Embed Size (px)
Citation preview

IgY antiporcine endothelial cell antibodieseffectively block human antiporcinexenoantibody binding
Fryer JP, Firca J, Leventhal JR, Blondin B, Malcolm A, Ivancic D,Gandhi R, Shah A, Pao W, Abecassis M, Kaufman DB, Stuart F,Anderson B. IgY antiporcine endothelial cell antibodies effectively blockhuman antiporcine xenoantibody binding. Xenotransplantation 1999; 6:98±109. # Munksgaard, Copenhagen.
Avian IgY antibodies are structurally different from mammalian IgGsand do not ®x mammalian complement components or bind human Fcreceptors. As these antibody-mediated interactions are believed to playsigni®cant roles in both hyperacute rejection (HAR) and acute vascularxenograft rejection (AVXR), IgY antibodies to xenoantigen targetepitopes may inhibit these rejection processes. In this report, we showthat chicken IgY antibodies to a-Gal antigen epitopes and to otherporcine aortic endothelial cell (PAEC) antigens block human xenor-eactive natural antibody binding to both porcine and rat cardiac tissuesand porcine kidney tissues. Chicken IgY antibodies blocked comple-ment-mediated lysis of PAECs by human serum, and inhibited antibody-dependent cell-mediated lysis of PAECs by heat-inactivated humanserum plus peripheral blood leukocytes. Binding of IgY to porcineendothelial cells did not affect cell morphology nor expression of E-selectin. These results suggest that avian IgYs could be of potential use ininhibiting pig-to-human xenograft rejection.
Jonathan Fryer1 Joseph Firca2
Joseph Leventhal1 Beth Blondin1
Andrew Malcolm3 David Ivancic1
Ripal Gandhi1 Ateet Shah1 WinniePao1 Michael Abecassis1 DixonKaufman1 Frank Stuart1 and ByronAnderson4
Department of 1Surgery, North-western UniversityMedical School, Chicago, IL, USA; 2OphidianPharmaceuticals, Madison, WI, USA; 3Departmentof Laboratory Medicine and Pathology, University ofAlberta, Edmonton, Alberta, Canada; 4Cell andMolecular Biology, North-western UniversityMedical School, Chicago, IL, USA
Key words: endothelial activation ± endothelialcells ± IgY ± xenoantibodies ±xenotransplantation
Address reprint requests to Dr Jonathan P. Fryer,Department of Surgery, Division of OrganTransplantation, North-western University, 303 E.Superior Street, Su 536, Chicago, IL 60611, USA.
Received 9 November 1999;
accepted 5 February 1999
Introduction
In many discordant species combinations, hyper-acute rejection of xenografts is mediated byxenoreactive natural antibodies (XNAs). In thepig-to-human system, the major antigen recog-nized by XNAs is the Gala1,3Galb1,R epitopesequence (aGal) [1,2], and it has been determinedthat 1±2% and 4±8% of human IgG and IgM,respectively, are reactive with aGal [3]. Theendothelial cell binding of XNAs, and subsequentbinding of complement proteins with activationof the classical complement system, results inxenograft endothelial cell activation and retrac-tion, platelet activation with formation ofthrombi, and graft in®ltration by neutrophils,natural killer (NK) cells and macrophages [4±6].
Thus, prevention of complement activation [7±9], modulation of the aGal epitope [10±12], andprevention of XNA binding to aGal [13±15] arestrategies that could potentially prevent hyper-acute rejection (HAR). If the HAR responseis suppressed, a delayed xenograft rejection(DXR) occurs [5,16]. Evidence from recentstudies suggests that the DXR response may bemediated by NK cells and monocytes bindingto endothelium-bound XNAs or directly toendothelial carbohydrate epitopes [16±20].Another factor that may contribute to DXR isendothelial cell activation caused by XNAbinding without the involvement of complement.Endothelial activation can lead to both morpho-logic and physiologic changes that can signi®-cantly alter interactions with other cells [21±23].
Xenotransplantation 1999: 56: 98±109
Printed in UK. All rights reserved
Copyright # Munksgaard 1999
XENOTRANSPLANTATIONISSN 0908±665X
98

Avian species, like humans, lack aGal epitopeexpression [24] and therefore should respondto immunization with aGal antigen. Indeed,Bouhours et al. [25] have shown that chickensproduce anti-aGal antibodies in response toimmunization with rabbit erythrocyte mem-branes containing aGal epitopes. The majorIgG-like antibody of the chicken is designatedIgY and is found in high quantities in the egg yolkfrom which it can be readily isolated by severalmethods [26±28]. IgY does not bind humancomplement or Fc receptors [28]. Thus, if thevasculature of xenograft tissues were coated withIgY antiendothelial antibodies, both HAR andDXR may be prevented.
In this report, using an in vitro porcine aorticendothelial cell (PAEC) system, we show thatprevention of complement-mediated lysis (CML)and antibody-dependent cell-mediated lysis(ADCC) by human serum (source of XNAsand complement) and peripheral blood leuko-cytes (PBLs, ADCC effector cells) can beinhibited in a dose-dependent manner by chickenIgY produced by immunization with aGalepitope or PAECs
Materials and methods
Antigen and antibody preparation
The aGal trisaccharide epitope sequencesGala1,3Galb1,4Gal and Gala1,3Galb1,4GlcNAc,purchased from V Labs (Covington, LA, USA),were conjugated to ovalbumin (Sigma, St Louis,MO, USA) by reductive amination withNaCNBH3 (Sigma) [29]. Using the phenol±sulfuricacid method for neutral sugar quantitation [30] andaGal as a standard, 8±10 trisaccharides were foundto be coupled per ovalbumin molecule.
The Gala1,3Galb1,4GlcNAc linked to bovineserum albumin (BSA) was used as the targetepitope in ELISAs. Four white Leghorn chickenswere immunized by biweekly intramuscular (i.m.)injection for 16 weeks with 125 mg ofaGal-ovalbumin. Two chickens were immunizedwith the Gala1,3Gal b1,4Gal conjugate and twowith the Gala1,3Galb1,4GlcNAc conjugate. Theantigens were emulsi®ed in Freund's completeadjuvant for the ®rst injections; all subsequentinjections were with antigens emulsi®ed inincomplete adjuvant. PAECs were obtainedfrom in vitro culture. Con¯uent cells werewashed with PBS and scrape harvested. Onemilligram of cell protein was injected i.m. intotwo chickens once biweekly. For an additional6 weeks, the chickens were immunized withPAECs that had been stimulated for 24 h with
10 ng/ml human tumor necrosis factor alpha(TNF-a) (Genzyme, Cambridge, MA, USA)(tnfPAEC). Egg collection was terminated 1and 2 months after the last immunization withPAECs and aGal conjugates respectively.
A method to purify lipid-free IgY from eggyolk was developed by modifying the proceduredescribed by Hatta et al. [27]. Egg yolks mixed1:1 with PBS were treated with 1.5 mg/mlpolysulfated carrageenan (Sigma) and the result-ing precipitate collected and discarded. IgY wasthen precipitated with 20% ammonium sulfate.The precipitate was collected, dissolved in PBSand ®ltered through diatomaceous earth followedby reprecipitation with 20% ammonium sulfate.The ®nal preparation was essentially lipid free asanalyzed by thin-layer chromatography devel-oped with a chloroform±methanol solvent andwas nearly 100% IgY by SDS±PAGE analysisshowing the IgY heavy and light chains at60 kDa and 25 kDa respectively. The IgY pre-parations were af®nity puri®ed using the aGalepitope trisaccharide (Gala1,3Galb1,4GlcNAc)linked to Synsorb beads (Alberta ResearchCouncil, Edmonton, Alta, Canada). Humanserum anti-aGal antibodies were also isolatedon the same af®nity matrix.
Enzyme-linked immunosorbent assay (ELISA)
Gala1,3Galb1, 4GlcNAc-BSA (aGal-BSA),100 ng in 100 ml of 0.1 M sodium bicarbonate,pH 9.0, was used to coat the wells of Titertek96-well ELISA plates (Titertek, ICNBiomedicals) at room temperature for 1 h (orovernight at 4 uC) and washed with PBS contain-ing 0.08% sodium azide and 0.08% Tween-20(Sigma), then backcoated with 300 ml of PBSA(1% BSA in PBS) per well at room temperaturefor 1 h (or overnight at 4 uC). The plate waswashed with PBS, and 100 ml per well of humanor avian antibodies in PBSA was incubated for1 h at room temperature. Alkaline phosphatase-labeled secondary antibodies, 100 ml, were addedfor 1 h at room temperature and the wells washedwith Tris-HCl buffer, pH 7.0. Sigma 104 alkalinephosphatase substrate, 100 ml in Tris buffer,pH 9.5, was added per well. The ODs at414 nm were obtained using a TitertekMultiskan plate reader (Flow Labs, McClean,VA, USA).
IgY preparations (100 ml) were added at0.5 mg/ml to wells coated with aGal-BSA asthe target antigen and incubated for 1 h at roomtemperature. Bound IgY was detected withalkaline phosphatase-conjugated rabbit anti-chicken IgY reagent, activity of the alkaline
Blocking of human antiporcine xenoantibody binding
99

phosphatase being determined with the substratep-nitrophenylphosphate and product p-nitrophe-nol being quantitated at 414 nm after 15 minincubation.
When PAECs were used as antigen target, thecells were cultured as described below in 48-welltissue culture plates until con¯uent. Plates werestored at 4 uC after the cells were ®xed with 2%glutaraldehyde for 10 min at room temperatureand washed with PBS. The ELISA was thenperformed as described above.
Complement-mediated lysis (CML) of PAECs.
PAECs were grown in 24- or 48-well Falconculture plates (Becton Dickinson, Lincoln Park,NJ, USA) coated by the addition of 0.5 ml/well of2% gelatin Type B from bovine skin (Sigma)diluted 1:10 in Hanks' buffered salt solution(HBSS, Gibco BRL, Grand Island, NY, USA)and incubated for 15 min at room temperature.The gelatin solution was removed and 125 3 103
PAECs in 2 ml of cell culture medium (CCM),Dulbecco's modi®ed Eagle medium (DMEM),with high glucose (Gibco BRL) containing 10±20% de®ned fetal bovine serum (FBS) (HyCloneLaboratories, Logan, UT, USA) that was heat-inactivated for 30 min at 56 uC, 1% (v/v) 200 mMl-glutamine (Gibco BRL) and penicillin Gsodium (10 000 units/ml)/streptomycin sulfate(10 000 mg/ml in 0.85% saline) (Gibco BRL)]were added per well. The plate was centrifugedfor 2 min to settle the cells onto the gelatin in aneven monolayer and the plate incubated at 37 uCunder 5% CO2/95% air. After 4 h, when cellattachment had occurred and the CCM wasremoved and replaced with 2 ml of CCM. In 2±4 days the PAECs had grown to 80±100%con¯uence and were used in the chromium releaseassays.
The PAECs were washed three times with 1 mlof serum-free medium (SFM, CCM minus theFBS). Chromium (51Cr,Na51CrO4, Amersham,Arlington Heights, IL, USA) was diluted in SFMto give 40 mCi/ml, and 100 ml was added to eachwell of the PAEC culture plate and the plateincubated for 2±3 h at 37 uC under 5% CO2±95%air. The wells were washed three times with SFM,and then 1 ml of PBS was added to each well andincubated at 37 uC for 30 min to help reduce the51Cr background.
After washing away excess 51Cr, 100 ml ofchicken IgY preparations plus 200 ml of SFMwere incubated with half of the cells for 1 h; incontrol wells, the cells were incubated for 1 h with300 ml of SFM. Undiluted human serum (HS,100 ml) was then added to each well and
incubated at 37 uC for 1 h. Supernatant (200 ml)was removed, added to 200 ml of scintillationcocktail (Optiphase `Supermix', Wallac, Turku,Finland) in each well of the terephthalate readingplate (Wallac), which was then sealed with anacetate plate sealer (Dynatech Laboratories,Chantilly, VA, USA) and placed on a shaker toobtain a homogeneous mixture. Radioactivity(CPMs) was determined using the 1450MicroBeta PLUS plate reader (Wallac).
Total 51Cr lysis controls were determined from200-ml aliquots of cells treated with 5% Triton-100. Total complement-mediated lysis (CML)was determined from cells treated with HS alone.Background lysis was de®ned by cells treated withSFM alone, heat-inactivated human serum(HIHS) alone or IgY preparation alone (thesevalues were similar). The average backgroundwas subtracted from all cell counts. The percen-tage inhibition of cell lysis by chicken IgY wascalculated as the total CML with HS alone(100%) minus the percentage CML in thepresence of IgY. Per cent inhibition was plottedagainst inhibitor concentrations.
Antibody-dependent cell-mediated lysis (ADCC) of PAECs
This assay was a modi®ed version of the CMLassay and similar to that described bySchaapherder et al. [18]. PAECs were culturedand labeled with 51Cr. PAECs in the 48-wellplates were incubated for 1 h with chicken IgYpreparations or with SFM alone, followed bysequential additions of HI-HS (100 ml) and ®nallyfreshly isolated PBLs (100 ml) in various ratios ofPBLs/PAECs. After 4 h of incubation, 200 mlof supernatant was removed, mixed with 200 mlof scintillation cocktail (Wallac) and CPMsrecorded on the 1450 MicroBetaPLUS platereader (Wallac).
Total lysis was determined by incubatingPAECs with 200 ml Triton X-100. Effector totarget (E/T) ratios (PBLs/PAECs) ranged from25:1 to 2.5:1. Background counts were deter-mined for PAECs incubated with each of thefollowing: SFM alone, HIHS alone, IgY aloneand PBLs alone. The percentage inhibition ofADCC was calculated and the data plotted in thesame manner, as described above.
Immuno¯uorescence (IF) studies of antibody binding to
porcine or rat tissues
Frozen sections of porcine heart, kidney and ratheart were cut into 7-mm sections and ®xed in ±20 uC acetone for 15 min. For GS-IB4 lectin(Griffonia simplicifolia-I lectin, B4 isoform)
Fryer et al.
100
staining, ¯uorescein-labeled GS-IB4 (Sigma)diluted 1:200 in PBS was added.
For chicken IgY or human antibody staining,frozen sections were coated with the antibodypreparations, then washed with PBS and coatedwith 10% host serum (from same source as thesecondary antibodies were derived) to reducenon-speci®c binding. The sections were washedwith PBS and ¯uorescein-labeled secondaryantibodies (rabbit anti-chicken IgY or goatanti-human Ig reagents, Sigma) were added andincubated for 1 h at room temperature.
For blocking of human XNA binding toanimal tissue, either chicken IgY was pre-incubated with the tissue for 1 h at roomtemperature, followed by incubation withhuman serum for 1 h, or the chicken IgY waspre-mixed with human serum and incubated withthe tissue. The tissue sections were washed andthen incubated with secondary ¯uorescein-labeled anti-human Ig reagent (goat antibodyspeci®c for either human IgM or IgG) todetermine blocking. Fluorescein intensities wereassessed visually by two pathologists, and their
estimates, which were within 10% of each other,were averaged.
IF studies for E-Selectin and VCAM expression on cultured
cells
Human aortic endothelial cells (HAECs)(Regional Organ Bank of Illinois), porcineaortic endothelial cells (PAECs) (Cell Systems,Kirkland, WA, USA) and human CD55/CD59transgenic PAECs (tg PAECs) (Nextran,Princeton, NJ, USA) were grown to con¯uencyon Falcon tissue culture slides (BectonDickinson) with endothelial growth media(Gibco BRL) plus 10% FCS.
Endothelial cells were incubated with SFM andor 10% sera for 30 min at 37 uC or 20 ng/mlhuman TNF-a for 4 h at 37 uC. The cells werewashed with PBS and immediately immersed in ±20 uC acetone for 15 min, then air dried for5 min. The cells were then blocked by 10% non-immune rat serum for 10 min at room tempera-ture. Primary antibodies: porcine cross-reactivemouse anti-human E-selectin (1.2B6) and mouse
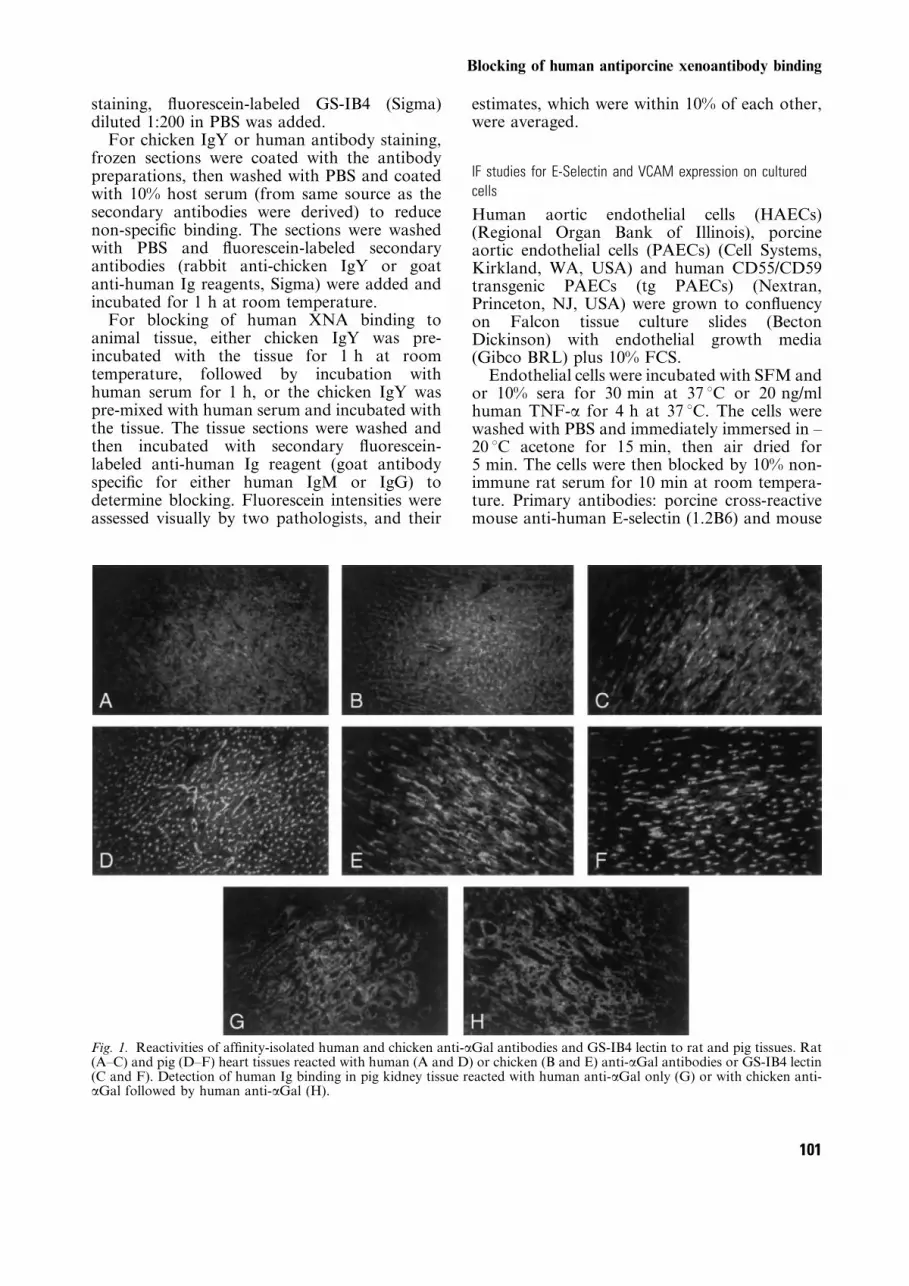
Fig. 1. Reactivities of af®nity-isolated human and chicken anti-aGal antibodies and GS-IB4 lectin to rat and pig tissues. Rat(A±C) and pig (D±F) heart tissues reacted with human (A and D) or chicken (B and E) anti-aGal antibodies or GS-IB4 lectin(C and F). Detection of human Ig binding in pig kidney tissue reacted with human anti-aGal only (G) or with chicken anti-aGal followed by human anti-aGal (H).
Blocking of human antiporcine xenoantibody binding
101

antiporcine VCAM (10.2B7) (kind gifts from DrDorian Haskard, Hammersmith Hospital,London, UK) and mouse anti-human VCAM(Ab-1) (Calbiochem, Cambridge, MA, USA)
were added at a dilution of 1:500 for 1 h atroom temperature. The cells were washed andsecondary antibody, biotinylated rat anti-mouseIgG (Zymed) at a dilution of 1:50, was added for1 h at room temperature and the cells washed.Fluorescein-labeled streptavidin (Zymed) wasadded at a dilution 1:100 for 15 min at roomtemperature. The slides were viewed under¯uorescence using a Zeiss Axioskop microscope(Carl Zeiss, Thornwood, NY, USA).
Evaluation of antibody induced apoptosis in endothelial cells
Endothelial cells will be evaluated for apoptosisafter incubation with antibodies for various timeperiods. Apoptosis will be evaluated using bothan Annexin V binding assay (TACS, Trevigen,Gaithersburg, MD, USA), which identi®es phos-phatidylserine exposure on the endothelial cellsurface ± an early phase in the apoptotic process,and by the Tunel assay (Apoptag, Oncor,Gaithersburg, MD, USA), which identi®esDNA fragmentation. These techniques will beperformed according to the manufacturers'recommendations.
Fig. 2. Titering of af®nity-isolated human and chicken anti-aGal to porcine aortic endothelial cells (PAECs).
Table 1. Blocking of human serum xenoreactive antibodies to porcine and rat heart,
and porcine kidney tissues by chicken IgY anti-7Symbol uGal (anti-Gal), anti-PAEC
and anti-tnfPAEC antibodies
Blocking
antibody Tissue
Human Ig
evaluated
Per cent
blocking
Anti-Gal Pig kidney 20±30
(af®nity puri®ed) Pig heart IgM 40±50
Pre-mixed Rat heart 10±20
Anti-Gal Pig kidney 90±100
(af®nity puri®ed) Pig heart IgM 90±100
Pre-incubated Rat heart 90±100
Anti-PAEC Pig kidney 80±90
Premixed Pig heart IgM 80±90
Rat heart 80±90
Anti-tnfPAEC Pig kidney IgM, IgG 80±90, 90±100
Premixed Pig heart IgM, IgG 70±80, 90±100
Rat heart IgM, IgG 80±90, 90±100
Anti-tnfPAEC Pig kidney 90±100
Pre-incubated Pig heart IgM 90±100
Rat heart 90±100
Fryer et al.
102

Results
Reactivities of af®nity-puri®ed human anti-aGal, chicken
anti-a Gal, and lectin GS-IB4 to rat and pig tissues
The af®nity-puri®ed human and chicken anti-aGal antibodies were incubated with cryostat-cutfrozen tissue sections of rat and pig heart, and thebound antibodies detected using ¯uoresceinatedanti-human or anti-chicken reagents. As shown inFig. 1 the patterns of binding of chicken orhuman anti-aGal and of the lectin, are similar.The vessels of the heart tissue sections arestrongly positive. The human anti-aGal demon-strated stronger binding to the porcine heartvessels than to the rat. The chicken anti-aGal andlectin also exhibited stronger reaction to theporcine heart tissue sections. Porcine kidneyincubated with the af®nity-isolated human anti-aGal showed the presence of the aGal epitope atthe luminal face of the renal tubules and noreaction with vessels.
Blocking of human XNA binding to porcine or rat tissues
with IgY preparations
It would be expected that if the human andchicken anti-aGal antibodies are reacting to thesame epitopes the chicken anti-aGal could blockhuman anti-aGal antibody binding. Therefore,pig kidney tissue sections were ®rst incubatedwith chicken IgY reagents followed by reactionwith human antibodies and the extent of anyreduction in human antibody binding estimatedby reduction in the ¯uorescence staining. As seenin Fig. 1, essentially complete inhibition ofhuman antibody binding to the renal tubuleswas achieved.
Additional blocking experiments were con-ducted using the chicken IgY preparations ofanti-aGal and anti-PAEC. Porcine or rat heartsor porcine kidney cryostat-cut tissue sectionswere incubated with a mixture of IgY preparationplus human serum, or incubated ®rst with IgYpreparation followed by human serum, and theamounts of human IgG or IgM binding to thetissue estimated (Table 1).
When af®nity-puri®ed anti-aGal IgY wasmixed with human serum, only 10±50% blockingof human IgG or IgM was obtained. There was
no discernible difference between the blocking ofthe human IgG or IgM class of XNAs. Theblocking effects of the IgY anti-aGal was greaterfor the porcine heart than for the porcine kidneyor rat heart and could be indicative that the aGalepitope sequence distributions differ in thosetissues. If the same af®nity-puri®ed IgY anti-aGalwas pre-incubated with the kidney or hearttissues, nearly all subsequent binding of humanIgM XNAs was blocked in the 1-h incubationtime with human serum; the effects on IgG XNAbinding were not determined.
The anti-PAEC and anti-tnfPAEC IgY pre-parations exhibited nearly complete blocking ofhuman IgM XNAs whether the tissues were pre-incubated with the IgY or whether the humanserum was pre-mixed with the IgY preparations.For the anti-tnfPAEC IgY pre-mixing withhuman serum experiment, both human IgG andIgM binding were examined and the IgG bindingwas found to be blocked completely.
Reactivities of human anti-aGal and chicken anti-aGal, anti-
PAEC and anti-TNF-a±PAEC antibodies to PAECs
The relative reactivities of antibodies to anantigen under equilibrium conditions and pro-portionality of binding to antibody concentrationre¯ect differences of the antibody binding af®-nities. Increasing quantities of the human andchicken af®nity puri®ed anti-aGal antibodieswere incubated with PAECs as target antigenfor 2 h to allow binding equilibrium. The chickenanti-aGal exhibited about three times greaterreactivity to PAECs with increasing antibodyconcentration than the human anti-aGal (Fig. 2).If it is assumed that the highest concentrationstested are at saturation of binding, then theestimated Kd values at half-maximal binding areabout 0.3 ? 10±6 M for human anti-aGal anti-bodies and 0.8 ? 10±7 M for the chicken anti-aGal antibodies.
The egg yolk IgY preparations from chickensimmunized with Gala1,3Galb1,4Gal-ovalbumin[anti-Gal(Gal)] or Gala1,3Galb1,4GlcNAc-oval-bumin [anti-7SymbolGal(GlcNAc)], and chick-ens immunized with PAECs or TNF-a-stimulatedPAECs were titered against PAECs as the antigentarget. As shown in Fig. 3, the reactivities of thetwo anti-aGal preparations are similar in bind-ing, as are the anti-PAEC and anti-tnfPAEC IgYpreparations. The last two IgY preparationsexhibited about 50% greater reactivity toPAECs than the anti-aGal IgY preparation atconcentrations of half-saturation of binding andabout 20±25% greater reactivities at near satura-tion amounts of the IgY preparations. These
Table 2. Reactivity of chicken anti-7Symbol uGal (anti-Gal) and anti-PAEC IgY
preparations to aGal epitope
IgY
preparation
Product formation
(ODs at 414 nm) Activity/mg IgY
anti-Gal 1.885 0.038
anti-PAEC 0.880 0.018
Blocking of human antiporcine xenoantibody binding
103

results are consistent with the expectation that theanti-PAEC IgYs contain antibodies to PAECcell-surface antigens other than the aGal epitopesequences.
The anti-aGal and anti-PAEC IgY prepara-tions were also tested for reactivity to the aGal-BSA antigen target under conditions of linearityof binding vs. IgY concentrations, and the anti-PAEC IgYs showedà 50% of the binding activityfor IgY concentration than the anti-aGal IgYs(Table 2). This result together with the bindingcurves of Fig. 3 indicate that about half of theanti-PAEC IgYs of those preparations aredirected to the aGal epitope sequences.
Chicken anti-aGal and anti-PAEC IgY pre-parations inhibit human XNA-initiatedcomplement-mediated and PBL-mediated lysisof PAECs. Because chicken IgYs can blockhuman XNA binding to porcine tissues (Fig. 1and Table 1) and because avian IgYs do not ®xand activate human complement, the IgY pre-parations should inhibit human XNA andcomplement mediated lysis of PAECs. As
shown in Fig. 4, this is the case. The anti-aGalIgY preparations (100 ml) inhibited up to 90% ofhuman serum mediated lysis at the 1.6 mg/ml IgYprotein concentration added to the incubationmixture with 100 ml of human serum. The anti-PAEC and anti-tnfPAEC IgY preparationsexhibited essentially 100% inhibitions at about5 mg/ml of the added IgYs. The total amount ofIgYs in the incubation mixture was 160 mg foranti-aGal and 500 mg for the anti-PAEC pre-parations or approximately twice the amountsthat yielded saturation of binding in the titeringexperiments shown in Fig. 3.
Inhibition of antibody-dependent cell-mediated cytotoxicity
(ADCC) using IgY
Because IgY does not interact with human Fcreceptors, ADCC experiments were performed tosee whether anti-Gal IgY would inhibit ADCC-mediated lysis of PAECs. As the data fromTable 3 show, IgY successfully inhibited ADCC-mediated lysis of PAECs. When PAECs were pre-
Fig. 3. Titering of anti-PAEC and anti-aGal IgY preparations to PAECs. Anti-tnfPAEC IgY, IgY antibodies isolated fromchickens immunized with TNF-a-stimulated PAECs. Anti-PAEC IgY, IgY antibodies isolated from chickens immunized withnon-stimulated PAECs. Anti-aGal(GlcNAc), IgY antibodies isolated from chickens immunized with Gala1,3Galb1,4GlcNAc-ovalbumin. Anti-aGal (Gal), IgY antibodies isolated from chickens immunized with Gala1,3Galb1,4Gal-ovalbumin.
Fryer et al.
104

incubated with anti-aGal(Gal) IgY (5 mg/ml at37 uC for 1 h) followed by HIHS and PBLs, theADCC-mediated lysis was signi®cantly inhibited.Furthermore, no PAEC lysis occurred wheneither HIHS or avian antibodies alone wereincubated with non-transgenic PAECs.
Effects of chicken IgY on PAEC activation and apoptosis
A number of investigators have shown thathuman XNA or GSI-B4 lectin binding toPAECs results in their activation and mayin¯uence adhesive interactions with circulating
PBLs [31±33]. We therefore asked what effectschicken IgY would have on PAECs.
E-selectin and VCAM expression were up-regulated on PAECs and TgPAECs when incu-bated for 1 h with human or porcine TNF-a(20 ng/ml), heat-inactivated pooled human ABserum (PHS) or af®nity-isolated human anti-aGal antibodies (AIHAG). No E-selectin orVCAM up-regulation was seen after 1-, 4- or24-h incubations with pig serum, chickenanti-aGal IgY (4 mg/ml), chicken anti-PAECIgY (2.66 mg/ml), chicken anti-tnfPAECs IgY(3.22 mg/ml), af®nity-puri®ed chicken anti-aGalIgY (9.2 mg/ml).
Previous studies have shown that human seracan induce apoptotic changes in PAECs [34]. Wehave established positive controls for apoptosisby incubating HAECs and PAECs with staur-osporin (0.5 mM at room temperature) for 4 h todetect annexin binding and for 24 h for a positiveresult using the Tunel assay. Additional studieswere performed to determine the in¯uence ofIgY antibody binding on apoptosis of PAECs.PAECs were incubated with the following anti-body sources: chicken anti-aGal IgY (4.0 mg/ml);chicken anti-PAEC IgY (2.66 mg/ml); chicken
Fig. 4. Inhibition of human antibody and serum-mediated lysis of PAECs by chicken IgY preparations.
Table 3 Inhibition of ADCC with avian IgY antibodies
E/T
ratio
hPBL/HIHS
no IgY
(% ADCC)
hPBL/HIHS
IgY
(% ADCC)
Inhibition of
ADCC (%)
25:1 22.8 ¡ 0.9 4.8 ¡ 2.5* 79
20:1 18.5 ¡ 3.1 6.0 ¡ 3.6* 68
15:1 11.4 ¡ 5.0 7.8 ¡ 5.5* 32
10:1 10.6 ¡ 3.4 6.9 ¡ 7.0 35
ADCC, antibody-dependent cell-mediated cytotoxicity; E/T, effector cell to target cellratio; hPBL, human peripheral blood leukocytes; HIHS, heat inactivated pooledhuman AB serum. *P-value , 0.05 compared with no IgY, using paired Student's t-test).
Blocking of human antiporcine xenoantibody binding
105

anti-tnfPAECs IgY (3.22 mg/ml); af®nity-isolated chicken anti-aGal IgY (9.2 mg/ml).Each IgY antibody source was incubated withPAECs for up to 96 h and none inducedapoptosis in the PAECs as detected by annexinbinding or by Tunel assay.
Discussion
Although pig-to-human xenotransplantationseems closer to reality than ever before, thesequelae of human xenoantibodies binding toporcine endothelial antigens continue to be asigni®cant barrier to success. Xenoantibody±antigen interactions appear to play a major rolein both HAR and DXR. Strategies that have beenattempted to block these interactions include:depleting preformed antibody [14,15,35], inhibit-ing new antibody formation [36, 37], andeliminating or modulating target antigens[11,12, 38].
Although xenoantibody±antigen interactionsoccurring in any tissue may in¯uence xenograftsurvival, the endothelial cell is clearly the focalpoint for DXR [39]. Column depletion of IgG,IgM [35] or anti-aGal antibodies [14,15] has beenshown to decrease endothelial deposition ofxenoantibodies and prolong xenograft survivalin pig-to-primate models. However, these deple-tion techniques cannot keep up with the largequantities of antibodies that are produced afterxenotransplantation [35,36]. Therefore, althoughall of these approaches have shown promise, nonehas completely overcome the problem.
With HAR, xenoantibody binding triggersactivation of the classical pathway of thecomplement cascade, which ultimately leads tosigni®cant endothelial injury and graft loss.Although successful complement inhibitory stra-tegies prevent HAR, they do not prevent DXR.This may be because the complement cascade isnot completely inhibited as some investigators[36] have shown that complement protein deposi-tion is still detectable. Furthermore, as mostcomplement inhibitory strategies are directed atthe level of C3 or more distal components of thecomplement cascade, the early components of theclassical pathway, which can also initiate proin-¯ammatory and procoagulant events [6], con-tinue to deposit uninhibited. Deposition ofsublytic quantities of complement proteins canpotentially increase endothelial binding of in¯am-matory cells expressing surface complementreceptors, and the cleavage products fromresidual complement activity can enhance localin¯ammation [6].
Other antibody-mediated mechanisms mayalso contribute to AVXR. Circulating leukocytespossessing Fc receptors, such as NK cells,monocytes, macrophages, neutrophils and plate-lets, can bind to the Fc region of endothelialbound xenoantibodies. These interactions canresult in direct endothelial injury by ADCC-related mechanisms [18], and the local release ofproin¯ammatory cytokines and procoagulantfactors [40] can predispose to thrombosis.
Also, xenoantibody binding alone can activateendothelial cells inducing proin¯ammatory andprocoagulant changes [31,32,41] to the endothe-lial surface. Up-regulation of E-selectin andVCAM [31]; down-regulation of thrombomodu-lin (TM) [41], release of IL-1a, IL-8 [39], tissuefactor (TF) [42], and plasminogen activatorinhibitor type 1 (PAI-1)[43] are among thechanges associated with endothelial activationthat could predispose to thrombosis.
The Gala1,3Galb1,R epitope (aGal) [1,2] isthe primary porcine endothelial target of humanpreformed xenoantibodies, and may also be theprimary target of induced antiporcine antibodies[44]. Furthermore, human NK cells and mono-cytes have been shown to bind directly to aGalantigens [17,20], thereby contributing to DXR.Although the exact mechanism of DXR hasnot been determined, as described above, mostof the suspected contributing mechanismsinvolve interactions with aGal. These mechan-isms, and therefore possibly DXR, may beinhibited by blocking all potential interactionswith aGal.
With the exception of humans and old worldmonkeys, all mammals constitutively expressaGal antigens on many of their tissues. We,and others have demonstrated that aGal antigenexpression varies depending on the tissue [10], thespecies [24], and it can even vary betweenindividuals of the same species [45]. However, itis not known whether organs expressing loweramounts of aGal antigen will have any survivaladvantage over those expressing higher amountsof antigen.
Although knock-out of the aGal gene has beenaccomplished in mice [38], it is currently notpossible in pigs because they have no establishedembryonic stem cell lines. A promising approachthat has been achieved in both pigs and mice isthe insertion of genes responsible for productionof fucosyl-transferase [11]. This enzyme success-fully competes for the same substrate as a-galactosyl-transferase and therefore divertsendothelial terminal carbohydrate antigen synth-esis towards production of H antigen, which isnot a target of human antibodies. Both of these
Fryer et al.
106

transgenic approaches used alone or in combina-tion in mice have not imparted any greaterxenograft survival advantage than that achievedwith transgenic complement regulatory proteins[46]. Pigs made transgenic for fucosyl-transferaseare also being developed, but it is not yet knownwhether their organs will survival longer thanorgans from pigs transgenic for human comple-ment regulatory proteins.
The results of these studies suggest thatalthough aGal is the most signi®cant antigenictarget in pig-to-human xenotransplantation it isnot the only one. In fact, it has been noted thatother pre-terminal carbohydrates, which are alsotargets of human xenoantibodies, are exposedwhen aGal is knocked out [1]. Proteins expressedon the PAEC surface likely induce antibodyresponses that contribute to the overall increasein anti-PAEC antibodies after xenotransplanta-tion. Thus, successful strategies may have toaddress interactions with endothelial antigensother than aGal.
If a complete inhibition of interactions with allendothelial antigens were achievable, it is notknown how long this would be necessary tosuccessfully avert DXR. Experience from ABOincompatible kidney allotransplantation, sug-gests that after a ®nite period of time, the graftbecomes resistant to antibody mediated rejection[47]. This phenomenon, known as `accommoda-tion' [48], has not been as clearly documented inxenograft models [49], although recent in vitroexperiments using PAECs have demonstratedthat resistance to complement-mediated lysis canbe induced after a 5-day incubation with anti-PAEC IgG antibodies [31]. Although themechanism of accommodation is not known, aninduction of cytoprotective genes has beenimplicated [40].
Blocking antibodies against aGal may pro-vide a ®nite period of protection for theendothelium by inhibiting the interactions thatcontribute to vascular rejection. To be effective,blocking antibodies would have to prevent theinteraction of all other ligands with the targetantigen without inducing proin¯ammatory/pro-coagulant changes in the target cell or inter-acting with other blood elements that couldinduce local proin¯ammatory/procoagulantchanges.
IgG may function as a blocking antibody forIgM as it has been shown that administration ofhuman IVIG prevent HAR of porcine xenograftsby primates [50]. IgG and IgM have similaraf®nities for aGal [51] and therefore IVIG, aconcentrated human IgG preparation containinganti aGal antibodies, will compete with IgM for
aGal binding sites. As IgM is much more ef®cientin activating the complement cascade and therebyinitiating HAR, its displacement by IgG couldslow the pace of a vascular rejection process. Thegraft loss that subsequently occurs may thereforeresult from a slower paced complement mediatedinjury facilitated by the IgG subtypes that dobind complement (IgG1, IgG3) or to an ADCC-related mechanism because IgGs bind Fc recep-tors better than IgM.
Avian species do not express aGal on theirendothelial surface and therefore, like humans,they have pre-formed anti-aGal antibodies.Since, owing to their structural differences,avian IgY antibodies bind neither human com-plement components nor human Fc receptors[28], anti-aGal IgYs are good candidates for useas blocking antibodies to inhibit the interactionsthat probably contribute to AVXR.
Antiavian antibody development will be afactor in determining how long a blockingstrategy will be effective. As with murine anti-bodies, it is possible that avian antibodies couldbe humanized to reduce their immunogenicity,although this may be counter-productive if theydo not retain the structure that prevents theirinteraction with complement or Fc receptors.
Unlike human anti-PAEC antibodies, aviananti-PAEC antibodies do not cause endothelialactivation. This is important, because ongoingendothelial activation may play a role in AVXR.Therefore, even if all other direct or indirectinteractions with the PAECs are prevented,AVXR may still occur if the blocking antibodiesinduce endothelial activation.
Long-term administration of IgY antibodieswill not be practical because they will certainly beimmunogenic. Also, they will need to be modi®edto bind more effectively and thereby avoid theneed to infuse massive amounts to successfullycompete for binding sites with recipient xenoan-tibodies. Additional studies are planned todetermine whether, as has been demonstratedwith human antiendothelial Igs [31,32], IgYsinduce protective changes in PAECs that mayobviate the need for their long-term administra-tion.
In summary, these studies show that avian IgYantibodies block human xenoantibody binding torat and porcine tissues. IgY can also bind PAECswithout activating them, and can thereby inhibitrecruitment of the complement system andin¯ammatory cells. These avian antibodies needto be evaluated in both small and large animalxenotransplant models to determine their phar-macokinetics, toxicity and ef®cacy.
Blocking of human antiporcine xenoantibody binding
107

References
1. SANDRIN MS, MCKENZIE IFC. Gala(1,3)Gal, the majorxenoantigen(s) recognized in pigs by human naturalantibodies. Immunol Rev 1994; 141: 169.
2. PALMETSHOFER A, GALILI U, DALMASSO AP, ROBSON SC,BACH FH. aGalactosyl epitope-mediated activation ofporcine aortic endothelial cells: type I activation.Transplantation 1998; 65: 844.
3. MCMORROW IM, COMRACK CA, SACHS DH, DERSIMONIAN
H. Heterogeneity of human anti-pig natural antibodiescross-reactive with the Gal(a1,3) Galactose epitope.Transplantation 1997; 64: 501.
4. LEVENTHAL JR, DALMASSO AP, SUN LH, REIF S, MATAS
AJ, PLATT JL. Immunopathology of guinea pig cardiacxenograft rejection. Transplantation 1994; 56: 363.
5. HANCOCK WW, BACH FH. The immunopathology ofdiscordant xenograft rejection. Xeno 1994; 2: 68.
6. BALDWIN WM III, PRUITT SK, BRAUER RB, DAHA MR,SANFILIPPO F. Complement in organ transplantation.Transplantation 1995; 59: 797.
7. DALMASSO AP. The complement system in xenotrans-plantation. Immunopharmacology 1992; 24: 149.
8. LEVENTHAL JR, DALMASSO AP, CROMWELL JW, PLATT JL,MANIVEL CJ, BOLMAN RM III, MATAS AJ. Prolongationof cardiac xenograft survival by depletion of comple-ment. Transplantation 1993; 55: 857.
9. CARRINGTON CA, RICHARDS AC, vAN DEN BOGAERDE J,TUCKER AW, WHITE DJG. Complement activation, itsconsequences, and blockage by gene transfer. WorldJ Surg 1997; 21: 907.
10. COOPER DKC. Xenoantigens and xenoantibodies.Xenotransplantation 1998; 5: 6.
11. SANDRIN MS, FODOR WL, MOUHTOURIS E, OSMAN N,COHNEY S, ROLLINS SA, et al.. Enzymatic remodeling ofthe carbohydrate surface of a xenogeneic cell substan-tially reduces human antibody binding and complementmediated cytolysis. Nature Medicine 1995; 1: 1261.
12. LAVECCHIO JA, DUNNE D, EDGE ASB. Enzymatic removalof a-galactosyl epitopes from porcine endothelial cellsdiminishes the cytotoxic effect of natural antibodies.Transplantation 1995; 60: 841.
13. SIMON PM, NEETHLING FA, TANIGUCHI S, GOODE PL,ZOPF D, HANCOCK WW, et al. Intravenous infusionof Gal(a1,3)Gal oligosaccharides in baboons delayshyperacute rejection of porcine heart xenografts.Transplantation 1998 65: 346.
14. XU Y, LORF T, SABLINSKI T, GIANELLO P, BAILIN M,MONROY R, et al. Removal of anti-porcine naturalantibodies from human and nonhuman primate plasmain vitro and in vivo by a Gala1±3Galb1±4bGlc-Ximmunoaf®nity column. Transplantation 1998; 65: 172.
15. LIN SS, WEIDNER BC, BYRNE GW, DIAMOND LE, LAWSON
JH, et al. The role of antibodies in acute vascularrejection of pig-to-baboon cardiac xenotransplants.J Clin Invest 1998; 101: 1745.
16. HANCOCK WW. Delayed xenograft rejection. WorldJ Surg 1997; 21: 917.
17. INVERARDI L, CLISSI B, STOLZER AL, BENDER JR, SANDRIN
MS, PARDI R. Human natural killer lymphocytesdirectly recognize evolutionary conserved oligosac-charide ligands expressed by xenogeneic tissues.Transplantation 1997; 63: 1318.
18. SCHAAPHERDER AF, DAHA MR, TE BULTE MT, vAN DER
WOUDE FJ, GOOSZEN HG. Antibody-dependent cell-mediated cytotoxicity against porcine endothelium
induced by a majority of human sera. Transplantation1994; 57: 1376.
19. FRYER JP, CHEN S, JOHNSON E, SIMONE P, SUN LH,GOSWITZ JJ, MATAS AJ. The role of monocytes andmacrophages in delayed xenograft rejection.Xenotransplantation 1997; 4 (1): 40.
20. ITESCU S, KWIATOWSKI P, WANG SF, BLOOD T, MINANOV
OP, ROSE S, MICHLER R. Circulating human mono-nuclear cells exhibit augmented lysis of pig endotheliumafter activation with interleukin-2. Transplantation1996; 62: 1927.
21. PALMETSHOFER A, GALILI U, DALMASSO AP, ROBSON SC,BACH FH. a-Galactosyl epitope-mediated activation ofporcine aortic endothelial cells-type I activation.Transplantation 1998; 65: 844.
22. PALMETSHOFER A, ROBSON SC, BACH FH. Tyrosinephosphorylation following mediated endothelial cellstimulation. Xenotransplantation 1998; 5: 61.
23. SAADI S, PLATT JL. Transient perturbation of endothelialintegrity induced by natural antibodies and comple-ment. J Exp Med 1995; 181 (1): 21.
24. ORIOL R, CANDELIER JJ, TANIGUCHI S, PETERS L, COOPER
Dkc. Major oligosaccharide epitopes found in tissues of23 animal species: Potential donors for organ xeno-transplantation. Transplant Proc 1996; 28: 794.
25. BOUHOURS JF, RICHARD C, RUVOEN N, BARREAU N,NAULET J, BOUHOURS D. Characterization of a polyclonalanti-Gala1±3Gal antibody from chicken.Glycoconjugate J 1998; 15: 93.
26. SVENDSEN L, CROWLEY A, OSTERGAARD LH, STODULSKI G,HAU J. Development and comparison of puri®cationstrategies for chicken antibodies from egg yolk. LabAnimal Sci 1995; 45: 89.
27. HATTA H, KIM M, YAMAMOTO T. A novel isolationmethod for hen egg yolk antibody, `IgY'. Agricul BiolChem 1990; 54: 2531.
28. LARSSON A, SJOQUIST J. Chicken IgY: utilizing theevolutionary difference. Comp Immunol Microbioland Inf Dis 1990; 13: 199.
29. ANDERAS PR, PAUL D, TOMITA JT, ANDERSON B.Preparation and speci®cities of antisera to b1,4 linkedN-acetyl-D-glucosamine oligosaccharides. Mol Immunol1979; 16: 341.
30. DUBOIS M, GILLES KA, HAMILTON JK, REBERS PA, SMITH
F. Colorimetric method for determination of sugars andrelated substances. Anal Chem 1956; 28: 350.
31. DORLING A, STOCKER C, TSAO T, HASKARD DO, LECHLER
RI. In vitro accommodation of immortalized porcineendothelial cells. Transplantation 1996; 62: 1127.
32. DALMASSO AP, HE T, BENSON BA. Human IgMxenoreactive natural antibodies can induce resistanceof porcine endothelial cells to complement mediatedinjury. Xenotransplantation 1996; 3: 54.
33. VANHOVE B, DE MARIN R, LIPP J, BACH F. Humanxenoreactive natural antibodies of the IgM isotypeactivate pig endothelial cells. Xenotransplantation 1994;1: 17.
34. YOUNG DS, KADOKURA M, BROCKHAUSEN I, KASHEF V,COLES JG. Human serum induced porcine endothelialcell apoptosis-another pathway to xenograft rejection.Transplant Proc 1996; 28: 611.
35. LEVENTHAL JR, JOHN R, FRYER JP, WITSON JC, DERLICH
JM, REMISZEWSKI J, et al. Removal of baboon andhuman antiporcine IgG and IgM natural antibodies byimmunoadsorption. Transplantation 1995; 59: 294.
36. WHITE DJ. HDAF transgenic pig organs: are theyconcordant for human transplantation? Xeno 1996; 4: 50.
Fryer et al.
108

37. SACHS DH, SABLINSKI T. Tolerance across discordantxenogeneic barriers. Xenotransplantation 1995; 2: 234.
38. TEARLE RG, TANGE MJ, ZANNETTINO ZL, KATERELOS M,SHINKEL TA, VANDENDEREN BJ, et al. The a1-3galacto-syltransferase knockout mouse: implications for xeno-transplantation. Transplantation 1996; 61: 13.
39. PLATT JL, LIN SS, MCGREGOR Cga. Acute vascularrejection. Xenotransplantation 1998; 5: 168.
40. BACH FH, FERRAN C, SOARES M, WRIGHTON CJ,ANRATHER J, WINKLER H, et al. Modi®cation of vascularresponses in xenotransplantation: in¯ammation andapoptosis. Nature Med 1997; 3: 944.
41. LAWSON JH, DANIELS L, PLATT JL. The evaluation ofthrombomodulin activity in porcine to human xeno-transplantation. Transplant Proc 1997h; 29: 884.
42. KOPP CW, SIEGEL JB, HANCOCK WW, ANRATHER J,WINKLER H, GECZY C, et al. Effect of porcine endothelialtissue factor pathway inhibitor on human coagulationfactors. Transplantation 1997; 63: 749.
43. BLAKELY ML, vAN DER WERF WJ, BERNDT MC, DALMASSO
AP, BACH FH, HANCOCK WW. Activation of intragraftendothelial and mononuclear cells during discordantxenograft rejection. Transplantation 1994; 58: 1059.
44. COTTERELL AH, COLLINS BH, PARKER W, HARLAND RC,PLATT JL. The humoral response in humans followingcross-perfusion of porcine organs. Transplantation1995; 60: 861.
45. ALVARADO CG, COTTERELL AH, MCCURRY KR, COLLINS
BH, MAGEE JC, BERTHOLD J, et al. Variation in the
level of xenoantigen expression in porcine organs.Transplantation 1995; 59: 1589.
46. COWAN PJ, CHEN CG, SHINKEL TA, FISICARO N, SALVARIS
E, AMINIAN A, et al. Knock out of a1,3galactosyltrans-ferase or expression of a1,2fucosyltransferase furtherprotects CD55- and CD59-expressing mouse hearts inan ex vivo model of xenograft rejection. Transplantation1998; 65: 1599.
47. ALEXANDRE GP, SQUIFFLET JP, DEBRUZERE M, LATINNE D,REDING R, GIANELLO P, et al. Present experiences in aseries of 26 ABO-incompatible living donor renalallografts. Transplant Proc 1987; 19: 4538.
48. WINKLER H, FERRAN C, BACH FH. Accommodation ofxenografts: a concept revisited. Xenotransplantation1995; 2: 53.
49. FISCHEL RJ, PLATT JL, MATAS AJ, PERRY E, DALMASSO
AP, MURIEL C, et al. Prolonged survival of a discordantcardiac xenograft in a rhesus monkey. Transplant Proc1991; 23: 59.
50. MAGEE JC, COLLINS BH, HARLAND RC, LINDMAN BJ,BOLLINGER RR, FRANK MM, PLATT JL. Immunoglobulinprevents complement mediated hyperacute rejectionin swine to primate xenotransplantation. J Clin Invest1995; 96: 2404.
51. YU PB, HOLZKNECHT ZE, BRUNO D, PARKER W, PLATT JL.Modulation of natural IgM binding and complementactivation by natural IgG antibodies. Journal ofImmunology 1996; 157: 5163.
Blocking of human antiporcine xenoantibody binding
109





























