Embed Size (px)
Citation preview

1 23
Marine BiologyInternational Journal on Life in Oceansand Coastal Waters ISSN 0025-3162 Mar BiolDOI 10.1007/s00227-012-2066-7
Temporal and spatial variability in coralrecruitment on two Indonesian coralreefs: consistently lower recruitment to adegraded reef
P. Salinas-de-León, C. Dryden,D. J. Smith & J. J. Bell

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL PAPER
Temporal and spatial variability in coral recruitment on twoIndonesian coral reefs: consistently lower recruitmentto a degraded reef
P. Salinas-de-Leon • C. Dryden • D. J. Smith •
J. J. Bell
Received: 25 January 2012 / Accepted: 30 August 2012
� Springer-Verlag 2012
Abstract Corals are the primary reef-building organisms,
therefore it is key to understand their recruitment patterns
for effective reef management. Coral recruitment rates and
juvenile coral abundance were recorded in the Wakatobi
National Marine Park, Indonesia, on two reefs (Sampela
and Hoga) with different levels of environmental degra-
dation (12.5 vs. 44 % coral cover with high and low sed-
imentation rates, respectively) to examine consistencies in
recruitment patterns between years and seasons. Recruit-
ment was measured on multiple panels at two sites on each
reef (6–7 m depth) and cleared areas of natural reef.
Although coral recruitment was twofold higher in
2008–2009 than in 2007–2008, and seasonal differences
were identified, consistent significant differences in
recruitment rates were found between the two reefs even
though they are separated by only *1.5 km. Recruitment
rates and juvenile abundance were lower on the more
degraded reef. These patterns are likely a consequence of
differential pre- and post-settlement mortality as a result of
the high sedimentation rates and degraded conditions and
possibly reduced larval supply.
Introduction
Understanding temporal and spatial variation in recruit-
ment patterns is critical for measuring population dynamics
and effectively managing marine populations, and for
understanding the resilience of coral reef communities to
disturbance (Hughes et al. 2010). Explaining such
dynamics are particularly important for coral reef systems
as they are in decline worldwide as a consequence of
natural and anthropogenic disturbance (e.g. Hughes 1994;
Bellwood et al. 2004; Mora 2008). For example, Indo-
Pacific coral reefs are seriously threatened, and recent
estimates show a 32 % region-wide decline in coral cover
since the 1970s (Bruno and Selig 2007).
Recruitment has long been recognised as one of the most
important factors driving the ecology of marine inverte-
brates and is critical for the maintenance of viable reef
populations and for promoting the recovery of coral reefs
after disturbance (Babcock and Mundy 1996; Connell et al.
1997; Hughes et al. 2000). Here, we define recruitment as
the number of new individuals that settle and survive until
the time of observation, which reflects patterns of larva
availability, larva substrate selection and post-settlement
mortality rates (Harrison and Wallace 1990; Connell et al.
1997).
Despite Burke et al. (2002) suggesting that [85 % of
Indonesia’s reefs are threatened by anthropogenic impacts
and Edinger et al. (1998) showing a drastic loss of coral
biodiversity in Indonesian reefs due to land-based
Communicated by J. P. Grassle.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00227-012-2066-7) contains supplementarymaterial, which is available to authorized users.
P. Salinas-de-Leon (&) � J. J. Bell
Centre for Marine Environmental and Economic Research,
School of Biological Sciences, Victoria University of
Wellington, PO Box 600, Wellington, New Zealand
e-mail: [email protected]
C. Dryden
School of Marine Science and Technology, Newcastle
University, Armstrong Building, Newcastle Upon
Tyne NE1 7RU, UK
D. J. Smith
Coral Reef Research Unit, Department of Biological Sciences,
University of Essex, Wivenhoe Park, Colchester CO4 3SQ, UK
123
Mar Biol
DOI 10.1007/s00227-012-2066-7
Author's personal copy

pollution, only a few coral recruitment studies have been
conducted in the Coral Triangle region (Tomascik et al.
1996; Reyes and Yap 2001; Fox et al. 2003; Fox 2004;
Schmidt-Roach et al. 2008). Of these studies, only Fox
et al. (2003) and Fox (2004) were conducted in more than
one locality and were temporally replicated. Therefore,
more information is needed on hard coral population
dynamics in this biodiverse region to assist in the devel-
opment of effective management plans.
Scleractinian coral recruitment is influenced by a num-
ber of factors including the amount of live coral cover in
source populations (Hughes et al. 2000), abundance and
diversity of coral larvae (Potts et al. 1985), hydrodynamic
variability (Amar et al. 2007), sedimentation (Babcock and
Davies 1991), temperature (Nozawa and Harrison 2007),
recruitment cues and inhibition from other benthic taxa
(Harrington et al. 2004), eutrophication levels (Tomascik
1991), grazing pressure (Sammarco 1980), light levels
(Mundy and Babcock 1998) and connectivity between reefs
(Roberts 1997). Some of these, such as the presence of
crustose coralline algae (CCA), positively affect coral
recruitment, while others, such as increased sedimentation
or competition with macroalgae and/or soft corals, nega-
tively affect recruitment (Benayahu and Loya 1985; Bab-
cock and Davies 1991; Maida et al. 1995).
In a preliminary 1-year study in 2007–2008 (Salinas-de-
Leon et al. 2011) on the same reefs used in the present
study (Hoga and Sampela reefs in Indonesia), we charac-
terised the benthic habitats with respect to coral taxonomic
composition and coverage by different organisms. We also
examined coral recruitment on cleared areas of vertical reef
wall and on concrete and terracotta settlement panels. From
this study, we concluded that there were significant dif-
ferences between the Hoga and Sampela sites with respect
to coral cover (*44 vs. 12.5 %, respectively) and taxo-
nomic composition, with Acroporidae and Poritidae being
most abundant at Hoga, and Faviidae and Poritidae being
most abundant at Sampela. Coral recruitment was similar
on the different substrates but differed significantly
between Hoga and Sampela reefs. We concluded that ter-
racotta panels secured to vertical reef faces provide a
suitable substrate for assessing relative recruitment rates in
this environment. Here, we extended this earlier study into
a second year (2008–2009) and also conducted a seasonal
study to determine whether the differences between the
Hoga and Sampela reefs were consistent. The objectives of
this study were to (1) compare coral recruitment rates to
reefs with different levels of environmental degradation in
the Wakatobi National Marine Park (WNMP), SE Sulaw-
esi, Indonesia (using the same sites as Salinas-de-Leon
et al. 2011); and (2) examine the consistency of any pat-
terns between 2 years and among three seasons. We used
three approaches to these objectives including measuring
coral recruitment rates to natural reef substrate (annual
patterns only) and artificial settlement panels and juvenile
coral abundance. The seasonal study in 2008–2009 exam-
ined three consecutive 4-month periods. Indonesia’s cli-
mate is dominated by two monsoon seasons each year,
driven by the Inter-tropical Convergence Zone, with the
north-west monsoon (wet season) occurring between
November and March and the south-east monsoon (dry
season) between May and September (Tomascik et al.
1997).
Materials and methods
Study sites
The WNMP is in the Tukang Besi Archipelago, an island
group off south-east Sulawesi in Indonesia (Fig. 1). The
park represents Indonesia’s third largest Marine Protected
Area, which was designated in 1992 and covers 1.39 mil-
lion hectares. The park lies at the heart of the ‘Coral Tri-
angle’ and supports extremely high coral diversity with
more than 390 hermatypic scleractinian coral species
belonging to 68 genera and 13 families (Turak 2003).
Sampling
Our study was conducted at four sites on two reefs (con-
sidered two replicate sites on each reef). Two sites (B3, B4;
terminology for sites is based on earlier studies in this
region) were on the Hoga reef (5�2803000S, 123�4501800E)
and two sites (S1, S2) on the Sampela reef (5�2805100S,
123�44’3900E); (Fig. 1). The reefs are *1.5 km apart and
Fig. 1 Sampling sites within Wakatobi National Marine Park in SE
Sulawesi, Indonesia. Sites B3 and B4 were on Hoga reef, and sites S1
and S2 on Sampela reef, adjacent to a Bajau village (represented on
map by three white houses). Black areas on map represent land and
grey areas show coral reefs. Map modified from Salinas-de-Leon
et al. (2011)
Mar Biol
123
Author's personal copy

separated by a deep channel ([60 m). Sites within reefs were
at least 250 m apart and were selected based on earlier ben-
thic surveys that revealed higher live coral coverage at Hoga
B3 and B4 (*44 %) compared to Sampela S1 and S2
(*12.5 %); (Salinas-de-Leon et al. 2011). Sites S1 and S2
are near the Bajau (Sea Gypsy) village of Sampela and are
subjected to severe anthropogenic disturbance, including
overfishing, blast and cyanide fishing, coral mining and
sedimentation due to untreated sewage discharge and man-
grove removal (Crabbe and Smith 2005).
Site characterisation
Benthic habitat composition was characterised in July 2007
at each site with respect to live coral cover, taxonomic
composition of the coral community and abundance of
other benthic taxa. This information was collected because
in some taxa, the relative abundance of spat populations is
similar to that of the adult populations (although not for all
species, reviewed by Pineda 2000), and the presence of
other benthic groups also influences coral recruitment
(Smith 1992; Harrington et al. 2004). Benthic habitat
composition was determined using the continual line
intercept method (English et al. 1997). Ten haphazardly
placed 10-m transects were sampled at 6 m depth using the
line intercept method at each of the four study sites. For
each transect, all coral colonies lying underneath the
transect tape were identified to genus level and recorded.
Other benthic categories included bare substrate, sand,
coral rubble, dead coral, soft coral, algae, crustose coralline
algae (CCA), sponges and ascidians.
Temperature was recorded throughout the study period
using HOBO pendant temperature loggers (Onset Corpora-
tion, USA) placed at 6 m depth at two sites (B3, S1), one on
each reef system. Temperature data were smoothed based on
mean data averaged over 10-min intervals, and averaged
across the four sites, by the logger software (Onset). Current
strength and direction were measured using a Valeport model
106 flow meter (Valeport Oceanographic instruments, UK)
positioned at 7–8 m depth. Only 3 9 24 h deployments of the
flow meter, during both neap and spring tides, at each of the
four study sites were conducted due to logistical limitations.
Sedimentation rates were estimated by placing three replicate
sediment traps (plastic cylinders 7 cm diameter and 20 cm tall
(English et al. 1997) at 6 m depth for 7 days at each site during
November 2008 and July 2009. Sediment was washed out of
the traps and filtered, and then oven dried for 24 h and
weighed to the nearest 0.01 g.
Recruitment
The panels deployed to monitor coral recruitment were
made of locally available unglazed terracotta
(20 9 10 9 0.7 cm). Panels were directly attached to the
reef substrate using a modification of the method of Mundy
(2000). Terracotta panels were cable tied to thin (0.5 cm)
hardwood planks. The wood was then drilled, and
recruitment panels were attached to vertical surfaces on the
reef using two galvanised nails, a material widely used in
previous recruitment studies (e.g. Harriott and Fisk 1987;
Mangubhai et al. 2007). A small piece of hardwood
(2 9 2 9 2 cm) was placed between each panel and the
reef to leave a 2-cm gap, as this is known to facilitate coral
recruitment (Harriott and Fisk 1987); (See ESM 1 for a
diagrammatic representation of the panel set up). Eight
panels were haphazardly placed between 6 and 7 m depth
at each of the four study sites, and at least 5 m separated
the panels from one another; panels were attached to ver-
tical reef surfaces based on Salinas-de-Leon et al. (2011).
Only corals recruiting to the back surface of the panels
(closest to the reef) were included in the analysis as 99 %
of all corals recruited to this side of the tiles.
Panels were submerged for two periods of time to record
yearly and seasonal coral recruitment rates and therefore
examine the consistency of the patterns on the two different
reefs over time. (A) Panels were deployed for 12 mo
between July 2007 and July 2008; these panels were a
subset of those used by Salinas-de-Leon et al. (2011). Upon
retrieval, new panels were placed during July 2008 and
retrieved 12 mo later in July 2009. (B) Panels were also
deployed in July 2008 to assess seasonal recruitment, and
were retrieved and replaced by 8 more panels every 4 mo,
providing three 9 4 mo recruitment data sets: July 2008 to
November 2008, November 2008 to March 2009 and
March 2009 to July 2009. These periods of time coincided
with major climatic changes within the study region.
When panels were collected, they were labelled and
transported back to the laboratory in sea water. They then
were photographed and panels were bleached in a chlorine
solution for 24 h, then rinsed in fresh water to remove
excess bleach before being air dried. Each panel was
searched twice by two different observers at two different
microscope magnifications (109 and 209), and all coral
recruits were recorded. The panel edges were not analysed
given their small area (i.e. panels were 0.7 cm thick).
Recruits were identified as belonging to the families
Acroporidae, Pocilloporidae and Poritidae based on pho-
tomicrographs in English et al. (1997) and Babcock et al.
(2003), with the remaining spat, including those too dam-
aged to identify, being designated as ‘others’.
Recruitment to natural reef cleared areas
Four reef areas (25 9 25 cm) were cleared of all living
organisms at each of the four sites, to determine coral
recruitment rates to natural substrata. Permanent cleared
Mar Biol
123
Author's personal copy

areas were created in July 2007 and July 2008, and were
surveyed after 12 mo in July 2008 and July 2009, respec-
tively. These areas were haphazardly located on vertical
walls near the recruitment panels and were permanently
marked with four galvanised nails. Cleared areas were at
least 5 m apart.
Cleared areas were surveyed by eye during the day and
at night. Initial surveys were conducted during the day, and
all coral recruits present in the quadrats were recorded and
identified to the lowest taxonomic level possible. Observed
recruits were marked using a small nail. A second survey
was conducted at night using the Underwater Fluorescence
Technique of Piniak et al. (2005). Permanent cleared areas
were resurveyed with a blue light (Nightsea Inc.) and with
a yellow barrier filter placed over the diver’s mask
(Nightsea Inc.) to facilitate the detection of coral recruits.
False positives can be caused by a number of other
organisms that also fluoresce, but we minimised this effect
by using a strong white light as described by Baird et al.
(2006). Recruits missed under daylight conditions were
recorded, measured and identified to the lowest taxonomic
level possible; results were presented as the total number of
recruits recorded. The data collected from areas cleared in
2007 and surveyed in 2008 were the same as those used by
Salinas-de-Leon et al. (2011) to compare with recruitment
rates on panels made of different materials.
Juvenile coral abundance and diversity
Juvenile coral abundance and diversity were assessed in
July 2008 using 20 0.5 m 9 0.5 m (0.25 m2) quadrats
positioned haphazardly along 50-m transects at 6 m depth
at each of the four sampling sites. Prior to surveying the
quadrats, sediment was removed from the quadrat by
gently wafting the quadrat area; this facilitated counts of
the small juvenile corals. Juvenile corals were defined as
colonies \40 mm in diameter that were attached to the
substratum and did not have the fractured surface charac-
teristic of asexual recruits (Edmunds 2000). Corals present
in the quadrats were identified to genus level.
Statistical analysis
Data were analysed by the statistical packages MINITAB
v16, PRIMER v6 and PERMANOVA v1.0.2. Differences
in benthic cover composition among sites were analysed
using a Kruskal–Wallis test for each of the benthic cate-
gories (as data were not normally distributed). The
recruitment and juvenile coral abundance data were not
normally distributed even after transformation and vari-
ances were unequal, so the data were analysed with a
permutational ANOVA. PERMANOVA is a permutation-
based version of analysis of variance (Anderson 2001). It
uses the distances between samples to partition variance
and randomisations or permutations of the data to produce
the p value for the hypothesis test. It is non-parametric and
is therefore robust to the assumption of multivariate nor-
mality, making it less prone to Type I errors. A preliminary
PERMANOVA analysis was used to investigate the dif-
ferences in recruitment rates to panels, cleared reef areas
and abundance of juvenile corals including the factors year
(season), reef (Hoga, Sampela) and site (B3, B4, S1, S2).
With this preliminary analysis, we found no significant
differences at the site level, so the factor site was removed
from the final analysis (data not shown).
Differences in the yearly recruitment rates to panels and
to cleared reef areas were analysed using a PERMANOVA
model with two fixed factors: year (two levels: 2008, 2009)
and reef (two levels: Hoga and Sampela). The PERMA-
NOVA seasonal recruitment data statistical design had two
fixed factors: season (three levels: November, March and
July) and reef (two levels: Hoga and Sampela). The PER-
MANOVA model created for the juvenile abundance data
included just the factor reef (two levels: Hoga and Sam-
pela), given that surveys were only conducted in 2008.
Results
Biological and physical characteristics of the study sites
The Hoga and Sampela reefs had significantly different
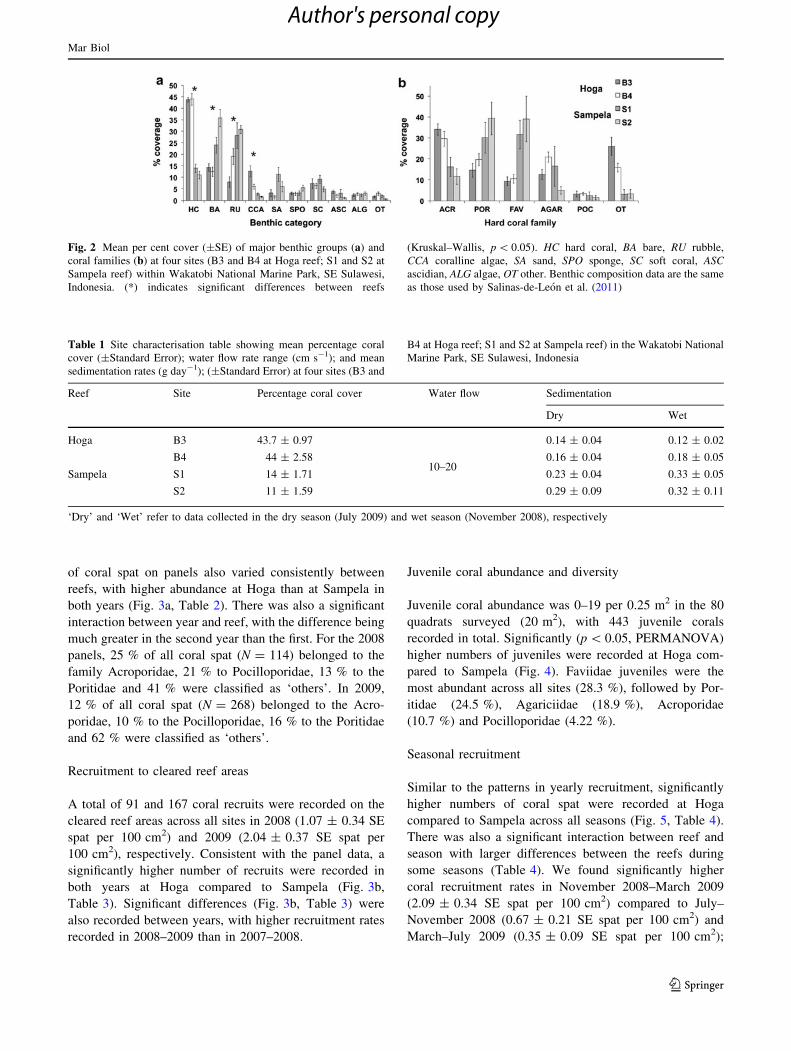
benthic substrata and community composition (Fig. 2a, b).
The Hoga reef was dominated by hard coral, while the
Sampela reef was dominated by bare substrate and coral
rubble (Table 1). There were also significant differences
(p \ 0.05, Kruskal–Wallis) in the percentage cover of
CCA and sand between the two reefs (Fig. 2a), with higher
CCA abundance at Hoga and higher sand cover at Sampela.
The Hoga reef coral assemblage (at 6-m depth) was dom-
inated by Acroporidae and Poritidae, while the Sampela
reef coral assemblage was dominated by members of the
family Faviidae and Poritidae (Fig. 2b).
The Hoga and Sampela reefs had similar water flow
rates and water temperatures (2 years range: 26.8–29.9 �C)
over the study period (Table 1). Sedimentation rates were
consistently higher on the Sampela reef, compared to the
Hoga reef during both the wet and dry seasons (Table 1).
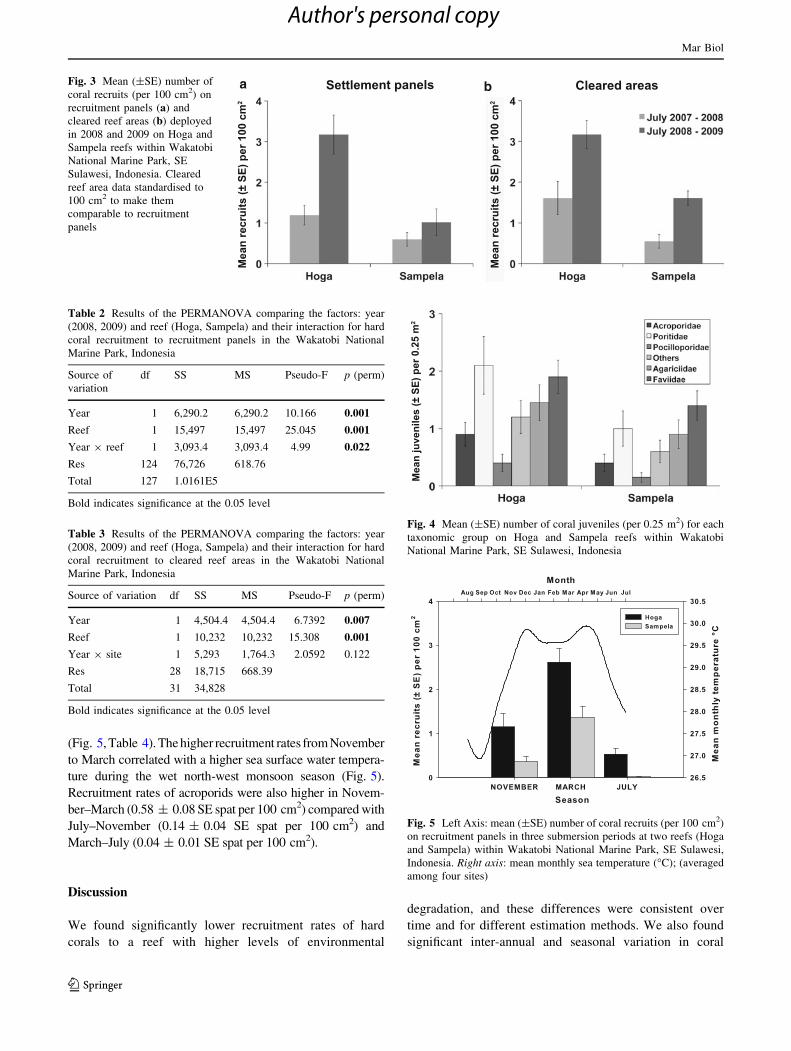
Annual recruitment
A total of 114 and 268 coral spat were recorded on the 32
panels across all sites in 2008 (0.89 ± 0.28 SE spat per
cm2) and 2009 (2.09 ± 0.57 SE spat per 100 cm2),
respectively. Inter-annual variation in coral recruitment
was highly significant (Fig. 3a, Table 2), and the number
Mar Biol
123
Author's personal copy
of coral spat on panels also varied consistently between
reefs, with higher abundance at Hoga than at Sampela in
both years (Fig. 3a, Table 2). There was also a significant
interaction between year and reef, with the difference being
much greater in the second year than the first. For the 2008
panels, 25 % of all coral spat (N = 114) belonged to the
family Acroporidae, 21 % to Pocilloporidae, 13 % to the
Poritidae and 41 % were classified as ‘others’. In 2009,
12 % of all coral spat (N = 268) belonged to the Acro-
poridae, 10 % to the Pocilloporidae, 16 % to the Poritidae
and 62 % were classified as ‘others’.
Recruitment to cleared reef areas
A total of 91 and 167 coral recruits were recorded on the
cleared reef areas across all sites in 2008 (1.07 ± 0.34 SE
spat per 100 cm2) and 2009 (2.04 ± 0.37 SE spat per
100 cm2), respectively. Consistent with the panel data, a
significantly higher number of recruits were recorded in
both years at Hoga compared to Sampela (Fig. 3b,
Table 3). Significant differences (Fig. 3b, Table 3) were
also recorded between years, with higher recruitment rates
recorded in 2008–2009 than in 2007–2008.
Juvenile coral abundance and diversity
Juvenile coral abundance was 0–19 per 0.25 m2 in the 80
quadrats surveyed (20 m2), with 443 juvenile corals
recorded in total. Significantly (p \ 0.05, PERMANOVA)
higher numbers of juveniles were recorded at Hoga com-
pared to Sampela (Fig. 4). Faviidae juveniles were the
most abundant across all sites (28.3 %), followed by Por-
itidae (24.5 %), Agariciidae (18.9 %), Acroporidae
(10.7 %) and Pocilloporidae (4.22 %).
Seasonal recruitment
Similar to the patterns in yearly recruitment, significantly
higher numbers of coral spat were recorded at Hoga
compared to Sampela across all seasons (Fig. 5, Table 4).
There was also a significant interaction between reef and
season with larger differences between the reefs during
some seasons (Table 4). We found significantly higher
coral recruitment rates in November 2008–March 2009
(2.09 ± 0.34 SE spat per 100 cm2) compared to July–
November 2008 (0.67 ± 0.21 SE spat per 100 cm2) and
March–July 2009 (0.35 ± 0.09 SE spat per 100 cm2);
Fig. 2 Mean per cent cover (±SE) of major benthic groups (a) and
coral families (b) at four sites (B3 and B4 at Hoga reef; S1 and S2 at
Sampela reef) within Wakatobi National Marine Park, SE Sulawesi,
Indonesia. (*) indicates significant differences between reefs
(Kruskal–Wallis, p \ 0.05). HC hard coral, BA bare, RU rubble,
CCA coralline algae, SA sand, SPO sponge, SC soft coral, ASCascidian, ALG algae, OT other. Benthic composition data are the same
as those used by Salinas-de-Leon et al. (2011)
Table 1 Site characterisation table showing mean percentage coral
cover (±Standard Error); water flow rate range (cm s-1); and mean
sedimentation rates (g day-1); (±Standard Error) at four sites (B3 and
B4 at Hoga reef; S1 and S2 at Sampela reef) in the Wakatobi National
Marine Park, SE Sulawesi, Indonesia
Reef Site Percentage coral cover Water flow Sedimentation
Dry Wet
Hoga B3 43.7 ± 0.97
10–20
0.14 ± 0.04 0.12 ± 0.02
B4 44 ± 2.58 0.16 ± 0.04 0.18 ± 0.05
Sampela S1 14 ± 1.71 0.23 ± 0.04 0.33 ± 0.05
S2 11 ± 1.59 0.29 ± 0.09 0.32 ± 0.11
‘Dry’ and ‘Wet’ refer to data collected in the dry season (July 2009) and wet season (November 2008), respectively
Mar Biol
123
Author's personal copy
(Fig. 5, Table 4). The higher recruitment rates from November
to March correlated with a higher sea surface water tempera-
ture during the wet north-west monsoon season (Fig. 5).
Recruitment rates of acroporids were also higher in Novem-
ber–March (0.58 ± 0.08 SE spat per 100 cm2) compared with
July–November (0.14 ± 0.04 SE spat per 100 cm2) and
March–July (0.04 ± 0.01 SE spat per 100 cm2).
Discussion
We found significantly lower recruitment rates of hard
corals to a reef with higher levels of environmental
degradation, and these differences were consistent over
time and for different estimation methods. We also found
significant inter-annual and seasonal variation in coral
a bFig. 3 Mean (±SE) number of
coral recruits (per 100 cm2) on
recruitment panels (a) and
cleared reef areas (b) deployed
in 2008 and 2009 on Hoga and
Sampela reefs within Wakatobi
National Marine Park, SE
Sulawesi, Indonesia. Cleared
reef area data standardised to
100 cm2 to make them
comparable to recruitment
panels
Table 2 Results of the PERMANOVA comparing the factors: year
(2008, 2009) and reef (Hoga, Sampela) and their interaction for hard
coral recruitment to recruitment panels in the Wakatobi National
Marine Park, Indonesia
Source of
variation
df SS MS Pseudo-F p (perm)
Year 1 6,290.2 6,290.2 10.166 0.001
Reef 1 15,497 15,497 25.045 0.001
Year 9 reef 1 3,093.4 3,093.4 4.99 0.022
Res 124 76,726 618.76
Total 127 1.0161E5
Bold indicates significance at the 0.05 level
Table 3 Results of the PERMANOVA comparing the factors: year
(2008, 2009) and reef (Hoga, Sampela) and their interaction for hard
coral recruitment to cleared reef areas in the Wakatobi National
Marine Park, Indonesia
Source of variation df SS MS Pseudo-F p (perm)
Year 1 4,504.4 4,504.4 6.7392 0.007
Reef 1 10,232 10,232 15.308 0.001
Year 9 site 1 5,293 1,764.3 2.0592 0.122
Res 28 18,715 668.39
Total 31 34,828
Bold indicates significance at the 0.05 level
Fig. 4 Mean (±SE) number of coral juveniles (per 0.25 m2) for each
taxonomic group on Hoga and Sampela reefs within Wakatobi
National Marine Park, SE Sulawesi, Indonesia
Fig. 5 Left Axis: mean (±SE) number of coral recruits (per 100 cm2)
on recruitment panels in three submersion periods at two reefs (Hoga
and Sampela) within Wakatobi National Marine Park, SE Sulawesi,
Indonesia. Right axis: mean monthly sea temperature (�C); (averaged
among four sites)
Mar Biol
123
Author's personal copy

recruitment rates in the WNMP. Understanding recruitment
dynamics and identifying the factors responsible for tem-
poral and spatial variation in such patterns are important
for effectively managing marine populations. The first
stage in understanding such dynamics is to describe vari-
ation patterns and propose hypotheses to explain the pat-
terns observed.
We propose a number of hypotheses to explain the
consistent differences in the number of coral recruits
between the Hoga and Sampela reefs. A likely explanation
for the differences we report is the effect of settlement-
related processes (such as habitat difference or larval
selection) and post-settlement mortality. Sedimentation
levels were much higher at Sampela reef during both the
dry (July) and wet (November) seasons. Sedimentation is
known to affect the growth and calcification (Tomascik and
Sander 1985), reproduction (Tomascik and Sander 1987b),
respiration, feeding and photosynthesis (Fabricius 2005),
fecundity (Tomascik and Sander 1987b), community
structure (Tomascik and Sander 1987a) and more impor-
tantly, the larval settlement of scleratinian corals (Hunte
and Wittenberg 1992). The effects of sedimentation vary
considerably among coral species, but also between sedi-
ment types and environmental conditions (Fabricius 2005).
The lower recruitment rates reported at Sampela might in
part be influenced by the higher sedimentation rates
recorded there. Sedimentation and associated turbidity are
known to affect planula settlement behaviour through
direct physical action in the water column (Tomascik 1991)
and by inhibiting actual planula settlement (Babcock and
Davies 1991). We found that the majority of the recruits
settled on the back (cryptic) side of the panels (Harriott and
Fisk 1987), and this habitat has low sediment accumulation
rates, but the effects of suspended sediment are still pres-
ent, particularly the light reduction, which is known to be
important for corals (Mundy and Babcock 1998).
Habitat differences might also explain the higher post-
recruitment mortality rates at Sampela compared to Hoga.
Our benthic surveys revealed higher levels of coral rubble
at Sampela, and rubble fields created by chronic blast
fishing are unsuitable for coral recruit survival. This is
because spat settling onto coral rubble are killed by the
periodic shifting of the rubble with strong currents (Fox
et al. 2003). Finally, differences in predation rates could
explain the recruitment rate differences between reefs, as
fish abundance (see Bell and Smith 2004) and assemblage
composition, along with other potential predators are likely
to be different between the sites, and predation influences
coral recruitment (e.g. Sammarco 1980). However, since
most coral recruits were on the backs of the panels, we
were able to compare recruitment between the two reefs. A
future experimental approach will investigate these factors
further to determine which factor or combination of factors
explains the patterns we have reported.
These recruitment differences could also be affected by
differences in the live coral coverage between the reefs,
being significantly lower at Sampela than at Hoga. How-
ever, while there is some evidence that a high proportion of
the coral larvae produced locally might be retained close to
the parental population (Sammarco and Andrews 1988;
Underwood et al. 2007; Underwood et al. 2009), the rela-
tively close proximity of the two reefs (1.5 km) also sug-
gests they may share a common pool of larvae, particularly
for species with long-lived pelagic larvae. In contrast to our
results, other authors have not found recruitment differ-
ences between reefs with different levels of coral cover or
taxonomic composition. For example, Fox (2004) found no
differences in coral recruitment rates to panels at blasted
and unblasted reefs, despite the reefs having greater spatial
separation than our sites; this suggests that low-coral-cover
sites are not necessarily limited by larval supply.
The number of coral spat per panel recorded at sites
within the Wakatobi is slightly lower than those recorded
in other recruitment studies within Indonesia (Manado and
Komodo), and much lower than those at many locations
along the central Great Barrier Reef (Baird and Hughes
2000; Fox 2004; Schmidt-Roach et al. 2008). However, it
is difficult to interpret differences in recruitment rates
because of methodological and site-specific differences
between studies, which are known to influence those rates
(Glassom et al. 2004). Importantly, the patterns we repor-
ted and the recruitment rates measured were consistent
between natural areas of reef and panels on the same reef
suggesting that the panels provided a suitable surrogate for
recruitment rates to these reefs (for further discussion, see
Salinas-de-Leon et al. 2011).
Recruitment was twofold higher during the second year
of our study compared to the first, and the differences
between reefs were also greater in year 2. Such inter-
annual variation has been recorded in other studies
including in the Komodo National Park in Indonesia and
also in other biogeographical regions, such as the Red Sea
Table 4 Results of the PERMANOVA comparing the factors: season
(November, March and July) and reef (Hoga, Sampela) and their
interactions for hard coral recruitment to terracotta recruitment panels
in the Wakatobi National Marine Park, Indonesia
Source of
variation
df SS MS Pseudo-F p (perm)
Season 2 55,222 27,611 46.959 0.001
Reef 1 29,616 29,616 50.369 0.001
Season 9 reef 2 4,204.9 2,102.4 3.5757 0.014
Res 186 1.09E ? 05 587.98
Total 191 1.98E ? 05
Bold indicates significance at the 0.05 level
Mar Biol
123
Author's personal copy

(Glassom et al. 2004) and the GBR, Australia (Harriott and
Banks 1995; Dunstan and Johnson 1998). This annual
variability might be caused by a number of factors, for
example, some corals show considerable variation in their
reproductive output between years (Hughes et al. 2000), or
by differences in the physical processes that influence
larval transport and supply (Pineda 2000).
Many of the seasonal differences reported in our study
are likely to be driven by differences between coral fami-
lies in their reproductive modes (i.e. brooding vs. broad-
casting) and because different species are likely to spawn at
different times of the year. Broadcasting corals often
exhibit spawning synchrony within populations (Harrison
et al. 1984). Richmond and Hunter (1990) predicted that
for equatorial regions, where sea surface temperature
variations and tidal amplitude are small, reproductive
seasonality and synchrony among species would be
reduced. In contrast to this prediction, recent work on coral
reefs close to equatorial regions, including Singapore
(Guest et al. 2005) and several locations within Eastern
Indonesia (Baird et al. 2009), has revealed that multiple
species show synchronous spawning or ‘mass’ coral
spawning. From these studies, it is now believed that
acroporid spawning occurs in Indonesia during the days
following the full moon between either March and April, or
October and November, depending on the species (Baird
et al. 2009). Although we cannot specify exactly when
acroporid spawning occurs in the WNMP (given that
panels were replaced every 4 months), the recruitment
results support a spawning season somewhere between
November and March. This seasonal peak coincided with
the period of warmest sea surface temperature during the
north-west monsoon season, and our results are similar to
those of Fox (2004) in Komodo (Central Indonesia).
In conclusion, we found consistent differences in coral
recruitment rates between two reef systems with different
levels of environmental degradation, which is likely to be
explained by different post-settlement mortality effects,
although we cannot rule out some differences in larval
supply as a partial explanation. Our results demonstrate
how major environmental remediation would be needed for
degraded reefs like Sampela to recover.
Acknowledgments P. Salinas-de-Leon was supported by a Victoria
University PhD Scholarship and wishes to thank the Salinas de Leon
foundation, the Centre for Marine Environmental and Economic
Research, the Centre for Biodiversity and Restoration, the New
Zealand Postgraduate Study Abroad scheme and Operation Wallacea
through collaboration with the Wakatobi Taman National for funding
this study. P. Salinas-de-Leon would also like to thank S. Rowley,
A. Costales-Carrera, Jufri, Magliani, Ludi, Ilu, Arif and other
volunteers at the Hoga Island research centre. Also, thanks to the
Wallacea Foundation and the Indonesian Institute of Sciences (LIPI
research permit granted to DJ Smith). J.J. Bell is also grateful to the
PADI Foundation for providing funding for travel.
References
Amar KO, Chadwick NE, Rinkevich B (2007) Coral planulae as
dispersion vehicles: biological properties of larvae released early
and late in the season. Mar Ecol Prog Ser 350:71–78
Anderson MJ (2001) Permutation tests for univariate or multivariate
analysis of variance and regression. Can J Fish Aquat Sci 58:626–639
Babcock R, Davies P (1991) Effects of sedimentation on settlement of
Acropora millepora. Coral Reefs 9:205–208
Babcock R, Mundy C (1996) Coral recruitment: consequences of
settlement choice for early growth and survivorship in two
scleractinians. J Exp Mar Biol Ecol 206:179–201
Babcock RC, Baird AH, Piromvaragorn S, Thomson DP, Willis BL
(2003) Identification of scleractinian coral recruits from Indo-
Pacific reefs. Zool Stud 42:211–226
Baird AH, Hughes TP (2000) Competitive dominance by tabular
corals: an experimental analysis of recruitment and survival of
understorey assemblages. J Exp Mar Biol Ecol 251(1):117–132
Baird AH, Salih A, Trevor–Jones A (2006) Fluorescence census
techniques for the early detection of coral recruits. Coral Reefs
25:73–76
Baird AH, Guest JR, Willis BL (2009) Systematic and biogeograph-
ical patterns in the reproductive biology of scleractinian corals.
Annu Rev Ecol Evol Syst 40:551–571
Bell JJ, Smith DJ (2004) Ecology of sponge assemblages (Porifera) in
the Wakatobi region, south-east Sulawesi, Indonesia: richness
and abundance. J Mar Biol Assoc UK 84:581–591
Bellwood DR, Hughes TP, Folke C, Nystrom M (2004) Confronting
the coral reef crisis. Nature 429:827–833
Benayahu Y, Loya Y (1985) Settlement and recruitment of a soft
coral: why is Xenia macrospiculata a successful colonizer? Bull
Mar Sci 36:177–188
Bruno JF, Selig ER (2007) Regional decline of coral cover in the
Indo-Pacific: timing, extent, and subregional comparisons. PLoS
ONE 2(8):e711. doi:10.1371/journal.pone.0000711
Burke L, Selig E, Spalding M (2002) Reefs at risk in south-east Asia.
World Resources Institute, Washington, DC
Connell JH, Hughes TP, Wallace CC (1997) A 30-year study of coral
abundance, recruitment, and disturbance at several scales in
space and time. Ecol Monogr 67:461–488
Crabbe MJC, Smith DJ (2005) Sediment impacts on growth rates of
Acropora and Porites corals from fringing reefs of Sulawesi,
Indonesia. Coral Reefs 24(3):337–441
Dunstan PK, Johnson CR (1998) Spatio-temporal variation in coral
recruitment at different scales on Heron Reef, southern Great
Barrier Reef. Coral Reefs 17:71–81
Edinger EN, Jompa J, Limmon GV, Widjatmoko W, Risk MJ (1998)
Reef degradation and coral biodiversity in Indonesia: effects of
land-based pollution, destructive fishing practices and changes
over time. Mar Pollut Bull 36:617–630
Edmunds PJ (2000) Patterns in the distribution of juvenile corals and
coral reef community structure in St. John, US Virgin Islands.
Mar Ecol Prog Ser 202:113–124
English S, Wilkinson C, Baker V (1997) Survey manual for tropical
marine resources. Australian Institute of Marine Science,
Townsville
Fabricius KE (2005) Effects of terrestrial runoff on the ecology of
corals and coral reefs: review and synthesis. Mar Pollut Bull
50:125–146
Fox HE (2004) Coral recruitment in blasted and unblasted sites in
Indonesia: assessing rehabilitation potential. Mar Ecol Prog Ser
269:131–139
Fox HE, Pet JS, Dahuri R, Caldwell RL (2003) Recovery in rubble
fields: long-term impacts of blast fishing. Mar Pollut Bull
46:1024–1031
Mar Biol
123
Author's personal copy

Glassom D, Zakai D, Chadwick-Furman NE (2004) Coral recruit-
ment: a spatio-temporal analysis along the coastline of Eilat,
northern Red Sea. Mar Biol 144:641–651
Guest JR, Baird AH, Goh BPL, Chou LM (2005) Reproductive
seasonality in an equatorial assemblage of scleractinian corals.
Coral Reefs 24:112–116
Harrington L, Fabricius K, De’Ath G, Negri A (2004) Recognition
and selection of settlement substrata determine post-settlement
survival in corals. Ecology 85:3428–3437
Harriott VJ, Banks SA (1995) Recruitment of scleractinian corals in
the Solitary Islands Marine Reserve, a high latitude coral-
dominated community in Eastern Australia. Mar Ecol Prog Ser
123:155–161
Harriott VJ, Fisk DA (1987) A comparison of settlement plate types
for experiments on the recruitment of scleractinian corals. Mar
Ecol Prog Ser 37:201–208
Harrison P, Wallace C (1990) Reproduction, dispersal, and recruit-
ment of scleractinian corals. In: Dubinsky Z (ed) Ecosystems of
the world: coral reefs. Elsevier, Amsterdam, pp 133–207
Harrison PL, Babcock RC, Bull GD, Oliver JK, Wallace CC, Willis
BL (1984) Mass spawning in tropical reef corals. Science
223:1186–1189
Hughes TP (1994) Catastrophes, phase shifts, and large-scale
degradation of a Caribbean coral reef. Science 265:1547–1551
Hughes TP, Baird AH, Dinsdale EA, Moltschaniwskyj NA, Pratchett
MS, Tanner JE, Willis BL (2000) Supply-side ecology works
both ways: the link between benthic adults, fecundity, and larval
recruits. Ecology 81:2241–2249
Hughes TP, Graham T, Jackson JBC, Mumby PJ, Steneck RS (2010)
Rising to the challenge of sustaining coral reef resilience. Trends
Ecol Evol 25:633–642
Hunte W, Wittenberg M (1992) Effects of eutrophication and
sedimentation on juvenile corals. II Settlement. Mar Biol
114:625–631
Maida M, Sammarco PW, Coll JC (1995) Effects of soft corals on
scleractinian coral recruitment.1. Directional allelopathy and
inhibition of settlement. Mar Ecol Prog Ser 121:191–202
Mangubhai S, Harrison PL, Obura DO (2007) Patterns of coral larval
settlement on lagoon reefs in the Mombasa Marine National Park
and Reserve, Kenya. Mar Ecol Prog Ser 348:149–159
Mora C (2008) A clear human footprint in the coral reefs of the
Caribbean. Proc R Soc Biol Sci Ser B 275:767–773
Mundy CN (2000) An appraisal of methods used in coral recruitment
studies. Coral Reefs 19:124–131
Mundy CN, Babcock RC (1998) Role of light intensity and spectral
quality in coral settlement: implications for depth-dependent
settlement? J Exp Mar Biol Ecol 223:235–255
Nozawa Y, Harrison PL (2007) Effects of elevated temperature on
larval settlement and post-settlement survival in scleractinian
corals, Acropora solitaryensis and Favites chinensis. Mar Biol
152:1181–1185
Pineda J (2000) Linking larval settlement to larval transport:
assumptions, potentials and pitfalls. Oceanogr East Pac I:84–105
Piniak GA, Fogarty ND, Addison CM, Kenworthy WJ (2005)
Fluorescence census techniques for coral recruits. Coral Reefs
24:496–500
Potts DC, Done TJ, Isdale PJ, Fisk DA (1985) Dominance of a coral
community by the genus Porites (Scleractinia). Mar Ecol Prog
Ser 23:79–84
Reyes MZ, Yap HT (2001) Effect of artificial substratum material and
resident adults on coral settlement patterns at Danjugan Island,
Philippines. Bull Mar Sci 69:559–566
Richmond RH, Hunter CL (1990) Reproduction and recruitment of
corals: comparisons among the Caribbean, the Tropical Pacific,
and the Red Sea. Mar Ecol Prog Ser 60:185–203
Roberts CM (1997) Connectivity and management of Caribbean coral
reefs. Science 278:1454–1457
Salinas-de-Leon P, Costales-Carrera A, Zeljkovic S, Smith DJ, Bell JJ
(2011) Scleractinian settlement patterns to natural cleared reef
substrata and artificial settlement panels on an Indonesian coral
reef. Estuar Coast Shelf Sci 93:80–85
Sammarco PW (1980) Diadema and its relationship to coral spat
mortality: grazing, competition, and biological disturbance.
J Exp Mar Biol Ecol 45:245–272
Sammarco PW, Andrews JC (1988) Localized dispersal and recruit-
ment in Great Barrier Reef corals: the Helix Experiment. Science
239:1422–1424
Schmidt-Roach S, Kunzmann A, Arbizu PM (2008) In situ observa-
tion of coral recruitment using fluorescence census techniques.
J Exp Mar Biol Ecol 367:37–40
Smith SR (1992) Patterns of coral recruitment and postsettlement
mortality on Bermudas reefs—comparisons to Caribbean and
pacific reefs. Am Zool 32:663–673
Tomascik T (1991) Settlement patterns of Caribbean scleractinian
corals on artificial substrata along a eutrophication gradient,
Barbados, West Indies. Mar Ecol Prog Ser 77:261–269
Tomascik T, Sander F (1985) Effects of eutrophication on reef
building corals. I. Growth rate of the reef building coral
Montastrea annularis. Mar Biol 87:143–155
Tomascik T, Sander F (1987a) Effects of eutrophication on reef
building corals. II. Structure of scleractinian coral communities
on fringing reefs, Barbados, West Indies. Mar Biol 94:53–75
Tomascik T, Sander F (1987b) Effects of eutrophication on reef
building corals. III. Reproduction of the reef building coral
Porites porites. Mar Biol 94:77–94
Tomascik T, vanWoesik R, Mah AJ (1996) Rapid coral colonization
of a recent lava flow following a volcanic eruption, Banda
Islands, Indonesia. Coral Reefs 15:169–175
Tomascik T, Mah AM, Nontji A, Moosa MK (1997) The ecology of
Indonesian seas. Oxford University Press, Singapore
Turak E (2003) Coral diversity and distribution in the Wakatobi
National Marine Park. In: Pet-Soede L, Erdmann MV (eds)
Rapid ecological assessment Wakatobi National Park, WWF and
TNC Joint Publication, Indonesia
Underwood JN, Smith LD, Van Oppen MJH, Gilmour JP (2007)
Multiple scales of genetic connectivity in a brooding coral on
isolated reefs following catastrophic bleaching. Mol Ecol
16:771–784
Underwood JN, Smith LD, van Oppen MJH, Gilmour JP (2009)
Ecologically relevant dispersal of corals on isolated reefs:
implications for managing resilience. Ecol Appl 19:18–29
Mar Biol
123
Author's personal copy





























