Embed Size (px)
Citation preview

Article No. jaut.1998.0268, available online at http://www.idealibrary.com on I It I: ~ 1 ®
JAi
Journal of Autoimmunity (1999) 13, 121-127
Natural Anti-endothelial Cell Antibodies (AECA)
Nicoletta Ronda 1, Silvia LeonardP, Guido Orlandini 2, Rita Gatti 2, Stefano Bellosta 3, Franco Bernini 4 and Alberico BorghettP
1Dipartimento di Clinica Medica, Nefrologia e Scienze della Prevenzione, Universita degli Studi di Parma, Parma, Italy 21stituto di Istologia, Universita degfi Studi di Parma, Parma, Italy 31stituto di Scienze Farmacologiche, Universita degli Studi di Milano, Milano, Italy 41stituto di Farmaco/ogia e Farmacognosia, Universita degli Studi di Parma, Parma, /ta/y
Key words: autoantibodies, endocytosis, endothelial cell, function, immunoglobulin G
Natural AECA constitute a pool of autoantibodies circulating in healthy subjects, which react with a restricted and conserved set of endothelial antigens and establish idiotypic interactions within the immunoglobulin networks. Normal IgG interacts with living endothelial cells and is internal- ized with a mechanism involving microtubules and resembling that of ligand-receptor internalization. IgG-endothelial cell interaction appears to be dependent on the variable region of antibodies and is followed by modifica- tions of endothelial cell function. Natural AECA increase anti-inflammatory properties of endothelial cells through the selective inhibition of thromboxane A2, endothelin and metalloproteinase-9 secretion, and also through the inhibition of endothelial cell proinflammatory response to TNF-~. We have gathered evidence demonstrating that natural AECA constitute a strictly controlled autoantibody pool, interact with living endothelial cells and take part in the regulation of endothelial function, through direct anti- inflammatory effects. © 1999 Academic Press
Introduction
The concept that physiological autoreactivity, that is controlled self-recognition, is one of the functions of the immune system has been widely accepted since it was proven that autoreactive lymphocytes and anti- bodies are active elements normally expressed in healthy subjects and play a role in maintaining the homeostasis of the system [1]. In fact, besides genetic factors, one of the mechanisms controlling autoreac- tivity is the ability of autoantibodies and autoreactive lymphocytes to establish variable region interactions. The resulting immune networks play an important role regulating the expansion and the final 'net' activity of single clones [2].
Natural autoantibodies towards several antigens have been described, but the occurrence of antibodies physiologically recognizing endothelial cells (EC) has not been previously considered. On the contrary a few reports described the presence of normal IgG in endothelial cells, but this was ascribed to pathological mechanisms such as passive diffusion in irreversibly damaged cells.
Instead, a pathologic humoral response against vascular tissue was suggested more than twenty years ago and since, a long series of reports has confirmed
Correspondence to: Nicoletta Ronda, Dipartimento di Clinica Medica, Nefrologia e Scienze della Prevenzione, V. Gramsci 14, 43100 Parma, Italy. Fax: +39 521 292627. E-mail: climed01 @ ipruniv.cle.u nipr.it
the occurrence and role of pathologic autoantibodies reacting with EC [3]. Thus the expression 'anti- endothelial cell antibodies' (AECA) has referred so far to a family of pathologic antibodies, recognizing various endothelial antigens, detectable in systemic inflammatory and autoimmune disorders.
In this paper we summarize the results of some studies that we have published previously and report original data to provide an overview on natural AECA. We gathered evidence indicating not only that natural AECA do occur, but also that they are strictly controlled in terms of activity and antigen specificity and that they establish V-region interactions with normal immunoglobulins. Natural AECA interact with living endothelial cells, and are internalized with a mechanism that is probably receptor-dependent and involves microtubules. Finally, natural AECA directly modulate endothelial function in vitro, enhancing the anti-inflammatory properties of EC.
121
Material and Methods
Sources of IgG and IgG fragments
We used pooled intravenous immunoglobulin (Wig) (Sandoglobulin, Novartis, Basel, Switzerland) and single healthy donor IgG, purified from the serum by precipitation with saturated ammonium sulphate and chromatography on protein G-Sepharose. F(ab')2 and Fc fragments were prepared by pepsin (2% w / w )
0896-8411/99/050121 + 07 $30.00/0 © 1999 Academic Press

122 N. Ronda etal,
and papain digestion respectively. The purity of IgG and F(ab')2 fragment preparations was verified by SDS-PAGE and the concentration of IgG was measured by single radial immunoprecipitation (Nor-Partigen IgG-HC, Behring S.P.A. Scoppito, L'Aquila, Italy).
Endothelial cell culture
EC were from human umbilical cord veins [4] if not stated otherwise. For particular experiments cells were obtained from human adult saphenous vein [5].
EC were grown on 25 cm 2 culture flasks before lysis for immunoblotting, on glass coverslides for confocal microscopy and on 24-well culture plates for stimu- lation experiments and EC product measurement.
Affinity chromatography
IgG from healthy donors were chromatographed on an affinity column of Sepharose-bound F(ab)2 frag- ments of IVIg, as previously described [6] to separate IgG reacting and non-reacting idiotypically with F(ab')2 fragments of IVIg.
Immunoblotting
Immunoblotting was performed using total endo- thelial protein extract to analyse antigenic reactivity of IgG in the serum of 12 healthy donors and in purified preparations. SDS-PAGE under reducing conditions on 10% acrylamide gels was followed by semi-dry electroblotting of proteins onto nitrocellulose membranes and incubation with the IgG solutions. Binding was revealed using an alkaline phosphatase- labelled anti-human IgG and the appropriate sub- strate. Densitometry and computerized analysis of data [7] allowed an extremely accurate evaluation of patterns of reactivity of antibody pools. Briefly, the light remission densitometric analysis of the blots using a high resolution CCD camera system (MasterScan, Scanalytics, Billerica, MA, USA) was followed by staining of the nitrocellulose membrane with a colloidal gold solution (Protogold, Biocell, Cardiff, UK) and by a second densitometric evaluation of gold stained proteins in the spaces between lanes of immunoblotting. The data acquired were analysed with IGOR software, to superimpose protein peaks in different lanes, to adjust possible migration distortions for each lane and to substract non-specific background reactivity.
Preparation of fluoresceinated IgG (IgG FITC), IgG fragments and insulin
To visualize IgG interactions with living EC, IVIg and single healthy donor IgG were coupled to FITC and purified by chromatography on a Sephadex G25 col- umn, as described [5]. F(ab')2 and Fc fragments of IgG,
monoclonal mieloma IgG and insulin (Sigma Aldrich Srl, Milan, Italy) used in control experiments, were fluoresceinated following the same procedure.
Confocal microscopy
A Molecular Dynamics Multiprobe 2001 equipped with an Argon-ion laser was used to observe the cells through an oil immersion x l00 objective (Nikon PlanApo, NA 1.4) with a confocal aperture (pinhole) of 100pm. Image processing and fluorescence measurements were performed on a Silicon Graphics Personal Iris workstation (Image Space Software, Molecular Dynamics, Sunnyvale, CA, USA).
The glass coverslide on which endothelial cells had been grown was mounted on a flow chamber provid- ing constant standard culture conditions and allowing the substitution of culture medium without changes in the plane of focus along x, y or z axes. Accurate visualization of the cell profile was obtained by counterstaining with I ~tM Calcein-AM after IgG-FITC signal acquisition. Calcein staining is an indicator of cell vitality and membrane integrity because in pres- ence of membrane damage the fluorescence drops immediately to zero even with residual cell estherase activity. Experimental procedures and technical aspects have ,been discussed in detail in previous publications [5, 8].
Evaluation of endothelial cell secretory activity
EC secretory activity was evaluated measuring secretion products in the supernatant of a constant number of confluent cells grown on 24-well plates. Endothelin-1 measurement in the supernatant of IgG treated and control EC was performed by radioimmunoassay [9].
Prostaglandin 6-keto-PGF1, ~ and thromboxane B 2 (TxB2) concentration was measured by radio- immunoassay after acidification of supernatants, supplementation with 3H-6-keto-PGFl~ to assess final recovery, extraction of 6-keto-PGFl~ and T xB 2 with Sep-Pak C 18 cartridges (Waters) and separation by HPLC. To study the mechanism of interference of IgG on EC prostaglandin secretion, EC were incubated with 3H-arachidonic acid, washed and incubated with IgG or control medium. Supernatants were then extracted with Sep-Pak C 18 and fractionated by HPLC to analyse 3H-prostaglandins [9].
Metalloproteinases (MMP) were measured by zymography [10]. Samples underwent electrophoresis at 4°C on 7.5% polyacrilamide gels containing 10% SDS and gelatin (1 mg/ml) under non-reducing con- ditions and without boiling. After electrophoresis, SDS was removed from gels with two washes in 2.5% Triton X-100 at room temperature. The gels were then incubated overnight at 37°C with gentle shaking in Tris 50 mmol/1 pH 7.5 containing NaC1 150 mmol/1, CaC12 10 mmol/1, ZnC12 1 mmol/1 to restore metallo- proteinase activity. For inhibition studies and to confirm the identity of MMPs, identical gels were
Natural anti-endothelial cell antibodies 123
incubated in the above buffer containing either EDTA 20 mmol/1, an inhibitor of MMPs, or PMSF I mmol/1, an inhibitor of serine proteinases. The addition of PMSF did not affect MMP activity, while the treatment with EDTA completely abolished it (data not shown). At the end of the incubation, the gels were stained with a solution of 0.1% Coomassie brilliant blue R-250 in 25% methanol and 7% acetic acid. Clear zones against the blue background indicated the presence of proteinolytic activity. For quantitation of zyrnograms, densitometric scanning was performed using a video camera and a computer analysis package (NIH Image 1.52 image analysis software). Each experiment was performed at least three times with different prep- arations of cells. The background was set for each gel and each lane was analysed sequentially. Results were normalized by cellular protein content and expressed as optical density (OD) units.
Interleukin-6 (IL-6) was measured in EC super- natant with a commercially available ELISA kit, fol- lowing manufacturer's instructions (Amersham Italia, Srl, Milan, Italy). The experimental procedure in this case was as follows: EC were incubated for 24 h with either Wig at 10 m g / m l in culture medium or with culture medium only. After collecting the super- natants, the cells were washed and incubated with TNF-a (Sigma Aldrich Chemical Co., Milan, Italy) (10ng/ml) or LPS (Sigma) (100ng/ml) or control medium for 6 h. Finally, after collecting the super- natants and washing the cells, standard culture medium was added to all wells and again collected after 16 h.
Statistical analysis
Data on the secretory activity of EC were expressed as mean+SD and analysed for statistical significance by the Whitney-Mann U test.
Results
Natural AECA antigen specificity
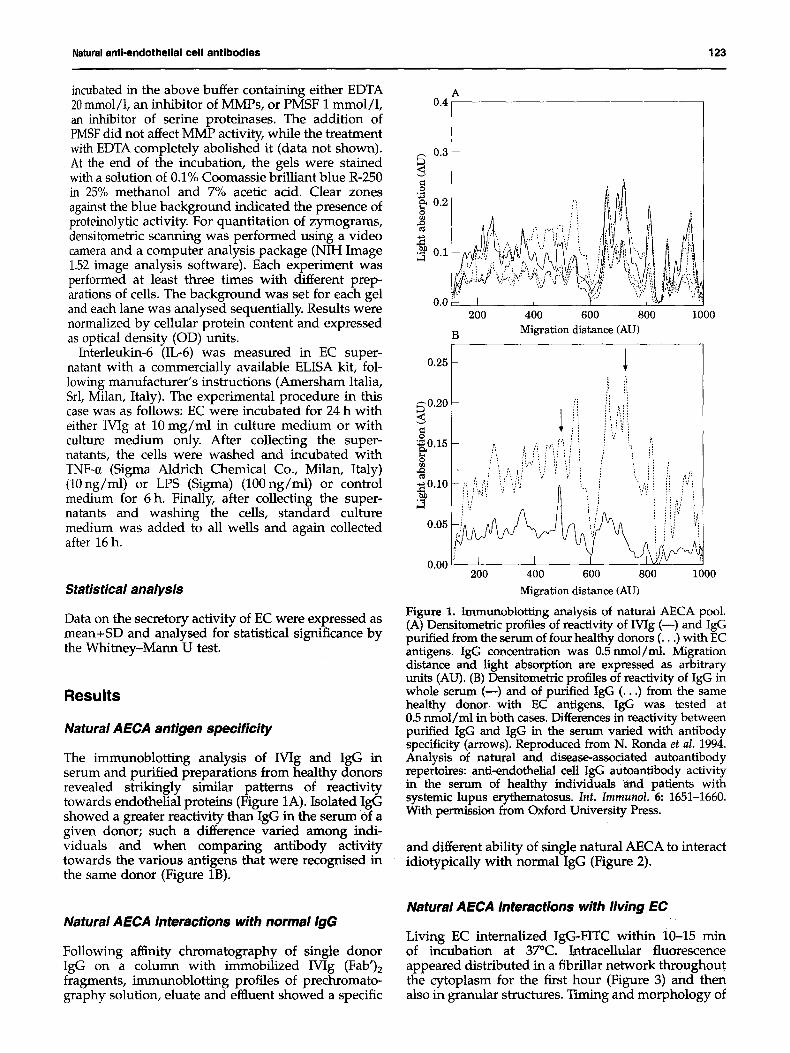
The immunoblotting analysis of Wig and IgG in serum and purified preparations from healthy donors revealed strikingly similar patterns of reactivity towards endothelial proteins (Figure 1A). Isolated IgG showed a greater reactivity than IgG in the serum of a given donor; such a difference varied among indi- viduals and when comparing antibody activity towards the various antigens that were recognised in the same donor (Figure 1B).
Natural AECA interactions with normal IgG
Following affinity chromatography of single donor IgG on a column with immobilized Wig (Fab')2 fragments, immunoblotting profiles of prechromato- graphy solution, eluate and effluent showed a specific
A 0.4 l
0.3
0.2
$5
~0.1
0.0
.,,-,
200 ""~::" i :;;,' ' j ,~
400 600 800 Migration distance (AU)
1000
0.25 -
~ 0,20 - /] ii
-e / 1 0 , 1 5 - ,.,-~ ~ :,'~ ,,.v~! ~i o ,;" i ~i !i ~ i :'
0.05 i ~,/ i ii ~ ',,: ;,: ! i V !
200 400 600 800 1000 Migration distance (AU)
Figure 1. Immunoblotting analysis of natural AECA pool. (A) Densitometric profiles of reactivity of Wig (--) and IgG purified from the serum of four healthy donors (...) with EC antigens. IgG concentration was 0.5 nmol/ml. Migration distance and light absorption are expressed as arbitrary units (AU). (B) Densitometric profiles of reactivity of IgG in whole serum (--) and of purified IgG (...) from the same healthy donor with EC antigens. IgG was tested at 0.5 nmol/ml in both cases. Differences in reactivity between purified IgG and IgG in the serum varied with antibody specificity (arrows). Reproduced from N. Ronda et aL 1994. Analysis of natural and disease-associated autoantibody repertoires: anti-endothelial cell IgG autoantibody activity in the serum of healthy individuals and patients with systemic lupus erythematosus. Int. Immunol. 6: 1651-1660. With permission from Oxford University Press.
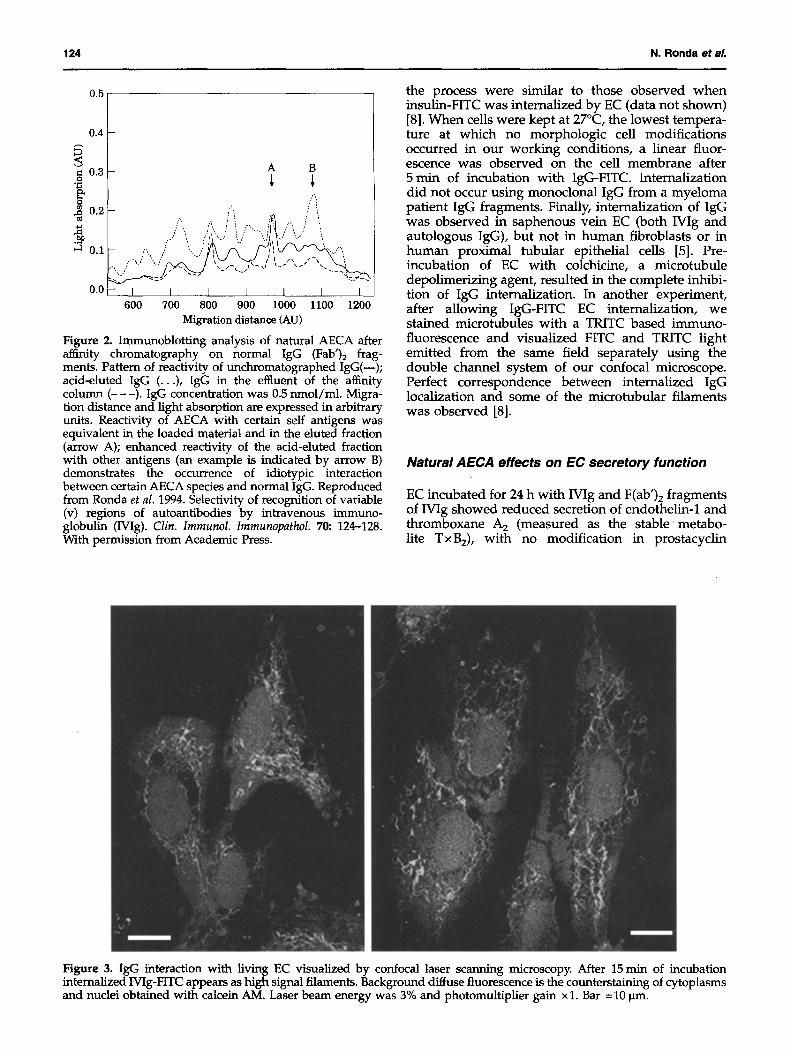
and different ability of single natural AECA to interact idiotypicaUy with normal igG (Figure 2).
Natural AECA interactions with living EC
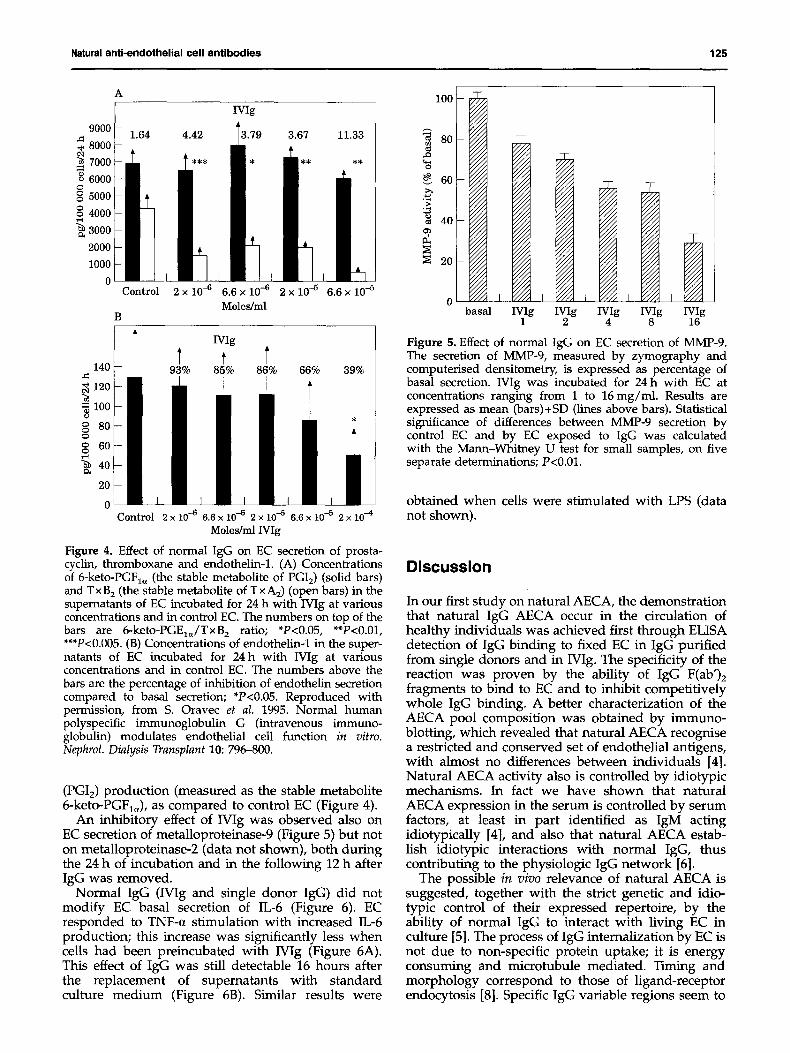
Living EC internalized IgG-FITC within 10-15 rain of incubation at 37°C. ~tracellular fluorescence appeared distributed in a fibrillar network throughout the cytoplasm for the first hour (Figure 3) and then also in granular structures. Timing and morphology of
124 N. Ronda et al.
0.5
0.4
"~ 0.3 A B o
0 f"i
: '. ,:f', ! ~.: "-",~ , / '...:
~ 0 . I " : ", : ' "",'",
0 . 0
600 700 800 900 1000 1100 1200 Migration distance (AU)
Figure 2. Immunoblotting analysis of natural AECA after affinity chromatography on normal IgG (Fab')2 frag- ments. Pattern of reactivity of unchromatographed IgG(--); acid-eluted IgG (...), IgG in the effluent of the affinity column (---) . IgG concentration was 0.5 nmol/ml. Migra- tion distance and light absorption are expressed in arbitrary units. Reactivity of AECA with certain self antigens was equivalent in the loaded material and in the eluted fraction (arrow A); enhanced reactivity of the acid-eluted fraction with other antigens (an example is indicated by arrow B) demonstrates the occurrence of idiotypic interaction between certain AECA species and normal IgG. Reproduced from Ronda e t aL 1994. Selectivity of recognition of variable (v) regions of autoantibodies by intravenous immuno- globulin (Wig). C l i n . I m m u n o l . I m m u n o p a t h o l . 70: 124-128. With permission from Academic Press.
the process were similar to those observed when insulin-FITC was internalized by EC (data not shown) [8]. When cells were kept at 27°C, the lowest tempera- ture at which no morphologic cell modifications occurred in our working conditions, a linear fluor- escence was observed on the cell membrane after 5 min of incubation with IgG-FITC. Internalization did not occur using monoclonal IgG from a myeloma patient IgG fragments. Finally, internalization of IgG was observed in saphenous vein EC (both Wig and autologous IgG), but not in human fibroblasts or in human proximal tubular epithelial cells [5]. Pre- incubation of EC with colchicine, a microtubule depolimerizing agent, resulted in the complete inhibi- tion of IgG internalization. In another experiment, after allowing IgG-FITC EC internalization, we stained microtubules with a TRITC based immuno- fluorescence and visualized FITC and TRITC light emitted from the same field separately using the double channel system of our confocal microscope. Perfect correspondence between internalized IgG localization and some of the microtubular filaments was observed [8].
Natural AECA effects on EC secretory function
EC incubated for 24 h with Wig and F(ab')2 fragments of IVIg showed reduced secretion of endothelin-1 and thromboxane A 2 (measured as the s tablemetabo- lite T xB2), with no modification in prostacyclin
Figure 3. IgG interaction with living EC visualized by confocal laser scanning microscopy. After 15 min of incubation internalized Mg-F1TC appears as high signal filaments. Background diffuse fluorescence is the counterstaining of cytoplasms and nuclei obtained with calcein AM. Laser beam energy was 3% and photomultiplier gain x 1. Bar -- 10 ~m.
Natural anti-endothelial cell antibodies 125
A
9000 8000
~7000 6000
o 5oo0 4000
~3000 2000 i000
0 Control
B
2x10 4 6.6x10 4 2x10 -5 6.6x10 -5 Moles/ml
M g
l t l 93% 85% 86% 140 66% 39%
,v 120
100 g~
ilJ ~ 60
~ 40
20
0 - __k__ ~__ ~ Control 2 x 10 4 6.6 x 10 4 2 x 10 . 5 6 .6 x 10 -5 2 x 10 .-4
Moles/ml IVIg
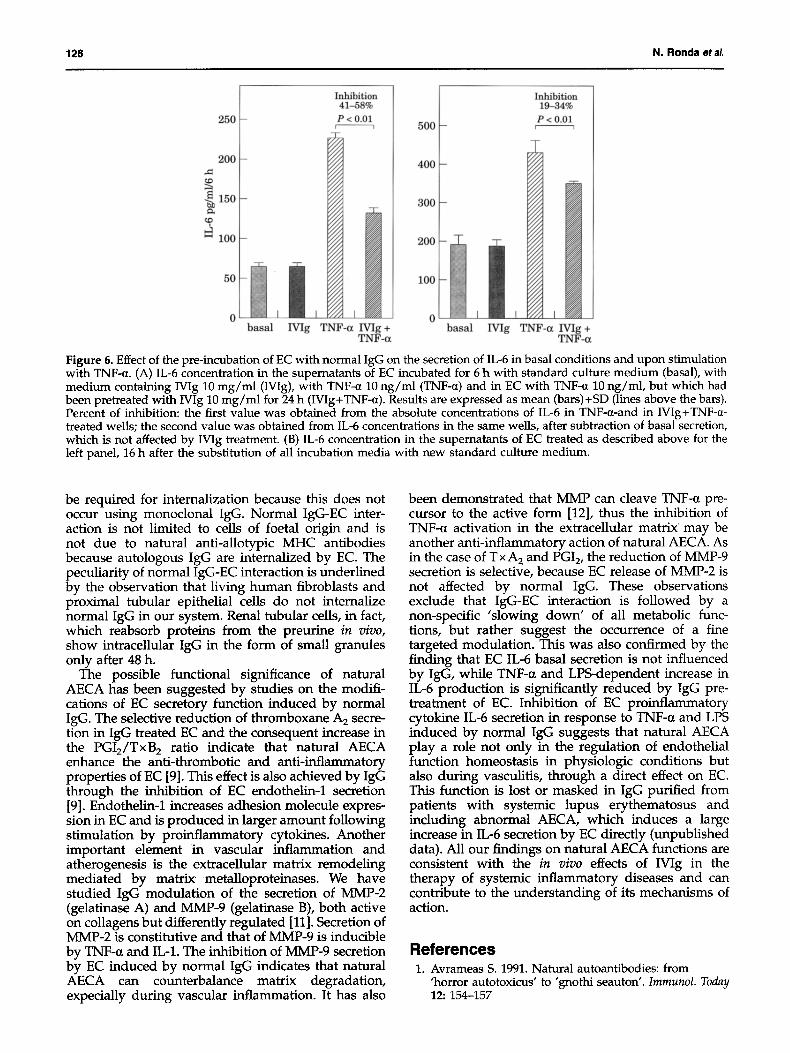
Figure 4. Effect of normal IgG on EC secretion of prosta- cyclin, thromboxane and endothelin-1. (A) Concentrations of 6-keto-PGF1, (the stable metabolite of PGI2) (solid bars) and T x B 2 (the stable metabolite of T x A2) (open bars) in the supematants of EC incubated for 24 h with IVIg at various concentrations and in control EC. The numbers on top of the bars are 6-keto-PGE1dTxB 2 ratio; *P<0.05, **P<0.01, ***P<0.005. (B) Concentrations of endothelin-1 in the super- natants of EC incubated for 24 h with IVIg at various concentrations and in control EC. The numbers above the bars are the percentage of inhibition of endothelin secretion compared to basal secretion; *P<0.05. Reproduced with permission, from S. Oravec et al. 1995. Normal human polyspecific immunoglobulin G (intravenous immuno- globulin) modulates endothelial cell function in vitro. Nephrol. Dialysis Transplant 10: 796-800.
(PGI2) production (measured as the stable metabolite 6-keto-PGFl~ ), as compared to control EC (Figure 4).
An inhibitory effect of IVIg was observed also on EC secretion of metalloproteinase-9 (Figure 5) but not on metalloproteinase-2 (data not shown), both during the 24 h of incubation and in the following 12 h after IgG was removed.
Normal IgG (Wig and single donor IgG) did not modify EC basal secretion of IL-6 (Figure 6). EC responded to TNF-a stimulation with increased IL-6 production; this increase was significantly less when cells had been preincubated with Wig (Figure 6A). This effect of IgG was still detectable 16 hours after the replacement of supernatants with standard culture medium (Figure 6B). Similar results were
,.Q
i °° 40
~ 20
0 ~ basal IVIg M g IVIg IVIg M g
1 2 4 8 16
Figure 5. Effect of normal IgG on EC secretion of MMP-9. The secretion of MMP-9, measured by zymography and computerised densitometry, is expressed as percentage of basal secretion. Wig was incubated for 24 h with EC at concentrations ranging from 1 to 16 mg/ml. Results are expressed as mean (bars)+SD (lines above bars). Statistical significance of differences between MMP-9 secretion by control EC and by EC exposed to IgG was calculated with the Mann-Whitney U test for small samples, on five separate determinations; P<0.01.
obtained when cells were stimulated with LPS (data not shown).
Discussion
In our first study on natural AECA, the demonstration that natural IgG AECA occur in the circulation of healthy individuals was achieved first through ELISA detection of IgG binding to fixed EC in IgG purified from single donors and in IVIg. The specificity of the reaction was proven by the ability of IgG F(ab')2 fragments to bind to EC and to inhibit competitively whole IgG binding. A better characterization of the AECA pool composition was obtained by immuno- blotting, which revealed that natural AECA recognise a restricted and conserved set of endothelial antigens, with almost no differences between individuals [4]. Natural AECA activity also is controlled by idiotypic mechanisms. In fact we have shown that natural AECA expression in the serum is controlled by serum factors, at least in part identified as IgM acting idiotypically [4], and also that natural AECA estab- lish idiotypic interactions with normal IgG, thus contributing to the physiologic IgG network [6].
The possible in vivo relevance of natural AECA is suggested, together with the strict genetic and idio- typic control of their expressed repertoire, by the ability of normal IgG to interact with living EC in culture [5]. The process of IgG internalization by EC is not due to non-specific protein uptake; it is energy consuming and microtubule mediated. Timing and morphology correspond to those of ligand-receptor endocytosis [8]. Specific IgG variable regions seem to
126 N. Ronda etal.
250
2OO
-~ 150 - e~
~D
i 0 0
i o ~ basal
I n h i b i t i o n 41 -58%
P < 0.01
7G, 9/,
>'/, >'/, 9"/,
I
IVIg TNF-a Mg + TNF-a
I n h i b i t i o n 19-34%
P < 0 . 0 1 500 - r - - ,
4 0 0 - Y / / / z l /
~//×
300 -
200
100
0 I I z//~/ I basal IVIg TNF-a Wig +
TNF-a
Figure 6. Effect of the pre-incubation of EC with normal IgG on the secretion of IL-6 in basal conditions and upon stimulation with TNF-a. (A) IL-6 concentration in the supernatants of EC incubated for 6 h with standard culture medium (basal), with medium containing IVIg 10 mg/ml (IVIg), with TNF-R 10 ng/ml (TNF-a) and in EC with TNF-a 10 ng/ml, but which had been pretreated with Wig 10 mg/ml for 24 h (IVIg+TNF-a). Results are expressed as mean (bars)+SD (lines above the bars). Percent of inhibition: the first value was obtained from the absolute concentrations of IL-6 in TNF-a-and in IVIg+TNF-(~- treated wells; the second value was obtained from IL-6 concentrations in the same wells, after subtraction of basal secretion, which is not affected by Wig treatment. (B) IL-6 concentration in the supernatants of EC treated as described above for the left panel, 16 h after the substitution of all incubation media with new standard culture medium.
be required for internalization because this does not occur using monoclonal IgG. Normal IgG-EC inter- action is not limited to cells of foetal origin and is not due to natural anti-allotypic MHC antibodies because autologous IgG are internalized by EC. The peculiarity of normal IgG-EC interaction is underlined by the observation that living human fibroblasts and proximal tubular epithelial cells do not internalize normal IgG in our system. Renal tubular cells, in fact, which reabsorb proteins from the preurine in vivo, show intracellular IgG in the form of small granules only after 48 h.
The possible functional significance of natural AECA has been suggested by studies on the modifi- cations of EC secretory function induced by normal IgG. The selective reduction of thromboxane A2 secre- tion in IgG treated EC and the consequent increase in the PGI2/TxB 2 ratio indicate that natural AECA enhance the anti-thrombotic and anti-inflammatory properties of EC [9]. This effect is also achieved by IgG through the inhibition of EC endothelin-1 secretion [9]. Endothelin-1 increases adhesion molecule expres- sion in EC and is produced in larger amount following stimulation by proinflammatory cytokines. Another important element in vascular inflammation and atherogenesis is the extracellular matrix remodeling mediated by matrix metalloproteinases. We have studied IgG modulation of the secretion of MMP-2 (gelatinase A) and MMP-9 (gelatinase B), both active on collagens but differently regulated [11]. Secretion of MMP-2 is constitutive and that of MMP-9 is inducible by TNF-a and IL-1. The inhibition of MMP-9 secretion by EC induced by normal IgG indicates that natural AECA can counterbalance matrix degradation, expecially during vascular inflammation. It has also
been demonstrated that MMP can cleave TNF-a pre- cursor to the active form [12], thus the inhibition of TNF-a activation in the extracellular matrix' may be another anti-inflammatory action of natural AECA. As in the case of T x A 2 and PGI 2, the reduction of MMP-9 secretion is selective, because EC release of MMP-2 is not affected by normal IgG. These observations exclude that IgG-EC interaction is followed by a non-specific 'slowing down" of all metabolic func- tions, but rather suggest the occurrence of a fine targeted modulation. This was also confirmed by the finding that EC IL-6 basal secretion is not influenced by IgG, while TNF-a and LPS-dependent increase in IL-6 production is significantly reduced by IgG pre- treatment of EC. Inhibition of EC proinflammatory cytokine IL-6 secretion in response to TNF-a and LPS induced by normal IgG suggests that natural AECA play a role not only in the regulation of endothelial function homeostasis in physiologic conditions but also during vasculitis, through a direct effect on EC. This function is lost or masked in IgG purified from patients with systemic lupus erythematosus and including abnormal AECA, which induces a large increase in IL-6 secretion by EC directly (unpublished data). All our findings on natural AECA functions are consistent with the in vivo effects of IVIg in the therapy of systemic inflammatory diseases and can contribute to the understanding of its mechanisms of action.
References 1. Avrameas S. 1991. Natural autoantibodies: from
'horror autotoxicus' to 'gnothi seauton'. Immunol. Today 12:154-157

Natural anti-endothelial cell antibodies 127
2. Varela F., Coutinho A. 1991. Second generation immune networks. Immunol. Today 12: 159- 166
3. Meroni P.L., Del Papa N., Raschi E., Tincani A., Balestrieri G., Youinou P. 1999. Antiendothelial cell antibodies (AECA): from a laboratory curiosity to another useful autoantibody. In The decade of autoimmunity. Y. Shoenfeld, ed. Elsevier, Amsterdam, pp. 285-294
4. Ronda N., Haury M., Nobrega A., Kaveri S.V., Coutinho A., Kazatchkine M.D. 1994. Analysis of natural and disease-associated autoantibody repertories: anti-endothelial cell IgG autoantibody activity in the serum of healthy individuals and patients with systemic lupus erythematosus. Intern. Immunol. 6:1651-1660
5. Ronda N., Gatti R., Orlandini G., Borghetti A. 1997. Binding and internalization of human IgG by living cultured endothelial cells. Clin. Exp. Immunol. 109: 211-216
6. Ronda N., Haury M., Nobrega A., Coutinho A., Kazatchkine M.D. 1994. Selectivity of recognition of variable (V) regions of autoantibodies by intravenous immunoglobulin (IVIg). Clin. Immun. Immunopatol. 70: 124-128
7. Haury M., Grandien A., Sundblad A., Coutinho A., Nobrega A. 1994. Global analysis of antibody repertoires. 1. An immunoblot method for the quantitative screening of a large number of reactivities. Scand. J. Immunol. 39:79-85
8. Orlandini G., Ronda N., Gatti R., Gazzola G.C., Borghetti A. 1999. Receptor-ligand internalization. Methods Enzymol 307:340-350
9. Oravec S., Ronda N., Carayon A., Milliez J., Kazatchkine M.D., Hornych A. 1995. Normal human polyspecific immunoglobulin G (intravenous immunoglobulin) modulates endothelial cell flanction in vitro. Nephrol. Dial. Transplant. 10:796--800
10. Bellosta S., Via D., Canavesi M., Pfister P., Fumagalli R., Bernini F. 1998. HMGCoA reductase inhibitors reduce MMP-9 secretion by macrophages. Arterioscl. Thromb. Vasc. Biol. 18; 1671-1678
11. Dollery C.M., McEwan J.R., Henney A.M. 1995. Matrix metalloproteinases and cardiovascular disease. Cir. Res. 77:863-868
12. Gearing A.J.H., Beckett P., Christodoulou M., Churchill M., Clements J., Davidson A.H., Drummond A.H., Galloway W.A., Gilbert R., Gordon J.L. 1994. Processing of tumor necrosis factor alpha precursor by metalloproteinases. Nature 370:555-557





























