Embed Size (px)
Citation preview

1991 78: 2715-2720
Volpi, C Patrono and E DejanaP Patrignani, A Del Maschio, G Bazzoni, L Daffonchio, A Hernandez, R Modica, L Montesanti, D lytic enzymesInactivation of endothelin by polymorphonuclear leukocyte-derived
http://bloodjournal.hematologylibrary.org/content/78/10/2715.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom

Inactivation of Endothelin by Polymorphonuclear Leukocyte-Derived Lytic Enzymes
By Paola Patrignani, Aldo Del Maschio, Gianfranco Bazzoni, Luisa Daffonchio, Alicia Hernandez, Rossana Modica, Lilli Montesanti, Daniele Volpi, Carlo Patrono, and Elisabetta Dejana
Cultured bovine aortic endothelial cells (BAEC) released endothelin-1 (ET-1) in the culture medium in a time- dependent fashion. Coincubation of fMLP-activated human polymorphonuclear leukocytes (PMN) with BAEC caused a fast (maximal activity was reached within 15 minutes) and cell number-dependent disappearance of ET-1 from the me- dium. This effect was direct t o ET-1, because it was also present when PMN were incubated with the synthetic pep- tide in the absence of BAEC. PMN-dependent disappearance of ET-1 was associated with loss of constrictor activity on isolated rabbit aorta. PMN-released products were responsi- ble for ET-1 degrading activity, because supernatants of activated PMN were equally effective as the intact cells. Resting PMN, in the same time frame, were uneffective. Eglin
NDOTHELIN-1 (ET-l), a 21-amino acid peptide re- E cently isolated from the medium of cultured endothe- lial cells,’ represents one of the most potent vasoconstric- tors known. Relatively high blood levels of this peptide have been found in different pathologies, including uremia,’~~ acute myocardial infarction: and acute renal failure?
ET-1 can also play a role in inflammatory conditions; indeed, interleukin-1 and tumor necrosis factor, two media- tors of the acute inflammatory response, were reported to stimulate endothelial cell ET-1 production in vitro and in vivo?’ Despite being devoid of any direct stimulatory effect on leukocyte activation,* ET-1 can exert an antiinflamma- tory action by suppressing enhanced vascular permeability induced by several inflammatory mediators.’
The interaction of polymorphonuclear leukocytes (PMN) with endothelial cells, one of the first events in the acute inflammatory response and tissue ischemia, induces pro- found changes in the biosynthesis of potent endothelial modulators of vascular tone and The aim of this work was to assess the effect of PMN on ET-1 production by endothelial cells. We report that addition of fMLP-activated PMN dramatically reduces ET-1 content and biologic activity in endothelial cell culture media. This effect was apparently due to degradation of ET-1 by PMN-derived lytic enzymes. We suggest that this is a physiologic mechanism of ET-1 inactivation in vivo.
MATERIALS AND METHODS
PMN were isolated from venous blood of healthy volun- teers according to the method of B0yum” and resuspended in Hepes-Tyrode buffer supplemented with 0.1% bovine serum albu- min (BSA). BAEC were cultured in Modified Eagle’s Medium (MEM) supplemented with 15% fetal bovine serum (GIBCO- Europe, Paisley, UK) as described,” and used between three and 10 passages. For the experiments described, bovine aortic endothe- lial cells (BAEC) detached by brief exposure to trypsin (0.25%)- EDTA (0.022%), were plated and grown to confluence in 24-well plates.
Aliquots of PMN (0.37 to 3 x 1@ cells in a final volume of 300 FL) were added to each BAEC well in the presence of fibrinogen (Kabi Vitrum, Stockholm, Sweden) and cytochalasin B (0.38 mg/mL and 2.5 Fg/mL, respectively). After
Cells.
Eiperimentalprocedures.
C, a potent blocker of PMN-derived elastase and cathepsin G, reversed the ET-1 inhibitory activity of fMLP-stimulated PMN and of their supernatant. Direct addition of elastase and cathepsin G to synthetic ET-1 destroyed its immunoreactivity and this effect was blocked by eglin C. High-performance liquid chromatography (HPLC) analysis supported the hypoth- esis that ET-1 degradation by PMN was due to enzymatic proteolysis. These data provide evidence that activated PMN are able to degrade ET-1 through the release of proteases. Because physiologic concentrations of PMN can destroy high amounts (up to 100 nmol/L) of ET-1 within a few minutes, we propose that this mechanism of ET-1 inactivation has bio- logic relevance. 0 1991 by The American Society of Hematology.
addition of lo-’ mol/L M L P (Sigma Chemical, St Louis, MO) or the equivalent volume of isotonic saline, plates were incubated at 37°C for the times indicated (15 to 180 minutes). At the end of the incubation, the assay medium was collected and centrifuged at 12,OOQg for 1 minute. Supernatants were then collected and stored at -20°C for ET-1 determination by radioimmunoassay.
In some experiments, either PMN (in the presence or absence of mol/L M L P ) or PMN-derived supernatants were incubated
(15 minutes at 37°C) with different concentrations of synthetic ET-1 (1 to 100 nmol/L). Samples were then centrifuged and the supernatants collected for measuring ET-1 recovery. PMN-derived supernatants were prepared by centrifuging 1.5 x lo7 cells/mL that had been previously incubated (37°C for 15 minutes) with 3 x lo-’ mol/L M L P or isotonic saline. In some experiments, supernatants were obtained following PMN stimulation with platelet-activating factor (PAF), leukotriene B, (LTB,), or zymosan-activated serum ( U S ) . ET-1 degrading activity of PMN and PMN-derived superna- tants was evaluated in presence of the following protease inhibi- tors: leupeptin (Sigma), phosphoramidon (Boehringer, Mannheim, Germany), eglin C (a generous gift of Dr H.P. Schnebli, Ciba- Geigy, Basel, Switzerland), and EDTA (Merck, Darmstadt, Ger- many). These compounds were added to PMN or PMN-derived supernatants before addition of synthetic ET-1 (100 nmol/L). Samples were then incubated at 3PC for 15 minutes, centrifuged, and subsequently assayed for ET-1 recovery.
In a set of experiments, ET-1 (100 nmol/L) was incubated (15 to 180 minutes at 3PC) in the absence or presence of 10 to 200 nmol/L purified human elastase
Studies with purijied proteases.
From the University of Chieti “G. D’Annunzio” School of Medicine, Chieti; Laboratory of Vascular Biology, Istituto di Ricerche Farmaco- logiche “Mario Negri; Milano; Institute of Pharmacological Sciences, University of Milano; and Farmitalia Carlo Erba, Milano, Italy.
Submitted December 24,1990; accepted July 18, 1991. Supported by the Italian National Research Council (CNR) (Project
CNR No. 89.01277.04). Address reprint requests to Dr Aldo Del Maschio, Istituto di Ricerche
Farmacologiche “Mario Negri;” Ea Eritrea 62-20157, Milano, Italy. The publication costs of this article were defrayed in pari by page
charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section I734 solely to indicate this fact. 0 1991 by The American Society of Hematology. 0006-4971 /91/ 7810-Oo21$3.O0/0
2715 Blood, Vol78, No 10 (November 15). 1991: pp 2715-2720
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom
2716 PATRIGNANI ET AL
(Elastin Products, Owensville, MO) or cathepsin G (a generous gift of Dr M. Chignard, Institut Pasteur, Pans, France) in 1 mL of Hepes-Tyrode buffer or human whole blood. At the end of the incubation, samples in buffer were immediately frozen; samples in blood were centrifuged at 4°C (3,000 rpm, 10 minutes), and plasma was collected and frozen. Samples were then assayed for ET-1 content by radioimmunoassay.
Radioimmunoassay technique. A radioimmunoassay technique for endothelin was developed. The assay used 2,000 dpm of lZSI-ET-l (Novabiochem, Laufelfingen, Switzerland) and an anti- ET-1 serum (Peninsula, Belmont, CA) diluted 1:100,ooO that showed 100% of cross-reactivitywith both ET-1 and ET-2,9% with ET-3, 1.5% with big ET-l(1-38), and 0.83% with big ET-l(1-39). Aliquots (100 pL) of unextracted incubation media were incubated at 4°C for 72 hours in 2.5 ml of 0.025 mol/L Tris-phosphate-HC1 (pH 9). Separation of the antibody-bound ET-1 from free ligand was achieved by rapidly adding 50 pL of charcoal-treated human plasma and 100 pL of a charcoal suspension (100 mg/mL) and subsequent centrifugation at 3,000 rpm for 10 minutes at 4°C. Supernatant solutions (antibody-bound ET-1) and pellets (charcoal- adsorbed ET-1) were counted in a Beckman gamma counter (Fullerton, CA). The IC,, value (concentration of unlabeled ET-1 that reduced by 50% the bound to free ratio of the homologous tracer) was 2.4 fmol/mL. The least detectable concentration that could be measured with 95% confidence (ie, 2 SD at zero) was 0.2 fmol/mL. Validation of ET-1 measurements was obtained by dilution and recovery studies.
In the experiments reported here, the recovery of synthetic ET-1 after 15 minutes of incubation averaged 80%. This was considered the maximal recovery (100%) and the results are referred to this value. This reduction of ET-1 recovery did not further decrease with time (up to 180 minutes) and probably reflected an aspecific ET-1 binding to plastic surfaces.
Male albino rabbits (2.0 to 2.2 kg; Charles River, Calco, Italy) were killed by excess urethane anesthesia and exsanguinated. The descending thoracic aorta was isolated, cut as spirals of approximately 30 mm in length, and the segments were mounted in an organ bath (20 mL) containing oxygenated (OJCO,: 95%/5%) Krebs-bicarbonate solution at 3PC under a load of 2 g. Samples (1 mL) containing synthetic ET-1 (100 nmol/L) preincubated (15 minutes at 37°C) with or without lo-’ mol/L fMLP-activated PMN (3 x lo6) were spun down and supernatants (200 pL) added to the aorta segments; the response to ET-1 (final concentration, 1 nmol/L) was recorded during 20 minutes. Changes in the length of tissues were measured isotoni- cally and the data were expressed as millimeters of contraction. All the preparations showed the same maximal contractile response to norepinephrine (3 x mol/L).
ET-1 (400 nmol/L) was incubated (15 minutes at 37°C) with 100 nmol/L cathepsin G or elastase or control buffer, in the presence or absence of 25 pg/mL eglin C. Samples were applied to SEP-PAK C18 cartridges (Waters, Milford, MA) pretreated with 10 mL methanol and 10 mL water. After washing with 10 mL of trifluoro- acetic acid (TFA, O.l%), the peptides were eluted with 3 mL of acetonitrile (60%) in 0.1% TFA. The eluted samples were concen- trated by evaporation under vacuum and reconstituted in 0.15 mL of 30% acetonitrile in 0.1% TFA and subjected to reverse-phase HPLC analysis by using a p-bondapack C18 column (0.39 X 30 cm; Waters). Elution was performed by using a binary gradient consist- ing of 10% acetonitrile in 0.1% TFA (solvent A) and 90% acetonitrile in 0.1% TFA (solvent B). The elution profile incorpo- rated a linear gradient from 10% to 80% solvent B in 60 minutes at a flow rate of 1 mL/min. Eluates were monitored by absorbance at 220 nm. Synthetic ET-1 eluted as a single peak with a retention
Bioassay of vasoconstrictor activiy.
High-performance liquid chromatography (HPLC) analysis.
time of 28.2 2 0.2 minutes (mean * SD, n = 4). ET-1 extraction by SEP-PAK C18 cartridges caused the appearance of an additional peak eluting at 26.6 f 0.2 minutes. Since both peaks were recognized by ET-1 antiserum, the peptide eluting at 26.6 minutes is likely to represent an aggregated form of ET-1, as recently r e~0r t ed . l~
RESULTS
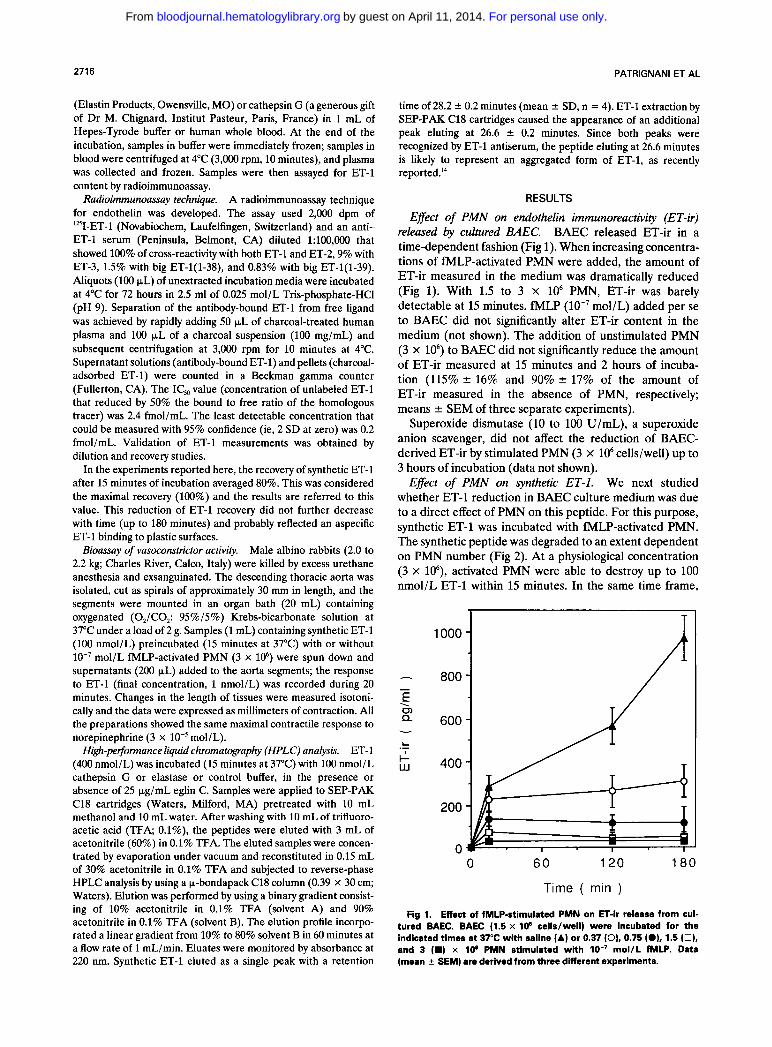
Effect of PMN on endothelin immunoreactivity (ET-ir) released by cultured BAEC. BAEC released ET-ir in a time-dependent fashion (Fig 1). When increasing concentra- tions of fMLP-activated PMN were added, the amount of ET-ir measured in the medium was dramatically reduced (Fig 1). With 1.5 to 3 x lo6 PMN, ET-ir was barely detectable at 15 minutes. fMLP (lo-’ mol/L) added per se to BAEC did not significantly alter ET-ir content in the medium (not shown). The addition of unstimulated PMN (3 x lo6) to BAEC did not significantly reduce the amount of ET-ir measured at 15 minutes and 2 hours of incuba- tion (115% r 16% and 90% * 17% of the amount of ET-ir measured in the absence of PMN, respectively; means r SEM of three separate experiments).
Superoxide dismutase (10 to 100 U/mL), a superoxide anion scavenger, did not affect the reduction of BAEC- derived ET-ir by stimulated PMN (3 X lo6 cells/well) up to 3 hours of incubation (data not shown).
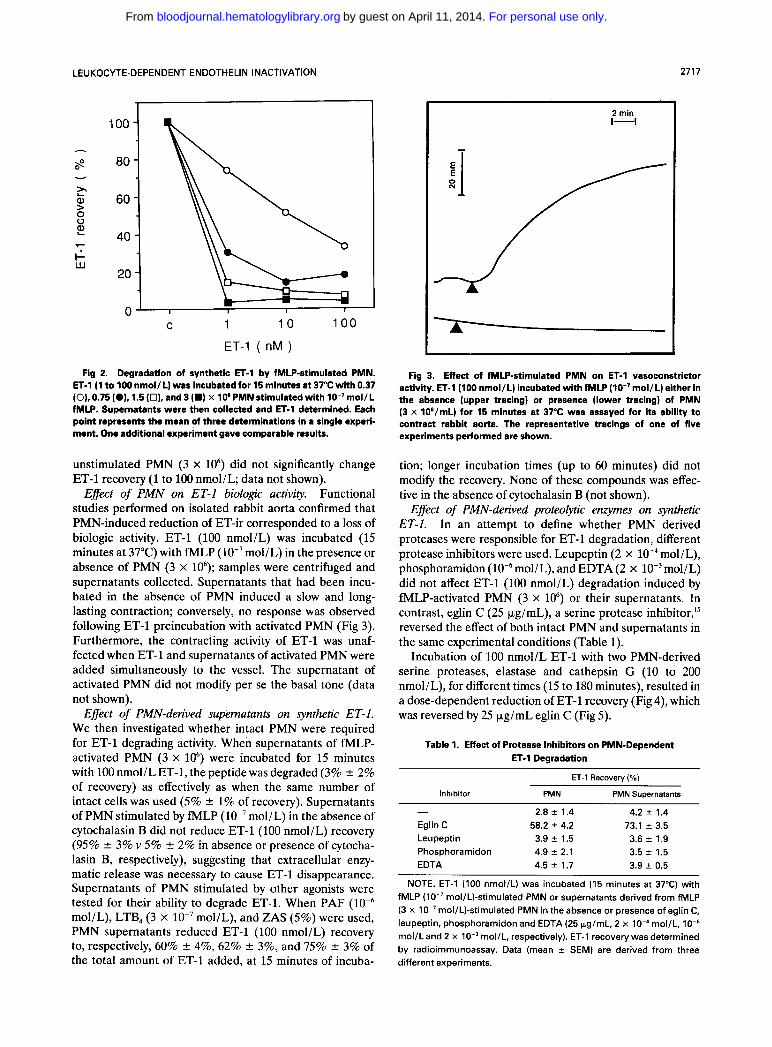
Effect of PMN on synthetic ET-1. We next studied whether ET-1 reduction in BAEC culture medium was due to a direct effect of PMN on this peptide. For this purpose, synthetic ET-1 was incubated with fMLP-activated PMN. The synthetic peptide was degraded to an extent dependent on PMN number (Fig 2). At a physiological concentration (3 x lo6), activated PMN were able to destroy up to 100 nmol/L ET-1 within 15 minutes. In the same time frame,
h
- E . 0) Q v
L .- I- w
1000
800
600
400
200
0 0 6 0 120 180
Time ( min )
Fig 1. Effect of fMLP-stimulated PMN on ET-ir release from cul- tured BAEC. BAEC (1.5 x I O 6 cells/well) were incubated for the indicated times at 37°C with saline (A) or 0.37 (0). 0.75 (OL1.5 (0). and 3 (W) x I O e PMN stimulated with IO-’ mol/L fMLP. Data (mean 2 SEM) are derived from three different experiments.
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom
LEUKOCYTE-DEPENDENT ENDOTHELIN INACTIVATION 2717
-- I \A? n ~ -
C 1 1 0
ET-1 ( nM )
100
Fig 2. Degradation of synthetic ET-1 by fMLP-stimulated PMN. ET-1 (1 to 100 nmol/L) was incubated for 15 minutes at 37°C with 0.37 (O), 0.75 IO), 1.5 (0). and 3 (W) x I O 6 PMN stimulated with mol/L fMLP. Supernatants were then collected and ET-1 determined. Each point represents the mean of three determinations in a single experi- ment. One additional experiment gave comparable results.
unstimulated PMN (3 x lo6) did not significantly change ET-1 recovery (1 to 100 nmol/L; data not shown).
Effect of PMN on ET-1 biologic activity. Functional studies performed on isolated rabbit aorta confirmed that PMN-induced reduction of ET-ir corresponded to a loss of biologic activity. ET-1 (100 nmol/L) was incubated (15 minutes at 37°C) with fMLP (lO-’mol/L) in the presence or absence of PMN (3 x 10‘); samples were centrifuged and supernatants collected. Supernatants that had been incu- bated in the absence of PMN induced a slow and long- lasting contraction; conversely, no response was observed following ET-1 preincubation with activated PMN (Fig 3). Furthermore, the contracting activity of ET-1 was unaf- fected when ET-1 and supernatants of activated PMN were added simultaneously to the vessel. The supernatant of activated PMN did not modify per se the basal tone (data not shown).
Effect of PMN-derived supematants on synthetic ET-1. We then investigated whether intact PMN were required for ET-1 degrading activity. When supernatants of fMLP- activated PMN (3 x lo6) were incubated for 15 minutes with 100 nmol/L ET-1, the peptide was degraded (3% f 2% of recovery) as effectively as when the same number of intact cells was used (5% +- 1% of recovery). Supernatants of PMN stimulated by fMLP (lo-’ mol/L) in the absence of cytochalasin B did not reduce ET-1 (100 nmol/L) recovery (95% f 3% v 5% 2 2% in absence or presence of cytocha- lasin B, respectively), suggesting that extracellular enzy- matic release was necessary to cause ET-1 disappearance. Supernatants of PMN stimulated by other agonists were tested for their ability to degrade ET-1. When PAF mol/L), LTB, (3 x lo-’ mol/L), and ZAS (5%) were used, PMN supernatants reduced ET-1 (100 nmol/L) recovery to, respectively, 60% 2 4%, 62% f 3%, and 75% f 3% of the total amount of ET-1 added, at 15 minutes of incuba-
2 min 1-1
Fig 3. Effect of fMLP-stimulated PMN on ET-1 vasoconstrictor activity. ET-1 (100 nmol/L) incubated with fMLP (IO-’mol/L) either in the absence (upper tracing) or presence (lower tracing) of PMN (3 x 106/mL) for 15 minutes at 37°C was assayed for its ability to contract rabbit aorta. The representative tracings of one of five experiments performed are shown.
tion; longer incubation times (up to 60 minutes) did not modify the recovery. None of these compounds was effec- tive in the absence of cytochalasin B (not shown).
Effect of PMN-derived proteolytic enzymes on synthetic ET-1. In an attempt to define whether PMN derived proteases were responsible for ET-1 degradation, different protease inhibitors were used. Leupeptin (2 x lo-, mol/L), phosphoramidon mol/L), and EDTA (2 x lO-’moI/L) did not affect ET-1 (100 nmol/L) degradation induced by fMLP-activated PMN (3 x lo6) or their supernatants. In contrast, eglin C (25 kg/mL), a serine protease inhibitor,” reversed the effect of both intact PMN and supernatants in the same experimental conditions (Table 1).
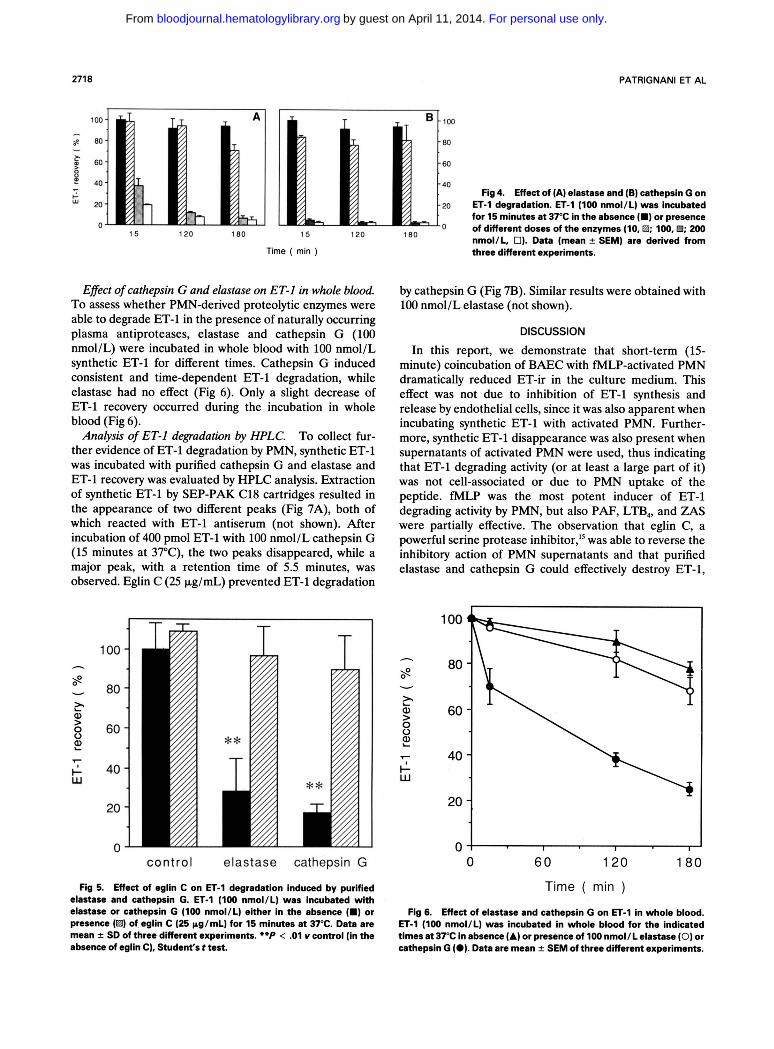
Incubation of 100 nmol/L ET-1 with two PMN-derived serine proteases, elastase and cathepsin G (10 to 200 nmol/L), for different times (15 to 180 minutes), resulted in a dose-dependent reduction of ET-1 recovery (Fig 4), which was reversed by 25 pg/mL eglin C (Fig 5).
Table 1. Effect of Protease Inhibitors on PMN-Dependent ET-I Degradation
ET-1 Recovery ( O h )
inhibitor PMN PMN Supernatants
- 2.8 2 1.4 4.2 * 1.4 Eglin C 58.2 4.2 73.1 2 3.5 Leupeptin 3.9 f 1.5 3.6 2 1.9 Phosphoramidon 4.9 2 2.1 3.5 f 1.5 EDTA 4.5 2 1.7 3.9 f 0.5
NOTE. ET-1 (100 nmol/L) was incubated (15 minutes at 37°C) with fMLP (lo-’ mol/L)-stimulated PMN or supernatants derived from fMLP (3 x IO-’ mol/L)-stimulated PMN in the absence or presence of eglin C, leupeptin, phosphoramidon and EDTA (25 pg/mL, 2 x lO-‘mol/L, mol/L and 2 x mol/L, respectively). ET-1 recovery was determined by radioimmunoassay. Data (mean f SEM) are derived from three different experiments.
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom
2718 PATRlGNANl ET AL
100 100 - $ 80 80 - F w 60
40 40 Fig 4. Effect of (A) elastase and (6) cathepsin G on G
u 2 0 20 ET-1 degradation. ET-1 (100 nmol/L) was incubated for 15 minutes at 37°C in the absence I.) or presence of different doses of the enzymes (10. B; 100,B; 200 nmol/L, 01. Data [mean -t SEMI are derived from
- n
180 15 120 180 15 120
Time ( min )
Effect of cathepsin G and elastase on ET-I in whole blood. To assess whether PMN-derived proteolytic enzymes were able to degrade ET-1 in the presence of naturally occurring plasma antiproteases, elastase and cathepsin G (100 nmol/L) were incubated in whole blood with 100 nmol/L synthetic ET-1 for different times. Cathepsin G induced consistent and time-dependent ET-1 degradation, while elastase had no effect (Fig 6). Only a slight decrease of ET-1 recovery occurred during the incubation in whole blood (Fig 6).
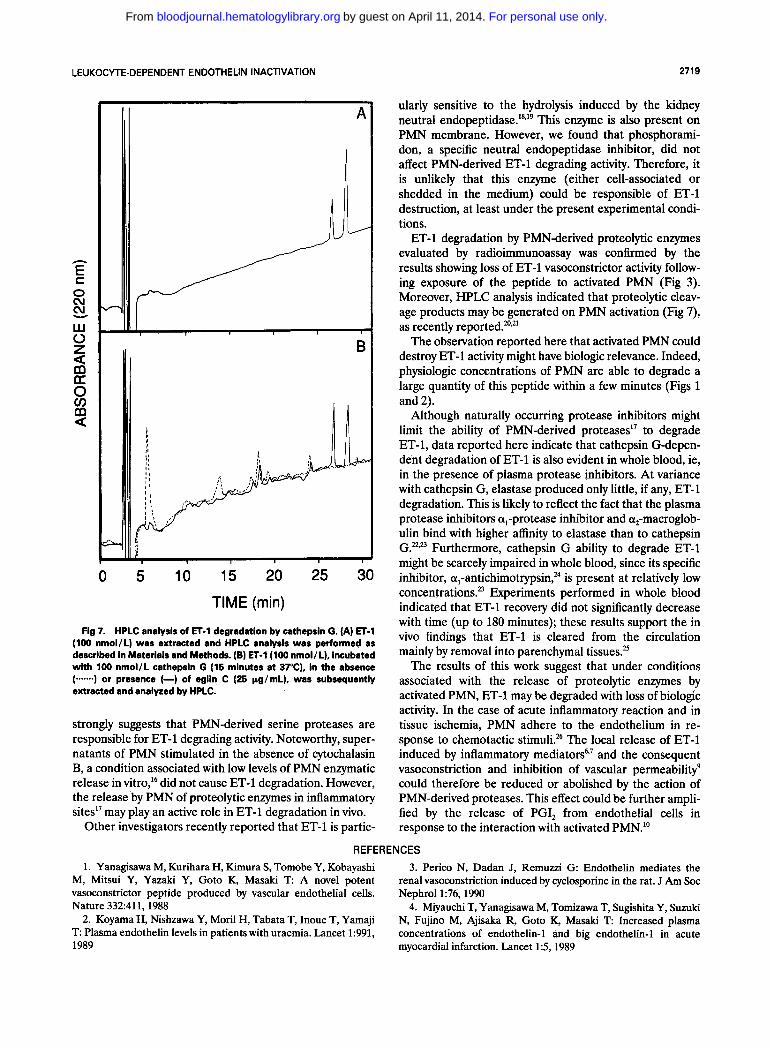
To collect fur- ther evidence of ET-l degradation by PMN, synthetic ET-l was incubated with purified cathepsin G and elastase and ET-1 recovery was evaluated by HPLC analysis. Extraction of synthetic ET-1 by SEP-PAK C18 cartridges resulted in the appearance of two different peaks (Fig 7A), both of which reacted with ET-1 antiserum (not shown). After incubation of 400 pmol ET-1 with 100 nmol/L cathepsin G (15 minutes at 37"C), the two peaks disappeared, while a major peak, with a retention time of 5.5 minutes, was observed. Eglin C (25 pg/mL) prevented ET-1 degradation
Anahsis of ET-I degradation by HPLC.
100 -
80 -
60 -
40 -
20 -
0-
T
control elastase cathepsin G
Fig 5. Effect of eglin C on ET4 degradation induced by purlfled elastase and cathepsin G. ET-1 (100 nmol/L) was incubated with elastase or cathepsin G (100 nmol/L) either in the absence (m) or presence (El) of eglin C (25 pg/mL) for 15 minutes at 37°C. Data are mean f SD of three different experiments. **P < .01 Y control (in the absence of eglin C), Student's t test.
three different expekents.
by cathepsin G (Fig 7B). Similar results were obtained with 100 nmol/L elastase (not shown).
DISCUSSION
In this report, we demonstrate that short-term (15- minute) coincubation of BAEC with MLP-activated PMN dramatically reduced ET-ir in the culture medium. This effect was not due to inhibition of ET-1 synthesis and release by endothelial cells, since it was also apparent when incubating synthetic ET-1 with activated PMN. Further- more, synthetic ET-1 disappearance was also present when supernatants of activated PMN were used, thus indicating that ET-1 degrading activity (or at least a large part of it) was not cell-associated or due to PMN uptake of the peptide. M L P was the most potent inducer of ET-1 degrading activity by PMN, but also PAF, LTB,, and Z A S were partially effective. The observation that eglin C, a powerful serine protease inhibitor," was able to reverse the inhibitory action of PMN supernatants and that purified elastase and cathepsin G could effectively destroy ET-1,
1 0 60 120 180
Time ( min )
Fig 6. Effect of elastase and cathepsin G on ET-1 in whole blood. ET-1 (100 nmol/L) was incubated in whole blood for the indicated times at 37°C in absence (A) or presence of 100 nmol/L elastase (0) or cathepsin G (0). Data are mean f SEM of three different experiments.
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom
LEUKOCYTE-DEPENDENT ENDOTHELIN INACTIVATION 2719
h
E
e!. C 0 cu
LU 0 z U 0 cn m a
3 B
0 5 10 15 20 25 30 TIME (min)
Fig 7. HPLC analysis of ET-1 degradation by cathepsin G. (A) ET-1 (100 nmol/L) was extracted and HPLC analysis was performed as described in Materials and Methods. (B) ET-1 (100 nmol/L), incubated with 100 nmol/L cathepsin G (15 minutes at 37"C), in the absence (.--) or presence (-) of eglin C (25 pg/mL), was subsequently extracted and analyzed by HPLC.
strongly suggests that PMN-derived serine proteases are responsible for ET-1 degrading activity. Noteworthy, super- natants of PMN stimulated in the absence of cytochalasin B, a condition associated with low levels of PMN enzymatic release in vitro,16 did not cause ET-1 degradation. However, the release by PMN of proteolytic enzymes in inflammatory sited7 may play an active role in ET-1 degradation in vivo.
Other investigators recently reported that ET-1 is partic-
ularly sensitive to the hydrolysis induced by the kidney neutral end~peptidase. '~~'~ This enzyme is also present on PMN membrane. However, we found that phosphorami- don, a specific neutral endopeptidase inhibitor, did not affect PMN-derived ET-1 degrading activity. Therefore, it is unlikely that this enzyme (either cell-associated or shedded in the medium) could be responsible of ET-1 destruction, at least under the present experimental condi- tions.
ET-1 degradation by PMN-derived proteolytic enzymes evaluated by radioimmunoassay was confirmed by the results showing loss of ET-1 vasoconstrictor activity follow- ing exposure of the peptide to activated PMN (Fig 3). Moreover, HPLC analysis indicated that proteolytic cleav- age products may be generated on PMN activation (Fig 7), as recently
The observation reported here that activated PMN could destroy ET-1 activity might have biologic relevance. Indeed, physiologic concentrations of PMN are able to degrade a large quantity of this peptide within a few minutes (Figs 1 and 2).
Although naturally occurring protease inhibitors might limit the ability of PMN-derived proteases" to degrade ET-1, data reported here indicate that cathepsin G-depen- dent degradation of ET-1 is also evident in whole blood, ie, in the presence of plasma protease inhibitors. At variance with cathepsin G, elastase produced only little, if any, ET-1 degradation. This is likely to reflect the fact that the plasma protease inhibitors &,-protease inhibitor and q-macroglob- ulin bind with higher affinity to elastase than to cathepsin G.".= Furthermore, cathepsin G ability to degrade ET-1 might be scarcely impaired in whole blood, since its specific inhibitor, a,-antichimotrypsin," is present at relatively low concentrations.= Experiments performed in whole blood indicated that ET-1 recovery did not significantly decrease with time (up to 180 minutes); these results support the in vivo findings that ET-1 is cleared from the circulation mainly by removal into parenchymal tissues.=
The results of this work suggest that under conditions associated with the release of proteolytic enzymes by activated PMN, ET-1 may be degraded with loss of biologic activity. In the case of acute inflammatory reaction and in tissue ischemia, PMN adhere to the endothelium in re- sponse to chemotactic stimuli?6 The local release of ET-1 induced by inflammatory mediators6s7 and the consequent vasoconstriction and inhibition of vascular permeability' could therefore be reduced or abolished by the action of PMN-derived proteases. This effect could be further ampli- fied by the release of PGI, from endothelial cells in response to the interaction with activated PMN."
REFERENCES 1. Yanagisawa M, Kurihara H, Kimura S , Tomobe Y, Kobayashi
M, Mitsui Y, Yazaki Y, Goto K, Masaki T A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 332:411,1988
2. Koyama H, Nishzawa Y, Moril H, Tabata T, Inoue T, Yamaji T Plasma endothelin levels in patients with uraemia. Lancet 1:991, 1989 myocardial infarction. Lancet 15,1989
3. Perico N, Dadan J, Remuzzi G: Endothelin mediates the renal vasoconstriction induced by cyclosporine in the rat. J Am SOC Nephrol1:76,1990
4. Miyauchi T, Yanagisawa M, Tomizawa T, Sugishita Y, Suzuki N, Fujino M, Ajisaka R, Goto K, Masaki T Increased plasma concentrations of endothelin-1 and big endothelin-1 in acute
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom

2720 PATRlGNANl ET AL
5. Tomita K, Ojile K, Nakanishi T, Tomura S, Matsuda 0, Ando K, Shichiri M, Hirata Y, Marumo F Plasma endothelin levels in patients with acute renal failure. N Engl J Med 321:1127, 1989
6. Yoshizumi M, Kurihara H, Morita T, Yamashita T, Oh-hashi Y, Sugiyama T, Takaku F, Yanagisawa M, Masaki T, Yazaki Y: Interleukin 1 increases the production of endothelin-1 by cultured endothelial cells. Biochem Biophys Res Commun 166:324, 1990
7. Vemulapalli S, Chin PJS, Rivelli M, Cedeno K, Coleman W, Sybertz EJ: Tumor necrosis factor and hemorrage produce eleva- tion of plasma endothelins in rats. FASEB J 4:A456,1990
8. Bazzoni G, Chen ZG, Mantovani A, Dejana E, Del Maschio A Effect of endothelin-1 on human polymorphonuclear leukocyte functions. Life Sci Adv 9:373, 1990
9. Chander CL, Howat DW, Moore AR, Colville-Nash PR, Desa FM, Braquet P, Willoughby D A Comparison of endothelin-1 and -3 on models of inflammation. Agents Actions 29:27,1990
10. Miller KD, Sadowski S, Sodeiman DD, Kuehl F A Endothe- lial cell prostacyclin production induced by activated neutrophils. J Biol Chem 260:1006,1985
11. Feinmark SJ, Cannon PJ: Endothelial cell leukotriene C, synthesis results from intercellular transfer of leukotriene A, synthesized by polymorphonuclear leukocytes. J Biol Chem 261: 16466,1986
12. B~yum A A one-stage procedure for isolation of granulo- cytes and lymphocytes from human blood. General sedimentation properties of white blood cells in a 1 g gravity field. Scand J Clin Lab Invest 2151, 1968 (suppl97)
13. Dejana E, Balconi G, De Castellarnau C, Barbieri B, Vergara-Dauden M, de Gaetano G: Prostacyclin production by human endothelial and bovine smooth muscle cells in culture. Effect of repeated stimulation with arachidonic acid, thrombin and ionophore A23187. Biochem Biophys Acta 750:261,1983
14. Bennes R, Calas B, Chabrier PE, Demaille J, Heitz F: Evidence for aggregation of endothelin-1 in water. FEBS Lett 276:21, 1990
15. Renesto P, Ferrer-Lopez P, Chignard M: Interference of recombinant eglin C, a proteinase inhibitor extracted from leeches,
with neutrophil-mediated platelet activation. Lab Invest 62:409, 1990
16. Zurier RB, Hoffstein S, Weissmann G: Cytochalasin B: effect on lysosomal enzyme release from human leukocytes. Proc Natl Acad Sci USA 70:844,1973
17. Weiss SJ: Tissue destruction by neutrophils. N Engl J Med 257:2318,1989
18. Sokolowski M, Galron R, Kloog Y, Bdolah A, Indig FE, Blumberg S, Fleminger G: Endothelins are more sensitive than sarafotoxins to neutral endopeptidase: Possible physiological signif- icance. Proc Natl Acad Sci USA 87:4702,1990
19. Vijayaraghavan J, Scicli AG, Carretero OA, Slaughter C, Moomaw C, Hersh LB: The hydrolysis of endothelins by neutral endopeptidase 24.11 (Enkephalinase). J Biol Chem 265:14150, 1990
20. Sessa WC, Kaw S, Hecker M, Vane JR: The biosynthesis of endothelin-1 by human polymorphonuclear leukocytes. Biochem Biophys Res Commun 174:613,1991
21. Yoshizumi M, Morita T, Kurihara H, Oh-hashi Y, Yanagi- sawa M, Inoue A, Masaki T, Yazaki Y: Regulatory effects of macrophage-derived cytokines and polymorphonuclear leukocytes on endothelin production and its metabolism. J Vasc Med Bioi 2: 177, 1990
22. Beatty K, Bieth J, Travis J: Kinetics of association of serine proteinases with native and oxidized a-1-proteinase inhibitor and a-1-antichymotrypsin. J Biol Chem 255:3931,1980
23. Travis J: Structure, function and control of neutrophil proteinases. Am J Med 84:37, 1988 (suppl6A)
24. Travis J, Bowen J, Baugh R: Human a-1-antichymotrypsin: interaction with chymotrypsin-like proteinases. Biochemistry 17: 5651,1978
25. Shiba R, Yanagisawa M, Miyauchi T, Ishii Y, Kimura S, Uchiyama Y, Masaki T, Goto K: Elimination of intravenously injected endothelin-1 from the circulation of the rat. J Cardiovasc Pharmacol13:S98,1989 (suppl5)
26. Bazzoni G, Dejana E, Del Maschio A: Adrenergic modula- tion of human polymorphonuclear leukocyte activation. Potentiat- ing effect of adenosine. Blood 77:2042,1991
For personal use only.on April 11, 2014. by guest bloodjournal.hematologylibrary.orgFrom











![Comparison of PAF- and fMLP-induced [Ca2+]i transients in human polymorphonuclear leukocytes](https://img.dokumen.tips/doc/110x75/63607be75e6ba0ceb50ee373/comparison-of-paf-and-fmlp-induced-ca2i-transients-in-human-polymorphonuclear.jpg)

















