Embed Size (px)
Citation preview

MARINE ECOLOGY PROGRESS SERIESMar Ecol Prog Ser
Vol 486 153ndash164 2013doi 103354meps10325
Published July 12
INTRODUCTION
The replacement of corals by macroalgae is a rela-tively common occurrence in the degradation of coralreefs (Hughes 1994 Bellwood et al 2004 Mumby etal 2006) This shift from coral to macroalgal domi-
nance often occurs as a result of disturbance and mor-tality to the coral community (Done 1992 Hughes1994) after which algae can proliferate through rapidrecruitment growth and early dominance of newlyavailable space (Littler amp Littler 1999 Diaz-Pulido ampMcCook 2002 Connell et al 2004) Two main pro-
copy Inter-Research 2013 middot wwwint-rescomEmail cdoropoulosuqeduau
Herbivores strongly influence algal recruitment inboth coral- and algal-dominated coral reef habitats
Christopher Doropoulos12 Glenn A Hyndes2 David Abecasis3 Adriana Vergeacutes245
1School of Biological Sciences and Australian Research Council Centre of Excellence for Coral Reef Studies The University of Queensland Brisbane 4072 Australia
2Centre for Marine Ecosystems Research School of Natural Sciences Edith Cowan University 270 Joondalup Drive Joondalup Western Australia 6027 Australia
3Centro de Ciecircncias do Mar da Universidade do Algarve Campus de Gambelas 8005-139 Faro Portugal4Centre for Marine Bio-Innovation and Evolution and Ecology Research Centre School of Biological
Earth and Environmental Sciences University of New South Wales Sydney New South Wales 2052 Australia5Sydney Institute of Marine Science Mosman New South Wales 2088 Australia
ABSTRACT Coral reefs can exist as coral- and macroalgae-dominated habitats often separatedby only a few hundred metres While herbivorous fish are known to depress the abundance ofalgae and help maintain the function of coral-dominated habitats less is known about their influence in algae-dominated habitats Here we quantified herbivorous fish and benthic algalcommunities over a 6 mo period in coral-dominated (back-reef) and algal-dominated (lagoon)habitats in a relatively undisturbed fringing coral reef (Ningaloo Western Australia) Simulta -neously we tested the effects of herbivorous fish on algal recruitment in both habitats usingrecruitment tiles and fish exclusion cages The composition of established algal communities differed consistently between habitats with the back-reef hosting a more diverse community thanthe Sargassum-dominated lagoon However total algal biomass and cover only differed betweenhabitats in autumn coinciding with maximum Sargassum biomass The back-reef hosted highcoral cover and a diverse herbivorous fish community with herbivore biomass an order of magni-tude greater than the lagoon Despite these differences in herbivore composition exclusion oflarge herbivores had a similar positive effect to foliose macroalgae recruitment on experimentaltiles in both back-reef and lagoon habitats Additionally territorial damselfish found in the back-reef increased turf algae cover and decreased crustose coralline algae cover on recruitment tilesCollectively our results show that disparate herbivorous fish communities in coral- and algae-dominated habitats are similarly able to limit the recruitment of foliose macroalgae but suggestthat when herbivorous fish biomass and diversity are relatively low macroalgal communities areable to escape herbivore control through increased growth
KEY WORDS Functional group middot Recruitment middot Macroalgae middot Sargassum middot Turf algae middot Crustosecoralline algae middot Ecosystem function middot Ningaloo
Resale or republication not permitted without written consent of the publisher
Mar Ecol Prog Ser 486 153ndash164 2013
cesses are often proposed to greatly influencemacroalgal abundance in degraded systems anthro-pogenic nutrient loading (bottom-up effects) andoverharvesting of herbivore populations (top-downeffects) (Burkepile amp Hay 2006) In coral reefs manystudies have investigated the relative importance ofthese 2 processes (eg Diaz-Pulido amp McCook 2003McClanahan et al 2003 Albert et al 2008 Smith etal 2010 Rasher et al 2012) and a meta-analysis of 54field experiments revealed that globally herbivoreshave a greater effect on algal production and distrib-ution than nutrient inputs (Burkepile amp Hay 2006)Herbivores can directly in fluence algal abundance byconsuming both algal recruits (Diaz-Pulido ampMcCook 2003) and adults (Hoey amp Bellwood 2010aBennett amp Bellwood 2011 Vergeacutes et al 2011) andtheir removal can cause algal proliferation that re-duces juvenile and adult coral abundance andgrowth (Jompa amp McCook 2002 Hughes et al 2007Birrell et al 2008 OrsquoLeary et al 2012)
Functional group identity of herbivorous fish is particularly important when assessing algae-grazerdynamics (Cheal et al 2010) Roving herbivorous fishare often classified into 3 major groups grazers anddetritivores (Acanthuridae most Siganidae) scrapersand excavators (Scaridae) and browsers (eg Nasospp Kyphosidae Green amp Bellwood 2009) Grazersand detritivores feed intensely on the epilithic algalmatrix (EAM which includes turf algae and detritus)(Wilson et al 2003 Marshell amp Mumby 2012) scra -pers and excavators also feed on the EAM butremove part of the substrate as they feed (Bellwood ampChoat 1990) and browsers feed on large foliosemacroalgae (Hoey amp Bellwood 2010a Bennett amp Bell-wood 2011 Vergeacutes et al 2012 Michael et al 2013)Territorial herbi vorous fish including many Poma-centridae and some Acanthuridae defend a definedarea of reef substratum against other herbivoresTheir behaviour promotes algal turf productivity anddiversity as well as altering successional trajectoriesof algal recruitment (Klumpp et al 1987 Hixon ampBrostoff 1996 Ceccarelli et al 2001 2005 2006 Gob-ler et al 2006 Hoey amp Bellwood 2010b)
While it is clear that high levels of algal cover are acharacteristic feature of degraded reefs recent stud-ies have challenged the view that intact reefs arecoral-dominated habitats with small algal popula-tions and propose that some healthy coral reef eco -systems may instead be regarded as adjacent coral-and algal-dominated benthic communities (Wismeret al 2009 Johansson et al 2010 Vroom amp Braun2010) Herbivory varies dramatically among theseadjoining habitats and is at least partly mediated by
structural complexity with the highest rates of algalconsumption occurring in the more complex coral-dominated habitats (Hay 1981 Hay et al 1983Vergeacutes et al 2011) The use of long-term experimen-tal herbivore exclusion cages has shown that herbi -vory is clearly one of the key processes that excludeslarge foliose algae from coral-dominated habitats(Hughes et al 2007 Burkepile amp Hay 2010 Rasher etal 2012) by limiting algal growth in both Caribbeanand Indo-Pacific coral reefs (reviewed by Roff ampMumby 2012) Similarly experiments assessing theimpact of herbivore exclusion on algal recruitment incoral-dominated habitats also have demonstratedthat high levels of herbivory can significantly reducethe recruitment of foliose algae to new substrata(Diaz-Pulido amp McCook 2003 Burkepile amp Hay 2006Smith et al 2010) In contrast to the well-establishedeffects of herbivores on coral-dominated habitatsour knowledge about the influence of these con-sumers on habitats where algae dominate is limitedHerbivore populations and herbivory rates are gen-erally low in these habitats (Fox amp Bellwood 2007Vergeacutes et al 2011) and it is thought that low con-sumption rates allow dense macroalgal stands toform Once high-density patches of algae developpositive feedback mechanisms inhibit further herbi -vory facilitating algal growth and persistence (Hoeyamp Bellwood 2011) However there is a lack of exper-imental evidence quantifying the effects of herbivo -res on algal recruitment in algal-dominated habitats
Algal recruitment in its broadest sense incorpo-rates the settlement and post-settlement survival andgrowth of algal propagules (sensu Diaz-Pulido ampMcCook 2003) Microscopic propagules are releasedinto the water column from gravid adult algae thesedisperse settle and then spend time in the EAMbefore developing into macroscopic individuals (Santelices 1990) Habitats with abundant adult algalpopulations may be expected to have higher algalrecruitment due to higher propagule supply Fordominant brown algae such as Sargassum spp dis-persal is highly localised with most propagules sett -ling within 1 m from the adult (Kendrick amp Walker1995) The post-settlement survival of algal recruits ishigher in areas with lower abundances of roving her-bivores (Diaz-Pulido amp McCook 2003) because algaeexperience little herbivory following recruitment tosuch areas (Lubchenco amp Gaines 1981 Diaz-Pulido ampMcCook 2003) Thus it may be expected that algalrecruitment would be greatest in areas with less her-bivores owing to (1) higher survival of microscopicsettled propagules in the EAM and (2) higher sur-vival of recruits that reach critical size-escape thresh-
154
Doropoulos et al Herbivore role in coral reef algal recruitment
olds from predation (Lubchenco amp Gaines 1981Diaz-Pulido amp McCook 2003) but this has never beentested
In this study we aimed to determine the effect oflarge herbivorous fish and habitat type on the re -cruiting algal community of a relatively intact coralreef ecosystem Ningaloo Reef (Western Australia)Over a 6 mo period we quantified changes in thebenthic community and the herbivorous fish commu-nity in back-reef and lagoon habitats 2 distinctneighbouring areas of the reef dominated by coraland algae respectively in terms of percentage coverof substratum Simultaneously we used settlementtiles and herbivore exclusion cages to test the effectsof habitat and large herbivorous fish on algal recruit-ment Based on a study by Vergeacutes et al (2011) whichshowed dominance of large macroalgae in thelagoon habitat and greater herbivore biomass andherbivory rates in the structurally complex coral-dominated back-reef habitat our a priori expectationwas that (1) algal recruitment would be greatest inthe lagoon where algae are more abundant and (2)algal recruitment would be reduced by herbivoryespecially in the more structured back-reef
MATERIALS AND METHODS
Study location
This study was conducted in the Ningaloo MarinePark Western Australia (see Fig S1 in the Supple-ment at wwwint-rescomarticlessupplm486 p153_supppdf) Ningaloo is a coral reef system that hasbeen managed as a marine park since 1987 and wasrezoned in 2005 to include 33 of the park as no-take sanctuary zones (CALM amp MPRA 2005) It isfound in an arid zone that is free of any major anthro-pogenic threats and is generally considered rela-tively undisturbed (Johansson et al 2010) NingalooReef is the largest fringing reef in Australia and theonly extensive fringing reef on the west coast of anycontinent (Collins et al 2003) It is a narrow fringingreef approximately 270 km in length that forms a dis-continuous barrier adjacent to the North West Capebetween the latitudes 21deg47rsquo S and 24deg 00rsquo S Theback-reef habitat is situated a few metres inshorefrom the reef crest at a depth of 15 m at the springlow tide and the lagoon habitat is situated furtherinshore at a similar depth Both habitats are equallyinfluenced by tidal movement which has a maxi-mum range of ~2 m during spring tides The back-reef is dominated by tabulate Acropora and the
lagoon is dominated by rubble sand and patches offoliose macroalgae (Cassata amp Collins 2008)
We conducted our study from November 2008 untilMay 2009 in the Mandu no-take sanctuary zone(22deg 08rsquo S 113deg45rsquo E) within Ningaloo Marine Park(Fig S1 in the Supplement) The Mandu SanctuaryZone is approximately 8 km long by 2 km wide andhas a lagoon ~600 m wide and a back-reef ~400 mwide We haphazardly selected 2 sites nested withineach habitat (lagoon and back-reef) with sitesbetween the lagoon and back-reef habitats sepa-rated by ~400 m and each site nested within a habi-tat separated by ~400 m (Fig S1 in the Supplement)
Benthic community characterisation
Benthic community cover and algal biomass weresurveyed at each site nested in the lagoon and back-reef habitats at 3 times of the year spring (November2008) summer (February 2009) and autumn (May2009) Benthic community cover was sampled using aline intercept method following Fox amp Bellwood(2007) A 10 m transect tape was laid haphazardlyacross the reef and a diver recorded the nature of thesubstrate directly touching the tape measure at 1 minterval points and at points 1 m perpendicular to theright and left of the tape (total of 30 points per re -plicate) The substrate categories used were livecoral crustose coralline algae (CCA) EAM (includesturf algae and detritus) foliose macroalgae (gt1 cmheight) and sand We conducted a total of 6 replicatetransects at each site in the lagoon and back-reefhabitats
Algal biomass was measured by clearing all macro -algae (thalli larger than 2 cm) in three 50 times 50 cmhaphazardly placed quadrats at each site-habitatcombination at every time point Algal samples werebagged and returned to the laboratory sorted togenus level where possible (a total of 17 genera wereidentified) rinsed with fresh water oven dried at60degC for ge48 h and weighed Algal taxa that wereun able to be identified were classified according tobroad functional groups including filamentousgreen filamentous red articulated coralline algaeand CCA
Herbivorous fish community characterisation
Herbivorous fish abundance and biomass werecharacterised for each site nested in the lagoon andback-reef habitats at the same 3 times of the year
155
Mar Ecol Prog Ser 486 153ndash164 2013
using the same transects in which benthic commu-nity cover and macroalgae were sampled (seeabove) A diver swam a 25 m transect at a constantspeed and counted the abundance and size class (to5 cm) of individual roving nominally herbivorous fish(sensu Choat et al 2002 2004) along 25 m on bothsides of the transect line Territorial damselfish abun-dance and size class were counted on the returnswim along each transect narrowing the width to 1 times25 m length At each site at each time period 4 repli-cate transects were conducted transects were sepa-rated from each other by at least 10 m Abundancedata were converted to biomass using the allometriclength-weight conversions W = a middotTLb where W isweight in g TL is total length in cm and parametersa and b are constants obtained from Fishbase (Froeseamp Pauly 2005) Fish surveys were always conductedat least 2 h after sunrise and 2 h before sunset Indi-viduals were recorded from the 4 major roving herbi-vore families (Acanthuridae Kyphosidae ScaridaeSiganidae) and territorial damselfish (Pomacentri-dae) to the lowest possible taxa (generally to speciesalways to genus) (see Table S1 in the Supplement)
Algal recruitment experimental design
To assess the influence of large herbivorous fish onalgal recruitment in habitats dominated either bymacro algae or coral cages were installed to excludeherbivores at each site-habitat combination We in -stalled 6 plots per treatment per site and these werescrubbed clean once a month throughout the experi-mental period The cages were triangular in shapeand measured 50 times 50 times 50 cm The corners weremarked with rebar to which PVC coated wire meshwas attached The mesh size used was 25 times 25 cmwhich excluded most herbivores but allowed somesmall fishes and invertebrates inside the cages In thecontext of this study large herbivorous fishes aretherefore considered as those excluded by the exper-imental cage mesh size In the caged plots the wirematerial covered all sides and the top of the plots Askirting edge on the sides of the cages was designedto prevent large benthic invertebrates from acces -sing the cages Open (uncaged) plots were markedwith the rebar and partial cages were used as a con-trol for cage artefacts and consisted of one side and aroof to maintain the potential influence of the cagewhile allowing access to all herbivores Some of theplots (70) from all 3 treatments in the back-reefhabitat became inhabited by territorial damselfishThese fish were included in analyses as a covariate
owing to their influence on benthic algal communi-ties (see review by Ceccarelli et al 2001) Urchins arenot abundant on the benthos in the back-reef orlagoon habitats (Johansson et al 2010) and theywere never found in the experimental plots
In each plot 2 recruitment tiles were installed inAugust 2008 and collected in February 2009 Thetiles measured 10 times 10 cm and were made from PVCAlthough PVC is not a natural substrate long-termalgal recruitment experiments have demonstratedthat algal community composition on this material issimilar to communities that recruit on tiles made ofnatural materials such as dead corals (Hixon amp Bros -toff 1996) The tiles were sanded to roughen the sur-faces which created irregular depressions to allowalgal attachment (Hixon amp Brostoff 1996 Smith et al2001 2010) Recruitment tiles were attached to apiece of horizontal wire mesh which was elevated10 cm from the benthos in each plot This standard-ised the attachment substrate and avoided smother-ing of the tiles by sediment
In February 2009 6 mo after deployment the 2 tilereplicates from each plot were carefully removed andplaced in individual Ziploc bags in situ After collec-tion the tiles were frozen until laboratory processingIn the laboratory each tile was thawed and rinsedwith seawater to remove any sediment and anysmall invertebrates were removed with forceps Todetermine the percent cover of the algal communityon the top surface of each tile a point-interceptmethod was used by placing a transparent plasticsheet with a grid of 25 dots on top of each tile Usinga dissecting microscope the algae or substrata pre-sent beneath each point were classified into one ofthe following categories bare tile CCA encrustingfleshy algae turf algae (red green or brown) andfoliose macroalgae (which included erect macro-phytes such as Dictyota Laurencia Lobophora Pad-ina and Sargassum)
Following classification algae from the tile werescraped and oven dried at 60degC for ge48 h after whichthe algae were weighed to 10minus4 g To estimate fleshyalgae biomass (including encrusting fleshy algae turfalgae and foliose macroalgae) HCl (10) was thenadded to the vials to remove any carbonate associatedwith coralline algae Vials were again rinsed placedin the oven at 60degC for ge48 h and re weighed
Data analyses
Separate principal coordinate analyses (PCO)were conducted to visualise differences in benthic
156
Doropoulos et al Herbivore role in coral reef algal recruitment
community cover algal community compositionand fish community composition Vector plots usingSpearman rankings based on correlations gt06 wereoverlaid on PCO plots to visualise relationshipsbetween variables and the ordination axes Differ-ences in total biomass of fleshy algae roving her-bivorous fish and damselfish were tested usingmixed effects analysis of variance (ANOVA) withhabitat (2 levels) and time (3 levels) as fixed factorsand site (2 levels) included as a random factornested in habitat Benthic cover and biomass ofalgal communities and roving herbi vorous fish wereused to assess patterns in their compositions usingseparate mixed effects permutational multivariateanalysis of variance (PERMANOVA) with the samemodel design described above
Mean percent cover on the top surface of the tileswas used to analyse the composition of the recruit-ing algal community Data collected from the repli-cate tiles within each plot were averaged and theaveraged data used as the residuals for all analyses(ie at the plot level) We used a mixed effects permutational multivariate analysis of covariance(PERMANCOVA) Habitat (2 levels) and herbivory(3 levels) were included as fixed factors with site(2 levels) in cluded as a random factor nested inhabitat Pomacentrus alexanderae (damselfish) werein cluded as a covariate because they establishedterritories in 70 of the experimental plots in theback-reef Following this the mean percent cover ofeach individual functional group and total fleshyalgal biomass were analysed using permutationalANCOVA using the same model
All data were evaluated using p-values generatedfrom 9999 permutations using Primer-E v6 (Clarkeamp Gorley 2006) with the PERMANOVA+ add-onpackage (Anderson et al 2008) When the numberof unique permutations was low the Monte Carloasymptotic p-value was used to assess statistical significance (Anderson 2005) Interaction or maineffect terms were pooled when they were highlynon-significant (p gt 025) (Underwood 1997) and significant differences (p lt 005) were investigatedwith pair-wise comparisons Euclidean distancewas the metric used to create resemblance matricesfor all univariate data analyses and Bray-Curtissimilarity was the metric used to create resem-blance matrices for all multivariate data analysesWhen necessary the data were transformed tomeet the assumptions of homogeneity (permuta-tional analysis of multi variate dispersion PERM -DISP) Transformations for relevant data ana ly sesare outlined in Tables S2minusS6 in the Supplement
RESULTS
Benthic community cover algal biomass and algal community composition
Multivariate analysis of the benthic communitycover revealed a significant distinction between thelagoon and back-reef habitats and between summerand autumn in both habitats (Table S2a in the Supp -lement at wwwint-rescomarticles suppl m486 p153_ supp pdf) The community composition was sepa-rated on the PCO1 axis (Fig 1a) which explained915 of the total variation (eigenvalue 49834) andthe PCO2 axis explained 84 of the total variation(eigenvalue 5498) Spearman correlations indicatedthat differences between habitats were clearly drivenby the dominance of foliose macroalgae and sand inthe lagoon and by dominance of live coral and EAMin the back-reef (Fig 1a) The lagoon had an averageof 29 macroalgae 63 sand and lt4 live coralEAM and CCA cover while the back-reef had anaverage of 48 live coral 28 EAM 16 sand 5macroalgae and 2 CCA cover
Total algal biomass showed a significant Habitat timesTime interaction (Table S2b) with biomass greaterin the lagoon compared with the back-reef duringautumn only (Fig 2a Table S2b) There was a mar ked increase in algal biomass in the lagoonfrom 168 g DW (dry weight) mminus2 in spring and sum-mer to 748 g DW mminus2 in autumn Benthic algal community composition displayed significant differ-ences be tween the lagoon and back-reef habitats(Table S2c) Spearman correlation indicated thatSargassum was highly correlated with the lagoonwhile no particular algae were correlated with theback-reef (Fig 1b) Throughout the study foliosealgae in the lagoon were dominated by Sargassum(88 algal community biomass) followed by Hyp-nea (9) In the back-reef Sargassum contri buted46 to the total algal community biomass 25Turbinaria 13 arti culated coralline algae 6Lobophora and 4 Dictyota
Herbivorous fish biomass and community composition
We recorded a total of 15 species of roving herbi -vorous fish from the Acanthuridae Kyphosidae Sca -ri dae and Siganidae families (see Table S1 in theSupplement) Initial phase parrotfish were recor dedas a single group (Scarus IP) Four species of terri -torial Pomacentridae that feed on benthic algae were
157
Mar Ecol Prog Ser 486 153ndash164 2013
recorded Hemiglyphidodon plagiometapon Poma-centrus alexanderae Stegastes fasciolatus and Sobreptus
Total biomass of roving herbivorous fish differedsignificantly between the lagoon and back-reef habi-tats where the biomass in the back-reef was at leastan order of magnitude higher than the lagoon(Fig 2b Table S3a) In contrast to the benthic algalcommunity we detected no significant differences intotal herbivorous fish biomass among times or sites(Table S3a) The herbivorous fish community compo-sition also differed significantly between the lagoonand back-reef habitats and we also detected differ-ences among sites but not among the times of theyear (Table S3b) Although there was some overlapthe separation of fish communities between the
158
Fig 1 Principal coordinate analyses (PCO) of (a) benthiccommunity cover (b) algal community composition and (c)herbivorous fish community composition from lagoon andback-reef habitats at Ningaloo Reef (M) la goon (S) back-reef Vector overlays represent correlations gt06 based onSpearman ranking CCA crustose coralline algae EAM
epilithic algal matrix
Fig 2 Mean biomass (plusmn SEM) of (a) fleshy algae and (b) her-bivorous fish in the lagoon and back-reef habitats duringspring summer and autumn at Ningaloo Reef Note the logarithmic scale on the y-axis for herbivorous fish biomass
Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Mar Ecol Prog Ser 486 153ndash164 2013
cesses are often proposed to greatly influencemacroalgal abundance in degraded systems anthro-pogenic nutrient loading (bottom-up effects) andoverharvesting of herbivore populations (top-downeffects) (Burkepile amp Hay 2006) In coral reefs manystudies have investigated the relative importance ofthese 2 processes (eg Diaz-Pulido amp McCook 2003McClanahan et al 2003 Albert et al 2008 Smith etal 2010 Rasher et al 2012) and a meta-analysis of 54field experiments revealed that globally herbivoreshave a greater effect on algal production and distrib-ution than nutrient inputs (Burkepile amp Hay 2006)Herbivores can directly in fluence algal abundance byconsuming both algal recruits (Diaz-Pulido ampMcCook 2003) and adults (Hoey amp Bellwood 2010aBennett amp Bellwood 2011 Vergeacutes et al 2011) andtheir removal can cause algal proliferation that re-duces juvenile and adult coral abundance andgrowth (Jompa amp McCook 2002 Hughes et al 2007Birrell et al 2008 OrsquoLeary et al 2012)
Functional group identity of herbivorous fish is particularly important when assessing algae-grazerdynamics (Cheal et al 2010) Roving herbivorous fishare often classified into 3 major groups grazers anddetritivores (Acanthuridae most Siganidae) scrapersand excavators (Scaridae) and browsers (eg Nasospp Kyphosidae Green amp Bellwood 2009) Grazersand detritivores feed intensely on the epilithic algalmatrix (EAM which includes turf algae and detritus)(Wilson et al 2003 Marshell amp Mumby 2012) scra -pers and excavators also feed on the EAM butremove part of the substrate as they feed (Bellwood ampChoat 1990) and browsers feed on large foliosemacroalgae (Hoey amp Bellwood 2010a Bennett amp Bell-wood 2011 Vergeacutes et al 2012 Michael et al 2013)Territorial herbi vorous fish including many Poma-centridae and some Acanthuridae defend a definedarea of reef substratum against other herbivoresTheir behaviour promotes algal turf productivity anddiversity as well as altering successional trajectoriesof algal recruitment (Klumpp et al 1987 Hixon ampBrostoff 1996 Ceccarelli et al 2001 2005 2006 Gob-ler et al 2006 Hoey amp Bellwood 2010b)
While it is clear that high levels of algal cover are acharacteristic feature of degraded reefs recent stud-ies have challenged the view that intact reefs arecoral-dominated habitats with small algal popula-tions and propose that some healthy coral reef eco -systems may instead be regarded as adjacent coral-and algal-dominated benthic communities (Wismeret al 2009 Johansson et al 2010 Vroom amp Braun2010) Herbivory varies dramatically among theseadjoining habitats and is at least partly mediated by
structural complexity with the highest rates of algalconsumption occurring in the more complex coral-dominated habitats (Hay 1981 Hay et al 1983Vergeacutes et al 2011) The use of long-term experimen-tal herbivore exclusion cages has shown that herbi -vory is clearly one of the key processes that excludeslarge foliose algae from coral-dominated habitats(Hughes et al 2007 Burkepile amp Hay 2010 Rasher etal 2012) by limiting algal growth in both Caribbeanand Indo-Pacific coral reefs (reviewed by Roff ampMumby 2012) Similarly experiments assessing theimpact of herbivore exclusion on algal recruitment incoral-dominated habitats also have demonstratedthat high levels of herbivory can significantly reducethe recruitment of foliose algae to new substrata(Diaz-Pulido amp McCook 2003 Burkepile amp Hay 2006Smith et al 2010) In contrast to the well-establishedeffects of herbivores on coral-dominated habitatsour knowledge about the influence of these con-sumers on habitats where algae dominate is limitedHerbivore populations and herbivory rates are gen-erally low in these habitats (Fox amp Bellwood 2007Vergeacutes et al 2011) and it is thought that low con-sumption rates allow dense macroalgal stands toform Once high-density patches of algae developpositive feedback mechanisms inhibit further herbi -vory facilitating algal growth and persistence (Hoeyamp Bellwood 2011) However there is a lack of exper-imental evidence quantifying the effects of herbivo -res on algal recruitment in algal-dominated habitats
Algal recruitment in its broadest sense incorpo-rates the settlement and post-settlement survival andgrowth of algal propagules (sensu Diaz-Pulido ampMcCook 2003) Microscopic propagules are releasedinto the water column from gravid adult algae thesedisperse settle and then spend time in the EAMbefore developing into macroscopic individuals (Santelices 1990) Habitats with abundant adult algalpopulations may be expected to have higher algalrecruitment due to higher propagule supply Fordominant brown algae such as Sargassum spp dis-persal is highly localised with most propagules sett -ling within 1 m from the adult (Kendrick amp Walker1995) The post-settlement survival of algal recruits ishigher in areas with lower abundances of roving her-bivores (Diaz-Pulido amp McCook 2003) because algaeexperience little herbivory following recruitment tosuch areas (Lubchenco amp Gaines 1981 Diaz-Pulido ampMcCook 2003) Thus it may be expected that algalrecruitment would be greatest in areas with less her-bivores owing to (1) higher survival of microscopicsettled propagules in the EAM and (2) higher sur-vival of recruits that reach critical size-escape thresh-
154
Doropoulos et al Herbivore role in coral reef algal recruitment
olds from predation (Lubchenco amp Gaines 1981Diaz-Pulido amp McCook 2003) but this has never beentested
In this study we aimed to determine the effect oflarge herbivorous fish and habitat type on the re -cruiting algal community of a relatively intact coralreef ecosystem Ningaloo Reef (Western Australia)Over a 6 mo period we quantified changes in thebenthic community and the herbivorous fish commu-nity in back-reef and lagoon habitats 2 distinctneighbouring areas of the reef dominated by coraland algae respectively in terms of percentage coverof substratum Simultaneously we used settlementtiles and herbivore exclusion cages to test the effectsof habitat and large herbivorous fish on algal recruit-ment Based on a study by Vergeacutes et al (2011) whichshowed dominance of large macroalgae in thelagoon habitat and greater herbivore biomass andherbivory rates in the structurally complex coral-dominated back-reef habitat our a priori expectationwas that (1) algal recruitment would be greatest inthe lagoon where algae are more abundant and (2)algal recruitment would be reduced by herbivoryespecially in the more structured back-reef
MATERIALS AND METHODS
Study location
This study was conducted in the Ningaloo MarinePark Western Australia (see Fig S1 in the Supple-ment at wwwint-rescomarticlessupplm486 p153_supppdf) Ningaloo is a coral reef system that hasbeen managed as a marine park since 1987 and wasrezoned in 2005 to include 33 of the park as no-take sanctuary zones (CALM amp MPRA 2005) It isfound in an arid zone that is free of any major anthro-pogenic threats and is generally considered rela-tively undisturbed (Johansson et al 2010) NingalooReef is the largest fringing reef in Australia and theonly extensive fringing reef on the west coast of anycontinent (Collins et al 2003) It is a narrow fringingreef approximately 270 km in length that forms a dis-continuous barrier adjacent to the North West Capebetween the latitudes 21deg47rsquo S and 24deg 00rsquo S Theback-reef habitat is situated a few metres inshorefrom the reef crest at a depth of 15 m at the springlow tide and the lagoon habitat is situated furtherinshore at a similar depth Both habitats are equallyinfluenced by tidal movement which has a maxi-mum range of ~2 m during spring tides The back-reef is dominated by tabulate Acropora and the
lagoon is dominated by rubble sand and patches offoliose macroalgae (Cassata amp Collins 2008)
We conducted our study from November 2008 untilMay 2009 in the Mandu no-take sanctuary zone(22deg 08rsquo S 113deg45rsquo E) within Ningaloo Marine Park(Fig S1 in the Supplement) The Mandu SanctuaryZone is approximately 8 km long by 2 km wide andhas a lagoon ~600 m wide and a back-reef ~400 mwide We haphazardly selected 2 sites nested withineach habitat (lagoon and back-reef) with sitesbetween the lagoon and back-reef habitats sepa-rated by ~400 m and each site nested within a habi-tat separated by ~400 m (Fig S1 in the Supplement)
Benthic community characterisation
Benthic community cover and algal biomass weresurveyed at each site nested in the lagoon and back-reef habitats at 3 times of the year spring (November2008) summer (February 2009) and autumn (May2009) Benthic community cover was sampled using aline intercept method following Fox amp Bellwood(2007) A 10 m transect tape was laid haphazardlyacross the reef and a diver recorded the nature of thesubstrate directly touching the tape measure at 1 minterval points and at points 1 m perpendicular to theright and left of the tape (total of 30 points per re -plicate) The substrate categories used were livecoral crustose coralline algae (CCA) EAM (includesturf algae and detritus) foliose macroalgae (gt1 cmheight) and sand We conducted a total of 6 replicatetransects at each site in the lagoon and back-reefhabitats
Algal biomass was measured by clearing all macro -algae (thalli larger than 2 cm) in three 50 times 50 cmhaphazardly placed quadrats at each site-habitatcombination at every time point Algal samples werebagged and returned to the laboratory sorted togenus level where possible (a total of 17 genera wereidentified) rinsed with fresh water oven dried at60degC for ge48 h and weighed Algal taxa that wereun able to be identified were classified according tobroad functional groups including filamentousgreen filamentous red articulated coralline algaeand CCA
Herbivorous fish community characterisation
Herbivorous fish abundance and biomass werecharacterised for each site nested in the lagoon andback-reef habitats at the same 3 times of the year
155
Mar Ecol Prog Ser 486 153ndash164 2013
using the same transects in which benthic commu-nity cover and macroalgae were sampled (seeabove) A diver swam a 25 m transect at a constantspeed and counted the abundance and size class (to5 cm) of individual roving nominally herbivorous fish(sensu Choat et al 2002 2004) along 25 m on bothsides of the transect line Territorial damselfish abun-dance and size class were counted on the returnswim along each transect narrowing the width to 1 times25 m length At each site at each time period 4 repli-cate transects were conducted transects were sepa-rated from each other by at least 10 m Abundancedata were converted to biomass using the allometriclength-weight conversions W = a middotTLb where W isweight in g TL is total length in cm and parametersa and b are constants obtained from Fishbase (Froeseamp Pauly 2005) Fish surveys were always conductedat least 2 h after sunrise and 2 h before sunset Indi-viduals were recorded from the 4 major roving herbi-vore families (Acanthuridae Kyphosidae ScaridaeSiganidae) and territorial damselfish (Pomacentri-dae) to the lowest possible taxa (generally to speciesalways to genus) (see Table S1 in the Supplement)
Algal recruitment experimental design
To assess the influence of large herbivorous fish onalgal recruitment in habitats dominated either bymacro algae or coral cages were installed to excludeherbivores at each site-habitat combination We in -stalled 6 plots per treatment per site and these werescrubbed clean once a month throughout the experi-mental period The cages were triangular in shapeand measured 50 times 50 times 50 cm The corners weremarked with rebar to which PVC coated wire meshwas attached The mesh size used was 25 times 25 cmwhich excluded most herbivores but allowed somesmall fishes and invertebrates inside the cages In thecontext of this study large herbivorous fishes aretherefore considered as those excluded by the exper-imental cage mesh size In the caged plots the wirematerial covered all sides and the top of the plots Askirting edge on the sides of the cages was designedto prevent large benthic invertebrates from acces -sing the cages Open (uncaged) plots were markedwith the rebar and partial cages were used as a con-trol for cage artefacts and consisted of one side and aroof to maintain the potential influence of the cagewhile allowing access to all herbivores Some of theplots (70) from all 3 treatments in the back-reefhabitat became inhabited by territorial damselfishThese fish were included in analyses as a covariate
owing to their influence on benthic algal communi-ties (see review by Ceccarelli et al 2001) Urchins arenot abundant on the benthos in the back-reef orlagoon habitats (Johansson et al 2010) and theywere never found in the experimental plots
In each plot 2 recruitment tiles were installed inAugust 2008 and collected in February 2009 Thetiles measured 10 times 10 cm and were made from PVCAlthough PVC is not a natural substrate long-termalgal recruitment experiments have demonstratedthat algal community composition on this material issimilar to communities that recruit on tiles made ofnatural materials such as dead corals (Hixon amp Bros -toff 1996) The tiles were sanded to roughen the sur-faces which created irregular depressions to allowalgal attachment (Hixon amp Brostoff 1996 Smith et al2001 2010) Recruitment tiles were attached to apiece of horizontal wire mesh which was elevated10 cm from the benthos in each plot This standard-ised the attachment substrate and avoided smother-ing of the tiles by sediment
In February 2009 6 mo after deployment the 2 tilereplicates from each plot were carefully removed andplaced in individual Ziploc bags in situ After collec-tion the tiles were frozen until laboratory processingIn the laboratory each tile was thawed and rinsedwith seawater to remove any sediment and anysmall invertebrates were removed with forceps Todetermine the percent cover of the algal communityon the top surface of each tile a point-interceptmethod was used by placing a transparent plasticsheet with a grid of 25 dots on top of each tile Usinga dissecting microscope the algae or substrata pre-sent beneath each point were classified into one ofthe following categories bare tile CCA encrustingfleshy algae turf algae (red green or brown) andfoliose macroalgae (which included erect macro-phytes such as Dictyota Laurencia Lobophora Pad-ina and Sargassum)
Following classification algae from the tile werescraped and oven dried at 60degC for ge48 h after whichthe algae were weighed to 10minus4 g To estimate fleshyalgae biomass (including encrusting fleshy algae turfalgae and foliose macroalgae) HCl (10) was thenadded to the vials to remove any carbonate associatedwith coralline algae Vials were again rinsed placedin the oven at 60degC for ge48 h and re weighed
Data analyses
Separate principal coordinate analyses (PCO)were conducted to visualise differences in benthic
156
Doropoulos et al Herbivore role in coral reef algal recruitment
community cover algal community compositionand fish community composition Vector plots usingSpearman rankings based on correlations gt06 wereoverlaid on PCO plots to visualise relationshipsbetween variables and the ordination axes Differ-ences in total biomass of fleshy algae roving her-bivorous fish and damselfish were tested usingmixed effects analysis of variance (ANOVA) withhabitat (2 levels) and time (3 levels) as fixed factorsand site (2 levels) included as a random factornested in habitat Benthic cover and biomass ofalgal communities and roving herbi vorous fish wereused to assess patterns in their compositions usingseparate mixed effects permutational multivariateanalysis of variance (PERMANOVA) with the samemodel design described above
Mean percent cover on the top surface of the tileswas used to analyse the composition of the recruit-ing algal community Data collected from the repli-cate tiles within each plot were averaged and theaveraged data used as the residuals for all analyses(ie at the plot level) We used a mixed effects permutational multivariate analysis of covariance(PERMANCOVA) Habitat (2 levels) and herbivory(3 levels) were included as fixed factors with site(2 levels) in cluded as a random factor nested inhabitat Pomacentrus alexanderae (damselfish) werein cluded as a covariate because they establishedterritories in 70 of the experimental plots in theback-reef Following this the mean percent cover ofeach individual functional group and total fleshyalgal biomass were analysed using permutationalANCOVA using the same model
All data were evaluated using p-values generatedfrom 9999 permutations using Primer-E v6 (Clarkeamp Gorley 2006) with the PERMANOVA+ add-onpackage (Anderson et al 2008) When the numberof unique permutations was low the Monte Carloasymptotic p-value was used to assess statistical significance (Anderson 2005) Interaction or maineffect terms were pooled when they were highlynon-significant (p gt 025) (Underwood 1997) and significant differences (p lt 005) were investigatedwith pair-wise comparisons Euclidean distancewas the metric used to create resemblance matricesfor all univariate data analyses and Bray-Curtissimilarity was the metric used to create resem-blance matrices for all multivariate data analysesWhen necessary the data were transformed tomeet the assumptions of homogeneity (permuta-tional analysis of multi variate dispersion PERM -DISP) Transformations for relevant data ana ly sesare outlined in Tables S2minusS6 in the Supplement
RESULTS
Benthic community cover algal biomass and algal community composition
Multivariate analysis of the benthic communitycover revealed a significant distinction between thelagoon and back-reef habitats and between summerand autumn in both habitats (Table S2a in the Supp -lement at wwwint-rescomarticles suppl m486 p153_ supp pdf) The community composition was sepa-rated on the PCO1 axis (Fig 1a) which explained915 of the total variation (eigenvalue 49834) andthe PCO2 axis explained 84 of the total variation(eigenvalue 5498) Spearman correlations indicatedthat differences between habitats were clearly drivenby the dominance of foliose macroalgae and sand inthe lagoon and by dominance of live coral and EAMin the back-reef (Fig 1a) The lagoon had an averageof 29 macroalgae 63 sand and lt4 live coralEAM and CCA cover while the back-reef had anaverage of 48 live coral 28 EAM 16 sand 5macroalgae and 2 CCA cover
Total algal biomass showed a significant Habitat timesTime interaction (Table S2b) with biomass greaterin the lagoon compared with the back-reef duringautumn only (Fig 2a Table S2b) There was a mar ked increase in algal biomass in the lagoonfrom 168 g DW (dry weight) mminus2 in spring and sum-mer to 748 g DW mminus2 in autumn Benthic algal community composition displayed significant differ-ences be tween the lagoon and back-reef habitats(Table S2c) Spearman correlation indicated thatSargassum was highly correlated with the lagoonwhile no particular algae were correlated with theback-reef (Fig 1b) Throughout the study foliosealgae in the lagoon were dominated by Sargassum(88 algal community biomass) followed by Hyp-nea (9) In the back-reef Sargassum contri buted46 to the total algal community biomass 25Turbinaria 13 arti culated coralline algae 6Lobophora and 4 Dictyota
Herbivorous fish biomass and community composition
We recorded a total of 15 species of roving herbi -vorous fish from the Acanthuridae Kyphosidae Sca -ri dae and Siganidae families (see Table S1 in theSupplement) Initial phase parrotfish were recor dedas a single group (Scarus IP) Four species of terri -torial Pomacentridae that feed on benthic algae were
157
Mar Ecol Prog Ser 486 153ndash164 2013
recorded Hemiglyphidodon plagiometapon Poma-centrus alexanderae Stegastes fasciolatus and Sobreptus
Total biomass of roving herbivorous fish differedsignificantly between the lagoon and back-reef habi-tats where the biomass in the back-reef was at leastan order of magnitude higher than the lagoon(Fig 2b Table S3a) In contrast to the benthic algalcommunity we detected no significant differences intotal herbivorous fish biomass among times or sites(Table S3a) The herbivorous fish community compo-sition also differed significantly between the lagoonand back-reef habitats and we also detected differ-ences among sites but not among the times of theyear (Table S3b) Although there was some overlapthe separation of fish communities between the
158
Fig 1 Principal coordinate analyses (PCO) of (a) benthiccommunity cover (b) algal community composition and (c)herbivorous fish community composition from lagoon andback-reef habitats at Ningaloo Reef (M) la goon (S) back-reef Vector overlays represent correlations gt06 based onSpearman ranking CCA crustose coralline algae EAM
epilithic algal matrix
Fig 2 Mean biomass (plusmn SEM) of (a) fleshy algae and (b) her-bivorous fish in the lagoon and back-reef habitats duringspring summer and autumn at Ningaloo Reef Note the logarithmic scale on the y-axis for herbivorous fish biomass
Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Doropoulos et al Herbivore role in coral reef algal recruitment
olds from predation (Lubchenco amp Gaines 1981Diaz-Pulido amp McCook 2003) but this has never beentested
In this study we aimed to determine the effect oflarge herbivorous fish and habitat type on the re -cruiting algal community of a relatively intact coralreef ecosystem Ningaloo Reef (Western Australia)Over a 6 mo period we quantified changes in thebenthic community and the herbivorous fish commu-nity in back-reef and lagoon habitats 2 distinctneighbouring areas of the reef dominated by coraland algae respectively in terms of percentage coverof substratum Simultaneously we used settlementtiles and herbivore exclusion cages to test the effectsof habitat and large herbivorous fish on algal recruit-ment Based on a study by Vergeacutes et al (2011) whichshowed dominance of large macroalgae in thelagoon habitat and greater herbivore biomass andherbivory rates in the structurally complex coral-dominated back-reef habitat our a priori expectationwas that (1) algal recruitment would be greatest inthe lagoon where algae are more abundant and (2)algal recruitment would be reduced by herbivoryespecially in the more structured back-reef
MATERIALS AND METHODS
Study location
This study was conducted in the Ningaloo MarinePark Western Australia (see Fig S1 in the Supple-ment at wwwint-rescomarticlessupplm486 p153_supppdf) Ningaloo is a coral reef system that hasbeen managed as a marine park since 1987 and wasrezoned in 2005 to include 33 of the park as no-take sanctuary zones (CALM amp MPRA 2005) It isfound in an arid zone that is free of any major anthro-pogenic threats and is generally considered rela-tively undisturbed (Johansson et al 2010) NingalooReef is the largest fringing reef in Australia and theonly extensive fringing reef on the west coast of anycontinent (Collins et al 2003) It is a narrow fringingreef approximately 270 km in length that forms a dis-continuous barrier adjacent to the North West Capebetween the latitudes 21deg47rsquo S and 24deg 00rsquo S Theback-reef habitat is situated a few metres inshorefrom the reef crest at a depth of 15 m at the springlow tide and the lagoon habitat is situated furtherinshore at a similar depth Both habitats are equallyinfluenced by tidal movement which has a maxi-mum range of ~2 m during spring tides The back-reef is dominated by tabulate Acropora and the
lagoon is dominated by rubble sand and patches offoliose macroalgae (Cassata amp Collins 2008)
We conducted our study from November 2008 untilMay 2009 in the Mandu no-take sanctuary zone(22deg 08rsquo S 113deg45rsquo E) within Ningaloo Marine Park(Fig S1 in the Supplement) The Mandu SanctuaryZone is approximately 8 km long by 2 km wide andhas a lagoon ~600 m wide and a back-reef ~400 mwide We haphazardly selected 2 sites nested withineach habitat (lagoon and back-reef) with sitesbetween the lagoon and back-reef habitats sepa-rated by ~400 m and each site nested within a habi-tat separated by ~400 m (Fig S1 in the Supplement)
Benthic community characterisation
Benthic community cover and algal biomass weresurveyed at each site nested in the lagoon and back-reef habitats at 3 times of the year spring (November2008) summer (February 2009) and autumn (May2009) Benthic community cover was sampled using aline intercept method following Fox amp Bellwood(2007) A 10 m transect tape was laid haphazardlyacross the reef and a diver recorded the nature of thesubstrate directly touching the tape measure at 1 minterval points and at points 1 m perpendicular to theright and left of the tape (total of 30 points per re -plicate) The substrate categories used were livecoral crustose coralline algae (CCA) EAM (includesturf algae and detritus) foliose macroalgae (gt1 cmheight) and sand We conducted a total of 6 replicatetransects at each site in the lagoon and back-reefhabitats
Algal biomass was measured by clearing all macro -algae (thalli larger than 2 cm) in three 50 times 50 cmhaphazardly placed quadrats at each site-habitatcombination at every time point Algal samples werebagged and returned to the laboratory sorted togenus level where possible (a total of 17 genera wereidentified) rinsed with fresh water oven dried at60degC for ge48 h and weighed Algal taxa that wereun able to be identified were classified according tobroad functional groups including filamentousgreen filamentous red articulated coralline algaeand CCA
Herbivorous fish community characterisation
Herbivorous fish abundance and biomass werecharacterised for each site nested in the lagoon andback-reef habitats at the same 3 times of the year
155
Mar Ecol Prog Ser 486 153ndash164 2013
using the same transects in which benthic commu-nity cover and macroalgae were sampled (seeabove) A diver swam a 25 m transect at a constantspeed and counted the abundance and size class (to5 cm) of individual roving nominally herbivorous fish(sensu Choat et al 2002 2004) along 25 m on bothsides of the transect line Territorial damselfish abun-dance and size class were counted on the returnswim along each transect narrowing the width to 1 times25 m length At each site at each time period 4 repli-cate transects were conducted transects were sepa-rated from each other by at least 10 m Abundancedata were converted to biomass using the allometriclength-weight conversions W = a middotTLb where W isweight in g TL is total length in cm and parametersa and b are constants obtained from Fishbase (Froeseamp Pauly 2005) Fish surveys were always conductedat least 2 h after sunrise and 2 h before sunset Indi-viduals were recorded from the 4 major roving herbi-vore families (Acanthuridae Kyphosidae ScaridaeSiganidae) and territorial damselfish (Pomacentri-dae) to the lowest possible taxa (generally to speciesalways to genus) (see Table S1 in the Supplement)
Algal recruitment experimental design
To assess the influence of large herbivorous fish onalgal recruitment in habitats dominated either bymacro algae or coral cages were installed to excludeherbivores at each site-habitat combination We in -stalled 6 plots per treatment per site and these werescrubbed clean once a month throughout the experi-mental period The cages were triangular in shapeand measured 50 times 50 times 50 cm The corners weremarked with rebar to which PVC coated wire meshwas attached The mesh size used was 25 times 25 cmwhich excluded most herbivores but allowed somesmall fishes and invertebrates inside the cages In thecontext of this study large herbivorous fishes aretherefore considered as those excluded by the exper-imental cage mesh size In the caged plots the wirematerial covered all sides and the top of the plots Askirting edge on the sides of the cages was designedto prevent large benthic invertebrates from acces -sing the cages Open (uncaged) plots were markedwith the rebar and partial cages were used as a con-trol for cage artefacts and consisted of one side and aroof to maintain the potential influence of the cagewhile allowing access to all herbivores Some of theplots (70) from all 3 treatments in the back-reefhabitat became inhabited by territorial damselfishThese fish were included in analyses as a covariate
owing to their influence on benthic algal communi-ties (see review by Ceccarelli et al 2001) Urchins arenot abundant on the benthos in the back-reef orlagoon habitats (Johansson et al 2010) and theywere never found in the experimental plots
In each plot 2 recruitment tiles were installed inAugust 2008 and collected in February 2009 Thetiles measured 10 times 10 cm and were made from PVCAlthough PVC is not a natural substrate long-termalgal recruitment experiments have demonstratedthat algal community composition on this material issimilar to communities that recruit on tiles made ofnatural materials such as dead corals (Hixon amp Bros -toff 1996) The tiles were sanded to roughen the sur-faces which created irregular depressions to allowalgal attachment (Hixon amp Brostoff 1996 Smith et al2001 2010) Recruitment tiles were attached to apiece of horizontal wire mesh which was elevated10 cm from the benthos in each plot This standard-ised the attachment substrate and avoided smother-ing of the tiles by sediment
In February 2009 6 mo after deployment the 2 tilereplicates from each plot were carefully removed andplaced in individual Ziploc bags in situ After collec-tion the tiles were frozen until laboratory processingIn the laboratory each tile was thawed and rinsedwith seawater to remove any sediment and anysmall invertebrates were removed with forceps Todetermine the percent cover of the algal communityon the top surface of each tile a point-interceptmethod was used by placing a transparent plasticsheet with a grid of 25 dots on top of each tile Usinga dissecting microscope the algae or substrata pre-sent beneath each point were classified into one ofthe following categories bare tile CCA encrustingfleshy algae turf algae (red green or brown) andfoliose macroalgae (which included erect macro-phytes such as Dictyota Laurencia Lobophora Pad-ina and Sargassum)
Following classification algae from the tile werescraped and oven dried at 60degC for ge48 h after whichthe algae were weighed to 10minus4 g To estimate fleshyalgae biomass (including encrusting fleshy algae turfalgae and foliose macroalgae) HCl (10) was thenadded to the vials to remove any carbonate associatedwith coralline algae Vials were again rinsed placedin the oven at 60degC for ge48 h and re weighed
Data analyses
Separate principal coordinate analyses (PCO)were conducted to visualise differences in benthic
156
Doropoulos et al Herbivore role in coral reef algal recruitment
community cover algal community compositionand fish community composition Vector plots usingSpearman rankings based on correlations gt06 wereoverlaid on PCO plots to visualise relationshipsbetween variables and the ordination axes Differ-ences in total biomass of fleshy algae roving her-bivorous fish and damselfish were tested usingmixed effects analysis of variance (ANOVA) withhabitat (2 levels) and time (3 levels) as fixed factorsand site (2 levels) included as a random factornested in habitat Benthic cover and biomass ofalgal communities and roving herbi vorous fish wereused to assess patterns in their compositions usingseparate mixed effects permutational multivariateanalysis of variance (PERMANOVA) with the samemodel design described above
Mean percent cover on the top surface of the tileswas used to analyse the composition of the recruit-ing algal community Data collected from the repli-cate tiles within each plot were averaged and theaveraged data used as the residuals for all analyses(ie at the plot level) We used a mixed effects permutational multivariate analysis of covariance(PERMANCOVA) Habitat (2 levels) and herbivory(3 levels) were included as fixed factors with site(2 levels) in cluded as a random factor nested inhabitat Pomacentrus alexanderae (damselfish) werein cluded as a covariate because they establishedterritories in 70 of the experimental plots in theback-reef Following this the mean percent cover ofeach individual functional group and total fleshyalgal biomass were analysed using permutationalANCOVA using the same model
All data were evaluated using p-values generatedfrom 9999 permutations using Primer-E v6 (Clarkeamp Gorley 2006) with the PERMANOVA+ add-onpackage (Anderson et al 2008) When the numberof unique permutations was low the Monte Carloasymptotic p-value was used to assess statistical significance (Anderson 2005) Interaction or maineffect terms were pooled when they were highlynon-significant (p gt 025) (Underwood 1997) and significant differences (p lt 005) were investigatedwith pair-wise comparisons Euclidean distancewas the metric used to create resemblance matricesfor all univariate data analyses and Bray-Curtissimilarity was the metric used to create resem-blance matrices for all multivariate data analysesWhen necessary the data were transformed tomeet the assumptions of homogeneity (permuta-tional analysis of multi variate dispersion PERM -DISP) Transformations for relevant data ana ly sesare outlined in Tables S2minusS6 in the Supplement
RESULTS
Benthic community cover algal biomass and algal community composition
Multivariate analysis of the benthic communitycover revealed a significant distinction between thelagoon and back-reef habitats and between summerand autumn in both habitats (Table S2a in the Supp -lement at wwwint-rescomarticles suppl m486 p153_ supp pdf) The community composition was sepa-rated on the PCO1 axis (Fig 1a) which explained915 of the total variation (eigenvalue 49834) andthe PCO2 axis explained 84 of the total variation(eigenvalue 5498) Spearman correlations indicatedthat differences between habitats were clearly drivenby the dominance of foliose macroalgae and sand inthe lagoon and by dominance of live coral and EAMin the back-reef (Fig 1a) The lagoon had an averageof 29 macroalgae 63 sand and lt4 live coralEAM and CCA cover while the back-reef had anaverage of 48 live coral 28 EAM 16 sand 5macroalgae and 2 CCA cover
Total algal biomass showed a significant Habitat timesTime interaction (Table S2b) with biomass greaterin the lagoon compared with the back-reef duringautumn only (Fig 2a Table S2b) There was a mar ked increase in algal biomass in the lagoonfrom 168 g DW (dry weight) mminus2 in spring and sum-mer to 748 g DW mminus2 in autumn Benthic algal community composition displayed significant differ-ences be tween the lagoon and back-reef habitats(Table S2c) Spearman correlation indicated thatSargassum was highly correlated with the lagoonwhile no particular algae were correlated with theback-reef (Fig 1b) Throughout the study foliosealgae in the lagoon were dominated by Sargassum(88 algal community biomass) followed by Hyp-nea (9) In the back-reef Sargassum contri buted46 to the total algal community biomass 25Turbinaria 13 arti culated coralline algae 6Lobophora and 4 Dictyota
Herbivorous fish biomass and community composition
We recorded a total of 15 species of roving herbi -vorous fish from the Acanthuridae Kyphosidae Sca -ri dae and Siganidae families (see Table S1 in theSupplement) Initial phase parrotfish were recor dedas a single group (Scarus IP) Four species of terri -torial Pomacentridae that feed on benthic algae were
157
Mar Ecol Prog Ser 486 153ndash164 2013
recorded Hemiglyphidodon plagiometapon Poma-centrus alexanderae Stegastes fasciolatus and Sobreptus
Total biomass of roving herbivorous fish differedsignificantly between the lagoon and back-reef habi-tats where the biomass in the back-reef was at leastan order of magnitude higher than the lagoon(Fig 2b Table S3a) In contrast to the benthic algalcommunity we detected no significant differences intotal herbivorous fish biomass among times or sites(Table S3a) The herbivorous fish community compo-sition also differed significantly between the lagoonand back-reef habitats and we also detected differ-ences among sites but not among the times of theyear (Table S3b) Although there was some overlapthe separation of fish communities between the
158
Fig 1 Principal coordinate analyses (PCO) of (a) benthiccommunity cover (b) algal community composition and (c)herbivorous fish community composition from lagoon andback-reef habitats at Ningaloo Reef (M) la goon (S) back-reef Vector overlays represent correlations gt06 based onSpearman ranking CCA crustose coralline algae EAM
epilithic algal matrix
Fig 2 Mean biomass (plusmn SEM) of (a) fleshy algae and (b) her-bivorous fish in the lagoon and back-reef habitats duringspring summer and autumn at Ningaloo Reef Note the logarithmic scale on the y-axis for herbivorous fish biomass
Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Mar Ecol Prog Ser 486 153ndash164 2013
using the same transects in which benthic commu-nity cover and macroalgae were sampled (seeabove) A diver swam a 25 m transect at a constantspeed and counted the abundance and size class (to5 cm) of individual roving nominally herbivorous fish(sensu Choat et al 2002 2004) along 25 m on bothsides of the transect line Territorial damselfish abun-dance and size class were counted on the returnswim along each transect narrowing the width to 1 times25 m length At each site at each time period 4 repli-cate transects were conducted transects were sepa-rated from each other by at least 10 m Abundancedata were converted to biomass using the allometriclength-weight conversions W = a middotTLb where W isweight in g TL is total length in cm and parametersa and b are constants obtained from Fishbase (Froeseamp Pauly 2005) Fish surveys were always conductedat least 2 h after sunrise and 2 h before sunset Indi-viduals were recorded from the 4 major roving herbi-vore families (Acanthuridae Kyphosidae ScaridaeSiganidae) and territorial damselfish (Pomacentri-dae) to the lowest possible taxa (generally to speciesalways to genus) (see Table S1 in the Supplement)
Algal recruitment experimental design
To assess the influence of large herbivorous fish onalgal recruitment in habitats dominated either bymacro algae or coral cages were installed to excludeherbivores at each site-habitat combination We in -stalled 6 plots per treatment per site and these werescrubbed clean once a month throughout the experi-mental period The cages were triangular in shapeand measured 50 times 50 times 50 cm The corners weremarked with rebar to which PVC coated wire meshwas attached The mesh size used was 25 times 25 cmwhich excluded most herbivores but allowed somesmall fishes and invertebrates inside the cages In thecontext of this study large herbivorous fishes aretherefore considered as those excluded by the exper-imental cage mesh size In the caged plots the wirematerial covered all sides and the top of the plots Askirting edge on the sides of the cages was designedto prevent large benthic invertebrates from acces -sing the cages Open (uncaged) plots were markedwith the rebar and partial cages were used as a con-trol for cage artefacts and consisted of one side and aroof to maintain the potential influence of the cagewhile allowing access to all herbivores Some of theplots (70) from all 3 treatments in the back-reefhabitat became inhabited by territorial damselfishThese fish were included in analyses as a covariate
owing to their influence on benthic algal communi-ties (see review by Ceccarelli et al 2001) Urchins arenot abundant on the benthos in the back-reef orlagoon habitats (Johansson et al 2010) and theywere never found in the experimental plots
In each plot 2 recruitment tiles were installed inAugust 2008 and collected in February 2009 Thetiles measured 10 times 10 cm and were made from PVCAlthough PVC is not a natural substrate long-termalgal recruitment experiments have demonstratedthat algal community composition on this material issimilar to communities that recruit on tiles made ofnatural materials such as dead corals (Hixon amp Bros -toff 1996) The tiles were sanded to roughen the sur-faces which created irregular depressions to allowalgal attachment (Hixon amp Brostoff 1996 Smith et al2001 2010) Recruitment tiles were attached to apiece of horizontal wire mesh which was elevated10 cm from the benthos in each plot This standard-ised the attachment substrate and avoided smother-ing of the tiles by sediment
In February 2009 6 mo after deployment the 2 tilereplicates from each plot were carefully removed andplaced in individual Ziploc bags in situ After collec-tion the tiles were frozen until laboratory processingIn the laboratory each tile was thawed and rinsedwith seawater to remove any sediment and anysmall invertebrates were removed with forceps Todetermine the percent cover of the algal communityon the top surface of each tile a point-interceptmethod was used by placing a transparent plasticsheet with a grid of 25 dots on top of each tile Usinga dissecting microscope the algae or substrata pre-sent beneath each point were classified into one ofthe following categories bare tile CCA encrustingfleshy algae turf algae (red green or brown) andfoliose macroalgae (which included erect macro-phytes such as Dictyota Laurencia Lobophora Pad-ina and Sargassum)
Following classification algae from the tile werescraped and oven dried at 60degC for ge48 h after whichthe algae were weighed to 10minus4 g To estimate fleshyalgae biomass (including encrusting fleshy algae turfalgae and foliose macroalgae) HCl (10) was thenadded to the vials to remove any carbonate associatedwith coralline algae Vials were again rinsed placedin the oven at 60degC for ge48 h and re weighed
Data analyses
Separate principal coordinate analyses (PCO)were conducted to visualise differences in benthic
156
Doropoulos et al Herbivore role in coral reef algal recruitment
community cover algal community compositionand fish community composition Vector plots usingSpearman rankings based on correlations gt06 wereoverlaid on PCO plots to visualise relationshipsbetween variables and the ordination axes Differ-ences in total biomass of fleshy algae roving her-bivorous fish and damselfish were tested usingmixed effects analysis of variance (ANOVA) withhabitat (2 levels) and time (3 levels) as fixed factorsand site (2 levels) included as a random factornested in habitat Benthic cover and biomass ofalgal communities and roving herbi vorous fish wereused to assess patterns in their compositions usingseparate mixed effects permutational multivariateanalysis of variance (PERMANOVA) with the samemodel design described above
Mean percent cover on the top surface of the tileswas used to analyse the composition of the recruit-ing algal community Data collected from the repli-cate tiles within each plot were averaged and theaveraged data used as the residuals for all analyses(ie at the plot level) We used a mixed effects permutational multivariate analysis of covariance(PERMANCOVA) Habitat (2 levels) and herbivory(3 levels) were included as fixed factors with site(2 levels) in cluded as a random factor nested inhabitat Pomacentrus alexanderae (damselfish) werein cluded as a covariate because they establishedterritories in 70 of the experimental plots in theback-reef Following this the mean percent cover ofeach individual functional group and total fleshyalgal biomass were analysed using permutationalANCOVA using the same model
All data were evaluated using p-values generatedfrom 9999 permutations using Primer-E v6 (Clarkeamp Gorley 2006) with the PERMANOVA+ add-onpackage (Anderson et al 2008) When the numberof unique permutations was low the Monte Carloasymptotic p-value was used to assess statistical significance (Anderson 2005) Interaction or maineffect terms were pooled when they were highlynon-significant (p gt 025) (Underwood 1997) and significant differences (p lt 005) were investigatedwith pair-wise comparisons Euclidean distancewas the metric used to create resemblance matricesfor all univariate data analyses and Bray-Curtissimilarity was the metric used to create resem-blance matrices for all multivariate data analysesWhen necessary the data were transformed tomeet the assumptions of homogeneity (permuta-tional analysis of multi variate dispersion PERM -DISP) Transformations for relevant data ana ly sesare outlined in Tables S2minusS6 in the Supplement
RESULTS
Benthic community cover algal biomass and algal community composition
Multivariate analysis of the benthic communitycover revealed a significant distinction between thelagoon and back-reef habitats and between summerand autumn in both habitats (Table S2a in the Supp -lement at wwwint-rescomarticles suppl m486 p153_ supp pdf) The community composition was sepa-rated on the PCO1 axis (Fig 1a) which explained915 of the total variation (eigenvalue 49834) andthe PCO2 axis explained 84 of the total variation(eigenvalue 5498) Spearman correlations indicatedthat differences between habitats were clearly drivenby the dominance of foliose macroalgae and sand inthe lagoon and by dominance of live coral and EAMin the back-reef (Fig 1a) The lagoon had an averageof 29 macroalgae 63 sand and lt4 live coralEAM and CCA cover while the back-reef had anaverage of 48 live coral 28 EAM 16 sand 5macroalgae and 2 CCA cover
Total algal biomass showed a significant Habitat timesTime interaction (Table S2b) with biomass greaterin the lagoon compared with the back-reef duringautumn only (Fig 2a Table S2b) There was a mar ked increase in algal biomass in the lagoonfrom 168 g DW (dry weight) mminus2 in spring and sum-mer to 748 g DW mminus2 in autumn Benthic algal community composition displayed significant differ-ences be tween the lagoon and back-reef habitats(Table S2c) Spearman correlation indicated thatSargassum was highly correlated with the lagoonwhile no particular algae were correlated with theback-reef (Fig 1b) Throughout the study foliosealgae in the lagoon were dominated by Sargassum(88 algal community biomass) followed by Hyp-nea (9) In the back-reef Sargassum contri buted46 to the total algal community biomass 25Turbinaria 13 arti culated coralline algae 6Lobophora and 4 Dictyota
Herbivorous fish biomass and community composition
We recorded a total of 15 species of roving herbi -vorous fish from the Acanthuridae Kyphosidae Sca -ri dae and Siganidae families (see Table S1 in theSupplement) Initial phase parrotfish were recor dedas a single group (Scarus IP) Four species of terri -torial Pomacentridae that feed on benthic algae were
157
Mar Ecol Prog Ser 486 153ndash164 2013
recorded Hemiglyphidodon plagiometapon Poma-centrus alexanderae Stegastes fasciolatus and Sobreptus
Total biomass of roving herbivorous fish differedsignificantly between the lagoon and back-reef habi-tats where the biomass in the back-reef was at leastan order of magnitude higher than the lagoon(Fig 2b Table S3a) In contrast to the benthic algalcommunity we detected no significant differences intotal herbivorous fish biomass among times or sites(Table S3a) The herbivorous fish community compo-sition also differed significantly between the lagoonand back-reef habitats and we also detected differ-ences among sites but not among the times of theyear (Table S3b) Although there was some overlapthe separation of fish communities between the
158
Fig 1 Principal coordinate analyses (PCO) of (a) benthiccommunity cover (b) algal community composition and (c)herbivorous fish community composition from lagoon andback-reef habitats at Ningaloo Reef (M) la goon (S) back-reef Vector overlays represent correlations gt06 based onSpearman ranking CCA crustose coralline algae EAM
epilithic algal matrix
Fig 2 Mean biomass (plusmn SEM) of (a) fleshy algae and (b) her-bivorous fish in the lagoon and back-reef habitats duringspring summer and autumn at Ningaloo Reef Note the logarithmic scale on the y-axis for herbivorous fish biomass
Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Doropoulos et al Herbivore role in coral reef algal recruitment
community cover algal community compositionand fish community composition Vector plots usingSpearman rankings based on correlations gt06 wereoverlaid on PCO plots to visualise relationshipsbetween variables and the ordination axes Differ-ences in total biomass of fleshy algae roving her-bivorous fish and damselfish were tested usingmixed effects analysis of variance (ANOVA) withhabitat (2 levels) and time (3 levels) as fixed factorsand site (2 levels) included as a random factornested in habitat Benthic cover and biomass ofalgal communities and roving herbi vorous fish wereused to assess patterns in their compositions usingseparate mixed effects permutational multivariateanalysis of variance (PERMANOVA) with the samemodel design described above
Mean percent cover on the top surface of the tileswas used to analyse the composition of the recruit-ing algal community Data collected from the repli-cate tiles within each plot were averaged and theaveraged data used as the residuals for all analyses(ie at the plot level) We used a mixed effects permutational multivariate analysis of covariance(PERMANCOVA) Habitat (2 levels) and herbivory(3 levels) were included as fixed factors with site(2 levels) in cluded as a random factor nested inhabitat Pomacentrus alexanderae (damselfish) werein cluded as a covariate because they establishedterritories in 70 of the experimental plots in theback-reef Following this the mean percent cover ofeach individual functional group and total fleshyalgal biomass were analysed using permutationalANCOVA using the same model
All data were evaluated using p-values generatedfrom 9999 permutations using Primer-E v6 (Clarkeamp Gorley 2006) with the PERMANOVA+ add-onpackage (Anderson et al 2008) When the numberof unique permutations was low the Monte Carloasymptotic p-value was used to assess statistical significance (Anderson 2005) Interaction or maineffect terms were pooled when they were highlynon-significant (p gt 025) (Underwood 1997) and significant differences (p lt 005) were investigatedwith pair-wise comparisons Euclidean distancewas the metric used to create resemblance matricesfor all univariate data analyses and Bray-Curtissimilarity was the metric used to create resem-blance matrices for all multivariate data analysesWhen necessary the data were transformed tomeet the assumptions of homogeneity (permuta-tional analysis of multi variate dispersion PERM -DISP) Transformations for relevant data ana ly sesare outlined in Tables S2minusS6 in the Supplement
RESULTS
Benthic community cover algal biomass and algal community composition
Multivariate analysis of the benthic communitycover revealed a significant distinction between thelagoon and back-reef habitats and between summerand autumn in both habitats (Table S2a in the Supp -lement at wwwint-rescomarticles suppl m486 p153_ supp pdf) The community composition was sepa-rated on the PCO1 axis (Fig 1a) which explained915 of the total variation (eigenvalue 49834) andthe PCO2 axis explained 84 of the total variation(eigenvalue 5498) Spearman correlations indicatedthat differences between habitats were clearly drivenby the dominance of foliose macroalgae and sand inthe lagoon and by dominance of live coral and EAMin the back-reef (Fig 1a) The lagoon had an averageof 29 macroalgae 63 sand and lt4 live coralEAM and CCA cover while the back-reef had anaverage of 48 live coral 28 EAM 16 sand 5macroalgae and 2 CCA cover
Total algal biomass showed a significant Habitat timesTime interaction (Table S2b) with biomass greaterin the lagoon compared with the back-reef duringautumn only (Fig 2a Table S2b) There was a mar ked increase in algal biomass in the lagoonfrom 168 g DW (dry weight) mminus2 in spring and sum-mer to 748 g DW mminus2 in autumn Benthic algal community composition displayed significant differ-ences be tween the lagoon and back-reef habitats(Table S2c) Spearman correlation indicated thatSargassum was highly correlated with the lagoonwhile no particular algae were correlated with theback-reef (Fig 1b) Throughout the study foliosealgae in the lagoon were dominated by Sargassum(88 algal community biomass) followed by Hyp-nea (9) In the back-reef Sargassum contri buted46 to the total algal community biomass 25Turbinaria 13 arti culated coralline algae 6Lobophora and 4 Dictyota
Herbivorous fish biomass and community composition
We recorded a total of 15 species of roving herbi -vorous fish from the Acanthuridae Kyphosidae Sca -ri dae and Siganidae families (see Table S1 in theSupplement) Initial phase parrotfish were recor dedas a single group (Scarus IP) Four species of terri -torial Pomacentridae that feed on benthic algae were
157
Mar Ecol Prog Ser 486 153ndash164 2013
recorded Hemiglyphidodon plagiometapon Poma-centrus alexanderae Stegastes fasciolatus and Sobreptus
Total biomass of roving herbivorous fish differedsignificantly between the lagoon and back-reef habi-tats where the biomass in the back-reef was at leastan order of magnitude higher than the lagoon(Fig 2b Table S3a) In contrast to the benthic algalcommunity we detected no significant differences intotal herbivorous fish biomass among times or sites(Table S3a) The herbivorous fish community compo-sition also differed significantly between the lagoonand back-reef habitats and we also detected differ-ences among sites but not among the times of theyear (Table S3b) Although there was some overlapthe separation of fish communities between the
158
Fig 1 Principal coordinate analyses (PCO) of (a) benthiccommunity cover (b) algal community composition and (c)herbivorous fish community composition from lagoon andback-reef habitats at Ningaloo Reef (M) la goon (S) back-reef Vector overlays represent correlations gt06 based onSpearman ranking CCA crustose coralline algae EAM
epilithic algal matrix
Fig 2 Mean biomass (plusmn SEM) of (a) fleshy algae and (b) her-bivorous fish in the lagoon and back-reef habitats duringspring summer and autumn at Ningaloo Reef Note the logarithmic scale on the y-axis for herbivorous fish biomass
Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013
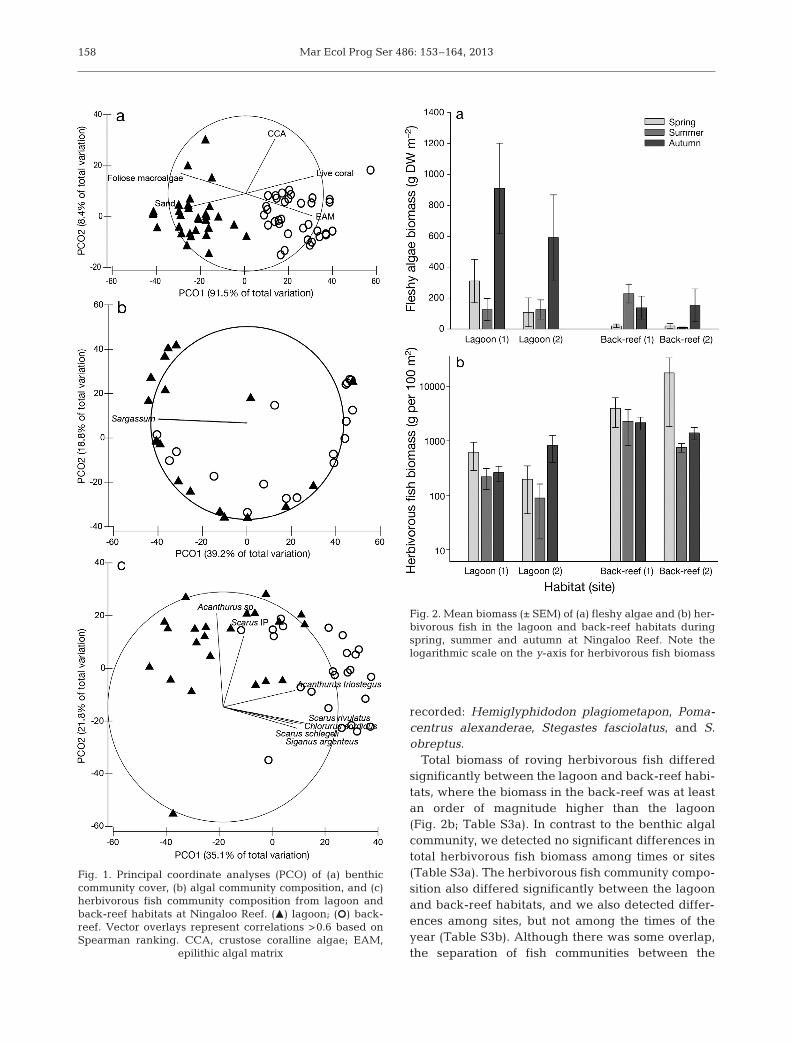
Mar Ecol Prog Ser 486 153ndash164 2013
recorded Hemiglyphidodon plagiometapon Poma-centrus alexanderae Stegastes fasciolatus and Sobreptus
Total biomass of roving herbivorous fish differedsignificantly between the lagoon and back-reef habi-tats where the biomass in the back-reef was at leastan order of magnitude higher than the lagoon(Fig 2b Table S3a) In contrast to the benthic algalcommunity we detected no significant differences intotal herbivorous fish biomass among times or sites(Table S3a) The herbivorous fish community compo-sition also differed significantly between the lagoonand back-reef habitats and we also detected differ-ences among sites but not among the times of theyear (Table S3b) Although there was some overlapthe separation of fish communities between the
158
Fig 1 Principal coordinate analyses (PCO) of (a) benthiccommunity cover (b) algal community composition and (c)herbivorous fish community composition from lagoon andback-reef habitats at Ningaloo Reef (M) la goon (S) back-reef Vector overlays represent correlations gt06 based onSpearman ranking CCA crustose coralline algae EAM
epilithic algal matrix
Fig 2 Mean biomass (plusmn SEM) of (a) fleshy algae and (b) her-bivorous fish in the lagoon and back-reef habitats duringspring summer and autumn at Ningaloo Reef Note the logarithmic scale on the y-axis for herbivorous fish biomass
Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Doropoulos et al Herbivore role in coral reef algal recruitment
lagoon and back-reef habitats was clustered pre-dominantly on the PCO1 axis (Fig 1c) which ex -plained 351 of the variation (eigenvalue 31923)The second PCO axis explained 218 and the thirdaxis explained 136 (eigenvalues 21084 for PCO2and 12520 for PCO3) Acanthurus triostegus Chloru-rus sordidus Scarus rivulatus S schlegeli and Siga -nus argenteus were correlated with the back-reefwhile Acanthurus spp and Scarus IP were correlatedwith both habitats
Total damselfish biomass differed significantly be -tween habitats (F1 = 282 p = 0003) but no significantdifferences were found among time or sites Totaldamselfish biomass per 100 m2 averaged 18 g in thelagoon and 369 g in the back-reef Pomacentrusalexanderae was the only damselfish observed estab-lishing territories in our experimental plots in back-reef habitats and was observed indiscriminatelyestablishing territories in all cage treatments (openpartial closed)
Recruitment tile algal community compositionand biomass
The community composition of the algae growingon the recruitment tiles was not significantly influ-enced by habitat but was instead strongly affectedby the experimental herbivore exclusion treatments(Table S4) Algal community composition was signif-icantly different between tiles ex cluded from rovingherbivores (closed treatments) compared with thoseexposed to roving herbivores (open and partial treat-ments post-hoc analysis Closed ne Open = Partial)These differences were characterised by signifi-cantly higher cover of foliose macroalgae (Fig 3b)and bare tile (Fig 3a) in the roving herbivore exclu-sion treatment in both habitats (Table S5) The pres-ence of Pomacentrus alexanderae damselfish signifi-cantly affec ted the community composition of thealgae present on the recruitment tiles (Table S4) Palexanderae territories were only established at plotsin the back-reef sites where their presence signifi-cantly in creased turf cover (Fig 3d) and decreasedCCA cover (Fig 3c) irrespective of roving herbivoreexclusion treatment (Table S5) There was significantvariability among sites in the multivariate analysis(Table S5) and for the CCA turf and foliose macro -algae cover in the univariate analyses (Table S5)
Following a similar pattern to total algal cover thebiomass of fleshy algae was significantly higher ontiles excluded from roving herbivores (closed treat-ments) compared with those exposed to roving herbi-
vores (open and partial treatments post-hoc analysisClosed ne Open = Partial) in both lagoon and back-reef habitats (Fig 3f Table S6) We found no signifi-cant differences in total algal biomass between habi-tats and high variability among sites within eachhabitat (Table S6)
DISCUSSION
Our study has highlighted 3 important aspects ofalgal dynamics in a relatively intact coral reef eco -system First contrary to our first hypothesis wefound no habitat effects on the recruiting algal com-munity Our a priori expectation was that algal re -cruitment on the experimental tiles would differbetween habitats due to large differences in the ben-thic community and herbivorous fish assemblagesdescribed in Vergeacutes et al (2011) Instead we foundsimilar recruiting algal communities in the 2 habitatsdespite large differences in the adult establishedcommunities of macroalgae Given the close proxi -mity of the 2 neighbouring habitats and the watermovement between the habitats that supports theexchange of particulate matter from the back-reef tothe lagoon (Wyatt et al 2010) this similarity inrecruitment is likely to be due to a comparable sup-ply of propagules Second in partial support of oursecond hypothesis algal recruitment was highlyinfluenced by herbivores but this effect was surpris-ingly similar for algal communities in both lagoonand back-reef habitats despite major differences intheir respective herbivorous fish communities withback-reef herbivorous fish biomass being an order ofmagnitude greater than in the lagoon Finally al -though we detected clear and constant differencesbetween lagoon and back-reef algal communitycomposition differences in algal biomass and coverwere restricted to certain times of the year
Similar to other recruitment studies that have iden-tified the role of roving herbivores in limiting foliosemacroalgal abundance in coral-dominated habitats(Belliveau amp Paul 2002 Diaz-Pulido amp McCook 2003Burkepile amp Hay 2006 Albert et al 2008) large her-bivores significantly influenced early algal commu-nity structure in both lagoon and back-reef habitatsof Ningaloo Reef Even though roving herbivoreabundance was significantly lower in the lagooncompared with the back-reef when herbivores wereallowed access to graze the recruitment tiles therewas no difference in algal cover or biomass betweenhabitats and when large roving herbivores wereexcluded macroalgal cover and biomass similarly
159
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013
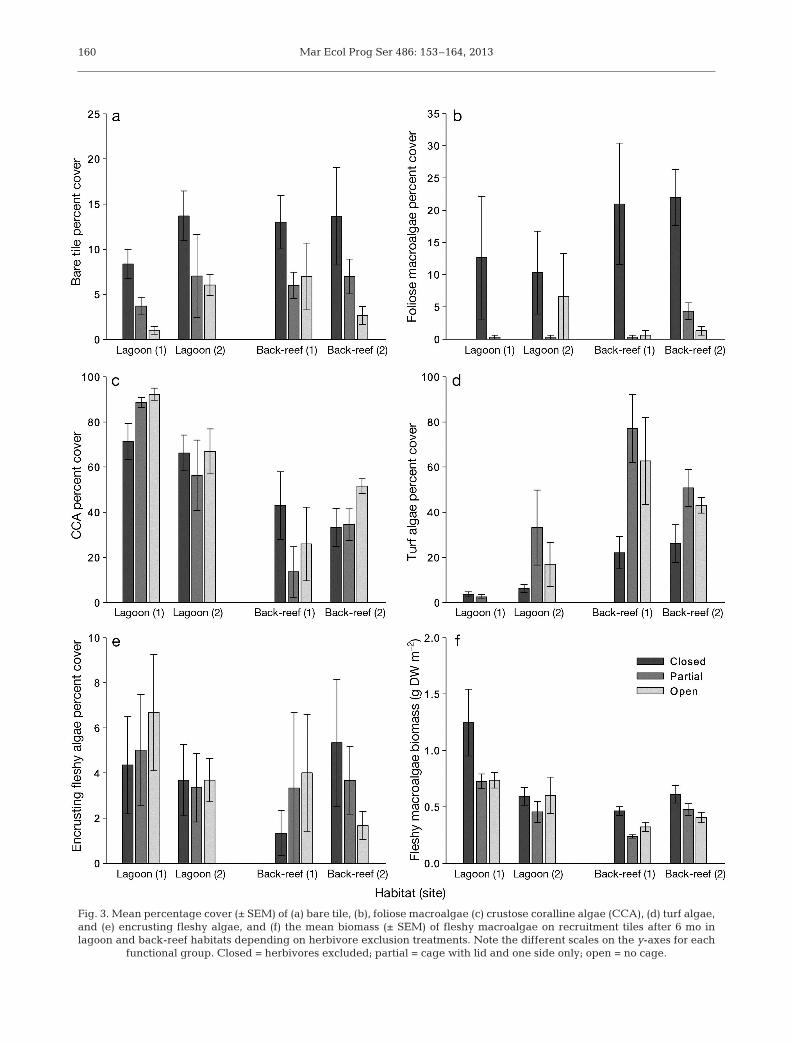
Mar Ecol Prog Ser 486 153ndash164 2013160
Fig 3 Mean percentage cover (plusmn SEM) of (a) bare tile (b) foliose macroalgae (c) crustose coralline algae (CCA) (d) turf algaeand (e) encrusting fleshy algae and (f) the mean biomass (plusmn SEM) of fleshy macroalgae on recruitment tiles after 6 mo inlagoon and back-reef habitats depending on herbivore exclusion treatments Note the different scales on the y-axes for each
functional group Closed = herbivores excluded partial = cage with lid and one side only open = no cage
Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Doropoulos et al Herbivore role in coral reef algal recruitment
increased in both habitats Although there was atrend towards higher biomass of foliose algae on therecruitment tiles in the lagoon this was not statisti-cally different at this early stage of development Theamount of bare space was higher when herbivoreswere excluded suggesting that perhaps the foliosealgae found on the recruitment tiles limited thegrowth of other algae (eg encrusting algae CCAturf) This may be due to competitive forces such asabrasion or shading as has been demonstrated whentemperate kelp limits algal understory growth (Ken -drick et al 1999) Our study clearly demonstrates thatlarge herbivorous fish play an important role in struc-turing the community composition of algal recruits inboth herbivore-rich and herbivore-depauperate coralreef habitats
While large roving herbivorous fish similarly affec -ted the cover and biomass of fleshy macroalgalrecruits on the experimental tiles in both habitatscomposition of algal recruits on the tiles was differ-entially influenced in the lagoon and back-reef bythe presence of smaller territorial damselfish in theexperimental plots located in the back-reef The pre -sence of territorial damselfish (Pomacentrus alexan-derae) increased turf algal cover however we cannotdistinguish whether this was caused by direct (ieconsumptive) or indirect (eg through increasednutrient supply via faeces) effects Other studieshave demonstrated that large roving herbivores havea significant influence on the broad benthic algalcommunity structure (Mumby et al 2006 Paddack etal 2006 Fox amp Bellwood 2007 Wismer et al 2009Vergeacutes et al 2011) while the farming practices of ter-ritorial damselfish can instead influence turf produc-tivity and diversity at smaller scales (Klumpp et al1987 Ceccarelli et al 2005 Gobler et al 2006 Hoeyamp Bellwood 2010b) Territorial damselfish often chaseaway larger roving herbivores to protect the turfalgal communities that they farm influencing pat-terns of algal recruitment and succession (Hixon ampBrostoff 1983 1996 Ceccarelli et al 2001 2006) Thefarming behaviour of damselfish also generally re -duces the establishment of large foliose macroalgaethrough their selective removal from the EAM (egHixon amp Brostoff 1996 Ceccarelli et al 2001 Gobleret al 2006 but see Hoey amp Bellwood 2010b) Al -though we did not quantify their effect on the estab-lished algal community the presence of territorialdamselfish in the back-reef sites presumably influ-enced the high EAM cover in that habitat Thesepatches add to the spatial heterogeneity of algae incoral-dominated reef habitats providing an impor-tant source of nutrition that supports a large portion
of the secondary production in reef systems (Smith etal 2001 Wilson et al 2003)
Although benthic algal community compositiondiffered markedly between habitats throughout the6 mo period total algal biomass and algal cover onlydiffered between habitats during the autumn whichwas characterised by a major increase in macroalgaein the lagoon Similar to studies from the Great Bar-rier Reef that find seasonal increases in algal cover inautumn (Schaffelke amp Klumpp 1997 Lefevre amp Bell-wood 2010) the increase of macroalgae found in thelagoon at Ningaloo Reef was dominated by Sargas-sum However the patterns observed in adult algalcommunities are not what we expected based on theresults from our recruitment experiment It appearsthat although the herbivorous fish found in thelagoon were able to limit the recruitment of foliosealgae in experimental plots they were not able toprevent the marked increase in Sargassum biomassobserved in autumn This may be due to a number ofreasons On the one hand algae may be escapingherbivory through size effects as recruits grow intoadult algal thalli they may become tougher harderless nutritious or more easily recognised as unpalat-able (Duffy amp Hay 1990) Indeed a previous studyfrom the same location shows that herbivores havenegligible effects on dominant adult algae such asSargassum myriocystum and Lobophora variegata inlagoon habitats (Vergeacutes et al 2011) On the otherhand the inability of lagoon herbivores to constrainthe periodic increase of adult foliose algae may bedue to a lsquoresource saturationrsquo effect comparable tothe predator-saturation hypothesis that explainsmass seeding events in plants (Janzen 1974 1976)Under conditions of high algal productivity herbi-vores may become satiated and unable to controlalgal growth a process that has also been suggestedto explain the limited role of herbivores in marginalcoral reefs with high algal densities (Bennett et al2010) In contrast the abundance and diversity of theherbivore community found in the back-reef habitatappears sufficient to consistently limit macroalgalgrowth Therefore it appears that while the herbivo-rous fish community in the lagoon was capable oflimiting foliose algae on the settlement tiles therewas a periodic increase in algal growth that escapedthis top-down effect
It is interesting to note that while the intense her-bivory found in the back-reef at Ningaloo Reef(Johansson et al 2010 Vergeacutes et al 2011) consis-tently limited macroalgal abundance throughout theperiod of this study it may also drive the higher algaldiversity found in that habitat compared with the
161
Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Mar Ecol Prog Ser 486 153ndash164 2013
lagoon Although algae were only identified at acoarse taxonomic level (genus or functional group)we were nevertheless able to identify clear differ-ences in diversity among habitats with 12 macro -algal taxa inhabiting the coral-dominated back-reefversus only 7 in the lagoon The enhanced algal di -versity found in the back-reef is likely to be driven bythe intense herbivory of roving herbivores and abun-dant damselfish following the intermediate distur-bance hypothesis (Hixon amp Brostoff 1983 1996) Ad -ditionally many of the taxa found in the back-reefhave physical or chemical defences (eg corallinealgae Dictoyota Lobophora Turbinaria) (Steinbergamp Paul 1990 Mumby et al 2005 Bittick et al 2010)as a means of deterring grazing from roving herbi -vorous fish
In our study algal communities varied in lagoonand back-reef habitats at different spatial and tem-poral scales Surprisingly despite large differencesin the herbivorous fish communities between lagoonand back-reef habitats at Ningaloo Reef we foundthat grazing from large roving herbivores equallyaffected algal recruitment in both habitats This sug-gests that relatively depauperate herbivorous fishcommunities such as those found in marginal or over-fished coral reefs may be enough to control algalrecruitment to the same levels as found in communi-ties characterised by diverse and abundant herbivo-rous fish However our results show that when her-bivorous fish biomass and diversity are low algaethat are established within a habitat are able to es -cape herbivore control during periods of maximumalgal growth In this context the role of browsers ieherbivorous fishes that are able to consume adultmacroalgae emerges as particularly important in theprevention of shifts towards macroalgal dominanceOur findings are therefore consistent with a growingnumber of studies that recommend the developmentof coral reef management and conservation pro-grammes that aim to specifically protect dominantbrowsing fishes (eg Naso unicornis Kyphosus spp)(Hoey amp Bellwood 2009 Vergeacutes et al 2012 Michaelet al 2013)
Acknowledgements We thank M J Anderson and J COrtiz for statistical advice and G Roff for thoughtful discus-sions and critical comments to the manuscript Four anony-mous reviewers provided critical comments that greatly im -proved the manuscript We are grateful to S Bennett ABostock J Eyres K Inostroza A Lemmon A Michael PMichael A Richards and J Santana and for their assistancewith field and laboratory work This study was funded bythe Western Australian Marine Science Institute (WAMSI)under the Ningaloo Research Program and the Department
of Environment and Conservation of Western Australia(DEC) provided the permits under which all work was con-ducted (CE002084 CE006457) This is publication 96 fromthe Sydney Institute of Marine Science
LITERATURE CITED
Albert S Udy J Tibbetts IR (2008) Responses of algal com-munities to gradients in herbivore biomass and waterquality in Marovo Lagoon Solomon Islands Coral Reefs27 73minus82
Anderson MJ (2005) PERMANOVA a FORTRAN computerprogram for permutational analysis of variance Depart-ment of Statistics University of Auckland
Anderson MJ Gorley RN Clarke KR (2008) PERMANOVA+for PRIMER guide to software and statistical methodsPRIMER-E Plymouth
Belliveau SA Paul VJ (2002) Effects of herbivory and nutri-ents on the early colonization of crustose coralline andfleshy algae Mar Ecol Prog Ser 232 105minus114
Bellwood DR Choat JH (1990) A functional-analysis of gra -zing parrotfishes (Family Scaridae) the ecological impli-cations Environ Biol Fishes 28 189minus214
Bellwood DR Hughes TP Folke C Nystrom M (2004) Con-fronting the coral reef crisis Nature 429 827minus833
Bennett S Bellwood DR (2011) Latitudinal variation inmacroalgal consumption by fishes on the Great BarrierReef Mar Ecol Prog Ser 426 241minus252
Bennett S Vergeacutes A Bellwood DR (2010) Branching coral asa macroalgal refuge in a marginal coral reef systemCoral Reefs 29 471minus480
Birrell CL McCook LJ Willis BL Diaz-Pulido GA (2008)Effects of benthic algae on the replenishment of coralsand the implications for the resilience of coral reefsOceanogr Mar Biol Annu Rev 46 25minus63
Bittick SJ Bilotti ND Peterson HA Stewart HL (2010)Turbinaria ornata as an herbivory refuge for associatealgae Mar Biol 157 317minus323
Burkepile DE Hay ME (2006) Herbivore vs nutrient controlof marine primary producers context-dependent effectsEcology 87 3128minus3139
Burkepile DE Hay ME (2010) Impact of herbivore identityon algal succession and coral growth on a Caribbeanreef PLoS ONE 5 e8963
CALM (Department of Conservation and Land Manage-ment) MPRA (Marine Parks and Reserves Authority)(2005) Management plan for the Ningaloo Marine Parkand Muiron Islands Marine Management Area 2005minus2015 management plan number 52 Government ofWestern Australia Perth
Cassata L Collins LB (2008) Coral reef communities habi-tats and substrates in and near sanctuary zones of Nin -galoo Marine Park J Coast Res 24 139minus151
Ceccarelli DM Jones GP McCook LJ (2001) Territorialdamselfishes as determinants of the structure of benthiccommunities on coral reefs Oceanogr Mar Biol AnnuRev 39 355minus389
Ceccarelli DM Jones GP McCook LJ (2005) Foragers versusfarmers contrasting effects of 2 behavioural groups ofherbivores on coral reefs Oecologia 145 445minus453
Ceccarelli DM Hughes TP McCook LJ (2006) Impacts ofsimulated overfishing on the territoriality of coral reefdamselfish Mar Ecol Prog Ser 309 255minus262
Cheal A MacNeil M Cripps E Emslie M Jonker M Schaf-
162
Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Doropoulos et al Herbivore role in coral reef algal recruitment
felke B Sweatman H (2010) Coralminusmacroalgal phaseshifts or reef resilience links with diversity and func-tional roles of herbivorous fishes on the Great BarrierReef Coral Reefs 29 1005minus1015
Choat J Clements K Robbins W (2002) The trophic status ofherbivorous fishes on coral reefs I Dietary analyses MarBiol 140 613minus623
Choat JH Robbins WD Clements KD (2004) The trophic sta-tus of herbivorous fishes on coral reefs II Food proces -sing modes and trophodynamics Mar Biol 145 445minus454
Clarke KR Gorley RN (2006) PRIMER v6 Plymouth MarineLaboratory Plymouth
Collins LB Zhu ZR Wyrvoll KH Eisenhauer A (2003) LateQuaternary structure and development of the northernNingaloo Reef Australia Sediment Geol 159 81minus94
Connell JH Hughes TE Wallace CC Tanner JE Harms KEKerr AM (2004) A long-term study of competition anddiversity of corals Ecol Monogr 74 179minus210
Diaz-Pulido G McCook LJ (2002) The fate of bleachedcorals patterns and dynamics of algal recruitment MarEcol Prog Ser 232 115minus128
Diaz-Pulido G McCook LJ (2003) Relative roles of herbivoryand nutrients in the recruitment of coral-reef seaweedsEcology 84 2026minus2033
Done TJ (1992) Phase-shifts in coral-reef communities andtheir ecological significance Hydrobiologia 247 121minus132
Duffy JE Hay ME (1990) Seaweed adaptations to herbivoy Chemical structural and morphological defenses areoften adjusted to spatial or temporal patterns of attackBioscience 40 368minus375
Fox RJ Bellwood DR (2007) Quantifying herbivory across acoral reef depth gradient Mar Ecol Prog Ser 339 49minus59
Froese R Pauly D (eds) (2005) Fishbase wwwfishbaseorgGobler CJ Thibault DB Davis TW Curran PB Peterson BJ
Liddle LB (2006) Algal assemblages associated with Ste-gastes sp territories on Indo-Pacific coral reefs charac-terization of diversity and controls on growth J Exp MarBiol Ecol 336 135minus145
Green AL Bellwood DR (2009) Monitoring functionalgroups of herbivorous fishes as indicators for coral reefresilience a practical guide for coral reef managers inthe Asia Pacific region IUCN Working Group on ClimateChange and Coral Reefs IUCN Gland
Hay ME (1981) Spatial patterns of agrazing intensity on aCaribbean barrier reef herbivory and algal distributionAquat Bot 11 97minus109
Hay ME Colburn T Downing D (1983) Spatial and temporalpatterns in herbivory on a Caribbean fringing reef theeffects of plant distribution Oecologia 58 299minus308
Hixon MA Brostoff WN (1983) Damselfish as keystone spe-cies in reverse intermediate disturbance and diversity ofreef algae Science 220 511minus513
Hixon MA Brostoff WN (1996) Succession and herbivory effects of differential fish grazing on Hawaiian coral-reefalgae Ecol Monogr 66 67minus90
Hoey AS Bellwood DR (2009) Limited functional redun-dancy in a high diversity system single species domi-nates key ecological process on coral reefs Ecosystems12 1316minus1328
Hoey AS Bellwood DR (2010a) Among-habitat variation inherbivory on Sargassum spp on a mid-shelf reef in thenorthern Great Barrier Reef Mar Biol 157 189minus200
Hoey AS Bellwood DR (2010b) Damselfish territories as arefuge for macroalgae on coral reefs Coral Reefs 29 107minus118
Hoey AS Bellwood DR (2011) Suppression of herbivory bymacroalgal density a critical feedback on coral reefsEcol Lett 14 267minus273
Hughes TP (1994) Catastrophies phase-shifts and large-scale degradation of a Caribbean coral-reef Science 265 1547minus1551
Hughes TP Rodrigues MJ Bellwood DR Ceccarelli D andothers (2007) Phase shifts herbivory and the resilienceof coral reefs to climate change Curr Biol 17 360minus365
Janzen DH (1974) Tropical blackwater rivers animals andmast fruiting by the Dipterocarpaceae Biotropica 6 69minus103
Janzen DH (1976) Why bamboos wait so long to flowerAnnu Rev Ecol Syst 7 347minus391
Johansson CL Bellwood DR Depczynski M (2010) Seaurchins macroalgae and coral reef decline a functionalevaluation of an intact reef system Ningaloo WesternAustralia Mar Ecol Prog Ser 414 65minus74
Jompa J McCook LJ (2002) Effects of competition and her-bivory on interactions between a hard coral and a brownalga J Exp Mar Biol Ecol 271 25minus39
Kendrick GA Walker DI (1995) Dispersal of propagules ofSargassum spp (Sargassaceae Phaeophyta) observa-tions of local patterns of dispersal and consequences forrecruitment and population structure J Exp Mar BiolEcol 192 273minus288
Kendrick GA Lavery PS Phillips JC (1999) Influence ofEcklo nia radiata kelp canopy on structure of macro-algalassemblages in Marmion Lagoon Western AustraliaHydrobiologia 398minus399 275minus283
Klumpp DW McKinnon D Daniel P (1987) Damselfish terri-tories zones of high productivity on coral reefs Mar EcolProg Ser 40 41minus51
Lefevre CD Bellwood DR (2010) Seasonality and dynamicsin coral reef macroalgae variation in condition and sus-ceptibility to herbivory Mar Biol 157 955minus965
Littler MM Littler DS (1999) Disturbances due to CycloneGavin parallel those caused by a ship grounding CoralReefs 18 146
Lubchenco J Gaines SD (1981) A unified approach tomarine plant-herbivore interactions I Populations andcommunities Annu Rev Ecol Syst 12 405minus437
Marshell A Mumby PJ (2012) Revisiting the functional rolesof the surgeonfish Acanthurus nigrofuscus and Cteno -chaetus striatus Coral Reefs 31 1minus9
McClanahan TR Sala E Stickels PA Cokos BA Baker ACStarger CJ Jones SH (2003) Interaction between nutri-ents and herbivory in controlling algal communities andcoral condition on Gloverrsquos Reef Belize Mar Ecol ProgSer 261 135minus147
Michael PJ Hyndes GA Vanderklift MA Vergeacutes A (2013)Identity and behaviour of herbivorous fish influencelarge-scale spatial patterns of macroalgal herbivory in acoral reef Mar Ecol Prog Ser 482227ndash240
Mumby PJ Foster NL Fahy EAG (2005) Patch dynamics ofcoral reef macroalgae under chronic and acute distur-bance Coral Reefs 24 681minus692
Mumby PJ Dahlgren CP Harborne AR Kappel CV and oth-ers (2006) Fishing trophic cascades and the process ofgrazing on coral reefs Science 311 98minus101
OrsquoLeary JK Potts DC Braga JC McClanahan TR (2012)Indirect consequences of fishing reduction of corallinealgae suppresses juvenile coral abundance Coral Reefs31 547minus559
Paddack MJ Cowen RK Sponaugle S (2006) Grazing pres-
163
Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013

Mar Ecol Prog Ser 486 153ndash164 2013
sure of herbivorous coral reef fishes on low coral-coverreefs Coral Reefs 25 461minus472
Rasher DB Engel S Bonito V Fraser GJ Montoya JP HayME (2012) Effects of herbivory nutrients and reef pro-tection on algal proliferation and coral growth on a trop-ical reef Oecologia 169 187minus198
Roff G Mumby PJ (2012) Global disparity in the resilience ofcoral reefs Trends Ecol Evol 27 404minus413
Santelices B (1990) Patterns of reproduction dispersal andrecruitment in seaweeds Oceanogr Mar Biol Annu Rev28 177minus276
Schaffelke B Klumpp DW (1997) Biomass and productivityof tropical macroalgae on three nearshore fringing reefsin the central Great Barrier Reef Australia Bot Mar 40 373minus383
Smith JE Smith CM Hunter CL (2001) An experimentalanalysis of the effects of herbivory and nutrient enrich-ment on benthic community dynamics on a Hawaiianreef Coral Reefs 19 332minus342
Smith JE Hunter CL Smith CM (2010) The effects of top-down versus bottom-up control on benthic coral reefcommunity structure Oecologia 163 497minus507
Steinberg PD Paul VJ (1990) Fish feeding and chemicaldefenses of tropical brown algae in Western AustraliaMar Ecol Prog Ser 58 253minus259
Underwood AJ (1997) Experiments in ecology their logisti-cal design and interpretation using analysis of varianceCambridge University Press Cambridge
Vergeacutes A Vanderklift MA Doropoulos C Hyndes GA(2011) Spatial patterns in herbivory on a coral reef areinfluenced by structural complexity but not by algaltraits PLoS ONE 6 e17115
Vergeacutes A Bennett S Bellwood DR (2012) Diversity amongmacroalgae-consuming fishes on coral reefs a transcon-tinental comparison PLoS ONE 7 e45543
Vroom PS Braun CL (2010) Benthic composition of a healthysubtropical reef baseline species-level cover with anemphasis on algae in the northwestern Hawaiian Is -lands PLoS ONE 5 e9733
Wilson SK Bellwood DR Choat JH Furnas MJ (2003) Detri-tus in the epilithic algal matrix and its use by coral reeffishes Oceanogr Mar Biol Annu Rev 41 279minus309
Wismer S Hoey AS Bellwood DR (2009) Cross-shelf benthiccommunity structure on the Great Barrier Reef relation-ships between macroalgal cover and herbivore biomassMar Ecol Prog Ser 376 45minus54
Wyatt ASJ Lowe RJ Humphries S Waite AM (2010) Partic-ulate nutrient fluxes over a fringing coral reef relevantscales of phytoplankton production and mechanisms ofsupply Mar Ecol Prog Ser 405 113minus130
164
Editorial responsibility Kenneth Heck Jr Dauphin Island Alabama USA
Submitted October 9 2012 Accepted March 9 2013Proofs received from author(s) June 18 2013





























