Embed Size (px)
Citation preview

ARTHRITIS & RHEUMATISMVol. 54, No. 10, October 2006, pp 3152–3162DOI 10.1002/art.22161© 2006, American College of Rheumatology
Oncostatin M Induces Angiogenesis and Cartilage Degradationin Rheumatoid Arthritis Synovial Tissue and
Human Cartilage Cocultures
Ursula Fearon,1 Ronan Mullan,1 Trevor Markham,1 Mary Connolly,1 Shane Sullivan,2
A. Robin Poole,3 Oliver FitzGerald,1 Barry Bresnihan,1 and Douglas J. Veale1
Objective. To investigate the role of oncostatin M(OSM) in cell adhesion, angiogenesis, and matrix de-gradation in rheumatoid arthritis (RA) synovial tissueand normal human cartilage.
Methods. Human dermal microvascular endothe-lial cell (HDMEC) and RA synovial fibroblast (RASF)proliferation and intercellular adhesion molecule 1(ICAM-1) and vascular cell adhesion molecule 1(VCAM-1) expression were assessed by a bromodeoxy-uridine proliferation assay and flow cytometry. HDMECtubule formation and migration were assessed by Ma-trigel culture and migration assay. Production of matrixmetalloproteinase (MMP) and tissue inhibitor of met-alloproteinases 1 (TIMP-1) in RA synovial explants, andproteoglycan/glycosaminoglycan (GAG) release, vascu-lar endothelial growth factor (VEGF), and angiopoietin2 production from RASF/normal cartilage cocultureswere assessed by enzyme-linked immunosorbent assayand immunohistology.
Results. HDMEC/RASF proliferation was in-duced by OSM and interleukin-1� (IL-1�), alone and incombination. OSM enhanced cell surface expression ofICAM-1, but not VCAM-1, on endothelial cells and
RASFs. OSM increased endothelial cell tubule forma-tion and migration. In RA synovial explants, OSMinduced production of MMP-1 and TIMP-1. When OSMwas combined with IL-1�, however, the MMP-1:TIMP-1ratio was significantly increased. OSM potentiated IL-1�–induced MMP-1 and MMP-13 expression in normalhuman cartilage/RASF cocultures, resulting in a signif-icant increase in the MMP:TIMP ratio. In OSM/IL-1�–stimulated cocultures, cartilage sections demonstrated sig-nificant proteoglycan depletion that was paralleled by asignificant increase in GAG release in supernatants. Fi-nally, compared with either cytokine alone, the combina-tion of OSM and IL-1� significantly induced VEGF pro-duction in RASF/cartilage cocultures.
Conclusion. These data suggest that OSM pro-motes angiogenesis and endothelial cell migration andpotentiates the effects of IL-1� in promoting extracel-lular matrix turnover and human cartilage degradation.Furthermore, the induction of VEGF in coculturessupports the hypothesis of a link between angiogenesisand cartilage degradation.
Rheumatoid arthritis (RA) is a chronic diseasecharacterized by synovial tissue proliferation and artic-ular cartilage degradation (1,2). In RA, angiogenesis isan early event required for pannus development, en-abling activated monocytes to enter the synovium viaendothelial cells by active recruitment (3). The newvessels support expansion of the synovial pannus overthe cartilage, facilitating RA synovial fibroblast (RASF)invasion and cartilage degradation by proteolytic cleav-age of both aggrecan and collagen (4). This processdepends on cytokines and growth factors to stimulatecell survival, proliferation, and extracellular matrix(ECM) degradation (5). Erosion is characterized by aloss of ECM by overexpression of matrix metalloprotein-
Supported by the Health Research Board of Ireland.1Ursula Fearon, PhD, Ronan Mullan, MD, Trevor Markham,
MD, Mary Connolly, BSc, Oliver FitzGerald, MD, FRCP, FRCPI,Barry Bresnihan, MD, FRCP, Douglas J. Veale, MD, FRCPI: St.Vincent’s University Hospital and the Conway Institute of Biomolec-ular and Biomedical Research, Dublin, Ireland; 2Shane Sullivan, MSc:St. Vincent’s University Hospital, Dublin, Ireland; 3A. Robin Poole,PhD, DSc: Shriners Hospitals for Children, Montreal, Quebec, Can-ada.
Address correspondence and reprint requests to Douglas J.Veale, MD, FRCPI, Department of Rheumatology, St. Vincent’sUniversity Hospital and the Conway Institute of Biomolecular andBiomedical Research, Elm Park, Dublin 4, Ireland. E-mail: [email protected].
Submitted for publication October 12, 2005; accepted inrevised form July 11, 2006.
3152

ase (MMP), the activity of which is modulated byspecific tissue inhibitors (e.g., tissue inhibitor of metal-loproteinases 1 [TIMP-1]) (6,7).
The cytokine oncostatin M (OSM) shares a com-mon receptor subunit, gp130 (8), with interleukin-6(IL-6); together, they constitute a family based onpredicted structural similarities and shared signal trans-ducer receptor components (9). OSM is raised in RAsynovial fluid and tissue, and the levels of OSM correlatewith the levels of markers of joint inflammation andcartilage turnover (10–12). OSM alone up-regulatesexpression of acute-phase proteins, increases expressionof angiogenic factors such as vascular endothelial growthfactor (VEGF), fibroblast growth factor (FGF), andchemokines, and stimulates proteoglycan and collagenrelease from cartilage (13–18). In vitro, OSM alonemodulates matrix turnover in synoviocytes, chondro-cytes, and cartilage explants, suggesting a pivotal role forOSM in regulating the balance between MMP andTIMP at the site of destruction (17–21). In addition,OSM has been demonstrated to act synergistically withproinflammatory cytokines such as IL-1�, tumor necro-sis factor � (TNF�), and IL-17 to induce MMP turnover(18,22,23).
In animal models, intraarticular injection or ad-enoviral overexpression of OSM was shown to result insynovial inflammation and cartilage destruction, furthersupporting a pivotal role for OSM in RA (24–27).Conversely, however, studies have demonstrated thatOSM may exert antiinflammatory effects by inhibitingthe effects of the IL-1–induced level of IL-8 andgranulocyte–macrophage colony-stimulating factor (28)and promoting TIMP expression (21).
Angiogenesis and cell migration are key pro-cesses in the proinflammatory response, leading to car-tilage invasion and joint destruction. We hypothesizedthat OSM directly induces angiogenesis and ECM de-gradation, which together, are responsible for cartilagedamage. In this study, we investigated the specific role ofOSM, alone and in combination with IL-1�, in angiogen-esis, cell migration, and ECM degradation, using humandermal microvascular endothelial cells (HDMECs), wholeRA synovial tissue explants, and cocultures of RASFs andnormal human cartilage.
PATIENTS AND METHODS
Patients and RA synovial tissue. All patients fulfilledthe 1987 American College of Rheumatology (formerly, theAmerican Rheumatism Association) criteria for a diagnosis ofRA (29) and had an actively inflamed knee joint. RA synovial
tissue was obtained at the time of arthroscopy, using localanesthesia as previously described (30), prior to the adminis-tration of disease-modifying therapy or oral corticosteroids.Fully informed written consent was obtained from each pa-tient. This study was approved by the St. Vincent’s UniversityHospital Ethics and Medical Research Committee.
Isolation and culture of RASFs and HDMECs. RASFswere obtained by enzymatic digestion of synovial biopsy spec-imens with 1 mg/ml of type 1 collagenase (WorthingtonBiochemical, Lakewood NJ) in RPMI (Gibco BRL, Paisley,UK) for 4 hours at 37°C in humidified air with 5% CO2.Dissociated cells were plated in RPMI 1640 supplemented with10% fetal calf serum (FCS; Gibco BRL), 10 ml of 1 mmole/liter HEPES (Gibco BRL), penicillin (100 units/ml), strepto-mycin (100 units/ml), and Fungizone (0.25 �g/ml) (all fromBiosciences, Plymouth, MN). The cells were incubated andgrown to confluence in T75 flasks (�10 days) at 37°C in ahumidified air with 5% CO2 before being harvested withtrypsin and passaged. RASFs between the fourth and eighthpassages were used for experiments. HDMECs (Clonetics, SanDiego, CA) were incubated in epithelial basal medium (EBM;Clonetics) supplemented with endothelial growth medium(EGM)–microvascular bullet-kit medium (25 ml FCS, 0.5 mlhuman epidermal growth factor, 0.5 ml hydrocortisone, 0.5 mlgentamicin, and 0.5 ml bovine brain extract) (Clonetics). Thecells were grown to confluence in T75 flasks at 37°C inhumidified air with 5% CO2 before being harvested withtrypsin–EDTA (Clonetics) and passaged. HDMECs betweenthe third and eighth passages were used for experiments.
Whole RA synovial tissue explant culture. We estab-lished an ex vivo RA synovial tissue explant model (31,32) toinvestigate the effect of OSM on matrix degradation in the RAjoint. This culture system allows the synovial architecture andcell–cell contact of many cell types to remain intact, and thusreflects more closely the in vivo joint environment. To reducethe effect of heterogeneity between biopsy specimens, we used1 biopsy specimen divided into 4 pieces for each experimentalcondition, thus providing cellular consistency. Each synovialtissue biopsy specimen was sectioned into 4 separate cubes,which were placed in 96-well plates in serum-free RPMI 1640supplemented with penicillin (100 units/ml) and streptomycin(100 units/ml) for 24 hours at 37°C in air with 5% CO2.Synovial explants were then stimulated (in triplicate) for 24hours with OSM (10 ng/ml) (R&D Systems, London, UK),IL-1� (5 ng/ml) (R&D Systems), and the combination of OSM(10 ng/ml) plus IL-1� (5 ng/ml). Following incubation, the wetweights of the biopsy specimens were obtained. The condi-tioned medium was aspirated, collected, and frozen at �80°Cuntil assayed for proMMP-1 and TIMP-1 by enzyme-linkedimmunosorbent assay (ELISA; R&D Systems).
Cell proliferation. A bromodeoxyuridine (BrdU) col-orimetric ELISA kit (catalog no. 11647229001; Roche, India-napolis, IN) was used to assess RASF and HDMEC prolifer-ation in response to OSM and IL-1�, alone and incombination. Briefly, RASFs and HDMECs were cultured andtrypsinized, and 3,000 cells per well were seeded in 48-wellplates in duplicate. Cells were grown in 5–10% FCS andallowed to adhere overnight. Cells were stimulated for 4 dayswith OSM (10 ng/ml), IL-1� (5 ng/ml), and OSM (10 ng/ml)plus IL-1� (5 ng/ml). The BrdU assay was carried out accord-ing to the manufacturer’s instructions. The proliferation rate
ROLE OF OSM IN RA 3153

was based on the measurement of BrdU incorporation duringDNA synthesis. The extent of cell proliferation was measuredby absorbance at 450 nm.
HDMEC tubule formation in vitro. Matrigel (BectonDickinson, Mountain View, CA) basement membrane matrixwas used to examine HDMEC tubule formation. Matrigel (50�l) was plated in 48-well culture plate slides after thawing onice and allowed to polymerize for 1 hour, at 37°C in humidifiedair with 5% CO2. HDMECs were removed from culture,trypsinized, and resuspended at a concentration of 4 � 104
cells/ml in full-strength EGM. Five hundred microliters of cellsuspension was added to each chamber in the presence ofOSM (10 ng/ml), IL-1� (5 ng/ml), and OSM (10 ng/ml) plusIL-1� (5 ng/ml), in duplicate. The chambers were then incu-bated for 24 hours at 37°C in humidified air with 5% CO2.Culture medium was aspirated off the Matrigel surface, andcells were fixed in methanol. Endothelial cell tubule formationwas assessed using phase-contrast microscopy and then photo-graphed. A connecting branch between 2 discrete endothelialcells was counted as 1 tubule and required a consistentintensity and thickness, as previously described (33). Thetubule analysis was determined from 5 sequential fields (40�magnification), focusing on the surface of the Matrigel.
Flow cytometric analysis of ICAM-1 and VCAM-1expression on HDMECs and RASFs. HDMECs and RASFs(cell count 5 � 104) were plated in 12-well plates (Falcon,Oxford, UK) and allowed to grow to confluence. HDMECsand RASFs were incubated in serum-reduced EBM (1% FCS)or serum-free RPMI for 24 hours prior to stimulation for 3–48hours with OSM (10 ng/ml), IL-1� (5 ng/ml) (Sigma, Poole,UK), and OSM (10 ng/ml) plus IL-1� (5 ng/ml). Cells werethen harvested with a cell scraper and transferred tofluorescence-activated cell sorting tubes (Becton Dickinson).Cells were washed and incubated with an optimal concentra-tion of phycoerythrin-conjugated monoclonal mouse anti–ICAM-1, anti–VCAM-1, and isotype-matched control (BectonDickinson) for 30 minutes at 4°C. Samples were then washedtwice with 1% phosphate buffered saline (PBS), and fixed in1% paraformaldehyde. Samples were assayed using a FACS-can flow cytometer (Becton Dickinson), using Lysis II soft-ware.
HDMEC migration. A quantitative InnoCyte Cell Mi-gration assay (Oncogene Research Products, Cambridge, MA)was used to assess endothelial cell migration in response tostimulation with OSM and IL-1�, alone and in combination.Cell migration was quantitatively assessed by staining (with afluorescent dye) those cells that migrated through the 8-�mpores and attached to the lower side of the membrane, asoutlined in the manufacturer’s instructions. Briefly, endothelialcells were grown to confluence and trypsinized. The cellnumber was adjusted to 500,000 cells/ml in EBM medium(Clonetics) containing 1% FCS, in the presence of OSM (10ng/ml) or IL-1� (5 ng/ml), alone or in combination, in dupli-cate. Following 24 hours of incubation at 37°C in air with 5%CO2, cell migration was quantitatively assessed by staining(with fluorescent dye) those cells that migrated through the8-�m pores and attached to the lower side of the Transwellmembrane. The extent of cell migration was measured byfluorescence at excitation 485 nm and emission 520 nm.
RASF and cartilage cocultures. RASFs were isolatedand cultured as described above. Normal human articular
cartilage was obtained from patients undergoing surgery fortraumatic fracture of the femoral neck, each of whom had nohistory or radiologic evidence of arthritis. Cartilage explantcultures were prepared using 3-mm punch biopsy specimens,thus ensuring that only full-depth cartilage biopsy sampleswere used in these experiments. Approximately 12,000 RASFswere cocultured with cartilage explants in 96-well plates inserum-free RPMI 1640 supplemented with 10% FCS, penicil-lin (100 units/ml), and streptomycin (100 units/ml), for 4 weeksin the presence of OSM (10 ng/ml) and IL-1� (5 ng/ml), aloneand in combination.
Culture supernatants were harvested every 5 days, andwells were replenished with fresh medium that was identical tothe medium used on day 1. Following 4 weeks of culture,supernatants were pooled and stored at �80°C for analysis ofproMMP-1, proMMP-13, TIMP-1, VEGF, angiopoietin 2(Ang-2), and glycosaminoglycan (GAG). Cartilage explantswere weighed and paraffin embedded for immunohistologicanalysis. Experiments were performed in duplicate. MMP andTIMP data are expressed as picograms per milliliter, in orderto compare cartilage and RASF culture models. There was nosignificant difference in cartilage weight between the 4 condi-tions, and when cultures were corrected for cartilage weight,similar significant increases in the responses to cytokine stim-ulation were demonstrated.
In further experiments, cartilage explants were cul-tured with either RASFs or with stimulated RASF–conditioned media from the same primary culture in thepresence of OSM (10 ng/ml) and IL-1� (5 ng/ml), alone and incombination. Culture supernatants were assessed forproMMP-1 to examine whether the further increases in theresponses to cytokine stimulation in cocultures were mediatedby soluble mediators or cell–cell/matrix interactions.
Histologic examination of human cartilage explants.Following 4 weeks of culture, human cartilage explants wereremoved and fixed overnight in 7% formaldehyde in PBS (pH7.4), decalcified in 10% EDTA in PBS for 10 days, andembedded in wax. Five-micrometer sections were stained withhematoxylin and eosin and examined microscopically. Foranalysis of proteoglycans, 5-�m sections were stained withSafranin O–fast green and counterstained with hematoxylin(18,34).
ProMMP-1, proMMP-13, TIMP-1, VEGF, Ang-2, andGAG quantification. Levels of proMMP-1, proMMP-13,TIMP-1, VEGF, and Ang-2 were assessed by specific ELISA(R&D Systems). The content and release of proteoglycan inconditioned culture media were determined as sulfated GAG,which is primarily a measure of proteoglycan aggrecan content,using a modification of the colorimetric dimethylmethyleneblue dye-binding assay (35).
Statistical analysis. Statistical analysis was performedusing SPSS 11 for Windows (SPSS, Chicago, IL). Nonparamet-ric and Student’s t-tests were used. P values less than 0.05 wereconsidered significant.
RESULTS
Induction of RASF and HDMEC proliferation byOSM and IL-1�. RASF and HDMEC proliferationincreased significantly in response to stimulation with
3154 FEARON ET AL

OSM alone and IL-1� alone, as demonstrated in Table1. The combination of OSM plus IL-1� produced afurther significant increase in proliferation comparedwith basal levels (Table 1).
Effect of OSM on ICAM-1 and VCAM-1 expres-sion on HDMECs and RASFs. OSM stimulation sig-nificantly increased the percentage of ICAM-1–positiveHDMECs, from a mean � SEM of 20.5 � 0.43% atbaseline to 38.4 � 8.0% (P � 0.05); stimulation withIL-1� increased the percentage to 46 � 5.03% (P �0.05) (Figure 1A). The combination of OSM and IL-1�
further increased the percentage of ICAM-1–positivecells to 63 � 3.7% (P � 0.05) (Figure 1A), demonstrat-ing an additive effect. Although OSM stimulation signif-icantly increased the number of ICAM-1–positive cells,no change in the mean fluorescence intensity (MFI) wasdemonstrated. IL-1� alone significantly increased theICAM-1 MFI, from 103 � 23 (basal) to 346 � 84 (P �0.05); the addition of OSM only slightly potentiated thiseffect (Figure 1B). Stimulation with OSM alone andIL-1� alone significantly increased the percentage ofICAM-1–positive RASFs (P � 0.05), similar to theincrease demonstrated in HDMECs, with the combina-tion of OSM and IL-1� having an additive effect.Although IL-1� stimulation significantly increasedVCAM-1 expression on both HDMECs and RASFs(P � 0.05), OSM, alone or in combination with IL-1�,had no effect on VCAM-1 expression.
OSM-induced angiogenesis in Matrigel in vitro.The induction of HDMEC tubule formation on Matrigelmatrices plated in 48-well culture plates was assessedfollowing stimulation by OSM (10 ng/ml) and IL-1� (5ng/ml), alone and in combination, compared with con-trol vehicle (PBS). Tube-like structures formed after 24hours of incubation with both OSM and IL-1�, whiletubule formation following stimulation with control ve-hicle was minimal (Figures 2A and B). Endothelial celltubule formation increased from a mean � SEM of 18 �2.2 at baseline to 45.8 � 5.1 in the presence of OSM (10ng/ml) (P � 0.05) and to 29.8 � 1.92 (P � 0.05) in thepresence of IL-1�. The combination of OSM and IL-1�
Table 1. Proliferation of RASFs and HDMECs in response to IL-1�(5 ng/ml) and OSM (10 ng/ml), alone and in combination, at 24 and 96hours*
24 hours 96 hours
RASFsBasal 0.1168 � 0.0032 0.2882 � 0.0119†IL-1� 0.1352 � 0.0046 0.5738 � 0.0901‡OSM 0.1345 � 0.0061 0.4472 � 0.0483‡OSM � IL-1� 0.1343 � 0.0067 0.6893 � 0.0117‡
HDMECsBasal 0.1412 � 0.004 0.3763 � 0.0106†IL-1� 0.1457 � 0.0048 0.5057 � 0.0323‡OSM 0.1403 � 0.0074 0.4597 � 0.0210‡OSM � IL-1� 0.3085 � 0.1050 0.5002 � 0.0220‡
* Values are the mean � SEM results from 6 experiments. Theproliferation rate was based on measurement of bromodeoxyuridineincorporation during DNA synthesis. The extent of cell proliferationwas measured by absorbance at 450 nm. RASFs � rheumatoid arthritissynovial fibroblasts; HDMECs � human dermal microvascular endo-thelial cells; IL-1� � interleukin-�; OSM � oncostatin M.† P � 0.05 versus 24 hours.‡ P � 0.05 versus basal and versus 24 hours.
Figure 1. Oncostatin M (OSM)– and interleukin-1� (IL-1�)–induced up-regulation of intercellular adhesionmolecule 1 (ICAM-1) on microvascular endothelial cells. Human dermal microvascular endothelial cells(HDMECs) were stimulated with OSM (10 ng/ml) and IL-1� (5 ng/ml), alone and in combination. The expressionof ICAM-1 was detected by flow cytometric analysis. A, ICAM-1 expression following incubation with OSM andIL-1�, alone and in combination. Values are the mean and SEM results from 4 experiments. � � P � 0.05 versusbaseline (Bas). B, Representative flow cytometry histogram demonstrating induced ICAM-1 expression onHDMECs following stimulation with OSM and IL-1�, alone and in combination. Pe � phycoerythrin.
ROLE OF OSM IN RA 3155
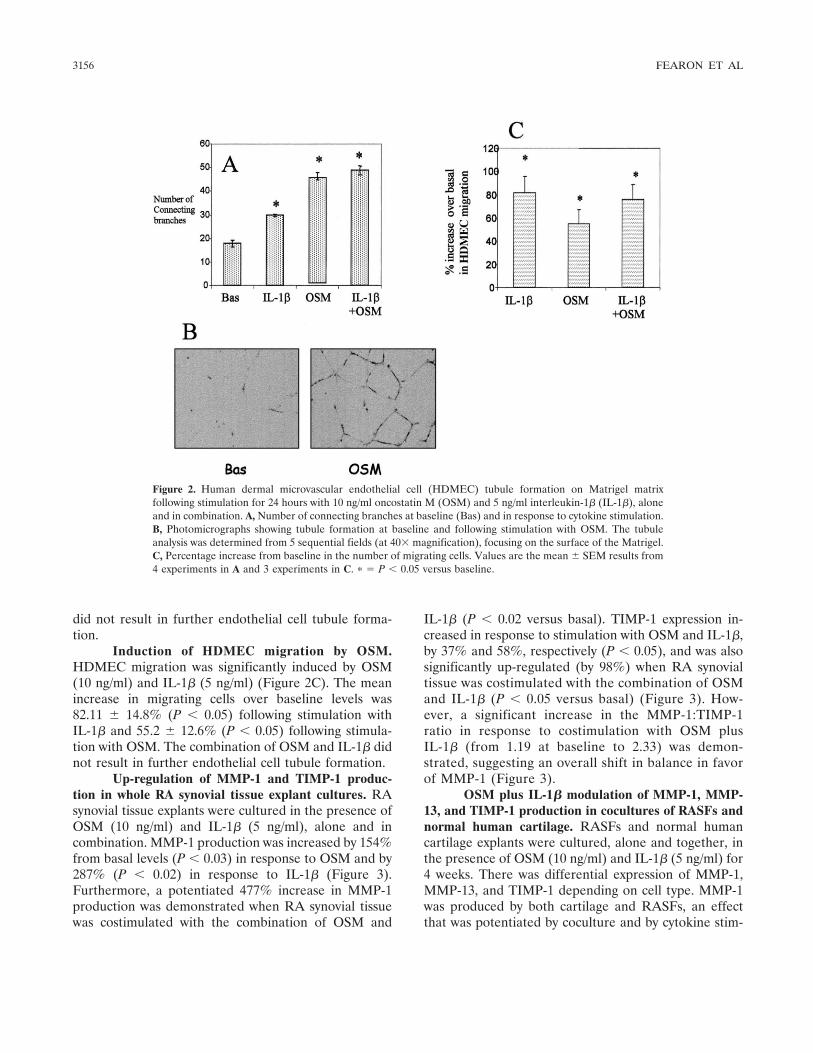
did not result in further endothelial cell tubule forma-tion.
Induction of HDMEC migration by OSM.HDMEC migration was significantly induced by OSM(10 ng/ml) and IL-1� (5 ng/ml) (Figure 2C). The meanincrease in migrating cells over baseline levels was82.11 � 14.8% (P � 0.05) following stimulation withIL-1� and 55.2 � 12.6% (P � 0.05) following stimula-tion with OSM. The combination of OSM and IL-1� didnot result in further endothelial cell tubule formation.
Up-regulation of MMP-1 and TIMP-1 produc-tion in whole RA synovial tissue explant cultures. RAsynovial tissue explants were cultured in the presence ofOSM (10 ng/ml) and IL-1� (5 ng/ml), alone and incombination. MMP-1 production was increased by 154%from basal levels (P � 0.03) in response to OSM and by287% (P � 0.02) in response to IL-1� (Figure 3).Furthermore, a potentiated 477% increase in MMP-1production was demonstrated when RA synovial tissuewas costimulated with the combination of OSM and
IL-1� (P � 0.02 versus basal). TIMP-1 expression in-creased in response to stimulation with OSM and IL-1�,by 37% and 58%, respectively (P � 0.05), and was alsosignificantly up-regulated (by 98%) when RA synovialtissue was costimulated with the combination of OSMand IL-1� (P � 0.05 versus basal) (Figure 3). How-ever, a significant increase in the MMP-1:TIMP-1ratio in response to costimulation with OSM plusIL-1� (from 1.19 at baseline to 2.33) was demon-strated, suggesting an overall shift in balance in favorof MMP-1 (Figure 3).
OSM plus IL-1� modulation of MMP-1, MMP-13, and TIMP-1 production in cocultures of RASFs andnormal human cartilage. RASFs and normal humancartilage explants were cultured, alone and together, inthe presence of OSM (10 ng/ml) and IL-1� (5 ng/ml) for4 weeks. There was differential expression of MMP-1,MMP-13, and TIMP-1 depending on cell type. MMP-1was produced by both cartilage and RASFs, an effectthat was potentiated by coculture and by cytokine stim-
Figure 2. Human dermal microvascular endothelial cell (HDMEC) tubule formation on Matrigel matrixfollowing stimulation for 24 hours with 10 ng/ml oncostatin M (OSM) and 5 ng/ml interleukin-1� (IL-1�), aloneand in combination. A, Number of connecting branches at baseline (Bas) and in response to cytokine stimulation.B, Photomicrographs showing tubule formation at baseline and following stimulation with OSM. The tubuleanalysis was determined from 5 sequential fields (at 40� magnification), focusing on the surface of the Matrigel.C, Percentage increase from baseline in the number of migrating cells. Values are the mean � SEM results from4 experiments in A and 3 experiments in C. � � P � 0.05 versus baseline.
3156 FEARON ET AL

Figure 4. OSM and IL-1� in combination significantly shift the MMP-1:TIMP-1 and MMP-13:TIMP-1 ratios in rheumatoidarthritis synovial fibroblast (RASF)/human cartilage explant cocultures. RASFs and human cartilage explants were culturedalone and together for 4 weeks in the presence of OSM (10 ng/ml) and IL-1� (5 ng/ml), alone and in combination, and culturemedia were analyzed for the expression of MMP-1, MMP-13, and TIMP-1 by enzyme-linked immunosorbent assay (A–C).Values are the mean and SEM results from 5 experiments. � � P � 0.05 versus baseline; �� � P � 0.05 versus IL-1� alone;§ � P � 0.05, coculture versus RASFs cultured alone and cartilage cultured alone; §§ � P � 0.05 versus OSM stimulation incartilage cultured alone; # � P � 0.05 versus either cytokine alone; ## � P � 0.05 versus OSM induction in cocultures. Dand E, MMP-1:TIMP-1 and MMP-13:TIMP-1 ratios in cartilage cocultures. � � P � 0.05 versus baseline and versus eithercytokine alone. Values are the mean and SEM results from 5 experiments. See Figure 3 for other definitions.
Figure 3. Oncostatin M (OSM) and interleukin-1� (IL-1�) in combination significantly shift the ratio of matrixmetalloproteinase 1 (MMP-1) to tissue inhibitor of metalloproteinases 1 (TIMP-1) in whole rheumatoid arthritis(RA) synovial explant cultures. Whole RA synovial tissue explants were serum-starved for 24 hours and incubatedwith OSM (10 ng/ml) and IL-1� (5 ng/ml), alone and in combination. Levels of MMP-1 and TIMP-1 in culturemedia were measured by enzyme-linked immunosorbent assay. Values are the mean and SEM results from 7experiments. � � P � 0.05 versus baseline (Bas).
ROLE OF OSM IN RA 3157
ulation (Figure 4A). MMP-13 was primarily produced bycartilage, and the effect was potentiated by coculturewith RASFs and cytokine stimulation (Figure 4B). Incontrast, TIMP-1 expression was produced by bothcartilage and RASFs at baseline; in response to cytokinestimulation, however, the expression of TIMP-1 wasreduced in cartilage but not in RASFs (Figure 4C).Furthermore, TIMP-1 production was not further po-tentiated in cytokine-stimulated cocultures. This demon-strates differential effects of cytokines on MMP-1 andTIMP-1 production in different cell culture models.
At baseline, MMP-1 production was increased incocultures of RASFs and normal human cartilage, com-pared with either RASFs or cartilage cultured alone. Incocultures, OSM alone had a greater effect on MMP-1and MMP-13 production compared with IL-1� (Figures4A and B). IL-1� potentiated the effect of OSM for bothMMP-1 and MMP-13 in the cocultures (P � 0.05).TIMP-1 expression was also up-regulated in response toOSM in cartilage alone, from a baseline level of 148 �32 ng/ml to 312 � 47 ng/ml (P � 0.05) (Figure 4C).However, when cartilage was incubated with the combi-
nation of OSM and IL-1�, TIMP-1 expression wasactually reduced, to 99.8 � 18.2 ng/ml. TIMP-1 expres-sion was up-regulated in RASFs in response to stimula-tion with OSM and IL-1�, alone and in combination(P � 0.05) (Figure 4C), an effect that was not potenti-ated in the RASF/cartilage cocultures. This resulted in adramatic shift in the balance of the MMP:TIMP-1 ratio,with a synergistic 3-fold increase in the MMP-1:TIMP-1ratio, from 0.50 at baseline to 1.4 (P � 0.05), and a 4-foldincrease in the MMP-13:TIMP-1 ratio, from 51.9 atbaseline to 226 (P � 0.05), in the cocultures in responseto the combination of OSM and IL-1�.
In further experiments, human cartilage was in-cubated with RASFs or RASF-conditioned media inorder to examine the involvement of soluble mediatorsor cell–cell/matrix interactions in the cocultures. Atbaseline, the mean � SEM level of MMP-1 in cartilagefollowing incubation with conditioned media was 105 �22.5 pg/ml compared with 130 � 31 pg/ml followingincubation with RASFs. Following stimulation with thecombination of IL-1� and OSM, the mean � SEM levelof MMP-1 was 456 � 48 pg/ml in the conditioned media
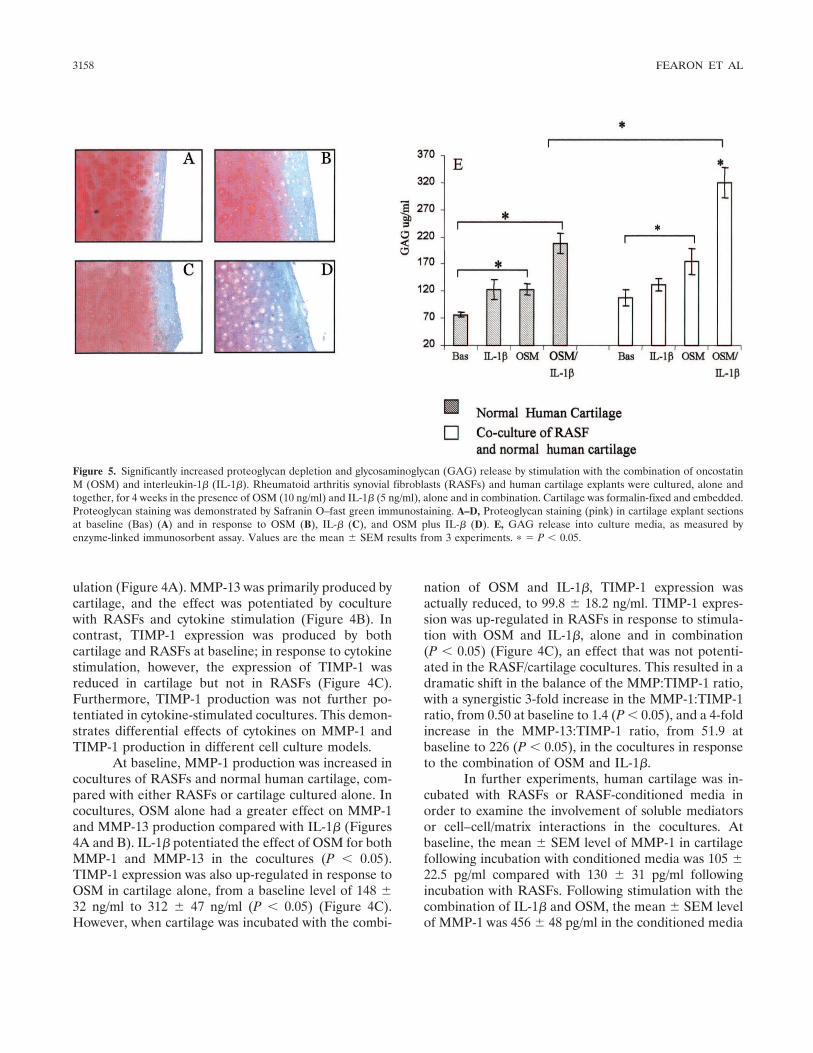
Figure 5. Significantly increased proteoglycan depletion and glycosaminoglycan (GAG) release by stimulation with the combination of oncostatinM (OSM) and interleukin-1� (IL-1�). Rheumatoid arthritis synovial fibroblasts (RASFs) and human cartilage explants were cultured, alone andtogether, for 4 weeks in the presence of OSM (10 ng/ml) and IL-1� (5 ng/ml), alone and in combination. Cartilage was formalin-fixed and embedded.Proteoglycan staining was demonstrated by Safranin O–fast green immunostaining. A–D, Proteoglycan staining (pink) in cartilage explant sectionsat baseline (Bas) (A) and in response to OSM (B), IL-� (C), and OSM plus IL-� (D). E, GAG release into culture media, as measured byenzyme-linked immunosorbent assay. Values are the mean � SEM results from 3 experiments. � � P � 0.05.
3158 FEARON ET AL

and 524 � 38.5 pg/ml in the RASF cocultures. Thissuggests that soluble mediators and, to a lesser extent,cell–cell contact are involved.
Finally, we examined the effects of IL-1� andOSM, alone and in combination, on VEGF and Ang-2production in cartilage, RASFs, and cocultures. IL-1�alone and OSM alone had minimal effect on VEGF incocultures, but the combination of OSM and IL-1�significantly increased VEGF production, from a basallevel of 316 � 62 pg/ml to 505 � 118 pg/ml (P � 0.05).No significant effect was observed for VEGF productionin either cartilage or RASF cultured alone. Neithercytokine, alone or in combination, had an effect onAng-2 expression in any culture model.
Effect of OSM and IL-1� on proteoglycan releasein cartilage explant cultures. At baseline, strong Safra-nin O staining was observed in cartilage explant sections,demonstrating minimal loss of proteoglycan (Figure5A). Cocultures incubated with IL-1� alone or OSMalone demonstrated a mild reduction in Safranin Ostaining (Figures 5B and C). However, when cocultureswere incubated with OSM in combination with IL-1�,almost complete depletion of proteoglycan was demon-strated in cartilage sections (Figure 5D).
Effect of OSM and IL-1� on GAG release fromRASF/cartilage explant cocultures. Minimal release ofGAG was detected in RASFs. In human cartilage cul-tured alone, a significant increase in GAG release wasdemonstrated in response to stimulation with OSMalone and IL-1� alone, with a potentiated response tothe combination of OSM and IL-1� (P � 0.05) (Figure5E). In RASF/cartilage cocultures, GAG release wasalso up-regulated in response to OSM alone and IL-1�alone; however, this effect was not significantly differentfrom that in cartilage alone. A significant increase inGAG release into the supernatants in response tostimulation with the combination of OSM and IL-1� wasobserved in the cocultures compared with cartilagealone (P � 0.05).
DISCUSSION
TNF� and IL-1� are central to the inflammatoryprocess in RA, increasing production of MMP andlinking inflammation to cartilage destruction. RA syno-vial tissue consists of a variety of cell types, the cell–cellinteractions of which contribute to this proinflammatoryresponse. In this study, we used an ex vivo RA synovialtissue culture and an RASF/normal cartilage coculturemodel to investigate the functional interaction of syno-vial cells and the cytokines OSM and IL-1�. We dem-
onstrated that OSM alone induced proliferation,ICAM-1 expression, endothelial cell migration, and tu-bule formation, thereby promoting endothelial cell mi-gration and angiogenesis.
Furthermore, in RA synovial tissue explant cul-tures ex vivo and RASF/human cartilage cocultures, thecombination of OSM and IL-1� promoted production ofMMP-1 and MMP-13, resulting in a dramatic shift in theMMP:TIMP-1 ratio in favor of MMP. This was paral-leled by OSM/IL-1�–induced proteoglycan depletionand GAG release in the RASF/normal human cartilagecoculture model. Finally, we demonstrated that thecombination of OSM and IL-1� significantly increasedVEGF production in the coculture model. DifferentialOSM and IL-1� interactions in the different culturemodel systems highlight the complex cytokine regulationof inflammation observed in vivo in the joint. These datasuggest that the cytokine OSM is pivotal in driving theproinflammatory response in the joint by promotingangiogenesis and cell activation and potentiating theeffect of IL-1� in promoting ECM turnover and degra-dation in human cartilage.
Angiogenesis is one of the earliest events in RA,facilitating leukocyte extravasation, synovial prolifera-tion, and pannus formation (3,4). Recruitment of in-flammatory cells into the joint is mediated by adhesionmolecule/ligand pairs and chemokines (3,4,32). In thisstudy, OSM induced significant endothelial cell prolifer-ation and migration and dramatically increased tubuleformation, suggesting a key role of OSM in these earlyevents of the synovial proinflammatory response. Theangiogenic response to OSM in this study was greaterthan that seen with IL-1�, and the combination of OSMand IL-1� produced little additive effect. This findingsupports previous studies using bovine endothelial cellsor smooth muscle cells that demonstrated the proangio-genic potential of OSM by inducing VEGF, bovine FGF,and chemokine expression (15,36,37). Vasse et al (37)demonstrated that the effect of OSM on bovine endo-thelial cell proliferation was greater than that of VEGF,FGF, leukemia inhibitory factor, and IL-6, while Faffe etal (36) demonstrated that OSM-induced VEGF produc-tion by smooth muscle cells was greater than that ofIL-1�. The use of anti-VEGF neutralizing antibodiesonly partially blocked OSM-induced endothelial cellactivation, suggesting that VEGF-independent mecha-nisms are also involved.
Leukocytes are recruited to the sites of inflam-mation through a series of interactions with activatedendothelium. Mediating this cascade is expression ofadhesion molecules such as ICAM and VCAM on the
ROLE OF OSM IN RA 3159

activated endothelial cells involved in monocyte, lym-phocyte, and neutrophil recruitment. We demonstratedOSM-induced ICAM-1 expression on endothelial cells,suggesting a potential mechanism whereby OSM pro-motes cell migration. This effect, combined with effectson angiogenesis, migration, chemokine (17,36), andP-selectin expression (38), suggests that OSM promotesleukocyte emigration into the perivascular space in RA.Cross et al (39) demonstrated rapid release of OSMfrom activated neutrophils, suggesting one mechanismwhereby OSM could induce endothelial cell activationand cell migration in an autocrine manner. This isconsistent with previous reports that cytokines releasedfrom activated T lymphocytes can induce phenotypicchange in normal endothelial cells (40). Furthermore, invivo studies have shown that OSM induces leukocyteinfiltration into joints of goats following intraarticularinjection of the cytokine (24). Interestingly, OSM had noeffect on VCAM-1 expression in our study; this may beexplained by differential cytokine signaling to homespecific leukocytes into the joint (41).
OSM-induced RASF proliferation and activationsuggest a role in lining layer activation and hyperplasiain the joint. Previous studies demonstrated that CD11a-positive T cells accumulate around ICAM-1–positivecells in the synovial lining layer, and ICAM-1 canmodulate the function of natural killer cells (42,43).Furthermore, increased ICAM-1 expression on RASFsmay potentiate interactions with T cells and, in turn,induce cytokine gene transcription (41–44). This2-directional cross-talk among adhesion molecules andcytokines is critically important for the initiation of T cellemigration into the joint and for T cell and fibroblastactivation, thus sustaining both the proinflammatory andinvasive processes.
In normal tissue/cartilage, a balance exists be-tween active MMP and TIMP (45). In the setting ofinflammation, however, this balance becomes disturbed,leading to an increase in the level of active MMPcompared with that of TIMP, an effect previously ob-served in RA synovial tissue and fluid (46). Using an exvivo RA synovial tissue explant culture model, we dem-onstrated that OSM, alone and in combination withIL-1�, increased MMP-1 production. This is consistentwith previous findings in other cell types, includingRASFs and bovine and porcine chondrocytes (16–19).The RA synovial tissue explant model provides furtherevidence that OSM potentiates the effect of othercytokines in promoting matrix turnover and furthersupports the relevance of this in the RA joint. AlthoughOSM and IL-1�, alone and in combination, also in-
creased TIMP-1 expression, the MMP-1:TIMP-1 ratioshifted significantly in favor of MMP-1. This supportschondrocyte culture and murine model studies, whichdemonstrated a reduction in the level of TIMP-1 inresponse to OSM combined with IL-1� or TNF�(18,25). Some studies have suggested that OSM isantiinflammatory through its ability to promote TIMP-1production (20,21); however, the effect of the combina-tion of OSM and IL-1� on TIMP-1 production may betransient, compared with a more sustained effect onMMP-1 (22). The model used in the current studyreflects the RA joint environment more closely anddemonstrates high MMP-1 and low TIMP-1 levels pro-duced by RA synovial tissue, which is consistent withprevious reports (46).
Increased production of MMP-1 and MMP-13and a reduction in the level of TIMP-1 in response tostimulation with the combination of OSM and IL-1�have been reported in bovine, porcine, and humancartilage and in chondrocyte monocultures (16,25). Be-cause RASFs are central to cartilage destruction in RA,we examined RASF/cartilage cocultures. We demon-strated a dramatic shift in the production of MMP toTIMP, both of which are key enzymes involved in theprimary cleavage and denaturation of type II collagen incartilage. In chondrocyte monoculture and in vivo mu-rine experiments, TIMP-1 production increased in re-sponse to OSM, while the combination of OSM andIL-1� or TNF� led to reduced TIMP-1 production(18,21,24).
We observed similar results in our monocultureexperiments; however, when cartilage was coculturedwith RASFs, the level of TIMP-1 did not significantlychange, in contrast to the levels of MMP-1 and MMP-13,which were markedly increased. This resulted in asignificant increase in the excess of MMP over TIMP, aneffect that may involve interaction of soluble mediators.The differential effects of OSM, alone and in combina-tion with IL-1�, on TIMP production in single-cell orcoculture systems may account for conflicting results inprevious studies. We also demonstrated that the combi-nation of OSM and IL-1� significantly promoted pro-teoglycan loss and GAG release. RA is characterized byprogressive loss of cartilage matrix mediated by cyto-kines such as IL-1� and TNF� (47,48). The mechanismsinvolved in cytokine-induced proteoglycan release arecomplex; however, it is likely mediated through induc-tion of both aggrecanase activity and MMP (48,49).
Finally, we demonstrated that the combination ofIL-1� and OSM significantly increased the production ofVEGF in cocultures. VEGF acts at an early point in the
3160 FEARON ET AL

time sequence of vascular morphogenic events and isconsidered one of the main “on switches” of the angio-genic cascade. It promotes endothelial cell activation,proliferation, and survival, which in turn enables acti-vated monocytes to enter the site of inflammation byactive recruitment involving interactions between adhe-sion molecules and their ligands (3,4). This furthersupports a role for the involvement of OSM in drivingthe proangiogenic response, both directly and indirectly.The increased levels of VEGF secreted from coculturesin response to IL-1� plus OSM may exacerbate theproinflammatory response by activating endothelialcells, promoting angiogenesis, and ultimately facilitatingthe vascular synovial pannus to advance and invadecartilage.
In conclusion, we have shown that OSM pro-motes angiogenesis and cell activation and migration,which suggests that OSM is a key cytokine and possibletherapeutic target in RA (50). Furthermore, we haveshown for the first time in multicell culture systems,which more closely reflect the in vivo environment, thatOSM potentiates ECM turnover, human cartilage de-gradation, and the induction of MMP by IL-1�.
REFERENCES
1. Arend WP, Dayer JM. Cytokines and cytokine inhibitors orantagonists in rheumatoid arthritis. Arthritis Rheum 1990;33:305–15.
2. Feldmann M, Brennan FM, Chantry D, Haworth C, Turner M,Abney E, et al. Cytokine production in the rheumatoid joint:implications for treatment [review]. Ann Rheum Dis 1990;49:480–6.
3. Szekanecz Z, Koch AE. Endothelial cells in inflammation andangiogenesis [review]. Curr Drug Targets Inflamm Allergy 2005;4:319–23.
4. Szekanecz Z, Kim J, Koch AE. Chemokines and chemokinereceptors in rheumatoid arthritis [review]. Semin Immunol 2003;1:15–21.
5. Sacre SM, Andreakos E, Taylor P, Feldmann M, Foxwell BM.Molecular therapeutic targets in rheumatoid arthritis. Expert RevMol Med 2005;16:1–20.
6. Poole AR, Alini M, Hollander AP. Cellular biology of cartilagedegradation. In: Henderson B, Edwards JC, Pettigher ER, editors.Mechanisms and models in rheumatoid arthritis. London: Aca-demic Press; 1995. p. 163–204.
7. Mauviel A. Cytokine regulation of metalloproteinase gene expres-sion [review]. J Cell Biochem 1993;53:288—95.
8. Gearing DP, Comeau MR, Friend DJ, Gimpel SD, Thut CJ,McGourty J, et al. The IL-6 signal transducer, gp130: an oncosta-tin M receptor and affinity converter for the LIF receptor. Science1992;255:1434–7.
9. Stahl N, Yancopoulos GD. The alphas, betas, and kinases ofcytokine receptor complexes [review]. Cell 1993,74:587–90.
10. Okamoto H, Yamamura M, Morita Y, Harada S, Makino H, OtaZ. The synovial expression and serum levels of interleukin-6,interleukin-11, leukemia inhibitory factor, and oncostatin M inrheumatoid arthritis. Arthritis Rheum 1997;40:1096–105.
11. Hui W, Bell M, Carroll G. Detection of oncostatin M in synovial
fluid from patients with rheumatoid arthritis. Ann Rheum Dis1997;56:184–7.
12. Manicourt DH, Poilvache P, van Egeren A, Devogelaer JP, LenzME, Thonar EJ. Synovial fluid levels of tumor necrosis factor �and oncostatin M correlate with levels of markers of the degrada-tion of crosslinked collagen and cartilage aggrecan in rheumatoidarthritis but not in osteoarthritis. Arthritis Rheum 2000;43:281–8.
13. Richards CD, Brown TJ, Shoyab M, Baumann H, Gauldie J.Recombinant oncostatin M stimulates the production of acutephase proteins in HepG2 cells and rat hepatocytes in vitro.J Immunol 1992;148:1731–6.
14. Cawston TE, Curry VA, Summers CA, Clark IM, Riley GP, LifePF, et al. The role of oncostatin M in animal and humanconnective tissue collagen turnover and its localization within therheumatoid joint. Arthritis Rheum 1998;41:1760–71.
15. Wijelath ES, Carlsen B, Cole T, Chen J, Kothari S, Hammond WP.Oncostatin M induces basic fibroblast growth factor expression inendothelial cells and promotes endothelial cell proliferation, mi-gration and spindle morphology. J Cell Sci 1997;110:871–9.
16. Cawston TE, Ellis AJ, Humm G, Lean E, Ward D, Curry V.Interleukin-1 and oncostatin M in combination promote therelease of collagen fragments from bovine nasal cartilage inculture. Biochem Biophys Res Comm 1995;215:377–85.
17. Langdon C, Leith J, Smith F, Richards CD. Oncostatin Mstimulates monocyte chemoattractant protein-1– and interleukin-1–induced matrix metalloproteinase-1 production by human syno-vial fibroblasts in vitro. Arthritis Rheum 1997;40:2139–46.
18. Hui W, Rowan AD, Richards CD, Cawston TE. Oncostatin M incombination with tumor necrosis factor � induces cartilage dam-age and matrix metalloproteinase expression in vitro and in vivo.Arthritis Rheum 2003;48:3404–18.
19. Li WQ, Dehnade F, Zafarullah M. Oncostatin M-induced matrixmetalloproteinase and tissue inhibitor of metalloproteinase-3genes expression in chondrocytes requires Janus kinase/STATsignaling pathway. J Immunol 2001;166:3491–8.
20. Nemoto O, Yamada H, Mukaida M, Shimmei M. Stimulation ofTIMP-1 production by oncostatin M in human articular cartilage.Arthritis Rheum 1996;39:560–6.
21. Richards CD, Shoyab M, Brown TJ, Gauldie J. Selective regula-tion of metalloproteinase inhibitor (TIMP-1) by oncostatin M infibroblasts in culture. J Immunol 1993;150:5596–603.
22. Rowan AD, Koshy PJ, Shingleton WD, Degnan BA, Heath J,Vernallis AB, et al. Synergistic effects of glycoprotein 130 bindingcytokines in combination with interleukin-1 on cartilage collagenbreakdown. Arthritis Rheum 2001;44:1620–32.
23. Koshy PJ, Henderson N, Logan C, Life PF, Cawston TE, RowanAD. Interleukin 17 induces cartilage collagen breakdown: novelsynergistic effects in combination with proinflammatory cytokines.Ann Rheum Dis 2002;61:704–13.
24. Bell MC, Carroll GJ, Chapman HM, Mills JN, Hui W. OncostatinM induces leukocyte infiltration and cartilage proteoglycan degra-dation in vivo in goat joints. Arthritis Rheum 1999;42:2543–51.
25. Rowan AD, Hui W, Cawston TE, Richards CD. Adenoviral genetransfer of interleukin-1 in combination with oncostatin M inducessignificant joint damage in a murine model. Am J Pathol 2003;162:1975–84.
26. Langdon C, Kerr C, Hassen M, Hara T, Arsenault AL, RichardsCD. Murine oncostatin M stimulates mouse synovial fibroblasts invitro and induces inflammation and destruction in mouse joints invivo. Am J Pathol 2000;157:1187–96.
27. Plater-Zyberk C, Buckton J, Thompson S, Spaull J, Zanders E,Papworth J, et al. Amelioration of arthritis in two murine modelsusing antibodies to oncostatin M. Arthritis Rheum 2001;44:2697–702.
28. Richards CD, Langdon C, Botelho F, Brown TJ, Agro A. Oncosta-tin M inhibits IL-1- induced expression of IL-8 and granulocyte-
ROLE OF OSM IN RA 3161

macrophage colony-stimulating factor by synovial and lung fibro-blasts. J Immunol 1996;156:343–9.
29. Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF,Cooper NS, et al. The American Rheumatism Association 1987revised criteria for the classification of rheumatoid arthritis.Arthritis Rheum 1988;31:315–24.
30. Veale DJ. The role of arthroscopy in early arthritis [review]. ClinExp Rheumatol 1999;17:37–8.
31. Fraser A, Fearon U, Reece R, Emery P, Veale DJ. Matrixmetalloproteinase 9, apoptosis, and vascular morphology in earlyarthritis. Arthritis Rheum 2001;44:2024–8.
32. McEvoy AN, Bresnihan B, FitzGerald O, Murphy EP. Cyclooxy-genase 2–derived prostaglandin E2 production by corticotropin-releasing hormone contributes to the activated cAMP responseelement binding protein content in rheumatoid arthritis synovialtissue. Arthritis Rheum 2004;50:1132–45.
33. Park CC, Morel JC, Amin MA, Connors MA, Harlow LA, KochAE. Evidence of IL-18 as a novel angiogenic marker. J Immunol2001;167:1644–53.
34. Van Meurs J, van Lent P, Stoop R, Holthuysen A, Singer I, BayneE, et al. Cleavage of aggrecan at the Asn341–Phe342 site coincideswith the initiation of collagen damage in murine antigen-inducedarthritis: a pivotal role for stromelysin 1 in matrix metalloprotein-ase activity. Arthritis Rheum 1999;42:2074–84.
35. Farndale RW, Buttle DJ, Barrett AJ. Improved quantitation anddiscrimination of sulphated glycosaminoglycans by use of dimeth-ylmethylene blue. Biochim Biophys Acta 1986;883:173–7.
36. Faffe DS, Flynt L, Mellema M, Whitehead TR, Bourgeois K,Panettieri RA, et al. Oncostatin M causes VEGF release fromhuman airway smooth muscle: synergy with IL-�. Am J PhysiolLung Cell Mol Physiol 2005;288:1040–8.
37. Vasse M, Pourtau J, Trochon V, Muraine M, Vannier JP, Lu H, etal. Oncostatin M induces angiogenesis in vitro and in vivo.Arterioscler Thromb Vasc Biol 1999;19:1835–42.
38. Yao L, Pan J, Setiadi H, Patel KD, McEver RP. Interleukin 4 oroncostatin M induces a prolonged increase in P-selectin mRNAand protein in human endothelial cells. J Exp Med 1996;184:81–92.
39. Cross A, Edwards SW, Bucknall RC, Moots RJ. Secretion ofoncostatin M by neutrophils in rheumatoid arthritis. ArthritisRheum 2004;50:1430–6.
40. Fiorelli V, Gendelman R, Samaniego F, Markham AD, Ensoli BJ.Cytokines from activated T cells induce normal endothelial cells to
acquire the phenotypic and functional features of AIDS-Kaposi’ssarcoma spindle cells. J Clin Invest 1995;95:723–34.
41. Nakatsuka K, Tanaka Y, Hubscher S, Abe M, Wake A, Saito K, etal. Rheumatoid synovial fibroblasts are stimulated by the cellularadhesion to T cells through lymphocyte function associated antigen-1/intercellular adhesion molecule-1. J Rheumatol 1997;24:458–64.
42. Cho DH, Song HK, Kang HS, Yoon SR, Lee HG, Pyun KH, et al.Ligation of ICAM-1 molecules inhibits target cell-induced granuleexocytosis of IL-12-activated natural killer cells. Cell Immunol2000;199:1–7.
43. Tanaka Y. The role of chemokines and adhesion molecules in thepathogenesis of rheumatoid arthritis. Drugs Today (Barc) 2001;37:477–84.
44. Hanyuda M, Kasama T, Isozaki T, Matsunawa MM, Yajima N,Uchida H, et al. Activated leucocytes express and secrete macro-phage inflammatory protein-1� upon interaction with synovialfibroblasts of rheumatoid arthritis via a �2-integrin/ICAM-1 mech-anism. Rheumatology (Oxford) 2003;42:1390–7.
45. Visse R, Nagase H. Matrix metalloproteinases and tissue inhibi-tors of metalloproteinases: structure, function, and biochemistry[review]. Cir Res 2003;92:5:827–39.
46. Ishiguro N, Ito T, Oguchi T, Kojima T, Iwata H, Ionescu M, et al.Relationships of matrix metalloproteinases and their inhibitors tocartilage proteoglycan and collagen turnover and inflammation asrevealed by analyses of synovial fluids from patients with rheuma-toid arthritis. Arthritis Rheum 2001;44:2503–11.
47. Fraser A, Fearon U, Billinghurst RC, Ionescu M, Reece R,Barwick T, et al. Turnover of type II collagen and aggrecan incartilage matrix at the onset of inflammatory arthritis in humans:relationship to mediators of systemic and local inflammation.Arthritis Rheum 2003;48:3085–95.
48. Kobayashi M, Squires GR, Mousa A, Tanzer M, Zukor DJ,Antoniou J, et al. Role of interleukin-1 and tumor necrosis factor� in matrix degradation of human osteoarthritic cartilage. ArthritisRheum 2005;52:128–35.
49. Hui W, Barksby HE, Young DA, Cawston TE, McKie N, RowanAD. Oncostatin M in combination with tumor necrosis factor �induces a chondrocyte membrane associated aggrecanase that isdistinct from ADAMTS aggrecanase-1 or -2. Ann Rheum Dis2005:64:1624–32.
50. Pelletier JP, Martel-Pelletier J. Oncostatin M: foe or friend?[editorial]. Arthritis Rheum 2003:48:3301–3.
3162 FEARON ET AL





























