Embed Size (px)
Citation preview

J. Microbiol. Biotechnol. (2012), 22(12), 1613–1620http://dx.doi.org/10.4014/jmb.1208.08072First published online October 6, 2012pISSN 1017-7825 eISSN 1738-8872
Biocontrol Potential of a Lytic Bacteriophage PE204 against Bacterial Wiltof Tomato
Bae, Ju Young1†
, Jing Wu2†
, Hyoung Ju Lee3, Eun Jeong Jo
1, Senthilkumar Murugaiyan
4, Eunsook Chung
1,
and Seon-Woo Lee1,3*
1Department of Medical Bioscience, Dong-A University, Busan 604-714, Korea2Department of Biological and Pharmaceutical Engineering, Wuhan Polytechnic University, Wuhan 430-023, China3Department of Applied Biology, Dong-A University, Busan 604-714, Korea4Department of Agricultural Microbiology, Tamil Nadu Agricultural University, Coimbatore 641-003, Tamil Nadu, India
Received: August 28, 2012 / Accepted: September 6, 2012
Bacterial wilt caused by Ralstonia solanacearum is a
devastating disease of many economically important
crops. Since there is no promising control strategy for
bacterial wilt, phage therapy could be adopted using
virulent phages. We used phage PE204 as a model lytic
bacteriophage to investigate its biocontrol potential for
bacterial wilt on tomato plants. The phage PE204 has a
short-tailed icosahedral structure and double-stranded
DNA genome similar to that of the members of
Podoviridae. PE204 is stable under a wide range of
temperature and pH, and is also stable in the presence of
the surfactant Silwet L-77. An artificial soil microcosm
(ASM) to study phage stability in soil was adopted to
investigate phage viability under a controlled system.
Whereas phage showed less stability under elevated
temperature in the ASM, the presence of host bacteria
helped to maintain a stable phage population. Simultaneous
treatment of phage PE204 at 108 PFU/ml with R.
solanacearum on tomato rhizosphere completely inhibited
bacterial wilt occurrence, and amendment of Silwet L-77
at 0.1% to the phage suspension did not impair the disease
control activity of PE204. The biocontrol activities of
phage PE204 application onto tomato rhizosphere before
or after R. solanacearum inoculation were also investigated.
Whereas pretreatment with the phage was not effective in
the control of bacterial wilt, post-treatment of PE204
delayed bacterial wilt development. Our results suggested
that appropriate application of lytic phages to the plant
root system with a surfactant such as Silwet L-77 could be
used to control the bacterial wilt of crops.
Keywords: Bacterial wilt, bacteriophage, biocontrol, Ralstonia
solanacearum
Bacterial wilt is a destructive plant disease caused by the
soilborne Gram-negative bacterium Ralstonia solanacearum.
R. solanacearum naturally infects over 200 plant species
belonging to more than 50 botanical families, and causes
lethal wilt [14]. This bacterial pathogen is highly
heterogenous and has been divided into five races and six
biovars based on host range and carbon source utilization
[13, 15, 16]. A new hierachical classification of this
heterogenous group of bacteria was also proposed based
on phenotypic and molecular typing, to provide four
phylotypes, corresponding to the geographical origin of
the strains [7].
In spite of the economic importance of the disease on
various crops in many tropical and subtropical areas, disease
management is highly limited. There are no effective
chemicals available to eradicate the bacterial pathogens
from soil or water, nor to cure infected plants. Soil
fumigation using methyl bromide is no longer approved
for use, having been banned since 2005. Cultivation of
resistant crops may be effective, although there are not
many resistant crops available, and the breeding of wilt-
resistant cultivars is difficult because the genetics of
resistance is particularly complex [30]. Furthermore, the
heterogenous nature of the soilborne pathogen makes it
difficult to control, and its persistence in soil and water for
long periods complicates disease management [11, 23, 37].
*Corresponding authorPhone: +82-51-200-7551; Fax: +82-51-200-7505;E-mail: [email protected]†Ju Young Bae and Jing Wu contributed equally to this study.

1614 Bae et al.
Recently, Shin and Yun [32] reported that accelerated
bacterial wilt progress was mainly due to the increased
temperature rather than the elevated CO2 conditions.
Therefore, there is an urgent need to develop a novel
method to manage the bacterial wilt of various crops.
Phage therapy uses an active bacteriophage as a
biocontrol agent to kill a disease-causing bacteria. Although
phage therapy has received attention since the discovery of
phages, antibiotics and chemical sterilization have been
much more widely used to control bacterial disease [24].
Phage therapy to control plant bacterial diseases has been
proposed [2], and phages infecting R. solanacearum are
possible control agents for wilt disease [9]. Phages are
prevalent in nature where there is a high density of
metabolically active bacteria. Various phages have been
used for the study of biological control of different plant
diseases [2]. Since phages infecting R. solanacearum have
been discovered in field soil and diseased tobacco plants in
Japan [34, 35], various kinds of phages infecting R.
solanacearum strains have been characterized [1, 8, 19, 20,
25, 38]. Only a few studies were conducted to investigate
the potential of phages to control bacterial wilt. Since
biocontrol attempts with multiple phages have been
proposed for investigation [34, 35], single unique phages
with a broad host range have showed promising biocontrol
activity, suggesting that use of specific phages such as
ΦRSL1 could be more useful for the control of bacterial
wilt than the use of phage cocktails with virulent phages
[9]. Considering the huge diversity of phages [28], the
potential of phages for the control of plant diseases, such
as bacterial wilt, has not been sufficiently investigated. The
characterization of various phages and suitable investigation
of biocontrol conditions of phages may provide simple and
effective methods for the management of bacterial wilt of
various crops.
Recently, Murugaiyan et al. [25] isolated 15 different
phages infecting R. solanacearum from wilt-infected plant
rhizospheres. Some of the phages were highly virulent to
R. solanacearum strains, and we used a phage PE204,
isolated from a pepper field, to investigate the biocontrol
potential of the phage under controlled conditions. Various
environmental factors affect phage viability in soil and
water. In this study, we aimed to examine some environmental
factors affecting the phage stability of PE204, a member of
Podoviridae, and to investigate the biological control
activity of the phage against bacterial wilt of tomato plants.
MATERIALS AND METHODS
Bacterial Strain, Bacteriophage, and Culture Condition
The host bacterial strain R. solanacearum SL341 [18] is a strain in
race 1, phylotype I, and was routinely grown at 28oC in casamino
acid peptone glucose (CPG) broth, or in CPG plus 0.005% 2,3,5-
triphenyltetrazolium chloride (TZC) agar medium [22]. R. solanacearum
SL341 inoculums for plant infection were prepared by growing the
bacteria in mannitol-glutamate broth medium [21] for 24 h at 30oC
at 150 rpm. The bacterial cells were harvested by centrifugation at
12,000 rpm for 10 min, and were resuspended in sterile distilled
water and adjusted to have a cell density of 2 × 108 CFU/ml.
Bacteriophage PE204 was routinely propagated using the bacterial
host strain SL341 [25]. When necessary, phage stability was
investigated using a plaque assay with a soft agar. Bacterial lawns
were prepared by the double-layer agar plate method [33]. Briefly,
3 ml of CPG soft agar (CPG with 0.6% agar) with bacterial
suspension in water (5 × 108 CFU) was mixed with phage solution
in SM buffer (50 mM Tris-HCl, pH 7.5, 100 mM NaCl, 10 mM
MgSO4, and 0.01% gelatin) and the soft agar mixture was subsequently
poured on a regular CPG plate. The plates were incubated at 30oC
for 24 h to assess plaque formation.
Electron Microscopy of Purified Bacteriophage
Phage purification was conducted by following a standard protocol
of polyethylene glycol precipitation [25, 31]. The purified phage
particles were suspended and maintained in SM buffer. To examine
phage morphology, a concentrated phage suspension, containing ca.
108 PFU/ml phages in SM buffer, was spotted on a Formvar-carbon-
coated copper grid and the phages were allowed to adsorb for
2 min. The phages were stained by the addition of 2% (w/v) sodium
phosphotungstate (pH 7.4), and the grid was allowed to air-dry for
10 min. The phages were observed under a transmission electron
microscope (TEM; JEM-2010; JEOL, Tokyo, Japan) at 200 kV.
Recombinant DNA Technology and Nucleic Acid Analysis of
Bacteriophage
DNA manipulations, such as plasmid preparation, restriction
endonuclease digestion, DNA ligation, plasmid DNA transformation,
agarose gel electrophoresis, and other recombinant DNA techniques,
were carried out following standard methods [31]. Nucleic acids of
phage PE204 were extracted from 10 ml of purified phage suspension
using proteinase K and phenol-chloroform-isoamyl alcohol. The
double-stranded DNA of phage PE204 was digested with either
EcoRI or PstI, and the resulting DNA fragments were cloned into
pUC119 digested with the same restriction enzymes. The phage
genomic DNA, randomly cloned in pUC119, was sequenced to
obtain partial DNA sequence information. DNA sequencing and
primer synthesis were performed commercially at the DNA
sequencing facility of Cosmogenetech Corp. (Seoul, Korea). DNA
sequences were analyzed with the BLAST program provided by the
National Center for Biotechnology Information (NCBI).
Phage PE204 Stability In Vitro and Viability in Soil Microcosm
To investigate phage stability in various temperatures, the double-
layer agar method was conducted to count plaque formation using
phage solutions incubated at various temperatures [3]. The 0.2 ml of
phage suspension in SM buffer (108
PFU/ml) was incubated at
various temperatures for 1 h. The phage suspension was subsequently
diluted serially with SM buffer, and 0.1 ml of diluted suspension
was mixed with 0.1 ml of host strain SL341 (5 × 108 CFU/ml). The
mixture was added to 3 ml of CPG soft agar to generate a bacterial
lawn on CPG plates, and the number of plaques was counted.

BIOCONTROL POTENTIAL OF PHAGE PE204 1615
Phage stability under various conditions of pH was also investigated
similarly. Various pH solutions of 2 M [sodium citrate buffer (pH
3-5), sodium phosphate buffer (pH 6 and 7), and Tris-HCl (pH 8-
11)] were prepared. The pH buffer (0.1 ml) was mixed with the
same volume of phage solution (108 PFU/ml) to give a final pH
buffer concentration of 1 M, and the mixture was incubated at 30oC
for 1 h. Phage viability based on plaque formation was counted by
the method described above. The phage stability assay included
three replications for pH and temperature. The effect of a surfactant,
Silwet L-77, on phage stability was also examined by incubating
phage suspensions in the presence of Silwet L-77 (Sigma Chemical
Co., St. Louis, MO, USA). Silwet L-77 was added to a phage
PE204 suspension (5 × 106 PFU/ml) in SM buffer, which was
incubated at 30oC for 24 h. Plaque formation was counted by the
double-layer agar method described above.
Phage viability in situ was also investigated at various temperatures
using an artificial soil microcosm (ASM; Ellis, 2004). A PE204
phage solution (107 PFU/ml) in SM buffer (0.4 ml) was added to
12.75 g of ASM, which was maintained at 25oC, 30oC, and 35oC for
a total period of 15 days. One gram of ASM containing PE204 in
SM buffer was taken at various intervals, and was suspended in SM
buffer. The suspension was serially diluted with SM buffer, and
plaque formation was enumerated by the double-layer agar plate
method described above. To investigate the effect of host bacteria on
phage viability in the soil microcosm, the same experiment was
conducted with 12.75 g of ASM supplemented with 0.2 ml of PE204
and 0.2 ml of R. solanacearum SL341 suspension (108 CFU/ml). Three
replicates were included to assess phage viability in the soil microcosm.
Application of PE204 to Control Bacterial Wilt of Tomato Plant
Tomato seeds (Solanum lycopersicum) of cultivar Seogun were
obtained from Sakata Korea Corp. (Seoul, Korea), the surface of
which was sterilized in 1.0% sodium hypochlorite solution for
5 min, and was then thoroughly rinsed with sterile distilled water
until all traces of disinfectant were removed. The surface-sterilized
seeds were germinated in moistened filter paper. Uncontaminated well
sprouted seedlings were transferred to pots containing commercial
horticultural nursery media soil (Punong Co., Ltd, Korea), and were
grown in a greenhouse for 3 weeks at 20oC to 30
oC. The tomato
plants were then transferred into in a growth chamber and were
maintained at 30oC for 12 h light, and at 25
oC for 12 h dark cycles,
before R. solanacearum inoculation. Four-week-old tomato plants
were inoculated with R. solanacearum SL341.
Bacterial inoculation onto tomato plants was carried out by
applying 107 CFU bacterial suspension per gram of soil to the plant
root system of each plant, as previously described [26]. To
investigate the biological control activity of PE204, 2 ml of phage
PE204 solution in SM buffer (108 PFU/ml) was added to the
bacterial suspension of SL341, and the mixture was immediately
applied to tomato plants. When necessary, PE204 solution was
applied to tomato plants either before or after bacteria inoculation.
In this case, 2 ml of the PE204 solution in SM buffer was mixed
with the same volume of sterile water used for bacterial suspension,
and was immediately treated to tomato plants. All inoculations
included eight to ten replicates and controls without inoculation.
Plants were monitored for disease progress over a 15 day period
after inoculation, and disease was rated using the following scale: 0,
no wilting; 1, 1-25% wilting; 2, 26-50% wilting; 3, 51-75%
wilting; 4, 76-100% wilt or dead [27].
RESULTS
General Characteristics of Bacteriophage PE204
Phage PE204 was previously isolated in pepper plant
rhizospheres infected with R. solanacearum [25]. The
phage PE204 showed relatively clear and large plaques on
bacterial lawn of R. solanacearum strains (data not
shown), and this phage was selected to investigate its
potential as a biocontrol agent of bacterial wilt on tomato
plants. First, we characterized PE204 using electron
microscopy and nucleic acid analysis. Electron microscopic
observations of phage PE204 are shown in Fig. 1. Phage
particles revealed short-tailed icosahedral structures with a
head of approximately 50-60 nm in diameter and a short
tail of 16-20 nm in length, confirming them as belonging
to the family Podoviridae.
A phage genome, extracted from PE204, was resistant
to RNase and S1 nuclease treatment (data not shown),
indicating that the phage genome might be a double-
stranded DNA. In fact, several restriction endonucleases,
such as EcoRI, BamHI, and PstI digested phage DNA to
generate a unique DNA fragmentation pattern. EcoRI-, or
PstI-digested PE204 DNA was cloned and sequenced
randomly. Although we have not sequenced a whole-length
phage genome, the initial DNA sequence information
revealed that the genome of PE204 was highly similar to
that of ΦRSB1 [20]. The total determined DNA sequence
of PE204 was 21 kb, and the partial genome organization
was identical to ΦRSB1. The determined DNA sequence
spanned orf1-4, orf9, orf17, orf22-32, orf37, and orf38 of
ΦRSB1 (data not shown). The deduced amino acid
sequence of ORFs of PE204 was highly similar to those of
Fig. 1. Electron micrographs showing the icosahedral head and ashort tail structure of bacterophage PE204 (A), and a singlephage particle of PE204 at high resolution (B). White bars, 50 nm.
1616 Bae et al.
ΦRSB1 at the identity range of 97-100%. Morphology
and genome analysis indicated that PE204 is almost
identical to ΦRSB1, which was isolated from a tomato
field in Japan [20].
Persistence of Phage PE204 Activity at Various
Temperatures and pH, and in Soil Microcosm
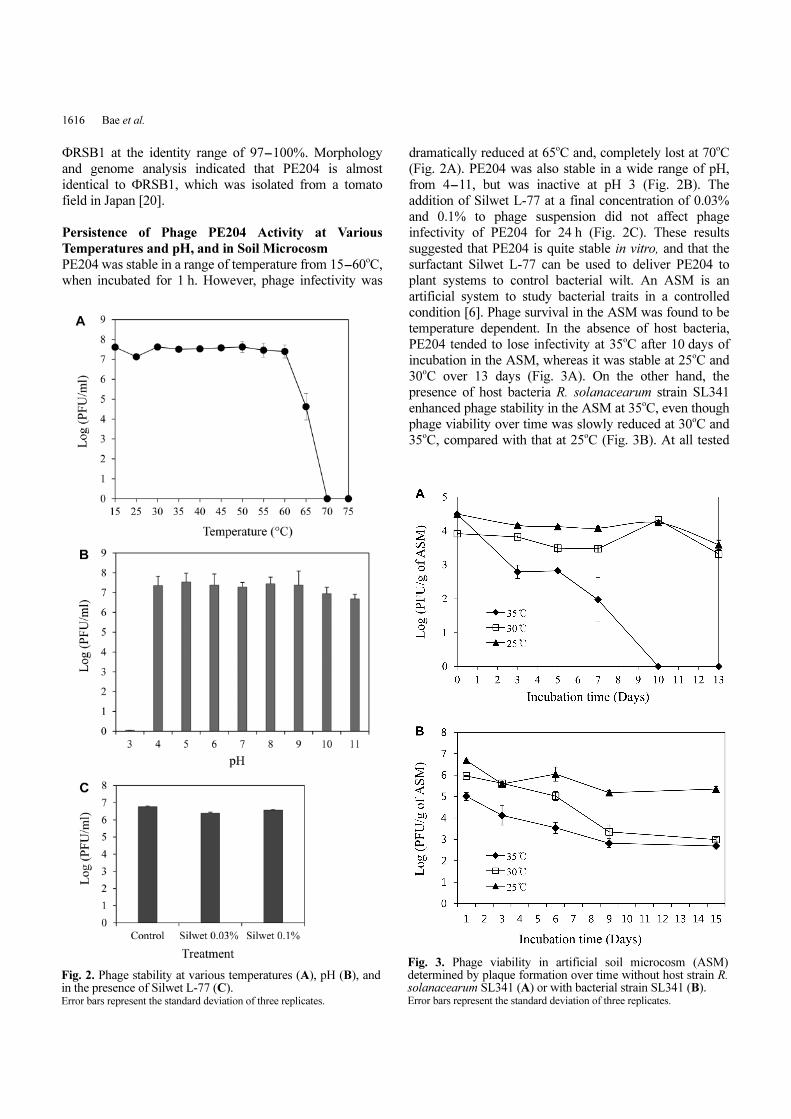
PE204 was stable in a range of temperature from 15-60oC,
when incubated for 1 h. However, phage infectivity was
dramatically reduced at 65oC and, completely lost at 70oC
(Fig. 2A). PE204 was also stable in a wide range of pH,
from 4-11, but was inactive at pH 3 (Fig. 2B). The
addition of Silwet L-77 at a final concentration of 0.03%
and 0.1% to phage suspension did not affect phage
infectivity of PE204 for 24 h (Fig. 2C). These results
suggested that PE204 is quite stable in vitro, and that the
surfactant Silwet L-77 can be used to deliver PE204 to
plant systems to control bacterial wilt. An ASM is an
artificial system to study bacterial traits in a controlled
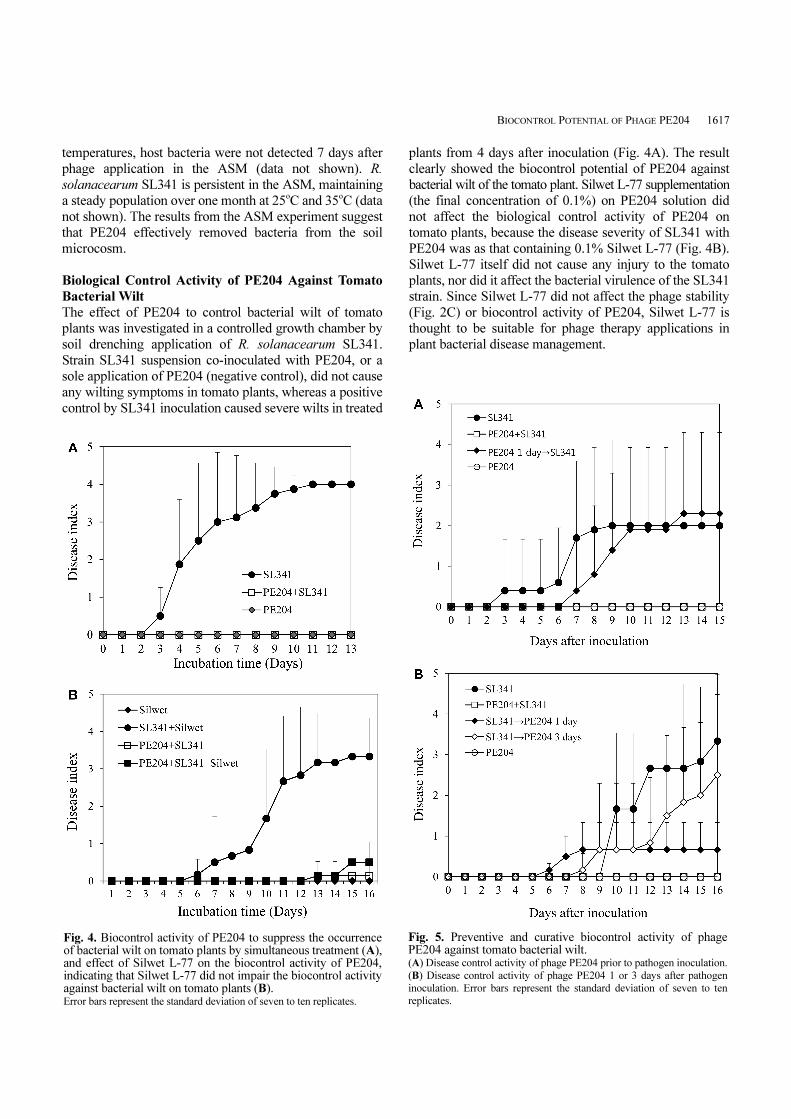
condition [6]. Phage survival in the ASM was found to be
temperature dependent. In the absence of host bacteria,
PE204 tended to lose infectivity at 35oC after 10 days of
incubation in the ASM, whereas it was stable at 25oC and
30oC over 13 days (Fig. 3A). On the other hand, the
presence of host bacteria R. solanacearum strain SL341
enhanced phage stability in the ASM at 35oC, even though
phage viability over time was slowly reduced at 30oC and
35oC, compared with that at 25oC (Fig. 3B). At all tested
Fig. 2. Phage stability at various temperatures (A), pH (B), andin the presence of Silwet L-77 (C). Error bars represent the standard deviation of three replicates.
Fig. 3. Phage viability in artificial soil microcosm (ASM)determined by plaque formation over time without host strain R.solanacearum SL341 (A) or with bacterial strain SL341 (B). Error bars represent the standard deviation of three replicates.
BIOCONTROL POTENTIAL OF PHAGE PE204 1617
temperatures, host bacteria were not detected 7 days after
phage application in the ASM (data not shown). R.
solanacearum SL341 is persistent in the ASM, maintaining
a steady population over one month at 25oC and 35oC (data
not shown). The results from the ASM experiment suggest
that PE204 effectively removed bacteria from the soil
microcosm.
Biological Control Activity of PE204 Against Tomato
Bacterial Wilt
The effect of PE204 to control bacterial wilt of tomato
plants was investigated in a controlled growth chamber by
soil drenching application of R. solanacearum SL341.
Strain SL341 suspension co-inoculated with PE204, or a
sole application of PE204 (negative control), did not cause
any wilting symptoms in tomato plants, whereas a positive
control by SL341 inoculation caused severe wilts in treated
plants from 4 days after inoculation (Fig. 4A). The result
clearly showed the biocontrol potential of PE204 against
bacterial wilt of the tomato plant. Silwet L-77 supplementation
(the final concentration of 0.1%) on PE204 solution did
not affect the biological control activity of PE204 on
tomato plants, because the disease severity of SL341 with
PE204 was as that containing 0.1% Silwet L-77 (Fig. 4B).
Silwet L-77 itself did not cause any injury to the tomato
plants, nor did it affect the bacterial virulence of the SL341
strain. Since Silwet L-77 did not affect the phage stability
(Fig. 2C) or biocontrol activity of PE204, Silwet L-77 is
thought to be suitable for phage therapy applications in
plant bacterial disease management.
Fig. 4. Biocontrol activity of PE204 to suppress the occurrenceof bacterial wilt on tomato plants by simultaneous treatment (A),and effect of Silwet L-77 on the biocontrol activity of PE204,indicating that Silwet L-77 did not impair the biocontrol activityagainst bacterial wilt on tomato plants (B). Error bars represent the standard deviation of seven to ten replicates.
Fig. 5. Preventive and curative biocontrol activity of phagePE204 against tomato bacterial wilt. (A) Disease control activity of phage PE204 prior to pathogen inoculation.
(B) Disease control activity of phage PE204 1 or 3 days after pathogen
inoculation. Error bars represent the standard deviation of seven to ten
replicates.

1618 Bae et al.
Application of PE204 to tomato plants 1 day prior to
SL341 inoculation did not effectively suppress the occurrence
of bacterial wilt (Fig. 5A), suggesting that there was no
preventive activity of PE204 against bacterial wilt on
tomato. However, PE204 application to tomato plants after
pathogen application exhibited some biocontrol activity
and a subsequent application on the following day resulted
in a much improved reduction in disease occurrence. Three
days later, an application of PE204 exhibited only slight
reduction in disease severity (Fig. 5B). The reduction of
disease severity by the delayed application of PE204
suggests a curative activity of PE204 against bacterial wilt
of tomato.
DISCUSSION
Bacteriophages infect bacterial hosts and cause either
lysogeny or lysis on the plant pathogenic bacteria. Since
management of bacterial diseases of plants is generally
limited, due to a variety of reasons, some of the lytic
phages have received attention for the control of bacterial
infection in plants [2]. The biological control of bacterial
wilt using phage PE204, infective to R. solanacearum, was
investigated in this study. Phage PE204 is a lytic phage,
which generates clear plaques on the bacterial lawn of
several strains of R. solanacearum. Analysis of phage
morphology and the genome of PE204 suggested that
PE204 is highly similar to ΦRSB1, although PE204 was
isolated in a pepper rhizosphere in Korea [25] and ΦRSB1
was isolated in a tomato field in Japan [20]. Recently,
Fujiwara et al. [9] reported the appearance of phage-
resistant R. solanacearum cells in a prolonged incubation
with ΦRSB1 in vitro. Therefore, the biocontrol potential of
ΦRSB1 for bacterial wilt has not been investigated.
However, another phage, ΦRSL1, with stable lysis activity
against R. solanacearum cells, displayed the effective
prevention of tomato wilt [9]. Therefore, biocontrol potential
using phages needs to be thoroughly explored to control
bacterial wilt of various plants.
The persistence of biocontrol agents in applied systems,
and their expression of biocontrol activity for extended
durations are a prerequisite for successful biocontrol of
plant disease [2]. PE204 was stable in a wide range of
temperature and pH conditions, suggesting its stability in
field application. In fact, PE204 exhibited strong persistence
in the ASM. The ASM provides a rather uniform soil
environment, which mimics natural soil systems [6]. Soils
are generally heterologous and complex in their compositions,
and therefore, the ASM will be a standard soil microcosm
to study phage survival in soils under various environmental
factors. The presence of host bacteria enhanced phage
stability in the microcosm system, suggesting that the
supplement of host bacteria will aid phage persistence in
soils. However, pathogenic bacteria R. solanacearum
cannot be applied with PE204 in soil, because there is a
possibility of development of phage-resistant bacteria in
soil. Use of nonpathogenic mutants of R. solanacearum
could be considered when PE204 is applied into soil,
to enhance PE204 viability in soil over an extended
period of time. Our preliminary investigation showed that
R. solanacearum mutant SL341E [36], defective in
exopolysaccharide (EPS) production, was susceptible to
PE204 (data not shown), although the mutant strain
SL341E completely lost bacterial virulence on all of
the host plants tested. An EPS-nonproducing mutant of
R. solanacearum could be extensively tested for its
enhancement of phage viability.
Addition of a surfactant, Silwet L-77, did not impair
phage PE204 stability in vitro, and its supplementation to a
PE204 suspension can hence be considered, although there
was no positive effect on biocontrol efficacy through
addition of the surfactant. Phage delivery to control plant
disease is another important issue, because appropriate
formulation and delivery will enhance phage persistence
and biocontrol activity when phages are subjected to
hostile environments. It has been reported that some
products, such as ionic copper [17], surfactants [5], and
iron chelators [29], adversely affected phages. Previous
results by Chattopadhyay et al. [5] showed that most tested
surfactants, such as a nonionic Triton X-100, an anionic
sodium dodecyl benzene sulfonate, a cationic hexadecyl
trimethylammonium bromide, and surfactin, significantly
reduced viral activity of T-2 and ΦX-174 below
concentrations of 0.1%. However, our result using Silwet
L-77 revealed that it does not affect phage stability, up to a
concentration of at least 0.1% (Fig. 2C), nor does it affect
biocontrol activity (Fig. 4B). Silwet L-77 is an organo-
silicone surfactant based on trisiloxane ethoxylate.
Because of its effect on the water/cuticle interface, this
surfactant is used at concentrations of 0.025% to 0.1% as a
component to enhance the delivery of agrochemicals,
including herbicides, insecticides, fungicides, plant growth
regulators, fertilizers, and micronutrients [4]. It has also
been reported that Silwet L-77 enhanced the entrance of
bacterial pathogen into plant leaf tissues [10, 12]. Therefore,
our result suggested that Silwet L-77 could be used as a
supplement for phage formulation as it might enhance
phage stability and phage delivery into plants.
Phage treatment on tomato plants revealed that it did not
show preventive effects, whereas phages may confer some
curative effect for bacterial wilt caused by R. solanacearum
(Fig. 5). It is not clear why pretreatment with the phage
was not effective in the control of disease. One of the
possible explanations is that PE204 was not stable in the
rhizosphere without host bacteria. Plant rhizosphere
environments are different from controlled environments
such as the ASM, and this will affect phage stability.

BIOCONTROL POTENTIAL OF PHAGE PE204 1619
Nonetheless, post-treatment of phage PE204 exhibited
biocontrol activity. Tomato plants not showing initial wilt
symptoms after phage treatment survived, without showing
further development of disease symptoms. Application of
phage to tomato plants at regular time intervals may
control bacterial wilt development and phage stability in
various conditions, suggesting that phage application
through irrigation should be considered for soils. In the
conductive soil, or at the earliest stage of bacterial
infection, the appropriate application of phages to tomato
plants may cure plants from the early stage of infection.
Fujiwara et al. [9] suggested that prolonged disease control
will be possible if phages are applied to plants at the
seedling stage. This is because phage ΦRSL1 was stable in
soil, and was recovered from roots of treated plants over 4
months after its application.
One of the limitations of practical biocontrol using phages
is their high host specificity. Application of multiple
phages with different host ranges was recommended for
practical use in biocontrol against bacterial wilt [35].
However, Fujiwara et al. [9] proposed the use of specific
phages, such as ΦRSL1 to control bacterial wilt, rather
than the use of cocktails of virulent phages. The
identification of bacteria-specific phages, and trials with
various phages solely or in combination, should be
extensively studied in order to realize the full potential of
phage therapy in bacterial wilt control. Here, we showed
that a virulent phage, PE204, could be used as a biocontrol
agent to control bacterial wilt on tomato plants. Although
PE204 does not have an extremely broad host range to be
used for practical application for biocontrol, this system
using both the ASM and in vivo growth chamber will be a
good model system to explore the biocontrol potential of
virulence phages.
Acknowledgment
This study was supported by the Dong-A University
Research Fund, in 2010.
REFERENCES
1. Askora, A., T. Kawasaki, S. Usami, M. Fujie, and T. Yamada.
2009. Host recognition and integration of filamentous phage
ΦRSM in the phytopathogen, Ralstonia solanacearum. Virology
384: 69-76.
2. Balogh, B., J. B. Jones, F. B. Iriarte, and M. T. Momol. 2010.
Phage therapy for plant disease control. Curr. Pharm.
Biotechnol. 11: 48-57.
3. Capra, M. L., A. Quiberoni, and J. A. Reinheimer. 2004.
Thermal and chemical resistance of Lactobacillus casei and
Lactobacillus paracasei bacteriophages. Lett. Appl. Microbiol.
38: 499-504.
4. Cating, R. A., M. A. Hoy, and A. J. Palmateer. 2010. Silwet L-
77 Improves the efficacy of horticultral oils for control of
boisduval scale Diaspis boisduvalii (Hemiptera: Diaspididae)
and the flat mite Tenuipalpus pacificus (Arachnida: Acari:
Tenuipalpidae) on orchids. Fla. Entomol. 93: 100-106.
5. Chattopadhyay, D., S. Chattopadhyay, W. G. Lyon, and J. T.
Wilson. 2002. Effects of surfactants on the survival and sorption
of viruses. Environ. Sci. Technol. 36: 4017-4024.
6. Ellis, R. J. 2004. Artificial soil microcosm: A tool for studying
microbial autecology under controlled conditions. J. Microbiol.
Methods. 56: 287-290.
7. Fegan, M. and P. Prior. 2005. How complex is the Ralstonia
solanacearum species complex?, pp. 449-462. In C. Allen, P.
Prior, and A. C. Hayward (eds.). Bacterial Wilt Disease and the
Ralstonia solanacearum Species Complex. APS Press, St. Paul.
8. Fujiwara, A., T. Kawasaki, S. Usami, M. Fujie, and T. Yamada.
2008. Genomic characterization of Ralstonia solanacearum
phage RSA1 and its related prophage (RSX) in strain GMI1000.
J. Bacteriol. 190: 143-156.
9. Fujiwara, A., M. Fujisawa, R. Hamasaki, T. Kawasaki, M. Fujie,
and T. Yamada. 2011. Biocontrol of Ralstonia solanacearum by
treatment with lytic bacteriophages. Appl. Environ. Microbiol.
77: 4155-4162.
10. Gottwald, T. R., J. H. Graham, and T. D. Riley. 1997. The
influence of spray adjuvants on exacerbation of citrus bacterial
spot. Plant Dis. 81: 1305-1310.
11. Grey, B. and T. R. Steck. 2001. The viable but nonculturable
state of Ralstonia solanacearum may be involved in long-term
survival and plant infection. Appl. Environ. Microbiol. 67:
3866-3872.
12. Gu, G., J. Hu, J. M. Cevallos-Cevallos, S. M. Richardson, J. A.
Bartz, and A. H. van Bruggen. 2011. Internal colonization of
Salmonella enterica serovar Typhimurium in tomato plants.
PLoS ONE 6: e27340.
13. Hayward, A. C. 1964. Characteristics of Pseudmonas solanacearum.
J. Appl. Bacteriol. 27: 265-277.
14. Hayward, A. C. 2000. Ralstonia solanacearum, pp. 32-42. In J.
Lederberg (ed.). Encyclopedia of Microbiology, Vol 4. Academic
Press, San Diego.
15. Hayward, A. C., H. M. El-Nashaar, U. Nydegger, and L. De
Lindo. 1990. Variation in nitrate metabolism in biovars of
Pseudomonas solanacearum. J. Appl. Bacteriol. 69: 269-280.
16. He, L. Y., L. Sequiera, and A. Kelman. 1983. Characteristics of
strains of Pseudomonas solacearum from China. Plant Dis. 67:
1357-1361.
17. Iriarte, F. B., B. Balogh, M. T. Momol, L. M. Smith, M.
Wilson, and J. B. Jones. 2007. Factors affecting survival of
bacteriophage on tomato leaf surfaces. Appl. Environ. Microbiol.
73: 1704-1711.
18. Jeong, Y., J. Kim, Y. Kang, S. Lee, and I. Hwang. 2007.
Genetic diversity and distribution of Korean isolates of Ralstonia
solanacearum. Plant Dis. 91: 1277-1287.
19. Kawasaki, T., S. Nagata, A. Fujiwara, H. Satsuma, M. Fujie, S.
Usami, and T. Yamada. 2007. Genomic characterization of the
filamentous integrative bacgeriophage ΦRSS1 and ΦRSM1,
which infect Ralstonia solanacearum. J. Bacteriol. 189: 5792-
5802.
20. Kawasaki, T., M. Shimizu, H. Satsuma, A. Fujiwara, M. Fujie,
S. Usami, and T. Yamada. 2009. Genomic characterization of

1620 Bae et al.
Ralstonia solanacearum phage ΦRSB1, a T7-like wide-host-
range phage. J. Bacteriol. 191: 422-427.
21. Keane, P. J., A. Kerr, and P. B. New. 1970. Crown gall of stone
fruit. II. Identification and nomenclature of Agrobacterium
isolates. Aust. J. Biol. Sci. 23: 585-595.
22. Kelman, A. 1954. The relationship of pathogenicity of
Pseudomonas solanacearum to colony appearance in a
tetrazolium medium. Phytopathology 44: 693-695.
23. Kelman, A. 1956. Survival of Pseudomonas solanacearum in
water. Phytopathology 46: 16-17.
24. Kropinski, A. M. 2006. Phage therapy. Everything old is new
again. Can. J. Infect Dis. Med. Microbiol. 17: 297-306.
25. Murugaiyan, S., J. Y. Bae, J. Wu, S. D. Lee, H. Y. Um, H. K.
Choi, et al. 2010. Characterization of filamentous bacteriophage
PE226 infecting Ralstonia solanacearum strain. J. Appl.
Microbiol. 110: 296-303.
26. Park, E. J., S. D. Lee, E. J. Chung, M. H. Lee, H. Y. Um, S.
Murugaiyan, et al. 2007. MicroTom - A model plant system to
study bacterial wilt by Ralstonia solanacearum. Plant Pathol. J.
23: 239-244.
27. Roberts, P. D., T. P. Denny, and M. A. Schell. 1988. Cloning of
the egl genes of Pseudomonas solanacearum and analysis of its
role in phytopathogenicity. J. Bacteriol. 170: 1445-1451
28. Rohwer, F. 2003. Global phage diversity. Cell 113: 141.
29. Romeo, A. M., L. Christen, E. G. Niles, and D. J. Kosman.
2001. Intracellular chelation of iron by bipyridyl inhibits DNA
virus replication: Ribonucleotide reductase maturation as a
probe of intracellular iron pools. J. Biol. Chem. 276: 24301-
24308.
30. Saddler, G. C. 2005. Management of bacterial wilt disease, pp.
121-132. In C. Allen, P. Prior, and A. C. Hayward (eds.).
Bacterial Wilt Disease and the Ralstonia solanacearum Species
Complex. APS Press, St. Paul.
31. Sambrook, J. and D. W. Russell. 2001. Molecular Cloning: A
Laboratory Manual. Cold Spring Harbor, New York.
32. Shin, J.-W. and S.-C. Yun. 2010. Elevated CO2 and temperature
effects on the incidence of four major chili pepper diseases.
Plant Pathol. J. 26: 178-184.
33. Svensson, U. and A. Christiansson. 1991. Methods for phage
monitoring. FIL-IDF Bull. 263: 29-39.
34. Tanaka, H., H. Negishi, and H. Maeda. 1990. Control of
tobacco bacterial wilt by an avirulent strain of Pseudomonas
solanacearum strain M4S and its bacteriophages. Ann. Phytopathol.
Soc. Jpn. 56: 243-246.
35. Toyoda, H., K. Kakutani, S. Ikeda, S. Goto, H. Tanaka, and
S. Ouchi. 1991. Characterization of deoxyribonucleic acid of
virulent bacteriophage and its infectivity to host bacteria,
Pseudomonas solanacearum. J. Phytopathol. 131: 11-21.
36. Um, H. Y., E. Chung, J. H. Lee, and S. W. Lee. 2011. Improved
antibiotic resistance gene cassette for marker exchange mutagenesis
in Ralstonia solanacearum and Burkholderia species. J. Microbiol.
49: 305-308.
37. Van Elsas, J. D., P. Kastelein, P. M. de Vries, and L. S. van
Overbeek. 2001. Effects of ecological factors on the survival
and physiology of Ralstonia solanacearum bv. 2 in irrigation
water. Can. J. Microbiol. 47: 842-854.
38. Yamada, T., T. Kawasaki, S. Nagata, A. Fujiwara, S. Usami,
and M. Fujie. 2007. New bacteriophages that infect the
phytopathogen Ralstonia solanacearum. Microbiology 153:
2630-2639.





























