Embed Size (px)
Citation preview

Forest Ecology and Management 262 (2011) 730–738
Contents lists available at ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/ locate/ foreco
Fire-vegetation interplay in a mosaic structure of Quercus suber woodlandsand Mediterranean maquis under recurrent fires
A. Schaffhauser a,b,⇑, T. Curt a, T. Tatoni b
a Cemagref UR EMAX Ecosystèmes méditerranéens et risques, 3275 Route de Cézanne CS 40061, 13182 Aix-en-Provence Cedex 5, Franceb Institut Méditerranéen d’Ecologie et de Paléoécologie, Aix-Marseille Université, IMEP (UMR 6116 CNRS/IRD), Université Paul Cézanne – Aix-Marseille III,Faculté des Sciences et Techniques de St Jérôme, Avenue Escadrille Normandie – Niémen – Case 462, 13397 MARSEILLE Cedex 20, France
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 February 2011Received in revised form 28 April 2011Accepted 2 May 2011Available online 28 May 2011
Keywords:Fire recurrenceStructural diversityAcidic soilsMaquisCork oak woodlands (Quercus suber L.)
0378-1127/$ - see front matter � 2011 Elsevier B.V. Adoi:10.1016/j.foreco.2011.05.003
⇑ Corresponding author at: Cemagref UR EMAX Ecrisques, 3275 Route de Cézanne CS 40061, 13182 Aix-
E-mail address: [email protected] (A. Sch
We examined the effects of fire recurrence on a mosaic structure of Quercus suber and Erica-Cistus shrub-land communities of Southeastern France in order to improve the understanding of fire-vegetation inter-play. Plant communities that were similar in 1959 (woodlands on shrublands called maquis on acidicsoils) were compared along a gradient of fire recurrence, from 0 to 4 fires, with different time intervalsbetween fires. The results showed that understory cover increased roughly with fire recurrence, whereastree height, cover, density, stand basal area and litter depth decreased. Different fire recurrences alongthe past decades led to different vegetation types. High fire recurrence corresponded to maquis andsparse cork-oak woodlands while pure oak woodlands established in the absence of fire during the sameperiod. In all, the diversity of tree diameter decreased with fire recurrence. High fire recurrence (3 or 4fires in 50 years) led to a simple vertical structure of vegetation with a mono-layered shrub cover andfew Quercus suber trees. In contrast, spatial connections between plants were maximal at longer timeintervals, leading to a multi-layered vegetation. We finally discuss the potential implications of past fireson the behavior of future fires in the perspective of a sustainable management of these Mediterraneanecosystems.
� 2011 Elsevier B.V. All rights reserved.
1. Introduction
Fire is a key disturbance in Mediterranean areas, which affectsconsiderably the composition and dynamics of ecosystems (Naveh,1994; Pausas et al., 1999). In a context of climatic and globalchanges (Mouillot et al., 2002; Westerling and Bryant, 2008), theresponse of ecosystems to different fire regimes is of importance.Actually, fire regime affects vegetation composition, diversity andstructure, which, in turn, affect fire behavior (Malamud andTurcotte, 1999). This dynamic interaction (i.e. interplay) relies onthe trait-based selection of plants by fires, the development ofadaptive traits (Pausas et al., 1999), and thus the structuration ofvegetation (Trabaud, 1984).
Fire regime is defined as the nature, pattern (i.e. size), season,intensity (energy released) and recurrence of the fires that prevailin an area (Gill, 1979). Among these components, those relating tofire recurrence play an important part, including the frequency (i.e.number of fires within a time span) and the mean fire return inter-val (i.e. the interval of time between two successive fires). In fire-prone Mediterranean areas, some plant and forest communities are
ll rights reserved.
osystèmes méditerranéens eten-Provence Cedex 5, France.affhauser).
subjected to high fire recurrence with a mean fire return interval ofless than 15–20 years (Eugenio Gosalbo and Lloret, 2004; Baezaet al., 2006), which may not allow the survival of many plants be-cause it is lower than the minimal time to recruit from seeds (Pausaset al., 1999; Curt et al., 2009). Fire recurrence is hypothesized toaffect plant communities through individual mortality, selectionand replacement (Gill and Groves, 1981; Trabaud, 1991; Lloret et al.,2003). Fire kills or weakens some species, while it stimulates theseed production or emergence of pyrophyte seeders (Kunholtz-Lordat,1938) and it favors the resprouting species such as Quercus suber(Pausas, 1997; Keeley et al., 2005). This selection by successivefires affects the structure of vegetation, especially its architecture(Barbero et al., 1987; Papio and Trabaud, 1991), phytomass(Trabaud, 1991), and its cover or density (Tavsanoglu and Gürkan,2005). Changes in vegetation structure after fire often occurthrough the shift of dominant species, such as in pine-oakwoodlands (Eugenio Gosalbo and Lloret, 2004). Within a samevegetation sequence such as in Ulex shrublands, recurrent fireslead to simpler vegetation structure with the predominance ofthe most adapted species, and to a decrease in vegetation coverand phytomass (Baeza et al., 2006).
As a consequence of vegetation changes with past fires, past firerecurrence would alter the behavior of future fires. Actually, plantsconstitute the wildland fuel that influence both fire ignition and

A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738 731
fire spread (Fernandes et al., 2000; Riccardi et al., 2007). Thebehavior of wildland fires (i.e. rate of spread, fireline intensityand energy released) is hypothesized to vary according to thestructure of fuels, multi- or mono-layered (Fernandes et al.,2000; Hély et al., 2000; Tanskanen et al., 2005). In Mediterraneanecosystems, fuels are often heterogeneous spatially and in termsof composition (Gill and Groves, 1981; Keeley et al., 2005). Firesimulations in Mediterranean heterogeneous fuels (Arca et al.,2007; Duguy and Vallejo, 2008) indicate that fire behavior at standscale depends on fuel composition, phytomass, density (De Luiset al., 2004; Sandberg et al., 2007), amount and structural arrange-ment of dead and live biomass and spatial connectivity betweenfuels (both vertically and horizontally), i.e. contacts or distance be-tween individuals in the same vegetation layer or between differ-ent layers inducing a ‘ladder effect’ in fire spread (Fernandes andRigolot, 2007). As an example, fire would propagate slowly withlow-intensity in dense and mature Quercus suber woodlands dueto a low understory, while it is faster and more intense in sparsestands with a dense understory made of shrubs (Fernandes andRigolot, 2007). Spatial metrics and indices of vertical and horizon-tal connectivity between plants can be of interest to describe veg-etation structure (Chessel et al., 1975; Staudhammer and Le May,2001; Kant, 2002).
On the basis of literature on vegetation composition, structure(Barbero et al., 1987; Malanson and Trabaud, 1987; EugenioGosalbo and Lloret, 2004), regeneration (Götzenberger et al., 2003),diversity (Moya et al., 2008) and dynamics after fire (Baeza et al.,2006; Capitanio and Carcaillet, 2008), we hypothesize that (i) theabundance and dimensions of trees and shrubs would vary inver-sely with fire recurrence; (ii) vertical structure should be simplifiedby high fire recurrence (mono-layered vegetation); and (iii) hori-zontal connections would increase with a longer time betweenfires (particularly in high shrublands). The responses of thesehypotheses will also be discussed in the perspective of fire behav-ior modeling.
2. Materials and methods
2.1. Study area
The study area is located in the Maures massif of south-easternFrance (43�3 N; 6�3 E), which has high ecological and patrimonialvalue (Loisel, 1971; Molinier, 1973). It is made of acidic soils devel-oped on granitic and gneissic bedrock (Gueirard, 1963). The areahas been affected by recurrent wildfires such as the large and in-tense fires of 1990 and 2003 that covered 25,000 and 14,000 ha,respectively. All selected plots were located in the eastern part ofthe massif, on an homogeneous granitic and gneissic basement(Ladier and Ripert, 1996). They are also similar in terms of soils (med-ium richness and 25 cm depth on average) and topography. Massifridges and steep valleys with shallow soils were discarded. The an-nual mean temperatures vary between 11 and 14 �C and precipita-tions between 900 and 1100 mm. The altitude of all study plotsvaries between 50 and 450 m. The main vegetation types are corkoak (Quercus suber L.) woodlands and shrublands called ‘maquis’ onacidic soils, with Erica arborea L., Calicotome spinosa (L.) Link., Arbu-tus unedo L. and Cistus spp. as the dominant species. The oak wood-lands are present in the vegetation dynamics since the earlyHolocene period (Dubar et al., 1995, 2004; Bergaglio et al., 2006).
2.2. Sampling
The fire history over the last 50 years is known thanks to Med-iterranean fire data sets (Prométhée, 2007), satellite images andaerial photographs taken since 1959 (Curt et al., 2008). Wildfires
were intense (i.e. they had high energy release, see Keeley, 2009)because they occurred in summer under harsh drought and strongwind, and they were assumed to have a similar and severe effect onthe ecosystem. This was confirmed by the fact that all plots had asimilar level of post-fire mortality of the fire-resistant and fire-resilient Quercus suber (see Curt et al., 2009). Fire contours havebeen drawn for the fires larger than 1 ha by using aerial photo-graphs and satellite images (SPOT 1–5 since 1986 CNES Isis, Land-sat and Aster), with a mean time interval of 5–10 years accordingto the availability of photographs and images. All fire contourshave been geometrically corrected and integrated into a GIS sys-tem (ArcGis™ 9.1).
The aerial photographs were interpreted in order to select plotsaccording to classes of fire recurrence. Aerial photographs and sa-tellite images were examined to check for the presence of wildfiresin these plots since 1959 (Fig. 1). Unburned areas were also iden-tified within the contour of major wildfires. Finally, the initial stateof vegetation in this area was identified since the 1950s. It corre-sponds to woodlands dominated by Quercus suber L. on maquis.
The sampling design was based on fire recurrence. Plots wereselected according to the number of fires since 1959, and to thetime since the last fire (see Table 1). Two main wildfires were se-lected because they were similar in terms of fire intensity and cov-ered a wide area in the Maures massif: the 1990 and the 2003 firesused as a reference for reconstructed shrubland vegetation afterfire (around 15 years, according to Trabaud, 1994) and recentburned vegetation, respectively. Sampling included the followingclasses of fire recurrence: class 1 (control plots not burned since1959), class 2 (plots burned once in 1990), class 3 (plots burned2–3 times before 1964 and before the 1990 fire), class 4 (plotsburned 2–3 times between 1970 and 1985 and before the 1990fire) and class 5 (plots burned 3–4 times before 1990 and 2003,shortest time elapsed since the last fire).
The five classes are displayed along an increasing gradient offire recurrence. Two extreme classes (1, 5) correspond to unburnedand recently burned plots, respectively. The other classes (2, 3 and4) concern plots all burned in 1990, with previous fires or not, anddifferent intervals between fires. Classes 3 and 4 are used to testthe hypothesis of the time interval between fires that we hypoth-esized to affect resilience (old fires in the 1960s or more recentfires in the 1970s and 1980s, with a total of 2–3 fires).
2.3. Field survey
The stand structure was studied in 2006 on 51 plots (at least 10repetitions per class) of 20 � 20 m. Each tree or shrub was mea-sured individually (height, crown diameter, circumference at1.30 m) and spatially located with an accuracy of some decimetersusing field decameters. Continuous and homogeneous fuel beds(i.e. non-woody shrublands) have been described using a ‘pointcontact method’ along parallel lines every two meters in x and y.At each point, we assessed the height and cover of the main spe-cies. The cover and depth of litter including the flammable parti-cles (e.g. leaves, needles, or twigs) were also measured on asquare grid of 81 points per plot. Three-dimension scenes werebuilt from field data (Fig. 2) using the R statistical computing soft-ware (2.4.1., R Development Core Team, packages akima, ellipseand rgl).
On the basis of our vegetation descriptions, six vegetation typeswere described according to cover and biomass of the differentvegetation layers. This classification is quite similar to Europeanclassifications, such as Prometheus (Riaño et al., 2003; Jappiotet al., 2007) and EUNIS (Davies et al., 2004). These are oak wood-lands (called here DenseWood), cork-oak woodlands on maquis(SparseWood), low maquis not exceeding 1 meter high (Low-Shrub), medium maquis with a mean height of 2 m, a high biomass

Fig. 1. Number of fires since 1959 on the Eastern part of the Maures massif (Var, Southern France). Fire history is based on Mediterranean fire data sets, satellite images andaerial photographs taken since 1959. Unburned areas were also identified within the outline of major wildfires.
Table 1Fire classes based on fire recurrence, from analysis on images and aerial photographsin the study area since 1959.
Firerecurrenceclass
Number of fireswithin the standsince 1959
Time(years)since thelast fire
Date ofthe lastfire
Time (years)since theprevious lastfire
Date of thepreviouslast fire
1 None, control 47 1959at least
Unknown Unknown
2 1 16 1990 31 1959 atleast
3 2 16 1990 26 19644 2–3 16 1990 5 19855 3–4 3 2003 13 1990
732 A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738
(MedShrubHigh) or a low biomass (MedShrubLow), and high ma-quis (HighShrub, 3–5 m) which constitute an intermediate struc-tural stage. The last type was dominated by dense, mature orsenescent Erica arborea, one of the main structuring shrub species.Thus, we observed some vegetation types with one layer (wood-lands, low maquis) and several layers (woodlands on maquis, med-ium and high maquis).
At last, we also estimated environmental (i.e. abiotic) factorssuch as altitude (m), slope (%), exposure (grades), litter type (loose,compact, or woody), soil depth (cm), presence of rocks (blocks>20 cm and stones <20 cm in size, respectively), and the groundcover composition or nature (nonvascular cryptogams, bare soil,
rock outcrop cover). These environmental data were used to checkfor a potential effect on post-fire vegetation recovery.
2.4. Vegetation structure, diversity and patterning
Our vegetation descriptions provided three complementarytypes of variables, including those describing phytomass, standstructure, and vegetation patterning. Phytomass was measuredfor litter and shrubs (estimated with forestry tables for trees).Stand structure comprised tree circumferences and basal area, treeheights, densities of trees and shrubs, and the distribution of spe-cies in each vegetation layer (density, height and cover). The spa-tial patterning of vegetation included measurements of spatialconnectivity among plants, both vertically and horizontally. Simplevertical indices (connectivity called ‘‘C’’, based on a simple calcula-tion between the height of individuals in the vegetation layer (cm),then averaged and classified (classes 1–5) from variation analysisby value histograms) were obtained using the base of tree crownand the height of shrubs or continuous fuel bed (Fig. 2). The Cts in-dex estimates the degree of connectivity between trees and shrubs,while Csb is for isolated shrubs and the continuous fuel bed, and Ctb
is for trees and shrubs within the continuous fuel bed. High valuesindicate high vertical connectivity between plants. Horizontal indi-ces (so-called ‘‘Interdist’’) were based on the computation of thedistance between tree crowns, shrubs, and the continuous fuelbed. The ‘‘Interdist’’ index has been calculated by a Delaunay’s tri-angulation (cm). High values indicate low horizontal connectivity
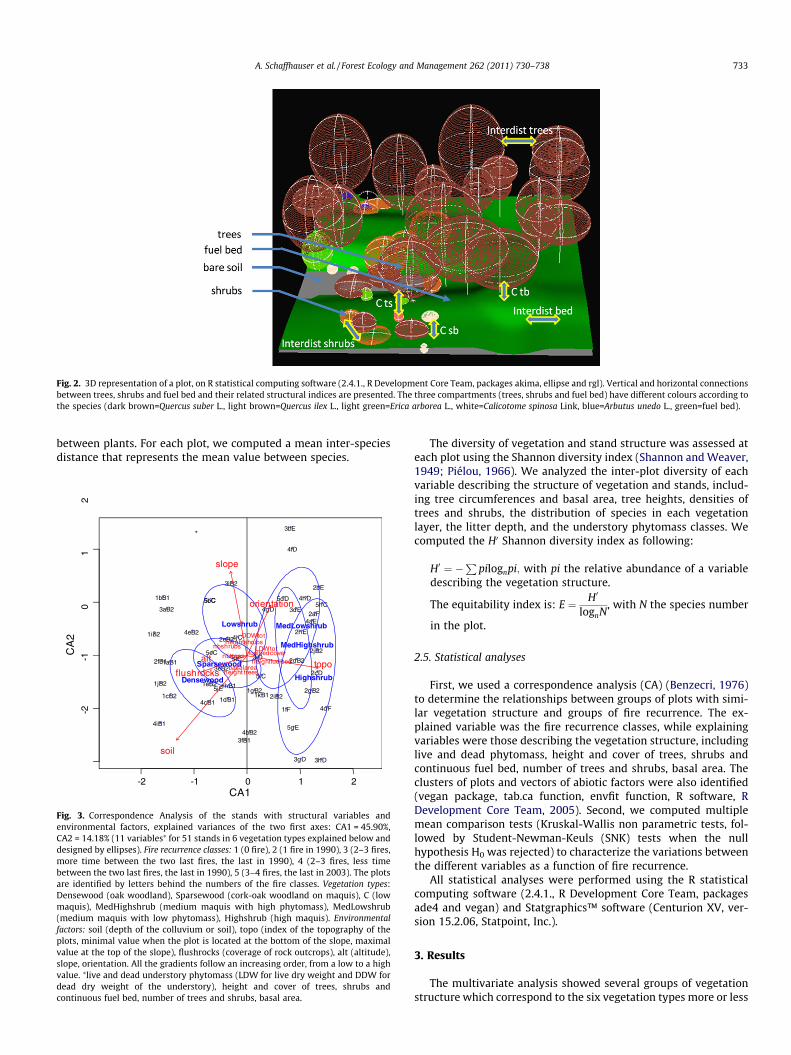
Fig. 2. 3D representation of a plot, on R statistical computing software (2.4.1., R Development Core Team, packages akima, ellipse and rgl). Vertical and horizontal connectionsbetween trees, shrubs and fuel bed and their related structural indices are presented. The three compartments (trees, shrubs and fuel bed) have different colours according tothe species (dark brown=Quercus suber L., light brown=Quercus ilex L., light green=Erica arborea L., white=Calicotome spinosa Link, blue=Arbutus unedo L., green=fuel bed).
A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738 733
between plants. For each plot, we computed a mean inter-speciesdistance that represents the mean value between species.
-2 -1 0 1 2
-2-1
01
2
CA1
CA
2
basalarea
nbtrees
nbshrubs
height trees
heightshrubs
heightfuelbedfuelbedcover
LDWtot
DDWtot
1aB1
1bB1
1cB2 1dB1
1eB2
1fF
1gB21hB1
1iB2
1jB2
1kB1
2aF
2bE
2cD
2dB2
2eB2
2fB1
2gB2
2hE
2iB2
2jB2
3aB2
3bE
3dE
3eB2
3fB1
3gD 3hD
3iB2
3jD
4aE
4bB2
4cB14dF
4eB2
4fD
4gD
4hD
4iB1
4jC
5bC5cC 5dD
5eC5fC
5gE
5hC
5iC
5jEDensewood
Sparsewood
Lowshrub
MedHighshrub
MedLowshrub
Highshrub
orientation
flushrockstopo
slope
soil
alt
Fig. 3. Correspondence Analysis of the stands with structural variables andenvironmental factors, explained variances of the two first axes: CA1 = 45.90%,CA2 = 14.18% (11 variables⁄ for 51 stands in 6 vegetation types explained below anddesigned by ellipses). Fire recurrence classes: 1 (0 fire), 2 (1 fire in 1990), 3 (2–3 fires,more time between the two last fires, the last in 1990), 4 (2–3 fires, less timebetween the two last fires, the last in 1990), 5 (3–4 fires, the last in 2003). The plotsare identified by letters behind the numbers of the fire classes. Vegetation types:Densewood (oak woodland), Sparsewood (cork-oak woodland on maquis), C (lowmaquis), MedHighshrub (medium maquis with high phytomass), MedLowshrub(medium maquis with low phytomass), Highshrub (high maquis). Environmentalfactors: soil (depth of the colluvium or soil), topo (index of the topography of theplots, minimal value when the plot is located at the bottom of the slope, maximalvalue at the top of the slope), flushrocks (coverage of rock outcrops), alt (altitude),slope, orientation. All the gradients follow an increasing order, from a low to a highvalue. ⁄live and dead understory phytomass (LDW for live dry weight and DDW fordead dry weight of the understory), height and cover of trees, shrubs andcontinuous fuel bed, number of trees and shrubs, basal area.
The diversity of vegetation and stand structure was assessed ateach plot using the Shannon diversity index (Shannon and Weaver,1949; Piélou, 1966). We analyzed the inter-plot diversity of eachvariable describing the structure of vegetation and stands, includ-ing tree circumferences and basal area, tree heights, densities oftrees and shrubs, the distribution of species in each vegetationlayer, the litter depth, and the understory phytomass classes. Wecomputed the H0 Shannon diversity index as following:
H0 ¼ �P
pilognpi; with pi the relative abundance of a variabledescribing the vegetation structure.
The equitability index is: E ¼ H0
lognN, with N the species number
in the plot.
2.5. Statistical analyses
First, we used a correspondence analysis (CA) (Benzecri, 1976)to determine the relationships between groups of plots with simi-lar vegetation structure and groups of fire recurrence. The ex-plained variable was the fire recurrence classes, while explainingvariables were those describing the vegetation structure, includinglive and dead phytomass, height and cover of trees, shrubs andcontinuous fuel bed, number of trees and shrubs, basal area. Theclusters of plots and vectors of abiotic factors were also identified(vegan package, tab.ca function, envfit function, R software, RDevelopment Core Team, 2005). Second, we computed multiplemean comparison tests (Kruskal-Wallis non parametric tests, fol-lowed by Student-Newman-Keuls (SNK) tests when the nullhypothesis H0 was rejected) to characterize the variations betweenthe different variables as a function of fire recurrence.
All statistical analyses were performed using the R statisticalcomputing software (2.4.1., R Development Core Team, packagesade4 and vegan) and Statgraphics™ software (Centurion XV, ver-sion 15.2.06, Statpoint, Inc.).
3. Results
The multivariate analysis showed several groups of vegetationstructure which correspond to the six vegetation types more or less

Fig. 4. Structural diversity (Shannon indices) for some structural variables by fire recurrence, means (lines with dots) and standard deviations. The circumferences and basalarea are for trees, whereas the heights and the densities are for trees and shrubs. Fire recurrence classes: 1 (0 fire), 2 (1 fire in 1990), 3 (2–3 fires, more time between the twolast fires, the last in 1990), 4 (2–3 fires, less time between the two last fires, the last in 1990), 5 (3–4 fires, the last in 2003).
734 A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738
(with an overlap for some of them). The two main axes of the cor-respondence analysis explain more than 50% of the variance of dis-tribution (Fig. 3). A major gradient exists among plots according tothe cover of understory shrubs and overstory trees. We observethis gradient from mature and mixed oak woodlands (generallyunburned since 1959) to cork oak woodlands (burned more fre-quently), and from low Cistus maquis (recurrently burned) to highErica maquis (low fire recurrence). Control plots (mixed oak wood-lands) had a maximal diversity (Fig. 4) as concerns tree circumfer-ence, stand basal area and density, and cover of vegetation layers.Conversely, they had a low diversity of tree and shrub height,understory phytomass and litter cover (Fig. 4). In the case of a un-ique fire in 1990 (class 2), understory diversity and tree dimen-sions are highly variable. The plots burned recurrently andrecently which are dominated by low Cistus maquis had a verylow variability of vegetation structure. In total, fire recurrenceaffected strongly the vegetation structure (Table 2) and the livephytomass (Fig. 5), in particular the medium particles (diameter6–25 mm) that contribute to the propagation of fire (data notshown). Understory cover and connectivity between trees and
Table 2Significant variations of structural variables in relation to fire recurrence classes. Kruskal-groups (letters a, b, c, d). ns: not significant. Units: cm (distances), % (cover), g m�2 (phytomthe two last fires, the last in 1990), 4 (2–3 fires, less time between the two last fires, thebetween the trees and the shrubs; Ctb: connectivity indice between the trees and the contbed; Interdist bed: distance between the diameters of individuals in continuous fuel bed.
Connectivity indices
Cts Ctb Csb Interdistbed(cm)
Treeheight(cm)
Treedensity(n. plot)
Fire recurrence classes 1 1.91 a 2.09 2.91 397.25 b 722.67 28.45 b2 1.8 a 3.1 3.8 113.16 a 591 14.2 a3 2.7 ab 3 3.2 191.53 a 578.98 23.4 ab4 2.7 ab 3 3 199.25 a 574.35 25.7 ab5 3.5 b 3.1 3 98.38 a 524.1 18 ab
Kruskal-Wallis test p < 0.05 ns ns 0.01 ns 0.05Khi2 value 9.62 13.02 9.72
shrubs increased, while tree height, cover and density, stand basalarea and litter depth decreased from the control plots to therecently and recurrently burned plots (Table 2). Fuel bed phytomasssignificantly increased from the low Cistus maquis recurrentlyburned to the high Erica maquis. Tree circumference was minimalwith several old fires and after 4 fires (Tables 2 and 3). It is also oneof the more affected variables by fire recurrence variables, withtree cover, live dry understory weight and litter depth (p values<0.001 or 0.01). Among the variables describing fire recurrence,time since last fire impacted vegetation slightly more than thenumber of fires (p values from 0.03 to <0.001), and much morethan the time since previous last fire (only two significant p values,see Table 3).
4. Discussion and conclusions
4.1. Fire recurrence affects vegetation structure
A diagram is proposed to demonstrate the relations betweenthe spatial patterning of vegetation and past fire recurrence, espe-
Wallis tests give the p value and Student-Newman-Keuls test give the homogeneousass). Fire recurrence classes: 1 (0 fire), 2 (1 fire in 1990), 3 (2–3 fires, more time betweenlast in 1990), 5 (3–4 fires, the last in 2003). Structural indices: Cts: connectivity indiceinuous fuel bed; Csb: connectivity indice between the shrubs and the continuous fuel
Dimensions
Treecover(%)
Treecircumference(cm)
Standbasal area(m2 ha�1)
Understorycover (%)
Understorylive dry weight(g m�2)
Litterdepth (cm)
109.12 b 57.85 b 30.03 b 62.18 a 1077.91 a 2.91 d35.65 a 57.76 b 14.16 a 75.88 ab 2451.84 b 1.94 b44.71 a 46.82 a 18.84 a 72.79 ab 1945.69 ab 2.34 c58.07 a 54.39 b 25.36 ab 77.83 ab 1805.11 ab 2.37 c29.36 a 54.11 b 14.77 a 85.57 b 1341.2 ab 1.17 a0.00 0.00 0.03 0.03 0.01 0.0019.07 25.33 11.03 10.40 14.71 307.03
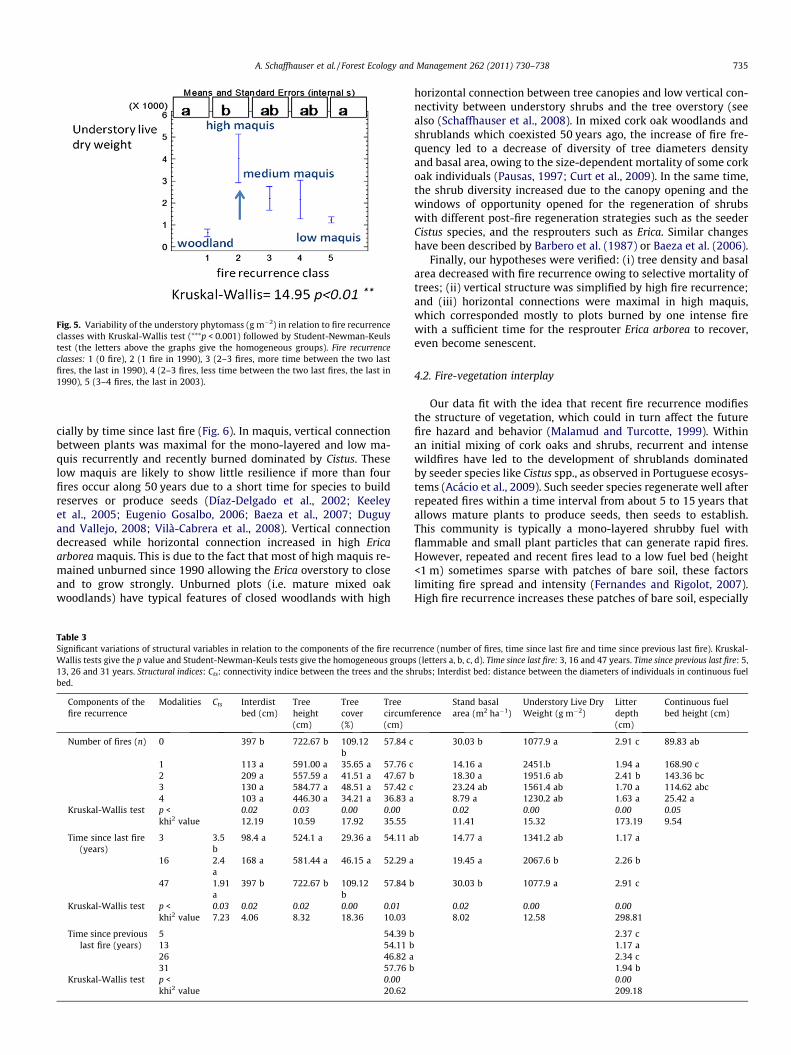
Fig. 5. Variability of the understory phytomass (g m�2) in relation to fire recurrenceclasses with Kruskal-Wallis test (⁄⁄⁄p < 0.001) followed by Student-Newman-Keulstest (the letters above the graphs give the homogeneous groups). Fire recurrenceclasses: 1 (0 fire), 2 (1 fire in 1990), 3 (2–3 fires, more time between the two lastfires, the last in 1990), 4 (2–3 fires, less time between the two last fires, the last in1990), 5 (3–4 fires, the last in 2003).
A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738 735
cially by time since last fire (Fig. 6). In maquis, vertical connectionbetween plants was maximal for the mono-layered and low ma-quis recurrently and recently burned dominated by Cistus. Theselow maquis are likely to show little resilience if more than fourfires occur along 50 years due to a short time for species to buildreserves or produce seeds (Díaz-Delgado et al., 2002; Keeleyet al., 2005; Eugenio Gosalbo, 2006; Baeza et al., 2007; Duguyand Vallejo, 2008; Vilà-Cabrera et al., 2008). Vertical connectiondecreased while horizontal connection increased in high Ericaarborea maquis. This is due to the fact that most of high maquis re-mained unburned since 1990 allowing the Erica overstory to closeand to grow strongly. Unburned plots (i.e. mature mixed oakwoodlands) have typical features of closed woodlands with high
Table 3Significant variations of structural variables in relation to the components of the fire recurWallis tests give the p value and Student-Newman-Keuls tests give the homogeneous group13, 26 and 31 years. Structural indices: Cts: connectivity indice between the trees and the shbed.
Components of thefire recurrence
Modalities Cts Interdistbed (cm)
Treeheight(cm)
Treecover(%)
Treecircum(cm)
Number of fires (n) 0 397 b 722.67 b 109.12b
57.84 c
1 113 a 591.00 a 35.65 a 57.76 c2 209 a 557.59 a 41.51 a 47.67 b3 130 a 584.77 a 48.51 a 57.42 c4 103 a 446.30 a 34.21 a 36.83 a
Kruskal-Wallis test p < 0.02 0.03 0.00 0.00khi2 value 12.19 10.59 17.92 35.55
Time since last fire(years)
3 3.5b
98.4 a 524.1 a 29.36 a 54.11 a
16 2.4a
168 a 581.44 a 46.15 a 52.29 a
47 1.91a
397 b 722.67 b 109.12b
57.84 b
Kruskal-Wallis test p < 0.03 0.02 0.02 0.00 0.01khi2 value 7.23 4.06 8.32 18.36 10.03
Time since previouslast fire (years)
5 54.39 b13 54.11 b26 46.82 a31 57.76 b
Kruskal-Wallis test p < 0.00khi2 value 20.62
horizontal connection between tree canopies and low vertical con-nectivity between understory shrubs and the tree overstory (seealso (Schaffhauser et al., 2008). In mixed cork oak woodlands andshrublands which coexisted 50 years ago, the increase of fire fre-quency led to a decrease of diversity of tree diameters densityand basal area, owing to the size-dependent mortality of some corkoak individuals (Pausas, 1997; Curt et al., 2009). In the same time,the shrub diversity increased due to the canopy opening and thewindows of opportunity opened for the regeneration of shrubswith different post-fire regeneration strategies such as the seederCistus species, and the resprouters such as Erica. Similar changeshave been described by Barbero et al. (1987) or Baeza et al. (2006).
Finally, our hypotheses were verified: (i) tree density and basalarea decreased with fire recurrence owing to selective mortality oftrees; (ii) vertical structure was simplified by high fire recurrence;and (iii) horizontal connections were maximal in high maquis,which corresponded mostly to plots burned by one intense firewith a sufficient time for the resprouter Erica arborea to recover,even become senescent.
4.2. Fire-vegetation interplay
Our data fit with the idea that recent fire recurrence modifiesthe structure of vegetation, which could in turn affect the futurefire hazard and behavior (Malamud and Turcotte, 1999). Withinan initial mixing of cork oaks and shrubs, recurrent and intensewildfires have led to the development of shrublands dominatedby seeder species like Cistus spp., as observed in Portuguese ecosys-tems (Acácio et al., 2009). Such seeder species regenerate well afterrepeated fires within a time interval from about 5 to 15 years thatallows mature plants to produce seeds, then seeds to establish.This community is typically a mono-layered shrubby fuel withflammable and small plant particles that can generate rapid fires.However, repeated and recent fires lead to a low fuel bed (height<1 m) sometimes sparse with patches of bare soil, these factorslimiting fire spread and intensity (Fernandes and Rigolot, 2007).High fire recurrence increases these patches of bare soil, especially
rence (number of fires, time since last fire and time since previous last fire). Kruskal-s (letters a, b, c, d). Time since last fire: 3, 16 and 47 years. Time since previous last fire: 5,rubs; Interdist bed: distance between the diameters of individuals in continuous fuel
ferenceStand basalarea (m2 ha�1)
Understory Live DryWeight (g m�2)
Litterdepth(cm)
Continuous fuelbed height (cm)
30.03 b 1077.9 a 2.91 c 89.83 ab
14.16 a 2451.b 1.94 a 168.90 c18.30 a 1951.6 ab 2.41 b 143.36 bc23.24 ab 1561.4 ab 1.70 a 114.62 abc8.79 a 1230.2 ab 1.63 a 25.42 a0.02 0.00 0.00 0.0511.41 15.32 173.19 9.54
b 14.77 a 1341.2 ab 1.17 a
19.45 a 2067.6 b 2.26 b
30.03 b 1077.9 a 2.91 c
0.02 0.00 0.008.02 12.58 298.81
2.37 c1.17 a2.34 c1.94 b0.00209.18

Fig. 6. Vegetation structure in relation to time since last fire and fire recurrence, with structural factors inducing changes in fire risk.
736 A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738
after soil erosion (De Luis et al., 2001). Recurrent fires with longertime intervals (typically 15–20 years) generally produced mediummaquis with mixing of Cistus and the resprouter Erica arborea, orErica high maquis that generate very high intensity fires. The de-crease of fire recurrence and the longer time since last fire allowsthe development of mixed maquis dominated by Erica, which be-comes dominant and senescent around 16 years after the last fire,likely to 17 for a gorse shrubland, i.e. dominated by Ulex parviflorus(Baeza et al., 2006) increasing the proportion of dead twigs withtime since last fire. This gradient from low to high maquis followsa double gradient of fire recurrence and of time since last fire, asthe post-fire fuel build-up is rapid in shrublands, especiallyfor resprouting species. Erica arborea, as a very efficient respr-outer and a long-lived and flammable species, entails a highfire hazard at landscape-scale because it covers most of the1990 fire outline, and it blocks the regeneration of oaks fromseeds. Mixing of cork oak and shrubs can correspond to variousfire recurrences: fires affect mostly the tree layer through themortality of some individuals, generally the smallest and thelargest ones, and the resprouting of others (Pausas, 1997; Silvaand Catry, 2006). The potential behavior of fire in these multi-layered fuels depends mostly on tree density and height thatchange the wind velocity and allow or not the torching andthe crowning of fire. Low fire recurrence with time intervalslonger than 50 years generally allows the establishment andthe maturation of mixed oak woodlands with Quercus suber,Q. ilex and Q. pubescens. These stands are dense and generallymono-layered with a high canopy and a clear understory becausemost fire-prone shrub species present in the study area areshade-intolerant. They exhibit a high diversity of tree circum-ferences and densities. In these woodlands, typical fires aresurface fires with a low rate of spread and a low intensity.
However, they also exhibit a deep and continuous litter layerthat favors fire ignition.
Among the variables of fire recurrence, time since last fire playsa major role in the structural patterns of vegetation. It especiallyimpacts the phytomass and the connectivity between species,and thus creates a gradient from low maquis to mature mixedwoodlands. Actually, time since last fire controls for a large partthe post-fire fuel build-up, especially for shrubs. Fire interval andnumber of fires along the past decades impact mostly regenerationtime, especially for trees by oaks (Curt et al., 2009). This clearly im-pacts the structure of vegetation and the live phytomass of med-ium particles such as twigs, which affect fire spread andintensity. Long fire intervals and low fire recurrence lead to treemono-layered fuels with an abundant litter production, whereasrecurrent fires at short intervals favor the shrub layer. Intermediatefire recurrence leads to multi-layered fuels with trees and shrubsinterconnected, inducing high fire risk, particularly for high maquis(Schaffhauser, 2009).
In addition to this study, some 3D vegetation scenes were real-ized according to the fire recurrence classes, revealing some impor-tant characteristics on fire-vegetation interplay that will be usefulto predict future fire behavior. Indeed, the final objective will be toimplement fire behavior models such as Behave, semi-empiricalmodel (Andrews et al., 2004), and FireTec, physical model (Linnet al., 2002; Pimont et al., 2006), for a sustainable managementof these Mediterranean ecosystems.
Acknowledgements
This work was financially supported by the ‘‘Conseil Régional ofthe PACA region’’ and the Cemagref through a PhD scholarship, andreceived the collaboration from several other projects (IRISE, MED-

A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738 737
DAD-DNP, FIRE PARADOX). We would like to thank Samuel Allea-ume, Jonathan Baudel, Laurent Borgniet, Simon Brewer, AminataN’Diaye Boubacar, Olivier Chandioux, Roland Estève, Anne Gantea-ume and Willy Martin for field survey. We are grateful to thereviewers for their comments which have been very helpful forrevising the manuscript.
References
Acácio, V., Holmgren, M., Rego, F., Moreira, F., Mohren, G.M.J., 2009. Are drought andwildfires turning Mediterranean cork oak forests into persistent shrublands?Agroforestry Syst. 76, 389–400.
Andrews, P.L., Bevins, C.D., Seli, R.C., 2004. BehavePlus fire modeling system, version3.0: User’s Guide. In. Dept. of Agriculture, Forest Service. Rocky MountainResearch Station, Ogden, UT, p. 132.
Arca, B., Duce, P., Laconi, M., Pellizzaro, G., Salis, M., Spano, D., 2007. Evaluation ofFARSITE simulator in Mediterranean maquis. International Journal of WildlandFire 16, 563–572.
Baeza, M., Raventós, J., Escarré, A., Vallejo, V., 2006. Fire risk and vegetationstructural dynamics in Mediterranean shrubland. Plant Ecol. 187, 189–201.
Baeza, M.J., Valdecantos, A., Alloza, J.A., Vallejo, R., 2007. Human disturbance andenvironmental factors as drivers of long-term post-fire regeneration patterns inMediterranean forests. J. Veg. Sci. 18, 243–252.
Barbero, M., Bonin, G., Loisel, R., Miglioretti, F., Quezel, P., 1987. Impact of forest fireson structure and architecture of Mediterranean ecosystems. EcologiaMediterranea 13, 39–50.
Benzecri, J.P., 1976. L’Analyse des données: L’analyse des Correspondances. 2eédition, Paris, Dunod.
Bergaglio, M., Talon, B., Médail, F., 2006. Histoire et dynamique des forêts de l’ubacdu massif des Maures au cours des derniers 8000 ans. Forêt méditerranéenne t.XXVII, n� 1.
Capitanio, R., Carcaillet, C., 2008. Post-fire Mediterranean vegetation dynamics anddiversity: a discussion of succession models. For. Ecol. Manag. 255, 431–439.
Chessel, D., Debouzie, D., Donadieu, P., Klein, D., 1975. Introduction à l’étude de lastructure horizontale en milieu steppique – échantillonnage systématique pardistance et indice de régularité. Oecologia Plantarum 10, 25–42.
Curt, T., Bertrand, R., Schaffhauser, A., Tatoni, T., 2008. Les communautés végétalesfortement perturbées par les feux deviennent-elles plus inflammables et pluscombustibles ? Un exemple dans les maquis et formations à chêne liège. In:Bonis, A. (Ed.), Actualité de la recherche en écologie des communautésvégétales. ECOVEG4 Congress. Col. TEC & DOC, Lavoisier, pp. 145–150.
Curt, T., Adra, W., Borgniet, L., 2009. Fire-driven oak regeneration in FrenchMediterranean ecosystems. For. Ecol. Manag. 258, 2127–2135.
Davies, C.E., Moss, D., Hill, M., 2004. Eunis habitat classification revised 2004. Reportto the European Environment Agency and the European Topic Centre on NatureProtection and Biodiversity. Accessed on November 12, 2007 at: <http://eunis.eea.europa.eu/>.
De Luis, M., Francisca Garcia-Cano, M., Cortina, J., Raventos, J., Carlos Gonzalez-Hidalgo, J., Rafael Sanchez, J., 2001. Climatic trends, disturbances and short-term vegetation dynamics in a Mediterranean shrubland. For. Ecol. Manag. 147,25–37.
De Luis, M., Baeza, M.J., Raventos, J., Gonzalez-Hidalgo, J.C., 2004. Fuelcharacteristics and fire behaviour in mature Mediterranean gorse shrublands.International Journal of Wildland Fire 13, 79–87.
R Development Core Team, 2005. R: A language and environment for statisticalcomputing, reference index version v. 2.5.1. R Foundation for StatisticalComputing, Vienna, Austria.
Díaz-Delgado, R., Lloret, F., Pons, X., Terradas, J., 2002. Satellite evidence ofdecreasing resilience in Mediterranean plant communities after recurrentwildfires. Ecology 83, 2293–2303.
Dubar, M., Ivaldi, J.P., Thinon, M., 1995. Feux de forêt méditerranéens: une histoirede pins. La recherche 26, 188–189.
Dubar, M., Mai, B.T., Pichard, S., Thinon, M., 2004. Etude palynologique du carottagede Pont d’Argens (Roquebrune-sur-Argens, Var): Histoire holocène de lavégétation en Provence cristalline; facteurs naturels et anthropiques. Ecol.Medit. 30.
Duguy, B., Vallejo, R., 2008. Land-use and fire history effects on post-fire vegetationdynamics in eastern Spain. J. Veg. Sci. 19, 97–108.
Eugenio Gosalbo, M., 2006. Fire recurrence effects on Pinus halepensis Mill.Communities of Catalonia. Ph.D. Thesis. Universitat Autonoma de Barcelona,p. 227.
Eugenio Gosalbo, M., Lloret, F., 2004. Fire recurrence effects on the structure andcomposition of Mediterranean Pinus halepensis communities in Catalonia(northeast Iberian Peninsula). Ecoscience 11 (4), 446–454.
Fernandes, P.M., Rigolot, E., 2007. The fire ecology and management of maritimepine (Pinus pinaster Ait.). For. Ecol. Manag. 241, 1–13.
Fernandes, P.M., Catchpole, W.R., Rego, F.C., 2000. Shrubland fire behaviourmodelling with microplot data. Canadian Journal of Forest Research-RevueCanadienne De Recherche Forestiere 30, 889–899.
Gill, A.M., 1979. Fire and the Australian landscape. Landscape Planning 6, 343–357.Gill, A.M., Groves, R.H., 1981. Fire regimes in heathlands and their plant ecological
effects. In: Specht, R.L. (Ed.), Ecosystems of the World 9B Heathlands andRelated Shrublands Analytical Studies. Elsevier Scientific, New York, pp. 61–84.
Götzenberger, L., Ohl, L., Hensen, I., Sánchez Gómez, P., Wesche, K., 2003. Postfireregeneration of a thermomediterranean shrubland area in south-eastern Spain.Anales de Biología 25, 21–28.
Gueirard, S., 1963. Le Massif des Maures. Harmann, Géologie régionale de la France,Paris.
Hély, C., Bergeron, Y., Flannigan, M.D., 2000. Effects of stand composition on firehazard in mixed-wood Canadian boreal forest. J. Veg. Sci. 11, 813–824.
Jappiot, M., Abdelmoula, K., Allgöwer, B., Borgniet, L., Curt, T., Ghosn, D., Gitas, I.,Kazakis, G., Koetz, B., Lampin, C., Marell, A., Morge, D., Morsdorf, F., Rigolot, E.,Sesbou, A., Vega, J.-A., Vigy, O., 2007. Georeferenced database of representativesites including ecological data. In: Deliverable D.3.4-1 of the Integrated project‘‘Fire Paradox’’, P.n.F.-., European Commission (Ed.), p. 73.
Kant, S., 2002. The marginal cost of structural diversity of mixed uneven-aged standof hard maple forests. Can. J. For. Res. 32, 616–628.
Keeley, J., 2009. Fire intensity, fire severity and burn severity: a brief review andsuggested usage. Int. J. Wildland Fire 18, 116–126.
Keeley, J., Fotheringham, C.J., Baer-Keeley, M., 2005. Determinants of postfirerecovery and succession in Mediterranean-climate shrublands of California.Ecol. Appl. 15, 1515–1534.
Kunholtz-Lordat, G., 1938. La terre incendiée. Maison Carrée, Nîmes.Ladier, J., Ripert, C., 1996. Les stations forestières de la Provence cristalline (Cap
Sicié, îles d’Hyères, Maures, Tanneron), Typologie des stations forestières. In.Rapport du CEMAGREF, Aix-en-Provence, p. 92 + annexes.
Linn, R.R., Reisner, J., Colman, J.J., Winterkamp, J., 2002. Studying wildfire behaviorusing FIRETEC. Int. J. Wildland Fire 11, 233–246.
Lloret, F., Pausas, J.G., Vilà, M., 2003. Responses of Mediterranean plant species todifferent fire frequencies in Garraf Natural Park (Catalonia, Spain): fieldobservations and modelling. Plant Ecol. 223, 235.
Loisel, R., 1971. Séries de végétation propres, en Provence, aux massifs des Maureset de l’Esterel (ripisilves exclues). Bulletin Société Botanique de France 118,203–236.
Malamud, B.D., Turcotte, D.L., 1999. Self-organized criticality applied to naturalhazards. Nat. Hazards 20, 93–116.
Malanson, G.P., Trabaud, L., 1987. Post-fire development of canopy structure in aMediterranean shrub, Quercus coccifera L. Phys. Geogr. 8, 266–274.
Molinier, R., 1973. Les études phytosociologiques en Provence cristalline. Bull. Mus.Hist. nat. Marseille 33, 7–45.
Mouillot, F., Rambal, S., Joffre, R., 2002. Simulating climate change impacts on firefrequency and vegetation dynamics in a Mediterranean-type ecosystem. GlobalChange Biol. 8, 423–437.
Moya, D., De las Heras, J., Lopez-Serrano, F.R., Condes, S., Alberdi, I., 2008. Structuralpatterns and biodiversity in burned and managed Aleppo pine stands. PlantEcol. 200, 217–228.
Naveh, Z., 1994. The role of fire and its management in the conservation ofMediterranean ecosystems and landscapes. In: Moreno, J.M., Oechel, W.C. (Eds.),The role of Fire in Mediterranean-Type Ecosystem, Vol. 107. Berlin, Springer-Verlag, pp. 163–185.
Papio, C., Trabaud, L., 1991. Comparative study of the Aerial Structure of five shrubsof Mediterranean shrublands. For. Sci. 37, 146–159.
Pausas, J.G., 1997. Resprouting of Quercus suber in NE Spain after fire. J. Veg. Sci. 8,703–706.
Pausas, J.G., Carbo, E., Caturla, R.N., Gil, J.M., Vallejo, R., 1999. Post-fire regen-eration patterns in the eastern Iberian Peninsula. Acta Oecologica 20, 499–508.
Piélou, E.C., 1966. Species-diversity and pattern-diversity in the study of ecologicalsuccession. J. Theor. Biol. 10, 370–383.
Pimont, F., Linn, R., Dupuy, J.L., Morvan, D., 2006. Effects of vegetation descriptionparameters on forest fire behavior with FIRETEC. In: Viegas, D.X. (Ed.), VInternational Conference on Forest Fire Research.
Prométhée, 2007. Banque de données sur les incendies de forêts en régionméditerranéenne en France. http://www.promethee.com/.
Riaño, D., Meier, E., Allgower, B., Chuvieco, E., Ustin, S.L., 2003. Modeling airbornelaser scanning data for the spatial generation of critical forest parameters in firebehavior modeling. Remote Sens. Environ. 86, 177–186.
Riccardi, C.L., Ottmar, R.D., Sandberg, D.V., Andreu, A., Elman, E., Kopper, K., Long, J.,2007. The fuelbed: a key element of the Fuel Characteristic ClassificationSystem. Can. J. For. Res. 37, 2394–2412.
Sandberg, D.V., Riccardi, C.L., Schaaf, M.D., 2007. Fire potential rating for wildlandfuelbeds using the fuel characteristic classification system. Can. J. For. Res.-Revue Canadienne De Recherche Forestiere 37, 2456–2463.
Schaffhauser, A., 2009. Impacts de la récurrence des incendies sur la végétation, soninflammabilité et sa combustibilité. Application à la Provence cristalline (massifdes Maures, Var, France). Thèse de doctorat. Aix-Marseille Université, Marseille,France, pp. 224 + annexes, 282 p.
Schaffhauser, A., Curt, T., Tatoni, T., 2008. The resilience ability of vegetation afterdifferent fire recurrences in Provence. FOREST FIRES congress, Toledo, Spain, pp.297–310.
Shannon, C., Weaver, W., 1949. The Mathematical Theory of Communication.University of Illinois Press, Urbana.
Silva, J.S., Catry, F., 2006. Forest fires in cork oak (Quercus suber L.) stands inPortugal. Int. J. Environ. Stud. 63, 3, 235–257(223).
Staudhammer, C.L., Le May, V.M., 2001. Introduction and evaluation of possibleindices of stand structural diversity. Can. J. For. Resour. 31 (7), 1105–1115.
Tanskanen, H., Venäläinen, A., Puttonen, P., Grandström, A., 2005. Impact of standstructure on surface fire ignition potential in Picea abies and Pinus sylvestrisforests in southern Finland. Can. J. For. Res. 35, 410–420.

738 A. Schaffhauser et al. / Forest Ecology and Management 262 (2011) 730–738
Tavsanoglu, C., Gürkan, B., 2005. Post-fire dynamics of Cistus spp. in a Pinus brutiaforest. Turk. J. Bot. 29, 337–343.
Trabaud, L., 1984. Fire adaptation strategies of plants in the French Mediterraneanarea. In: M. Greece, N.S., Arianoustou-Faraggitaki, M., Oechel, W.C. (Eds.),Proceedings International symposium adaptations terrestrial environment. DrW. Junk Publishers, Halkidiki, Greece, pp. 63–69.
Trabaud, L., 1991. Fire regimes and phytomass growth dynamics in a Quercuscoccifera garrigue. J. Veg. Sci. 2, 307–314.
Trabaud, L., 1994. Wildland fire cycles and history in Central Southern France.In: International Conference on Forest Fire Research 2, Coimbra, pp. 545–556.
Vilà-Cabrera, A., Saura-Mas, S., Lloret, F., 2008. Effects of fire frequency onspecies composition in a Mediterranean shrubland. Ecoscience 15, 519–528.
Westerling, A.L., Bryant, B.P., 2008. Climate change and wildfire in California. Clim.Change 87, S231–S249.





























