Embed Size (px)
Citation preview

Algae (The Korean Journal of Phycology)Volume l2(3): 1,47 -157, 7997
lReview Articlel
Algal Phenotypic Plasticity: Its Importance in Developing New ConceptsThe Case for Scenedesmus
Eduardo A. Morales* and Francis R. Trainor
Depnrtment of Ecology and Euoltttionary Biology, Uniuersity of ConnectiuttStorrs, CT06269-3043, USA
"The scientist's world is qualitatively transformed as well as quantitatively enriched by funda-mental novelties of either fact or theorv." T. Ku}rrr
The phycological literature contains several accounts for the occurrence of phenotypic plasticity, briefly defined as
morphological variations induced by changing environmental conditions. However, the potential impact of plastici-ty on algal systematics has been underestimated. Irr the past, conventional clissification schemes have not been suf-
ficiently flexible to incorporate a rather extensive phenotypic plasticity, resulting in the application of differentnames to morphs (ecomorphs) of the same species Research on the green algal genus Scenedesmus has demonstrated
the impact of this morphological plasticity on taxonomy at the species level, suggesting possible pathways toward
the development of a more natural classification system based on multidisciplinary information.
Key Words: algae, phenotypic plasticity, Scenetlesmus, taxonomy and systematrcs
INTRODUCTION
One of the paramount problems in phycology is the
polyphyletic (several origins) character of the aIgae,
which leads to a great diversity of forms, physiologies,
and ways to deal with the nutritional environment. Such
diversity led early phycologists to develop techniques
providing analysis of the greatest number of specimens
in the shortest time possible. Thus it was - and still is -an accepted procedure to use preserved material in the
description of new species. With little recognition and
acceptance of developments in other sciences such as
ecology and the culture of organisms in the laboratory,
algal taxonomy and systematics matured within a rigidframework focused on "appearance", and thus lacked
concepts of "function". Now we appreciate the enor-
mous value of laboratory algal cultures for the investiga-
tion of life histories and physiological responses of the
organisms, including recognition of phenotypic plastici-
ty in algae, a common phenomenon that traditionai field
nAddress for reprint request (eam [email protected]
edu).
techniques hid from us.
Some terminology and examples of phenotypic plastic-
itySeveral terms have been developed to differentiate
among types of phenotypic plasticity and distinguish itfrom other responses of the genotype, not necessarily
induced by environmentai factors. Phycologists do not
usually refer to particular types of phenotypic plasticity,
but rather make use of this term in a general sense. The
most accepted definition seems to be the capnbility oJ a
genotype to produce sezteral phenotypes tmder changing enut-
ronmentnl conditiotts. The environment plays a key role in
being the changing variable inducing the genotype to
adapt physiologically to the new set of conditioris in the
medium, Environmental stimuli may include physical,
chemical, and biological parameters. The resulting phys-
iological adaptation is translated into a change in the
appearance of the organism.
One of the most common examples of phenotypic
plasticity described in the literature in cyclomorphosis' Ithas been defined as cyclic changes in the phenotype of a par-
ticular genotype (orgnnism) triggered by the presence of a
predator. Such phenotypic variation is actually the resuit

148 Algae Yol. 72(3), 7997
of the interaction of organic substances (released bypredators to the medium), and the genotype of the cyclo-morphic individual. The examples oi Ceratitrm andDaphnia are classic within this category (Black andSIob odkin 1 98 7; Dodso n 19 89 ; Hutchinson 19 67 ; T olluian7994).
The ordered sequence of ecomorph formation in species ofScerLedesmus constitutes another example of phenotypicplasticity, very closely related to cyclomorphosis (see
later). Although a cyclic sequence of phenotypes may be
produced, each one of them is able to restart thesequence by producing unicells if favorable conditionsare given. This peculiarity has not been described forcyclomorphic changes in either Ceratium or Daphnia,butthe hypothesis may be raised. Phenotypic plasticity inScenedesmus may be predator or physico-chemicallyinduced (Hessen and van Donk 1993; Trainor 1963,7995;
Trainor and Hilton 1963). Accordingly, a more broaddefinition of cyclomorphosis may be in order (Black and
Slobodkin 1987).
Another very closely related term is polyphenism, a
more general term which may be synonymous with phe-notypic plasticity. Stearns (1989) defines it as the produc-
tion of truo or more discrete phenotypes by n single genotype,
in response to an enaironmentnl signnl.
The occurrence of phenotype changes in other groups
of algae has not received close attention, and these
occurrences have been regarded simply as cases of phe-
notypic plasticity. For example, salinity-related morpho-Iogical changes in the centric diatom Cyclotella (Schultz
7971), changes in chrysophyte scale morphology trig-gered by silica availability (Sandgren et nl.1996), changes
in the stipe morphology of some brown algae (Chapman
1973), morphological changes related to daylength and
temperature in red algae (Carbary et aL 1978), consider-
able morphological differences in xenic and axenic cul-tures of green algae (Bonneau 1977; Nakanishi andNishijima 7996), etc.
In one sense, the most common example of phenotypicplasticity is exemplifiedby alternntion of generations rn
various plants. Such is the case of the red aigae belong-
ing to the genus Bangin. In the life history of severalspecies of Bangia there are two phases. One is character-
rzedby a macroscopic foliose thallus, and the second bymicroscopic filaments. Before discovery of the connec-
tion between these two phases (Drew 1949), each was
known as a different genus: Bangia and Conchocelis,
respectively. Subsequent studies dealing with otherspecies demonstrated that Conchocelis was but a stage
through which several species of Bangia pass duringtheir life histories (Conway and Cole 1977).In 1969,
Dixon and Richardson (1969) determined that the change
from one phase to the other was influenced by the pho-
toperiod. Later studies concluded that species in othergenera (such as Porphyra) share "conchocelis" stages
(Fukuhara 1968), just as various Scenedesmus species pass
through the quadricaudate phase (Trainor 1991).
Although some investigators refer to alternation ofheteromorphic generations in the algae as ploidy effects,
there is no evidence showing that experimental changes
in the ploidy of gametophytes would produce sporo-ph)rtic individuals. Evidence indicates that rather thanploidy effects, phase specific cDNAs (derived frommRNA) might be controlling polymorphic differencesbetween gametophytes and sporophytes (Liu et aI. 1994).
Then, it might be better to regard variations betweenheteromorphic generations as physiological (Kain 1996).
As discussed previously in this paper, phenotypic plas-
ticity changes are the outcome of physiological adapta-
tions to changing envirclnments.
Another example of phenotypic plasticity is the lifehistory of ferns and related taxa. In the specific case ofEqtrisetum, the sporophyte phase is the one used in the
identification of the organism, but a gametophyte phase
also exists, the prothallus, completely different in mor-
phology and capable of surviving by itself.
Within higher plants, one of the classical examples ofphenotypic plasticity is Ihe dimorphisnt of leaaes ol the
aquatic p),anl Proserpinaca. DeepIy lobed leaves are pro-duced when individuals grow underwater. When the
piant reaches the water surface and grows above it, ser-
rate leaves are produced. Morphology is determined by
the environment.Embryology and studies related to growth and devel-
opment in higher plants have demonstrated that embry-onic and meristematic cells in animals and plants are
capable of a high degree of plasticity. In plants, differen-tiation of these cells may produce any cell to be found inadult tissues (totipotency). Biotechnological investiga-
tions have revealed that some plant cells are able torestart their meristematic ability to produce a complete
new individual.There are additional examples in the animal kingdom.
Within reptiles for example, Ford and Seigel (1989)
dernonstrated the occurrence of phenotypic plasticity insome reproductive traits of the snake Thamnophis mar-
cinnus. Number of offspring and clutch mass were signif-icantly affected by changes in food availability. This par-
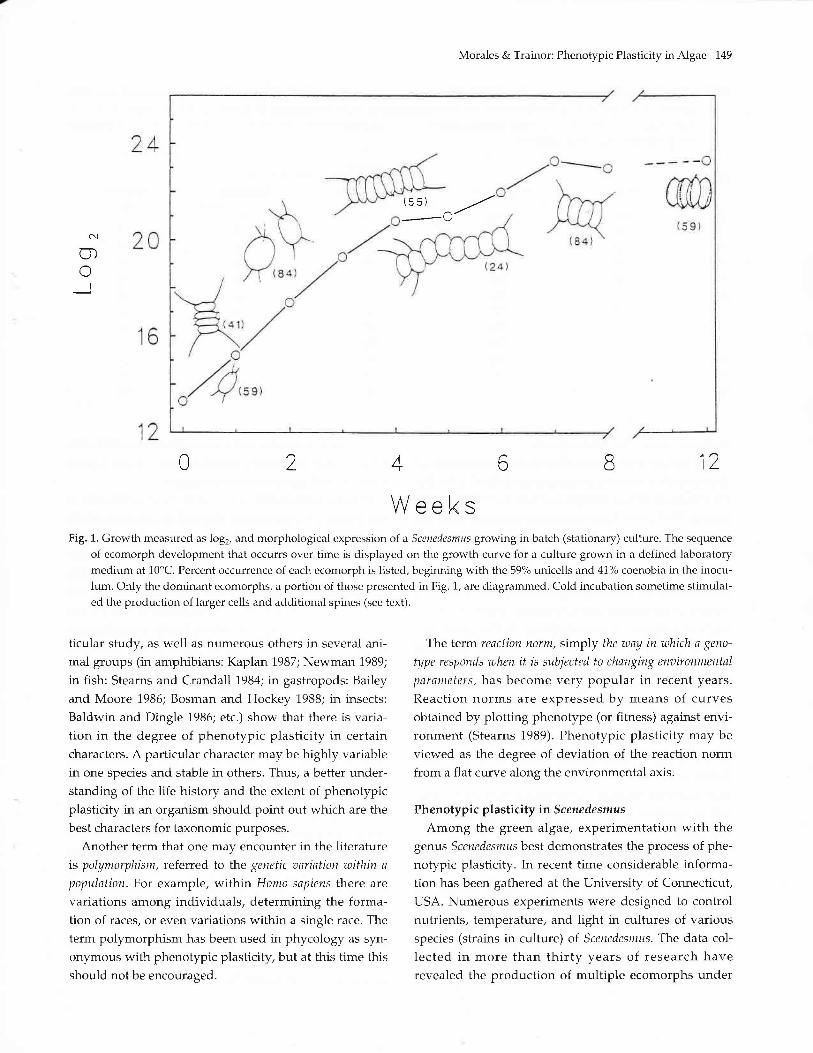
Morales & Trainor: Phenotypic Plasticity in Algae 149
N
O)o
J
12
WeeksFig. 1. Growth measured as log2, and morphological expression oI a Scenedesmtts growtng in batch (stationary) culture. The sequence
of ecomorph development that occurrs over time is displayed on the growth curve for a culture grown in a defined laboratorymedium at 10"C. Percent occurrence of each ecomorph is listed, beginning with the 59% urLicells and 41% coenobia in the inocu-lum. Only the dominant ecomorphs, a portion of those presented in Fig. 1, are diagrammed. Cold incubation sometime stimulat-ed the production of larger cells and additional spines (see text).
B6/'-.r2tl
ticular study, as well as numerous others in several ani-
mal groups (in amphibians: Kaplan 1987; Newman L989;
in fish: Stearns and Crandail 1984; in gastropods: Bailey
and Moore 1986; Bosman and Hockey 1988; in insects:
Baldwin and Dingle 1986; etc.) show that there is varia-tion in the degree of phenotypic plasticity in certaincharacters. A particular character may be highly variablein one species and stable in others. Thus, a better under-standing of the life history and the extent of phenotypicplasticity in an organism should point out which are the
best characters for taxonomic purposes.
Another term that one may encounter in the literature
rs polymorphism, referred to lhe genetic unriation within a
population. For example, within Homo sapiens there are
variations among individuals, determining the forma-
tion of races, or even variations within a single race. The
term polymorphism has been used in phycology as syn-
onymous with phenotypic plasticity, but at this time thisshould not be encouraged.
The term reactiott norm, simply the way in zuhich a geno-
type responds ruhen it is subjected to changing enaironmental
parameters, has become very popular in recent years.
Reaction norms are expressed by means of curvesobtained by plotting phenotype (or fitness) against envi-
ronment (Stearns 1989). Phenotypic plasticity may be
viewed as the degree of deviation of the reaction normfrom a flat curve along the environmental axis.
Phenotypic plasticity in S c ene de smu s
Among the green algae, experimentation with the
gents Scenedesmus best demonstrates the process of phe-
notypic plasticity. In recent time considerable informa-tion has been gathered at the University of Connecticut,
USA. Numerous experiments were designed to control
nutrients, temperature, and light in cultures of various
species (strains in culture) of Scenedesmus. The data col-
lected in more than thirty years of research haverevealed the production of multiple ecomorphs under
,tt, ,/

150 Algae Yol. 12(3), 7997
Table 1. Gradual production of ecomorphs ir UTEX 831 (a multispined species). Note that the experiments were initiated with 100%unicells. There is a shift in dominance of different ecomorphs as we advance along the growth curve. For example, on dayseven there are three morphs present (unicells, quadricaudate and multispined), but only the multispined morph is dorninant(61%). Modified from Trainor (1995).
EcomorphDays
Unicell Multispined Bicaudate AcaudateQuadricaudate
0
3
5
7
9
11
IJ
15
77
1,9
27
28
100
65
45
77
27
6
7
7
5
2
2
1
0
35
55
61
44
55
J/
38
20
18
16
10
0
0
0
22
30
37
37
40
51
60
67
56
0
0
0
0
0
0
0
0
6
1
3
3
0
0
0
0
5
2
79
15
18
19
18
30
changing environmental conditions.In S. armatus for example, there is an ordered sequence
of ecomorph formation, a sequence postulated to be truefor all species within the genus (Egan and Trainor 1989;
Trainor 7993, 1995). Unicellular organisms are producedat the beginning of the growth curve, becoming gradual-ly replaced by bicellular coenobia, long-spined quadri-caudate coenobia, and short-spined quadricaudatecoenobia toward the completion of log phase. Finally, at
the end of the growth curve, spineless coenobia possess-
ing heavy ridges are produced (Fig. 1). The productionof these morphs is often not instantaneous for there maybe transitional stages. As cultures become older newforms appear, and at any given time populations may be
composed of several morphs, only one of which is domi-nant. Clearly, dominant morphs found at the beginningof the growth curve are not dominant late in stationarygrowth (Table 1). Specifically, unicells present in muchhigher cell densities at the beginning, are present in verysmall numbers - if at all - at the end of the growth curve,
where spineless coenobia with heavy ridges dominate.The intermediate forms between unicells and coenobia
with ridges are also characterized by a small reductionin cell size.
The ordered sequence of ecomorph formation is
dependent on physico-chemical and biological changes
occurring in aquatic ecosystems (Fig. 2). This means thatphenotypic plasticity ensures survival in unstable condi-tions. There is an adaptive advantage to remainingbuoyant in the plankton when nutrient levels are high,
and to seeking another niche when environmental condi-tions change. Instability is characteristic of the aquaticmedium, and many other environments.
In temperate aquatic systems, few nutrients exist inthe water column toward the end of autumn, whilesimultaneously temperature and photoperiod change
drastically. Those factors trigger the production ofcoenobia with short spines and/or spineless coenobia
(bearing ridges in some species). These morphs offer less
resistance to sedimentation, precipitating toward the
bottom of lakes and ponds where they become dormant(Conway and Trainor 7972). Most of the cell foodreserves are used during the period of dormancy; how-
ever, the photosynthetic apparatus remains intact. As
soon as light and nutrient conditions are favorable, the
cells regain vitality, reproduce, and seek another niche.
Metabolic reactivation in dormant cells tends to occur
at the begining of the spring season, when bicellular and
tetracellular coenobia are produced. Because bothmorphs bear long spines (increased frictional resistance
to sinking), they can be easily transported by water cur-
rents to the upper portion of the water column.At the end of winter stratification, fresh nutrients from
the sediments enter the photic zone. In this abundance of
nutrients (much higher in eutrophic conditions) large
unicell populations are supported. Unicells are producedfrom bicellular and tetracellular long-spined morphs,
and not directly from dormant cells (Egan and Trainor1989). Intraspecific and interspecific competition, as wellas quick nutrient depletion in the water column, soon
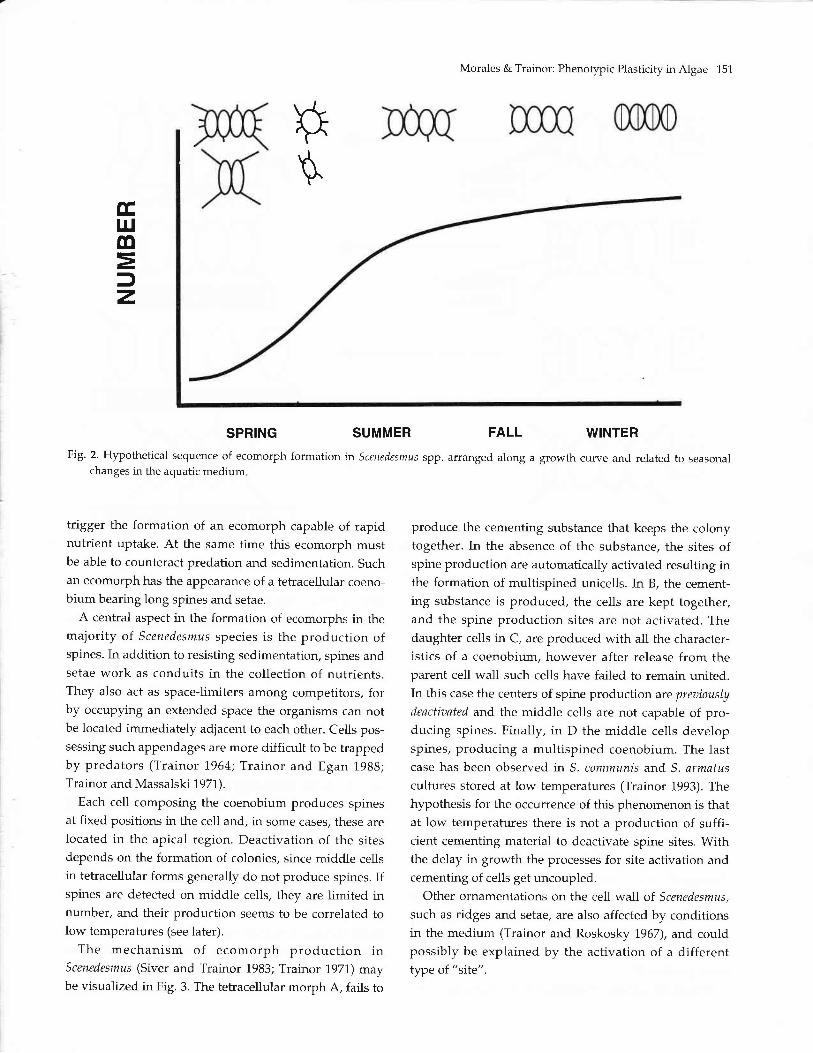
Morales & Trainor: Phenotypic Plasticity in Algae 151
SPRING SUMMER FALL WINTER
Fig. 2. Hypothetical sequence of ecomorph formation in Scenedesmus spp. arranged along a growth curve and related to seasonalchanges in the aquatic medium
*
uEIIJo=fz
trigger the formation of an ecomorph capable of rapidnutrient uptake. At the same tirne this ecomorph mustbe able to counteract predation and sedimentation. Suchan ecomorph has the appearance of a tetracellular coeno-bium bearing long spines and setae.
A central aspect in the formation of ecomorphs in themajority of Scenedesmas species is the production ofspines. In addition to resisting sedimentation, spines andsetae work as conduits in the collection of nutrients.They also act as space-limiters among competitors, forby occupying an extended space the organisms can notbe located immediately adjacent to each other. Cells pos-sessing such appendages are more difficult to be trappedby predators (Trainor 7964; Trainor and Egan 1988;
Trainor and Massalskr 1977).
Each cell composing the coenobium produces spinesat fixed positions in the cell and, in some cases, these are
located in the apical region. Deactivation of the sitesdepends on the formation of colonies, since middle cellsin tetracellular forms generally do not produce spines. Ifspines are detected on middle cells, they are limited innumber, and their production seems to be correlated tolow temperatures (see later).
The mechanism of ecomorph production inScenedesmus (Siver iand Trainor 1983; Trainor 7977) maybe visualized in Fig. 3. The tetracellular morph A, fails to
produce the cementing substance that keeps the colonytogether. In the absence of the substance, the sites ofspine production are automatically activated resulting inthe formation of multispined unicells. In B, the cement-ing substance is produced, the cells are kept together,and the spine production sites are not activated. Thedaughter cells in C, are produced with all the character-istics of a coenobium, however after release from theparent cell wall such cells have failed to remain united.In this case the centers of spine production are preoiously
deactiaated and the middle cells are not capable of pro-ducing spines. Finally, in D the middle cells developspines, producing a multispined coenobium. The lastcase has been observed in S. commlLnis and S. rtrmatus
cultures stored at low temperatures (Trainor 1993). Thehypothesis for the occurrence of this phenomenon is thatat low temperatures there is not a production of suffi-cient cementing material to deactivate spine sites. Withthe delay in growth the processes for site activation andcementing of cells get uncoupled.
Other ornamentations on the cell wall of Scenedesmus,
such as ridges and setae, are also affected by conditionsin the medium (Trainor and Roskosky 7967), and couldpossibly be explained by the activation of a differenttvpe of "site".
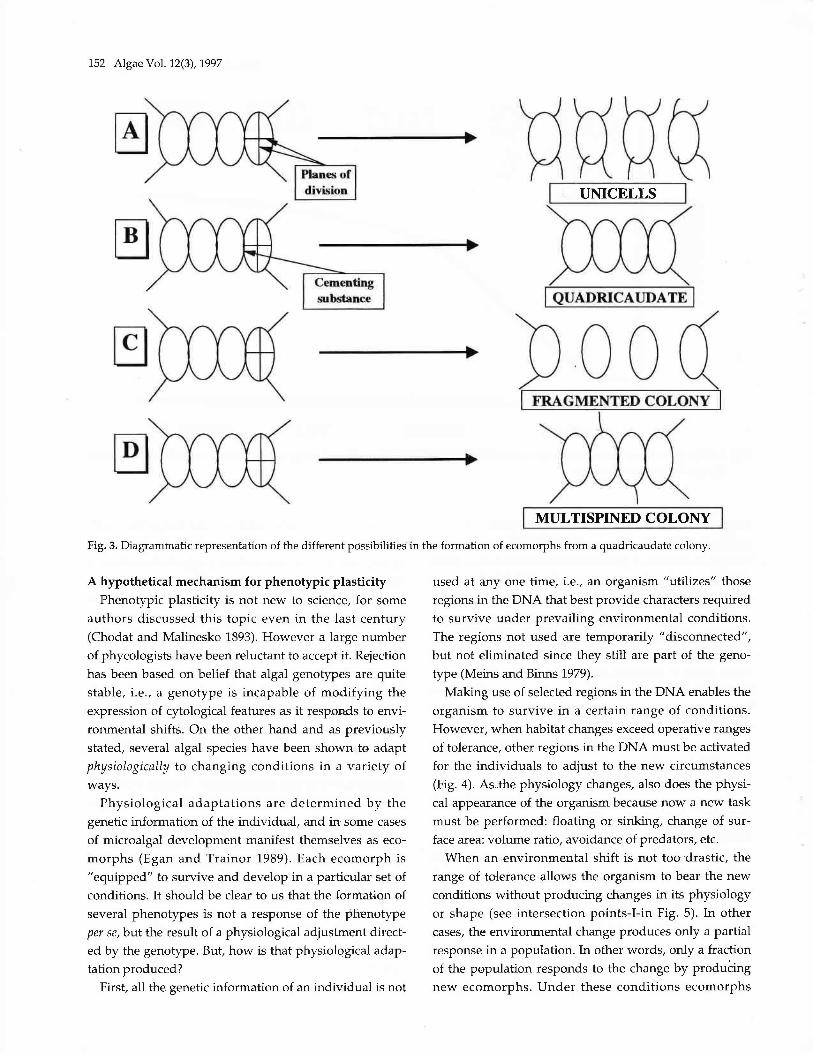
152 Algae YoL. 12(3), \997
UI\IICELLS
MT]LTISPINED COLOI\TY
Fig. 3. Diagrammatic representation of the different possibiLitieS in the formation of ecomorphs from a quadricaudate colony.
A hypothetical mechanism for phenotypic plasticityPhenotypic plasticity is not new to science, for some
authors discussed this topic even in the last century(Chodat and Malinesko 1893). However a large numberof phycologists have been reluctant to accept it. Rejection
has been based on belief that algal genotypes are quitestable, i.e., a genotype is incapable of modifying the
expression of cytological features as it responds to envi-ronmental shifts. On the other hand and as previouslystated, several algal species have been shown to adaptphysiologically to changing conditions in a variety ofways.
Physiological adaptations are determined by thegenetic information of the individual, and in some cases
of microalgal development manifest themsel.ves as eco-
rnorphs (Egan and Trainor 1989). Each ecomorph is"equipped" to survive and develop in a particular set ofconditions. It should be clear to us that the formation ofseveral phenotypes is not a response of the phenotype
per se,but the result of a physiological adjustrnent direct-
ed by the genotype. But, how is that physiological adap-
tation produced?
First, all the genetic hformation of an individual is not
used at any one time, i.e., an organism "utilizes" those
regions in the DNA that best provide characters required
to survive urxder prevailing environmental conditions.
The regions not used are temporarily "disconnected",
but not eliminated since they stilt are part of the geno-
type (Meins and Binns 1979).
Making use of selected regions in the DNA enables the
organism to survive in a certain range of conditions.However, when habitat changes exceed operative ranges
of tolerance, other regions in the DNA must be activated
for the individuals to adjust to the new circumstances
Gig. ). As.the physiology changes, also does the physi-
cal appearance of the organism because now a new task
must be performed: floating or sinking, change of sur-
face area: volume ratio, avoidance of predators, etc.
When an environmental shift is not too drastic, the
range of tolerance allows the organism to bear the new
conditions without producing changes in its physiology
or shape (see intersection points-I-in Fig. 5). In other
cases, the environmental change produces only a partialresponse in a population. In other words, only a fraction
of the population responds to the change by produiingnew ecomorphs. Under these conditions ecomorphs
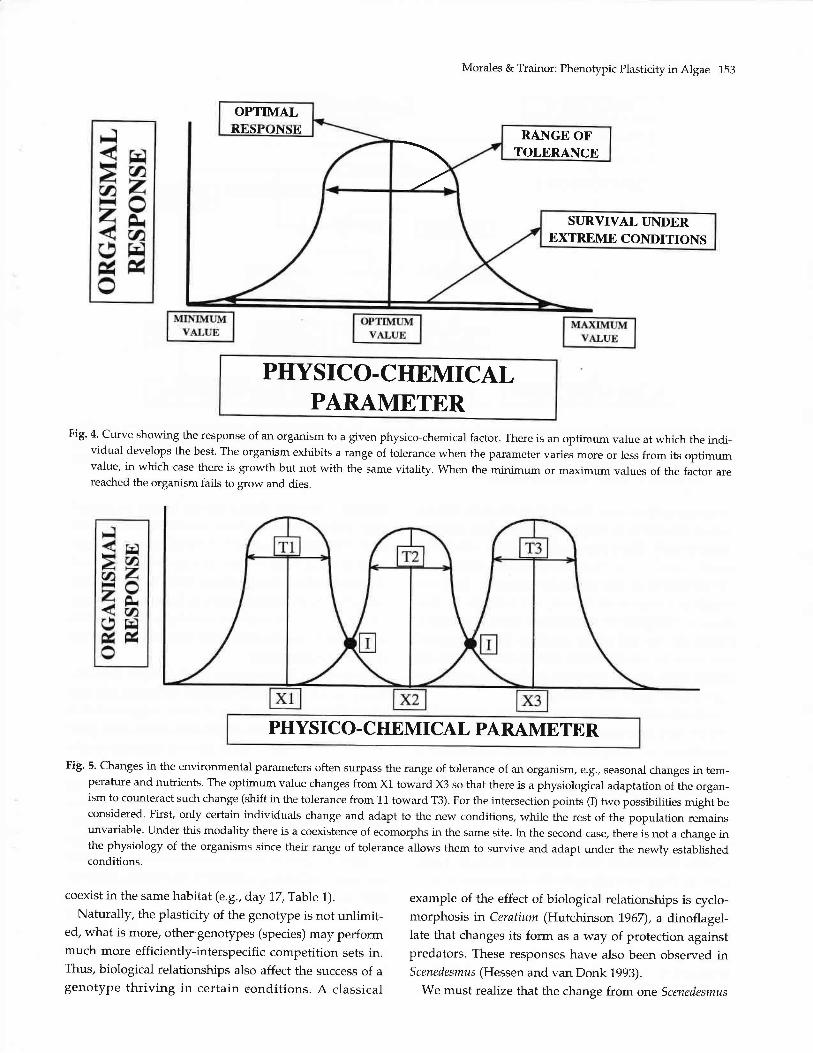
Morales & Trainor: Phenotypic Plasficity in Algae 153
PHYSICO.CHEMICALPARAMETER
Fig' 4' Cuwe showing the response of an organism to a given physico-chemical factor. There is an optimum value at which the indi-vidual develops the best. The organism exhibits a range of tolerance when the parameter varies more or less from its optimumvalue, in which case there is growth but not with the same vitality. When the minimum or maximum values of the factor arereached the organism fails to grow and dies.
PHYSICO- CHEMICAL PARAMETBR
Fig. 5. Changes in the environmental parameters often surpass the range of tolerance of an organism, e.g., seasonal changes in tem-Perature and nutrients. The optimum value charges from X1 toward X3 so that there is a physiological adaptation of the orgal-ism to counteract such change (bhift in the tolerance from T1 toward T3). For the intersection points (I) two possibilities might beconsidered. First, only certain individuals chalge and adapt to the new conditions, while the rest of the population remainsunvariable. Under this modality there is a coexistence of ecomorphs in the same site. hr the second case, there is not a change inthe physiology of the organisms since their range of tolerance allows them to survive and adapt under the newlv establishedconditions.
coedst in the same habitat (e.g., day 77,Table 7).
Naturally, the plasticity of the genotype is not unlimit-ed, what is more, other'genotypes (species) may performmuch more efficiently-interspecific competition sets in.Thus, biological relationships also affect the success of agenotype thriving in certain eonditions. A classical
example of the effect of biological relationships is cyclo-morphosis in Ceratium (Hutchinson 7967), a dinoflagel-late that changes its form as a way of protection againstpredators. These responses have also been observed inScenedesmus (Hessen and van Donk 1993).
We must realize that the change from one Scenedesmus
OPTIMAL|(I'JT(JNSIi; RANGE OF
TOLERANCE\-/
SURVWAL UNDEREXTREME COI\DITIONS

154 Algae Yol. 12(3\, 1q97
FIELD DATA LABORATORY DATA
Fig. G. Multidisciplinary character that taxonomic studies should possess. Most artificial schemes concentrate only on field data dur-
ing the identification and/or classification of organisms (A). More natural classification systems must consider laboratory stud-
ies in addition to field data. The sy'nthesis of data produced in this manner (B) will result irr more accurate identification and
classification processes (Trainor and Morales 1997).
morph to another is not developed in the same individ-ual, i.e., the change in morphology occurs from parent to
daughter cells. Then, phenotypic plasticity is linked to
sexual and asexual reproductive Processes, and thus, to
the life history of any particular organism. Each eco-
morph is a stage in life history, and each one of these
stages is "programed" to execute a set of functions to
ensure the survival of the genotype. This 'tprogtam"
may work for a long time if habitat conditions do not
change drastically, or may serve as a base for a new Pro-gram responding to nevv conditions.
A broader scope. Thinking in these terms, it is not diffi-
cult to find similarities among the life history of
Scenedesmus, cyclomorpho sis in Ceratium, and alternation
of generations in Bangia, fungi, mosses, ferns, etc. Then
we realize that phenotypic plasticity is not a phenome-
non restricted to algae, but a common process in nature.
Considerations of taxonomic inconsistencies
Many implications may be derived from investiga-
tions on phenotypic plasticity (For Scenedesrtus refer Io
Trainor and Egan 7990a,1990b). Traditional science
tends to discard the validity of selected experiments
arguing that they are simply organismal responses to
laboratory conditions. As stated previously, a general
dogma exists among phycologists to accept stability of
morphological characters. In fact, much of the early clas-
sification is founded on such dogma. But can we support
many of the basic assumptions, can evidence be accumu-
lated? The conditions created in the laboratory, and spe-
cially those in certain dilute medium cultures, are not
that different from the conditions found in natural envi-
ronments. The morphs produced in Scenedesm;us cultures
are definitely not examples of teratology, an early accu-
sation. Such questions may be raised, but soon we
deserve supporting data.
Considering the resistance to phenotypic plasticity
theory which periodically emerges, we might ask our-
selves: Have all these millions of years of algal evolution
merely resulted in survival Processes which take place
only in the laboratory? What selective pressures would
induce such changes? Obviously, phenotypic plasticity is
not a process "used" by algae to "confuse" us in our
research. \A4rat is more, it is not difficult to hypothesize
and experiment in order to understand possible implica-
tions of phenotypic plasticity, whether we explore phe-
nomena occurring in natural environments or organisms
growing in laboratory containers. The aquatic environ-
ment is constantly changing, with fluctuations in nutri-
ents, changes in the position of the organism due to
water movements and gravity, presence of predators,
and so forth. These are but some of the factors that might
be affecting adaptability of the biota. Actually, we might
postulate that most of the phenotypically plastic
LIGHT MICROSCOPY
FLAGELLARAPPARATUS
DNA AND RNASEOUENCING
MITOSIS ANDCYTOKINESIS
B: SYNTIIESIS OF DATA: IDENTIFICATIONA: IDENTIFICATION

responses are triggered by a combination of these factorsrather than a single one.
Knowing that there is phenotypic plasticity inScettedesnnts, rapid identification of any species becomes
almcst impossible. The landscape is much more compli-cated by the fact that many unicellular morphs had been
previously classified as other genera such as Chodatella
and Francein (Swale 7967; Fott 1968). Moreover, because
of an assumed stability of form most of the species inScerrcdesmus are not well defined, and application of dif-ferent specific names to morphs of a single species has
not been an infrequent event. So then, what can we - the
scientists not involved in systematics, or newcomers tothe phycological field, do to solve these problems?
Artificial vs. natural classification systems
Classification systems are constantly being trans-formed and updated. This is the result of the multidisci-plinary character that every classification model shouldhave in order to express natural (or at least the most nat-ural) relationships among organisms. At higher levelseach time we read a new paper dealing with algal sys-
tematics we get confused by the presentation of a newclassification scheme. When it becomes necessary toidentify an organism, we do not always know which sys-
tern of classification is more appiicable, updated, or evencorrect.
Certainly the causes of these complications concerningalgal classification systems are numeror,rs. After theinvention of the microscope a large diversity of microal-gal forms was discovered. The description of such vari-ability was time-demanding, so affinities among thespecimens were rapidly resolved based on their habitat,m-orphology, and/ or color. Phycology matured so quick-ly that when people thought of more natural ways ofclassification, there were already too many species tomake a detailed examilation of each Despite these facts,
scientific papers continued to be published, and moretaxa were described.
Each time a new publication appeared in the librariesit was considered as a definitive work. In the last centurythe authoritative nature of such papers was enhanced bya sense of respect and prestige. During this time utiliza-tion of discussion as a means of scientific progress sel-
dom occurred. In this manner artificial systems became
widely adopted mainly because they were practical.However the apparent practicality of artificial systems
makes the establishment of a natural approach more dif-ficult. In the case of diatoms, for instance, identification
Morales & Trainor: Phenotlpic Plasticity in Algae 155
of acid clean material would be considered more labori-ous, if their classification was not based on frustule mor-phology. When experimentation deals with ecologicalassessments, it becomes impossible to culture everyorganism in order to apply a proper name.
Concatenating criteria is an arduous and complexprocess when we are dealing with a limited number ofscientists. This fact becomes more conspicuous whenthere are differences in culture and language amongthose investigators. Each culture has its own "cosmovi-
sion" affecting the way in which a particular observation.is understood and transmitted. This is, the manner inwhich a scientist sees a particular issue and communi-cates it to colleagues depends on the culture in whichthat person has been educated and performed hisresearch. This peculiarity leads to formation of groups orscientific schools.
When a new publication appears on the market itreflects the thinking of the school to which it belongs,
and does not necessarily share the criteria of otherschools. When we make use of a paper or book in the
identification of organisms, we are not aware of all the
background details that led to its publication, and thus,
the names that we use may be interpreted in differentways by the reader. This process is even more obvious in"third world" countries in which preference of a particu-lar iiterature is suppressed by its availability. There, the
researcher uses publications from different schools foridentification of organisms even within a single genus.
This problem is not restricted to algae, but it is also
extended to ecological studies; descriptive Europeanmethodologies are not always shared by NorthAmerican Schools.
A momentary solutionBecause we can not simply leave other tasks to con-
centrate only cln algal taxonomy, we must find a way tosort out the obstacles imposed by the historical develop-ment of Phycology. We can not deny the practicality ofexistent classification schemes; neither we can ignore the
fact that diversity of thinking and scientific activitiescontribute to the development of a science with a more
secure foundation. However, we are still compelled to
accurately name the organisms we use in our studies.
The only viable way, at least until further research pre-sents an alternative, is to cite the references used duringthe identification process and/or state that our organismreseftfules another that has been described in the litera-ture.

156 Algae YoL 72(3),1997
Consolidation of natural classification systems
With the development of new techniques and method-
ologies such as electron microscopy, it is clear that lightmicroscopy is not the only path leading to the descrip-
tion of new taxa and establishment of expected natural
schemes. Moreover, examples provided by other sci-
ences such as Bacteriology, which makes extensive use
of biochemical and physiological characteristics in the
identification of bacteria (suggested by Chodat 100 years
ago), give us another point of view concerning the best
modern approach to algal classification.
In the future, no doubt more functional schemes ought
to be produced by multidisciplinary studies linking both
field and laboratory approaches (Fig. 6). Lately utlliza-tion of molecular techniques (e.g., DNA and RNAsequencing) seems to be a key step in the elucidation ofrelationships and identities within ScenedesmtLs (Kessler
et aI. 1996). Various phases have to be considered duringsuch multidisciplinary studies. Light microscopy is a
useful approach when it is combined with ultrami-croscopy in studies of life cycles (including aspects ofreproduction, mitosis, and cytokinesis), and morphologi-
cal studies on vegetative and reproductive cells (analysis
of plastids, cell walls, flagellar apparatus, etc.).Integration of the information produced in such a waywill be, without any doubt, of much help in clarifyingaffinities among algae.
ACKNOWLEDGEMENTS
We thank Dr. Peter H. Rich and Juan Dupuy for their
valuable suggestions and corrections during the elabora-
tion of this text.
REFERENCES
Bailey R. and Moore G. 1986 Reproduction of a fingernail clam
in contrasting habitats: life-history tactics? Cnn. I Zool.64:
770r-L704
Baldwin J and Dingle H 1986. Geographic variation in the
effects of temperature on life-history traits in the large
milkweed bug Oncopeltus fasciatus ()ecologia (Berlin) 59: 64-
77.
Black R. and Slobodkin L 7987. What is cyclomorphosis?F r e shutater Bio l . 18: 37 3-37 8
Bonneau E. 7977. Polymorphic behavior of Ulun Inctuca
(Chlorophyta) in axenic cuiture. I. Occurrence ofEnteromorphn-like plants in haploid clones. I Phycol. 13:.
133-140.
Bosman A. and Hockey P. 1988 Life-history patterns of popula-
tions of the limpet Patella granularls: the dominant roles offood supply and mortality rate. Oecologia (Berlin) 75: 412-
479.
Chapman A. 1973. Phenetic variability of stipe morphology inrelation to season, exposure, and depth in the non-digitate
complex of Lominaria Lamour (Phaeophyta, Laminariales)
in Nova Scotia Phycologia 12:53-57.
Chodat R. and Malinesko O. 1893. Sur le polymorphisme de
Scenedesmus ncuttLs Bull. Herb. Boiss.7: L84-790.
Conway E and Cole K 1977. Studies in the Bangiaceae: struc-
ture and reproduction of the conchocelis of Porphyrn and
Bangia in culture (Bangiales, Rhodophyceae). Phycologin 16:
205-216.
Conway K. and Trainor F. 7972. Scenedesmus morphology and
flotation. J. Phycol.8: 138-143.
Dixon P. and Richardson N. 1969. The life histories of Bangin
and Porplryra and the photoperiodic control of spore pro-
duction. Proc httl Senz,ueed Symp 6:133-139
Drew K 7949 Conchocelis-phase in the life history oI Porphyra
ttmbilicalis (L.) Kiitz N atttre'164: 7 48.
Egan P and Trainor F. 1989 The role of unicells in the poly-morphic Scenedesmus armatus (Chlorophyceae). l. Phycol.
252 65-70.
Ford N. and Seigel R. 1989 Phenotypic plasticity in reproduc-
tive traits: evidence from a viviparous snake. Ecology 70:
1768-1774.
Fott B. 7968. Chodatella stages rn Scenedesmtrs. Actn Uniaer.
Car olinae -Biolo gicn 1957 : 1,89 -79 6.
Fukuhara E 7968. Studies on the taxonorny and ecology of
Porphyn from Hokkaido, and its adjacent walers Bttll.
Hokknido Fish Res. Lnb.34:40-99.
Garbary D., Grund D. and Mclachlan I L978. The taxonomic
status of Ceramium rttbrum (Huds.) C. Ag. (Ceramiales,
Rhodophyceae) based on culture experiments. PlLycologia
17:85-94.
Hessen D and van Donk E.7993 Morphological changes in
Scenedesmus induced by substances released from Daphnia.
Ar ch. Hy drobiol 127 : 729-740.
Hutchinson E. 1967 . A treatise in Limnology. Il. Introduction tolake Biology and the limnoplankton. John Wiley & Sons.
1115 pp.Kain J. 1996. Photoperiodism in Delesseria sangtrinen
(Ceramiales, Rhodophyta) 1. The phases and sexes differ.
Phycologin 35:44o-455
Kaplan R. 1987. Developmental plasticity and maternal effects
of reproductive characteristics in the frog Bomhinn orientnl-
is. O e colo g in (Berlin) 7 l: 27 3-279.
Kessler E., Schdfer M., Htimmer C., Kloboucek A. and Huss A.
1996. Physiological, biochemical, and molecular characters
for the taxonomy of three subgenera of Scenedesmus
(Chlorococcales, Chlorophy ta) . Botanica Acfa (in press)
Liu Q , van der Meer J. and Reith M. 1994.Isolation and charac-
terization of phase-specific complementary DNAs from
sporophytes and gametophytes of Porphyrn purpurea
(Rhodophyta) using subtracted complementary DNAlibraries. L Phucol 30: 513-520

Morales & Trainor: Phenotypic Plasticity in Algae 157
Meins F. and Binns A.7979. CelI determination in plant devel- tures. Amer.l. Bot.51,:995-1001opment. Bioscience 292 227-225 Trainor F.797L. Development of form in Scenedesmus. In: Parker
Nakanishi K. and Nishijima M. 1996. Bacteria that induce mor- B. and Brown R (Eds.), ContribtLtions in Phycology ATlenphogenesis in Ulua pertusn (Chlorophyta) grown under Press Inc., p.8I-92.axenic conditions. l. Phacol. 32: 47 4-482 Trainor F. 1991. To which species of Scenedesmus should the
Newman R. 1989. Developmental plasticity of Scaphiopus cochii name S. quadricauda apply? Quadricaudate ecomorphs oftadpoles in an unpredictable environemt Ecology 70 L775- spiny species. Taxon 40:453-457.1787 Trainor F. 7992. Cyclomorphosis in Scenedesmus communis
Raven P., Everet R. and Eichhorn S 1986. Biology of plants. Hegew Ecomorph expression at low temperature. BrFourth ed. Worth Publishers, Inc. 775 pp. Phycol.l 27:75-81
Sandgren C., Hall S. and Barlow S. 1996. Siliceous scale produc- Trainor F. 1993. Cyclomorpho sis in Scenedesmus subspicntus. R.tion in chrysophyte and synurophyte algae. I. Effects of sil- Chod. Cell behavior during the unicell-colony transforma.-icalimited growth on cell silica content, scale morphology, tion of a phenotypicly plastic organism. Arch. Protistenkd.and the construction of the scale layer of Synura petersenii. 143:55-61.
l. Phycol.32:675-692 Trainor F. 1993. Cyclomorphosis tn Scenedesmus stLbspicatusSchr,rltz M.797L. Salinity-related polymorphism in the brackish- (Chlorococcales, Chlorophyta): stimulation of colony
waterdiatomCyclotellacrypticn Cnnl.Bot.49:7285-7289 developmentatlowtemperatnrc.Phycologia32:429-433.Sormus L and Bicudo C. 7974 Polymorphism in the desmid Trainor F. 1995. The sequence of ecomorph formation in a phe-
Micrasterias pinnatifida and its taxonomical implications. ,l notypicly plastic, multispined Scenedesmrrs specresPhycol 10:274-279 (Chlorophyce ae). Ar ch. Hy dr obiol. 133: 767 -77 1,.
Stearns S. 1989. The evolutionary significance of phenotypic Trainor F. and Egan P 1988. The role of bristles in the distribu-plasticitv. Bioscience 392 436-445 tion of a Scenedesmus (Chlorophyce ae). Br. Phy col l. 23; 735-
Stearns S. and Crandall R 1984. Plasticity for age and size at I41,.
sexual maturity: a life-history response to unavoidable Trainor F. 1990a. The implications of polymorphism for the sys-stress. In: Potts C. and Wooten R (eds), Fish reprodtLction: tematics of Scenedesmus.Br.Phqcol 1.25:275-279.strategies and tactics. Academic Press, pp 13-34. Trainor F 1990b. Phenotypic plasticity in Scenedesmus
Stoermer E.7967. Polymorphism in Mnstogloia. I. Phycol.3:73- (Chlorophyta) with special reference to S armatus unicells.77. Phycologia 29: 461-469
Siver P. and Trainor F. 1983 Effect of growth rate on unicell Trainor F. and Hilton R. 1963. Culture oI Scenedesmus longus.production in two strains of Scenedesmus (Chlorophyta). Bttll.TorreyBot.ChtbgO:407-472.P h y c ol o g i a 222 727 -737. Trainor F. and Massalsky A. 1971. Ultrastructure of Scenedesmus
Swale E. 7967. A clone of Scenedesmus with Chodntelln staqes. strain 614 bristles. Can [. Bot. 49:7273-1276.Brit. Phycol BulI 3: 287-293 Trainor F. and Morales E. 1997. Recent trends in the taxonomv
Tollrian R. 1994. Fish-kairomone induced morphological of selected Chlorococcalean algae. (in press)changes in Daphnin lumholtzi (Sars). Arch Hydrobiol. 130: Trainor F and Roskosky F.7967 Spine patterns in several69-75.
Trainor F.7963. The occurrence of a Dactyloccocus-like stage inclones of a Scenedesmus. Trarts Amer. Microsc. Soc.86:76-27.
an axenic culture of a Scenedesmtts. Cat. J Bot 47:967-968. Accepted 20 August 1997
Trainor F. 7964. Spine distribution in several Scenedesmus cul-





























