Embed Size (px)
Citation preview

MOLECULAR APPROACHES TOMICROBIOLOGICAL MONITORING:
FECAL SOURCE DETECTION
KATHARINE G. FIELD1*, ANNE E. BERNHARD2, AND TIMOTHY J. BRODEUR1
1Department of Microbiology, Oregon State University, Corvallis, OR 97331;
2Department of Civil and Environmental Engineering, University of Washington, Seattle, WA 98195-2700
*(author for correspondence, e-mail: [email protected])
Abstract. Molecular methods are useful both to monitor natural communities of bacteria, and totrack specific bacterial markers in complex environments. Length-heterogeneity polymerase chainreaction (LH-PCR) and terminal restriction fragment length polymorphism (T-RFLP) of 16S rDNAsdiscriminate among 16S rRNA genes based on length polymorphisms of their PCR products. Withthese methods, we developed an alternative indicator that distinguishes the source of fecal pollutionin water. We amplify 16S rRNA gene fragments from the fecal anaerobic genus Bacteroides withspecific primers. Because Bacteroides normally resides in gut habitats, its presence in water indi-cates fecal pollution. Molecular detection circumvents the complexities of growing anaerobic bacte-ria. We identified Bacteroides LH-PCR and T-RFLP ribosomal DNA markers unique to either rumi-nant or human feces. The same unique fecal markers were recovered from polluted natural waters.We cloned and sequenced the unique markers; marker sequences were used to design specific PCRprimers that reliably distinguish human from ruminant sources of fecal contamination. Primers formore species are under development. This approach is more sensitive than fecal coliform assays, iscomparable in complexity to standard food safety and public health diagnostic tests, and lends itselfto automation and high-throughput. Thus molecular genetic markers for fecal anaerobic bacteriahold promise for monitoring bacterial pollution and water quality.
Keywords: Microbiological monitoring, fecal contamination, fecal source discrimination, Bacteroi-des, anaerobic bacteria, LH-PCR, T-RFLP, bacterial communities, molecular markers
1. Introduction
Fecal contamination of aquatic environments afflicts many regions of the U.S.,with associated human health risks and environmental damage. Human fecal pol-lution spreads many dangerous bacterial pathogens and viruses such as hepatitis,while other pathogens such as Cryptosporidium parvum, Giardia lamblia, Salmo-nella spp., and Escherichia coli (E. coli) O157:H7 are associated with animal fe-cal pollution. Fecal bacteria, pathogenic protista and viruses may be highly con-centrated in sediments (Pommepuy et al., 1992) and in shellfish (DePaola et al.,1990). In 1995, 4.9 million acres of shellfish-growing waters were restricted na-tionally due to water pollution. Additionally, in 1996, over 2,500 beaches in theU.S. were posted or closed due to microbial contamination.
According to the Clean Water Action Plan of the U.S. EPA, “after 25 years,the clean water program has addressed many of the major pipe discharges ofsewage and industrial waste. By far the predominant source of remaining wa-ter polluting problems is from urban and agricultural lands and facilities suchas animal feeding operations...”
Environmental Monitoring and Assessment 81: 313–326, 2003.c©2003 Kluwer Academic Publishers. Printed in the Netherlands.

314 FIELD, BERNHARD, AND BRODEUR
Often the source of fecal contamination in water cannot be determined. Forexample, failing septic systems, overloads at sewage treatment facilities, and run-off from non-point sources such as farm manure may all be candidates. In addi-tion, although sediments are known to be a source of human pathogens, the contri-bution of bacterial pollution “stored” in sediments and re-suspended during stormevents is unknown relative to other sources. In order to adequately assess humanhealth risks and develop management plans for watersheds providing drinkingwater and supporting recreation and fisheries, it is necessary to know the sourcesof fecal contamination.
The standard method of measuring fecal pollution does not distinguish betweenhuman and animal sources. The method requires growing fecal coliforms or Es-cherichia coli from water samples to estimate their concentrations. Because thesebacteria are found in a variety of warm-blooded animals and even reptiles (Harwoodet al., 1999), this method does not distinguish between human and animal sources.In addition, coliform-based methods can be confounded by growth of coliforms inreceiving water and sediments (Gerba and McLeod, 1976; Hood and Ness, 1982;Howell et al., 1996; LaLiberte and Grimes, 1982). Coliforms adhere to sedimentsand are transported in and out of the water column, affecting the measure of fecalpollution in unknown ways (Weiskel et al., 1996; Gary and Adams, 1985; Gerbaand McLeod, 1976; Hood and Ness, 1982; Howell et al., 1996; LaLiberte andGrimes, 1982; Sherer et al., 1988). It is remarkable that although coliforms are thegold standard for measuring fecal pollution, the influences of these important fac-tors are still so poorly understood.
2. Methods of detecting the source of fecal pollution
Several methods of discriminating the source of fecal pollution have been proposed,but each has drawbacks. The ratio of fecal streptococci to fecal coliforms has beenused to differentiate human fecal contamination from that of other animals (Sinton etal., 1993). But because strains of coliforms and streptococci have different survivalrates, the ratio changes in complex ways over time, making it unreliable.
Antibiotic resistance patterns of fecal streptococci or coliforms can distinguishamong sources of fecal pollution (Andrews et al., 1997; Wiggins, 1996; Wigginset al., 1999). Because this method requires culturing a large number of fecal strep-tococcal isolates, screening them for resistance against a panel of antibiotics, andanalyzing the results by discriminant analysis, it is labor-intensive and time-con-suming. When sets of isolates from a single type of feces have been analyzed,rates of correct classification have varied from about 64 to 87%. However, whenindividual isolates from mixed fecal sources were analyzed, rates of correct classi-fication were lower. Thus the accuracy of this method may be unacceptably low.
E. coli ribotyping (Carson et al., 2001; Parveen et al., 1999) and rep-PCR DNAfingerprinting (Dombek et al., 2000) are genotyping methods that depend on grow-

MOLECULAR APPROACHES TO MICROBIOLOGICAL MONITORING: FECAL SOURCE DETECTION 315
ing a large number of isolates. Ribotyping correctly discriminates between humanand non-human fecal isolates with a high rate of success, but is less successful atidentifying animal sources (Carson et al., 2001; Parveen et al., 1999). A ribotypinganalysis is estimated to take two weeks to complete (M. Samadpour, pers. comm.).DNA fingerprinting utilizes complex, tedious-to-interpret patterns that don’t al-ways correctly group isolates, even after statistical analysis (Dombek et al., 2000).Both methods require massive local collections of strains for comparison. Althoughsuch collections are underway in several locales, the expense of maintaining themoften puts them out of reach of anyone except private consulting companies.
Phage can be used as indicators of fecal pollution (Goyal et al., 1980; Havelaaret al., 1993; Osawa et al., 1981; Palmateer et al., 1991; Paul et al., 1997; Singh andGerba, 1983; Tartera and Jofre, 1987; Tartera et al., 1989). Coliphage abundanceis well correlated with sewage effluent (Paul et al., 1997); coliphage are also asso-ciated with non-point discharge (Paul et al., 1997). RNA phages, which constituteonly a minor fraction of human coliphages, show specificity in host distribution(Osawa et al., 1981). Although coliphage may indicate the presence of humanenteric viruses in polluted water (Havelaar et al., 1993; Palmateer et al., 1991;Sobsey, 1989), their use may be limited since only a small portion of fecal bacteriamay contain phages (Osawa et al., 1981), and virus concentrations in sewagesamples are highly variable (Havelaar et al., 1993).
All of these methods of fecal source discrimination are time consuming, laborintensive, require specialized equipment or extensive strain collections, are sub-ject to culture bias, and may have unacceptably low rates of accuracy. A method isneeded that:
• Is quick and reliable;
• Does not require culturing isolates;
• Does not require a large library of reference strains;
• Is flexible for easy field handling and storage of samples; and
• Requires a minimum of specialized equipment.
Modern methods of identifying microbes in the environment, foods, and clini-cal diagnosis rely on genetic markers, not growing the microbes. We adapted thesemethods to source discrimination of fecal pollution.
3. Markers from Fecal Bacteroidesto Distinguish the Source of Fecal Pollution
To distinguish the source of fecal pollution in fresh and marine waters, we havedeveloped a new Polymerase Chain Reaction (PCR)-based indicator system, uti-lizing molecular markers from the Bacteroides group of fecal anaerobic bacteria(Bernhard and Field, 2000a; Bernhard and Field, 2000b). The method differs fromexisting methods of detecting fecal pollution in that it detects genetic marker se-

316 FIELD, BERNHARD, AND BRODEUR
quences that are not only specific to fecal bacteria, but are also specific to the hostspecies that produced the feces, allowing discrimination among different potentialsources. Furthermore, the method does not require isolating and growing the indi-cator bacteria, and it is rapid and accurate.
Bacteroides is a genus of non-spore-forming, obligate anaerobes that make upone-third of the human fecal flora, far outnumbering coliforms (Holdeman et al.,1976; Moore and Holdeman, 1974; Salyers, 1984). The most abundant Bacteroi-des species in human feces are either human-specific, or present only at very lowlevels in other species (Allsop and Stickler, 1985; Straub and Dixon, 1997). Inaddition, Bacteroides and its close relatives in the genus Prevotella are found ex-clusively in feces, animal rumens, the mouth and teeth, and other cavities withinanimals and humans (Paster et al., 1994). When found in water, they are invariablydiagnostic of pollution. Because they are strict anaerobes, they are thought not tosurvive very long once released into receiving waters (Avelar et al., 1998; Kreader,1998). Bacteroides has been identified from environmental water samples for sev-eral days after dispersal in water, (Kreader, 1995; Straub and Dixon, 1997). It isdetectable by PCR for up to 14 days at 4°C, even in the presence of predators,while at higher temperatures more typical of natural waters (14°C), Bacteroides isdetectable for 4–5 days (Kreader, 1998).
In the past, Bacteroides has had limited use as an indicator because of the rela-tive complexity of growing strict anaerobes. We circumvent this by using molecu-lar, rather than culture-based, methods to detect the bacteria. Molecular approachesmake it possible to use difficult-to-grow, but diagnostic, microbes in fecal pollu-tion assays. They also avoid cultivation bias, which is the tendency to preferen-tially grow weedy, easy-to-grow species and strains, thus not getting an accuraterepresentation of the actual population.
To establish diagnostic markers, we used recently developed technologies thatdiscriminate among mixtures of bacterial gene sequences by detecting sequencelength differences in a particular gene fragment (Avaniss-Aghajani et al., 1994;Brunk et al., 1996). Length Heterogeneity Polymerase Chain Reaction (LH-PCR;Rappé et al., 1997; Suzuki et al., 1997) and Terminal Restriction Fragment LengthPolymorphism (T-RFLP; Bruce, 1997; Clement et al., 1998; Liu et al., 1997) ana-lyze differences in lengths of gene fragments due to insertions and deletions andpoint mutations affecting restriction recognition sites, and estimate the relativeabundance of each fragment.
To find the Bacteroides marker sequences that discriminate fecal sources, weuse the Polymerase Chain Reaction (PCR, a method of making millions of copiesof a specified gene in a test tube) to amplify specific Bacteroides 16S rDNA mark-ers from DNAs extracted from feces. Amplified fragments were cut with restric-tion enzymes and screened by LH-PCR and T-RFLP using GeneScan on an auto-mated DNA sequencer (Bernhard and Field, 2000a). These methods allowed us to
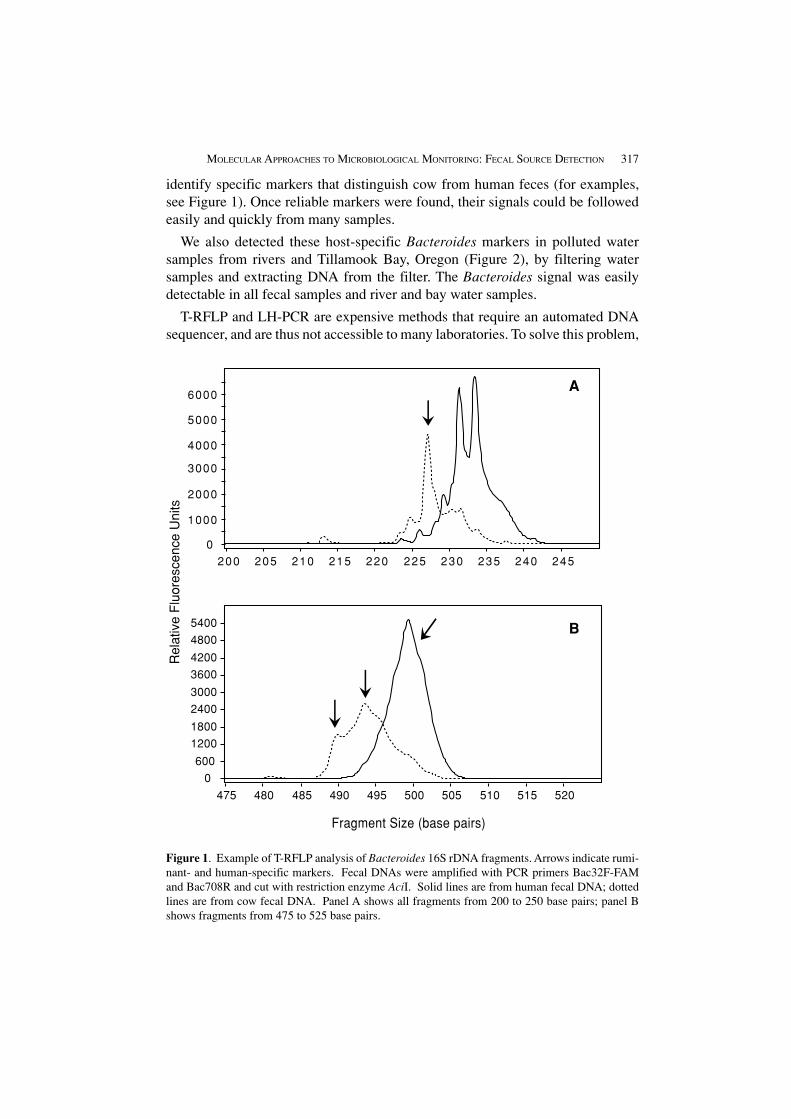
MOLECULAR APPROACHES TO MICROBIOLOGICAL MONITORING: FECAL SOURCE DETECTION 317
identify specific markers that distinguish cow from human feces (for examples,see Figure 1). Once reliable markers were found, their signals could be followedeasily and quickly from many samples.
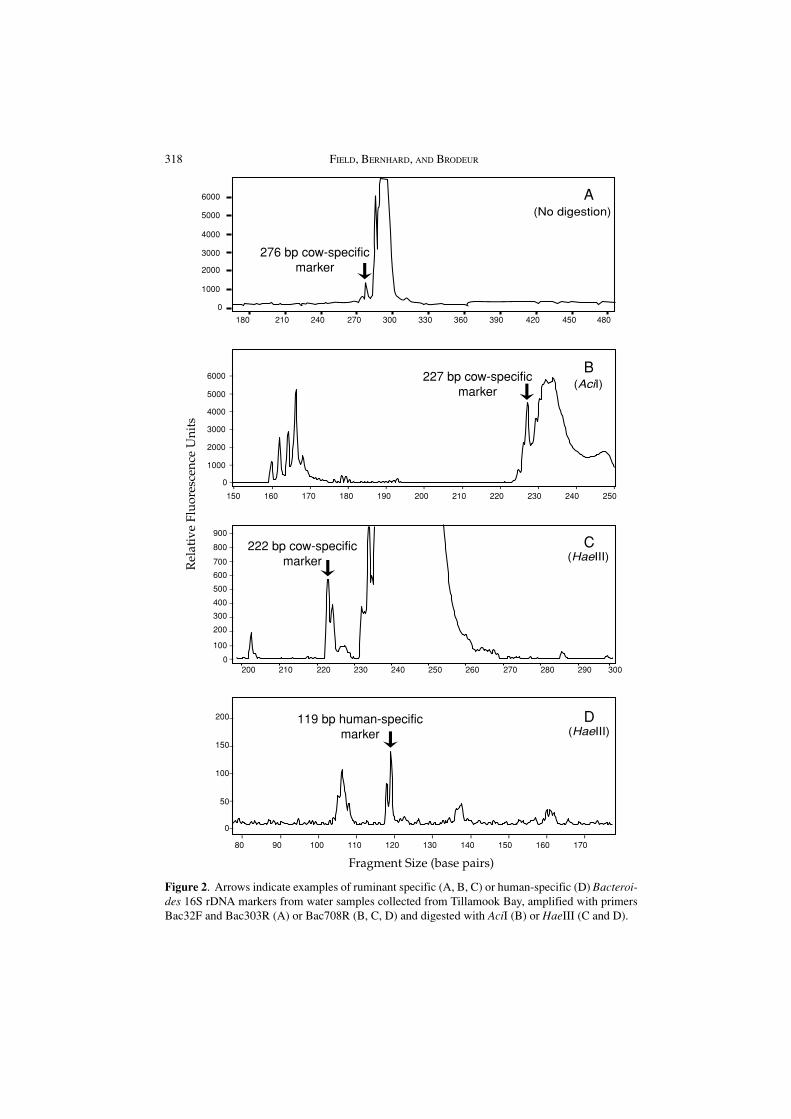
We also detected these host-specific Bacteroides markers in polluted watersamples from rivers and Tillamook Bay, Oregon (Figure 2), by filtering watersamples and extracting DNA from the filter. The Bacteroides signal was easilydetectable in all fecal samples and river and bay water samples.
T-RFLP and LH-PCR are expensive methods that require an automated DNAsequencer, and are thus not accessible to many laboratories. To solve this problem,
Figure 1. Example of T-RFLP analysis of Bacteroides 16S rDNA fragments. Arrows indicate rumi-nant- and human-specific markers. Fecal DNAs were amplified with PCR primers Bac32F-FAMand Bac708R and cut with restriction enzyme AciI. Solid lines are from human fecal DNA; dottedlines are from cow fecal DNA. Panel A shows all fragments from 200 to 250 base pairs; panel Bshows fragments from 475 to 525 base pairs.
Fragment Size (base pairs)
0
1000
2000
3000
4000
5000
6000
200 205 210 215 220 225 230 235 240 245
A
0
600
1200
1800
2400
3000
3600
4200
4800
5400
475 480 485 490 495 500 505 510 515 520
B
Rel
ativ
e F
luor
esce
nce
Uni
ts
318 FIELD, BERNHARD, AND BRODEUR
Figure 2. Arrows indicate examples of ruminant specific (A, B, C) or human-specific (D) Bacteroi-des 16S rDNA markers from water samples collected from Tillamook Bay, amplified with primersBac32F and Bac303R (A) or Bac708R (B, C, D) and digested with AciI (B) or HaeIII (C and D).
1000
0
2000
3000
4000
5000
6000
150 160 170 180 190 200 210 220 230 240 250
80 90 100 110 120 130 140 150 160 170
Fragment Size (base pairs)
Rel
ativ
e Fl
uore
scen
ce U
nits
200 210 220 230 240 250 260 270 280 290 300
100
200
300
400
500
600
700
800
900
0
0
50
100
150
200
A
B
D
C
0
1000
180 210 240 270 300 330 360 390 420 450 480
2000
3000
4000
5000
6000
276 bp cow-specificmarker
227 bp cow-specificmarker
119 bp human-specificmarker
222 bp cow-specificmarker
(No digestion)
(HaeIII)
(HaeIII)
(AciI)
MOLECULAR APPROACHES TO MICROBIOLOGICAL MONITORING: FECAL SOURCE DETECTION 319
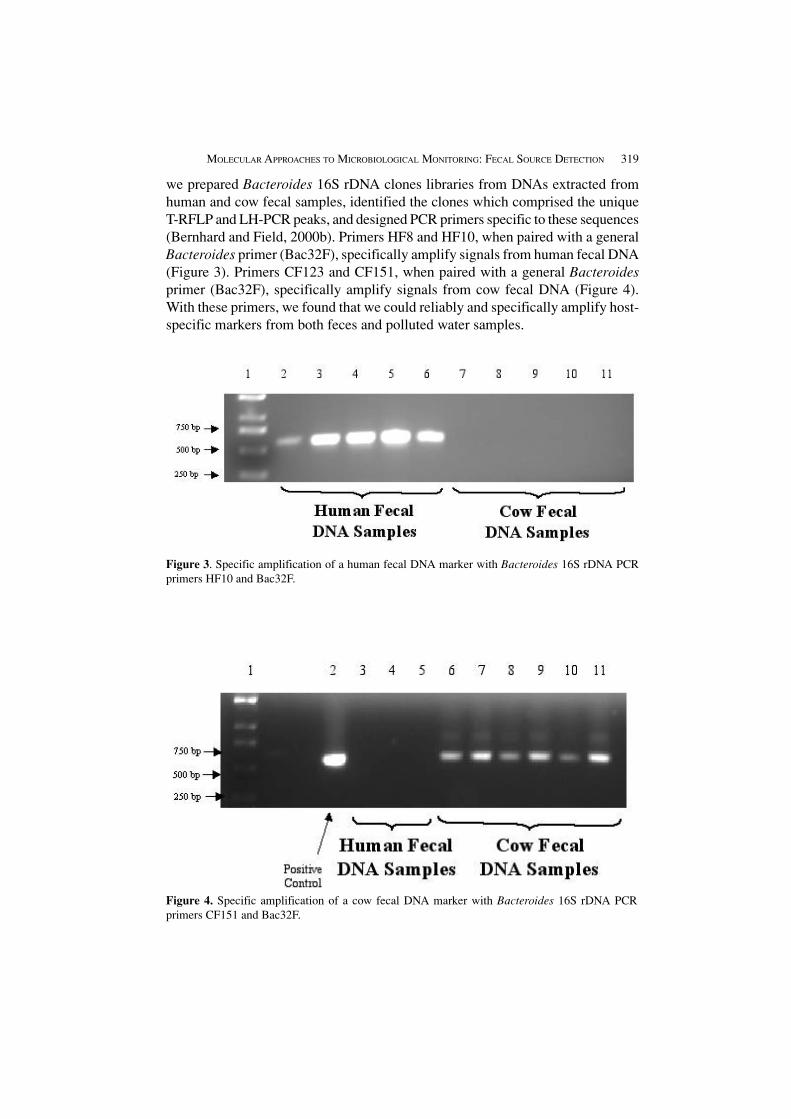
we prepared Bacteroides 16S rDNA clones libraries from DNAs extracted fromhuman and cow fecal samples, identified the clones which comprised the uniqueT-RFLP and LH-PCR peaks, and designed PCR primers specific to these sequences(Bernhard and Field, 2000b). Primers HF8 and HF10, when paired with a generalBacteroides primer (Bac32F), specifically amplify signals from human fecal DNA(Figure 3). Primers CF123 and CF151, when paired with a general Bacteroidesprimer (Bac32F), specifically amplify signals from cow fecal DNA (Figure 4).With these primers, we found that we could reliably and specifically amplify host-specific markers from both feces and polluted water samples.
Figure 3. Specific amplification of a human fecal DNA marker with Bacteroides 16S rDNA PCRprimers HF10 and Bac32F.
Figure 4. Specific amplification of a cow fecal DNA marker with Bacteroides 16S rDNA PCRprimers CF151 and Bac32F.

320 FIELD, BERNHARD, AND BRODEUR
Figure 5. Detection of fecal contamination with PCR/gel analysis using general Bacteroides primerscompared to fecal coliform counts (CFU) by membrane filtration. Cow feces were mixed with waterand diluted. Triplicate subsamples of each dilution were used for membrane filtration of fecal coliformsand DNA extraction followed by PCR detection. 1g wet fecal mass = 1.4 x 10-1 g dry fecal mass.
Using general Bacteroides PCR primers, fecal pollution can be detected withgreater sensitivity than by using membrane filtration of fecal coliforms. Figure 5shows cow fecal dilutions analyzed by PCR of Bacteroides markers and mem-brane filtration. Bacteroides markers were detected to well below the 14 CFU/100ml cutoff, above which shellfish cannot be harvested. However, the irregular-ity of the PCR bands at the lowest dilutions emphasizes that fecal material in thedilutions was somewhat unevenly dispersed.
The source-specific Bacteroides primers are somewhat less sensitive than thegeneral Bacteroides primers, because the bacterial groups they sample make uponly part of the total Bacteroides present. Nevertheless, they are of comparablesensitivity to standard coliform tests (Bernhard and Field, 2000b).
4. Are Genetic Markers from Fecal Bacteroides Universal?
The utility of this method of fecal source discrimination will depend on the rangeof applicability of the markers. Markers represent particular related groups of fe-cal bacteria, found in one host organism but not another. How widespread willthese groups be? Three lines of evidence suggest that the markers will be univer-sally distributed among members of each host group.
First, there is no overlap between cow and human Bacteroides sequences. Al-though LH-PCR and T-RFLP Bacteroides peaks from human and cow fecal DNAslargely overlapped, implying that the two hosts share many species, analysis of alarge number of Bacteroides gene sequences from cow and human feces found nosequence overlap between the two host species (Figure 6) (Mintie et al., 2000(abstract). Sequences from the human-specific markers were closely related to

MOLECULAR APPROACHES TO MICROBIOLOGICAL MONITORING: FECAL SOURCE DETECTION 321
Figure 6. Phylogenetic relationships among partial 16S rDNA sequences of clones from human(HF) and cow (CF) feces. The tree was inferred by neighbor joining. Numbers above internal branchesare bootstrap values; values below 50% are not shown.
Bacteroides fragilisBacteroides thetaiotaomicron
Bacteroides ovatusBacteroides caccae
Prevotella intermediaPrevotella melaninogenica
Prevotella ruminicolaPorphyromonas gingivalis
Porphyromonas endodontalis
Flavobacter aquatileCytophaga fermentans
Sphingobacter mizutaeFlexibacter canadensis
CF17CF157CF123
CF26CF138
CF68CF151
CF46
Bacteroides vulgatusHF145HF8HF117
HF102HF74
Bacteroides eggerthiiBacteroides uniformis
HF10Bacteroides stercoris
Bacteroides acidofaciens
.10
HF8 cluster
CF123 cluster
CF151 cluster
100
100
100
100
100 100
100
50
51
81
92
5059
60
77
99
76
76
86
8885
74
known human Bacteroides isolates such as B. uniformis. Sequences from the cow-specific markers formed a previously unknown lineage within the Cytophaga/Flexibacter/Bacteroides (CFB) division. Marker clones recovered from watersamples were members of the same lineages as the human and cow fecal markers.
Second, these markers occur throughout a broad phylogenetic group (ruminants),suggesting that they co-evolved with host groups. To determine the host specific-ity of the primers, we tested them with fecal DNAs collected from a broad range ofanimals (Bernhard and Field, 2000b). Although the human primers only amplified

322 FIELD, BERNHARD, AND BRODEUR
markers from human fecal DNA, the cow primers amplified markers from all ru-minant animals tested, included deer, elk, goat, and sheep, and even thepseudoruminant, llama. This suggests that the unique group of Bacteroides (Fig-ure 6) found in cows evolved in ruminants in general. If all ruminants have thisunique group, then it is virtually certain that cows in different geographic areaswill have it. Depending on choice of primer, either very general or very specificBacteroides groups can be differentiated.
Third, fecal samples from widespread areas of the United States have proven tohave the marker bacterial sequences. For example, primers developed for Oregoncow and human feces also amplify DNA markers from the Northeastern U. S.(Figures 3 and 4 are from a Northeastern U.S. study).
5. Protocol for Source Detection
A principal advantage of this approach to source discrimination is that it does notrequire culturing bacterial isolates. As a result, the method can be very rapid, witha turnaround time of a day or less. Conversely, samples can be stored before analy-sis if necessary or convenient, since the analysis does not require living cells. Thisallows for convenient and flexible field handling. In addition, scoring does notrequire complex pattern interpretation. Either the specific primers amplify a band,in which case that specific type of fecal pollution is present in the sample, or thereis no band, in which case the specific source of fecal pollution is absent or presentat a level below detection. A summary of the protocol is as follows:
1. Collect water samples (50 mL);
2. Filter samples at time of collection, store filters in lysis buffer;
3. Extract DNA from filters with commercial kit;
4. Perform PCR with diagnostic primers; and
5. Score PCRs: if there is a product, sample is positive.
Instead of filtering water samples in the field and storing the filters in preserva-tive, samples can be returned to the lab, filtered and either stored or analyzedimmediately. In that case, we always filter and process water samples within sixhours of collection.
The major pieces of equipment needed are a thermal cycler, for PCR amplifica-tion, and equipment to inspect the PCR products. The commonest and least expen-sive method of visualizing PCR products is gel electrophoresis utilizing a DNAstain. This requires a gel box, power source, light or UV light source, and cameraor gel documentation system. Also required is filtration equipment (disposablesyringes and filters in the field, or vacuum flasks and a vacuum source in the lab)and a microfuge and lab ware for DNA extraction.

MOLECULAR APPROACHES TO MICROBIOLOGICAL MONITORING: FECAL SOURCE DETECTION 323
This method of source discrimination can be conveniently adapted to high-throughput analysis. For example, DNA extraction, PCR, and scoring (by fluores-cence) can all be done by robot.
6. Future Experimental Work
We are currently developing primers for other animal species, and for distinguish-ing wild from domestic ruminants. In addition, in both lab and field trials, we areestablishing the comparative sensitivity of each primer, relative to general Bacteroi-des primers, each other, and coliforms and E. coli-based methods. Finally, we aredeveloping a quantitative assay, using real-time quantitative PCR.
Acknowledgements
We are grateful for assistance from Caragwen Bracken. This work was partiallysupported by Grant NA76RG0476 (project no. R/ECO-04) from the National Oce-anic and Atmospheric Administration to the Oregon Sea Grant College Program,by appropriations made by the Oregon State Legislature, by Grant R827639-01-0from the U. S. Environmental Protection Agency, and by Grant 00-51130-9818from the U. S. Department of Agriculture.
ReferencesAllsop, K., and Stickler, J.D.: 1985, ‘An assessment of Bacteroides fragilis group organisms as
indicators of human faecal pollution’, J. Appl. Bacteriol. 58, 95–99.
Andrews, R. W., Conway, R. A., Corr, C. L., Dobratz, E. J., Dougherty, D. P., Eppard, J. R., Knupp,S. R., Limjoco, M. C., Mettenburg, J. M., Rinehardt, J. M., Sonsino, J., Torrijos, R. L.,Zimmerman, M. E., and Wiggins, B. A.: 1997, ‘Classification of fecal streptococci iso-lated from potential sources of fecal pollution using discriminant analysis: supporting evi-dence from large datasets’, Abstracts, American Society of Microbiology General Meet-ing, Miami Beach, Florida.
Avaniss-Aghajani, E. A., Jones, K., Chapman, D., and Brunk, C.: 1994, ‘A molecular technique foridentification of bacteria using small subunit ribosomal RNA sequences’, BioTechniques17, 144–149.
Avelar, K. E. S., Morales, S. R., Pinto, L. J. F., Silva e Sousa, R. M. C. P. D., and Ferreira, M. C. d. S.:1998, ‘Influence of stress conditions of Bacteroides fragilis survival and protein profiles’,Zent. bl. Bakteriol. 287, 399–409.
Bernhard, A. E., and Field, K.G.: 2000a, ‘Identification of nonpoint sources of fecal pollution incoastal waters by using host-specific 16S ribosomal DNA genetic markers from fecal anaer-obes’, Appl. Environ. Microbiol. 66, 1587–1594.
Bernhard, A. E., and Field, K. G.: 2000b, ‘A PCR assay to discriminate human and ruminant feces onthe basis of host differences in Bacteroides-Prevotella genes encoding 16S rRNA’, Appl.Environ. Microbiol. 66, 4571–4574.
Bruce, K. D.: 1997, ‘Analysis of mer gene subclasses within bacterial communities in soils andsediments resolved by fluorescent-PCR-restriction fragment length polymorphism profil-ing’, Appl. Environ. Microbiol. 63, 4914–4919.

324 FIELD, BERNHARD, AND BRODEUR
Brunk, C. F., Avaniss-Aghajani, E., and Brunk, C. A.: 1996, ‘A computer analysis of primer andprobe hybridization potential with bacterial small-subunit rRNA sequences’, Appl. Environ.Microbiol. 62, 872–879.
Carson, C. A., Shear, B. L., Ellershiek, M. R., and Asfaw, A.: 2001, ‘Identification of fecal Escherichiacoli from humans and animals by ribotyping’, Appl. Environ. Microbiol. 67, 1503–1507.
Clement, B. G., Kehl, L. E., DeBord, K. L., and Kitts, C. L.: 1998, ‘Terminal restriction fragmentpatterns (TRFPs), a rapid, PCR-based method for the comparison of complex bacterialcommunities’, J. Microbiol. Methods 31, 135–142.
DePaola, A., Hophins, L. H., Peeler, J. T., Wentz, B., and McPhearson, R. M.: 1990, ‘Incidence ofVibrio parahaemolyticus in U. S. coastal waters and oysters’, Appl. Environ. Microbiol.56, 2299–2302.
Dombek, P. E., Johnson, L. K., Zimmerley, S. T., and Sadowsky, M. J.: 2000, ‘Use of repetitive DNAsequences and the PCR to differentiate Escherichia coli isolates from human and animalsources’, Appl. Environ. Microbiol. 66, 2572–2577.
Gary, H. L., and Adams, J. C.: 1985, ‘Indicator bacteria in water and stream sediments near thesnowy range in southern Wyoming’, Water, Air, Soil Pollut. 25, 133–144.
Gerba, C. P., and McLeod, J. S.: 1976, ‘Effect of sediments on the survival of Escherichia coli inmarine waters’, Appl. Environ. Microbiol. 32, 114–120.
Goyal, S. M., Zerda, K. S., and Gerba, C. P.: 1980, ‘Concentration of coliphage from large volumesof water and wastewater’, Appl. Environ. Microbiol. 59, 2956–2962.
Harwood, V. J., Butler, J., Parrish, D., and Wagner, V.: 1999, ‘Isolation of fecal coliform bacteriafrom the diamondback terrapin (Malaclemys terrapin centrata)’, Appl. Environ. Microbiol.65, 865–867.
Havelaar, A. H., van Olphen, M., and Drost, Y. C.: 1993, ‘F-specific RNA bacteriophages are ad-equate model organisms for enteric viruses in fresh water’, Appl. Environ. Microbiol. 59,2956–2962.
Holdeman, V., Cato, E. P., and Moore, W. E. C.: 1976, ‘Human fecal flora: variation in bacterialcomposition within individuals and a possible effect of emotional stress’, Appl. Environ.Microbiol. 31, 359–375.
Hood, M. A., and Ness, G. E.: 1982, ‘Survival of Vibrio cholerae and Escherichia coli in estuarinewaters and sediments’, Appl. Environ. Microbiol. 43, 578–584.
Howell, J. M., Coyne, M. S., and Cornelius, P. L.: 1996, ‘Effect of sediment particle size and tem-perature on fecal bacteria mortality rates and the fecal coliform/fecal streptococci ratio’, J.Environ. Qual. 25, 1216–1220.
Kreader, C. A.: 1995, ‘Design and evaluation of Bacteroides DNA probes for the specific detectionof human fecal pollution’, Appl. Environ. Microbiol. 61, 1171–1179.
Kreader, C. A.: 1998, ‘Persistence of PCR-detectable Bacteroides distasonis from human feces inriver water’, Appl. Environ. Microbiol. 64, 4103–4105.
LaLiberte, P., and Grimes, D. J.: 1982, ‘Survival of Escherichia coli in lake bottom sediment’, Appl.Environ. Microbiol. 43, 623–628.
Liu, W.-T., Marsh, T. L., Cheng, H., and Forney, L. J.: 1997, ‘Characterization of microbial diversityby determining terminal restriction fragment length polymorphisms of genes encoding16S rRNA’, Appl. Environ. Microbiol. 63, 4516–4522.
Mintie, A., Bernhard, A. E., Trippet, D., Cooper, B., Buccafurni, D., and Field, K. G.: 2000 (ab-stract), ‘Phylogenetic diversity of Bacteroides-Prevotella 16S rRNA genes from cow andhuman feces’, American Society for Microbiology, Los Angeles, CA.

MOLECULAR APPROACHES TO MICROBIOLOGICAL MONITORING: FECAL SOURCE DETECTION 325
Moore, W. E. C., and Holdeman, L. V.: 1974, ‘Human fecal flora: the normal flora of 20 Japanese-Hawaiians’, Appl. Microbiol. 27, 552–555.
Osawa, S., Furuse, K., and Watanabe, I.: 1981, ‘Distribution of ribonucleic coliphages in animals’,Appl. Environ. Microbiol. 41, 164–168.
Palmateer, G. A., Dutka, E. M., Jantzen, E. M., Meissner, S. M., and Sakeellaries, M. G.: 1991,‘Coliphage and bacteriophage as indicators of recreational water quality’, Water Research25, 355–357.
Parveen, S., Portier, K. M., Robinson, K., Edmiston, L., and Tamplin, M.: 1999, ‘Discriminate analysisof ribotype profiles of Escherichia coli for differentiating human and nonhuman sourcesof fecal pollution’, Appl. Environ. Microbiol. 65, 3142–3147.
Paster, B. J., Dewhirst, F. E., Olsen, I., and Fraser, G. J.: 1994, ‘Phylogeny of Bacteroides, Prevotella,and Porphyromonas spp. and related bacteria’, J. Bacteriol. 176, 725–732.
Paul, J. H., Rose, J. B., Jiang, S. C., London, P., Xhou, X., and Kellogg, C.: 1997, ‘Coliphage andindigenous phage in Mamala Bay, Oahu, Hawaii’, Appl. Environ. Microbiol. 63, 133–138.
Pommepuy, M., Guillaud, J. F., Dupray, E., Derien, A., Le Guyader, F., and Cormier, M.: 1992,‘Enteric bacteria survival factors’, Water Sci. Technol. 12, 93–103.
Rappé, M. S., Kemp, P. F., and Giovannoni, S. J.: 1997, ‘Phylogenetic diversity of marine coastalpicoplankton 16S rRNA genes cloned from the continental shelf off Cape Hatteras, NorthCarolina’, Limnol. Ocean. 42, 811–826.
Salyers, A. A.: 1984, ‘Bacteroides of the human lower intestinal tract’, Ann. Rev. Microbiol. 38,293–313.
Sherer, B. M., Miner, J. R., Moore, J. A., and Buckhouse, J. C.: 1988, ‘Resuspending organisms froma rangeland stream bottom’, Trans. Amer. Soc. Ag. Eng. 31, 1217–1222.
Singh, S. N., and Gerba, C. P.: 1983, ‘Concentration of coliphages from water and sewage withcharge-modified filter aid.’, Appl. Environ. Microbiol. 45, 232–237.
Sinton, L. W., Donnison, A. M., and Hastie, C. M.: 1993, ‘Faecal streptococci as faecal pollutionindicators: a review. Part II: Sanitary significance, survival, and use’, New Zealand J. MarineFreshwater Res. 27, 117–137.
Sobsey, M. D.: 1989, ‘Inactivation of health-related microorganisms in water by disinfection pro-cesses’, Water Sci. Technol. 21, 179–195.
Straub, D. V., and Dixon, B.: 1997, ‘Bacteroides vulgatus, an alternative indicator for the assessmentof fecal contamination of shellfish and estuarine waters’, 38th Annual Western Fish Dis-ease Workshop, Bodega Marine Laboratory, Bodega Bay, CA.
Suzuki, M. T., Rappé, M. S., Haimberger, Z. W., Winfield, H., Adair, N., Ströbel, J., and Giovannoni,S. J.: 1997, ‘Bacterial diversity among small-subunit rRNA gene clones and cellular iso-lates from the same seawater sample’, Appl. Envir. Microbiol. 63, 983–989.
Tartera, C., and Jofre, J.: 1987, ‘Bacteriophages active against Bacteroides fragilis in sewage pol-luted waters’, Appl. Environ. Microbiol. 53, 1632–1637.
Tartera, C., Lucena, F., and Jofre, J.: 1989, ‘Human origin of Bacteroides fragilis bacteriophagespresent in the environment’, Appl. Environ. Microbiol. 55, 2696–2701.
Weiskel, P. K., Howes, B. L., and Heufelder, G. R.: 1996, ‘Coliform contamination of a coastalembayment: sources and transport pathways’, Environ. Sci. Technol. 30, 1872–1881.
Wiggins, B. A.: 1996, ‘Discriminant analysis of antibiotic resistance patterns in fecal streptococci, amethod to differentiate human and animal sources of fecal pollution in natural waters’,Appl. Environ. Microbiol. 62, 3997–4002.

326 FIELD, BERNHARD, AND BRODEUR
Wiggins, B. A., Andrews, R. W., Conway, R. A., Corr, C. L., Dobratz, E. J., Dougherty, D. P., Eppard,J. R., Knupp, S. R., Limjoco, M. C., Mettenburg, J. M., Rinehardt, J. M., Sonsino, J.,Torrijos, R. L., and Zimmerman, M. E.: 1999, ‘Use of antibiotic resistance analysis toidentify nonpoint sources of fecal pollution’, Appl. Environ. Microbiol. 65, 3483–3486.




























