Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, Feb. 2011, p. 1528–1540 Vol. 85, No. 40022-538X/11/$12.00 doi:10.1128/JVI.01709-10Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Kaposi’s Sarcoma-Associated Herpesvirus ORF57 Interacts withCellular RNA Export Cofactors RBM15 and OTT3 To
Promote Expression of Viral ORF59�†Vladimir Majerciak,1 Hiroaki Uranishi,2 Michael Kruhlak,3 Guy R. Pilkington,4
Maria Julia Massimelli,1 Jenifer Bear,4 George N. Pavlakis,2Barbara K. Felber,4 and Zhi-Ming Zheng1*
Tumor Virus RNA Biology Section, HIV and AIDS Malignancy Branch,1 Human Retrovirus Section2 andHuman Retrovirus Pathogenesis Section,4 Vaccine Branch, and Experimental Immunology Branch,3
Center for Cancer Research, National Cancer Institute, Bethesda, Maryland 20892
Received 12 August 2010/Accepted 17 November 2010
Kaposi’s sarcoma-associated herpesvirus (KSHV) encodes ORF57, which promotes the accumulation ofspecific KSHV mRNA targets, including ORF59 mRNA. We report that the cellular export NXF1 cofactorsRBM15 and OTT3 participate in ORF57-enhanced expression of KSHV ORF59. We also found that ectopicexpression of RBM15 or OTT3 augments ORF59 production in the absence of ORF57. While RBM15 promotesthe accumulation of ORF59 RNA predominantly in the nucleus compared to the levels in the cytoplasm, wefound that ORF57 shifted the nucleocytoplasmic balance by increasing ORF59 RNA accumulation in thecytoplasm more than in the nucleus. By promoting the accumulation of cytoplasmic ORF59 RNA, ORF57offsets the nuclear RNA accumulation mediated by RBM15 by preventing nuclear ORF59 RNA from hyper-polyadenylation. ORF57 interacts directly with the RBM15 C-terminal portion containing the SPOC domainto reduce RBM15 binding to ORF59 RNA. Although ORF57 homologs Epstein-Barr virus (EBV) EB2, herpessimplex virus (HSV) ICP27, varicella-zoster virus (VZV) IE4/ORF4, and cytomegalovirus (CMV) UL69 alsointeract with RBM15 and OTT3, EBV EB2, which also promotes ORF59 expression, does not function likeKSHV ORF57 to efficiently prevent RBM15-mediated nuclear accumulation of ORF59 RNA and RBM15’sassociation with polyadenylated RNAs. Collectively, our data provide novel insight elucidating a molecularmechanism by which ORF57 promotes the expression of viral intronless genes.
Kaposi’s sarcoma-associated herpesvirus (KSHV) ORF57 isa viral early protein. Like its homologues herpes simplex virustype 1 (HSV-1) ICP27 (38), Epstein-Barr virus (EBV) EB2(40), human cytomegalovirus (HCMV) UL69 (47), and vari-cella-zoster virus (VZV) IE4/ORF4 (32), KSHV ORF57 reg-ulates virus gene expression at the posttranscriptional level(23). The KSHV genome with a disrupted ORF57 gene cannotefficiently express a subset of viral lytic genes (20). ORF57functions as a viral splicing factor and promotes viral RNAsplicing (19, 21, 23). ORF57 also promotes the expression ofviral intronless genes, such as ORF59, a viral DNA polymeraseprocessivity factor. The specific ORF57 interactions with itstargeted RNAs are facilitated by cellular proteins (23). It hasbeen proposed that ORF57 stimulates RNA export via itsinteraction with Aly/REF, a cellular RNA-binding proteinserving as an adaptor for the nuclear RNA export receptorNXF1/TAP (24). However, recent reports indicate that Aly/REF-ORF57 interaction does not play an important role in theORF57-mediated enhancement of ORF59 expression, sinceAly/REF knockdown in host cells did not affect the function of
ORF57 (22, 29). In contrast, a recent study indicates thatORF57 appears to recruit the entire TREX through its inter-action with Aly/REF to facilitate the export of a viral latetranscript, ORF47 (2). Since ORF57 specifically binds toORF59 RNA only in the presence of cellular proteins, it isthought that the cellular adaptors may act as cofactors. Insupport of this, the posttranscriptional regulator EB2 of EBV,a member of the gammaherpesvirus subfamily, was reported tointeract with OTT3 (also referred to as RNA binding motifprotein 15B [RBM15B]) and OTT3 was shown to have post-transcriptional regulatory function (6).
OTT3 (6, 26), RNA binding motif protein 15 (RBM15[OTT1]) (16, 26), and SHARP (SMRT/HDAC1-associated re-pressor protein) (30, 41) are members of the SPEN (split ends)protein family. These nuclear proteins share a domain struc-ture comprising three highly conserved N-terminal RNA-bind-ing motifs (RNA recognition motif [RRM]) and a highly con-served C-terminal SPOC (Spen paralogue and orthologueC-terminal) domain, with 52%, 68%, and 82% sequence iden-tity for RRM1, RRM2, and RRM3, respectively, and 58%amino acid (aa) sequence identity in their SPOC domain (6,16, 37, 50). SPEN homologs are conserved in eukaryotes andare found from Caenorhabditis elegans to humans but showdistinct functional properties (6, 13, 49). While SHARP (3,664aa residues) acts at the transcriptional level (3, 10, 12, 30, 31,39, 41) and has also been found in association with spliceo-somes (51), both of the two smaller proteins, RBM15 (957 to977 aa residues for the different isoforms) and OTT3 (890 aa
* Corresponding author. Mailing address: HIV and AIDS Malig-nancy Branch, Center for Cancer Research, NCI/NIH, 10 CenterDrive, Room 6N106, Bethesda, MD 20892-1868. Phone: (301) 594-1382. Fax: (301) 480-8250. E-mail: [email protected].
† Supplemental material for this article may be found at http://jvi.asm.org/.
� Published ahead of print on 24 November 2010.
1528

residues), act at the posttranscriptional level (6, 13, 49). Weand others have reported that the SPOC-containing C-termi-nal region of RBM15 and OTT3 mediates the RNA exportfunction (6, 13, 49), while SHARP lacks this function, support-ing divergent functions of RBM15 and OTT3. Ectopically ex-pressed OTT3 was reported to act as a splicing regulator (6).RBM15 was determined to bind directly to the RNA transportelement (RTE) (13), which is exported via the NXF1 pathway(42). RTE is present in murine intracisternal A-particle retro-elements (28, 43) and is essential for the activity of the retro-transposon (52). RBM15 and OTT3 were shown to promotethe export and expression of RTE-containing transcripts (13,49). Both proteins localize to the nucleus and associate withthe splicing factor compartment (SC35-containing speckles)and the nuclear envelope but do not shuttle to the cytoplasm(6, 13, 49). Both proteins can also interact with each other (49).RBM15 and OTT3 act as cofactors to the nuclear export re-ceptor NXF1 and interact directly with NXF1 via the C-termi-nal region (13, 49). We recently reported that RBM15 alsoprovides a direct molecular link between NXF1 and DEADfamily RNA helicase DBP5 (53), supporting the model inwhich RBM15 acts locally at the nuclear pore complex, facil-itating the recognition of NXF1-mRNP complexes by DBP5during translocation and thereby contributing to efficientmRNA export. Genetic knockout of RBM15 is embryonic le-thal in homozygous mice (35, 36, 49), indicating that OTT3cannot compensate for the loss of RBM15. These findingssupport the model that, in addition to shared functions, thesefactors have distinct biological roles in animal development. Inthe study presented herein, we demonstrated that bothRBM15 and OTT3, like KSHV ORF57, promote KSHVORF59 expression. We identified the direct interaction ofKSHV ORF57 with RBM15 and OTT3 and showed that theinteraction with RBM15 and OTT3 is conserved amongORF57 homologs of four other herpesvirus family members.
MATERIALS AND METHODS
Cell cultures and cotransfection. HEK293 and HeLa cell lines from ATCC(Manassas, VA) were cultivated in Dulbecco’s modified Eagle’s medium supple-mented with 10% fetal bovine serum as described before (22). The cells, at 2.5 �105 per ml in a 60-cm dish, were cotransfected with plasmid DNA at 20, 50, or100 ng for the ORF57 expression vector, at 20 or 100 ng for the RBM15expression vector, at 100 ng for the OTT3 expression vector, and at 300 ng for theORF59 expression vector in the presence of Lipofectamine 2000 (Invitrogen,Carlsbad, CA) or Fugene HD (Roche Applied Science, Indianapolis, IN).
Plasmids. Vector pEGFP-N1 was obtained from Clontech (Mountain View,CA). All other plasmids used in this study were previously described, as follows:pVM18 (FLAG-tagged KSHV ORF59), pVM68 (3�FLAG-tagged KSHVORF57), pVM90 (3�FLAG-tagged KSHV ORF57 mutant mtNLS2 � 3), pVM8(green fluorescent protein [GFP]-tagged wild-type [wt] ORF57) and its mutantpVM36 (mtNLS2 � 3) (22), FLAG-tagged RBM15 and OTT3 or hemagglutinin(HA)-tagged OTT3, as well as their truncation mutants (13, 49), or HA-taggedRef1-II (53). Enhanced GFP (EGFP)-tagged ICP27 (a gift from R. Sandri-Goldin), Myc-tagged EB2 (a gift from S. Swaminathan), VZV IE4-V5 (a giftfrom C. Sadzot), and FLAG-tagged UL69 (a gift from T. Stamminger) wereused.
RNA interference (RNAi). HeLa cells were transfected with 40 nM ON-TARGETplus SMARTpool small interfering RNA (siRNA) (Thermo Scientific,Waltham, MA) targeting endogenous RBM15 protein or a nonspecific siRNA.At 24 h after siRNA transfection, the cells were transfected with 500 ng ofORF59 expression vector (pVM18) and incubated for an additional 24 h. Totalprotein and RNA were isolated as described previously (22) for Western andNorthern blot analysis, respectively.
Western blot analysis. Protein extracts were prepared by direct cell lysis inSDS protein sample buffer and analyzed in Western blots as described previously(22). The following monoclonal antibodies were used in the study: anti-GFP (BDBiosciences, Mountain View, CA), anti-FLAG (Sigma, St. Louis, MO), anti-myc(Sigma), anti-HA (Sigma), anti-tubulin (Sigma), anti-RBM15 (Protein TechGroup, Chicago, IL), anti-V5 horseradish peroxidase (HRP) (Invitrogen), andanti-ORF59 (Advanced Biotechnologies, Columbia, MD). Rabbit anti-ORF57was previously described (20).
Isolation of nuclear and cytoplasmic RNA. Cytoplasmic and nuclear totalRNAs were fractionated as described previously (22). Briefly, after transfection,the cells were trypsinized, washed with phosphate-buffered saline (PBS), andresuspended in cold buffer A (50 mM Tris, pH 8.0, 140 mM NaCl, 1.5 MgCl2,
0.2% NP-40, 1 mM dithiothreitol [DTT], and 200 U RNasin). After 5 min ofincubation on ice, the nuclei were pelleted by spinning for 2 min at 3,000 � g at4°C. Both the supernatant for the cytoplasmic fraction and the pellet for thenuclear fraction were collected for RNA isolation by using TRIzol reagent(Invitrogen).
Northern blot analysis. RNA (�5 �g) was separated in agarose gel andanalyzed in Northern blots as described previously (22). The following oligonu-cleotide probes labeled with �-32P were used in this study: oVM73 (5�-GTCCACCCTGACCCCATAGT-3�) for ORF59, oZMZ270 (5�-TGAGTCCTTCCACGATACCAAA-3�) for glyceraldehyde-3-phosphate dehydrogenase (GAPDH),oVM11 (5�-CTCGTCTTCCAGTGTCGGTG-3�) for ORF57, and oST197 (5�-AAAATATGGAACGCTTCACGA-3�) for U6 snRNA. The hybridization sig-nal was captured using a Molecular Dynamics PhosphorImager Storm 860 andanalyzed with ImageQuant software.
Indirect immunofluorescence staining. For indirect immunofluorescencestaining (IFA), cells growing on coverslips were fixed with 4% paraformaldehydefor 15 min at room temperature, permeabilized for 10 min at room temperaturewith 0.5% Triton X-100, and blocked with 2% bovine serum albumin (BSA) inPBS with the addition of 0.02% Tween 20 (blocking buffer) for 1 h at 37°C.Primary antibodies were diluted in blocking buffer and incubated with samplesfor 1 h at 37°C, followed by 3 washes with PBS. Secondary antibodies conjugatedwith Alexa Fluor 546 dye (Molecular Probes, Eugene, OR) were diluted 1:500 inblocking buffer and incubated for 1 h at 37°C. After a final 3 washes with PBS,the cell nuclei were stained with DAPI (4�,6�-diamidino-2-phenylindole) and allsamples were mounted in ProLong Gold mounting medium (Invitrogen). Con-focal fluorescence images were collected using a Zeiss LSM510 META (CarlZeiss MicroImaging, Inc., Thornwood, NY) laser scanning microscope, andcolocalization analysis was performed using the colocalization module of Imarisversion 6.0 software (Bitplane) as described previously (21).
Immunoprecipitation. For coimmunoprecipitation (co-IP) assays, HEK293cells were transfected with the indicated expression constructs and harvested 2days later. The cells were extracted under stringent conditions (400 mM NaCl, 50mM KCl, 0.2% Triton X-100, 15 mM HEPES, pH 7.9, 10% glycerol, proteaseinhibitors) (13). The extracts were treated with RNase A (0.5 mg/ml) for 10 minat room temperature, incubated with anti-FLAG M2 or anti-HA antibody beadsfor 1 h, and subsequently washed with lysis buffer containing 2 M urea, asdescribed previously (49). Co-IPs under physiological conditions were performedin the same buffer (0.2% Triton X-100, 15 mM HEPES, pH 7.9, 10% glycerol,protease inhibitors) containing 150 mM NaCl, using a wash buffer which did notcontain urea. The protein complexes were eluted by boiling the beads in SDS-PAGE loading buffer. The complexes were subjected to Western blot analysis,and the blots were probed with anti-GFP, anti-FLAG, anti-HA, anti-V5, oranti-myc antibodies. In vitro protein binding assays were performed (48) usingEscherichia coli-expressed purified glutathione S-transferase (GST)-taggedRBM15 (aa 530 to 977), GST-tagged OTT3 (aa 488 to 890), or GST only andmetabolically labeled, reticulocyte-produced ORF57, UAP56, or firefly lucifer-ase protein, respectively.
RNase H digestion. The length of the mRNA poly(A) tail was determined byRNase H digestion as described by Murray and Schoenberg (27), with smallmodifications. Briefly, 10 �g of RNA in a total volume of 7 �l was denatured for5 min at 85°C. Two microliters of 5 nmol oligo(dT16) (Applied Biosystems,Carlsbad, CA) was added to RNA and annealed for 10 min at 42°C. Each RNAsample was supplemented with 1 �l of 10� RNase H buffer (200 mM Tris, pH8.0, 500 mM KCl, 100 mM MgCl2, 10 mM DTT) and split into two tubes (5 �leach). One tube of the RNA was digested with 10 U of RNase H (Fermentas,Glen Burnie, MD) for 30 min at 30°C, and the other tube of the RNA served asan undigested RNA control. After digestion, 15 �l of formaldehyde loading dyewas added directly to each sample and the samples were incubated for 15 min at75°C and then analyzed by Northern blotting as described above.
UV cross-linking and isolation of polyadenylated RNA. The detection ofproteins associated with polyadenylated RNAs was performed as described pre-
VOL. 85, 2011 RNA EXPORT COFACTORS AND KSHV ORF57 1529

viously (33). Briefly, transfected cells washed with PBS were exposed to UV light(4,800 mJ/cm2) and resuspended in PBS. One third of the cell suspension wasspun, and the cell pellet was lysed directly in SDS protein sample buffer to serveas an input control. The rest of the cells were used for the isolation of polyade-nylated RNAs using an Illustra QuickPrep micro mRNA purification kit (GEHealthcare, Piscataway, NJ) according to the manufacturer’s instructions. Theeluted mRNA-protein complexes were treated with RNase T1 (5 U/sample) at37°C for 15 min and analyzed by Western blotting.
UV cross-linking and immunoprecipitation (CLIP). HEK293 cells (5 � 105
cells/well in a 6-well plate) were cotransfected with RBM15-FLAG and ORF59-FLAG, 500 ng each, in the presence or absence of ORF57-GFP. At 24 h aftercotransfection, the cells were washed with PBS and UV irradiated (4,800 mJ/cm2). Total cell extracts after UV cross-linking were prepared by lysis directly inradioimmunoprecipitation (RIPA) buffer with protease inhibitors added. Pro-tein-RNA complexes were immunoprecipitated as previously described (22) us-ing anti-FLAG M2 mouse monoclonal antibody. Nonspecific mouse IgG wasused as a negative control. Immunoprecipitated RNA digested by proteinase Kwas amplified by RT-PCR with ORF59-specific oligonucleotides (5� primeroST179, 5�-TAATACGACTCACTATAGG/GTCACCTGATTGGCAGCC-3�,and 3� primer oST180, 5�-AACGCATCGAG/CGGTGACTGTGTCTGTCAGC-3�; slashes separate the nonspecific sequences from the specific viral se-quences) or with GAPDH-specific oligonucleotides (21) and analyzed in agarosegel.
Quantitative RT-PCR and RNA stability analysis. RNA samples obtainedfrom CLIP assays were treated with Turbo DNase I (Ambion, Austin, TX)according to the manufacturer’s instructions and subjected to TaqMan quanti-tative reverse transcription-PCR (qRT-PCR) using TaqMan reverse transcrip-tion reagents and TaqMan universal master mix (Applied Biosystems). KSHVORF59 transcripts were amplified and detected by using the following TaqManprimers: ORF59 probe (5� 56-FAM-AAACCGATCTGTGTCTGCCGAGG-3IABkFG 3�), ORF59 primer 1 (5� TTAGAAGTGGAAGGTGTGCC 3�), andORF59 primer 2 (5� TCCTGGAGTCCGGTATAGAATC 3�). After back-ground subtraction, the cycle threshold (CT) values of the qRT-PCR data from2 repeats, each in triplicate, were analyzed by the 2���CT or 2��CT method (14)and are presented as bar graphs with means standard deviations.
For RNA stability assays, HEK293 cells were transfected with ORF59 expres-sion vector pVM18 plus FLAG-tagged ORF57 expression vector pVM7, FLAG-tagged RBM15 or empty pFLAG-CMV-5.1 vector control. Since the expressionlevel of ORF59 is very low in the absence of ORF57 or RBM15, HEK293 cells(1 � 106) in 60-mm dishes were cotransfected with 2 �g of ORF59 expressionplasmid plus the appropriate amount of pFLAG-CMV-5.1 vector to keep theratio as 5:1 or 15:1 as a control for the ORF57 or RBM15 expression vectors,using FuGene HD (Roche). Eighteen hours after transfection, actinomycin D(ActD) was added to the cells at a final concentration of 10 �g/ml, and total cellRNA was extracted at the designated time points by using an RNeasy kit (Qia-gen), followed by poly(A) mRNA enrichment with an Oligotex mini mRNA kit(Qiagen). Approximately 100 to 200 ng of poly(A) mRNA was reverse tran-scribed with a poly(T) oligonucleotide. Real-time PCR for ORF59 RNA andGAPDH RNA was carried out on the cDNA in duplicates by using TaqManUniversal Master mix (Applied Biosystems). The GAPDH RNA level in eachreaction mixture, determined by using TaqMan GAPDH control probes (catalognumber 402869; Applied Biosystems), served as a loading control and was usedto quantify the ORF59 RNA. The relative level of expression of ORF59 wasdetermined using the CT method as described above and converted to a per-centage, with the amount at the moment of the addition of ActD (time zero), setas 100%, representing the percentage of remaining mRNA. A nonlinear regres-sion analysis on the raw data (fold percent) was performed, choosing an expo-nential decay model {[fold % � � exp(� � t)], where � is the intercept at timezero � is the decay rate, and t is time}. The � was constrained to be either lessthan or equal to 100% (� � 100%) or equal to 100% (� 100%), and the � tobe less than or equal to zero (� � 0). Weighted linear regression results weregraphed, and a regression curve was employed for half-life calculation. Compar-ison of parameter estimates (� only if � equals 100%, and � and � if � is less thanor equal to 100%) between the treatments was performed by using a model witha dummy variable.
RESULTS
RBM15 promotes ORF59 protein expression in the absenceor presence of ORF57. As ORF57 promoted intronless ORF59expression in our previous studies (19, 20, 22), we testedwhether RBM15 and OTT3 would potentiate ORF57 to en-
hance ORF59 expression upon cotransfection of HEK293cells. Unexpectedly, we found that cotransfection of theORF59 plasmid with RBM15 alone led to increased levels ofboth ORF59 protein and RNA in the absence of ORF57 (Fig.1A and B, compare lanes 1 and 2 as well as the correspondingbar graphs of RNA levels). The enhanced expression ofORF59 protein and RNA was observed in cells transfectedwith 20 ng of the RBM15 expression vector. Comparing theexogenous to the endogenous RBM15 levels in transfectedHEK293 cells showed that the level of exogenous RBM15 withthe 20-ng transfection was about 30% less than its endogenouslevel in our experimental conditions (see Fig. S1 in the sup-plemental material). Our data indicate that ectopic RBM15near its endogenous level was enough to further augmentORF59 expression in the absence of ORF57.
We next tested whether RBM15 at this low dose (20 ng)could synergize with ORF57 to further augment ORF59 ex-
FIG. 1. Ectopic RBM15 promotes and cooperates with ORF57 inORF59 expression. HEK293 cells at 2.5 � 105 per ml were cotrans-fected with 20 ng of RBM15-FLAG expression vector in combinationwith 300 ng of ORF59-FLAG expression vector in the absence orpresence of an increased amount of ORF57-FLAG expression vectoras indicated. Protein and RNA samples were prepared 24 h aftercotransfection and analyzed, respectively, by Western blot (WB) andNorthern blot (NB) assays. Tubulin served as a protein loading controlfor Western blotting, and GAPDH RNA served as an RNA loadingcontrol for Northern blotting. (A) RBM15 regulation of ORF59 ex-pression in the absence or presence of ORF57. Relative ORF59 pro-tein levels (fold) were quantified based on each band’s density afternormalization to that of tubulin as a sample loading control, with theORF59 protein level in lane 1 being set as 1. (B) Bar graphs showrelative level of ORF59 RNA in each sample from Northern blotanalysis in panel A after being normalized to the level of the corre-sponding GAPDH RNA for each sample loaded. Shown above the bargraph is the relative ratio of ORF59 RNA in the presence versus theabsence of RBM15 for each pair of samples.
1530 MAJERCIAK ET AL. J. VIROL.
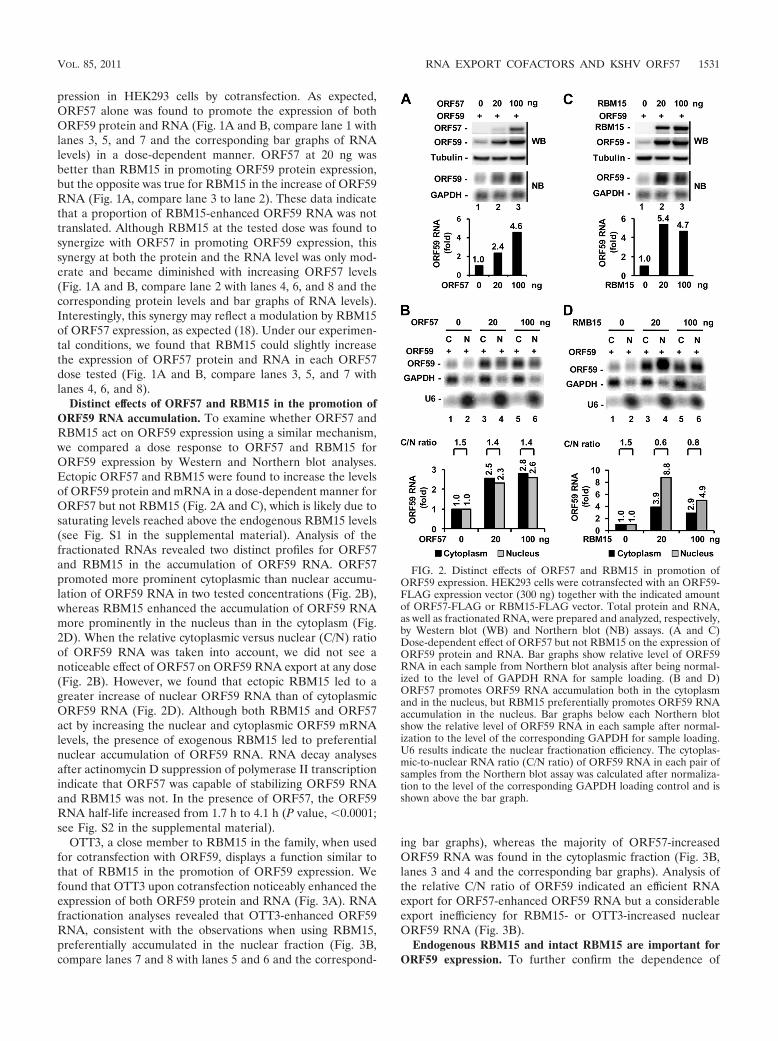
pression in HEK293 cells by cotransfection. As expected,ORF57 alone was found to promote the expression of bothORF59 protein and RNA (Fig. 1A and B, compare lane 1 withlanes 3, 5, and 7 and the corresponding bar graphs of RNAlevels) in a dose-dependent manner. ORF57 at 20 ng wasbetter than RBM15 in promoting ORF59 protein expression,but the opposite was true for RBM15 in the increase of ORF59RNA (Fig. 1A, compare lane 3 to lane 2). These data indicatethat a proportion of RBM15-enhanced ORF59 RNA was nottranslated. Although RBM15 at the tested dose was found tosynergize with ORF57 in promoting ORF59 expression, thissynergy at both the protein and the RNA level was only mod-erate and became diminished with increasing ORF57 levels(Fig. 1A and B, compare lane 2 with lanes 4, 6, and 8 and thecorresponding protein levels and bar graphs of RNA levels).Interestingly, this synergy may reflect a modulation by RBM15of ORF57 expression, as expected (18). Under our experimen-tal conditions, we found that RBM15 could slightly increasethe expression of ORF57 protein and RNA in each ORF57dose tested (Fig. 1A and B, compare lanes 3, 5, and 7 withlanes 4, 6, and 8).
Distinct effects of ORF57 and RBM15 in the promotion ofORF59 RNA accumulation. To examine whether ORF57 andRBM15 act on ORF59 expression using a similar mechanism,we compared a dose response to ORF57 and RBM15 forORF59 expression by Western and Northern blot analyses.Ectopic ORF57 and RBM15 were found to increase the levelsof ORF59 protein and mRNA in a dose-dependent manner forORF57 but not RBM15 (Fig. 2A and C), which is likely due tosaturating levels reached above the endogenous RBM15 levels(see Fig. S1 in the supplemental material). Analysis of thefractionated RNAs revealed two distinct profiles for ORF57and RBM15 in the accumulation of ORF59 RNA. ORF57promoted more prominent cytoplasmic than nuclear accumu-lation of ORF59 RNA in two tested concentrations (Fig. 2B),whereas RBM15 enhanced the accumulation of ORF59 RNAmore prominently in the nucleus than in the cytoplasm (Fig.2D). When the relative cytoplasmic versus nuclear (C/N) ratioof ORF59 RNA was taken into account, we did not see anoticeable effect of ORF57 on ORF59 RNA export at any dose(Fig. 2B). However, we found that ectopic RBM15 led to agreater increase of nuclear ORF59 RNA than of cytoplasmicORF59 RNA (Fig. 2D). Although both RBM15 and ORF57act by increasing the nuclear and cytoplasmic ORF59 mRNAlevels, the presence of exogenous RBM15 led to preferentialnuclear accumulation of ORF59 RNA. RNA decay analysesafter actinomycin D suppression of polymerase II transcriptionindicate that ORF57 was capable of stabilizing ORF59 RNAand RBM15 was not. In the presence of ORF57, the ORF59RNA half-life increased from 1.7 h to 4.1 h (P value, 0.0001;see Fig. S2 in the supplemental material).
OTT3, a close member to RBM15 in the family, when usedfor cotransfection with ORF59, displays a function similar tothat of RBM15 in the promotion of ORF59 expression. Wefound that OTT3 upon cotransfection noticeably enhanced theexpression of both ORF59 protein and RNA (Fig. 3A). RNAfractionation analyses revealed that OTT3-enhanced ORF59RNA, consistent with the observations when using RBM15,preferentially accumulated in the nuclear fraction (Fig. 3B,compare lanes 7 and 8 with lanes 5 and 6 and the correspond-
ing bar graphs), whereas the majority of ORF57-increasedORF59 RNA was found in the cytoplasmic fraction (Fig. 3B,lanes 3 and 4 and the corresponding bar graphs). Analysis ofthe relative C/N ratio of ORF59 indicated an efficient RNAexport for ORF57-enhanced ORF59 RNA but a considerableexport inefficiency for RBM15- or OTT3-increased nuclearORF59 RNA (Fig. 3B).
Endogenous RBM15 and intact RBM15 are important forORF59 expression. To further confirm the dependence of
FIG. 2. Distinct effects of ORF57 and RBM15 in promotion ofORF59 expression. HEK293 cells were cotransfected with an ORF59-FLAG expression vector (300 ng) together with the indicated amountof ORF57-FLAG or RBM15-FLAG vector. Total protein and RNA,as well as fractionated RNA, were prepared and analyzed, respectively,by Western blot (WB) and Northern blot (NB) assays. (A and C)Dose-dependent effect of ORF57 but not RBM15 on the expression ofORF59 protein and RNA. Bar graphs show relative level of ORF59RNA in each sample from Northern blot analysis after being normal-ized to the level of GAPDH RNA for sample loading. (B and D)ORF57 promotes ORF59 RNA accumulation both in the cytoplasmand in the nucleus, but RBM15 preferentially promotes ORF59 RNAaccumulation in the nucleus. Bar graphs below each Northern blotshow the relative level of ORF59 RNA in each sample after normal-ization to the level of the corresponding GAPDH for sample loading.U6 results indicate the nuclear fractionation efficiency. The cytoplas-mic-to-nuclear RNA ratio (C/N ratio) of ORF59 RNA in each pair ofsamples from the Northern blot assay was calculated after normaliza-tion to the level of the corresponding GAPDH loading control and isshown above the bar graph.
VOL. 85, 2011 RNA EXPORT COFACTORS AND KSHV ORF57 1531
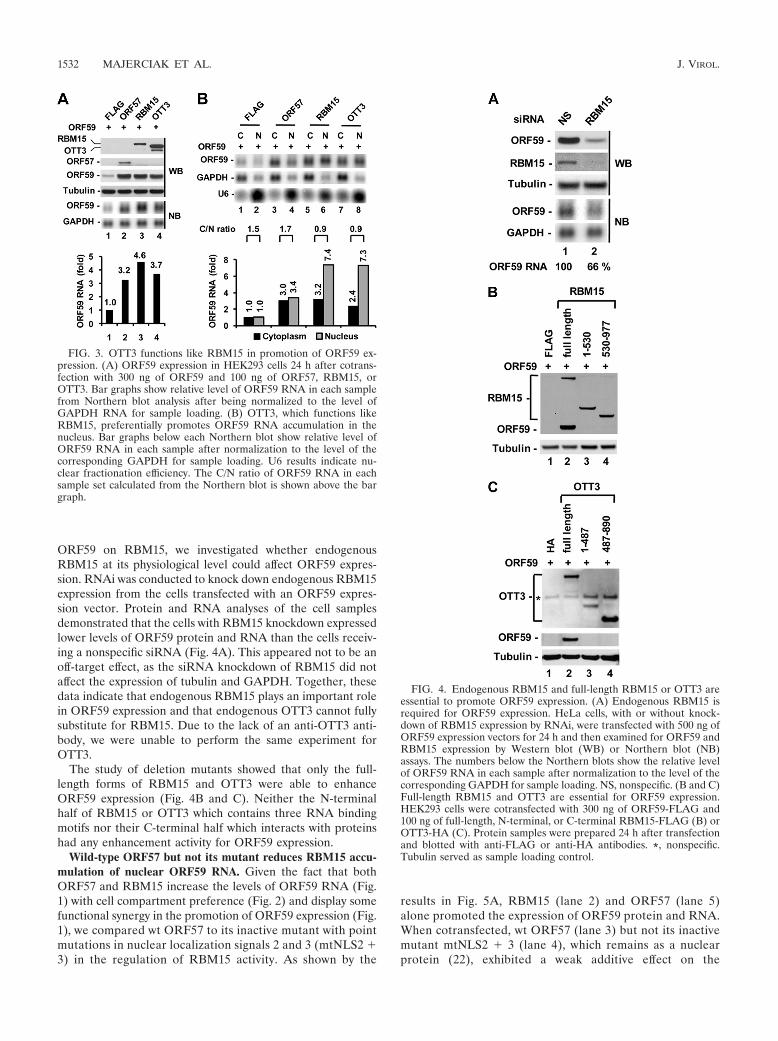
ORF59 on RBM15, we investigated whether endogenousRBM15 at its physiological level could affect ORF59 expres-sion. RNAi was conducted to knock down endogenous RBM15expression from the cells transfected with an ORF59 expres-sion vector. Protein and RNA analyses of the cell samplesdemonstrated that the cells with RBM15 knockdown expressedlower levels of ORF59 protein and RNA than the cells receiv-ing a nonspecific siRNA (Fig. 4A). This appeared not to be anoff-target effect, as the siRNA knockdown of RBM15 did notaffect the expression of tubulin and GAPDH. Together, thesedata indicate that endogenous RBM15 plays an important rolein ORF59 expression and that endogenous OTT3 cannot fullysubstitute for RBM15. Due to the lack of an anti-OTT3 anti-body, we were unable to perform the same experiment forOTT3.
The study of deletion mutants showed that only the full-length forms of RBM15 and OTT3 were able to enhanceORF59 expression (Fig. 4B and C). Neither the N-terminalhalf of RBM15 or OTT3 which contains three RNA bindingmotifs nor their C-terminal half which interacts with proteinshad any enhancement activity for ORF59 expression.
Wild-type ORF57 but not its mutant reduces RBM15 accu-mulation of nuclear ORF59 RNA. Given the fact that bothORF57 and RBM15 increase the levels of ORF59 RNA (Fig.1) with cell compartment preference (Fig. 2) and display somefunctional synergy in the promotion of ORF59 expression (Fig.1), we compared wt ORF57 to its inactive mutant with pointmutations in nuclear localization signals 2 and 3 (mtNLS2 �3) in the regulation of RBM15 activity. As shown by the
results in Fig. 5A, RBM15 (lane 2) and ORF57 (lane 5)alone promoted the expression of ORF59 protein and RNA.When cotransfected, wt ORF57 (lane 3) but not its inactivemutant mtNLS2 � 3 (lane 4), which remains as a nuclearprotein (22), exhibited a weak additive effect on the
FIG. 3. OTT3 functions like RBM15 in promotion of ORF59 ex-pression. (A) ORF59 expression in HEK293 cells 24 h after cotrans-fection with 300 ng of ORF59 and 100 ng of ORF57, RBM15, orOTT3. Bar graphs show relative level of ORF59 RNA in each samplefrom Northern blot analysis after being normalized to the level ofGAPDH RNA for sample loading. (B) OTT3, which functions likeRBM15, preferentially promotes ORF59 RNA accumulation in thenucleus. Bar graphs below each Northern blot show relative level ofORF59 RNA in each sample after normalization to the level of thecorresponding GAPDH for sample loading. U6 results indicate nu-clear fractionation efficiency. The C/N ratio of ORF59 RNA in eachsample set calculated from the Northern blot is shown above the bargraph.
FIG. 4. Endogenous RBM15 and full-length RBM15 or OTT3 areessential to promote ORF59 expression. (A) Endogenous RBM15 isrequired for ORF59 expression. HeLa cells, with or without knock-down of RBM15 expression by RNAi, were transfected with 500 ng ofORF59 expression vectors for 24 h and then examined for ORF59 andRBM15 expression by Western blot (WB) or Northern blot (NB)assays. The numbers below the Northern blots show the relative levelof ORF59 RNA in each sample after normalization to the level of thecorresponding GAPDH for sample loading. NS, nonspecific. (B and C)Full-length RBM15 and OTT3 are essential for ORF59 expression.HEK293 cells were cotransfected with 300 ng of ORF59-FLAG and100 ng of full-length, N-terminal, or C-terminal RBM15-FLAG (B) orOTT3-HA (C). Protein samples were prepared 24 h after transfectionand blotted with anti-FLAG or anti-HA antibodies. *, nonspecific.Tubulin served as sample loading control.
1532 MAJERCIAK ET AL. J. VIROL.

RBM15-enhanced expression of both ORF59 protein andRNA. The RNA fractionation analysis shown in Fig. 5Bexhibited a similar correlation of the cytoplasmic ORF59RNA level with the observed ORF59 protein and total RNAexpression shown in Fig. 5A in response to RBM15 and/orORF57. Despite the RBM15-mediated accumulation of nu-clear ORF59 RNA, as expected (Fig. 5B, lane 2), the cellscotransfected with RBM15 and ORF57 showed that a largeproportion of ORF59 RNA was accumulated in the cyto-plasm in the presence of wt ORF57 but not in the presenceof its inactive mutant mtNLS2 � 3 (Fig. 5B, compare lane 3to lane 4 and the corresponding bar graphs), which does notinteract with RBM15 and OTT3 (see Fig. 8 below). Analysis ofthe relative ORF59 RNA C/N ratio indicates that ectopic RBM15specifically led to more nuclear ORF59 RNA (Fig. 5B), as shownin Fig. 2D, but the presence of ORF57 reversed the C/N ratioback to normal. These data clearly indicate that one of KSHVORF57’s functions is to reduce ectopic-RBM15-mediated nu-clear accumulation of ORF59 RNA.
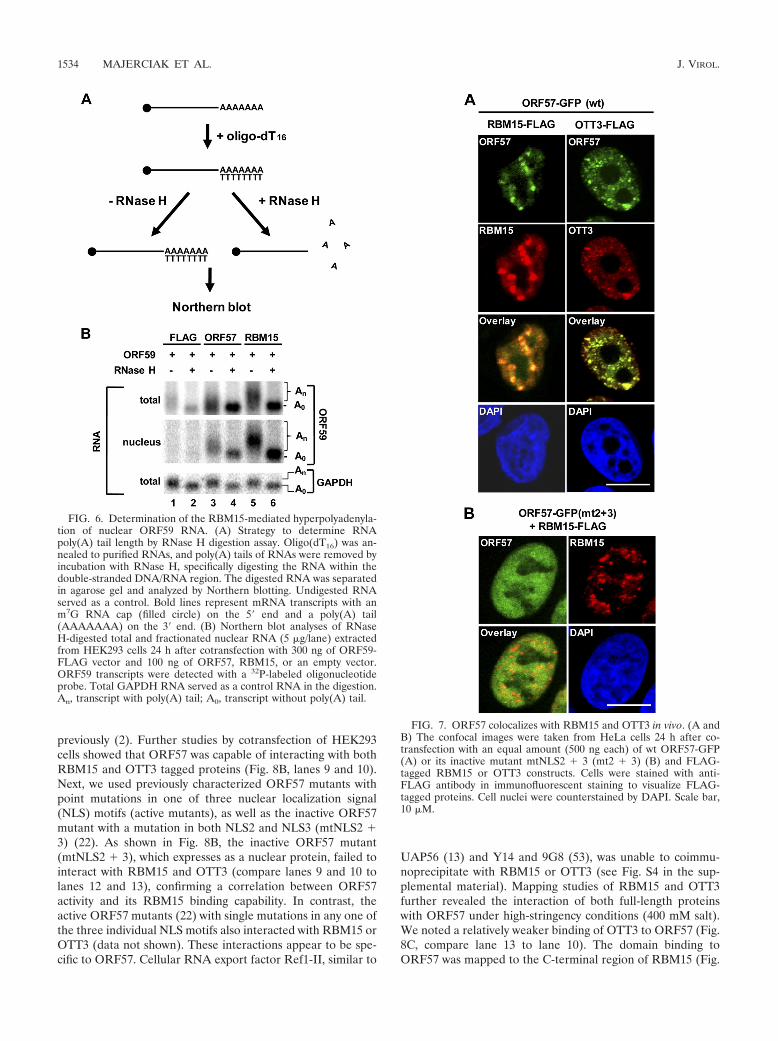
ORF57 prevents RBM15-induced accumulation of hyper-polyadenylated nuclear ORF59 RNA. Investigating how ec-
topic expression of RBM15 could lead to nuclear accumulationof ORF59 RNA, we noticed in our Northern blot analyses thatthe RBM15- or OTT3-induced nuclear ORF59 RNA migratedmore slowly than its counterpart in the cytoplasm in electro-phoresis (Fig. 3B, lanes 6 and 8, and Fig. 5B, lanes 2 and 4),indicating that the accumulated nuclear ORF59 RNA is of alarger size. Interestingly, ORF57 was found not only to reducethe RBM15-mediated nuclear accumulation of ORF59 RNAbut also to eliminate the heterogeneity of ORF59 RNAs,thereby leading them to migrate as its normal size in electro-phoresis (Fig. 5B, compare lane 3 to lane 5). We hypothesizedthat the longer nuclear ORF59 RNAs that are accumulated inthe presence of RBM15 might be hyperpolyadenylated RNAs.To determine whether hyperpolyadenylation was a cause of theincrease in length of nuclear ORF59 RNA, we extracted totalor fractionated ORF59 RNAs from HEK293 cells 24 h aftercotransfection with ORF59 and RBM15 or ORF57 and con-ducted RNase H digestion analyses after RNA hybridizationwith an oligo(dT16). As revealed by their slow migration inelectrophoresis, shown in Fig. 6, the RBM15-accumulated nu-clear ORF59 RNAs in the total RNA preparation (Fig. 6B,lane 5, top blot) and the nuclear fractionated RNA prepara-tion (Fig. 6B lane 5, middle blot) had a larger size thanORF57-enhanced ORF59 RNAs (compare lanes 5 with lanes3). However, all ORF59 RNAs analyzed by RNase H diges-tion, including total RNA and the fractionated nuclear ORF59RNA, migrated by electrophoresis at the same position afterthe RNA poly(A) tail removal mediated by oligo(dT16) andRNase H digestion, indicating that the more slowly migratingnuclear ORF59 RNA was indeed hyperpolyadenylated in thepresence of ectopic RBM15. RBM15’s hyperpolyadenylationeffect on ORF59 RNA in the nucleus also occurred with theinactive ORF57 mtNLS2 � 3, whose expression could be in-creased by ectopic RBM15 (see Fig. S3 in the supplementalmaterial), but this effect did not occur with GAPDH RNA(Fig. 6B, compare lane 5 with lane 3 for total ORF59 andGAPDH RNA).
ORF57 colocalizes and interacts with RBM15 and OTT3.Since ORF57, RBM15, and OTT3 are nuclear proteins (7, 22,49), we visualized their expression and localization in cotrans-fected HeLa cells. Consistent with other reports (6, 7, 13, 49),we show that the majority of RBM15 and OTT3 localized tothe nucleus. In support of the findings described above, wefound that ORF57 colocalized with RBM15 and OTT3 indistinct, speckle-like nuclear structures (Fig. 7A). The inactiveORF57 mutant (mtNLS2 � 3) lacking the ability to promoteORF59 expression (22) exhibited a diffuse distribution in thenucleus and did not display much colocalization with RBM15(Fig. 7B). Thus, the cellular colocalization results suggest thepresence of biochemical interaction of functional ORF57 withRBM15 and OTT3.
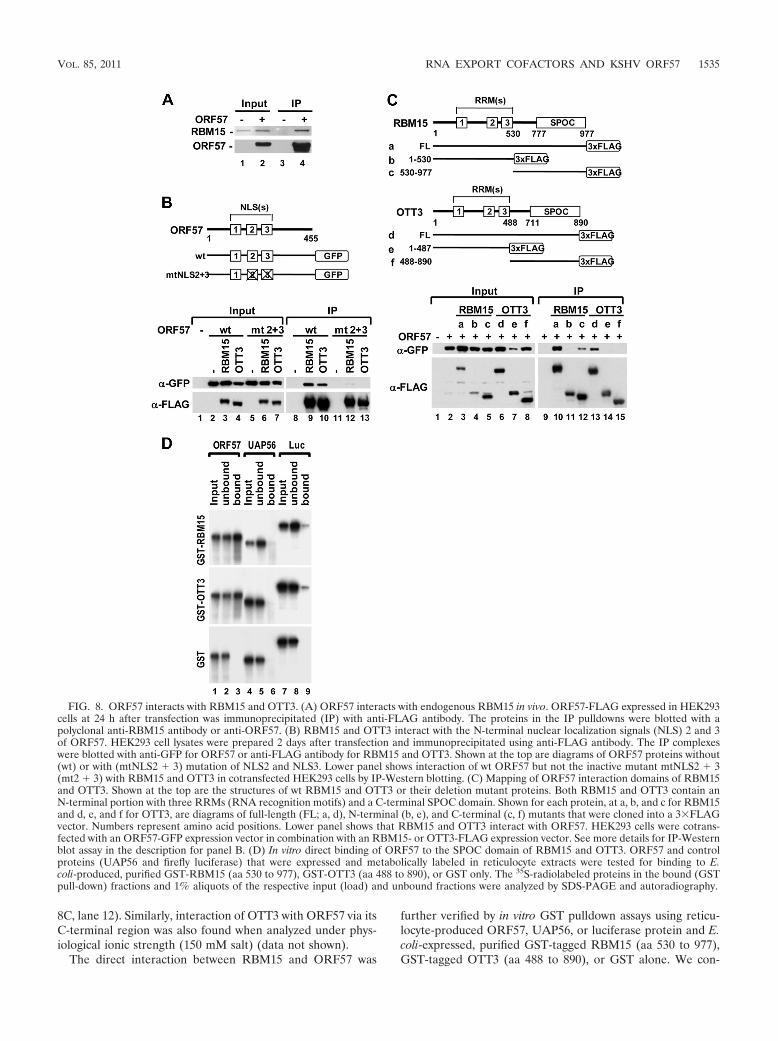
Next, we examined the possible interaction of ORF57 pro-tein with RBM15 and OTT3. We first immunoprecipitatedtagged ORF57 protein from extracts of transfected HEK293cells. The extracts were digested with RNase A prior to IP toexclude the possibility of RNA-mediated protein-protein in-teraction in the IP pull-down assays. We found that, inHEK293 cells under physiological conditions (150 mM salt),ORF57 was associated with endogenous RBM15 (Fig. 8A) butnot with endogenous UAP56 (data not shown), as reported
FIG. 5. ORF57 offsets the nuclear RNA accumulation activity ofRBM15 in promoting ORF59 expression. (A) ORF57 coordinates withRBM15 to additively promote ORF59 expression. (B) ORF57 pro-motes ORF59 RNA accumulation in the cytoplasm by offsetting thenuclear RNA accumulation mediated by ectopic RBM15. HEK293cells were cotransfected with 300 ng of ORF59-FLAG in combinationwith or without 20 ng of RBM15-FLAG in the presence or absence ofthe indicated ORF57-GFP expression vector (100 ng). mtNLS2 � 3(mt2 � 3), an inactive ORF57 mutant with point mutations in itsnuclear localization signals 2 and 3, served as a control. A GFP-onlyplasmid was used as a vector control. Protein samples, total RNA, andfractionated RNA were prepared 24 h after cotransfection. Proteinsamples were examined by Western blot (WB) assay with anti-FLAGand anti-GFP antibodies (A, upper panel). Tubulin served as sampleloading control. All RNA samples were analyzed by Northern blot(NB) assay (A, lower panel; B, upper panel). Bar graphs below eachNorthern blot in panels A and B show the relative level of ORF59 RNAin each sample after normalization to the level of the correspondingGAPDH for sample loading. U6 results show nuclear fractionation effi-ciency. Relative C/N ratio of ORF59 RNA in each sample set calculatedfrom the Northern blot in panel B is shown above the bar graph.
VOL. 85, 2011 RNA EXPORT COFACTORS AND KSHV ORF57 1533
previously (2). Further studies by cotransfection of HEK293cells showed that ORF57 was capable of interacting with bothRBM15 and OTT3 tagged proteins (Fig. 8B, lanes 9 and 10).Next, we used previously characterized ORF57 mutants withpoint mutations in one of three nuclear localization signal(NLS) motifs (active mutants), as well as the inactive ORF57mutant with a mutation in both NLS2 and NLS3 (mtNLS2 �3) (22). As shown in Fig. 8B, the inactive ORF57 mutant(mtNLS2 � 3), which expresses as a nuclear protein, failed tointeract with RBM15 and OTT3 (compare lanes 9 and 10 tolanes 12 and 13), confirming a correlation between ORF57activity and its RBM15 binding capability. In contrast, theactive ORF57 mutants (22) with single mutations in any one ofthe three individual NLS motifs also interacted with RBM15 orOTT3 (data not shown). These interactions appear to be spe-cific to ORF57. Cellular RNA export factor Ref1-II, similar to
UAP56 (13) and Y14 and 9G8 (53), was unable to coimmu-noprecipitate with RBM15 or OTT3 (see Fig. S4 in the sup-plemental material). Mapping studies of RBM15 and OTT3further revealed the interaction of both full-length proteinswith ORF57 under high-stringency conditions (400 mM salt).We noted a relatively weaker binding of OTT3 to ORF57 (Fig.8C, compare lane 13 to lane 10). The domain binding toORF57 was mapped to the C-terminal region of RBM15 (Fig.
FIG. 6. Determination of the RBM15-mediated hyperpolyadenyla-tion of nuclear ORF59 RNA. (A) Strategy to determine RNApoly(A) tail length by RNase H digestion assay. Oligo(dT16) was an-nealed to purified RNAs, and poly(A) tails of RNAs were removed byincubation with RNase H, specifically digesting the RNA within thedouble-stranded DNA/RNA region. The digested RNA was separatedin agarose gel and analyzed by Northern blotting. Undigested RNAserved as a control. Bold lines represent mRNA transcripts with anm7G RNA cap (filled circle) on the 5� end and a poly(A) tail(AAAAAAA) on the 3� end. (B) Northern blot analyses of RNaseH-digested total and fractionated nuclear RNA (5 �g/lane) extractedfrom HEK293 cells 24 h after cotransfection with 300 ng of ORF59-FLAG vector and 100 ng of ORF57, RBM15, or an empty vector.ORF59 transcripts were detected with a 32P-labeled oligonucleotideprobe. Total GAPDH RNA served as a control RNA in the digestion.An, transcript with poly(A) tail; A0, transcript without poly(A) tail.
FIG. 7. ORF57 colocalizes with RBM15 and OTT3 in vivo. (A andB) The confocal images were taken from HeLa cells 24 h after co-transfection with an equal amount (500 ng each) of wt ORF57-GFP(A) or its inactive mutant mtNLS2 � 3 (mt2 � 3) (B) and FLAG-tagged RBM15 or OTT3 constructs. Cells were stained with anti-FLAG antibody in immunofluorescent staining to visualize FLAG-tagged proteins. Cell nuclei were counterstained by DAPI. Scale bar,10 �M.
1534 MAJERCIAK ET AL. J. VIROL.
8C, lane 12). Similarly, interaction of OTT3 with ORF57 via itsC-terminal region was also found when analyzed under phys-iological ionic strength (150 mM salt) (data not shown).
The direct interaction between RBM15 and ORF57 was
further verified by in vitro GST pulldown assays using reticu-locyte-produced ORF57, UAP56, or luciferase protein and E.coli-expressed, purified GST-tagged RBM15 (aa 530 to 977),GST-tagged OTT3 (aa 488 to 890), or GST alone. We con-
FIG. 8. ORF57 interacts with RBM15 and OTT3. (A) ORF57 interacts with endogenous RBM15 in vivo. ORF57-FLAG expressed in HEK293cells at 24 h after transfection was immunoprecipitated (IP) with anti-FLAG antibody. The proteins in the IP pulldowns were blotted with apolyclonal anti-RBM15 antibody or anti-ORF57. (B) RBM15 and OTT3 interact with the N-terminal nuclear localization signals (NLS) 2 and 3of ORF57. HEK293 cell lysates were prepared 2 days after transfection and immunoprecipitated using anti-FLAG antibody. The IP complexeswere blotted with anti-GFP for ORF57 or anti-FLAG antibody for RBM15 and OTT3. Shown at the top are diagrams of ORF57 proteins without(wt) or with (mtNLS2 � 3) mutation of NLS2 and NLS3. Lower panel shows interaction of wt ORF57 but not the inactive mutant mtNLS2 � 3(mt2 � 3) with RBM15 and OTT3 in cotransfected HEK293 cells by IP-Western blotting. (C) Mapping of ORF57 interaction domains of RBM15and OTT3. Shown at the top are the structures of wt RBM15 and OTT3 or their deletion mutant proteins. Both RBM15 and OTT3 contain anN-terminal portion with three RRMs (RNA recognition motifs) and a C-terminal SPOC domain. Shown for each protein, at a, b, and c for RBM15and d, e, and f for OTT3, are diagrams of full-length (FL; a, d), N-terminal (b, e), and C-terminal (c, f) mutants that were cloned into a 3�FLAGvector. Numbers represent amino acid positions. Lower panel shows that RBM15 and OTT3 interact with ORF57. HEK293 cells were cotrans-fected with an ORF57-GFP expression vector in combination with an RBM15- or OTT3-FLAG expression vector. See more details for IP-Westernblot assay in the description for panel B. (D) In vitro direct binding of ORF57 to the SPOC domain of RBM15 and OTT3. ORF57 and controlproteins (UAP56 and firefly luciferase) that were expressed and metabolically labeled in reticulocyte extracts were tested for binding to E.coli-produced, purified GST-RBM15 (aa 530 to 977), GST-OTT3 (aa 488 to 890), or GST only. The 35S-radiolabeled proteins in the bound (GSTpull-down) fractions and 1% aliquots of the respective input (load) and unbound fractions were analyzed by SDS-PAGE and autoradiography.
VOL. 85, 2011 RNA EXPORT COFACTORS AND KSHV ORF57 1535

firmed that ORF57, like NXF1 (49), interacted specificallywith RBM15 and OTT3, whereas UAP56 or luciferase proteindid not (Fig. 8D). Collectively, these data indicate that ORF57acts by promoting ORF59 mRNA expression through directprotein-protein interactions with RBM15 and OTT3.
The ORF57-RBM15 interaction interferes with RBM15RNA binding. To determine whether the ORF57 colocaliza-tion and interaction with RBM15 would affect RBM15 bindingto ORF59 RNA and thereby prevent ORF59 RNA from hy-perpolyadenylation in the nucleus, we took two different ap-proaches. First, we examined whether ORF57 could affectRBM15’s association with polyadenylated mRNAs in HEK293cells. As shown in Fig. 9A, FLAG-tagged RBM15 could be UVcross-linked to polyadenylated mRNAs in the cells (comparelane 1 to lane 2), but the RBM15 cross-linking to the poly-adenylated mRNAs appeared reduced in the presence of wtORF57 over the presence of mutant ORF57 (compare lane 3to lane 4). When a truncated RBM15 (aa 1 to 530), lacking aC-terminal SPOC domain and deficient in ORF57 interaction(Fig. 8C) but bearing all three RRMs, was used in the assay,this mutant RBM15 had the same RNA binding activitywhether in the presence of wt or mutant ORF57 (Fig. 9B,compare lane 3 to lane 4). These data indicate that the ORF57reduction of RBM15-mRNA interaction requires ORF57-RBM15 interaction. This reduction appeared to be unrelatedto ORF59 protein. We found that only RBM15, and notORF59, could be pulled down along with polyadenylatedmRNAs (Fig. 9C). The associated RBM15 level on polyade-nylated mRNA was reduced in the presence of wt ORF57compared to the level in the presence of the mutant ORF57that was incapable of interacting with RBM15 (Fig. 9C, com-pare lane 5 to lane 6).
Next, we performed CLIP assays to examine the specificeffect of ORF57 on RBM15-ORF59 RNA interactions.HEK293 cells were cotransfected with FLAG-tagged RBM15and FLAG-tagged ORF59 in the absence or presence of GFP-tagged wt or mutant ORF57 and UV irradiated. The protein-RNA complexes from the cells were coimmunoprecipitated byanti-FLAG antibody and digested with proteinase K. TheRNAs in the digested complex were extracted and amplified byRT-PCR for ORF59 RNA as described previously (22). Usingthis strategy, we first confirmed that RBM15 binds ORF59RNA in the cells in the absence of ORF57 (Fig. 9D, lane 2).Further analysis demonstrated that RBM15 bound ORF59RNA in the presence of mutant ORF57, but this protein-RNAinteraction was reduced in the presence of wt ORF57 (Fig. 9E,right panel, compare lane 3 to lane 1). Quantitative analysis ofthe ORF59 RNA levels from these two samples by qRT-PCRindicated a significant (P value, 0.024) reduction of ORF59
FIG. 9. ORF57 affects RBM15-RNA interactions in vivo. (A andB) ORF57-RBM15 SPOC domain interaction interferes withRBM15’s association with polyadenylated RNA. HEK293 cells (5 �105/well in a 6-well plate) were cotransfected with the following ex-pression vectors (500 ng each): FLAG-tagged ORF59, FLAG-taggedfull-length RBM15 (A) or SPOC domain deletion mutant (aa 1 to 530)(B) and GFP-tagged wild-type ORF57 (wt) or NLS2 � 3 mutant (mt).Empty vector pEGFP-N1 served as an ORF57 negative control. At24 h after transfection, the cells were UV irradiated, and the poly-adenylated [poly(A)] RNA transcripts were isolated. After RNasedigestion, the proteins associated with poly(A) RNAs were detected byWestern blotting by using anti-FLAG antibody for RBM15. Total celllysates prior to poly(A) RNA selection served as input for each samplein Western blotting. (C to F) ORF57 interaction with RBM15 affectsthe binding of RBM15 to ORF59 RNA. HEK293 cells 24 h aftercotransfection with RBM15 and ORF59 in the absence or presence ofwt or mutant ORF57 were examined by poly(A) selection (C) or CLIPassays (D to F) followed by Western blot (C) or RT-PCR (D to F)analysis. (C) RBM15 protein but not ORF59 associates withpoly(A) RNAs, which is preventable by wt ORF57. Lanes 3 and 4contain cell lysate input prior to poly(A) mRNA selection, showingectopic expression of FLAG-tagged RBM15 and ORF59. (D) RBM15protein binds ORF59 RNA in the absence of ORF57. The RBM15-ORF59 RNA complex in the CLIP assay, after proteinase K digestion,was analyzed by RT-PCR analysis for ORF59 RNA or GAPDH RNAas a nonspecific RNA control. Preimmune rabbit serum IgG served asan IP control. RT, reverse transcriptase; M, 100-bp DNA ladder;DNA, ORF59 plasmid DNA for size control. (E and F) Coexpression
of wt ORF57 with RBM15 reduces RBM15 binding to ORF59 RNA.See other details in the panel D legend. (E, left) Results show the sameamount of ORF59 RNA from the input cell extracts as used for CLIP.GAPDH RNA in the extracts served as a loading control. (E, right)Results show that wt ORF57 but not ORF57 mtNLS2 � 3 preventsRBM15 association with ORF59 RNA in CLIP assays. (F) Bar graphswith means standard deviations show the ORF59 RNA level fromeach anti-FLAG CLIP, quantified by qRT-PCR and analyzed by two-tailed Student’s t test. n 2, each in triplicate. Ab, antibody.
1536 MAJERCIAK ET AL. J. VIROL.

RNA on RBM15 protein in the presence of wt ORF57 (Fig.9F). Collectively, these data provide compelling evidence thatwt ORF57’s interaction with RBM15 affects the association ofRBM15 with ORF59 RNA.
The ORF57 homologs EB2, ICP27, UL69, and IE4 also in-teract with RBM15 and OTT3. As EBV EB2 was reported tointeract with OTT3 and RBM15 (6), we examined whether theinteraction of KSHV ORF57 with RBM15 is a conserved func-tion among other homologs in the herpesvirus family. Wetransfected HEK293 cells with RBM15 or OTT3 expressionvectors in combination with EBV EB2 (6), HSV-1 ICP27 (38),VZV IE4 (32), or CMV UL69 (47) and analyzed the total cellextracts by co-IP. We found that EB2 (Fig. 10A, lanes 5 and 6),ICP27 (Fig. 10B, lanes 5 and 6), IE4 (Fig. 10C, lanes 6 and 8),and UL69 (Fig. 10D, lanes 5 and 6), consistent with the resultsfor ORF57 (Fig. 8C, lanes 10 and 13), all interact with RBM15and OTT3. As described for ORF57 (Fig. 8C) and consistentwith the report by Hiriart et al. (6), we found that EB2 inter-acts with the C-terminal portion of RBM15 (data not shown).These findings indicate that the interaction of RBM15 orOTT3 with ORF57 is indeed a highly conserved functionamong ORF57 homologs in the herpesvirus family.
EBV EB2 does not function like KSHV ORF57 to preventRBM15-mediated nuclear accumulation of ORF59. As EBVEB2 is the closest homologue to KSHV ORF57, we examinedwhether EB2 functions mechanistically like ORF57. We firstdetermined EB2 colocalization with RBM15 by transienttransfection of HeLa cells (Fig. 11A) and then compared EB2with ORF57 and RBM15 in promoting ORF59 expression. Wefound that EB2 protein in HEK293 cells exhibited a function
similar to that of ORF57 and RBM15 in promoting ORF59expression at both the protein and RNA level (Fig. 11B), asdescribed previously (22), despite the fact that EB2 does notcomplement ORF57 function in an ORF57-null KSHV ge-nome (4), nor does it complement ectopic RBM15 inBac36�57 (our unpublished data). Further analyses of frac-tionated nuclear versus cytoplasmic RNA revealed that EB2,unlike ORF57, was unable to efficiently prevent RBM15-me-diated accumulation of nuclear ORF59 RNA by cotransfection(Fig. 11C, compare lane 4 to lanes 2 and 3). Furthermore, EB2exhibited no effect on RBM15’s association with polyadeny-lated RNAs (Fig. 11D). Together, these data indicate thatEB2, which interacts with RBM15 (Fig. 10A), may functiondifferently from ORF57 in the promotion of ORF59 expres-sion.
DISCUSSION
RNA transport from the nucleus to the cytoplasm is anessential step in the expression of mammalian and viral genesand requires several nuclear export proteins, including UAP56,URH49, Aly/REF, SRp20, 9G8, SF2/ASF, RBM15, OTT3,and the RNA export receptor NXF1/TAP (5, 8, 9, 11, 34, 46).These RNA binding and export factors, which function asRNA export adaptors, have been identified in two nuclearprotein complexes, including the spliceosome and TREX com-plexes, to interact with the NXF1/p15 heterodimer, a majorRNA exporter. Together, these complexes deliver the bound,export-ready RNA to the nuclear pore complex and, therefore,are responsible for the tightly coupled events of RNA tran-scription, splicing, and export in eukaryotic gene expression(15, 25, 45, 51). In this study, KSHV ORF57 was found tointeract with the cellular RNA export cofactors RBM15 andOTT3. This protein-protein interaction affects the RNA bind-ing activity of RBM15 in the nucleus and prevents RBM15-mediated nuclear accumulation of hyperpolyadenylated, in-tronless viral ORF59 RNA.
We have found that RBM15 and OTT3 themselves canfunction similarly to KSHV ORF57 to promote the expressionof KSHV ORF59. The observed ability of RBM15 to promotethe accumulation of ORF59 RNA in both cytoplasmic andnuclear fractions in the absence of ORF57 was unexpectedsince RBM15, and the related OTT3, were reported to partic-ipate in RNA export (6, 13, 49, 53). In contrast to ORF57,which promotes ORF59 expression in a dose-dependent man-ner, the maximum effect of ectopic RBM15 on ORF59 expres-sion was found at a tested low dose (20 ng) that producedabout 70% more RBM15 than the endogenous RBM15 level inHEK293 cells. Because RBM15 at the tested low dose alsoshowed a moderate effect on ORF57 expression, the synergis-tic effect of ORF57 with RBM15 on ORF59 expression ob-served in this study could be the result of RBM15’s effect onORF57 expression, consistent with our previous observationthat ORF57 expression depends on endogenous RBM15 andOTT3 (18). However, the function of ORF57 in ORF59 RNAexpression differs from that of RBM15. The former accumu-lates more ORF59 RNA in the cytoplasm, but the latter accu-mulates the RNA more in the nucleus than in the cytoplasm.Therefore, ORF57 has a greater effect than RBM15 on the net
FIG. 10. The interaction with RBM15 and OTT3 proteins is con-served among ORF57 homologs. (A to C) Myc-tagged EBV EB2 (A),GFP-tagged HSV-1 ICP27 (B), and V5-tagged VZV IE4 (C) wereexpressed in HEK293 cells alone or together with FLAG-taggedRBM15 or OTT3, and the complexes were coimmunoprecipitated withanti-FLAG antibody under stringent salt conditions (400 mM). The IPcomplexes were blotted with anti-myc for EB2 (A), with anti-GFP forICP27 (B), with anti-V5 to detect IE4 (C), and with anti-FLAG forRBM15 and OTT3 (A, B, and C, lower panels). (D) To determine theinteraction of CMV UL69 with RBM15 and OTT3 in vivo, HEK293cells were cotransfected with FLAG-tagged UL69 and HA-taggedRBM15 or HA-tagged OTT3. The protein complexes were coimmu-noprecipitated with anti-FLAG antibody (UL69) under stringent saltconditions (400 mM) and blotted with anti-HA to detect RBM15 andOTT3 and with anti-FLAG to detect UL69.
VOL. 85, 2011 RNA EXPORT COFACTORS AND KSHV ORF57 1537

outcome of ORF59 protein production when cotransfected at20 ng.
As the increase of ORF59 RNA expression by RBM15 is atthe posttranscriptional level and RBM15 was unable to stabi-lize ORF59 RNA in this study, the cytoplasmic accumulationof ORF59 RNA by RBM15 could result from RBM15’s pro-motion of RNA export (13, 18, 49). However, when ectopicallyexpressed beyond its physiological level in the absence ofORF57, RBM15 adversely affects the RNA export machinery,leading to nuclear accumulation of ORF59 RNA, while a smallproportion of the RNA remains exportable (Fig. 12). Theeffect of overexpressed RBM15 on ORF59 RNA hyperpoly-adenylation also occurred with the ORF57 mtNLS2 � 3, whichis a functionally inactive nuclear protein and lacks interactionwith RBM15 and OTT3 (Fig. 8B). Other studies show thatexcess UAP56, another RNA export factor, inhibits the re-cruitment of Aly/REF to the spliced mRNP (15) and is toxic inC. elegans (17) and yeast (44). The mechanism by whichRBM15 promotes the nuclear accumulation of ORF59 is now
partially understood due to the findings in this report. Weobserved that the nuclear ORF59 RNA in the presence ofectopic RBM15 migrated slightly more slowly in electrophore-sis due to its hyperpolyadenylation. Although it remains to beinvestigated whether this modification of ORF59 RNA is medi-ated directly by RBM15 or is the consequence of RBM15-medi-ated nuclear accumulation of ORF59 RNA, it is discernible thatORF57 can prevent ORF59 RNA from hyperpolyadenylation inthe presence of ectopic RBM15 and reduce the RNA accumula-tion in the nucleus (Fig. 5B). Given the facts that RBM15 andOTT3 are RNA binding proteins (13, 49) and that the endog-enous level of RBM15 promotes only limited expression ofORF59 protein in the absence of ORF57, our data indicatethat RBM15 and OTT3 act as molecular links for ORF57,consistent with our previous reports of ORF57 binding specif-ically to its target RNAs only in the presence of cellular factors(22, 23) (Fig. 12). The inherent advantage of studying theKSHV ORF59 mRNA regulation rather than the previouslydescribed RTE-containing transcript (13, 49) lies in the fact
FIG. 11. EBV EB2 does not prevent RBM15-mediated nuclear accumulation of ORF59 RNA and does not block RBM15-RNA interactions.(A) Colocalization of EB2-myc and RBM15-FLAG in HeLa cells by cotransfection. Scale bar, 10 �m. (B and C) EB2 promotes ORF59 expressionbut does not prevent RBM15-mediated nuclear accumulation of ORF59 RNA. HEK293 cells (2.5 � 105 cells/ml) were transfected with 300 ng ofORF59-FLAG with or without 20 ng RBM15-FLAG in addition to 100 ng empty vector, ORF57-FLAG, or EB2-myc. Twenty-four hours aftertransfection, the cells were collected for total protein and RNA analyses by Western blot (WB) and Northern blot (NB) assays, respectively (B).Fractionated cytoplasmic and nuclear RNA was also analyzed in Northern blot assays by using a 32P-labeled oligonucleotide probe specific forORF59 RNA (C). (D) EB2 does not prevent RBM15’s association with polyadenylated RNAs. HEK293 cells (5 � 105) were transfected with 500ng each of ORF59-FLAG and RBM15-FLAG vectors together with 500 ng of an empty FLAG vector, ORF57-FLAG, or EB2-myc. Twenty-fourhours after transfection, the cells were exposed to UV light and polyadenylated RNAs were isolated. RBM15 proteins bound onto polyadenylatedRNAs were analyzed by Western blotting. SE, short exposure; LE, long exposure.
1538 MAJERCIAK ET AL. J. VIROL.

that the ORF59 mRNA requires both cellular (RBM15 andOTT3) and viral (ORF57) factors, whereas RTE requires onlycellular factors (RBM15, OTT3, and NXF1), in order to effi-ciently export from the nucleus. The ability to dissect the spe-cific contributions of both the viral and cellular factors inORF59 regulation contributes to our understanding of RNAexport.
ORF57 protein interacts with RBM15 and OTT3. The N-terminal NLS2 and NLS3 regions of ORF57 (22) are impor-tant for the high-affinity interaction with full-length RBM15and OTT3. ORF57 interacts with the C-terminal regions ofRBM15 and OTT3, which were previously shown to mediateinteraction with the nuclear RNA export receptor NXF1 (13,49). Interestingly, in addition to the interaction of the RBM15or OTT3 protein with KSHV ORF57 in this study and thepreviously reported interaction with EBV EB2 (6), we foundthat posttranscriptional regulators of the other herpesvirusfamily members, such as the HSV-1 ICP27, the VZV IE4, andthe CMV UL69, interact with the RBM15 or OTT3 proteins.The conservation of these interactions further supports theirbiological relevance. In vivo, we found ORF57 to colocalizewith RBM15 and OTT3, and several reports have shown thatthese factors, when exogenously expressed, also colocalize withSC35 (1, 21, 49). Our previous studies demonstrated thatORF57 mainly associates with cellular protein-RNA com-plexes (23). Recently, we showed that ORF57-RBM15 inter-
action is required for ORF57 expression and functions inKSHV-infected cells (18). However, RBM15 is incapable ofcomplementing ORF57’s function in stable Bac36�57 cellscontaining an ORF57-null KSHV genome (our unpublisheddata). In this report, the biological relevance of the ORF57interaction with RBM15 or OTT3 was further supported by theobservation that ORF57-RBM15 interactions interfered withRBM15 binding to polyadenylated mRNAs and ORF59 RNAand, therefore, prevented the RBM15-/OTT3-mediated nu-clear accumulation of ORF59 RNA. Interestingly, we foundthat EBV EB2, although it interacts with RBM15 and OTT3,does not function like KSHV ORF57 to prevent RBM15-RNAinteraction and, thereby, RBM15-mediated nuclear accumula-tion of ORF59. Despite the fact that ORF57 interacts withRNA export factors Aly/REF (2, 22, 24) and CBP80 (2) andother cellular factors (23), the ORF57-Aly/REF interactionwas found not to be important for ORF57-mediated ORF59expression (22, 29). Thus, the finding that ORF57 interactswith RNA export cofactors RBM15 and OTT3 and offsets theRBM15- and OTT3-mediated nuclear accumulation of ORF59RNA provides further insight into how ORF57 promotes theexpression of intronless viral genes.
ACKNOWLEDGMENTS
We thank R. Sandri-Goldin, S. Swaminathan, C. Sadzot, and T.Stamminger for plasmids and A. S. Zolotukhin for discussions. We alsothank David Liewehr and Seth Sternberg for statistical analyses.
This study was supported by the Intramural Research Program ofNIH, National Cancer Institute, Center for Cancer Research.
We declare that we have no conflicts of interest.
REFERENCES
1. Bello, L. J., et al. 1999. The human herpesvirus-8 ORF 57 gene and itsproperties. J. Gen. Virol. 80:3207–3215.
2. Boyne, J. R., K. J. Colgan, and A. Whitehouse. 2008. Recruitment of thecomplete hTREX complex is required for Kaposi’s sarcoma-associated her-pesvirus intronless mRNA nuclear export and virus replication. PLoSPathog. 4:e1000194.
3. Doroquez, D. B., T. L. Orr-Weaver, and I. Rebay. 2007. Split ends antago-nizes the Notch and potentiates the EGFR signaling pathways during Dro-sophila eye development. Mech. Dev. 124:792–806.
4. Han, Z., and S. Swaminathan. 2006. Kaposi’s sarcoma-associated herpesvi-rus lytic gene ORF57 is essential for infectious virion production. J. Virol.80:5251–5260.
5. Hautbergue, G. M., M. L. Hung, A. P. Golovanov, L. Y. Lian, and S. A.Wilson. 2008. Mutually exclusive interactions drive handover of mRNA fromexport adaptors to TAP. Proc. Natl. Acad. Sci. U. S. A. 105:5154–5159.
6. Hiriart, E., et al. 2005. Interaction of the Epstein-Barr virus mRNA exportfactor EB2 with human Spen proteins SHARP, OTT1, and a novel memberof the family, OTT3, links Spen proteins with splicing regulation and mRNAexport. J. Biol. Chem. 280:36935–36945.
7. Juillard, F., et al. 2009. Epstein-Barr virus protein EB2 contains an N-terminal transferable nuclear export signal that promotes nucleocytoplasmicexport by directly binding TAP/NXF1. J. Virol. 83:12759–12768.
8. Kapadia, F., A. Pryor, T. H. Chang, and L. F. Johnson. 2006. Nuclearlocalization of poly(A)� mRNA following siRNA reduction of expression ofthe mammalian RNA helicases UAP56 and URH49. Gene 384:37–44.
9. Kota, K. P., S. R. Wagner, E. Huerta, J. M. Underwood, and J. A. Nickerson.2008. Binding of ATP to UAP56 is necessary for mRNA export. J. Cell Sci.121:1526–1537.
10. Kuroda, K., et al. 2003. Regulation of marginal zone B cell development byMINT, a suppressor of Notch/RBP-J signaling pathway. Immunity 18:301–312.
11. Lai, M. C., Y. H. Lee, and W. Y. Tarn. 2008. The DEAD-box RNA helicaseDDX3 associates with export messenger ribonucleoproteins as well as tip-associated protein and participates in translational control. Mol. Biol. Cell19:3847–3858.
12. Lin, H. V., et al. 2003. Splits ends is a tissue/promoter specific regulator ofWingless signaling. Development 130:3125–3135.
13. Lindtner, S., et al. 2006. RNA-binding motif protein 15 binds to the RNAtransport element RTE and provides a direct link to the NXF1 exportpathway. J. Biol. Chem. 281:36915–36928.
FIG. 12. Proposed model for the effects of RBM15 and ORF57 onORF59 expression. (A) RBM15 at physiological level promotes theexpression of ORF59 at the early stage of KSHV lytic replication whenORF57 is at low concentration. Although ORF57 promotes ORF59expression, ORF57 expression is also dependent on RBM15 (18). At alater stage of infection when ORF57 is high, its interaction withRBM15 prevents RBM15’s association with ORF59 RNA and, conse-quently, reduces the function of RBM15 to promote ORF59 RNAexport. ORF57 accumulates ORF59 RNA by increasing ORF59 RNAstability by an unknown mechanism. (B) Overexpression of RBM15preferentially increases the nuclear over the cytoplasmic level ofORF59 RNA and stimulates hyperpolyadenylation of nuclear ORF59RNA. In the presence of ORF57, ORF57 interacts with RBM15 andprevents the RBM15-mediated nuclear accumulation of ORF59 RNA.
VOL. 85, 2011 RNA EXPORT COFACTORS AND KSHV ORF57 1539

14. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method.Methods 25:402–408.
15. Luo, M. L., et al. 2001. Pre-mRNA splicing and mRNA export linked bydirect interactions between UAP56 and Aly. Nature 413:644–647.
16. Ma, Z., et al. 2001. Fusion of two novel genes, RBM15 and MKL1, in thet(1;22)(p13;q13) of acute megakaryoblastic leukemia. Nat. Genet. 28:220–221.
17. MacMorris, M., C. Brocker, and T. Blumenthal. 2003. UAP56 levels affectviability and mRNA export in Caenorhabditis elegans. RNA 9:847–857.
18. Majerciak, V., M. Deng, and Z. M. Zheng. 2010. Requirement of UAP56,URH49, RBM15, and OTT3 in the expression of Kaposi sarcoma-associatedherpesvirus ORF57. Virology 407:206–212.
19. Majerciak, V., M. Kruhlak, P. K. Dagur, J. P. McCoy, Jr., and Z. M. Zheng.2010. Caspase-7 cleavage of Kaposi sarcoma-associated herpesvirus ORF57confers a cellular function against viral lytic gene expression. J. Biol. Chem.285:11297–11307.
20. Majerciak, V., N. Pripuzova, J. P. McCoy, S. J. Gao, and Z. M. Zheng. 2007.Targeted disruption of Kaposi’s sarcoma-associated herpesvirus ORF57 inthe viral genome is detrimental for the expression of ORF59, K8alpha, andK8.1 and the production of infectious virus. J. Virol. 81:1062–1071.
21. Majerciak, V., et al. 2008. Kaposi’s sarcoma-associated herpesvirus ORF57functions as a viral splicing factor and promotes expression of intron-con-taining viral lytic genes in spliceosome-mediated RNA splicing. J. Virol.82:2792–2801.
22. Majerciak, V., K. Yamanegi, S. H. Nie, and Z. M. Zheng. 2006. Structuraland functional analyses of Kaposi sarcoma-associated herpesvirus ORF57nuclear localization signals in living cells. J. Biol. Chem. 281:28365–28378.
23. Majerciak, V., and Z. M. Zheng. 2009. Kaposi’s sarcoma-associated herpes-virus ORF57 in viral RNA processing. Front. Biosci. 14:1516–1528.
24. Malik, P., D. J. Blackbourn, and J. B. Clements. 2004. The evolutionarilyconserved Kaposi’s sarcoma-associated herpesvirus ORF57 protein interactswith REF protein and acts as an RNA export factor. J. Biol. Chem. 279:33001–33011.
25. Masuda, S., et al. 2005. Recruitment of the human TREX complex tomRNA during splicing. Genes Dev. 19:1512–1517.
26. Mercher, T., et al. 2001. Involvement of a human gene related to the Dro-sophila spen gene in the recurrent t(1;22) translocation of acute megakaryo-cytic leukemia. Proc. Natl. Acad. Sci. U. S. A. 98:5776–5779.
27. Murray, E. L., and D. R. Schoenberg. 2008. Assays for determining poly(A)tail length and the polarity of mRNA decay in mammalian cells. MethodsEnzymol. 448:483–504.
28. Nappi, F., et al. 2001. Identification of a novel posttranscriptional regulatoryelement by using a rev- and RRE-mutated human immunodeficiency virustype 1 DNA proviral clone as a molecular trap. J. Virol. 75:4558–4569.
29. Nekorchuk, M., Z. Han, T. T. Hsieh, and S. Swaminathan. 2007. Kaposi’ssarcoma-associated herpesvirus ORF57 protein enhances mRNA accumu-lation independently of effects on nuclear RNA export. J. Virol. 81:9990–9998.
30. Oswald, F., et al. 2002. SHARP is a novel component of the Notch/RBP-Jkappa signalling pathway. EMBO J. 21:5417–5426.
31. Oswald, F., et al. 2005. RBP-Jkappa/SHARP recruits CtIP/CtBP corepres-sors to silence Notch target genes. Mol. Cell Biol. 25:10379–10390.
32. Ote, I., et al. 2009. Varicella-zoster virus IE4 protein interacts with SRproteins and exports mRNAs through the TAP/NXF1 pathway. PLoS ONE.4:e7882.
33. Pinol-Roma, S., S. A. Adam, Y. D. Choi, and G. Dreyfuss. 1989. Ultraviolet-induced cross-linking of RNA to proteins in vivo. Methods Enzymol. 180:410–418.
34. Pryor, A., et al. 2004. Growth-regulated expression and G0-specific turnoverof the mRNA that encodes URH49, a mammalian DExH/D box protein thatis highly related to the mRNA export protein UAP56. Nucleic Acids Res.32:1857–1865.
35. Raffel, G. D., et al. 2009. Ott1 (Rbm15) is essential for placental vascularbranching morphogenesis and embryonic development of the heart andspleen. Mol. Cell Biol. 29:333–341.
36. Raffel, G. D., et al. 2007. Ott1(Rbm15) has pleiotropic roles in hematopoieticdevelopment. Proc. Natl. Acad. Sci. U. S. A. 104:6001–6006.
37. Rebay, I., et al. 2000. A genetic screen for novel components of the Ras/mitogen-activated protein kinase signaling pathway that interact with the yangene of Drosophila identifies split ends, a new RNA recognition motif-containing protein. Genetics 154:695–712.
38. Sandri-Goldin, R. M. 2008. The many roles of the regulatory protein ICP27during herpes simplex virus infection. Front. Biosci. 13:5241–5256.
39. Schreiber, S. L., A. Preiss, A. C. Nagel, I. Wech, and D. Maier. 2002. Geneticscreen for modifiers of the rough eye phenotype resulting from overexpres-sion of the Notch antagonist hairless in Drosophila. Genesis 33:141–152.
40. Sergeant, A., H. Gruffat, and E. Manet. 2008. The Epstein-Barr virus (EBV)protein EB is an mRNA export factor essential for virus production. Front.Biosci. 13:3798–3813.
41. Shi, Y., et al. 2001. Sharp, an inducible cofactor that integrates nuclearreceptor repression and activation. Genes Dev. 15:1140–1151.
42. Smulevitch, S., et al. 2006. RTE and CTE mRNA export elements synergis-tically increase expression of unstable, Rev-dependent HIV and SIVmRNAs. Retrovirology 3:6.
43. Smulevitch, S., et al. 2005. Structural and functional analysis of the RNAtransport element, a member of an extensive family present in the mousegenome. J. Virol. 79:2356–2365.
44. Strasser, K., and E. Hurt. 2001. Splicing factor Sub2p is required for nuclearmRNA export through its interaction with Yra1p. Nature 413:648–652.
45. Strasser, K., et al. 2002. TREX is a conserved complex coupling transcrip-tion with messenger RNA export. Nature 417:304–308.
46. Tintaru, A. M., et al. 2007. Structural and functional analysis of RNA andTAP binding to SF2/ASF. EMBO Rep. 8:756–762.
47. Toth, Z., and T. Stamminger. 2008. The human cytomegalovirus regulatoryprotein UL69 and its effect on mRNA export. Front. Biosci. 13:2939–2949.
48. Tretyakova, I., et al. 2005. Nuclear export factor family protein participatesin cytoplasmic mRNA trafficking. J. Biol. Chem. 280:31981–31990.
49. Uranishi, H., et al. 2009. The RNA binding motif protein 15B (RBM15B/OTT3) acts as co-factor of the nuclear export receptor NXF1. J. Biol. Chem.284:26106–26116.
50. Wiellette, E. L., et al. 1999. spen encodes an RNP motif protein that interactswith Hox pathways to repress the development of head-like sclerites in theDrosophila trunk. Development 126:5373–5385.
51. Zhou, Z., L. J. Licklider, S. P. Gygi, and R. Reed. 2002. Comprehensiveproteomic analysis of the human spliceosome. Nature 419:182–185.
52. Zolotukhin, A. S., et al. 2008. The RNA transport element RTE is essentialfor IAP LTR-retrotransposon mobility. Virology 377:88–99.
53. Zolotukhin, A. S., et al. 2009. Nuclear export factor RBM15 facilitates theaccess of DBP5 to mRNA. Nucleic Acids Res. 37:7151–7162.
1540 MAJERCIAK ET AL. J. VIROL.





























