Embed Size (px)
Citation preview

Journal of Clinical Immunology, Vol. 18, No. I, 1998
HIV-1 gp120 Accelerates Fas-Mediated Activation-InducedHuman Lamina Propria T Cell Apoptosis
MONICA BOIRIVANT,1,3 MARINA VIORA,1 LUCIANA GIORDANI,1 ALMA L. LUZZATI,1
ANNA MARIA PRONIO,2 CHIARA MONTESANI,2 and ORSOLA PUGLIESE1
Accepted: August 22, 1997
Intestinal mucosa represents an important portal of entry ofHIV and a site of virus reservoir and active replication.Recently, in HIV patients, an early depletion of intestinallamina propria T lymphocytes (LPT) has been described.HIV-1 gp120 has been demonstrated to promote apoptosis innoninfected isolated peripheral blood T cells, therefore weinvestigated whether gp120 modulates apoptosis of normalhuman intestinal lamina propria T cells. Purified T cellswere obtained by immunomagnetic negative selection fromhuman lamina propria mononuclear cells isolated fromsurgical specimens by enzymatic procedure. Cells wereincubated with or without recombinant gp120 (10 xg/ml)and cultured either in the absence of any stimulus or in thepresence of plate-bound anti-CD3 Ab (OKT3) or solubleanti-CD2 Ab (T112 + T113). Apoptosis was assessed byflow cytometric analysis after propidium iodide staining. Wedemonstrated that preincubation of normal LPT cells withHIV-1 gp120 accelerates the apoptosis observed duringCD2-pathway stimulation of LPT cells. This process ismediated by Fas/Fas ligand interaction and related to anincreased induction of Fas ligand mRNA by gp120. There-fore HIV-1 gp120 could contribute to the depletion ofnoninfected LPT cells inducing a premature cell death.
HIV-infected patients undergoing endoscopic examina-tion for gastrointestinal symptoms (2). Recently, abnor-malities in subset distribution, activation, and differenti-ation of T cells isolated from large intestine biopsies inHIV infection have been described. In particular, areduction of activated and resting CD4 T cells withnormal CD8 T cells has been observed in AIDS patientsand in HIV-infected subjects (3, 4). The early depletionof CD4+ lymphocytes involves mainly rectal laminapropria lymphocytes rather than lymphoid aggregatesand it is not closely associated with the CD4+ lympho-cyte depletion in blood (4). Intestinal T cells have aunique state of activation and differentiation which mightspecifically affect or be affected by HIV infection. Themajority of these cells shows CD45RO bright expressionand exhibits increased expression of surface markers ofcell activation (5-8). Furthermore, lamina propria T(LPT) cells show a distinctive pattern of activation andlymphokine production. LPT cells display proliferativehyporesponsiveness when stimulated via the TCR/CD3signaling pathway but relatively normal responsivenesswhen stimulated via the accessory CD2 signaling path-way compared to peripheral blood T (PBT) cells. Nev-ertheless, they produce normal or increased amounts oflymphokine under various stimulation conditions com-pared to PBT cells (9-12). In addition, in recent studieswe demonstrated that resting LPT cells exhibit an in-creased level of apoptosis, greatly enhanced by stimula-tion via the CD2 pathway but not via the TCR/CD3pathway (13). Apoptosis has been postulated as animportant factor in AIDS pathogenesis and activation-induced apoptosis occurs in noninfected T cells (14, 15).In particular, it has been demonstrated that the cross-linking of the CD4 molecule by HIV-1 gp120 triggersapoptosis in uninfected CD4+ T cells (16), possiblycontributing to CD4+ T cell depletion. Evidence thatsuch a mechanism may operate in vivo comes from the
390271-9142/98/0100-0039$15.00/0 & 1998 Plenum Publishing Corporation
KEY WORDS: Apoptosis; T lymphocytes; human; intestinal laminapropria; regional immunity; HIV; gp120.
INTRODUCTION
Intestinal mucosa represents an important portal of entryof HIV and there is evidence that the HIV burden issignificantly higher in the intestine and in other lymphoidtissues compared to peripheral blood (1). Using a sensi-tive RT-PCR technique it is, indeed, possible to detectHIV RNA in nearly all endoscopic bioptic specimens of
1immunology Department, Istituto Superiore di Sanita, Viale ReginaElena 299, 00161 Rome, Italy.
2VI Clinica Chirurgica, Universita "La Sapienza," Rome, Italy.3To whom correspondence should be addressed.

40 BOIRIVANT ET AL.
observations made in lymphoid tissue from HIV-1-infected individuals. Apoptosis in lymph nodes of HIV-1-infected individuals has been observed in noninfectedcells (17), where free gp120 was identified on the surfaceof CD4+ apoptotic T cells (18). Moreover preliminaryreports indicate an increased apoptosis in the rectallamina propria in HIV-infected subjects (19).
In the present study we investigated whether HIV-1gp120 is able to affect apoptosis of lymphocytesisolated from human intestinal lamina propria. Wedemonstrated that preincubation of normal LPT cellswith HIV-1 gp120 accelerates the apoptosis associatedwith CD2-pathway stimulation of LPT cells. Thisprocess is mediated by Fas/Fas ligand interaction andrelated to an increased induction of Fas ligand mRNAby gp120.
MATERIALS AND METHODS
Intestinal and Peripheral Mononuclear Cells
Intestinal mononuclear cells were extracted from sur-gical specimens obtained from 10 HIV-negative surgicalpatients admitted for bowel resection to the VI ClinicaChirurgica, Universita Degli Studi "La Sapienza," Rome,for malignant and nonmalignant conditions. Amongthese, six had sigmoid carcinoma, two had right coloncarcinoma, and the remaining two had diverticular dis-ease. Mucosal samples were obtained from macroscopi-cally and microscopically unaffected areas.
Lamina propria mononuclear cells (LPMC) were iso-lated from freshly resected mucosa using a previouslydescribed DTT-EDTA-collagenase method (20). Inbrief, strips of mucosa (6- to 8-g total weight) werewashed in HBSS free of calcium and magnesium(HBSS-CMF; Hyclone, Europe ltd., Cramlington, NE).They were then washed in HBSS-CMF containing 1 mMDTT (Sigma Chemical Co., St. Louis, MO) and antibi-otics (penicillin, 100 U/ml; streptomycin, 100 jug/ml;gentamicin, 50 ig/ml; and fungizone, 25 ag/ml) for 15min at room temperature. After three washings in HBSS-CMF, the mucosal strips were chopped into pieces(approximately 3 x 3 mm) and incubated four or fivetimes in HBSS-CMF containing 0.75 mM EDTA, 10mM HEPES buffer, and antibiotics for 45 min at 37°C ina humidified 5% CO2 atmosphere to remove epithelialcells. After two washes, the pieces were incubated for10-13 hr at 37°C in a humidified 5% CO2 atmosphere incomplete medium [RPMI 1640 plus 10 mM HEPESbuffer, 2 mM L-glutamine, 10% heat-inactivated FCS(Hyclone), and antibiotics] containing 25 U/ml collage-nase V (Sigma). After incubation, the supernatant was
collected and washed twice in HBSS-CMF, and LPMCwere isolated on a Percoll (Sigma) gradient.
Autologous PBMC were isolated from venous hepa-rinized blood obtained from individuals immediatelyprior to surgery. Mononuclear cells were separated fromother cellular elements by layering on a Ficoll-Paquedensity gradient. Purified T cells were obtained fromPBMC and LPMC by immunomagnetic negative selec-tion using immunomagnetic beads coated with anti-CD 14 and anti-CD20 mouse anti-human mAb (Dynal,Oslo, Norway). The resulting (unbound) T cell popula-tion contained greater than 95% CD3 + cells as assessedby flow cytometric analysis. In some experiments PBTcells were further depleted of CD45RA+ cells usingimmunomagnetic beads coated with goat anti-mouse IgGafter incubation of cells with anti-CD45RA mouse anti-human mAb (clone 2H4; Coulter Immunology, Hialeah,FL).
Cell Cultures
T cells (1 X 106) were resuspended in 1 ml ofcomplete medium and cultured for various lengths oftime in 24-well plates (Falcon) in a humidified 5%CO2 incubator. Cells were preincubated for 1 hr at37°C with gp120 (10 ug/ml) (recombinant gp120,HIV-1 strain IIIB, from the baculovirus expressionsystem; Intracel, London) or in medium alone and thencultured either in the absence of any stimulus or in thepresence of plate-bound anti-CD3 Ab (OKT3) orsoluble anti-CD2 (T112 + T113). The anti-CD3 wasobtained as murine ascites produced by the hybridomacell line OKT3 (available from the American TypeCulture Collection, Rockville, MD) and was affinity-purified using protein G columns (Pierce Immunotech-nology, Rockford, IL). Anti-CD3 mAb was used undercross-linking conditions, i.e., culturing cells in wellsprecoated with Ab (anti-CD3 at 10 ag/ml in carbonatebuffer). The anti-CD2 Ab pair (T112 and T113) werekindly provided by Dr. Ellis Reinherz (Dana FarberCancer Institute, Boston, MA) and were used insoluble form at a 1:1000 final dilution.
In some experiments, T lymphocytes were preincu-bated for 1 hr at 37°C in the presence or in the absenceof gp120, with a blocking anti-Fas IgG mAb (finalconcentration, 10 jug/ml) (clone M3; kindly provided byImmunex, Seattle, WA) which blocks apoptosis inducedby a second anti-Fas IgM mAb (clone CH-11; UBI, LakePlacid; NY) (21). Apoptosis induced by CH-11 anti-FasmAb (final concentration, 200 ng/ml) was also evaluated.
Journal of Clinical Immunology, Vol. 18, No. 1, 1998

HIV gp120 AND HUMAN LP T CELL APOPTOSIS 41
In some experiments soluble CD4 (ABT, Cambridge,MA) at a final concentration of 0.2 xg/ml was added toprevent gp120-CD4 membrane-bound ligation.
Assessment of Apoptosis
Apoptosis was measured by flow cytometry as de-scribed previously (13). Briefly, at the end of the cultureperiod, cells were washed twice in PBS and resuspendedin hypotonic fluorochrome solution [propidium iodide 50mg/ml (Sigma) in 0.1 % sodium citrate plus 0.1 % TritonX-100], kept 4-8 hr at 4°C in the dark, and analyzed byFACScan cytofluorimeter. The percentage of apoptoticcells was determined by evaluating hypodiploid nucleiafter proper gating on DNA content according to previ-ous studies (13, 22).
Apoptosis was also assessed using acridine orange/ethidium bromide fluorescence staining (23) and usingAnnexin V staining (ApoAlert Annexin V Apoptosis Kit,Clontech Laboratories, Inc., Palo Alto, CA) (24).
RT-PCR
Cellular Fas ligand mRNA was evaluated using semi-quantitative RT-PCR as described previously (23).Briefly, after RNA extraction using a guanidinium-thiocyanate method (24), 1 /ug of extracted RNA resus-pended in 14 al of water was reverse-transcribed adding16 al of a reaction buffer. The buffer consisted of 1.5 jalM-MMLV reverse transcriptase, 200 U/ml (BRL), 1 IURnasin, 40 U/ml (Promega), 6 ul 5 x RT buffer, 250 mMTris-HCl, pH 8.3, 375 mM KC1, 50 mM DTT, 15 mMMgCl2, (BRL), 3 jal acetyl-BSA, 1 mg/ml (Sigma), 1.5jal of a mix of dATP, dTTP, dCTP, and dGTP, 10 mMeach (Boeringer Mannheim), and 3 /ul of oligo(DT) (0.5mg/ml; Sigma). Reverse transcription was carried out at39°C for 45 min. For PCR, 5 jal of a 10-fold dilution ofeach cDNA (undiluted, 1:10, and 1:100) was added to 45jal of a buffer consisting of 5 jal of 10X PCR buffer [100mM Tris-HCl, pH 8.3, 500 mM KC1, 15 mM MgCl2,0.1% (w/v) gelatin], 2 /il of a mix of dATP, dTTP,dCTP, and dGTP, 10 mM each, 2.5 a1 each of the 20 mM5' primer and the 20 mM 3' primer, and 31.75 xl ofwater. Taq DNA polymerase (Boeringer; 0.25 jal) wasthen added.
The following oligonucleotide primers were used:human Fas ligand—5', 5'-CAG CTC TTC CAC CTACAG AAG GAG-3'; 3', 5'-CAG AGA GAG CTC AGATAC GTT GAC-3'; and GAPDH—5', 5'-GTC TTCACC ATG GAG AAG GTC-3'; 3', 5'-CAT GCC AGTGAG CTT CCC GTT CA-3'.
Amplification was carried out with a thermal cycler(Perkin Elmer Cetus) for 30 cycles, each cycle consistingof a denaturation step at 94°C for 30 sec, an annealingstep at 58°C for 20 sec, and an extension step at 72°C for20 sec. The 30 cycles were followed by an additionalextension step at 72°C for 7 min. Eighteen microliters ofeach PCR product was mixed with 2 jal of sampleloading buffer and electrophoresed on a 3% agarose gelcontaining 1 ag/ml ethidium bromide (Sigma).
RESULTS
LPT Cells Preincubated with gp120 Show IncreasedApoptosis When Stimulated via the CD2 SignalingPathway
We first determined the susceptibility of PBT cells andLPT cells to apoptosis after preincubation with gp120.As already observed (13) and confirmed in this study,after 18 hr of culture CD2 pathway-stimulated LPT cellscontained a significantly greater fraction of apoptoticcells compared both to unstimulated and to TCR/CD3pathway-stimulated LPT cells. Moreover, LPT cellspreincubated with soluble gp120 followed by CD2-pathway stimulation, showed a significant increase in thepercentage of apoptotic cells over the value observed inthe CD2 pathway-stimulated LPT cells (P = 0.001 bypaired Student's t test) (Table I). Preincubation withsoluble gp 120 did not affect the percentage of apoptoticcells in unstimulated LPT cells, while an increase in thepercentage of apoptotic cells was observed in TCR/CD3pathway-stimulated LPT cells in three of the six exper-iments in which it was evaluated, therefore not reachingthe statistical significance (Table I). In two experiments,the above results were confirmed using acridine orange/ethidium bromide staining in the evaluation of apoptosis(see Table I). As also shown previously (13), CD3- orCD2-pathway stimulation of PBT cells did not induceany variation in the percentage of apoptotic cells, andpreincubation of PBT cells with soluble gp120 did notaffect the percentage of PBT apoptotic cells, which inboth cases was equal to that of resting cells (data notshown). Since up to 90% of LPT cells are CD45RO+ (8),we determined apoptosis also in purified PB CD45RO+T cells to compare the effect of gp120 on phenotypicallyequivalent cell population. We found that gp120 prein-cubation followed by stimulation via TCR/CD3 or CD2pathways did not affect the level of apoptosis of CD45R0+ PB T cells (data not shown). Since the effect ofgp120 on CD3 stimulated LPTC was not observed in allthe experiments, we focused on CD2 stimulated LPTC to
Journal of Clinical Immunology, Vol. 18, No. 1, 1998

42 BOIRIVANT ET AL.
characterize better the features of the gpl20-inducedapoptosis in LPT cells.
Time-Course Study of gp120-Induced Apoptosis of LPTCells
As shown in Fig. 1, LPT cells preincubated withgp120, followed by stimulation via the CD2 signalingpathway, showed a significant increase (P - 0.02 by thepaired Student's t test) in the percentage of apoptoticcells after 18 hr of culture compared to CD2-stimulatedcells. An increase in the mean level of apoptosis was stillobservable at 24 and 48 hr of cultures, although it wasnot present in each experiment performed. Therefore,from these data it appears that gp120 mainly acceleratesthe time course of CD2-induced apoptosis. This featureis better illustrated by evaluating apoptosis using annexin
Fig. 1. Percentage of apoptotic LPT cells at different time points afteranti-CD2 stimulation. The relative proportion of apoptotic cells wasquantitated by propidium iodide staining followed by FACS analysis.Cells were preincubated for 1 hr with (O) medium alone or (•) gp120.Data represent the mean ± SD of three experiments. *P = 0.002 LPTcells preincubated with gp120 followed by stimulation via CD2 vs.CD2 stimulation only.
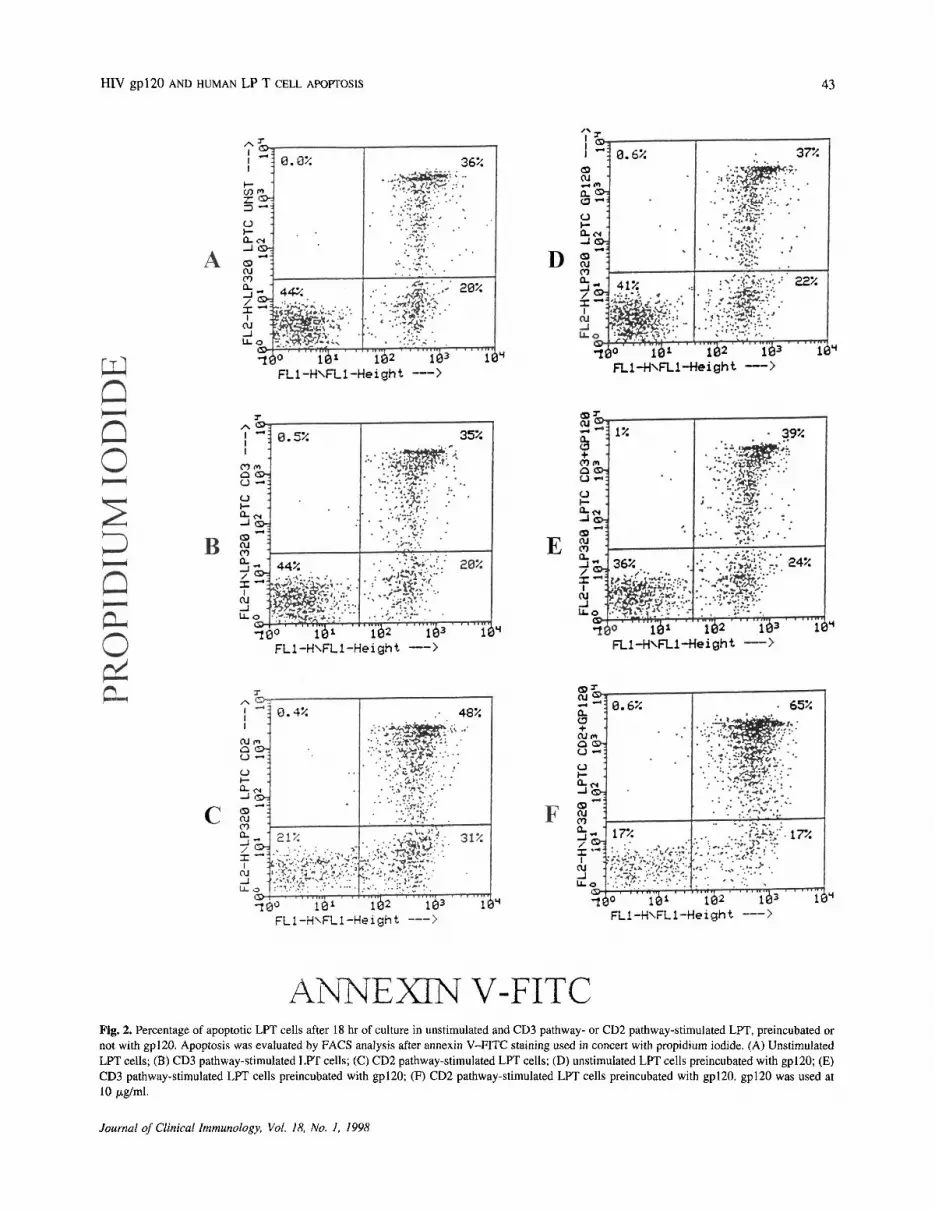
V/propidium iodide staining. By this method it is possi-ble to discriminate between the early stage of apoptosis,in which only membrane alterations consisting in thetranslocation of the membrane phospholipid phosphati-dylserine from the inner face of the plasma membrane tothe cell surface are present, without functional impair-ment of membrane permeability and later stages ofapoptosis, when impairment of membrane permeabilitybecomes apparent. The first condition is marked bymembrane green fluorescence staining due to the ligationof FITC-annexin V to the phosphatidylserine; the secondcondition is characterized by double-fluorescent cells(green on the surface and red inside the cell) due toimpaired membrane permeability, which allows the pro-pidium iodide to penetrate the cells. As shown in Fig. 2,after 18 hr of culture an increase in the percentage ofdouble-positive cells is present in the gp120-preincu-bated cells followed by CD2 stimulation compared to theCD2-stimulated LPT cells, thus indicating a later stage ofapoptosis in these cultures.
Modulation of CD2 Pathway-Induced Apoptosis byPreincubation with gp120 Is Abrogated by FasBlockade
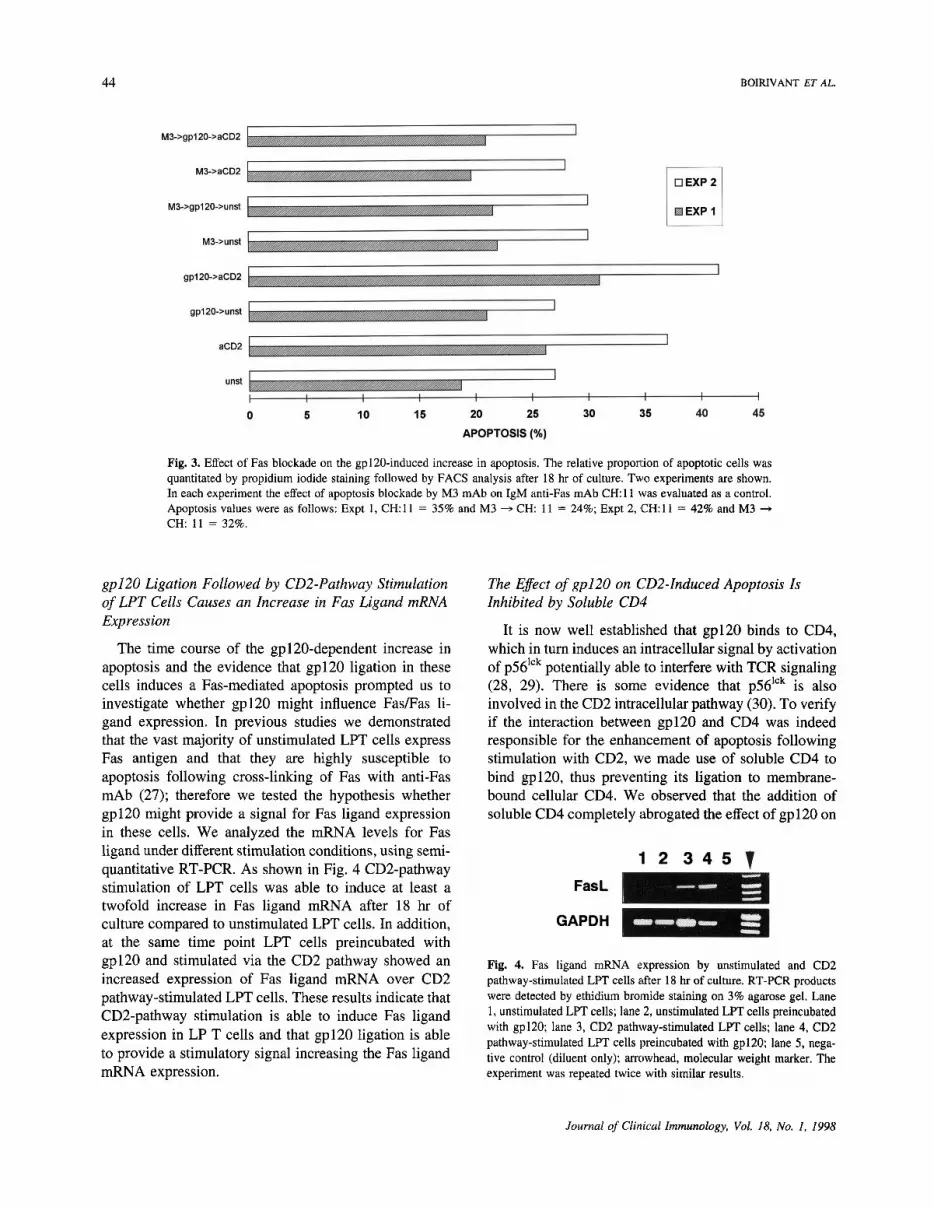
In previous studies (13), we observed that in humanLPT cells, CD2-pathway stimulation-mediated apoptosiswas Fas mediated. Therefore, we determined whether theincreased apoptosis induced by gp120 was Fas mediated.To this purpose, we used a blocking IgGl anti-Fas mAb(M3 mAb; kindly provided by Immunex, Seattle, WA)able to block apoptosis induced by a second IgM anti-FasmAb (clone CH-11). As shown in Fig. 3 preincubation ofLPT cells with M3 mAb reduced apoptosis to levels ofunstimulated cells, both in CD2 pathway-stimulated Tcells and in gp120-preincubated LPT cells. Thus, gp120ligation in these cells induces apoptotic death in aFas-dependent manner. No effect was observed when M3mAb was used on unstimulated T cells.
Journal of Clinical Immunology, Vol. 18, No. 1, 1998
Table I. Effect of gp120 on Apoptosis of LPT Cellsa
ExptNo.
1234567
Unstim
1120(15)271725 (13)
77
Apoptosis (%)
gp120 — » unstim
1623 (18)271225 (9)
88
Anti-CD3
1524(16)
ND1428 (19)76
gp120 -» anti-CD3
1935 (20)
ND1427 (18)179
Anti-CD2
2242 (27)372145 (27)1513
gp120 -» anti-CD2
3054 (48)413360 (46)2018
aApoptosis was evaluated as the percentage of hyplodiploic nuclei after PI staining and by acridine orange- ethidium bromide staining. Values inparentheses represent the percentage of apoptotic cells as assessed by acridine orange- ethidium bromide staining. P = 0.001, gp120 — > anti-CD2vs anti-CD2; P = 0.08, gp120 -» anti-CD3 vs anti-CD3 (paired Student's t test). ND, not done.
HIV gp120 AND HUMAN LP T CELL APOPTOSIS
Fig. 2. Percentage of apoptotic LPT cells after 18 hr of culture in unstimulated and CD3 pathway- or CD2 pathway-stimulated LPT, preincubated ornot with gp120. Apoptosis was evaluated by FACS analysis after annexin V-FITC staining used in concert with propidium iodide. (A) UnstimulatedLPT cells; (B) CD3 pathway-stimulated LPT cells; (C) CD2 pathway-stimulated LPT cells; (D) unstimulated LPT cells preincubated with gp120; (E)CD3 pathway-stimulated LPT cells preincubated with gp120; (F) CD2 pathway-stimulated LPT cells preincubated with gp120. gp120 was used at10 ug/ml.
Journal of Clinical Immunology, Vol. 18, No. 1, 1998
43
44 BOIRIVANT ET AL.
Fig. 3. Effect of Fas blockade on the gp120-induced increase in apoptosis. The relative proportion of apoptotic cells wasquantitated by propidium iodide staining followed by FACS analysis after 18 hr of culture. Two experiments are shown.In each experiment the effect of apoptosis blockade by M3 mAb on IgM anti-Fas mAb CH: 11 was evaluated as a control.Apoptosis values were as follows: Expt 1, CH:11 = 35% and M3 -» CH: 11 = 24%; Expt 2, CH:11 = 42% and M3 -»CH: 11 = 32%.
gp120 Ligation Followed by CD2-Pathway Stimulationof LPT Cells Causes an Increase in Fas Ligand mRNAExpression
The time course of the gp120-dependent increase inapoptosis and the evidence that gp120 ligation in thesecells induces a Fas-mediated apoptosis prompted us toinvestigate whether gp120 might influence Fas/Fas li-gand expression. In previous studies we demonstratedthat the vast majority of unstimulated LPT cells expressFas antigen and that they are highly susceptible toapoptosis following cross-linking of Fas with anti-FasmAb (27); therefore we tested the hypothesis whethergp120 might provide a signal for Fas ligand expressionin these cells. We analyzed the mRNA levels for Fasligand under different stimulation conditions, using semi-quantitative RT-PCR. As shown in Fig. 4 CD2-pathwaystimulation of LPT cells was able to induce at least atwofold increase in Fas ligand mRNA after 18 hr ofculture compared to unstimulated LPT cells. In addition,at the same time point LPT cells preincubated withgp120 and stimulated via the CD2 pathway showed anincreased expression of Fas ligand mRNA over CD2pathway-stimulated LPT cells. These results indicate thatCD2-pathway stimulation is able to induce Fas ligandexpression in LP T cells and that gp120 ligation is ableto provide a stimulatory signal increasing the Fas ligandmRNA expression.
The Effect of gp120 on CD2-Induced Apoptosis IsInhibited by Soluble CD4
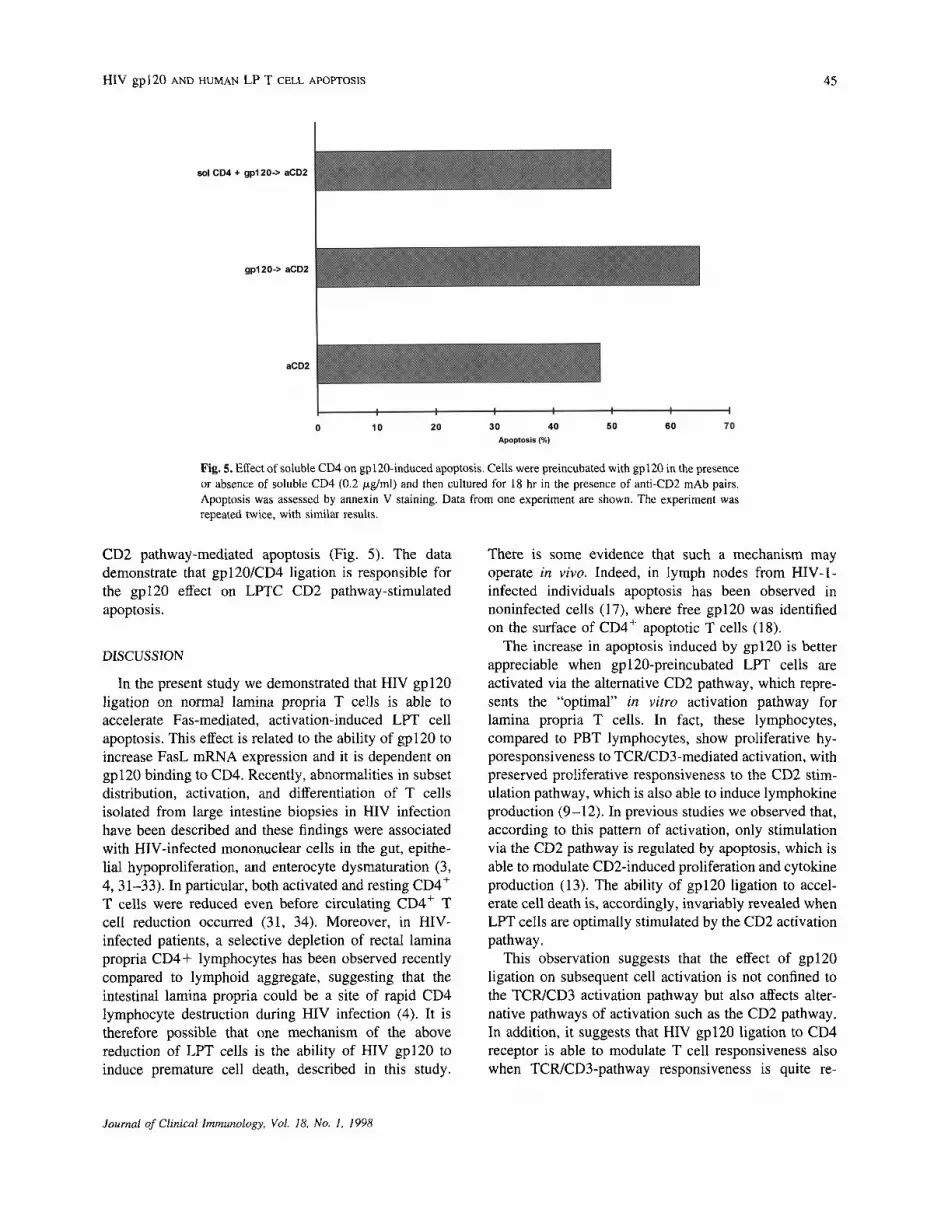
It is now well established that gp120 binds to CD4,which in turn induces an intracellular signal by activationof p56lck potentially able to interfere with TCR signaling(28, 29). There is some evidence that p56lck is alsoinvolved in the CD2 intracellular pathway (30). To verifyif the interaction between gp120 and CD4 was indeedresponsible for the enhancement of apoptosis followingstimulation with CD2, we made use of soluble CD4 tobind gp120, thus preventing its ligation to membrane-bound cellular CD4. We observed that the addition ofsoluble CD4 completely abrogated the effect of gp120 on
Fig. 4. Fas ligand mRNA expression by unstimulated and CD2pathway-stimulated LPT cells after 18 hr of culture. RT-PCR productswere detected by ethidium bromide staining on 3% agarose gel. Lane1, unstimulated LPT cells; lane 2, unstimulated LPT cells preincubatedwith gp120; lane 3, CD2 pathway-stimulated LPT cells; lane 4, CD2pathway-stimulated LPT cells preincubated with gp120; lane 5, nega-tive control (diluent only); arrowhead, molecular weight marker. Theexperiment was repeated twice with similar results.
Journal of Clinical Immunology, Vol. 18, No. 1, 1998
HIV gp120 AND HUMAN LP T CELL APOPTOSIS 45
Fig. 5. Effect of soluble CD4 on gp120-induced apoptosis. Cells were preincubated with gp120 in the presenceor absence of soluble CD4 (0.2 ug/ml) and then cultured for 18 hr in the presence of anti-CD2 mAb pairs.Apoptosis was assessed by annexin V staining. Data from one experiment are shown. The experiment wasrepeated twice, with similar results.
CD2 pathway-mediated apoptosis (Fig. 5). The datademonstrate that gp120/CD4 ligation is responsible forthe gpl20 effect on LPTC CD2 pathway-stimulatedapoptosis.
DISCUSSION
In the present study we demonstrated that HIV gp120ligation on normal lamina propria T cells is able toaccelerate Fas-mediated, activation-induced LPT cellapoptosis. This effect is related to the ability of gp120 toincrease FasL mRNA expression and it is dependent ongp120 binding to CD4. Recently, abnormalities in subsetdistribution, activation, and differentiation of T cellsisolated from large intestine biopsies in HIV infectionhave been described and these findings were associatedwith HIV-infected mononuclear cells in the gut, epithe-lial hypoproliferation, and enterocyte dysmaturation (3,4, 31-33). In particular, both activated and resting CD4+
T cells were reduced even before circulating CD4+ Tcell reduction occurred (31, 34). Moreover, in HIV-infected patients, a selective depletion of rectal laminapropria CD4+ lymphocytes has been observed recentlycompared to lymphoid aggregate, suggesting that theintestinal lamina propria could be a site of rapid CD4lymphocyte destruction during HIV infection (4). It istherefore possible that one mechanism of the abovereduction of LPT cells is the ability of HIV gp120 toinduce premature cell death, described in this study.
There is some evidence that such a mechanism mayoperate in vivo. Indeed, in lymph nodes from HIV-1-infected individuals apoptosis has been observed innoninfected cells (17), where free gp120 was identifiedon the surface of CD4+ apoptotic T cells (18).
The increase in apoptosis induced by gp120 is betterappreciable when gp120-preincubated LPT cells areactivated via the alternative CD2 pathway, which repre-sents the "optimal" in vitro activation pathway forlamina propria T cells. In fact, these lymphocytes,compared to PBT lymphocytes, show proliferative hy-poresponsiveness to TCR/CD3-mediated activation, withpreserved proliferative responsiveness to the CD2 stim-ulation pathway, which is also able to induce lymphokineproduction (9-12). In previous studies we observed that,according to this pattern of activation, only stimulationvia the CD2 pathway is regulated by apoptosis, which isable to modulate CD2-induced proliferation and cytokineproduction (13). The ability of gp120 ligation to accel-erate cell death is, accordingly, invariably revealed whenLPT cells are optimally stimulated by the CD2 activationpathway.
This observation suggests that the effect of gp120ligation on subsequent cell activation is not confined tothe TCR/CD3 activation pathway but also affects alter-native pathways of activation such as the CD2 pathway.In addition, it suggests that HIV gp120 ligation to CD4receptor is able to modulate T cell responsiveness alsowhen TCR/CD3-pathway responsiveness is quite re-
Journal of Clinical Immunology, Vol. 18, No. 1, 1998

46 BOIRIVANT ET AL.
duced as in LPT cells. This effect is probably due to theability of CD4 intracellular signaling to interfere with theCD2 intracellular pathway. As already reported, gp120ligation of CD4 induces p56lck activation which is able tointerfere with TCR signaling (28, 29). Similar mecha-nisms may also be operating in modulating the CD2pathway. Indeed an involvement of p56lck has also beenobserved in the CD2 intracellular pathway (30). Ourobservation that preincubation with soluble CD4 pre-vents the effect of gp120 on CD2-activated LPT cellspoints in that direction.
Further insight into the possible stimulatory activity ofgp120 ligation comes from observations made in thisstudy that gp120-preincubated T cells express more Fasligand mRNA, after CD2-pathway stimulation, than Tcells stimulated via the CD2 pathway only. A similarfinding has been reported with regard to the ability ofHIV-1 tat and gp120 or CD4 stimulation to accelerateCD95-mediated, activation-induced apoptosis in PBMCby induction of Fas ligand mRNA (35). More recently,direct evidence that gp120 can induce Fas-L expression,analyzed by immunofluorescence, Western blot, ELISA,and bioassay, in T cell clones has been provided (36).This underscores that CD4 engagement is able to coop-erate in the induction of an activation signal for Fasligand mRNA production, therefore contributing to in-crease activation-induced apoptosis in susceptible cellssuch as LPT cells. Indeed, recently it has been demon-strated that pp56lck affects TCR-induced cell death byregulating Fas ligand expression (37).
The above data, along with the evidence provided inthe present work that interaction between gp120 andCD4 is necessary to increase the apoptosis of CD2pathway-stimulated LPT cells, lead to the suggestion thatthe signal provided by gp120-CD4 ligation, whichinvolves pp56lck, might indeed contribute to the in-creased Fas ligand expression observable in CD2 path-way-stimulated LPT cells preincubated with gp120.Functional consequences of the premature cell death atthe mucosal site might involve mainly lymphokine mod-ulation since it is well-known that one of the majorfeatures of LPT cells is their ability to produce lympho-kines to provide help for Ig synthesis (11, 12). Studiesare in progress to examine this issue.
ACKNOWLEDGMENTS
We acknowledge the kind gift of anti-T112 + anti-T113 from Dr. Ellis Reinherz (Dana Farber Institute,Boston, MA). The authors also thank Dr. Maria SaveriaGilardini Montani (Universita La Tuscia, Viterbo, Italy)for kindly providing Fas ligand primers and Immunex
(Seattle, WA) for kindly providing the Fas M3 mAb.This work was supported by Ministry of Health ISSGrant 940-W.
REFERENCES
1. Schneider T, Ullrich R, Zeitz M: Immunopathology of humanimmunodeficiency virus infection in the gastrointestinal tract.Spinger Semin Immunopathol 18:515-533, 1997
2. Kotler DP, Reka S, Borchic A, Cronin WJ: Detection, localizationand quantitation of HIV-associated antigens in intestinal biopsiesfrom patients with HIV. Am J Pathol 139:823-830, 1991
3. Schneider T, Ullrich R, Bergs C, Schmidt W, Riechen EO, ZeitzM. The Berlin Diarrhoea/Wasting Syndrome Study Group: Abnor-malities in subset distribution, activation and differentiation of Tcells isolated from large intestine biopsies in HIV infection. ClinExp Immunol 95:430-435, 1994
4. Clayton F, Snow G, Reka S, Kotler DP: Selective depletion ofrectal lamina propria rather than lymphoid aggregate CD4 lym-phocytes in HIV infection. Clin Exp Immunol 107:288-292, 1997
5. Pallone F, Fais S, Squarcia O, Biancone L, Pozzilli P, Boirivant M.Activation of peripheral and intestinal lamina propria lymphocytesin Crohn's disease. "In vivo" state of activation and "in vitro"response to stimulation as defined by the expression of earlyactivation antigens. Gut 28:745-753, 1987
6. Peters MG, Secrist H, Anders KR, Nash GS, Rich SR,MacDermott RP: Normal human intestinal lymphocytes. Increasedactivation compared with the peripheral blood. J Clin Invest83:1827-1833, 1989
7. De Maria R, Fais S, Silvestri M, Frati L, Pallone F, Santoni A,Testi R: Continuous in vivo activation and transient hyporespon-siveness to TCR/CD3 triggering of human gut lamina proprialymphocytes. Eur J Immunol 23:3104-3108, 1993
8. Schieferdecher HL, Ullrich R, Hierselandh H, Zeitz M: T celldifferentiation antigens on lymphocytes in the human intestinallamina propria. J Immunol 149:2816-2822, 1992
9. Pirzer UC, Schurmann G, Post S, Betzler M, Meuer SC: Differ-ential responsiveness to CD3-Ti vs. CD2-dependent activation ofhuman intestinal T lymphocytes. Eur J Immunol 20:2339-2342,1990
10. Qiao L, Schurmann G, Betzler M, Meuer SC: Activation andsignaling status of human lamina propria T lymphocytes. Gastro-enterology. 101:1529-2389, 1991
11. Targan SR, Deem RL, Liu M, Wang S, Nel A: Definition of laminapropria T cell responsive state. Enhanced cytokine responsivenessof T cells stimulated through the CD2-pathway. J Immunol154:664-675, 1995
12. Boirivant M, Fuss I, Fiocchi C, Klein JS, Strong SA, Strober W:Hypo-proliferative human lamina propria T cells retain the capac-ity to secrete lymphokines when stimulated via the CD2/CD28accessory signaling pathways. Proc Assoc Am Phys 108:55-67,1996
13. Boirivant M, Pica R, De Maria R, Testi R, Pallone F, Strober W:Stimulated human lamina propria T cells manifest enhancedFas-mediated apoptosis. J Clin Invest 98:2616-2622, 1996
14. Ameisen JC: Programmed cell death (apoptosis) and AIDS patho-genesis. AIDS Res Hum Retrovir 10:S3-S5, 1994
15. Pantaleo G, Fauci AS: Apoptosis in HIV infection. Nature Med1:118-120, 1995
16. Banda NK, Bernier J, Cuhara DK, Kurrle R, Haigwood N, SekalyRP, Finkel TH: Crosslinking CD4 by human immunodeficiency
Journal of Clinical Immunology, Vol. 18, No. 1, 1998

HIV gp120 AND HUMAN LP T CELL APOPTOSIS 47
virus gp120 primes T cells for activation-induced apoptosis. J ExpMed 176:1099-1106, 1992
17. Finkel TH, Tudor-Williams G, Banda NK, et al: Apoptosis occurspredominantly on bystander cells and not in productively infectedcells of HIV- and SIV-infected lymph nodes. Nature Med 1:129-134, 1995
18. Sunila I, Vaccarezza M, Pantaleo G, Fauci AS, Orenstein JM:gp120 is present on the plasma membrane of apoptotic CD4 cellsprepared from lymph nodes of HIV-1-infected individuals: Animmunoelectron microscopic study. AIDS 11:27-32, 1997
19. Reka S, Kotler DP: Apoptosis in rectal lamina propria: Variationwith disease stage. J Inv Med 43S:290, 1995 (abstr)
20. Bull DM, Bookmann MA: Isolation and functional characteriza-tion of human mucosal lymphoid cells. J Clin Invest 59:966-974,1977
21. Alderson MR, Tough TW, Braddy S, et al.: Regulation ofapoptosis and T cell activation by Fas-specific mAB. Int Immunol6:1799-1806, 1994
22. Zamai L, Falcieri E, Zauli G, Cataldi A, Vitale M: Optimaldetection of apoptosis by flow cytometry depends on cell morphol-ogy. Cytometry 14:891-897, 1993
23. Duke RC, Cohen JJ: Morphological and biochemical assays ofapoptosis. In Current Protocols in Immunology, JE Coligan, AMKruisbeek, DH Margulies, EM Shevach, W Strober (eds). NewYork, Greene Associates and J. Wiley & Sons, 1992, pp 1-3
24. Vermes I, Haanen C, Steffens-Nakken H, Reutelingspergwe C: Anovel assay for apoptosis flow cytometric detection of phosphati-dylserine expression on early apoptotic cells using fluoresceinlabelled Annexin V. J Immunol Methods 184:39-51, 1995
25. Luzzati AL, Boirivant M, Giacomini E, Giordani L, Di ModugnoF, Chersi A: Interferon-gamma (IFN-y) can counteract the in vitroinhibitory effect of an HIV p24 immunosuppressive heptapeptide.Clin Exp Immunol 105:403-408, 1996
26. Chomczynski P, Sacchi N: Single-step method of RNA isolationby acid guanidium thiocyanate-phenol-chloroform extraction. AnalBiochem 162:156-159, 1987
27. De Maria R, Boirivant M, Cifone MG, Roncaioli P, Hahne M,Tschopp J, Pallone F, Santoni A, Testi R: Functional expression ofFas and Fas ligand on human gut lamina propria T lymphocytes: A
potential role for the acidic sphingomyelinase pathway in normalimmunoregulation. J Clin Invest 97:316-322, 1996
28. Goldman F, Jensen WA, Johnson GL, Heasley L, Cambier JC:gp120 ligation of CD4 induces p56lck activation and TCR desen-sitization independent of TCR tyrosine phosphorylation. J Immu-nol 153:2905-2917, 1994
29. Hubert P, Bismuth G, Korner M, Debre P; HIV-1 glycoproteingp120 disrupts CD4-p56lck/CD3-T cell receptor interactions andinhibits CD3 signaling. Eur J Immunol 25:1417-1425, 1995
30. Danielian S, Fagard R, Alcover A, Acuto O, Fischer S: Thelymphocyte specific protein tyrosine kinase p56lck is hyperphos-phorilated on serine and tyrosine residues within minutes afteractivation via T cell receptor or CD2. Eur J Immunol 19:2183-2189, 1989
31. Lim SG, Condez A, Lee CA, Johonson MA, Elia C, Poulter LW:Loss of mucosal CD4 lymphocytes is an early feature of HIVinfection. Clin Exp Immunol 92:448-454, 1993
32. Ullrich R, Zeitz M, Heise W, Lage M, Hoffken G, Rieken EO:Small intestinal structure and function in patients infected withhuman immunodeficiency virus (HIV): Evidence for HIV-inducedenteropathy. Ann Intern Med 111:15-21, 1989
33. Ullrich R, Zeitz M, Heise W, et al: Mucosal atrophy is associatedwith loss of activated T cells in the duodenal mucosa of humanimmunodeficiency virus (HIV)-infected patients. Digestion 46(Suppl 2):302-307, 1990
34. Schneider T, Ullrich R, Zeitz M: The immunological aspect ofhuman immunodeficiency virus infection in the gastrointestinaltract. Semin Gastroint Dis 7:19-29, 1996
35. Westendorp MO, Frank R, Ochsenbauer C, et al: Sensitization ofT cells to CD95-mediated apoptosis by HIV-1 Tat and gp120.Nature 375:497-500, 1995
36. Accornero P, Radrizzani M, Delia D, Gerosa F, Kurrle R, ColomboMP: Differential susceptibility to HIV gp120 sensitized apoptosisin CD4+ T cell clones with different T-helper phenotypes: Role ofCD95/CD95L interaction. Blood 89:558-569, 1997
37. Gonzalez-Garcia A, R.-Borlado L, Leonardo E, Merida I, MartinezAC, Carrera AC: Lck is necessary and sufficient for Fas-Ligandexpression and apoptotic cell death in mature cycling T cells.J Immunol 158:4104-4112, 1997
Journal of Clinical Immunology, Vol. 18, No. 1, 1998





























