Embed Size (px)
Citation preview

Volume 1 1 Number 18 1983 Nucleic Acids Research
Isolation of cDNA clones for basal lamina components: type IV procollagen
Markku Kurkinen, Denise P.Barlow, David M.Helfman*, Jeffrey G.Williams and BrigidL.M.Hogan
Imperial Cancer Research Fund, Mill Hill Laboratories, Burtonhole Lane, London NW7 lAD, UK,and *Cold Spring Harbor Laboratory, P.O. Box 100, Cold Spring Harbor, NY 11724, USA
Received 28 July 1983; Accepted 6 September 1983
ABSTRACT
We have isolated cDNA clones for mouse type IV procollagen from alibrary constructed from total poly A+RNA of 13.5 day mouse embryoparietal endoderm (PE) cells. In Northern analysis these clones hybridiseto a 6.8 kb RNA which is abundant in embryonic PE cells and indifferentiated F9 teratocarcinoma cells. Hybrid selection and in vitrotranslation of the cDNA specific mRNA produced a single polypeptide ofMr = 165 000. This polypeptide was specifically immunoprecipitated withmouse type IV procollagen antisera and comigrated on SDS-gel electro-phoresis with one of the two in vitro synthesised chains of type IVprocollagen. Undifferentiated F9 teratocarcinoma cells can be induced byretinoic acid and dibutyrylcAMP to differentiate in vitro into endoderm-like cells which resemble mouse PE cells in synthesising large amountsof basement membrane proteins, including type IV procollagen. Here weshow, using one of the cDNA clones as a probe for type IV procollagen,that an increase in cellular concentration of type IV procollagen mRNAoccurs within 24 to 48 hours of induction, reaching a constant high levelby 72 hours.
NTRODUCT I ON
Parietal endoderm (PE) cells of the mouse embryo are specialisedfor producing a thick basal lamina known as Reichert's membrane. They
synthesise large amounts of several well characterised, high molecular
weight proteins, including type IV procollagen, laminin, entactin and
heparan sulphate proteoglycan (1-3). PE cells therefore provide a model
system in which to study the developmental regulation of the genes for
basal lamina structural components, as well as other proteins involved in
their processing, assembly, and turnover. The differentiation of PE cells
in vivo can be mimicked in vitro by treating undifferentiated F9
teratocarcinoma cells with retinoic acid and dibutyryl cAMP (4,5). This
results in a large increase in the synthesis of type IV procollagen,
laminin and entactin by F9 cells (5,6). For example, PE cells devote
approximately 15?6 of their total protein synthesis to type IV procollagen,
©) I R L Press Limited, Oxford, England. 6199

Nucleic Acids Research
laminin and entactin combined. The corresponding values for differentiated
and undifferentiated F9 cells are 2% and 0.15%6, respectively. We have
previously reported the in vitro synthesis of type IV procollagen (7) and
laminin and entactin (8) polypeptides. The difference in the rate of
synthesis of these basal lamina components between the three cell types
(PE, differentiated F9 and undifferentiated F9 cells) was shown to be due
to a difference in the level of translatable mRNA (7,8 and unpublishedresults).
In order to obtain cDNA clones for basal lamina components, as well
as other PE cell specific proteins, we constructed cDNA libraries starting
from total poly(A)+RNA of PE mouse embryo cells. "Dot blots" of plasmid
DNA isolated from 1300 clones were screened by their differential
hybridisation to cDNA-probes prepared from RNA of mouse embryo PE and
undifferentiated F9 cells. Based on this analysis we identified 80 PE
specific cDNA clones. One of these (pPE 1180) contained a 340 bp sequence
derived from type IV procollagen mRNA and was used to isolate two other
plasmids from the PE cDNA libraries which contained a 1400 bp (pPE 76)
and a 1500 bp (pPE 41) sequence.
MATERIALS AND METHODS
Vanadyl-ribonucleoside complex (9) was obtained from Bethesda
Research Laboratories, AMV reverse transcriptase was from Life Sciences
Inc. E.coli DNA polymerase I was from Boehringer Mannheim. The source
of other materials used here has been described (7,8,10).Cells
Mouse F9 teratocarcinoma cells were maintained and subcultured
as described (7). Undifferentiated cells, termed F9 (-), were induced to
differentiate into parietal eridoderm-like cells, termed F9 (+), by
treatment for 5 days with 5 x 10-8 M retinoic acid, 104 dibutyryl cyclic-4AMP and 10 M isobutyl methylxanthine (4). Parietal endoderm (PE) cells
attached to Reichert's membrane were dissected from 13.5 day C3H/He mouse
embryos and used immediately for RNA extraction.
RNA isolation
Total cellular RNA from F9 cell cultures and embryonic PE cells was
isolated using guanidinium isothiocyanate extraction and caesium chloride
centrifugation (11) as described (7). Poly(A)+RNA was prepared as detailed
(12) except that PE poly(A)+RNA was prepared by three cycles of oligo (dT)
cellulose chromatography. RNA concentration was estimated by spectro-
6200

Nucleic Acids Research
photometry (1 A260 unit = 40 pg/mi).
Construction of a parietal endoderm cDNA library
Starting from total poly(A)+RNA of parietal endoderm cells a cDNA
library was constructed by the double-linker method exactly as described
(10) except that cDNA was inserted in both the vectors pUC8 and pUC9 (13).
In the former case the double stranded cDNA has an EcoRl linker at its 5'
end and a Sall linker at its 3' end. In the latter case the position of
these linkers is reversed. Both types of cDNA libraries were established
in the E.coli strain DH-1 and colonies were stored on nitrocellulose
filters placed between sterile 3mm Whatman paper at -700C,Screening by differential hybridisation
Plasmid DNA was isolated using a rapid, small scale boiling method
(14) from 1300 colonies picked from the parietal endoderm cDNA library
constructed with the vector pUC9. After denaturation by boiling for 5 min
in 0.3 NaOH, then neutralisation with an equimolar amount of HCl, plasmid
DNA was dotted in duplicates onto two sets of nitrocellulose filters.
Filters were air dried, soaked in 2 x SSC (1 x SSC is 0.15 M NaCl, 0.015 M
trisodium citrate) blotted dry and kept at 800C for 2 hours. The duplicate
filters were prehybridised overnight at 420C with 50% formamide, 5 x SSPE
(0.9 M NaCl, 50 mM NaH2PO4, 5 mM EDTA), 5 x Denhardt's solution (0.2% BSA,
0.2% PVP, 0.2% FICOLL) 0.1 SDS and 0.1 mg/ml denatured salmon sperm DNA,
and then hybridised for 3 days at 420C with cDNA-probes in the above
buffer (1 x 104 Cerenkov cpm/10 p1/cm2). One set of filters was hybridised
with cDNA prepared from the poly(A)+RNA of PE mouse embryo cells and the
other set was hybridised with cDNA prepared from the total poly(A) RNA of
undifferentiated F9 cells. Filters were washed at room temperature first
in 2 x SSC and 0.l15 SDS and then in 0.1 x SSC and 0.l1? SDS and auto-
radiographed for 5 days at -70°C with Dupont Cronex Lightning plus X-ray
intensifying screens.
Northern analysis
The cDNA clones displaying a PE specific hybridisation signal in the
DNA dot blots were further analysed by Northern hybridisation. TotalRNA was electrophoresed on formaldehyde 0.7% agarose gels. After electro-
phoresis gels were rinsed twice in distilled water, treated with 50 mM
NaOH, 10 mM NaCl for 40 min and then neutralised with 0.1 M Tris pH 7.5
for 30 min. Gels were then stained for 10 min with 0.5 pg/ml ethidium
bromide in 0.1 M Tris pH 7.5 and photographed under UV to locate the 28S
and 18S ribosomal RNA bands. RNA was transferred to nitrocellulose filters
6201

Nucleic Acids Research
using 20 x SSC as the blotting buffer for 16 hours. After transfer filterswere rinsed briefly in 20 x SSC, dried at room temperature and kept at800C for 2 hours. The filters were prehybridised overnight at 420C in 50Oformamide, 5 x SSPE, 5 x Denhardt's solution, 0.1% SDS and 0.1 mg/mldenatured salmon sperm DNA. Hybridisation with nick-translated (15) CsClpurified plasmid DNA was carried out in the above buffer for 24 hours,using 1-10 x 106 Cerenkov cpm/ml. Filters were washed first in 2 x SSC 0.1lSDS and then in 0.1 x SSC, 0.1% SDS at room temperature. RNA sizes were
estimated using vitellogenin mRNA of 6.7 Kb (16), and 28S and 18S ribosomalRNAs as markers. In order to rehybridise to the same filter, the probe was
removed by soaking the filter in distilled H20 at 750C for 7 min and thefilter pre-hybridised and used as above.
In vitro translation of hybrid selected mRNA
Hybrid selection of mRNA specific to cDNA clones (16,21) was performedusing 1 mg/ml of total RNA from PE cells. In vitro translation of thehybrid selected mRNA was carried out in the nuclease-treated rabbitreticulocyte lysate and analysed by immunoprecipitation and SDS-gelelectrophoresis (7). The type IV collagen antisera used here has beendescribed previously and has been shown to be specific to type IVprocollagen polypeptides (7).Rescreening the PE cDNA library
Plasmids containing longer cDNA inserts hybridising to the pPE 1180cDNA were identified by rescreening the pUC8 and pUC9 libraries with theisolated, nick translated pPE 1180 insert, using conditions identical tothose used for the initial differential screen. Two positive signals wereidentified which contained cDNA inserts of 1.4 kb (pPE 76) and 1.5 kb(pPE 41). Both these clones recognised the pPE 1180-specific 6.8 kb RNAon Northern analysis.
RESULTS
A mouse embryo parietal endoderm (PE) cDNA library was constructedfrom total poly(A)+RNA using the double linker method and the expressionvectors pUC8 and pUC9 (10). This approach was adopted in the hope that cDNAclones expressing sequences of laminin, entactin and type IV procollagen,or other abundant parietal endoderm proteins could be isolated by antibody-screening (10). Initial attempts to identify cDNA clones expressing lamininor entactin sequences failed and because of this, we next screened the PEcDNA library by the differential hybridisation technique (17). Plasmid
6202

Nucleic Acids Research
l 9 a i 1C 1 2 2 4 8 9 K
.: '
*:.~ ~.*v_* lb
* 0'z*
Fig.1. Differential screening of PE cDNA library. Plasmid DNA wasprepared from 48 clones and dotted in duplicates onto two nitrocellulosefilters. Filter A was hybridised with a cDNA probe prepared from totalpoly(A)+RNA of undifferentiated F9 teratocarcinoma cells. Filter B washybridized with a cDNA probe prepared from total poly(A)+RNA of PE cells.PE specific clones were identified by visual comparison between the twofilters.
DNA isolated from 1300 clones was dot blotted in duplicate onto nitro-cellulose filters. One set of filters was hybridised with a cDNA-probeprepared from the total poly(A)+RNA of PE cells, and the other with a cDNA-
probe prepared from the total poly(A)+RNA of undifferentiated F9 cells.As described in the Introduction, we have estimated that PE cells synthesiseabout 100-fold more laminin, entactin and type IV procollagen than the F9(-) cells (7,8). Based on the results of the differential screening weidentified 19 clones that gave strong and 60 clones that gave less strongPE specific signals. As an example, Fig. 1 shows the signals obtained for48 clones. In addition to the one strong and three weak PE specific signals,three apparently F9 (-) specific signals were also identified on thisfilter.
We next analysed all the 19 strongly PE specific cDNA clones by
Northern hybridisation, to confirm the results obtained from the preliminarydifferential screen and to identify those cDNA clones which hybridised toan RNA of sufficient size to encode a type IV procollagen polypeptide of1741 amino acids (19). One plasmid, pPE 1180, which contained a 340 bpcDNA insert, showed the required behaviour. Fig. 2A shows that pPE 1180hybridised to a single RNA species of 6.8 kb present only in total cellularRNA from differentiated F9 cells (F9+) and not detectable in totalcellular RNA from undifferentiated F9 cells (F9-). Fig. 2B shows the same
filter reprobed with pDM238 (18), a plasmid containing a fragment of
D.melanogaster ribosomal DNA that recognises mouse 28S and 18S ribosomalRNA. This figure shows that equal amounts of ribosomal RNA were present
6203

Nucleic Acids Research
A B Northern hybridisation. Totalcellular RNA (15 'g) of differentiated(lane F9+) and undifferentiated (lane
F9+ F9- F9+ F9- F9-) F9 cells was analysed on aformaldehyde-agarose gel and blottedonto a nitrocellulose filter. Panel A:filter probed with a PE specificplasmid pPE 1180. This probe hybridisedto a single RNA of 6.8 kb whichappeared only in the F9+ lane. PanelB: the same filter reprobed with a
* plasmid containing a fragment of the4 28s 0 D.melanogaster ribosomal DNA (pDm 238).
This probe was used to show that the418ss _ _ nitrocellulose filter contained
comparable amounts of ribosomal RNAin lane F9+ and F9-.
in the F9+ and F9- lanes and that the absence of the 6.8 kb RNA in the F9-
lane is not due to difference in the amount of total cellular RNA present
in each lane.
The data presented in Fig. 2 show that pPE 1180 recognises an
abundant mRNA species present in differentiated F9 but not undifferentiated
F9 cells. This RNA is of sufficient size to encode the type IV procollagen
polypeptide. However studies (7,8), using the differentiating F9 terato-
carcinoma system have shown that other large polypeptides are developmentally
regulated. These include entactin, an Mr = 150 000 glycoprotein, and
laminin, a multisubunit glycoprotein consisting of an A chain, Mr = 360 000,
and two B chains Mr = 225 000 and 205 000. A third laminin B chain is also
seen after in vitro translation of laminin mRNA (8). In addition type IV
procollagen migrates as two polypeptides termed al (Mr = 185 000) and
a2(Mr = 170 000). All these polypeptides are modified in vivo and
consequently show a reduced molecular weight after in vitro translation.
Thus, the correct identity of pPE 1180 can only be ascertained by selection
of the specific mRNA and in vitro translation of its product. Results of
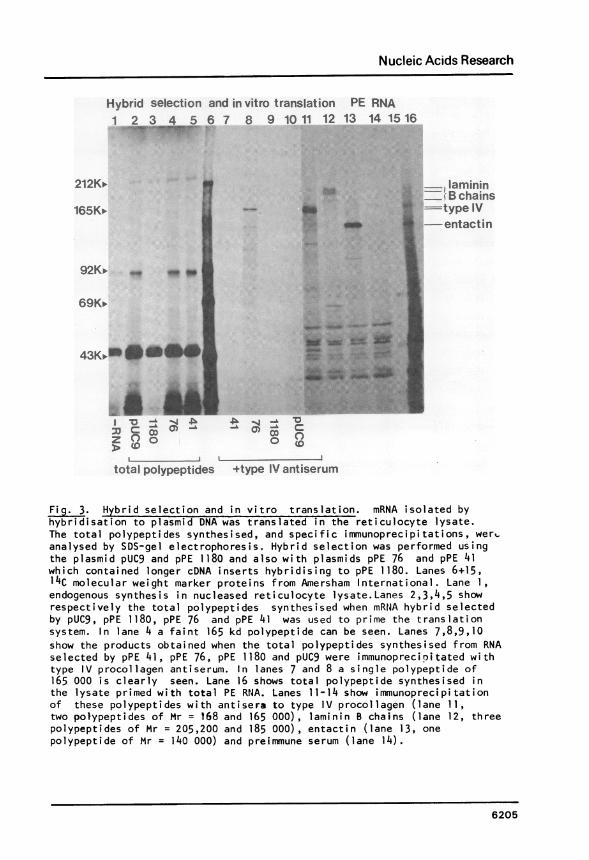
this selection are presented in Fig. 3 which shows the migration, on a
5-10% gradient SDS-polyacrylamide gel, of hybrid selected and control in
vitro synthesised polypeptides. Hybrid selection and in vitro translation
were performed as described (16,7). Fifty pg of total plasmid DNA was
6204
Nucleic Acids Research
Hybrid selection and in vitro translation PE RNA1 2 3 4 5 6 7 8 9 10 11 12 13 14 1516
v ,llt,,,,!PlllLvr
212Kw __ laminin22'r __ Bchains
165KPF p type IVannul. - entact i n
92Kw .,,
69K..
43KU-S _fl :ue:
4 14D COO 0
total polypeptides +type IV antiserum
Fig. 3. Hybrid selection and in vitro translation. mRNA isolated byhybridisation to plasmid DNA was translated in the reticulocyte lysate.The total polypeptides synthesised, and specific immunoprecipitations, wert,analysed by SDS-gel electrophoresis. Hybrid selection was performed usingthe plasmid pUC9 and pPE 1180 and also with plasmids pPE 76 and pPE 41which contained longer cDNA inserts hybridising to pPE 1180. Lanes 6+15,14C molecular weight marker proteins from Amersham International. Lane 1,endogenous synthesis in nucleased reticulocyte lysate.Lanes 2,3,4,5 showrespectively the total polypeptides synthesised when mRtJA hybrid selectedby pUC9, pPE 1180, pPE 76 and pPE 41 was used to prime the translationsystem. In lane 4 a faint 165 kd polypeptide can be seen. Lanes 7,8,9,10show the products obtained when the total polypeptides synthesised from RNAselected by pPE 41, pPE 76, pPE 1180 and pUC9 were immunoprecipitated withtype IV procollagen antiserum. In lanes 7 and 8 a single polypeptide of165 000 is clearly seen. Lane 16 shows total polypeptide synthesised inthe lysate primed with total PE RNA. Lanes 11-14 show immunoprecipitationof these polypeptides with antisera to type IV procollagen (lane 11,two polypeptides of Mr = 168 and 165 000), laminin B chains (lane 12, threepolypeptides of Mr = 205,200 and 185 000), entactin (lane 13, onepolypeptide of Mr = 140 000) and prei mmune serum (lane 14).
6205

Nucleic Acids Research
applied to a 3 mm2 nitrocellulose filter and used to hybrid selection
specific mRNA from PE total cellular RNA. Hybridisation was carried out at
500 for 3 hours in 50 il hybridisation mixture which contained 50 ig RNA
in 65% formamide (21). Hybrid selection was performed using pPE 1180,
pPE 76 and pPE 41. These plasmids contained cDNA inserts of 340 bp,
1400 and 1500 bp respectively. Fig. 3 lanes 3,4 and 5 show the total
polypeptides synthesised in the reticulocyte lysate primed with mRNA hybrid
selected by pPE 1180 (lane 3), pPE 76 (lane 4) and pPE 41 (lane 5).
Comparison with the endogenously synthesised polypeptides from this system
(lane 1) and those synthesised from mRNA hybrid selected by the plasmid
pUC9 (lane 2) shows that only pPE 76 selected mRNA that directed the
synthesis of a polypeptide (Mr = 165 000) which could be seen above the
background polypeptide synthesis. However,if the total polypeptides
synthesised from the hybrid selected mRNAs are immunoprecipitated with
antiserum that specifically reacts with type IV procollagen, this
Mr = 165 000 polypeptide is more clearly seen. In lanes 7,8,9 and 10,
the total polypeptides synthesised by hybrid selected mRNA from pPE 41(lane 7), pPE 76 (lane 8), pPE 1180 (lane 9) and pUC9 (lane 10) were
immunoprecipitated with type IV procollagen antisera. A Mr = 165 000
polypeptide is clearly seen in lanes 7 and 8.Total polypeptide synthesis directed by the PE RNA used for the
hybrid selections is shown in lane 16, and it can be seen that many large
polypeptides are present. Immunoprecipitation of the translation products
shows that only type IV procollagen antiserum(lane 11) specifically
immunoprecipitates the al and a2 doublet of Mr = 165 000 and 168 000 (notethat the unmodified polypeptides are smaller than those synthesised in vivo,
ref 7). These polypentides are not seen if the immunoprecipitation is done
with anti B-laminin serum (lane 12), anti entactin serum (lane 13) or
preimmune serum (lane 14). Thus Fig. 3 shows that hybrid selection using
plasmids pPE 41 and pPE 76, which contain longer cDNA inserts hybridising
to pPE 1180, can yield specific mRNA, encoding a Mr = 165 000 polypeptide
that comigrates with the smaller in vitro synthesised type IV procollagen
chain and is immunoprecipitated with antiserum specific to type IV
procol lagen.
Quantitation of type IV procollagen polypeptide production (7) has
shown that synthesis is approximately seven-fold higher in PE cells than in
differentiated F9 cells induced for 5.5 days, and approximately fifteen-
fold higher in differentiated F9 cells when compared to undifferentiated
6206
Nucleic Acids Research
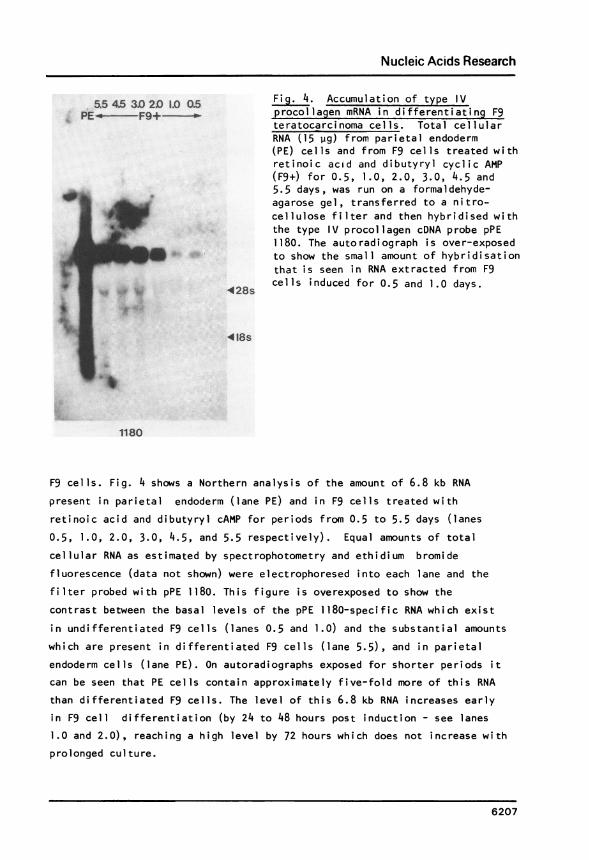
a5 45 31) 2D 1.0 0.5PE - F9+- --
428s
Fig. 4. Accumulation of type IVprocollagen mRNA in differentiating F9teratocarcinoma cel ls. Total cellularRNA (15 pg) from parietal endoderm(PE) cells and from F9 cells treated withretinoic acid and dibutyryl cyclic AMP(F9+) for 0.5, 1.0, 2.0, 3.0, 4.5 and5.5 days, was run on a formaldehyde-agarose gel, transferred to a nitro-cellulose filter and then hybridised withthe type IV procollagen cDNA probe pPE1180. The autoradiograph is over-exposedto show the small amount of hybridisationthat is seen in RNA extracted from F9cells induced for 0.5 and 1.0 days.
418s
1180
F9 cells. Fig. 4 shows a Northern analysis of the amount of 6.8 kb RNA
present in parietal endoderm (lane PE) and in F9 cells treated with
retinoic acid and dibutyryl cAMP for periods from 0.5 to 5.5 days (lanes0.5, 1.0, 2.0, 3.0, 4.5, and 5.5 respectively). Equal amounts of total
cellular RNA as estimated by spectrophotometry and ethidium bromide
fluorescence (data not shown) were electrophoresed into each lane and the
filter probed with pPE 1180. This figure is overexposed to show the
contrast between the basal levels of the pPE 1180-specific RNA which exist
in undifferentiated F9 cells (lanes 0.5 and 1.0) and the substantial amounts
which are present in differentiated F9 cells (lane 5.5), and in parietal
endoderm cells (lane PE). On autoradiographs exposed for shorter periods it
can be seen that PE cells contain approximately five-fold more of this RNA
than differentiated F9 cells. The level of this 6.8 kb RNA increases earlyin F9 cell differentiation (by 24 to 48 hours post induction - see lanes
1.0 and 2.0), reaching a high level by 72 hours which does not increase with
prolonged culture.
6207

Nucleic Acids Research
DISCUSSION
We have shown that the plasmid pPE 1180 contains a 340 bp cDNA insert
derived from the mRNA encoding the Mr = 165 000 polypeptide of mouse type
IV procollagen. At present it is not known whether this unmodified 165 kd
polypeptide corresponds to the al (Mr = 185 000) or a2 (Mr = 170 000)
modified procollagen polypeptide.
pPE 1180 recognises a 6.8 kb RNA species on Northern analysis. However,
since type IV procollagen contains 1740 amino acids (19) which corresponds
to 5.2 kb of RNA, there must be 1.6 kb of untranslated sequence in this RNA.
Type IV procollagen has two chains, al and a2, which are similar in their
properties but are encoded by separate mRNAs (7). Limited amino acid
sequences have been determined (23) and show that each polypeptide consists
of a central helical region, containing multiple repeats of the triplet
amino acid structure Gly-X-Y bounded by linear peptides at the amino and
carboxy termini. Clones pPE 76 and pPE 41, isolated from the original
cDNA library by hybridisation with pPE 1180, contain cDNA inserts of 1.4 kb
and 1.5 kb respectively. The sequence of 150 bp of the most 5' region of
these inserts has been determined by the Maxam-Gilbert technique (22) and
does not contain regions which could encode the known helical pattern of
amino acids (data not shown). cDNA inserts containing sequences which
correspond to the helical region of type IV procollagen would be expected
to hybridise to RNAs encoding both the al and a2 polypeptides. The fact
that this was not seen in the hybrid selection data presented in Fig. 3
is probably because the cDNA clones described here represent sequences
derived from the 3' non-coding region of the mRNA. The increased
proportion of AT sequences found in most 3' non-coding regions most likely
also explains the lack of selection by the 340 bp cDNA insert of pPE 1180
under the stringent conditions used here.
It is known that increased synthesis of type IV procollagen is induced
in F9 cells by retinoic acid and dibutyryl cAMP (4). Here we show, using a
cDNA probe to type IV procollagen (pPE 1180), that this is due to an
increase in the abundance of the type IV procollagen mRNA. This increased
cellular concentration was detected early, within 24 to 48 hours, and the
RNA accumulates to a constant high level by 72 hours post-induction (Fig. 4).Early changes in the morphology of F9 cells in response to these inducers
have been described (4) but, by both morphological and biochemical criteria,
the F9 phenotype continues to alter up to six days post-induction. It will
therefore be of interest to see if all the PE-specific cDNA clones isolated
6208

Nucleic Acids Research
from the parietal endoderm bank will hybridise to RNAs that behave in a
simi lar manner in the differentiating F9 system.
ACKNOWLEDGEMENTS
M.K. was supported in part by an EMBO short-term fellowship. We thank
David Ish-Horowicz for many helpful discussions, and also D. Glover for
the pDm 238 clone.
REFERENCES
1. Hogan, B.L.M. and Cooper, A.R. (1982) In 'New Trends in BasementMembrane Research' ed. K. Kuehn, H. Schoene and R. Timpl. Raven Press,New York, 245-255.
2. Cooper, A.R., Kurkinen, M., Taylor, A. and Hogan, B.L.M. (1981) Eur. J.Biochem. 119, 189-197.
3. Hogan, B.L.M., Taylor, A. and Cooper, A.R. (1982) Dev. Biol. 90, 210-214.
4. Strickland, S., Smith, K.K. and Marotti, K.R. (1980) Cell 21, 347-355.5. Hogan, B.L.M., Barlow, D.P. and Tilly, R. (1983) Cancer Surveys 2,
115-140.6. Cooper, A.R., Taylor, A. and Hogan, B.L.M. (1983) Dev. Biol. In the
press.7. Kurkinen, M., Foster, L., Barlow, D.P. and Hogan, B.L.M. (1982) J. Biol.
Chem. 257, 15151-15155.8. Kurkinen, M., Barlow, D.P., Jenkins, J.R. and Hogan, B.L.M. (1983)
J. Biol. Chem. 258, 6543-6548.9. Puskas, R.S., Manley, N.R., Wallace, D.M. and Berger, S.L. (1982)
Biochemistry 21, 4602-4608.T0. Helfman, D.M., Feramisco, J.R., Fiddes, J.C., Thomas, G.P. and
Hughes, S.H. (1983) Proc. Nat. Acad. Sci. USA 80, 31-35.11. Chirgwin, J.M., Przyoyla, A.E., MacDonald, R.J. and Rutter, W.J. (1979)
Biochemistry 18, 5294-5299.12. Blumberg, D.D. and Lodish, H.F. (1980) Dev. Biol. 78, 268-284.13. Vieira, J. and Messing, J. (1982) Gene 19, 259-268.14. Holmes, D.S. and Quigley, M. (1981) Anal. Biochem. 114, 193-197.15. Rigby, P.W.J., Dieckman, M., Rhodes, C. and Berg, P. (1977) J. Mol.
Biol. 113, 237-251.16. Parnes, J.R., Velan, B., Felsenfeld, A., Ramanathon, L., Ferrini, U.,
Apella, E. and Seidman, J.G. (1981) Proc. Nat. Acad. Sci. USA 78,2253-2257.
17. Williams, J.G. and Lloyd, M.M. (1979) J. Mol. Biol. 129, 19-35.18. D. Glover, personal communication.19. Dean, D.C., Barr, J.F., Freytag, J.W. and Hudson, B.G. (1983) J.Biol.
Chem. 258, 590-596.20. Trieb, G., Grobli, B., Spiess, M., Odermatt, B.F. and Winterhalter,
K.H. (1982) J. Biol. Chem. 257, 5239-5245.21. Maniatis, T., Fritsch, E.F. and Sambrook, J. (1982) Molecular Cloning,
A Laboratory Manual, CSH Publication U.S.A. p.330.22. Maxam, A. and Gilbert, W. (1980) Methods in Enzymology 65 (part 1)
497-559.23. Schuppen, D., Glanville, R.W. and Timpl, R. (1982) Eur. J. Biochem.
123, 505-512.
6209





























