Embed Size (px)
Citation preview

Y.V. Mendez-Fernandez Enhanced binding of low-affinity antibodiesR.P. Pogulis interacting simultaneously with targeted cellM.S. BlockA.J. Johnson surface molecules and Fc receptorS.T. KuhnsK.S. AllenM.J. HansenL.R. Pease
Key words:antibodies; antigens/ peptides/ epitopes;Fc receptors; lymphocytes; MHC
Received 2 July 2002, revised 6 September 2002,accepted for publication 15 September 2002
Copyright c Blackwell Munksgaard 2002Tissue Antigens . 0001-2815
Tissue Antigens 2002 60: 515–525Printed in Denmark . All rights reserved
515
Abstract: The possibility that different forms of class I molecules mightbe expressed on the cell surface of lymphocytes has been investigatedperiodically over the past several decades. A series of majorhistocompatibility complex (MHC) class I-specific monoclonal antibodies,including the commonly used antibodies 64-3-7 and 25-D1.16, bind B cellsdifferentially, suggesting the existence of differentially expressed class I-associated cell surface determinants on B lymphocytes. However, the abilityof antibodies to bind cells is determined by the sum of interactions betweenthe antibodies and the molecules expressed on the cell surface. Theinteractions of class I-specific antibodies with B cells were dissected,revealing dual specificity of the antibodies for the targeted class Imolecules, as well as to Fc receptors preferentially expressed by B cells. Wedemonstrate that antibodies simultaneously bind targeted class I moleculesand Fc receptors expressed on the surface of B cells. Simultaneous bindingto two cell surface structures significantly enhances the class I-specificbinding pattern of certain antibodies by increasing their avidity, leading toapparent cell-specific differences in MHC expression patterns. We concludethat no differences in MHC structures need be postulated to account for theobserved binding patterns.
The anti-Db antibody H141-31 reportedly binds a form of the H-2Db
molecule that is primarily expressed by B lymphocytes in the mouse
spleen (1). In the course of staining for major histocompatibility
complex (MHC) class I molecules in mouse splenocytes, we have
identified several additional monoclonal antibodies (mAb) that ap-
pear to bind preferentially to B lymphocytes. L368 is a monoclonal
antibody originally described as specific for human beta-2-micro-
globulin (hb2m) (2). This antibody was found to be cross-reactive
with mouse MHC class I and to bind to the B cell population in the
spleen. Two additional mAb, 64-3-7 and 25-D1.16, specific for Ld
and Kb, respectively (3,4), follow the same binding pattern seen for
H141-31 and L368.
Evidence that heterogeneous forms of class I molecules are ex-
Authors’ affiliations:
Y.V. Mendez-Fernandez,R.P. Pogulis,M.S. Block,A.J. Johnson,S.T. Kuhns,K.S. Allen,M.J. Hansen,L.R. Pease
Department of Immunology,Mayo Medical and GraduateSchools, Mayo Clinic,Rochester, MN, USA
Correspondence to:Larry R. Pease, PhDDepartment of ImmunologyMayo Graduate and MedicalSchoolsMayo Clinic200 First Street SWRochester, MN 55905USATel: 507-284-8177Fax: 507-266-0981e-mail:pease.larry/mayo.edu

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
pressed in the spleen has been previously presented (5–7). The finding
that structurally distinct class I molecules are differentially expressed
on lymphocytes would be of considerable interest. Consequently, we
investigated the hypothesis that B lymphocytes preferentially ex-
press alternative forms of class I molecules relative to class I mol-
ecules expressed on the surface of T cells. Here we report that the spe-
cific binding of these antibodies to B cells results from the simul-
taneous interaction of the antibody binding (Fab) and constant (Fc)
portions with both the specific epitope and the Fc gamma receptor,
respectively. We conclude that these binding patterns do not provide
evidence for alternative class I structures. Rather they reflect a
greater avidity of the antibody for B cells resulting from a low affinity,
yet specific interaction of the antibody binding site with a specific
antigen, together with a low affinity binding of the constant region of
the antibody with the Fc gamma receptor IIB (FcgRIIB).
Materials and methods
Mice
C57BL/6 J, BALB/cJ, C57BL/10, and B10.BR mice were obtained
from Jackson Laboratory (Bar Harbor, ME). B6.K1, B6.K2, B6.K3
and B6.K4 mice were provided by L. Flaherty (Wadsworth Center
for Laboratories and Research, Albany, NY). B10.R40 and B10.SP2
mice were provided by K. Fischer Lindahl (University of Texas
South-western Medical Center, Dallas, TX). The Kbm3, FVB/Kb and
FVB/Db transgenic mice were produced in the Mayo Immunogen-
etics colony (Mayo Clinic, Rochester, MN). Class I -/- mice were ob-
tained with permission from J. Forman (University of Texas South-
western Medical Center). Two or more individuals of each strain
were analyzed in all cases.
Antibodies
Anti-mouse H-2 monoclonal antibodies B8-24–3 (IgG1, anti-Kb),
K10.56 (IgG2b, anti-Kb) and Y-3 (IgG2b, anti-Kb), 25-D1.16 (IgG1, anti-
Kb-OVA), 28–11–5 s (IgM, anti Db) and 28–14–8 s (IgG2a, anti-Db), 66-
3-5 (IgG2a, anti-Ld), and 64-3-7 (IgG2b), which reacts with the a1 do-
main of open Ld (3), were used in the form of cultured supernatants or
ascites (K10.56). The hybridoma cell lines for the mAbs B8-24–3, 28–
11–5s, Y-3, and 28–14–8s were obtained from the American Type Cul-
ture Collection. The anti-Ld hybridomas 66-3-5 and 64-3-7 were ob-
tained from K. Ozato (National Institutes of Health, Deutsches
Krebsforschungszentrum, Heidelberg, Germany). The K10.56 hy-
bridoma was a gift of G. Hammerling (Sloan Kettering Institute, Beth-
esda, MD). 25-D1.16 obtained from J. Yewdell (National Institutes of
516 Tissue Antigens 2002: 60: 515–525
Health) and was purified and biotinylated using the FluoReporter
Mini-Biotin-XX Protein Labeling Kit from Molecular Probes. Mono-
clonal L368 (IgG1, Eugene, OR), originally described as specific for
hb2m (2), was obtained from American Type Culture Collection
(ATCC, Manassas, VA) and used in the form of supernatant or puri-
fied antibody labeled with fluorescein isothiocyanate (FITC, St. Louis,
MO) using the Fluorotag FITC Conjugation Kit from Sigma. Biotinyl-
ated rat antimouse Fc gamma receptor III/II (CD16/CD32,2.4G2,
IgG2b) was obtained from PharMingen (San Diego, CA).
Cell lines
The thymoma-derived EG7 line expressing the ovalbumin (OVA)
protein was obtained from ATCC.
Peptides
The SIINFEKL peptide from chicken ovalbumin amino acids 257–
264, and the control SIYRYYGL peptide were synthesized and puri-
fied at the Mayo Protein Core Facility. Peptide was used at a concen-
tration of 100 mM.
Flow cytometry
Mouse spleens were homogenized in 1 ml of RPMI, incubated with
ACK buffer for 5 min at room temperature for lysis of red blood cells,
centrifuged and resuspended in FACS medium containing 1% BSA
and 0.02% sodium azide in HBSS. Mouse splenocytes were incubated
with 50 ml of supernatant derived from hybridoma cell cultures,
washed with FACS medium and incubated with FITC-conjugated
goat antimouse IgG F(Ab)2ƒ fragment (JAXELL Accurate Chemical
and Scientific Group, Westburg, NY). For three-color analysis, these
incubations were followed with staining for B220 with phycoerythrin
(PE)-conjugated mAb RA3–6B2 (PharMingen, Carlsbad, CA), and for
CD3e with biotinylated mAb 145–2C11 (PharMingen, Carlsbad, CA)
followed by streptavidin/RED-613 (Gibco-BRL). Stained cells were
fixed in 2% paraformaldehyde and analyzed by flow cytometry. Flow
cytometry was performed on either a FACScan or FACSCalibur
(Beckton Dickinson). Data was analyzed using Cell Quest (Becton
Dickinson). Nonviable cells were excluded based on the forward scat-
ter profile. Fluorescence channel data is shown in a log4 scale.
Blocking experiments
For the blocking experiments, splenocytes were stained for B220
and CD3e, washed and incubated with blocking supernatant or
anti-CD16/CD32 (1 : 100 dilution) for 30 min on ice. Without washing

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
between incubations, labeled L368 was added and incubated for
another 30 min on ice. Cells were washed with FACS medium, fixed
in 2% paraformaldehyde and analyzed by flow cytometry as de-
scribed earlier.
EG7 cells were pulsed with peptide for 1 h at 37æC in RPMI-10
medium and washed for incubation with antibody. Peptide-pulsed
or non-pulsed EG7 cells were incubated with 2.4G2 antibody for
10 min, then biotinylated 25-D1.16 was added for 20 min. Cells were
washed twice with FACS medium, incubated with streptavidin-PE
(Pharmingen), washed again, and fixed in 2% paraformaldehyde
for flow cytometry analysis.
Results
Monoclonal antibody L368 detects an epitope that is
preferentially expressed by mouse B lymphocytes
Monoclonal antibody L368 was derived from mice immunized with
the human B cell line RPMI-8866, and was shown to be specific for
human b2m (2,8). In the course of staining for the product of a
human b2m transgene in C57BL/6 mice, we observed a biphasic
staining profile in control C57BL/6 splenocyte populations (Fig. 1).
In an attempt to identify the cross-reactive splenocyte popula-
tion(s), we analyzed C57BL/6 splenocytes by three-color flow cyto-
Fig.1. L368 antibody binds differentially to two cell populationsin the spleen. Freshly isolated splenocytes from a C57BL/6 mouse wereincubated with L368-FITC antibody and analyzed by flow cytometry. Histo-gram shows analysis of gated lived cells.
517Tissue Antigens 2002: 60: 515–525
metry, staining with L368 in combination with a B cell-specific anti-
body that binds B220 (CD45/RO) and a T cell-specific antibody that
binds CD3e. As shown in Fig. 2(A), the L368 epitope in the mouse
is preferentially expressed on B cells. The L368 staining profile of
the B220–CD3– population is indistinguishable from that of CD3π
T cells (data not shown).
Analysis of the L368/B220/CD3 staining pattern in congenic mouse
lines showed that the L368 epitope is H-2 dependent, with minimal
expression on B10.BR (H-2k) and B10.S (H-2s), but strong expression
in B10.A (H-2a) and B10.D2 (H-2d) mice (data not shown). C57BL/
6 ¿ C3H/HJ (BXH) recombinant inbred lines confirmed the H-2 de-
pendence and the preferential B-cell reactivity of the L368 antibody
(data not shown). Analysis of the H-2 recombinant congenic lines
B6.K1, B6.K2, B6.K3, B6.K4, B10.R40, and B10.SP2 indicated that the
H-2 gene responsible for the preferential staining of B cells mapped
between TNFa and Q1 (Table 1). A prime candidate gene in this re-
Fig.2. Detection of B cell-associated epitopes by major histocom-patibility complex class I-specific monoclonal antibodies. Splenocytesfrom C57BL/6 were isolated and analyzed by three-color flow cytometry for theexpression of the L368 epitope, B220-PE (B lymphocyte marker) and CD3e-Red613 (T lymphocyte marker). Histograms show analysis of PEπ/FITC π double-positive (B cells) or Red 613π/FITC π double-positive (T cells)live cells. (A) Staining of CD3π and B220π splenocyte populations from a B6mouse with the L368 antibody. (B) Staining of CD3π and B220π splenocytepopulations from a B6 mouse with the anti-Db antibody 28–14–8s. Negativestaining shows autoflourescence of cells incubated with FACS medium only.

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
gion of the MHC was Db. However, staining with mAbs specific for
the a3 domain of Db demonstrated that the molecule itself is not pref-
erentially expressed on B cells (Fig. 2B and data not shown).
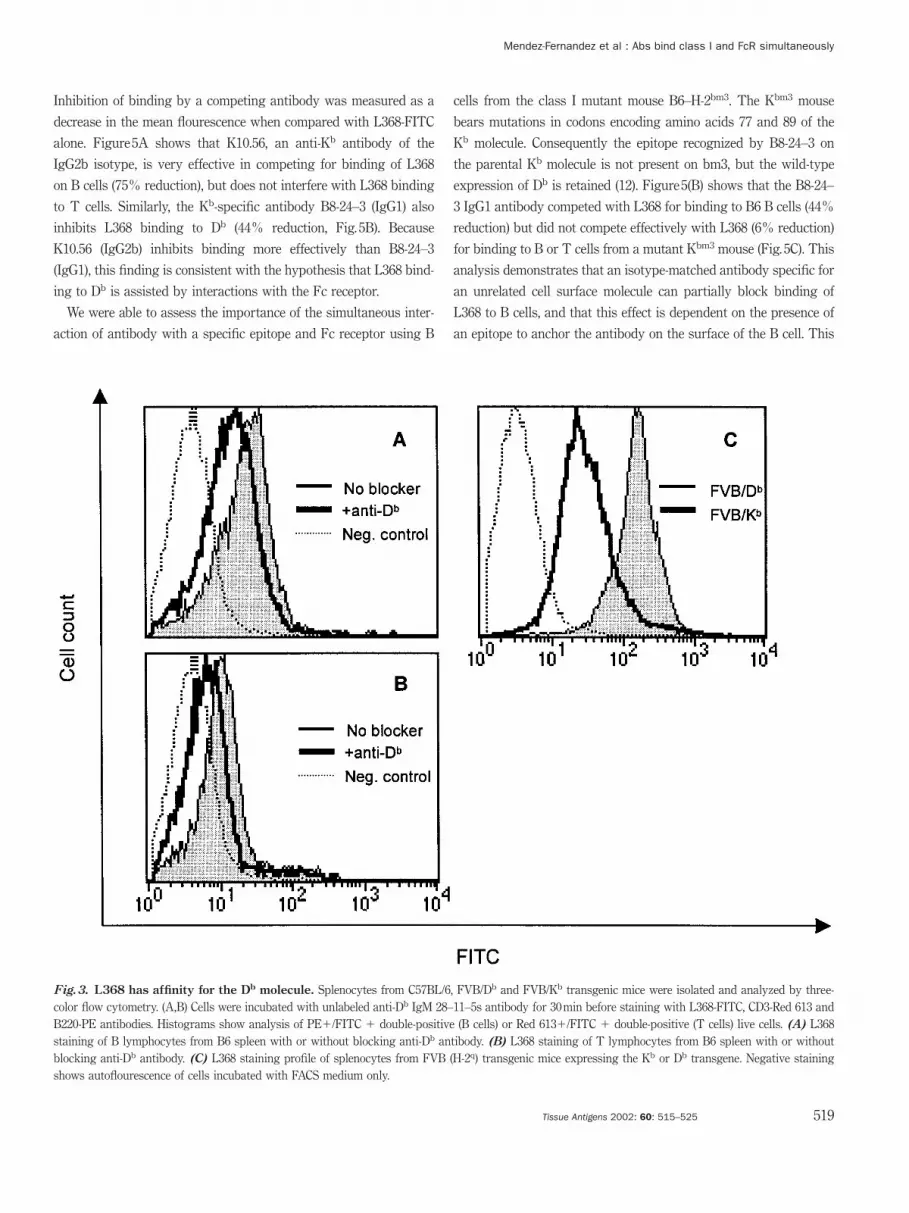
L368 has affinity for H-2Db
In order to further define the epitope specificity of L368, we per-
formed blocking experiments using reagents specific for the H-2Db
molecule. Mouse splenocytes from C57BL/6 mice (H-2b) were stained
with anti-CD3 and B220 antibodies and incubated with IgM antibody
from the 28–11–5s hybridoma cells that is specific for Db. After a 30-
min incubation with unlabeled antibody, cells were stained with L368-
FITC and analyzed by three-color flow cytometry. Inhibition of bind-
ing was measured as a decrease in the mean channel fluorescence.
The monoclonal antibody 28–11–5s was able to partially block bind-
ing of L368 to both B and T lymphocytes (Fig. 3A,B). However, while
28–11–5s was able to reduce L368 binding in T lymphocytes to near
background levels, L368 binding to B cells was only partially blocked
(Fig. 3A). These results demonstrate that L368 has affinity for the Db
molecule and is consistent with the hypothesis that B lymphocytes
may be expressing an alternate binding site that is not recognized by
the antibody 28–11–5s. Definitive proof that L368 recognizes an epi-
tope associated with the Db molecule comes from the analysis of mice
expressing a Db transgene (Fig. 3C). L368 antibody stains B cells iso-
lated from FVB-Db mice 6-fold more intensely than B cells isolated
from FVB-Kb mice, demonstrating directly that the antibody recog-
nizes the Db class I molecule.
L368 binding to the Fc receptor gamma II on B cells
accounts for differential staining
Our results indicate that the biphasic staining profile of L368 is the
result of the recognition of an epitope that is preferentially or
L368 epitope maps near H-2D
Strain* K 1 D Q T M bkgd.† L368‡
B6 b b b b b b B6 π
B6.K1 b b b k k k B6 π
B6.K2 b b b b k k B6 π
B6.K3 k k k b a (a) B6 – loª1
B6.K4 k k k k a (a) B6 – loª1
B10 b b b b b b B10 π
B1O.R40 b b sp2 sp2 sp2 sp2 B10 – loª1
B1O.SP2 sp2 sp2 sp2 sp2 sp2 sp2/b B10 π
* b, k, a and sp2 indicate the haplotypes from which the various major histocompatibility complex regions (K, I, D, Q, T, M) originate in the recombinant strains listed.† Genetic background of the recombinant strains listed is either C57BL/6 (B6) or C57BL/10 (B10).‡ Expression of the L368 epitope on B220 π splenocytes was assessed as described in Fig.2. Any strain displaying significantly reduced L368 reactivity compared with C57BL/6 is listed as - loª1.
Table1
518 Tissue Antigens 2002: 60: 515–525
uniquely expressed on B lymphocytes. The nature of the interaction
of L368 with B cells was further investigated. In contrast to T cells,
B lymphocytes constitutively express the Fc receptor gamma II (9–
11). Therefore, additional interactions of an antibody with a receptor
that is differentially expressed in two populations of cells could
account for observed increased binding.
To evaluate the possibility that the Fc receptor was responsible
for differential staining of B and T cells, possible interactions with
the Fc gamma receptor II (FcgIIRB) were blocked using 2.4G2 anti-
body. Pre-incubation with the anti-Fc receptor antibody partially
blocks binding of L368 to B lymphocytes (Fig. 4A), while it does not
have any effect on binding of L368 to T lymphocytes (Fig. 4B). Thus,
we conclude that binding of L368 to B cells is enhanced and stabil-
ized by interactions of the Fc portion of the antibody with the
FcgIIRB. As blocking the interactions of neither the Db molecule nor
the FcgIIRB with the L368 antibody reduced staining to background
levels, we combined the blocking strategies to determine whether
we could account for all the binding activity. As shown in
Figs 4(C,D), the ability of antibody L368 to stain Db on B cells was
completely blocked by the combined treatments in splenocytes from
both B6 and FVB/Db mice.
Interaction of antibodies with the Fc receptor is enhanced
by antigen-specific binding on the cell surface
To explore further the simultaneous binding of antibody to a speci-
fic epitope and Fc receptor on the same cell, mouse splenocytes
were preincubated for 30 min with unlabeled antibodies containing
different Fc gamma regions before labeling with L368-FITC. IgG
isotypes have decreasing affinities for the Fc receptor gamma II in
the following order: IgG2b � IgG2a �� IgG1 ��� IgG3. We tested
the ability of an IgG2b mAb specific for a different target on the
cell surface (Kb) to block the Db-specific binding of L368 to B cells.
Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
Inhibition of binding by a competing antibody was measured as a
decrease in the mean flourescence when compared with L368-FITC
alone. Figure 5A shows that K10.56, an anti-Kb antibody of the
IgG2b isotype, is very effective in competing for binding of L368
on B cells (75% reduction), but does not interfere with L368 binding
to T cells. Similarly, the Kb-specific antibody B8-24–3 (IgG1) also
inhibits L368 binding to Db (44% reduction, Fig. 5B). Because
K10.56 (IgG2b) inhibits binding more effectively than B8-24–3
(IgG1), this finding is consistent with the hypothesis that L368 bind-
ing to Db is assisted by interactions with the Fc receptor.
We were able to assess the importance of the simultaneous inter-
action of antibody with a specific epitope and Fc receptor using B
Fig.3. L368 has affinity for the Db molecule. Splenocytes from C57BL/6, FVB/Db and FVB/Kb transgenic mice were isolated and analyzed by three-color flow cytometry. (A,B) Cells were incubated with unlabeled anti-Db IgM 28–11–5s antibody for 30 min before staining with L368-FITC, CD3-Red 613 andB220-PE antibodies. Histograms show analysis of PEπ/FITC π double-positive (B cells) or Red 613π/FITC π double-positive (T cells) live cells. (A) L368staining of B lymphocytes from B6 spleen with or without blocking anti-Db antibody. (B) L368 staining of T lymphocytes from B6 spleen with or withoutblocking anti-Db antibody. (C) L368 staining profile of splenocytes from FVB (H-2q) transgenic mice expressing the Kb or Db transgene. Negative stainingshows autoflourescence of cells incubated with FACS medium only.
519Tissue Antigens 2002: 60: 515–525
cells from the class I mutant mouse B6–H-2bm3. The Kbm3 mouse
bears mutations in codons encoding amino acids 77 and 89 of the
Kb molecule. Consequently the epitope recognized by B8-24–3 on
the parental Kb molecule is not present on bm3, but the wild-type
expression of Db is retained (12). Figure 5(B) shows that the B8-24–
3 IgG1 antibody competed with L368 for binding to B6 B cells (44%
reduction) but did not compete effectively with L368 (6% reduction)
for binding to B or T cells from a mutant Kbm3 mouse (Fig. 5C). This
analysis demonstrates that an isotype-matched antibody specific for
an unrelated cell surface molecule can partially block binding of
L368 to B cells, and that this effect is dependent on the presence of
an epitope to anchor the antibody on the surface of the B cell. This

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
finding delineates the underlying mechanism that enhances binding
of antibodies similar to L368 to the surface of B cells.
Detection of a B cell-associated epitope by other MHC class
I-specific monoclonal antibodies
To extend the observation of preferential B-cell staining by MHC
class I-specific antibodies, we have analyzed a variety of antibodies
for reactivity with splenocyte populations by three-color flow cyto-
Fig.4. L368 binding to the FcgIIR on B cells accounts for differential staining. Mouse splenocytes from C57BL/6 and FVB/Db transgenic micewere incubated with the the anti-CD16/CD32 Fcg receptor antibody 2.4G2 or 2.4G2 plus the anti-Db antibody 28–11–5s for 30 min before staining with L368-FITC, CD3-Red 613 and B220-PE antibodies. Histograms show analysis of PEπ/FITC π double-positive (B cells) or Red 613π/FITC π double-positive (Tcells) live cells. (A) L368 staining of B lymphocytes blocked with the anti-Fcg receptor antibody. (B) L368 staining profile of T lymphocytes blocked withthe anti-Fcg receptor antibody. Negative staining shows autoflourescence of cells incubated with FACS medium only. (C) L368 staining of B lymphocytesfrom B6 spleen blocked with the anti-CD16/CD32 antibody and 28–11–5s. L368 staining of splenocytes from Class I -/- mice blocked with 2.4G2 and 28–11–5s served as the negative control. (D) L368 staining of B lymphocytes from FVB/Db blocked with the anti-CD16/CD32 antibody and 28–11–5s. L368 stainingof splenocytes from FVB/Kb mice blocked with 2.4G2 and 28–11–5s served as the negative control.
520 Tissue Antigens 2002: 60: 515–525
metry. The mAb 64-3-7 has been shown to be specific for an imma-
ture intracellular precursor of the fully folded mature form of the
Ld molecule (3). BALB/c (H-2d) splenocytes display a 64-3-7 staining
profile very similar to that observed with L368 (Fig. 6A), although
Ld expression as judged by the staining pattern of the Ld-specfic
antibody 66-3-5 is nearly equivalent in B- and T-cell populations
(Fig. 6B). The antibody 25-D1.16 has demonstrable specificity for
the Kb molecule (4). Similar to the Db-specific antibody L368 and
the Ld-specific antibody 64-3-7, the Kb-binding antibody 25-D1.16

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
binds H-2b spleen cells with a biphasic profile, preferentially stain-
ing B lymphocytes (Fig. 6C). However, Fig. 6(D) shows that Kb ex-
pression is equal in both T and B lymphocytes as indicated by
staining with the Kb-specific antibody Y-3. Together these data re-
veal a consistent pattern of class I-related epitopes preferentially
detected on B cells.
Specific binding of antibody 25-D1.16 to Kb class I
molecules expressing the peptide SIINFEKL is enhanced by
interaction with FcR
The antibody 25-D1.16 is one of the few antibodies that differentially
recognizes class I molecules when loaded with a specific peptide. Al-
though the antibody binds weakly to Kb molecules that are folded
with a wide spectrum of peptides, it binds strongly to Kb molecules
loaded with the peptide SIINFEKL (4). Accordingly, this antibody has
Fig.5. Antibody interactions with the Fc receptor require the presence of the specific epitope on the cell surface. Splenocytes from C57BL/6 and Kbm3 mice were isolated and incubated with the irrelevant anti-Kb IgG2b antibody K10.56 or the anti-Kb IgG1 antibody B8-24–3 for 30 min beforestaining with L368-FITC, CD3-Red 613 and B220-PE. Histograms show analysis of PEπ/FITC π double-positive (B cells) or Red 613π/FITC π double-positive (T cells) live cells. Upper panel shows L368-FITC binding on B lymphocytes; lower panel shows staining of T lymphocytes. (A) L368 staining ofB6 splenocytes with and without preincubation with the anti-Kb IgG2b antibody K10.56. (B) L368 staining of B6 splenocytes with and without preincubationwith the anti-Kb IgG1 antibody B8-24–3. (C) L368 staining of lymphocytes from Kbm3 spleen with and without preincubation with anti-Kb IgG1 antibodyB8-24–3. Numbers show percent of inhibition of binding by blocking IgG on B cells as calculated by the following formula: [(MFI B cellsno blocker–MFI Bcellsblocker)/(MFI B cellsno blocker–MFI T cellsno blocker)] ¿ 100. MFI � mean fluorescence intensity. Negative staining shows autoflourescence of cells incubatedwith FACS medium only. Staining profiles are representative of two mice tested.
521Tissue Antigens 2002: 60: 515–525
been very useful in studies targeting peptide presentation at the cell
surface of cells coexpressing the Kb class I molecule and the chicken
ovalbumin-derived peptide, SIINFEKL. During the course of our
analysis, we found that the antibody specifically binds cells express-
ing chicken ovalbumin only when the Kb molecule is coexpressed
with the FcgIIR. Some EL-4 derivatives expressing a transfected oval-
bumin gene (EG7 sublines) express Fc receptor, while others do not
(Fig. 7A). The antibody 25-D1.16 preferentially binds EG7 cells that
express Fc receptor in comparison to EG7 cells that do not (Fig. 7B).
The binding of 25-D1.16 antibody to EG7-Fc receptor positive cells
can be blocked by preincubating the cells with anti-FcgR antibody
(Fig. 7C). When the EG7 Fc receptor negative cells were pulsed with
the OVA peptide, binding of 25-D1.16 substantially increased
(Fig. 7D). In addition to confirming our observations that low-affinity
antibody interactions with specific epitopes on the cell surface are sta-
bilized by interactions with the Fc receptor, these findings illustrate

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
that the ability of the 25-D1.16 antibody to detect the presence of the
SIINFEKL peptide derived from the intracellular processing of endo-
genously expressed ovalbumin is influenced by the concomitant ex-
pression of FcgIIR.
Discussion
The existence of isoforms of the Db class I molecule that are ex-
pressed primarily by B cells has been previously reported (1). The
binding of two monoclonal anti-Db antibodies, H141-31 and B22-
249, to T and B lymphocytes from mouse spleen were compared.
The H141-31 antibody bound to B lymphocytes preferentially while
the B22-249 antibody bound to both B and T cells equivalently. The
Fig.6. Detection of a B-cell associated epitope by other major histocompatibility complex class I-specific monoclonal antibodies. Spleno-cytes from Balb/c (H-2d) and C57BL/6 mice were isolated and analyzed for binding of MHC class I, CD3-Red 613 and B220-PE. Histograms show analysis ofFITCπ/PE π double-positive (B cells) and FITCπ/Red 613 π (T cells) live cells. (A) Staining of Balb/c splenocytes with anti-Ld alt 64-3-7. (B) Staining ofBalb/c splenocytes with anti-Ld antibody 66-3-5. (C) Profile of B6 splenocytes stained with the anti-Kb/OVA antibody 25-D1.16. (D)Profile of B6 splenocytesstained with the anti-Kb antibody Y-3.
522 Tissue Antigens 2002: 60: 515–525
authors reported that the H141-31 antibody bound three- to four-
fold less to B cells than the B22-249 antibody. It was concluded that
B cells express a heterogeneous class I population with different
glycosylation patterns. Similarly, the distinct binding pattern we
observed using the L368 antibody led us to the hypothesis that B
lymphocytes might express an alternate isoform of the Db molecule
that is not expressed by T lymphocytes. Subsequently, we found
that antibodies to other class I molecules, including Ld and Kb, also
preferentially binding to B cells isolated from the spleen. In this
report we have shown that the antigen-specific preferential binding
of antibody to B cells is caused by a secondary interaction of the
bound antibodies with Fc receptors expressed on B cells. Conse-
quently, these binding patterns are not evidence for differential ex-
pression of class I isoforms on B cells.

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
The mAb 64-3-7 has been a useful tool in the study of class I
intracellular processing pathways. It binds to the Ld unfolded intra-
cellular precursor termed Ldalt (3). The antibody is also reactive
to a subset of Ld molecules that are expressed on the cell surface
independent of peptide and b2-microglobulin (3). Staining of BALB/
c splenocytes with 64-3-7 shows a biphasic profile similar to that
observed using L368, with higher binding to B lymphocytes. Reac-
Fig.7. Specific binding of antibody 25-D1.16 to Kb class I molecules expressing the SIINFEKL peptide is enhanced by interaction withthe FcR when peptide density is low. Two different EG7 sublines expressing the OVA protein, EG7M and EG7R, were analyzed for the expression ofthe FcgIII/IIR and binding of the anti-Kb/OVA antibody 25-D1.16. (A) Analysis of EG7 sublines for the expression of the Fcg receptor. (B) EG7M and EG7Rcells were analyzed for binding of the anti-Kb OVA antibody 25-D1.16. (C) The EG7 cell line expressing the Fcg receptor was preincubated with anti-Fcreceptor antibody 2.4G2 before staining with 25-D1.16-SA-PE. (D) EG7 cells negative for the expression of the FcR were pulsed with the SIINFEKL (fromOVA) or SIYRYYGL irrelevant peptide and stained with 25-D1.16. Irrelevant isotype control antibody did not stain the cell lines shown. The fluorescentprofile of cells stained with isotype control antibodies was equivalent to the gray profile shown in panel C.
523Tissue Antigens 2002: 60: 515–525
tivity of 64-3-7 can be reduced to T-cell levels by blocking with
2.4G2 (data not shown).
The monoclonal antibody 25-D1.16 was obtained from mice im-
munized with TAP-2-deficient RMA-S (H-2b) cells loaded with the
SIINFEKL peptide from chicken ovalbumin (4). This antibody has
proven to be very useful in studying antigen processing and presen-
tation as well as identification of antigen-presenting cells in situ. We

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
found that 25-D1.16 binds normal C57BL/6 splenocytes (in the ab-
sence of the SIINFEKL peptide), preferentially staining B lympho-
cytes. As previously reported, 25-D1.16 can stain splenocytes in the
absence of the peptide SIINFEKL with very low intensity, but the
interaction requires Kb expression (4). We found that the interaction
of 25-D1.16 with Kb-SIINFEKL on EG7 cells is completely blocked
by the FcgRII/III antibody 2.4G2. Furthermore, 25-D1.16 does not
bind to EG7 OVA-expressing sublines that do not express the Fc
receptor. However, when cells are pulsed with excess SIINFEKL
peptide, 25-D1.16 stains EG7 FcgR-negative cells with high inten-
sity. This finding demonstrates that binding of 25-D1.16 is depend-
ent on the presence of FcgRII/III when Kb-SIINFEKL complexes
available on the surface are not abundant. Background staining of
Kb-expressing splenocytes in the absence of SIINFEKL may also be
the result of interactions of the antibody with the Kb molecule and
the FcgRII on B cells.
The affinity of 25-D1.16 for Kb-SIINFEKL has a Kd of approxi-
mately 7 ¿ 10ª8 M (4). This is considered low for antibody affinity
interactions. The anti-Db antibody H141-31 was also found to bind
with low affinity to B cells and to bind to a lower number of Db
molecules (1). Because 64-3-7 and L368 bind with low intensity to
mouse splenocytes and their binding is enhanced by interactions
with FcR, we suspect that they too are low-affinity reagents. What
is common among this set of antibodies is that they have led investi-
gators astray by displaying apparent cell-type binding specificity.
All these antibodies show increased binding to B lymphocytes as
compared with T lymphocytes. Here we show that the increased
affinity of these class I antibodies for B cells results from interac-
References
1. 5. 9.O’Neill HC. Identification of class I H-2Db Demant P, Ivanyi D, Oudshoorn-Snoek M, Lynch RG, Mathur A, Waldschmidt TJ,molecules primarily expressed by B lympho- Calafat J, Roos MH. Molecular heterogeneity Sandor M, Mueller A, Johanns P. Immuno-cytes in murine spleen. Immunol Cell Biol of H-2 antigens. Immunol Rev 1981: 60: 5– globulin (Fc) receptors on murine T- and B-1991: 69: 95–102. 22. lymphocytes: Investigations using tumor
2. 6. models. Mol Immunol 1988: 25: 1151–7.Lampson LA, Fisher CA, Whelan JP. Strik- Ivanyi D, Demant P. One (H-2D2b) of theing paucity of HLA-A, B, C and beta 2- three Db region-controlled molecules (H- 10. Lynch RG, Sandor M, Waldschmidt TJ et al.microglobulin on human neuroblastoma cell 2D1b, H-2D2b, H-2Lb) is not detected in Lymphocyte Fc receptors: expression,lines. J Immunol 1983: 130: 2471–8. bm13 mutant. J Immunol 1983: 131: 1080– regulation and function. Mol Immunol 1990:
3. 4.Lie WR, Myers NB, Connolly JM, Gorka J, 27: 1167–79.Lee DR, Hansen TH. The specific binding of 7. Ivanyi D, Snoek M, Demant P. H-2L: demon- 11. Sandor M, Lynch RG. Lymphocyte Fc recep-peptide ligand to Ld class I major histocom- stration of four new allelic products and in- tors. The special case of T cells. Immunolpatibility complex molecules determines dependence of H-2D and H-2L molecules. Today 1993: 14: 227–31.their antigenic structure. J Exp Med 1991: Tissue Antigens 1979: 14: 233–50. 12. Pullen JK, Hunt HD, Horton RM, Pease LR.173: 449–59. 8. Lampson LA, Levy R. Two populations of The functional significance of two amino
4. Ia-like molecules on a human B cell line. JPorgador A, Yewdell JW, Deng Y, Bennink acid polymorphisms in the antigen-present-JR, Germain RN. Localization, quantitation, Immunol 1980: 125: 293–9. ing domain of class I MHC molecules. Mol-and in situ detection of specific peptide- ecular dissection of Kbm3. J Immunol 1989:MHC class I complexes using a monoclonal 143: 1674–9.antibody. Immunity 1997: 6: 715–26.
524 Tissue Antigens 2002: 60: 515–525
tions with the FcgRIIB, which is expressed on resting B cells but
not resting T cells (9–11).
We find that the interactions of IgG1 antibody with the FcgRIIB
is substantially enhanced by the presence of the specific epitope on
the surface of the cell, providing the antibody-binding profile with
an element of epitope-specificity. This underscores the significance
of the dual interaction of a low-affinity antibody molecule with the
Fab portion binding with the specific class I epitope and the Fc
portion of the antibody binding to the FcgRIIB in close proximity
on the surface of the B cell. Our results support old studies that
claimed that Fc receptors, B- and T-cell receptors, CD-2, and class I
molecules are arranged on the cell membrane such that antibodies
bound to these targets can interact with Fc receptors on the same
cells (13–17). We propose that whenever there is a low density of
epitopes on the surface, low-affinity antibodies stabilize their inter-
action with targeted surface molecules by binding to the Fc receptor
through their Fc region. This type of interaction would not occur
when antibody affinity for a surface antigen is high or in some
cases when epitope density is plentiful. Previous studies cotargeting
B and NK receptors along with Fc receptors indicate that juxtapo-
sition of Fc receptor with the specifically bound ligands can have
profound functional consequences (15,16). Cross-linking MHC mol-
ecules with Fc receptors using this approach can also induce intra-
cellular signals (17). Consequently, the interaction of surface-bound
antibody with Fc receptors on the same cell can have widespread
implications in studies of antibody-binding specificity, as well as in
studies using antibody to manipulate cell physiology.

Mendez-Fernandez et al : Abs bind class I and FcR simultaneously
13. 14. 16.Smith JD, Lie WR, Gorka J, Kindle CS, My- Tyan ML. Evidence for clustering of H-2K, Anasetti C, Martin PJ, June CH et al. Induc-ers NB, Hansen TH. Disparate interaction H-2D, and the Fc receptor on the membranes tion of calcium flux and enhancement ofof peptide ligand with nascent versus ma- of B cells. Proc So Exp Biol Medical 1976: cytolytic activity in natural killer cells byture class I major histocompatibility com- 151: 526–31. cross-linking of the sheep erythrocyte bind-plex molecules. Comparisons of peptide 15. Parker DC. Induction and suppression of ing protein (CD2) and the Fc-receptorbinding to alternative forms of Ld in cell polyclonal antibody responses by anti-Ig re- (CD16). J Immunol 1987: 139: 1772–9.lysates and the cell surface. J Exp Med 1992: agents and antigen-nonspecific helper fac- 17. Benichou G, Voisin GA. Antibody bipolar175: 191–202. tors: a comparison of the effects of anti-fab, bridging: Isotype-dependent signals given
anti-IgM, and anti IgD on murine B cells. to guinea pig alveolar macrophages by anti-Immunol Rev 1980: 52: 115–39. MHC alloantibodies. Cell Immunol 1987:
106: 304–17.
525Tissue Antigens 2002: 60: 515–525





























