Embed Size (px)
Citation preview

Survey, Molecular Systematics and Nanobiotechnological Potentials of Marine Cyanobacteria and Diatom
Thesis submitted to Bharathidasan University
For the award of the degree of
DOCTOR OF PHILOSOPHY IN MICROBIOLOGY
By
Mr. D. MUBARAK ALI
(Ref. No. 022323/Ph.D1/Microbio/FT/Oct 2008)
Under the tutelage of
Dr. N. THAJUDDIN
Department of Microbiology School of Life Sciences
Bharathidasan University Tiruchirappalli – 620024
Tamil Nadu, India
May 2012

Dr. N. THAJUDDIN Associate Professor
UNIVERSITY DEPARTMENT OF MICROBIOLOGY
SCHOOL OF LIFE SCIENCES
21.05.2012
Certificate
This is to certify that this thesis entitled “Survey, Molecular Systematics and
Nanobiotechnological Potentials of Marine Cyanobacteria and Diatom” submitted
by Mr. D. Mubarak Ali, for the degree of Doctor of Philosophy in Microbiology, to
the Bharathidasan University is based on the results of studies carried out by him
under my guidance and supervision. This thesis or any part of thereof has not been
submitted anywhere else for the award of degree, diploma, Associateship, fellowship
or other similar titles to any candidates.
(N. THAJUDDIN)
Tiruchirappalli – 620 024, India, Phone: +91 431 2407082; Mobile +91 098423 79719; E-mail: [email protected]

Mr. D. MUBARAK ALI
Doctoral Research Scholar & CSIR – SRF
DECLARATION
I do hereby declare that this work has been originally done by me under the tutelage
of Dr. N. Thajuddin, Associate Professor, Department of Microbiology, Bharathidasan
University, Tiruchirappalli and this work has not been submitted elsewhere for any other
degree, diploma or other similar titles.
(D. MUBARAK ALI)

Dedicated this thesis to Almighty…
This is not the end; this is not even beginning of the end; but it perhaps. This
is end of the beginning. - Winston Churchill

ACKNOWLEDGEMENT
“Omnipotent and Omnipresent, in the of name of Allah”
The writing of this thesis has been an incredible journey and a monumental milestone in my academic life. I could not have embarked on this expedition and traveled this far without the passionate and continued support of advisors, colleagues, friends and my family.
This is my respectful commitment to thank Dr. N. Thajuddin, Associate Professor, Department of Microbiology, Bharathidasan University, Tiruchirappalli for his responsibility of guidance and supervision in my research carrier. His independency on my research work, friendly approaches and his kind support are the secret of successful completion of my work on time. He is a great ambassador of real meaning of guide and supervisor and also thank his family for its love and affection.
Dr. Maggy F. Lengke, Professor, Department of Earth Sciences, University of Western Ontario, London, helped me in many ways to enable and shape my work and continued to provide me firm support and constructive feedback along the way. For that, I will always be grateful.
Dr. S. Maruthamuthu, Scientist, Central Electrochemical Research Institute (CECRI), Karaikudi, India. He is the one who initiated my research on this topic at first and I thank him for his moral support.
Dr. K. Jeganathan Associate Professor, Co-ordinator, Centre for Nanoscience and Nanotechnology, BDU. Its not only my doctoral committee member, but also has enlightened me in so many aspects about innovations, ideas and other academic matters. He is a wonderful guide and ally whom one has the good fortune to know.
I wish to express my sincerest thanks to Dr. K. Natarajaseenivasan, Associate Professor and Head, Dr. V. Rajeshkannan, Dr. G. Muralitharan and Dr. D. Dhanasekaran, Assistant Professors, Department of Microbiology and Dr. Joseph Selvin, Professor and Head, Department of Bioinformatics, BDU for their support and encouragement.
This is my duty to thank Mr. Vincent, St. Joseph College, Trichirappalli, Dr. S. Shanthi and Dr. Sivakumar, Annamalai University, Chithambaram, Mr. S. Ravishanker, Central Electro Chemical Research Institute, Karaikudi, Mrs. Dr. Pushpa Viswanathan, Cancer Institute, Chennai for their timely help and providing me instrumentation facility.
I wish to record my heartful thanks to R. Mahesh, Post. Doc. Kyung Hee University, South Korea, Dr. Kasthuri, Post Doc, and Dr. M. I. Mohd Ershath, Post Doc. National University of Singapore, Singapore. Dr. Mr. M. Rahuman Sheriff, Researcher, Max Planck’s Institute of Molecular Physiology, Germany, Mr. K. Harish Nag, Researcher, Institute of Botany and Microbiology, Belgium, Mr. Satish,

Researcher, Delhi University, New Delhi and Mr. S. Chandru, Researcher, New Delhi for their encouragement and timely help whenever I approached.
My sincere gratitude also goes to Professors Dr. C. Thangamuthu and Dr. M. Ponnavaikko, Former Vice Chancellors, Bharathidasan University, Dr. G. Subramanian, Founder and former Director, NFMC, Dr. M. A. Akbersha, Director, Mahatma Gandhi Dorenkamp Centre (MGDC), Dr. M. Gunasekaran, Professor in Biology, Fisk University, USA, Dr. L. Uma, Professor and Director, NFMC, Dr. D. Prabakaran Associate Professor, NFMC, Dr. A. Pannerselvam, Associate Professor, Dept. of Botany and Microbiology, A.V.V.M Sri Pushpam College, Tanjore, Dr. K. Kathiresan, Professor, CAS in Marine Biology, Annamalai University, Chithambaram, Dr. S. Ravikumar, Dept. of Oceanography and CAS, Alagappa University, Karaikudi, Dr. N. Anand, Professor and Dr. Rengasamy, Professor and Director, CAS in Botany, University of Madras, Chennai and Dr. B. G. Raghavan, Chennai without their encouragement I could not have come this far.
I also express my thanks to Dr. M. B. Viswanathan, Professor, Dept. of Plant Science, Dr. S. Ravikumar, Asst Professor, Dept. of Biochemistry, Dr. P. Santhanam Asst. Professor, Dept. of Marine Sciences, BDU, our collaborators Dr. N. Rameshbabu and Dr. C. Velmathi Asst. Professors, National Institute of Technology, Trichirappalli and Dr. Prabakaran, Sri Gowri Biotech, Tanjore for their constant support and encouragement.
I express my heartfelt thanks to my initial advisors, Mr. K. Kandasamy, Mr. A. K. Saravanan, Mr. S. Anbalagan, Mrs. S. Shahitha Begam, Ms. Nashima Begam, Mrs. Stella Baby, Mr. Dhanasekaran and Librarians from Muthayammamal College of Arts and Sciences, Rasipuram, Nammakkal for their encouragement and proper guidance.
I would also like to thank the non-teaching technical community, especially Mrs. M. Umarani, Mr. S. Elangovan, Mr. K. Vijayakumar, Mr. Arumugam, Mr. G. Durairajan, Mr. G. Pazhanivel, Mr. K. Gobikanna, Mr. S. Kathiresan from Department of Microbiology, Mr. Stephen, Mrs. Malathy, Mr. Kaviyarasan, Admn building, BDU, among many others, for their professional and administrative backup and for securing a place where we can rest our fatigued minds.
A book with thousand pages starts from single dot. This is my intense to thank my teacher, Mrs. G. Porselvi Prakasham, G. B. H. S. School, Thammampatti (1999-2003). She was the one, who sowed research seed in my mind during my school hood and I also thank to Dr. N. Geetha Achunathan for her motivation and encouragement from my childhood on wards.
I express my heartfelt thanks to Ms. C. Divya, Mr. M. Jayarajan, Mrs. Mari Nivetha, Mrs. T. Shenbagavalli and Mr. D. Pandiaraj, Researchers, Dept of Microbiology, BDU Mr. V. Gopinath, Researcher, SRM University, Chennai and

Mr. K. Venteshwarlu, Researcher, National Institute of Technology, Trichirappalli for their moral support and care.
I place my heartfelt thanks to Mr. R. Praveen Kumar for his cooperation in research and his love and affection towards me and my work and also I place heartfelt thanks to Mrs. A. Ilavarasi Krishnan for her inevitable support and encouragement.
I cannot overlook the warmth that my labmates gave to me and the humour and smiles of friendship that they brought to me; Dr. C. Nithya, Dr. M. Shatheesh Kumar, R. Arvind Kumar A. Suresh, D. Vijayan, A. Parveez Ahamed, M. Abhijith, N. Reehana, the list is endless…
I place my sincere thanks to juniors M. Leo Antony, K.A. Sheik Syed Ishack, K. Kannan, E. Baldev, F. Lewis Oscar, D. Sanker Ganesh, C. Anchana Devi for their constant encouragement and love and affection towards me and my work.
I wish to record the support from my friends Mr. J Arunkumar, Mr. P. Prabakaran, Mr. M. Sundaravadivel, Mrs. S. Manupriya, Mrs. Sudha, Mr. T. Vinodh Kumar, Mr. V. Purusothaman Mr. K. P. Prakash, Mr. R. Suresh, Mr. R.V., Dr. M. Jaccob, and Mr. S. Karthik for their love cheered me on.
This is my immense pleasure to thank my fine arts masters Mr. M. Saravanan, Dr. C. Lalithambal, Mrs. S. Sri Vidya and Mr. R. Venkatesh for their encouragement and timely help. I place my sincere thanks to Mr. Abdul Azees, Karaikudi for his timely help and love and affection.
As the whole, my heartfelt thanks to all my seniors, juniors, colleagues, Ph.D and M. Phil Scholars of Dept. of Microbiology, Eco-Biotechnology, Env. Management, Biotechnology, Marine Biotechnology, Animal Biotechnology, Plant Biotechnology, Marine Sciences, Bioinformatics, Biochemistry, Biomedical Sciences, Physics, Chemistry, Geography and English BDU for their prayers and wishes which made me complete this thesis well and good.
I gratefully acknowledge the financial support offered by the Council for Scientific and Industrial Research (CSIR-SRF Fellowship), Department of Biotechnology (DBT-JRF Fellowship) and University Grant Commission (UGC-PF), Govt. of India, funded my research work and I also thank Department of Science and Technology (ITS), INSA–CICS (TG) for their travel grant to participate in the overseas conferences at South Korea and Singapore respectively and made my stay in the Bharathidasan University possible and to the Staff at Vaigai, Porunai and Cauvery Hostel, who, though were so willing all the time, to attend to my needs any way.
As personal note, I thank everyone in my family and PG, UG and School friends for their endless support, encouragement, care, love and affection. Because of them only, my work sailed in smooth way.
To all, I say, thank you and God bless!!!
Mubarak Ali, D
Contents Page No.
Survey and Molecular Systematics of marine Cyanobacteria and Diatom
Chapter I
Introduction 1
Review of Literature 6
Materials and Methods 36
Results and Discussion 42
Partial Biochemical Characterization of C-Phycobiliproteins
Chapter II
Introduction 47
Review of Literature 50
Materials and Methods 60
Results and Discussion 64
Nanobiotechnological Potentials of Marine Cyanobacteria and Diatom
Chapter III
Introduction 72
Review of Literature 78
Materials and Methods 101
Results and Discussion 107
Annexure
Summary 124
References 126
List of GenBank Accession (NCBI) 162
Publications

Chapter I
Survey and Molecular Systematics of Marine Cyanobacteria and Diatoms

1
INTRODUCTION
As primary producers in the worlds oceans, they are keystone species in global
nutrient cycling. About 20% of the total carbon and silica sequestered are fixed by less
than a few hundred species in the marine plankton (Guillard and Kilham 1978;
Goldman 1993; Hasle and Syversten 1996; Mann 1999). Knowledge of the geological
age of the diatoms gives palaeoclimatologists a clue as to how long these organisms
have been modifying the biosphere (Kooistra and Medlin 1996; Siever 1991). The cell
wall structure is species specific, demonstrating that diatom silica morphogenesis is
genetically encoded.
Cyanobacteria are prokaryotic oxygenic phototrophs found in almost every
conceivable habitat on earth (Ferris et al. 1996; Ward et al. 1997; Nubel et al. 1999,
2000; Abed and Garcia-Pichel 2001; Garcia-Pichel and Pringault 2001). They exist in
different morphologies including unicellular and filamentous forms (Castenholz 2001).
While unicellular types exist as single cells, suspended or benthic, or aggregates,
filamentous types may be thin or thick, single trichome or bundles either with or
without a sheath. Cyanobacteria are able to perform different modes of metabolism
with the capacity to switch from one mode to another (Stal, 1995).
All cyanobacteria carry out oxygenic photosynthesis but some cyanobacterial
species can switch to the typical bacterial anoxygenic photosynthesis using sulfide as
electron donor (Cohen et al. 1986). Under anoxic conditions and during the dark,
cyanobacteria carry out fermentation (Stal 1997). Some cyanobacteria form heterocysts
and have the ability to fix atmospheric nitrogen (Capone et al. 2005). Phylogenetic
analysis of cyanobacteria based on 16S rDNA genes showed that they are a diverse,
monophyletic phylum of organisms within the bacterial radiation. Research on
cyanobacteria in the last decades focused largely on their ecology, morphology,
physiology and 16S rDNA-based phylogeny but relatively little has been done on their
potential uses in biotechnology. The overwhelming available knowledge on the
diversity and physiology of cyanobacteria serves as an excellent base for exploring
their applications in biotechnology (Thajuddin and Subramanian, 2005).

2
In the last few years, cyanobacteria have gained much attention as a rich source
of bioactive compounds and have been considered as one of the most promising groups
of organisms to produce them (Bhadury et al. 2004; Dahms et al. 2006). These
cyanobacterial metabolites include antibacterial (Jaki et al. 2000 and MubarakAli et al.,
2008), antifungal (Kajiyama et al. 1998), antiviral (Patterson et al. 1994), anticancer
(Gerwick et al. 1994), antiplasmodial (Papendorf et al. 1998), algicide (Papke et al.
1997) and immunosuppressive agents. Screening of cyanobacteria for antibiotics has
opened a new horizon for discovering new drugs. Some cyanobacteria have also been
found to intracellularly accumulate polyhydroxyalkanoates (PHA), which are
comparable in properties to polyethylene and polypropylene (Steinbuchel et al. 1997).
These biodegradable plastics could replace oil-derived thermoplastics in some fields.
Recent research on cyanobacteria has demonstrated that they form ideal consortia with
chemotrophic bacteria and can effectively be used to cleanup oil contaminated
sediments and wastewaters (Abed and Koster 2005).
Cyanobacterial taxonomy has been established based on morphological features,
such as the shape and dimensions of the cells, presence of structurally differentiated
cells, and whether the cells grow as solitary cells or in colonies (Paerl, 1988).
Molecular methods have become an indispensable tool for characterization of
cyanoprokaryotes and the assessment of evolutionary relations among them in recent
decades. The direct sequencing of various genes is the most common method used in
taxonomy of cyanobacteria. However, RFLP (Restriction Fragment Length
Polymorphism) is also widely applied, especially for more detailed examination of the
genetic variability of closely related taxa (Ernst et al., 1995) or to infer the extent of
cyanobacterial diversity in nature. Also random amplified polymorphic DNA (RAPD)
analysis is sometimes used in order to discriminate between genotypes of close
relatives. Much less common are the allozyme or the whole-cell protein analysis
(Neilan, 1995).
Diatoms are single celled photosynthesizing eukaryotic algae that produce
intricately structured cell wall made of nanopatterned silica (SiO2) and are found in

3
almost every aquatic environment including fresh and marine water, soil, in fact almost
anywhere in moist. They are non motile, or capable of only limited movement along a
substrate by secretion of mucilaginous material along a slit like groove or channel
called a raphe, both benthic and planktonic forms exist. Diatoms are formally classified
as belonging to the division Chrysophyta, Class Bacillariophyceae. The Chrysophyta
are algae which form endoplasmic cysts, store oil rather than starch, posses a bipartite
cell wall and secrete silica at some stages of their life cycle. Diatoms are nearly all
autotrophic, heterotrophic members are rare (Li and Volcani 1987).
Diatoms are commonly between 20-200 microns in diameter or length, although
sometimes they can be up to 2 millimeter long first appeared more than 180 million
years ago. First recorded occurence of diatoms are from Jurassic however, these are
uncertain and the earliest recorded well preserved diatoms are centric forms from
Aptian-albian stages of the Cretaceous. At first diatoms are thought to be animals
because what are now called raphid diatom move at a speed of upto 25µms-1 when
attached to surfaces. Chris Bowler (2001) explains that molecular phylogeny and
morphological studies suggest that diatoms originated probably as the result of a
eukaryote being invaded or engulfed by a photosynthetic eukaryote, most probably a
red algae.
Diatoms are phylogenetic similarity to green algae and higher plants is derived
from the primary endosymbiotic event, which is thought to have occurred at least 700
million years ago (Kowallik, 1992). Therefore, diatom cells have a range of features
that make them highly divergent from the classical cellular structure of higher plants,
including:
(a) The use of the brown carotenoid pigment, fucoxanthin for light energy transfer
within the light harvesting complexes of photosystems I and II
(b) The presence of four membranes around their plastids. The inner two
membranes are equivalent to the membranes that normally surround higher
plant chloroplasts, whereas the second membrane (as counted from the outside)

4
is thought to be derived from the endosymbiont’s plasma membrane, and the
outer membrane is continuous with the endoplasmatic reticulum of the host cell.
(c) The presence of a rigid cell wall composed largely of amorphous silica (i.e.
glass). The exquisite lacework-like patterning of diatom cell walls is reproduced
with high fidelity from generation to generation and is species specific. For
these reasons, it has been used since the last century for taxonomic
classification.
Current phylogenetic trees place diatoms close to Alveolata lineages and far
from the green and red lineages of other photosynthetic eukaryotes (Baldauf et al.,
2000). The expressed sequence tags (EST) program has revealed the presence of genes
encoding enzymes involved in cAMP metabolism, as well as genes encoding the
components of the animal extracellular matrix, such as fibronectins, elastins, and
tenascins, none of which appear to be present in higher plants. As a consequence, one
could almost consider diatoms as photosynthetic animals rather than unicellular plants.
The rigid part of the cell wall (frustule) of diatoms is composed of amorphous
silica (Pickett-Heaps et al., 1990; Parkinson and Gordon 1999; Vrieling et al., 2000),
located in the girdle bands and two valves: the epitheca consisting of an epivalve and
epicingulum, and the hypotheca consisting of hypovalve and hypocingulum (Round et
al., 1990). The timing of diatom valve formation has not been directly determined in an
individual cell, however, some estimates were made based on measurements on whole
synchronized cultures. For instance, 2D valve formation in Melosira varians Agardh
takes approximately 8 minutes. Cell wall morphogenesis in Ditylum brightwellii (West)
Grunow, and in Navicula pelliculosa (Kutz) Hilse occurs during respectively a 53 min
and a 2-3 h period of the cell cycle (Sullivan and Volcani, 1981).
Commonly, in pennate diatoms the development of the new hypovalve starts
from the axial area and grows towards the outer cellular edges (Pickett-Heaps et al.,
1990). Subsequently, the frustule thickens in the third dimension and is completed by
coverage with a casing followed by exocytosis of the valve and girdle bands, and

5
finally cell separation (Darley et al. 1976; Darley and Volcani 1971; Pickett-Heaps et
al., 1990).
Application of the M13 PCR fingerprinting method was useful for almost all
forms of cyanobacteria strain and species discrimination. PCR amplification
techniques viz., repetitive DNA element PCR (REP-PCR), short randomly repeated
repetitive PCR (STRR-PCR) and arbitrarily primed PCR (RAPD-PCR) were used for
the taxonomic discrimination among the strains of the unicellular cyanobacterium,
Synechococcus elongatus collected across the coastal regions of the Indian
subcontinent. In comparison with the STRR and RAPD, the REP primer set generates
fingerprints of lower complexity, but still the phenogram clearly differentiated the
strains. In conclusion, described PCR fingerprinting methods can be considered as
promising tools for the differentiation at the strain level of cyanobacteria from the same
species (Muralitharan and Thajuddin, 2010). 151 unique cyanobacterial genes in 8
studied genomes and found a few examples of largely conserved gene order, which
could prove useful for solving problems of cyanobacterial evolution on a larger scale
(Martin et al., 2003).
Molecular approaches have been divided into two classes: PCR independent and
PCR based approaches. Molecular assessment of cyanobacterial biodiversity has been
studied by using markers like 16S rDNA, phycocyanin locus, nif gene, rpo gene, ITS
region etc. A general overview of biodiversity assessment, molecular techniques and
markers used for biodiversity assessment recommends combinatorial approach with
different molecular markers. It is likely to improve the degree of resolution and provide
as possible the broadest picture and in depth information about biodiversity
documentation (Kumari et al., 2009).
Due to innate biotechnological potentials and most abundantly available in
marine ecosystems, both cyanobacteria and diatoms has been selected in the present
study.

6
REVIEW OF LITERATURE
Diatom Biology
Diatoms were first discovered by light microscopy and mistakenly addressed as
unicellular animals due to their brownish color and their capability to move on
substrates (Lind et al., 1997). According to the species concept, organisms mating with
each other belong to the same species. Diatom taxonomy thus was based mainly either
on the identification of ribosomal sequences (Medlin et al., 1996) or more classically
on the morphology and the shape of the frustules, the extracellular silica cell walls.
These frustules, which are formed by two valves that fit together like Petri dishes, often
are highly ornamented and show species-specific structures.
The ability of diatoms to genetically define these structures makes them highly
interesting for nanotechnological applications (Drum and Gordon, 2003). Because of
their siliceous composition frustules were often well preserved in fossil deposits
(Damste et al., 2004) and strong sedimentation of diatom shells in ancient oceans led to
the deposition of siliceous earth or diatomite, a material of high importance for
industrial uses (Harwood, 1999).
Diatoms play a significant ecological role; about half of the global annual net
primary production in the oceans is due to phytoplankton which is dominated by
diatoms (Falkowski et al., 1998). Diatoms thus produce and represent the main input
into the marine food web. They are not only strongly involved in fixation of CO2, but
also in cycling of soluble silicates by integrating them into the shells and by releasing
parts of them after decomposition at the bottom of the oceans (Bidel and Azam, 1999).
It is not yet fully understood why diatoms filled aquatic niches so successfully, even
though they have the capability to grow at a wide range of light intensities (Falkowski
et al., 2004) and the apparent ability to perform C4 photosynthesis (Reinfelder, 2004).
Cell Wall Structure, Cell Cycle and Sexual Reproduction
The diatom cell wall consists of two halves of identical structure, although one
half (epitheca) is slightly larger and overlaps the other half (hypotheca). Together the

7
epitheca and hypotheca completely enclose the protoplast. Each theca is composed of a
valve displaying ornate structures with often elaborate pore patterns, and several girdle
bands exhibiting generally uniform pore patterns. The terminal girdle band of each
theca is termed the pleural band and is located in the region where the epitheca and the
hypotheca overlap (Fig 1).
Fig 1: Schematic overview of the siliceous components diatom cell wall
Diatoms can be divided into three major groups according to their shape and
symmetry of their cell walls: radial centrics, polar centrics, and pennates. Radial
centrics have a petri dish-like architecture with a circular center of symmetry (annulus)
in the middle of the valve, and rows of pores (striae) radiating from the annulus. Polar
centrics have bi or multipolar valves with an elongated or distorted annulus. Pennate
diatoms are bilaterally symmetrical, and instead of an annulus possess a rib (sternum)
running along the longitudinal axis of each valve.
In raphid pennates the sternum contains a slit (raphe) that is interrupted by the
central nodule in the center of the valve. Raphid pennates are able to move on surfaces
by means of traction generated by adhesive mucilage (polysaccharides or
proteoglycans) that is secreted through the raphe (Edgar and Pickett-Heaps, 1984). The
traction force appears to be generated by an actin-myosin based system that is located
8
precisely underneath the raphe on the cytosolic side of the plasma membrane (Poulsen
et al., 1999). Araphid pennates, radial centrics, and polar centrics are nonmotile.
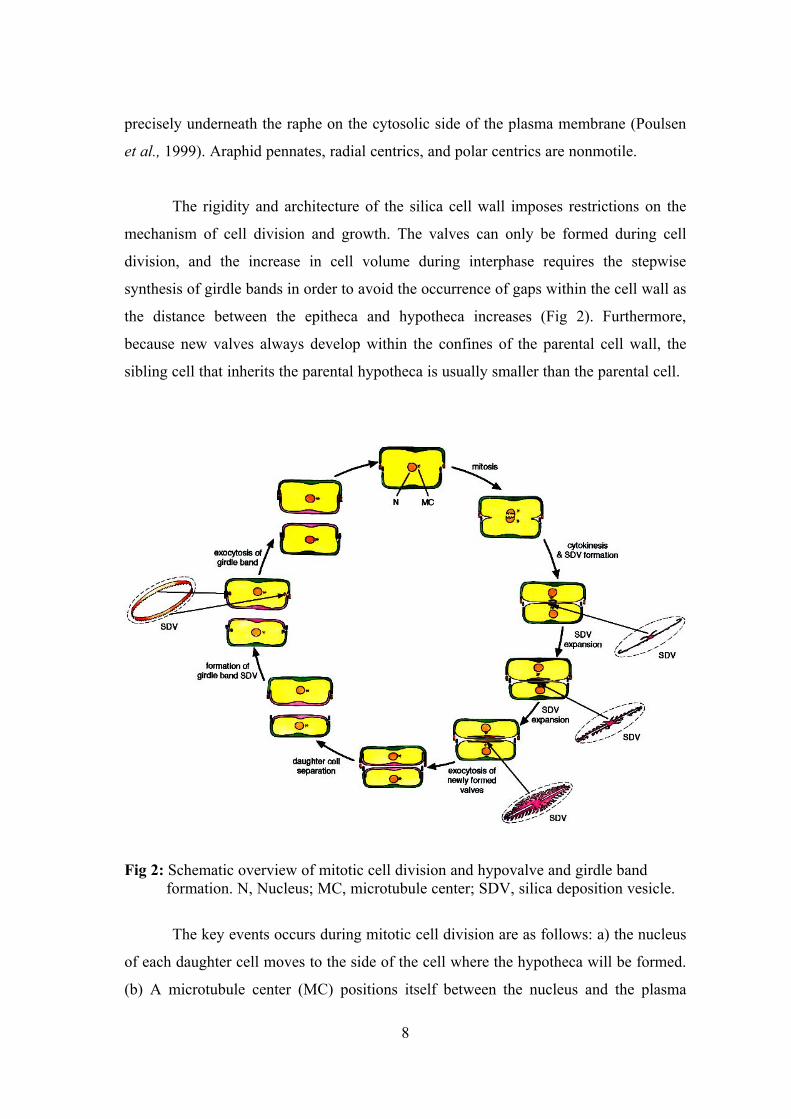
The rigidity and architecture of the silica cell wall imposes restrictions on the
mechanism of cell division and growth. The valves can only be formed during cell
division, and the increase in cell volume during interphase requires the stepwise
synthesis of girdle bands in order to avoid the occurrence of gaps within the cell wall as
the distance between the epitheca and hypotheca increases (Fig 2). Furthermore,
because new valves always develop within the confines of the parental cell wall, the
sibling cell that inherits the parental hypotheca is usually smaller than the parental cell.
Fig 2: Schematic overview of mitotic cell division and hypovalve and girdle band formation. N, Nucleus; MC, microtubule center; SDV, silica deposition vesicle.
The key events occurs during mitotic cell division are as follows: a) the nucleus
of each daughter cell moves to the side of the cell where the hypotheca will be formed.
(b) A microtubule center (MC) positions itself between the nucleus and the plasma

9
membrane above which the hypotheca will eventually be placed. (c) A specialized
vesicle known as the silica deposition vesicle (SDV) forms between the MC and the
plasma membrane in a region that becomes the “pattern center.”(d) The SDV elongates
into a tube and then spreads along one side of the cell. (e) A new valve is formed within
the SDV by the targeted transport of silica, proteins, and polysaccharides. During this
process, the SDV becomes acidic as a result of the silica polymerization process
(Vrieling et al., 1999). Some of the organic components eventually form a coat around
the silica framework, whereas others are involved in silica deposition. (f) Once valve
biogenesis is complete, it is exocytosed by fusion of the SDV membrane (the
silicalemma) with the plasma membrane. As a consequence, the inner face of the
silicalemma is thought to become the new plasma membrane. (g) Following separation,
the daughter cells can expand unidirectionally along the cell division axis by the
biogenesis of girdle bands. These structures are also formed within SDVs in an
analogous way.
As a consequence, the average cell size in a diatom population gradually
decreases with continued vegetative growth (the MacDonald-Pfizer rule). Ultimately,
this size reduction would result in cells too small to be viable and the diatom population
would die. The only way to escape this fate is by sexual reproduction (Chepurnov et al.,
2004). During this process meiotic cell divisions occur, and the resulting gametes shed
their cell walls. Immediately after gamete fusion a specialized zygote (termed
auxospore) is formed, which is covered by an organic cell wall, and undergoes an
enormous increase in volume within a relatively short time (hours to a few days).
Already during the first division of the auxospore, silicified valves are formed
that exhibit the species-specific shape and silica patterns.The molecular details of this
amazing process are almost completely unexplored with the exception of a study by
Armbrust, who discovered a new gene family (Sig) that is expressed during the onset of
sexual reproduction in T. weissflogii (Armbrust, 1999). Sig genes have also been
identified in other Thalassiosira species, yet the function of the Sig-encoded proteins is
still unknown (Armbrust and Galindo, 2001).

10
The size of some of the best-studied diatoms growing in laboratory culture (e.g.,
Cylindrotheca fusiformis, Phaeodactylum tricornutum, Thalassiosira pseudonana)
remains constant, and sexual stages of these organisms have never been observed. The
mechanism by which these species avoid size reduction was not yet fully understood,
but appears to involve the ability to form expandable girdle bands (Hildebrand et al.,
2007).
Organic components of the cell wall
Early light-microscopy studies of diatom cell walls using ruthenium red
presented evidence that organic material is intimately associated with the silica
structures (Pickett-Heaps et al., 1990). Later, imaging of ultrathin-sections after
anhydrous hydrogen fluoride (HF) treatment (i.e., dissolution of silica) followed by
transmission electron microscopy indicated the presence of organic material both on the
silica surface and inside the silica (Pickett-Heaps et al., 1990, Volcani, 1981). Based on
the ruthenium red staining, it was assumed that the silica-associated organic
components are polysaccharides (pectin).
Indeed, to date a plethora of extracellular polysaccharides have been
characterized from diatoms, which appear to play a role in cell adhesion to surfaces,
gliding of pennate diatoms, and protection against desiccation (Hoagland et al., 1993).
While these polysaccharides are relatively loosely attached to the diatom cell wall, and
can often be extracted by treatment with hot water or mildly alkaline solutions,
polysaccharides have recently been characterized that are much more tightly bound to
or even embedded within the diatom silica.
These silica-associated polysaccharides are composed of 3-linked mannans that
are highly polyanionic due to the attachment of numerous uronic acid residues and or
sulfate groups (Chiovitti et al., 2003). It was unclear if the polyanionic mannans are
involved in silica biosynthesis as no functional studies have been performed.
Furthermore, the polyanionic mannans might be protein-linked in vivo, since silica-
embedded highly mannosylated and sulphated glycoproteins have been identified from

11
the diatoms T. pseudonana and C. fusiformis (Poulsen and Kroger, 2003, Poulsen et al.,
2003).
Cyanobacteria
Cyanobacteria or cyanoprokaryota are oxygen producing, photosynthesizing,
gram-negative prokaryotes which had a main part in the evolution of the Earth´s
atmosphere. Cyanobacteria thrive in a wide range of mostly aquatic habitats. The
cellular organization and basic functions regarding growth and photosynthesis have
been comprehensively described (Sandgren, 1988) and since in late 1990s, the
sequencing of entire genomes of or smaller specific regions of within a genome such as
toxin production has allowed more extensive understanding of their unique lifestyle and
metabolic traits (Kaneko et al., 2001).
While cyanobacteria generally have relatively simple basic metabolic
requirements consisting of carbon dioxide, light, water, and inorganic nutrients and
they possess efficient uptake and retention mechanisms for phosphate, nitrate and
bicarbonate. In addition to the above mechanisms, cyanobacteria are capable of
producing high affinity iron chelators, known as siderophores, under iron deficient
conditions to alleviate iron stress and some filamentous cyanobacterial genera possess
heterocysts, cells specialized to fix atmospheric nitrogen. In total, cyanobacteria bear
unique traits that allow them to dominate many phytoplankton communities as a result
of the interactions between their unique physiological traits and the physical and
chemical characteristics of the aquatic system itself (Ritchie et al., 1997).
Biological diversity
Although cyanobacteria probably evolved as a group of organisms about 2,000
million years before the advent of eukaryotes, they comprise fewer taxa than eukaryotic
microalgae (Bisby, 1995). The concept of species in the cyanobacteria has, however, no
distinct boundaries. The situation is similar for most organisms, except for those that
are sexually reproductive depending on the classification system used; the number of
species recognised varies greatly. Based on the International Code of Botanical
Nomenclature the class Cyanophyceae, for example, contains about 150 genera and

12
2,000 species (Hoek et al., 1995). Chemotaxonomic studies include the use of markers,
such as lipid composition, polyamines, carotenoids and special biochemical features.
The resulting data support the more traditional examinations of phenotypic and
ecological characteristics. Physiological parameters are conveniently studied using
laboratory cultures (Packer and Glazer, 1988). The diversity of cyanobacteria can be
seen in the multitude of structural and functional aspects of cell morphology and in
variations in metabolic strategies, motility, cell division, developmental biology, etc.
The production of extracellular substances and cyanotoxins by cyanobacteria illustrates
the diverse nature of their interactions with other organisms (i.e. allelopathy) (Rizvi and
Rizvi, 1992).
A molecular approach to the systematics of cyanobacteria may be most fruitful
for inferring phylogenetic relationships. Macromolecules, such as nucleic acids and
proteins, are copies or translations of genetic information. The methods applied involve
direct studies of the relevant macromolecules by sequencing, or indirectly by
electrophoresis, hybridisation, or immunological procedures (Wilmotte, 1994). Nucleic
acid technologies, especially the polymerase chain reaction (PCR), have advanced to
the point that it is feasible to amplify and sequence genes and other conserved regions
from a single cell. To date, 16S rRNA has given the most detailed information on the
relationships within the cyanobacteria (Rudi et al., 1997). However, the molecular
results obtained should be integrated with other characteristics as the base for a
polyphonic taxonomy (Vandamme, et al., 1996). A considerable morphological, as well
as a genotypical, polymorphy exists in the cyanobacteria, although as data from rRNA
sequencing indicates they are correlated to a high degree.
The phylogenetic relationship of cyanobacteria is the rationale behind the
meaningful systematic groupings. However, it is difficult to set up a system of
classification that serves both the everyday need for practical identification, and offers
an expression of the natural relationship between the organisms in question (Mayr,
1981). Meanwhile, it will be necessary to use the available manuals and reference
books to help in these investigations and with the proper identification of the
cyanobacteria. They are photoautotrophs, cyanobacteria can be grown in simple

13
mineral media. Vitamin B12 is the only growth factor that is known to be required by
some species. Media must be supplemented with the essential nutrients needed to
support cell growth, including sources of nitrogen, phosphorus, trace elements, etc.
Toxigenic strains of cyanobacteria are deposited in international-type culture
collections (Rippka, 1988; Sugawara et al., 1993). Clonal cultures are distributed for
research, taxonomic work and teaching purposes.
Molecular Systematics:
Microbial Systematics has long remained an enigma. Conceptual advances in
microbiology during the twentieth century included the realization that a discontinuity
exists between those cellular organisms that are prokaryotic (i.e. whose cells have no
nucleus) and those that are eukaryotic (i.e. more complexly structured cells with a
nucleus) within the organization of their cells. The microalgae investigated by
phycologists under the International Code of Botanical Nomenclature (ICBN) (Greuter
et al., 1994) included organisms of both eukaryotic and prokaryotic cell types. The
cyanobacteria (Geitler, 1932) constituted the largest group of the latter category. The
prokaryotic nature of these organisms and their fairly close relationship with eubacteria
made work under provisions of the International Code of Nomenclature of Bacteria
(ICNB) (Sneath, 1992) more appropriate (Rippka et al., 1979; Waterbury, 1992).
The prevailing systematic view is that comparative studies of the genetic
constitution of the cyanobacteria will now contribute significantly to the revision of
their taxonomy. Relevant classification should reflect as closely as possible the
phylogenetic. The integration of phenotypic, genotypic and phylogenetic information
renders possible a consensus type of taxonomy known as polyphasic taxonomy
(Vandamme et al., 1996). The names "cyanobacteria" and "blue-green algae"
(Cyanophyceae) are valid and compatible systematic terms. This group of micro-
organisms comprises unicellular to multicellular prokaryotes that possess chlorophyll a
and perform oxygenic photosynthesis associated with photosystems I and II
(Castenholz and Waterbury, 1989).

14
Occurrence in nature
The majority of Cyanobacteria are aerobic photoautotrophs. Their life processes
require only water, carbon dioxide, inorganic substances and light. Photosynthesis is
their principal mode of energy metabolism. In the natural environment, however, it is
known that some species are able to survive long periods in complete darkness.
Furthermore, certain cyanobacteria show a distinct ability for heterotrophic nutrition
(Fay, 1965). Cyanobacteria are often the first plants to colonize bare areas of rock and
soil. Adaptations, such as ultraviolet absorbing sheath pigments, increase their fitness in
the relatively exposed land environment.
Many species are capable of living in the soil and other terrestrial habitats,
where they are important in the functional processes of ecosystems and the cycling of
nutrient elements (Whitton, 1992). The prominent habitats of cyanobacteria are limnic
and marine environments. They flourish in water that is salty, brackish or fresh, in cold
and hot springs, and in environments where no other microalgae can exist. Most marine
forms (Humm and Wicks, 1980) grow along the shore as benthic vegetation in the zone
between the high and low tide marks. The cyanobacteria comprise a large component of
marine plankton with global distribution (Wille, 1904; Gallon et al., 1996).
A number of freshwater species are also able to withstand relatively high
concentrations of sodium chloride. It appears that many cyanobacteria isolated from
coastal environments tolerate saline environments (i.e. are halotolerant) rather than
require salinity (i.e. are halophilic). As frequent colonisers of euryhaline (very saline)
environments, cyanobacteria are found in salt works and salt marshes, and are capable
of growth at combined salt concentrations as high as 3-4 molar mass (Reed et al.,
1984). Freshwater localities with diverse trophic states are the prominent habitats for
cyanobacteria.
Numerous species characteristically inhabit, and can occasionally dominate,
both near surface epilimnic and deep, euphotic, hypolimnic waters of lakes (Whitton,
1973). Others colonize surfaces by attaching to rocks or sediments, sometimes forming
mats that may tear loose and float to the surface. Cyanobacteria have an impressive

15
ability to colonize infertile substrates such as volcanic ash, desert sand and rocks (Jaag,
1945; Dor and Danin, 1996). They are extraordinary excavators, boring hollows into
limestone and special types of sandstone (Weber et al., 1996). Another remarkable
feature is their ability to survive extremely high and low temperatures. Cyanobacteria
are inhabitants of hot springs (Castenholz, 1973), mountain streams (Kann, 1988),
Arctic and Antarctic lakes (Skulberg, 1996a) and snow and ice (Kol, 1968; Laamanen,
1996). The cyanobacteria also include species that run through the entire range of water
types, from polysaprobic zones to katharobic waters (Van Landingham, 1982).
Cyanobacteria also form symbiotic associations with animals and plants.
Symbiotic relations exist with, for example, fungi, bryophytes, pteridophytes,
gymnosperms and angiosperms (Rai, 1990). The hypothesis for the endosymbiotic
origin of chloroplasts and mitochondria should be mentioned in this context. The
evolutionary formation of a photosynthetic eukaryote can be explained by a
cyanobacteria being engulfed and co developed by a phagotrophic host (Douglas,
1994). Fossils of what were almost certainly prokaryotes are present in the 3,450
million year old Warrawoona sedimentary rock of north-western Australia.
Cyanobacteria were among the pioneer organisms of the early earth (Brock
1973; Schopf, 1996). These photosynthetic micro-organisms were, at that time,
probably the chief primary producers of organic matter, and the first organisms to
release elemental oxygen into the primitive atmosphere. Sequencing of
deoxyribonucleic acid (DNA) has given evidence that the earliest organisms were
thermophilic and thus able to survive in oceans that were heated by volcanoes, hot
springs and bolide impacts (Holland, 1997).
Organization, function and behavior:
The structure and organization of cyanobacteria are studied using light and
electron microscopes. The basic morphology comprises unicellular, colonial and
multicellular filamentous forms. Unicellular forms, for example in the order
Chroococcales, have spherical, ovoid or cylindrical cells. They occur singly when the
daughter cells separate after reproduction by binary fission. The cells may aggregate in

16
irregular colonies, being held together by the slimy matrix secreted during the growth
of the colony. By means of a more or less regular series of cell division, combined with
sheath secretions, more ordered colonies may be produced.
A particular mode of reproduction, which may supplement binary fission,
distinguishes cyanobacteria in the order Chamaesiphonales and Pleurocapsales. In the
Chamaesiphonales exospores are budded off from the upper ends of cells. In the second
order, the principal mode of replication is by a series of successive binary fissions
converting a single mother cell into many minute daughter cells (baeocytes or
endospores).
Filamentous morphology is the result of repeated cell divisions occurring in a
single plane at right angles to the main axis of the filament. The multicellular structure
consisting of a chain of cells is called a trichome. The trichome may be straight or
coiled. Cell size and shape show great variability among the filamentous cyanobacteria.
Species in the order Oscillatoriales, with unseriated and unbranched trichomes, are
composed of essentially identical cells. The other orders with a filamentous
organization (orders Nostocales and Stigonematales) are characterized with trichomes
having a heterogeneous cellular composition. Vegetative cells may be differentiated
into heterocysts (having a thick wall and hyaline protoplast, capable of nitrogen
fixation) and akinetes (large thick-walled cells, containing reserve materials, enabling
survival under unfavourable conditions). In the order Stigonematales, the filaments are
often multiseriated, with genuine branching. Both heterocysts and akinetes are present.
The only means of reproduction in cyanobacteria is asexual. Filamentous forms
reproduce by trichome fragmentation, or by formation of special hormogonia.
Hormogonia are distinct reproductive segments of the trichomes.
They exhibit active gliding motion upon their liberation and gradually develop
into new trichomes. In contrast to eukaryotic microalgae, cyanobacteria do not possess
membrane-bound sub-cellular organelles; they have no discrete membrane-bound
nucleus; they possess a wall structure based upon a peptidoglycan layer; and they
contain 70 S rather than 80 S ribosomes (Fay and Van Baalen, 1987; Bryant, 1994).

17
The photosynthetic pigments of cyanobacteria are located in thylakoids that lie free in
the cytoplasm near the cell periphery. Cell colours vary from blue-green to violet-red.
The green of chlorophyll a is usually masked by carotenoids (e.g. beta-carotene)
and accessory pigments such as phycocyanin, allophycocyanin and phycoerythrin
(phycobiliproteins). The pigments are embodied in phycobilisomes, which are found in
rows on the outer surface of the thylakoids (Douglas, 1994). All cyanobacteria contain
chlorophyll a and phycocyanin. The basic features of photosynthesis in cyanobacteria
have been well described (Ormerod, 1992). Cyanobacteria are oxygenic phototrophs
possessing two kinds of reaction centres, PS I and PS II, in their photosynthetic
apparatus. With the accessory pigments mentioned above, they are able to use
effectively that region of the light spectrum between the absorption peaks of
chlorophyll a and the carotenoids.
The ability for continuous photosynthetic growth in the presence of oxygen,
together with having water as their electron donor for CO2 reduction, enables
cyanobacteria to colonize a wide range of ecological niches (Whitton, 1992).
Phycobiliprotein synthesis is particularly susceptible to environmental influences,
especially light quality. Chromatic adaptation is largely attributable to a change in the
ratio between phycocyanin and phycoerythrin in the phycobilisomes. Thus,
cyanobacteria are able to produce the accessory pigment needed to absorb light most
efficiently in the habitat in which they are present.
Cyanobacteria have a remarkable ability to store essential nutrients and
metabolites within their cytoplasm. Prominent cytoplasmic inclusions for this purpose
can be seen with the electron microscope (e.g. glycogen granules, lipid globules,
cyanophycin granules, polyphosphate bodies, carboxysomes) (Fay and Van Baalen,
1987). Reserve products are accumulated under conditions of an excess supply of
particular nutrients. For example, when the synthesis of nitrogenous cell constituents is
halted because of an absence of a usable nitrogen source, the primary products of
photosynthesis are channeled towards the synthesis and accumulation of glycogen and
lipids. Dinitrogen fixation is a fundamental metabolic process of cyanobacteria, giving

18
them the simplest nutritional requirements of all living organisms. By using the enzyme
nitrogenase, they convert N2 directly into ammonium (NH4) (a form through which
nitrogen enters the food chain) and by using solar energy to drive their metabolic and
biosynthetic machinery, only N2, CO2, water and mineral elements are needed for
growth in the light.
Nitrogen-fixing cyanobacteria are widespread among the filamentous,
heterocyst forming genera (e.g. Anabaena, Nostoc) (Stewart, 1973). However, there are
also several well documented examples of dinitrogen fixation among cyanobacteria not
forming heterocysts (e.g. Trichodesmium) (Carpenter et al., 1992). Under
predominantly nitrogen limited conditions, but when other nutrients are available,
nitrogen fixing cyanobacteria may be favored and gain growth and reproductive
success. Mass developments (often referred to as "blooms") of such species in limnic
(e.g. eutrophic lakes) and marine environments (e.g. the Baltic Sea) are common
phenomena world-wide. Many species of cyanobacteria possess gas vesicles. These are
cytoplasmic inclusions that enable buoyancy regulation and are gas-filled, cylindrical
structures. Their function is to give planktonic species an ecologically important
mechanism enabling them to adjust their vertical position in the water column (Walsby,
1987). To optimize their position, and thus to find a suitable niche for survival and
growth, cyanobacteria use different environmental stimuli (e.g. photic, gravitational,
chemical, thermal) as clues.
Gas vesicles become more abundant when light is reduced and the growth rate
slows down. Increases in the turgor pressure of cells, as a result of the accumulation of
photosynthate, cause a decrease in existing gas vesicles and therefore a reduction in
buoyancy. Cyanobacteria can, by such buoyancy regulation, poise themselves within
vertical gradients of physical and chemical factors. Other ecologically significant
mechanisms of movement shown by some cyanobacteria are photomovement by slime
secretion or surface undulations of cells (Häder, 1987; Paerl, 1988).

19
Cyanobacterial Taxonomy:
A characteristic cyanobacterial membrane lipid has been extracted from late
Archean sedimentary rocks dated to 2.65 Ga .The microfossils found at the Apex Chert
in Western Australia, believed to be cyanoprokaryotes, are even 800 million years
older. A minimum date for the evolution of heterocytic forms is set to 1.5 billion years
ago; to the period the oldest fossils interpreted as akinetes have been dated (Summons
et al., 1999).
For more than 150 years cyanobacteria were considered to be eukaryotic algae,
with botanists and phycologists placing them into the Cyanophyceae or cyanobacteria.
Thus, initial classifications followed the International Code of Botanical Nomenclature.
Traditional techniques for cyanobacteria identification and systematics have relied
essentially on the observation of morphological characteristics. The confirmation by
molecular methods that these denominated cyanobacteria, where in fact photosynthetic
bacteria and their transfer from Cyanophyceae to Cyanobacteria were of most
importance (Rasmussen and Svenning, 1998).
Evolutionary Markers:
The assessment of the phylogeny of organisms through gene sequence analysis
has increased dramatically since the advent of PCR and automated sequencing. A
number of genes have been used as evolutionary markers for inferring phylogenetic
relations and delineation of cyanobacterial taxonomy, being the 16S rDNA gene
analyzed most extensively (Nubel et al., 1997).
Phylogenetic studies using this marker helped to clarify the phylogenetic
relationships among cyanobacteria, revealed the structure and intraspecific diversity of
cyanobacterial communities and provide further evidence to the cyanobacterial origin
of chloroplasts and the existence of a divergent evolutionary pathway among bacteria
(Bergsland & Haselkorn, 1991). Bacterial rRNA genes are commonly organized in an
operon in the order 16S rRNA - 3S rRNA - 5S rRNA, each rRNA gene being separated
by an internal transcribed spacer (ITS) region. The amplification of the 16S - 23S
rRNA internal transcribed spacer (ITS) in cyanobacteria have shown to present

20
different sizes and also to be more variable in sequence even within closely related
taxonomic groups than the 16S rRNA (Iteman et al., 2002).
Other Markers
In some cases other sequences have also been used for phylogenetic inferences:
the non-coding intergenic spacer of the phycocyanin operon (PC-IGS); other DNA-
dependent RNA polymerase regions, rpoB and rpoD; the gene encoding a serine-type
of protease with a regulatory role in the differentiation process of heterocysts (hetR);
nitrogen fixation (nif) genes; carbon-fixation-associated gene (RubisCO spacer)
(rbcLX); and the subunit B protein of DNA gyrase (gyrB) (Robertson et al., 2001).
C-Phycocyanin gene:
Cyanobacteria are the only microorganisms to produce significant quantities of
Phycocyanin (PC) and its derivative Allophycocyanin. This distribution of PC in
aquatic microorganisms makes the study of PC gene sequence heterogeneity ideal for
the classification of freshwater cyanobacteria. The entire PC operon contains genes
coding for two bilin subunits and three linker polypeptides. The intergenic spacer (IGS)
between the two bilin subunit genes, designated b (cpcB) and a (cpcA), of the PC
operon was chosen as a potentially highly variable region of DNA sequence useful for
the identification of cyanobacteria to the strain level. C-phycocyanin is one of the major
photosynthetic biliproteins and also one of the important components in the electron
transfer of photosynthesis. It has some functions that are good for human health, such
as antioxidant, radical scavenging, anti-inflammatory and anti-cancer properties, and
also used in the food, biotechnology, and cosmetic industry because of their color,
florescent and antioxidant properties. So in recent years it has been drawn more and
more attention (Reddy et al., 2003, Sekar and Chandramohan, 2007).
The Phycobilisome components (phycobiliproteins) are responsible for the blue-
green pigmentation of most cyanobacteria. All phycobiliproteins are water-soluble and
therefore cannot exist within the membrane as do carotenoids, but aggregate forming
clusters that adhere to the membrane called phycobilisomes. Phycocyanin absorbs
orange and red light, particularly near 620 nm (depending on which specific type it is),

21
and emits fluorescence at about 650 nm (also depending on which type it is).
Allophycocyanin absorbs and emits at longer wavelengths than Phycocyanin. A major
advantage of using molecular techniques instead of microscopic methods is the ability
to enhance the taxonomic resolution from genus-level to genotype-level, which is not
possible to attain through other methods (Ouellette and Wilhelm, 2003).
The C-phycocyanin gene sequence was first reported (Pilot & Fox, 1984), in
which the oligonucleotide synthesis of alpha and beta subunits of cpc gene of a
freshwater cyanobacterum Agmenellum quadruplicatum was described and also
reported the cpcA and cpcB gene sequence in Spirulina platensis. The cpc gene
sequences of some Spirulina strains are used as a site to analyze their phylogenetic
relationships. There is no more detailed information about the regulatory sequence and
cpc operon (Yu et al., 2002).
Molecular characterization of ten marine cyanobacterial isolates belonging to
the order Oscillatoriales was carried out using the phycocyanin locus (cpcBA-IGS) and
the 16S-23S internally transcribed spacer region. DNA sequences from the
phycocyanin operon discriminated ten genotypes, which corresponded to seven
morphotypes identified by traditional microscopic analysis. The cpcB coding region
revealed 17% nucleotide variation, while cpcA exhibited 29% variation across the
studied species. Phylogenetic analyses support the conclusion that the Phormidium and
Leptolyngbya genera are not monophyletic. The nucleotide variations were
heterogeneously distributed with no or minimal informative nucleotides. Our results
suggest that the discriminatory power of the phycocyanin region varies across the
cyanobacterial species and strains. The DNA sequence analysis of the 16S - 23S
internally transcribed spacer region also supports the polyphyletic nature of the studied
Oscillatorian cyanobacteria. This study demonstrated that morphologically very similar
strains might differ genotypically. Thus, molecular approaches comprising different
gene regions in combination with morphological criteria may provide better
taxonomical resolution of the order Oscillatoriales (Premanandh et al., 2006).

22
Cyanobacterial Features:
Cyanobacterial morphology ranges from simple unicellular, colonial and
multicellular filamentous forms. The vegetative cell wall is of gram-negative type but
in some species the peptidoglycan layer is considerably thicker than in other bacteria.
Minute pores are present in regular or scattered order in the wall of all cyanobacteria,
but the arrangement varies greatly. Many unicellular and filamentous cyanobacteria
possess an “envelope” outside the lipopolysaccharide (LPS) “outer membrane”, which
is called: sheath, glycocalyx, or capsule
The photosynthetic apparatus of cyanobacteria contains photosystem I and
photosystem II as found in higher plants with Chlorophyll a and specific accessory
pigments, including Allophycocyanin, Phycocyanin and Phycoerythrin. Cyanobacteria
possess the ability to use low light intensities effectively, since they are able to produce
the accessory pigments needed to adsorb light most efficiently in the habitat in which
they are present, providing them a great advantage for the colonization of their wide
range of ecological niches. Phycobiliprotein synthesis is particularly susceptible to
environmental influences, especially light quality. The chromatic adaptation is largely
attributable to a change in the ratio between Phycocyanin and Phycoerythrin in the
Phycobilisomes. The photosynthetic pigments are located in thylakoids that are free in
the cytoplasm near the cell periphery. Cell colours vary from blue green to violet-red
due to the chlorophyll a masking by the carotenoids and accessory pigments. The
pigments are involved in phycobilisomes, which are found in rows on the outer surface
of the thylakoids.
Cyanobacteria get their name from the bluish pigment phycocyanin, which they
use to capture light for photosynthesis. In some cyanobacteria, the color of light
influences the composition of phycobilisomes. In green light, the cells accumulate more
phycoerythrin, whereas in red light they produce more phycocyanin. Thus the bacteria
appear green in red light and red in green light. This process is known as
complementary chromatic adaptation.

23
Cyanobacteria are also able of storing essential nutrients and metabolites within
their cytoplasm. The occurrence of fimbriae (pili) is abundant in many cyanobacteria
with varying patterns. Some filamentous forms are also able of gliding (sliding), using
mucilaginous excretions as propellant. Some cyanobacteria have evolved specialized
cells for nitrogen fixation (heterocytes), survival in stressed conditions (akinetes), and
dispersion (hormogonia).
Cyanobacteria have many fascinating features such as buoyancy,
photosynthesis, fixation of atmospheric nitrogen and production of a wide variety of
bioactive compounds. In addition, cyanobacteria form symbiosis with several
eukaryotic hosts such as plants, fungi, and protists. Probably owing to their
physiological flexibility and long evolutionary history, cyanobacteria inhabit a large
variety of terrestrial and aquatic habitats from deserts to lakes as well as hot springs and
glaciers. Cyanobacteria form biofilms (microbial mats) on shores and on the surface of
stones, plants, and artificial objects. Cyanobacterial blooms are frequently toxic and
thus pose a health risk for humans and animals, cause an aesthetic problem, and reduce
the recreational value of water (Mur et al., 1999).
Limitations of phenotypic characters in cyanobacterial identification led to the
development of molecular techniques, including DNA base composition, DNA and
RNA hybridizations, gene sequences and PCR fingerprinting methods for
cyanobacteria taxonomy. However, it has been difficult to define taxonomic or
phylogenetic relationships within the cyanobacteria because of the scarcity of distinct,
consistent characters that support a taxonomic scheme. The problems of cyanobacterial
taxonomy are name changes of some strains, besides the misidentification issue of
others. Consequently, an ever changing classification system and a lack of a consensus
phylogeny are the proofs of the unresolved evolutionary relationships among
cyanobacteria.
A molecular approach to the systematic of cyanobacteria may be most fruitful
for inferring phylogenetic relationships. Macromolecules, such as nucleic acids and
proteins, are copies or translations of genetic information. The methods applied involve

24
direct studies of the relevant macromolecules by sequencing, or indirectly by
electrophoresis, hybridization, or immunological procedures (Wilmotte, 1994). Nucleic
acid technologies, especially the polymerase chain reaction (PCR), have advanced to
the point that it is feasible to amplify and sequence genes and other conserved regions
from a single cell. However, the molecular results obtained have integrated with other
characteristics as the base for polyphasic taxonomy (Vandamme et al., 1996).
Rapid developments in genomic and other molecular research technologies and
developments in information technologies have combined to produce a tremendous
amount of information related to molecular biology by the application of
bioinformatics. Common activities in bioinformatics include mapping, analyzing DNA
and protein sequences, then for aligning different DNA and protein sequences to
compare, create and to view 3-D models of protein structures. Major research efforts in
the field include sequence alignment, gene finding, genome assembly, drug design,
drug discovery, protein structure alignment, protein structure prediction, prediction of
gene expression and protein-protein interactions, genome-wide association studies and
the modeling of evolution. Software tools were used for sequence analysis and structure
prediction includes BLAST, protein structure viewing, NCBI, EMBL etc (Mount,
2002).
Protein has three main structures: primary structure which is essentially the
linear amino acid sequence and usually represented by a one letter notation. Alpha
helices, beta sheets, and loops are formed when the sequences of primary structures
tend to arrange themselves into regular conformations; these units are known as
secondary structure. Protein folding is the process that results in a compact structure in
which secondary structure elements are packed against each other in a stable
configuration. This three-dimensional structure of the protein is known as the protein
tertiary structure. However, loops usually serve as connection points between alpha-
helices and beta-sheets, they do not have uniform patterns like alpha-helices and beta-
sheets and they could be any other part of the protein structure rather than helices or
strands (Kendrew et al., 1960).

25
In Bioinformatics, BLAST is an algorithm for comparing primary biological
sequence information, such as the amino-acid sequences of different proteins or the
nucleotides of DNA sequences. A blast search enables a researcher to compare a query
sequence with a library or database of sequences, and identify library sequences that
resemble the query sequence above a certain threshold. Molecular phylogenetics is the
analysis of hereditary molecular differences, mainly in DNA sequences, to gain
information on an organism's evolutionary relationships. The result of a molecular
phylogenetic analysis is expressed in a phylogenetic tree. Molecular phylogenetics is
one aspect of molecular systematics, a broader term that also includes the use of
molecular data in taxonomy and biogeography.
Molecular modelling is used to model the molecules. Both theoretical and
computational techniques are used in molecular modelling. Molecular modelling
methods are used to investigate the structure, dynamics and thermodynamics of
inorganic, biological, and polymeric systems. The types of biological activity that have
been investigated using molecular modelling include protein folding, enzyme catalysis,
protein stability, conformational changes associated with biomolecular function, and
molecular recognition of proteins, DNA, and membrane complexes. Cyanobacteria, our
model organisms, are regarded as an origin of producing phycocyanin pigments;
therefore they are considered as excellent candidates to study molecular techniques to
resolve many of the issues and problems in cyanobacterial taxonomy. Thus, the work is
mainly focused on the following objectives.
Molecular Modeling of C-Phycocyanin:
Molecular modeling is a powerful methodology for analyzing the three
dimensional structure of biological macromolecules. There are many ways in which
molecular modeling methods have been used to address problems in structural biology.
It is not widely appreciated that modeling methods are often an integral component of
structure determination by NMR spectroscopy and X-ray crystallography. The overall
aim of modeling methods will often be to try to relate biological activity to structure.
An important step towards this goal is to be able to compute the potential energy of the
molecule as a function of the positions of the constituent atoms. The common feature of

26
molecular modeling techniques is the atomistic level description of the molecular
systems; the lowest level of information is individual atoms or a small group of atoms.
This is in contrast to quantum chemistry which is also known as electronic structure
calculations, where electrons are considered explicitly. The benefit of molecular
modeling is that it reduces the complexity of the system, allowing many more atoms to
be considered during simulations (Foster, 2002).
Structural modeling of the gene:
Alpha-helix is spiral turns of amino acids while a beta-sheet is flat segments or
strands of amino acids formed usually by a series of hydrogen bonds. As the
polypeptide chain coils in, the CO and NH groups of residues form hydrogen bonds
which stabilize the helix. Most of the residues in a helix are bonded in this way, making
it somewhat a rigid unit of structure with a little free space in its core. A helix and can
have 4 - 50 residues and makes a whole turn every 3.6 residues.
Beta-strands are the most regular form of extended polypeptide chain in protein
structures. Like alpha-helices, beta-sheets are stabilized by hydrogen bonds between
CO and NH groups, but they are distantly separated along the chain. Because of the
geometry of the peptide backbone, the amino acid side chains of beta-strands alternate
on either side of the sheet.
Loops usually serve as connection points between alpha-helices and beta-sheets,
they do not have even patterns like alpha-helices and beta-sheets and they could be any
other part of the protein structure. They are recognized as random coil and not
classified as protein secondary structure. When the polypeptide chain makes very sharp
changes in direction using as few as four residues by means of hydrogen bond, it forms
turns. These secondary structures commonly contain proline or glycine or both residues
(Hutchinson and Thornton, 1994).
Tertiary Structure:
The three-dimensional structure of the protein, which is formed from the
secondary structures as subunits elements, is known as the protein’s tertiary structure.

27
Protein folding is the process that results in a compact structure in which secondary
structure elements are packed against each other in a stable configuration.
Hydrogen bonds, van der Waals forces, and oppositely charged amino acid side-
chains are other interactions that help to stabilize the fold. Folds are considered as sets
of connected secondary structure elements, so they are known as topologies. Longer
polypeptide chains that are usually clearly distinguished by a naked eye as self-
contained units of structure, and have distinct hydrophobic cores, are known as
domains. Homology modeling can produce high-quality structural models when the
target and template are closely related, which has inspired the formation of a structural
genomics consortium dedicated to the production of representative experimental
structures for all classes of protein folds (Dill, 1990).
The Crystal structure of the light-harvesting protein phycocyanin from the
Cyanobacterium Cyanidium caldarium with novel crystal packing has been solved at
1.65-Å resolution. The structure has been refined to an R value of 18.3% with excellent
backbone and side-chain stereochemical parameters. In crystals of phycocyanin used in
this study, the hexamers are offset rather than aligned as in other phycocyanins that
have been crystallized to date. Analysis of this crystal’s unique packing leads to a
proposal for phycobilisome assembly in vivo and for a more prominent role for
chromophore b-155. This new role assigned to chromophore b-155 in phycocyanin
sheds light on the numerical relationships among and function of external
chromophores found in phycoerythrin and phycoerythrocyanin (Stec et al., 1999).
Impact of Molecular Methods on Cyanobacterial Taxonomy:
Molecular methods have had a great impact on every level of cyanobacterial
taxonomy. Prochlorophytes, which were considered a special group of oxyphototroph
prokaryotes for the lack of phycobiliproteins and the presence of chlorophyll b, have
been shown to be polyphyletic according to the 16S rDNA and scattered throughout the
cyanobacterial lineage. But contradictory to the general concept, a phycobiliprotein
gene, similar to that of the marine Synechococcus, has been detected in
Prochlorococcus marinus CCMP 1375 strain (Hess et al., 1996).

28
Another important task solved by 16S rDNA sequencing was the origin of
plastids that were proven to have descended from cyanobacteria. This was later
confirmed also by tufA gene sequence. Plastids form a monophyletic group within
cyanobacterial lineage, but no strong candidate for the sister taxon to plastids exists at
present (Delwiche et al., 1995 and Turner, 1997). Heterocytic cyanobacteria were
confirmed and the orders Chroococcales and Oscillatoriales were shown to be
polyphyletic. The baeocyte-forming order Pleurocapsales seemed to be monophyletic at
first, but it turned out to be polyphyletic too, with Chroococcidiopsis thermalis being a
close relative of heterocytous cyanobacteria. The same may be stated for heterocytous
order Stigonematales - it has been shown recently, on the basis of both 16S rDNA and
nif genes, that it is polyphyletic as well (Giovannoni et al., 1988 and Zehr et al., 1997).
The situation on the generic and subgeneric level is even more confused. The
ecological studies concerning diversity of cyanoprokaryotes in various habitats are
many, but it is often not clear what is meant under the taxonomic designations assigned
to the studied organisms.
Many misidentified strains in culture collections are available and no
morphological data for a bulk of available cyanobacterial sequences. However, some
work has been done on the cytomorphological and polyphasic characterization of
chroococcalian "Synechocystis", "Synechococcus" and a few other unicellular strains as
well as of hetercytous Aphanizomenon/Anabaena/Nostoc strains. The studies on
filamentous "Phormidium" and "Oscillatoria" genera are few. Although there is often
no correlation between morphological and molecular traits, especially for taxa with
very simple morphology, some morphologically well defined genera were shown to be
monophyletic. These include Microcystis, Planktothrix or marine Trichodesmium
species. Notwithstanding the unreliability of traditional morphological criteria, some
cytomorphological and ultra structural characters were found to correlate well with
molecular data. This concerns e.g. the cell division type and especially the thylakoid
arrangement, which seems to have substantial taxonomic value. Some other traits, such
as perforation-patterns in the cell wall of cyanobacteria may prove useful on certain
taxonomic levels (Pffeifer and Palinska, 2002 and Palinska and Krumbein, 2000).

29
Relationship between Genotypic and Phenotypic Characters
Cyanobacteria are morphologically diverse in comparison to the rest of bacteria.
Nevertheless, only a quite restricted number of morphotypes can be recognized.
Molecular methods enabled revelation of cryptic genetic, physiological and ecological
diversity among them. Rather broadly defined species Phormidium retzii was shown to
be quite variable on molecular level. Although there were some morphological
distinctions between individual populations, they did not correlate with genetic
similarity. Strains morphologically corresponding to the genus Geitlerinema sp.
generated very different restriction patterns (Casamatta et al., 2003).
The studies on cyanobacterial communities of both geothermal springs in the
Philippines and Lake Fryxell in Antarctica revealed significantly higher degree of
diversity by molecular methods than by light microscopy. Genetically distinct toxic
Microcystis and Planktothrix populations were found in different parts of the same
lake. However, it is also possible that the strains, closely related on molecular level,
show substantial phenotypic variability, as shown for several Merismopedia isolates.
Co-existing Prochlorococcus ecotypes were detected, almost identical genetically, but
possessing very different light-dependent physiologies (Taton et al., 2003, Palinska et
al., 1996, Moore et al., 1998).
Interesting topic is the correlation between phylogenetic relatedness and
ecological or ecophysiological characters. Unicellular cyanobacteria from hypersaline
habitats in various geographical regions form a well defined cluster on the basis of their
16S rDNA that was denominated Halothece cluster. Moderately halophilic, benthic
strains with very thin trichomes also form a distinct cluster in the phylogenetic tree and
were assigned to a new genus Halomicronema (Garcia-Pichel et al., 1998).
An interesting fact that the only available 16S rDNA sequences of
cyanobacteria from stone surfaces of buildings group with those of desert strains from
distant geographic region, while sequence homology with the strains from other

30
habitats is quite low. The sequence similarity somehow reflects the capacity to survive
in such extreme environments (Crispim and Gaylarde, 2005)
The analysis of the hli gene family, which has to do with adaptation to high
light intensities, revealed that some groups of this gene family are specific either for
marine or freshwater cyanobacteria .A nice example of ecological divergence of
morphologically and phylogenetically closely related, but distinct genera
Trichodesmium and Hydrocoleum (Blennothrix) was documented. The first one is
planktonic, the latter is the most common mat-forming cyanobacterium in tropical
oceans (Bhaya et al., 2002 and Abed et al., 2006).
Biotechnological Applications of cyanobacteria:
Cyanobacteria constitute a resource for several applications such as aquaculture,
food, feed, fuel, fertilizer, medicine, industry and even in combating pollution (WHO
1999).
Cyanobacterial bioactive compounds Cyanobacteria have been identified as a
new and rich source of bioactive compounds (Abarzua et al. 1999; Shimizu 2003;
Bhadury et al. 2004; Dahms et al. 2006). Isolated compounds belong to groups of
polyketides, amides, alkaloids, fatty acids, indoles and lipopeptides (Abarzua et al.
1999; Burja et al. 2001).
The literature review showed that to date up to 19 cyanobacterial strains
produce more than 20 different bioactive compounds. Most of the bioactive compounds
isolated from cyanobacteria tend to be lipopeptides, i.e. they consist of an amino acid
fragment linked to a fatty acid portion. The range of biological activity of secondary
metabolites isolated from cyanobacteria includes antibacterial, antifungal, antialgal,
antiprotozoan, and antiviral activities.
Only few cyanobacteria produce bioactive compounds that show a broad
spectrum of biological activities. For example, the cyanobacterium Phormidium sp. has
been reported to inhibit growth of different Grampositive and Gram-negative bacterial

31
strains, yeasts, and fungi (Bloor and England 1989). Another example is Lyngbya
majuscula (Burja et al. 2001) that produces numerous chemicals including nitrogen-
containing compounds, polyketides, lipopeptides, cyclic peptides and many others
(Shimizu 2003).
The biological activity of these compounds is also diverse and includes protein
kinase C activators and tumour promoters, inhibitors of microtubulin assembly,
antimicrobial and antifungal compounds and sodium-channel blockers. Secondary
metabolites with antibacterial activity are widely produced by cyanobacteria (Dahms et
al. 2006). These compounds are effective against Gram positive and ⁄ or Gram-negative
bacteria. Both toxic and nontoxic strains of cyanobacteria are producers of antibacterial
compounds that are distinct from cyanotoxins. Antifungal compounds include
fisherellin A, hapalindole, carazostatin, phytoalexin, tolytoxin, scytophycin,
toyocamycin, tjipanazole, nostocyclamide and nostodione produced by cyanobacteria
belonging to Stigonematales, Nostocales and Oscillatoriales. Additionally,
cyanobacteria produce a broad spectrum of antialgal compounds, which may be used to
control algal blooms.
Cyanobacteria probably use these compounds in order to out-compete other
microorganisms. Antialgal compounds produced by cyanobacteria inhibit growth of
algae, their photosynthesis, respiration, carbon uptake, enzymatic activity and induce
oxidative stress (Dahms et al. 2006). In contrast to the large amount of antibacterial and
antialgal compounds isolated from cyanobacteria there are only a few compounds that
show antiviral properties, although 2–10% of extracts of different cyanobacterial
species have been shown to have antiviral activity (Cohen 1999). These include
acetylated sulfoglyco-lipids from Oscillatoria raoi (Reshef et al. 1997) and spirulan
from Spirulina platensis (Hayashi et al. 1991). The compounds isolated from Lyngbya
lagerhaimanii and Phormidium tenue has been shown to have anti-HIV activity
(Rajeev and Xu 2004). Gamma linolenic acid (GLA) found rich in S. platensis and
Arthrospira sp. is medically important since it is converted in the human body into
arachidionic acid and then into prostaglandin E2. This compound has a lowering action
on blood pressure and plays an important role in lipid metabolism. Some of the marine

32
cyanobacteria constitute potential sources for large-scale production of vitamins, such
as vitamins B and E (Plavsic et al. 2004).
Piechula et al. (2001) demonstrated that some cyanobacteria can produce
thermostable enzymes. Out of 21 endonucleases from Phormidium, four enzymes that
catalyse the hydrolysis of DNA and RNA were active in a wide range of temperatures
from 15 to 60oC. Phenyltransferases enzymes catalysing the consecutive condensation
of homoallylic diphosphate of isopentenyl diphosphates at temperatures above 60oC
have been isolated from the thermophilic cyanobacterium, Synechococcus elongates
(Ohto et al. 1999). Thermostable polyphosphate kinase from the thermophilic
cyanobacterium Thermosynechococcus elongatus BP-1 was successfully employed in
an ATP regeneration system that could be used at high temperatures for the effective
production of d-amino acid dipeptides (Sato et al. 2007). These enzymes and other heat
stable bioactive compounds are of great interest in biotechnology. So far none of the
isolated compounds have demonstrated anti-larval activity and inhibition of settlement
of larvae and algal spores (Dobretsov et al. 2006).
Cyanobacteria have been used to synthesize isotopically labelled compounds
such as sugars, lipids and amino acids, which are nowadays commercially available
(Patterson 1996). This is achieved by growing the cyanobacetria in photobioreactors
and allowing them to photosynthetically transform simple labelled compounds such as 14CO2,
13CO2, 33H2O and 15NO3 into complex organics cyanobacteria as a healthy food
source Strains of Spirulina, Anabaena and Nostoc are consumed as human food in
many countries including Chile, Mexico, Peru and Philippines. Arthrospira platensis
(misidentified as S. platensis) is grown in large scale using either outdoor ponds or
sophisticated bioreactors but marketed in the form of powder, flakes, tablets and
capsules. It is used as a food supplement because of its richness in nutrients and
digestibility. It contains more than 60% proteins and is rich in beta-carotene, thiamine
and riboflavin and is considered to be one of the richest sources of vitamin B12. Nostoc
commune is rich in fibres and proteins and can play an important physiological and
nutritional role in the human diet. Aphanizomenon sp. is collected from natural blooms
in the Lake Klamath (Oregon, USA) to be used as healthy food. Marine nitrogen-fixing

33
cyanobacteria have also been tested to feed fishes in aquacultures. The Tilapia fish
showed high growth rates when fed with marine cyanobacteria in indoor and outdoor
cultures (Mitsui et al. 1983). Phormidium valderianum has been used in India to serve
as a complete aquaculture feed source based on its nutritional value and nontoxic
nature. In view of the cyanobacterial significance as a food source, very little research
has been performed and published about this.
Cyanobacterial alternative energy sources Cyanobacteria have been used to
produce hydrogen gas that constitutes an alternative future energy source to the limited
fossil fuel resources (Dutta et al. 2005). The advantages of using biological hydrogen
as a fuel are its eco-friendly nature, efficiency, renewability and the absence of carbon
dioxide emission during its production and utilization (Lindbald 1999).
Cyanobacteria produce hydrogen either as a byproduct of nitrogen fixation,
when nitrogenase-containing heterocystous cyanobacteria are grown under nitrogen
limiting conditions, or by the reversible activity of hydrogenases enzymes.
Heterocystous cyanobacteria are thus more efficient in hydrogen production than
nonheterocystous types (Pinzon-Gamez et al. 2005). More than 14 cyanobacterial
genera including Anabaena, Calothrix, Oscillatoria, Cyanothece, Nostoc,
Synechococcus, Microcystis, Gloeobacter, Aphanocapsa, Chroococcidiopsis and
Microcoleus are known for their ability to produce hydrogen gas under various culture
conditions (Lambert and Smith 1977; Sveshnikov et al. 1997; Masukawa et al. 2001).
Anabaena spp. is able to produce significant amounts of hydrogen. Nitrogen- starved
Anabaena cylindrica cells produce the highest amount of hydrogen (30 ml of H2 per lit
culture per hour) (Jefferies et al. 1978). Gloeocapsa alpicola showed increase in
hydrogen production under sulfur starvation (Antal and Lindblad 2005) whereas S.
platensis could produce hydrogen under complete dark and anoxia (Aoyama et al.
1997).
Large-scale hydrogen production by Spirulina and Anabaena spp. has been tried
using different types of bioreactors that included vertical column reactor, tubular type
and flat panel photobioreactor (Dutta et al. 2005). These reactors were designed to

34
make use of solar light for illumination, to maximize the area for incident light (high
surface to volume ratio) and to allow sterilization and hydrogen collection with
convenience and ease. Nevertheless, the reactors are subjected to continuous
modification in order to increase their productivity and to decrease costs of
maintenance and production. So far, these modifications succeeded in bringing down
the cost of biologically produced hydrogen to $25 per m-3 compared to $170 per m-3 for
the hydrogen produced by splitting of water (Block and Melody 1992).
Increased production of hydrogen was also achieved by genetically modifying
the nitrogenase enzyme in hydrogen-producing strains. The ongoing research on
hydrogen production by cyanobacteria focuses on finding new strains with higher
potential to produce hydrogen, optimizing the mass production of hydrogen in
bioreactors and modifying the physiology and the genetic system of H2-producing
cyanobacteria to ensure maximum production of hydrogen Cyanobacteria as
biofertilizers. Heterocystous cyanobacteria and several nonheterocystous cyanobacteria
are known for their ability to fix atmospheric nitrogen (Capone et al. 2005). The
fertility of many tropical rice field soils has been mainly attributed to the activity of
nitrogen-fixing cyanobacteria.
Species of cyanobacteria, namely strains of Spirulina (Arthrospira) platensis,
Nostoc commune and Aphanizomenon flos-aquae, are currently used as food and
supplements due to their amounts of proteins (e.g. up to 70% of Spirulina dry weight),
lipids, chlorophyll, carotenoids, vitamins, minerals, unique pigments and also due to
their potential prebiotic effects. Spirulina is also used as an aquaculture and animal feed
source. As a crop, Spirulina has the advantage of growing well in saline ponds in arid
environments, places that few other crops could support (Pulz and Gross, 2004).
Several fine chemicals such as pigments (carotenoids and phycobiliproteins),
vitamins (B-complex group and vitamin E) and enzymes with varied applications from
cyanobacteria are being commercialized. The pigments are used as natural food
colourants, food additives, and cosmetics and as enhancers of ornamental fish colour.
Isotopically labelled (14C, 15C, 3H and 15N) cyanobacterial metabolites such as sugars,

35
lipids and aminoacids are commercially available. Several endonucleases produced
from cyanobacteria are being marketed, e.g. AcyI (Anabaena cylindrica).
Cyanobacteria produce numerous bioactive compounds. Some secondary
metabolites are potentially of therapeutical importance, such as antiviral compounds,
immunomodulators, inhibitors, or cytostatics. N2-fixing cyanobacteria are used as
biofertilizers, for instance in the rice paddies where nitrogen to support the rice plants
comes from fixation by free-living and symbiotic cyanobacteria, besides being also
used in other tropical and subtropical agriculture.
Reports concerning the advantageous effects of cyanobacteria inoculation on
other crops such as barley, oats, tomato, radish, cotton, sugarcane, maize, chili and
lettuce have been described (Thajuddin and Subramanian, 2005). It has been reported
that the marine cyanobacterium, Oscillatoria formosa NTDM02 was efficiently
decolorized textile dyes and laboratory dye (MubarakAli et al., 2010).
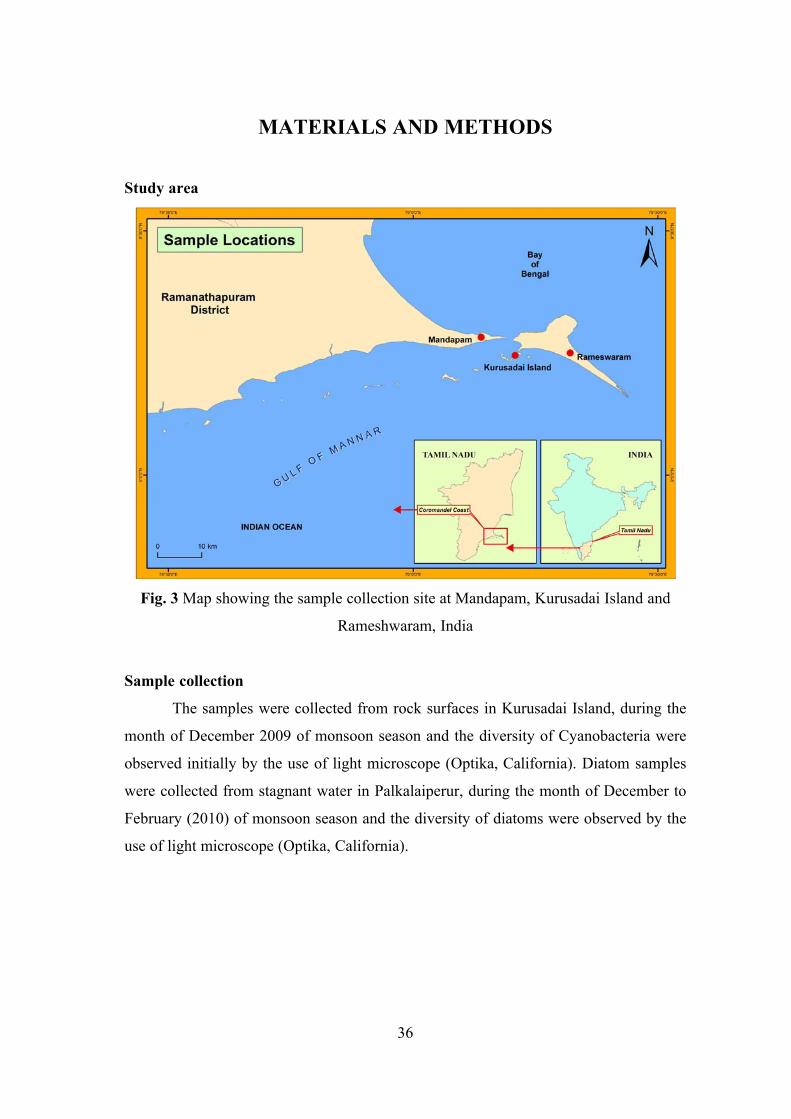
36
MATERIALS AND METHODS
Study area
Fig. 3 Map showing the sample collection site at Mandapam, Kurusadai Island and
Rameshwaram, India
Sample collection
The samples were collected from rock surfaces in Kurusadai Island, during the
month of December 2009 of monsoon season and the diversity of Cyanobacteria were
observed initially by the use of light microscope (Optika, California). Diatom samples
were collected from stagnant water in Palkalaiperur, during the month of December to
February (2010) of monsoon season and the diversity of diatoms were observed by the
use of light microscope (Optika, California).

37
Isolation of diatoms
The diatoms were isolated by using enriched sea water medium F/2 (Guillard
and Ryther, 1962) by serial dilution, capillary or micropipette method and streak plate
method. The cultures were grown under the fluorescent light of 1500 lux, 12:12
light:dark (L:D) condition at 25±2ºC. After incubation the cultures were examined at
regular intervals under light microscope.
Isolation of Cyanobacteria:
The Cyanobacteria were isolated by using enriched sea water medium MN and
ASN-III (Rippika, 1988) by serial dilution and streak plate method. The cultures were
grown under the fluorescent light of 1500 lux, 12:12 light: dark (L: D) condition at
25±2ºC. After incubation the cultures were examined at regular intervals under light
microscope.
Morphological characterization of diatom and marine Cyanobacteria
The isolated diatom and marine Cyanobacteria were examined carefully by using
light microscope and microphotograph was taken (Optika Photomicroscope Unit) to
determine the morphology. The identification of Cyanobacteria in marine water
samples was achieved using standard monograph (Geitler, 1932; Desikachary, 1959).
The diatom strains were genus was identified based on frustules morphology by using
the atlas of diatoms (Desikachary, 1989). The isolated cyanobacterial strains were
maintained as axenic cultures in the germplasm of Department of Microbiology,
Bharathidasan University.
Molecular characterization of diatom
DNA extraction from diatom (Grachev et al., 2006)
A grown culture was centrifuged to obtain about 50µl of packed biomass. Cells
were suspended in 100µl of a lysis buffer (100 mM Tris-HCl, pH 8.0,1.4 M NaCl, 2%
CTAB, 20mM EDTA), homogenized with a pestle at 60ºC for 10 minutes, combined
with 150µl of the lysis buffer, and incubated at 60ºC for 30 minutes with occasional
shaking. Cell debris was removed by centrifugation (3 minutes), and the supernatant
was transferred into another tube. The pellet was washed with 100µl of the lysis buffer,

38
and the supernatant was added to the first one. The pooled supernatant was combined
with 7.5µl of RNase, incubated at 60ºC for 30 minutes, combined with 3µl of
proteinase K, incubated again at 60ºC for 30 minutes, and combined with 1ml of 1%
CTAB (aqueous solution) at room temperature.
The resulting suspension was stirred and frozen-thawed. The pellet was
collected by centrifugation (15 minutes) and washed twice with 300µl of 1% cetavlon
(aqueous solution). The supernatant was carefully removed by overturning the tube on
filter paper. The pellet was extracted twice with 400µl of ethanol at 60ºC for 10
minutes. The supernatant (ethanol solution of the CTAB salt of DNA) was collected by
centrifugation (5 minutes) and combined with 80µl of 3M sodium acetate and kept at -
20ºC overnight. The precipitate (DNA sodium salt) was collected by centrifugation,
washed with ethanol, and air-dried. DNA was dissolved in 50µl of a PCR buffer and
stored at -20°C.
The extracted DNA were estimated directly from ethidium bromide
fluorescence in agarose gel (0.8% in 1X TBE buffer) against a standard marker,
(Finzymes, Finland) by using a gel documentation with associated software.
Oligonucleotide primers and PCR conditions:
All PCR reactions were carried out in a 40µl volume containing (i)20µl of 2x
prime Taq premix ( GENET BIO, Korea) which contains 1unit Taq DNA Polymerase,
20mM Tris-Hcl, 80mM KCl, 4mM MgCl2, enzyme stabilizer, sediment, loading dye,
pH 9.0, 0.5mM of each dATP, dCTP, dGTP, dTTP. (ii) 1µl of upstream primer (10
pmoles/µl) (iii) 1µl of downstream primer (10pmoles/µl) (iv) 1µl (50ng) of extracted
DNA from diatom (v) 7 µl of sterilized distilled water.
PCR amplification of the small subunit of 18S rRNA gene (SSU rDNA) was
done in a thermal cycler gradient (Applied biosystems, Germany) with the primer pairs
1F and 1528R (Medlin et al. 1988). Conditions for PCR were as follows: initial
denaturation at 94ºC for 5 mins, followed by 36 cycles of 94ºC for 2 min, 54ºC for 4
min, and 72ºC for 2 min, and a final extension at 72ºC for 10 min.

39
18S rDNA gene amplification:
A fragment of 18S rDNA gene was amplified as mentioned above (Ulf Karsten
et al., 2006). After the reaction was completed, 10µl of amplified DNA was separated
on 1.2% agarose, stained with ehidium bromide and recorded using a CCD camera in
UVP Imager (UVP Bioimaging, England). A ready to use DNA size standard samples
(Finzymes, Finland) was added in the gel to determine the molecular weight of the
marker.
Primer Sequence 5´→3’ Author
1F AAC CTG GTT GAT CCT GCC AGT
Medlin et al.,1988
1528 R TGA TCC TTC TGC AGG TTC ACC
TAC
Molecular characterization of marine Cyanobacteria:
DNA extraction:
The extraction of genomic DNA from cyanobacterial isolates were carried out
(Smoker and Barnum 1988). Briefly, 1ml of overnight grown cultures was centrifuged
at 10,000 rpm for 5 minutes and the pellet was collected, then collected pellet was
washed using STE buffer (NaCl – 50mM; Tris-HCl - 50mM; EDTA - 5mM) and
suspended in 500μl of STE buffer. Followed by 20μl of lysozyme (10mg/ml) was
added and incubated in water bath at 55°C for 30 minutes. After incubation, 10μl of
proteinase K (10mg/ml) and 20μl of 10% SDS was added and incubated in water bath
at 55°C for 30 minutes. The mixture was then cooled in ice and extracted with equal
volume of phenol: chloroform: Isoamyl alcohol mixture (25:24:1) and centrifuged at
12,000 rpm for 10 minutes. After centrifugation, the supernatant was separated with
care and equal volume of 4M Ammonium acetate and two volumes of Isopropanol
were added. Mixture was again centrifuged at 14,000 rpm for 15 minutes and the
supernatant was decanted and the pellet was washed with 70% ethanol, dried. Finally,
the DNA pellet was dissolved in 100μl of TE buffer and stored at 20°C until further
use. The loading dye was added to each DNA samples and the samples were transferred
to separate wells in 0.8% agarose gel and after loading, the electrophoresis was carried

40
out using 1X TAE buffer at a constant supply of 100 V for 30 minutes. The bands were
documented under transilluminator.
16S rDNA gene amplification:
PCR amplification was performed for the respective two samples of purified
DNA using CYA 106 (5’ - CGG ACG GGT GAG TAA CGC GTGT- 3’) and CYA 781
(5’ - GAC TAC TGG GGT ATC TAA TCC CA T - 3’) primers. The polymerase chain
reactions conditions include initial denaturation of template DNA was achieved at 94oC
for 3 minutes. Further denaturation was carried out at 94oC for 45 sec; annealing at
56oC for 30 sec, elongation at 72oC for 1min and a final elongation at 72oC for 7 min
for 30 cycles. Amplified products were isolated by electrophoresis on 1.2% agarose gel
using 1X TAE buffer at a constant supply of 100 V for 30 minutes. Again, resolved
bands were documented under Uv transilluminator.
C- Phycocyanin (PC) Gene amplification:
PCR amplification of C-Phycocyanin was performed for the respective two
samples of purified DNA using PCβ F (GGC TGC TTG TTT ACG CGA CA) and
PCαR (CCA GTA CCA CCA GCA ACT AA) primers (Neilan, 2002). The polymerase
chain reactions conditions include initial denaturation of template DNA was achieved
at 94oC for 5 minutes. Further denaturation was carried out at 940C for 2 mins;
annealing at 550 C for 1min, elongation at 720C for 3 mins and a final elongation at
720C for 8 min for 40 cycles. The reaction mixture of amplification of PC-IGS region
contains 40µl.
Sequencing and Construction of phylogeny:
Sequencing was done with amplified samples with respective forward and
reverse primers and sequences. The query sequence was uploaded in alignment box and
the query was submitted to BLAST analysis for pair-wise sequence alignment. The
evolutionary distances were computed using the Maximum Composite Likelihood
method (Tamura et al., 2004) and are in the units of the number of base substitutions
per site and it was computed in MEGA 5 software.

41
Molecular modeling of C-Phycocyanin gene:
Prediction of primary, secondary and tertiary structure of C-Phycocyanin and
determination of conserved domains of the C-phycocyanin sequences were analyzed
using online and offline softwares for molecular modeling of the protein.
Tertiary Structure:
Retrieval of target protein sequences from NCBI in Fasta format and save as in
notepad. The template structure was searched through Blastp algorithm against PDB
database. Target sequence alignment and template protein structure were derived for
Structure prediction using spdV software. The modelled structures were subjected to fit
the models and to optimize the stereochemistry atoms of molecules. The final modelled
structure to predict 3D structure profiles and it was visualized.
Conserved Domain determination (CDD):
CDD is a protein annotation resource that consists of a collection of well-
annotated multiple sequence alignment models for ancient domains and full-length
proteins. These are available as position-specific score matrices (PSSMs) for fast
identification of conserved domains in protein sequences via RPS-BLAST. CDD
content includes NCBI-curated domains, which use 3D-structure information to
explicitly define domain boundaries and provide insights into sequence / structure /
function relationships, as well as domain models imported from a number of external
source databases (Pfam, SMART, COG, PRK, TIGRFAM)
Identify the putative function of a protein sequence, identify the amino acids in
a protein sequences that are putatively involved in functions such as binding or
catalysis, as mapped from conserved domain annotations to the query sequences. View
a query protein sequence embedded within the multiple sequence alignment of a
domain model, interactively views the 3D structure of a conserved domain, Find other
proteins with similar domain architecture and interactively view the phylogenetic
sequence tree for a conserved domain model of interest with or without a query
sequence embedded (Marchler-Bauer et al. 2011).

42
RESULTS AND DISCUSSION
Totally, 50 cyanobacterial species and 38 diatom species were recorded. They
were identified taxonomically with standard monographs. The schematic representation
of collection and marine cyanobacterial diversity were shown (Fig. 4). There are five
family from cyanobacterial diversity such as Chroococcaceae (7), Nostacaceae (4),
Oscillatoriaceae (37), Pseudanabaenaceae (1) and Mastigocladaceae (1) respectively,
where as in diatom were found to be in thirteen families such as Achanthaceae (2),
Bacillariaceae (6), Catenulaceae (1), Cymbellaceae (2) Desmidiaceae (1),
Fragilariaceae (4), Gomphonemataceae (1), Mastoglioaceae (2), Naviculaceae (8),
Pinnulariaceae (4), Pleurosigmataceae (4), Stauroneidaceae (1), Stephanodiscaceae
(2) (Table 1a &b).
The morphological evidence of the cyanobacteria and diatom were shown (Fig.
5a & b and 6). Cyanobacterial taxonomy follows the International Code of
Nomenclature of Bacteria (Stainer et al., 1978) and published a taxonomy of the
cyanobacteria that was based on physiological, morphological and some genetic criteria
(Rippka et al., 1979). The revision of the status of the nomenclature of Cyanobacteria
under the Bacteriological Code, only 13 names of cyanobacterial species have been
proposed and since the corresponding genera name had not been validly published
according to the International Code of Nomenclature of Bacteria. Although at the
present there has been an attempt to classify cyanobacteria using both Botanical and
Bacteriological Code of Nomenclature (Tindall, 2006).
The traditional observation of the morphological characters by light microscopy
requires considerable expertise to identify species. Moreover, some phenotypic
features, such as gas vacuoles or akinetes etc. may vary with environmental or growth
conditions and even their features may lost during repeated subcultures (Rudi et al.,
1997). Although cyanobacterial identification at genus level is easier to be reached,
particularly where morphological characteristics are significantly different from other
genera, there are some cases where this separation is not clear, namely the delineation
of Microcystis from Synechocystis, Anabaena from Nostoc, as well as Anabaena from
Aphanizomenon and Nodularia.
Figure 4 Schematic representation of cyanobacterial sample collection and morphological divergence of marine Cyanobacteria

43
Table 1a. Microbial diversity of Cyanobacteria Chroococcaceae
1. Chroococcus minor Kutzing
2. Chroococcus sp. Kutzing
3. Gleocapsa sp. Kutzing
4. Synechococcus elongatus Nageli
5. Synechocystis sp. Sauvageau
6. Microcystis marginata Menedhini
7. Merismopedia convvoluta Brebisson
Nostacaceae
8. Nodularia spumigena Mertens
9. Nodularia sp. Mertens
10. Anabaena spiroides Klebahn
11. Anabaena cylindrica Lemmermann
Oscillatoriaceae
12. Phormidium tenue Gomant
13. Phormidium valderianum Gomant
14. Phormidium chlorinum Kutzing
15. Phormidium tenue Meneghini
16. Phormidium fragile Gomant
17. Phormidium subtilissma Kutzing
18. Phormidium angusitissimum
19. Phormidium corium Agardh
20. Phormidium tenue Gomant
21. Oscillatoria subrevis Schmidle
22. Phormidium willei Gardner
23. Oscillatoria formosa Vincent
24. Oscillatoria boryana Bory
25. Oscillatoria subrevis Schmidle
26. Oscillatoria princeps Rabenhorst
27. Oscillatoria tenius Agardh
28. Oscillatoria amphibium Anagnostidis
29. Oscillatoria laete-viriens Crouan
30. Oscillatoria limosa Agardh
31. Oscillatoria princeps Roberhorst
32. Oscillatoria limosa Agardh
33. Oscillatoria curviceps Agardh
34. Oscillatoria boryana Bory and Gomant
35. Oscillatoria princeps Rabenhorst
36. Oscillatoria rubencens De Candolle
37. Oscillatoria tenue Gomant
38. Oscillatoria sp. Bory and Gomant
39. Oscillatoria chalybya Metens
40. Oscillatoria acuminata Gomant
41. Oscillatoria salina Biswas
42. Jaaginema pseudogeminatum Schmid
43. Spirulina subsalsa Oersted
44. Spirulina princeps West and West
45. Spirulina meneghiniana Zanardini
46. Lyngbya majuscula Harvey
47. Lyngbya allergii Kutzing
48. Lyngbya confervoides Agardh
Pseudanabaenaceae
49. Pseudoanabaena sp Lauterborn
Mastigocladaceae
50. Hapalosiphon welwitschii West and
West
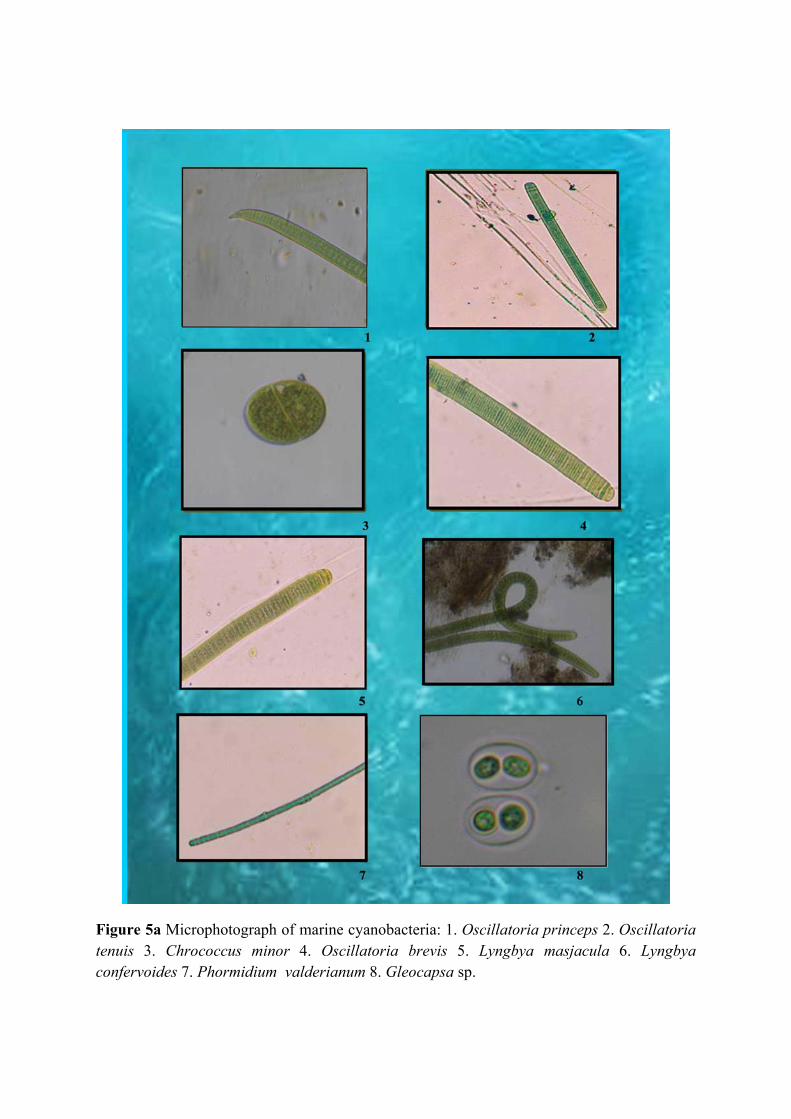
Figure 5a Microphotograph of marine cyanobacteria: 1. Oscillatoria princeps 2. Oscillatoria tenuis 3. Chrococcus minor 4. Oscillatoria brevis 5. Lyngbya masjacula 6. Lyngbya confervoides 7. Phormidium valderianum 8. Gleocapsa sp.
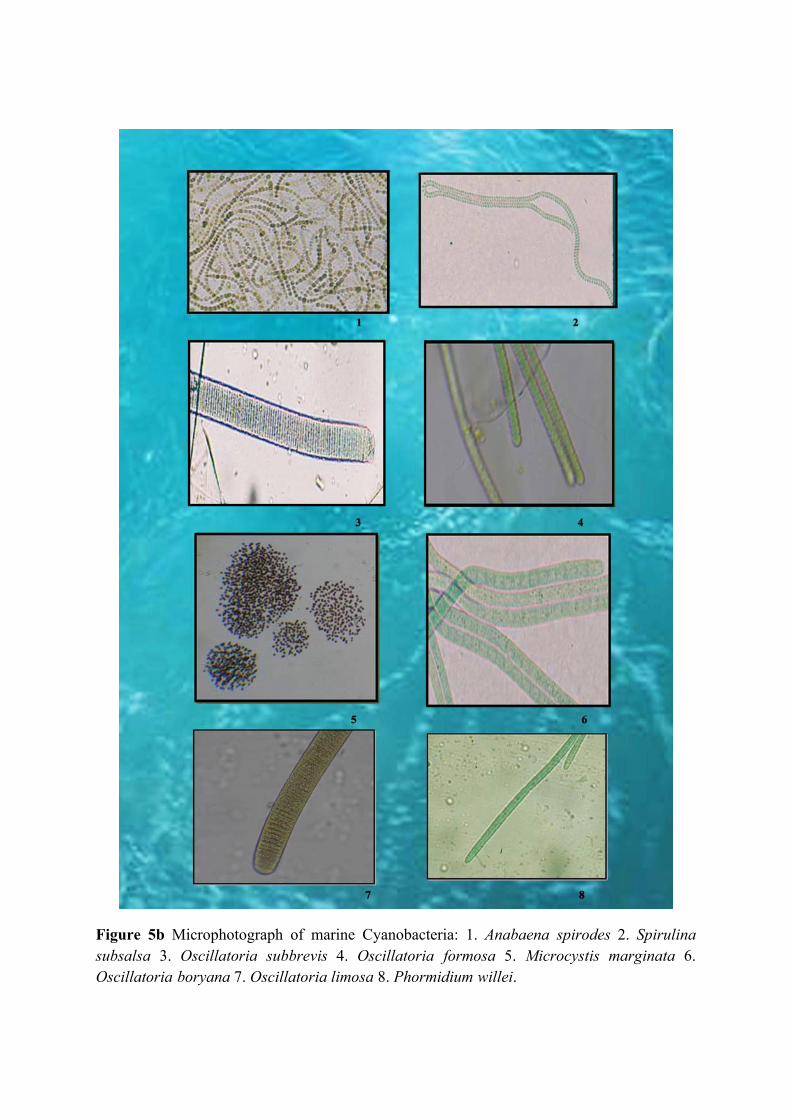
Figure 5b Microphotograph of marine Cyanobacteria: 1. Anabaena spirodes 2. Spirulina subsalsa 3. Oscillatoria subbrevis 4. Oscillatoria formosa 5. Microcystis marginata 6. Oscillatoria boryana 7. Oscillatoria limosa 8. Phormidium willei.

44
Table 1b. Microbial diversity of Diatoms
Achanthaceae
1. Achnanthes gibberula Bory De Saint
2. Achnanthes minutissima Bory
Bacillariaceae
3. Denticula vanheurcki Brun
4. Nitzchia vasnii Smith
5. Nitzchia sp. Smith
6. Nitzchia subtilis Grunow
7. Nitzchia palea Smith
8. Nitzchia sigmoidea Smith
Catenulaceae
9. Amphora sp. Smith
Cymbellaceae
10. Cymbella tumida Agardh
11. Cymbella affinis Agardh
Desmidiaceae
12. Docidium baculum Playfair
Fragilariaceae
13. Fragilaria construens Grunnow
14. Fragilaria sp. Lyngbye
15. Synedra ulna Ehrenberg
16. Synedra acus Ehrenberg
Gomphonemataceae
17. Gomphoneis herculeana Ehrenberg
Mastoglioaceae
18. Mastogloia smithii Smith
19. Matogloia sp. Cleve
Naviculaceae
20. Caloneis silicula Cleve
21. Caloneis lewisii Patrick
22. Navicula andium Muller.
23. Navicula capsidata Kutzing
24. Navicula seminulum Grunow
25. Navicula andium Muller
26. Navicula viridula Ehrenberg
27. Navicula capsidata Kutzing
Pinnulariaceae
28. Pinnularia burkii Ehrenberg
29. Pinnularia subcaptata Gregory
30. Pinnularia burkii Pat
31. Pinnularia sp. Ehrenberg
Pleurosigmataceae
32. Gyrosigma sp. Hassall
33. Pleurosigma acuminatum Grunow
34. Pleurosigma sp. Smith
35. Pleurosigma angulatum Queckett
Stauroneidaceae
36. Stauroneis sp. Ehrenberg
Stephanodiscaceae
37. Cyclotella sp. Kutzing
38. Cyclotella sp. Kutzing

Figure 6 Microphotograph of Diatoms: 1. Stauroneis sp. 2. Navicula palea 3. Synedra ulna 4. Navicula vasni 5. Pinnularia burkii 6. Navicula 7. Navicula andium 8. Gyrosigma 9. Pinnularia subcaptata 10. Gomphonema herculeana 11. Synedra acus 12. Nitzchia subtilis 13. Caloneis silicula 14. Cymbella affinis 15. Caloneis lewisii 16. Cymbella 17. Fragilaria construns 18. Navicula seminulum 19. Achnathes minutissima 20. Navicula virudula 21. Pleurosigma acuminatum 22. Pinnularia sp. 23. Pleurosigma 24. Pleurosigma angulatum

45
In addition, it is difficult to accurately identify cyanobacteria of the order
Oscillatoriales. The identification problems increase further at species level, being of
course impossible to differentiate strains through this approach. Sometimes, even for
someone with experience, misidentification can be obtained, as occurred with an
Aphanizomenon strain, originally classified as Aphanizomenon flos-aquae and later re-
classified as Aph. issatschenkoi with the help of the 16S rRNA phylogenetic
positioning (Neilan, 2002).
Molecular characterization of the marine cyanobacteria and diatoms were done
with 16S rDNA and 18S rDNA sequence. Amplification of 18S rDNA and 16S rDNA
gel picture was shown (Fig. 7), in which clear resolved bands at 2.1kb and 1.6kb were
found in diatom and cyanobacteria respectively. The evolutionary relationships of the
isolates were found out with phylogenetic tree, in which the isolated strain were closely
associated with marine forms of cyanobacteria and some of new reports were found in
case of diatom (Fig 8 & 9).
Molecular phylogenetics is the analysis of hereditary differences in DNA which
gave the information on an organism's evolutionary relationships. The result of a
molecular phylogenetic analysis is expressed in a phylogenetic tree. The first molecular
and phylogenetic characterization of two species of the genus Nostoc - Nostoc linckia
and Nostoc punctiforme based on the cpcB-IGS-cpcA locus of the phycocyanin operon
(Teneva et al., 2005). The phylogenetic position of these two species within order
Nostocales as well as within division Cyanoprokaryota has been determined. The
results indicate that genus Nostoc is heterogeneous. Analysis of the IGS region between
cpcB and cpcA showed that Nostoc and Anabaena are distinct genera, while the
taxonomic status of the genera Aphanizomenon and Anabaena is still unclear, since the
similarity is up to 96.3%. Therefore, additional analyses and revision are required.
Reported molecular and phylogenetic data will be useful to solve other problematic
points in the taxonomy of genera Aphanizomenon, Anabaena and Nostoc.
The ribosomal sequences were deposited to GenBank (NCBI) to record our
isolated diatom and cyanobacterial strains. Accession numbers for the each organism
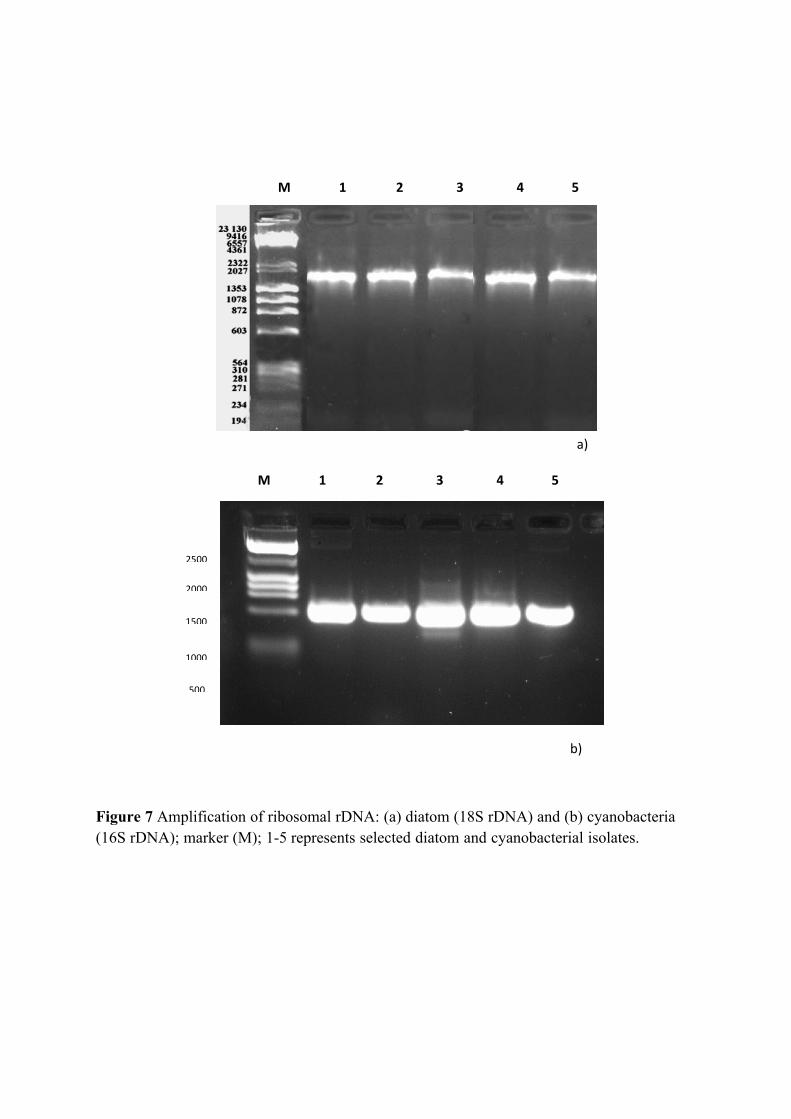
a)
b)
Figure 7 Amplification of ribosomal rDNA: (a) diatom (18S rDNA) and (b) cyanobacteria (16S rDNA); marker (M); 1-5 represents selected diatom and cyanobacterial isolates.
M 1 2 3 4 5
M 1 2 3 4 5
2500
2000
1000
1500
500

Figure 8 Evolutionary relationship of diatom based on the 18S rDNA sequences. Organisms indicated by red color are the selected isolates while black colored ones are retrieved from GenBank (NCBI). Differences between sequences are indicated by the sum of the horizontal length; the scale bar is in fixed nucleotide substitutions per sequence position.

Figure 9 Evolutionary relationship of marine Cyanobacteria based on the 16S rDNA sequences by Maximum Likelihood Method. Organisms indicated by red color are the selected isolates while black colored ones are retrieved from GenBank (NCBI). Differences between sequences are indicated by the sum of the horizontal length; the scale bar is in fixed nucleotide substitutions per sequence position.
EF654085.1| Y18795.1| Spirulina sp. strain MPI S2 16S rRNA gene
JQ347352.1| Uncultured bacterium clone AP60 16S ribosomal RNA gene partial sequence
EF188837.1| Phormidium boryanum BDU 92181 16S ribosomal RNA gene partial sequence
GU585847.1| Phormidium tenue NTDM05 16S ribosomal RNA gene partial sequence
HQ738569.1| Oscillatoria boryana BRIP02 16S ribosomal RNA gene partial sequence
JQ867393 Oscillatoria boryana NTDM11
DQ845200.1| Oscillatoria boryana BDU92181 16S ribosomal RNA gene partial sequence
Leptolynbya valderiana
GU812858.1| Phormidium willei NTDM01 16S ribosomal RNA gene partial sequence
EF654085.1| Phormidium persicinum SAG 80.79 16S ribosomal RNA gene partial sequence
JQ867392 Leptolynbya valderiana NTDM10
GU186898.1| Leptolyngbya valderiana BDU 41001 16S ribosomal RNA gene partial sequence
Y18792.1| Spirulina sp. strain MPI S4 16S rRNA gene
GU585848.1| Spirulina subsalsa NTDM21 16S ribosomal RNA gene partial sequence77 67
69
35 49
29
24
27 16
0.2
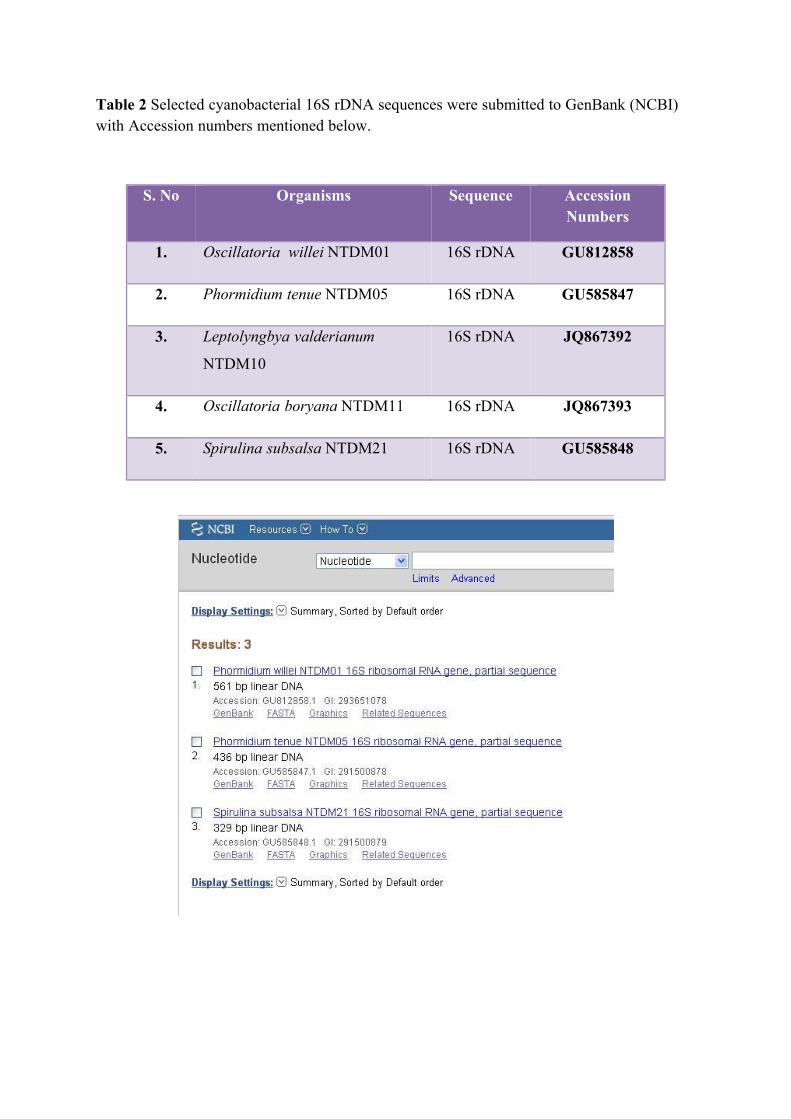
Table 2 Selected cyanobacterial 16S rDNA sequences were submitted to GenBank (NCBI) with Accession numbers mentioned below.
S. No Organisms Sequence Accession Numbers
1. Oscillatoria willei NTDM01 16S rDNA GU812858
2. Phormidium tenue NTDM05 16S rDNA GU585847
3. Leptolyngbya valderianum
NTDM10
16S rDNA JQ867392
4. Oscillatoria boryana NTDM11 16S rDNA JQ867393
5. Spirulina subsalsa NTDM21 16S rDNA GU585848
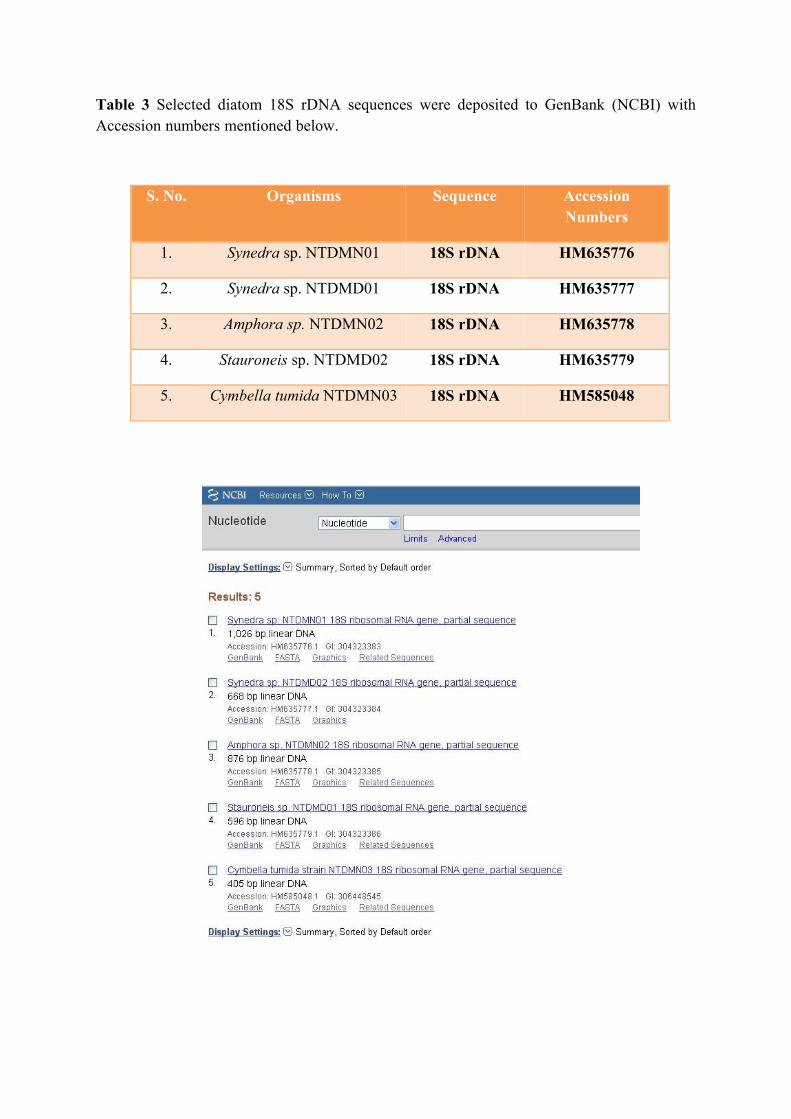
Table 3 Selected diatom 18S rDNA sequences were deposited to GenBank (NCBI) with Accession numbers mentioned below.
S. No. Organisms Sequence Accession Numbers
1. Synedra sp. NTDMN01 18S rDNA HM635776
2. Synedra sp. NTDMD01 18S rDNA HM635777
3. Amphora sp. NTDMN02 18S rDNA HM635778
4. Stauroneis sp. NTDMD02 18S rDNA HM635779
5. Cymbella tumida NTDMN03 18S rDNA HM585048

46
have received from GenBank were tabulated (Table 2 & 3). In addition, C-phycocyanin
gene sequence also been deposited to GenBank with translated protein sequence
(Table 4).
Protein conformational structures are primary, secondary and tertiary structures
were predicted. Among which tertiary structure gives three-dimensional structures in
different isoforms or different structural elucidation such ribbon model, ball and stick
model, space-filling model and so on. A represenatative of the molecular modeling
studies ribbon model and ball and stick models were elucidated with amino acid
sequence (Fig. 11). In this protein model, sequence start with Met aminoacids and ends
with Ile aminoacids.
Conservation domains were analysed in which the query sequence highly
matched with phycoblisome superfamily (Fig 12). It was further analyzed for the
identity with cyanophyceae and chlorphyceae phycocyanin family. Flow-chart for the
determination of cyanobacterial Phycocyanin gene was shown (Fig 13). Molecular
modelling methods are now routinely used to investigate the structure, dynamics,
surface properties and thermodynamics of inorganic, biological and polymeric systems.
The types of biological activity that have been investigated using molecular modelling
include protein folding, enzyme catalysis, protein stability, conformational changes
associated with biomolecular function, and molecular recognition of proteins, DNA,
and membrane complexes (Foster, 2002). The query sequence was closely identical
with cyanobacterial species with 75 % identity, whereas 37% of identity was observed
chlorphyceae algal groups; hence, perhaps it may be used as genetic marker for
development of DNA barcodes for the Cyanobacteria (Fig 14). After performing the
structure prediction of SPDBV software, the final 3D Structure calculated for modelled
structure reactivity, which can provide valuable information of protein structure and
function. The in silico design of drug formulations, plays crucial roles in the biomedical
research. Computational research on biomedics has generated a lot of information
concerning its protein structure and reactivity.
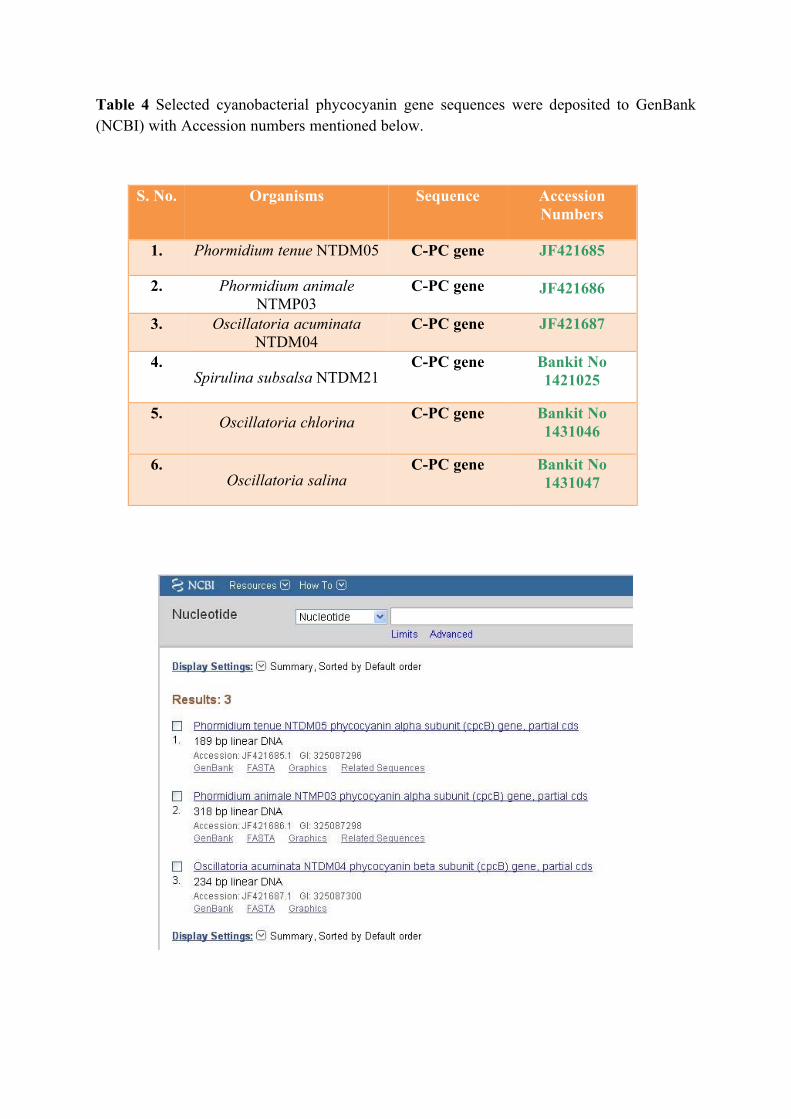
Table 4 Selected cyanobacterial phycocyanin gene sequences were deposited to GenBank (NCBI) with Accession numbers mentioned below.
S. No. Organisms Sequence Accession Numbers
1. Phormidium tenue NTDM05 C-PC gene JF421685
2. Phormidium animale NTMP03
C-PC gene JF421686
3. Oscillatoria acuminata NTDM04
C-PC gene JF421687
4. Spirulina subsalsa NTDM21
C-PC gene Bankit No 1421025
5. Oscillatoria chlorina
C-PC gene Bankit No 1431046
6. Oscillatoria salina
C-PC gene Bankit No 1431047
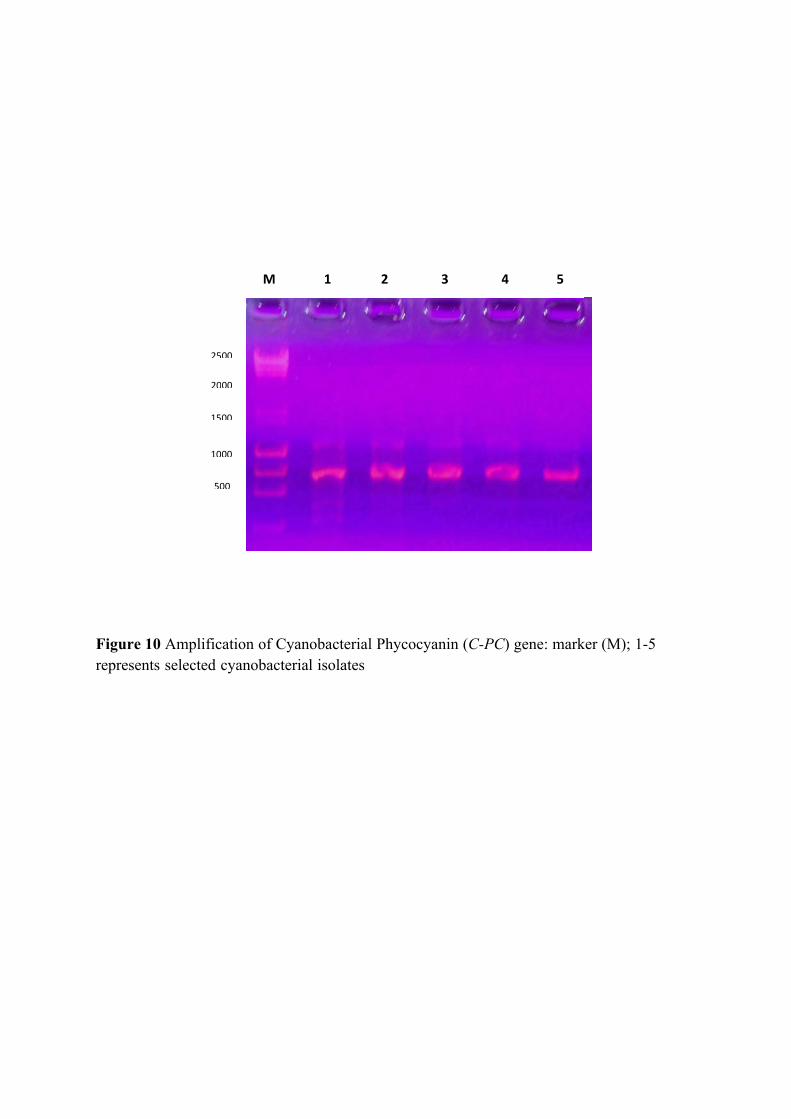
Figure 10 Amplification of Cyanobacterial Phycocyanin (C-PC) gene: marker (M); 1-5 represents selected cyanobacterial isolates
M 1 2 3 4 5
2500
2000
1000
1500
500

a)
b)
Figure 11 Three- Dimensional structure of C-Phycocyanin gene: ribbon model (a) and Ball and stick model with 63 amino acid alpha chain (b)
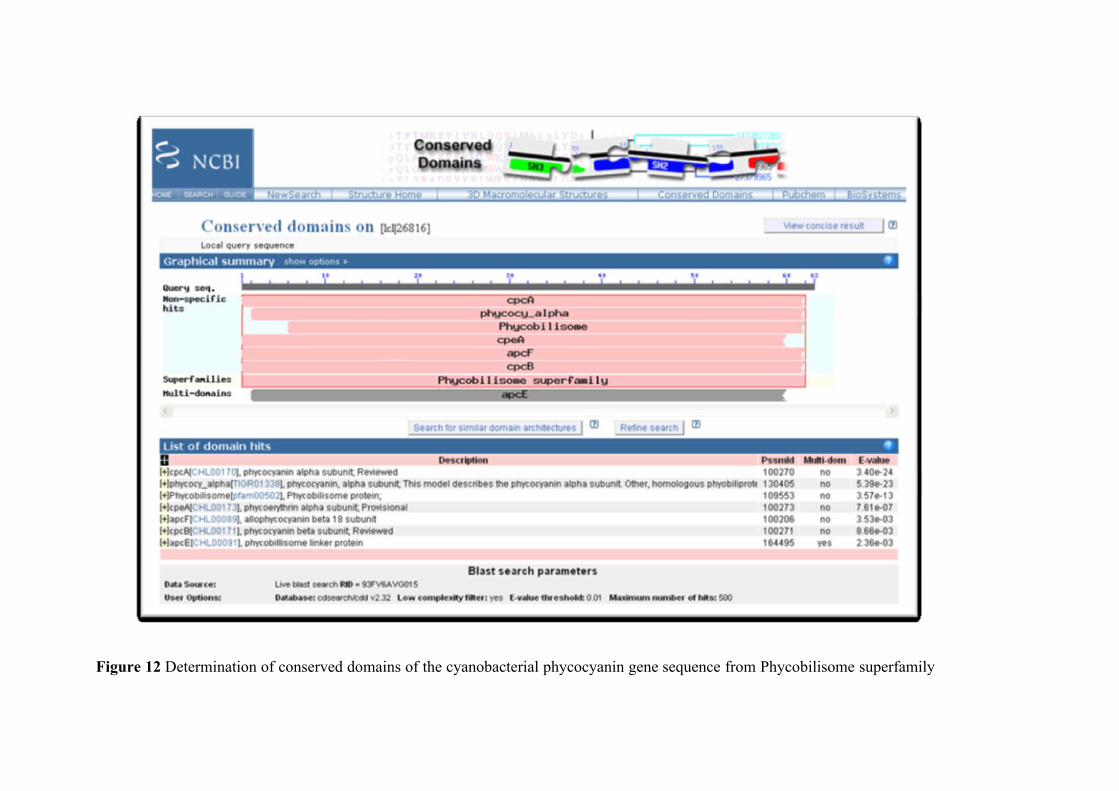
Figure 12 Determination of conserved domains of the cyanobacterial phycocyanin gene sequence from Phycobilisome superfamily
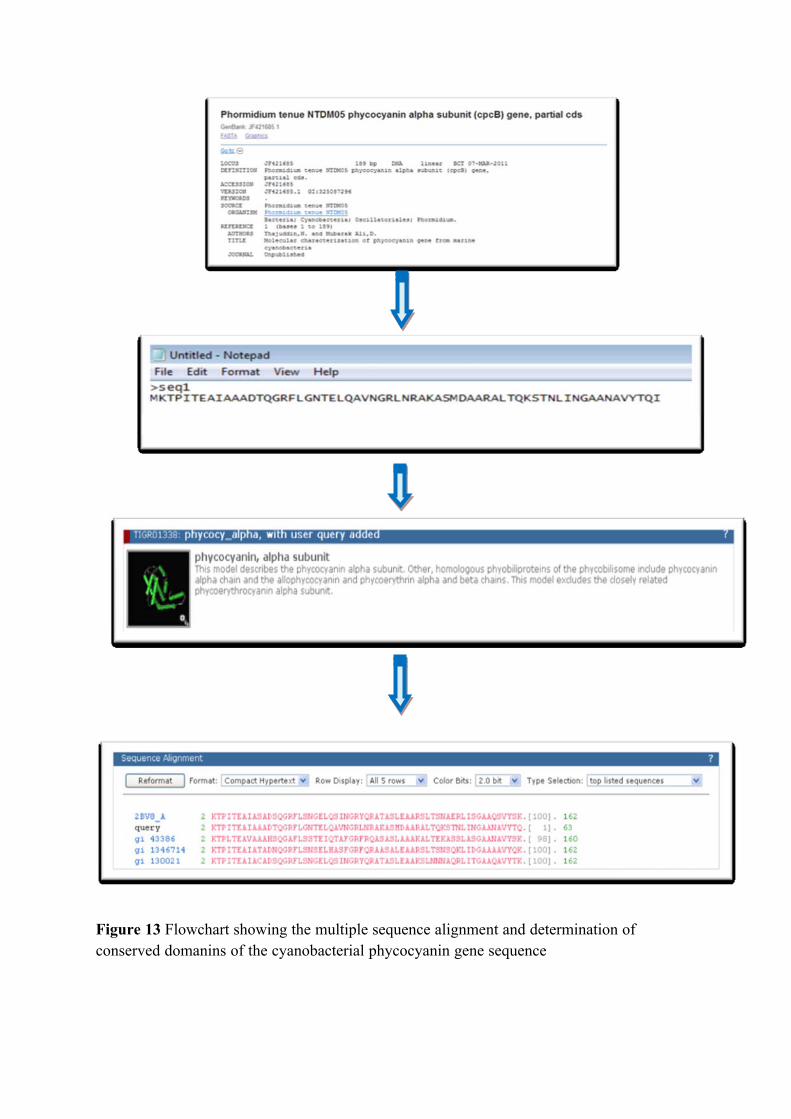
Figure 13 Flowchart showing the multiple sequence alignment and determination of conserved domanins of the cyanobacterial phycocyanin gene sequence
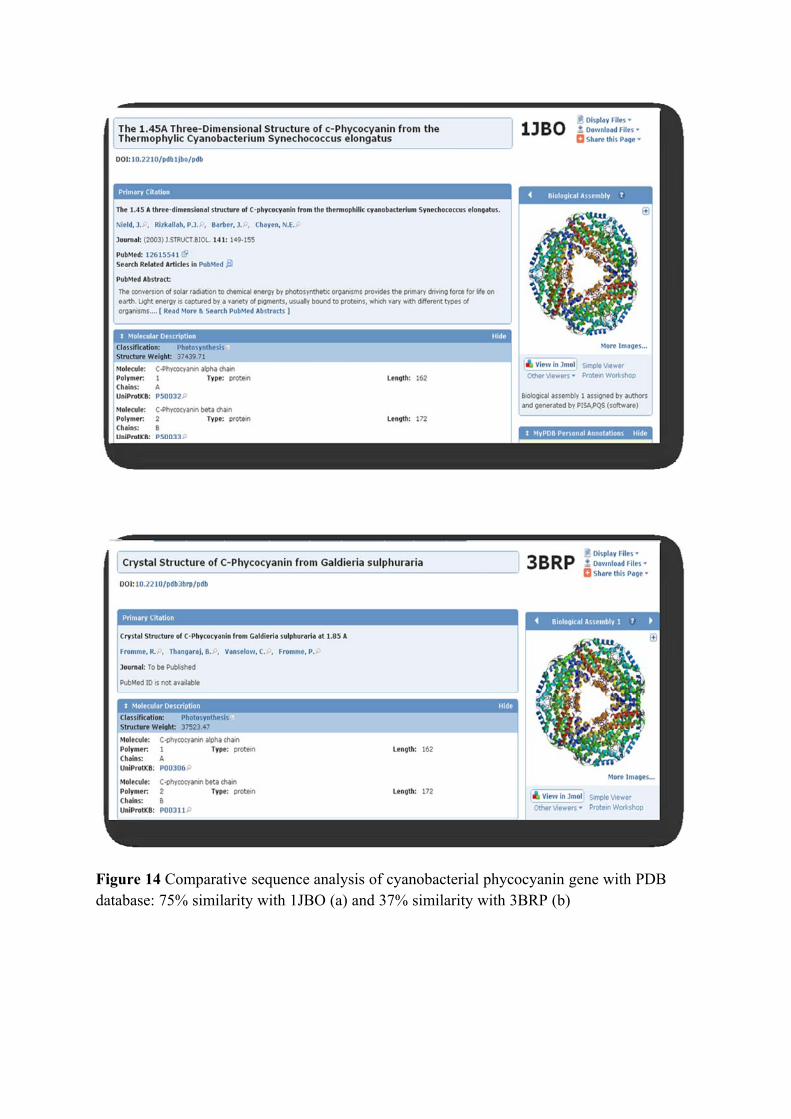
Figure 14 Comparative sequence analysis of cyanobacterial phycocyanin gene with PDB database: 75% similarity with 1JBO (a) and 37% similarity with 3BRP (b)

Chapter II
Partial Biochemical Characterization of Cyanobacterial Phycobiliproteins

47
INTRODUCTION
Cyanobacteria are photosynthetic microorganisms, which carry out
photosynthesis similar to higher plants. However, instead of chloroplasts they contain
membranous structures called thylakoids, where phycobilisomes are found. These
microorganisms only contain chlorophyll a and almost 50% of the light required is
captured by the phycobiliproteins. Phycobiliproteins are antenna pigments, light
harvesting, and consist of chromophores called bilins, which are attached to cysteine
residues of the apoprotein (Samsonoff and MacColl, 2001).
The assembly of the phycobilisome is stabilized by a group of polypeptides
named ‘linker’ polypeptides (L). They induce a face-to-face aggregation of
phycoerythrin or phycoerythrocyanin and phycocyanin trimers. They additionally cause
the tail-to-tail joining of hexameric assemblies of these biliproteins to form larger
aggregates such as peripheral rods and core cylinders. Allophycocyanin assemble into
phycobilisome core with the assistance of three types of linker polypeptides: the large
core-membrane linker polypeptide (LCM) involved in the interaction of PBS core with
the thylakoids, rod-core linker polypeptides (LRC) that mediate the attachment of the
peripheral rods to the PBS core and the small core-linker polypeptides (LC) which
participate in the assembly of core substructure.
The peripheral rods of the phycobilisomes are composed of phycoerythrin or
phycoerythrocyanin and phycocyanin in association with the appropriate rod-linker
polypeptides (LR). Phycoerythrin hexamers of some marine cyanobacteria and red
algae contain a third type of subunit γPE in the central cavity of the hexamer. These
subunits are bifunctional phycobiliproteins that act as light harvesting phycobiliproteins
and linker polypeptides (Sidler 1994). The morphology of phycobilisomes varies with
the organisms. These particles may be ellipsoidal, hemidiscoidal, or bundles of rod
shaped elements. The differences in gross morphology do not reflect fundamental
differences in the placement of the major phycobiliproteins or in the fundamental
properties of the particle (Glazer 1989).

48
In contrast to the proteins of the cyanobacteria and of the red algae, the
cryptomonad phycobiliproteins are unusual in several ways such as (1) phycobilisomes
are lacking due to the formation of tetrameric complexes and therefore not possible to
form phycobilisomes, (2) each species of cryptomonad contains only one type of
phycobiliproteins either phycoerythrin or phycocyanin, (3) the proteins are found on the
lumenal rather than the stromal side of the thylakoid membrane, (4) the proteins form
dimers, rather than trimers or hexamers, and (5) they contain several unusual bilins, for
example in the cryptophycean biliprotein phycocyanin 645, three chemically different
bilins such as Cys-bilin 618, DiCys-bilin 584 and Cys-bilin 584 are present (Becker et
al. 1998; Wedemayer et al. 1991).
Cyanobacterial phycobiliproteins have gained importance in the commercial
sector, as they have several applications. The primary potential of these molecules are
as natural dyes but a number of investigations have shown on their health-promoting
properties and broad range of pharmaceutical applications. Thus, one of the
applications of phycocyanin is to use as food pigment replacing current synthetic
pigments. They are used as a colourant in chewing gum, ice sherberts, popsicles,
candies, soft drinks, dairy products and cosmetics like lipstick and eyeliners. In
addition, phycobiliproteins are widely used in clinical and immunological research
laboratories (Spolaore et al. 2006). They serve as labels for antibodies, receptors and
other biological molecules in a fluorescence-activated cell sorter, and they are used in
immunolabelling experiments, fluorescence microscopy and diagnostics.
The major organisms exploited for production are the cyanobacterium,
Spirulina for phycocyanin and the red alga Phorphyridium for phycoerythrin (Roman et
al. 2002). The purpose of the review is to assess the wealth of knowledge on the
applications of phycobiliproteins and the technological backup available for
commercial venturing. This can be envisaged by analyzing application oriented
research, patents and commercial activities. A compilation of this knowledge and its
analysis will be of immense use for attaining further improvements.

49
The phycocyanins have an apparent molecular mass of 140–210 kDa and two
subunits, α and β. Studies of crystal structure in phycocyanin have shown that all of
these have a very similar three-dimensional structure. The subunits are associated in an
(α β) protomers, which in turn can be associated in trimmers (α β)3 and hexamers (α β)6
(Stec et al., 1999). The chromophores confer a characteristic colour of each
phycobiliprotein: phycocyanin (blue brilliant), phycoerythrin (red) and allophycocyanin
(green–blue). The characteristic coloration of the phycobiliproteins, make them
attractive to food and cosmetic industries as natural pigments (Santos et al., 2004).
Objectives:
In this chapter, work is emphasized on the biochemical characterization of C-
Phycobiliprotein (specifically, C-PE) with following process.
• Selection of cyanobacterial strains for the pigment production
• Extraction of C-Phycobiliproteins from selected Cyanobacteria
• Spectrometric quantification of C-Phycobiliproteins
• Optimization of physical and chemical methods
• Study on effect of preservatives on the C-Phycoerythrin
• Biomedical application of C-Phycoerythrin

50
REVIEW OF LITERATURE
Cyanobacterial Pigments:
Cyanobacteria are the pioneer oxygenic phototrophs on earth whose distribution
around the world is surpassed only by bacteria (Adams et al., 2000). Worldwide
attention is drawn towards cyanobacteria for their possible use in mariculture, food,
feed, biopolymer, biofuel and bio-fertilizer, production of various secondary
metabolites including vitamins, toxins, enzymes, pharmaceuticals, pharmacological
probes and pollution abatement (Thajuddin and Subramanian, 2005 and Shrivastav et
a., 2010). Phycobiliproteins, including phycoerythrin, phycocyanin and
allophycocyanin are a class of chromo-protein bearing covalently bound linear
tetrapyrole chromophore.
Phycobiliproteins are the major photosynthetic accessory pigments in
cyanobacteria (prokaryotic); rhodophytes (red algae, eukaryotic); cryptomonads
(biflagellate unicellular eukaryotic algae) and cyanelles (endosymbiotic plastid-like
organelles). Phycobiliproteins are brilliant-colored and water-soluble antennae-protein
pigments organized in supramolecular complexes, called phycobilisomes (PBSs),
which are assembled on the outer surface of the thylakoid membranes. The colors of
phycobiliproteins originate mainly from covalently bound prosthetic groups that are
open-chain tetrapyrrole chromophores bearing A, B, C and D rings named phycobilins.
There are four main classes of phycobiliproteins – allophycocyanin (APC bluish green),
phycocyanin (PC: deep blue), phycoerythrin (PE: deep red) and phycocyanobilin (PCB:
orange) (Hemlata et al., 2011).
Classification of Phycobiliprotein:
They are classified by their spectral properties as Phycoerythrobilin (PEB,
Amax 560 nm), Phycocyanobilin (PCB, Amax 620–650 nm), Phycobiliviolin (PXB,
Amax 575 nm) and Phycourobilin (PUB, Amax 498 nm). There are three main types of
phycobiliproteins, phycoerythrin (PE), phycocyanin (PC), and allophycocyanin (APC).
PE is at the tip of the rod-like phycobilisomes, PC is in the middle, while APC forms a
core attached to the reaction and energy transfer proceeds successively from

51
PE→PC→APC→chlorophyll a. Phycobiliproteins are chromo-proteins that have anti-
inflammatory, hepato-protective and antioxidant properties, it can also play important
role in photodynamic action on cancerous tumors and leukemia treatment (Khan et al.,
2005; Kulshreshtha et al., 2008; Vadiraja et al., 1998; Patel et al., 2006 and Paul et al.,
2006).
Cyanobacteria with high levels of specific phycobiliproteins are of commercial
interest. The primary potential of these molecules are as natural dyes in food industry.
A number of investigations have shown their health-promoting properties and
pharmaceutical applications.
Phycocyanin and phycoerythrin are the two currently used natural pigmented
proteins having versatile applications. They are stable at low temperature with some
preservative like citric acid, in acidic and basic solutions and therefore, they could be
utilized as food colorant (chewing gum, jellies), health drinks, beverages and coloring
agent in sweet confectionary and cosmetics (Patel., 2004; Mishra et al., 2008 and
Eriksen, 2008). Of late, more attention is given to the application utilizing its
biochemical and biophysical structural properties. These natural pigments are also used
in fluorescent labeling of antibodies that are applied in diagnostic kits in immunology,
cell biology and biomedical research (Sekar and Chandramohan, 2008).
Nutritional requirements for production of c-Phycobiliproteins:
Macronutrients such as nitrogen and phosphorus and micronutrients such as iron
and molybdenum are known to play a large part in regulating the metabolism and
ultimately the growth of cyanobacteria (Carr and Whitton 1982, Carpenter 1983).
Nitrogen is an essential element for plant growth, required for the synthesis of
amino acids, proteins, nucleotides, nucleic acids, coenzymes, chlorophyll and other
photosynthetic pigments (Raven et al., 1992). Most cyanobacteria are nitrogen fixers,
converting atmospheric nitrogen to ammonia via the enzyme nitrogenase (Postgate,
1987). The resulting ammonia is assimilated into glutamate through the reductive
amination of α-ketoglutarate, by the enzyme glutamate synthase (GOGAT) (Marscher,

52
1995). The glutamate is subsequently catalysed by glutamine synthetase (GS) and
transformed to glutamine, which is then utilised in the synthesis of organic compounds
(Garrett & Grisham, 1999).
Phosphorus is essential for energy-carrying phosphate compounds (ATP, ADP,
NADPH and NADP), nucleic acids, several essential coenzymes and phospholipids
(Marsher 1995). The orthophosphate ion, PO4 3-, is the principal form of phosphorus
taken up by marine plants. Other sources of phosphorus include inorganic
polyphosphates and organic-phosphorus compounds; however, both of these forms
require complex extracellular cleavage and hydrolysis, leaving orthophosphate ions as
the preferred source of phosphorus for uptake (Lobban & Harrison, 1994). Upon
uptake, the orthophosphate ion is esterified with hydroxyl groups of sugars and
alcohols or bound to another phosphate group (Mengel & Kirkby, 1987).
Uses of Phycobiliproteins
As colorant
There is an increasing demand for natural colors which are of use in food,
pharmaceuticals, cosmetics, textiles and as printing dyes. However, their utility is
limited to few of these since the natural dyes have low tinctorial values and persistence.
Due to the toxic effect of several synthetic dyes, there is an increasing preference to use
natural colors for various end uses. Phycobiliproteins are used as a natural protein dye
in the food industry (C-phycocyanin) and in the cosmetic industry (C-phycocyanin and
R-phycoerythrin).
Phycocyanin derived from Spirulina platensis is used as a natural pigment in
food such as chewing gum, dairy products and jellies (Santos et al. 2004). Despite its
lower stability to heat and light, phycocyanin is considered more versatile than gardenia
and indigo, showing a bright blue color in jelly gum and coated soft candies (Lone et
al. 2005). They are also used in coloring of many other food products such as
fermented milk products, ice creams, soft drinks, desserts, sweet cake decoration, milk
shakes and cosmetics. The shade of blue color produced from the red microalga
Phorphyridium aerugineum does not change with pH. The color was stable under light,

53
but sensitive to heat. Within a pH range of 4 to 5, the blue color produced is stable at
60°C for 40 min. This property was important for food uses, since many food items are
acidic, particularly drink and confections. The blue color was added to beverages
without heat application (Pepsi® and Bacardi Brezzer®) which did not lose their color
for at least 1 month at room temperature. The color was very stable in dry preparations.
Sugar flowers for cake decoration maintained their colors for years of storage. Foods
prepared with the phycobiliproteins include gelatin and ice cream. In addition to its
coloring properties, phycoerythrin possesses a yellow fluorescence.
Opportunities for exploiting this property for special effects were studied. A
range of foods that fluoresce under natural light and UV light were prepared and tested.
These include transparent lollipops made from sugar solution, dry sugar-drop candies
for cake decoration (that fluoresce under UV light), and soft drinks and alcoholic
beverages that fluoresce at pH 5–6. Fluorescent color has also been added to alcoholic
beverages containing up to 30% alcohol but the shelf life for such products is short
(Dufosse et al. 2005).
As fluorescent agent
Phycobiliproteins play an important role in fluorescent based detection systems,
particularly for flow cytometry. The spectral properties, such as (1) excitation and
emission at the red end of the spectrum, where interference from biological matrices
tend to be less, (2) a large Stokes shift, so that interference from Rayleigh and Raman
scatter and other fluorescing components is less significant or nonexistent, (3)
immunity from quenching by naturally occurring biological substances, (4) high
solubility in aqueous environment so that non-specific binding effects are minimal, and
(5) fluorescence quantum yield independent of pH of phycobiliproteins, particularly
R-phycoerythrin (R-PE) and allophycocyanin (APC), have made them dominant
reagents in this class of flurochromes (Kronick and Grossman 1983).
Synthesis of conjugates of phycobiliproteins with molecules having biological
specificity, like immunoglobulins, protein A, biotin and avidin, were reported and
showed that phycobiliproteins conjugates are excellent reagents for two color

54
fluorescence analysis of single cells using fluorescence activated cell sorter (FACS) (Oi
et al. 1982). The stabilized phycobilisomes designated PBXL-3L was accessed as a
fluorochrome for flow cytometric immunodetection of surface antigens on immune
cells (Telford et al. 2001b). The lesser-known but potentially important series of low-
molecular weight cryptomonad-derived phycobiliproteins are evaluated for their
applicability to flow cytometry both in extracellular and intracellular labeling
applications (Telford et al. 2001a).
Fluorescent labeling reagents are an essential component of a huge industry
built on sensitive fluorescence detection. This powerful multi-chromophore protein-
labeling reagent has changed the flow cytometry industry and permitted sensitive two-
color lymphocyte subset analysis with a single argon ion laser. Phycoerythrin excites
very strongly at 488 nm but the emission is shifted to 580 nm by energy transfer events
between chromophores within the protein.
Phycoerythrin as a second color works well with fluorescein- labeled
antibodies. They can also be excited at 488 nm but are detected separately at 525 nm to
provide the second colour of detection. HIV monitoring and cancer diagnostics were a
strong driving force for the growth of this reagent. Three- and four-color analyses with
a single laser are now commonplace with phycobiliprotein-based energy transfer
reagents. Low molecular weight fluorescent labels such as cyanine dyes Cy5
(Indodicarbocyanine) and Cy7 (Indotricarbocyanine) are good acceptors from excited
PE and shift emission farther to red and deep red wavelengths and 11- color analyses
are now possible (Waggoner 2006).
PE is also a very important reagent in proteomics and genomics and form the
basis of the detection system in Affymetrix chips (DNA microarrays). Phycoerythrin
labeled streptavidin is added after complete binding and produces a strong signal from
array elements containing the biotin-labeled DNA or protein probes (De Rosa et al.
2003).

55
As pharmaceutical agent
In the last decade, the screening of micro algae, especially the cyanobacteria,
for antibiotics and pharmaceutically active compounds has received ever increasing
interest (Borowitzka 1995). The pharmacological property attributed by phycocyanin
includes antioxidant, anti-inflammatory, neuroprotective and hepatoprotective activity.
When it was evaluated as an antioxidant in vitro, it was able to scavenge alkoxyl,
hydroxyl and peroxyl radicals, and inhibits microsomal lipid peroxidation induced by
Fe+2 –ascorbic acid or the free radicals initiators 2, 2′ Azobis (2- amidinopropane)
dihydrochloride, (AAPH).
They also reduced edema, histamine (Hi) release, myeloperoxide (MPO)
activity and the levels of prostaglandin (PGE2) and leukotriene (LTB4) in the inflamed
tissues. Phycocyanin also reduces the levels of tumor necrosis factor (TNF-α) in the
blood serum of mice-treated with endotoxin and it showed neuroprotective effects in
the rat cerebellar granule cell cultures. Aphanizomenon flos-aquae (AFA) as a source of
phycocyanin have been described as a strong antioxidant (Bhat and Madyastha 2000;
Romay et al. 2003). The involvement of phycocyanin in the antioxidant protection of
the AFA extract against the oxidative damage was demonstrated in vitro (Benedetti et
al. 2004). In red blood cells, oxidative hemolysis and lipid peroxidation induced by
aqueous peroxyl radical generator, AAPH were significantly lowered. In plasma
samples, they inhibited the extent of lipid oxidation induced by pro-oxidant agent
cupric chloride (CuCl2). Such findings about the phycocyanin consider the potential
benefits in the prevention of many pathological disorders associated with oxidative
stress and inflammation. Allophycocyanin was found to inhibit enterovirus 71- induced
apoptosis. At concentration nontoxic to the host cells (0.045±0.012 μM),
allophycocyanin was found to inhibit enterovirus 71-induced cytopathic effects, viral
plaque formation, and viral-induced apoptosis (Shih et al. 2003).
In addition, inhibitory effect of phycocyanin from Spirulina platensis on the
growth of human leukemia K562 cells in a dose- and time-dependent manner was
reported (Liu et al. 2000). R-PE subunits were indicated as an attractive option for
improving the selectivity of photodynamic therapy (PDT). Mouse tumor cells S180 and

56
human liver carcinoma cells SMC 7721 cells was treated with RPE subunits (Bei et al.
2002). C-Phycocyanin derived from Spirulina platensis powerfully influenced serum
cholesterol concentrations and imparted a stronger hypocholesterolemic activity
(Nagaoka et al. 2005). Phycocyanin also exerts hepatoprotective and anti-inflammatory
effects in a human hepatitis animal model. It reduced alanine amino transferase (ALT),
aspartate amino transferase (AST) and malondialdehyde (MDA) in the serum
(Gonzalez et al. 2003).
In addition, at a concentration of 0.25 mg/ml, influence of C-phycocyanin on
hepatocellular parameters related to liver oxidative stress and kupffer cell functioning
indicated that C-phcocyanin elicite a concenteration-depedent inhibition of carbon
phagocytosis and carbon-induced O2 uptake by perfused livers, with a 52% diminution
in the carbon-induced sinusoidal release of lactate dehydrogenase. C-phycocyanin also
suppressed the 3,3′,5-triiodothyronine (T3) induced increase in serum nitrite levels and
in the activity of hepatic nitric oxide synthase (NOS) (Remirez et al. 2002).
Importance of c-phycobiliproteins:
Phycoerythrin is good for human health, having antioxidant, radical scavenging,
anti-inflammatory and anticancer properties. C-PE can be used as nutrient ingredients
and natural dyes for food and cosmetics besides, being potential therapeutic agent in
oxidative stress-induced diseases and as fluorescent marker in biomedical research and
currently used as a natural food colorant in China (Li and Qi, 1997; Romay et al., 1998;
Gonzalez et al., 1999; Olvera-Ramirez et al., 2000; Bhat and Madhyastha, 2000;
Estrada et al., 2001; Tseng, 2001; Soni et al., 2008 and Mishra, 2007). Although,
several methods have been developed for the separation and purification of C-PE from
cyanobacteria, the purity and recovery is relatively low because, it is highly sensitive to
light, oxygen and moisture, hence, it is needed to process it along with efficient
preservatives. Sodium azide (NaN3) and dithiothreitol (DTT) are commonly used as
preservatives in phycoerythrin for analytical purpose, but they are toxic; so for
developing process of food grade C-PE only edible preservatives with unique
properties can be used. It is therefore, desired to develop a simple, but more well-

57
organized method for the separation, purification and stabilization of the food grade C-
PE from cyanobacteria (Mishra et al., 2010).
Extraction of Phycobiliproteins:
Each microorganism has particular characteristics referring to the location of
intracellularly produced proteins, meaning that the molecule of interest might be
located in the cytoplasm, periplasm or even be stored in some cellular organelle, such
as in the mitochondria. Hence, the extraction protocol could vary according to the
desired protein. In general, the extraction method is the key for maximum recovery of
phycobiliproteins in the natural state from algae (Niu et al., 2006).
The extraction of phycobiliproteins involves cell rupture and release of these
proteins from within the cell. The cell walls of cryptophytes are easily disrupted, but
those of cyanobacteria are extremely resistant (Siegelman and Kycia, 1978). Thus, the
use of variations in the osmotic pressure, abrasive conditions, chemical treatment,
freezing-thawing and sonication, amongst other disruption methods, are necessary.
Mechanical cell disintegration methods are currently preferred for large-scale
operations (Gacesa and Hubble, 1990; Kula and Schütte, 1987) since a complete
disintegration of the biomass is desired, with high product and activity yields. Some
papers report C-PC extraction from cyanobacterium. Moraes et al. (2010) and Silveira
et al. (2007) studied the optimization of extraction from dried biomass.
The reported methods to extract C-PC from wet biomass include freezing and
thawing (Sony et al., 2008; Sarada et al., 1999; Abalde et al., 1998; Bermejo et al.,
2006), sonication (Bermejo et al., 2006; Abalde et al., 1998), homogenization (Sarada
et al., 1999), lysozyme treatment (Bermejo et al., 2006; Stewart and Farmer, 1984) and
acid treatment (Sarada et al., 1999; Bermejo et al., 2006).
Preservation of Phycobiliproteins:
Although, several methods have been developed for the separation and
purification of C-PC from cyanobacteria, the purity and recovery is relatively low
because, it is highly sensitive to light, oxygen and moisture, hence, it is needed to

58
process it along with efficient preservatives. Sodium azide (NaN3) and dithiothreitol
(DTT) are commonly used as preservatives in C-PC for analytical purpose, but they are
toxic; so for developing process of food grade C-PC only edible preservatives with
specific properties can be used. It is therefore, desired to develop a simple, but more
efficient method for the separation, purification and stabilization of the food grade C-
PC from cyanobacteria (Patel et al., 2005). C-PC can be used as nutrient ingredients
and natural dyes for food and cosmetics besides, being potential therapeutic agent in
oxidative stress-induced diseases and as fluorescent marker in biomedical research and
currently used as a natural food colorant in China (Li and Qi, 1997; Romay et al., 1998;
Romay et al., 1998; Romay et al., 2001; Gonzalez et al., 1999; Bhat and Madhyastha,
2000; Hirata et al., 2000; Estrada et al., 2001 and Tseng, 2001).
Phycobiliproteins extracted from cyanobacteria have received a great attention
due to its potential in biotechnology and biomedical sciences (therapeutic agent and
immunodiagnostic probes) because of their versatile properties, i.e. its fluorescent
nature and radical scavenger activity, which is suitable in photodynamic therapy for
tumor cells (Paul et al., 2006). Their radical scavenger activity makes them potent
antioxidant and anticancer agent; lipid peroxidase activity as an antidote for toxic
metals like chromium; immunomodulator and bio-enhancer activity makes them anti-
inflammatory and antidiabetic (Mishra, 2007; Mishra et al., 2005; Patel et al., 2006
and Patel et al., 2004). Preservatives are required to ensure that manufactured foods or
additives remain safe and unspoiled for a long time (shelf life). The present study
describes the effect of selected food grade preservatives for C-PE in order to develop a
stable product with long shelf life.
Food additives have been used by mankind for centuries, e.g. salt, sugar and
vinegar were among the first to be used as preservatives in food. In the past 30 years,
however, with the advent of processed foods, there has been a massive explosion in the
chemical adulteration of foods with additives. Considerable controversy has been
associated with the potential threats and possible benefits of food additives. Some food
additives are known to be carcinogenic or toxic. Hyperactivity in children, allergies,
asthma, and migraines are often associated with adverse reactions to food additives.

59
Preservative for the food grade C-phycoerythrin is indispensable for making the
process commercially viable due to its sensitivity.
Preservatives are required to ensure that manufactured foods or additives remain
safe and unspoiled for a long time (self life). The present study describes the effect of
selected food grade preservatives for phycoerythrin in order to develop a stable product
with long shelf life. Food additives have been used by mankind for centuries, e.g. salt;
sugar and vinegar were among the first to be used as preservatives in food.
Phycoerythrin can be used as a food colorant which is not only safe but, has an extra
advantage of being potential antioxidant. Preservative for the food grade phycoerythrin
is indispensable for making the process commercially viable due to its extreme
sensitivity towards light oxygen and temperature under aqueous condition.

60
MATERIALS AND METHODS
Materials:
All the chemicals used in this study were procured from Sigma, Hi-Media and
Qualigens. All buffers and reagents used in this study were prepared in double distilled
water.
Selection of Cyanobacterial Strain:
Among isolated marine cyanobacteria described in Chapter I, preliminary
screening was done for pigment production. Among them, a marine cyanobacterium,
Phormidium tenue NTDM05 has been selected for quantification of phycobiliproteins
and further studies. A marine cyanobacterium was mass cultured raised in ASN-III
Medium (Stanier et al., 1971). Culture was allowed to grow in light intensity provided
by cool-white fluorescent tubes of 50 µmol photons.m-2.s-1 following 12:12 hour, light
and dark regime at 30oC ±1oC.
Extraction and quantification of Phycobiliproteins:
Cyanobacterial cells were harvested by centrifugation (10,000g for 15 min).
Adhered salts were removed by washing with double distilled water and then biomass
was dried (50ºC). Biomass was homogenized with phosphate buffer and repeated
freezing and thawing was done in dark. The mixture was subsequently centrifuged
(10,000g for 20 min, 4ºC) to separate the phycobiliprotein containing clear supernatant.
Absorbance of supernatants was measured at wavelength 620, 652, and 562 nm
(Optizen Scan UV–VIS spectrophotometer) for C-phycocyanin, allophycocyanin and
phycoerythrin respectively (Siegelman and Kycia. 1978).
The amount of C-PC, C-PE and C-APC in the sample was calculated using
simultaneous equations of Bennet and Bogorad (1973) as follows:
Phycocyanin (mgmL−1) = [A620 −0.474 (A652)]/5.34
Allophycocyanin (mgmL−1) = [A652 −0.208 (A620)]/5.09
Phycoerythrin (mgmL−1) = [A568 −2.41 (PC) −0.849 (APC)]/9.62

61
Extraction of C-Phycoerythrin:
Cyanobacterial cells harvested by whatman filter paper (No.1) filtration were
washed with phosphate buffer (pH 7.2) and lyophilized. Two grams of freeze dried
biomass was suspended in 50 ml of sodium–phosphate buffer (0.1 M, pH 7.2,).
Repeated freezing at −20 ◦C and thawing at room temperature 25±5 ◦C was done in the
dark, The mixture was subsequently centrifuged at 10,000×g for 30 min at 0±5 ◦C and a
phycoerythrin containing clear supernatant was collected and precipitated by
ammonium sulphate fractional precipitation (25 and 60%), The pellet was suspended in
water followed by dialysis against distilled water at 0±5 ◦C for 48 h in dark, by
intermittent changing of distilled water at regular intervals and freeze dried. The
complete process for extraction of phycoerythrin is shown in Fig. 1.
Phormidium tenue NTDM05
Grown in ASN and MN-III medium
Harvesting the biomass for extraction
Extraction in Phosphate Buffer system by simple freeze and thaw method
(12h freeze and 12h thawing)
Cell debris removal by centrifugation
Ammonium sulphate precipitation
Dialysis
Stored at -4oC
Lyophilized
Figure 1 Process for extraction of C-Phycoerythrin from Phormidium tenue NTDM05

62
Ammonium sulfate fractionation:
The crude extract to obtain 20% saturation of finely powdered ammonium
sulfate was gradually added with continuous stirring for 1 h. The resulting solution was
kept overnight and centrifuged at 17,000 x g for 20 min. The supernatant was pooled
and subjected to 70% ammonium sulfate saturation in a manner similar to that of 20%
saturation. After overnight incubation, the solution was centrifuged at 10,000 rpm for
20 min. The pellets were resuspended in a small quantity of 20 mM Tris–Cl buffer (pH
8.1) and subjected to dialysis for 48 h against 100 times volume of the same buffer,
with a change of buffer three times.
Optimization of the production of C-Phycobiliproteins:
Physical and chemical methods were adopted for the higher production of c-
phycobiliproteins in selected cyanobacteria.
Physical methods:
In this method, different methods were adopted, firstly, extraction methods such
as effect of different cell disruption methods like homogenization (Oranda et al. 1978),
sonication (Furuki et al. 2003), heat shock (Soni et al. 2006) and freezing and thawing
(Soni et al. 2006) on phycobiliprotein yield. Secondly, temperature, where 20 – 60oC
were adopted.
Chemical methods:
In this method, micro nutrients such as Magnesium sulphate (4, 4.5, 5, 5.5,
6mg/l), Sodium carbonate (20, 40, 60, 80, 100mg/l), Potassium phosphate (20, 40, 60,
80, 100mg/l), Sodium Nitrate (1.5, 2.25, 3., 3.75, 4.5, 5.25, 6mg/l) are important
nutrients for the production of phycobiliproteins specially, C-phycoerythrin, in buffer
system Sodium phosphate buffer (pH-7.0) of 0.1M, Tris buffer (pH-7.2), Phosphate
buffered saline, HEPES and double distilled water was used. Finally, pH (6.0, 6.5, 7.0,
7.5 & 8.0) of medium was checked on phycobiliprotein yield.

63
Experimental selection of effective preservatives:
The stability of C-phycoerythrin in aqueous forms was checked by adding
different preservatives viz. Sucrose, Calcium chloride, Sodium chloride and Citric acid,
kept in two different conditions,
(1) At 0±5 ◦C in the refrigerator
(2) At room temperature 35±5oC
For checking efficacy of preservatives sucrose (0.1 g), Calcium chloride (0.1 g),
Citric acid (0.1 g), Sodium chloride (0.1 g) was added in 25 ml phycoerythrin solution
[5mg of freeze dried c-phycoerythrin was dissolved in 25 ml of sodium phosphate
buffer (0.1 M, pH 7.2)] separately and kept at 0±5 ◦C and 35±5 ◦C for study.
The absorbance of C-phycoerythrin solution containing preservative along with
the control without preservative was measured after regular interval, up to 30 days, on a
Optizen Scan UV-Vis, NIR spectrophotometer at wavelength 250–700nm for
calculating the concentration of C-Phycoerythrin using appropriate equation (Bennetts
and Bogorad, 1973).
Antibacterial activity of extracted C-Phycobiliprotein:
Antibacterial activity of extracted C-Phycobiliprotein was analyzed by well –
diffusion method with human pathogens such as E. coli, Enterobacter sp.,
Pseudomonas sp., Vibrio cholerae, Klebsiella pneumoniae. 24h cultures of human
pathogens were streaked on Muller Hinton Agar (MHA) and wells were formed in
15mm depth on the agar plate. Extracted phycobiliprotein was inoculated on the well in
different concentration such as 1mg/ml, 10mg/ml and 100mg/ml. The plates were
incubated at 37oC for 24h and extended to 48h. The antibacterial effect of extracted
phycobiliprotein were documented and discussed.
Determination of fluorescence property:
Fluorescence property of extracted C-Phycobiliprotein was analyzed in
Fluorescence spectroscopy and Uv-Transilluminator. An aliquot of extracted C-
phycobiliprotein from Phormidium tenue NTDM05 was tested for fluorescence
property for the application biolabelling purpose.

64
RESULTS AND DISCUSSION
One of the most important requirements for obtaining phycobiliproteins from
Cyanobacteria is the selection of extraction and purification protocol. An extraction
and purification procedure that works well for a phycobiliprotein from one organism
may not be the method of choice for the corresponding phycobiliprotein from another
organism. In this study, this reasons the phycobiliprotein extraction and
optimization and preservation methods were compared in Phormidium tenue
NTDM05. Hemlata et al., (2011) also described these extraction and purification
methods in Anabaena.
Uv – Vis Spectrum of phycobiliprotein showed peaks at 540 nm in higher
percentage (120mg/g), which indicates that the phycoerythrin is a major constituents
among phycobiliproteins that produced by Phormidium tenue NTDM05 (Fig 2).
Whereas other phycobiliproteins like phycocyanin and allophycocyanin present in the
meager amount of 70mg/g and 20mg/g respectively. Hence, the works have focused on
the extraction, production and optimization of C-Phycoerythrin.
The data of quantitative evaluation of phycobiliproteins in the selected
cyanobacterial species revealed C-PE as the major component with respect to other
phycobiliproteins, i.e., PC and APC, which makes the purification process easy. Since
APC is present in very low amount, this molecule does not interfere in the purification
process. This may be due to both the strain specificity in the phycobiliprotein content
and the culture conditions of each cyanobacterial strains.
In the first set of experiments for the optimization of nutrients, culture was
grown under increasing concentration of carbonate. The results showed an increase in
the biomass coupled with an increase in photosynthetic activity, indicated by increased
photosynthetic pigments like chlorophyll-a, carotenoids, and phycobilins.
A culture was grown under increasing concentration of Magnesium sulphate for
the optimization of production (Fig 3). The highest biomass was observed in the culture
a)
b)
Figure 2 Small–scale culturing of Phormidium tenue NTDM05 for pigment production (a) and microphotograph of Phormidium tenue NTDM05 with red color pigmentation (b)
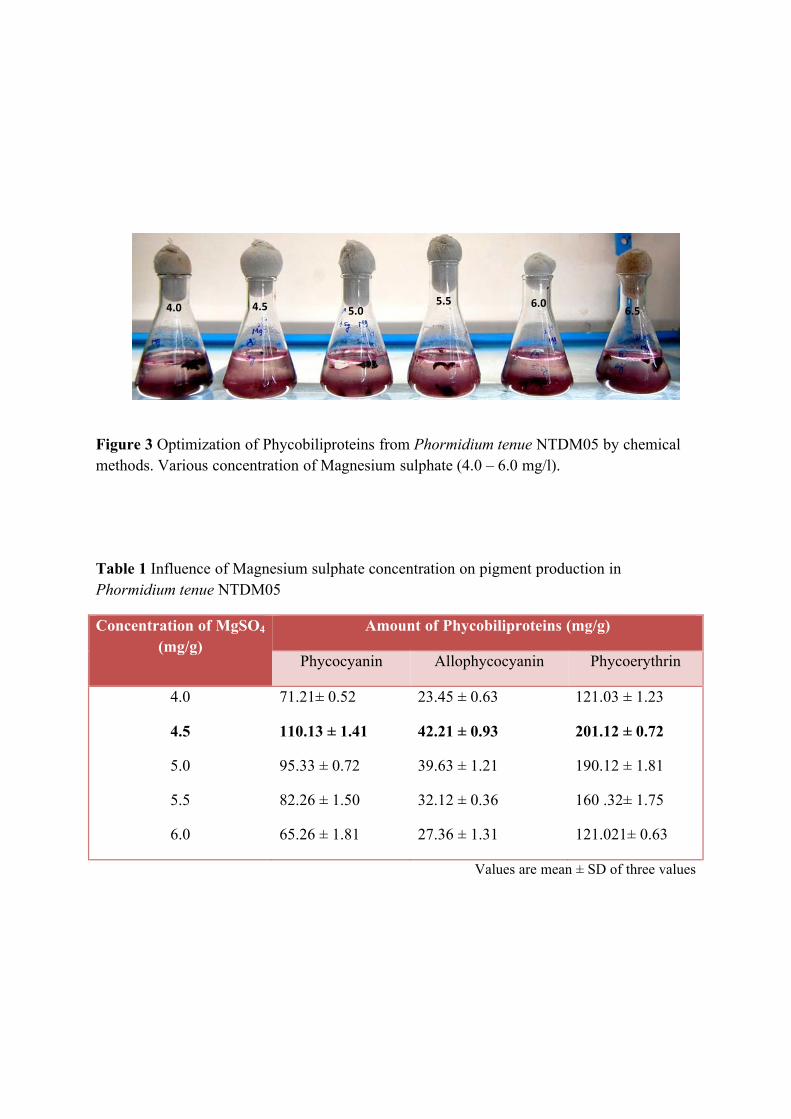
Figure 3 Optimization of Phycobiliproteins from Phormidium tenue NTDM05 by chemical methods. Various concentration of Magnesium sulphate (4.0 – 6.0 mg/l).
Table 1 Influence of Magnesium sulphate concentration on pigment production in Phormidium tenue NTDM05
Concentration of MgSO4 (mg/g)
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
4.0
4.5
5.0
5.5
6.0
71.21± 0.52
110.13 ± 1.41
95.33 ± 0.72
82.26 ± 1.50
65.26 ± 1.81
23.45 ± 0.63
42.21 ± 0.93
39.63 ± 1.21
32.12 ± 0.36
27.36 ± 1.31
121.03 ± 1.23
201.12 ± 0.72
190.12 ± 1.81
160 .32± 1.75
121.021± 0.63
Values are mean ± SD of three values
4.0 4.5 5.0 5.5 6.0
6.5
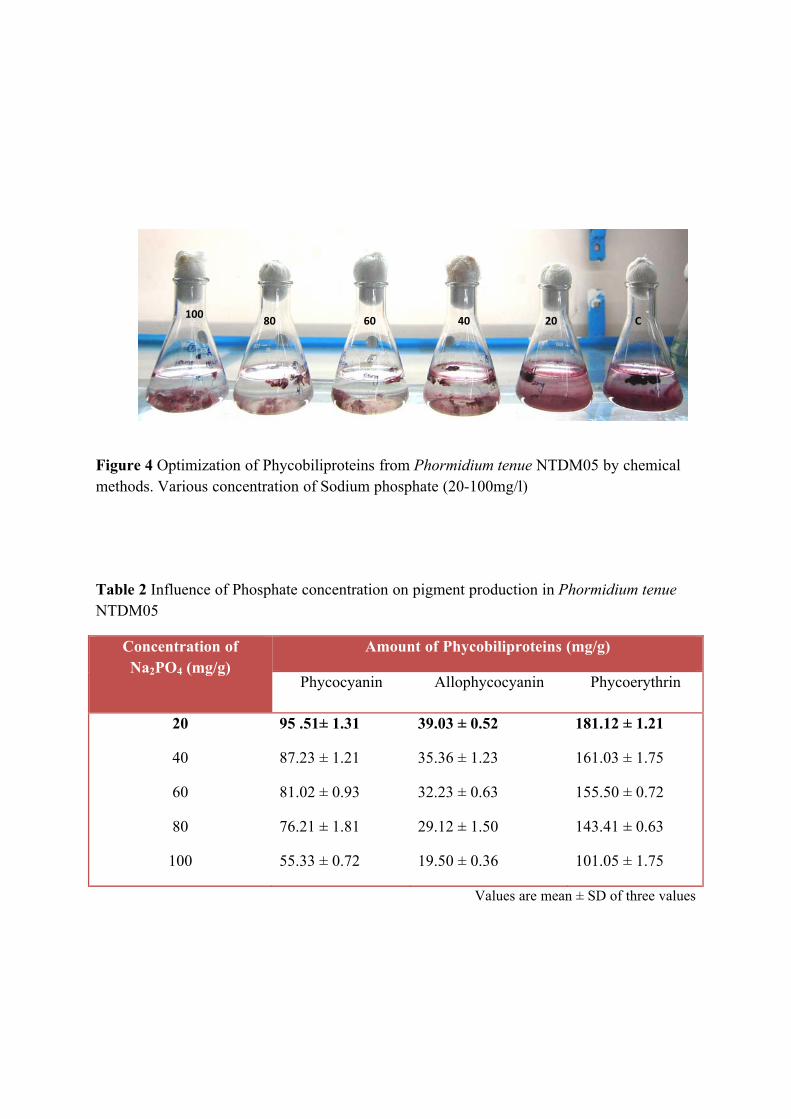
Figure 4 Optimization of Phycobiliproteins from Phormidium tenue NTDM05 by chemical methods. Various concentration of Sodium phosphate (20-100mg/l)
Table 2 Influence of Phosphate concentration on pigment production in Phormidium tenue NTDM05
Concentration of Na2PO4 (mg/g)
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
20
40
60
80
100
95 .51± 1.31
87.23 ± 1.21
81.02 ± 0.93
76.21 ± 1.81
55.33 ± 0.72
39.03 ± 0.52
35.36 ± 1.23
32.23 ± 0.63
29.12 ± 1.50
19.50 ± 0.36
181.12 ± 1.21
161.03 ± 1.75
155.50 ± 0.72
143.41 ± 0.63
101.05 ± 1.75
Values are mean ± SD of three values
100 80 60 40 20 C

65
containing 4.5mg/l of Magnesium sulphate, which was increased concentration than the
control (4mg/l) and the concentration of C-phycoerythrin was found to be 201.12mg/g
(Table 1). The same type of observation was also found in Sodium phosphate, in which
various concentration of Sodium phosphate (20-100 mg/l) (Fig 4) amended in the
culture medium, where 20mg/l of sodium phosphate showed significantly increased the
production of C-phycoerythrin maximum of 181.12mg/g of biomass. In other hands,
when increase the concentration of sodium carbonate and potassium phosphate from
the control concentration, there was slow growth compared with control culture
(Table 2).
The highest biomass was observed in the culture containing 20mg/l of Sodium
carbonate, which was normal concentration as control (20mg/l) and the concentration
of C-phycoerythrin was found to be 303.26mg/g (Table 3). Eiler et al., (2003) showed
that bacterial biomass production was limited by supply of carbon. These results
showed that there was a correlation between substrate carbon and growth. However,
there was a stagnation of biomass production at a carbon concentration that range from
20-100 mg/l.
On optimization of one of the nutrients was Sodium nitrate on the production of
phycoerythrin also been investigated in which 3.75mg/l of NaNO3 showed significantly
increased production of C-Phycoerythrin. It was increased concentration than the
control (3mg/l) and the concentration of C-Phycoerythrin was found to be 261.61mg/g
(Table 4)
There are several reports on the growth limitations of microbial communities.
Bacterial biomass, production, and rate of growth of all responded to organic
enrichments, indicating that bacterial growth was constrained primarily by the
availability of dissolved organic matter (Church et al., 2000) Carbonate and
bicarbonate ions are the most easily available forms of inorganic carbon to the marine
Cyanobacteria. In the present study, sodium carbonate (Na2CO3) was used as the source
of carbon in the medium. Farjala et al., (2006) have suggested that the low availability
of phosphate and nitrogen coupled with excess organic carbon was the main factor
Table 3 Influence of carbonate concentration on pigment production in Phormidium tenue NTDM05
Concentration of Na2CO3 (mg/g)
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
20
40
60
80
100
164.01 ± 1.50
155.25 ± 0.93
131.03 ± 0.72
111.37 ± 1.81
103.04 ± 0.52
71.32 ± 1.21
63.41 ± 0.52
59.04 ± 1.41
51.62 ± 1.75
45.81 ± 0.63
303.26 ± 1.31
291.67 ± 0.36
249.24± 1.75
201.13 ± 1.21
195.35 ± 1.23
Values are mean ± SD of three values
Table 4 Influence of nitrate concentration on pigment production in Phormidium tenue NTDM05
Concentration of NaNO3 (mg/g)
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
1.5
2.25
3.0
3.75
4.5
5.25
6.0
71.21 ± 1.75
73.35 ± 0.63
85.19 ± 0.52
138.43 ± 0.72
120.18 ± 0.36
115.21 ± 1.50
95.36 ± 1.81
29.16± 1.21
31.27 ± 0.93
35.36 ± 1.81
55.15 ± 1.23
53.67 ± 1.12
49.23 ± 1.31
45.21 ± 0.63
121.36 ± 0.52
132.21± 1.23
163.15 ± 0.36
261.61 ± 1.41
221.57 ± 1.75
211.12± 1.21
191.02 ± 0.93
Values are mean ± SD of three values

66
responsible for the relatively low bacterial utilization of DOC. Badger and Andrews
(1982) stated that during steady-state photosynthesis in seawater equilibrated with air,
HCO- uptake into cell is the primary source of internal inorganic carbon for
cyanobacteria. Sultemeyer et al., (1998) have stated the presence of inducible high
affinity HCO- and CO2 transporter at the chloroplast envelope membrane of
microalgae. Use of Chlorophyll-a fluorescence measurements for ascertaining the
photosynthetic condition of entire phytoplankton communities across large temporal
(Suggett et al., 2006a) and spatial (Behrenfeld et al., 1996; Suzuki et al., 2002; Moore
et al., 2005, 2006a,b and Suggett et al., 2006b). Chlorophyll-fluorescence was used to
study the relation between increased carbon substrate and growth of the cyanobacteria.
Fox (1996) has observed the presence of carbon supplement enhancing chlorophyll-a
content ranging from 6.1 to 7.6 mg/l in Spirulina sp.
The present study also analyzed the effect of carbon on other pigments such as
phycocyanin and allophycocyanin. Fox (1996) has observed the influence of carbon
content level of carotenoids in Spirulina and reported that it’s ranged from 2.9 to
4.0mg/g. In the present study when 400mg/l carbonate was provided, the carotenoids
production was 12.7µg/l, which is far higher than the earlier reports. Whereas in
Phycocyanin content was found to be highest (7.2 µg/l) at 450mg/l which lesser than
the reports of Palaniselvam et al., (1998), in which they observed 13ug/l level of
phycocyanin in the Phormidium tenue NTDM05 at carbonate concentration of 20mg/l.
the allophycocyanin and phycoerythrin showed the maximum concentration of 11.2
µg/l and 3.3 µg/l at 400mg/l of carobonate respectively. In earlier studies Fatima et al.,
(1998) have recorded the phycocyanin level of 13mg/g in Phormidium tenue in the
presence of carbonate. Sakthivel (2004) reported that the level of phycocyanin varies
from 2.99 to 14.63 mg/g respectively in Oscillatoria cortiana and Phormidium tenue.
It was observed that there is no significant increased production of
phycobiliprotein was found in pH optimization study (Table 5). At pH 8 showed
comparatively higher amount of phycoerythrin (110.22mg/g). When pH increase
production of pigments also decreased and after the incubation period, pH of the
medium has been changed into neutral in all the experiment. The cellular mechanism of
Table 5 Influence of pH on pigment production in Phormidium tenue NTDM05
pH
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
4.0 ± 0.5
6.0 ± 0.5
8.0 ± 0.5
10.0 ± 0.5
12.0 ± 0.5
22.03 ± 1.75
37.63 ± 1.21
65.33 ± 0.63
61.22 ± 0.93
45.36 ± 1.81
9.0.12 ± 0.72
13.03 ± 0.52
29.23 ± 1.41
24.11 ± 1.23
19.01± 0.36
39.33 ± 1.31
71.45 ± 1.21
110.22 ± 1.50
103.63 ± 1.81
81.03 ± 0.63
Values are mean ± SD of three values
Table 6 Influence of temperature on the production of pigment from Phormidium tenue NTDM05
Temperature (oC)
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
15
25
35
45
55
41.11 ± 1.50
55.36 ± 1.31
73.33 ± 1.23
71.22 ± 1.75
69.12 ± 0.72
17.01 ± 1.23
21.03 ± 1.81
29.36 ± 0.93
27.33 ± 0.63
28.30 ± 1.21
71.36 ± 0.52
90.33 ± 0.72
125.02 ± 1.23
123.45 ± 1.21
118.61 ± 0.52
Values are mean ± SD of three values

67
the cyanobacteria have played significant role in the alteration of pH in the medium. In
temperature study 35oC showed significant results in the production of pigments
(125.02mg/g) of phycoerythrin. Both pH and temperature of the medium not showed
tremendous changes in the production. Concentration of the pigments was same that of
pigments produced in normal condition and no significant on the production of the
biomass (Table 6).
During the process of extraction, cyanobacterial cells were lysed by freezing at
20oC and thawing at room temperature, which also allows APC and PE with other
proteins and nucleic acids to be extracted along with CPC. The absorption spectra of
crude extract of Phormidium tenue NTDM05, showed the presence of C-PC, APC,
other proteins, and nucleic acids corresponding to their maximum absorption at 620,
652, 280, and 260 nm wavelengths, respectively (Fig 5).
Extraction was considered to be important method to extract C-Phycoerythrin
very effectively. Among tested methods, freeze thawing process was considered to be
the best extraction method (Table 7). It extracts nearly 120.56mg/g of C-Phycoerythrin
pigment. Whereas in other methods, there was considerable amount of pigment loss and
needs skilled personnel to extract them. During sonication process for extraction, the
temperature was raised about 50oC; in this temperature all pigment protein gets
precipitated and denatured. It extracts 51.22mg/g of c-phycoerythrin. In grinding
method, phycobiliprotein losses its color and it was very difficult to purify the C-
Phycoerythrin and other cellular protein. It gives 49.32 mg/g of C-Phycoerythrin (Fig
6).
Selection of suitable buffer for maximum phycobiliproteins extraction is very
crucial (Viskari and Colyer, 2003) as these ensure that proteins are not denatured due
to shift in pH (Roe, 2000). During comparison of various buffers, sodium
phosphate buffer was found as best buffer with 130.68mg/g phycobiliprotein. Whereas,
extraction in double distilled water was found to be 115 .02mg/g. The lowest
yield in distilled water may be due to its incapability to lyse the cell wall. Hemlata
et al., (2011) reported that sodium phosphate buffer was best for extraction of 100 mg/g
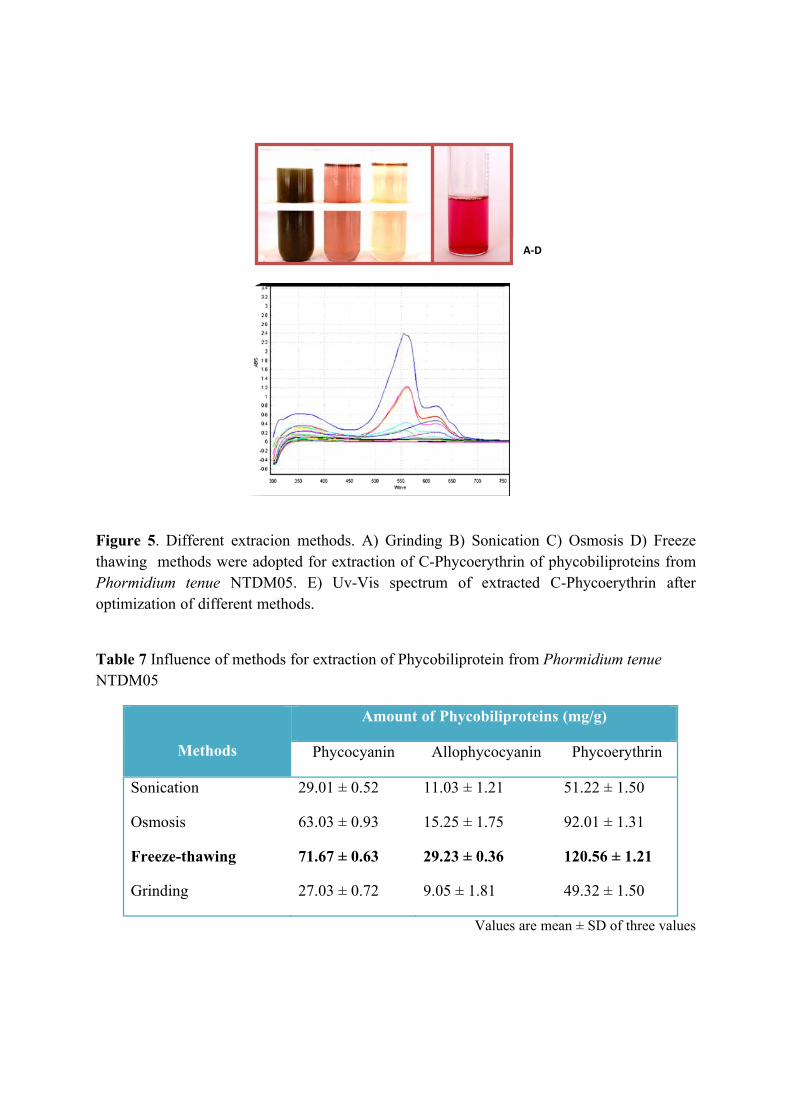
Figure 5. Different extracion methods. A) Grinding B) Sonication C) Osmosis D) Freeze thawing methods were adopted for extraction of C-Phycoerythrin of phycobiliproteins from Phormidium tenue NTDM05. E) Uv-Vis spectrum of extracted C-Phycoerythrin after optimization of different methods.
Table 7 Influence of methods for extraction of Phycobiliprotein from Phormidium tenue NTDM05
Methods
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
Sonication
Osmosis
Freeze-thawing
Grinding
29.01 ± 0.52
63.03 ± 0.93
71.67 ± 0.63
27.03 ± 0.72
11.03 ± 1.21
15.25 ± 1.75
29.23 ± 0.36
9.05 ± 1.81
51.22 ± 1.50
92.01 ± 1.31
120.56 ± 1.21
49.32 ± 1.50
Values are mean ± SD of three values
A-D

68
of phycobiliprotein. Soni et al., (2006) a l so found ou t the p resence o f
Sod ium phospha te showed h igher phycocyan in in phycocyanin in
Oscillatoria and Spirulina Sodium phosphate buffer is also being reported to have
inhibition property to metabolic enzymes. Cyanobacterial cell walls are thick and
extremely resistant t o e x t r e me c o n d i t i o ns (Stewart & Farmer, 1984). It acts as
physical barrier fo r the release of phycobiliprotein into the extracting
medium. So, optimization of cell disruption method is also very important for the
maximum extraction (Table 8).
Fourier transform infrared spectra (FT-IR spectra) IR spectra of freeze-dried C-
PE (Fig. 7) of Phormidium tenue NTDM05 showed the protein specific amide I band at
1637 cm-1 (C=O stretching) and amide II at 1365 cm-1. Position and shape of amide I
band are employed for the analysis of protein secondary structure. The sharp amide I
band at 1637 cm-1 for C-PC indicates the a-helix as the major element of its secondary
structure. It was found that IR spectrum of C-PC of Phormidium, Spirulina and
Lyngbya sp showed same type peak pattern. Besides their experiment, IR spectra of
freeze-dried C-PC further ensure its purity by the absence of inorganic sulfates and
phosphates (represent intense band at 1043 and 1015 cm-1) (Patel et al., 2005).
The absorption spectrum was recorded with preservative and without
preservative at 4oC and 35oC. It was found that the loss of C-PE content in aqueous
solution (i.e. buffer of pH 7.0) at 0±5oC is less than 35±5oC. Loss of C-PE content
observed after addition of different preservatives at 0±5oC is very less, ranging from
2.5 to 40 mg/g while at 35±5oC. C-PE content loss is 5-10mg/g after 45 days as shown
in Table 9. C-Phycoerythrin in aqueous phase was found to be more stable with citric
acid (1 mg/ml) as preservative at 35±5oC, as observed through the spectral analysis, no
discoloration of the C-Phycoerythrin was observed after 45 days both at 0±5 ◦C and
35±5oC with citric acid as preservative and showing only 9.55mg/g loss of C-PE
content at 35±5oC after 45 days, while only 5mg/g C-PE loss was observed at 0±5oC
after 45 days. Even after 45 days C-PE in aqueous solution with citric acid at 35±5oC
retained its color as shown in Fig. 8 C-PE solution without any preservative (control)
showed discoloration after 5 days with 25mg/g loss of C-PE content and completely

Figure 6 Display of phycobiliprotein extracted from Phormidium tenue NTDM05 after optimized production: phycoerythrin (red) and Phycocyanin (blue) extracted from Spirulina subsalsa NTDM21
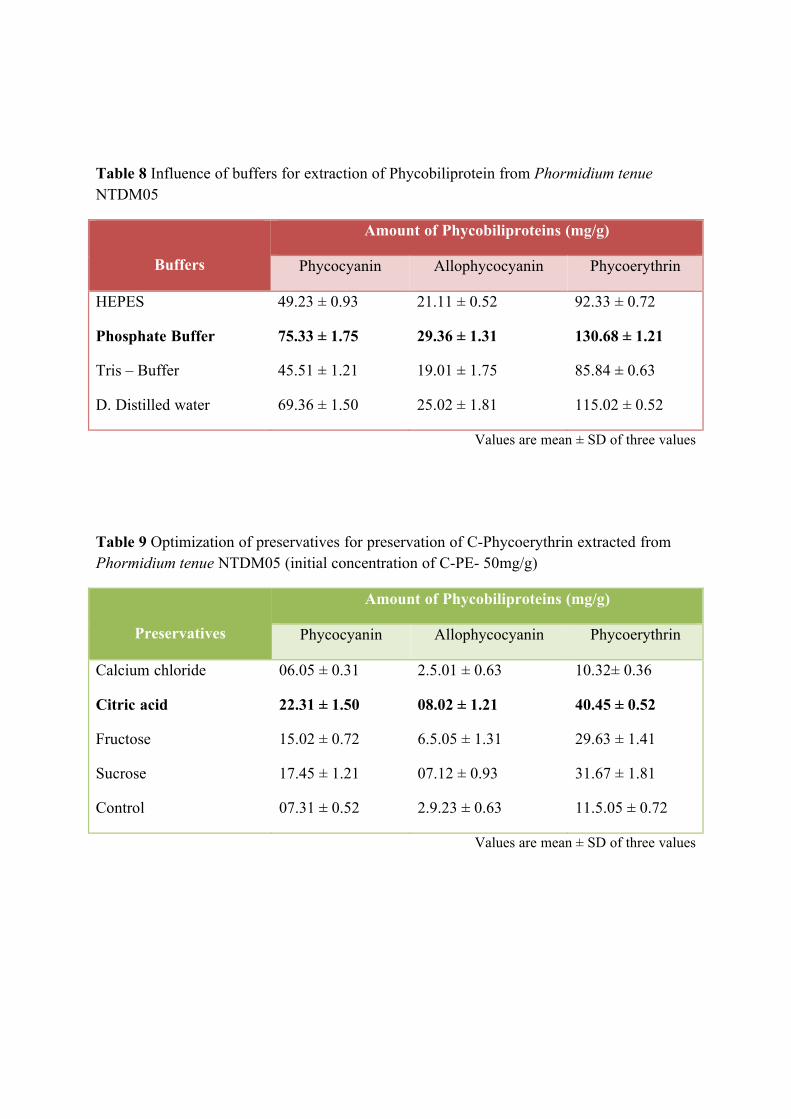
Table 8 Influence of buffers for extraction of Phycobiliprotein from Phormidium tenue NTDM05
Buffers
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
HEPES
Phosphate Buffer
Tris – Buffer
D. Distilled water
49.23 ± 0.93
75.33 ± 1.75
45.51 ± 1.21
69.36 ± 1.50
21.11 ± 0.52
29.36 ± 1.31
19.01 ± 1.75
25.02 ± 1.81
92.33 ± 0.72
130.68 ± 1.21
85.84 ± 0.63
115.02 ± 0.52
Values are mean ± SD of three values
Table 9 Optimization of preservatives for preservation of C-Phycoerythrin extracted from Phormidium tenue NTDM05 (initial concentration of C-PE- 50mg/g)
Preservatives
Amount of Phycobiliproteins (mg/g)
Phycocyanin Allophycocyanin Phycoerythrin
Calcium chloride
Citric acid
Fructose
Sucrose
Control
06.05 ± 0.31
22.31 ± 1.50
15.02 ± 0.72
17.45 ± 1.21
07.31 ± 0.52
2.5.01 ± 0.63
08.02 ± 1.21
6.5.05 ± 1.31
07.12 ± 0.93
2.9.23 ± 0.63
10.32± 0.36
40.45 ± 0.52
29.63 ± 1.41
31.67 ± 1.81
11.5.05 ± 0.72
Values are mean ± SD of three values
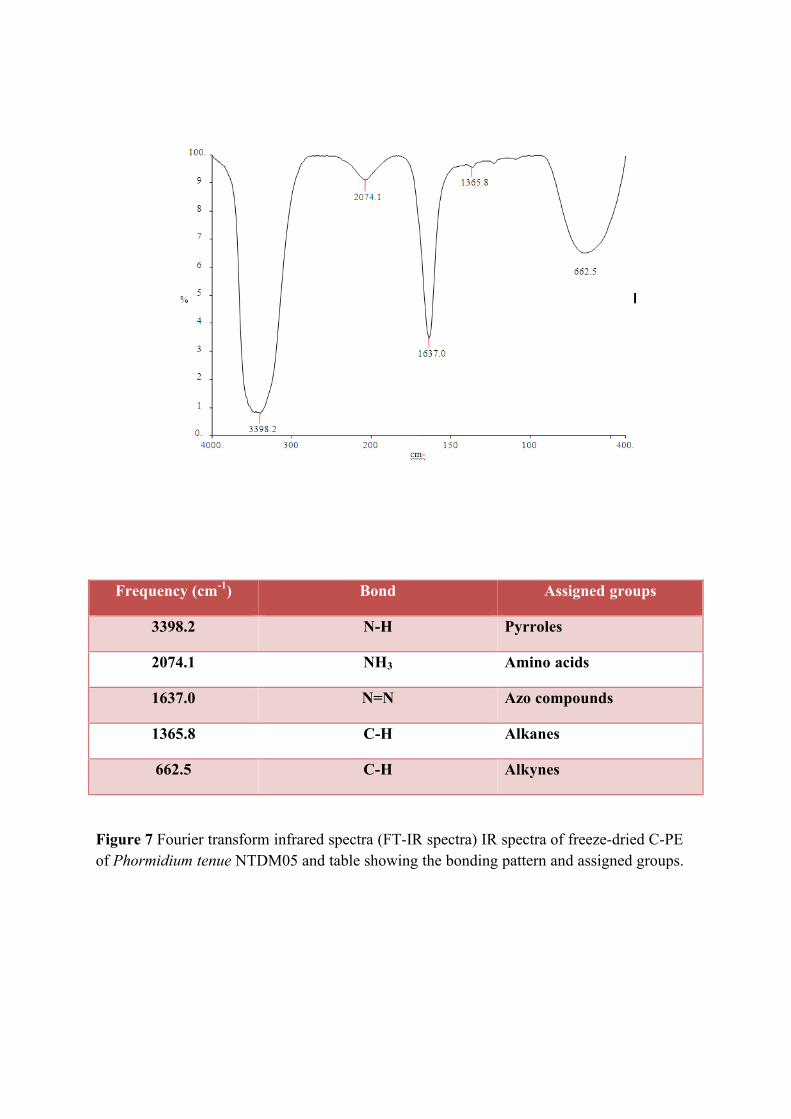
Frequency (cm-1) Bond Assigned groups
3398.2 N-H Pyrroles
2074.1 NH3 Amino acids
1637.0 N=N Azo compounds
1365.8 C-H Alkanes
662.5 C-H Alkynes
Figure 7 Fourier transform infrared spectra (FT-IR spectra) IR spectra of freeze-dried C-PE of Phormidium tenue NTDM05 and table showing the bonding pattern and assigned groups.

69
discolored after 45 days showing 47.5mg/g loss of C-PE content at 35±5 ◦C, while no
discoloration was observed in C-PE sample even after 45 days at 0±5 oC and C-PE
content loss observed was only 39mg/g, because C-PE is heat sensitive at pH 7.2.
C- Phycoerythrin was found to be sensitive to heat and light in aqueous
solution; C-PE is insoluble in acidic solution (pH 3.0) and is highly soluble and stable
at pH 7.0 which is the reason for its efficient extraction in buffer of pH 7.0. Hence,
after extraction and purification of the product, immediate addition of preservative is
crucial prior to lyophilization and further packing in the amber/dark colored bottles to
protect from light. It denatures at temperature above 45 ◦C at pH 7.0, leading to
discoloring.
The stability of food grade C-PE with citric acid as preservative was found to be
more than sodium chloride, calcium chloride and sucrose at 35±5 ◦C which is
comparable to its stability at 0±5 ◦C; the pH 7.0 is found to be most suitable for
maintaining its unique properties; where it is in hexameric form citric acid acts as a
chelator and lowers the pH preventing the degradation of protein (Jespersen et al., 2005
and Edwards et al., 1997). The present work leads to the conclusion that out of the
three tested edible preservatives-(citric acid, sucrose, sodium chloride and calcium
chloride). Citric acid (conc. 1mg/ml) is one of the most effective preservative for
maintaining the stability of C-Phycoerythrin in aqueous phase at 35±5 ◦C even for 45
days with minimum loss at room temperature 35±5 ◦C.
There was no antibacterial property was observed in extracted phycobiliprotein
against tested organisms. This gives another way to exploit the protein for food industry
for colorant. It has been reported that the effect of various concentrations of C-PE on
three human cell lines using MTS assay which measures the cell viability on the basis
of mitochondrial dehydrogenase activity. In this assay, the C-PE concentrations used
did not adversely affect formazan formation.
Based on the previous hypothesis, phycoerythrin has an impact on the defense
against oxidative stress and to examine the relationship between intracellular ROS-
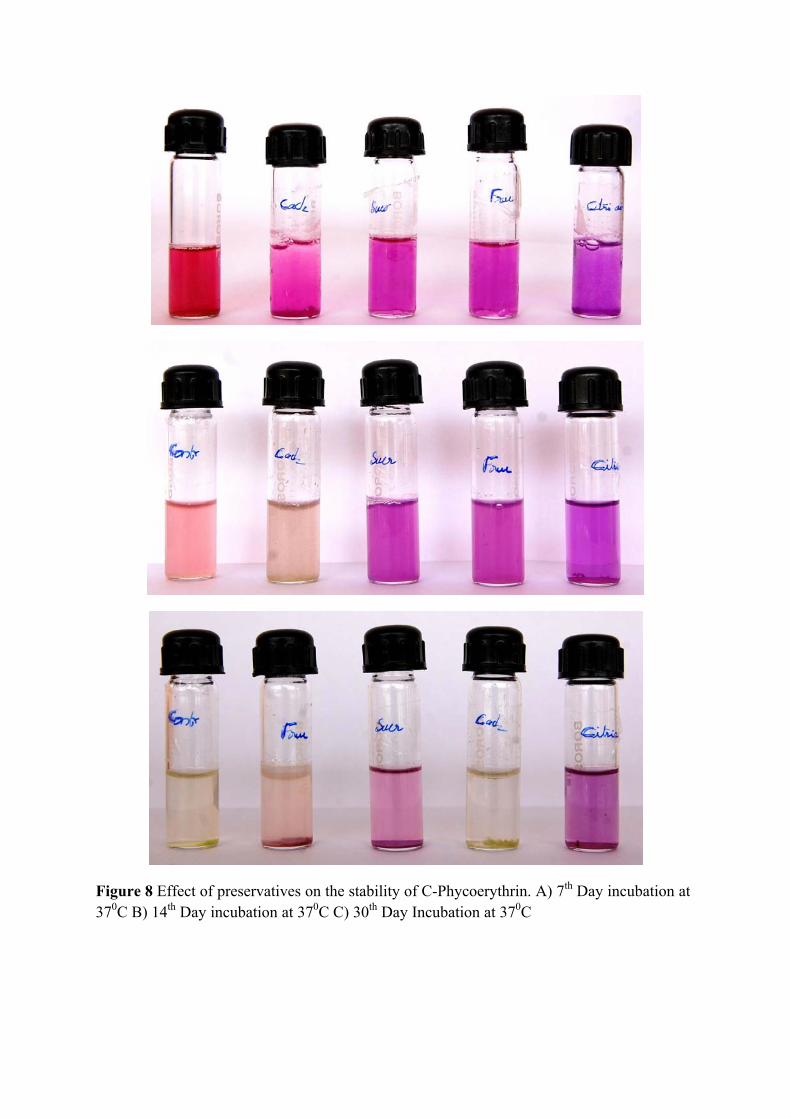
Figure 8 Effect of preservatives on the stability of C-Phycoerythrin. A) 7th Day incubation at 370C B) 14th Day incubation at 370C C) 30th Day Incubation at 370C

70
inducing action of permanganate and oxidative DNA damage, and further assessed the
protective actions of C-PE. The oxidative DNA damage generated by KMnO4 was
measured by comet assay in fibroblast cells. Various concentrations of KmnO4 were
checked for generation of fibroblast DNA damage (data not shown). The 25 mM
KMnO4 treatment at 4 °C was found to generate consistent comets and was kept
constant throughout the experiment. As compared with the positive control, the
increased tail movements caused by permanganate-induced oxidative DNA damage
were not observed in the cells treated with C-PE.
The fluorescence property of C-phycoerythrin was observed and the absorption
and emission spectra of the purified PE from growth phases of a culture was recorded.
The absorption spectrum of the purified PE from a young culture was same with a
sharp peak at 562 nm but it varied for PE from old culture, whereas the emission
wavelength for purified PE was found to be 580 nm (Fig. 9).
In PE purified from young culture of 25 days of Phormidium tenue NTDM05,
a sharp absorption peak was observed at 562 nm whereas for PE purified from 45 days
old culture a shoulder peak at 542 nm along with an absorption peak at 562 nm was
observed. However, when the same proteins were compared for their emission
wavelength, no difference was found as both the proteins emitted light at 580 nm. This
proved that PE with single subunit was equally functional as PE with two subunits.
It has been reported that the purified PE from young culture of Halomicronema
sp. A32DM showed an absorption peak at 562 nm whereas for PE from old culture, the
absorption peak shifted to 542 nm along with a shoulder peak at 562 nm. On comparing
for their emission wavelength, a similar result to that of Phormidium sp. A27DM was
observed thus proving PE with single subunit to be functional in this culture also (Rigbi
et al., 1980). On the other hand, PE purified, from both young and old cultures
of Lyngbya sp. A09DM showed an absorption peak at 562 nm. However, there was an
increase in the shoulder peak at 620 nm for the PE from old culture. It was suggested
that the culture might have started producing phycoerythrocyanin (PEC) for its survival
in nutrient depletion conditions rather than cleavage of PE observed in the other two

(A)
(B)
Figure 9 Fluorescence property of C-Phycoerythrin extracted from Phormidium tenue NTDM05. A) C-Phycocyanin and C-Phycoerythrin illuminated with Uv-Transilluminator showing bright orange color fluorescence B) fluorescence emission overlay spectra of purified C-Phycoerythrin showing fluorescence property.
Intensity
Wavelength

71
cultures. When compared for emission wavelength, both the proteins emitted light at
580 nm proving both of them to be functional. Phycoerythrin generally emits light at
580 nm (Rigbi et al., 1980). PE purified from all the cultures at all growth phases
emitted light at around 580 nm, thus proving them to be functional.
The results indicated the successful development of an efficient method for the
extraction of C-phycoerythrin from an isolate Phormidium tenue NTDM05. The culture
has the ability to produce high amount of C-PE when grown under normal white light,
suggesting it to be a good candidate for the production and extraction of phycoerythrin.
Freezing–thawing is a mild, nondenaturing condition for the extraction of
phycobiliproteins as compared to other methods we studied. This extraction process
may need to be studied for each strain of cyanobacteria depending upon the cell wall
rigidity, but once the required freezing and thawing temperature are standardized, it
could give very good reproducibility in each batch of extraction.
On the whole, in this chapter a cyanobacterial species was selected among
isolated species such as Phormidium tenue NTDM05 based on the pigment production.
It was found that micronutrients are played an important role in the production of
phycobiliprotein in general and phycoerythrin in particular. The extraction methods are
crucial step to extract the pigmented protein from the cell, in which freeze-thawing was
the best method of extraction. Regarding the preservation of the pigments, citric acid
was found to be best preservatives, which preserves upto one month without losing its
color at 4oC and minimum loss of colors at 37oC. It was found that there was no
antibacterial activity observed in the protein; it can be used for the food additives as
colorant and protein supplements.

Chapter III
Nanobiotechnological Potentials of Marine Cyanobacteria and Diatoms

72
INTRODUCTION
Study of Nanobiotechnology:
Nanotechnology is enabling technology that deals with nano-meter sized
objects. It is expected that nanotechnology will be developed at several levels such as
materials, devices and systems. Synthesis of nanoparticles using biological entities has
great interest due to their unique shape dependent optical, electrical and chemical
properties have potential application in nanobiotechnology. In addition,
nanobiotechnology allows fabrication, synthesis and polymerization of nano and micro
scale structure and architecture of various biomolecules including protein and nucleic
acids with and without organic and inorganic materials.
One key aspects of nanotechnology concerns the development of reliable
experimental protocols for the synthesis of nanomaterials over a range of chemical
composition size and high monodispersity. In the context of the current drive to
develop green technologies in materials synthesis, this aspect of nanotechnology
assumes considerable importance. In utilizing is microorganism for the synthesis of
nanoparticles.
Nanoparticles are now considered much more important due to physical,
chemical and biological properties than that is exhibit in the bulk material. Synthesis of
nanomaterials by physical and chemical ways were studied past decade with impact of
environmental pollution, later researchers developed their synthesis by biological
entities is much simpler and eco-friendly. Most of the prokaryotic and eukaryotic
microorganisms were studied in this direction. Microbial cells are highly organized
units, regarding morphology and metabolic pathways, capable of synthesizing
reproducible particles with well defined size and structure, more importantly; biogenic
nanoparticles often exhibit water-soluble and biocompatible properties, which are
essential for many applications.
Nature has devised various processes for the synthesis of nano and micro scaled
inorganic materials which have contributed to the development of relatively new and

73
largely unexplored area of research based on the biosynthesis of nanomaterials (Sastry
et al., 2004). The synthesis and assembly of nanoparticle would benefit from the
development of clean, non-toxic and environmentally acceptable “green chemistry”
procedures, probably involving organism ranging from bacteria to fungi and even
plants (Bhattacharya and Rajinder 2005 and Sastry et al., 2004). Hence both unicellular
and multicellular organisms are known to produce inorganic material either intra of
extracellularly (Mann, 1996).
Biosynthesis of Nanoparticles:
The development of simple and versatile methods for the preparation of
nanoparticles in a anisotropic size or shape selected and controlled manner has been a
challenging but intellectually rewarding task (Xia et al., 2009 and Sau et al., 2010). The
nanoparticles morphology often emerges as a result of a competitive growth of different
crystallographic surfaces. This is typically achieved by altering the relative growth rates
of different facets by the selective localization of surface-modifying or capping agents,
but also by the modulation of nucleation and reaction parameters such as time,
temperature, reagent concentration, and pH (Sohn et al., 2009).
There are several reports in the literature on the cell-associated biosynthesis of
metallic nanoparticles using several microorganisms. Also, for the past few years,
various rapid chemical methods have been developed for the synthesis of metallic
nanoparticles in general and silver nanoparticles in particular. The synthesis of silver
nanoparticles has attracted much attention because their unique shape-dependent
optical, electrical and chemical properties have potential applications in
Nanotechnology. Silver nanoparticles are used in photographic reactions, catalysis,
chemical analysis, exhibit strong toxicity to a wide range of micro organism and
antibacterial applications, therefore development of silver nanoparticles are expected to
open avenues to fight and prevent diseases (MubarakAli et al., 2011)
Silver nanoparticles production as the nanoparticles possess more surface atom
than a micro particle which greatly improve its characteristics. The biosynthesis of
silver nanoparticles of different sizes ranging from 1- 70nm and shapes, including

74
spherical, triangular and hexagonal has been conducted using bacteria, fungi, plants and
plant extracts.
Metal nanoparticles, particularly gold, are being considered in wide ranging
applications, such as photonics, information storage, electronic and optical detection
systems, therapeutics, diagnostics, photovoltaics, and catalysis (Murphy et al., 2005;
Burda et al., 2005; Sau et al., 2010; Talapin et al., 2010; Cuenya, 2010; Daniel and
Astuc, 2004; Eustis and El-Sayed, 2006; Sardar et al., 2009; Zhou et al., 2009; Rosi
and Mirkin, 2005 and Giljohama et al., 2010). While the chemical composition of a
nanoparticle is important, even more important is the morphology (size and shape) of
the nanoparticles and its surface / colloidal properties.
Biosynthesis of Nanocomposites:
The unicellular eukaryotic algae known as diatoms make cell walls out of silica
with an amazing diversity of structures on the nanoscale (Round et al., 1990). Because
the complexity of diatom silica structures can exceed that possible using synthetic
material production approaches, and diatoms reproduce these structures faithfully, in
enormous numbers, and inexpensively, diatoms are being developed as a source of
nanostructured materials (Sandhage et al., 2005). Facilitating this techniques diatoms
can be utilizing as templates for functionalized coatings (Payne et al., 2005; Rosi et
al., 2004; Weatherspoon et al., 2007), or convert their silica entirely to other molecules
while maintaining nanoscale features (Bao et al., 2007; Kusari et al., 2007; Sandhage
K.H. et al., 2002; Sandhage et al., 2005; Unocic R.R. et al., 2004; Weatherspoon et al.,
2006; Zhao J.P. et al., 2005). In addition, the development of diatoms as a source of
nanostructured materials, it would be beneficial to develop approaches whereby
structure could be genetically tailored to suit particular applications (Hildebrand, 2005;
Sandhage et al., 2005). This requires better understanding of the silicification process.
Understanding the process of diatom silica cell wall synthesis requires elucidation of
the underlying cellular and molecular biology, especially, important is to know how
genes and organic molecules (proteins, polyamines, carbohydrates, and lipids) are
involved in formation of specific mineral substructures.

75
The cell wall structure is a species-specific characteristics demonstrating that
diatom silica morphogenesis is genetically encoded. It is expected that these structures
exhibit superior properties in a wide range of applications. The fabricated and self
assembled of semiconductor nanostructures that possess unique optical and electronic
properties (Rorrer, 2005). The supramolecular chemistry and catalysis have led to novel
surface and size dependent chemistry such as enation selective catalysis at the surface.
The ability of diatoms to make complex, nanoscaled, three dimensional silica shells
called “frustules” offers attractive possibilities for their application to
nanobiotechnology (Hildebrand. 2005).
Bioengineered nanosystem is extremely difficult to realize on the basis of
current scientific knowledge and bottom up assembly is very powerful in creating of
identical structures with atomic precision, such as supramolecular functional entities in
living organisms. In this regards diatoms, a prolific class of single celled algae that
make micro scale biosilica or frustules with intricate submicron features dominated by
two dimensional pore arrays, have been advertized as a paradigm for the controlled
production of nanostructured silica with interesting properties (Sumper, 2006). The
fabrication of novel biomaterials through molecular self assembly is studied currently
(Zhang, 2003).
Biosynthesis of Semiconductor:
CdS is an important semiconductor owing to its unique electronic and optical
properties, and its potential applications in solar energy conversion, nonlinear optical,
photo electrochemical cells and heterogeneous photocatalysis (Hu et al., 1998 and
Weller et al., 1993). The synthesis of CdS nanoparticles has been tried by various
methods such as the direct reaction of metals with sulfur powders under high
temperature (Kaito et al., 1989), the thermal decomposition of molecular precursors
containing M–S bonds (Milhemy et al., 1984 and Ludoph and Malik, 1998) or the use
of poisonous H2S as the sulphur source at higher temperature (Pickeet, 1996) and
chemical precipitation method involving the precipitation of metal ions with Na2S as
the source of S2- ions (Herron, 1990 and Chen, 1996), in which the homogeneity at an
early stage results in a broadening of the product particle size distributions.

76
Developing reliable protocols for the synthesis of nanometer scale
semiconductor particles is a problem of great importance, (Shipway, 2000 and
Alivisatos, 1996) resulting in researchers seeking inspiration from biological systems
where biominerals are routinely synthesized (Addadi, 1992 and Mann, 1996). Both
prokaryotic (bacteria) (Labrenz, et al., 2000 and Holmes, et al., 1995) and eukaryotic
organisms such as yeast (Dameron, et al., 1989) have been found to produce
semiconductor nanoparticles within the cell wall of the microorganisms.
Cadmium sulfide nanoparticles (CdS NPs) are one of the widely studied
semiconductor nanoparticles that have unique photochemical and photo physical
properties. The semiconductor nanoparticles were often referred as Quantum Dots
(QDs), these particles when embedded within an appropriate matrix act as potential
wells that confine and stabilize electrons in discrete energy levels. The technologically
useful properties of CdS QDs are due in part to the fact that the band-gap is tunable
over a range of 1.5 to 3.5eV (Flenniken, et al., 2004).
Biological synthesis of CdS NPs has been studied effectively by wide range of
organisms for instance algae (Hosea, 1986 and Scarano and Morelli, 2003), fungi
(Ahmed, 2002), yeast (Dameron, et al., 1989 and Prasad and Anal Jha, 2010) and
bacteria (Bai, et al., 2009 and Prasad and Anal Jha. 2010). More lately, apart from the
synthesis of such CdS NPs, the stabilization of synthesized particles were mediated by
extracted and purified active compounds from microbes is efficient method to
understand the stabilizing agent present in the microorganism.
A deserve mention that the cyanobacteria and diatom species selected for this
purpose is easily and abundantly available in eutrophic ponds and marine ecosystem of
India as well as in different parts of the world. Some other characteristics of these
organisms are efficient and rapid biosorption of metals, low production cost, ease in
separation of compounds from the culture media. In order to utilizing the efficiency of
nanomaterials synthesis in cyanobacteria and diatom, the work have framed with the
following objectives:

77
To synthesis silver and gold nanoparticles using selected marine
Cyanobacteria
To synthesis and stabilize CdS nanoparticles by extracted c-phycoerythrin
from Cyanobacteria
To synthesis silicon-germanium oxide (Si-Ge) nanocomposites using
Diatom
To characterize the synthesized nanocomposites and nanoparticles by TEM,
HRSEM, EDAX, EDS, FTIR, FS, Uv-Vis spectroscopic studies
To evaluate the antimicrobial properties towards multi drug resistant (MDR)
clinical human pathogens.
To evaluate the antioxidant property and fluorescence property of
synthesized nanoparticles.

78
REVIEW OF LITERATURE
The study of nanometer size crystallites provides an opportunity to observe the
evolution of material properties with size. The study of intermediate size regime gives
the information about the collective behavior of bulk materials which emerges from the
discrete nature of molecular properties.
Physical and Chemical Properties of Nanoparticles:
The principal parameter of nanoparticles is their shape (including aspect ratios
where appropriate), size and the morphological sub-structure of the substance.
Nanoparticles are presented as an aerosol (mostly solid or liquid phase in air), a
suspension (mostly solids in liquids) or an emulsion (two liquid phases). In the
presence of chemical agents (surfactant), the surface and interfacial properties may be
modified. Indirectly such agents can stabilize against coagulation or aggregation by
conserving particle charge and by modifying the out most layer of the particle.
Depending on the growth history and the lifetime of nanoparticles, very
complex compositions, possibly with complex mixtures of adsorbents, have to be
expected. In the typical history of combustion nanoparticles, for example, many
different agents are prone to condensation on the particle while of processes are to be
expected and have been identified only for a small number of particulate model
systems. At the nanoparticles – liquid interface, polyelectrolyte have been utilized to
modify surface properties and the interaction between particle and their environment.
They have been used in a wide range of technologies, including adhesion, lubrication,
stabilization, and controlled flocculation of colloidal dispersions (Liufu et al., 2004).
At some point between the Angstrom level and the micrometer scale, the simple
picture of nanoparticles as a ball or droplet changes. Both physical and chemical
properties are derived from atomic and molecular origin in a complex way. For
example, the electronic and optical properties and chemical reactivity of small cluster
are completely different from better known property of each component in the bulk or
at extended surface. Complex quantum mechanical models are required to predict the

79
evolution of such properties with particle size, and typically very well defined
conditions are needed to compare experiments and theoretical predictions.
Nanoparticle – Nanoparticle Interaction:
At the nanoscale, particle-particle interaction is either dominated by weak van
der walls forces, stronger polar and electrostatic interaction or covalent interactions.
Depending on the viscosity and polarisability of the fluid, particle aggregation is
determained by the interparticle interaction. By the modification of the surface layer,
the tendency of a colloid to coagulate can be enhanced or hindered. For nanoparticles
suspended in air, charges can be accumulated by physical processes such as glow
discharge or photoemission. In liquids, particle charge can be stabilized by
electrochemical processes at surfaces. Fluid interactions are of key importance to
describe physical and chemical processes, and the temporal evolution of free
nanoparticles. They remain difficult to characterize due to the small amount of
molecules involved in the surface layer. Surface energy, charge and salvation are
relevant parameters to be considered due to the crucial role of the nanoparticles.
Nanoparticles interaction and the nanoparticles are fluid interaction; the term
free nanoparticles can be easily misunderstood. The interaction forces either attractive
or repulsive, crucially determine the fate of individual and collective nanoparticles
resulting in aggregates and or agglomerates may influence on their behavior. In gas
suspensions, aggregation is crucially determined by the size and diffusion, and
coagulation typically occurs in fasters then in the liquid phase as the sticking
coefficient is closer to unity than in liquids 3.5 sources of free nanoparticles.
Nanoparticles are formed through the natural or human mediated disintegration
of larger structure or by controlled assembly processes. The associated processes occur
either in the gas phase, in a plasma, in a vacuum phase or in the liquid phase, eventually
followed by the intentional or unintentional transfer into one or more relevant fluid
media and then to an individual receptor in an exposure setting.

80
Formation of Nanoparticles in Liquid phase:
Defined bottom up production of nanoparticles on the liquid phase with respect
to particle size, chemical composition, surface and change properties occurs mainly
through controlled chemical reactions (Frens, 1973), and self assembly processes have
evolved by controlling growth conditions. In view of the ecological cycling of
nanomaterials, some emphasis has to be given to the corrosion and disintegration of
bulk material, where little knowledge is currently available (Oberdorster, et al., 2005).
Naturally occurring processes generating nanosized structures in the liquid
phase include erosion and chemical disintegration of organic (plant and micro organism
debris) or geological (clays) parent materials. In all these types of disintegration
process, the surface properties and their change through chemical reaction are critical in
determining whether individual nanoparticles will be formed in the respective medium
(Boyle et al., 2005). Schematic representation of formation of silver nanoparticles in
liquid phase by chemical process (Fig. 1).
Figure 1 Schematic representation of formation of silver nanoparticles in liquid phase by chemical process
Formation of Nanoparticles in the Gas phase:
The main route of bottom- up formation of nanoparticle in the gas-phase is by a
chemical reaction leading to a non-volatile product, which undergoes homogeneous
nucleation followed by condensation and growth. Recently, this has become an

81
important pathway for the industrial production of nanoparticle powders, which may be
of metal, oxides, semiconductors, polymers and various forms of carbon, and which
may be in the form of spheres, wires, needles, tubes, platelets, or other shapes. This is
also the unintentional pathway by which nanoparticles are formed following the
oxidation of gas-phase precursors in the atmosphere, in volcanic plumes, in natural and
man-made combustion processes, or in fumes associated with any man-made process
involving volatilizable material at elevated temperature, such as welding or smelting,
polymer fabrication, or even cooking .
As with the liquid phase case, disintegration processes of parent materials
provide a pathway which only leads to nanoparticles suspended in the gas phase under
special conditions. While in the liquid phase the presence of emulsifying agents
accompanying an erosion or chemical disintegration process could support the
suspension process, the dispersion of nanoparticles into a gas from liquid emulsions or
dry powders is severely limited by the strong adhesive forces between individual
nanoparticles. Therefore, any mechanically induced stress on the parent material mostly
leads to particles in the micrometer range and above. Only under accidental conditions,
example in the case of uncontrolled release of a powder or an emulsion from a highly
pressurized vessel could strong shear forces overcome these adhesive forces (Reeks and
Hall, 2001). In contrast, the spraying of liquids containing nanoparticles or soluble
material at very low concentrations, followed by drying of the solvent, can lead to the
resuspension of nanoparticles or to the formation of new nanoparticles from the solutes.
This can lead to redistribution of nanoparticles, biological material or toxic substances
into nanoparticulate airborne form.
Environmental Sources of Airborne Nanoparticles:
The amount of nanoparticles in the air can be surprisingly similar in urban and
rural areas, with as much as 106 to 108 nanoparticles per liter of air depending on
conditions. In rural areas; nanoparticles mostly originate from the oxidation of volatile
compounds of biogenic or anthropogenic origin, including secondary organic aerosols.
In urban areas, the primary sources of these particles are diesel engines (Schneider et al
2005) or cars with defective or cold catalytic converters (Zhiqiang et al., 2000). Photo-

82
oxidation processes also lead to significant numbers of nanoparticles in urban areas.
Real-time measurements show that exhaust aerosol concentrations range between104 to
106 particles cm-3 with the majority of the particles by number being less than 50nm in
diameter. The highest particle number concentrations and smallest particle size are
associated with high-speed road traffic, presumably due to the subtle conditions during
concomitant cooling and dilution of the exhaust gases. Emission factors for gasoline
vehicles ranged from 1.9 to 9.9x1014 particles.km-1 and 2.2x1015 to 1.1x1016
particles km-1 fuel (Kittelson et al., 2003a,b).
The awareness that combustion processes significantly contribute to the
nanoparticles load by number has been rising recently and has provided a new
motivation for airborne particle research (Donaldson et al., 2001a). Wieser and
Gaegauf (2000) have evaluated different wood combustion systems with respect to
emissions. Particle sizes were mainly in the range of 30 to 300 nm. Particles of less
than 300 nm did not add much to the total particle count in flue gas. The particle
distribution of manually operated appliances varied during a burn cycle, while
continuous fed wood combustion systems show a fairly constant particle size
distribution. Total particle numbers for automatically fed burners were smaller than
with manual operation. Around 95% of the particles were smaller than 400 nm. The
most frequent size of the particle number concentrations for batch operated appliances
is approximately 110 nm, whereas the particle distribution changes significantly during
the burn cycle. Silver nanoparticles array on Track Etch membrane support as flow-
through optical sensors for quality control (Julian and et al., 2007).
Biosynthesis of Nanoparticles using Cyanobacteria:
Morphological control over the shape of gold nanoparticles has been achieved
by using Plectonema boryanum UTEX 485, a filamentous cyanobacterium. When it
was reacted with aqueous Au(S2O3)23- and AuCl4
- solutions at 25-100oC for up to 1
month and at 200oC for 1 day resulted in the precipitation of cubic gold nanoparticles
and octahedral gold platelets, respectively (Lengke et al., 2006).
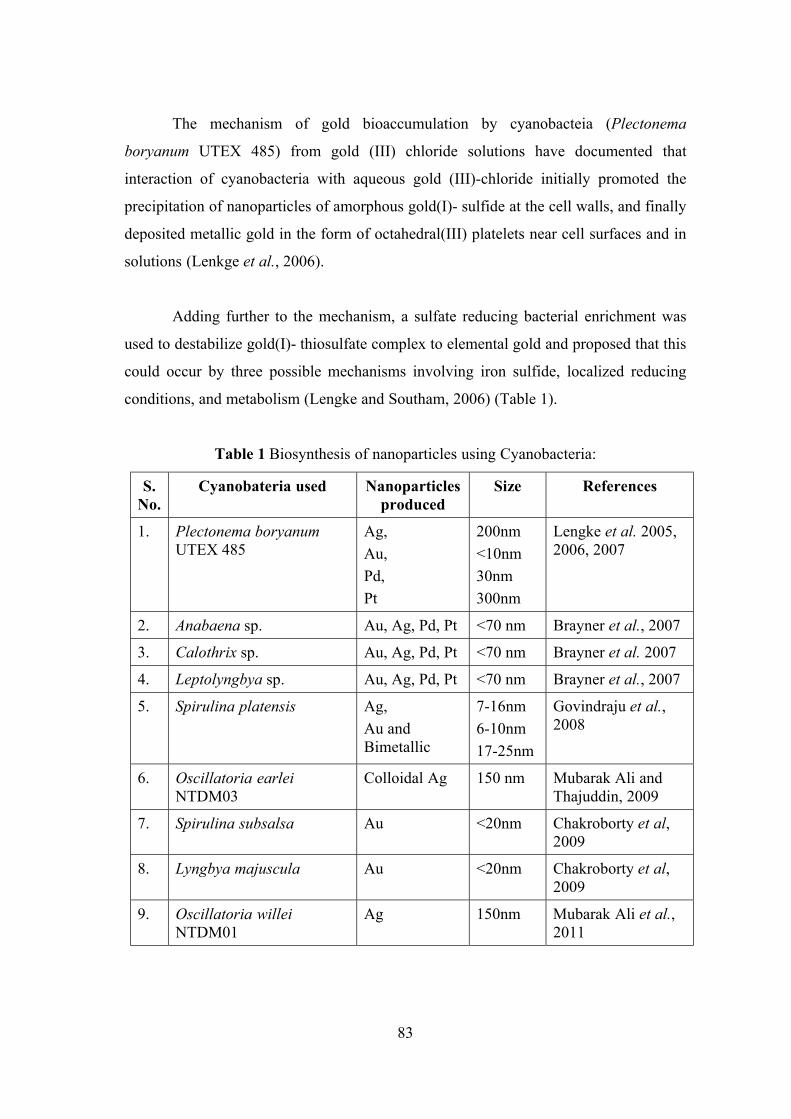
83
The mechanism of gold bioaccumulation by cyanobacteia (Plectonema
boryanum UTEX 485) from gold (III) chloride solutions have documented that
interaction of cyanobacteria with aqueous gold (III)-chloride initially promoted the
precipitation of nanoparticles of amorphous gold(I)- sulfide at the cell walls, and finally
deposited metallic gold in the form of octahedral(III) platelets near cell surfaces and in
solutions (Lenkge et al., 2006).
Adding further to the mechanism, a sulfate reducing bacterial enrichment was
used to destabilize gold(I)- thiosulfate complex to elemental gold and proposed that this
could occur by three possible mechanisms involving iron sulfide, localized reducing
conditions, and metabolism (Lengke and Southam, 2006) (Table 1).
Table 1 Biosynthesis of nanoparticles using Cyanobacteria:
S. No.
Cyanobateria used Nanoparticles produced
Size References
1. Plectonema boryanum UTEX 485
Ag,
Au,
Pd,
Pt
200nm
<10nm
30nm
300nm
Lengke et al. 2005, 2006, 2007
2. Anabaena sp. Au, Ag, Pd, Pt <70 nm Brayner et al., 2007
3. Calothrix sp. Au, Ag, Pd, Pt <70 nm Brayner et al. 2007
4. Leptolyngbya sp. Au, Ag, Pd, Pt <70 nm Brayner et al., 2007
5. Spirulina platensis Ag,
Au and Bimetallic
7-16nm
6-10nm
17-25nm
Govindraju et al., 2008
6. Oscillatoria earlei NTDM03
Colloidal Ag 150 nm Mubarak Ali and Thajuddin, 2009
7. Spirulina subsalsa Au <20nm Chakroborty et al, 2009
8. Lyngbya majuscula Au <20nm Chakroborty et al, 2009
9. Oscillatoria willei NTDM01
Ag 150nm Mubarak Ali et al., 2011
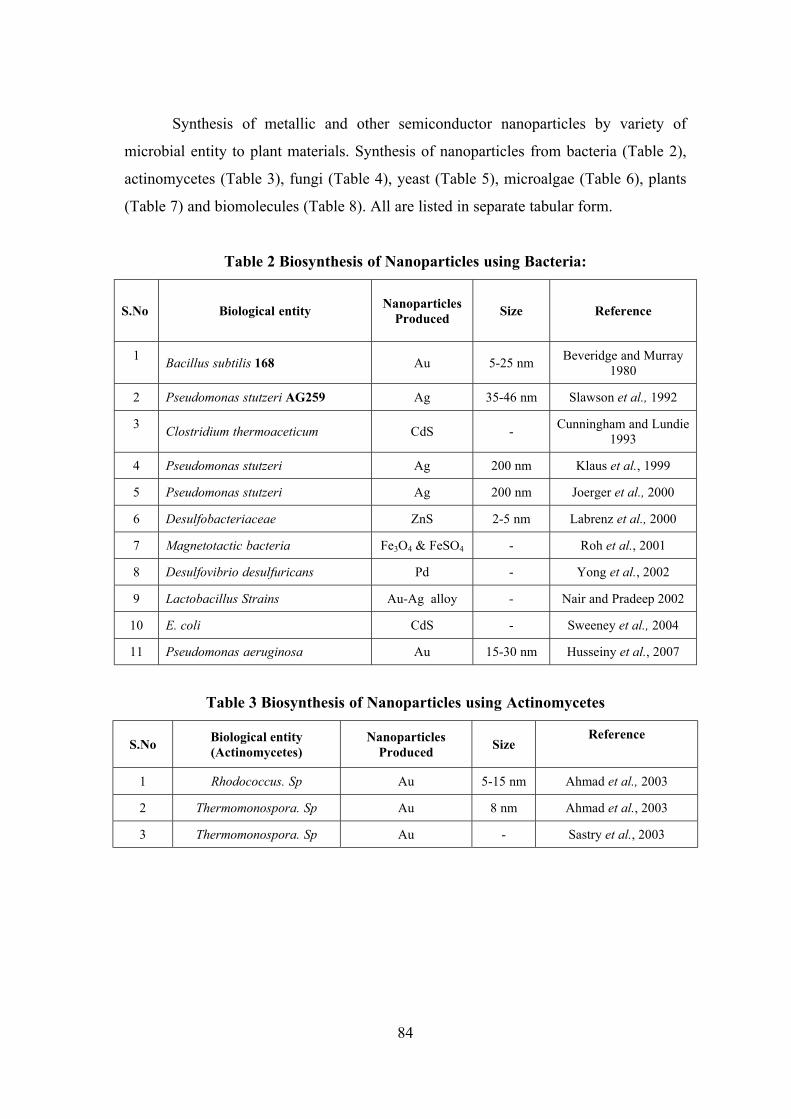
84
Synthesis of metallic and other semiconductor nanoparticles by variety of
microbial entity to plant materials. Synthesis of nanoparticles from bacteria (Table 2),
actinomycetes (Table 3), fungi (Table 4), yeast (Table 5), microalgae (Table 6), plants
(Table 7) and biomolecules (Table 8). All are listed in separate tabular form.
Table 2 Biosynthesis of Nanoparticles using Bacteria:
S.No Biological entity Nanoparticles
Produced Size Reference
1 Bacillus subtilis 168 Au 5-25 nm
Beveridge and Murray 1980
2 Pseudomonas stutzeri AG259 Ag 35-46 nm Slawson et al., 1992
3 Clostridium thermoaceticum CdS -
Cunningham and Lundie 1993
4 Pseudomonas stutzeri Ag 200 nm Klaus et al., 1999
5 Pseudomonas stutzeri Ag 200 nm Joerger et al., 2000
6 Desulfobacteriaceae ZnS 2-5 nm Labrenz et al., 2000
7 Magnetotactic bacteria Fe3O4 & FeSO4 - Roh et al., 2001
8 Desulfovibrio desulfuricans Pd - Yong et al., 2002
9 Lactobacillus Strains Au-Ag alloy - Nair and Pradeep 2002
10 E. coli CdS - Sweeney et al., 2004
11 Pseudomonas aeruginosa Au 15-30 nm Husseiny et al., 2007
Table 3 Biosynthesis of Nanoparticles using Actinomycetes
S.No Biological entity (Actinomycetes)
Nanoparticles Produced
Size Reference
1 Rhodococcus. Sp Au 5-15 nm Ahmad et al., 2003
2 Thermomonospora. Sp Au 8 nm Ahmad et al., 2003
3 Thermomonospora. Sp Au - Sastry et al., 2003
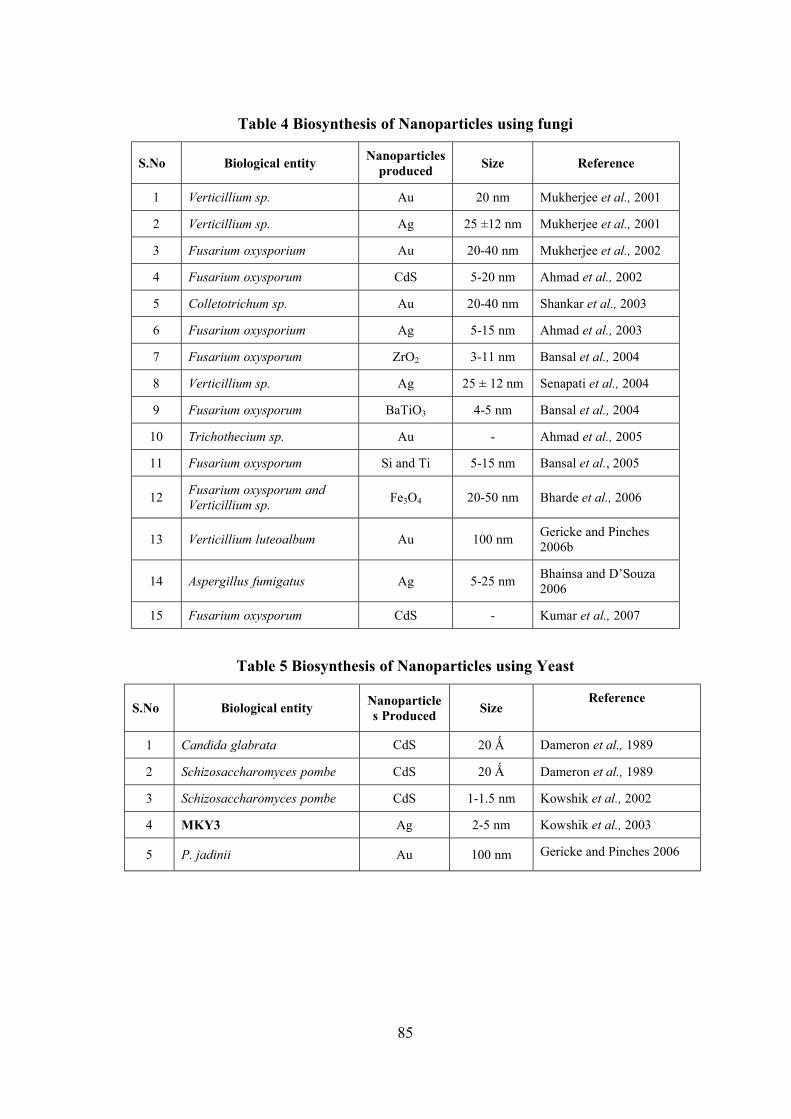
85
Table 4 Biosynthesis of Nanoparticles using fungi
S.No Biological entity Nanoparticles
produced Size Reference
1 Verticillium sp. Au 20 nm Mukherjee et al., 2001
2 Verticillium sp. Ag 25 ±12 nm Mukherjee et al., 2001
3 Fusarium oxysporium Au 20-40 nm Mukherjee et al., 2002
4 Fusarium oxysporum CdS 5-20 nm Ahmad et al., 2002
5 Colletotrichum sp. Au 20-40 nm Shankar et al., 2003
6 Fusarium oxysporium Ag 5-15 nm Ahmad et al., 2003
7 Fusarium oxysporum ZrO2 3-11 nm Bansal et al., 2004
8 Verticillium sp. Ag 25 ± 12 nm Senapati et al., 2004
9 Fusarium oxysporum BaTiO3 4-5 nm Bansal et al., 2004
10 Trichothecium sp. Au - Ahmad et al., 2005
11 Fusarium oxysporum Si and Ti 5-15 nm Bansal et al., 2005
12 Fusarium oxysporum and Verticillium sp.
Fe3O4 20-50 nm Bharde et al., 2006
13 Verticillium luteoalbum Au 100 nm Gericke and Pinches 2006b
14 Aspergillus fumigatus Ag 5-25 nm Bhainsa and D’Souza 2006
15 Fusarium oxysporum CdS - Kumar et al., 2007
Table 5 Biosynthesis of Nanoparticles using Yeast
S.No Biological entity Nanoparticles Produced
Size Reference
1 Candida glabrata CdS 20 Ǻ Dameron et al., 1989
2 Schizosaccharomyces pombe CdS 20 Ǻ Dameron et al., 1989
3 Schizosaccharomyces pombe CdS 1-1.5 nm Kowshik et al., 2002
4 MKY3 Ag 2-5 nm Kowshik et al., 2003
5 P. jadinii Au 100 nm Gericke and Pinches 2006
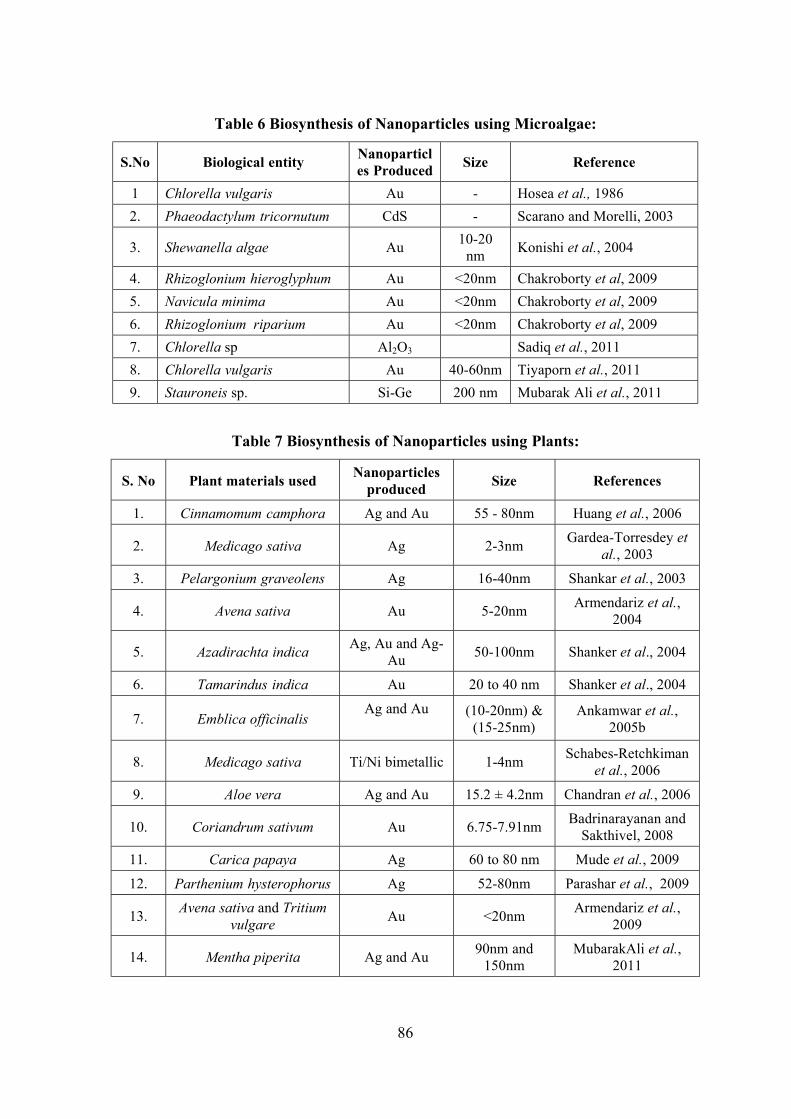
86
Table 6 Biosynthesis of Nanoparticles using Microalgae:
S.No Biological entity Nanoparticles Produced
Size Reference
1 Chlorella vulgaris Au - Hosea et al., 1986
2. Phaeodactylum tricornutum CdS - Scarano and Morelli, 2003
3. Shewanella algae Au 10-20
nm Konishi et al., 2004
4. Rhizoglonium hieroglyphum Au <20nm Chakroborty et al, 2009
5. Navicula minima Au <20nm Chakroborty et al, 2009
6. Rhizoglonium riparium Au <20nm Chakroborty et al, 2009
7. Chlorella sp Al2O3 Sadiq et al., 2011
8. Chlorella vulgaris Au 40-60nm Tiyaporn et al., 2011
9. Stauroneis sp. Si-Ge 200 nm Mubarak Ali et al., 2011
Table 7 Biosynthesis of Nanoparticles using Plants:
S. No Plant materials used Nanoparticles
produced Size References
1. Cinnamomum camphora Ag and Au 55 - 80nm Huang et al., 2006
2. Medicago sativa Ag 2-3nm Gardea-Torresdey et
al., 2003
3. Pelargonium graveolens Ag 16-40nm Shankar et al., 2003
4. Avena sativa Au 5-20nm Armendariz et al.,
2004
5. Azadirachta indica Ag, Au and Ag-
Au 50-100nm Shanker et al., 2004
6. Tamarindus indica Au 20 to 40 nm Shanker et al., 2004
7. Emblica officinalis Ag and Au
(10-20nm) &
(15-25nm) Ankamwar et al.,
2005b
8. Medicago sativa Ti/Ni bimetallic 1-4nm Schabes-Retchkiman
et al., 2006
9. Aloe vera Ag and Au 15.2 ± 4.2nm Chandran et al., 2006
10. Coriandrum sativum Au 6.75-7.91nm Badrinarayanan and
Sakthivel, 2008
11. Carica papaya Ag 60 to 80 nm Mude et al., 2009
12. Parthenium hysterophorus Ag 52-80nm Parashar et al., 2009
13. Avena sativa and Tritium
vulgare Au <20nm
Armendariz et al., 2009
14. Mentha piperita Ag and Au 90nm and
150nm MubarakAli et al.,
2011
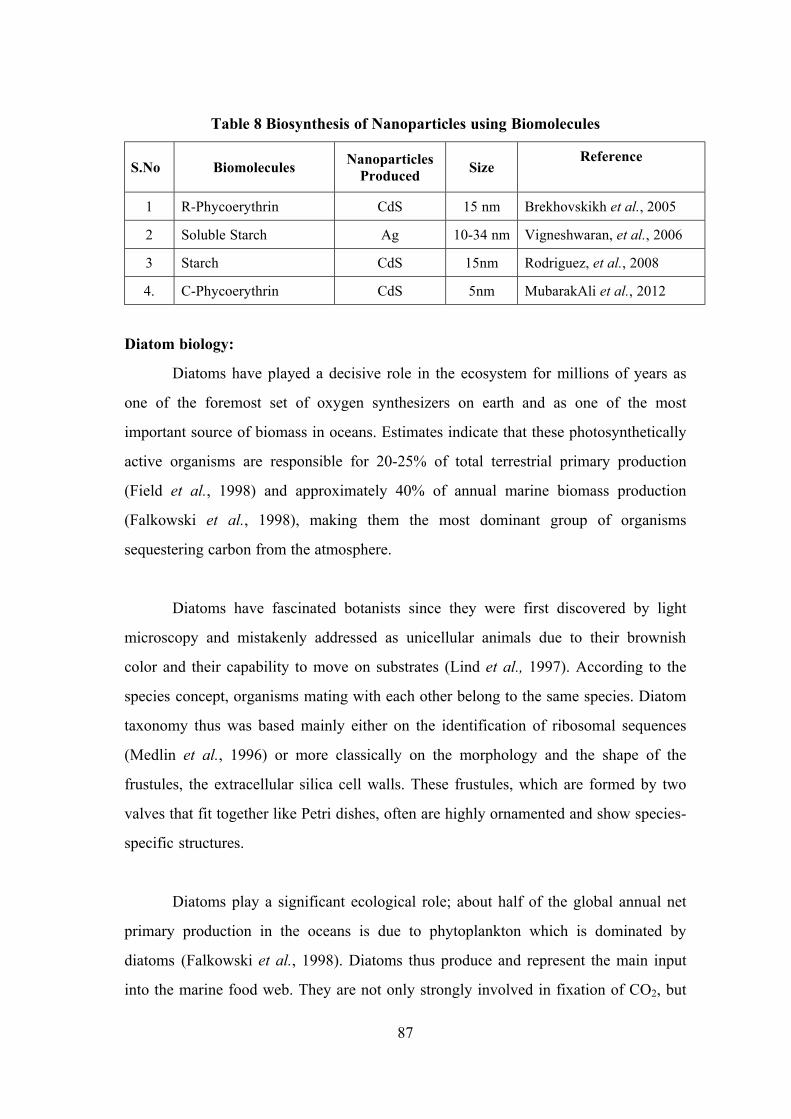
87
Table 8 Biosynthesis of Nanoparticles using Biomolecules
S.No Biomolecules Nanoparticles
Produced Size
Reference
1 R-Phycoerythrin CdS 15 nm Brekhovskikh et al., 2005
2 Soluble Starch Ag 10-34 nm Vigneshwaran, et al., 2006
3 Starch CdS 15nm Rodriguez, et al., 2008
4. C-Phycoerythrin CdS 5nm MubarakAli et al., 2012
Diatom biology:
Diatoms have played a decisive role in the ecosystem for millions of years as
one of the foremost set of oxygen synthesizers on earth and as one of the most
important source of biomass in oceans. Estimates indicate that these photosynthetically
active organisms are responsible for 20-25% of total terrestrial primary production
(Field et al., 1998) and approximately 40% of annual marine biomass production
(Falkowski et al., 1998), making them the most dominant group of organisms
sequestering carbon from the atmosphere.
Diatoms have fascinated botanists since they were first discovered by light
microscopy and mistakenly addressed as unicellular animals due to their brownish
color and their capability to move on substrates (Lind et al., 1997). According to the
species concept, organisms mating with each other belong to the same species. Diatom
taxonomy thus was based mainly either on the identification of ribosomal sequences
(Medlin et al., 1996) or more classically on the morphology and the shape of the
frustules, the extracellular silica cell walls. These frustules, which are formed by two
valves that fit together like Petri dishes, often are highly ornamented and show species-
specific structures.
Diatoms play a significant ecological role; about half of the global annual net
primary production in the oceans is due to phytoplankton which is dominated by
diatoms (Falkowski et al., 1998). Diatoms thus produce and represent the main input
into the marine food web. They are not only strongly involved in fixation of CO2, but

88
also in cycling of soluble silicates by integrating them into the shells and by releasing
parts of them after decomposition at the bottom of the oceans (Bidel and Azam, 1999).
It is not yet fully understood why diatoms filled aquatic niches so successfully, even
though they have the capability to grow at a wide range of light intensities (Falkowski
et al., 2004) and the apparent ability to perform C4 photosynthesis (Reinfelder, 2004).
Diatom nanotechnology:
The ability of diatoms to genetically define these structures makes them highly
interesting for nanotechnological applications (Drum and Gordon, 2003). Because of
their siliceous composition frustules were often well preserved in fossil deposits
(Damste et al., 2004) and strong sedimentation of diatom shells in ancient oceans led to
the deposition of siliceous earth or diatomite, a material of high importance for
industrial uses (Harwood, 1999).
There is a tremendous research interest in the area of nanotechnology to develop
reliable processes for the synthesis of nanomaterials over a range of sizes and chemical
compositions. Although the conventional methods of synthesis of metal sols, known
since the times of Michael Faraday, continue to be used for generating metal
nanoparticles, there have been several improvements and modifications in the methods
which provide a better control over the size, shape, and other characteristics of the
nanoparticles (Burda, et al., 2005 and Sun and Xia, 2002). These developments have
enabled studies of quantum confinement as well as other properties dependent on size,
shape, and composition (Rao, et al., 2002).
Ligating nanoparticles with organic molecules and assembling these in one,
two, or three-dimensional mesostructures have added another dimension to this field
where in collective properties of nanoparticles have been of special interest
(Kakkassery et al., 2004). The exciting potential of nanomaterials can be routed to nano
device applications, only with a combination of nano building units and strategies for
assembling them.

89
Synthesis and assembly strategies of nanoparticles mostly accommodate
precursors from liquid, solid or gas phase; employ chemical or physical deposition
approaches and similarly rely on either chemical reactivity or physical compaction to
integrate the nanostructure building blocks within the final material structure.
Bottom up approach:
The bottom up approach of nanomaterials synthesis first forms the
nanostructured building blocks (nanoparticles) and then assembles these into the final
material. An example of this approach is the formation of powder components through
aerosol and sol-gel techniques and then the compaction of the components into the final
material. Nanoparticles with diameters ranging from 1 nm to 10 nm with consistent
crystal structure, surface derivatisation, and a high degree of monodispersity can be
processed by these techniques (Murray et al., 1993).
Top-down approach
The top-down approach begins with a suitable starting material and then sculpts
the functionality from the material. This technique is similar to the approach used by
the semiconductor industry in forming devices out of an electronic substrate (silicon),
utilizing pattern formation (such as electron beam lithography) and pattern-transfer
processes (such as reactive-ion etching) that have the requisite spatial resolution to
achieve creation of structures at the nanoscale. This particular area of nanostructure
formation has tremendous scope and is a driving issue for the electronics industry.
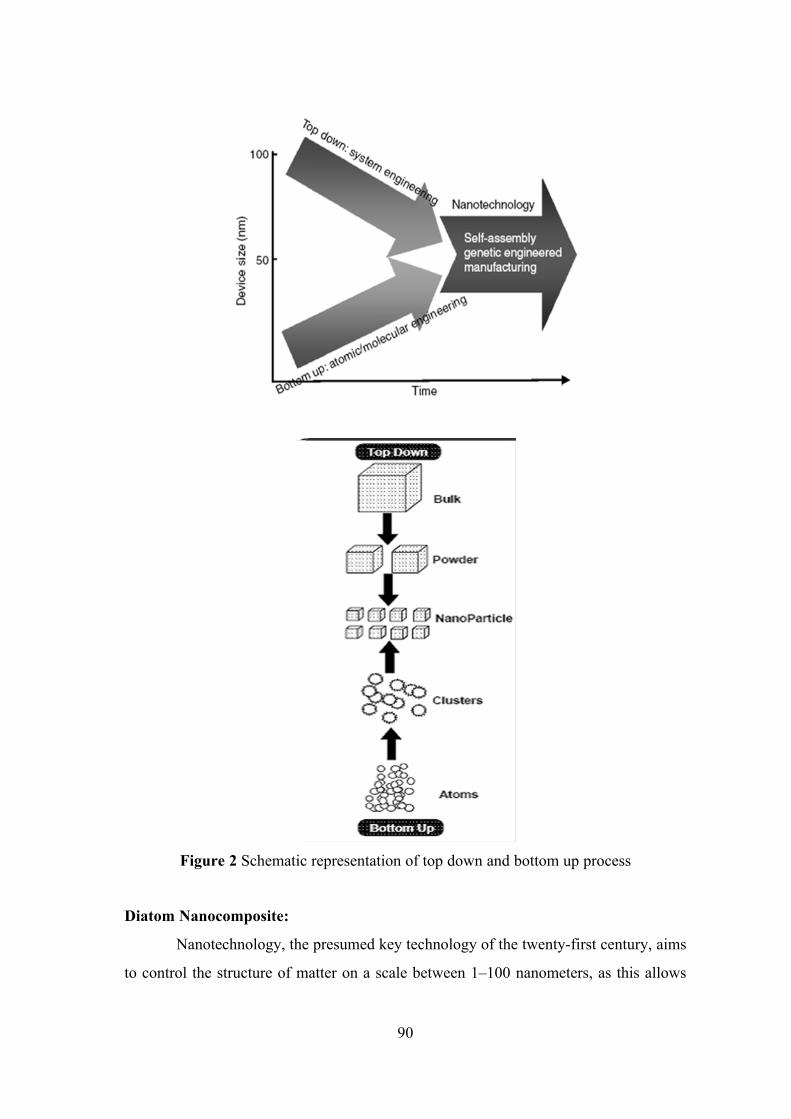
90
Figure 2 Schematic representation of top down and bottom up process
Diatom Nanocomposite:
Nanotechnology, the presumed key technology of the twenty-first century, aims
to control the structure of matter on a scale between 1–100 nanometers, as this allows

91
exploitation of new physical phenomena (Poole and Owens, 2003). The establishment
of an industry that produces nanotechnology based devices critically depends on
methods for inexpensive manufacturing of nanoscale materials on a massively parallel
scale; yet current nanomanufacturing methods are slow and require expensive
equipment.
It has recently been suggested that the silica structures produced by diatoms
may be particularly useful for nanotechnological applications, because they exhibit
highly controlled nanopatterns that can be produced by biological self-assembly in
large quantities and at low cost (Parkinson and Gordon, 1999 and Sandhage et al.,
2005). A variety of applications have been suggested for diatom silica, including
membranes for biomolecule separations (Losic et al., 2006; Parkinson and Gordon,
1999), size-selective biosensors (Parkinson and Gordon, 1999), microelectromechanical
systems (Drum and Gordon, 2003), and masks for microfabrication (Parkinson and
Gordon, 1999).
To fully exploit the potential of diatom silica, methods have been recently
developed to introduce additional functionalities. The chemical attachment of single-
stranded DNA (ssDNA) to cleaned diatom silica enabled the diatom silica to be
covered with a dense layer of gold particles or the fluorescent dye Cy3 through
hybridization with Au-labeled and Cy3-labeled complementary ssDNA. The modified
diatom silica may be useful for catalysis and optics, specifically as substrates for
Surface Enhanced Raman Scattering (SERS) (Rosi et al., 2004).
Recently, an alternative method for biomolecule attachment to diatom silica has
been established by Kroger and coworkers. This method is based on molecular genetic
engineering of T. pseudonana and enables immobilization of the enzyme
hydroxylaminobenzene mutase (HabB) within the diatom silica by expressing a tpSil3-
HabB fusion protein. Due to the presence of the silaffin domain, the fusion protein
became incorporated into diatom silica in vivo, remained attached to the silica after

92
isolation of the cell wall, and the HabB moiety was catalytically active (Poulsen et al.,
2007).
The method is likely applicable to a large variety of enzymes or receptor
proteins, and represents a new paradigm for utilizing biomineralization for the
sustainable production of nanomaterials with tailored functionalities. Because of the
excellent mechanical (Hamm et al., 2003) and mass transport properties of diatom
silica (Losic et al., 2006), enzyme-functionalized diatom cell walls may be attractive
materials for microfluidic-based devices.
The applications for diatom cell walls are limited by the chemical and physical
properties of silica. Therefore, it would be desirable to convert diatom silica into other
materials without changing the morphology. Indeed, the pioneering work of Sandhage
and co-workers has resulted in a number of protocols that yielded exact replicas of
diatom cell walls consisting entirely of MgO, TiO2, BaTiO3, elemental Si, and several
other materials.
These materials have a wide variety of properties and potential applications. For
example the fluorine-doped TiO2 replicas of diatom silica were highly efficient
catalysts for the rapid hydrolysis of organophosphate-based pesticides (Lee et al.,
2007), and the Si replicas represented ultra rapid, low-voltage NO sensors (Bao et al.,
2007).In a different approach, Mirkin and co-workers have used diatom silica as
sacrificial template to generate 30-nm thick silver microshells that preserved the sub-
100-nm topological features of diatom silica and could be used as SERS substrates for
the sensitive detection of organic molecules (Payne et al., 2005).
Nanofabrication:
Currently, there is enormous interest in bioinspired processes for the fabrication
and self-assembly of semiconductor nanostructures that possess unique optical and
electronic Although biomimetic approaches typically focus on biomolecule-mediated

93
processes, living cells also have the potential to direct the fabrication of nanostructured
materials, particularly for photonic device applications (Parker and Townley, 2007).
Diatoms are nature’s priceless manufactures of 3D intricate nanocomposites
(Sandhage et al., 2002). It has been reported that the fabrication of a photosynthetic
biodevice using living diatoms (Leabeau et al., 2002). They revealed that the diatom
cell can be cultured on agar films that are prepared on a glass surface, and these cells
can perform photosynthesis. Although this was an important study on developing
biodevices by using living diatoms, no microscopic characterization of the device was
included. To date no other study has reported the development of biodevices by using
living diatoms.
Biological processes have been developed to incorporate atoms other than
silicon into the silica diatom frustules while maintaining the frustules detailed
morphology. This process has allowed us to fabricate silicon-germanium (Si-Ge) oxide
nanocomposites that possess optoelectronic properties (Rorrer et al., 2005). Diatoms
carry out this process under ambient conditions of temperature and pressure and
produce no environmental pollutants. In that study, several electron microscopy and
microanalysis techniques were used to characterize the morphologies and elemental
composition of the processed diatoms and more importantly to assess and direct the
optimization of the fabrication of the nanocomposites.
The electron microscopy characterization and microanalysis results suggest that
germanium doped into the silica frustules of the diatom has an upper threshold that if
exceeded introduces structural aberrations and the assimilated germanium is randomly
distributed within a diatom frustules. Soluble germanium can be metabolically inserted
into the biosilica of the pennate diatom Pinnularia sp., by a two stage bioreactor
cultivation process, where the incorporation of Ge into the frustules biosilica reduced
the frustule pore diameter but otherwise did not affect the frustules morphology
(Jeffreys et al., 2006).
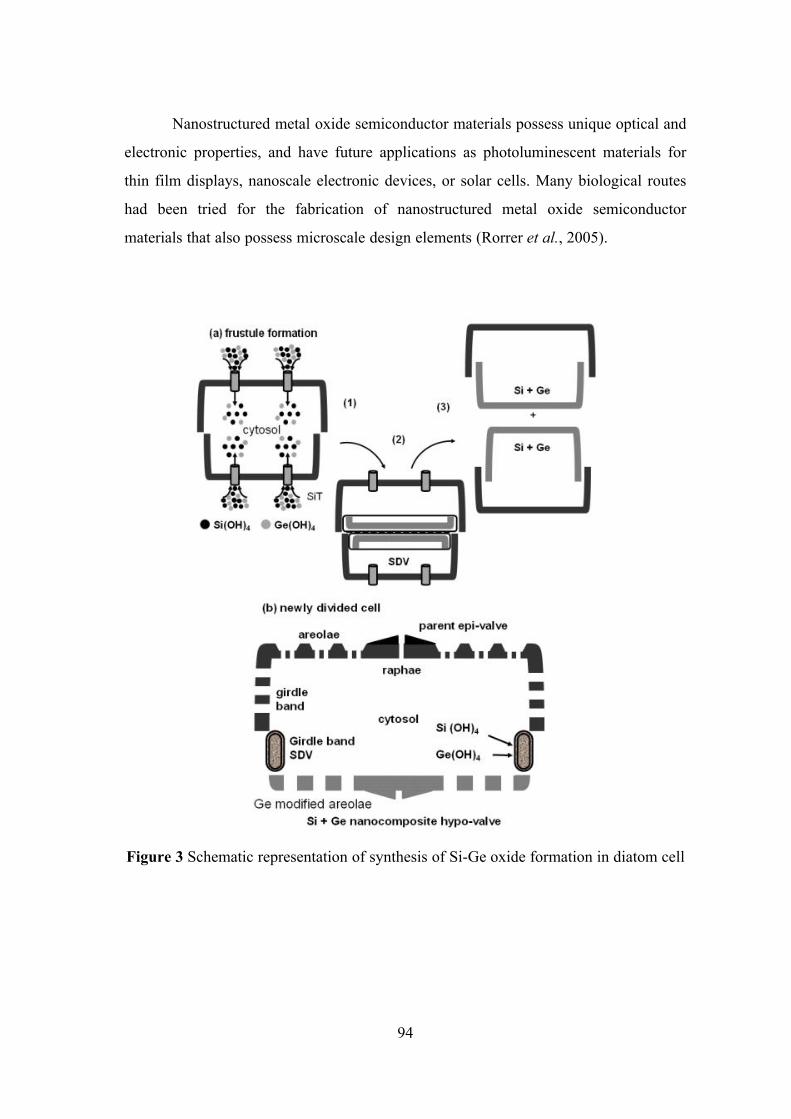
94
Nanostructured metal oxide semiconductor materials possess unique optical and
electronic properties, and have future applications as photoluminescent materials for
thin film displays, nanoscale electronic devices, or solar cells. Many biological routes
had been tried for the fabrication of nanostructured metal oxide semiconductor
materials that also possess microscale design elements (Rorrer et al., 2005).
Figure 3 Schematic representation of synthesis of Si-Ge oxide formation in diatom cell

95
Applications of Nanoparticles:
Figure 4 Diagramatic representation of application of nanoparticles in different fields
Nanomaterials have been blossoming in the field of nanotechnology. Their
unique size-dependent properties make these materials superior and in dispensable in
many areas of human activity (Murray et al., 2000).
The nanomaterials level is the most advanced at present, both in scientific
knowledge and in commercial applications. The fact that the nanoparticles exist in the
same size domain as proteins makes nanoparticles suitable for biotagging or labeling.
However, size is just one of many characteristics of nanoparticles that it is rarely
sufficient if one is to use nanoparticles as biological tags. The bioinorganic interface
should be attached to the nanoparticles for the good interactions with the biological
target (Bruchez et al., 1998).

96
Nanoparticles usually form the core of nanobiomaterial. It can be used as a
convenient surface for molecular assembly and may be composed of inorganic or
polymeric materials. It can also be in the form of nanovesicle surrounded by a
membrane or a layer. The shape is more spherical but cylindrical, plate-like and other
shapes are possible. The size and size distribution might be important in some cases, for
example, if penetration through a pore structure of a cellular membrane is required. The
size and size distribution are becoming extremely critical when quantum sized effects
are used to control material properties. A tight control of the average particle size and a
narrow distribution of sizes allow creating very efficient fluorescent probes that emit
narrow light in a very wide range of wavelengths. This helps with creating biomarkers
with many and well distinguished colors. The core itself might have several layers and
be multi functional.
Tissue Engineering:
Natural bone surface is quite often contains features that are about 100 nm
across. If the surface of an artificial bone implant were left smooth, the body would try
to reject it. Because of that smooth surface is likely to cause production of a fibrous
tissue covering the surface of implant. This layer reduces the bone-implant contact,
which may result in loosening of the implant and further inflammation (De La et al.,
2003).
In order to reduce the bone implant rejection as well as to induce the production
of osteoblast, the nanosized features can be created on the surface of hip or knee
prosthesis. The osteoblasts are the cells responsible for the growth of the bone matrix
and are found on the advancing surface of the developing bone. More than 90% of the
human bone cells from suspension adhere to the nanostructured metal surface. It was
shown that biomimetic approach a slow growth of nanostructured appetite film from
the stimulated body fluid-resulted in the formation of a strongly adherent, uniform
nanoporous layer. The layer was found to be built of 60 nm crystallites and posses a
stable nanoporous structure and activity (Ma et al., 2003).

97
Cancer Therapy:
Photo dynamic cancer therapy is based on the destruction of the cancer cells by
laser generated atomic oxygen, which is cytotoxic. A greater quantity of a special dye
that is used to generate atomic oxygen is taken in by cancer cells when compared with
the healthy tissue. Hence, only the cancer cells are destroyed then exposed to a laser
radiation. Unfortunately the remaining molecules migrate to the skin and the eyes and
make the patient very sensitive to the day light exposure. This effect can last for up to
six weeks (Yoshida et al., 1999).
To avoid these side effects, the hydrophobic version of the dye molecule was
enclosed inside a porous nanoparticle (Roy et al., 2003). The dye stayed trapped inside
the ormosil nanoparticle and didn’t spread to the other parts of the body. At the same
time its oxygen generating ability has not been affected and the pore size of about 1nm
freely allowed for the oxygen diffuse out.
Multicolor Optical Coding For Biological Assays:
The ever increasing research in proteomics and genomics generates escalating
number of sequence data and requires development of high throughputs screening
technologies. Realistically various array technologies that are currently used in parallel
analysis are likely to reach saturation when a number of array elements exceed several
millions.
The selection of nanoparticles used in those experiments had six different colors
as well as ten intensities. It is enough to encode over 1 million combinations. The
uniformity and reproducibility of beads was high letting for the bead identification
accuracies of 99.99 % (Han et al., 2001).
Manipulation of Cells and Bio Molecules:
Functionalized magnetic nanoparticles have found many applications including
cell separation and probing. For example, prophyrins with thiol or carboxyl linkers
were simultaneously attached to the gold or nickel segments respectively. Thus it is
possible to produce magnetic nanowires with spatially segregated fluorescent parts. In

98
addition because of the large aspect ratios the residual magnetization of these
nanowires can be high. Hence weaker magnetic fields can be used to drive them. It has
been show that a self assembly of magnetic nanowires in suspension can be controlled
by weak external magnetic fields. This would potentially allow controlling cell
assembly in different shapes and forms (Reich et al., 2003).
Protein Detection:
Gold nanoparticles are widely used in immune histochemistry to identify
protein-protein interactions. The group of Mirkin has designed a sophisticated multi
functional probe that is built around a 13 nm gold nanoparticle. The nanoparticles are
coated with hydrophilic oligo nucleotides containing a Raman dye at one end and
terminally capped with a small molecule recognition element (e.g. biotin).
Moreover this molecule is catalytically active and will be coated with silver in
the solution of Silver (I) and hydroquinine. After the probe is attached to a small
molecule or an antigen it is designed to detect the substrate which is exposed to silver
and hydroquinine solutions. The protein can be detected by modifying the probe to
contain antibodies on the surface (Nam et al., 2003).
Electrochemical Application:
The bioreduction of aqueous gold chloride ions by the bacterium E.coli DH5α
assist the formation of gold nanoparticles. The nanoparticles are bound to the surface of
the bacteria and this composite may be used for application in realizing the direct
electrochemistry of hemoglobin. The bacteria-mediated production of gold
nanoparticles is a green chemistry synthesis procedure and also provides a new
approach to achieve the direct electrochemistry of proteins (Du et al., 2007).
Nanoparticles as ROS Scavenger:
Major problem with T2DM patients is delayed healing of wound. Several
literatures show this is because of increase in oxidative stress and decreased
antioxidants level content leads to increased inflammation, which results huge amount
of ROS production that leads to premature apoptosis of inflammatory cells.

99
Recently, nanotechnology and nanoparticle field is a fast-growing research area.
Research has already led to significant breakthrough and several products are available
commercially. Recently, role of silver nanoparticle in diabetic wound healing therapy
has also been considered. However, the complete mechanisms of silver nanoparticle in
bacteriostatic phenomena in wound healing remain unknown.
It has been reported a novel Ag+-loaded zirconium phosphate nanoparticle
plays crucial role in diabetic wound healing (Manish Mishra et al., 2008). Some of the
nanoparticle now a day are preparing in a manner that act as free radical scavenger
(Wasrheit, 2004). Moreover, reports reveal Cerium oxide (CeO2, ceria) plays major
active role because of its excellent free radical scavenging potentials (Chung, 2003).
This metal oxide is monodisperse particles with single crystals and few twin boundaries
(Zhang et al., 2002) as well as expanded lattice parameter (Perebeinos et al., 2002).
Moreover, CeO2 tends to be a nonstoichiometric compound. Cerium atom
characterized by both +4 and +3 oxidation states.
Literatures shows X-ray photoelectron spectroscopy and X-ray absorption near
edge spectroscopy suggests that the concentration of Ce3+ relative to Ce4+ increases as
particle size decreases with a conservative [Ce3+] minimum of 6% in 6 nm
nanoparticles and 1% in 10 nm particles (Zhang et al., 2004). This dual oxidation state
means that these nanoparticles have oxygen vacancies (Robinson et al., 2002). The loss
of oxygen and the reduction of Ce4+ to Ce3+ is accompanied by creation of an oxygen
vacancy. This property is responsible for the interesting redox chemistry exhibited by
CeO2 nanoparticles and makes them attractive for wound healing process followed by
scavenging properties.
Nanoparticles made of other metal oxides were also considered for its potential
scavenger behavior. These included particles aluminum oxide (Al2O3), commonly
known as alumina. That differs in crystal structure and yttrium oxide (Y2O3, yttria).
Moreover, Alumina is also thermodynamically stable at all temperatures and has a
corundum like structures with oxygen atoms adopting hexagonal close-packing and

100
Al3+ ions, filling 2/3 of the octahedral sites in the lattice. However, yttria has a cubical
structure (Kingery et al., 1976). Yttrium oxide is now a day considered most significant
due to its highest free energy of oxide formation from elemental yttrium among known
metal oxides (Kilboun et al., 1994).
It is characterized by only small changes from stoichiometry under normal
conditions of temperature and pressure (Kofstad, 1972) and by atmospheric absorption
of H2O and CO2 (Kilbourn et al., 1986). The particular polymorph of it is the
monoclinic B form, which is closely related to it’s A - form having hexagonal close-
packing (hcp). Unlikely the A form which has only sevenfold coordination states, it has
six fold (Kofstad, 1972). The B form structure is slightly less dense than the A form
variant and has the yttrium cations in nonequivalent sites in the crystal (Atou et al.,
1990). These groups of nanoparticles are relatively nontoxic to neutrophiles and
macrophages, and that CeO2 and Y2O3 particles protect cells from death due to
oxidative stress. This protection is due to the direct antioxidant properties of the
nanoparticles.
Cerium and yttrium oxide nanoparticles are able to rescue cells from oxidative
stress-induced cell death in a manner that appears to be dependent upon the structure of
the particle but independent of its size within the range of 6–1000 nm. This might be
useful for the diabetic wound healing. Furthermore, three alternative explanations were
demonstrated that the cerium oxide and yttrium oxide particles protect from oxidative
stress. They may act as direct antioxidants, block ROS production, which inhibit
programmed cell death pathway and they may directly cause a low level of ROS
production, which rapidly induces a ROS defense system before the glutamate-induced
cell death program is complete. The latter is a form of preconditioning that could be
caused by the exposure of cells to particulate material known to induce low levels of
ROS (Becker et al., 2002). Therefore it could be suggested that CeO2, alumina and
yttrium may be useful for diabetic wound healing mediated be ROS scavenging
potentials of above nanoparticles and have diabetic wound healing therapy.

101
MATERIALS AND METHODS
Selection of Cyanobacteria and diatom
Cyanobacteria culture:
The filamentous marine cyanobacteria, Phormidium willei NTDM01 and
Phormidium tenue NTDM05 were selected for this study and were grown in batch
culture in ASN - III. The pH of the medium was adjusted to 8.0 using a 1M NaOH
solution before experimentation; the culture were transferred and grown for
approximately 6-8 weeks to reach a stationary growth phase. It was then centrifuged
and washed three times with sterile deionized water to remove the salts and trace metals
from the medium before being used for the cyanobacterial experiments.
Diatom Culture:
A diatom species such as Stauroneis sp. has been chosen for the study. It was
grown in F/2 medium. The pH of the medium was adjusted to 8.0 using a 1M NaOH
solution before experimentation; the culture was transferred and grown for
approximately 2-3 weeks to reach a stationary growth phase. It was then centrifuged
and washed three times with sterile distilled water to remove the salts and trace metals
from the medium before being used for the diatom experiments.
These strains were selected to explore nanobiotechnological potentials. In this
regards, Phormidium willei NTDM01 has been chosen for the synthesis of silver and
gold nanoparticles and Phormidium tenue NTDM05 has been selected for the synthesis
of CdS nanoparticles and finally, a diatom Stauroneis sp. also been selected to adopt for
the synthesis of Si-Ge oxide nanocomposites. The detailed experimentation for the
synthesis of nanomaterials have described below.
Biosynthesis of Nanoparticles:
Synthesis of silver nanoparticles using cyanobacteria have adopted a method
that described earlier by Lengke et al., (2005) such as biotic and abiotic experiment and
for the gold nanoparticles modified method of Lengke et al., (2006) have been adopted.

102
Silver nanoparticles:
a. Biotic experiment:
The cyanobacterial experiments were conducted to examine the role of
cyanobacteria in the synthesis of silver nanoparticles from aqueous solution of AgNO3.
The experiments were conducted using washed cyanobacteria and without the presence
of the growth medium.
To initiate the experiment 5mL of the silver solution (~560mg/l) was added to
5mL of washed Cyanobacteria culture. The experiment were conducted at 25 oC for 28
days after the incubation period with silver solution and maintained in the dark, and
cyanobacteria populations were measured with time. pH, cell viability and precipitation
were measured with time.
b. Abiotic experiments:
Abiotic control experiments consisting of ~560mg/l in silver were conducted
using AgNO3 solution without addition of cyanobacteria.
Gold nanoparticles:
a. Biotic experiment:
The cyanobacterial experiments were conducted to examine the role of
cyanobacteria in the synthesis of gold nanoparticles from aqueous solution of HAuCl4.
The experiments were conducted using washed cyanobacteria and without the presence
of the growth medium.
To initiate the experiment 100mL of the Chloroauric acid (1mM) was added to
5g of washed cyanobacteria culture. The experiment were conducted at 25oC for 5 days
and maintained in the dark, and cyanobacteria populations were measured with time
and the pH, cell viability and precipitation were measured with time.
b. Abiotic experiments:
Abiotic control experiments consisting of Au2+ (1mM) were conducted using
HAuCl4 solution without addition of the presence of Cyanobacteria.

103
Characterization of silver and gold nanoparticles:
Uv-Vis spectrum analysis:
Aqueous silver nitrate solution (10-3M) taken in a clean conical flask inoculated
with isolated cyanobacteria. The reduction of the Ag+ ions by biological species in the
solution was monitored by sampling the aqueous component and measuring the Uv -
Vis Spectrum of solutions. The Uv - Visible spectra of this solution was recorded in
Lambda 35 in absorbance mode from 300 to 900nm.
Fourier Transform Infra Red analysis:
The experimental system after incubation of AgNO3 solution to form slime
sediment (brown color). This sediment characterization analysis by employing FTIR
(Fourier Transform Infra Red). The silver nanoparticles prepared as described above
were analyzed by Spectrum RX1. The FTIR spectrum was taken in the mid IR region
of 400-4000 cm-1. The spectrum was recorded using ATR (Attenuated Total
Reflectance) technique. The sample was directly placed in the Potassium bromide
crystal and the spectrum was recorded in the transmittance mode.
Scanning Electron Microscope with Energy Dispersive Spectrum analysis:
A drop of aqueous solution containing the silver nanomaterials (brown color)
was placed on the sterile stains steel coupon and it air dried to applied gold sputtering
(model). These plates were examined at different magnifications (10,000X, 40,000X)
by scanning electron microscope (Model – JEOL Instrument JSM-5610). The natures
of elements were identified by EDX.
Transmission Electron Microscopic study:
A drop of the aqueous solution of silver and gold nanoparticles were separately
placed onto a carbon coated copper grid, and air-dried and then analyzed in a
transmission electron microscope (TEM).
Biosynthesis of Nanocomposites:
Si-Ge Oxide Nanocomposites:
The two stage cultivation methods are followed for the synthesis of silicon-
germanium nanocomposites as described (Qin et al., 2008 and MubarakAli et al., 2011)

104
1. In first stage, the cells were grown in nutrient medium containing sufficient
silicon concentration for its growth. The silicon concentration (30mg/l) and cell
density is monitored at regular intervals. When all the silicon was used up and
the cell density had become constant, the cells were then considered to be
starved of silicon, this condition prevails one week.
2. In the second stage, the silicon and germanium (1:0.5) is added to the liquid
medium for the biosynthesis of Silicon –Germanium nanocomposites. After 7
days the cells were harvested from the medium and washed thrice in deionized
water in order to remove surface adsorbed germanium and other trace elements.
Then the sample was centrifuged at 2000rpm for 5mins and the pellet was
collected.
Characterization of Si-Ge Oxide Nanocomposites:
The collected pellet was dispersed on the thin slide and allowed to dry in hot air
oven. The small portion of slide was placed onto carbon tape affixed to a stub, the gold
was coated before introduction to the Scanning Electron Microscopy equipped with
EDS. (Model – JEOL Instrument JSM-5610). The natures of elements were identified
by EDS.
Biosynthesis of Semiconductor:
Synthesis of CdS nanoparticles:
CdS nanoparticles were synthesized by reacting Cd2+ with extracted c-
phycoerythrin. The experiment were conducted wherein aqueous solution containing a
mixture of the salt CdCl2 and Na2S in the concentration of 0.25mM and 1mM
respectively.
The reaction volume was kept for incubation and regular monitoring of color
changes; temperature and pH of the solutions were analyzed periodically for 5days
(MubarakAli et al., 2012).
Characterization of CdS nanoparticles:
The appearance of nanoparticles was gauzed in a UV- vis spectrophotometer, in
the wavelength range 200 to 900nm, periodically. Once the nanoparticles were formed,

105
the preparation was lyophilized and analyzed adopting FTIR and HRSEM and EDAX,
with Li drift Si detector (Thermo elution corporation, USA). Finally, a drop of the
sample was placed onto a carbon coated copper grid, and air-dried and then analyzed in
a transmission electron microscope (TEM).
Biomedical application:
The synthesized nanoparticles were analyzed for antibacterial and antioxidant
property with standard protocols.
Antibacterial activity
The multi drug resistant (MDR) pathogens were isolated from hospitalized
patients. Following pathogens were selected for the antibacterial property such as
E.coli, Staphyloccocus aureus, Proteus vulgaris, Klebsiella pneumoniae and
Pseudomonas aeruginosa by well–diffusion method.
The pathogenic cultures were bringing into broth culture for antibacterial assay.
Approximately 7-mm diameter of well was made on Muller Hinton Agar plate with the
help of gel puncture. The cultures were swabbed on test media with sterile cotton swab.
25μl of synthesized particles were inoculated to the well, and then the plates were
incubated in incubator for 37 °C for 24 h, the zones of inhibition was tabulated and
discussed.
Antioxidant property
The effect of extracts on DPPH radical was estimated by DPPH radical
scavenging assay (Pathiranan and Shahidi, 2005). A solution of 0.135 mM DPPH in
methanol was prepared and 1.0 ml of this solution was mixed with 1.0 ml (1:1 ratio) of
nanoparticle suspension in methanol containing (50, 100, 150, 200, 250 µg/ml) of the
nanoparticle. The reaction mixture was vortexed thoroughly and kept in the dark at
room temperature for 30 min. The absorbance of the mixture was measured
spectrophotometrically at 517 nm. Citric acid was used as reference. The ability to
scavenge DPPH radical was calculated by the following equation:

106
DPPH radical scavenging activity (%) = [(Abs*control – Abs*sample)] × 100
(Abs*control)]
Where.,
Abs*control is DPPH solution + methanol (1:1 ratio)
Abs*sample is the absorbance of DPPH radical + synthesized nanoparticles

107
RESULTS AND DISCUSSION
Biosynthesis of nanoparticles:
Silver nanoparticles:
In cyanobacterial experiment with Phormidium willei NTDM01, soluble silver
was completely precipitated from solutions within 28days. A grayish- black silver
precipitate on cyanobacteria was observed in experiment macroscopically. The pH was
constant at 5.0 and 4.7. In abiotic experiments using silver nitrate, total soluble silver
concentration and pH were constant until the completion of experiment (Fig. 4a & b).
The UV-vis Spectrum reveals that the band observed at 450 nm due to the
plasmon resonance of the silver nanoparticles and low wavelength region recorded
from the reaction medium exhibited an absorption band at ca.265 nm and it was
attributed to aromatic amino acids of proteins (Fig. 4c). It is well known that the
absorption band at ca 265 nm arises due to electronic excitation in tryptophan and
tyrosine residue in the protein.
The FTIR spectrum was recorded from the film of silver nanoparticles, formed
after several days of incubation with the bacteria (Fig.5). The bands seen at 3280 cm−1
and 2924 cm−1 were assigned to the stretching vibrations of primary and secondary
amines respectively. The corresponding bending vibrations were seen at 1651 cm−1 and
1548 cm−1, respectively. The two bands observed at 1379 cm−1 and 1033 cm−1 can be
assigned to the C–N stretching vibrations of aromatic and aliphatic amines,
respectively. The presence of protein as the stabilizing agent surrounded the silver
nanoparticles (Muherjee et al., 2001). The silver ions were reduced in the presence of
nitrate reductase, leading to the formation of a stable silver hydrosol 10-25 nm in
diameter and stabilized by the capping peptide.
SEM observation on products of marine cyanobacterium, Phormdium willei
NTDM01 in AgNO3 experiment is presented (Fig.6a). At room temperature, the
addition of AgNO3 to the cyanobacteria caused the precipitation of silver nanoparticles
at cell surfaces. Small spherical silver nanoparticles with size ranging from 100nm to
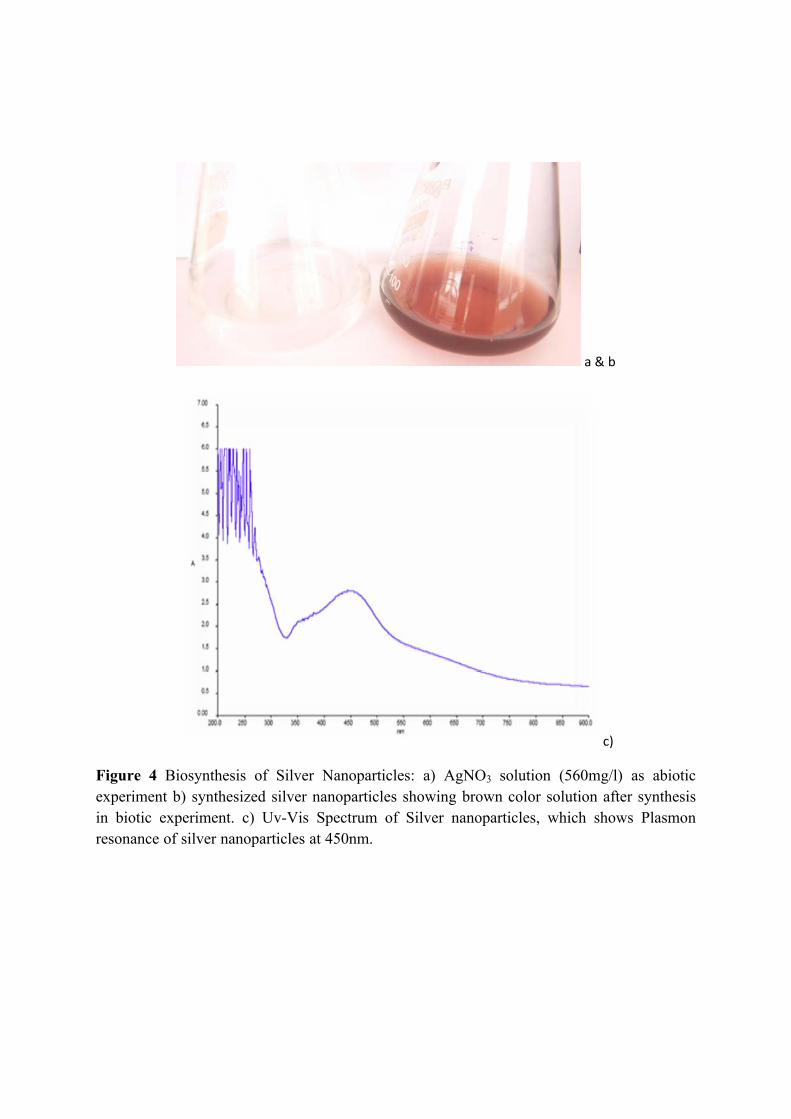
a & b
c)
Figure 4 Biosynthesis of Silver Nanoparticles: a) AgNO3 solution (560mg/l) as abiotic experiment b) synthesized silver nanoparticles showing brown color solution after synthesis in biotic experiment. c) Uv-Vis Spectrum of Silver nanoparticles, which shows Plasmon resonance of silver nanoparticles at 450nm.
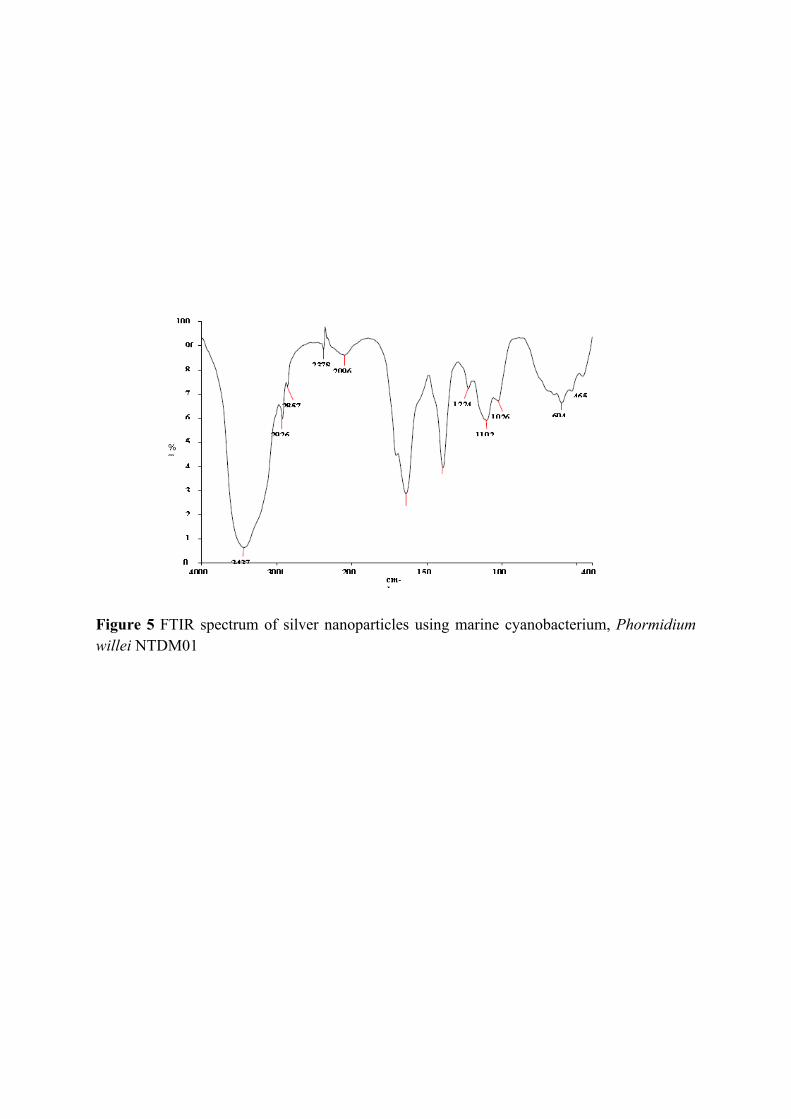
Figure 5 FTIR spectrum of silver nanoparticles using marine cyanobacterium, Phormidium willei NTDM01

108
200nm (extracellularly) were also precipitated in solution silver nanoparticles were
deposited at cell surfaces. EDS showed the occurrence of silver particle in higher
amount with trace of magnesium, calcium and chloride. In this analysis by EDS of the
silver nanoparticles was confirmed the presence of elemental silver signal (Fig.6b).
TEM micrograph of a thin section of cyanobacteria showed nanoparticles of
silver deposited inside the cell in spherical shaped nanoparticles size is about 20 nm.
Octahedral, spherical, and anhedral nanoparticles of silver precipitated in solution (Fig.
7).
The reaction of cyanobacteria with AgNO3 solutions results in the formation of
silver nanoparticles. The presence of spherical silver nanoparticles was observed in
experiment. The precipitation of silver nanoparticles was not observed for abiotic
experiment that were run under similar condition and duration, that suggesting that
cyanobacteria were required for silver precipitation and the precipitation processes
were not controlled by inorganic chemical reactions.
A brief overview of the use of micro organisms such as cyanobacteria, yeast,
algae, fungi and actinomycetes in the biosynthesis of metal nanoparticles has been
described. In the present study, spectrum, UV-vis observation indicated the release of
protein into the culture medium indicting the electronic excitation of aromatic amino
acids. A similar observation has been made in Fusarium oxysporum and suggests a
possible mechanism for the reduction of the metal ions present in the solution. The
overall peaks from FTIR observation confirm the presence of protein in the samples of
silver nanoparticles. It can be assumed that nitrate reducing bacteria to produce nitrate
reductase enzyme (protein). This reduces to silver nitrate solution form silver
nanoparticles. The synthesis of silver nanoparticles using ß-NADPH-dependent nitrate
reductase and phytochelatin in vitro has been demonstrated (Shanker, 2003).
In this study, cyanobacterial cells were encrusted with silver nanoparticles and
separation of some filaments into their constituent cells were still observed. The
nanoparticles were not in direct contact even within the aggregates, indicating
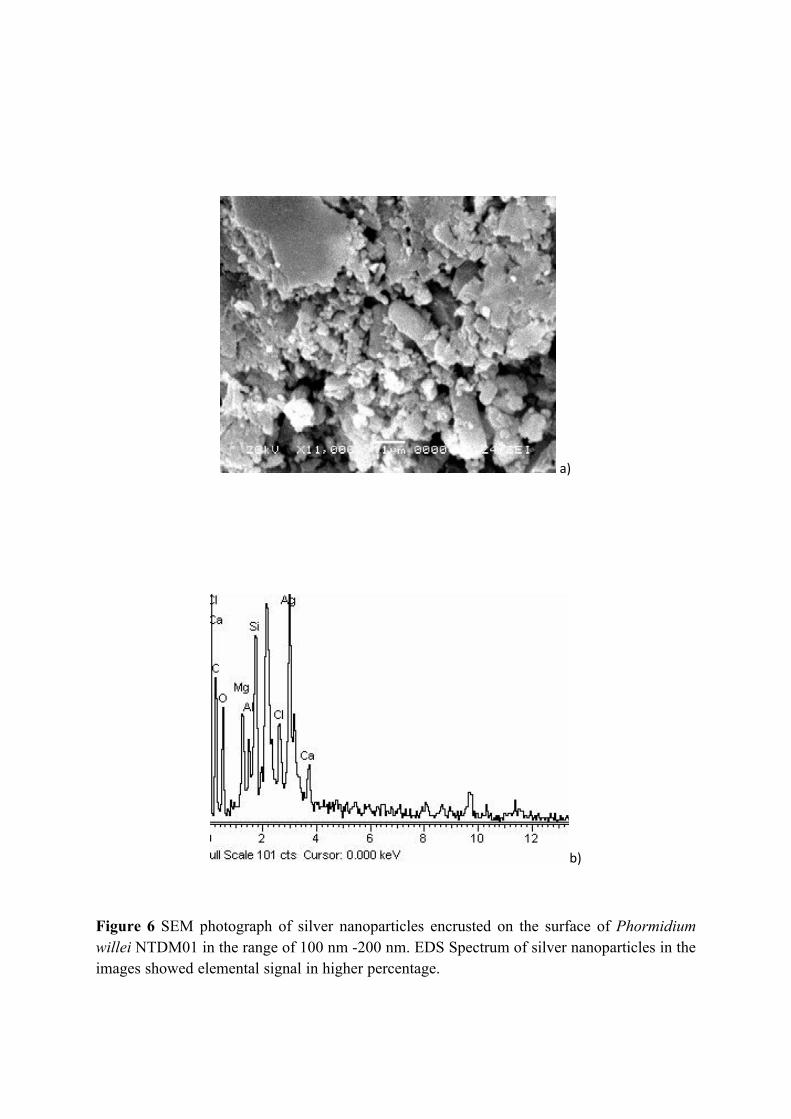
a)
b)
Figure 6 SEM photograph of silver nanoparticles encrusted on the surface of Phormidium willei NTDM01 in the range of 100 nm -200 nm. EDS Spectrum of silver nanoparticles in the images showed elemental signal in higher percentage.
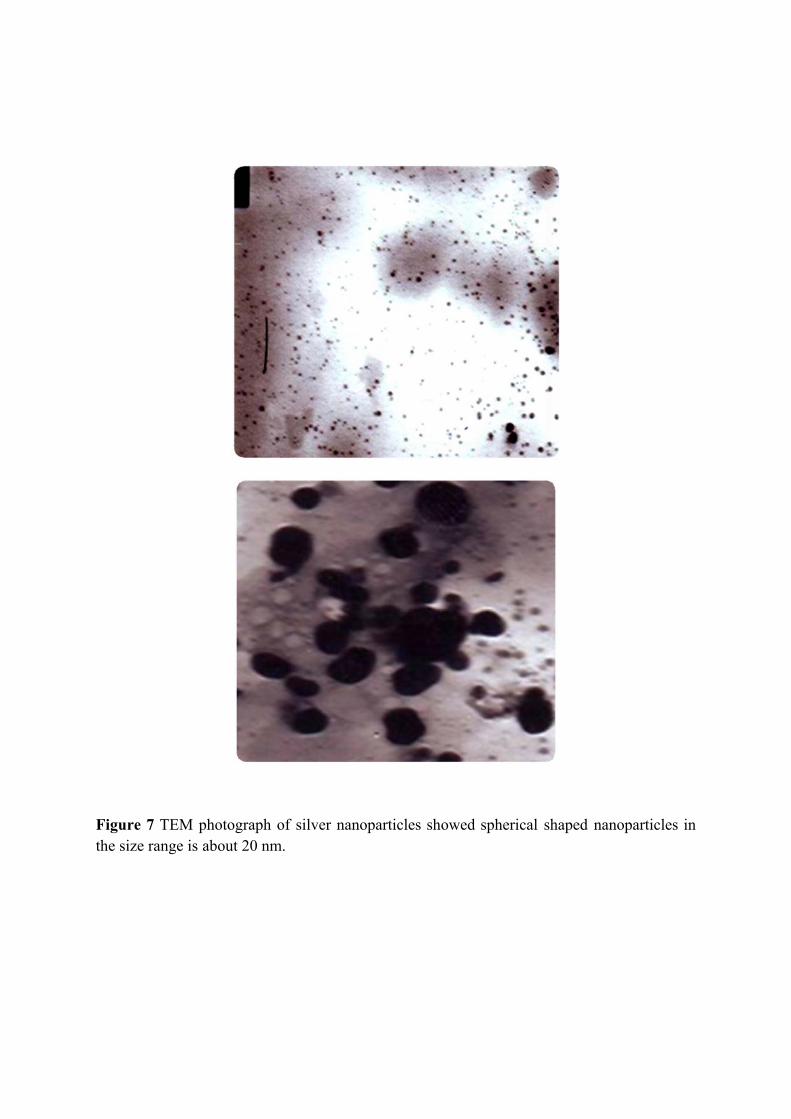
Figure 7 TEM photograph of silver nanoparticles showed spherical shaped nanoparticles in the size range is about 20 nm.

109
stabilization of the nanoparticles by capping agent. This corroborates with the previous
observation in F. oxysporum (Shanker, 2003). Cyanobacteria commonly use nitrate as
the major source of nitrogen for three different purposes including growth, generation
of metabolic energy, and redox balancing (Suzuki et al., 1995). The presence of silver
nanoparticles scattered throughout the cells suggests that Ag+ and NO3 entered in the
cyanobacterial cells through a transport system and degraded in solution. The presence
of silver nanoparticles inside the cytoplasm, Ag+ is presumably reduced to Ago Because
AgNO3, a toxic reagent, was used in metabolic processes, it ultimately killed the cells.
During the death of cyanobacteria, nanoparticles of silver produced inside the
cells were release through the cell membrane into solution, as indicated by the
precipitation of silver nanoparticles around the cells. The dead cyanobacteria also
released organics (protein, and other biochemical) that caused further precipitation of
silver from solution outside the cells. The protein molecules act as reducing agent for
silver nanoparticles. The protein molecule made up of different functional group in
amino acid sequences such as amino, carboxyl, sulfate groups present in the
cyanobacterial protein favors the formation of extremely small-sized silver
nanoparticles with narrow size distribution. Hydroxyl and sulfonic groups are
beneficial to synthesis of silver nanoparticle with a slightly larger particle size in a
weak reducing environment. It has already been reported that the silver nanoparticles
can be stabilized by functional groups, such as carboxyl, hydroxyl, and amido and thiol
group, except the steric effect arising from the large molecular structures of the organic
modifier (protein) on the surface of the particles (Temgire, 2004 and Vigneshwaran et
al., 2007).
The microorganisms used in the synthesis of nanomaterials as a possible viable
alternative them, the more popular physical and chemical methods. The use of
cyanobacteria as a source of enzymes that can catalyze specific reactions leading to
inorganic nanoparticles is a new and rational biosynthesis strategy that is being
developed. Extracellular secretion of enzymes offers the advantages of obtaining large
quantities in a relatively pure-state, free from other cellular proteins associated with the

110
organism and can be easily processed by filtering of the cells and isolating the enzyme
for nanoparticles synthesis from cell-free filtrate.
Gold nanoparticles:
Synthesis of gold nanoparticles was confirmed by changes of pale yellow color
chloroauric acid into ruby red colored solution, which indicates that formation of gold
nanoparticles by the action of cyanobacterial proteins present in the reaction vessel
(Fig. 9a). The UV-Vis spectrum reveals that the band observed at 540nm due to the
plasmon resonance of the gold nanoparticles and low wavelength region recorded from
the reaction medium exhibited an absorption band at ca.265nm and it was attributed to
aromatic amino acids of proteins. It is well known that the absorption band at ca 265nm
arises due to electronic excitation in tryptophan and tyrosine residue in the protein. It is
also gave an idea for the synthesized nanoparticles due to the action of protein that
released from cyanobacteria and the nanoparticles surrounded by the extracellular or
intracellular protein (Fig. 9b).
In this study, cyanobacterial and chlorophycean biomass treated with 1mM of
HAuCl4 solution started to turn purple within 72 h of exposure, in contrast to the
control sets which remained the same (Fig. 8a & b). The UV-visible spectra of the
extracted purple solutions of gold nanoparticles are shown in Fig 9. It showed
absorbance at about 525 nm, it was reported that the UV–visible spectroscopy of
nanoparticle solutions obtained from P. tenue and Microcoleus biomass. The
characteristic absorption bands at around 530nm at all pHs. In the case of P. tenue,
maximum absorption was observed at 529nm (pH 5 and pH 7) or 532nm (pH 9),
whereas Microcoleus showed absorption maxima at 534nm (pH 5 and pH 7) and at
530nm (pH 9). Ulva showed absorbance peaks at 542 nm, 541nm and 542nm at pHs 5,
7 and 9, respectively and Rhizoclonium showed prominent peaks at 537 nm, 529nm and
517 nm. No absorption band beyond 600nm was visible in any of these cases (Parial et
al., 2012).
In this study, FTIR spectrums of gold nanoparticles were performed, in which
after the experimentation; powdered samples was analyzed for FTIR spectrum to gold
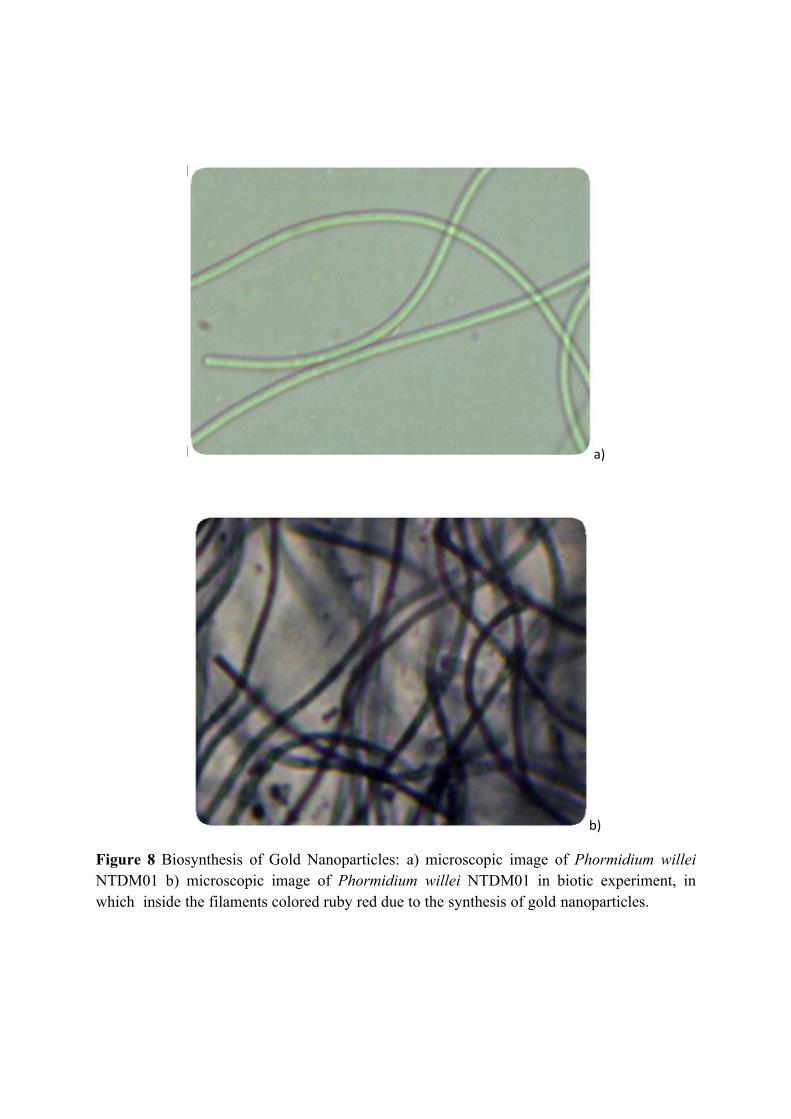
a)
b)
Figure 8 Biosynthesis of Gold Nanoparticles: a) microscopic image of Phormidium willei NTDM01 b) microscopic image of Phormidium willei NTDM01 in biotic experiment, in which inside the filaments colored ruby red due to the synthesis of gold nanoparticles.
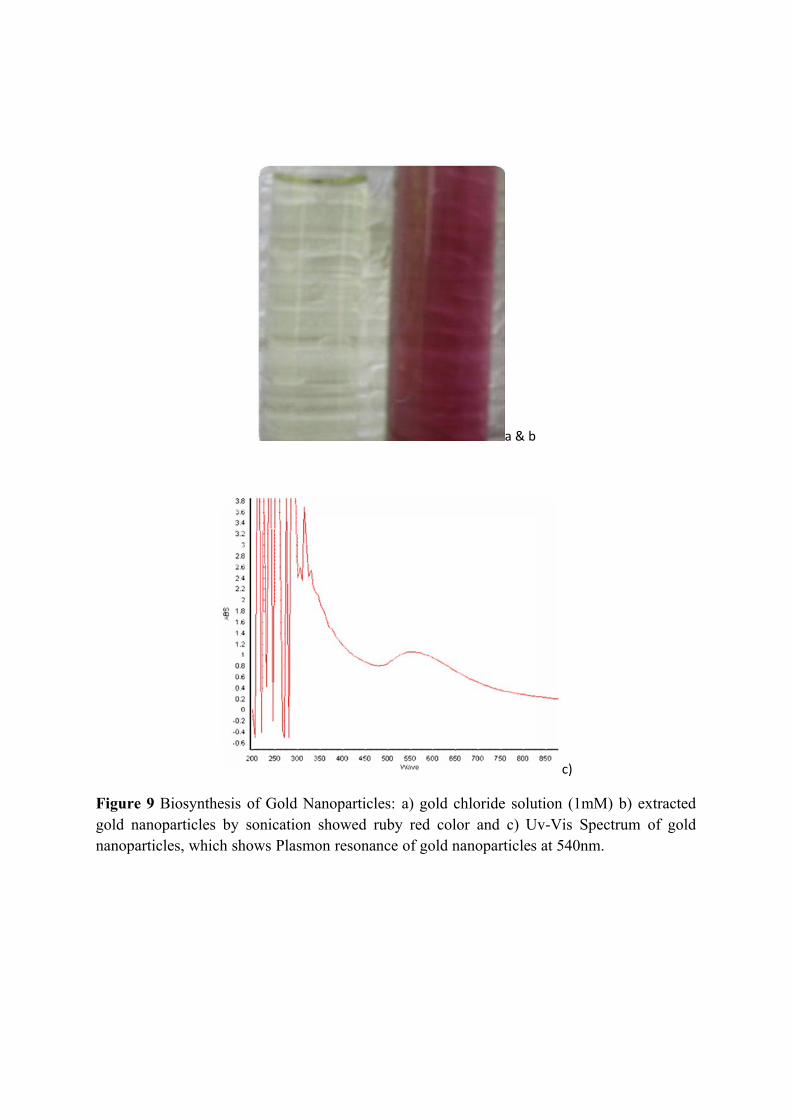
a & b
c)
Figure 9 Biosynthesis of Gold Nanoparticles: a) gold chloride solution (1mM) b) extracted gold nanoparticles by sonication showed ruby red color and c) Uv-Vis Spectrum of gold nanoparticles, which shows Plasmon resonance of gold nanoparticles at 540nm.
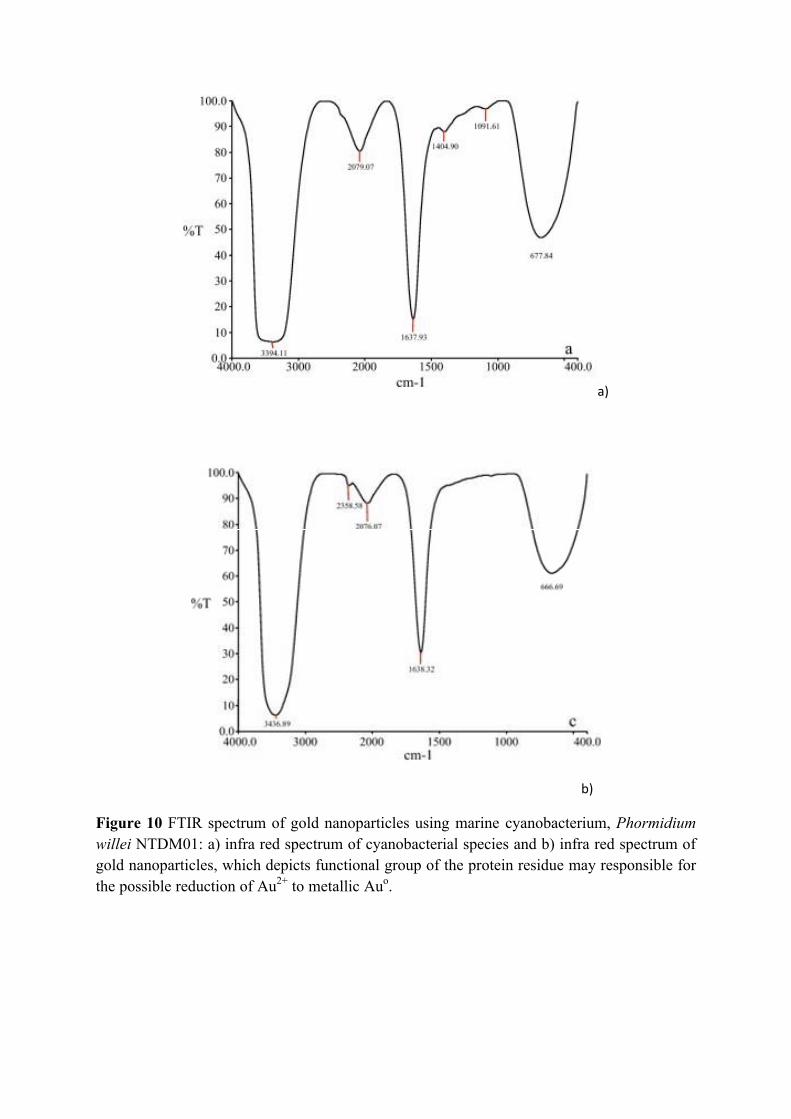
a)
b)
Figure 10 FTIR spectrum of gold nanoparticles using marine cyanobacterium, Phormidium willei NTDM01: a) infra red spectrum of cyanobacterial species and b) infra red spectrum of gold nanoparticles, which depicts functional group of the protein residue may responsible for the possible reduction of Au2+ to metallic Auo.

111
nanoparticles. The peaks were observed at 3394.11 cm-1, 2079.07 cm-1, 1607.93 cm-1,
1404.90 cm-1, 1091.61 cm-1 and 677.84 cm-1 in pure cyanobacterial sample. Whereas in
gold nanoparticles experiments, the peak was observed at 3436.89 cm-1, 2358.58 cm-1,
2076.07 cm-1, 1638.32 cm-1, 666.69 cm-1. The presence and absence of peaks and
formation of peaks, which reveals that chemical modification was happened in order to
formation of nanoparticles by the action of cyanobacterial protein and the metallic
nanoparticles were combined with functional group of the protein and stabilized the
synthesized nanoparticles (Fig. 10a & b).
The results of HRSEM of gold nanoparticles showed that spherical shaped
uniform sizes of nanoparticles were observed. The size of the particles was found to be
10 nm. The gold nanoparticles encrusted on surface of the cyanobacterial cells. EDAX
peaks also confirmed that presence of gold signal in the sample in higher percentage
(Fig. 11a & b). The results obtained from TEM gave clear indication about the size and
shape of the nanoparticles. The solution extracted from gold-loaded biomass of P.
willei treated showed the presence of spherical particles of 10 nm diameter, along with
nanotriangles of approximately 15 nm (Fig. 12). It has already been reported that the
colour of gold nanoparticles can vary from red to blue, depending upon the shape and
size of the particles (Sharma et al., 2009).
The gold nanoparticles solutions extracted in the present study were purple,
because of the excitation of surface plasmon vibrations in the gold nanoparticles
(Mulvaney, 1996). After Au (III) exposure, the purple, ruby red or blue colorations of
microorganisms are due to the reduction of Au (III) to Au (0) and subsequent formation
of Au nanoparticles intra- and/or extracellularly (Druff et al., 1987; Chow & Zukoski,
1994; Lujan et al., 1994; Chakraborty et al., 2009; Parial et al., 2012). The present
study showed that the Phormidium willei NTDM01 have potential as bionano-
factories, and this is evident from the visual transformation of the cyanobacteria from
green to deep purple. The biosynthesized gold nanoparticles were extracted
successfully by sonicating the gold loaded biomass of the experimental taxa.

112
It is highly desirable to synthesize nanorods in a clean and non-toxic way,
because a toxic chemical CTAB (cetyl trimethylammonium bromide) is used in most of
the conventional chemical methods of nanorod synthesis (Johnson et al., 2002); this
chemical could lead to undesirable functional aberrations in target cells, when
nanoparticles are used in photothermal therapy or in a drug delivery system. CTAB
helps synthesis in two ways: firstly, it can give a directional template promoting
formation of rods and secondly, it is attached to the surface of the nanoparticles and
gives the nanoparticles colloidal stability in solution. Therefore, complete wash-out of
CTAB from the surface of nanoparticles results in the loss of colloidal stability. It has
been recently demonstrated a green chemical approach for the controlled biosynthesis
of gold nanorods by Nostoc ellipsosporum without any shape anisotropy (Parial et al.,
2012).
The surface plasmon band for gold nanoparticles usually has a range of 510–
560nm in aqueous solutions, depending on the nanoparticle morphology. Non-spherical
gold nanoparticles exhibit multiple absorption bands, correlated with their multiple
axes, in comparison to the single one for isodiametric particles. In the present study two
discrete surface plasmon bands were observed in the case of P. willei indicating
possible synthesis of non-spherical gold nanoparticles and it was later confirmed that
these were nanotriangles by TEM study.
Other studies have also indicated that pH plays a key role in controlling particle
morphology. Brown et al. (2000) reported changes in the shape of gold crystals with
changes in pH while studying the role of polypeptides on the resulting gold crystals.
Gericke & Pinches (2006) observed that the pH influences the reduction rate which
regulates the synthesis of different shapes and sizes of gold nanoparticles and further
suggested that spherical particle formation is favoured at low reduction rates whereas
high reduction rates resulted in the formation of nanorods and platelets.
The present study, suggests that a high reduction rate at lower pH may lead to
the formation of nanotriangles. TEM analysis of extracted nanoparticle solutions of P.
willei showed the production of mostly spherical particles. In algae, both processes
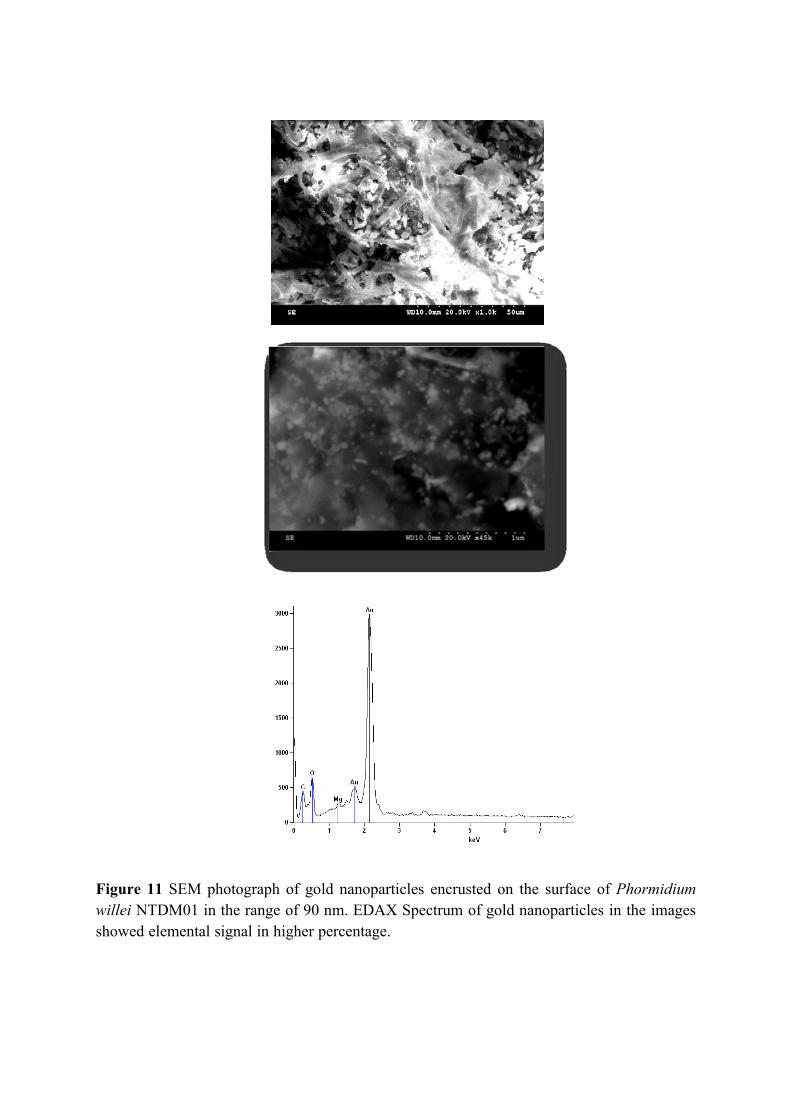
Figure 11 SEM photograph of gold nanoparticles encrusted on the surface of Phormidium willei NTDM01 in the range of 90 nm. EDAX Spectrum of gold nanoparticles in the images showed elemental signal in higher percentage.

Figure 12 TEM photograph of gold nanoparticles showed different shaped such as spherical (5 nm), hexagonal (10 nm) and triangles (15 nm).

113
occur, as gold nanoparticles are recorded either intracellularly or extracellularly in the
medium, as in Lyngbya majuscule and Spirulina subsalsa (Chakraborty et al., 2009).
Interestingly, it has been observed that gold solution incubated with dried,
powdered algal biomass did not turn purple over time (one week), which confirms that
the reduction of gold is associated with cellular metabolism and presumably involves
reducing enzymes or synthesis of other metabolites. In the experiment, cyanobacterial
cells were poisoned and died after converting Au (III) to Au (0). In conclusion, it seems
that synthesis of fairly mono-dispersed gold nanoparticles by the reaction of aqueous
chloroaurate ions with cyanobacterial biomass represents a more eco-friendly and cost-
effective method than the conventional approach using toxic chemicals. The aqueous
gold(III)-chloride solutions at different concentrations investigated were stable for one
week in abiotic experiments (without the presence of cyanobacteria) under similar
conditions.
On the addition of HAuCl4 to the cyanobacteria, precipitation of nanoparticles
(<20 nm) containing gold was observed by HRSEM-EDS at the cell surfaces and at the
interface between the cell wall and solution within several minutes. These nanoparticles
were amorphous to electron diffraction. After 17 h, the dead cyanobacteria released
cytoplasmic material around the cells, causing further precipitation of metallic gold
nanoparticles, with octahedral and sub-octahedral shapes and diameters up to 6 nm. A
tendency for the frequency of larger octahedral platelets to increase with run duration
indicated a weak Ostwald ripening effect (Parial et al., 2012).
Gold concentrations in river and marine waters range up to about 50 ppb, while
near gold deposits; soluble gold concentrations can be up to 1 ppm (Korobushkina et
al., 1983 and McHugh, 1988). Thus, the gold concentrations in natural environments
are 2-3 orders of magnitude lower than those used in this study. This study showed that
the gold reduction process from HAuCl4 to metallic gold by cyanobacteria was
facilitated by higher gold concentrations. In natural environments where gold
concentrations were relatively low, the reduction of HAuCl4 to metallic gold by
cyanobacteria is presumably a slow process. In addition, at low gold concentrations (at

114
ppb level), gold toxicity would not kill all viable bacterial cells; therefore, the
remaining cells would be able to grow and probably adapt to the presence of gold
(Reith, 2006). Passive gold accumulation via organic sulfur present in the
cyanobacterial outer membrane or periplasma may explain the mechanisms of gold
bioaccumulation by cyanobacteria.
The recovery of gold from solutions has been actively studied for industrial
purposes (Nakajima et al., 2003; Pethkar, et al., 1998 and Romero-Gonzalez, 2003).
Mining, mineral processing, and metallurgical industries generate billions of tons of
metal waste every year (Eccles, 1995). Conventional methods for removing metals
from ore processing solutions, including chemical precipitation, chemical oxidation or
reduction, ion exchange, filtration, electrochemical treatment, membrane technologies,
and evaporation, have been used for gold recovery (Volesky, 1990). Because these
processes are extremely expensive, alternative methods for metal recovery using living
or dead microorganisms have been actively investigated. The present study indicate that
the interaction of HAuCl4 with cyanobacteria can be used as a promising method for
recovery of gold particles from ores.
Biosynthesis of Semiconductor:
CdS nanoparticles:
In the earlier study, cytotoxicity of C-PE was evaluated in human cell lines by
assessment of cell proliferation and neutral red uptake; it was found that the C-PE is not
cytotoxic (Soni et al., 2010). Among various concentration of CdCl2 and Na2S, CaCl2
at 0.25mM and Na2S at 1mM proved to be most optimum concentration for the
synthesis of CdS nanoparticles. The first step in this reaction sequence in the
nanoparticles synthesis was formation of C-PE- Cd2+ complex, then this complex
reacted with Na2S to produce CdS nanoparticles, when the color reaction mixture
changed to yellow to orange (Fig. 13a). CdS nanoparticles thus synthesized measured
about 5 nm and, this size remains unchanged well over 8 months of storage. In an
earlier investigation using R-PE, the solution was contains only coarse CdS
nanoparticles.
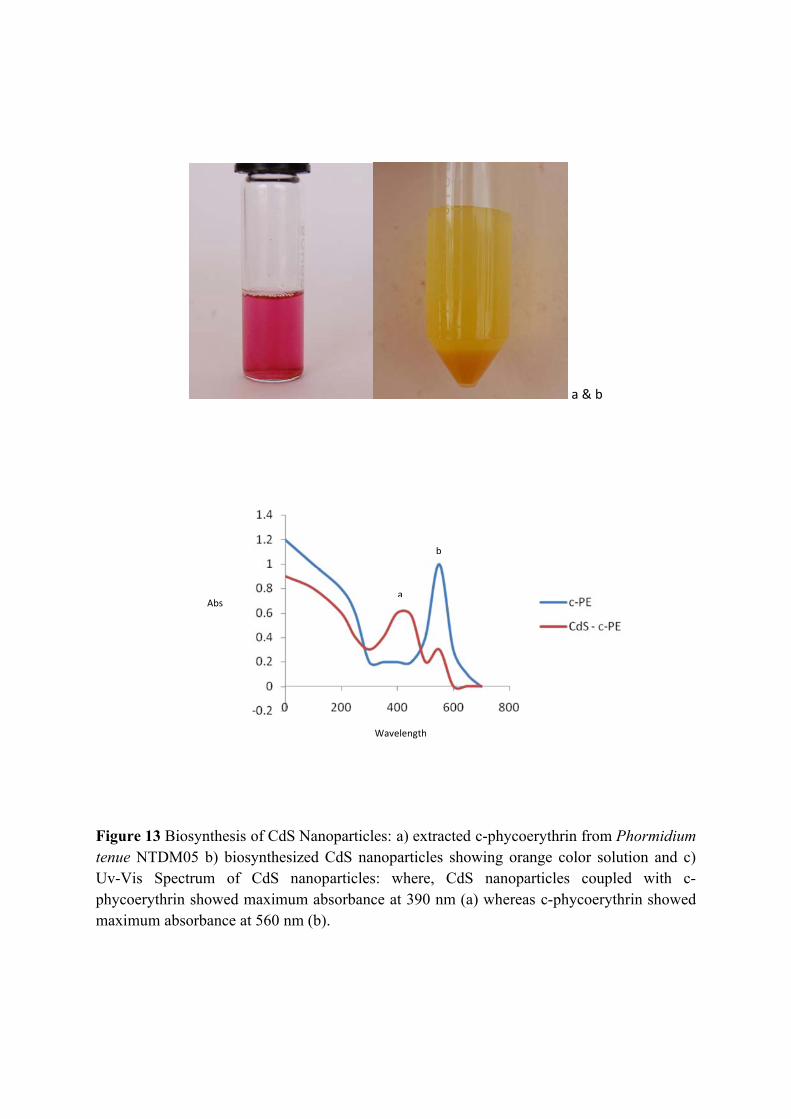
a & b
Figure 13 Biosynthesis of CdS Nanoparticles: a) extracted c-phycoerythrin from Phormidium tenue NTDM05 b) biosynthesized CdS nanoparticles showing orange color solution and c) Uv-Vis Spectrum of CdS nanoparticles: where, CdS nanoparticles coupled with c-phycoerythrin showed maximum absorbance at 390 nm (a) whereas c-phycoerythrin showed maximum absorbance at 560 nm (b).
Abs
Wavelength
a
b

115
It also found that R-PE prevents the growth and aggregation of nanoparticles
(Brekhovskikh and Bekasova., 2005). A similar experiment in higher pH the charged
capping agent found to be very high, which might be the basic of the colloidal stability
of QDs. Also, the size of the Cd-R-PE complex was primarily depending on the
concentration of salts, pH of the reaction mix and the concentration of the pigments.
(Liji Shobhana, et al., 2011).
Soluble starch has also been also used as a capping agent in the synthesis of
CdS and CdSe nanoparticles, ranging from 8-15nm (Wei et al., 2004 and Li et al.,
2007). The hydroxyl group of the starch polymer acts as passivation contacts for the
stabilization of the CdS nanoparticles in aqueous solution (Rodriguez, 2008). The
synthesis of CdS nanoparticles occurs due to pH sensitive membrane-bound oxido-
reductase and carbon source-dependent rH2 in the culture solution (Prasad et al., 2010).
This work also emphasizes that the fluorescent protein in the pigment is mainly due to
tyrosine, phenylalanine and tryptophan moieties, which would be the basis of the
formation of CdS nanoparticles very efficiently and stabilizing it to prevent
agglomeration.
The presence of CdS nanoparticles was confirmed by the absorption peak at
470nm. The maximum absorption peak position of hollow nanoparticle chains is at
487nm with the band gap at 2.56 eV (Liu et al., 2007). Compared with that of bulk
material (515nm, 2.42 eV), the absorption peak was a blue shift at 28 nm and its band
gap enlarged about 0.14eV. This is indication of quantum size effect (Gao et al., 2002).
It was reported that the band gap value 2.57 eV is shifted compared with the bulk value
and this could be a consequence of a size quantization effect in the sample (Castanon et
al., 2005) and the absorption peak of c-PE was 550 nm (Fig. 13b).
The FTIR spectrum of the c- phycoerthrin is shown in Fig. 14a. The
characteristic bands due to S-H stretching, appearing at 3398, 2074, 1637, 1365 cm-1
for pure C-PE were not seen in capped CdS, which is due to the interaction of thiol
group of the capping agent with CdS. The characteristics band such as S-H stretching
was found in the entire capping agent that was not found in all capped CdS. It could be
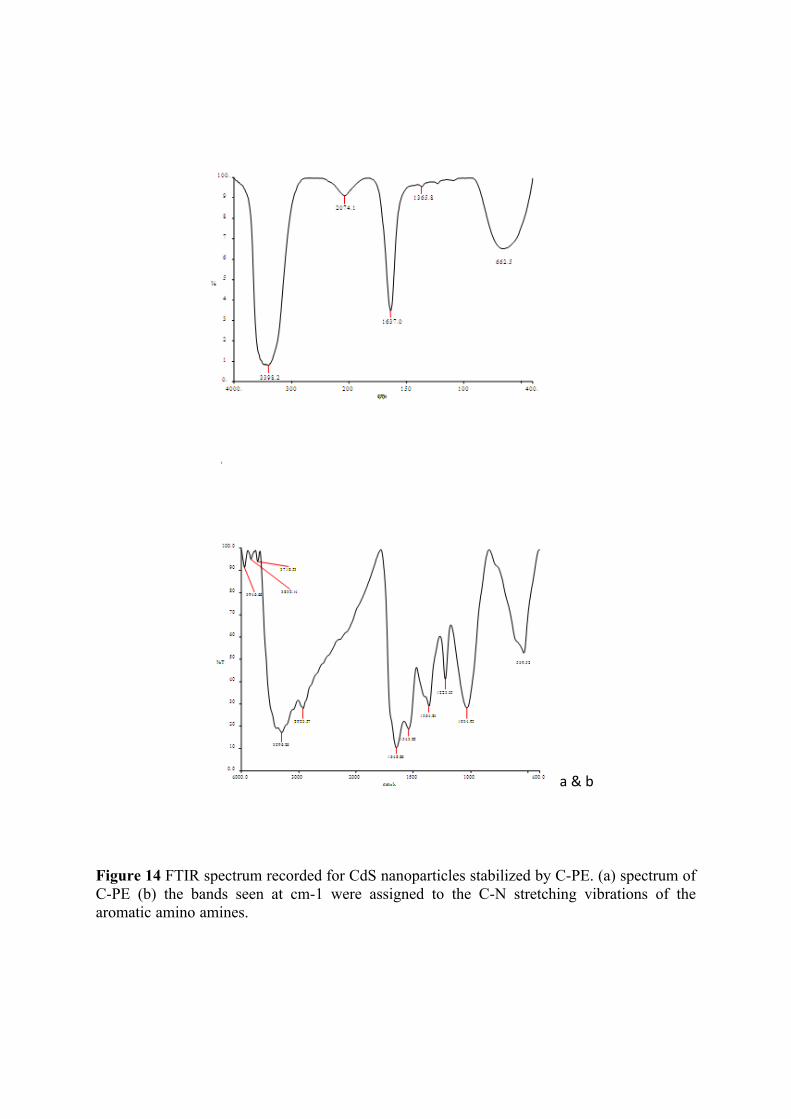
a & b
Figure 14 FTIR spectrum recorded for CdS nanoparticles stabilized by C-PE. (a) spectrum of C-PE (b) the bands seen at cm-1 were assigned to the C-N stretching vibrations of the aromatic amino amines.

Figure 15 HRSEM photograph of CdS nanoparticles encrusted on the surface of Phormidium willei NTDM01 in the range of 5 nm - 30 nm. EDAX Spectrum of CdS nanoparticles in the images showed elemental signal in higher percentage.

116
formed due to bonding with CdS NPs (Liji Shobhana, et al., 2011). In the interaction
between carboxymethyl chitosan (CMCH) and CdS NPs, there were two kinds of
bonding; carboxymethyl group was strongly bound by coordination with Cd ions on to
the particle surface and the unsubstituted other groups were linked to the particle
surface via hydrogen bonding (Li et al., 2003). From this observation, it was suggested
that the biological molecules can possibly perform the function for formation as well as
stabilization of the CdS nanoparticles in aqueous medium (Bai et al., 2009).
The peak at 1543, 1364, 1223 cm-1 appeared only in CdS-CPE complex (Fig.
14b). This new band was not observed in unbound CdS, which indicates a new
vibration from the CPE due to C-S band at another possible scheme of bonding
between CPE and CdS. The interaction mechanism is mainly due to hydrogen bonding.
Generally CdS QDs grow well in a solution that may has a large amount of hydroxyl
groups and in water-bound surface, which are active sites for the formation of hydrogen
bonding with the surrounding molecules (Ladizhansky, et al., 1998).
The morphology and size of the CdS nanoparticles were observed adopting
HRSEM. It was found that CdS nanoparticles encrusted on the surface of the cell in the
range of 5 nm to 30nm (Fig 15). The morphology and size of the CdS nanoparticles
were observed adopting TEM. It was found that the nanoparticles were of uniform size,
at about 5 nm. The EDAX revealed the presence of both Cd and S in the nanoparticles
synthesized (Fig. 16). The lifetime of capped CdS QDs was higher than that of
uncapped CdS QDs. The results also indicated that the lifetime of the particles
increased with increase in size, which may be due to the uniform passivation caused
MSA on the surface of CdS particles (Liji Shobhana et al., 2011). Based on the studies
carried out hypothetical mechanism was proposed for the formation of CdS
nanoparticles by the action of functional amino acids present in the C-Phycoerythrin
protein extracted from cyanobacteria (Fig. 17).
In the present study the particle size was 5.1±0.2nm. This size range may
possibly be due to the formation of nanoparticles over a time period. In support of this
inference, an earlier report also found difference in size of the particles over the
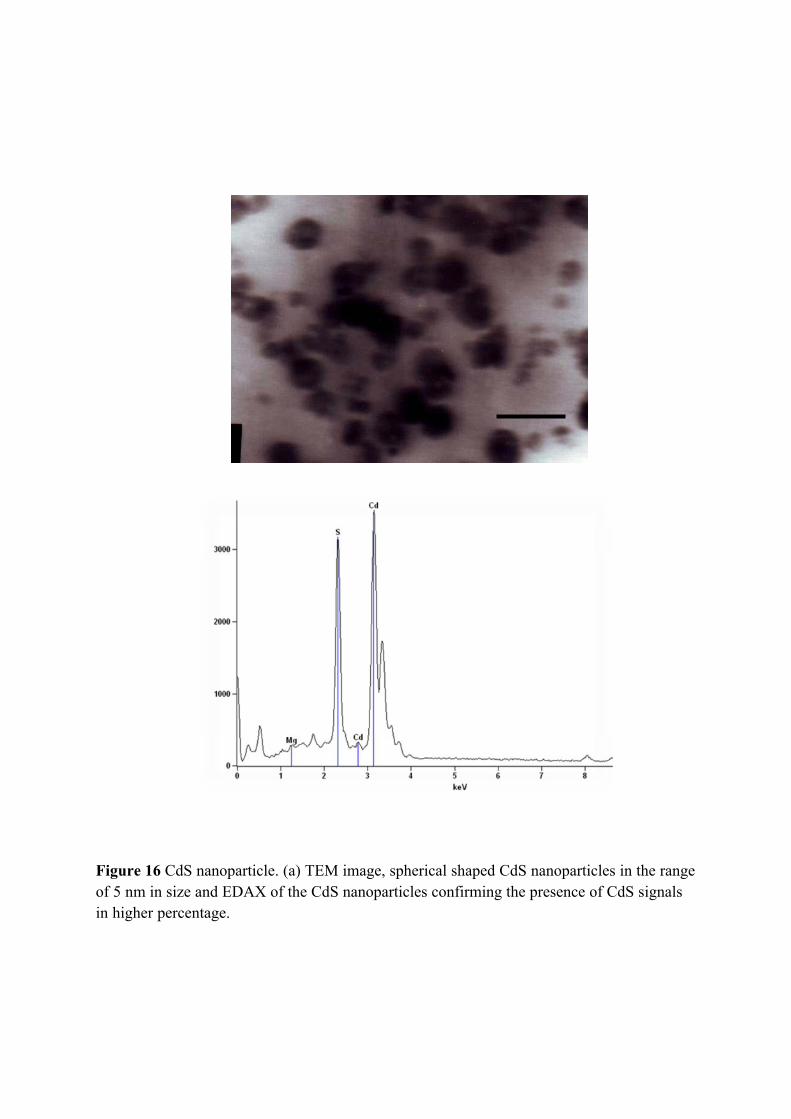
Figure 16 CdS nanoparticle. (a) TEM image, spherical shaped CdS nanoparticles in the range of 5 nm in size and EDAX of the CdS nanoparticles confirming the presence of CdS signals in higher percentage.
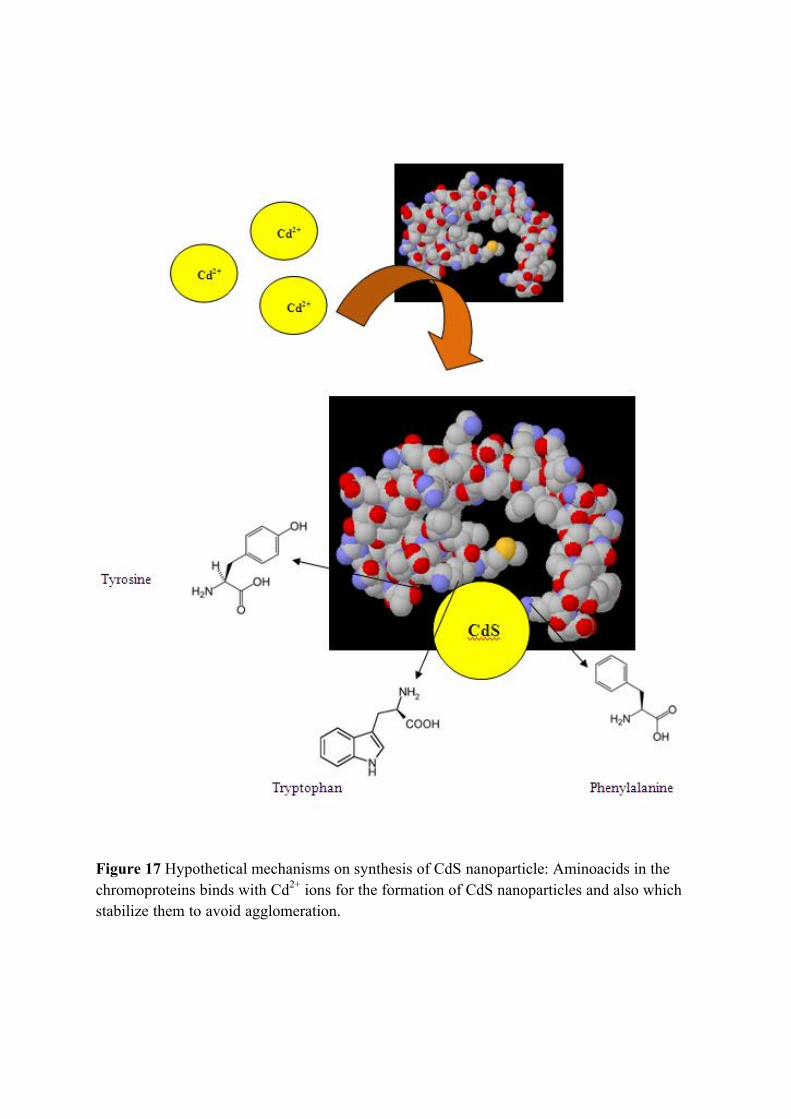
Figure 17 Hypothetical mechanisms on synthesis of CdS nanoparticle: Aminoacids in the chromoproteins binds with Cd2+ ions for the formation of CdS nanoparticles and also which stabilize them to avoid agglomeration.

117
incubation time (Prasad et al., 2010). Since CdS nanoparticles, synthesized in aqueous
phase, could be a novel fluorescence probe for ultrasensitive detection of DNA as a
biolabel (Wang et al., 2010), the enhanced synthesis of CdS nanoparticles using C-PE,
are the unique properties of the particles might prove to the particles that prove to be of
great advantage.
Biosynthesis of Nanocomposits:
Si-Ge Oxide Nanocomposites:
A diatom, Stauroneis sp. culture was treated with germanium in two different
ways and they were periodically monitored for structural changes in their frustules
morphology. The pH was also monitored and changes were not observed. The
fabrications of germanium in diatom frustules were examined by electron microscopy
characterization.
The ability of diatoms to make complex, nanoscaled, three-dimensional silica
shells called “frustules” offers attractive possibilities for their applications to
nanobiotechnology (Hildebrand, 2005) and synthesis of new materials. For example,
silicon in the shell can be replaced with other elements by chemical processes while
preserving the intricate patterns of the frustule (Sandhage et al., 2002).
Scanning Electron Microscopy (SEM) and Energy Dispersive X ray
Spectroscopy (EDS) was used to study the integrity of the sample. SEM was to study
the surface morphology of the sample and EDS was used to study the structural
properties and chemical composition of the diatom frustules. Here the EDS spectrum
scan was used to locate and determine the distribution of Si-Ge nanocomposites in the
frustules.
The microscopy results showed that the Stauroneis sp. grown in an enriched sea
water medium, normal growth different view of the diatoms was documented for this
study (Fig. 18). The microscopy results showed that the Stauroneis sp. grown in an
enriched sea water medium along with germanium and the same culture that grown in a
silica and germanium alone did not showed any structural or morphological changes
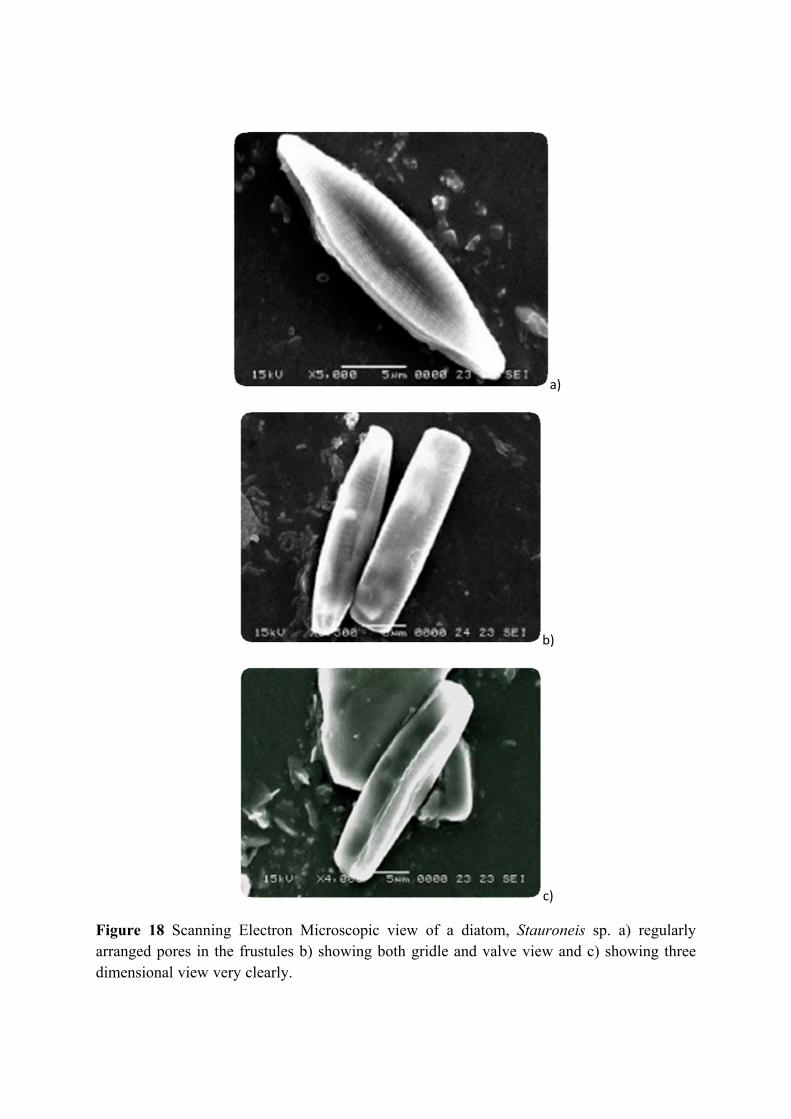
a)
b)
c)
Figure 18 Scanning Electron Microscopic view of a diatom, Stauroneis sp. a) regularly arranged pores in the frustules b) showing both gridle and valve view and c) showing three dimensional view very clearly.

118
(Fig. 19). In Stauroneis, the cultures that grown in nutrients along with germanium did
not showed any structural changes (Fig. 19a) and the nanostructures such as pores and
raphe were observed clearly on the frustules. But with the same culture that grown in
germanium and silicate alone shows some aberrations in their frustules morphology
(Fig. 19b). The concentration of silicon and germanium used in all those cultures was
0.5mM: 0.1mM.
The incorporation of germanium into the diatom frustules was analysed by EDS
spectrum analysis. Characteristic energy peaks of germanium and silicate were
observed at 1 to 22 keV. In EDS spectrum of Stauroneis that grown in enriched sea
water medium and cultures with germanium and silicate alone, have no incorporation
of germanium and they are not fabricated with germanium and they could not used as
nanocomposite.
The EDS spectrum of Stauroneis culture that grown in enriched seawater
medium along with germanium showed that the germanium was incorporated into the
diatom frustules and the amount was very low and the distribution profile of
germanium in the diatom frustules is similar to the silicate although the intensity was
quite different. The maximum peak was obtained in silicate signal than germanium
signal. The results indicate that the Si-Ge nanocomposites were not homogenously
distributed within the diatom frustules (Fig. 20).
In this present study, scanning electron microscopy and microanalysis
techniques were used to characterize the morphologies and elemental composition of
the processed and unprocessed diatoms. Microscopic results show that the diatom cells
suffered varying degrees of morphological changes, probably from the incorporation of
germanium.
It is also possible that the morphological changes could have been caused by the
availability of insufficient silicon for the normal growth of the structural organs. The
samples with high germanium levels have been found to consist of more broken pieces
of diatom frustules than samples with no germanium. The difference in bond lengths
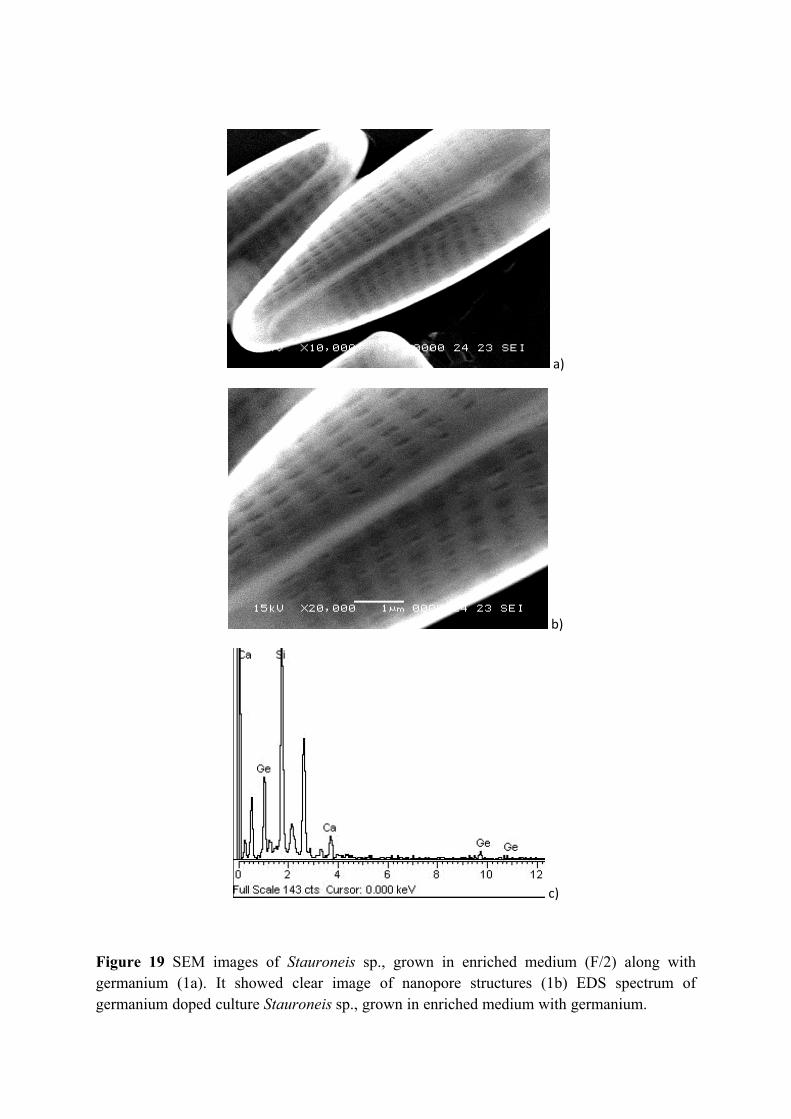
a)
b)
c)
Figure 19 SEM images of Stauroneis sp., grown in enriched medium (F/2) along with germanium (1a). It showed clear image of nanopore structures (1b) EDS spectrum of germanium doped culture Stauroneis sp., grown in enriched medium with germanium.
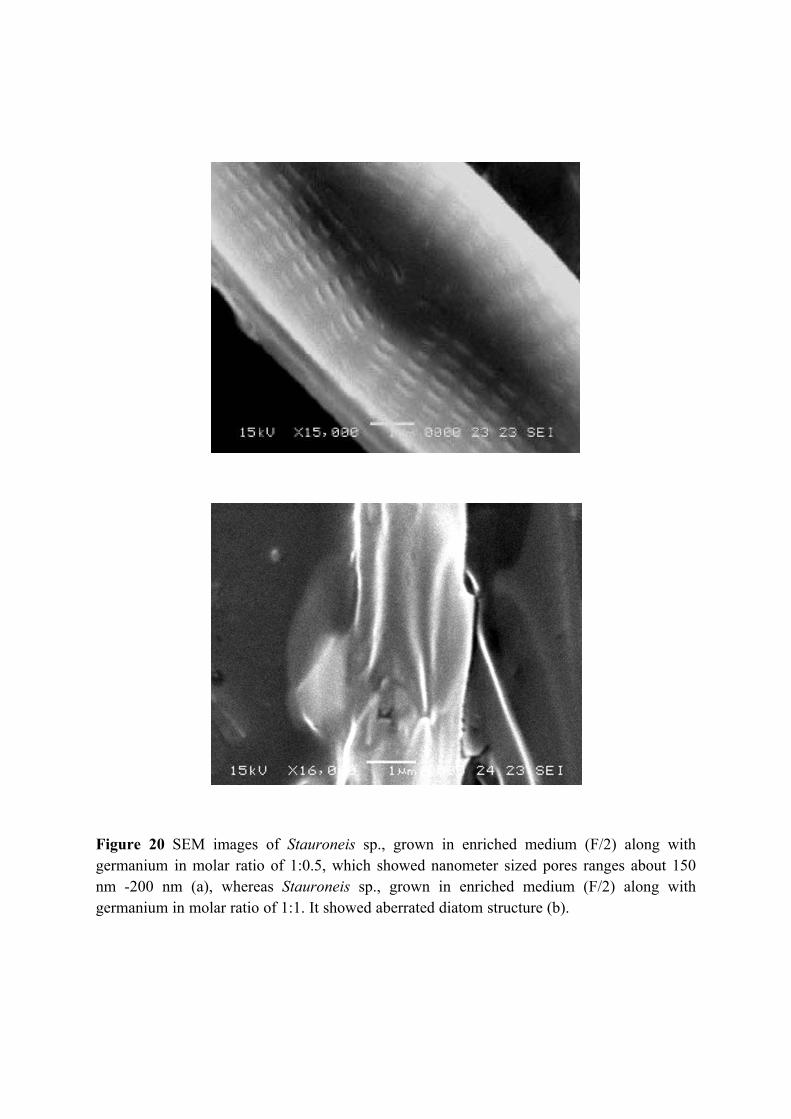
Figure 20 SEM images of Stauroneis sp., grown in enriched medium (F/2) along with germanium in molar ratio of 1:0.5, which showed nanometer sized pores ranges about 150 nm -200 nm (a), whereas Stauroneis sp., grown in enriched medium (F/2) along with germanium in molar ratio of 1:1. It showed aberrated diatom structure (b).

119
between silicon oxide and germanium oxide probably weakens the diatom frustules and
cause them to break easily during the processing of the cells or to collapse when
exposed to a high energy beam of electrons (Gutu and Jiao, 2006).
The results indicate that the Si-Ge nanocomposite was not distributed within the
diatom frustules. This was because the amount of germanium incorporated into the
diatom cells was very low. Experimental parameters such as molar ratio of silicon to
germanium determine amount of germanium assimilated by diatom cells.
Biomedical application:
Antibacterial property against multidrug resistant clinical pathogens:
Synthesized silver nanoparticles active against clinically isolated multi drug
resistant bacterial pathogens and a fungal pathogen namely, Aspergillus flavus. Zone of
inhibition of the isolates were tabulated (Table 8). All isolated pathogens are multi drug
resistant for the 15 antibiotics tested. It was quite interesting that the bacterial and
fungal isolates which were originally sensitive towards synthesized silver nanoparticles.
More specifically, E.coli and Pseudomonas aeruginosa showed good antibacterial
activity after 24 h and 48 h incubation respectively multi drug resistant. In contrast,
Proteus mirabilis and Streptococcus pyogens had antibacterial activity but not like
other organism, which maintain the zone pattern until 24 h and 48 h incubation. The
enhanced antibacterial effects of silver nanoparticles are characterized and also stated
that once inside the cell, nanoparticles would interfere with the bacterial growth
signaling pathway by modulating tyrosine phosphorylation of putative peptides
substrate critical for cell viability and division and the nanoparticles were not in direct
contact even within the aggregates, indicating stabilization of the nanoparticles by a
capping agent (Shrivasatava et al., 2007 and Duran et al., 2005).
Shahverdi et al. (2007) reported that the silver nanoparticles have an
antimicrobial effect on S. aureus and E. coli. The oligodynamic effect of silver has
antibacterial activity against microorganisms. The changes in the local electronic
structure on the surface of the smaller sized particles lead to the enhancement of their
chemical reactivity leading to bactericidal effect (Thiel et al., 2004). It was suggested
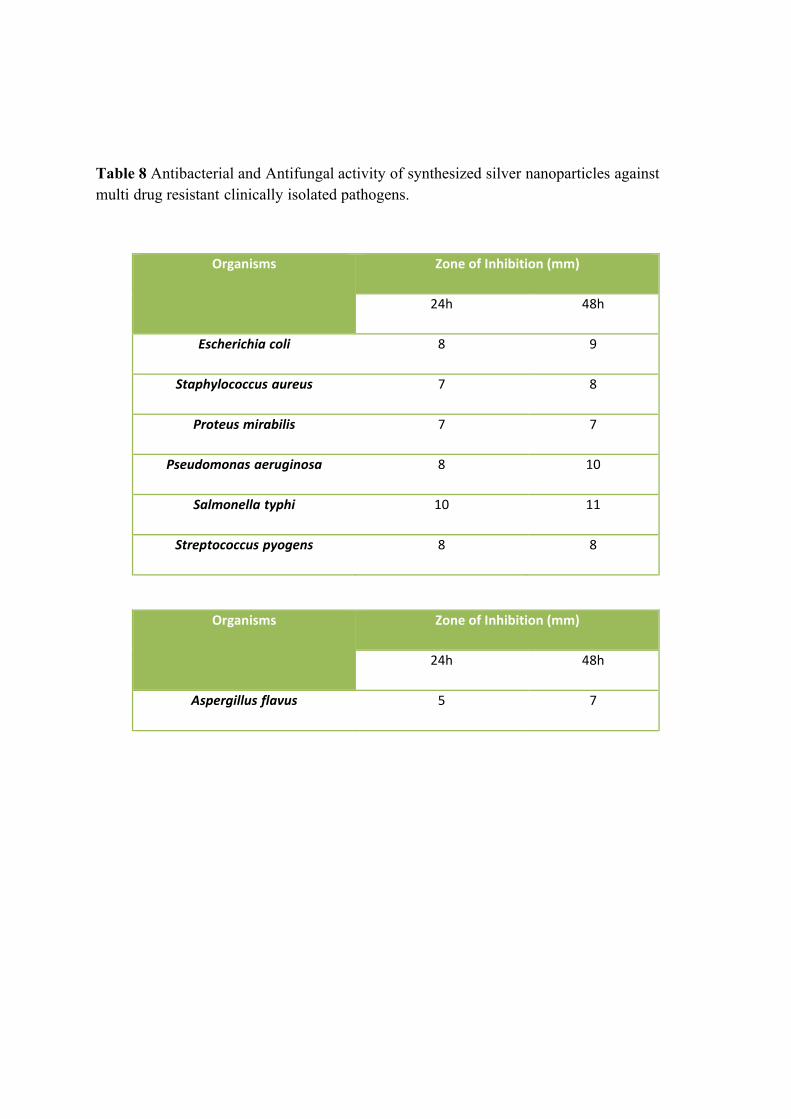
Table 8 Antibacterial and Antifungal activity of synthesized silver nanoparticles against multi drug resistant clinically isolated pathogens.
Organisms Zone of Inhibition (mm)
24h 48h
Escherichia coli 8 9
Staphylococcus aureus 7 8
Proteus mirabilis 7 7
Pseudomonas aeruginosa 8 10
Salmonella typhi 10 11
Streptococcus pyogens 8 8
Organisms Zone of Inhibition (mm)
24h 48h
Aspergillus flavus 5 7

120
that the pre and post washed silver nanoparticles delayed bacteria growth at the
interface of silver nanoparticles as reported by Morones et al. (2005) and direct usage
of the silver nanoparticles from the native reaction solution warranted more effective
(Sondi et al., 2004).
In fact, it is well known that Ag ions and Ag-based compounds are highly toxic
to microorganisms, showing strong biocidal effects on 12 species of bacteria including
E. coli (Zhao, 1998). Recently, it has been showed that hybrids of Ag nanoparticles
with amphiphilic hyperbranched macromolecules exhibited effective antimicrobial
surface coating agents (Aymonier et al., 2002). Reducing the particle size of materials
is an efficient and reliable tool for improving their biocompatibility. In fact,
nanotechnology helps in overcoming the limitations of size and can change the outlook
of the world regarding science (Mirkin, 2000). Furthermore, nanomaterials can be
modified for better efficiency to facilitate their applications in different fields such as
bioscience and medicine.
It is well known that Ag ions and Ag-based compounds have strong
antimicrobial effects (Furno et al., 2004), and many investigators are interested in using
other inorganic nanoparticles as antibacterial agents (Furno et al., 2004; Abuskhuna et
al., 2004 and Hamouda et al., 2000). These inorganic nanoparticles have a distinct
advantage over conventional chemical antimicrobial agents. The most important
problem caused by the chemical antimicrobial agents is multidrug resistance.
Generally, the antimicrobial mechanism of chemical agents depends on the specific
binding with surface and metabolism of agents into the microorganism.
Various microorganisms have evolved drug resistance over many generations.
Thus far, these antimicrobial agents based on chemicals have been effective for
therapy; however, they have been limited to use for medical devices and in prophylaxis
in antimicrobial facilities. Therefore, an alternative way to overcome the drug
resistance of various microorganisms is needed desperately, especially in medical
devices, etc. Ag ions and Ag salts have been used for decades as antimicrobial agents in
various fields because of their growth-inhibitory capacity against microorganisms.

121
Also, many other researchers have tried to measure the activity of metal ions against
microorganisms.
Studies of Russel and Hugo (1994) on the antimicrobial properties of Ag and
Cu and Marsh (2002) on Zn were reported. However, Ag ions or salts has only limited
usefulness as an antimicrobial agent for several reasons, including the interfering
effects of salts and the antimicrobial mechanism of [the continuous release of enough
concentration of Ag ion from the metal form.] In contrast, these kinds of limitations can
be overcome by the use of Ag nanoparticles. However, to use Ag in various fields
against microorganisms, it is essential to prepare the Ag with cost-effective methods
and to know the mechanism of the antimicrobial effect. Besides, it is important to
enhance the antimicrobial effect.
When compares to the chemical and physical synthesis, biological synthesis of
Ag nanoparticles can be prepared cost effectively and thus synthesized. These Ag
nanoparticles are homogeneous and stable. The nanosize allowed expansion of the
contact surface of Ag with the microorganisms, and this nanoscale has applicability for
medical devices by surface coating agents. In this study, to evaluate the antimicrobial
effects against various microorganisms, we used seven representative microorganisms,
E. coli and S. aureus. P.mirabilis, P. aeruginosa, S. typhi and S. pyogens. There were
distinct differences among them. When Ag nanoparticles were tested in yeast and E.
coli, they effectively inhibited bacterial growth. Gram-positive and gram-negative
bacteria have differences in their membrane structure, the most distinctive of which is
the thickness of the peptidoglycan layer. It is concluded that the lower efficacy of the
Ag nanoparticles against S. aureus may derive from the difference as a point of
membrane structure.
In contrast, Sondi and Salopek-Sondi (2004) reported that the antimicrobial
activity of silver nanoparticles on Gram-negative bacteria was dependent on the
concentration of Ag nanoparticle, and was closely associated with the formation of
dpitsT in the cell wall of bacteria. Then, Ag nanoparticles accumulated in the bacterial
membrane increased the permeability, resulting in cell death. The experimental

122
evidences are insufficient to explain the antimicrobial mechanism of positively charged
Ag nanoparticles.
Amro et al. (2000) suggested that metal depletion may cause the formation of
irregularly shaped pits in the outer membrane and change membrane permeability,
which is caused by progressive release of lipopolysaccharide molecules and membrane
proteins. Also, Sondi and Salopek-Sondi (2004) speculate that a similar mechanism
may cause the degradation of the membrane structure of E. coli during treatment with
Ag nanoparticles (Sondi and Salopek-Sondi, 2004). Although their interference
involved some sort of binding mechanism, still unclear is the mechanism of the
interaction between Ag nanoparticles and molecules of the outer membrane. Recently,
Danilczuk and co-workers (2006) reported Ag-generated free radicals through the ESR
study of Ag nanoparticles.
It is expected that the antimicrobial mechanism of Ag nanoparticles is related to
the formation of free radicals and subsequent free radical–induced membrane damage.
In conclusion, Ag nanoparticles prepared by the cost effective reduction method
described here have great promise as antimicrobial agents. Applications of Ag
nanoparticles based on these findings may lead to valuable discoveries in various fields
such as medical devices and antimicrobial systems. Silver and silver-based compounds
have been in use for centuries in the treatment of burns and chronic wounds (Castellano
et al., 2007).
Antioxidant property of synthesized nanoparticles:
The synthesized silver and gold nanoparticles showed antioxidant property,
when increase concentration of nanoparticles then the scavenging activity also is
increased. This gradual increase was observed in both of the nanoparticles (Fig 21). It is
known that gold nanoparticles exhibit high catalytic activity in some oxidation
reactions, such as low temperature CO oxidation, whereas gold as a compact metal is
chemically inactive because of both the size effect and the character of interaction of
the metal nanoparticles and support accompanied by the charge transfer between them.
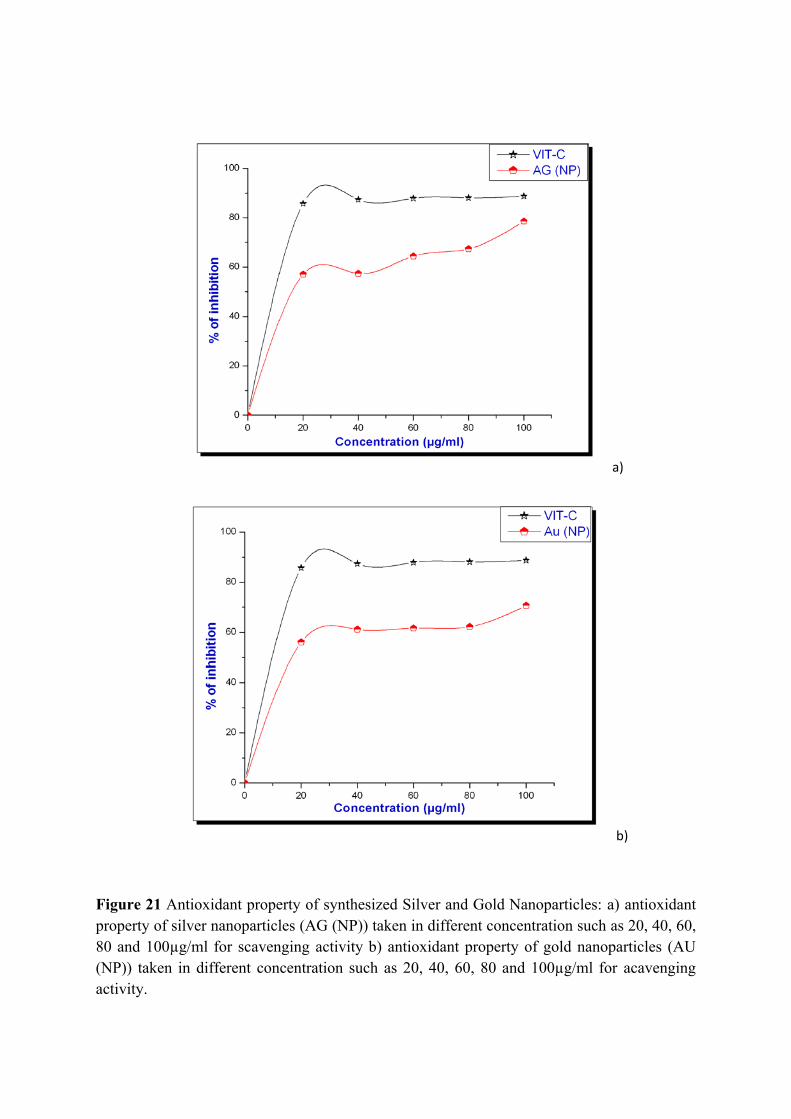
a)
b)
Figure 21 Antioxidant property of synthesized Silver and Gold Nanoparticles: a) antioxidant property of silver nanoparticles (AG (NP)) taken in different concentration such as 20, 40, 60, 80 and 100µg/ml for scavenging activity b) antioxidant property of gold nanoparticles (AU (NP)) taken in different concentration such as 20, 40, 60, 80 and 100µg/ml for acavenging activity.

123
The antioxidant activity of the gold nanoparticles in chitosan oligomer solutions
was studied by ESR spectroscopy from their ability to deactivate hydroxy radicals. The
•OH radicals were generated in a hydrogen peroxide ascorbic acid redox system (molar
ratio 1: 1). It was impossible to detect the PBN adduct directly with the hydroxy radical
because of its short lifetime. For instance, at pH 7.4 in an aqueous medium the halved
lifetime (1/2) of the PBN-OH adduct is 38s (Kotake and Janzen, 1991).
Therefore, we used the secondary radical spintrapping technique. Dimethyl
sulfoxide was added to the system, and its interaction with the hydroxy radicals
afforded Me• radicals. The Me• radicals were detected by their stable adduct with PBN
characterized (Takeshita et al., 2004) by the following HFC constants: aN = 1.6 mT
and а H = 0.36 mT. This method finds wide use in studies of the formation of hydroxyl
radicals (Kadiidka and Mason, 2002 and Burkitt and Mason, 1991).

Annexure
Summary, References, List of GenBank Accessions and Publications

124
SUMMARY
On the whole, the research work on survey, molecular systematics and
nanobiotechnological potentials of diatom and marine cyanobacteria has been
summarized in three chapters.
Chapter I
Totally, 50 cyanobacterial species and 38 diatom species were recorded. They
were identified taxonomically with standard monographs.
Five cyanobacterial and five diatoms were phylogenetically characterized using
16S and 18S rDNA sequencing.
16S and 18S rDNA sequences were deposited in GenBank (NCBI) with
accession numbers. Molecular modeling was done with C-phycocyanin (PC) gene
sequence isolated from cyanobacteria.
Chapter II
A marine cyanobacterium Phormidium tenue NTDM05 was selected for the
production of phycobiliproteins. Extraction and quantification of C-Phycobiliprotein
was done from selected Cyanobacteria
Optimum conditions were found to the production of C-phycoerythrin by
physical and chemical methods such as temperature, pH, extraction methods, buffer
system and nutrients.
Citric acid was found to the best preservative to preserve C-phycoerythrin and
antimicrobial property and biochemical composition of the pigments were studied for
food additive

125
Chapter III
Silver and Gold nanoparticles has been synthesized using marine
cyanobacterium, Phormidium willei NTDM01 and characterized with TEM, SEM,
EDS, EDAX, FS, FTIR and Uv-vis spectrum.
It was found to be 20nm range of silver nanoparticles have synthesized
intracellularly and 150nm size of particles synthesized extracellularly. Anisotropic gold
nanoparticles synthesized by the action intracellular proteins in the cyanobacteria.
Antimicrobial activity of silver nanoparticles was analyzed against multidrug
resistant clinical pathogens. In vitro antioxidant property and Fluorescence property
were studied for application of biolabelling process.
Diatom cell was selected for the nanofabrication of Si-Ge nanocomposite in
ambient condition. By bottom up process, the Si-Ge nanocomposite formed in the
diatom, Stauroneis sp., was characterized by SEM and EDS.
CdS nanoparticles were synthesized by biological technique, using c-
phycoerythrin as capping agent. This present study average particle come out to be
5.1±0.2nm for C-PE assisted CdS nanoparticles after the statistical analysis. The
difference in size may possibly due to the fact that nanoparticles are being formed at
different times. The earlier report also found that difference size of particles due to the
incubation time.
This simplest and possible procedure may apply to the commercial production
of uniform and stable CdS nanoparticles effectively. Hypothetical mechanism for the
synthesis of CdS nanoparticle has been confirmed with previous reports.
This work emphasizes on the synthesis and application of nanomaterials using
green nanotechnology procedure for the development of eco-friendly, less-laborious
and inexpensive materials without affecting their properties and with increased
efficiency.

126
REFERENCES
Abalde, J., Betancourt, L., Torres, E., Cid, A. and C. Barwell. 1998. Purification and
Characterization of Phycocyanin from the Marine Cyanobacterium Synechococcus sp.
IO9201. Plant Science, 136 (1) 109.
Abarzua, S., Jakubowski, S., Eckert, S. and Fuchs, P. 1999. Biotechnological investigation for
the prevention of marine biofouling II. Blue-green algae as potential producers of
biogenic agents for the growth inhibition of microfouling organisms. Botanica Mar 42,
459–465
Abed, R.M.M. and Garcia-Pichel, F. 2001. Long-term compositional changes after transplant in
a microbial mat cyanobacterial community composition revealed using a polyphasic
approach. Environ Microbiol 3, 53–62.
Abed, R.M.M. and Koster, J. 2005. The direct role of aerobic heterotrophic bacteria associated
with cyanobacteria in the degradation of oil compounds. Int Biodeterior Biodegrad 55,
29–37.
Abed, R.M.M., Palinska, K.A., Camoin, G. and Golubic, S. 2006. Common evolutionary origin
of planktonic and benthic nitrogen-fixing oscillatoriacean cyanobacteria from tropical
oceans – FEMS Microbiol. Lett., 260 (2): 171-177.
Abuskhuna S, Briody J, McCann M, Devereux M, Kavanagh K, Fontecha JB, et al. 2004.
Synthesis, structure and anti-fungal activity of dimeric Ag(I) complexes containing bis-
imidasole ligands. Polyhedron. 23:1249- 55.
Adams, D.G. in: Y.V. Brun, L.J. Shimkets (Eds.), Prokaryotic Development, ASM Press, Washington, DC, 2000, 51.
Addadi, L. Weiner, S. 1992. Control and design principles in biological mineralization. Angew.
Chem., Int. Ed. Engl. 31, 153
Ahmad, A., Mukherjee, P., Mandal, D., Senapati, S. Khan, M.I., Kumar, R., Sastry, M. 2002.
Enzyme mediated extracellular synthesis of CdS nanoparticles by the fungus Fusarium
oxysporum’, J.Am.Chem.Soc., 124: 12108-12109.
Ahmad, A., Mukherjee, P., Senapati, S., Mandal, D., Khan, M.I., Kumar, R., Sastry, M. 2003. Extracellular biosynthesis of silver nanoparticles using the fungus Fusarium
oxysporum. Colloids Surf B, 28: 313-318.
Ahmad, A., Senapati, S., Khan, M.I., Kumar, R., Ramani, R., Srinivas, V., Sastry, M. 2003.
Intracellular synthesis of gold nanoparticles by a novel alkalotolerant actinomycete,
Rhodococcus species’, Nanotechnology,14: 824-828.

127
Ahmad, A., Senapati, S., Khan, M.I., Kumar, R., Sastry, M. 2003. ‘Extracellular biosynthesis of
monodisperse gold nanoparticles by a novel extremophilic actinomycete,
Thermomonospera sp’, Langmuir, 19: 3550-3553.
Ahmed, A., P. Mukherjee, D. Mandal, S. Senapati, M. I. Khan, R. Kumar, M. Sastry. 2002.
Enzyme Mediated Extracellular Synthesis of CdS Nanoparticles by the
Fungus, Fusarium oxysporum.J. Am. Chem. Soc. 124: 12108- 12109.
Ahmed, A., Satyajyoti, S. Khan, M.I. Rajiv, K. Sastry, M. 2005. Extra-/intracellular
biosynthesis of gold nanoparticles by an alkalotolerant fungus, Trichothecium sp., J.
Biomed Nanotech. 1, 47-53.
Alivisatos, A. P. 1996. Semiconductor clusters, nanocrystals, and Quantum Dots. Science. 271:
933.
Amro N.A, Kotra L.P, Wadu-Mesthrige K, Bulychev A, Mobashery S, Liu G. 2000. High-
resolution atomic force microscopy studies of the Escherichia coli outer membrane:
structural basis for permeability. Langmuir. 16: 2789- 2796
Ankamwar B, Damle C, Ahmad A and Sastry M, 2005. Biosynthesis of gold and silver nanoparticles using Emblica officinalis fruit extract, their phase transfer and
transmetallation in an organic solution. J Nanosci Nanotechnol 5: 1665–1671.
Ankamwar B, Damle C, Ahmed A, Sastry, M. 2005b. Biosynthesis of Gold and silver
nanoparticles using Emblica officinalis fruit extract their phase transfer and
transmetallation in an organic solution. J. Nanosci. Nanotechnol. 5: 1665
Antal, T.K. and Lindblad, P. 2005. Production of H2 by sulphur-deprived cells of the
unicellular cyanobacteria Gloeocapsa alpicola and Synechocystis sp. PCC 6803 during
dark incubation with methane or at various extracellular pH. J Ferment Bioeng 98,
114–120.
Aoyama, K., Uemura, I., Miyake, J. and Asada, Y. 1997. Fermentation metabolism to produce
hydrogen gas and organic compounds in a cyanobacterium. Spirulina platensis J
Ferment Bioeng 83:17–20.
Armbrust, E.V, and Galindo, H.M. 2001. Rapid evolution of a sexual reproduction gene in
centric diatoms of the genus Thalassiosira. Appl. Environ. Microbiol. 67:3501–13.
Armbrust, E.V. 1999. Identification of a new gene family expressed during the onset of sexual
reproduction in the centric diatom Thalassiosira weissflogii. Appl. Environ. Microbiol.
65:3121-28.
Armendariz V, Isaac Herrera, Jose R, Peralta-Videa, Yacaman MJ, Troiani H, Santiago P,
Jorge, L, Gardea-Torresdey. 2004. Size controlled gold nanoparticles formation by
Avena sativa biomass: use of plants in Nanobiotechnology. J. Nanopar. Res. 6: 377.
Armendariz V, Parsons JG, Lopez ML, Peralta-Videa JR, Yacaman MJ, Jorge L, Gardea-
Torresdey. 2009. The extraction of gold nanoparticles from Oat and Wheat biomasses

128
using Sodium citrate and cetyltrimethylammoniumbromide, studied by X-ray
absorption spectroscopy, high-resolution transmission electro microscopy, and Uv-
Visible spectroscopy. Nanotech. 20: 105607.
Atou, T., K. Kusaba, K. Fukuoka, M. Kikuchi, Y. Syono, 1990. Shock induced phase transition
of M203 (M = Sc, Y, Sm, Gd, and In)-type compounds, J. Solid State Chem, 89, 378.
Aymonier C, Schlotterbeck U, Antonietti L, Zacharias P, Thomann R, Tiller JC, et al. 2002.
Hybrids of silver nanoparticles with amphiphilic hyperbranched macromolecules
exhibiting antimicrobial properties. Chem Commun. 24: 3018
Badger, M.R and T.J. Andrews. 1982. Photosynthesis and inorganic carbon usage by the
marine cyanobacterium, Synechococcus sp. Plant Physiol., 70: 517-523.
Badrinarayanan K, Sakthivel N. 2008. Coriander leaf mediated biosynthesis of gold
nanoparticles. Mate Lett. 62: 4588.
Bai, H.J., Z. M. Zhang, Y. Guo and G. E. Yang. 2009. Biological synthesis of size-controlled
cadmium sulfide nanoparticles by photosynthetic bacteria Rhodopseudomonas
palustris. Colloids Surf. B. 70: 142-146.
Baldauf,S.L., Roger,A.J., Wenk-Siefert.I., Doolittle,W.F. 2000. A kingdom-level phylogeny of
Eukaryotes based on combined protein data. Science 290: 972–977.
Bansal V, Rautaray D, Ahmad A, Sastry M. 2004. Biosynthesis of zirconia nanoparticles using
the fungus Fusarium oxysporum. J Mater Chem 14: 3303–3305.
Bansal, V., Rautaray, D., Bharde, A., Ahire, K., Sanyal, A., Ahmad, A., Sastry, M. 2005.
Fungus-mediated biosynthesis of silica and titania particles. J Mater Chem, 15: 2583–
2589.
Bao,Z., Weatherspoon,M.R., Shian,S., Cai,Y and Graham,P.D. 2007. Chemical reduction of
three-dimensional silica microassemblies into microporous silicon replicas. Nature
446:172–75.
Becker M, Stubbs, M.T and R. Huber. 1998. Crystallization of phycoerythrin 545 of
Rhodomonas lens using detergents and on usual additivies. Protein Sci 7:580–586.
Becker, S., J. M. Soukup, J. E. Gallagher, 2002. Differential particulate air pollution induced
oxidant stress in human granulocytes, monocytes and alveolar macrophages, Toxicol. In
Vitro, 16, 209
Bei, H., Guang-Ce, W., Chen-Kul Z et al. 2002. The experimental research of R- phycoerythrin
subunits on cancer treatment - A new photosensitizer in PDT. Cancer Biother
Radiopharm 17(1):35–42.
Benedetti, S., Benvenuti F, Pagliarani S et al. 2004. Antioxidant properties of a novel
phycocyanin extract from the blue-green alga Aphanizomenon flos-aquae. Life Sci
75:2353–2362.

129
Bennett, A. and Bogorad, L.1973. Complementary chromatic adaptation in a filamentous blue-
green alga. J.Cell Biol. 58:419-435.
Bergsland, K.J. and Haselkorn, R. 1991. Evolutionary relationships among Eubacteria,
Cyanobacteria, and Chloroplasts: evidence from the rpoC1 gene of Anabaena sp. strain
PCC 7120. J. Bacteriol., 173: 3446-3455.
Bermejo, R., Felipe, M. A., Talavera, E. M. and J.M. Alvarez-Pez. 2006. Expanded Bed
Adsorption Chromatography for Recovery of Phycocyanins from the
Microalga Spirulina platensis. Chromatograph, 63: 59.
Beveridge TJ, Murray RGE. 1980. Site of metal deposition in the cell wall of Bacillus subtilis.
J Bacteriol 141:876–887
Bhadury, P. and Wright, P.C. 2004. Exploitation of marine algae: biogenic compounds for
potential antifouling applications. Planta 219, 561–578.
Bhainsa, K.C., D’Souza, S.F., 2006. Extracellular biosynthesis of silver nanoparticles using
the fungus Aspergillus fumigatus, Colloids Surf B:Biointerf., 47: 160-164.
Bharde, A., Rautaray, D., Bansal, V., Ahmad, A., Sarkar, I., Yusuf, S.M., Sanyal, M., Sastry,
M. 2006. Extracellular biosynthesis of magnetite using fungi, Small, 2:135-141.
Bhat, V.B and K.M. Madyastha. 2000. C-phycocyanin : a potent peroxyl radial scavenger in
vivo and in vitro. Biochem Biophys Res Comm 275(1):20–25.
Bhattacharya D, Rajinder G. 2005. Nanotechnology and potential of microorganisms. Crit Rev
Biotechnol 25:199– 204
Bhaya, D., Dufresne, A., Vaulot, D. and Grossman, A. 2002. Analysis of the hli gene family in
marine and freshwater cyanobacteria - FEMS Microbiology Letters., 215 (2): 209-219.
Bidle,K.D and Azam,F. 1998. Accelerated dissolution of diatom silica by marine bacterial
assemblages. Nature 397: 508-512.
Bisby, F.A. 1995 Characterization of biodiversity. In: V.H. Heywood and R.T. Watson [Eds]
Global Biodiversity Assessment. Cambridge University Press, Cambridge, 21-106
Block, D.L. and Melody, I. 1992. Efficiency and cost goals for photoenhanced hydrogen
production processes. Int J Hydrogen Energy 17: 853–861.
Bloor, S. and England, R.R. 1989. Elucidation and optimization of the medium constituents
controlling antibiotic production by the cyanobactgerium Nostoc muscorum. Enzyme
Microb Tech 13: 76–81.
Borowitzka,M.A. 1995. Microalgae as sources of pharmaceuticals and other biologically active
compounds. J Appl Phycol 7:3–15.
Boyle JF, Manas-Zloczower I and Feke DL.2005. Hydrodynamic analysis of the mechanisms
of agglomerate dispersion. Powder Technol. 153: 127-133.

130
Brayner R Baberhousee H., hernadi M Djedjat and Yepremain C. Coradin T et al., 2007.
Cyanobacteria as bioreactor for the synthesis of Au, Ag, Pd and Pt nanoparticles via an
enzyme mediated route. J. Nanosci and Nanotech., 7: 2696-2708.
Brekhovskikh, A.A and O.D. Bekasova. 2005. CdS Nanoparticles in R-Phycoerythrin, a Protein
Matrix. Inorg. Mater. 41: 331-337.
Brock. T.D. 1973 Evolutionary and ecological aspects of the cyanophytes. In: N.G. Carr and
B.A. Whitton [Eds] The Biology of the Blue-Green Algae. Blackwell Scientific
Publications, Oxford, 487-500.
Brown, S., Sarikaya, M. & Johnson, E. 2000. A genetic analysis of crystal growth. J. Molecul.
Biol. 299: 725–735.
Bruchez, M., Moronne, M., Gin, P., Weiss, S., Alivisatos, A.P. 1998. Semiconductor
nanocrystals as fluorescent biological labels, Science, 281: 2016-2018.
Bryant, D.A. [Ed.] 1994 The Molecular Biology of Cyanobacteria. Kluwer Academic
Publishers, Dordrecht, 879 pp.
Burda, C., Chen, X., Narayanan,R and El-Sayed,M.A. 2005. Chemistry and properties of
nanocrystals of different shapes. Chem Rev. 105: 1025-102.
Burja, A.M., Banaigs, B., Abou-Mansour, E., Burgess, J.G. and Wright, P.C. 2001. Marine
cyanobacteria – a prolific source of natural products. Tetrahed. 57: 9347–9377.
Burkitt, M.J., R. P. Mason, Direct evidence for in vivo hydroxyl-radical generation in
experimental iron overload: An ESR spin-trapping investigation. Proc. Natl. Acad. Sci.,
1991, 88, 8440.
Cai,Y and Sandhage,K.H. 2005. Zn2SiO4-coated microparticles with biologically-controlled 3D
shapes. Phys. Stat. Sol. 202:105–7.
Capone, D.G., Burns, J.A., Montoya, J.P., Subramaniam, A., Mahaffey, A.C., Gunderson, T.,
Michaels, A.F. and Carpenter, E.J. 2005. Nitrogen fixation by Trichodesmium spp.: an
important source of new nitrogen to the tropical and subtropical North Atlantic Ocean.
Global Biogeochem Cycles. 19: 17.
Carmichael, W.W. and Gorham, P.R. 1980. Freshwater Cyanophyte Toxins. Algae Biomass.
pp. 437–448. New York: Elsevier.
Carpenter, E.J. 1983. Physiol.ogy and ecology of manne Oscillatoria (Trichodesmlum). Mar
Biol Lett 4.69-85
Carpenter, E.J., Capone, D.G. and Reuter, J.G. [Eds] 1992 Marine PelagicCyanobacteria:
Trichodesmium and other Diazotrophs. NATO ASI Series C
Carr, N.G and B.A. Whitton. 1982. The biology of cyanobacteria. Blackwell Scientific
publication, Oxford.

131
Casamatta, D.A., Vis, M.L. and Sheath R.G. 2003. Cryptic species in cyanobacterial
systematics: a case study of Phormidium retzii (Oscillatoriales) using RAPD molecular
markers and 16S rDNA sequence data - Aquatic Botany., 77 (4): 295-309.
Castanon GAM, Loredo MGS, Mendoza JRM, Ruiz F. 2005. Synthesis of CdS nanoparticles: a
simple method in aqueous media, Adv Tech Mat Mat Proc. 7:171–4.
Castellano JJ, Shafii SM, Ko F, Donate G, Wright TE, Mannari RJ, Payne WG, Smith DJ,
obson MC 2007. Comparative evaluation of silver-containing antimicrobial dressing
and drugs. Inter. Wound J. 4(2):144-22.
Castenholz, R.W. 1973 Ecology of blue-green algae in hot springs. In: N.G. Carr and B.A.
Whitton [Eds] The Biology of Blue-Green Algae. Blackwell Scientific Publications,
Oxford, 379-414.
Castenholz, R.W. 2001. Phylum BX. Cyanobacteria. Oxygenic photosynthetic bacteria. In
Bergey’s Manual of Systematic Bacteriology. The Archaea and the Deeply Branching
and Phototropic Bacteria ed. Garrity, G., Boone, D.R. and Castenholz, R.W. pp. 474–
487. New York: Springer-Verlag.
Castenholz, R.W. and Waterbury, J.B. 1989 In: J.T. Staley, M.P. Bryant, N. Pfennig and J.G.
Holt [Eds] Bergey's Manual of Systematic Bacteriology. Vol. 3, Williams & Wilkins,
Baltimore, 1710-1727.
Chakraborty, N., A. Banerjee, S. Lahiri A. Panda, A. N. Ghosh and Ruma Pal. 2009.
Biorecovery of gold using cyanobacteria and an eukaryotic alga with special reference
to nanogold formation – a novel phenomenon. J. Appl. Phycol. 21 (1): 145-152.
Chandran SP, Chadudhary M, Pasricha R, Ahamad A, Sastry M. Nanotriangles and silver
nanoparticles using Aloe vera plant extract. Biotechnol. Prog, 2006; 22: 577.
Chen, W., Z. Liu, Z. Wang, 1996. Some new observation on the formation and optical
properties of CdS clusters in zeolite-Y. Solid State Commun. 100: 101-104.
Chepurnov,V.A, Mann,D.G, Sabbe.K and Vyverman.W. 2004. Experimental studies on sexual
reproduction in diatoms. Int. Rev. Cytol. 237:91–154.
Chiara, Z and C. Bowler 2001. Exploring Bioinorganic Pattern Formation in Diatoms. A Story
of Polarized Trafficking. Plant Physiology, 127: 1339–1345.
Chow, M.K. & Zukoski, C.F. 1994. Gold sol formation mechanisms: role of colloidal stability. J. Colloid. Inter. Sci. 165: 97–109.
Chung, D., 2003. Nanoparticles have health benefits too, New Scientist, 179; 2410–2416.
Church, M.J., Hutchins, D.A and H.W. Ducklow. 2000. Limitation of bacterial growth by
dissolved organic matter and iron in the southern ocean. Appl. Environ. Microbiol.,
66(2): 455-466.

132
Cohen, Y., Jørgensen, B.B., Revsbech, N.P. and Poplawski, R. 1986. Adaptation to hydrogen
sulfide of oxygenic and anoxygenic photosynthesis among cyanobacteria. Appl Environ
Microbiol 51, 398–407.
Crispim, C.A. and Gaylarde, C.C. 2005. Cyanobacteria and biodeterioration of cultural
heritage: A review. Microbial Ecol. 49 (1): 1-9.
Cuenya, B. R, 2010. Synthesis and catalytic properties of metal nanoparticles: Size, shape,
support, composition, and oxidation state effects. Thin Solid Films, 518 (12), 3127
Cunningham DP, Lundie LL. 1993. Precipitation of cadmium by Clostridium thermoaceticum.
Appl Environ Microbiol 59:7–14.
Dahms, H.-U., Xu, Y. and Pfeiffer, C. 2006. Antifouling potential of cyanobacteria: a mini-
review. Biofouling 22, 317–327.
Dameron, C., R. N. Reese, R. K. Mehra., A. R. Kortan, P. J. Carroll, M. L. Steigerwald, L. E.
Brus and D. R. Winge. 1989. Biosynthesis of cadmium sulphide quantum
semiconductor crystallites. Nature. 338: 596- 597
Damste,J.S., Muyzer,G., and Abbas, B. 2004. The rise of the rhizosolenid diatoms. Science
304: 584-587.
Daniel, M.C., D. Astruc, 2004. Gold nanoparticles: assembly, supramolecular chemistry,
quantum-size-related properties, and applications toward biology, catalysis, and
nanotechnology. Chem. Rev. 104 (1), 293.
Danilczuk M, Lund A, Saldo J, Yamada H and Michalik J. 2006. Conduction electron spin
resonance of small silver particles. Spectrochim. Acta Part A. 63:189 - 91.
Darley,W.M and Volcani,B.E. 1971. Synchronised cultures: Diatoms. Methods Enzymol., 23:
85-96.
Darley,W.M., Sullivan,C.W and Volcani,B.E. 1976. Studies on the biochemistry and fine
structure of silica shell formation in diatoms. Planta, 130: 159-167.
De La Isla, A., Brostow, W., Bujard, B., Estevez, M., Rodriguez, J.R., Vargas, S., Castano,
V.M. 2003. Nanohybrid scratches resistant coating for teeth & bone visco elasticiy
manifested in tribology, Mat.Resr.Innovat, 7:110-114.
De Rosa, S.C., Brenchley, J.M and M. Roederer. 2003. Beyond six colours : a new era in flow cytometry. Nat Med, 9:112–117.
Delwiche, C.F., Kuhsel, M. and Palmer, J.D. 1995. Phylogenetic analysis of tufA sequences
indicates a cyanobacterial origin of all plastids – Molecul. Phylogen. Evolu. 4 (2): 110-
128.
Desikachary, T. V. 1959. Cyanophyta Indian Council of Agricultural Research New Delhi
India.

133
Desikachary,T.V. 1989. Marine diatoms of the Indian ocean region, Atlas of diatoms. Madras
scientific foundation.
Dill, K. A. 1990. Dominanat Forces in Protein Folding. Biochem. 29: 7133–7155.
Dobretsov, S., Dahms, H.-U. and Qian, P.-Y. 2006. Inhibition of biofouling by marine
microorganisms and their metabolites. Biofouling 22, 43–54.
Donaldson, K., Stone, V., Clouter, A., Renwick, L., MacNee, W. 2001. Ultrafine particles. Occup Environ Med, 58: 211-216.
Dor, I. and Danin, A. 1996 Cyanobacterial desert crusts in the Dead Sea Valley, Israel. Arch.
Hydrobiol. Suppl. 117, Algological Studies, 83: 197-206.
Douglas, S.E. 1994 Chloroplast origins and evolution. In: D.A. Bryant [Ed.] The Molecular
Biology of Cyanobacteria., Kluwer Academic Publishers, Dordrecht. 91-118
Druff, D.G., Curtis, A.C., Edwards, P.P., Jefferesen, D.A., Johnson, B.F., Kirkland, A.I. &
Logan, D.E. 1987. The morphology and microstructure of colloidal silver and gold.
Ang. Chem. Inter. Edi. 26: 676–678.
Drum,R.W and Gordon,R. 2003. Star Trek replicators and diatom nanotechnology. Trends
Biotechnol. 21: 325-328
Du TG, Schmid M, Jansen RP. 2007. Why cells move messages: the biological functions of
mRNA localization. Semin Cell Dev Biol 18: 171
Dufosse, L., Galaup, P., Yarnon, A et al. 2005. Microorganisms and microalgae as source of
pigments for use: a scientific oddity or an industrial reality? Trends Food Sci Technol
16: 389 -406.
Duran N, Marcato PD, Alves OL, De souza G, Esposito E. 2005. Mechanistic aspects of
biosynthesis of silver nanoparticles by several Fusarium oxysporum strains. J.
Nanobiotech. 3:1
Dutta, D., De, D., Chaudhuri, S. and Bhattacharya, S.K. 2005. Hydrogen production by
cyanobacteria. Microb cell Fact 4: 36.
Eccles, H. 1995. Removal of heavy metals from effluent streams – why select a biological
process? Int. Biodeterior. Biodegrad. 35, 5-16.
Edgar, L.A. and Pickett-Heaps, J.D. 1984. Diatom locomotion. In Progress in Phycological
Research, Vol. 3 (Round, F.E. & Chapman, G., editors), 47-88. , Bristol: Biopress.
Edwards,M.R. Hauer,C., Stack, R.F., Eisele, L.F and R. MacColl, 1997. Thermophilic C-
phycocyanin: effect of temerature, monomer stability, and structure, Biochim. Biophys.
Acta 1321. 157–164.
Eiler A., Langenheder, S., Bertilsson, S and L.J. Tranvik. 2003. Heterotrophic bacterial growth
efficiency and community structure at different natural organic carbon concentrations.
Appl. Environ. Microbiol., 69: 3701-3709.

134
Eriksen, N.T., Production of phycocyanin--a pigment with applications in biology,
biotechnology, foods and medicine. 2008. Appl. Microbiol. Biotechnol. 80:1–14.
Ernst, A., Marschall, P. and Postius, C. 1995. Genetic diversity among Synechococcus spp.
(Cyanobacteria) isolated from the pelagial of Lake Constance – FEMS Microbiology
Ecology., 17 (3): 197-203.
Estrada, J.E.P, Bescoc, P.B., Villar del Fresno, A.M. 2001. Antioxidant activity of different
fractions of Spirulina platensis protean extract. II Farmaco. 56:497–500.
Eustis,S., M. A. El-Sayed, 2006. Why gold nanoparticles are more precious than pretty gold:
Noble metal surface plasmon resonance and its enhancement of the radiative and
nonradiative properties of nanocrystals of different shapes. Chem. Soc. Rev. 35, 209.
Falkowski,P.C, Katz,M.E and Knoll, A.H. 2004. The evolution of modern eukaryotic
phytoplankton. Science. 305: 354-360.
Falkowski,P.G., Barber,R.T and Smetacek,V.V. 1998. Biogeochemical controls and feedbacks
on ocean primary production. Science 281:200–207.
Fatma, T., Sarada, R and L.V. Venkataraman. 1999. Evalution of selected species of Spirulina
for their constituents. In: Fatma T, (ed), Cyanobacterial and algal metabolism and
Environmental Biotechnology, narosa Publishing House, New Delhi. 113-119.
Fay, P. 1965 Heterotrophy and nitrogen fixation in Chlorogloea fritschii. J. Gen. Microbiol. 39,
11-20.
Fay, P. and Van Baalen, C. [Eds] 1987 The Cyanobacteria. Elsevier, Amsterdam, 534 pp.
Ferris, M.J., Muyzer, G. and Ward, D.M. 1996. Denaturing gradient gel electrophoresis profiles
of 16S rRNA-defined populations inhabiting a hot spring microbial mat community.
Appl Environ Microbiol 62, 340–346.
Field,C.B., Behrenfeld,M.J., Randerson,J.T and Falkowski,P. 1998. Primary production of the
biosphere: integrating terrestrial and oceanic components. Science 281:237–240.
Flenniken, M., M. Allen and T. Drouglas. Microbe Manufacturers of Semiconductors.
Chemistry & Biology 11 (2004) pp. 1478-1480.
Foster, M.J. 2002. Molecular modelling on structural biology. Micron., 33 :365 -384.
Fox, R.D. 1996. Spirulina: Production and potential. Editsud. Aixen Provence, France.
Frens, G. 1973. Controlled nucleation for regulation of particle-size in monodisperse gold
suspension. Nature- Physical Science, 241: 20-22.
Furno F, Morley KS, Wong B, Sharp BL, Arnold PL, Howdle SM, et al. Silver nanoparticles
and polymeric medical devices: a new approach to prevention of infection. J
Antimicrob Chemother 2004; 54:1019- 24.

135
Furuki, T., Schuichi, M. , Imajo, S., Hiroi, T., Amaya, T., Hirokawa, T., Ito, K and
Nozawa. 2003. Rapid and selective extraction of phycocyanin from Spirulina
platensis with ultrasonic cell disruption. J. Appl Phycol. 15: 319-324
Gacesa, P. and Hubble, J., Tecnología de las Enzimas. Editorial Acribia, Zaragoza. 1900
Gallon, J.R., Jones, D.A. and Page, T.S. 1996 Trichodesmium, the paradoxical diazotroph.
Arch. Hydrobiol. Suppl., Algological Studies, 83,215-243.
Gao, F., Q. Y. Lu, S. H. Xie and D. Y. Zhao. 2002. A Simple Route for the Synthesis of Multi-
Armed CdS Nanorod-Based Materials. Adv. Mater 14 : 1537-1540.
Garcia-Pichel, F. and Pringault, O. 2001. Cyanobacteria track water in desert soils. Nature 413,
380–381.
Gardea-Torresdey JL, Gomez E, Perlata-Videa JR, Parsons JG, Troiani H, Yacamen MJ.
Alfalfa sprouts: A Natural source for the synthesis of silver nanoparticles. Langumir.
2003; 19:1357
Garrett, R.H. & Grisham, C.M. 1999. Biochemistry. Saunders College Publishing, Sydney
Geitler, L. 1932. Cyanophyceae, In : Rabenhorst's Kryptogamenflora. Akademische
Verlagsgesellschaft Lepzig. 14: 1- 1196.
Gericke, M., Pinches, A. 2006b. Microbial production of gold nanoparticles, Gold Bull., 39: 22-
28.
Gericke, M., Pinches, A., 2006a. Biological synthesis of metal nanoparticles, Hydrometalurgy,
83:132-140.
Gerwick, W.H., Roberts, M.A., Proteau, P.J. and Chen, J.L. 1994. Screening cultured marine microalgae for anticancer-type activity. J Appl Phycol. 6: 143–149.
Giljohann, D.A., D. S. Seferos, W. L. Daniel, M. D. Massich, P. C. Patel, C. A. Mirkin, 2005.
Gold Nanoparticles for Biology and Medicine. Angew. Chem., Int. Ed. 49 (19), 3280.
Giovannoni, S.J., Turner, S., Olsen, G.J., Barns, S., Lane, D.J. and Pace, N.R. 1988.
Evolutionary relationships among cyanobacteria and green chloroplasts. J.Bacteriol.,
170: 3584-3592.
Glazer, A.N. 1989. Light Guides - Directional energy transfer in a photosynthetic antenna. J
Cell Biol 264(5):1–4
Goldman,J.C. 1993. Potential role of large oceanic diatoms in new primary production. Deep
sea Research, 40:159-168.
Gonzalez, R., Gonzalez, A and D. Remirez. 2003. Protective effects of phycocyanin on
galactosamine - induced hepatitis in rats. Biotecnol Appli. 20:107–110.

136
Gonzalez, R., Rodriguez, S., Romay, C., Ancheta, O., Gonzalez, A., Armesto, J., Remirez, D
and N. Merino. 1999. Anti-inflammatory activity of phycocyanin extract in acetic acid-
induced colitis in rats. Pharmacol Res. ;39:55–9.
Govindaraju, K.,, S. Khaleel Basha, V. Ganesh Kumar and G. Singaravelu. 2008. Silver, gold
and bimetallic nanoparticles production using single-cell protein (Spirulina platensis )
Geitler. J. Mater. Sci. 43 (15):5115-5122.
Grachev,M.A., Kuzn.sova,S.Yu and Sherbakova,T.A. 2006. A method for the isolation of pure
DNA for PCR. Molecular Biology,40.1: 159-161.
Greuter, W., Barrie, F., Burdet, H.M., Chaloner, W.G., Demoulin, V., Hawksworth, D.L.,
Jørgensen, P.M., Nicholson, D.H., Silva, P.C., Trehane, P. and McNeill, J. [Eds] 1994
International Code of Botanical Nomenclature (Tokyo Code). (Regnum Vegetabile No
131), Koeltz Scientific Books, Königstein. Geitler, L. 1932 Cyanophyceae. In: L.
Rabenhorst [Ed.] Kryptogamen-Flora. 14. Band. Akademische Verlagsgesellschaft,
Leipzig, 1196.
Guillard,R.R and Ryther,J.H. 1962. Studies of marine planktonic diatoms.I. Cyclotella nana Hustet and Detonula confervacea Cleve. Can J .Microbiol 8: 229-239
Guillard,R.R.L and Kilham,P. 1978. The ecology of marine planktonic diatoms. In: The
biology of diatoms (D.werner,ed). Botanical monographs, University of California
Press, Berkeley, California, USA. 13: 372-469
Gutu, T and Jun Jiao. 2006. Biosynthesis and Electron microscopy of Diatom Nanocomposites.
Material Research Society. Vol.901E.
Häder, D.P. 1987 Photomovement. In: P. Fay and C. Van Baalen [Eds] The Cyanobacteria.
Elsevier, Amsterdam, 325-345
Hamm,C.E., Merkel,R., Springer,O., Jurkojc,P and Maier,C. 2003. Architecture and material
properties of diatom shells provide effective mechanical protection. Nature 421:841–
43.
Hamouda T, Myc A, Donovan B, Shih A, Reuter JD, Baker Jr JR. 2000. A novel surfactant
nanoemulsion with a unique non-irritant topical antimicrobial activity against bacteria,
enveloped viruses and fungi. Microbiol Res. 156:1 -7.
Han, M., Gao, X., Su, J.Z., Nei, S. 2001. Quantum dot tagged microbeads for multiplexed
optical coding of biomolecules, Nature Biotech. 19: 631-635.
Harwood,D.M. Diatomite. In: Stoermer EF, Smol JP, eds. 1999. The Diatoms: Applications for
the Environmental and Earth Sciences. Cambridge University Press, 436-443
Hasle,G. and Syvertsen,E. 1996. Marine Diatoms. In: Identifying marine diatoms and
dinoflagellates (C.R Tomas, ed), Academic Press, Inc. San Diego, California. 5-385
Hayashi, K., Hyashi, T. and Kojima, J. 1991. A natural sulfated polysaccharide, calcium
spirulan, isolated from Spirulina platensis: in vitro and ex vivo evaluation of anti-

137
herpes simplex virus and anti-human immunodeficiency virus activities. AIDS Res
Hum Retrovir 12, 1463– 1471
Hemlata . Gunjan Pandey . fareha Bano . Tasneem Fatma. 2011. Studies on Anabaena sp.
NCCU-9 with special reference to phycocyanin. J. Algal Biomass Utln. 2 (1): 30 – 51
Herron, N., Y. Wang, 1990. Synthesis and Characterization of Surface-Capped, Size-Quantized
CdS Clusters. Chemical Control of Cluster Size. J. Am. Chem. Soc. 112: 1322.
Hess, W.R., Partensky, F., Vanderstaay, G.W.M., Garciafernandez, J.M., Borner, T. and
Vaulot, D. 1996. Coexistence of phycoerythrin and a chlorophyll a/b antenna in a
marine prokaryote - Proceedings of the National Academy of Science of the United
States of America.93 (20): 11126 - 11130.
Hildebrand, M., 2005. Prospects of manipulating diatom silica nanostructure. J. Nanosci.
Nanotech. 5, 146-157.
Hildebrand,M., Frigeri,L.G and Davis,A.K. 2007. Synchronized growth of Thalassiosira
pseudonana (Bacillariophyceae) provides novel insights into cell-wall synthesis
processes in relation to the cell cycle. J. Phycol. 43:730–40.
Hirata, T., Tanaka, M., Ooike, M., Tsunomura, T and M. Sakaguchi. 2000. Antioxidant
activities of phycocyanobilin prepared from Spirulina platensis. J Appl Phycol. 12:435–
9.
Hoagland, K.D., Rosowski, J.R., Gretz, M.R and Roemer, S.C. 1993. Diatom extracellular
polymeric substances : function, fine structure, chemistry and physiology. J. Phycol.,
29: 537-566.
Hoek, C. van den, Mann, D.G and Jahns, H.M. [Eds] 1995 Algae. An Introduction to Phycology. Cambridge University Press, Cambridge, 623.
Holland. H.D. 1997 Evidence for life on earth more than 3,850 million years ago. Science, 275,
38-39.
Holmes, J. D. Smith, P. R. Evans-Gowing, R. Richardson, D. J. Russell, D. A. Sondeau, J. R.
1995. Energy-dispersive X-ray analysis of the extracellular cadmium sulfide
crystallites of Klebsiella aerogenes. Arch. Microbiol. 163: 143.
Hosea, M., B. Greene, R. McPherson, M. Henzl, AM. Dale and DW. Darnall. Accumulation of
elemental gold on the alga Chlorella vulgaris. Inorg. Chim. Acta. 123 (1986) 253-261.
Hu, K., M. Brust, A. Bard, 1998. Characterization and Surface Charge Measurement of Self-
Assembled CdS Nanoparticle Films. Chem. Mater. 10: 1160.
Huang J, Li Q, Sun D, Lu Y, Su Y, Yang X, Wang H, Wang Y, Shao W, He N, Hong J, Chen
C. 2006. Biosynthesis of silver and gold nanoparticles by novel sundried Cinnanomum
camphora leaf. Nanotech. 18: 105104.

138
Humm, H.J. and Wicks, S.R. 1980 Introduction and Guide to the Marine Bluegreen Algae.
John Wiley & Sons, New York, 194 pp.
Husseiny, M.I., E1aziz, M.A., Badr, Y., Mahmoud, M.A. 2007. Biosynthesis of gold
nanoparticles using Pseudomonas aeruginosa, Spectrochim Acta A: Mol. Biomol.
Sectrosc., 67: 1003-1006.
Hutchinson, E.G. and Thornton, J.M. 1994. A Revised Set Of Potentials For Beta-Turn
Formation in Proteins. Protein Sci., 3: 2207-2216.
Iteman, I., Rippka, R., Tandeau de Marsac, N. and Herdman, M. 2002. rDNA analyses of
planktonic heterocystous cyanobacteria, including members of the genera
Anabaenopsis and Cyanospira. Microbiol., 148: 481-496.
Jaag, O. 1945 Untersuchungen über die Vegetation und Biologie der Algen des nackten
Gesteins in den Alpen, im Jura und im schweizerischen Mittelland. Kryptogamenflora
der
Jaki, B., Heilmann, J. and Sticher, O. 2000. New antibacterial metabolites from the
cyanobacterium Nostoc commune (EAWAG 122b). J Nat Prod. 63, 1283–1285.
Jefferies, T.W., Timourien, H. and Ward, R.L. 1978. Hydrogen production by Anabaena
cylindrica: effect of varying ammonium ad ferric ions, pH and light. App Environ
Microbiol 35, 704–710.
Jeffryes, C., Gutu, T., Jiao, J., and Rorrer, G.L. 2008. Peptide-Mediated Deposition of
Nanostructured TiO2 into the Periodic Structure of Diatom Biosilica. J. Mater. Res.
23, 3255-3262
Joerger, R., Klaus, T., Granquist, C.G. 2000. Biologically produced silver-carbon composite materials for optically functional thin film coatings, 12: 407- 409.
Johnson, C.J., Dujardin, E., Davis, S.A., Murphy, C.J. and Mann, S. 2002. Growth and form of
gold nanorods prepared by seed-mediated, surfactant-directed synthesis. J. Mater.
Chem. 12: 1765–1770.
Julian, S., Taurozzi., Voloymyr, V., Tarabara. 2007. Silver nanoparticle array on Track Etch
Membrane Support as Flow-T hrough Optical Sensors for water quality control,
Environ. Engin. Sci., 24: 122- 137.
Kadiiska, M.B., R. P. Mason, 2002. In vivo copper-mediated free radical production: an ESR spin trapping study. Spectrochim Acta, Part A, 58: 1227.
Kaito, C., Y. Saito, K. Fujita, 1989. Studies on the structure and morphology of ultrafine
particles of metallic sulfides. J. Cryst. Growth 94: 967.
Kajiyama, S., Kanazaki, H., Kawazu, K. and Kobayashi, A. 1998. Nostifungicidine, an
antifungal lipopeptide from the field-grown terrestrial blue-green algae Nostoc
commune. Tetrahedron Lett 39, 3737–3740

139
Kakkassery,J.J., Abid,J.P., Carrara,M and Fermin,D.J. 2004. Electrochemical and optical
properties of two dimensional electrostatic assembly of Au nanocrystals. Faraday
Discussions 125: 157-70.
Kaneko, T., Nakamura, Y., Wolk, C.P., Kuritz, T., Sasamoto, S. and Watanabe, A. 2001.
Complete genomic sequence of the filamentous nitrogen-fixing Cyanobacterium
Anabaena sp. strain PCC 7120. DNA Research., 8: 205-213.
Kann, E. 1988 Zur Autøkologie benthischer Cyanophyten in reinen europäischen Seen und
Fliessgewässern. Arch. Hydrobiol. Suppl. 80, Algological Studies, 50-53, 473-495.
Kendrew, J.C., Dickerson R.E., Strandberg, B.E., Hart, R.G. and Davies D.R. 1960. Structure
Of Myoglobin. Nature., 185: 422-427.
Khan Z, Bhadouria P, Bisen PS, 2005. Nutritional and therapeutic potential of Spirulina. Curr
Pharm Biotechnol, (5) 373-379.
Kilbourn, B. T., Yttria, in: M. B. Bever (Ed.), Encyclopedia of Materials Science and
Engineering Pergamon press, Ltd., Oxford, 7, 5509 (1986).
Kilbourn, B. T., Yttrium oxide, in: D. Bloor, R.J. Brook, M.C. Flemings, S. Mahajan (Eds.),
Pergamon Press, Ltd., Oxford, 4, 2957 (1994).
Kingery, W. D., H.K. Bowen, D.R. Uhlman, Introduction to Ceramics, second ed., John Wiley,
New York, 1976.
Kittelson, DB,. Watts, WF., Johnson, JP., Zarling, D., Schauer, J., Kasper, A., Baltensperger,
U., Burtscher, H. 2003b. Gasoline vehicle exhaust particle sampling study; Proceeding
of U.S. Department of Energy 9th Diesel Engine Emision Reduction Conference.
Kittelson, DB., Watts, WF., Johnson, JP. 2003a. On-Road Nanoparticle Measurements, 8th
International Conference On Environmental Science and Technology, Lemnos, Greece.
Klaus, T., Joerger, R., Olsson, E., Granquist, C.G. 1999. ‘Silver based crystalline nanoparticles,
microbially fabricated’ Proc.Natl.Acad.Sci., 96: 13611-13614.
Kofstad, P. Nonstoichiometry, Di.usion, and Electrical Conductivity in Binary Metal Oxides.,
Wiley Interscience, New York, 1972.
Kol, E. 1968 Kryobiologie. I. Kryovegetation. In: H.J. Elster and W. Ohle [Eds] Die
Binnengewässer, Band XXIV. E. Schweizerbart'sche Verlagsbuchhandlung, Stuttgart,
216 pp.
Konishi, Y., Nomura, T., Tsukiyama, T., Saitoh, N. 2004. Microbial preparation of gold
nanoparticles by anaerobic bacterium, Trans.Mater.Res.Soc.Jpn., 29: 2341-2343.
Kooistra,W.H.C.F and Medlin,L.K. 1996.The evolution of the diatoms (Bacillariophyta)
IV. A reconstruction of their age from small subunit rRNA coding regions and the
fossil record. Molecular Phylo. Evolut.6:391-407.

140
Korobushkina, E. D.; Karavaiko, G. I., Korobushkin, I. M. In Environmental Biogeochemistry;
Hallberg, R., Ed.; Economic Bulletin: Stockholm, 1983; Vol. 35, pp 325-333.
Kotake, Y., E. G. Janzen, Studies on the structure and morphology of ultrafine particles of
metallic sulfides. J. Am. Chem. Soc., 1991, 113, 9503.
Kowallik,K.V. 1992. Origin and Evolution of Plastids from Chlorophyll-a+c-Containing Algae:
Suggested Ancestral Relationships to Red and Green Algal Plastids. Chapman and
Hall, New York.
Kowshik, M., Ashtaputre, S., Kharrazi, S., Vogel, W., Urban, J., Kulkarni, SK., Paknikar,
KM. 2003. Extracellular synthesis of silver nanoparticles by a silver-tolerant yeast
strain MKY3. Nanotech.. 14: 95-100.
Kowshik, M., Deshmukh, N., Vogel, W., Urban, J., Kulkarni, S.K., Paknikar, K.M. 2002.
Microbial synthesis of semiconductor CdS nanoparticles, their characterization and
their use in the fabrication of an ideal diode, Biotechnol. Bioeng., 78: 583-588.
Krimm, S and J. Bandekar, 1986. Vibrational spectroscopy and conformation of peptides,
polypeptides and proteins, Adv. Protein Chem. 38:181–364.
Kronick, M.N and P.D. Grossman. 1983. Immunoassay techniques with fluorescent
phycobiliprotein conjugates. Clin Chem 29(9):1582– 1588.
Kula, M. R. and H. Schütte. 1987. Purification of Proteins and the Disruption of Microbial Cell.
Biotechnol. Prog. 3(1) 31.
Kulshreshtha, A., Jacharia, A.J. Jarouliya, U., Bhadauriya, P., Prasad, G.B.K.S and P.S. Bisen,
2008. Spirulina in health care management. Curr. Pharm. Biotechnol. 9: 400–405.
Kumar, S.A., Ayoobul, A.A., Absar, A., Khan, M.I., 2007. Extracellular biosynthesis of CdSe
quantum dots by the fungus Fusarium oxysporum, J.Biomed.Nanotechnol., 3: 190- 194.
Kumari, N., Srivastava, A.K., Bhargava, P. and Rai, L. 2009. Molecular approaches towards
assessment of cyanobacterial biodiversity. African J. Biotechnol. 8(18): 4284-4298
Kusari, U., Z. Bao, Y. Cai, G. Ahmad, K.H. Sandhage, and L.G. Sneddon, 2007. Formation of
nanostructured, nanocrystalline boron nitride microparticles with diatom-derived 3-D
shapes. Chem. Comm. 11, 1177-1179.
Laamanen, M. 1996 Cyanoprokaryotes in the Baltic Sea ice and winter plankton. Arch. Hydrobiol. Suppl. 117, Algological Studies, 83, 423-433.
Labrenz M, Druschel GK, Thomsen-Ebert T, Gilbert B, Welch SA Kemner KM, Logan GA,
Summons RE, Stasio GD, Bond PL, Lai B, Kelly SD, Banfield JF. 2000. Formation of
sphalerite (ZnS) deposits in natural biofilms of sulfate-reducing bacteria. Science
290:1744–1747.
Ladizhansky, V., G. Hodes and S. Vega. 1998. Surface Properties of Precipitated CdS
Nanoparticles Studied by NMR. J. Phy. Chem. B. 102: 8505-8509.

141
Lambert, G.R. and Smith, G.D. 1977. Hydrogen formation by marine blue-green algae. FEBS
Microbiol. Lett 83: 159–162.
Leabeau,T., Gaudin,P., Moan,R and Robert,J.M. 2002. A new photobioreactor for continuous
marenin Production with a marine diatom: influence of the light intensity and the
immobilised-cell matrix (alginate beads or agar layer). Appl Microbial biotech 59:
153-159.
Lee,S.J., Huang,C.H., Shian,S and Sandhage,K.H. 2007. Rapid hydrolysis of
organophosphorous esters induced by nanostructured, fluorine-doped titania replicas of
diatom frustules. J. Am. Ceram. Soc. 90:1632–36.
Lengke MF, Ravel B, Fleet ME, Wanger G, Gordon RA, Southam G. 2007. Precipitation of
gold by reaction of aqueous gold(III)-chloride with cyanobacteria at 25–80 °C, studied
by X-ray absorption spectroscopy. Can J Chem 85: 1–9.
Lengke MF, Southam G. 2005. The effect of thiosulfate-oxidizing bacteria on the stability of
the gold-thiosulfate complex. Geochim Cosmochim Acta 69: 3759–3772.
Lengke, M., Fleet, M.E., Southam, G. 2006. Morphology of gold nanoparticles synthesized by filamentous cyanobacteria from gold(I)-thiosulfate & gold(III)-chloride complexes.
Langmuir, 22: 2780-2787.
Lengke, M., Ravel, B., Fleet, M.E.,Wanger, G., Gordon, R.A., Southam, G. 2006. Mechanisms
of gold bioaccumulation by filamentous cyanobacteria from gold (III)-chloride
complex, Environ.Sci.Technol., 40: 6304-6309.
Lengke, M., Southam, G. 2006. Bioaccumulation of gold by sulfate-reducing bacteria cultured
in the presence of gold (I)-thiosulfate complex, Geochim Cosmodium Acta, 70: 3646-
3661.
Li CW, Volcani BE. 1987. Four new apochlorotlc diatoms. Br Phycol J. 22:375-382
Li, D.M and Y.Z. Qi. 1997. Industry in China: present status and future prospects. J Appl
Phycol. 9:25– 8.
Li, J.H., C. L. Ren, X. Y. Liu, Z. D. Hu, D. S. Xue, 2007. Green synthesis of starch capped
CdSe nanoparticles at room temperature. Mater. Sci. Eng. A 458: 319-322.
Li, Z and Y. Du. 2003. Biomimic synthesis of CdS nanoparticles with enhanced luminescence.
Mat. Lett. 57: 2480-2484.
Li. Du, H Jiang, Liu. X and Wang. E . 2007. Biosynthesis of gold nanoparticles assigned by
E.coli DH5α and its application on direct electrochemistry haemoglobin. Electrochem.
Commun. 9: 1165-1170.
Liji Shobhana, S.S., M. Vimala Devi, T. P. Sastry and A. S. Mandal. 2011. CdS quantum dots
for measurement of the size-dependent optical properties of thiol capping. J. Nanopart.
Res. 13 1747-1757.

142
Lind,J.L., Heimann,E.A and Vliet,C. 1997. Substratum adhesion and gliding in a diatom are
mediated by extracellular proteoglycans. Planta 203:213-221.
Lindbald, P. 1999. Cyanobacterial H2 metabolism: knowledge and potential ⁄ strategies for a
photobiotechnological production of H2. Biotechnologia Aplicada 16, 141–144.
Liu, J.K., C. X. Luo, X. H. Yang and X. Y. Zhang. 2009. Ultrasonic-template method synthesis
of CdS hollow nanoparticle chains. Mat. Lett. 63: 124-126.
Liu, Y., Xu, L., Cheng, N et al. 2000. Inhibitory effect of phycocyanin from Spirulina platensis
on the growth of human leukemia k562 cells. J Appl Phycol. 12:125–130.
Liufu, SC., Xiao, HN., Li, YP. 2004. Adsorption of cationic polyelectrolyte at the solid/liquid
interface and dispersion of nanosized silica in water. J Coll Interface Sci, 285: 33-40.
Liyana-Pathirana, C.M. and F. Shahidi. 2005. Antioxidant activity of commercial soft and hard
wheat (Triticum aestivum L.) as affected by gastric pH conditions. J. Agricul. Food
Chem.53: 2433–2440
Lobban, C.S, and P.J. Harrison. Cambridge, USA: Cambridge University Press; 1994.Seaweed
ecology and physiology.
Lone, J., Lene, D.S., Karsten, O et al. 2005. Heat and light stability of three natural blue
colourants for use in confectionery and beverages. Eur Food Res Technol 220(3–
4):261–266.
Losic,D., Rosengarten,G., Mitchell,J.G and Voelcker,N.H. 2006. Pore architecture of diatom
frustules: potential nanostructured membranes for molecular particle separation. J.
Nanosci. Nanotech. 5 :5-14.
Losic,D., Mitchell,J.G and Voelcker,N.H. 2005. Complex gold nanostructures derived by
templating from diatom frustules. Chem Commun. 4905-4907.
Luangpipat, T., Isabel R Beattie, Yusuf Chisti and Richard G. Haverkamp. 2011. Gold
nanoparticles produced in a microalga. J. Nanopart. Res. 13 (12): 6439-6445.
Ludoph, B., M.A. Malik, 1998. Novel single molecule precursor routes for the direct synthesis
of highly monodispersed quantum dots of cadmium or zinc sulfide or selenide. Chem.
Commun. 17: 1849.
Lujan, J.R., Darnall, D.W., Stark, P.C., Rayson, G.D. and Gardea-Torresdey, J.L. 1994. Metal ion binding by alga and higher plant tissues: a phenomenological study of solution pH
dependence. Sol. Ext. Ion Exchan. 12: 803–816.
Ma, I., Wong, H., Kong, L.B., Peng, K.W. 2003. Biomimetic processing of nanocrystallite
bioactive apatite coating on titanium, Nanotech., 14: 619-623.
Manish Mishra, Hemant Kumar, Kamlakar Tripathi. 2008. Diabetic delayed wound healing and
the role of silver nanoparticles Digest J. Nanomat. Biostru. 3(2), 49.

143
Mann, S.; Ozin, G. A. 1996. Synthesis of inorganic materials with complex form. Nature, 382:
313.
Mann,D.G. 1999. The species concept in diatoms. Phycologia, 38: 437-495.
Marchler-Bauer A et al. 2011. CDD: a Conserved Domain Database for the functional
annotation of proteins. Nucleic Acids Res.39: 225-9.
Marscher, H. 1995. Mineral Nutrition of Higher Plants. Academic Press, New York
Marsh PD. 1992. Microbiological aspects of the chemical control of plaque and gingivitis. J
Dent Res. 71:1431- 8.
Martin, K.A., Siefer J.L., Yerrapragada, S., Lu, Y., Mcneill, T.Z., Moreno, P.A., Weinstock,
G.M., Widger, W.R. and Fox, G.E. 2003. Cyanobacterial signature genes –
Photosynthesis Research., 75 (3): 211-221.
Masukawa, H., Nakamura, K., Mochimaru, M. and Sakurai, H. 2001. Photobiological
Hydrogen Production and Nitrogenase Activity in Some Heterocystous Cyanobacteria.
BioHydrogen II, pp. 63–66. Oxford, UK: Elsevier. Mathematical and Physical
Sciences, Vol. 362. Kluwer Academic Publishers, Dordrecht
Mayr, E. 1981 Biological classification: toward a synthesis of opposing methodologies.
Science, 214, 510-516.
McHugh, J. B. 1988. Concentration of gold in natural waters. J.Geochem. Explor. 30, 85-94.
Medlin, L.K., Kooistra,W.H.C.F and Gersonde,R. 1996. Evolution of the diatoms
(Bacillariophyta). II. Nuclear-Encoded small-subunit rRNA sequence comparisons
confirm a paraphyletic origin for the centric diatoms. Mol Biol Evol 13: 67-75.
Medlin,L.K., Elwood,H.J., Stickel,S and Sogin,M.L. 1988. The characterization of
enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 71:491–499.
Mengel, K. and E.A. Kirkby. 1987. Principles of Plant Nutrition. 4th Edition. International
Potash Institute. IPI, Bern, Switzerland. 685
Milhemy, D. M., E. Matijevic, 1984. Preparation and properties of monodispersed spherical-
colloidal particles of zinc sulphide. J. Chem. Soc., Faraday Trans.80: 563-570.
Mirkin CA, Taton TA. 2000. Semiconductors meet biology. Nature. 405: 626- 7.
Mishra, S. K., Shrivastav, A and Mishra, S. 2008. Effect of preservatives for food grade C-PC
from Spirulina platensis. Process Biochem.43 (4), 339-345.
Mishra, S. Phycobiliproteins from microalgae recent advances on applied aspects of Indian
marine algae with reference to Global scenario, vol. 2; 2007. p. 273–84.
Mishra, S., Chundawat, R.S., Sood, P.P and S. Mishra. 2005. Efficacy of C-phycocyanin
against Cr(IV) toxicity-induced lipid peroxidation. J Cell Tissue Res. 5(2):513–4.

144
Mishra, S.K., Shrivastav, A., Pancha, I., Jain, D and S. Mishra. 2010. Effect of preservatives for
food grade C-Phycoerythrin, isolated from marine cyanobacteria Pseudanabaena sp.
Int. J. Biol. Macromol. 47. 597–602.
Mitsui, A., Enternmann, B. and Gill, K. 1983. Indoor and outdoor cultivation of Tipapia in
seawater with algae as a sole food source. In Proceedings of the Second North Pacific
Aquaculture System. pp. 323–340, Japan: Tokyo University.
Moares, C. C., Burkert, J. F. M. and S.J. Kalil. 2010. C-Phycocyanin Extraction Process for
Large-Scale Use. J. Food Biochem. 34 (1): 133.
Moore, L.R., Rocap, G. and Chisholm, S.W. 1998. Physiology and molecular phylogeny of
coexisting Prochlorococcus ecotypes. Nature. 393 (6684): 464-467.
Morones, J.R., J.L. Elechiguerra, A. Camacho, K. Holt, J.B. Kouri, J.L. Ramirez, M.J.
Yacaman, 2005. The bactericidal effect of silver nanoparticles. Nanotech.16: 2346.
Mount, David W. 2002. Bioinformatics: Sequence and Genome Analysis. Spring Harbor Press.,
ISBN 0-87969-608-7.
MubarakAli, D and N. Thajuddin. 2009. Biosynthesis of Colloidal silver nanoparticles using
fresh water cyanobacterium, Oscillatoria earlei NTDM03. Seaweed Res. Util.31 (1&2):
219-226.
MubarakAli, D, Suresh, A, Praveen Kumar, R, Gunasekaran, M and N. Thajuddin. 2010.
Efficiency of textile dye decolorization by marine cyanobacterium, Oscillatoria
formosa NTDM02. African J. Basic Appli. Sci., 3(1): 9-13
MubarakAli, D., Arun Kumar, J., Suriya, K.R., Sheik Syed Ishack, K.A and N. Thajuddin.
2012. Molecular Modeling and phylogenetic analysis of C-Phycocyanin gene sequence
from marine cyanobacterium, Phormidium tenue NTDM05. Seaweed Res. Util.
(Accepted)
MubarakAli, D., Divya, C., Gunasekaran, M and N. Thajuddin. 2011. Biosynthesis and
characterization of Silicon-Germanium oxide nanocomposite by Diatom. Digest J.
Nanomater. Biostruc. 6(1): 117-120.
MubarakAli, D., Divya, C., Praveen Kumar, R., Parveez Ahamed and N. Thajuddin. 2010.
Scanning Electron Microscopic structural studies of Diatoms. Inter. J. Microbiol. Res.
1(2): 72-74.
MubarakAli, D., Gopinath, V, Rameshbabu, N and N. Thajuddin. 2012. Synthesis and
characterization of CdS nanoparticles using c-phycoerythrin extracted from marine
cyanobacteria. Mater. Lett. 74: 8-11.
MubarakAli, D., N. Thajuddin, K. Jeganathan, M. Gunasekaran. 2011. Plant extract mediated
synthesis of silver and gold nanoparticles and its antibacterial activity against clinically
isolated pathogens. Colloids and Surfaces B: Biointerfaces 85 (2011) 360–365.

145
MubarakAli, D., Sasikala, M., Gunasekaran, M and N. Thajuddin. 2011. Biosynthesis and
characterization of silver nanoparticles using marine cyanobacterium, Oscillatoria
willei NTDM01. Digest J. Nanomater. Biostruc. 6(2): 415-420.
MubarakAli, D., Vinodh Kumar, T, and D. Thajuddin. 2008. Screening of some selected
hypersaline cyanobacterial isolates for biochemical and antibacterial activity. Indian
Hydrobiol. 11(2): 241-246.
Mude N, Avinash I, Aniket G, Mahendra R. 2009. Synthesis of Silver Nanoparticles Using
Callus Extract of Carica papaya – A First Report. J. Plant Biochem. Biotech. 18 (1):
0971.
Mukherjee, P. Ahmad, A., Mandal, D., Senapati, S., Sainkar, S.R., Khan, M.I., Ramani, R.,
Parischa, R., Kumar, P.A.V., Alam, M., Sastry, M., Kumar, R. 2001. Bioreduction of
AuCl4 ions by the fungus, Verticillium sp. And the surface trapping of gold
nanoparticles formed. Angew.Chem.Int.Ed., 40: 3585-3588.
Mukherjee, P., A. Ahmed, D. Mandal, S. Senapati, S.R. Sainkar, M.I. Khan, R. Parischa, P.V.
Ajayakumar, R. Alam kumar, M. Sastry, 2001. Fungus-Mediated Synthesis of Silver
Nanoparticles and Their Immobilization in the Mycelial Matrix: A Novel Biological
Approach to Nanoparticle Synthesis. Nano Lett, 1: 515.
Mukherjee, P., Senapati, S., Mandal, D., Ahmad, A., Khan, M.I., Kumar, R., Sastry, M. 2002.
Extracellular synthesis of gold nanoparticles by the fungus Fusarium oxysporum,
Chem.Bio.Chem., 3: 461-463.
Mulvaney, P. 1996. Surface plasmon spectroscopy of nanosized metal particles. Langmuir, 12:
788–800.
Mur, L.R., Skulberg, O.M., and Utkilen, H. 1999. Cyanobacteria in the environment. In Toxic
Cyanobacteria in Water - A Guide to Their Public Health Consequences, Monitoring
and Management. Chorus, I., and Bartram, J. (eds). London, UK: Spon Press. 15-40.
Muralitharan, G. and N. Thajuddin. 2011. Rapid differentiation of phenotypically and
genotypically similar Synechococcus elongatus strains by PCR fingerprinting. Biologia,
66 (2): 238 – 243.
Muralitharan, G. and Thajuddin, N. 2010. M13-based genotyping of marine cyanobacterial
strains from the Indian subcontinent and maintained in the NFMC germplasm
collection. J. Appl. Phycol. 22(6): 709 – 716.
Murphy, C. J., T. K. San, A. M. Gole, C. J. Orendorff, J. X. Gao, L. Gou, S. E. Hunyadi, T. Li,
2005. Anisotropic Metal Nanoparticles: Synthesis, Assembly, and Optical
Applications. J. Phys. Chem. B, 109 (29), 13857
Murray, C.B., Kagan, C.R., Bawendi, M.G.., 2000. ‘Synthesis and characterization of
monodisperse nanocrystals and close-packed nanocrystal assemblies, Annv.
Rev.Mater.Sci., 30: 545-610.

146
Murray,C.B., Norris, D.J and Bawendi,M.G. 1993. Synthesis and characterization of nearly
monodisperse CdE (E = sulfur, selenium, tellurium) semiconductor nanocrystallites. J.
Amer. Chem. Soc 115: 8706-715.
Nagaoka, S., Shimizu, K., Kaneko, H et al. 2005. A novel protein C -phycocyanin plays a
crucial role in the hypocholesterolemic action of Spirulina platensis concentrate in rats.
J Nutr. 135: 2425 –2430.
Nair, B., Pradeep, T. 2002, Coalescence of Nanoclusters and Formation of Submicron
Crystallites Assisted by Lactobacillus Strains. Cryst. Growth Des., 2: 293-298.
Nakajima, A. Accumulation of gold by microorganisms. World J. Microbiol. Biotechnol. 2003,
19, 369-374.
Nam J M, Thaxton C S and Mirkin C A 2003. Nanoparticle-Based Bio-Bar Codes for the
Ultrasensitive Detection of Proteins. Science 301 1884.
Neilan, B. A. 2002. The molecular evolution and DNA profiling of toxic cyanobacteria. Curr.
Issues Mol. Biol., 4: 1-11.
Neilan, B.A. 1995 Identification and phylogenetic analysis of toxigenic Cyanobacteria by
multiplex Randomly Amplified Polymorphic DNA PCR - Applied and Environmental
Microbiology., 61 (6): 2286 - 2291.
Niu, J.-F., Wang, G.-C. and Tseng, C.-K. 2006. Method for large-scale isolation and
purification of R-phycoerytrin red alga Polysiphonia urceolata Grev. Protein Expr.
Purif. 49, 23–31.
Nubel, U., Garcial-Pichel, F. and Muyzer, G. 1997. PCR primers to amplify 16S rRNA genes
from Cyanobacteria. Appl. Environ. Microbiol., 63: 3327–3332.
Nubel, U., Garcia-Pichel, F., Clavero, E. and Muyzer, G. 2000. Matching molecular diversity
and ecophysiology of benthic cyanobacteria and diatoms in communities along a
salinity gradient. Environ Microbiol 2: 217–226.
Nubel, U., Garcia-Pichel, F., Ku¨hl, M. and Muyzer, G. 1999. Quantifying microbial diversity:
morphotypes, 16S rRNA genes, and carotenoids of oxygenic phototrophs in microbial
mats. Appl Environ Microbiol 65: 422–430.
Oberdörster, G., Oberdörster, E., Oberdörster, J. 2005, Nanotoxicology: An emerging discipline
evolving from studies of ultrafine particles. Environ Health Perspect, 113: 823-839.
Ohto, P, Ishida, C., Nakane, H., Muramatsu, M., Nishino, T. and Obata, S. 1999. A
thermophilic cyanobacterium Synechococcus elongatus has three different Class I
prenyltransferase genes. Plant Mol Biol 40, 1573–1628
Oi, V.T., Glazer, A.N and L. Stryer. 1982. Fluorescent phycobiliproteins conjugates for
analysis of cells and molecules. J Cell Biol. 93:981–986.

147
Olvera-Ramirez, R., Coria-Cedillo, M., Canizares-Villanueva, R.O., Martinez-Jeronimo, F.,
Ponce-Noyola, T., Rios-Leal, E. 2000. Growth evaluation and bioproducts
characterization of Calothrix sp. Bioresour. Technol. 72 121–124.
Oranda, H.W.K., Mercedes, R.L and S.B. Donald. 1975. Physical Chemical properties of C-
PC isolated from an acid thermophilic eukaryote Cyanidium caldaricum. Biochem J.
147:63-70.
Ormerod, J.G. 1992 Physiology of the photosynthetic prokaryotes. In: N.H. Mann and N.G.
Carr [Eds] Photosynthetic Prokaryotes. Plenum Press, New York, 93-120.
Ouellette, A.J.A., and Wilhelm, S.W. 2003. Toxic cyanobacteria: the evolving molecular
toolbox. Front. Ecol. Environ. 1: 359-366.
Oyle, JF., Manas-Zloczower, I., Feke DL. 2005, Hydrodynamic analysis of the mechanisms of
agglomerate dispersion. Powder Technol..153, 127-133.
Packer, L. and Glazer, A.N. [Eds] 1988 Cyanobacteria. Methods in Enzymology. Volume 167,
Academic Press, New York, 915 pp.
Paerl, H.W. 1988 Growth and reproductive strategies of freshwater blue-green algae. In: C.D.
Sandgren [Ed.] Growth and Reproductive Strategies of Freshwater Phytoplankton.
Cambridge University Press, Cambridge, 261-315.
Palinska, K.A., Liesack, W., Rhiel, E. and Krumbein, W.E. 1996. Phenotype variability of
identical genotypes: The need for a combined approach in cyanobacterial taxonomy
demonstrated on Merismopedia-like isolates. Arch. Microbiol. 166 (4): 224-233.
Papendorf, O., Ko¨nig, G.M. and Wright, A.D. 1998. Hirridin B and 2,4-dimethoxy-6
heptadecylphenol, secondary metabolites from the cyanobacterium Phormidium
ectocarpi with antiplasmodial activity. Phytochem. 49: 2383–2386.
Papke, U., Gross, E.M. and Francke, W. 1997. Isolation, identification and determination of the
absolute configuration of Fischerellin B. A new algicide from the freshwater
cyanobacterium Fischerella muscicola (Thuret). Tetrahedron Lett 38, 379–382.
Parashar V, Parashar R, Sharma B, Pandey AC. 2009. Parthenium leaf extract mediated
synthesis of silver nanoparticles : A Novel approach towards weed utilization. Digest J.
Nanomat. Biostru. 4(1):45.
Parial, D., Patra, H.K., Roychoudhury, P., Dasgupta, A.K. and Pal, R. 2012. Gold nanorod production by cyanobacteria – a green chemistry approach. J. Appl. Phycol. 24: 55–60.
Parker,A.R. and Townley, H. E. 2007. Biomimetics of Photonic Nanostructures. Nat.
Nanotechnol. 347–353.
Parkinson,J., Brechet.Y and Gordon,R. 1999a. Centric diatom morphogenesis: a model based
on a DLA algorithm investigating the potential role of microtubules. Biochim. Biophys.
Acta 1452:89–102.

148
Parkinson.J, and Gordon.R 1999b. Beyond micromachining: The potential of diatoms.Trends
Biotechnol. 17:190–96.
Patel, A., Mishra, S and P.K. Ghosh. 2006. Antioxidant potential of C-Phycocyanin isolated
from three cyanobacterial species: Lyngbya, Phormidium and Spirulina spp.. Indian J
Biochem. Biophy.43:31–5.
Patel, A., Mishra, S., Pawar, R and P.K. Ghosh. 2005. Purification and characterization of C-
phycocyanin from cyanobacterial species of marine and freshwater habitat. Prot Expr
Purif. 40:248–55.
Patel, A., Pawar, R., Mishra, S., Sonawane, S and P.K. Ghosh. 2004. Kinetic studies on thermal
denaturation of C-Phycocyanin. Indian J Biochem Biophy.41:254–7.
Patterson, G.M.L., Larsen, L.K. and Moore, R.E. 1994. Bioactive natural products from blue-
green algae. J Appl Phycol 6: 151–157.
Paul, B.T., Selvam, G., Murugesan, R., Patel, A., Mishra, S and P.K. Ghosh. 2006.
Photodynamic action of C-phycocyanins obtained from marine and fresh water
cyanobacterial cultures: a comparative study using EPR-spin trapping technique. Free
Radical Res. 40(8):821–5.
Payne, E.K., N.L. Rosi, C. Xue, and C.A. Mirkin, 2005. Sacrificial biological templates for the
formation of nanostructured metallic microshells. Angew. Chemie Ind Ed. 44: 5064-
5067.
Perebeinos, V., S.-W. Chan, F. Zhang, 2002. Madelung-Model Prediction for the Lattice
Constant Scaling with the Size of Ionic Nanocrystals of CeO2 and BaTiO3. Solid State
Commun, 123: 295.
Pethkar, A. V. Paknikar, K. M. 1998. Recovery of gold from solutions using Cladosporium
cladosporioides biomass beads. J. Biotechnol. 63, 121-136.
Pfeiffer, C. and Palinska, K.A. 2002. Characterization of marine Phormidium isolates -
conformity between molecular and ecophysiological results. Journal Of Plant
Physiology., 159 (6): 591-598.
Pickeet, N.L., D.F. Foster, D.J. Cole Hamilton. 1996. Syntheses of Semiconductor
Nanoparticles Using Single-Molecular Precursors J. Mater. Chem. 6 : 507.
Pickett-Heaps,J., Schmid,A.M.M and Edgar,L.A. 1990. The cell biology of diatom valve formation. In Progress in Phycological Research, ed. FE Round, DJ Chapman, 7:1–
169.
Piechula, S., Waleron, K., S ´ wiatek, W., Biedrzycka, I. and Podhajska, A.J. 2001. Mesophilic
cyanobacteria producing thermophilic restriction endonucleases. FEMS Microb Letter
198, 135–140.

149
Pilot, T.J., and Fox, J.L. 1984. Cloning and sequencing of the genes encoding the α and β
subunits of C-phycocyanin from the cyanobacterium Agmenellum quadruplilcatum.
Proc. natl. Acad. Sci. USA., 81: 6983–6987.
Pinzon-Gamez, N.M., Sundaram, S. and Ju, L.K. 2005. Heterocyst differentiation and H2
production in N2-fixing cyanobacteria. In AIChE Annual Meeting, Conference
Proceedings. Cincinnati, OH. 8949–8951
Planiselvam, V. 1998. Epiphytic cyanobacteria of mangroves: ecological, physiological and
biochemical studies and their utility as biofertilizers and shrimp feed. Ph.D thesis.
Annamalai University. India.
Plavsic, M., Terzic, S., Ahel, M. and van den Berg, C.M.G. 2004. Folic acid in coastal waters
of the Adriatic Sea. Mar Freshw Res 53, 1245–1252
Poole,C.P and Owens,F.J. 2003. Introduction to Nanotechnology. Hoboken, NJ:Wiley-Intersci.
Postgate, J. 1987. Nitrogen Fixation. Edward Arnold Publishing, London. 77.
Poulsen, N., Sumper,M and Kroger,N. 2003. Biosilica formation in diatoms: characterization of
native silaffin-2 and its role in silica morphogenesis. Proc. Natl. Acad. Sci. USA
100:12075-80.
Poulsen,N and Kroger,N. 2004. Silica morphogenesis by alternative processing of silaffins in
the diatom Thalassiosira pseudonana. J. Biol. Chem. 279: 42993-99.
Poulsen,N and Kroger,N. 2005.A new molecular tool for transgenic diatoms: Control of mRNA
and protein biosynthesis by an inducible promoter-terminator cassette. FEBS J. 272:
3413-3423.
Poulsen,N., Berne,C., Spain,J and Kroger,N. 2007. Silica immobilization of an enzyme
through genetic engineering of the diatom Thalassiosira pseudonana. Ang. Chem. Int.
Ed. 46:1843–46.
Poulsen,N.C., Spector,I., Spurck,T.P., Schultz,T.F and Wetherbee,R. 1999. Diatom gliding is
the result of an actin-myosin motility system. Cell Motil. Cytoskelet. 44:23–33.
Prasad, K and Anal K. Jha. 2010. Biosynthesis of CdS nanoparticles: An improved green and
rapid procedure. J. Col. Inter. Sci. 342 : 68-72.
Premanandh, J., Priya, B., Teneva, I., Dzhambazov, B., Prabaharan, D. and Uma L. 2006. Molecular Characterization of Marine Cyanobacteria from the Indian Subcontinent
Deduced from Sequence Analysis of the Phycocyanin Operon (cpcB-IGS-cpcA) and
16S-23S ITS Region. The J Microbiol: 44(6): 607-616.
Pulz, O and Gross, 2004. Valuable products from biotechnology of microalgae. Appl.
Microbiol. Biotechnol. 65:635-648.

150
Qin, T., T. Gutu, J. Jiao, C-H. Chang, G.L. Rorrer, 2008. Photoluminescence of silica
nanostructures from bioreactor culture of marine diatom Nitzschia frustulum. J.
Nanosci. Nanotechnol. 8: 2392.
Rai, A.N. 1990 CRC Handbook of Symbiotic Cyanobacteria. CRC Press, Boca Raton, 253 pp.
Rajeev, K.J. and Xu, Z. 2004. Biomedical compounds from marine organisms. Mar Drugs 2,
123–146.
Rao,C.N.R.., Kulkarni,G.U., Thomas,P.J and Edwards,P.P.2002. Size-dependent chemistry:
Properties of nanocrystals. Chem. Eur. J. 8:28-35.
Rasmussen, U., Svenning, M.M. 1998. Fingerprinting of cyanobacteria based on PCR with
primers derived from short and long tandemly repeated repetitive sequences. Appl.
Environ. Microbiol., 64: 265-272.
Raven, P.H., R.F. Evert and S.E. Eichhorn. 1992. Biology of Plants, 5th ed., Worth Publishers,
Inc., New York. Chapter 15, pp. 298-316.
Reddy, M.C., Subhashini, J. and Mabipal, S.V. 2003. C-Phycocyanin, a selective
cyclooxygenase-2 inhibitor, induces apoptosis in lipopolysaccharide-stimulated RAW
264.7 macrophages. Biochem biophys. Res Commun., 304: 385–392.
Reed, R.H., Chudek, J.A., Foster, R. and Stewart, W.D.P. 1984 Osmotic adjustment in
cyanobacteria. Arch. Microbiol., 138, 333-337.
Reeks, MW., Hall, D., 2001. Kinetic models for particle resuspension in turbulent flows: theory
and measurements. J Aerosol Sci., 32: 1-31.
Reich, D.H., Tanase., M. Hultgren, A., Bauer, L.A., Chen, C.S., Meyer, G.J. 2003. Biological
applications of multifunctional magnetic nanowires, J.Appl.Phys., 93: 7275-7280.
Reinfelder,J.R., Milligan,A.J and Morel,F.M. 2004. The role of the C4 pathway in carbon
accumulation and fixation in a marine diatom. Plant Physiol 135: 2106-2111.
Reith, F. Rogers, S. L.McPhail, D. C. Webb,D. 2006. Biomineralization of gold: Biofilms on
bacterioform gold. Science, 313: 233-236.
Remirez, D, Fernandez, V., Tapia, G et al. 2002. Influence of Cphycocyanin on hepatocellular
parameters related to liver oxidative stress and kupffer cell functioning. Inflamm Res
51:351–356.
Reshef, V., Mizrachi, E., Maretzki, T., Silberstein, C., Loya, S., Hizi, A. and Carmeli, S. 1997.
New acetylated known glycolipids from cyanobacteria with potential to inhibit the
reverse transcriptase of HIV-1. J Nat Prod 60: 1251–1260
Rigbi, M., Rosinski, J., Siegelman, H.W and J.C. Sutherland. 1980. Cyanobacterial
Phycobilisomes: selective dissociation monitored by fluorescence and circular
dichroism Proc. Natl. Acad. Sci., 77:1961–1965.

151
Rippka, R. 1988 Recognition and identification of cyanobacteria. In: L. Packer and A.N. Glazer
[Eds] Cyanobacteria. Methods in Enzymology, Volume 167, Academic Press, New
York, 28-67.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M. and Stanier, R.Y. 1979. Generic
assignments, strains histories and properties of pure cultures of cyanobacteria. J. Gen.
Microbiol., 111: 1-61.
Ritchie, R.J., Trautman, D.A., and Larkum, A.W.D. 1997. Phosphate uptake in the
cyanobacterium Synechococcus R-2 PCC 7942. Plant and Cell Physiology., 38: 1232-
1241.
Rizvi, S.J.H. and Rizvi, V. 1992 Allelopathy. Basic and Applied Aspects. Chapman & Hall,
London, 480.
Robertson, B. R., Tezuka, N. and Watanabe, M.M. 2001. Phylogenetic analyses of
Synechococcus strains (cyanobacteria) using sequences of 16S rDNA and part of the
phycocyanin operon reveal multiple evolutionary lines and reflect phycobilin content.
Int. J. Syst. Evol. Microbiol., 51: 861-871.
Robinson, R. D., J.E. Spanier, F. Zhang, S.-W. Chan, I. P. Herman, 2002. Visible thermal
emission from sub-band-gap laser excited cerium dioxide particles. J. Appl. Phys, 92,
1936.
Rodriguez, P., N. Munoz-Aguirre, E. S-M. Martinez, G. G. Cruz, S. A. Tomas and O. Z. Angel.
2008b. Synthesis and spectral properties of starch capped CdS nanoparticles in aqueous
solution. J. Cryst. Growth 310: 160-164.
Rodriguez, P., N. Munoz-Aguirre, E. S-M. Martinez, G. Gonzalez, O. Zelaya and J. Mendoza.
2008a. Formation of CdS nanoparticles using starch as capping agent. App. Surf. Sci.
225 740-742.
Roh, Y., Lauf, R.J., McMillan, A.D., Zhang, C., Rawn, C.J., Bai, J., Phelps, T.J. 2001.
Microbial synthesis and the characterization of metal-substituted magnetites, Solid
State Commun., 118: 529-534.
Roman, B.R., Alvarez-Pez, J.M., Fernandez, A et al. 2002. Recovery of pure B- Phycoerythrin
from the microalga Porphyridium cruentum. J Biotechnol 93:73–85
Romay C, Ledon N, Gonzalez R., 1998(b). Further studies on anti-inflammatory activity of
phycocyanin in some animal models of inflammation. Inflamm Res;47:334- 338.
Romay, C., Armesto, J., Remirez, D., Gonzalez, R., Ledon, N and I. Garcia. 1998a. Antioxidant
and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm
Res. 47:36–41.
Romay, C., Delgado, R., Remirez, D., Gonzalez, R and A. Rojas. 2001. Effects of phycocyanin
extract on tumor necrosis factor-a and nitrite levels in serum of mice treated with
endotoxin. Arzneim Forsh/Drug Res. 51: 733–6.

152
Romay, Ch., González, R., Ledon, N., Remirez, D and Rimbau, V., 2003. C-phycocyanin A
Biliprolein with antioxidant, ant-inflammatory and neuroprotective effects. Current
Protein Peptide Sci 4:207–216.
Romero-Gonzalez, M. E.; Williams, C. J.; Gardiner, P. H. E.;Gurman, S. J.; Habesh, S. 2003.
Spectroscopic Studies of the Biosorption of Gold(III) by Dealginated Seaweed Waste.
Environ. Sci. Technol. 37: 4163-4169.
Rorrer Gregory, L., Chang, Chih-Hung, Liu Shu-Hong, Jeffryes Clayton , Jiao Jun, Hedberg
and James A. 2005. Biosynthesis of Silicon–Germanium Oxide Nanocomposites by the
Marine Diatom Nitzschia frustulum. J Nanosci. Nanotech.5: 41-49
Rosi, N.L., C. A. Mirkin, 2005. Nanostructures in biodiagnostics. Chem. Rev. 105: 1547.
Rosi, N.L., C.S. Thaxton, and C.A. Mirkin, 2004. Control of nanoparticle assembly by using
DNA-modified diatom templates. Angew. Chemie 43, 5500-5503.
Round F.E., Crawford R.M., and Mann D.G., 1990. The diatoms: Biology and morphology of
the genera. Cambridge University Press, Bath. pp.747.
Round, F.E., Crawford, R.M and Mann, D.G. 1990. The diatoms. Biology and Morphology of
the genera. Cambridge University Press, Cambridge, U.K. .747.
Roy, I., Ohulchanskyy, T.Y., Pudavar, H.E., Bergey, E.J., Oseroff, A.R., Morgan, J.,
Dougherty, I.J., Prasad, P.N. 2003. Ceramic based nanoparticles entrapping water-
insoluble photosensitizing anticancer drugs: novel drug-carrier system for
photodynamic therapy, J.Am.Chem.Soc., 25: 7860-7865.
Rudi, K., Skulberg, O.M., Larsen, F. and Jakobsen, K.S. 1997 Strain characterization and
classification of oxyphotobacteria in clone cultures on the basis of 16S rRNA
sequences from the variable regions V6, V7 and V8. Appl. Environ. Microbiol., 63(7),
2593-2599.
Russel AD, Hugo WB. 1994. Antimicrobial activity and action of silver. Prog Med Chem
31:351- 70.
Sadiq, I. Pakrashi, Sunandan Chandrasekaran, N Mukherjee, Amitava. 2011. Studies on
toxicity of aluminum oxide (Al2O3) nanoparticles to microalgae
species: Scenedesmus sp. and Chlorella sp. J. Nanopart. Res.,13 (8): 3287-3299.
Sakthivel, K. 2004. Studies on marine cyanobacteria from mangrove environment. Ph.D thesis. Annamalai University. India.
Samsonoff, W.Aand R. MacColl. 2001. Biliproteins and phycobilisomes from cyanobacteria
and red algae at the extremes of habitat. Arch Microbiol. 7:1–10.
Sandgren, C.D. (ed) 1988. Growth and reproductive strategies of freshwater phytoplankton:
Cambridge University Press.

153
Sandhage K.H., Dickerson M.B., Huseman P.M., Caranna M.A., Clifton J.D., Bull T.A.,
Heibel T.J., Overton W.R., and Schoenwaelder M.E.A., 2002. Novel, bioclastic route
to self-assembled, 3D, chemically tailored meso/nanostructures: Shape- preserving
reactive conversion of biosilica (diatom) microshells. Adv. Mater. 14, 429-433.
Sandhage, K.H., S.M. Allan, M.B. Dickerson, C.S. Gaddis, S. Shian, M.R. Weatherspoon, Y.
Cai, G. Ahmad, M.S. Haluska, R.L. Snyder, R.R. Unocic, F.M. Zalar, Y. Zhang, R.A.
Rapp, M. Hildebrand, and B.P. Palenik, 2005. Merging biological self-assembly with
synthetic chemical tailoring: The potential for 3-D genetically-engineered micro/nano-
devices (3-D GEMS). Int. J. Appl. Ceram. Tech. 2, 317-326.
Santos MC, Noyalo PT, Olvera RR et al. 2004. Extraction and purification of phycocyanin
from Calothrix spp. Process Biochem 39(12):2047–2052.
Sarada, R., Pillai, M. G. and G.A. Ravishankar. 1999. Phycocyanin from Spirulina sp.:
Influence of Processing of Biomass on Phycocyanin Yield, Analysis of Efficacy of
Extraction Methods and Stability Studies on Phycocyanin. Process Biochem. 34(8):
795.
Sardar, R., A. M. Funston, P. Mulvaney, R. W. Murray, 2009. Gold Nanoparticles: Past,
Present, and Future. Langmuir. 25 (24): 13840.
Sastry, M., Ahmad, A., Islam, NI and Kumar, R. 2003. Biosynthesis of metal nanoparticles
using fungi and actinomycete. Current Sci., 85:162-170.
Sastry, M., Ahmad, A., Khan, M.I., Kumar, R. 2004. Microbial nanoparticle production,
Nanotechnology, Wiley-VCH, Weinheim, Germany, 126-135.
Sato, M., Masuda, Y., Kirimura, K. and Kino, K. 2007. Thermostable ATP regeneration system
using polyphosphate kinase from Thermosynechococcus elongatus BP-1 for d-amino
acid dipeptide synthesis. J Biosci Bioeng 103, 179–184.
Sau, T. K., A. L. Rogach, F. Jackel, T. A. Klar, J. Feldmann, 2010. Properties and Applications
of Colloidal Nonspherical Noble Metal Nanoparticles. Adv. Mater.22 (16), 1805.
Scarano G, Morelli E. 2003. Properties of phytochelatin-coated CdS nanocrystallites formed in
a marine phytoplanktonic alga (Phaeodactylum tricornutum, Bohlin) in response to Cd.
Plant Sci 165:803–810.
Schabes-Retchkiman PS, Canizal G, Herrera-Becerra R, Zorrilla C, Liu HB, Ascencio JA.
2006. Biosynthesis and characterization of Ti/Ni bimetallic nanoparticles. Optic.
Mater. 29(1): 95-99.
Schneider, J., Hock, N., Weimer, S., Borrmann, S. 2005. Nucleation particles in diesel exhaust:
Composition inferred from in situ mass spectrometric analysis. Environ Sci and Tech.,
35: 6153-6161.

154
Schopf, J.W. 1996 Cyanobacteria. Pioneers of the early Earth. In: A.K.S.K, Prasad, J.A.
Nienow and V.N.R Rao [Eds] Contributions in Phycology. Nova Hedwigia, Beiheft
112, J. Cramer, Berlin, 13-32.
Schweiz, Band IX, Heft 3. Kommissionsverlag Buchdruckerei Büchler and Co., Bern.
Sekar, S and M. Chandramohan, 2008. Phycobiliproteins as a commodity: trends in applied
research, patents and commercialization. J. Appl. Phycol. 20: 113–136.
Senapati, S., Mandal, D., Ahmad, A., Khan, M.I., Sastry, M., Kumar, R. 2004. Fungus
mediated synthesis of silver nanoparticles: a novel biological approach, Indian J.Phys.,
78:101-105.
Shahverdi AR, Fakhimi A, Shaverdi HR, Minaian S. 2007. Synthesis and effect of silver
nanoparticles on the antibacterial activity of different antibiotics against
Staphylococcus aureus and Escherichia coli. Nanomedcine. 3: 168-71.
Shankar S, Ahmad A, Sastry M. 2003. Geranium Leaf Assisted Biosynthesis of Silver
Nanoparticles. Biotechnol. Prog. 19:1627.
Shankar SS, Ahmad A, and Sastry M. 2003. Geranium leaf assisted biosynthesis of silver
nanoparticles. Biotechnol Prog. 19(6):1627-31
Shankar SS, Rai A, Ahmad A, Sastry M. 2004. Rapid synthesis of Au, Ag, and bimetallic Au
core-Ag shell nanoparticles using Neem (Azadirachta indica) leaf broth. J Colloid
Interface Sci.. 275(2):496-502
Shankar, SS., Ahmad, A., Pasrichaa, R., Sastry, M. 2003a. Bioreduction of chloroaurate ions by
geranium leaves and its endophytic fungus yields gold nanoparticles of different
shapes. J Mater Chem 13:1822–1826.
Shanker SS, Rai A, Ankamwar B, Singh A, Ahmed A, Sastry M. 2004. Biological synthesis of
triangular gold nanoprisms. Nat. Mater. 3: 482.
Shanker, S.S., A. Rai, B. Ankamwar, A. Singh, A. Ahmed, M. Sastry . 2004. Biological
synthesis of triangular gold nanoprisms. Nat. Mater., 3 :482 – 488.
Sharma, V., Park, K. and Srinivasarao, M. 2009. Colloidal dispersion of gold nanorods:
historical background, optical properties, seed-mediated synthesis, shape separation and
self-assembly. Mater. Sci. Engin. R, 65: 1–3.
Shih, S.R., Tsai, K.N., Li, Y.S et al. 2003. Inhibition of enterovirus 71- induced apoptosis by
Allophycocyanin isolated from a gluegreen alga Spirulina platensis. J Med Virol.
70:119–125.
Shimizu, Y. 2003. Microalgal metabolites. Curr Opin Microbiol 6, 236–243
Shipway, A. N.; Katz, E.; Willner, I. 2000. Nanoparticle Arrays on Surfaces for Electronic,
Optical, and Sensor Applications. Chem Phys Che. 1: 18.

155
Shrivastav, A., S.K. Mishra, S. Mishra. 2010. Polyhydroxyalkanoate (PHA) synthesis
by Spirulina subsalsa from Gujarat coast of India. Int. J. Biol. Macromol. 46:255–260.
Shrivastava S, Bera T, Roy A, Sing G, Ramachandrarao P, Dash D. 2007. Characterization of
enhaced antibacterial effects of novel silver nanoparticles. Nanotech. 18:225103
Sidler, W.A. 1994. Phycobilisome and phycobiliproteins structures. In: Bryant DA (ed) The
molecular biology of cyanobacteria. Kluwer, Netherlands, pp 139–127
Siegelman, H.W and J.H. Kycia. 1978. Algal biliproteins. In: Hellebust JA, Craige JS (ed)
Hand book of phycological methods – physiological and biochemical methods.
Cambridge University Press, Cambridge, pp 72–79
Siever,R. 1991. Silica in the oceans: Biological- Geochemical interplay. In: Scientists on Gaia
(Stephen H. Schneider and Penelope J.Boston, eds), MIT Press, Cambridge, Ma.287-
295.
Silveira, S. T., Burkert, J. F. M., Costa, J. A. V., Burkert, C. A. V. and S.J. Kalil. 2007.
Optimization of Phycocyanin Extraction from Spirulina platensis Using Factorial
Design. Biores. Technol. 98 (8): 1629.
Skulberg, O.M. 1996a Terrestrial and limnic algae and cyanobacteria In: A. Elvebakk and P.
Prestrud [Eds] A Catalogue of Svalbard Plants, Fungi, Algae and Cyanobacteria. Part
9, Norsk Polarinstitutt Skrifter 198: 383-395.
Slawson, R.W., Van Dyke, M.I., Trevors, I.T., Lee, H. 1992. Germanium and silver resistance-
Accumulation & toxicity in microorganisms, Plasmid, 27:72-79.
Smoker, J.A., and Barnum, S.R. 1988. Rapid small-scale DNA isolation from filamentous
cyanobacteria, FEMS Microbiolo Lett. 56(1): 119 – 122.
Sneath, P.H.A. 1992 International Code of Nomenclature of Bacteria, 1990 Revision. American
Society for Microbiology, Washington, D.C.
Sohn, K., F. Kim, K. C. Pradel, J. S. Wu, Y. Peng, F. M. Zhou, J. X. Huang, 2009. Construction
of Evolutionary Tree for Morphological Engineering of Nanoparticles. ACS Nano, 3
(8), 2191.
Sondi I, Salopek-Sondi B.2004. Silver nanoparticles as antimicrobial agent: a case study on E.
coli as a model for Gram-negative bacteria.J Colloid Interface Sci. 275:177 - 82.
Soni, B, Kalavadia B, Trivedi U and D. Madamwar. 2006. Extraction purification
characterization of phycocyanin from Oscillatoria quadripunctula isolated from the
rockyshores of Bet Dwarka Gujarat India. Process Biochem. 41:2017-2023.
Soni, B, U. Trivedi, D. Madamwar. 2008. A Novel Method of Single Step Hydrophobic
Interaction Chromatography for the Purification of Phycocyanin from Phormidium
fragile and its Characterization for Antioxidant Property. Bioresour. Technol. 99 (1):
188–194.

156
Soni, B., N. P. Visavadiya, N. Dalwadi, D. Madamwar, C. Winder and C. Khalil, 2010. Purified
c-phycoerythrin: safety studies in rats and protective role against permanganate-
mediated fibroblast-DNA damage. J. App. Toxicol. 30 : 542-550.
Spolaore, P., Cassan, C.J, Duran, E et al. 2006. Commercial application of microalgae. J Biosci
Bioeng 101(2):87–96.
Stal, L.J. 1995. Physiological ecology of cyanobacteria in microbial mats and other
communities. New Phytol 131: 1–32.
Stal, L.J. and Moezelaar, R. 1997. Fermentation in cyanobacteria. FEMS Microbiol Rev 21:
179–211.
Stanier, R. Y., Sistrom, W.R., Hansen, T.A., Whitton, B.A., Castenholz, R.W., Pfennig, N.,
Gorlenko, V.N., Kondratieva, E.N., Eimhjellen, E., Whittenbury, R., Gherna, R.L.,
Trüper, H.G. 1978. Proposal to place the nomenclature of the cyanobacteria blue-green
algae under the rules of the International Code of Nomenclature of Bacteria. Int. J.
Syst. Bacteriol., 28: 335-336.
Stanier, R.Y., Kunisawa, R., Mandel, M.D and G. Cohen-Bazire. 1971. Purification and properties of unicellular blue green algae (order chroococcales). Bacterial
Reviews 35:171
Stec, B., Troxler, R.F and M.M. Teeter. 1999. Crystal structure of C-phycocyanin from
Cyanidium caldarium provides a new perspective on phycobilisome assembly.
Biochem Biophys Res Commun. 76:2912–21.
Steinbuchel, A., Fuchtenbusch, B., Gorenflo, V., Hein, S., Jossek, R., Langenbach, S. and
Rehm, B.H.A. 1997. Biosynthesis of polyesters in bacteria and recombinant organisms.
Polymer Degrad Stabil 59, 177–182.
Stewart, D.E and F.H.Farmer. 1984. Extraction identification and quantification of
phycobiliprotein pigment from phototrophic plankton. Limnol. Oceonogr. 29: 392-
397.
Stewart, W.D.P. 1973 Nitrogen fixation by photosynthetic microorganisms. Ann. Rev.
Microbiol., 27,283-316.
Sugawara, H., Ma, J., Miyazaki, S., Shimura, J. and Takishima, Y. [Eds] 1993 World Directory
of Collections of Cultures of Microorganisms. ISBN 4-930813-56-5, 1152 pp.
Suggett, D.J., Maberly, S.C and R.J. Geider. 2006a. Gross photosynthesis and lake community
metabolism during the spring phytoplankton bloom. Limnol. Oceanogra., 51: 2064-
2076.
Suggett, D.J., Moore, C.M., Maranon, E., Omachi, C., Varela, R.A., Aiken, J and P.M.
Holligan. 2006b. Photosynthetic electron turn over in the tropical and subtropical
atlantic ocean. Deep Sea Res. II. 53: 1573-1592.

157
Sullivan,C.W and Volcani,B.E. 1981. Silicon in the cellular metabolism of diatoms. New York:
Springer Verlag pp. 15–42.
Sultemeyer, D., Klughammer, B., Badger, M.R and G.D. Price. 1998. Fast induction of high
affinity HCO3- transport in cyanobacteria. Plant Physiol. 116: 183-192.
Summons, R.E., Jahnke, L.L., Hope, J.M. and Logan G.A. 1999. 2-Methylhopanoids as
biomarkers for cyanobacterial oxygenic photosynthesis. Nature. 400 (6744): 554-557.
Sumper, M., E. Brunner, 2006. Learning from Diatoms: Nature’s Tools for the Production of
Nanostructured Silica. Adv. Funct. Mater. 16: 17.
Sun,M., Qian,K. and Su,N. 2003. Foot-and-mouth disease virus VPl protein fused with cholera
toxin B subunit expressed in Chlamydomonas reinhardtii chloroplast. Biotechnol Lett
25:1087-1092.
Sun,Y and Xia,Y. 2002. Shape-controlled synthesis of gold and silver nanoparticles. Science
the diatom Thalassiosira pseudonana. J. Biol. Chem. 279:42993–99.
Susi, H. 1972. Infrared spectroscopy conformation, Methods Enzymol. 22: 455–472.
Suzuki, I., H. Kikuchi, S. Nakanishi, Y. Fujita, T. Sugiyama, T. Omata, 1995. A novel nitrite
reductase gene from the cyanobacterium Plectonema boryanum. J. Bacteriol, 177: 137.
Suzuki, K., Liu, H., Saino, T., Obata, H et al., 2002. East-West Gradients in the photosynthetis
potential of phytoplankton and iron concentration in the subarctic pacifi ocean during
early summer. Limnol. Oceanogra., 47: 1581-1594.
Sveshnikov, D.A., Sveshnikova, N.V., Rao, K.K. and Hall, D.O. 1997. Hydrogen metabolism
of mutant forms of Anabaena variabilis in continuous cultures and under nutritional
stress. FEBS Microbiol Lett 147, 297–301.
Sweeny, R.Y., Mao, C., Gao, X., Burt, J.L., Belcher, A.M., Georgiou, G., Iverson, B.L.
2004. Bacterial biosynthesis of cadmium sulfide nanocrystals, Chem.Biol., 11: 1553-
1559.
Takeshita, K., K. Fujii, K. Anzai, T. Ozawa, 2004. In vivo monitoring of hydroxyl radical
generation caused by x-ray irradiation of rats using the spin trapping/epr technique.
Free Radi. Biolo. Medicine, 36: 1134.
Talapin, D. V., J. S. Lee, M. V. Kovalenko, E. V. Shevchenko, 2010. Prospects of colloidal nanocrystals for electronic and optoelectronic applications. Chem. Rev. 2010, 110 (1),
389.
Tamura. K., Nei, M and S. Kumar. 2004. Prospects for inferring very large phylogenies by
using the neighbor-joining method. Proc. Natl. Acad. Sci., 101. 11030-11599.
Taton, A., Grubisic, S., Brambilla, E., Dewit, R. and Wilmotte, A. 2003. Cyanobacterial
diversity in natural and artificial microbial mats of Lake Fryxell (McMurdo dry valleys,

158
Antarctica): A morphological and molecular approach – Appl. Environ Microbiol. 69
(9): 5157-5169.
Telford WG, Moss MW, Moreseman JP et al. 2001b. Cyanobacterial stabilized phycobilisomes
as fluorochromes for extracellular antigen detection by flow cytometry. J Immunol
Meth 54:13–30.
Telford, W.G., Moss, M.W., Moreseman, JP et al. 2001a. Cryptomonad algal phycobiliproteins
as fluorochromes for extracellular and intracellular antigen detection by flow
cytometry. Cytometry. 44:16–23.
Temgire, M. K., S.S. Joshi, 2004. Optical and structural studies of silver nanoparticles. Rad
Phys, 71: 1039
Teneva, I., Mladenov, R., Popov, N. and Dzhambazov, B. 2005. Phylogenetic Characterization
Of Nostoc Linckia And Nostoc Punctiforme (Cyanoprokaryota) Based On Cpcb-Cpca
Gene Sequences. Proceedings Of The Balkan Scientific Conference Of Biology In
Plovdiv (Bulgaria)., 349–364
Thajuddin, N and G. Subramanian, 2005. Biodiversity and biotechnological potential application of cyanobacteria. Curr. Sci. 89: 47–57.
Thiel J, Pakstis L, Buzby S, Raffi M, Ni C, Pochan DJ, Ismat Shah S. Antibacterial Properties
of Silver-Doped Titania. Small. 2007; 3(5): 799.
Tindall, B. J., Kampfer, P., Euzéby, J.P. and Oren, A. 2006. Valid publication of names of
prokaryotes according to the rules of nomenclature: past, history and current practice.
Int. J. Syst. Evol. Microbiol.,56: 2715-2720.
Tiyaporn, Luangpipat, Isabel R Beattie, Yusuf Chisti, Richard G Haverkamp. 2011. Gold nanoparticles produced in a microalga. J. Nanopar. Res. 13(12):6439-6445
Tseng, C.K. 2001. Algal biotechnology industries and research activities in China. J Appl
Phycol. 13:375–80.
Turner. S. 1997. Molecular systematics of oxygenic photosynthetic bacteria – Plant Systematics
and Evolution Suppl., 117:13-52.
Ulf Karsten, Rhena Schumann, Susanne Rothe, Ines Jung and Linda Medlin. 2006.
Temperature and light requirements for growth of two diatom species
(Bacillariophyceae) isolated from an Arctic macroalga. Polar Biol 29: 476-486.
Unocic R.R., Zalar F.M., Sarosi P.M., Cai Y., and Sandhage K.H., 2004. Anatase assemblies
from algae: coupling biological self-assembly of 3-D nanoparticles. Chem Commu. 7:
796
Vadiraja, B.B., Gaikwad, N.W and K. M. Madyastha. 1998. Hepatoprotective effect of C-
phycocyanin: protection for carbon tetrachloride and R-(+)-pulegone-mediated
hepatotoxicity in rats. Biochem. Biophys. Res. Commun. 249: 428–431.

159
Van Landingham, S.L. 1982 Guide to the Identification, Environmental Requirements and
Pollution Tolerance of Freshwater Blue-Green Algae (Cyanophyta). United States
Environmental Protection Agency, Cincinnati, Ohio, 341 pp.
Vandamme, P., Pot, B., Gillis, M., De Vos, P., Kersters, K. and Swings, J. 1996 Polyphasic
taxonomy, a consensus approach to bacterial systematics. Microbiol. Rev., 60, 407-438.
Vigneshwaran, N., A.A. Kathe, P.V. Varatharajan, R. P. Nachane, R. H. Balasubramanya,
2007. Silver-protein (core-shell) nanoparticle production using spent mushroom
substrateLangumir 23: 7113.
Vigneshwaran, N., R P Nachane, R H Balasubramanya, P V Varadarajan. 2006. A novel one-
pot 'green' synthesis of stable silver nanoparticles using soluble starch. Carbohy. Res..
341(12): 2012-2018.
Volcani,B.E. 1981. Cell wall formation in diatoms: morphogenesis and biochemistry. New
York: Springer Verlag. 157-200
Volesky, B. 1990. Biosorption of Heavy Metals; CRC Press Inc.: Boca Raton, FL.
Vrieling,E.G., Beelen,T.P.M, Van Santen,R.A and Gieskes,W.W.C. 2000. Nanoscale
uniformity of pore architecture in diatomaceous silica, a combined small and wide
angle X-ray scaterring study. J. Phycol. 36: 146-159.
Vrieling,E.G., Gieskes,W.W.C and Beelen,T.P.M. 1999. Silicon deposition in diatoms: control
by the ph inside the silicon deposition vesicle. J.Phycol. 35: 548–559.
Waggoner, A. 2006. Fluorescent labels for proteomies and genomics. Curr. Opin. Chem Biol
10:62–66.
Walsby, A.E. 1978. The gas vesicles of aquatic prokaryotes. In: R.Y. Stanier, H.J. Rogers and
J.B. Ward [Eds] Relations Between Structure and Functioning in the Prokaryotic Cell.
28th Symposium of the Society for General Microbiology. Cambridge University
Press, 338 pp.
Wang L, Fie J, Chen H, Liang A, Qian B, Ling B, Zhou C. 2010. One-step aqueous synthesis of
CdS nanoparticles as a novel fluorescence probe for the ultrasensitive detection of
DNA. J Lumines.130: 845–850.
Ward, D.M., Santegoeds, C.M., Nold, S.C., Ramsing, N.B., Ferris, M.J. and Bateson, M.M.
1997. Biodiversity within hot spring microbial mat communities: molecular monitoring
of enrichment cultures. Anton. Van Leeuwen. 71: 143–150.
Warheit, D.B., 2004. Nanoparticles: health impacts? Mater. Today 32–35.
Waterbury, J.B. 1992 The cyanobacteria isolation, purification and identification. In: A.
Balows, H.G., M. Trüper, M. Dworkin, W. Harder and K.H. Schleifer [Eds] The
Prokaryotes. Second edition, Volume II, Springer-Verlag, New York, 2058-2078.

160
Weatherspoon, M.R., M.B. Dickerson, G. Wang, Y. Cai, S. Shian, C. Jones, S.R. Marder, and
K.H. Sandhage, 2007. Thin, conformal, and continuous SnO2 coatings on three-
dimensional biosilica templates through hydroxy-group amplification and layer-by-
layer alkoxide deposition. Angew. Chem, Int. Ed. 46, 5724-5727.
Weatherspoon, M.R., M.S. Haluska, Y. Cai, J.S. King, C.J. Summers, R.L. Snyder, and K.H.
Sandhage, 2006. Phosphor microparticles of controlled three-dimensional shape from
phytoplankton. J. Electrochem. Soc. 153: 34-37.
Weber, B., Wessels, D.C.J. and Büdel, B. 1996 Biology and ecology of cryptoendolithic
cyanobacteria of a sandstone outcrop in the Northern Province, South Africa. Arch.
Hydrobiol., Suppl. 117, Algolog. Stud. 83: 565-579.
Wedemayer, G.J., Wemmer, D.E., Glazer, A.N. 1991. Phycobilins Of cryptophycean algae -
structures of novel bilins with acrylolyl substituents from phycoerythrin. J Biol Chem
266(8):4731–4741.
Wei, Q., S-Z. Kang and J. Mu. 2004. Green synthesis of starch capped CdS nanoparticles.
Physiochem. Eng. A. 247: 125-127.
Weller, H., 1993. Colloidal Semiconductor Q-Particles: Chemistry in the Transition Region
Between Solid State and Molecules. Angew. Chem., Int. Ed. 32: 41.
Whitton, B.A. 1973 Freshwater plankton, In: G.E. Fogg, W.D.P. Stewart, P. Fay and A.E.
Walsby [Eds] The Blue-Green Algae. Academic Press, London, 353-367.
Whitton, B.A. 1992 Diversity, ecology and taxonomy of the cyanobacteria. In: N.H. Mann and
N.G. Carr [Eds] Photosynthetic Prokaryotes. Plenum Press, New York, 1-51.
WHO. 1999. Chapter 2. Cyanobacteria in the environment. In Toxic Cyanobacteria in Water: A guide to their public health consequences, monitoring and management. I. Chorus,
Bartram, J. (Ed). World Health Organization.
Wieser, U., Gaegauf, CK. 2000, Nanoparticle emissions of wood combustion processes. Centre
of Appropriate Technology and Social Ecology; Laboratories for sustainable Energy
Systems, Switcher land, 1st World conference and Exhibition on Biomass for Energy
and Industry.
Wille, N. 1904 Die Schizophyceen der Plankton Expedition. Ergebnisse der Plankton
Expedition der Humboldt-Stiftung. Bd. II. M.f. Verlag von Lipsius & Tischer, Kiel, 88
pp.
Wilmotte, A. 1994 Molecular evolution and taxonomy of the cyanobacteria. In: D.A.Bryant
[Ed.] The Molecular Biology of Cyanobacteria. Kluwer Academic Publishers,
Dordrecht, 1-25.
Xia, Y., Y. J. Xiong, B. Lim, S. E. Skrabalak, 2009. Shape-Controlled Synthesis of Metal
Nanocrystals: Simple Chemistry Meets Complex Physics? Angew. Chem., Int. Ed. 48
(1), 60.

161
Yong, P., Rowsen, N.A., Farr, J.P.G., Harri, I.R., Macaskie, L.E. 2002. Bioreduction &
biocrystallization of palladium by Desulfovibrio desulfuricans NCIMB 8307, Bioeng.,
80: 369-379.
Yoshida, J., Kobayashi, T., 1999. Intracellular hyperthermia for cancer using magnetite cationic
liposomes, J.Magn.Magn Mater., 94:176-184.
Yu, P., Li, J.R., and Cen., P.L. 2002. Cloning and sequencing of the phycocyanin gene from
Spirulina maxima and its evolutionary analysis. J. appl. Phycol., 14: 307–310.
Zehr, J.P., Mellon, M.T. and Hiorns, W.D. 1997. Phylogeny of cyanobacterial nifH genes:
Evolutionary implications and potential applications to natural assemblages -
Microbiology-UK., 143(4): 1443-1450.
Zhang, F., P. Wang, J. Koberstein, S. Khalid, S.-W. Chan, 2004. Cerium oxidation state in ceria
nanoparticles studied with X-ray photoelectron spectroscopy and absorption near edge
spectroscopy. Surf. Sci, 563: 74.
Zhang, F., S.-W. Chan, J.F. Spanier, F. Apak, Q. Jin, R. Robinson, I.P. Herman, 2002. Cerium
oxide nanoparticles: size-selective formation and structure analysis. Appl. Phys. Lett,
80, 127
Zhang, S., 2003. Fabrication of novel biomaterials through molecular self-assembly. Nat.
Biotechnol. 21: 1171.
Zhao G, Stevens Jr SE. 1998. Multiple parameters for the comprehensive evaluation of the
susceptibility of Escherichia coli to the silver ion. Biometals. 11:27 - 32.
Zhao J.P., Gaddis C.S., Cai Y., and Sandhage K.H., 2005. Free-standing microscale structures
of nanocrystalline zirconia with biologically replicable three- dimensional shapes. J.
Mater. Res. 20: 282-287.
Zhiqiang, Q., Siegmann, K., Keller, A., Matter, U., Scherrer, L., Siegmann, HC. 2000.
Nanoparticle air pollution in major cities and its origin. Atmos. Environ. 34: 443-451.
Zhou, J.F., J. Ralston, R. Sedev, D. A. Beattie, 2009. Functionalized gold nanoparticles:
Synthesis, structure and colloid stability. J. Colloid Interface Sci. 331 (2): 251.
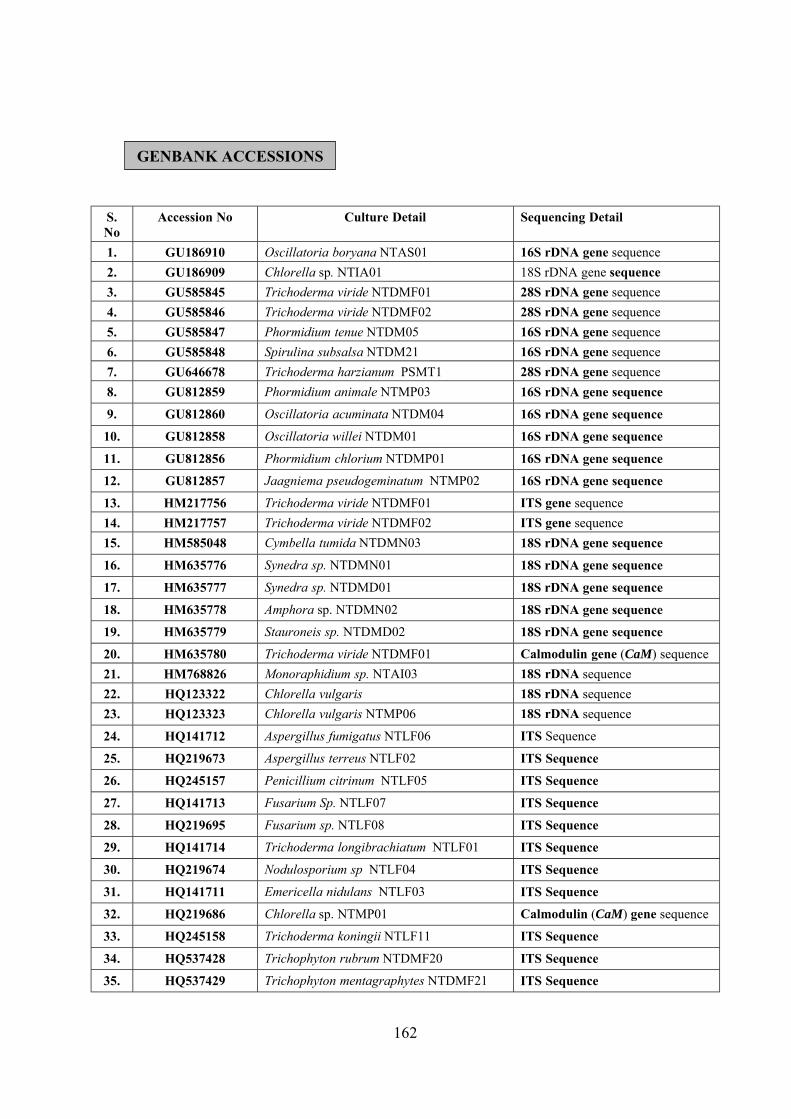
162
S. No
Accession No Culture Detail Sequencing Detail
1. GU186910 Oscillatoria boryana NTAS01 16S rDNA gene sequence
2. GU186909 Chlorella sp. NTIA01 18S rDNA gene sequence
3. GU585845 Trichoderma viride NTDMF01 28S rDNA gene sequence
4. GU585846 Trichoderma viride NTDMF02 28S rDNA gene sequence
5. GU585847 Phormidium tenue NTDM05 16S rDNA gene sequence
6. GU585848 Spirulina subsalsa NTDM21 16S rDNA gene sequence
7. GU646678 Trichoderma harzianum PSMT1 28S rDNA gene sequence
8. GU812859 Phormidium animale NTMP03 16S rDNA gene sequence
9. GU812860 Oscillatoria acuminata NTDM04 16S rDNA gene sequence
10. GU812858 Oscillatoria willei NTDM01 16S rDNA gene sequence
11. GU812856 Phormidium chlorium NTDMP01 16S rDNA gene sequence
12. GU812857 Jaagniema pseudogeminatum NTMP02 16S rDNA gene sequence
13. HM217756 Trichoderma viride NTDMF01 ITS gene sequence
14. HM217757 Trichoderma viride NTDMF02 ITS gene sequence
15. HM585048 Cymbella tumida NTDMN03 18S rDNA gene sequence
16. HM635776 Synedra sp. NTDMN01 18S rDNA gene sequence
17. HM635777 Synedra sp. NTDMD01 18S rDNA gene sequence
18. HM635778 Amphora sp. NTDMN02 18S rDNA gene sequence
19. HM635779 Stauroneis sp. NTDMD02 18S rDNA gene sequence
20. HM635780 Trichoderma viride NTDMF01 Calmodulin gene (CaM) sequence
21. HM768826 Monoraphidium sp. NTAI03 18S rDNA sequence
22. HQ123322 Chlorella vulgaris 18S rDNA sequence
23. HQ123323 Chlorella vulgaris NTMP06 18S rDNA sequence
24. HQ141712 Aspergillus fumigatus NTLF06 ITS Sequence
25. HQ219673 Aspergillus terreus NTLF02 ITS Sequence
26. HQ245157 Penicillium citrinum NTLF05 ITS Sequence
27. HQ141713 Fusarium Sp. NTLF07 ITS Sequence
28. HQ219695 Fusarium sp. NTLF08 ITS Sequence
29. HQ141714 Trichoderma longibrachiatum NTLF01 ITS Sequence
30. HQ219674 Nodulosporium sp NTLF04 ITS Sequence
31. HQ141711 Emericella nidulans NTLF03 ITS Sequence
32. HQ219686 Chlorella sp. NTMP01 Calmodulin (CaM) gene sequence
33. HQ245158 Trichoderma koningii NTLF11 ITS Sequence
34. HQ537428 Trichophyton rubrum NTDMF20 ITS Sequence
35. HQ537429 Trichophyton mentagraphytes NTDMF21 ITS Sequence
GENBANK ACCESSIONS
163
36. HQ537430 Epidermophyton floccosum NTDMF22 ITS Sequence
37. ADR65079 Chlorella sp. NTMP01 Calmodulin (CaM) protein sequence
38. JF421687 Oscillatoria acuminata NTDM04 Phycocyanin gene Sequence
39. JF421685 Phormidium tenue NTDM05 Phycocyanin gene Sequence
40. JF421686 Phormidium animale NTMP03 Phycocyanin gene Sequence
41. Bankit No 1431047 Oscillatoria salina Phycocyanin gene Sequence
42. Bankit No 1431046 Oscillatoria chlorina Phycocyanin gene Sequence
43. Bankit No 1421025 Spirulina subsalsa NTDM21 Phycocyanin gene Sequence
44. Bankit No 1448107 Phormidium tenue NTSM01 16S rDNA gene sequence
45. JN402964 Phormidium tenue NTSM02 16S rDNA gene sequence
46. Bankit No 1448113 Aspergillus sydowii NTSMP01 ITS Sequence
47. Bankit No 1448113 Penicillium cinnamopurpureum NTSMP02
ITS Sequence
48. Bankit No 1448113 Cladosporium sp. ITS Sequence
49. JN402974 Anabaena variabilis NTVMK01 16S rDNA gene sequence
50. JN402975 Anabaena sp. NTVMK02 16S rDNA gene sequence
51. JN402976 Westiellopsis Sp. NTVMK03 16S rDNA gene sequence
52. JN402980 Nocardiopsis sp. NTRJM01 16S rDNA gene sequence
53. JN402977 Nocardiopsis sp. NTRJM03 16S rDNA gene sequence
54. JN402979 Streptomyces sp. NTRJM05 16S rDNA gene sequence
55. JN216833 Aspergillus niger NTGMP01 ITS Sequence
56. JN216834 Aspergillus fumigatus NTGMP02 ITS Sequence
57. JN216835 Penicillium sp. NTGMP03 ITS Sequence
58. JN216836 Fusarium sp. NTGMP04 ITS Sequence
59. JN216837 Aspergillus terreus NTGMP05 ITS Sequence
60. JN66080 Trichoderma viride NTSA01 ITS sequence
61. JN66081 Trichoderma viride NTSA02 ITS sequence
62. JN66082 Trichoderma viride NTSA03 ITS sequence
63. JN402978 Rhodococcus sp NTRJM04 16S rDNA gene sequence
64. JN402981 Rhodococcus sp NTRJM04 16S rDNA gene sequence
65. JN402965 Poterioochromonas sp. NTNV01 18S rDNA gene sequence
66. JN402966 Poterioochromonas sp. NTNV02 18S rDNA gene sequence
67. JN402967 Poterioochromonas sp. NTNV03 18S rDNA gene sequence
68. JN402968 Poterioochromonas sp. NTNV04 18S rDNA gene sequence
69. JN402969 Poterioochromonas sp. NTNV07 18S rDNA gene sequence
70. JN402970 Botrycoccus sp NTNV08 18S rDNA gene sequence
71. JN402971 Hartmannella vermiformis NTNV09 18S rDNA gene sequence
72. JN402972 Poterioochromonas sp. NTNV04 18S rDNA gene sequence
73. JN402973 Poterioochromonas sp. NTNV05 18S rDNA gene sequence
164
74. JN601431 Staphyloccocus aureus NTMUR01 16S rDNA gene sequence
75. BankIt 1474793 Klebsiella pneumoniae NTMUR02 16S rDNA gene sequence
76. BankIt 1474793 Moraxella cattarhalis NTMUR03 16S rDNA gene sequence
77. JN601432 Candida albicans NTMUR04 18S rDNA gene sequence
78. JQ731679 Coelastrella sp. NTEB01 18S rDNA gene sequence
79. JQ796863 Botryococcus sp. NTAI06 18S rDNA gene sequence
80. JQ796861 Acharochaet rupens NTAI07 18S rDNA gene sequence
81. JQ796860 Tetradesmus sp NTAI05 18S rDNA gene sequence
82. JQ796862 Tetradesmus sp NTAI04 18S rDNA gene sequence
83. JQ867392 Leptolynbya valderiana NTDM10 16S rDNA gene sequence
84. JQ867393 Oscillatoria boryana NTDM11 16S rDNA gene sequence
85. JQ867394 Planktothrix pseudoadardhii NTRD01 16S rDNA gene sequence
http://www.ncbi.nlm.nih.gov/

Digest Journal of Nanomaterials and Biostructures Vol. 6, No 2, April-June 2011, p. 385-390
BIOSYNTHESIS AND CHARACTERIZATION OF SILVER NANOPARTICLES USING MARINE CYANOBACTERIUM, OSCILLATORIA WILLEI NTDM01
D. MUBARAK ALI., M. SASIKALA., M. GUNASEKARAN, N. THAJUDDIN* Department of Microbiology, School of Life Sciences, Bharathidasan University, Tiruchirappalli 620024, India Department of Biology, Fisk University, Nashville, TN 37208, USA Pioneering of reliable and eco-friendly process for synthesis of metallic nanoparticles biologically is an important step in the field of application of nanobiotechnology. This paper reports the extracellular biosynthesis of silver nanoparticles using marine cyanobacterium, Oscillatoria willei NTDM01 which reduces silver ions and stabilizes the silver nanoparticles by a secreted protein. The silver nitrate solution incubated with washed marine cyanobacteria changed to a yellow color from 72h onwards, indicating the formation of silver nanoparticles. The characteristics of the protein shell at 265nm were observed in Ultra violet spectrum for the silver nanoparticles in solution. While Fourier Transform Infra Red (FTIR) confirmed the presence of a protein shell which are responsible for the nanoparticles biosynthesis. Scanning Electron Microscopy (SEM) studies showed that the formation of agglomerated silver nanoparticles due to the capping agent in the range of 100 - 200nm. EDS spectrum of the silver nanoparticles was confirmed the presence of elemental silver signal in high percentage. Apart from Eco-friendliness and easy availability and low cost cyanobacterial biomass production will be more advantageous when compared to other classes of micro organism. Biosynthesis nanoparticles would have greater commercial viability if the nanoparticles could be synthesized more rapidly in the reaction vessel. (Received January 10, 2011; accepted February 2, 2011) Keywords: Marine cyanobacteria, silver nanoparticles, Oscillatoria willei NTDM01, FTIR, EDS
1. Introduction Nanotechnology is enabling technology that deals with nano-meter sized objects. It is
expected that nanotechnology will be developed at several levels: materials, devices and systems. Synthesis of nanoparticles using biological entities has great interest due to their unique shape dependent optical, electrical and chemical properties have potential application in nanobiotechnology. The synthesis and assembly of nanoparticle would benefit from the development of clean, nontoxic and environmentally acceptable "green chemistry" approaches for nanoparticles.
The biosynthesis of silver nanoparticles of different sizes, ranging from 1- 70nm, and shapes, including spherical, triangular and hexagonal has been conducted using bacteria, fungi, plant extracts (1-4). The mechanisms for the bioreduction of silver by bacteria involve reducing and other proteins in which sulphur and carboxylate group from cell wall (2-4). The silver nitrate caused the reduction of silver through a nitrate dependent reductase and synthesized the nanosized materials in Fusarium oxysporum (3). In this study, the formation of silver nanoparticles was investigated using silver nitrate in the presence of the marine cyanobacterium, Oscillatoria willei NTDM01. Marine cyanobacteria are one of the largest, photoautotrophic bacteria in marine ecosystem and are known to have high affinity transport system for nitrate.
*Corresponding author: [email protected]
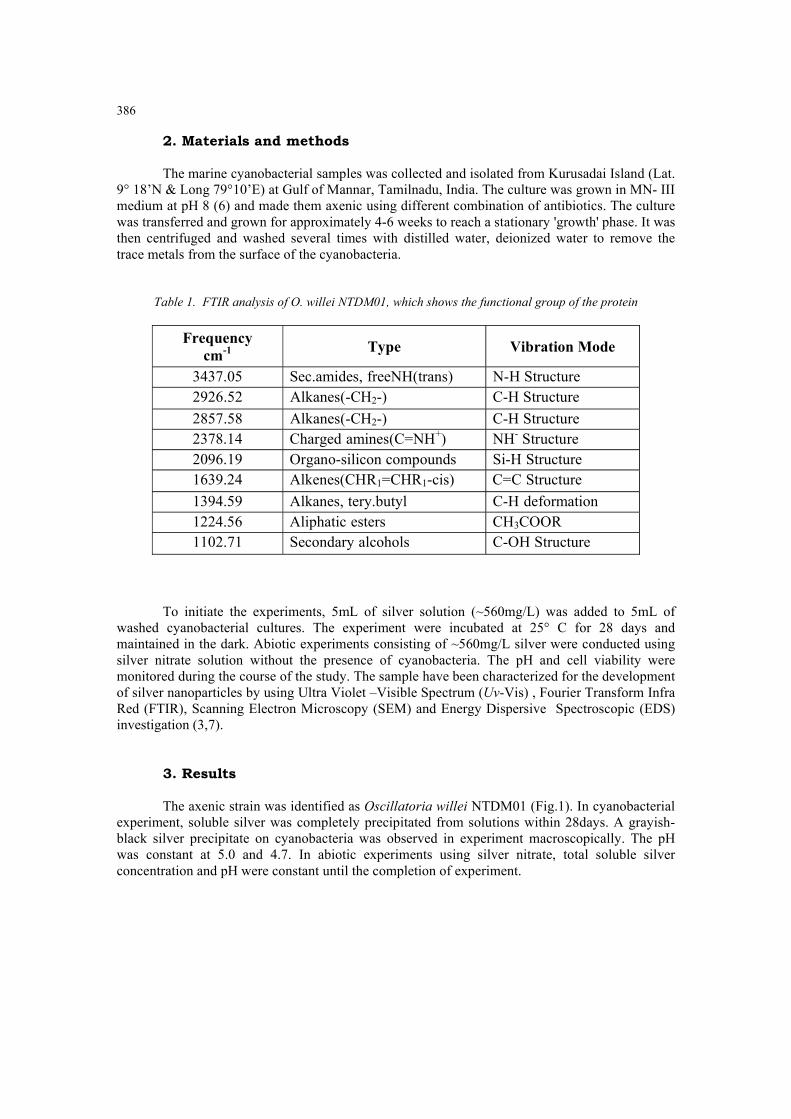
386
2. Materials and methods The marine cyanobacterial samples was collected and isolated from Kurusadai Island (Lat.
9° 18�N & Long 79°10�E) at Gulf of Mannar, Tamilnadu, India. The culture was grown in MN- III medium at pH 8 (6) and made them axenic using different combination of antibiotics. The culture was transferred and grown for approximately 4-6 weeks to reach a stationary 'growth' phase. It was then centrifuged and washed several times with distilled water, deionized water to remove the trace metals from the surface of the cyanobacteria.
Table 1. FTIR analysis of O. willei NTDM01, which shows the functional group of the protein
Frequencycm-1 Type Vibration Mode
3437.05 Sec.amides, freeNH(trans) N-H Structure 2926.52 Alkanes(-CH2-) C-H Structure 2857.58 Alkanes(-CH2-) C-H Structure 2378.14 Charged amines(C=NH+) NH- Structure 2096.19 Organo-silicon compounds Si-H Structure 1639.24 Alkenes(CHR1=CHR1-cis) C=C Structure 1394.59 Alkanes, tery.butyl C-H deformation 1224.56 Aliphatic esters CH3COOR 1102.71 Secondary alcohols C-OH Structure
To initiate the experiments, 5mL of silver solution (~560mg/L) was added to 5mL of
washed cyanobacterial cultures. The experiment were incubated at 25° C for 28 days and maintained in the dark. Abiotic experiments consisting of ~560mg/L silver were conducted using silver nitrate solution without the presence of cyanobacteria. The pH and cell viability were monitored during the course of the study. The sample have been characterized for the development of silver nanoparticles by using Ultra Violet �Visible Spectrum (Uv-Vis) , Fourier Transform Infra Red (FTIR), Scanning Electron Microscopy (SEM) and Energy Dispersive Spectroscopic (EDS) investigation (3,7).
3. Results The axenic strain was identified as Oscillatoria willei NTDM01 (Fig.1). In cyanobacterial
experiment, soluble silver was completely precipitated from solutions within 28days. A grayish- black silver precipitate on cyanobacteria was observed in experiment macroscopically. The pH was constant at 5.0 and 4.7. In abiotic experiments using silver nitrate, total soluble silver concentration and pH were constant until the completion of experiment.
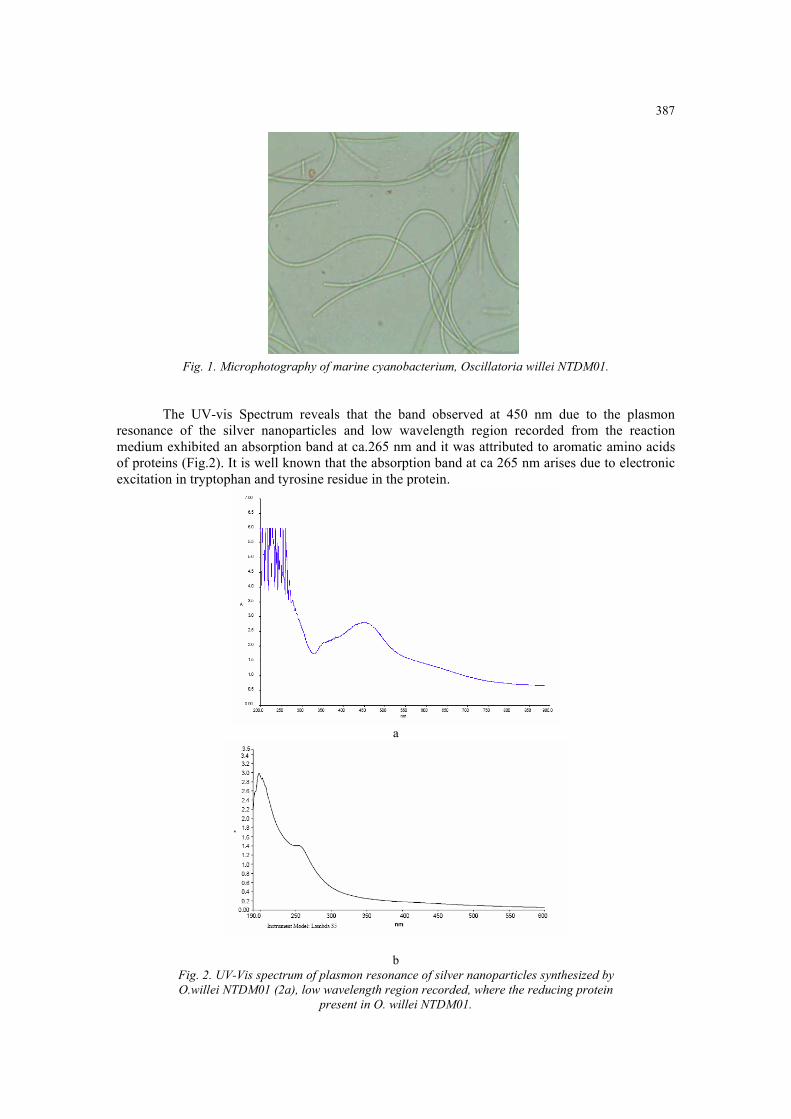
387
Fig. 1. Microphotography of marine cyanobacterium, Oscillatoria willei NTDM01.
The UV-vis Spectrum reveals that the band observed at 450 nm due to the plasmon
resonance of the silver nanoparticles and low wavelength region recorded from the reaction medium exhibited an absorption band at ca.265 nm and it was attributed to aromatic amino acids of proteins (Fig.2). It is well known that the absorption band at ca 265 nm arises due to electronic excitation in tryptophan and tyrosine residue in the protein.
a
b Fig. 2. UV-Vis spectrum of plasmon resonance of silver nanoparticles synthesized by O.willei NTDM01 (2a), low wavelength region recorded, where the reducing protein
present in O. willei NTDM01.
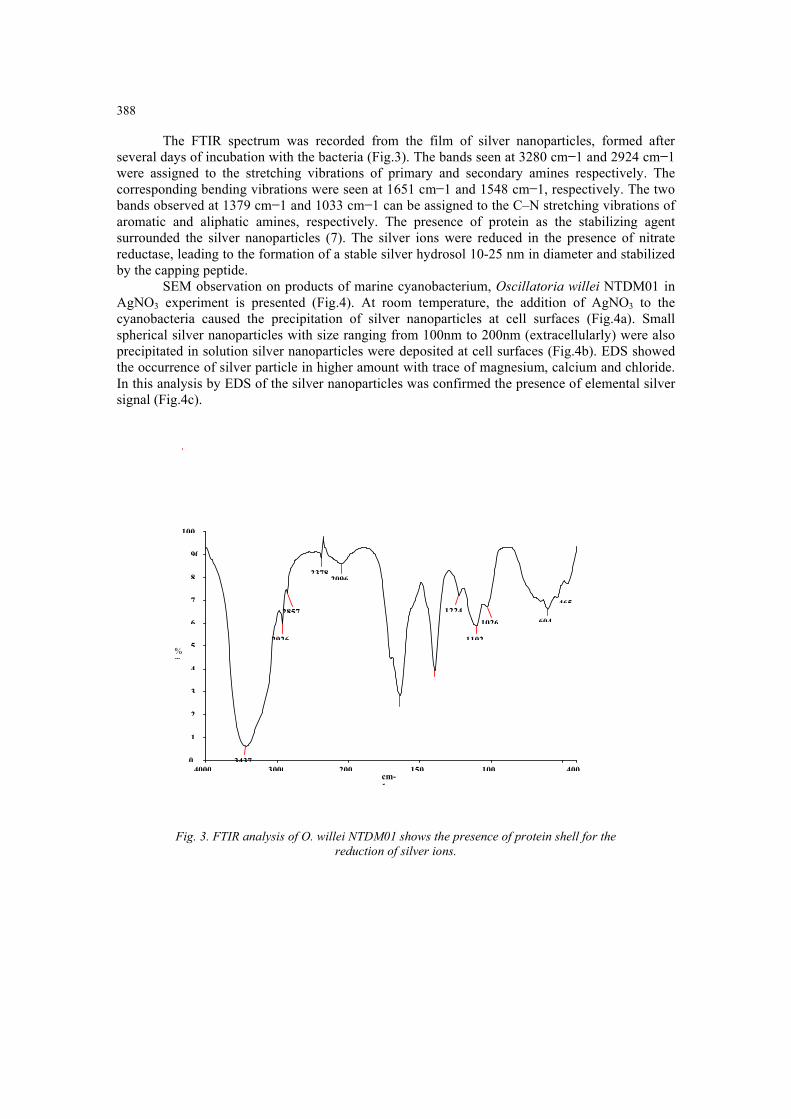
388
The FTIR spectrum was recorded from the film of silver nanoparticles, formed after several days of incubation with the bacteria (Fig.3). The bands seen at 3280 cm 1 and 2924 cm 1 were assigned to the stretching vibrations of primary and secondary amines respectively. The corresponding bending vibrations were seen at 1651 cm 1 and 1548 cm 1, respectively. The two bands observed at 1379 cm 1 and 1033 cm 1 can be assigned to the C�N stretching vibrations of aromatic and aliphatic amines, respectively. The presence of protein as the stabilizing agent surrounded the silver nanoparticles (7). The silver ions were reduced in the presence of nitrate reductase, leading to the formation of a stable silver hydrosol 10-25 nm in diameter and stabilized by the capping peptide.
SEM observation on products of marine cyanobacterium, Oscillatoria willei NTDM01 in AgNO3 experiment is presented (Fig.4). At room temperature, the addition of AgNO3 to the cyanobacteria caused the precipitation of silver nanoparticles at cell surfaces (Fig.4a). Small spherical silver nanoparticles with size ranging from 100nm to 200nm (extracellularly) were also precipitated in solution silver nanoparticles were deposited at cell surfaces (Fig.4b). EDS showed the occurrence of silver particle in higher amount with trace of magnesium, calcium and chloride. In this analysis by EDS of the silver nanoparticles was confirmed the presence of elemental silver signal (Fig.4c).
100
90
Fig. 3. FTIR analysis of O. willei NTDM01 shows the presence of protein shell for the
reduction of silver ions.
4000 3000 200 150 100 4000
1
2
3
4
5
6
7
8
cm-1
%T
3437
2926
2857
23782096
1224465
1102
1026 604

389
a
b
Fig. 4a. Precipitation of silver nanoparticles on the cyanobacterial surfaces. b. SEM micrograph of cyanobacteria cells with nanoparticles silver deposited outside the cell with size
range of 150nm c. EDS of the silver nanoparticles was confirmed the presence of elemental silver signal in high percentage.
The reaction of cyanobacteria with AgNO3 solutions results in the formation of silver
nanoparticles. The presence of spherical silver nanoparticles was observed in experiment. The precipitation of silver nanoparticles was not observed for abiotic experiment that were run under similar condition and duration, that suggesting that cyanobacteria were required for silver precipitation and the precipitation processes were not controlled by inorganic chemical reactions.
4. Discussion A brief overview of the use of micro organisms such as cyanobacteria, yeast, algae, fungi
and actinomycetes in the biosynthesis of metal nanoparticles has been described. Our UV-vis observation indicated the release of protein into the culture medium indicting the electronic excitation of aromatic amino acids. A similar observation has been made in Fusarium oxysporum and suggests a possible mechanism for the reduction of the metal ions present in the solution (8). The overall peaks from FTIR observation confirm the presence of protein in the samples of silver nanoparticles. It can be assumed that nitrate reducing bacteria to produce nitrate reductase enzyme (protein). This reduces to silver nitrate solution form silver nanoparticles. The synthesis of silver nanoparticles using ß-NADPH-dependent nitrate reductase and phytochelatin in vitro has been demonstrated for the first time (9). The cyanobacterial cells were encrusted with silver nanoparticles and separation of some filaments into their constituent cells were still observed. The nanoparticles were not in direct contact even within the aggregates, indicating stabilization of the nanoparticles by capping agent. This corroborates with the previous observation in F. oxysporum (8). Cyanobacteria commonly

390
use nitrate as the major source of nitrogen for three different purposes including growth, generation of metabolic energy, and redox balancing (10). The presence of silver nanoparticles throughout the cells suggests that Ag+ and NO3 entered the cyanobacteria cells through a transport system and degraded in solution. The presence of silver nanoparticles inside the cytoplasm, Ag+ is presumably reduced to Ago Because AgNO3, a toxic reagent, was used in metabolic processes, it ultimately killed the cells. During the death of cyanobacteria, nanoparticles of silver produced inside the cells were release through the cell membrane into solution, as indicated by the precipitation of silver nanoparticles around the cells. The dead cyanobacteria also released organics (protein, and other biochemical) that caused further precipitation of silver from solution outside the cells. The protein molecules act as reducing agent for silver nanoparticles. The protein molecule made up of different functional group in amino acid sequences such as amino, carboxyl, sulfate groups present in the cyanobacterial protein favor the formation of extremely small-sized silver nanoparticulates with narrow particle size distribution and hydroxyl and sulfonic groups are beneficial to synthesis of silver nanoparticle with a slightly larger particle size in a weak reducing environment. It has already been certified that the silver nanoparticles can be stabilized and are well functional groups, such as carboxyl, hydroxyl, and amido and thiol group, except the steric effect arising from the large molecular structures of the organic modifier (protein) on the surface of the particles (11,12). The micro organisms in the synthesis of nanomaterials as a possible viable alternative to the more popular physical and chemical methods, currently in vogue. The use of cyanobacteria as a source of enzymes that can catalyze specific reactions leading to inorganic nanoparticles is a new and rational biosynthesis strategy that is being developed. Extracellular secretion of enzymes offers the advantages of obtaining large quantities in a relatively pure-state, free from other cellular proteins associated with the organism and can be easily processed by filtering of the cells and isolating the enzyme for nanoparticles synthesis from cell-free filtrate.
Acknowledgments The authors wish to acknowledge support from University Grant commission (UGC) for
their financial assistance. References
[1] K. Abe, T. Hanada, Y. Yoshida, N. Tanigaki, H. Takiguchi, H. Nagasawa, M. Nakamoto, T. Yamaguchi, K. Yase, Thin Solid Films 327, 524 (1998). [2] A. Ahmed, P. Mukherjee, S. Senapati, D. Mandal, M.I. Khan, R. Kumar, M. Sastry, Colloids Surf B 28, 313 (2003). [3] S. Anil Kumar, M.K. Abyanesh, S. W. Gosavi, S.K. Kulkarni, R. Pasricha, M. Ahmed, M.I Khan, Biotechnol Lett 29: 439 (2007). [4] K.C. Bhainsa, K. De�Souza, Colloids Surf B, 47:160 (2006). [5] N. Duran, P.D. Marcato, O.L. Alves, G.I.H. De souza, E. Esposito, J.Nanobiotechnol, 3: 1 (2005). [6] M. Lenkge, M.E. Fleet, G. Southam, Langmuir, 23: 2694 (2007). [7] P. Mukherjee, A. Ahmed, D. Mandal, S. Senapati, S.R. Sainkar, M.I. Khan, R. Parischa, P.V. Ajayakumar, R. Alam kumar, M. Sastry, Nano Lett, 1: 515 (2001). [8] R. Rippika, Arch. Microbiol, 111: 93 (1972). [9] S.S. Shankar, A. Ahmed, S. Murali, J. Mater Chem. 13:1822 (2003). [10] I. Suzuki, H. Kikuchi, S. Nakanishi, Y. Fujita, T. Sugiyama, T. Omata, J. Bacteriol, 177, 6137 (1995). [11] M. K. Temgire, S.S. Joshi, Rad Phys, 71: 1039 (2004).[12] N. Vigneshwaran, A.A. Kathe, P.V. Varatharajan, R. P. Nachane, R. H. Balasubramanya, Langumir 23: 7113 (2007). [13] B. Yin, H. Ma, S. Wang, S.J. Chen, Phys. Chem 107: 8898 (2003).

Digest Journal of Nanomaterials and Biostructures Vol. 6, No 1, January-March 2011 p. 117 - 120
BIOSYNTHESIS AND CHARACTERIZATION OF SILICON-GERMANIUM OXIDE NANOCOMPOSITE BY DIATOM
D. MUBARAK ALI, C. DIVYA, M. GUNASEKARANa, N. THAJUDDIN Department of Microbiology, School of Lifescience Bharathidasan University, Tiruchirappalli, Tamil Nadu, IndiaaDepartment of Biology, Fisk University, Nashville Tennessee, USA
Diatoms are single cell photosynthesizing eukaryotic algae that produce intricately structured cell wall made of nano-patterned silica. The biologically fabricated nanostructures offer substantially different properties related adhesion, tribology, optic and electronic behavior. In this study biosynthesis and characterization of Silicon- Germanium (Si-Ge) oxide nanocomposite in the diatom, Stauroneis sp., by two stage cultivation process. The exponential growth of Stauroneis sp., inoculated with enrichment medium amended with Germanium. The silicon to germanium concentration molar ratio was 1:0.5. Scanning electron microscopic studies reveals that no structural change is observed when germanium concentration is low, the increased concentration of germanium led to structural changes of the diatoms. The Si-Ge oxide nanocomposite was characterized by Electron Microscopy equipped with energy dispersive spectroscopy to evaluate the chemical composition and structural properties of the diatoms.
(Received December 1, 2010; accepted January 14, 2011)
Keywords: Nanocomposites, silicon, germanium, diatom, Stauronesis sp., EDS
1. Introduction
The cell wall structure is a species-specific characteristics demonstrating that diatom silica morphogenesis is genetically encoded. It is expected that these structures exhibit superior properties in a wide range of applications. The fabricated and self assembled of semiconductor nanostructures that possess unique optical and electronic properties [1]. The supramolecular chemistry and catalysis have led to novel surface and size dependent chemistry such as enation selective catalysis at the surface. The ability of diatoms to make complex, nanoscaled, three dimensional silica shells called “frustules” offers attractive possibilities for their application to nanobiotechnology [2]. Bioengineered nanosystem is extremely difficult to realize on the basis of current scientific knowledge and bottom up assembly is very powerful in creating of identical structures with atomic precision, such as supramolecular functional entities in living organisms. In this regards diatoms, a prolific class of single celled algae that make micro scale biosilica or frustules with intricate submicron features dominated by two dimensional pore arrays, have been advertized as a paradigm for the controlled production of nanostructured silica with interesting properties [3]. The fabrication of novel biomaterials through molecular self assembly is studied currently [4]. In this study we report that the fabrication of Silicon- Germanium oxide nanocomposites characterized by Electron Microscopy and EDS.
______________________ *Corresponding author: [email protected]

118
2. Materials and methods The photosynthetic freshwater diatom, Stauroneis sp., was isolated from the freshwater
ponds from Bharathidasan University Campus and maintained in F/2 medium and cultivated in appropriate growth conditions such as light (2000lux), pH (8.0±0.2). The two stage cultivation methods is follwed for the synthesis of silicon-germanium nanocomposites as described [5]. In first stage, the cells were grown in nutrient medium containing sufficient silicon concentration for its growth. The silicon concentration (30mg/l) and cell density is monitored at regular intervals. When all the silicon was used up and the cell density had become constant, the cells were then considered to be starved of silicon, this condition prevails one week. In the second stage, the silicon and germanium (1:0.5) is added to the liquid medium for the biosynthesis of Silicon –Germanium nanocomposites. After 7 days the cells were harvested from the medium and washed thrice in deionized water in order to remove surface adsorbed germanium and other trace elements. Then the sample was centrifuged at 2000rpm for 5mins. The pellet was collected and dispersed on the thin slide and allowed to dry in hot air oven. The small portion of slide was placed onto carbon tape affixed to a stub, the gold was coated before introduction to the Scanning Electron Microscopy equipped with EDS.
3. Results and discussion
The incorporation of the Germanium into diatom cells was achieved and nanostructured pores could be found without morphological aberrations (Fig. 1). In this study the freshwater diatom, Stauroneis sp., was used to fabricate Silicon- Germanium by its metabolic activity. The SEM was used to assess and validate the integrity of the diatoms. The diatom, Stauroneis sp., frustules is observed in different growth conditions. After separation of diatom frustules from the organic matter, the sample was dispersed in alcohol, and then placed on the stub [6]. The characteristics peaks of Germanium were observed at 1.22 keV, 9.8 keV and 10. 4keV. The maximum peak of silicon was observed at 1.9keV (Fig. 2). It has been reported that the distribution profile of germanium in the diatom frustules is similar to that of silicon intensity is quite different [6]. It has been reported that silica from bioreactor cultured Nitzschia frustulumcells possessed blue photoluminescence, where the luminescence intensity and wavelength were dependent on the change in frustules nanosturucture as the cell culture moved from the exponential to the stationary phase of growth [5]. The soluble Germanium can be metabolically inserted into biosilica of the pennate diatom, Pinnularia sp., by two stage bioreactor process [7]. The metabolic incorporation of germanium into the biosilica of the diatom Nitzschia frustulum is described for the biological fabrication.
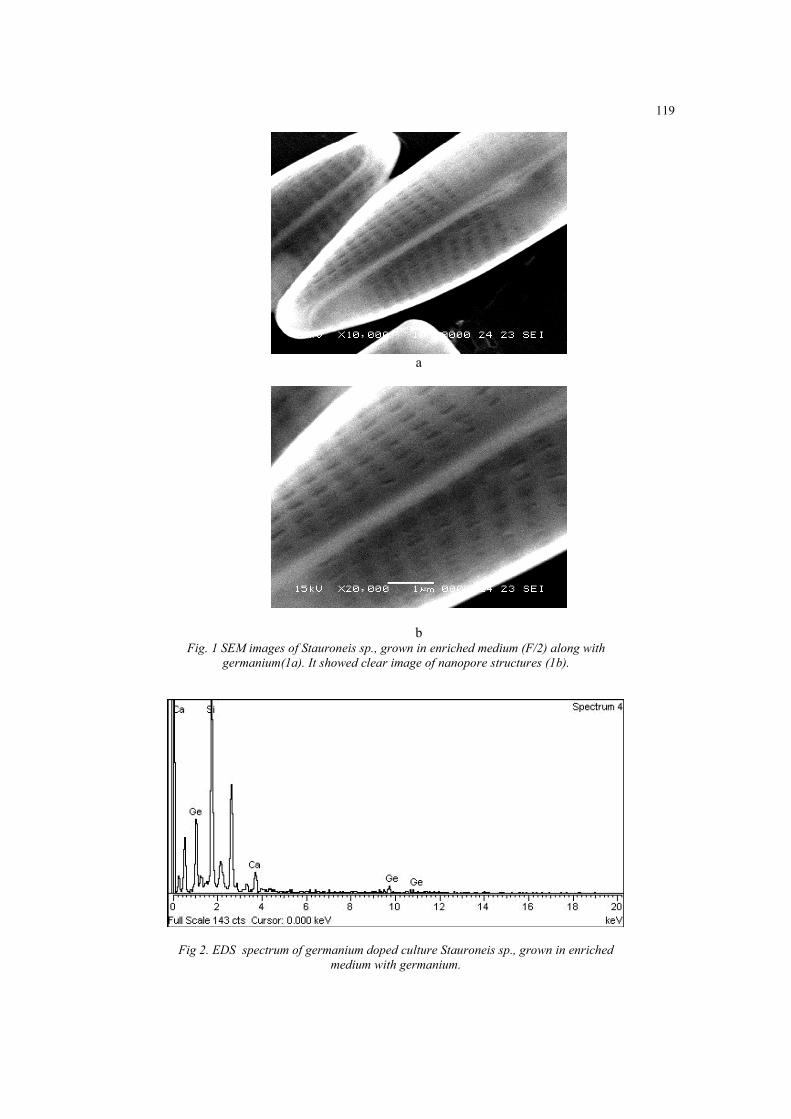
119
a
bFig. 1 SEM images of Stauroneis sp., grown in enriched medium (F/2) along with
germanium(1a). It showed clear image of nanopore structures (1b).
Fig 2. EDS spectrum of germanium doped culture Stauroneis sp., grown in enriched medium with germanium.

120
Fig 3. SEM images of Stauroneis sp., grown in enriched medium (F/2) along with germanium in molar ratio of 1:1. It showed aberrated diatom structure.
Fig. 3 shows that due to the equal volume of Silicon- Germanium, the diatom cells are fully aberrated. An Electron Microscopic study reveals that due to the incorporation of germanium, the diatom cells lead to various degrees of aberrations and also unavailability of sufficient silicon for the cell wall biosynthesis. The Stauroneis sp., with high percentage of germanium concentration has been found to cause cell aberrations than with the low percentage. The difference in bond length between silicon oxide and germanium oxide probably weakens the diatom frustules and cause them to break easy during the processing of the cells [6].
In this study incorporation of germanium into diatom cells were achieved. Several workers also reported that there was no Silicon- Germanium homogeneity in the distribution of the diatom cells. The experimental protocol should be standardized to achieve the homogeneity intricate Silicon-Germanium nanocomposites. These optoelectronic properties of the nanocomposites, which depend on the amount of Germanium assimilation can therefore be controlled.
Acknowledgement
The authors are grateful thanks to the University Grant Commission (UGC) for their grant.
References
[1] G. L. Rorrer, C-H. Chang, S.H. Liu, C. Jeffryes, J. Jiao, J. A. Hedberg, J. Nanosci. Nanotechnol. 5, 41 (2005). [2] M. Hildebrand. J. Nanosci. Nanotechnol. 5, 146 (2005). [3] M. Sumper, E. Brunner, Adv. Funct. Mater. 16, 17 (2006). [4] S. Zhang, Nat. Biotechnol. 21, 1171 (2003). [5] T. Qin, T. Gutu, J. Jiao, C-H. Chang, G.L. Rorrer, J. Nanosci. Nanotechnol. 8, 2392 (2008). [6] T. Gutu, L. Dong, J. Jiao, G.L. Rorrer, C-H. Chang, C. Jeffryes, Q. Tian. Microc Microanal.
11, 1958 (2005). [7] C. Jeffryes, T. Gutu, J. Jiao, G. L. Rorrer. Mater. Sci. Eng. C. 28, 107 (2008).

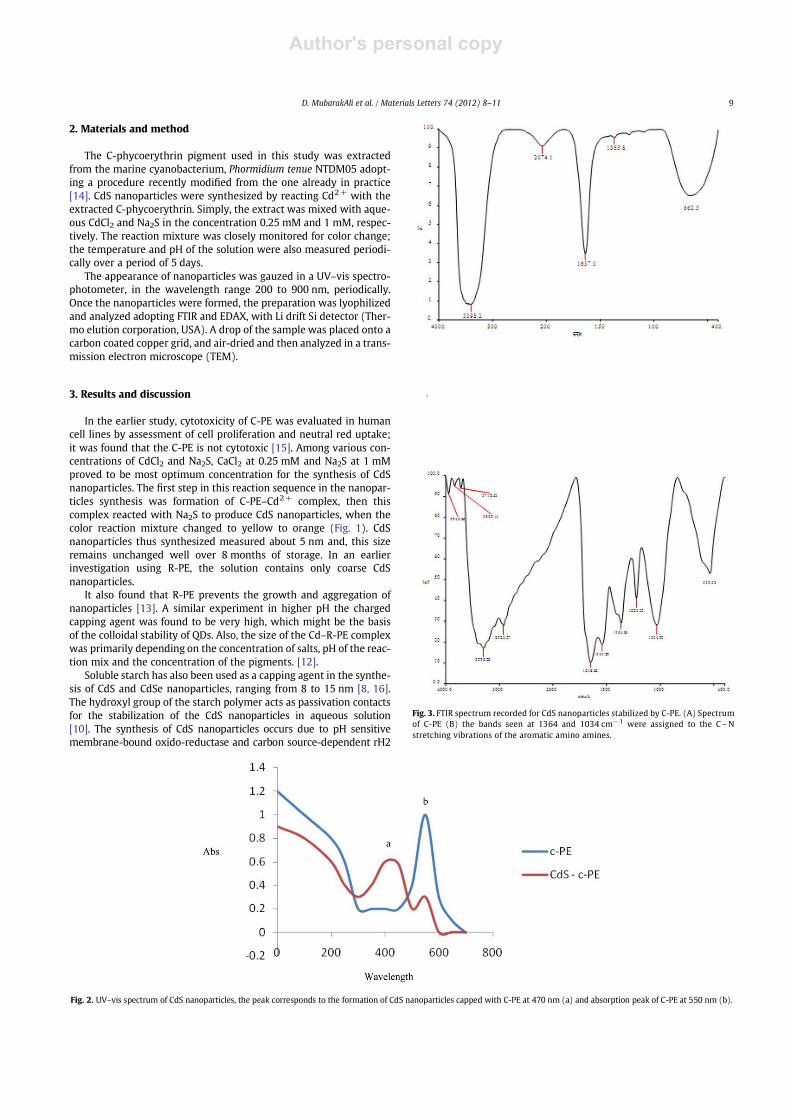
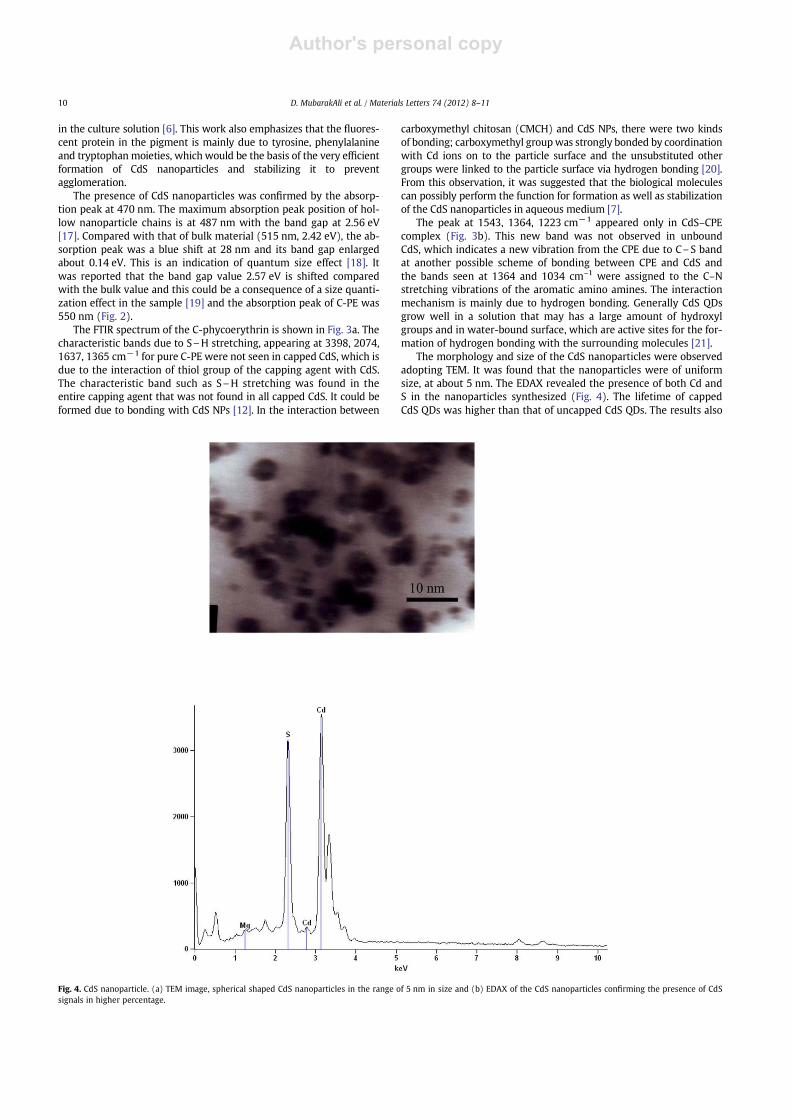



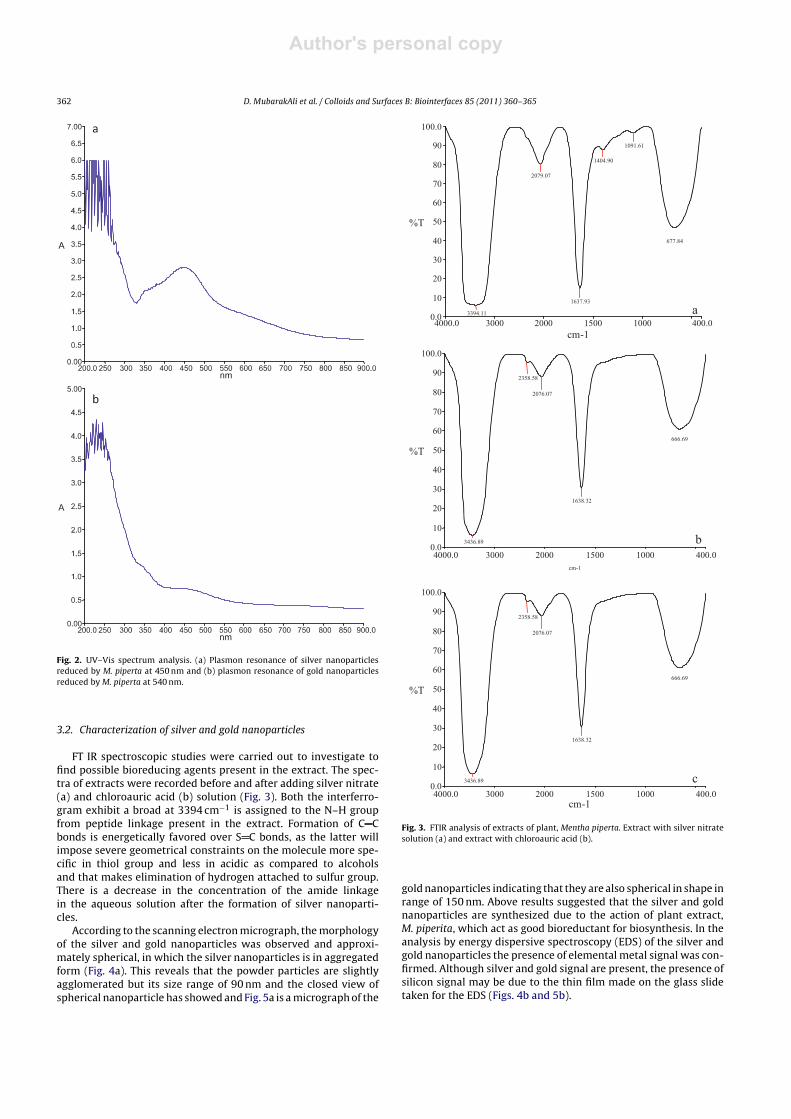
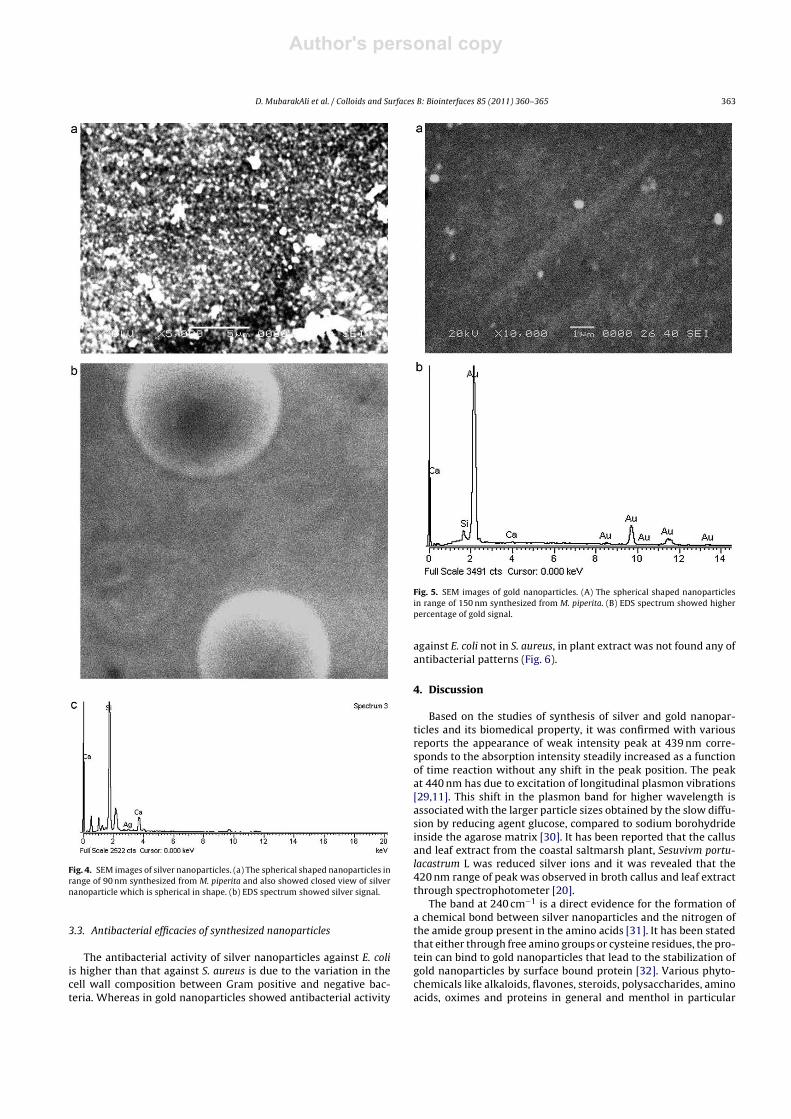
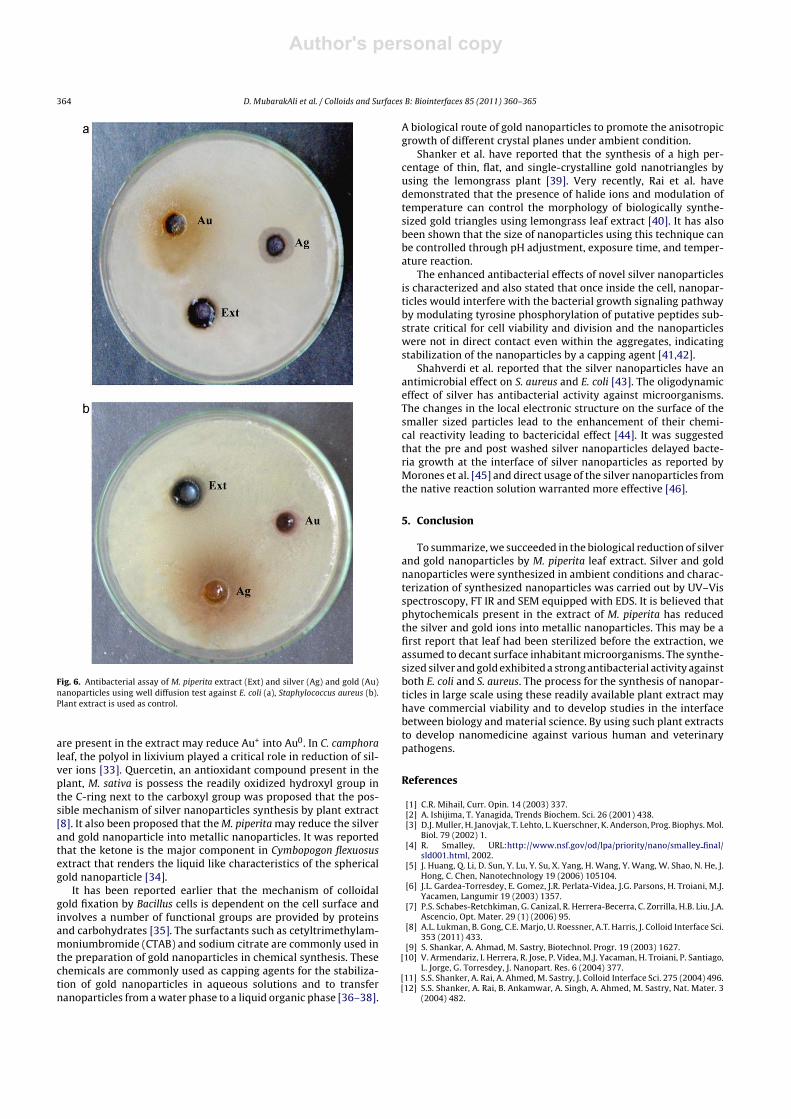


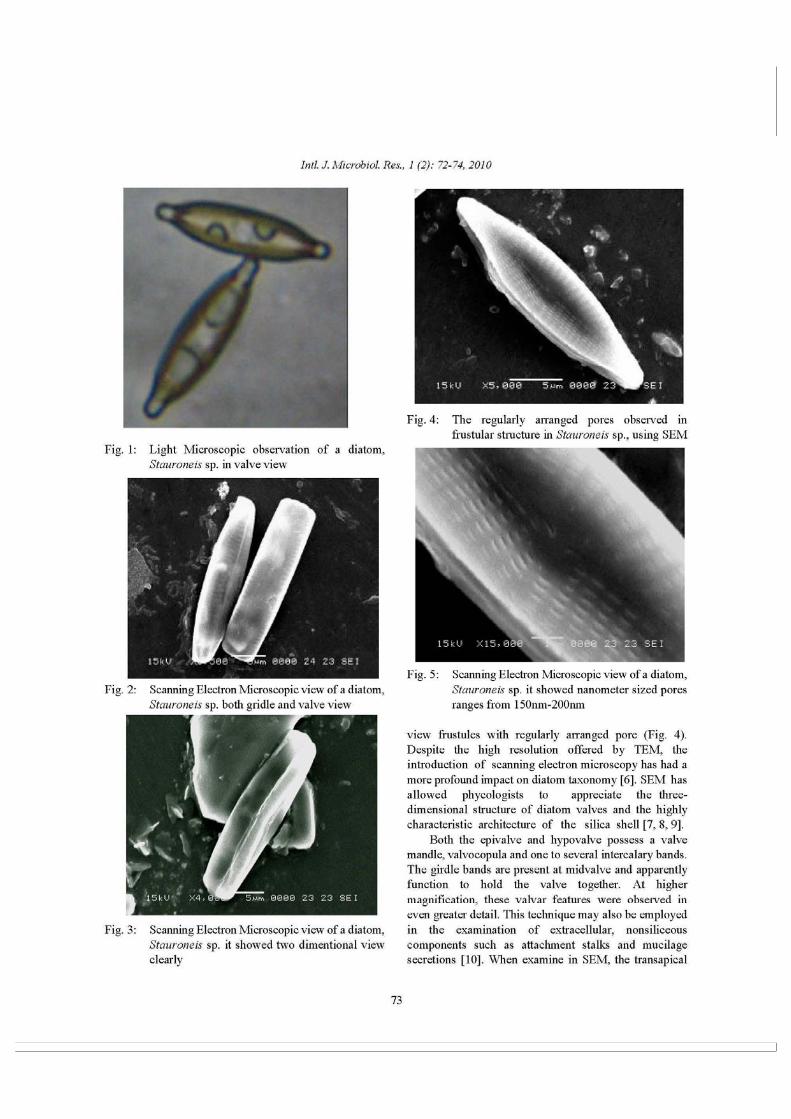







African Journal of Basic & Applied Sciences 3 (1): 09-13, 2011ISSN 2079-2034© IDOSI Publications, 2011
Corresponding Author: Dr. N. Thajuddin, Department of Microbiology, Bharathidasan University, Tiruchirappalli-620 024, India, Tel: +91-431-2407028, E-mail:[email protected].
9
Efficiency of Textile Dye Decolorization by Marine Cyanobacterium, Oscillatoria formosa NTDM02
D. Mubarak Ali, A. Suresh, R. Praveen Kumar, M. Gunasekaran and N. Thajuddin1 1 1 2 1
Department of Microbiology, School of Life sciences, Bharathidasan university, Tiruchirappalli-620 024, India
Department of Biology, Fisk University, Nashville Tennessee, USA 1
Abstract: Cyanobacteria are the Gram negative, oxygen evolving photosynthetic prokaryotes andhave wide variety of application in biotechnological research. Releasing of textile dye effluents intogeneral water bodies is a major environmental and health problem. Micro algae and cyanobacteria areconsidered as an important source for decolorizing dye and textile effluent. The dye Amido Black and textiledye effluent is chosen for this investigation and the marine cyanobacterium, Oscillatoria formosa NTDM02was used for the decolorization process. Chlorophyll-a content of this organism was tested before and after thetreatment. The experimental results showed that the marine cyanobacterium, Oscillatoria formosa NTDM02,which decolorize the textile effluent efficiently in short period of time. This can be used for the bioremediationof dye effluents.
Key words: Oscillatoria % Effluent % Dye % Chlorophyll-a % Cyanobacteria % Bioremediation
INTRODUCTION MATERIALS AND METHODS
Microorganism, as bioremediation agents in the Culture Conditions: The marine cyanobacterium,treatment of waste water containing textile dyes [1]. Oscillatoria formosa NTDM02 (obtained from theCyanobacteria have a ubiquitous distribution [2-5] but Microbial Culture Collections of Microbiologytheir role in functioning of ecosystems, including Department at Bharathidasan University, Tamilnadu,degradation of recalcitrant compounds including India) was maintained at 25°C in MN III [15] medium withdyes [6-9]. Azo-aromatic dyes are the major group of light/dark cycle of 16/8 h under 6000 Lux illumination. Fortextile dye stuffs. When these colored effluents enter the experiment, 7-day-old logarithmic growing cells ofwater bodies they may either inhibit the biological activity Oscillatoria formosa NTDM02 was used.or cause water-borne disorders such as nausea,hemorrhage, ulceration of skin and mucous membrane, Analysis of [H ] concentration (pH): pH of the mediumdermatitis, perforation of nasal septum, severe irritation of and effluent were monitored at every stages of the studyrespiratory tract or cancer [10]. These structures can be using pH meter (Elico) to analyze the variation during thereductively cleaved into colorless amines by several decolorization process. bacterial species [11, 12]. The cleavage of azo bonds is agratuitous process which can occur when the micro Estimation of Chlorophyll-a: Chlorophyll a was measuredorganisms using a reduced carbon compounds as thegrowth substrate [13]. The marine cyanobacterium,Phormidium valderianum produces hydrogen and canremove dyes from the solution [14]. Microbialdecolorization processes have the advantage of beingsimple in design and low cost compared withconventional treatments.
+
spectrophotometrically at 665nm from the cell mass beforeand after the treatment to compare the growth [16].
Study of Dye Decolorization: The dye effluent wascollected from textile industry at Karur, Tamilnadu, Indiaand Amido Black G200 was used for the study ofdecolorization control. For this study five set of


Phykos 42 (1): 10 14 (2012) Ilavarasi, et al., Production of FAME from freshwater microalgae
©Phycological Society, India
10
Production of FAME from freshwater microalgae and profiling of fatty acids for biodiesel feedstock
Ilavarasi, A., MubarakAli, D., Parveez Ahamed, A and N. Thajuddin*
Department of Microbiology, School of Life Sciences, Bharathidasan University,
Tiruchirappalli 620024, Tamil Nadu, India.
*Corresponding author: E. mail: [email protected]
Abstract
Due to diminishing fossil fuel reserves, it is necessary to shift for an alternative fuel source that could be sustainable and eco-friendly. Microalgae produce different ratios of lipids, proteins and carbohydrates. The lipid composition of microalgae is similar to vegetable oil which has already been used as biodiesel so that it can potentially be employed for biodiesel production. In present study has demonstrated the total lipid content, fatty acid profile and biodiesel production from a naturally isolated fresh water strain of Chlorella sp. and the total lipid content was found to be 8% under normal nutrient conditions. Gas Chromatography of FAME was analyzed, it was found to be the major fatty acids were palmitic acid, stearic acid, oleic acid, linoleic acid and alpha linolenic acid and pH of the biodiesel was found to be 8.1.
Keywords: Microalgae, Chlorella sp., BG-11, Lipid, FAME, GC, Biodiesel.
Introduction:
Robust population explosion, receding fossil fuel reserve, enhanced GHG, larger emission of flue gases etc have brought the world scientists and biotechnologists to the threshold of exploring alternative energy sources especially biodiesel. Biodiesel is characterized as a renewable, biodegradable and eco-friendly fuel which has attracted wide attention. Most recently, research effort has been aimed at identifying suitable biomass species which can provide high energy outputs to replace the fossil fuels. Many of the researchers have attempted to produce biodiesel from non-edible sources like frying oil, greases, tallow, jatropha, mahua and soy bean oils. Besides these non-edible sources we can use microalgae as feed stock for biodiesel.
Microalgae are oxygenic photosynthetic organisms which inhabit almost all moist and lighted environments. They are veritable miniature biochemical factories, appeared to be more photosynthetically efficient than terrestrial plants and are efficient CO2 fixers. Many of them are exceedingly rich in oil, fast growing and so they are used in biodiesel production. They have the ability to replace the fossil fuel (Chisti, 2007). Even, there is a pressing need to improve the biodiesel production from microalgae and that too on selection of suitable biomass species. The nutrients deprivations would be influence on the lipid accumulation in a dominant indigenenous microalga, Chlorella sp., BUM11008 for biodiesel production (Praveenkumar et al., 2012). In the present study, fresh water microalga, Chlorella sp. NTAI01 was
isolated and identified. The total lipid was characterized both quantitatively and qualitatively in biofuels aspects.
Materials and Methods: Samples:
The microalgal samples were collected from stagnant fresh water body in Bharathidasan University campus, Tiruchirappalli, India.
Isolation, identification and growth condition of microalgae:
The collected samples were enriched initially in BG -11broth in conical flasks (250ml) at 24±2°C under 37.5µmol-1m2s-1 intensity with 16:8 hours photoperiod for 10 days. Then the enriched culture samples were spread on BG 11 agar plates and incubated at the above said conditions (Ilavarasi et al., 2011). After incubation, individual colonies were picked and transferred to the same media for purification in 250ml conical flask. The culture broth was shaken manually for five to six times a day. The purity of the culture was monitored by regular observation under microscope. The isolated microalgae were identified microscopically using light microscope with standard manual for algae (Shashikant and Gupta, 1998).
Measurement of growth:
The growth of the algal biomass was assessed by means of optical density with five days intervals from 5th



























