in the level of NKCC-1 in adultbrain using mouse derived sequence to design probes and
w xantibodies, which were used in rat brain 37 . Subsequentstudies have demonstrated that these regions in mouse are
w xextensively processed by alternative splicing 40 . In con-trast, the experiments presented here used a rat derived
Fig. 5. Developmental expression of KCC-2 in cortical rat brain. In situ hybridization analysis of KCC-2 expression confirms the time course of expressionŽ . Ž . Ž .indicated by RPA see Fig. 4 . Panel A shows the lack of hybridization signal at E18. Positions of the cortical plate CP and ventricular zone VZ are
indicated. Hybridization with a sense strand probe in a section equivalent to A is presented in Panel B. Low to moderate levels of signal were observed byŽ . Ž .P0 C with a significant increase in hybridization seen at P7 D . High levels of KCC-2 hybridization were observed within the cortex throughout the rest
Ž . Ž .of postnatal development P21, E and into adulthood F . Hybridization signal was notably lacking in predominantly glial areas. All scale barss200 mm;bar in A applies to B and bar in C applies to D, E, and F.

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292 287

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292288
Ž .partial cDNA clone see Section 2 as a probe, whichincluded essential regions that would not be involved in
w xsplicing 15 . Therefore, these results demonstrate thatNKCC-1 expression continued to increase in adult life,even under conditions where the glial components con-tribute increasingly to the composition of brain RNA.
3.3. Localization of NKCC-1 NKCC-2, and NCC-1 by insitu hybridization
In situ hybridization methods allowed the cell specificexpression of NKCC-1, NKCC-2, and NCC-1 to be exam-ined in developing rat brain. In E15 brain, the NKCC-1hybridization signal first appeared in the region of cortical
Žplate and was not associated with ventricular zone data.not shown . By E18 increasing NKCC-1 hybridization
signal was present in cortical plate but remained absent inŽboth ventricular zone and the sub-ventricular zone see
.Fig. 2a and c . Throughout development the most intensesignal was associated with neurons as has been demon-strated by others. Non-neuronal regions of neocortex ex-
Ž .hibit no NKCC-1 hybridization signal Fig. 2f with theexception of occasional cells at early ages which are likelyneurons known to migrate through various glial regionsduring development. These results are consistent with the
Ž .RPA findings Fig. 1a of increasing levels of NKCC-1extending into adult life in rat brain. In embryonic brain,only modest NKCC-1 signal was detected in choroid plexusŽ .Fig. 2A ; however, prominent expression was observed in
Ž .postnatal choroid plexus data not shown as has beenw xpreviously reported 37 . In contrast, in situ hybridization
did not detect either NKCC-2 or NCC-1 in cortical sec-Ž .tions from either E15 or adult animals data not shown ,
which is consistent with the lack of evidence for expres-sion of these two cotransporters demonstrated by RPAŽ .Fig. 3 .
3.4. Expression of outwardly directed cation Cl y cotrans-porters
In mature neurons outwardly directed cation Cly co-transport is a striking feature of chloride homeostasis. Twomembers of a recently described subgroup of the cation–Cly cotransporter gene family are known to be expressedin rat brain and have been shown to depend heavily on theoutwardly directed Kq gradient for transport of Cly
w x12,35 . Potentially, developmentally regulated control ofexpression of these cotransporter genes could explain the
w yxprogressive decrease in Cl within developing cortexinw x31 and the reversal of GABA mediated depolarizationA
w xthat occurs around the first week of postnatal life 3,4,6,29 .In order to address this hypothesis the overall abundanceof KCC-1 and KCC-2 was examined by RPA methods.
As shown in Fig. 4, KCC-2 is absent in embryonicbrain, initiating expression perinatally with significant in-creases occurring by the second week of postnatal life,which is consistent with the observed timing of changes inchloride homeostasis and GABA response in cortex andA
Fig. 6. Analysis of KCC-1 expression in developing rat brain. RPA was performed by hybridizing 10 mg of total RNA with 32 P-labeled riboprobes whichŽ . Ž .protected unique regions of KCC-1 A and the cellular housekeeping gene cyclophilin B . Lanes 1–5 demonstrate results of hybridization with total RNA
from E15, P0, P7, P21, and adult rat brains. Lane 6 contains an equal amount of total RNA from rat kidney for use as a positive control because thiscotransporter was so difficult to detect in brain. Normalization of bands in ‘A’ to cyclophilin did not alter the general impression of the results presented.Expression of KCC-1 remains low throughout development with little change in expression. In situ hybridization analysis shows KCC-1 expression at a
Ž .representative age P7 in Panel C with a sense strand control presented in panel D. Note the extremely low levels of signal seen by both methods ofanalysis which was characteristic throughout development. All scale barss200 mm; bar in B applies to C; asterisks layer 1 of cortex, Vs layer 5 ofcortex, and arrowheads remnants of layer 6b, a known geminative zone present at postnatal ages.

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292 289
hippocampus. In contrast however, KCC-1 was demon-strated to be expressed at very low levels throughoutdevelopment with only a slight maturational increase evi-
Ž .dent by 3 weeks Fig. 6 .
3.5. Cell specific expression of KCC-1 and KCC-2
In situ hybridization methods were used to determinethe distribution of expression of KCC-1 and KCC-2 at thecellular level. KCC-1 in situ hybridization signal wasrelatively weak at all ages examined consistent with thelow levels of expression observed by RPA analysis. Thestrongest hybridization signal was observed in neuronsŽ .Fig. 6c demonstrating a broad distribution but moreevident in cortical layer V and VIb. Signal was not de-tected in predominantly glial regions; however, low levelexpression might not have been detectable for this lowabundance transcript. This distribution and level of expres-
Ž .sion continued into adulthood data not shown .KCC-2 expression was absent from the early develop-
Ž . Ž .ing brain cf E18, Fig. 5a however by P0 Fig. 5c low tomoderate levels of signal were evident within all layers ofcortex including prominent labeling of layer VIb. Neonatalappearance of KCC-2 correlates well with RPA resultsŽ .see Fig. 4 . A significant increase in cortical expression
Ž .occurs by postnatal day 7 P7, Fig. 5d however, layers Vand VIb remained the most heavily labeled. Further devel-opmental increases occurred with a peak in expressionbeing reached between postnatal day 14 and 21 where
Ž .levels remained stable into adulthood see Fig. 5e and f .KCC-2. In adult cortex the intensity of in situ hybridiza-tion signal did vary with the large pyramidal-like neuronsof layer V standing out amidst lower levels of expressionwithin other layers of cortex.
4. Discussion
This study examined the expression of known membersof the cation–Cly cotransporter gene family during braindevelopment, focusing on events in neocortex. These find-ings provide molecular insight into the known functionalchanges in neuronal chloride homeostasis and GABA func-
w xtion that are observed during cortical development 24,31 .Expression of NKCC-1, KCC-1, and KCC-2 was detectedin neurons, although the pattern and timing of their expres-sion varied significantly. Our results show a temporalcorrelation between the expression of the highly abundantoutwardly directed cotransporter KCC-2 and the timing ofa known decrease in intracellular chloride that is observed
w xpostnatally 31 . The inwardly directed cotransporterNKCC-1 has been found to be expressed during a similartime frame and maintains stable levels in the adult. NKCC-1 is not expressed within the embryonic ventricular zone,an area in which neuronal precursors are known to accu-mulate Cly to levels that are higher than predicted bypassive equilibrium by furosemide inhibitable mechanisms
w x31 . KCC-1 is expressed at barely detectable levels perina-tally and achieves only a small increase in expression byadulthood. The broad distribution evident in cortex and itspresence in a wide variety of tissues as determined by
w xothers 12,35 suggests that KCC-1 may serve a role inneurons other than the maintenance of Cly homeostasis.
4.1. Inwardly directed cation–Cl y cotransporters in de-Õeloping rat brain
A prior analysis of NKCC-1 expression in developingrat brain suggested a marked postnatal attenuation of
w xNKCC-1 expression 36 . The studies of Plotkin et al.utilized probe sequences and generated antibodies againsta fusion protein from a region of murine NKCC-1 that wassubsequently found to be involved in alternative splicingw x40 . The functional significance of alternative NKCC-1mRNA processing remains unknown, but has been well
w xdescribed for the renal NKCC-2 gene 34 . The murinederived probe and antibody were used in developmentalstudies of rat brain, where splicing patterns are unknownand sequence data for comparison with mouse has notbeen obtained. To avoid these potential problems, ourstudies used probes derived from a partial rat cDNA thatincludes the cation-binding site, which is likely to be
w xconserved in all expressed transcripts 15 . Our data clearlydemonstrate that following the initial appearance ofNKCC-1 signal in cortical plate the developmental expres-sion of NKCC-1 increases in rat brain into adult life. Thedifferences in our respective results suggest that age re-lated splicing may occur in rat as reported in mouse.However, the more detailed analysis of splice variants inrat will require cloning of full length cDNA for rat.
[ y]4.2. Maturational changes in Cl are consistent within
the deÕelopmental timing of expression of the outwardlydirected cation–Cl y cotransporter, KCC-2
High embryonic intracellular Cly levels decline signifi-cantly with maturity via two major shifts. The first transi-tion occurs as neuronal precursors move from the ventricu-
w xlar zone to the cortical plate 31 . In rat cortex, the onset ofNKCC-1 expression occurs within newly formed corticalplate and is correlated with the initial transition to a
y Ž .neuronal Cl concentration of 30 mM see Fig. 2a and c .The turning on of NKCC-1 expression with attainment offinal position in cortical plate also coincides with theexpression of a member of the voltage-gated Cly channel
w xfamily, ClC-2 7 . De novo expression of ClC-2 in neuronswhich accumulate Cly has been demonstrated to be suffi-cient to reduce intracellular Cly levels to concentrations
w xthat approach the passive equilibrium potential, E 46 .Cl
The coincidental expression of these two opposing Cly
homeostatic mechanisms therefore could explain why onlyw yxmodest attenuation of Cl is observed in this region ofin
developing brain.

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292290
Changes in cortical Cly homeostasis occurs more grad-ually in the first weeks of postnatal life and results inintracellular Cly concentrations that approach levels found
w xin mature neurons 31 . This process has been suggested toinvolve a class of cation–Cly cotransporters that dependprimarily on the potassium gradient for net outward move-
y Ž .ment of Cl KCC subfamily . In this study, the twoknown members of this class of Kq dependent Cly co-transporters, KCC-1 and KCC-2, demonstrated very differ-ent timing of expression. The expression of significantlevels of KCC-2, which bases its identification as a Kq
w xdependent cotransporter on homology to KCC-1 35 , wasw xdelayed relative to NKCC-1 and ClC-2 7 until early
Ž .postnatal dates see Fig. 4a and Fig. 5a and c . ClC-2 hasbeen shown to prevent Cly accumulation and by thetiming of its expression is likely to play a role in the
w yxmaturational decline in Cl . A developmentally delayedin
postnatal shift in neuronal Cly concentration to net extru-sion within neocortex occurs along a time course similar tothat of KCC-2. These data therefore provide good correla-tive evidence of a major role for KCC-2 mediated outwardCly cotransport in the attainment of mature neuronal Cly
levels.KCC-1 was expressed at very low levels throughout
development with only a modest increase with maturityŽ .see Fig. 6 . Other reports have demonstrated that KCC-1has a broad tissue distribution and has been identifiedwithin primary cultures of cerebral astrocytes as well as
w xC6 glioma cells 12,35 . Our in situ hybridization detectedlow levels of KCC-1 signal within neurons of neocortexŽ .Fig. 6b . Low level expression within glial regions couldhave been below the limits of detection of these methods.However, the combined evidence suggests a broad distri-bution for KCC-1 within brain. The pattern and levels ofexpression observed do not support a major role in thetransitions in Cly homeostasis observed in developingbrain. The apparent ubiquitous nature of KCC-1 is consis-tent with a proposed function in maintenance of cell
w xvolume 12 . Indeed, when KCC-1 was expressed in HEK-293 cells hypo-osmotic challenge and subsequent cellswelling activated the cotransporter providing functional
w xsupport for a role in regulatory volume decrease 12 .
4.3. Absence of known cation–Cl y cotransporters in Õen-tricular zone and glial structures
The actions of exogenously applied GABA agonistsA
on neuronal precursors and immature neurons differ signif-icantly from that observed in adult neurons. Cortical pre-cursors of ventricular zone depolarize in response toGABA agonists which is the result of a high intracellularA
Cly concentration maintained by a furosemide-sensitivew x yprocess 24 . In the ventricular zone, intracellular Cl has
been estimated to accumulate to a concentration of 37mM; however, the extensive connection of these cells bygap-junctions presents technical difficulties to blocking
cationic currents, which may lead to an underestimate ofthe Cly concentration. Chloride accumulation in the ven-tricular zone is inhibitable by the loop-diuretic furosemideand its analog bumetinide, which is a feature of cation–Cly
w x qcotransporters 31 . Because of their dependence on Naeither NKCC-1, NKCC-2, or NCC-1 could mediate Cly
accumulation by virtue of an inwardly directed gradient forthis cation. However, none of the known cation–Cly
cotransporters were detected in ventricular zone suggestingthe possibility that additional members of the gene familymay be responsible for accumulation of Cly within thisregion.
The other striking result from in situ hybridizationstudies was the lack of signal from regions of brain thatcontain exclusively glia and the clear association of hy-bridization signal with morphologically identifiable neu-
Ž .ronal profiles. Fig. 2f, 5f, and 6c . Functional evidencestrongly supports the expression of cation–Cly cotrans-porters in both astrocytes and oligodendrocytesw x11,14,18,19,27 . However, much of the functional charac-terization has been performed using glial cells in culture,which raises the possibility of alteration in expression bygrowth in tissue culture. The significance of this potentialproblem is supported by observation that glial cell line
Ž w x.cultures express NKCC-1 Fig. 1c and 8 while neitherthis study nor previous reports have detected significant
w xexpression in vivo 8,36 . A reported exception to the lackof in vivo expression of NKCC-1 is the transient expres-sion in subpopulation of oligodendrocytes which raises thepossibility of developmental control of expression in vivow x37 . The biologic significance of the expression in oligo-dendrocytes in vivo remains unclear, since expression wasregionally restricted and confined to a period of few daysin postnatal life. The lack of apparent expression of knowncotransporters in glia in vivo suggests that additional mem-bers of the cation–Cly gene family may function in thesecells as well.
4.4. Regulation of cation–Cl y cotransport and changingresponse to GABA actiÕationA
The simultaneous expression of both inwardly and out-wardly directed cation–Cly cotransport mechanisms inneurons has been long suggested based on functional
w xstudies 25 . The expression of NKCC-1 and KCC-2 in thesame cell populations suggests that mechanisms of differ-ential activation must function to avoid the energy cost ofunregulated flux through competing cotransport mecha-nisms. In red blood cells it has been shown that cotrans-porters that functionally resemble KCC-1 and NKCC-1 arereciprocally regulated by activation of second-messenger
w xsystems in response to osmotic challenge 12,13,41 . Suchcontrol mechanisms may also function in neurons. Inaddition, the differential localization of cation–Cly co-transporters with different cationic dependence could allowCly gradients to be established in neurons. Changes in

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292 291
mRNA expression, as investigated in these experiments,are not the only mechanisms by which Cly homeostasismay be modulated. Mechanisms such as alternative mRNAsplicing, changes in protein expression, post-translationalprocessing, and modulation by second-messenger systemsall may have functional effects on Cly homeostasis.
Cation–Cly cotransport mechanisms are only one ofthe components used by neurons to achieve chloride home-ostasis. Voltage gated chloride channels can also con-tribute to chloride homeostasis as has been shown for
w x w xClC-2 45,46 , for review see 16 , which by the timing ofits expression has also been proposed as a mediator of the
w yx w xmaturational change in Cl and GABA function 7 .in A
Anion exchange mechanisms may also significantly con-tribute to the developmental changes in Cly homeostasiswithin the nervous system. These proteins have been iden-tified in neuronal and non-neuronal cells, have been shownto be developmentally regulated in some populations, andhave been suggested to play a role in neural inductionw x20,21,23,39,44 .
As shown in this report and prior studies of NKCC-2w x y37 significant changes in the expression of cation Clcotransporters occur during cortical development. Thesestudies have implicated two members of the cation Cly
gene family, NKCC-1, and KCC-2, as likely primarymediators of Cly homeostasis in the postnatal and adultbrain. Their simultaneous expression would suggest recip-rocal regulation likely plays a role in setting intraneuronalCly concentrations and in determining responses toGABA activation. The data presented also provides evi-A
dence for constitutive expression of KCC-1, which hasw xbeen proposed to serve as a cell volume regulator 12 .
Lack of expression of any of the known inwardly directedcotransporters in cells known to accumulate Cly by
Ž .furosemide inhibitable processes ventricular zone and gliaimply as yet undiscovered members of this gene family areresponsible. Knowledge gained by these studies placed inthe context of data describing other chloride homeostaticmechanisms is a first step toward understanding how thebrain regulates Cly levels and in turn modulates GABAfunction and demonstrate that at least some of the regula-tion of neuronal Cly homeostasis must function at thelevel of gene expression. These studies demonstrate strik-ing changes in the molecular expression of cation–Cly
cotransporter genes during development. The correlation offunctional changes in chloride homeostasis with demon-stration of expression in individual neurons would providefurther support of a definitive role for members of thisgene family in the ontogeny of Cly homeostasis andGABA function. Future development of means for block-ing the expression or activity of individual members of thecation–Cly cotransporter gene family will enable func-tional studies to be done to verify the role that eachmember plays in maintaining or modulating neuronal Cly
homeostasis. Major questions remain regarding the poten-tial role of additional family members of these gene fami-
lies and the more complex issue of differential regulationof multiple chloride homeostatic mechanisms.
Acknowledgements
This work was supported in part by a training grantŽfrom the National Institutes of Health to GHC T32-
.NS07321-08 , and grants to RLS from the March of DimesŽ .a1-0454 and from United Cerebral Palsy.
References
w x y1 F.J. Alvarez-Leefmans, Intracellular Cl regulation and synapticinhibition in verterbrate and invertebrate neurons, in: F. Alvarez-
Ž .Leefmans and J. Russel Eds. , Chloride channels and carriers innerve muscle and glial cells, Plenum, New York and London, 1990,pp. 133–158.
w x2 F.J. Alvarez-Leefmans, S.M. Gamino, F. Giraldez, I. Nogueron,Intracellular chloride regulation in amphibian dorsal root ganglionneurones studied with ion-selective microelectrodes, J. Physiol. 406Ž .1988 225–246.
w x3 Y. Ben-Ari, E. Cherubini, R. Corradetti, J.L. Gaiarsa, Giant synapticpotentials in immature rat CA3 hippocampal neurons, J. Physiol. 416Ž .1989 303–325.
w x4 Y. Ben-Ari, V. Tseeb, D. Raggozzino, R. Khazipov, J.L. Gaiarsa,Ž .Gamma-aminobutyric acid GABA : a fast excitatory transmitter
which may regulate the development of hippocampal neurones inŽ .early postnatal life, Prog. Br. Res. 102 1994 261–273.
w x5 S. Brandt, T.J. Jentsch, ClC-6 and ClC-7 are two novel broadlyexpressed members of the ClC chloride channel family, FEBS Lett.
Ž .377 1995 15–20.w x6 E. Cherubini, J.L. Gaiarsa, Y. Ben-Ari, GABA: an excitatory trans-
Ž .mitter in early postnatal life, TINS 14 1991 515–519.w x7 G.H. Clayton, K.J. Staley, C.L. Wilcox, G.C. Owens, R.L. Smith,
Developmental expression of ClC-2 in the rat nervous system, Dev.Ž .Brain Res. 108 1998 307–318.
w x8 E. Delpire, M.I. Rauchman, D.R. Beier, S.C. Hebert, S.R. Gullans,Molecular cloning and chromosome localization of a putative baso-lateral Na–K–2Cl cotransporter from mouse inner medullary collect-
Ž . Ž .ing duct mIMCD-3 cells, J. Biol. Chem. 269 1994 25677–25683.w x9 G. Gamba, A. Miyansohita, M. Lombardi, J. Lyttton, W.-S. Lee,
M.A. Hediger, S.C. Hebert, Molecular cloning, primary structure,and characterization of two members of the mammalian electroneu-
Ž .tral sodium– potassium –chloride cotransporter family expressed inŽ .kidney, J. Biol. Chem. 269 1994 17713–17722.
w x10 H.M. Geller, M. Dubois-Dalcq, Antigenic and functional characteri-zation of a rat central nervous system-derived cell line immortalized
Ž .by a retroviral vector, J. Cell Biol. 107 1988 1977–1986.w x11 P. Gilbert, H. Kettenmann, M. Schachner, Gamma-aminobutyric
acid directly depolarizes cultured oligodendrocytes, J. Neurosci. 4Ž .1984 561–569.
w x12 C.M. Gillen, S. Brill, J.A. Payne, B. Forbush III, Molecular cloningand functional expression of the K–Cl cotransporter from rabbit, rat,
Ž .and human, J. Biol. Chem. 271 1996 16237–16244.w x13 M. Haas, T.J. McManus, Effect of norepinephrine on swelling-in-
duced potassium transport in Duck red cells, J. Gen. Physiol. 85Ž .1985 649–667.
w x y14 D. Hoppe, H. Kettenmann, GABA triggers a Cl efflux fromŽ .cultured mouse oligodendrocytes, Neurosci. Lett. 97 1989 334–339.
w x15 P. Isenring, B. Forbush III, Ion and bumetinide binding by theŽ .Na–K–Cl cotransporters, J. Biol. Chem. 272 1997 24556–24562.
w x16 T.J. Jentsch, K. Steinmeyer, G. Schwarz, Primary structure of

( )G.H. Clayton et al.rDeÕelopmental Brain Research 109 1998 281–292292
Torepedo marmorata chloride channel isolated by expression cliningŽ .in Xenopus oocytes, Nature 348 1990 510–514.
w x17 M.U.S. Kawasaki, T. Monkawa, A. Miyawaki, K. Mikoshiba, F.Marumo, S. Sasaki, Cloning and expression of a protein kinaseC-regulated chloride channel abundantly expressed in rat brain
Ž .neuronal cells, Neuron 12 1994 597–604.w x18 H. Kettenmann, K.H. Backus, M. Schachner, Gamma-aminobutyric
Ž .acid opens Cl-channels in cultured astrocytes, Br. Res. 404 19871–9.
w x19 H.K. Kimelberg, M.V. Frangakis, Furosemide- and bumetanide-sen-sitive ion transport and volume control in primary astrocyte cultures
Ž .from rat brain, Br. Res. 361 1985 125–134.w x20 S. Kobayashi, C.W. Morgans, J.R. Casey, R.R. Kopito, AE3 anion
exchanger isoforms in the vertegrate retina: Developmental regula-tion and differential expression in neurons and glia, J. Neurosci. 14Ž .1994 6266–6279.
w x21 R.R. Kopito, B.S. Lee, D.M. Simmons, A.E. Linsey, C.W. Morgans,K. Schneider, Regulation of intracellular pH by a neuronal homolog
Ž .of the eryuthrocyte anion exchanger, Cell 59 1989 927–937.w x22 J.M. Lauder, V.K.M. Han, P. Henderson, T. Verdoorn, A.C. Towle,
Prenatal ontogeny of the GABAergic system in the rat brain: AnŽ .immunocytochemical study, Neuroscience 19 1986 465–493.
w x23 A.E. Lindsey, K. Schneider, D.M. Simmons, R. Baron, B.S. Lee,R.R. Kopito, Functional expression and subcellular localization of ananion exchanger cloned from choroid plexus, Proc. Natl. Acad. Sci.
Ž .87 1990 5278–5282.w x24 J.J. LoTurco, D.F. Owens, M.J.S. Heath, M.B.E. Davis, A.R. Krieg-
stein, GABA and glutamate depolarize cortical progenitor cells andŽ .inhibit DNA synthesis, Neuron 1995 1287–1298.
w x25 H.J. Luhmann, D.A. Prince, Postnatal maturation of the GABAergicŽ .system in rat neocortex, J. Neurophysiol. 65 1991 247–263.
w x26 W. Ma, P.A. Saunders, R. Somogyi, M.O. Poulter, J.L. Barker,Ontogeny of the GABA receptor subunit mRNAs in rat spinal cordA
Ž .and dorsal root ganglia, J. Comp. Neurol. 338 1993 337–359.w x27 B.A. MacVicar, F.W.Y. Tse, S.A. Crichton, H. Kettenmann,
GABA-activated Cly channels in astrocytes of hippocampal slices,Ž .J. Neurosci. 9 1989 3577–3583.
w x28 U. Misgeld, R.A. Deisz, H.U. Dodt, H.D. Lux, The role of chloridetransport in postsynaptic inhibition of hippocampal neurons, Science
Ž .232 1986 1413–1415.w x29 C.B. Murray, M.M. Morales, T.R. Flotte, S.A. McGrath-Morrow,
W.B. Guggino, P.L. Zeitlin, ClC-2: A developmentally dependentchloride channel expressed in the fetal lung and downregulated after
Ž .birth, Am. J. Respir. Cell Mol. Biol. 12 1995 597–604.w x30 K. Obrietan, A.N. van den pol, GABA neurotransmission in the
hypothalamus: Developmental reversal from Ca2q elevating to de-Ž .pressing, J. Neurosci. 15 1995 5065–5077.
w x31 D.F. Owens, L.H. Boyce, M.B.E. Davis, A.R. Kriegstein, ExcitatoryGABA responses in embryonic and neonatal cortical slices demon-strated by gramicidin perforated patch recordings and calcium imag-
Ž .ing, J. Neurosci. 16 1996 6414–6423.w x32 J.G. Parnavelas, M.E. Cavanagh, Transient expression of neurotrans-
Ž .mitters in the developing neocortex, TINS 11 1988 92–93.w x33 J.A. Payne, Functional characterization of the neuronal-specific K–Cl
w q xcotransporter: implications for K regulation, Am. J. Physiol. 273Ž .1997 C1516–C1525.
w x34 J.A. Payne, B. Forbush III, Alternatively spliced isoforms of theputative renal Na–K–Cl cotransporter are differentially distributed
Ž .within the rabbit kidney, Proc. Natl. Acad. Sci. 91 1994 4544–4548.w x35 J.A. Payne, T.J. Stevenson, L.F. Donaldson, Molecular characteriza-
tion of a putative K–Cl cotransporter in rat brain, J. Biol. Chem. 271Ž .1996 16245–16252.
w x36 M.D. Plotkin, M.R. Kaplan, L.N. Peterson, S.R. Gullans, S.C.Hebert, E. Delpire, Expression of the Naq–Kq –2Cly cotransporter
Ž .BSC2 in the nervous system, Am. Physiol. Soc. 272 1997 C173–C183.
w x37 M.D. Plotkin, E.Y. Snyder, S.C. Hebert, E. Delphire, Expression ofthe Na–K–2Cl cotransporter is developmentally regulated in postna-tal rat brains: a possible mechanism underlying GABA’s excitatory
Ž .role in immature Brain, J.Neurobiol. 33 1997 781–795.w x38 S. Porter, L. Glaser, R.P. Bunge, Release of autocrine growth factor
by primary and immortalized schwann cells, Proc. Natl. Acad. Sci.Ž .84 1987 7768–7772.
w x39 K.M. Raley-Susman, R.M. Sapolsky, R.R. Kopito, ClyrHCO y3
exchange function differs in adult and fetal rat hippocampal neurons,Ž .Br. Res. 614 1993 308–314.
w x40 J. Randall, T. Thorne, E. Delphire, Partial cloning and charateriza-tion of Slc12a2: the gene encoding the secretory Naq, Kq –2Cly
Ž .cotransporter, Am. J. Physiol 273 1997 C1267–C1277.w x41 J.R. Sachs, D.W. Martin, The role of ATP in swelling-stimulated
K–Cl cotransport in human red cell ghosts, J. Gen. Physiol. 102Ž .1993 551–573.
w x42 H. Sakamoto, M. Kawasaki, S. Uchida, S. Sasaki, F. Marumo,Identification of a new outwardly rectifying Cly channel that be-longs to a subfamily of the ClC Cly channels, J. Biol. Chem. 271Ž .1996 10210–10216.
w x43 J. Sambrock, E.F. Fritich, T. Maniatis, Molecular Cloning: A Labo-ratory Manual, Cold Spring Harbor Press, New York, 1989.
w x44 A.K. Sater, J.M. Alderton, R.A. Steinhardy, An increase in intra-cellular pH during neural induction in Xenopus, Development 120Ž .1994 433–442.
w x45 R.L. Smith, G.H. Clayton, C.L. Wilcox, K.W. Escudero, K.J. Staley,Differential expression of an inwardly rectifying chloride conduc-tance in rat brain neurons: A potential mechanism for cell-specific
Ž .modulation of postsynaptic inhibition, J. Neurosci. 15 1995 4057–4067.
w x46 K. Staley, R. Smith, J. Schaack, C. Wilcox, T. Jentsch, Alteration ofGABA-A receptor function following gene transfer of the ClC-2
Ž .chloride channel, Neuron 17 1996 543–551.w x47 K.J. Staley, The role of an inwardly rectifying chloride conductance
Ž .in postsynaptic inhibition, J. Neurophysiol. 72 1994 273–284.w x48 K. Steinmeyer, B. Schwappach, M. Bens, A. Vandewalle, T.J.
Jentsch, Cloning and functional expression of rat ClC-5, a chlorideŽ .channel related to kidney disease, J. Biol. Chem. 270 1995 31172–
31177.w x49 A. Thiemann, S. Grunder, M. Pusch, T.J. Jentsch, A chloride
channel widely expressed in epithelial and non-epithelial cells, Na-Ž .ture 356 1992 57–60.
w x50 C.G. Van Eden, L. Mrzljak, P. Voorn, H.B.M. Uylings, Prenataldevelopment of GABA-ergic neurons in the neocortex of the rat, J.
Ž .Comp. Neurol. 289 1989 213–227.w x51 M.A. van Slegtenhorst, M.T. Bassl, G. Borsani, M.C. Wapenaar,
G.B. Ferrero, L. de Concillis, E.L. Rugarli, A. Grillo, B. Franco,H.Y. Zoghbi, A. Ballabio, A gene from the Xp22.3 region shareshomology with voltage-gated chloride channels, Hum. Mol. Gen. 3Ž .1994 547–552.
w x52 J. Xu, C. Lytle, T.T. Zhu, J.A. Payne, E. Benz, B. Forbush III,Molecular cloning and functional expression of the bumetanide-sen-
Ž .sitive Na–K–Cl cotransporter, Proc. Natl. Acad. Sci. 91 19942201–2205.




























