Embed Size (px)
Citation preview

© 2010 Gebrüder Borntraeger, 70176 Stuttgart, Germany www.borntraeger-cramer.deDOI: 10.1127/0340 - 269X/2010/0040 - 0451 0340-269X/10/0040-0451 $ 5.40
Phytocoenologia, 40 (4), 255–266Stuttgart, December 30, 2010
Factors affecting the fl oristic relationships among Mexican pinyon woodlands
by Mario LUNA-CAVAZOS, Angélica ROMERO-MANZANARES and Edmundo GARCÍA-MOYA, México
with 3 fi gures and 5 tables
Abstract. We analyzed, by indirect and direct gradient analysis, the fl oristic relationships among different Mexi-can pinyon woodland communities dominated by Pinus cembroides, P. johannis, P. remota, P. maximartinezii, P. monophylla, P. quadrifolia, P. nelsonii, P. culminicola, and P. lagunae, belonging to distinct biogeographic provinces. We elaborated a presence-absence data matrix of 1153 taxa and 32 woodlands, and analyzed the pinyon woodland relationships in relation to various environmental factors by using Detrended Correspondence Analysis (DCA) and Canonical Correspondence Analysis (CCA). The fl oristic differences among pinyon communities were tested by using Multiresponse Permutation Procedure (MRPP) analysis. Our results showed that the fi rst DCA axis distin-guished Operational Geographic Units (OGUs) based on a longitudinal and latitudinal patterns, while the second axis is related to mean annual temperature. Based on fl oristic relationships, three main groups were detected by DCA: a) pinyons from the California province; b) pinyons from Sierra Madre Oriental and Altiplano Norte and c) pinyons from Sierra Madre Occidental and Altiplano Sur. P. lagunae, from Del Cabo province, is more similar to this last group. Axis one of CCA correlated with longitude and latitude, axis two with elevation, mean annual precipita-tion, and aridity index. Overall, our data indicate that P. quadrifolia and P. monophylla communities are the pinyon woodlands fl oristically least similar to the other pinyon woodlands.
Keywords: operational geographic unit (OGU), ordination, pine, Pinus, plant community types, plant-soil relations.
Introduction
The pinyon woodlands are mainly found from the southwestern United States to north and central México (Passini 1987, Perry 1991, Farjon & Styles 1997). The pinyon pines are particularly diverse in Mexico (Lanner 1981) where they grow in large ex-panses across the country, distributed between 18° to 32°45’ N and 97° to 117° W (Farjon & Styles 1997). Pinyon woodlands fl ourish in dry climates, and are generally surrounded by grasslands, shrublands, and oak woodlands (Rzedowski 1978, Perry 1991).
Probably the climatic conditions prevailing dur-ing the Holocene have changed the Mexican pinyon pine woodlands seen today. At the start of the Holo-cene, 10 000 years ago, the southwestern U.S.A., and northern Mexico had a humid climate with a mesic forest of pinyon-juniper or Mexican pinyon-oak for-est that migrated to mountains with higher elevations when climate became drier (Van Devender 1977, Van Devender & Spaulding 1979, Van Devender et al. 1987, Axelrod 1981, Miller & Wigand 1994). In this period, the dominant vegetation in the Chihuahuan Desert during the different phases of the Holocene corresponded initially to a juniper forest, thence ocurred a transition to grassland, and fi nally to the establishment of a shrubland. In the Sonoran Desert, Pinus monophylla communities were co-dominant with juniper and oak, which have since moved al-titudinally from 600 to 1500 m a.s.l. These phyto-geographycal processes probably defi ned the plant communities extant. According to Van Devender &
Spaulding(1979), the actual climate and vegetation have been relatively stable since c.a. 8 000 years ago.
The pinyon woodlands are important in terms of economy, ecology, and phytogeography. Pinyons yield seeds for human consumption and marketing, as well as offering fi rewood and timber for people, and forage and shelter for indigenous wildlife (Robert 1973, Romero-Manzanares et al. 1999). Ecologically, the pinyon woodlands contribute to maintaining wa-tersheds and natural habitats for many fl oristic and faunistic species (Caballero & Ávila 1989, Binnema 1996). Various pinyon pine species and companion taxa are endemic to Mexico (Pinkava 1984), and they should be conserved. Previous studies in Mexican pinyon woodlands have included fl oristic descrip-tions (García & González 1991, González et al. 1993) and quantitative analysis of the fl oristic similarities and differences among pinyon woodlands at the lo-cal level (Passini 1982, Hernández-Reyna & García-Moya 1985, Passini 1994, Romero-Manzanares 2001). However, no previous study has fully analyzed the infl uence of environmental factors on the fl oristic re-lationships among different pinyon woodlands dis-tributed throughout the Mexican territory.
The analysis of fl oristic similarity among commu-nities allows researchers to examine the degree of iso-lation between taxa (McCoy & Heck 1987), as well as to identify, classify, and quantitatively analyze fl o-ristic areas and elements (McLaughlin 1994). Floris-tic phytogeography can also help prioritize areas for conservation by assessing the relative importance of different communities (Justus & Sarkar 2002), while

256 M. Luna-Cavazos et al.
quantitative fl oristic analyses can be used to group taxa of similar distributions and to correlate them with climatic variables (Birks 1987). Multivariate analysis is an important tool for community ecology, for the analysis of relationships between different sites based on the structural and fl oristic composi-tions of their communities (e.g. Dzwonko & Loster 2000, De Oliveira & Nelson 2001, Moora et al. 2007).
Here, we use ordination analysis of the fl oristic affi nities among Mexican pinyon woodlands. We believe that this work contributes to an understand-ing of the fl oristic relationship among these pinyon woodlands, and provides new insight into which en-vironmental factors help determine the fl oristic com-position of the Mexican pinyon woodlands.
Materials and methods
Floristic data
This study of fl oristic affi nities among the pinyon woodlands was based on the fl oristic records of vas-cular plants listed in Pinkava (1984), Passini (1985, 1994), Hernández-Reyna (1985), Passini & Pinel (1989), Suzán (1990), García & González (1991), González et al. (1993), Peinado et al. (1994), Villar-
real (1994), Reyes et al. (1996), González (1998) and Romero-Manzanares (2001). The pinyon woodlands included in this study are characterized by species such as Pinus cembroides Zucc., P. johannis M.-F. Robert, P. remota (Little) Bailey & Hawkswork, P. maximartinezii Rzedowski, P. monophylla Torr., P. quadrifolia Sudw., P. nelsonii Shaw., P. culminicola Andresen & Beaman, and P. lagunae M.-F. Passini. The locations and descriptive characteristics of the 32 Mexican pinyon woodlands studied are shown in Fig. 1, and Table 1. These woodlands were chosen to include a representative distribution of pinyon in Mexico, and each pinyon was considered an Opera-tional Geographic Unit (OGU, Crovello 1981) for the purpose of this work. There are pinyon wood-lands distributed farther south (to 18° N), but they were not included in our study due to a lack of fl o-ristic data. Each OGU was related to corresponding biogeographic provinces as described by Comisión Nacional para el Conocimiento y Uso de la Biodiver-sidad (CONABIO 1997).
We recorded a total of 1261 plant species from all the pinyon pine woodlands, which were examined for synonymy in the Integrated Taxonomic Informa-tion System (www.itis.gov) and Sistema Integrado de Información Taxonómica-México (www.siit.conabio.gob.mx); a total of 108 taxa were synonymous with
Fig. 1. Geographic distribution of the pinyon woodlands. Additional details appear in Table 1.

Floristic relationships among Mexican pinyon woodlands 257
other species, so the fi nal data matrix included 32 OGU-by-1153 taxa. These data were used to obtain a presence-absence data matrix, upon which indirect and direct gradient analysis of the fl oristic affi nities among 32 OGUs were performed.
Climatic and soil data
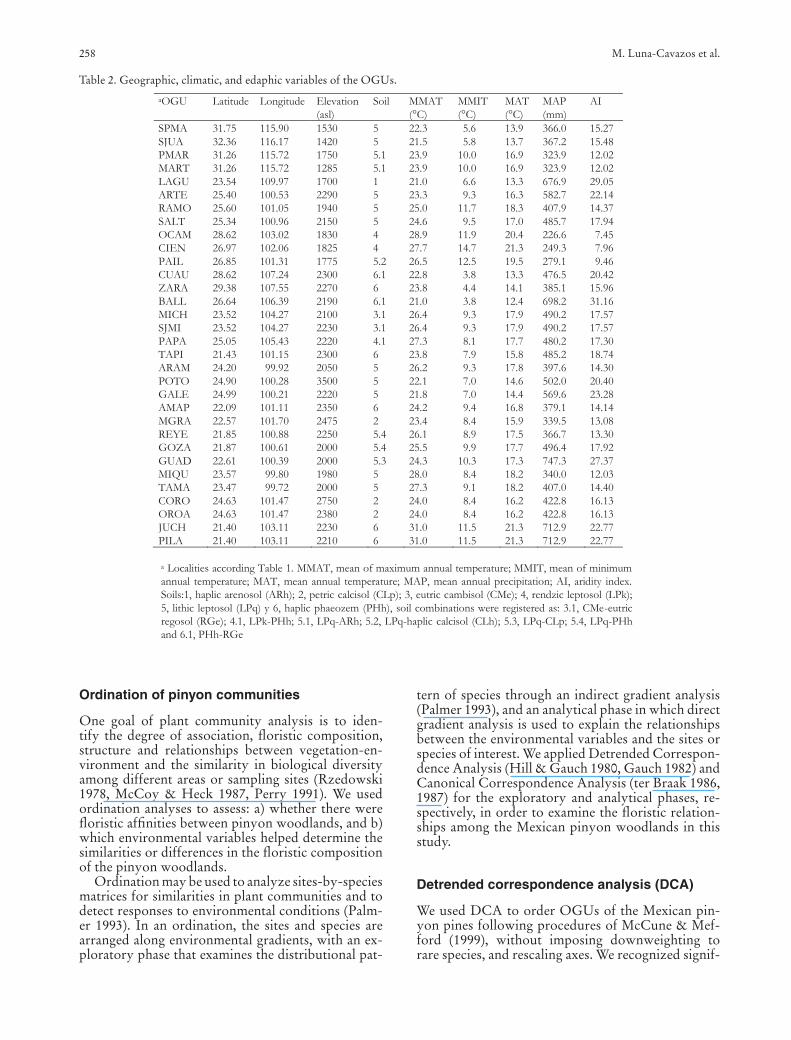
The climatic variables of this study (Table 2) were derived from the records of the National Meteoro-logical Service (www.smn.cna.gob.mx). We recorded climatological data from one or more meteorologi-cal stations near or within each study site; data of two or more stations for one OGU were averaged. Climatic data were based on records of 10 – 25 years. The climatic variables included: elevation (asl), mean maximum annual temperature (°C), mean minimum annual temperature (°C), mean annual temperature (°C), mean annual precipitation (mm), and an aridity index according to Martonne (1926). The geographic
data included longitude and latitude in decimal de-grees for the center of the area for each pinyon pine community. The soil data were gathered from the soil maps of Mexico (FAO-UNESCO-ISRIC 1988). We recorded the soil type of the each OGU and the main soil unit the type was a part of the 13 soil types fell into six major soil units and seven combinations of them: 1, haplic arenosol (ARh); 2, petric calcisol (CLp); 3, eutric cambisol (CMe); 4, rendzic leptosol (LPk); 5, lithic leptosol (LPq) y 6, haplic phaeozem (PHh), while the soil combinations were registered as: 3.1, CMe-eutric regosol (RGe); 4.1, LPk-PHh; 5.1, LPq-ARh; 5.2, LPq-haplic calcisol (CLh); 5.3, LPq-CLp; 5.4, LPq-PHh and 6.1, PHh-RGe (Table 2). Most OGUs had a principal soil unit, but for some OGUs there was an important contribution from an-other soil type that justifi ed a combination of units.
1. SPMA: Sierra San Pedro Mártir, Baja California: P. quadrifolia Sudw. - P. monophylla Torr.; 31°08’-31°59’ N, 115°30’-115°33’ W.
2. SJUA: Sierra de Juárez, Baja California P. quadrifolia; 32°04’-32°33’ N, 115°51’-116°07’ W. 3. PMAR: Sierra San Pedro Mártir, Baja California: P. quadrifolia; 30°15’-32°50’ N, 115°00’-116°05’ W. 4. MART: Sierra San Pedro Mártir, Baja California: P. monophylla; 30°15’-32°50’ N, 115°00’-116°05’ W. 5. LAGU: Sierra de la Laguna, Baja California Sur: P. lagunae M.-F. Passini; 23°29-23°35’ N, 109°53’-
110°01’ W. 6. ARTE: Arteaga, Coahuila: P. cembroides Zucc.; 25°11’-25°27’ N, 100°23’-100°46’ W. 7. RAMO: Ramos Arizpe, Coahuila: P. cembroides; 25°30’-25°56’ N, 100°40’-101°34’W. 8. SALT: Saltillo, Coahuila: P. cembroides; 24°54’-25°23’ N, 100°23’-101°30’ W. 9. OCAM: Villa de Ocampo, Coahuila: P. cembroides; 28°07’-29°02’ N, 101°28’-103°04’ W. 10. CIEN: Cuatro Ciénegas, Coahuila: P. johannis M.-F. Robert - P. remota (Little) Bailey & Hawksworth;
26°59’ N, 102°02 W. 11. PAIL : Sierra de La Paila, Coahuila: P. cembroides; 25°45’-26°16’ N, 101°25’-101°49’ W. 12. CUAU : Cuauhtémoc, Chihuahua: P. cembroides; 28°23’-29°09’ N, 106°47’-107°36’ W. 13. ZARA: Ignacio Zaragoza, Chihuahua: P. cembroides; 29°35’-29°45’ N, 107°30’-107°57’ W. 14. BALL: San Pablo Balleza, Chihuahua: P. cembroides; 26°47’-26°57’ N, 106°10’-106°47’ W. 15. MICH : Reserva de la Michilía, Durango: P. cembroides; 23°15’-23°35’ N, 104°00’-104°20’ W. 16. SJMI : San Juan de Michis, Durango: P. cembroides; 23°23’-23°35’ N, 104°01’-104°10’ W. 17. PAPA: Santiago Papasquiaro, Durango: P. cembroides; 25°02’-25°08’ N, 105°18’-105°38’ W. 18. TAPI: El Cubo, San Felipe, Guanajuato: P. cembroides: 21°26’-21°35’ N, 100°57’-101°12’ W. 19. ARAM: Aramberri, Nuevo León: P. cembroides; 23°58’-24°28’ N, 99°53’-100°03’ W. 20. POTO: El Potosí, Galeana Nuevo León: P. culminicola Andresen & Beaman; 24°50’-24°53’ N, 100°13’-
100°15’ W. 21. GALE: Galeana, Nuevo León: P. cembroides; 24°31’-25°16’ N, 99°59’-101°35’ W. 22. AMAP: La Amapola, San Luis Potosí, San Luis Potosí: P. cembroides - P. johannis: 22°00’-22°04’ N,
101°04’-101°09’ W. 23. MGRA: Sierra Monte Grande, Charcas, San Luis Potosí: P. johannis; 23°17’-23°21 N, 101°07’-101°11’
W. 24. REYE: Villa de Reyes, San Luis Potosí: P. cembroides; 21°48’-22°02 N, 100°03’-101°48’ W. 25. GOZA: Villa de Zaragoza, San Luis Potosí: P. cembroides; 21°55’-22°00’ N, 100°30’-100°38’ W. 26. GUAD: Guadalcazar, San Luis Potosí: P. nelsonii; 22°34’-22°36’ N, 100°25’-100°27’ W. 27. MIQU: Miquihuana, Tamaulipas: P. nelsonii - P. cembroides; 23°30’-23°37’ N, 99°37’-99°46’ W. 28. TAMA: Miquihuana, Tamaulipas: P. nelsonii; 23°16’-23°40’ N, 99°40’-99°53’ W. 29. CORO: Concepción del Oro, Zacatecas: P. cembroides - P. johannis; 24°17’-24°37’ N, 101°06’-101°28’ W. 30. OROA: Concepción del Oro, Zacatecas: P. johannis; 24°37’ N, 101°25’ W. 31. JUCH: Juchipila, Zacatecas: P. maximartinezii; 21°20’-21°21’ N, 103°13’-103°14’ W. 32. PILA: Juchipila, Zacatecas: P. maximartinezii; 21°21’-21°22’ N, 103°13’-103°14’ W.
Table 1. Description of the Mexican pinyon woodlands studied.
258 M. Luna-Cavazos et al.
Ordination of pinyon communities
One goal of plant community analysis is to iden-tify the degree of association, fl oristic composition, structure and relationships between vegetation-en-vironment and the similarity in biological diversity among different areas or sampling sites (Rzedowski 1978, McCoy & Heck 1987, Perry 1991). We used ordination analyses to assess: a) whether there were fl oristic affi nities between pinyon woodlands, and b) which environmental variables helped determine the similarities or differences in the fl oristic composition of the pinyon woodlands.
Ordination may be used to analyze sites-by-species matrices for similarities in plant communities and to detect responses to environmental conditions (Palm-er 1993). In an ordination, the sites and species are arranged along environmental gradients, with an ex-ploratory phase that examines the distributional pat-
tern of species through an indirect gradient analysis (Palmer 1993), and an analytical phase in which direct gradient analysis is used to explain the relationships between the environmental variables and the sites or species of interest. We applied Detrended Correspon-dence Analysis (Hill & Gauch 1980, Gauch 1982) and Canonical Correspondence Analysis (ter Braak 1986, 1987) for the exploratory and analytical phases, re-spectively, in order to examine the fl oristic relation-ships among the Mexican pinyon woodlands in this study.
Detrended correspondence analysis (DCA)
We used DCA to order OGUs of the Mexican pin-yon pines following procedures of McCune & Mef-ford (1999), without imposing downweighting to rare species, and rescaling axes. We recognized signif-
aOGU Latitude Longitude Elevation (asl)
Soil MMAT (°C)
MMIT (°C)
MAT (°C)
MAP (mm)
AI
SPMA 31.75 115.90 1530 5 22.3 5.6 13.9 366.0 15.27 SJUA 32.36 116.17 1420 5 21.5 5.8 13.7 367.2 15.48 PMAR 31.26 115.72 1750 5.1 23.9 10.0 16.9 323.9 12.02 MART 31.26 115.72 1285 5.1 23.9 10.0 16.9 323.9 12.02 LAGU 23.54 109.97 1700 1 21.0 6.6 13.3 676.9 29.05 ARTE 25.40 100.53 2290 5 23.3 9.3 16.3 582.7 22.14 RAMO 25.60 101.05 1940 5 25.0 11.7 18.3 407.9 14.37 SALT 25.34 100.96 2150 5 24.6 9.5 17.0 485.7 17.94 OCAM 28.62 103.02 1830 4 28.9 11.9 20.4 226.6 7.45 CIEN 26.97 102.06 1825 4 27.7 14.7 21.3 249.3 7.96 PAIL 26.85 101.31 1775 5.2 26.5 12.5 19.5 279.1 9.46 CUAU 28.62 107.24 2300 6.1 22.8 3.8 13.3 476.5 20.42 ZARA 29.38 107.55 2270 6 23.8 4.4 14.1 385.1 15.96 BALL 26.64 106.39 2190 6.1 21.0 3.8 12.4 698.2 31.16 MICH 23.52 104.27 2100 3.1 26.4 9.3 17.9 490.2 17.57 SJMI 23.52 104.27 2230 3.1 26.4 9.3 17.9 490.2 17.57 PAPA 25.05 105.43 2220 4.1 27.3 8.1 17.7 480.2 17.30 TAPI 21.43 101.15 2300 6 23.8 7.9 15.8 485.2 18.74 ARAM 24.20 99.92 2050 5 26.2 9.3 17.8 397.6 14.30 POTO 24.90 100.28 3500 5 22.1 7.0 14.6 502.0 20.40 GALE 24.99 100.21 2220 5 21.8 7.0 14.4 569.6 23.28 AMAP 22.09 101.11 2350 6 24.2 9.4 16.8 379.1 14.14 MGRA 22.57 101.70 2475 2 23.4 8.4 15.9 339.5 13.08 REYE 21.85 100.88 2250 5.4 26.1 8.9 17.5 366.7 13.30 GOZA 21.87 100.61 2000 5.4 25.5 9.9 17.7 496.4 17.92 GUAD 22.61 100.39 2000 5.3 24.3 10.3 17.3 747.3 27.37 MIQU 23.57 99.80 1980 5 28.0 8.4 18.2 340.0 12.03 TAMA 23.47 99.72 2000 5 27.3 9.1 18.2 407.0 14.40 CORO 24.63 101.47 2750 2 24.0 8.4 16.2 422.8 16.13 OROA 24.63 101.47 2380 2 24.0 8.4 16.2 422.8 16.13 JUCH 21.40 103.11 2230 6 31.0 11.5 21.3 712.9 22.77 PILA 21.40 103.11 2210 6 31.0 11.5 21.3 712.9 22.77
a Localities according Table 1. MMAT, mean of maximum annual temperature; MMIT, mean of minimum annual temperature; MAT, mean annual temperature; MAP, mean annual precipitation; AI, aridity index. Soils:1, haplic arenosol (ARh); 2, petric calcisol (CLp); 3, eutric cambisol (CMe); 4, rendzic leptosol (LPk); 5, lithic leptosol (LPq) y 6, haplic phaeozem (PHh), soil combinations were registered as: 3.1, CMe-eutric regosol (RGe); 4.1, LPk-PHh; 5.1, LPq-ARh; 5.2, LPq-haplic calcisol (CLh); 5.3, LPq-CLp; 5.4, LPq-PHh and 6.1, PHh-RGe
Table 2. Geographic, climatic, and edaphic variables of the OGUs.

Floristic relationships among Mexican pinyon woodlands 259
icant patterns based on percent of variance explained by the ordination. Also, we estimated fl oristic pat-terns among OGUs by means of ordination scores. Patterns were explained based on the environmental factors changing in the same direction as the DCA gradients. Ordination diagrams of the 32 OGUs were also elaborated.
Multiresponse permutation procedure (MRPP)
The MRPP analysis was carried out to test fl oristic differences among the groups of pinyon according to McCune & Grace (2002) under the null hypothesis of no fl oristic differences between groups revealed in DCA. Details of the MRPP procedure can be found in Mielke (1991), Mielke & Berry (1976), and Zimmer-man et al. (1985). We used the Sorensen index since, according to Boyce & Ellison (2001), this coeffi cient is more effective when: a) weighs joint occurrences of species in different sites, b) emphasizes similarity, and c) there are few species in common among samples, the statistic T describe the separation between groups in a n-dimensional space; if T is more negative, then the differences among groups will be greater (Lesica et al. 1991, McCune & Grace 2002).
Canonical correspondence analysis (CCA)
We applied CCA, as described by ter Braak (1986, 1987), to arrange OGUs along environmental space and to determine which environmental factor drives the fl oristic relationships among the Mexican pin-yon woodlands. We hypothesized that the common patterns in species distributions might explain the fl oristic affi nities observed between pinyon wood-lands. CCA was applied to relate a 32 OGU-by-1153 plant species data matrix, with a 32 OGU-by-9 en-vironmental and geographical data matrix, as shown in Table 2. The environmental and geographic data were standardized using logarithmic transformation (McCune & Mefford 1999) to remove arbitrariness in the units of measure, and to make comparable the canonical coeffi cients (ter Braak 1986). The species-environment correlation was obtained, and the per-centage of accumulated variance for the relation spe-cies-environment was estimated. The Monte Carlo permutation test was used to indicate the signifi cance of the canonical axes, and ninety-nine permutations were required in all tests.
All types of analyses were performed using PC-ORD version 4, MjM Software Design, Oregon, U.S.A. (McCune & Mefford 1999).
Results
Plant associations on the pinyon pine woodlands
The pinyon-pine woodlands are associated with other species that defi nes their physiognomy. The pinyon-pine of P. quadrifolia, is associated with Adenostoma fasciculatum Hook. & Arn., Arctostaphylos patula Greene, Juniperus californica Carr., Opuntia acan-thocarpa Engelm. & Bigelow, Quercus dumosa Nutt. and Yucca schidigera Roezl ex Ortgies. P. monophylla woodlands share some of the above mentioned spe-cies as well as Quercus turbinella Greene and Rhus ovata S. Wats. and more diversity of Opuntia, such as O. chlorotica Engelm. & Bigelow, O. echinocar-pa Engelm. & Bigelow, and O. erinacea Engelm. & Bigelow ex Engelm. P. lagunae has Rhus integrifolia (Nutt.) Benth. & Hook., and such grass species as Muhlenbergia and Aristida.
The P. cembroides woodlands have more species diversity. In the Sierra Madre Oriental these pinyon woodlands are associated with J. deppeana Steud., J. fl accida Schlecht. and J. monosperma (Engelm.) Sarg., Rhus and Bouteloua spp. In Altiplano Mexicano, P. cembroides coexists with P. johannis, Quercus potosi-na Trel, and Q. crassifolia Humb. & Bonpl., Yucca fi -lifera Chabaud, Y. carnerosana (Trel.), and the grasses Bouteloua gracilis Lag. and B. curtipendula (Michx.) Torr.; whereas in Sierra Madre Occidental P. cembroi-des is associated with Quercus chihuahuensis Trel., Q. emoryi Torr., Q. grisea Liebm. and Q. rugosa Née, Calliandra humilis Benth., Stevia serrata Cav. and Dichondra argentea Humb. & Bonpl. ex Willd.
Pinus culminicola is associated with Holodiscus dumosus (Nutt. ex Hook.) Heller, and Symphoricar-pos microphyllus Kunth; in open sites it is associated with Stachys vulnerabilis Rzed. & Calderón and Sal-via microphylla HBK. In the P. nelsonii woodlands, the most common species are Brickellia veronicifolia (Kunth) Gray, Gymnosperma glutinosum (Spreng.) Less., Rhus virens Lindheimer ex Gray, and Quer-cus pringlei Seemen. Finally, the community of P. maximartinezii is associated with oaks such as Quer-cus castanea Née and Q. praecox Trel., and species of Muhlenbergia, Tagetes, and Penstemon.
Detrended Correspondence Analysis
The eigenvalues for the fi rst three ordination axes were 0.931, 0.693, and 0.478, indicating that they explained most of the variation in the species data. Axes accounted for 27% of the biological variation. The high eigenvalue (0.931) in axis one indicates a high correlation between species and sites. Length of the gradient for the fi rst axis (8.187 units) is notable, suggesting a signifi cant turnover in species composi-tion among OGUs along environmental gradients on axis one. The gradient length for axes two and three were 5.73 and 2.73, respectively. Since axis 3 included

260 M. Luna-Cavazos et al.
the least change in the fl oristic composition among OGUs, this axis will not be considered in the subse-quent discussion.
Figure 2 shows dispersion of OGUs on axes one and two. We observe on axis one a longitudinal pat-tern. On the right side of this axis are four OGUs located in the western part of the study area (SPMA, SJUA, PMAR, MART) belonging to the California province and dominated by P. quadrifolia and P. monophylla. On the left side of this axis are OGUs located in the middle and eastern parts of the study area: Sierra Madre Occidental (SMOCC), Altiplano Norte (ALTNTE), Altiplano Sur (ALTSUR), and Sierra Madre Oriental (SMO) provinces, which are dominated by pinyon species such as P. cembroi-des, P. johannis, P. maximartinezii, and others. The OGUs from the western tip are communities of low-er altitude (c.a. 1490 asl), with mean annual precipita-tion ~ 345 mm, moderate mean annual temperature of 15.3ºC, and a low average aridity index (13.7). In contrast, OGUs in other provinces have a higher average altitude, 2200 asl, annual precipitation of 400- 700 mm, mean annual temperature of 16 – 18ºC, and an aridity index of 15.4. Axis two separated two groups: a) OGUs from SMO (RAMO, GALE, SALT, ARAM- P. cembroides; MIQU, TAMA, GUAD- P. nelsonii and POTO- P. culminicola) joined to com-munities from ALTNTE (CIEN, PAIL, OCAM) plus two communities from ALTSUR (OROA, CORO), and b) communities of SMOCC (JUCH, PILA- P. maximartinezii; CUAU, ZARA, BALL, PAPA, MICH, SJMI- P. cembroides) and of ALT-SUR (AMAP, REYE, TAPI, GOZA, MGRA), plus the community LAGU belonging to Del Cabo prov-
ince. The pinyon pines from SMO-ALTNTE are on leptosols and have higher mean annual temperatures and lower precipitation than OGUs from SMOCC-ALTSUR, which are on pheozems and cambisols and experience 100 mm more rain and lower mean annual temperatures.
Multiresponse permutation procedure
The communities of the three main groups of pin-yons (G1, SMO-ALTNTE; G2, SMOCC-ALTSUR-Del Cabo; and G3, California) are fl oristically differ-ent based on the results of MRPP (shown in Table 3): we observe a high negative value of T. Based upon MRPP, the null hypothesis of no differences between groups is rejected. In other words, there are notable differences among species compositions among these groups. Also, when we compared the fl oras of G1 vs G2 vs G3, we observed differences in species compo-sition among these communities (Table 3).
The percent of common species among biogeo-graphic provinces is shown in Table 4. Pinyon pines from California and Del Cabo provinces shared a low number of species compared to other localities. SMO, SMOCC, ALTNTE, and ALTSUR provinces have a higher percent of shared taxa, especially be-tween SMOCC and ALTSUR.
Canonical correspondence analysis
The results of the CCA indicate that the three main axes explain 17.1% of the variance; the fi rst axis ac-
Fig. 2. Ordination of the Mexican pinyon woodlands on axes 1 and 2, by detrended correspondence analysis. Names of OGUs are in Table 1.

Floristic relationships among Mexican pinyon woodlands 261
counts for 6.8%; the second one, 5.2% and the third one, 5.1%. Based on a Monte Carlo permutation test, the correlation species-environment with the three fi rst axes was r = 0.991 p < 0.01, r = 0.985 p < 0.02, and 0.961 p < 0.18. Interset correlations between en-vironmental variables and ordination axes indicate that axis one showed greatest correlation with lon-gitude, elevation, and latitude. Axis two has greatest correlation with aridity index, mean annual precipita-tion, and elevation. Axis three is related mainly with latitude and mean annual precipitation (Table 5).
When we examined relationships of pinyon wood-lands along axes one and two with the environmental variables (Fig. 3), we found that sites located at higher longitudes (axis one) were separated from the others, were dominated by P. monophylla and, P. quadrifolia, and were characterized by lower annual precipitation (~ 345 mm), and elevation (~ 1400 m), in contrast to pinyon woodlands of Sierra Madre Occidental, Si-erra Madre Oriental, and Altiplano, all dominated by other pinyon-pine species. These OGUs receive more precipitation (400 – 500 mm; up to 700 mm in some cases) and are located at elevations > 2000 m (up to 3500 in the case of POTO).
Axis two (Fig. 3) separated pinyon woodlands with lower annual precipitation and lower aridity in-dices (CIEN, PAIL) from JUCH, PILA, PAPA, and others. Axis two also separated pinyon woodlands at elevations < 2000 m (CIEN, PAIL, RAMO OCAM, GUAD, GOZA, MIQU, and TAMA) from those at elevations > 2000 m. Other trends could be ob-served along axis two: pinyon woodlands from Sierra Madre Oriental dominated by P. cembroides (PAIL, RAMO, OCAM, ARAM) are somewhat separated from OGUs in the Sierra Madre Occidental (CUAU, ZARA, BALL, PAPA, MICH, SJMI) and Altiplano Sur (AMAP, REYE, TAPI), which are dominated by P. cembroides. Two pinyon woodlands dominated by P. nelsonii (TAMA, MIQU) are close to each other
but GUAD (P. nelsonii) is not very close; however, all of them were close to pinyon woodlands dominated by P. cembroides. The pinyon woodlands of P. johan-nis (OROA, MGRA) were close to each other and closer to P. cembroides woodlands. On axis two, the P. maximartinezii woodlands were separated along a humidity gradient: they are the most humid pinyon woodlands of all those we studied.
Discussion
The ordination analysis detected meaningful pat-terns among the fl oristic compositions of the pinyon woodlands, and showed that the main patterns of variation could be primarily attributed to longitude, latitude, and elevation. The results of CCA further confi rmed the trend of these gradients. The longest gradient (axis one) was a longitudinal one, whereas humidity and elevation predominated on axis two. The robustness of our fi ndings is indicated by the fact that the results of these different analyses all showed that longitude and elevation were the main environ-mental factors responsible for determining the fl oral affi nities among pinyon woodlands.
Biological diversity frequently varies within a site on the local scale, between sites within a regional mesoscale, and between regions on a global or geo-graphical scale (Menge & Olson 1990). Thus, we dis-cuss the fl oristic affi nities among pinyon woodlands either separately, or (as appropriate) grouped, as indi-cated by our analysis.
Pinus monophylla, P. quadrifolia and P. lagunae woodlands
Both DCA and CCA demonstrated that longitude was a main factor separating pinyon woodlands lo-cated in the western tip of the study area. Myklestad
ξ (delta) Observed Expected Variance T p Three groups 0.8228 0.9008 0.0003419 -13.3306 0.00000000 G1 vs G2 0.8327 0.8758 0.0002002 -9.6268 0.000000001 G1 vs G3 0.8206 0.8915 0.0007761 -8.0506 0.00002243 G2 vs G3 0.8097 0.8850 0.001127 -7.0961 0.00004178
Table 3. Test of differences of species composition among three groups of pinyon pines as observed in DCA, based on MRPP analysis. G1, SMO + ALTNTE; G2, SMOCC + ALTSUR + Del Cabo; G3, California.
California Del Cabo SMO AN SMOCC AS California ------ Del Cabo 0.52 ----- SMO 1.86 1.87 ----- ALTNTE 1.47 1.61 15.96 ----- SMOCC 1.56 2.99 12.00 10.68 ----- ALTSUR 1.40 3.39 17.97 15.05 22.68 -----
Table 4. Percent of shared species among Mexican biogeographic provinces, based on taxa located in pinyon pine communities.

262 M. Luna-Cavazos et al.
t-values Variance Variables Axis 1 Axis 2 Axis 3 Axis 1 Axis 2 Axis 3 Latitude -0.614 0.364 -0.439 0.029 -0.059 -0.924 0.0487 Longitude -0.922 -0.083 0.073 -0.942 -0.465 -0.097 0.0176 Elevation 0.657 -0.643 -0.201 0.066 -0.776 -0.994 0.0758 Soil 0.002 -0.050 -0.255 -0.164 -0.253 -0.331 0.1540 Mean of maximum annual
temperature 0.353 0.316 0.216 -0.807 -3.235 -0.922 0.0426
Mean of minimum annual temperature
0.268 0.559 -0.049 -1.564 -3.142 -2.192 0.1400
Mean annual temperature 0.340 0.509 0.058 1.037 7.611 7.798 0.0628 Mean annual precipitation 0.109 -0.675 0.326 4.078 -5.702 -17.915 0.1360 Aridity index 0.010 -0.714 0.268 -4.621 6.520 20.828 0.1570
Table 5. Canonical Correspondence Analysis. The most signifi cant t-values for axes 1 and 2 are bold-faced.
Fig. 3. Ordination of the Mexican pinyon woodlands on axes 1 and 2, by canonical correspondence analysis. Names of OGUs are in Table 1.
& Birks (1993) found longitudinal variation in the dis-tribution of Salix species, and they noted that climate varies with longitude, infl uencing plant distribution patterns. A similar effect may account for some of the variation seen in the Mexican pinyon woodlands on the western side of the study area (OGUs from the California province), where rainfall is lower than in the eastern part of the study area (Table 2). Latitude also has a large effect on most climates, particularly in terms of temperature and rainfall, and thus it also
has an effect on fl oristic composition (Bowman et al. 1985). Consistent with this, we found latitude-asso-ciated regional differences among these three pinyon woodlands. The P. lagunae woodland is located in Sierra de la Laguna, Baja California Sur, c.a. 800 km south of the other two pinyon woodlands (Arriaga et al. 1992) and while the P. lagunae woodlands are found in a mesic area receiving 670 mm annual pre-cipitation, the P. monophylla and P. quadrifolia wood-lands inhabit much drier areas receiving ~ 345 mm

Floristic relationships among Mexican pinyon woodlands 263
annual precipitation (Table 2). At the local level, the fl oristic differences between P. quadrifolia and P. monophylla woodlands are probably due to climate and aspect, since P. monophylla has a more defi ned xeric preference and grows almost exclusively along the drier eastern slopes of Sierra de Juárez and Sierra de San Pedro Mártir (Peinado et al. 1994, Minnich & Franco-Vizcaíno 1998). In contrast, P. quadrifolia is restricted to the western semidry slopes and it is fre-quently allopatric with P. monophylla.
Pinus culminicola woodland
Previous studies (Hernández-Reyna & García-Moya 1985, Romero-Manzanares 2001) reported fl oris-tic similarities between geographically close pin-yon woodlands containing different pinyon species. However, this is not the case for the P. culminicola woodland, which is fl oristically distinct from near-by pinyon woodlands dominated by P. cembroides mainly GALE (Table 1) both belonging to SMO province. This difference may be due to the relatively high elevation of P. culminicola woodland (3500 m), in contrast to that of P. cembroides woodland (1700 – 2700 m). Elevation generally affects tempera-ture. In Mexico, the lapse rate is ~ 0.6 °C per 100 m rise in elevation (Ortiz-Solorio 1987). Thus the fl ora of the P. culminicola woodland is exposed to colder climatic conditions than those in the lower elevations of P. cembroides woodlands.
Pinus cembroides woodlands
As reported by Rzedowski (1978), P. cembroides woodlands are widely distributed across most dry climates, and are thus exposed to different environ-mental conditions that may infl uence their fl oristic compositions. The CCA results revealed that latitude (axis one), which directly affects the thermal gradient (Menge & Olson 1990), infl uenced the fl oristic dif-ferences observed between P. cembroides woodlands in the northern and southern regions of the study area. This is consistent with the fi ndings of Robert (1977), who reported that the tree layers of P. cem-broides woodlands in the north are different from those found in the south. In addition, the CCA re-sults further indicated that rainfall (axis two) corre-lated with the differences among pinyon woodlands, probably explaining the separation of P. cembroides communities of the Sierra Madre Occidental Range (ZARA, BALL, CUAU, PAPA, MICH, SJMI) from those of the Sierra Madre Oriental Range (such as RAMO, ARAM, MIQU, TAMA, Fig. 3). Passini (1983) indicated that the common species in P. cem-broides woodlands from western Mexico are differ-ent from those in eastern Mexico, and she reported that temperature and rainfall are the most important factors determining the fl oristic differences among P. cembroides woodlands. A relevant result is the high-
est fl oristic similarity among OGUs of SMOCC and ALTSUR (Fig. 2 and 3, Table 4), probably because they share similar climatic conditions, which have favored species migration between both provinces. Passini (1983) indicated that – in the zone at the west side of Altiplanicie Central and Sierra Madre Occi-dental- P. cembroides communities are fl oristically more homogeneous. A similar case is reported by Tu-omisto et al. (2003), who observed a greater fl oristic affi nity among nearby geographic sites with similar environmental conditions. Martinez (1995) analyzed the distribution of species of Phaseolus in Mexico, and he concluded that a high number of these species were common among OGUs from Sierra Madre Oc-cidental and Altiplanicie Range.
Pinus johannis and P. remota woodlands
The P. johannis woodlands showed contrasting pat-terns based on regional location, with the Cuatro Cienegas (CIEN) pinyon woodlands from ALTNTE of the northern area (Table 1, Fig. 2 and 3) differing from the rest of the P. johannis woodlands (AMAP, MGRA, CORO, OROA) from ALTSUR, which were more similar among them. The distinct na-ture of the Cuatro Cienegas woodland may be due to isolation, altitude, and soil type. This woodland, which contains both P. johannis and P. remota, is more or less geographically isolated from all other pinyon woodlands, leading to a higher percentage of endemism. Pinkava (1984) reported that the Cuatro Cienegas region has one of the richest fl oras in the Chihuahuan Desert, including many endemic taxa, some of which are located in the oak-pinyon wood-lands. In addition, the Cuatro Cienegas study site is located at 1800 m altitude, much lower than the 2400 – 2500 m typical of other P. johannis communi-ties. Perry (1991) mentioned that P. remota inhabits the lowest elevations of all pinyons. Lastly, the as-sociation between P. johannis and P. remota could be an indication of edaphic factors, since Malusa (1992) and Farjon & Styles (1997) both reported that these pinyon woodlands grew on basic, limestone-derived calcareous soils.
The P. johannis woodlands with P. cembroides (Fig. 2 and 3) have a high degree of fl oristic similar-ity. Romero-Manzanares et al. (1996) indicated that P. cembroides and P. johannis are sympatric at drier locations in the Altiplano Range, while Perry (1991) described P. cembroides woodlands of the Sierra Madre Oriental Range as being associated with P. jo-hannis. Although various P. johannis and P. cembroi-des woodlands grouped together, we did note some fl oristic differences at the local level. The differences could be due to soil characteristics, as Romero-Man-zanares et al. (1996) reported that P. johannis is gen-erally restricted to sedimentary substrates, whereas P. cembroides has a preference (but no strict require-ment) for igneous substrates. Other studies have suggested that local fl oristic differences between P.

264 M. Luna-Cavazos et al.
cembroides and P. johannis woodlands could be the result of micro-environmental variations in moisture and aspect. Romero-Manzanares (2001) noted that P. cembroides community was found at drier sites with southern exposures, whereas nearby P. johannis and associated species preferred mesic, north-facing sites.
Pinus nelsonii woodlands
The analyses showed that the species composition of P. nelsonii woodlands is most similar to P. cembroi-des (Fig. 2 and 3). Previous reports have identifi ed scattered populations of P. nelsonii in dry, semidry, and temperate environments along the Sierra Madre Oriental Range in northeast México. In some of these cases, P. nelsonii was associated with P. cembroides (Suzán 1987, Perry 1991). Suzán (1990) used multi-variate analysis to examine fl ora and vegetation of different sites containing P. nelsonii, and found that these sites were fl oristically similar. Also, Hernán-dez-Reyna & García-Moya (1985) found greater fl o-ristic similarities among geographically closer com-munities of P. nelsonii and P. cembroides woodlands and lower fl oristic similarities between more distant populations of P. cembroides.
Pinus maximartinezii woodlands
The ordinations revealed that the species composi-tion of P. maximartinezii (JUCH, PILA) communi-ties were similar to those of other pinyon woodlands from the Sierra Madre Occidental Range, such as SJMI, PAPA, CUAU, and ZARA (Fig. 2), indicating strong affi nities among OGUs in this physiographic province. As shown in Fig. 3 and Table 5, rainfall was correlated with axis 2, the P. maximartinezii wood-land receive ~ 700 mm of rain. Gentry (1988) men-tioned that rainfall is a climatic factor particularly responsible for species distribution in semi-arid parts of North America. Since P. maximartinezii tends to occupy higher pluvial regimes in contrast to other pinyon woodlands, the fl ora of these communities are typical of more humid sites, in contrast to the fl ora of pinyon woodlands from drier regions.
Acknowledgements. We acknowledge Professors M.G. Bar-bour and R.A. Minnich, and an anonymous reviewer, who re-viewed and improved this manuscript.
References
Arriaga, L., Díaz, S. & Mercado, C. (1992): The oak and oak-pine forest of Baja California Sur: present state and manage-ment perspectives. – In: Ecology and management of oak and associated woodlands: perspectives in the Southwest-ern United States and Northern Mexico, General Technical Report RM-218, pp 41 – 68. – Forest Service, USDA. Sierra Vista, Arizona.
Axelrod, D.I. (1981): Holocene climatic changes in relation to vegetation disjunction and speciation. – Amer. Nat. 117: 847 – 870.
Binnema, T. (1996): Presettlement rangeland management on the Northern plains. – Rangelands 18: 217 – 218.
Birks, H.J.B. (1987): Recent methodological developments in quantitative descriptive biogeography. – Ann. Zool. Fennici 24: 165 – 178.
Bowman, R.A., Mueller, D.M. & McGinnies, W.J. (1985): Soil and vegetation relationships in Central Plains saltgrass meadow. – J. Range Manage 38: 325 – 328.
Boyce, R.L. & Ellison, P.C. (2001): Choosing the best similarity index when performing fuzzy set ordination on binary data. – J. Veg. Sci. 12: 711 – 720.
Caballero, D.M. & Ávila, R. (1989): Importancia actual y po-tencial de los pinos piñoneros en México. – In: J.D. Flores, J. Flores, E. García & R.H. Lira (eds.): Memorias del III Sim-posio nacional sobre pinos piñoneros, pp 18 – 22. – Universi-dad Autónoma Agraria Antonio Narro/ Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. México.
Comisión Nacional para el Conocimiento y Uso de la Biodi-versidad-CONABIO (1997): “Provincias biogeográfi cas de México”. Escala 1: 4 000 000. México.
Crovello, T.J. (1981): Quantitative biogeography: an overview. Taxon 30: 563 – 575.
De Oliveira, A.A. & Nelson, B.W. (2001): Floristic relationships of terra fi rme forest in the Brazilian Amazon. – For. Ecol. Manag. 146: 169 – 179.
Dzwonko, Z. & Loster, S. (2000): Syntaxonomy and phytogeo-graphical differentiation of the Fagus woods in the south-west Balkan Penninsula. – J. Veg. Sci. 11: 667 – 678.
FAO-UNESCO-ISRIC (1988): Mapa de Suelos Dominantes. 1a. aproximación, SEMARNAP-CP-INEGI, México, D.F
Farjon, A. & Styles, B.T. (1997): Pinus (Pinaceae). – In: Flora Neotropica Monograph 75, pp 10 – 67. – The New York Bo-tanical Garden.
García, A. & González, S. (1991): Flora y vegetación de la cima del Cerro Potosí, Nuevo León, México. – Acta Bot. Mex. 13: 53 – 74.
Gauch, H.G. (1982): Multivariate analysis in community ecol-ogy. – Cambridge Univ. Press, New York.
Gentry, A.H. (1988): Changes in plant community diversity and fl oristic composition environmental and geographical gradi-ents. – Ann. Missouri Bot. Gard. 75: 1 – 34.
González, J. (1998): Los bosques piñoneros de México, estudio del bosque de Pinus johannis M-F. Robert, en Concepción del Oro, Zacatecas. – Tesis de Maestría en Ciencias. Univer-sidad Autónoma Chapingo, México.
González, S., González, M. & Cortés, A. (1993): Vegetación de la reserva de la biosfera “La Michilía, Durango, México. – Acta Bot. Mex. 22: 1 – 104.
Hernández-Reyna, A. (1985): Análisis estructural de los piño-neros del Altiplano Potosino-Zacatecano. – Tesis de Maestría en Ciencias. Colegio de Postgraduados, Chapingo, México.
Hernández-Reyna, A. & García-Moya, E. (1985): Análisis es-tructural de los piñonares del Altiplano Potosino. – Agro-ciencia 62: 7 – 20.
Hill, M.O. & Gauch, H.G. (1980): Detrended correspondence analysis, an improved ordination technique. – Vegetatio 42: 47 – 58.
Justus, J & Sarkar, S. (2002): The principle of complementarity in the design of reserve networks to conserve biodiversity: A preliminary history. – J. Biosc. 27: 421 – 435.
Lanner, R.M. (1981): The pinyon pine: a natural and cultural history. University of Nevada Press, Reno.

Floristic relationships among Mexican pinyon woodlands 265
Lesica, P., McCune, B., Cooper, S.V. & Hong, W.S. (1991): Differences in lichen and bryophyte communities between old-growth and managed second-growth forests in the Swan Valley, Montana. – Can. J. Bot. 69: 1745 – 1755.
Malusa, J. (1992): Phylogeny and biogeography of pinyon pines (Pinus subsect. Cembroides). – Syst. Bot. 17: 42 66.
Martinez, J.V. (1995): Fitogeografía de los taxones silvestres de Phaseolus en México y Guatemala. – Tesis de Maestro en Ciencias. Colegio de Postgraduados, Montecillo, México
Martonne, E.D. (1926): Aréisme et indices d’aridité. Académie des Sciences, Paris. – Comptes Rendus 182: 1395 – 1398.
McCoy, E.D. & Heck, K.L. (1987): Some observations on the use of taxonomic similarity in large-scale biogeography. – J. Biogeog. 14: 79 – 87.
McCune, B & Grace, J.B. (2002): Analysis of ecological com-munities. – MjM Software Design, Gleneden Beach, Oregon, USA.
McCune, B. & Mefford, M.J. (1999): PC-ORD. Multivariate analysis of ecological data, version 4. – MjM Software De-sign, Gleneden Beach, Oregon, USA.
McLaughlin, S.P. (1994): Floristic plant geography: the classifi -cation of fl oristic areas and fl oristic elements. – Progr. Phys. Geogr. 18: 185 – 208.
Menge, B.A. & Olson, A.M. (1990): Role of scale and envi-ronmental factors in regulation of community structure. – TREE 5 (2): 52 – 57.
Mielke, P.W. (1991): The application of multivariate permuta-tion methods based on distance functions in the earth sci-ences. – Earth-Science Reviews 31: 55 – 71.
Mielke, P.W. & Berry, K.J. (1976): Multi-response permutation procedures for a priori classifi cations. – Commun. Statist. Theor. Meth. A5:1409 – 1424.
Miller, R.F. & Wigand, P.E. (1994): Holocene Changes in semi-arid pinyon-juniper woodlands. – BioScience 44: 465 – 474.
Minnich, R.A. & Franco-Vizcaíno, E. (1998): Land of chamise and pines: historical descriptions of vegetation in northern Baja California. – University of California Publications in Botany, v. 80 p.1 – 166.
Moora, M., Daniell, T., Kalle, H., Liira, J., Püssa, K., Roosaluste, E., Öpik, M., Wheatley, R. & Zobel, M., (2007): Spatial pat-tern and species richness of boreonemoral forest understo-rey and its determinants – A comparison of differently man-aged forests. – For. Ecol. Manag. 250: 64 – 70.
Myklestad, A. & Birks, H.J.B. (1993): A numerical analysis of the distribution patterns of Salix L. species in Europe. – J. Biogeog. 20: 1 – 32.
Ortiz-Solorio, C.A. (1987): Elementos de agrometeorología cu-antitativa. – Universidad Autónoma Chapingo, México.
Palmer, M.W. (1993): Putting things in even better order: the advantages of canonical correspondence analysis. – Ecology 74: 2215 – 2230.
Passini, M.-F. (1982): Les forêts de Pinus cembroides au Mex-ique. Mission Archaeological et ethnologique Française au Mexique, Études Mesoaméricaines II-5. – Editions Recher-che sur les Civilisations. Cahier No. 9. Paris.
Passini, M.-F. (1983): Un exemple de forêt tropicale du Mex-ique: la forêt de Pinus cembroides Zucc. – Bull Soc. Bot. France 130: 69 – 80.
Passini, M.-F. (1985): Structure et régénération des formations ligneuses á Pinus maximartinezii Rzed., Mexique. – Bull. Soc. Bot. France 4/5: 327 – 339.
Passini, M.-F. (1987): Análisis de la distribución geográfi ca de los pinos piñoneros en Europa y Asia. – In: M.F. Passini, D. Cibrián & T. Eguiluz (eds.): II Simposio nacional sobre pinos piñoneros, pp 65 – 81. – Centre d’Etudes Mexicaines et Centraméricaines, México, D.F. Universidad Autónoma
Chapingo, México – Centro de Genética Forestal, A. C., Chapingo, México.
Passini, M.-F. (1994): Releves fl oristiques des forets de pins pi-gnons au Mexique (Florística de los piñoneros en México). – Université Pierre et Marie Curie, Paris.
Passini, M.-F. & Pinel, N. (1989): Ecology and distribution of Pinus lagunae, in the Sierra de La Laguna, Baja California Sur, México. – Madroño 36: 84 – 92.
Peinado, M., Bartolomé, C., Delgadillo, J. & Aguado, I. (1994): Pisos de vegetación de la Sierra de San Pedro Mártir, Baja California, México. – Acta Bot Mex 29: 1 – 30.
Perry, J.P. (1991): The pines of Mexico and Central America. – Timber Press, Portland, Oregon.
Pinkava, D.J. (1984): Vegetation and fl ora of the Bolsón of Cu-atro Cienégas region, Coahuila, Mexico: IV. Summary, ende-mism and corrected catalogue. – J. Arizona-Nevada Acad. Sci. 19: 23 – 47.
Reyes, J.A., González, F. & García, J. (1996): Flora vascular de la Sierra Monte Grande, municipio de Charcas, S.L.P., Méxi-co. – Bol. Soc. Bot. México 58: 31 – 42.
Robert, M.-F. (1973): Contribution l’etude des forêts de Pinus cembroides dans lést du Mexique. Thése de spécialité (écolo-gie). – Montpellier.
Robert, M.-F. (1977): Aspects phytogéographiques et écologiques des forêts de Pinus cembroides. I. Les forêts de l’est et du nord-est du Mexique. – Bull. Soc. Bot. France 124: 197 – 216.
Romero-Manzanares, A. (2001): Historia natural, ecología de poblaciones y fi tosociología de Pinus cembroides y Pinus johannis (piñoneros) del centro de México. Tesis de Docto-rado. – Facultad de Ciencias, Universidad Nacional Autóno-ma de México, México.
Romero-Manzanares, A. & García-Moya, E. (2002): Estabilidad y elasticidad de la composición fl orística de los piñonares de San Luis Potosí, México. – Agrociencia 36: 243 – 254.
Romero-Manzanares. A., García-Moya, E. & Passini, M.-F. (1996): Pinus cembroides s.l. y Pinus johannis del Altiplano Mexicano: una síntesis. – Acta Bot. Gallica 143: 681 – 693.
Romero-Manzanares, A., Aguirre-Rivera, J.R. & García-Moya, E. (1999): El papel del hombre en la evolución de los piñone-ros americanos. – Agrociencia 33: 473 – 481.
Rzedowski, J. (1978): Vegetación de México. – Limusa, México.Suzán, H. (1987): Estudios ecológicos en Pinus nelsonii de Tam-
aulipas. In: M.F. Passini, D. Cibrián & T. Eguiluz (eds.): II Simposio nacional sobre pinos piñoneros, pp 199 – 210. – Centre d’Etudes Mexicaines et Centraméricaines, México, D.F. Universidad Autónoma Chapingo, México – Centro de Genética Forestal, A. C., Chapingo, México.
Suzán, H. (1990): Estructura de un bosque de Pinus nelsonii Shaw en Tamaulipas. – BIOTAM 1: 29 – 35.
ter Braak, C.J.F. (1986): Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. – Ecology 67: 1167 – 1179.
ter Braak, C.J.F. (1987): The analysis of vegetation-environment relationships by canonical correspondence analysis. – Veg-etatio 69: 69 – 77.
Tuomisto, H.K., Ruokolainen, M. & Yli-Halla, M. (2003): Dispersal, environmental, and fl oristic variation of western Amazonian forest. – Science 299: 241 – 244.
Van Devender, T.R. (1977): Holocene woodlands in the south-western deserts. – Science 198: 189 – 192.
Van Devender, T.R. & Spaulding, W.G. (1979): Development of vegetation and climate in the southwestern United States. – Science 204: 701 – 710.
Van Devender, T.R., Thompson, R.S. & Betancourt, J.L. (1987): Vegetation history of the deserts of southwestern North

266 M. Luna-Cavazos et al.
America; the nature and timing of the Late Wisconsin-Ho-locene transition. In: W.F. Ruddiman & H.E. Wright (eds.): North America and adjascent oceans during the last degla-ciation, pp 323 – 352. – Geological Society of America, Boul-der, Colorado.
Villarreal, J.A. (1994): Flora vascular de la Sierra de la Paila, Coahuila, México. – SIDA 16: 109 – 138.
Zimmerman, G.M., Goetz, H. & Mielke, P.W. (1985): Use of an improved statistical method for group comparisons to study effects of prairie fi re. – Ecology 66: 606 – 611.
Address of the authorsDr. Mario Luna-Cavazos, Dra. Angélica Romero-Manzanares and Dr. Edmundo García-MoyaBotánica, Colegio de Postgraduados, Campus Montecillo. Km 36.5, carretera México-Texcoco, Montecillo, Texcoco, Edo. de México, C.P. 56230. MÉXICO. E-mails: [email protected]; [email protected]; [email protected]





























