Embed Size (px)
Citation preview

Journal of Cellular Biochemistry 102:925–935 (2007)
ARTICLES
Characterization of Anti-OsteopontinMonoclonal Antibodies: Binding Sensitivity toPost-Translational Modifications
Christian C. Kazanecki,1 Aaron J. Kowalski,2 Tony Ding,1 Susan R. Rittling,3 and David T. Denhardt1*1Department of Cell Biology and Neuroscience, Rutgers University, Piscataway New Jersey2Juvenile Diabetes Research Foundation, New York, New York3The Forsyth Institute, Boston, Massachusetts
Abstract Osteopontin (OPN) is primarily a secreted phosphoglycoprotein found in a variety of tissues and bodyfluids. It has awide rangeof reported functions,manyofwhich are affectedby thedegreeof post-translationalmodification(PTM) of the protein. These PTMs include phosphorylation, glycosylation, and cross-linking by transglutaminase.Herewedescribe the generation of unique monoclonal antibodies raised against recombinant OPN utilizing the OPN knockoutmouse. The antibodies exhibit differential binding to OPN produced by different cell lines from the same species, aswell to themultipleOPN forms in human urine.Most of the antibodies generated are able to recognizeOPN produced byras-transformedmouse fibroblasts, however only one antibody recognizes the more phosphorylated protein produced bythe differentiating pre-osteoblast murine cell line MC3T3E1. Using a novel biopanning procedure combining T7 phagegene fragment display and protein G precipitation, we have epitope-mapped these antibodies. Several of the antibodiesbind to regions of theOPNmolecule that are phosphorylated, and one binds the region ofOPN that is glycosylated. Usingphosphorylated and non-phosphorylated peptides, we show that the binding of two antibodies to the C-terminal end ofOPN is inhibited by phosphorylation of this region. In addition, these two antibodies are able to inhibit cell adhesion torecombinant andweaklymodifiedOPN. The antibodies described hereinmay proveuseful in determining the presence ofmodifications at specific sites and for identifying structural forms of OPN. Also, the sensitivity of these antibodies to PTMssuggests that caution must be taken when choosing anti-OPN monoclonal antibodies to detect this highly modifiedprotein. J. Cell. Biochem. 102: 925–935, 2007. � 2007 Wiley-Liss, Inc.
Key words: gene fragment display; phage display; epitope mapping; T7 phage; phosphorylation; cell adhesion;protein G
The secreted phosphoglycoprotein osteopon-tin (OPN) is often considered to be a cellattachment protein due to its ability to bind toa number of integrin receptors via its RGDdomain or a cryptic SVVYGLR site that isexposed after cleavage of OPN by thrombin.However, OPN can exist as a soluble cytokine aswell as amineralized-matrix associated protein.There is now considerable evidence that OPN isa ligand for certain variants of the hyaluronatereceptor CD44 [Weber et al., 1996], but thisinteraction may require the presence of the b1integrin [Katagiri et al., 1999]. An intracellularform of OPN has also been discovered thatcolocalizeswithCD44 at the inner surface of thecell membrane [Zohar et al., 1997; Zhu et al.,2004]. OPN has been associated with a varietyof functions including cell adhesion, migration,and survival, as well as activation of immunecells, inhibition of nitric oxide production, and
� 2007 Wiley-Liss, Inc.
Abbreviations used: GFD, gene fragment display; OPN,osteopontin; PTM, post-translational modification; mAb,monoclonal antibody; PAGE, polyacrylamide gel electro-phoresis; PVDF, polyvinylidene fluoride; ELISA, enzyme-linked immunosorbent assay; GST, glutathione S-trans-ferase; PBS, phosphate buffered saline.
Christian C. Kazanecki and Aaron J. Kowalski contributedequally to this study.
Grant sponsor: NIH; Grant numbers: #AR 44434, ES06897, #DK 067685; Grant sponsor: Busch BiomedicalResearch Award; Grant sponsor: National MultipleSclerosis Society; Grant sponsor: Rutgers TechnologyCommercialization Fund; Grant sponsor: National ScienceFoundation IGERT; Grant number: #DGE 0333196.
*Correspondence to: David T. Denhardt, Nelson Laborato-ries, 604 Allison Road, Piscataway, NJ 08854.E-mail: [email protected]
Received 14 June 2007; Accepted 18 June 2007
DOI 10.1002/jcb.21487

regulation of crystal formation and growth[reviewed in Denhardt et al., 2001a,b; Sodeket al., 2006]. Interestingly, OPN expression isupregulated in a number of pathological con-ditions, during tumorigenesis and in responseto injury or inflammation.
Post-translational modification (PTM) ofOPN, notably serine phosphorylation, plays animportant role in many of OPN’s functions.Other modifications include glycosylation,sulfation, and sialylation. Many sites of PTMsare conserved across species analyzed; howeverthe degree of modification of the proteinvaries depending on the source tissue, cell line,or differentiation state of the cells used. Forexample, both thebovineandhumanmilk formshave a largenumber of phosphorylated residues(�28 and 32 respectively) that are located inclusters [Sørensen et al., 1995; Christensenet al., 2005]. In contrast, rat bone OPN (afterpurification) contains only 10–11 phosphates[Keykhosravani et al., 2005]. In all cases, thedegree of phosphorylation of any individual siteis variable, so that the overall phosphorylationis extremely heterogeneous.
A great deal of evidence suggests that phos-phorylation of OPN plays a large role in itsphysiological functions. Phosphorylation ofOPN has been demonstrated to be required forits ability to inhibit smooth muscle cell calcifi-cation [Jono et al., 2000], to promote migrationof cancer cells [Al-Shami et al., 2005] as well asadhesion and bone resorption by osteoclasts[Ek-Rylander et al., 1994; Razzouk et al., 2002].Evidence has also suggested that phosphoryla-tion of OPN determines its interactionswith calcium phosphate (hydroxyapatite) oroxalate crystals and subsequent regulation ofcrystal growth and nucleation [Hoyer et al.,2001;Pampenaetal., 2003;Gericke et al., 2005].OPN produced by different sources may havealterations in specific modifications which canaffect function. Crawford et al. [1998] deter-mined that tumor-derived OPN inhibits macro-phage function and increases the survival ofmetastasizing cells, whereas host cell-derivedOPN is a more effective macrophage chemo-attractant. Glycosylation of OPN may also playa role in modulating OPNs function, but muchless is known about this potential mechanism.
Recent articles have described the creation ofanti-OPNmonoclonal antibodies raised againstfull-length protein or specific peptides [Hottaet al., 1999;Konet al., 2000, 2002].However, the
ability of these antibodies to bind OPN frommultiple species (i.e., mouse and human) andtissue sources has yet to be well established.Here we describe the creation and character-ization of novel mouse anti-mouse OPN andmouse anti-human OPNmonoclonal antibodiesemploying OPN-deficient mice. We took advan-tage of the OPN knockoutmice as naı̈ve hosts toproduce antibodies to this highly conservedprotein [Declerck et al., 1995]: this approachgenerated highly specific antibodies that re-cognize conserved regions of the moleculeand hence detect OPN from multiple speciesat functional locations. We developed a noveltechnique employing a protein G immunopreci-pitation procedure with T7 bacteriophage genefragment display (GFD) to rapidly determinethe OPN epitope recognized by the monoclonalantibodies. Some of the described antibodies aresensitive toPTMof their epitopesandareable toinhibit cell adhesion to OPN. These antibodiesmay be useful in determining the degree of PTMof OPN at specific sites.
MATERIALS AND METHODS
Anti-Osteopontin MonoclonalAntibody Production
Antibodies were prepared as described byHarlow and Lane [1999] and D’Alonzo et al.[2002]. Approximately 20 mg of recombinanthistidine-tagged human OPN (prepared inE. coli and purified using hisBind resin (Nova-gen, Madison WI)) or recombinant glutathioneS-transferase (GST)-tagged mouse OPN (pre-pared in E. coli and purified using glutathioneSepharose-4B [GE Healthcare, Piscataway,NJ]) mixed with Freund’s complete adjuvantwas injected subcutaneously into female OPNknockout mice [Rittling et al., 1998]. Two and5 weeks later the mice were boosted with anequal amount of protein in Freund’s incompleteadjuvant. Three weeks later (week 8) the micewere given a final boost 3 days prior to sacrifi-ce and splenectomy. Spleen cells were fused toSP2/0 cells and fused cells were selected withhypoxanthine–aminopterin–thymidinemedium. Positive clones were identified byenzyme-linked immunosorbent assay (ELISA)and Western blotting.
Antibodies were purified from hybridomacell culture medium that was collected afterallowing the cells to grow until completesaturation in flasks (approximately 2 weeks).
926 Kazanecki et al.

The cells were pelleted and the medium filteredthrough a 0.45 mm filter (Millipore 0.45 mmWhiteHAWP, 26mm).Antibodieswere purifiedusing Immobilized Protein G (Pierce Biotech,Inc., Rockford, IL).
Human OPN Gene Fragment Library
A human OPN gene fragment library wasconstructed employing the Novagen T7SelectPhage Display system according to the manu-facturer’s instructions (Novagen). Briefly, anOPN plasmid which encodes the full lengthhuman OPNmolecule (OPN1b/Harpo4) [Younget al., 1990; Rollo, 1995] was digested withDNase I. DNA fragments between approxi-mately 50–150 bp were ligated into theT7Select415-1b vector using EcoRI adapters.In vitro packaging reactions were performed asdescribed in the Novagen T7Select SystemManual. PCR was performed using the primersT7SelectUP (50-GGAGCTGTCGTATTCCAGT-C-30) and T7Select Down (50-AACCCCTCAA-GACCCGTTTA-30), which flank the T7Select-1b multicloning site. Positive clones weredefined as those with PCR products >30 bplarger than products from an empty vector. Theconcentration of unique phage in the T7 humanOPN library was determined to be 8� 105.
Biopanning and Epitope Determination
For epitope determination, the NovagenT7Select protocol was followed, and a modifiedbiopanning protocol was developed (see Fig. 1).Approximately 1012 phage were pre-incubatedat room temperature in hypoxanthine–thymi-dinemediumwith 10%ProteinG-agarose beads(Pierce Biotech) to remove non-specific bindingphage. Antibody-containing supernatants weresimultaneously mixed at room temperature for30minwith a concentration of 10% (v/v) ProteinG beads. Both mixtures were then centrifugedat �3,000g. The phage-containing supernatantwas added to the precipitated antibody-proteinGpellet and incubatedwithmixing for 1 h.Afterincubation and multiple phosphate bufferedsaline (PBS) washes, the mixture was centri-fugedaspreviously and thepellet addeddirectlyto log phase BL21 E. coli. The bacteria wereimmediately added to 3 ml of molten top-agarose and plated. Positive plaques wereidentified by incubating plaque lifts withthe desired antibody. Positive plaques weredispersed in 10 mM EDTA, pH 8.0 and heatedfor 10 min at 658C to disrupt the phage. The
mixture was then clarified by centrifugation at14,000g and used for PCR amplification of theinsertion regionwith theT7SelectUpandDownprimers. PCR products were sequenced andOPN amino acid sequences were determinedand aligned.
Cell Culture
MC3T3E1 subclone four cells (kind giftfrom Dr. R. Franceschi, University of Michigan)were maintained in a-MEM (Invitrogen Corp.,Carlsbad, CA) with 10% FBS (Hyclone, Logan,UT), 5 mg/ml penicillin, 5U/ml streptomycin, and2 mM glutamine. For differentiation, cells weregrown until confluent then switched to growthmedium above containing 100 mg/ml ascorbicacid and 10 mM b-glycerophosphate (Sigma-Aldrich, St. Louis, MO) for an additional 10–12 days before generating conditioned medium.Ras-transformed fibroblasts (275-3-2), or the
Fig. 1. Cartoon of novel biopanning protocol for antibodyepitope determination using T7 phage and protein G beads.ProteinGagarosebeadsareused topre-clear theT7phage libraryof non-specific binding phage (left). In a separate reaction,protein G beads are used to bind the antibody being assayed(right). The pre-cleared phage and the antibody-bead complexesare then incubated together to allow antibody-phage binding.The resulting complexes arewashed, addeddirectly to E. coli andplated for plaque formation. Positive plaques are identified byWestern blotting, and the region containing the OPN insert isamplified by PCR and sequenced to determine the peptideexpressed. [Color figure can be viewed in the online issue,whichis available at www.interscience.wiley.com.]
Anti-Osteopontin Monoclonal Antibodies 927

parental non-transformed cells 3T3-275 [Wuet al., 2000], were maintained in DMEM (Medi-atech, Inc., Herndon, VA) with 10% FBS(Hyclone), 5 mg/ml penicillin, 5 U/ml strepto-mycin, and 2 mM glutamine. Conditionedmediumwas generated from these confluent cellcultures by overnight incubation with serum-free medium.
Western Blotting
Freshly collected conditioned medium wasused for Western blotting of OPN producedby cell lines. Typically 10–20 ml/lane of con-ditioned medium was fractionated by SDS–polyacrylamide gel electrophoresis (PAGE)with 12% gels. For purified proteins, equalamounts (typically 50 ng) in each lane wereused. Protein was transferred to polyvinylidenefluoride (PVDF) membranes (Millipore, Bill-erica, MA), which were cut into strips andblotted with 1 mg/ml of purified monoclonalantibodies. Antibodies were purified fromhybridoma-conditioned medium using proteinG-agarose beads following the manufacturer’sinstructions (Pierce Biotech). Human urinewascollected and dialyzed extensively against 0.1MNaCl, then concentrated approximately tenfoldwith Centriprep spin columns (Millipore). Theequivalent of 50 ml of urine/lane was assayed bySDS–PAGE as above.
Peptide Affinity Assay
Biotin-tagged OPN peptides (kind gift fromDr. Lawrence Steinman, Stanford University)were added to Neutra-Avidin coated 96-wellplates (Pierce Biotech) at 10 mg/ml. Anti-OPNmonoclonal antibodies were added at 5 mg/mland detected with a Alexafluor 594-conjugatedanti-mouse IgG (Invitrogen) at 2 mg/ml. Fluo-rescence was detected using excitation/emission wavelengths of 584/612 nm with aFluoroskan-Ascent fluorometer (ThermoFisherScientific, Inc., Waltam, MA).
Cell Adhesion Assay
Flat-bottom 96-well tissue culture-treatedpolystyrene microtiter plates (Corning, NY)were coated with 100 ml recombinant his-tagged human OPN [Rollo, 1995] (5 mg/ml) orfibronectin (2.5 mg/ml) in PBS at 48C overnightand blocked with 1% BSA. MDA-MB-435 or275-3-2 cells were trypsinized, washed, andresuspended in Dulbecco’s modified Eagle’smedium containing 1 mg/ml BSA. Cells
(5� 104) were added to coatedwells and allowedto adhere for 3 or 3.5 h. Non-adhered cells wereremoved as described by Goodwin and Pauli[1995] with slight modifications. Cells werewashed twice by pipetting 75ml Percoll washsolution (73% Percoll [Sigma-Aldrich], 0.9%NaCl) slowly down the sides of the wells andadherent cellswerefixedbyadding50ml fixative(10% glutaraldehyde in Percoll) in the samemanner. The wash and fixative solutions werethen washed from the wells with 2–3 washes of100 ml PBS. Fixed cells were stained with 100 ml0.1% crystal violet (25 min), washed withtap water and solubilized in 50 ml 0.5% TritonX-100 at least 1 h before reading at 570 nm in aMRX Revelation Reader (Thermo Labsystems).
RESULTS
Creation of Monoclonal AntibodyProducing Hybridomas
OPN-deficient mice created in our laboratorymounted a strong immune response after immu-nization with recombinant OPN. Monoclonalantibody (mAb)-producing hybridoma cell linescreated from multiple fusions were screenedfor their ability to bind OPN via ELISA. Over1,000 clones were screened and 20þ lines werepositive in our initial ELISA screen. Seven ofthese anti-OPN monoclonal antibodies will bedetailed here andwere selected based upon theirperformance in various immunoassays and theirbinding locations.
Epitope Determination
We used a GFD strategy to determine theepitopes of OPN bound by the antibodieswhich were positive in the initial screening[Kowalski, 2005]. We chose to employ theNovagen T7Select415 vector due to the robustnature of the T7 phage and high copy number(415) of the gene fragment on the phage surface.A humanOPN expression plasmidwasDNAse1digested and fragments of 50–150 bps werecloned into the T7Select415 vector. A library ofapproximately 8� 105 clones was created andamplified.
Initially, the standard manufacturer’s bio-panning protocol (Novagen) in which the anti-bodies were coated onto microtiter plates andthen subjected to library panning, elution, andamplification of bound phage was employed.This method was successful for three of themonoclonal antibodies chosen for further study.
928 Kazanecki et al.

Since the highest affinity binders are themost difficult to elute, and to minimize back-ground non-specific phage binding, wemodifiedthe biopanning protocol. In the modifiedprotocol, selection for phage displaying anti-body epitopes is performed in solution utilizingantibody-protein G agarose beads to isolatebound phage. These precipitated complexescan then be mixed directly with the hostbacteria, and plated to form plaques (Fig. 1),thusavoiding the elution step.Weobserved overa 100-fold increase in positive binding phageand were able to epitope map antibodies thatresulted in no positive clones utilizing theimmobilized antibody methodology.In total,wehavemapped the epitopes of seven
anti-OPNmonoclonal antibodies. Interestingly,five of the seven antibodies mapped to thecarboxy terminal half of the OPN molecule,and one antibody (AK1G4)mapped to the signalsequence (Fig. 2A). The antibodies AK1H3 andAK1G4 only recognize human OPN, while the
other antibodies are able to bind both humanandmurineOPN.Figure2Bshows the results ofthe phage screening assay for antibody AK2A1.The screening yielded multiple peptides thatwere aligned to determine the minimal epitopein this case, PVA. Two of the antibodies, AK3D9and AK7B4 recognized the same region inthe extreme carboxy terminus of the molecule.The antibody AK10F6 did not yield multipleoverlapping peptides from the T7 phage libraryscreening, but yielded the same peptidemultiple times.
Antibody Binding to Osteopontin
As the OPN-knockout mice were originallyimmunized with recombinant OPN, we hypo-thesized that the presence of PTMs on nativeOPN may prevent binding of some of theantibodies. The ras-transformed fibroblast cellline (275-3-2) produces abundant amounts ofOPN (Fig. 3A), and this protein was recognizedby all the antibodies tested (AK1G4, which
Fig. 2. Localization of monoclonal anti-OPN antibody epitopes. A: Alignment of the mouse and humanOPN sequences showing the determined epitopes of our monoclonal antibodies. Antibody recognition sitesare underlined with the antibody name below. Post-translational modifications of human OPN are takenfrom Christensen et al. [2005]. Phosphorylated residues are highlighted in gray. Glycosylated residues arewritten in gray. B: Peptides resulting from phage display screening demonstrating the determination of theepitope for antibody AK2A1.
Anti-Osteopontin Monoclonal Antibodies 929
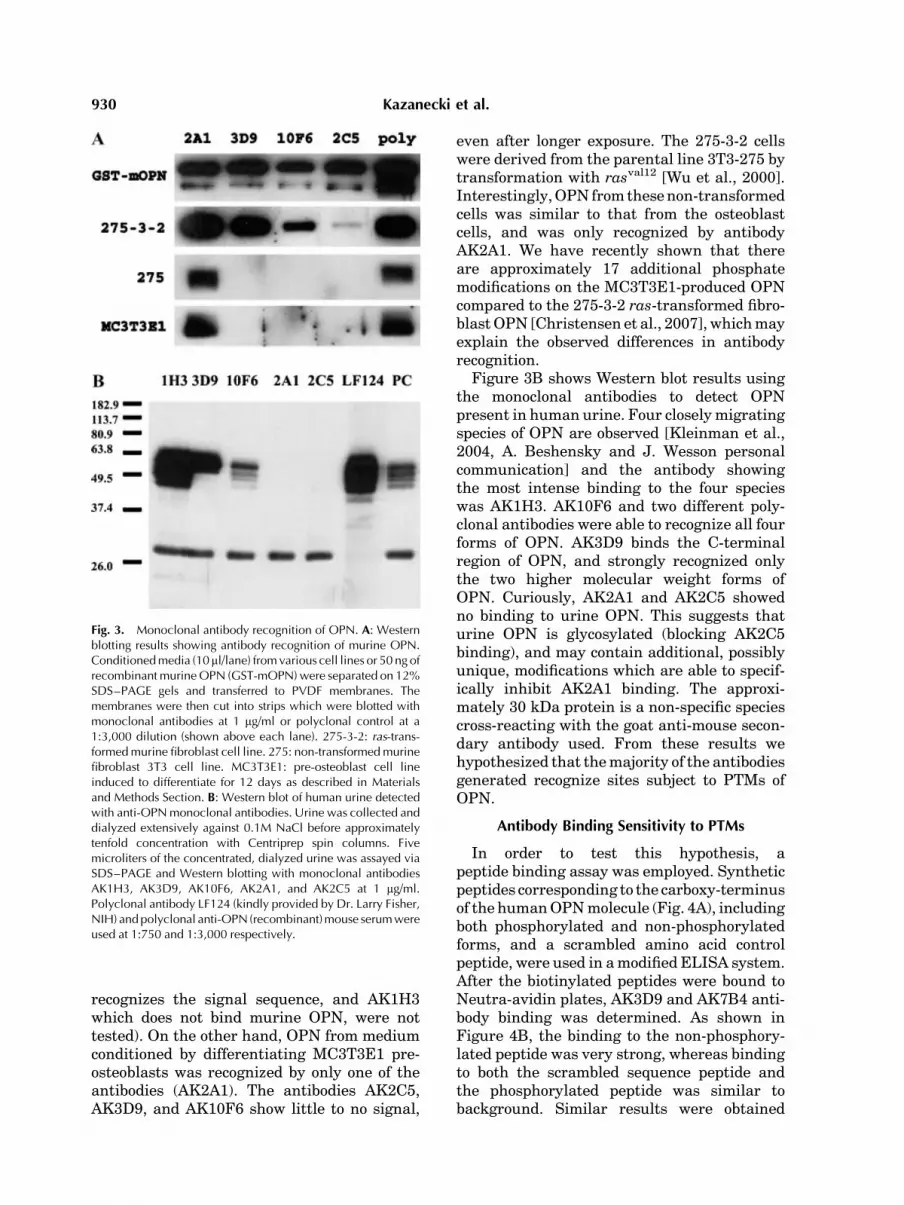
recognizes the signal sequence, and AK1H3which does not bind murine OPN, were nottested). On the other hand, OPN from mediumconditioned by differentiating MC3T3E1 pre-osteoblasts was recognized by only one of theantibodies (AK2A1). The antibodies AK2C5,AK3D9, and AK10F6 show little to no signal,
even after longer exposure. The 275-3-2 cellswere derived from the parental line 3T3-275 bytransformation with rasval12 [Wu et al., 2000].Interestingly,OPN from thesenon-transformedcells was similar to that from the osteoblastcells, and was only recognized by antibodyAK2A1. We have recently shown that thereare approximately 17 additional phosphatemodifications on the MC3T3E1-produced OPNcompared to the 275-3-2 ras-transformed fibro-blastOPN [Christensen et al., 2007],whichmayexplain the observed differences in antibodyrecognition.
Figure 3B shows Western blot results usingthe monoclonal antibodies to detect OPNpresent in human urine. Four closely migratingspecies of OPN are observed [Kleinman et al.,2004, A. Beshensky and J. Wesson personalcommunication] and the antibody showingthe most intense binding to the four specieswas AK1H3. AK10F6 and two different poly-clonal antibodies were able to recognize all fourforms of OPN. AK3D9 binds the C-terminalregion of OPN, and strongly recognized onlythe two higher molecular weight forms ofOPN. Curiously, AK2A1 and AK2C5 showedno binding to urine OPN. This suggests thaturine OPN is glycosylated (blocking AK2C5binding), and may contain additional, possiblyunique, modifications which are able to specif-ically inhibit AK2A1 binding. The approxi-mately 30 kDa protein is a non-specific speciescross-reacting with the goat anti-mouse secon-dary antibody used. From these results wehypothesized that themajority of the antibodiesgenerated recognize sites subject to PTMs ofOPN.
Antibody Binding Sensitivity to PTMs
In order to test this hypothesis, apeptide binding assay was employed. Syntheticpeptides corresponding to the carboxy-terminusof the humanOPNmolecule (Fig. 4A), includingboth phosphorylated and non-phosphorylatedforms, and a scrambled amino acid controlpeptide, were used in amodified ELISA system.After the biotinylated peptides were bound toNeutra-avidin plates, AK3D9 and AK7B4 anti-body binding was determined. As shown inFigure 4B, the binding to the non-phosphory-lated peptide was very strong, whereas bindingto both the scrambled sequence peptide andthe phosphorylated peptide was similar tobackground. Similar results were obtained
Fig. 3. Monoclonal antibody recognition of OPN. A: Westernblotting results showing antibody recognition of murine OPN.Conditionedmedia (10 ml/lane) fromvarious cell lines or 50 ng ofrecombinantmurineOPN (GST-mOPN) were separated on 12%SDS–PAGE gels and transferred to PVDF membranes. Themembranes were then cut into strips which were blotted withmonoclonal antibodies at 1 mg/ml or polyclonal control at a1:3,000 dilution (shown above each lane). 275-3-2: ras-trans-formedmurine fibroblast cell line. 275: non-transformedmurinefibroblast 3T3 cell line. MC3T3E1: pre-osteoblast cell lineinduced to differentiate for 12 days as described in Materialsand Methods Section. B: Western blot of human urine detectedwith anti-OPNmonoclonal antibodies. Urine was collected anddialyzed extensively against 0.1M NaCl before approximatelytenfold concentration with Centriprep spin columns. Fivemicroliters of the concentrated, dialyzed urine was assayed viaSDS–PAGE and Western blotting with monoclonal antibodiesAK1H3, AK3D9, AK10F6, AK2A1, and AK2C5 at 1 mg/ml.Polyclonal antibody LF124 (kindly provided by Dr. Larry Fisher,NIH) andpolyclonal anti-OPN (recombinant)mouse serumwereused at 1:750 and 1:3,000 respectively.
930 Kazanecki et al.

using phosphorylated and non-phosphorylatedmurine peptides (data not shown).Figure 2A also shows the sequence of human
milk OPN with PTMs noted as determined byChristensen et al. [2005]. The AK2A1 antibodyis the only one whose epitope does notcontain sites of PTM in the mature protein.The AK2C5 antibody binds an area that isO-glycosylated, explaining why this antibodyexhibits decreased binding affinity to allforms of native OPN assayed. The remainingantibodies have been determined to bind toregions of the OPN molecule containing phos-phorylations that may interfere with epitoperecognition by these antibodies.
Inhibition of Cell Adhesion
Since many of the antibodies recognize the C-terminal half of the OPN molecule, where theCD44 receptor has been shown to mediateadhesion, the ability of our antibodies toinhibit the adhesion of cells to wells coated
with recombinant OPN was then assessed.Antibodies were added to wells pre-coatedwith human recombinant his-tagged OPN(hisOPN) and allowed to bind OPN. The wellswere thenwashed prior to adding humanMDA-MB-435 breast cancer cells or 275-3-2 murineras-transformed fibroblasts. The ability ofthe antibodies to block adhesion of thesecells to hisOPN was assessed by comparingthe adhesion to that in wells blocked with non-specific mouse IgG (Fig. 5). The resultsshow that for both cell lines examined, theAK3D9 and AK7B4 antibodies are able toinhibit cell adhesion by approximately 40–50%, possibly by interfering with the CD44receptor. Similar results were obtained withplates coated with native OPN purifiedfrom medium conditioned by ras-transformedfibroblasts (data not shown), which is weaklyphosphorylated, containing an average of fourphosphates per molecule [Christensen et al.,2007].
Fig. 4. Phosphorylation blocks 3D9 binding. A: Peptides usedfor the antibody-peptide binding assay kindly provided byDr. Larry Steinman. B: Binding of antibodies AK3D9 andAK7B4 to synthetic human OPN peptides. The biotinylatedpeptides were coated onto Neutra-avidin plates at 10 mg/mland detected with 5 mg/ml AK3D9 or AK7B4 monoclonal
antibody following the manufacturer’s instructions (PierceBiotech). The secondary antibody used was Alexafluor 594 goatanti-mouse IgG (Molecular Probes) and fluorescence wasdetected using excitation/emission wavelengths of 584/612 nm.Data shown are representative of three independent experiments(n¼ 2/exp).
Anti-Osteopontin Monoclonal Antibodies 931

DISCUSSION
Phage display is a powerful technique thatallows for rapid protein–protein interactionand provides a direct link between thephage phenotype and genotype [Dunn, 1996;Smothers et al., 2002]. Biopanning has thepotential to enrich for a phage of interest thatis rare in an initial library. However, highstringency conditions are required to enrich forphage with the highest affinities and to avoid
low affinity phage and background [Smotherset al., 2002]. In many cases these elutionconditions fail to release the most stronglybound phage. Some phage display vectors haveincorporated protease cleavage sites to over-come this problem [Jestin et al., 2001]. Thiswork describes a novel modification of thestandard protocol for mapping linear epitopesof monoclonal antibodies using phage display.While others have combined protein G pre-cipitation with phage display, this was not for
Fig. 5. Antibody inhibition of cell adhesion to recombinanthuman OPN. Tissue culture treated 96-well plates were coatedwith 150 mM human recombinant his-tagged OPN, thenblocked with 1% BSA. Antibodies were then added at 125 mMand allowed to bind OPN for 2 h. The wells were then washedand 5� 104 MDA-MB-435 (A) or 275-3-2 (B) cells were added
and allowed to adhere for 3 or 3.5 h respectively. Non-adherentcells were removed by washing and adherent cells werequantitated by staining with crystal violet. Data are representa-tive of four independent experiments for the MDA-MB-435 cellline and two independent experiments for the 275-3-2 cell line(n¼4). *P< 0.001 Student’s t-test.
932 Kazanecki et al.

the purpose of epitope mapping, or to isolateantibody-bound phage [Cui et al., 2003]. Themodification described here has several benefitscompared to the standard microplate bio-panning protocol. First, the use of protein G toprecipitate antibodies allows for the use ofcomplex mixtures such as ascites or hybridomasupernatant directly, without additional anti-body purification. Second, phage can be pre-cleared to lower non-specific binding, and anti-body-phage complexes are allowed to form insolution, decreasing background. Third, sincethe bound phage remain infective, the proteinG-antibody-phage complexes can be directlyadded to bacterial cultures and plated forplaque formation, thereby eliminating thetroublesome elution step. Finally, this methodis much faster than the standard protocol. Thismodified technique allowed for the identifica-tion of the epitopes of anti-OPN antibodieswhich were not identified using the standardmicroplate biopanningprotocol, suggesting thatthis modification lends increased sensitivity tothe protocol.The antibodies described hereinmay be of use
in determining the phosphorylation or glycosy-lation state of various regions of the OPNmolecule. This has been demonstrated in theWestern blots of conditioned medium frommultiple cell lines shown in Figure 3. All of theantibodies in our panel are able to bind to OPNin medium conditioned by ras-transformedfibroblasts (275-3-2), however only AK2A1 isable to recognize OPN in medium conditionedby a non-transformed fibroblast line or fromdifferentiated MC3T3E1 osteoblasts. Thesedata suggest that the OPN produced by theoverexpressing ras-transformed cells, is lesspost-translationally modified than proteinmade by non-ras transformed cells. OPN froma similar set of non-transformed and ras-trans-formed mouse NIH3T3 fibroblasts [Chamberset al., 1993] reacted similarly with the panel ofantibodies as the 275and275-3-2 cells.OPNhasbeen shown to be upregulated in a variety ofcancers, and our data suggest that the OPNproduced by certain tumors may have signifi-cantly fewer PTMs than that made by normalcells.The antibodies described were also able to
distinguish differences in OPN species found inhuman urine. For instance, the C-terminalantibody AK3D9 did not recognize the lowertwo of the four bands, suggesting that these
bands are either more phosphorylated thanthe protein in the upper bands, or representC-terminal truncated fragments of OPN.More work is required to distinguish thesepossibilities, but the ability of these antibodiesto identify these subtle differences in proteinstructure highlights their usefulness.
Antibodies recognizing the extreme C-terminal region of the OPN molecule (AK3D9andAK7B4)werealso able to inhibit adhesionofa mouse and human cell line to recombinanthuman OPN. This region of OPN has not beenpreviously implicated as having a role in celladhesion, and may represent the bindingsite of the CD44 receptor to the C-terminalthrombin fragment of OPN, which has not beenlocalized [Weber et al., 1996; Katagiri et al.,1999]. The AK3D9 binding region on OPN hasbeen shown to be differentially phosphorylateddepending on the cell line [Christensen et al.,2007], suggesting that phosphorylation of thisregion may have a role in regulating CD44binding.
Uede and colleagues have developed anti-bodies raisedagainstdefined regions of theOPNbackbone; one (1B20) recognizes the same areaas the AK3D9 antibody described herein[Kon et al., 2000, 2002]. Like AK3D9, 1B20shows similar differences in its ability torecognize the various forms of OPN containedin human urine. Interestingly, the C-terminal1B20 antibody was demonstrated to bindOPN produced by transfected CHO cells bothbefore and after acid-phosphatase treatment,suggesting that the C-terminal region ofthe OPN produced by these cells is not phos-phorylated.
Recent work has suggested that differentELISA assays have variable sensitivities forplasma and urine OPN [Kon et al., 2000;Vordermark et al., 2006]. Our results suggestthat some of this variability may result fromthe effects of PTMs on antibody reactivity, aswell as changes in protein structure resultingfrom proteolytic cleavage. Our results provideimportant new reagents to begin to address thiscomplication. Overall this study emphasizesthe importance of the heterogenous nature ofOPN PTMs, especially when comparing OPNproduced by various sources, and stressesthe need for careful selection of monoclonalantibodies used for the detection of OPN,particularly when using ELISA systems forquantitation.
Anti-Osteopontin Monoclonal Antibodies 933

ACKNOWLEDGMENTS
This work was funded in part by NIH grant#AR 44434 and ES 06897 (DTD), a BuschBiomedical Research Award (DTD), theNational Multiple Sclerosis Society, theRutgers Technology Commercialization Fund(DTD), theNational ScienceFoundation IGERTfellowship #DGE 0333196 (CCK), and NIHgrant #DK 067685 (SRR).
REFERENCES
Al-Shami R, Sorensen ES, Ek-Rylander B, Andersson G,Carson DD, Farach-Carson MC. 2005. Phosphorylatedosteopontin promotes migration of human choriocarci-noma cells via a p70 S6 kinase-dependent pathway.J Cell Biochem 94:1218–1233.
Chambers AF, Hota C, Prince CW. 1993. Adhesion ofmetastatic, ras-transformed NIH 3T3 cells to osteopon-tin, fibronectin, and laminin. Cancer Res 53:701–706.
Christensen B, Nielsen MS, Haselmann KF, Petersen TE,Sørensen ES. 2005. Post-translationally modified resi-dues of native human osteopontin are located in clusters:Identification of 36 phosphorylation and five O-glycosy-lation sites and their biological implications. Biochem J390:285–292.
Christensen B, Kazanecki CC, Petersen TE, Rittling SR,Denhardt DT, Sorensen ES. 2007. Cell-type specific post-translational modifications of mouse osteopontin areassociated with different adhesive properties. J BiolChem 282:19463–19472.
Crawford HC,Matrisian LM, Liaw L. 1998. Distinct roles ofosteopontin in host defense activity and tumor survivalduring squamous cell carcinoma progression in vivo.Cancer Res 58:5206–5215.
Cui X, Negesha HS, Holmes IH. 2003. Identification ofcrucial residues of conformational epitopes on VP2protein of infectious bursal disease virus by phagedisplay. J Virol Methods 109:75–83.
D’Alonzo RC, Kowalski AJ, Denhardt DT, Nickols GA,Partridge NC. 2002. Regulation of collagenase-3 andosteocalcin gene expression by collagen and osteopontinin differentiating MC3T3-E1 cells. J Biol Chem 277:24788–24798.
Declerck PJ, Carmeliet P, VerstrekenM, De Cock F, CollenD. 1995. Generation of monoclonal antibodies againstautologous proteins in gene-inactivated mice. J BiolChem 270:8397–8400.
Denhardt DT, Giachelli CM, Rittling SR. 2001a. Role ofosteopontin in cellular signaling and toxicant injury.Annu Rev Pharmacol Toxicol 41:723–749.
Denhardt DT, NodaM, O’Regan AW, Pavlin D, Berman JS.2001b. Osteopontin as a means to cope with environ-mental insults: Regulation of inflammation, tissueremodeling, and cell survival. J Clin Invest 107:1055–1061.
Dunn IS. 1996. Phage display of proteins. Curr OpinBiotechnol 7:547–553.
Ek-Rylander B, Flores M, Wendel M, Heinegard D,Andersson G. 1994. Dephosphorylation of osteopontinand bone sialoprotein by osteoclastic tartrate-resistant
acid phosphatase. Modulation of osteoclast adhesionin vitro. J Biol Chem 269:14853–14856.
Gericke A, Qin C, Spevak L, Fujimoto Y, Butler WT,Sorensen ES, Boskey AL. 2005. Importance of phosphor-ylation for osteopontin regulation of biomineralization.Calcif Tissue Int 77:45–54.
Goodwin AE, Pauli BU. 1995. A new adhesion assay usingbouyancy to remove non-adherent cells. J ImmunolMethods 187:213–219.
Harlow E, Lane D. 1999. ‘‘Using Antibodies: A laboratorymanual’’. Cold Spring Harbor: Cold Spring HarborLaboratory Press.
Hotta H, Kon S, Katagiri YU, Tosa N, Tsukamoto T,Chambers AF, Uede T. 1999. Detection of variousepitopes of murine osteopontin by monoclonal antibodies.Biochem Biophys Res Commun 257:6–11.
Hoyer JR, Asplin JR, Otvos L. 2001. Phosphorylatedosteopontin peptides suppress crystallization by inhibit-ing the growth of calcium oxalate crystals. Kidney Int60:77–82.
Jono S, Peinado C, Giachelli CM. 2000. Phosphorylation ofosteopontin is required for inhibition of vascular smoothmuscle cell calcification. J Biol Chem 275:20197–20203.
Jestin JL, Volioti G, Winter G. 2001. Improving the displayof proteins on flamentous phage. Res Microbiol 152:187–191.
Katagiri YU, Sleeman J, Fujii H, Herrlich P, Hotta H,Tanaka K, Chikuma S, Yagita H, Okumura K,MurakamiM, Saiki I, Chambers AF, Uede T. 1999. CD44 variantsbut not CD44s cooperate with beta1-containing integrinsto permit cells to bind to osteopontin independently ofarginine-glycine-aspartic acid, thereby stimulating cellmotility and chemotaxis. Cancer Res 59:219–226.
Keykhosravani M, Doherty-Kirby A, Zhang C, Brewer D,Goldberg HA, Hunter GK, Lajoie G. 2005. Comprehen-sive identification of post-translational modifications ofrat bone osteopontin bymass spectrometry. Biochemistry44:6990–7003.
Kleinman JG, Wesson JA, Hughes J. 2004. Osteopontinand calcium stone formation. Nephron Physiol 98:43–47.
Kon S, Maeda M, Segawa T, Hagiwara Y, Horikoshi Y,Chikuma S, Tanaka K, Rashid MM, Inobe M, ChambersAF, Uede T. 2000. Antibodies to different peptides inosteopontin reveal complexities in the various secretedforms. J Cell Biochem 77:487–498.
Kon S, Yokosaki Y, Maeda M, Segawa T, Horikoshi Y,Tsukagoshi H, RashidMM,Morimoto J, InobeM, ShijuboN, Chambers AF, Uede T. 2002. Mapping of functionalepitopes of osteopontin by monoclonal antibodies raisedagainst defined internal sequences. J Cell Biochem84:420–432.
Kowalski A. 2005. Creation, characterization, and applica-tion of novel anti-osteopontin monoclonal antibodies.Doctoral Dissertation, Department of Microbiology andMolecular Genetics. Rutgers University, the State Uni-versity of New Jersey.
Pampena DA, Robertson KA, Litvinova O, Lajoie G,Goldberg HA, Hunter GK. 2003. Inhibition of hydrox-yapatite formation by osteopontin phosphopeptides.Biochem J 378(Pt 3):1083–1087.
Razzouk S, Brunn JC, Qin C, Tye CE, Goldberg HA, ButlerWT. 2002. Osteopontin posttranslational modifications,possibly phosphorylation, are required for in vitro boneresorption but not osteoclast adhesion. Bone 30:40–47.
934 Kazanecki et al.

Rittling SR, Matsumoto HN, McKee MD, Nanci A, AnXR, Novick KE, Kowalski AJ, Noda M, DenhardtDT. 1998. Mice lacking osteopontin show normal devel-opment and bone structure but display altered osteo-clast formation in vitro. J Bone Miner Res 13:1101–1111.
Rollo EE. 1995. Inhibition of nitric oxide production andmacrophage-mediated cytotoxicity by recombinanthuman osteopontin. Doctoral Dissertation, Departmentof Microbiology and Molecular Genetics. Rutgers, theState University of New Jersey.
Smothers JF, Henikoff S, Carter P. 2002. Tech. Sight.Phage display. Affinity selection from biological libraries.Science 298:621–622.
Sodek J, Batista Da Silva AP, Zohar R. 2006. Osteopontinand mucosal protection. J Dent Res 85:404–415.
Sørensen ES, Hojrup P, Petersen TE. 1995. Posttransla-tional modifications of bovine osteopontin: Identificationof twenty-eight phosphorylation and three O-glycosyla-tion sites. Protein Sci 4:2040–2049.
Vordermark D, Said HM, Katzer A, Kuhnt T, Hansgen G,Dunst J, Flentje M, Bache M. 2006. Plasma osteopontinlevels in patients with head and neck cancer and cervix
cancer are critically dependent on the choice of ELISAsystem. BMC Cancer 6:207.
Weber GF, Ashkar S, Glimcher MJ, Cantor H. 1996.Receptor-ligand interaction between CD44 and osteo-pontin (Eta-1). Science 271:509–512.
Wu Y, Denhardt DT, Rittling SR. 2000. Osteopontin isrequired for full expression of the transformed phenotypeby the ras oncogene. Br J Cancer 83:156–163.
Young MF, Kerr JM, Termine JD, Wewer UM, Wang MG,McBride OW, Fisher LW. 1990. cDNA cloning, mRNAdistribution and heterogeneity, chromosomal location,and RFLP analysis of human osteopontin (OPN).Genomics 7:491–502.
Zhu B, Suzuki K, Goldberg HA, Rittling SR, Denhardt DT,McCulloch CA, Sodek J. 2004. Osteopontin modulatesCD44-dependent chemotaxis of peritoneal macrophagesthrough G-protein-coupled receptors: Evidence of a rolefor an intracellular form of osteopontin. J Cell Physiol198:155–167.
Zohar R, Lee W, Arora P, Cheifetz S, McCulloch C, Sodek J.1997. Single cell analysis of intracellular osteopontin inosteogenic cultures of fetal rat calvarial cells. J CellPhysiol 170:88–100.
Anti-Osteopontin Monoclonal Antibodies 935





























