Embed Size (px)
Citation preview

Ecology, 92(12), 2011, pp. 2285–2298� 2011 by the Ecological Society of America
Habitat biodiversity as a determinant of fish community structureon coral reefs
VANESSA MESSMER,1,2,3,6 GEOFFREY P. JONES,1,2 PHILIP L. MUNDAY,1,2 SALLY J. HOLBROOK,4,5 RUSSELL J. SCHMITT,4,5
AND ANDREW J. BROOKS4
1School of Marine and Tropical Biology, James Cook University, Townsville, Queensland 4811 Australia2ARC Centre of Excellence for Coral Reef Studies, James Cook University, Queensland 4811 Australia
3USR 23278 CNRS-EPHE, Centre de Recherches Insulaires et Observatoire de l’Environnement (CRIOBE),Universite de Perpignan, BP 1013, 98729 Moorea, French Polynesia
4Marine Science Institute, University of California, Santa Barbara, California 93106 USA5Department of Ecology, Evolution and Marine Biology, University of California, Santa Barbara, California 93106 USA
Abstract. Increased habitat diversity is often predicted to promote the diversity of animalcommunities because a greater variety of habitats increases the opportunities for species tospecialize on different resources and coexist. Although positive correlations between thediversities of habitat and associated animals are often observed, the underlying mechanismsare only now starting to emerge, and none have been tested specifically in the marineenvironment. Scleractinian corals constitute the primary habitat-forming organisms on coralreefs and, as such, play an important role in structuring associated reef fish communities.Using the same field experimental design in two geographic localities differing in regional fishspecies composition, we tested the effects of coral species richness and composition on thediversity, abundance, and structure of the local fish community. Richness of coral speciesoverall had a positive effect on fish species richness but had no effect on total fish abundanceor evenness. At both localities, certain individual coral species supported similar levels of fishdiversity and abundance as the high coral richness treatments, suggesting that particular coralspecies are disproportionately important in promoting high local fish diversity. Furthermore,in both localities, different microhabitats (coral species) supported very different fishcommunities, indicating that most reef fish species distinguish habitat at the level of coralspecies. Fish communities colonizing treatments of higher coral species richness represented acombination of those inhabiting the constituent coral species. These findings suggest thatmechanisms underlying habitat–animal interaction in the terrestrial environment also apply tomarine systems and highlight the importance of coral diversity to local fish diversity. The lossof particular key coral species is likely to have a disproportionate impact on the biodiversity ofassociated fish communities.
Key words: biodiversity; climate change; coral reefs; diversity patterns; Great Barrier Reef, Australia;habitat–animal interactions; habitat loss; Kimbe Bay, northern Papua New Guinea; reef fish; resources;species richness.
INTRODUCTION
A fundamental issue in ecology is to understand the
key processes that establish and maintain patterns in
biodiversity. Although many hypotheses have been put
forward to explain differences in biodiversity at large
spatial scales, such as climatic factors, environmental
stability, land area, habitat heterogeneity, historical
influences, and energy availability (Kerr and Packer
1997), the underlying mechanisms remain largely
untested. Yet, biodiversity is increasingly argued to be
crucial in providing and maintaining ecosystem services
(Loreau et al. 2001, Hooper et al. 2005, Mora et al.
2011). Hence, understanding the causes and consequenc-
es of biodiversity loss requires urgent attention. Habitat
loss is widely recognized as a main driver of declining
biodiversity, particularly for terrestrial environments
(Vitousek et al. 1997, Fahrig 2001, Laurance 2007). In
many instances, habitat is transformed to a state of
lower diversity and complexity, rather than being lost
completely. Consequently, determining to what extent
the local diversity of animal communities is dependent
on habitat diversity is critical for predicting the outcome
of ongoing habitat modification and degradation (Tews
et al. 2004).
Positive correlations between the diversities of habitat
and associated animal communities have often been
observed (Lawton 1983, Tews et al. 2004, Kissling et al.
2008, Hortal et al. 2009), although occasionally no
(Currie 1991) or negative relationships (Ralph 1985)
have also been reported. In the terrestrial environment,
Manuscript received 7 January 2011; revised 10 May 2011;accepted 10 June 2011. Corresponding Editor: J. R. Rooker.
6 Present address: ARC Centre of Excellence for CoralReef Studies, James Cook University, Queensland 4811Australia. E-mail: [email protected]
2285

these relationships are usually based on plants, which
act as the primary habitat-forming organisms, and their
associated animals, often herbivores (Lawton 1983,
Currie 1991, Tews et al. 2004, Wolters et al. 2006, Jetz
et al. 2009), but cascading effects across trophic levels
can also occur (Knops et al. 1999, Crutsinger et al.
2006). For example, increased genotypic diversity in
plants was found to have positive effects on the diversity
of herbivorous insects, which in turn was correlated with
predator diversity (Crutsinger et al. 2006).
Attempts to explain the underlying mechanisms for
positive habitat–animal diversity relationships have
produced three main hypotheses (Kissling et al. 2008,
Jetz et al. 2009). (1) The ‘‘producer–consumer hypoth-
esis’’ assumes that animal species compete for food and
usually display some degree of resource specialization.
Higher diversity in food species therefore would
promote niche diversification and coexistence of diverse
consumers (Hutchinson 1959, Chesson 2000, Novotny et
al. 2006). (2) The ‘‘vegetation (habitat) structure
hypothesis’’ predicts that plant diversity may increase
structural or habitat complexity, thereby providing more
physical niches for animals to coexist (Tews et al. 2004).
(3) The habitat–animal diversity relationship may have
no direct causality and instead may arise from both
groups responding similarly to external factors, such as
environmental conditions (Hawkins and Porter 2003).
These mechanisms are not mutually exclusive and may
indeed act synergistically. However, a direct causal effect
vs. similar responses to environmental variables has very
different implications for conservation planning and is
fundamental to understanding how ecosystems work.
To date, few studies have attempted to experimentally
resolve the mechanisms underlying positive relationships
between habitat and animal diversity, and almost all of
this work has been conducted in terrestrial systems.
Tropical coral reefs present an ideal system to test the
proposed explanations because they are one of the most
diverse ecosystems in existence and large-scale correla-
tions between species richness of corals (the primary
habitat-forming organisms) and species richness of fishes
have been well documented along latitudinal and
longitudinal gradients (Briggs 1999, Hughes et al.
2002, Bellwood and Meyer 2009). The global degrada-
tion of coral reefs and detrimental effects on the
diversity and abundance of fish communities as a
consequence of loss in coral cover and structural
complexity (McClanahan 2002, Jones et al. 2004,
Graham et al. 2006, Wilson et al. 2006, Pratchett et al.
2008) highlight the importance of live corals for
associated animal communities. A better understanding
is necessary if we are to design effective conservation
strategies. Although it is highly unlikely that a single
process determines patterns of biodiversity on coral
reefs, the relative contribution of habitat variables in
shaping correlation among species patterns compared to
external variables, such as energy, climate, and historical
factors, has received very little attention. One exception
assessed how the mid-domain effect (overlap between
species ranges), energy supply (sea surface temperatures
and primary productivity), environmental variability
(changes in sea surface temperatures), and habitat
availability (habitat area, coastline length, and number
of coastal islands) predicted latitudinal diversity gradi-
ents in shore fishes using spatial autocorrelations (Mora
and Robertson 2005). In contrast to a number of
terrestrial studies (Kerr and Packer 1997, Hawkins and
Porter 2003, Jetz et al. 2009), environmental variability
and energy supply were not found to influence fish
diversity gradients. The mid-domain effect seemed to be
the strongest determinant for widespread species, and
habitat availability played an important role in small-
range species (Mora and Robertson 2005). However, the
effects of habitat diversity were not addressed and its
functional role in maintaining the diversity of associated
animals in the marine environment is unknown and has
never been experimentally tested.
We expect coral biodiversity to play an important role
in promoting diverse fish assemblages. Live coral cover
and topographic complexity of reef habitat, in particu-
lar, are critical for associated fishes and appear to have a
significant positive influence on reef fish diversity
(Luckhurst and Luckhurst 1978, Bell and Galzin 1984,
Roberts and Ormond 1987, Ohman and Rajasuriya
1998, Holbrook et al. 2008), but the effects of coral
diversity are less well known. Niche differentiation is
likely to be common among reef fishes, as varying
degrees of specialization on corals as a resource for food
or shelter exist between reef fish species (Munday 2004,
Cole et al. 2008, Pratchett and Berumen 2008).
Approximately 9–11% are strictly dependent on live
coral (Jones et al. 2004, Pratchett et al. 2008), and two-
thirds of coral reef fish are only found at sites with some
live coral (Bell and Galzin 1984). The strongest
associations with a particular coral species are generally
found for coral reef fish with specialized resource
requirements, such as obligate coral-dwelling (e.g.,
species of Gobiodon) (Munday et al. 1997) or coral-
feeding fishes (e.g., many species of Chaetodon) (Pratch-
ett and Berumen 2008). Other fish species may simply
prefer certain coral species as important shelter or
recruitment sites (Holbrook et al. 2002, Jones et al.
2004). Some fishes only associate with particular coral
species (Sale 1991, Munday 2004, Gardiner and Jones
2005) and different coral species have been shown to
support different fish communities (Holbrook et al.
2002, Feary et al. 2007). High levels of dependency on
the coral habitat suggest that niche differentiation (in
both food and shelter) is common among reef fishes, and
we would therefore predict a direct causal effect of coral
diversity on fish diversity.
In this study, field experiments were used to assess for
the first time the potential causal relationship between
the local biodiversity of corals and their associated fish
assemblages by isolating this effect from any environ-
mental variables. Specifically, the following two hypoth-
VANESSA MESSMER ET AL.2286 Ecology, Vol. 92, No. 12

eses were tested. (1) There is a direct causal link between
habitat diversity and the diversity of associated animals
on coral reefs, i.e., coral species richness is a primary
determinant of local reef fish species richness, diversity,
and abundance. (2) Niche partitioning is an importantprocess underlying the relationship between habitat
diversity and associated animal diversity, i.e., different
coral species support different fish assemblages, with the
consequence that fish community composition is depen-
dent on the presence of particular coral species. If the‘‘producer–consumer hypothesis’’ prevails, we would
only expect species directly feeding on coral to respond
to habitat diversity. In the ‘‘habitat structure hypothe-
sis,’’ habitat selection for shelter and living space would
play a major role, whereas a similar response to
environmental factors (hypothesis three) may be as-sumed if no causal link is found. To determine whether
local patterns were robust to regional differences in fish
species composition or fish–habitat interactions, the
experiment was repeated in two locations; Kimbe Bay in
Papua New Guinea, and Lizard Island on the GreatBarrier Reef, Australia. Similarly high levels of coral
and fish diversity, but substantial differences in fish
community structure, characterize the two locations.
METHODS
Experimental design and protocols
To test the effects of coral diversity on fish commu-
nities, experiments were conducted in the lagoons ofLizard Island on the Great Barrier Reef, Australia
(148410 S, 1458270 E) and Schumann Island in Kimbe
Bay, northern Papua New Guinea (58310 S, 150850 E)
(Appendix : Fig. A1). In each location, 45 patch reefs
were constructed using a total of six common, coexistingcoral species that had a branching morphology (Table
1). To examine the effects of coral species richness on
fish community characteristics, individual patch reefs
were composed of one, three, or six coral species. To
examine the effects of coral species composition on fish
assemblages, the single-species treatment was repeated
for each of the six coral species, and two medium-
diversity treatments were established using two different
combinations of three coral species, with each coral
species assigned to only one of the medium treatments
(see Table 1 for species combinations). All nine
treatments were replicated five times. The volume of
live coral was kept as constant as possible across
treatments. Four species were the same at both locations
(Acropora nasuta, Pocillopora damicornis, Porites cylin-
drica, and Seriatopora hystrix). Two similar species were
used to represent staghorn Acropora (A. muricata at
Lizard Island; A. grandis at Kimbe Bay) and bottlebrush
Acropora (A. loripes at Lizard Island; A. carduus in
Kimbe Bay); see Table 1. For simplicity, we refer to the
different treatments with codes: Acropora nasuta (An),
bottlebrush Acropora (Bb), Porites cylindrica (Pc),
Pocillopora damicornis (Pd), Seriatopora hystrix (Sh),
staghorn Acropora (St), medium combination A (MA),
medium combination B (MB), and high (H).
Patch reefs, each 100 cm in diameter and 50 cm high,
were built at 3–7 m depth on large, flat, sandy areas
where no other habitat structure was present. Reefs were
placed 15 m apart from each other and from any
neighboring reef structures to limit fish movement
between reefs. The base of each patch reef consisted of
dead coral rubble, which was covered with the same
amount of live coral for each patch reef (90% live coral
cover). Patch reefs were established in April 2007 in
Kimbe Bay and in November 2007 at Lizard Island.
Fish were allowed to naturally colonize the patch reefs
over a 12-month period and the patch reefs were
surveyed four times, i.e., every 3–5 months. For each
survey, the abundance of every fish species present on
each reef was recorded. Recorded fish species included
all those associated with the patch reefs, but did not
include larger mobile species that were observed to move
on a regular basis between reefs. Minor repairs to the
reefs were carried out where necessary after each survey.
Commencement of the experiment at each location was
timed to match the start of the respective recruitment
TABLE 1. List of the coral species and number of coral species used in each treatment at Lizard Island, Great Barrier Reef,Australia, and at Kimbe Bay, Papua, New Guinea.
Treatment CodeDiversity(no. spp.) Lizard Island Kimbe Bay
Single species
A. nasuta An 1 Acropora nasuta Acropora nasuta‘‘Staghorn’’ St 1 Acropora muricata Acropora grandis‘‘Bottlebrush’’ Bb 1 Acropora loripes Acropora carduusP. damicornis Pd 1 Pocillopora damicornis Pocillopora damicornisP. cylindrica Pc 1 Porites cylindrica Porites cylindricaS. hystrix Sh 1 Seriatopora hystrix Seriatopora hystrix
Medium A MA 3 A. loripes, P. damicornis, P. cylindrica A. carduus, P. damicornis, P. cylindricaMedium B MB 3 A. nasuta, A. muricata, S. hystrix A. nasuta, A. grandis, S. hystrixHigh H 6 A. nasuta, A. muricata, A. loripes, P.
damicornis, P. cylindrica, S. hystrixA. nasuta, A. grandis, A. carduus, P.
damicornis, P. cylindrica, S. hystrix
Notes: Patch reefs were characterized by three different levels of coral species richness (1, 3, and 6 species). The single-speciestreatments were repeated for all six coral species, and two combinations of three coral species were used for the medium-diversitytreatments. The high-diversity treatment included all six coral species used at each location.
December 2011 2287CORAL REEF HABITAT AND FISH DIVERSITY

seasons. Recruitment is highest during the dry winter
season in Kimbe Bay, whereas a distinct recruitment
peak occurs over summer at Lizard Island. Fish species
richness and abundance patterns were mostly estab-
lished after 2 months at both locations (V. Messmer,
unpublished data). Because the patch reefs in Kimbe Bay
were in a deteriorated state during the last survey, results
presented here are from the survey carried out in late
November 2007 (8 months), coinciding with the end of
the recruitment season. The 12-month survey at Lizard
Island (early December 2008) coincided with the peak of
the recruitment season and was therefore the most
comparable survey.
Statistical analyses
Fish communities at Lizard Island and Kimbe Bay
were very different, with only 24.2% of recorded species
shared between locations (Appendix : Fig. A2). Analy-
ses were therefore carried out separately for each
location. Mean fish species richness and abundance
were based on the total number of species or individuals
observed on each replicate reef during the last survey
and were averaged (1) for each level of coral species
richness or (2) for each treatment. First, the effects of
coral species richness on fish species richness, evenness,
and abundance were compared between three richness
levels of coral species (low, medium, high). For this
analysis, fish species richness and abundance were
pooled and averaged across the six single-coral species
treatments (low) and for both three-coral-species treat-
ments (medium) and for the six-coral-species plots
(high). Second, to test the effects of coral identity on
fish assemblages, fish species richness, evenness, and
abundance were analyzed separately for each treatment
(nine levels). One-way ANOVA, followed by Tukey’s
hsd post hoc tests, compared differences in fish species
richness, arcsine-transformed fish species evenness
(Shannon evenness index, J ), and total fish abundance.
To determine the influence of different coral species
and coral species richness on the composition of fish
communities (i.e., species composition and relative
species abundance), we used canonical analyses of
principal coordinates, CAP (Anderson and Willis
2003), and multivariate regression trees, MRT (De’ath
2002). CAP was used to examine patterns of community
differences between locations as well as between
treatments at each location. CAP is a constrained
ordination technique that further analyzes the results
of a principal coordinates analysis (PCO), for which the
type of ecological distance can be chosen. It enables
testing of significant grouping structure within the
ordination by using permutation tests to assign a P
value to the a priori hypothesis that the probability of
the grouping found in the analysis could be due to
chance alone by ‘‘leave-one-out’’ allocations. MRTs
were then conducted to test the differences and
similarities between groups. This multivariate discrimi-
nation technique constructs a hierarchical tree by
creating splits, which minimize the dissimilarity of
groups within clusters. Both CAP and MRT analyseswere based on the Bray-Curtis dissimilarity measure of
log-transformed abundance data (ln (x þ 1)) of the fishon each replicate reef during the last survey. Lognormal
transformations were applied to reduce the emphasis ofhighly abundant species, which would otherwise drivemost of the observed patterns, and Bray-Curtis distances
are generally considered well suited for abundance data.Recruitment of apogonids (cardinalfish) was very high
at Lizard Island, with many reefs receiving hundreds ofindividuals. Because this group of fish is known to
influence patterns in fish communities, apogonid specieswere excluded from CAP and MRT analyses to enable
differences in the majority of fish species betweentreatments to be detected. Rare species (fewer than five
individuals sighted over 12 months) were also excluded.The number of permutations in the CAP analyses was
set to 100. The default was selected for the number ofmeaningful PCO axes (m), which chooses the optimal
number of axes in order to provide the best distinctionbetween groups and maximizes the proportion of correct
allocations to the grouping variable and minimizesmisclassification error (Anderson and Willis 2003). The
first two axes, which explained most of the variation,were illustrated in an ordination plot. Dispersion ellipsesusing 0.9 confidence limits of the standard deviation of
point scores were also plotted. Species showing thestrongest indication of difference between treatments
(i.e., correlation with axis 1 and/or axis 2 . 0.2) wereplotted separately and listed in the Appendix: Table A1.
In the MRT analyses, the best tree size was chosen bycross-validation and the 1 SE rule. The relative error
corresponds to the amount of variation among samplesnot explained by the tree (De’ath 2002). All analyses and
plots were coded in R 2.10.0 (R Development CoreTeam 2009) using the R statistical packages vegan,
BiodiversityR, MASS, and mvpart.
RESULTS
Regional difference in local species composition
In total, 150 fish species colonized the patch reefs atLizard Island and 122 species were recorded in Kimbe
Bay. Of the overall total of 219 fish species, 53 (24.2%)were observed at both locations, representing ;35% of
the Lizard Island fish community and ;43% of theKimbe Bay community. The composition of fish
communities was very different between the twolocations (Appendix: Fig. A2).
Effects of coral species richness on fish species richness,evenness, and abundance
Fish species richness significantly increased with
increasing coral species richness in Kimbe Bay, withthe single-coral treatments supporting significantlylower fish species richness (13.6 fish species) than the
medium- (17.7 fish species) and high- (20.0 fish species)diversity coral treatments (Fig. 1B, Table 2A). At Lizard
VANESSA MESSMER ET AL.2288 Ecology, Vol. 92, No. 12

Island, mean fish species richness increased from 21.0 to
25.0 species with increasing coral species richness, but
here the difference was not statistically significant (Fig.
1A, Table 2A). Evenness of the fish communities did not
differ among different levels of coral species richness at
either location (Fig. 1C, D, Table 2A).
No significant differences in mean total fish abun-
dance were observed between different levels of coral
species richness (Fig. 1E, F, Table 2A). Pooled mean
abundances of fish were higher at Lizard Island (273.7 6
28.7 fish, mean 6 SE) than in Kimbe Bay (120.0 6 29.2
fish; ANOVA: F1,88 ¼ 24.75, P , 0.001), which was
primarily driven by the presence of large schools of
apogonids at Lizard Island. Distributions of Chromis
viridis (a damselfish) were patchy, as this species was
either absent or occurred in large schools. Pooled mean
fish abundances not including apogonids and C. viridis
were more similar between both locations, although still
significantly higher at Lizard Island than in Kimbe Bay
(81.2 6 4.3 fish and 60.8 6 4.3 fish, respectively;
ANOVA: F1,88¼ 12.16, P , 0.001). There was no effect
of coral species richness on abundance when apogonids
and C. viridis were excluded from analysis (Fig. 1G, H,
Table 2A).
FIG. 1. Effects of three levels of coral species richness (low¼ 1, medium¼ 3, high¼ 6 species) on fish communities of LizardIsland (Great Barrier Reef, Australia) and Kimbe Bay (Papua, New Guinea): (A, B) fish species richness (all species); (C, D)Shannon evenness index (all species); (E, F) total fish abundance per patch reef, ;1 m2 (all species); and (G, H) fish abundance perpatch reef, not including apogonids (cardinalfish) or Chromis viridis (a damselfish). Values are shown as mean 6 SE; see Table 2A.Lowercase letters (panel B) indicate significant differences (P , 0.05) identified by Tukey’s hsd post hoc tests.
December 2011 2289CORAL REEF HABITAT AND FISH DIVERSITY

Effects of coral species composition on fish species
richness, evenness, and abundance
Fish species richness differed between treatments at
both locations (Fig. 2A, B, Table 2B). The high-diversity
(H) and medium B-diversity (MB) treatments supported
the highest number of fish species in both locations.
However, some single-coral treatments (An and Sh)
supported similarly high fish species richness (Fig.
2A, B, Table 2B), and these coral types probably
contributed to the high species richness seen in the
combination treatments medium B and high. In
contrast, other coral species tended to support low
species richness at one (e.g., Bb and Pd in Kimbe) or
both locations (e.g., Pc and St), of which Pc, Bb, and Pd
form the medium A treatment, where similarly low fish
species richness was observed. Evenness was similar
across treatments at Lizard Island, with no significant
differences between treatments (Fig. 2C, Table 2B), but
it differed statistically among treatments in Kimbe Bay
(Fig. 2D, Table 2B). Low evenness in the An and
medium B treatments was driven by the presence of
large schools of Chromis viridis on some replicates.
Removal of C. viridis from the analysis caused evenness
to be similar across most treatments, with only Pd
showing significantly lower values than Pc (P ¼ 0.010).
In contrast to the low abundances and species richness
observed in the treatments St and Pc, values of evenness
were among the highest in these corals. Total fish
abundance, including all fish species per reef, did not
differ between treatments at Lizard Island (Fig. 2E,
Table 2B), whereas in Kimbe Bay, total abundances
were found to be markedly higher on patch reefs of the
treatments An and medium B (Fig. 2F, Table 2B). The
high variation in abundance of some treatments at both
sites was largely driven by large schools of Chromis
viridis, which were found on three of the six coral species
at Lizard Island (Acropora nasuta, Pocillopora damicor-
nis, Seriatopora hystrix), but on just one in Kimbe Bay
(A. nasuta). When apogonids and C. viridis were
excluded from the analyses, mean abundances at Lizard
Island were significantly lower in the Pc and St
treatments than in Bb, Pd, and Sh (Fig. 2G, Table
2B). In Kimbe Bay, mean fish abundances were also
significantly lower in Pc and St than in An, Bb, Pd, and
medium B (Fig. 2H, Table 2B).
Effects of coral species on composition of fish community
Clear differences in the composition of fish communi-
ties between treatments were observed for both locations.
TABLE 2. For Lizard Island (LI) and Kimbe Bay (KB), one-way ANOVA testing the effects of (A) coral species richness and (B)reef treatment on fish species richness, Shannon evenness index, total fish abundance, and fish abundance excluding apogonids(cardinalfish) and Chromis viridis (a damselfish).
Effect SS effect df effect SS residuals df residuals F P
A) Coral richness
Fish species richness
LI 73.9 2 876.4 42 1.77 0.183KB 254.5 2 619.3 42 8.63 ,0.001
Fish evenness
LI 0.0 2 0.3 42 1.98 0.150KB 0.1 2 1.7 42 0.84 0.438
Total fish abundance
LI 33 757 2 812 250 42 0.87 0.425KB 15 689 2 1 027 759 42 0.32 0.728
Fish abundance excluding two taxa
LI 318 2 35 929 42 0.19 0.831KB 415 2 36 976 42 0.24 0.791
B) Reef treatment
Fish species richness
LI 503.1 8 447.2 36 5.06 ,0.001KB 517.0 8 356.8 36 6.52 ,0.001
Fish evenness
LI 0.0 8 0.3 36 0.72 0.674KB 0.9 8 0.9 36 4.50 ,0.001
Total fish abundance
LI 106 774 8 739 233 36 0.65 0.731KB 502 680 8 540 768 36 4.18 0.001
Fish abundance excluding two taxa
LI 16 701 8 19 547 36 3.84 0.002KB 21 217 8 16 173 36 5.90 ,0.001
Notes: The Shannon evenness index was arcsine square-root transformed. Effects of coral richness (1, 3, and 6 species) are shownin Fig. 1. Coral reef treatments were low-diversity (single coral species An, Bb, St, Pc, Pd, Sh); medium A and medium B (twogroups of three species); and high (six coral species). Effects are shown in Fig. 2.
VANESSA MESSMER ET AL.2290 Ecology, Vol. 92, No. 12

At Lizard Island, fish communities inhabiting each coral
species were quite distinct; the six single-species treat-
ments (An, Bb, Pc, Pd, Sh, and St) formed clusters in the
ordination graph with little overlap (Fig. 3A). In contrast,
fish communities on the treatments of higher coral species
richness seemed to represent a mixture of those found on
each of its constituent coral species; their clusters
overlapped with their constituent single-species treat-
FIG. 2. Effects of coral species composition on fish communities of Lizard Island and Kimbe Bay for each treatment: (A, B) fishspecies richness (all species); (C, D) Shannon evenness index (all species); (E, F) total fish abundance (all species); and (G, H) fishabundance excluding apogonids and Chromis viridis. These two taxa were excluded because of their patchy occurrence due to highrecruitment on some reefs and schooling behavior. Values are shown as means 6 SE (see Table 2B). Lowercase letters indicatesignificant differences (P , 0.05) identified by Tukey’s hsd post hoc tests. Treatment patch reefs were low diversity (single coralspecies: An, Bb, Pd, Sh, St), medium diversity (two combinations of three different coral species each: MA, MB), or high diversity(six coral species: H); see Table 1 for full scientific names.
December 2011 2291CORAL REEF HABITAT AND FISH DIVERSITY

ments. For example, medium A overlapped largely with
Bb, whereas An and Sh are both constituents of medium
B, which was embedded within their cluster. The high-
diversity treatment was located in the middle of the plot,
overlapping with most treatments.
The distinction between fish communities of different
treatments using CAP was even stronger in Kimbe Bay
(Fig. 4A). Similarities between fish communities on the
high-diversity coral species richness treatments and
those of their constituent coral species were also
observed there. Medium A slightly overlapped with
Pd, which constitutes one of its species. The An cluster
overlapped to some degree with medium B and the Sh
cluster was situated in close proximity. High coral
species richness appeared to promote a mixture of the
fish communities found on the constituent coral species,
as suggested by the relatively central location of the
high-diversity treatment and dispersed spread of this
cluster in the ordination plot.
At Lizard Island, many fish species preferred partic-
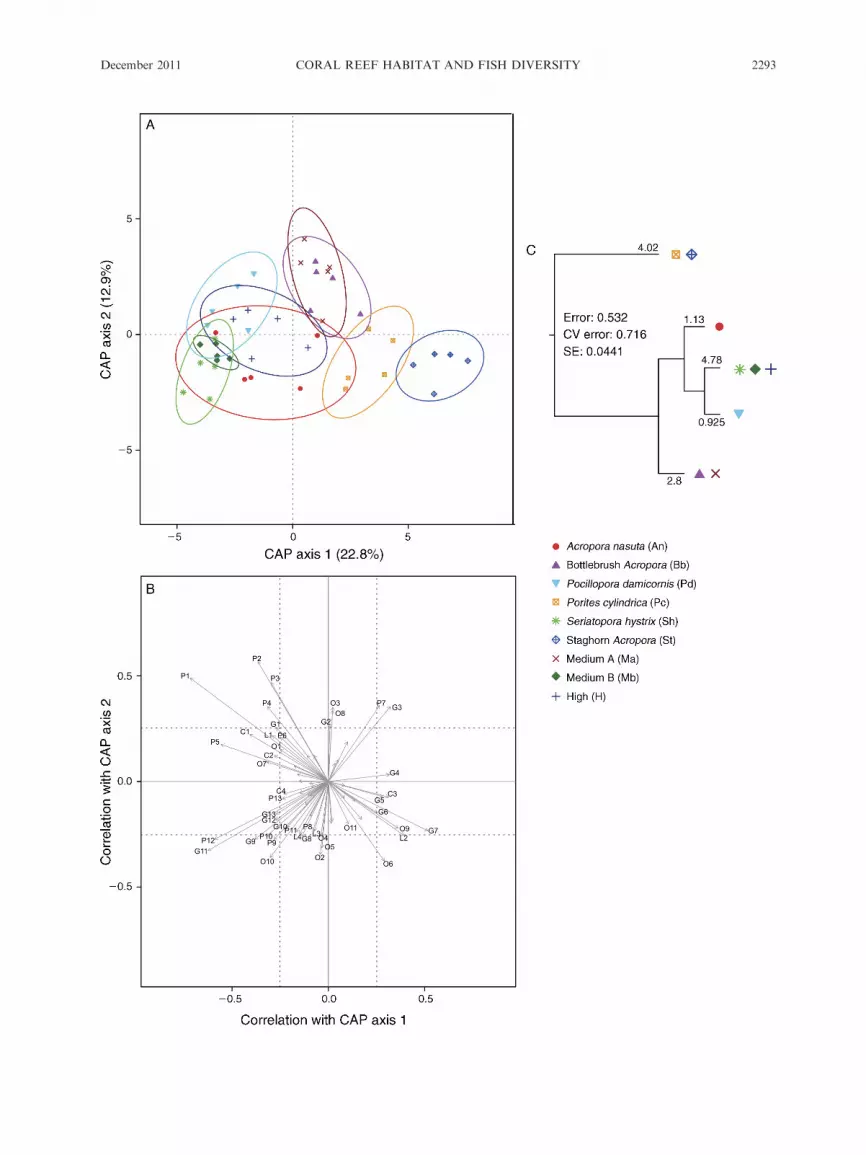
ular coral species; 66% of the species used in the analysis
were strongly correlated (.0.2) with one or both CAP
axes (Fig. 3B; also see the Appendix: Table A1). The
differences between fish communities on different coral
species were driven by a variety of fish species covering a
range of reef fish families, but consisted in particular of
gobies, damselfishes, and butterflyfishes (Fig. 3B;
Appendix: Table A1). Many of these species preferred
An, Sh, and Pd over other corals (V. Messmer,
unpublished data). Very similar patterns were observed
in Kimbe Bay, where 62% of the species were strongly
correlated with one or both of the CAP axes (Fig. 4B;
Appendix: Table A1). A considerable proportion of the
species driving the differences in fish communities
between treatments was also accounted for by gobies,
whereas only a few damselfish species showed a
preference for particular coral species. In contrast to
Lizard Island, only a few species preferred Pd and the
distinction of the fish community on St seemed to be
largely driven by the absence or low abundances of
many species, as no fish species preferred this coral
species (Fig. 4B; Appendix: Table A1).
The distinction between fish communities from
different coral species at Lizard Island found in the
CAP analysis was supported by the MRT, with 46.8% of
the total variation explained by the treatments (Fig. 3C).
The MRT provided information on similarities/dissim-
ilarities among the fish assemblages. Those on Pc and St
were most different from all others, as they formed the
first split of the tree, explaining most of the variation
between groups. The second split based on the
remaining variation separated Pd and medium A, with
Pd being part of medium A. Next, An split from the
remaining treatments and the final split distinguished Pd
from the cluster Sh, medium B, and H, Sh being part of
medium B. Very similar results were also found for
Kimbe Bay, equally supporting the CAP analysis (Fig.
4C). The treatments explained 46.8% of the total
variation. Pc, St, and Bb formed the first split, with fish
assemblages on St and Pc being more similar. These
three treatments were most distinct from all other
treatments, explaining most of the variation. The first
split within the other major branch of the tree separated
Pd with medium A and H, with Pd being part of medium
A. Of the remaining treatments, Sh split from the cluster
An and medium B, with An being part of medium B.
DISCUSSION
Through experimentally minimizing the effects of
environmental variables, our results directly demon-
strated the importance of microhabitat (coral) species
richness and composition to the local species richness,
composition, and abundance of reef fish communities.
The component of fish diversity that was most affected
by the experimentally imposed variation in habitat was
species richness. Coral species richness promoted local
fish species richness, tended to increase fish evenness, but
had little effect on the overall abundance of fishes.
Furthermore, different microhabitats were found to
strongly influence the structure of the associated animal
communities. Certain coral species supported signifi-
cantly more abundant and diverse fish assemblages than
others, but had little effect on evenness, suggesting that
habitat diversity and composition do not necessarily
influence the relationship between fish species richness
and fish abundance. There were also substantial
differences in the composition of fish communities
associated with different coral species, and a habitat
patch composed of several coral species supported a fish
assemblage that reflected the particular composition of
microhabitat (coral) types. These results were remark-
ably consistent between the two geographic locations,
despite having ,25% of fish species in common. The
consistency in the response of fish communities to coral
identity and species richness, regardless of differences in
regional species composition or other environmental
!FIG. 3. Lizard Island, December 2008. (A) Canonical analysis of principal coordinates (CAP) ordination plot (Bray-Curtis) of
fish assemblage data showing treatment effects; each point represents a separate patch reef. CAP groupings were stronglysupported, with 66.7% correct allocations (P ¼ 0.001). Six PCO axes produced the best result, accounting for 68.0% of the totalvariation. PCO axes 1 and 2 explained 22.8% and 12.9% of the total variation, respectively. (B) Species scores for CAP plots and(C) multivariate regression tree (MRT) based on a Bray-Curtis dissimilarity matrix of log-transformed fish abundance data, usingtreatments as groupings. In panel (B), letter-number codes are provided for each fish species showing correlations of r � 0.20 witheither canonical axis (C, chaetodontids; L, labrids; P, pomacentrids; G, gobies; O, other fish groups). Species names and r values arespecified in Appendix A: Table A1.
VANESSA MESSMER ET AL.2292 Ecology, Vol. 92, No. 12
December 2011 2293CORAL REEF HABITAT AND FISH DIVERSITY

features, highlights the importance of diversity in
habitat-forming species in promoting and maintaining
local fish diversity on coral reefs.
Using correlative data, positive relationships between
coral and fish diversity have also been found at small
(Luckhurst and Luckhurst 1978, Bell and Galzin 1984,
Roberts and Ormond 1987, Ohman and Rajasuriya
1998, Komyakova 2009) and large spatial scales
(Hughes et al. 2002, Bellwood and Meyer 2009), yet
the relationship had never been experimentally verified
for coral reefs. Such positive correlations in species
richness between taxa are commonly observed across
ecosystems (Lawton 1983, Currie 1991, Tews et al. 2004,
Wolters et al. 2006, Jetz et al. 2009), but the underlying
processes are still poorly understood (Rahbek and
Graves 2001). Although habitat diversity has long been
recognized as a potential driver of animal diversity
(‘‘producer–consumer hypothesis’’ and ‘‘habitat struc-
ture hypothesis’’) (Hutchinson 1959), climate and other
environmental variables (e.g., energy) also are often
invoked in a third hypothesis as principal factors
producing positive relationships (Hawkins and Porter
2003). Few attempts have been made to disentangle the
relative roles of these underlying processes and the
results are not always consistent. No positive effects of
habitat diversity on the diversity of associated animals
have been found in some correlative studies after
environmental variables were accounted for (Hawkins
and Porter 2003, Jetz et al. 2009), whereas in others the
contribution of habitat diversity was strong (Kerr and
Packer 1997, Marquez et al. 2004, Novotny et al. 2006,
Kissling et al. 2007, Menendez et al. 2007, Kissling et al.
2008, Hortal et al. 2009, Qian et al. 2009). The
advantage of experimental studies is that the influence
of environmental variables can be minimized, which
allows for direct testing of the importance of habitat
diversity itself. We are only aware of two such studies,
both conducted in terrestrial systems: in each of them,
plant species richness positively affected the species
richness of insects (Siemann et al. 1998, Haddad et al.
2001). The positive relationship between coral and fish
species richness found here supports these findings and
suggests that direct positive effects of habitat diversity
can also play an important role in the marine
environment. Although environmental variables (hy-
pothesis three) are also likely to play an important role
in general, particularly at large spatial scales, they did
not contribute to the positive effects of coral diversity on
fish diversity observed here. Instead, our results indicate
that the ‘‘producer–consumer hypothesis’’ and/or the
‘‘habitat structure hypothesis’’ clearly influence local
patterns of fish and coral diversity. To what degree
habitat diversity may work synergistically with other
proposed processes (Bellwood and Wainwright 2002,
Mora and Robertson 2005) in producing known
congruent species patterns among coral reef organisms
at very large spatial scales is yet to be tested.
The positive influence of plant species richness on the
diversity of associated animal communities, in particular
that of specialist herbivores, has often been attributed to
the availability of a greater diversity of resources, which
are thought to provide opportunities for niche parti-
tioning and the coexistence of species (Hutchinson 1959,
Murdoch et al. 1972, Chesson 2000, Novotny et al. 2006,
Kissling et al. 2007). Processes of habitat selection and
resource specialization are well known in the terrestrial
environment and play a fundamental role in structuring
ecological communities (Futuyma and Moreno 1988,
Morris 2003). Resource partitioning can occur either in
food resources (producer–consumer hypothesis)
(Hutchinson 1959, Chesson 2000, Novotny et al. 2006)
or in the provision of habitat (habitat structure
hypothesis) (Tews et al. 2004). These patterns of
habitat–animal associations also appear to apply to
coral reef systems. Like plants, corals provide both food
and habitat in which associated communities reside. On
coral reefs, strong, even obligate, associations occur
between reef fish species and certain corals (Munday et
al. 1997, Pratchett 2005), although less specialized
species also often exhibit some level of preference for
certain coral species (Munday et al. 1997, Gardiner and
Jones 2005, Cole et al. 2008, Wilson et al. 2008).
The concept of niche partitioning as a driving
mechanism for positive habitat–animal relationships is
supported by our results, as clear patterns of habitat
selection by fish species among the six coral species
investigated were confirmed by the multivariate analy-
ses. However, other post-settlement processes, such as
predation, cannot be excluded. Nearly two-thirds of the
species at both locations were strongly associated with
microhabitats, suggesting that habitat selection is
deterministic in structuring fish communities on coral
reefs. Clear choices for a particular coral species as food
or shelter, in particular Acropora nasuta, Seriatopora
hystrix, and Pocillopora damicornis, were often observed
in highly specialized fishes. Habitat selection in coral-
feeding fishes, e.g., butterflyfishes (Chaetodon spp.)
(Pratchett and Berumen 2008), supported the ‘‘produc-
er–consumer hypothesis.’’ However, the ‘‘habitat struc-
ture hypothesis’’ best explained our results, as the
!FIG. 4. Kimbe Bay, November 2007. (A) Canonical analysis of principal coordinates (CAP) ordination plot (Bray-Curtis) of
fish assemblage data showing treatment effects; each point represents a separate patch reef. Groupings were strongly supported,with 93.3% correct allocations (P¼ 0.001). The best result consisted of 13 principal coordinates axes, accounting for 96.1% of thevariation. Axes 1 and 2 explained 19.7% and 14.1% of the variation, respectively. (B) Species scores for CAP plots and (C)multivariate regression tree (MRT) constructed on a Bray-Curtis dissimilarity matrix of log-transformed fish abundance data,using treatments as groupings. Codes are as in Fig. 3.
VANESSA MESSMER ET AL.2294 Ecology, Vol. 92, No. 12

December 2011 2295CORAL REEF HABITAT AND FISH DIVERSITY

majority of species displaying habitat preferences
selected particular coral species for living space and
shelter, e.g., coral-dwelling gobies (Munday 2000), but
so did less specialized planktivorous pomacentrids. By
contrast, staghorn Acropora and Porites cylindrica
seemed to be actively avoided by a number of reef
fishes, whereas other fish species were found exclusively
on these corals. Not surprisingly, and consistent with the
concept that particular organisms are associated with
particular microhabitats (Bernays and Graham 1988),
the patch reefs characterized by higher coral species
richness supported fish communities that represented a
mixture of those found on the constituent coral species.
Although habitats characterized by higher coral diver-
sity did not necessarily support more diverse or
abundant fish communities than did some single-coral-
species patches, a diverse habitat contains more micro-
habitats that are likely to be preferred or strongly
selected for by a significant number of different coral
reef fishes and should result in overall higher animal
diversity.
In addition to better understanding the processes
producing positive relationships between habitat and
animal diversity, our results also highlight the critical
importance of particular components of the habitat (in
this case, individual coral species) in establishing this
relationship. The relationship was strikingly dependent
on the particular corals included in the diversity
treatments, suggesting that particular coral species,
and not necessarily coral diversity per se, are critical
for sustaining diverse and abundant fish communities.
Variation in fish diversity and community structure
between coral species is not unexpected, as coral species
vary in morphology and many fish species preferentially
associate with certain coral species (Hixon and Menge
1991, Munday et al. 1997, Holbrook et al. 2002). In our
experiment, Porites cylindrica and staghorn Acropora
were characterized by consistently low fish species
richness. In Kimbe Bay, bottlebrush Acropora, and
Pocillopora damicornis, which together with Porites
cylindrica formed the medium A treatment, displayed
similarly low fish species richness, and this was mirrored
by low fish species richness in the medium A treatment.
In contrast, fish species richness of the medium B
treatment was equal to the high-diversity treatment
because they both shared coral species that supported
high fish species richness, in particular Acropora nasuta
and Seriatopora hystrix. The discrepancy in diversity
between the medium diversity treatments thus is likely to
reflect the ‘‘performance’’ of their constituent coral
species in terms of fish species richness. Differences in
the ‘‘performance’’ of corals were also reflected in
patterns of total fish abundance, which were little
affected by coral species richness, but varied significantly
with coral identity and composition. Patterns of mean
abundance (not including apogonids and Chromis
viridis) were strikingly similar between locations and,
importantly, also mirrored patterns of fish species
richness. Coral identity played an important role in
maintaining abundant fish communities, with Porites
cylindrica and staghorn Acropora consistently support-
ing the lowest fish abundances. This may have been due
to the more open branching structure of these corals
compared to the other coral species, potentially provid-
ing less appropriate shelter relative to the size of the
patch reefs. At both sites, mean fish abundances were
high in the treatments of higher coral species richness,
but not more so than particular coral species, such as
Acropora nasuta, bottlebrush Acropora, Pocillopora
damicornis, and Seriatopora hystrix.
Differences in the capacity of different coral species to
promote and maintain abundant and diverse fish
communities is of great concern, because the health of
coral reefs is declining at a global scale (Hoegh-
Guldberg 1999, McClanahan 2002, Gardner et al.
2003, Wilkinson 2004) and increasing numbers of coral
species are expected to undergo reductions in abun-
dance, at least at local scales. The clear preference of
many fish species for particular types of coral suggest
that certain microhabitats, such as Acropora nasuta and
Seriatopora hystrix, play a particularly important role in
promoting diverse local fish communities, but these
particular coral species are also known to be highly
susceptible to disturbances (Marshall and Baird 2000,
Loya et al. 2001, McClanahan et al. 2007). Loss of these
species could have a disproportionately strong impact
on local fish communities. Similarly, Tews et al. (2004)
reviewed a number of studies that identified crucial
keystone structures in the vegetation to which different
species groups were closely linked and that dispropor-
tionately influenced animal species. A shift in the habitat
community structure therefore has the potential to result
in less diverse and abundant animal communities and
significant alterations in their composition.
The remarkable congruence in the response of fish
communities to coral diversity and identity between the
two locations highlights the strength and nature of the
relationship between fish and corals. The effects of coral
diversity and identity on fish composition, diversity, and
abundance were similar, despite the differences in the
fish communities and overall higher fish diversity and
abundance observed at Lizard Island. The use of a field
experiment enabled us to investigate for the first time the
specific influence of specific coral species and coral
diversity on the structure of coral reef fish communities,
the nature of the relationship between coral diversity
and fish diversity, and the potential consequences of
declines in coral diversity. A direct causal link between
local coral and fish diversity has been established, which
was best explained by habitat selection for shelter, but in
some cases also for food. Our results also highlighted the
variable contributions that different species of corals
make in maintaining patterns of species richness and
community structure of the associated fish assemblages.
Although many coral reef fish may be able to use a
number of microhabitats, selective preference for specific
VANESSA MESSMER ET AL.2296 Ecology, Vol. 92, No. 12

coral species by some species was clearly evident in our
study. The overall potential for a strong, positive
relationship between coral diversity and fish diversity
has the consequence that ongoing degradation of coral
reefs worldwide will probably greatly alter the diversity
and structure of associated fish communities, in partic-
ular due to the loss of specialized species with distinct
resource requirements.
ACKNOWLEDGMENTS
We thank the Lizard Island Research Station (LIRS),Mahonia Na Dari, and the Moorea Coral Reef LTER forlogistical support; K. Chong-Seng, S. Tang Smith, J. Johans-son, P. Saenz Agudelo, M.-E. Portwood, K. Markey, N.Crawley, G. Vima, and M. Giru for assistance in the field; andJ. Claudet for statistical help. Funding was provided by an IanPotter Foundation Fellowship at LIRS and Graduate ResearchScheme (JCU) to V. Messmer; ARC Centre of Excellence forCoral Reef Studies funding to G. P. Jones, and PLM and USANSF funding to R. J. Schmitt, S. J. Holbrook, and A. J.Brooks. This work was carried out under James CookUniversity Ethics Approval No. A1207. Comments fromanonymous reviewers greatly improved the manuscript.
LITERATURE CITED
Anderson, M. J., and T. J. Willis. 2003. Canonical analysis ofprincipal coordinates: A useful method of constrainedordination for ecology. Ecology 84:511–525.
Bell, J. D., and R. Galzin. 1984. Influence of live coral cover oncoral-reef fish communities. Marine Ecology Progress Series15:265–274.
Bellwood, D. R., and C. P. Meyer. 2009. Searching for heat in amarine biodiversity hotspot. Journal of Biogeography36:569–576.
Bellwood, D. R., and P. C. Wainwright. 2002. The history andbiogeography of fishes on coral reefs. Pages 5–32 in P. F.Sale, editor. Coral reef fishes: dynamics and diversity in acomplex ecosystem. Academic Press, San Diego, California,USA.
Bernays, E., and M. Graham. 1988. On the evolution of hostspecificity in phytophagous arthropods. Ecology 69:886–892.
Briggs, J. C. 1999. Coincident biogeographic patterns: Indo-West Pacific Ocean. Evolution 53:326–335.
Chesson, P. 2000. Mechanisms of maintenance of speciesdiversity. Annual Review of Ecology and Systematics31:343–366.
Cole, A. J., M. S. Pratchett, and G. P. Jones. 2008. Diversityand functional importance of coral-feeding fishes on tropicalcoral reefs. Fish and Fisheries 9:286–307.
Crutsinger, G. M., M. D. Collins, J. A. Fordyce, Z. Gompert,C. C. Nice, and N. J. Sanders. 2006. Plant genotypic diversitypredicts community structure and governs an ecosystemprocess. Science 313:966–968.
Currie, D. J. 1991. Energy and large-scale patterns of animal- andplant-species richness. American Naturalist 137:27–49.
De’ath, G. 2002. Multivariate regression trees: a new techniquefor modeling species–environment relationships. Ecology83:1105–1117.
Fahrig, L. 2001. How much habitat is enough? BiologicalConservation 100:65–74.
Feary, D. A., G. R. Almany, G. P. Jones, and M. I.McCormick. 2007. Coral degradation and the structure oftropical reef fish communities. Marine Ecology ProgressSeries 333:243–248.
Futuyma, D. J., and G. Moreno. 1988. The evolution ofecological specialization. Annual Review of Ecology andSystematics 19:207–233.
Gardiner, N. M., and G. P. Jones. 2005. Habitat specialisationand overlap in a guild of coral reef cardinalfishes (Apogo-nidae). Marine Ecology Progress Series 305:163–175.
Gardner, T. A., I. M. Cote, J. A. Gill, A. Grant, and A. R.Watkinson. 2003. Long-term region-wide declines in Carib-bean corals. Science 301:958–960.
Graham, N. A. J., S. K. Wilson, S. Jennings, N. V. C. Polunin,J. P. Bijoux, and J. Robinson. 2006. Dynamic fragility ofoceanic coral reef ecosystems. Proceedings of the NationalAcademy of Sciences USA 203:8425–8429.
Haddad, N. M., D. Tilman, J. Haarstad, M. Ritchie, and J.Knops. 2001. Contrasting effects of plant richness andcomposition on insect communities: a field experiment.American Naturalist 158:17–35.
Hawkins, B. A., and E. E. Porter. 2003. Does herbivorediversity depend on plant diversity? The case of Californiabutterflies. American Naturalist 161:40–49.
Hixon, M. A., and B. A. Menge. 1991. Species diversity: preyrefuges modify the interactive effects of predation andcompetition. Theoretical Population Biology 39:178–200.
Hoegh-Guldberg, O. 1999. Climate change, coral bleaching andthe future of the world’s coral reefs. Marine and FreshwaterResearch 50:839–866.
Holbrook, S. J., A. J. Brooks, and R. J. Schmitt. 2002.Variation in structural attributes of patch-forming corals andin patterns of abundance of associated fishes. Marine andFreshwater Research 53:1045–1053.
Holbrook, S. J., R. J. Schmitt, and A. J. Brooks. 2008.Resistance and resilience of a coral reef fish community tochanges in coral cover. Marine Ecology Progress Series371:264–271.
Hooper, D. U., et al. 2005. Effects of biodiversity on ecosystemfunctioning: a consenus of current knowledge. EcologicalMonographs 75:3–35.
Hortal, J., K. A. Triantis, S. Meiri, E. Thebault, and S.Sfenthourakis. 2009. Island species richness increases withhabitat diversity. American Naturalist 174:E205–E217.
Hughes, T. P., D. R. Bellwood, and S. R. Connolly. 2002.Biodiversity hotspots, centres of endemicity, and the conser-vation of coral reefs. Ecology Letters 5:775–784.
Hutchinson, G. E. 1959. Homage to Santa Rosalia or why arethere so many kinds of animals? American Naturalist 93:145–159.
Jetz, W., H. Kreft, G. Ceballos, and J. Mutke. 2009. Globalassociations between terrestrial producer and vertebrateconsumer diversity. Proceedings of the Royal Society B276:269–278.
Jones, G. P., M. I. McCormick, M. Srinivasan, and J. V. Eagle.2004. Coral decline threatens fish biodiversity in marinereserves. Proceedings of the National Academy of SciencesUSA 101:8251–8253.
Kerr, J. T., and L. Packer. 1997. Habitat heterogeneity as adeterminant of mammal species richness in high-energyregions. Nature 385:252–254.
Kissling, W. D., R. Field, and K. Bohning-Gaese. 2008. Spatialpatterns of woody plant and bird diversity: functionalrelationships or environmental effects? Global Ecology andBiogeography 17:327–339.
Kissling, W. D., C. Rahbek, and K. Bohning-Gaese. 2007.Food plant diversity as broad-scale determinant of avianfrugivore richness. Proceedings of the Royal Society B274:799–808.
Knops, J. M. H., D. Tilman, N. M. Haddad, S. Naeem, C. E.Mitchell, J. Haarstad, M. E. Ritchie, K. M. Howe, P. B.Reich, E. Siemann, and J. Groth. 1999. Effects of plantspecies richness on invasion dynamics, disease outbreaks,insect abundances and diversity. Ecology Letters 2:286–293.
Komyakova, V. 2009. Habitat characteristics as determinantsof the local diversity and structure of coral reef fishcommunities. Thesis. James Cook University, Townsville,Australia.
December 2011 2297CORAL REEF HABITAT AND FISH DIVERSITY

Laurance, W. F. 2007. Have we overstated the tropicalbiodiversity crisis? Trends in Ecology and Evolution 22:65–70.
Lawton, J. H. 1983. Plant architecture and the diversity ofphytophagous insects. Annual Review of Entomology 28:23–39.
Loreau, M., S. Naeem, P. Inchausti, J. Bengtsson, J. P. Grime,A. Hector, D. U. Hooper, M. A. Huston, D. Raffaelli, B.Schmid, D. Tilman, and D. A. Wardle. 2001. Biodiversityand ecosystem functioning: current knowledge and futurechallenges. Science 294:804–808.
Loya, Y., K. Sakai, K. Yamazato, Y. Nakano, H. Sambali, andR. van Woesik. 2001. Coral bleaching: the winners and thelosers. Ecology Letters 4:122–131.
Luckhurst, B. E., and K. Luckhurst. 1978. Analysis of theinfluence of substrate variables on coral reef fish communi-ties. Marine Biology 49:317–323.
Marquez, A. L., R. Real, and J. M. Vargas. 2004. Dependenceof broad-scale geographical variation in fleshy-fruited plantspecies richness and disperser bird species richness. GlobalEcology and Biogeography 13:295–304.
Marshall, P. A., and A. H. Baird. 2000. Bleaching of corals onthe Great Barrier Reef: differential susceptibilities amongtaxa. Coral Reefs 19:155–163.
McClanahan, T. R. 2002. The near future of coral reefs.Environmental Conservation 29:460–483.
McClanahan, T. R., M. Ateweberhan, N. A. J. Graham, S. K.Wilson, C. R. Sebastian, M. M. M. Guillaume, and J. H.Bruggemann. 2007. Western Indian Ocean coral communi-ties: bleaching responses and susceptibility to extinction.Marine Ecology Progress Series 337:1–13.
Menendez, R., A. Gonzalez-Megias, Y. Collingham, R. Fox,D. B. Roy, R. Ohlemuller, and C. D. Thomas. 2007. Directand indirect effects of climate and habitat factors on butterflydiversity. Ecology 88:605–611.
Mora, C., et al. 2011. Global human footprint on the linkagebetween biodiversity and ecosystem functioning in reef fishes.PLoS Biology 9(4):e1000606.
Mora, C., and D. R. Robertson. 2005. Causes of latitudinalgradients in species richness: A test with fishes of the tropicalEastern Pacific. Ecology 86:1771–1782.
Morris, D. W. 2003. Toward an ecological synthesis: a case forhabitat selection. Oecologia 136:1–13.
Munday, P. L. 2000. Interactions between habitat use andpatterns of abundance in coral-dwelling fishes of the genusGobiodon. Environmental Biology of Fishes 58:355–369.
Munday, P. L. 2004. Habitat loss, resource specialization, andextinction on coral reefs. Global Change Biology 10:1642–1647.
Munday, P. L., G. P. Jones, and M. J. Caley. 1997. Habitatspecialisation and the distribution and abundance of coral-dwelling gobies. Marine Ecology Progress Series 152:227–239.
Murdoch, W. W., F. C. Evans, and C. H. Peterson. 1972.Diversity and pattern in plants and insects. Ecology 53:819–829.
Novotny, V., P. Drozd, S. E. Miller, M. Kulfan, M. Janda, Y.Basset, and G. D. Weiblen. 2006. Why are there so manyspecies of herbivorous insects in tropical rainforests? Science313:1115–1118.
Ohman, M. C., and A. Rajasuriya. 1998. Relationships betweenhabitat structure and fish communities on coral. Environ-mental Biology of Fishes 53:19–31.
Pratchett, M. S. 2005. Dietary overlap among coral-feedingbutterflyfishes (Chaetodontidae) at Lizard Island, northernGreat Barrier Reef. Marine Biology 148:373–382.
Pratchett, M. S., and M. L. Berumen. 2008. Interspecificvariation in distributions and diets of coral reef butterfly-fishes (Teleostei: Chaetodontidae). Journal of Fish Biology73:1730–1747.
Pratchett, M. S., P. L. Munday, S. K. Wilson, N. A. J. Graham,J. E. Cinner, D. R. Bellwood, G. P. Jones, N. V. C. Polunin,and T. R. McClanahan. 2008. Effects of climate-inducedcoral bleaching on coral-reef fishes; ecological and economicconsequences. Oceanography and Marine Biology: AnAnnual Review 46:251–296.
Qian, H., W. D. Kissling, X. L. Wang, and P. Andrews. 2009.Effects of woody plant species richness on mammal speciesrichness in southern Africa. Journal of Biogeography36:1685–1697.
R Development Core Team. 2009. R 2.10.0. R Foundation forStatistical Computing, Vienna, Austria. hhttp://cran.r-project.org/bin/i
Rahbek, C., and G. R. Graves. 2001. Multiscale assessment ofpatterns of avian species richness. Proceedings of theNational Academy of Sciences USA 98:4534–4539.
Ralph, C. J. 1985. Habitat association patterns of forest andsteppe birds of northern Patagonia, Argentina. Condor87:471–483.
Roberts, C. M., and R. F. G. Ormond. 1987. Habitat complexityand coral reef fish diversity and abundance on Red Seafringing reefs. Marine Ecology Progress Series 41:1–8.
Sale, P. F. 1991. Habitat structure and recruitment in coral reeffishes. Pages 197–210 in S. S. Bell, E. D. McCoy, and H. R.Mushinsky, editors. Habitat structure, the physical arrange-ment of objects in space. Chapman and Hall, London, UK.
Siemann, E., D. Tilman, J. Haarstad, and M. Ritchie. 1998.Experimental tests of the dependence of arthropod diversityon plant diversity. American Naturalist 152:738–750.
Tews, J., U. Brose, V. Grimm, K. Tielborger, M. C. Wichmann,M. Schwager, and F. Jeltsch. 2004. Animal species diversitydriven by habitat heterogeneity/diversity: the importance ofkeystone structures. Journal of Biogeography 31:79–92.
Vitousek, P. M., H. A. Mooney, J. Lubchenco, and J. M.Melillo. 1997. Human domination of Earth’s ecosystems.Science 277:494–499.
Wilkinson, C. R. 2004. Status of coral reefs of the world.Australian Institute of Marine Science, Townsville, Queens-land, Australia.
Wilson, S. K., S. C. Burgess, A. J. Cheal, M. J. Emslie, R.Fisher, I. Miller, N. V. C. Polunin, and H. P. A. Sweatman.2008. Habitat utilization by coral reef fish: implications forspecialists versus generalists in a changing environment.Journal of Animal Ecology 77:220–228.
Wilson, S. K., N. A. J. Graham, M. S. Pratchett, G. P. Jones,and N. V. C. Polunin. 2006. Multiple disturbances and theglobal degradation of coral reefs: Are reef fishes at risk orresilient? Global Change Biology 12:2220–2234.
Wolters, V., J. Bengtsson, and A. S. Zaitsev. 2006. Relationshipamong the species richness of different taxa. Ecology87:1886–1895.
APPENDIX
Additional figures and tables, including a map showing study locations, a CAP analysis demonstrating differences in fishcommunities between locations, and the species lists for Figs. 3B and 4B (Ecological Archives E092-198-A1).
VANESSA MESSMER ET AL.2298 Ecology, Vol. 92, No. 12





























