Embed Size (px)
Citation preview

Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372www.elsevier.nl/locate/palaeo
Early Cretaceous calcareous nannofossils from high latitudes:implications for palaeobiogeography and palaeoclimate
Jorg Mutterlose *, Kai KesselsInstitut fur Geologie, Mineralogie und Geophysik, Ruhr-Universitat Bochum, Universitatsstr. 150, 44801 Bochum, Germany
Received 5 August 1999; received in revised form 26 January 2000; accepted for publication 2 March 2000
Abstract
In order to better understand the palaeoceanography and palaeoclimate of the Early Cretaceous, nannofossil datahave been obtained from high-latitudinal sites from offshore mid-Norway and from the Barents Sea. No consistentnannofossil data are yet available for Early Cretaceous high latitudes, which should more clearly reflect possiblepalaeoclimatic changes, as recorded in fluctuations of diversity and abundance. Existing data from France, Italy,Romania, Poland and NW Europe (Germany, North Sea, England) have been complemented by material from higherlatitudes. In order to record the Arctic–Boreal nannofossil assemblages, 400 samples from the Norwegian Shelf andthe Barents Sea have been analysed. Sixty samples, covering the Berriasian–Barremian interval, yielded calcareousnannofossils. These are derived from cores between 63 and 73°N of present-day latitudes. The nannofossil assemblagesrecorded are generally of a low diversity and characterized by abundant Watznaueria barnesae and Crucibiscutumsalebrosum, whereas Biscutum constans is less common. C. salebrosum, rare or absent at low latitudes, is extremelycommon in the Norwegian samples. It shows a bipolar distribution especially during Valanginian–Hauterivian times.In the early Valanginian, a distinctive latitudinal gradient in the abundance of C. salebrosum, impoverished in lowlatitudes and abundant in high latitudes, possibly reflects latitudinal differences in temperature. Restricted palaeoceano-graphic settings, caused by a sea-level lowstand, may have amplified these thermal gradients. Palaeoclimatically thisimplies the existence of climatic belts throughout parts of the Early Cretaceous (early Valanginian, Hauterivian),resulting from considerable temperature gradients from north to south. The palaeobiogeographic patterns discussedresulted in the formation of distinctive latitudinally bound nannofossil zones, which, to a certain extent, are similarto those of modern oceans. The palaeobiogeographic patterns described support the idea of an ice-house phase forthe early Valanginian and disagree with the suggestion of a globally warm equable climate during that time. © 2000Elsevier Science B.V. All rights reserved.
Keywords: C. salebrosum; calcareous nannofossils; Early Cretaceous; high latitudes; palaeobiogeography; palaeoclimate
1. Introduction Indo-Pacific area. They show a distinctivepalaeobiogeographic distribution pattern forthe Berriasian–Barremian and Aptian–AlbianEarly Cretaceous calcareous nannofossil assem-intervals. The Berriasian–Barremian period isblages show significant differences in abundancecharacterized by provinciality on the generic level.and diversity between the Boreal, Tethys andIt is possible to differentiate between a Boreal andTethyan Realm. Different and independent* Corresponding author. Fax: +49-234-7094571.biostratigraphic schemes were developed for theE-mail address: [email protected]
(J. Mutterlose) two areas using Tethyan and Boreal markers.
0031-0182/00/$ - see front matter © 2000 Elsevier Science B.V. All rights reserved.PII: S0031-0182 ( 00 ) 00082-1

348 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Endemic floras and faunas appear to develop Norwegian Shelf and the Barents Sea have beenanalysed. In addition, ODP samples from lowcoincidentally with periods of sea-level lowstand.
This is best demonstrated in the Berriasian and palaeolatitudes in the northern hemisphere (ODPSite 603) were studied for a direct comparisonBarremian, when the sea-level was lowest.
Conversely, periods of transgression and high- with the high-latitudinal nannofossil assemblages.The following problems are addressed in this study:stands of sea-level led to an exchange of floras
between the Tethys and Boreal and vice versa $ Quantitative and qualitative composition of cal-careous nannofossils from the Norwegian Shelf(Mutterlose, 1992). In addition to these
migrations, the Boreal Realm shows a general and the Barents Sea and a palaeoecologicalinterpretation of the assemblages.decrease of species towards higher latitudes.
The Tethyan Realm is subdivided into a $ Influx and timing of the immigration of Tethyantaxa to high palaeolatitudes. The impact ofMediterranean Province and an Indo-Pacific
Province. The Mediterranean Province is domi- third order sea-level fluctuations (0.5–5 Ma) onthe composition of nannofossils at highnated by cosmopolitan and Tethyan flora, while
the latter is absent from the Indo-Pacific Province. latitudes.$ Comparison of high-latitudinal nannofossilThe Aptian–Albian period was marked by a floral
(and faunal ) change, causing major shifts of biota assemblages with coeval low-latitudinalassemblages.worldwide. The differences between the Boreal and
Tethyan Realm vanished, as well as the differences $ Reconstruction of Early Cretaceous latitudinalnannofossil gradients in order to decipher thebetween the two provinces within the Tethyan
Realm. This change predates the early Aptian controlling mechanisms. The palaeoclimate andpalaeogeography in particular must be viewedoceanic anoxic subevent (OAE 1a; Jenkyns, 1980;
Arthur et al., 1990) and the associated positive as possible causes for the palaeobiogeographicdifferentiation.carbon-isotope excursion. Such a world-wide shift
of marine biota requires palaeoceanographicchanges in the mid-Cretaceous oceans on aglobal scale. 2. Geological background
In addition to this pattern, it is possible torecognize two high-latitudinal bipolar nannofossil 2.1. Palaeobiogeography and sea-levelbelts: one in the Indo-Pacific and the other in theBoreal Realm during the Valanginian–Hauterivian The interval under discussion (Berriasian–
Albian) is marked by a pronounced change of theand Albian–Cenomanian intervals. These high-latitudinal belts are characterized by certain species palaeogeography and sea-level. Latest Jurassic–
earliest Cretaceous times (Tithonian–Berriasian)absent or extremely rare in low-latitudinal sites(Mutterlose, 1992). experienced a distinctive sea-level lowstand that
was initiated by low spreading rates, resulting inThe information and data gathered so far fromthe Boreal Realm are mainly based on material large semi-restricted epicontinental seas (Haq
et al., 1988; Ziegler, 1988; Hardenbol et al., 1998);from NW Germany, the North Sea area andEngland: none of these areas reflects high palaeo- only the central part of the Atlantic was already
open. The latest Jurassic and earliest Cretaceouslatitudinal regions (Mutterlose, 1991). Fewdetailed data are available for high latitudes, which were periods not only of sea-level lowstand but
also of widespread deposition of non marine sedi-are generally characterized by assemblages of lowdiversity and abundance (Smelror et al., 1998). ments (e.g. the ‘Wealden facies’ in S England, N
France and NW Germany). Siliciclastic sedimenta-Data available from France, Italy, Romania,Poland and NW Europe (Germany, North Sea, tion prevailed during this interval, coincident along
with a high rate of endemic marine biota (e.g.England) need to be complemented by materialfrom high palaeolatitudes. To record the Arctic– ammonites; Casey, 1973; Hoedemaeker, 1990;
Rawson, 1994). Semi-restricted epicontinental seasBoreal nannofossil assemblages, samples from the

349J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
favoured the in-situ evolution of marine biota, tions, which prevailed during the Barremian, wereincluding planktonic, nektonic and benthic taxa. replaced by more open oceanic conditions. A gene-A relationship between sea-level rise and increasing ral increase in sea-level occurred throughout thecosmopolitanism of marine floras and faunas has Aptian and Albian and reached its maximumbeen described by various authors (e.g. Hallam, during the late Cenomanian–early Turonian (e.g.1994). Haq et al., 1988; Larson, 1991a,b; Hallam, 1992).
The subsequent Valanginian and Hauterivian Submarine volcanic eruptions on a large scale tookstages were characterized by widespread transgres- place in the Pacific in mid-Cretaceous times duringsions resulting in more open oceanic conditions the time interval 125–80 Ma (Larson, 1991a;(Rawson and Riley, 1982; Haq et al., 1988; Ziegler, Winterer, 1991). This proposed mantle plume in1988). Siliciclastic sedimentation prevailed in the the Pacific, which has been questioned by Hellermid-high-latitudes, while the Tethys was charac- et al. (1996), may have caused an exponentialterized by carbonate rich-pelagic sedimentation. increase in the production of oceanic crust. BothThe non-marine Wealden basins of the preceding normal sea-floor spreading and mantle plumingTithonian–Berriasian interval were then partly caused the major Aptian–Cenomanian rise in sea-flooded. Sea-floor spreading remained fairly stable level. Spreading rates of 2.3 cm/year (N Atlantic),from the latest Jurassic (1.8 cm/year, N Atlantic; 2.4 cm/year (S Atlantic), 3.0 cm/year (Australia),1.3 cm/year, Australia) to Hauterivian and 2.0 cm/year (Africa) have been suggested for(1.2 cm/year, N Atlantic; 1.5 cm/year, Australia; the Aptian (Kominz, 1984: table 1). These oceanic2.5 cm/year, Afrika; Kominz, 1984: table 1). This changes possibly caused a major shift of marineallowed an increasing exchange of marine floras floras and faunas, which occurred simultaneouslyand faunas from the Tethys to the Boreal Realm in the Tethyan and Boreal Realms and in the Indo-and vice versa; both realms still remained quite Pacific Province. Various floral and faunal ele-distinct from one another. This is best demon- ments became extinct in the Aptian in both thestrated for the Valanginian when Boreal ammo-
Tethyan and Boreal Realms, whereas new generanites and nannofossils were found in the Tethys
and species also evolved (Mutterlose, 1998).and vice versa (Kemper et al., 1981; Mutterlose,Marked changes in the diversification rate were1991). These patterns of floral and faunal homoge-observed for organisms with a different mode ofnization can be observed in all groups of organismslife (plankton, nekton, benthos; Mutterlose, 1998).(plankton, nekton, benthos).
During latest Jurassic to Early CretaceousSignificant palaeogeographic and palaeoceano-times, the studied area (Norwegian Shelf, Barentsgraphic changes occurred at the Hauterivian/Sea) is characterized by relatively low hinterlandBarremian boundary; the overall regressive naturerelief. Fine-grained sediments were deposited inof the Barremian is often quoted (e.g. Ruffell,the basins off mid-Norway and in the Barents Sea,1991). Slightly lower sea-floor spreading rates havewhile the structural highs are characterized bybeen calculated for the Barremian (0.7 cm/year, Ncondensed carbonates. A widespread hiatus in theAtlantic; 1.5 cm/year, Australia; 2.9 cm/year, Searliest Cretaceous time is thought to be due toAtlantic; Kominz, 1984: table 1). Lacustrine sedi-differential subsidence, with local uplifts and ero-mentation prevailed in Poland, and the North Seasion. The rapid global rise in sea-level duringbecame a marginal sea with the deposition ofAptian through Cenomanian/Turonian times ledorganic carbon-rich sediments. These relativeto widespread transgressions along the margin,changes are reflected in distinct biogeographicaugmented by vigorous local subsidence andrealms as restricted epicontinental seas favour thedifferential block-faulting in Albian throughevolution of endemic taxa. This is recorded in bothConiacian times. During the latter period, thethe Tethyan and Boreal Realms.palaeowater depth in many sub-basins offshoreThe Barremian/Aptian boundary interval isNorway increased to bathyal (Gradstein et al.,marked by a major change of the palaeoceano-
graphic setting. Restricted oceanographic condi- 1999).

350 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
2.2. Palaeoclimate gesting Early Cretaceous icehouse interludes forthe late Early Cretaceous.
The current ideas concerning palaeoclimate and $ Isotope studies on belemnites from the Loweroceanic circulation patterns for the Cretaceous Cretaceous of Svalbard (Ditchfield, 1997).oceans are contradictory. Some authors (e.g. Podlaha et al. (1998), who studied the isotopeFrakes, 1979; Hallam, 1981, 1985) consider the signature of belemnites from NW Germany andCretaceous to be a period of great warmth over Speeton (NE England), propose warm marinethe globe. Tropical–subtropical conditions pre- palaeotemperatures (20°C) for the Berriasian, availed to at least 45°N and possibly to 70°S, with decrease to 15°C in Valanginian and coldestwarm to cool–temperate climates extending to the palaeotemperatures (10°C) in early Hauterivianpoles (Frakes, 1979; Hallam, 1981). Mean annual times. These data are confirmed by Price et al.temperatures were siginificantly higher and latitu- (2000). Hallam et al. (1991) and Ruffell anddinal gradients only about half those of today. Batten (1990) suggest a Barremian–Aptian aridd18O palaeotemperature data from the low-latitude phase in northwest Europe, resulting in typicaloceans (Douglas and Woodruff, 1981) indicate clay mineralogical assemblages.that the Early Cretaceous was only 2–4°C cooler Many geologists argue for a climatic changethan the Cenomanian. Based on a northward shift from an icehouse world in Early Cretaceous timeof Eurasian floras through time, Vakhrameev (Berriasian–Barremian) to a greenhouse world in(1964, 1991) concluded that there was a global mid-Cretaceous time (Aptian–Turonian; e.g.warming from the Early Jurassic to the mid- Larson, 1991a,b). This period of global change inCretaceous. The Cretaceous has been generally the mid-Cretaceous may be viewed as a result ofviewed as a period free of ice-caps (Hallam, 1985; increased CO2 production caused by increasedFrancis and Frakes, 1993), and cooling has been submarine volcanism in the Pacific region.only postulated for Maastrichtian times. Extensive outgassing of CO2 would have caused a
A more varied climate history for theclimatic shift towards an Aptian–Turonian green-
Cretaceous has been suggested in more recenthouse world. This induces global warming, astudies (e.g. Kemper, 1987; Weissert and Lini,decrease of temperature gradients between low and1991; Stoll and Schrag, 1996). For the Earlyhigh latitudes, high humidity and sluggish oceanicCretaceous, Kemper (1987) proposed ice-ages forcirculation. These global processes influence boththe Valanginian and Aptian/Albian that lastedsedimentary and biotic patterns. This mid-from a few thousand to 2 million years. FrakesCretaceous scenario resulted in accelerated carbonand Francis (1988), Frakes et al. (1992) andcycling, increased availibility of nutrients and con-Ditchfield (1997) postulate at least seasonally coldsequently oceanic anoxic conditions. Widespreadocean temperatures and limited polar ice caps fororganic carbon-rich sediments associated withthe Early Cretaceous. In particular, during partsocean anoxic events (OAEs) were deposited simul-of the Early Cretaceous (Berriasian, Valanginian),taneously in different parts of the world (Schlangerice-house conditions, or at least cool climates, haveand Jenkyns, 1976; Jenkyns, 1980; Arthur et al.,been proposed. Early Cretaceous glaciations are1990). The early Aptian OAE 1a represents thesuggested by:earliest of the mid-Cretaceous anoxic events and$ The findings of glendonites in the Valanginianis considered to mark the onset of the greenhouseof the Sverdrup Basin from Arctic Canadaworld. Anoxic events also occur in the early Albian( Kemper, 1987).(OAE 1b), late Albian (OAE 1c, OAE 1d) and$ The changing composition of marine floras andclose to the Cenomanian/Turonian boundaryfaunas (Kemper, 1987).(OAE 2), causing the mid-Cretaceous to become$ The occurrence of ice-rafted deposits in Siberia,a ‘black shale world’. These environmental changesAustralia and Spitzbergen (Frakes andmight help to explain global turnovers in marineFrancis, 1988).
$ Isotope studies (Weissert and Lini, 1991) sug- biota during the Early Cretaceous.

351J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
3. Material and methods are based on personal core descriptions madein 1994.
A total of 400 samples of Early Cretaceous age4.1. Core 6307/07-U-02were studied with respect to their calcareous nan-
nofossil contents. Simple smear-slide preparationsGrid reference: 63° 27∞54.35◊ N, 07° 14∞44.26◊were examined under the light microscope using a
E; off mid-Norway (off the Smøla island); southernmagnification of 1500×. The abundance of calcar-part of the Hitra Basin. Twenty-seven of 66eous nannofossils in the samples studied is vari-samples yielded calcareous nannofossils.able. Based on simple estimates, calcareousReference: Skarbø et al. (1988). Palaeolatitude:nannofossils may constitute anything from 0 to53°N. Subsequently, the core will be referred to as20% of the whole rock. The abundance for each6307/07.sample was determined by counting at least 300
The 190.1 m thick sequence consists ofspecimens or all specimens in at least 200 fieldsQuaternary sediments (0–13.6 m) and of lowerof view.Valanginian to upper Volgian/upper Kimmer-Preservation is indicated in the following way:idgian claystones and marls (13.6–190.1 m). TheE1 (slightly etched), E2 (moderately etched), andage assignments, which are based on palyno-E3 (heavily etched). Although it occurs, over-morphs and foraminifera (Skarbø et al., 1988),growth was not reported. Bibliographic referenceshave not been subsequently revised. From top tofor the determined taxa encountered are given inbottom, the following lithological units can bePerch-Nielsen (1985) and Bown (1998).differentiated:Carbonate measurements were made using
13.6–36.4 m (22.9 m); red and grey marls, Langeatomic absorption spectrometry (AAS).Fm.; CaCO3 content 1.1–39%. 13.6–24.6 mNannofossil slides are housed at the Museum of(11.0 m); early Valanginian age; samples 1–16.Natural History and Archaeology, NTNU in24.6–36.4 m (11.8 m); earliest Valanginian–latestTrondheim.Berriasian age; samples 17–34.36.4–104.8 m (68.4 m); dark claystones, SpekkFm.; late Berriasian–early Volgian age; samples
4. Localites and lithology 34–66.104.8–183.5 m (78.7 m); fine grained sandstones,
Sediments from six cores, drilled offshore mid- Rogn Fm.; early Volgian–late Kimmeridgian age.and north Norway, were situated between 53 and 183.–190.1 m (6.6 m); dark clays, Spekk Fm.;63°N during the earliest Cretaceous. These sec- Kimmeridgian age.tions, which encompass the Barremian–Volgianinterval, were studied with respect to their contentsin calcareous nannofossils. The studied cores were 4.2. Core 7320/03-U-01drilled in the course of the SINTEF PetroleumResearch (former IKU ) ‘Shallow Drilling Project’ Grid reference: 73° 51∞15.20◊N, 20° 55∞00.80◊E;between 1985 and 1991. In 1994, the cores were western part of the Barents Shelf, northernlogged lithologically and sampled for this study. Bjørnøya Basin. All 40 samples studied were
Fig. 1 shows the geographic position of the barren of calcareous nannofossils. References:cores studied. The stratigraphic range of the sec- Fjerdingstad et al. (1985), Arhus et al.tions and the main lithological subdivision encoun- (1990), Arhus (1991), Smelror et al. (1998).tered are given in Figs. 2 and 3. Fig. 3 lists the Palaeolatitude: 62°N. Subsequently, the core willamount of samples studied from each core. A total be referred to as 7320/03.of 400 samples have been studied, from which only A section of 36.2 m of Quaternary sediments60 samples turned out to be nannofossil bearing. (0–2.1 m) and of lower Barremian to upperThe lithological data of the nannofossil bearing Volgian claystones, limestones and siltstones (2.1–
36.2 m) was cored. Since the samples are barrenintervals, of the four cores considered in this study,
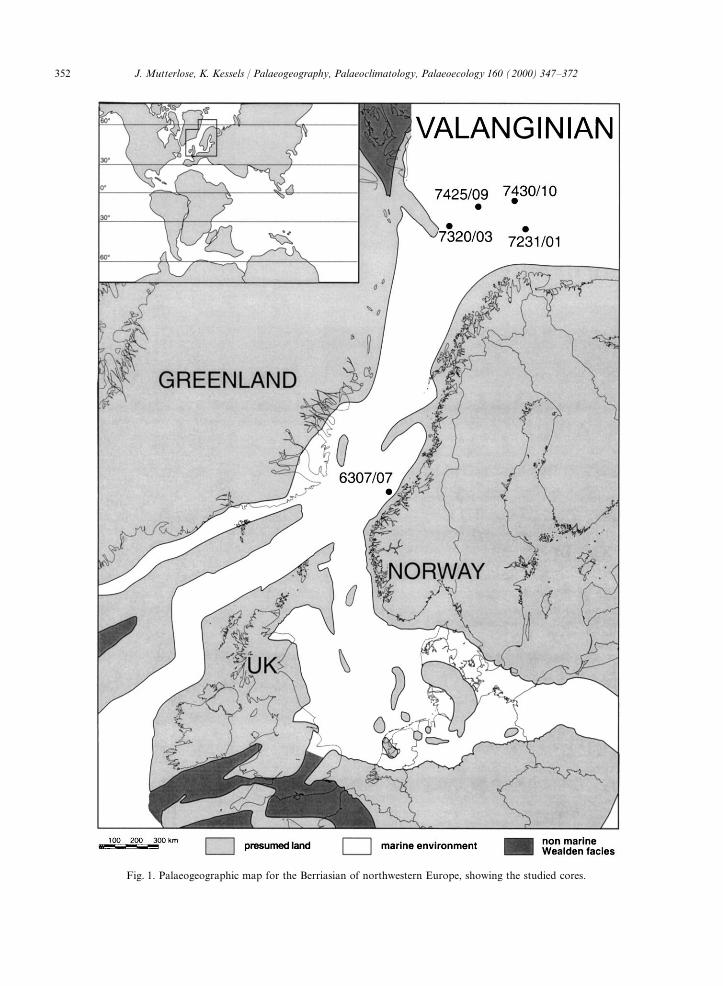
352 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 1. Palaeogeographic map for the Berriasian of northwestern Europe, showing the studied cores.

353J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 2. Stratigraphic ranges of the studied cores.
of calcareous nannofossils, the lithology of this Palaeolatitude: 63°N. Subsequently, the core willbe referred to as 7425/09.core is not discussed here any further.
This core recovered 64.4 m of Quaternary sedi-ments (0–49.9 m) and of lower Barremian to upper4.3. Core 7425/09-U-01Berriasian claystones and marls (49.9–64.4 m).The original age assignments (Fjerdingstad et al.,Grid reference: 74° 29∞25.30◊ N, 25° 46∞19.30◊
E; northwestern part of the Barents Shelf, west- 1985) have recently been revised by Smelror et al.(1998), using palynomophs and calcareous nanno-ern Bjarmeland Platform. Fifteen of 24 sam-
ples studied, yielded calcareous nannofossils. fossils. The latter scheme is followed here. Fromtop to bottom, the following units can beReferences: Fjerdingstad et al. (1985), Arhus et al.
(1990), Arhus (1991), Smelror et al. (1998). differentiated:
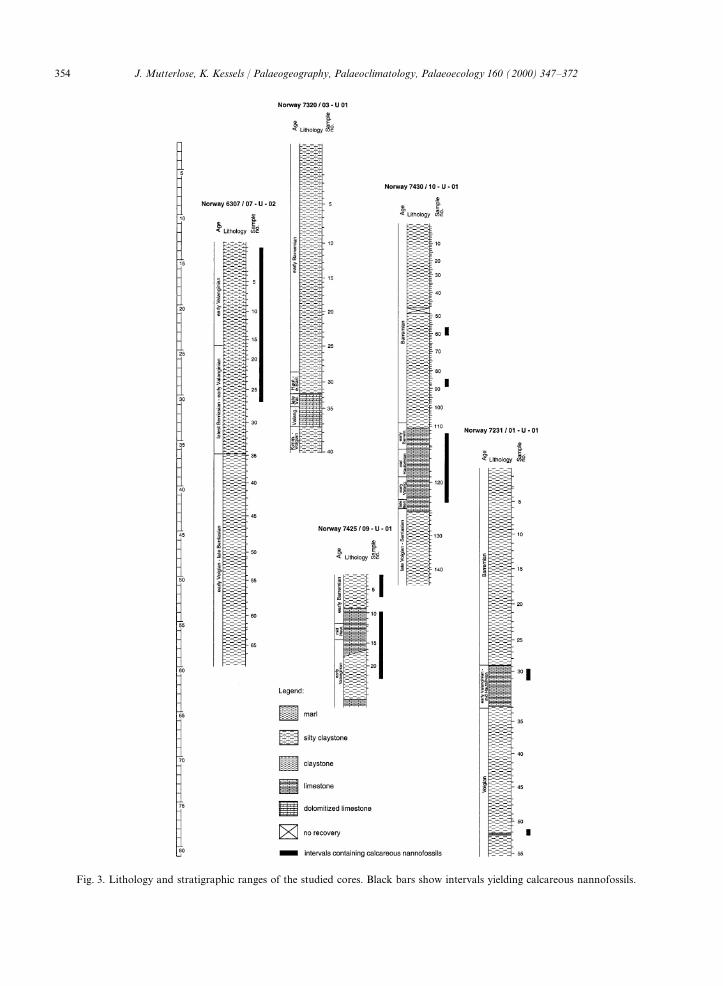
354 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 3. Lithology and stratigraphic ranges of the studied cores. Black bars show intervals yielding calcareous nannofossils.

355J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
49.9–53.5 m (3.6 m); grey claystones, Kolje Fm.; Basin. Only three of the 55 samples studied yieldedCaCO3 content 0–4%; early Barremian age; calcareous nannofossils. References: Arhus et al.samples 1–9. (1987), Arhus (1991), Smelror et al. (1998).53.5–58.8 m (5.3 m); marly limestones, Klippfisk Palaeolatitude: 62°N. Subsequently, the core willFm.; boundary against underlying unit gradual; be referred to as 7231/01.CaCO3 content 9.4–61%; early Barremian, mid-/ About 93 m of Quaternary sediments (0–early Hauterivian to early Valanginian age; 36.5 m) and of lower Barremian tosamples 10–18. Kimmeridgian/Oxfordian claystones and marls58.8–63.7 m (4.9 m); brown claystones, (36.5–93 m) were recovered. The age assignmentsHekkingen Fm.; CaCO3 content 1.7–25%; early used here are based on palynomorphs (Arhus,Valanginian age; samples 19–24. 1991). From top to bottom, the following units63.7–64.4 m (0.7 m); green limestones; earliest can be differentiated:Valanginian–latest Berriasian age; no samples. 36.5–59.8 m (23.3 m); dark grey claystones, Kolje
Fm.; Barremian age; samples 1–29.59.8–64.5 m (4.7 m); marly limestones, Klipp-
4.4. Core 7430/10-U-01 fisk Fm.; CaCO3 content 9.1–39.1%; mid-Hauterivian–early Valanginian age; samples 30–
Grid reference: 74° 12∞47.79◊ N, 30° 14∞44.22◊ 32.E; central part of the Barents Shelf, Bjarmeland 64.5–93 m (28.5 m); dark grey clays, HekkingenPlatform. Fifteen of the 165 samples studied Fm.; late Volgian–late Oxfordian age; samplesyielded calcareous nannofossils. References: Bugge 33–55.et al. (1988), Arhus (1991), Smelror et al. (1998).Palaeolatitude: 63°N. Subsequently, the core willbe referred to as 7430/10. 5. Results
Approximately 67.6 m of sediment were cored,consisting of Quaternary sediments (0–10.5 m) and
5.1. Diversity and preservationof upper Barremian to Kimmeridgian claystonesand marls (10.5–67.6 m). By using palynomophs
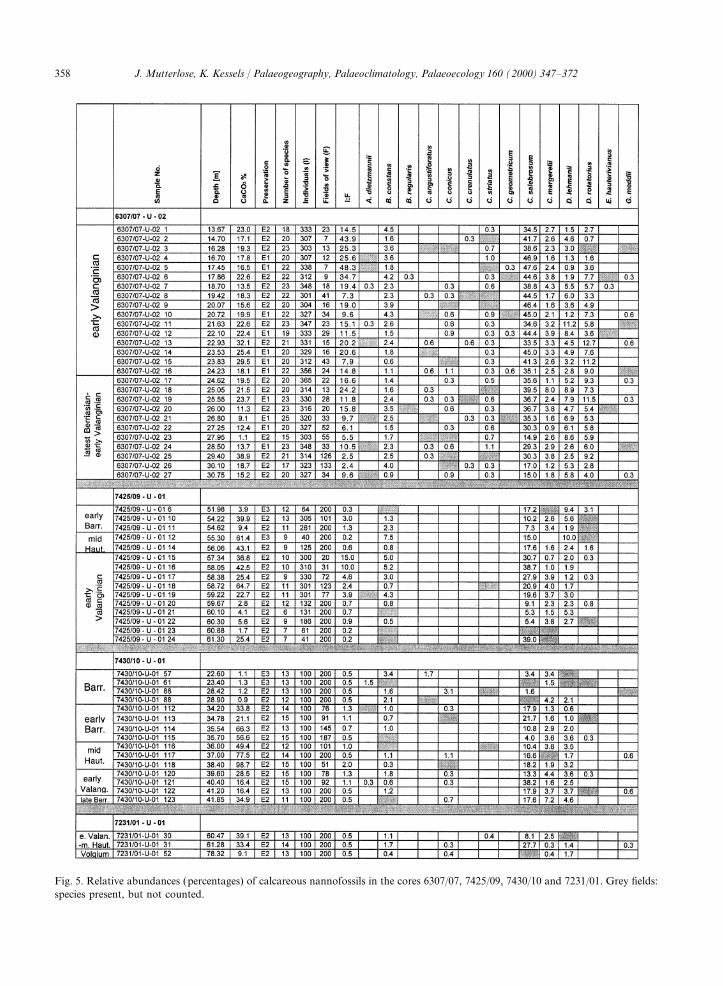
Figs. 4 and 5 include lists of relative abundancesand calcareous nannofossils, Smelror et al. (1998)of all samples with calcareous nannofossils. Of themodified the earlier age assignments of Bugge et al.40 species encountered, the most common include(1988). This modified zonation is used here. FromWatznaueria barnesae, Crucibiscutum salebrosum,top to bottom, the following units can beBiscutum constans and Rhagodiscus asper.differentiated:Together, they make up approximately 80–90% of10.5–33.7 m (23.2 m); dark grey claystones, Koljethe assemblages. Other common taxa includeFm.; CaCO3 content 1%; Barremian age; samplesCyclagelosphaera margerelii, Diazomatolithus leh-1–111.manii, Microstaurus chiastius, Rotelapillus laffittei,33.7–42.9 m (9.2 m); marly limestones, KlippfiskSollasites horticus, Vekshinella stradneri andFm.; CaCO3 content 16–99%; early Barremian,Zeugrhabdotus spp. The diversity (=number ofmid-Hauterivian – early Valanginian – latespecies) varies from 7 to 25 species/sample; 10–20Berriasian age; samples 112–125.species/sample are more often encountered.42.9–67.6 m (24.7 m); black shales, Hekkingen
The marly, carbonate-rich sediments of the ear-Fm.; late Berriasian–Kimmeridgian age; samplesliest Cretaceous (early Valanginian, mid-126–143.Hauterivian) have higher diversities than those ofthe Barremian. A comparison of the four nanno-fossil-bearing cores (Figs. 4 and 5) show a distinc-4.5. Core 7231/01-U-01tive decrease in diversity from south to north: 15–25 species were observed in the southernmost coreGrid reference: 72° 45∞12.45◊ N, 31° 07∞30.21◊
E; central part of the Barents Sea, Nordkapp 6307/07, seven to 13 species in core 7425/09, and
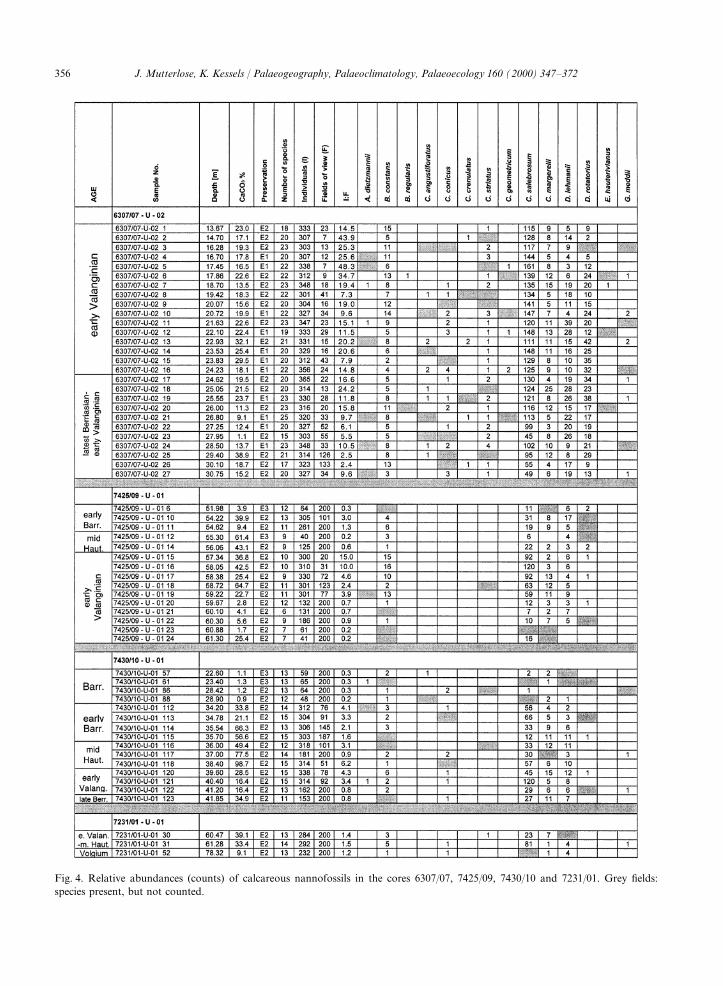
356 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 4. Relative abundances (counts) of calcareous nannofossils in the cores 6307/07, 7425/09, 7430/10 and 7231/01. Grey fields:species present, but not counted.
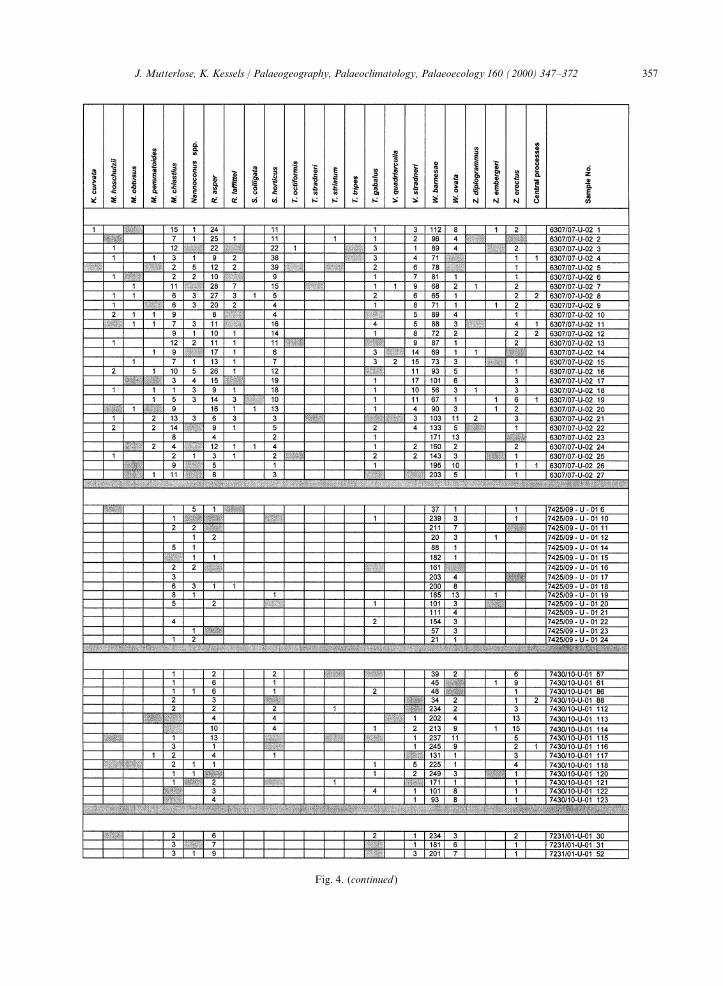
357J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 4. (continued )
358 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
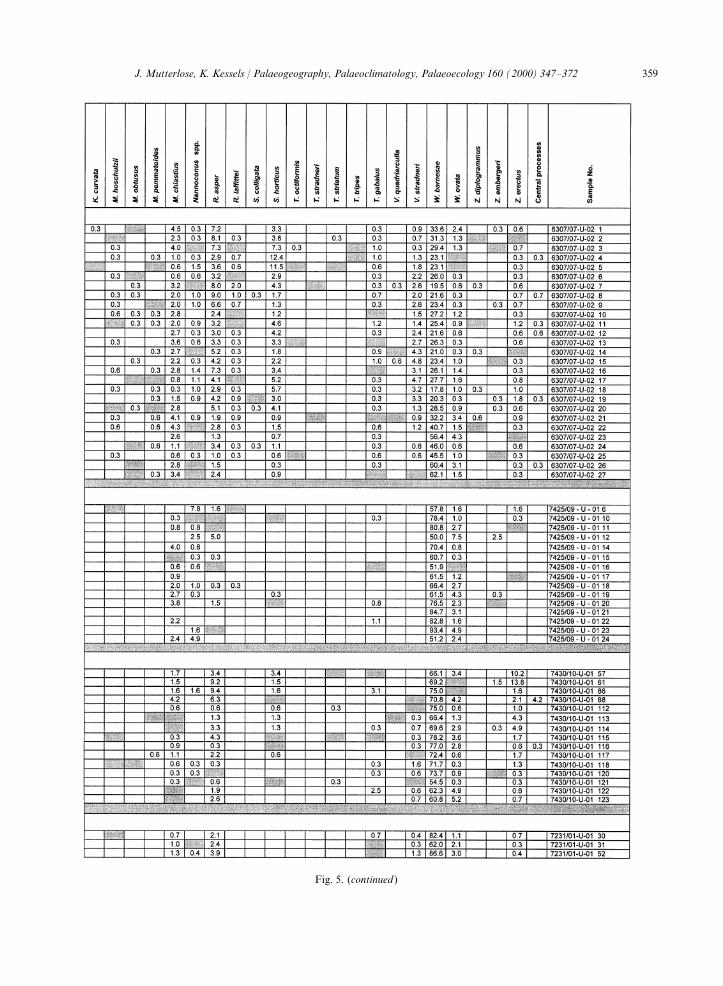
Fig. 5. Relative abundances (percentages) of calcareous nannofossils in the cores 6307/07, 7425/09, 7430/10 and 7231/01. Grey fields:species present, but not counted.
359J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 5. (continued )
360 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
11–15 species in the northernmost core 7430/10. fore, often interpreted as a measure of preservationof a given nannofossil assemblage (Roth andA comparison of abundance in individual time
slices also indicates that diversity is controlled by Bowdler, 1981; Roth and Krumbach, 1986).Assemblages containing more than 40% W.palaeogeography. Early Valanginian assemblages,
which were encountered in three cores, show a barnesae are thought to be heavily altered bydiagenesis (Roth and Bowdler, 1981). Otherdecrease in diversity from south to north.
The pattern described above may, however, also authors, however, consider W. barnesae to be aeurytopic species (Mutterlose, 1991), indicatingreflect different preservation modes and/or the
different number of samples studied for each core. assemblage alteration only after reaching percent-ages >70% (Williams and Bralower, 1995). It isFigs. 4 and 5 show a good preservation for the
southernmost core 6307/07, and moderate preser- remarkable that cores 7425/09 and 7430/10, situ-ated furthest north, yield rich assemblages of C.vation for the cores further north. Thus, a diage-
netic overprint cannot be ruled out. This is salebrosum. The presence of this small and delicate,dissolution-susceptible species indicates a prestinesupported by the fact that samples with a good
preservation show low percentages of W. barnesae signal. Though diagenetic overprint probablyinfluenced the composition of the assemblages in(core 6307/07), and those with a moderate to poor
preservation yield high percentages of W. barnesae the northernmost cores, the relevant groups ofcalcareous nannofossils, which are here under dis-(cores 7425/09, 7430/10, 7231/01). W. barnesae is
considered to be most resistant to diagenesis (Roth cussion, are still sufficiently well represented toallow a palaeoecological interpretation.and Bowdler, 1981). Mass abundances are, there-
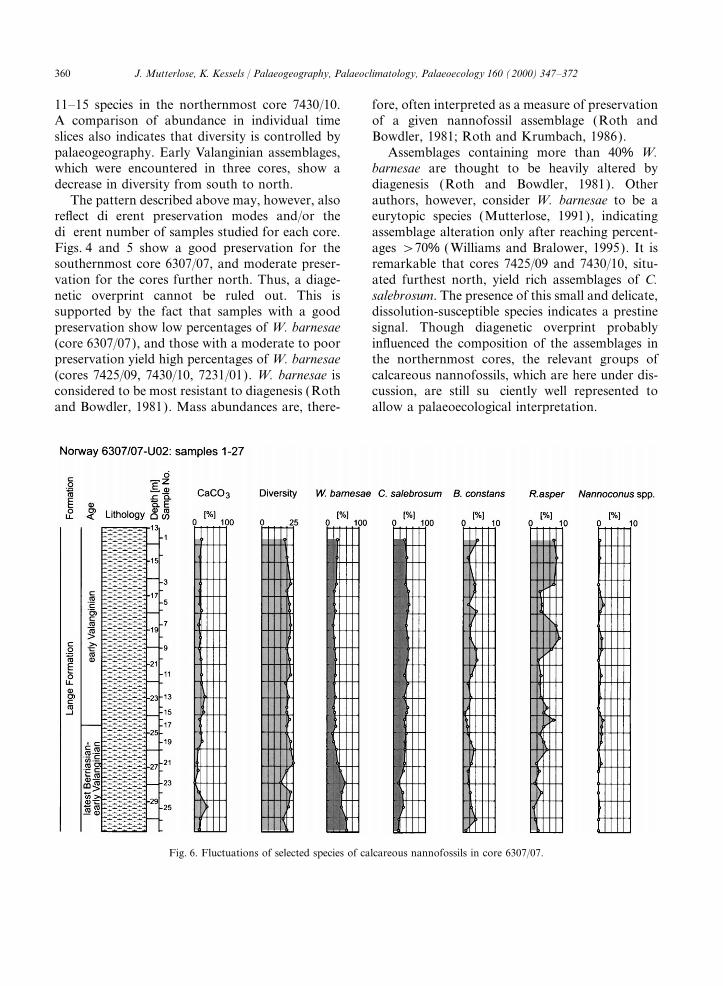
Fig. 6. Fluctuations of selected species of calcareous nannofossils in core 6307/07.
361J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
5.2. Abundance samples investigated. This species is less abundantin samples from the earliest Cretaceous of the low-latitudinal core 6307/07. In the lower ValanginianIt is difficult to make objective estimates of
abundances by using smear slides, due to the of core 6307/07, W. barnesae furnishes only 19–34% of the total abundance. Further north in corespreparation technique. Thus, the thickness of a
slide is a major factor influencing the abundance. 7425/09 and 7430/10, however, W. barnesae makesup more than 50–93 and 54–78% of the abun-Nevertheless, in Figs. 4 and 5, the columns I (=
individuals) and I: F (=individuals per field of dance. Vertically, the abundance of W. barnaesevaries in core 6307/07 from 18–62% in the upper-view) represent estimates of abundances. The data
obtained clearly support a significant north–south most Berriasian–lowermost Valanginian to 19–34% in the lower Valanginian. This pattern impliesgradient. Core 6307/07 quite far south yields sig-
nificantly more specimens than those cores fur- either a diagenetic overprint or an increase ofecologically robust taxa towards the north.ther north.
Selected common species were chosen from Biscutum constans, a small form of up to 4 mmlength, is constantly present with a relative abun-three cores (6307/07; 7425/09; 7430/10) for a more
detailed discussion (Figs. 6–8). These include W. dance of 1–8%. The abundance of B. constans isfairly stable over the latitudinal and stratigraphicbarnesae, B. constans, C. salebrosum, R. asper and
Nannoconus spp. interval studied, implying stable conditions for thisspecies over the studied area.W. barnesae shows high abundances in all
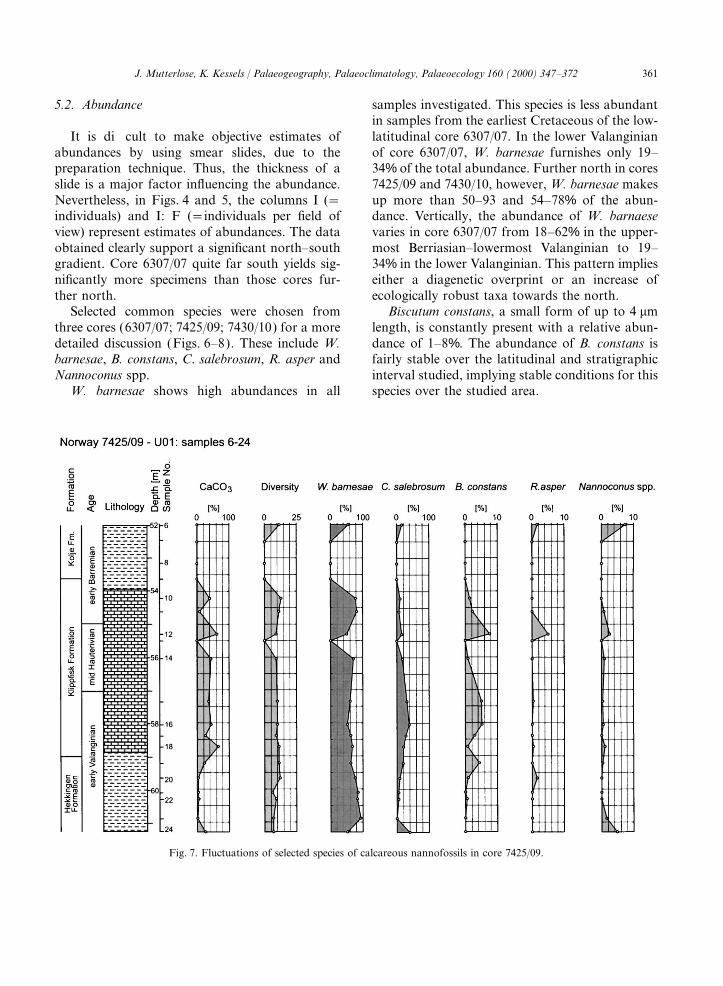
Fig. 7. Fluctuations of selected species of calcareous nannofossils in core 7425/09.

362 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig. 8. Fluctuations of selected species of calcareous nannofossils in core 7430/10.
C. salebrosum is most common in the low- Hauterivian sample of core 7425/09, whileNannoconus is rare in all samples of core 7430/10latitudinal site 6307/07 (15–48%) and less so in
the high-latitudinal sites 7425/09 (5–39%) and (0.3–1.6%).7430/10 (1–38%). The abundance of this speciesin core 6307/07 is rather stable, while siginificant 5.3. CaCO
3content and depositional environment
vertical fluctuations were observed in the high-latitudinal sites. The CaCO3 content varies considerably in the
cores studied from 1 to 98% (Figs. 4–8). The earlyRhagodiscus asper, a cosmopolitan species,shows a relatively low abundance of 1–9% in site Valanginian marls (=Lange Formation) of core
6307/07 vary from 1–39%, and CaCO3 values from6307/07. It is slightly less common in sites 7425/09(0.3–5%) and 7430/10 (0.3–9%). Vertical fluctua- 15 to 25% are most common. These data correlate
with well-preserved assemblages of a rather hightions in core 6307/07 are insignificant.Tethyan-derived Nannoconus spp. are usually diversity, with W. barnesae and C. salebrosum each
making up about 40% of the abundance. Therare. Nannoconids remain relatively constant incore 6307/07 (0.3–1.5%), whereas their abundance Lange Formation is considered to have been depos-
ited under open marine, aerobic conditionsvaries in core 7425/09 (0.3–8%). It is interestingto note that exceptionally high abundances of 5 (Skarbø et al., 1988).
Cores 7231/01, 7320/3, 7425/09 and 7430/10and 8% are recorded in one Valanginian and one

363J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
show a distinctive threefold subdivision: the upper blages of Crucibiscutum salebrosum from the earlyValanginian of the Moray Firth area off N Scotland.claystone sequence (Kolje Fm.), poor in CaCO3, The species is common in the late Valanginian andis underlain by marls and limestones ( Klippfiskearly Hauterivian interval. It decreases in lateFm.) with a high content of CaCO3. Dark clay-Hauterivian/early Barremian times, where it has itsstones (Hekkingen Fm.), poor in CaCO3, formlast occurrence. Williams and Bralower (1995)the lowermost member. The biomicrites of theobserved the following trends for the North Sea.carbonate rich Klippfisk Fm. were deposited underW. barnesae decreases from about 60% in theshallow marine conditions of platform areas andValanginian to 40–50% in the Hauterivian andstructural highs in the Barents Shelf area (Smelrorabout 30% in the Barremian. B. constans, however,et al., 1998). The highly condensed Klippfisk Fm.shows a steady increase from 5–10% in Valanginian(7231/01=4.7 m; 7320/3=3.3 m; 7425/09=5.3 m;times to 20–30% in the Barremian. Rhagodiscus spp.7430/10=9.2 m) range in age from late Berriasian(R. asper, R. splendens) are rare in the Valanginianto earliest Barremian. The carbonate is derived(0–5%), variable in the Hauterivian (0–30%) andfrom shell debris in particular Inoceramus prisms.common in the Barremian (about 10%). OtherIntervals containing calcareous nannofossils arecommon taxa, not considered here, includemost abundant in the Klippfisk Fm. NannofossilsVekshinella stradneri, Zeugrhabdotus spp., andare only absent from the carbonates of core 7320/3,Corollithion ellipticum. Crucibiscutum salebrosum is
where the Klippfisk Fm. is dolomitized and repre- not reported in this study.sented by coquina beds. The high CaCO3 content From further south, Mutterlose (1991) com-of the Klippfisk Fm. explains why the nannoflora piled the following data for NW Germany. W.is often moderately or heavily overgrown and thus barnesae is the most common species both inis much poorer preserved than that of core 6307/07. basinal and marginal settings, showing a steadyIt seems unlikely that the high abundances of C. decrease of abundance from 75% (Valanginian) tosalebrosum in the Klippfisk Fm. of the Barents 50% (Hauterivian), 40% (upper Barremian) andShelf reflect a regional carbonate platform–basin 20% (Aptian). Abundances in basinal environ-signal for two reasons. The sediments of core ments are generally higher than in marginal set-6307/7, deposited under a hemipelagic regime, tings. B. constans is the second most commonyield nannofossil assemblages of similar abun- species. In contrast to W. barnesae, the basin anddances. The highest abundances of C. salebrosum margin environments show an increase in abun-seem to occur simultaneously in the early dance through the Early Cretaceous. The speciesValanginian of the northen and southern hemi- is rare in the Valanginian (<5%), increasing to
20% in the uppermost Barremian and 25% in thesphere (see Section 6.3).Aptian. C. salebrosum is rare throughout the EarlyCretaceous: in most samples, it varies from 0.3 to1% and rarely exceeds 2%. Few samples from the6. Nannofossil abundance of other areasHauterivian yield abundances up to 6%. R. aspershows a steady increase in abundance throughoutIn order to show latitudinally influenced fluctu-the Valanginian–Aptian interval. Abundances ofations of abundances, additional data from further3% characterize the Valanginian; 6% have beensouth were compiled from the literature.encountered in the Hauterivan, 11% in theBarremian and 25% in the late Aptian.6.1. NW EuropeNannoconids are rare in most samples; their abun-dance varies between 0 and 10%. Major peaks inThe NW European area (England, North Seaabundance only occur in the Barremian.and Germany) has been well studied, and plenty
of data are readily available for the Berriasian– 6.2. Low latitudesAlbian interval (e.g. Jakubowski, 1987; Crux, 1989;Mutterlose, 1991; Williams and Bralower, 1995). The Berriasian–Aptian interval in Italy
(Lombardy) has been studied by Erba and QuadrioJakubowski (1987) reports abundant assem-

364 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
(1987). According to these data, W. barnesae is 6.3. Antarcticaabundant to common, B. constans is rare to absent,
Data from the Antarctic region have been com-C. salebrosum is absent, R. asper is rare to few,piled by Mutterlose and Wise (1990). ODP Holeand Nannoconus spp. is common to rare (Erba692 B yields fairly rich nannofossil assemblages ofand Quadrio, 1987).Valanginian and Hauterivian age. This site has aData for Umbria (Italy) have been compiled byValanginian palaeolatitude of about 70°S, reflect-Bralower (1987). W. barnesae is abundant toing an antipode position of the Norwegian setting.common, B. constans is absent, C. salebrosum isW. barnesae is by far the most common species inabsent, R. asper and Nannoconus spp. are commonthe Valanginian (75–90%) and Hauterivian (50–to rare. Neither of the data sets from Italy can be80%). B. constans is present in most samples. Itdirectly correlated with the Norwegian results,varies from 0.3 to 14% in the Valanginian–since no exact counts, only estimates, were made.Hauterivian interval, with the most common abun-Thus, the results from these two Italian areas allowdances between 1 and 5%. C. salebrosum is theonly an indirect comparison.second most common species in the interval underSpecimen counts on DSDP Site 603 (off Capdiscussion. It is present in any of the samples andHatteras, USA) gave the following abundances forranges between 1 and 28%, with most commonthe Berriasian–Aptian interval. W. barnesae isabundances between 3 and 10%. R. asper is rarecommon to abundant (4–69%), and B. constans isin all samples. Abundances between 0.3 and 4%few to common (0.3–13%). Only 15 out of 78characterize the Valanginian and Hauterivian.samples yielded C. salebrosum, which is rare (0–Nannoconids were not found in the samples1%). R. asper, present throughout the core, isanalysed.common to abundant (4–20%), with the most
common abundances between 8 and 13%.Nannoconus spp. is absent in most samples, its
7. Discussionabundance varies from 0.2 to 8%.
Low-latitude diversity can be summarized as7.1. Palaeoecology of calcareous nannofossils
generally high. For DSDP Site 603, the followingdiversities have been observed (20 species in the Keeping in mind that calcareous nannofossilsBerriasian; 59 species in the Valanginian/ are dissolution-susceptible, they are otherwise anHauterivian; 55 species in the Barremian). A ideal proxy for recording palaeooceanographictotal of 67 species were encounterd in the conditions of surface waters. A variation in theirBerriasian–Barremian interval. Abundances are abundance and composition may reflect changeshigh as well. The assemblages are dominated in in palaeoclimate, nutrient supply, detrital inputorder of abundance by W. barnesae, R. asper, C. and surface-water salinity. In addition, six or sevenmargerelii and Nannoconus spp. Rare species, taxa that usually make up 80–90% of the nannofos-however, include B. constans and C. salebrosum. sil assemblages may be used for palaeoecologicalThe latter species, which also rarely (0–0.3%) interpretations. These are mainly based on theoccurs in the Valanginian of DSDP Site 416B off following observations:Morocco (S. Kruse, pers. commun.), is rather $ In recent settings, a high diversity is indicativesmall and easily may have been overlooked in of stable conditions, in particular oligotrophicthe Italian sections. Alternatively, the absence warm surface waters (McIntyre and Be, 1967;of C. salebrosum in the Italian sections might McIntyre et al., 1970; Brand, 1994).reflect a preservation problem: the Italian $ Low-diversity assemblages, however, are con-Maiolica is characterized by dissolution-resistant sidered to be typical for unstable conditionstaxa (E. Erba, pers. commun.). However, the and cool surface waters enriched with nutrientsrare findings prove its presence in the (Okada and Honjo, 1973; Brand, 1994).
$ In the fossil record, W. barnesae is consideredValanginian of low-latitudinal sites.

365J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
to be a eurytopic species that is very resistant seem to be controlled primarily by autoecologicalfactors. This is supported by high abundances ofagainst dissolution and early-diagenetic alter-
ation. A dominance of this species can be used small, dissolution susceptible taxa (e.g. C. salebro-sum) in all cores. Rare specimens of Corollithionas a signal for diagenetic overprint (Roth, 1986;
Roth and Krumbach, 1986). However, W. geometricum, a delicate form, also support thisscenario. Diagenetic overprint and/or regionalbarnesae is an ecologically robust form that is
one of the first species to settle in new biotopes factors controlling the composition of the assem-blages can be excluded. The high abundance of C.(Mutterlose, 1991).
$ A dominance of B. constans and Zeugrhabdotus salebrosum (15–48%) is thus viewed as a pristinesignal. High abundances of C. salebrosum upspp. (20–30%) is considered to be indicative of
a high nutrient supply (Roth and Bowdler, to 39%, observed in cores 7425/09, 7430/10, con-firm this early Valanginian C. salebrosum peak.1981; Roth, 1986; Roth and Krumbach, 1986;
Watkins, 1986, 1989; Erba, 1987, 1989; Erba Both B. constans and Zeugrhabdotus spp. are rela-tively rare in the Norwegian samples, withet al., 1992). Such a B. constans–Zeugrhabdotus
spp. assemblage is usually considered to indicate Zeugrhabdotus spp. rarely exceeding 2% and B.constans attaining its maximum with 7.5%. Theseupwelling of cold water rich in nutrients.
$ Crucibiscutum salebrosum, which is more abundances correspond with those observed insediments considered to be deposited under low–common in the high latitudes, is viewed as a
cool-water species (Mutterlose, 1992). moderate fertility in NW Germany. The values aredefinitely lower than those observed for the mid-$ Rhagodiscus asper, Nannoconus spp.,
Micrantholithus spp. and Conusphaera spp. are Cretaceous along continental margins, as given inRoth and Bowdler (1981), Roth (1986) and Erbainterpreted as thermophile warm water taxa
(Erba, 1987; Mutterlose, 1991; Erba et al., et al. (1992), who quote abundances between 20and 40%. The high abundance of Crucibiscutm1992). An increase in the abundance of R. asper
is, by some authors (Erba, 1987; Erba et al., salebrosum otherwise may indicate cool–cold sur-face waters.1992), seen as an indicator of warm surface
water poor in nutrients. In the middle Aptian,R. asper is frequently associated with the
7.2.2. Mid-HauterivianTethyan genus Nannoconus.
The nannofossil assemblages of the BarentsShelf are characterized by high abundances of W.barnesae (50–80%). C. salebrosum is still abundant7.2. Norwegian settings(15–20%) but less common than in the earlyValanginian. The values for B. constans andNone of the Norwegian cores studied yields
nannofossil assemblages as diverse as those of Zeugrhabdotus spp. correspond with those of theearly Valanginian. These data may reflect low–lower latitudes. A comparison of abundances of
high-, mid- and low latitudes clearly show maxima moderate fertility with a slight increase in surfacewater temperatures.for each stage in the Tethys. This implies assem-
blages of a lower diversity for the high latitudesand of a higher diversity for the lower latitudes in
7.2.3. Barremianthe Early Cretaceous. Thus, the low diversity
The major change in the composition of thereflects the unstable conditions in the high-latitudi-
assemblages occurs to C. salebrosum. This speciesnal settings.
decreases steadily to 2–17% in the earliestBarremian (core 7430/10; 33.00–36.00 m; core7425/09; 52.00–55.00 m). The earliest Barremian7.2.1. Early Valanginian
Due to the relatively low abundance of W. of core 7430/10 has been tentatively assigned tothe S. variabilis ammonite zone of the NWbarnesae (19–34%) in most samples of core
6307/07, the nannofossil assemblages of this site European scheme. This interval has hitherto been

366 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
included in the uppermost Hauterivian, for which brosum was able to migrate from one floral zoneto another.abundances of 1–5% have been observed in NW
Germany. The dramatic decrease in C. salebrosum $ In the lower latitudes, C. salebrosum is repre-sented by a non-calcareous generation that ismight reflect the onset of the Barremian–Aptian
arid phase in NW Europe. not preserved.Since rare specimens of C. salebrosum have been
observed in the Valanginian of DSDP sites 416B7.3. Global patternsand 603, the second explanation is favouredhere.C. salebrosum, rare or absent in the low lati-
tudes, is extremely common in the Norwegian and Similar patterns are known from recent coccol-iths in the Atlantic. Presently, Coccolithus pelagicusAntarctic samples. It shows a bipolar distribution
especially during early Valanginian and mid- is only common at high northern latitudes in theNorth Atlantic, where it dominates the assem-Hauterivian times (Figs. 9 and 10). The extreme
rarity of C. salebrosum in the low latitudes may blages (McIntyre and Be, 1967; Baumann et al.,in press). According to Samtleben et al. (1995), itbe explained by one of the following arguments:
$ C. salebrosum is a coldwater morphotype of a belongs to the ‘Polar group’ and is restricted tocold–temperate waters. It has a positive correlationtropical species.
$ In the lower latitudes, C. salebrosum is only rare. to low water temperatures ranging from −1 to14°C with an optimum at about 8°C (Samtleben$ During phases of cooler temperatures, C. sale-
Fig. 9. Palaeobiogeography of C. salebrosum in Valanginian–Hauterivian times. Compiled from numerous sources; see text. 1, 7425/09;2, 7430/10; 4, 6307/07; 5, Moray Firth; 6, Speeton; 7, NW Germany; 8, Wawal, Poland; 9, DSDP 398D (only in Fig. 10); 10, DSDP603B; 11, Lombardy; 12, Umbria; 13, DSDP 105; 14, DSDP 416; 15, DSDP 370; 16, DSDP 534; 17, DSDP 535; 18, DSDP 391; 19,DSDP 368; 20, DSDP 367 (only in Fig. 10); 21, DSDP 511; 22, ODP 692; 23, DSDP 261; 24, ODP 765; 25, ODP 761; 26, ODP 762;27, ODP 763.
367J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
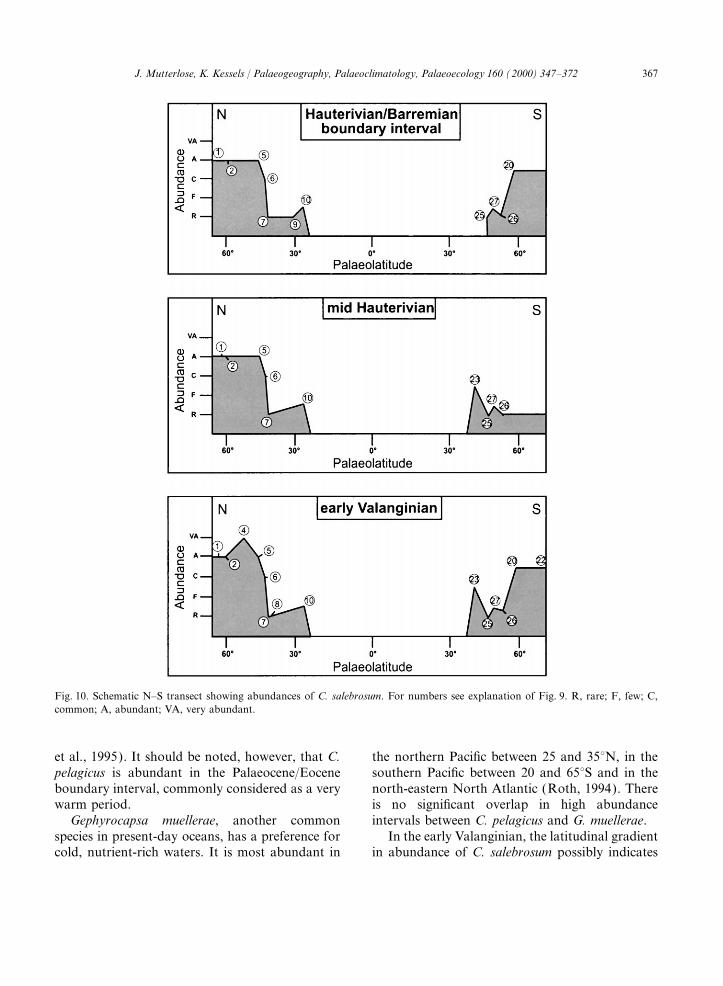
Fig. 10. Schematic N–S transect showing abundances of C. salebrosum. For numbers see explanation of Fig. 9. R, rare; F, few; C,common; A, abundant; VA, very abundant.
et al., 1995). It should be noted, however, that C. the northern Pacific between 25 and 35°N, in thesouthern Pacific between 20 and 65°S and in thepelagicus is abundant in the Palaeocene/Eocene
boundary interval, commonly considered as a very north-eastern North Atlantic (Roth, 1994). Thereis no significant overlap in high abundancewarm period.
Gephyrocapsa muellerae, another common intervals between C. pelagicus and G. muellerae.In the early Valanginian, the latitudinal gradientspecies in present-day oceans, has a preference for
cold, nutrient-rich waters. It is most abundant in in abundance of C. salebrosum possibly indicates

368 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
latitudinal differences in temperature. The palaeo- correspond to the recent E. huxleyi–G. muelleraeassemblage of the temperate floral zone.ceanographic settings may have strengthened these
thermal gradients. A significant physical separation $ W. barnesae–Nannoconus spp.–R. asper assem-blage: low latitudes; warm surface waters, poorof Tethyan and Boreal Realms in the Berriasian
and Valanginian restricted the exchange of water in nutrients; this assemblage may correspond tothe recent E. huxleyi–G. oceanica assemblage ofand biota and thereby led to the formation of
different climatic floral zones. The late Valanginian the subtropical and tropical floral zones.transgression put an end to this situation.
For this reason, the relatively low abundanceof R. asper in the Norwegian sites is held here to 8. Conclusionsbe an indicator for cool surface waters. The migra-tion of Tethyan nannoconids into the Boreal In the early Valanginian, the high abundances
of C. salebrosum indicate cool to cold surfaceRealm was only possible during times of suitablepalaeoceanographic settings. The significant maxi- water in the high latitudes (Fig. 11). The restricted
palaeoceanographic setting may have reinforcedmum in Barremian sediments in core 7425/09 (8%)may include endemic boreal taxa such as the thermal gradient. A significant separation of
the Tethyan and Boreal Realm in the BerriasianNannoconus abundans and Nannoconus borealis.These are quite common in NW Europe and and Valanginian reduced the exchange and mixing
of oceanic waters. This increased the formation ofcharacterize the restricted epicontinental NorthSea to Barents Sea. different climatic zones. The late Valanginian
transgression put an end to this biogeographicThe patterns observed in the Norwegian sitesclearly reflect a W. barnesae–C. salebrosum assem- isolation. The high abundance of C. salebrosum in
the Hauterivian of the Norwegian samples indi-blage, which is considered here to be typical forhigh latitudes. Since the eutrophic cool-water cates, however, the continuation of high-latitudinal
floral zones.species, B. constans, is relatively rare in mostsamples, nutrients may be disregarded as a major It is difficult to sort out a single controlling
factor for the composition described above. Bothfactor controlling the assemblage composition.This is supported by the scarcity of Zeugrhabdotus sea-level changes and palaeoclimate may have con-
trolled the high abundances of C. salebrosum inspp. The abundance of C. salebrosum, a smalldelicate form, indicates the absence of dissolution the Norwegian samples. The Berriasian/
Valanginian sea-level lowstand may have causedor diagenetic overprint. Its common occurrence inhigh latitudes either may indicate cool–cold surface endemic evolution of calcareous nannofossil taxa,
possibly allowing the radiation of C. salebrosum.waters poor in nutrients or may have occupied theniche that B. constans occupies further south. This This evolutionary pattern may be compared to the
in-situ evolution of N. abundans in the NWW. barnesae–C. salebrosum assemblage of the highlatitudes may be compared to the recent E. hux- European Barremian. This, however, is contra-
dicted by the presence of C. salebrosum in twoleyi–C. pelagicus assemblage of the subarctic floralzone. According to current data, it seems possible high latitudiunal belts both in the north and in the
south. Furthermore, one might expect a decreaseto recognize the following assemblages for theEarly Cretaceous: of C. salebrosum in the Hauterivian. A palaeocli-
matic control implies the existence of climatic belts$ W. barnesae–C. salebrosum–S. horticus assem-blage: high latitudes; bipolar; cool–cold surface throughout most of the Early Cretaceous, resulting
in considerable temperature gradients from northwaters, poor in nutrients; this assemblage maycorrespond to the recent E. huxleyi–C. pelagicus to south. The Early Cretaceous biogeographic
pattern is very similar to that found in modernassemblage of the subarctic floral zone.$ W. barnesae–B. constans–Zeugrhabdotus spp. oceans. From north to south, the modern Atlantic
Ocean consists of a subarctic, northern temperate,assemblage: mid–low latitudes; cool surfacewaters, rich in nutrients; this assemblage may northern subtropical, tropical, southern sub-
369J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Fig
.11.
Synt
hesi
sof
dist
ribu
tion
patt
erns
ofva
riou
sgr
oups
ofca
lcar
eous
nann
ofos
sils
and
thei
rpa
laeo
ecol
ogic
alin
terp
reta
tion
.A
bsol
ute
ages
acco
rdin
gto
Gra
dste
inan
dO
gg(1
996
),se
a-le
vel
curv
efr
omH
arde
nbol
etal
.(19
98),
ocea
n-cr
ust
prod
ucti
onfr
omL
arso
n(1
991a
,b),
ice
raft
edde
posi
tsfr
omK
empe
r(1
987)
and
Fra
kes
and
Fra
ncis
(198
8),d13
Cfr
omW
eiss
ert
and
Lin
i(1
991)
,d18
Ohi
ghla
titu
des
from
Pod
laha
etal
.(1
998
),an
dd18
Olo
wla
titu
des
from
Dou
glas
and
Woo
druff
(198
1).

370 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
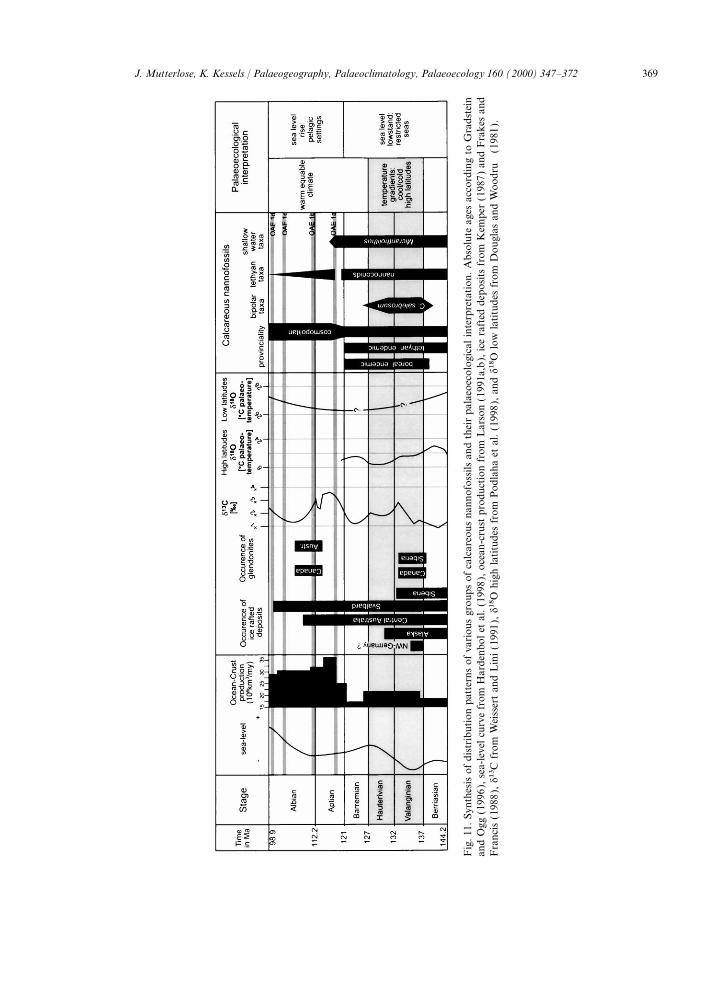
1990. Stratigraphy, geochemistry and paleoceanography oftropical, southern temperate and subantarctic belt.organic carbon-rich Cretaceous sequences. In: Ginsburg,The palaeobiogeographic patterns described sug-R.N., Beaudoin, B. (Eds.), Cretaceous Resources Eventsgest considerable thermal gradients for the Early and Rhythms. Kluwer, Dordrecht, pp. 75–119.
Cretaceous. The data gained from calcareous nan- Baumann, K.H., Andruleit, H., Samtleben, C., press. Coccoli-nofossils support the idea of an ice-house world thophores in the Nordic Seas: Comparison of living commu-
nities with surface sediment assemblages. Deep Sea Res.,or a cold mode for the early Valanginian andin press.early–mid-Hauterivian and clearly disagree with a
Bown, P.R., 1998. Calcareous Nannofossil Biostratigraphy.warm equable climate for the earliest CretaceousChapman & Hall, London. 314 pp.
(Valanginian–Hauterivian) as suggested, for exam- Bralower, T.J., 1987. Valanginian to Aptian calcareous nanno-ple, by Frakes (1979) and Hallam (1981, 1985). fossil stratigraphy and correlation with the upper
M-sequence magnetic anomalies. Mar. Micropaleontol.Our findings are supported by Strontium and11, 293–310.d18O data, suggesting periods of low temperatures
Brand, L.E., 1994. Physiological ecology of marine coccolitho-or high ice volume or both, for the latest Berriasianphores. In: Winter, A., Siesser, W.G. (Eds.), Coccolitho-
and early Valanginian (Stoll and Schrag, 1996). phores. Cambridge University Press, Cambridge, pp. 39–50.Bugge, T., Elvebakk, G., Bakke, S., Fanavoll, S., Lippard, S.,
Leith, T.L., Mangerud, G., Moller, N., Nilsson, I., Rømuld,A., Schou, L., Vigran, J.O., Weiss, H.M., Arhus, N., 1988.AcknowledgementsShallow drilling Barents Sea. IKU-Report. Trondheim.309 pp.
Financial support by the Deutsche Casey, R., 1973. The ammonite succession at the Jurassic–Cret-Forschungsgemeinschaft (Mu 667/6; 667/14-1, -2) aceous boundary in eastern England. In: Casey, R., Rawson,is gratefully acknowledged. SINTEF Petroleum P.F. (Eds.), The Boreal Lower Cretaceous, Geol. J. 5,
193–266.Research (Trondheim) kindly supplied the samplesCrux, J.A., 1989. Biostratigraphy and palaeogeographical appli-and gave access to unpublished reports. Atle Mørk,
cations of Lower Cretaceous nannofossils from north-west-Morten Smelror (Trondheim) and H. Weissertern Europe. In: Crux, J.A., van Heck, S.E. (Eds.),
(Zurich) made valuable comments and helped Nannofossils and their Applications. Ellis Horwood,improve the manuscript. D. Grocke (Oxford) criti- Chichester, UK, pp. 143–211.
Ditchfield, P.W., 1997. High northern palaeolatitude Juras-cally reviewed the manuscript, suggested consider-sic–Cretaceous palaeotemperature variation: New dataable changes and improved the English. H.from Kong Karls Land, Svalbard. Palaeogeogr. Palaeocli-Andruleit (Hannover) made valuable commentsmatol. Palaeoecol. 130, 163–175.
on the biogeography of recent coccoliths and A. Douglas, R.G., Woodruff, F., 1981. Deep sea benthic foramini-Bornemann (Bochum) compiled the figures. We fera, Emiliani, C. (Ed.), The Sea Vol. 7. Wiley-Interscience,
New York, pp. 1233–1327.thank E. Erba (Milano) and K. von Salis (Zurich)Erba, E., 1987. Mid-Cretaceous cyclic pelagic facies from thefor critical reviews and many useful suggestions.
Umbrian–Marchean Basin: What do calcareous nannofos-sils suggest? Int. Nannofossil Assoc. Newslett. 9, 52–53.
Erba, E., Quadrio, B., 1987. Biostratigrafia a Nannofossili calc-References arei, Calpionellidi e Foraminiferi planctonici della Maiolica
(Titoniano superiore–Aptiano) nelle Prealpi Bresciane(Italia settentrionale). Riv. Ital. Paleontol. Stratigraf. 93,Arhus, N., Bugge, T., Elvebakk, G., Fjerdingstad, V., Leith,3–108.T.L., Mørk, A., Rendall, H., Skarbø, O., Weiss, H.M., 1987.
Erba, E., 1989. Upper Jurassic to Lower Cretaceous Nannoco-Shallow drilling Elf Aquitaine Norsk Hydro Statoil Barentsnus distribution in some sections from Northern and CentralSea. IKU-Report. Trondheim. 50 pp.Italy. Mem. Sci. Geol. 41, 255–261.Arhus, N., Kelly, S.R.A., Collins, J.S.H., Sandy, M.R., 1990.
Erba, E., Castradori, D., Guasti, G., Ripepe, M., 1992. Calcare-Systematic palaeontology and biostratigraphy of two Earlyous nannofossils and Milankovitch cycles: The example ofCretaceous condensed sections from the Barents Sea. Polarthe Albian Gault Clay Formation (southern England).Res. 8, 165–194.Palaeogeogr. Palaeoclimatol. Palaeoecol. 93, 47–69.Arhus, N., 1991. The transition from deposition of condensed
Fjerdingstad, V., Arhus, N., Bugge, T., Elvebakk, G., Fanavoll,carbonates to dark claystones in the Lower Cretaceous suc-S., Helgesen, I.K., Leith, T.L., Schou, L., Sættem, J.,cession of the southwestern Barents Sea. Norsk Geol. Tids-Vigvan, J.O., 1985. Shallow Drilling Barents Sea 1985. IKU-skrift 71, 259–263.
Arthur, M.A., Brumsack, H.-J., Jenkyns, H.C., Schlanger, S.O., Report. Trondheim.

371J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
Frakes, L.A., 1979. Climates Through Geological Time. Elsev- Larson, R.L., 1991a. Latest pulse of Earth: Evidence for a mid-Cretaceous superplume. Geology 19, 547–550.ier, Amsterdam. 310 pp.
Frakes, L.A., Francis, J.E., 1988. A guide to Phanerozoic cold Larson, R.L., 1991b. Geological consequences of superplumes.Geology 19, 963–966.polar climates from high latitude ice-rafting in the Creta-
ceous. Nature 333, 547–549. McIntyre, A., Be, A.W.H., 1967. Modern Coccolithophoridaeof the Atlantic Ocean—I. Placoliths and Cyrtoliths. DeepFrakes, L.A., Francis, J.E., Syktus, J.I., 1992. Climate Modes
of the Phanerozoic. Cambridge University Press, Cam- Sea Res. 14, 561–597.McIntyre, A., Allan, W.H., Roche, M.B., 1970. Modern pacificbridge. 274 pp.
Francis, J.E., Frakes, L.A., 1993. Cretaceous climates. In: Coccolithophorida: A paleontological thermometer. NYAcad. Sci. Trans. 32, 720–731.Wright, V.P. (Ed.), Sedimentology Review 1. Blackwell,
Oxford, pp. 17–30. Mutterlose, J., Wise Jr., S.W., 1990. Lower Cretaceous nanno-fossil biostratigraphy of ODP LEG 113 Holes 692B andGradstein, F.M., Ogg, J.G., 1996. A Phanerozoic time scale
(plus chart). Episodes 19, 3–5. 693A, Continental slope off east Antarctica, Weddell Sea.Proc. ODP, Sci. Res. 113, 325–351.Gradstein, F.M., Kaminski, M.A., Agterberg, F.P., 1999. Bio-
stratigraphy and paleoceanography of the Cretaceous Mutterlose, J., 1991. Das Verteilungs- und Migrationsmusterdes kalkigen Nannoplanktons in der borealen Unterkreideseaway between Norway and Greenland. Earth Sci. Rev.
46, 27–98. (Valangin-Apt) NW-Deutschlands. Palaeontographica B221, 27–152.Hallam, A., 1981. Facies Interpretation and the Stratigraphic
Record. Freeman and Company, Oxford. 291 pp. Mutterlose, J., 1992. Biostratigraphy and palaeobiogeographyof Early Cretaceous calcareous nannofossils. CretaceousHallam, A., 1985. A review of Mesozoic climates. J. Geol. Soc.
Lond. 142, 433–445. Res. 13, 167–189.Mutterlose, J., 1998. The Barremian–Aptian turnover of biotaHallam, A., Grose, J.A., Ruffell, A.H., 1991. Palaeoclimatic
significance of changes in clay mineralogy across the Juras- in northwestern Europe: evidence from belemnites. Palaeo-geogr. Palaeoclimatol. Palaeoecol. 144, 161–173.sic–Cretaceous boundary in England and France. Palaeo-
geogr. Palaeoclimatol. Palaeoecol. 81, 173–187. Okada, H., Honjo, S., 1973. The distribution of oceanic coccoli-thophorids in the Pacific. Deep Sea Res. 20, 355–374.Hallam, A., 1992. Phanerozoic Sea-level Changes. Columbia
University Press, New York. 266 pp. Perch-Nielsen, K., 1985. Mesozoic calcareous nannofossils. In:Bolli, H.M., Saunders, J.B., Perch-Nielsen, K. (Eds.),Hallam, A., 1994. An outline of phanerozoic biogeography,
Oxford Biogeography Series 10, 246 pp. Plankton Stratigraphy. Cambridge University Press, Cam-bridge, pp. 329–426.Haq, B.U., Hardenbol, J., Vail, P.R., 1988. Mesozoic and Ceno-
zoic chronostratigraphy and cycles of sea-level change. Spec. Podlaha, O.G., Mutterlose, J., Veizer, J., 1998. Preservation ofd18O and d13C in belemnite rostra from the Jurassic/EarlyPubl. Soc. Econ. Petr. Mineral. 42, 71–108.
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., de Graci- Cretaceous successions. Am. J. Sci. 298, 324–347.Price, G.D, Ruffell, A.H., Jones, C.E., Kalin, R.M., Mutterlose,ansky, P.C., Vail, P.R., 1998. Mesozoic and Cenozoic
sequence chronostratigraphic framework of European J., 2000. Isotopic evidence for temperature variation duringthe Early Cretaceous ( late Ryazanian–mid Hauterivian).basins. Soc. Sed. Geol. Spec. Publ. 60, 3–13.
Heller, P.L., Anderson, D.L., Angevine, C.L., 1996. Is the J. Geol. Soc. Lond. 157, 335–343.Rawson, P.F., Riley, L.A., 1982. Latest Jurassic–Early Creta-middle Cretaceous pulse of rapid sea-floor spreading real or
necessary? Geology 24, 491–494. ceous events and the ‘late Cimmerian unconformity’ inNorth Sea area. AAPG 66, 2628–2648.Hoedemaeker, P.J., 1990. The Neocomian boundaries of the
Tethyan Realm based on the distribution of ammonites. Cre- Rawson, P.F., 1994. Sea-level changes and their influence onammonite biogeography in the European Early Cretaceous.taceous Res. 11, 331–342.
Jakubowski, M., 1987. A proposed Lower Cretaceous calcare- Palaeopelagos Spec. Pub. 1, 317–326.Roth, P.H., 1986. Mesozoic palaeoceanography of the Northous nannofossil zonation scheme for the Moray Firth area
of the North Sea. Abh. Geol. B.-A 39, 99–119. Atlantic and Tethys Oceans. In: Summerhayes, C.P., Shack-leton, N.J. (Eds.), North Atlantic Palaeoceanography, Geol.Jenkyns, H.C., 1980. Cretaceous anoxic events from continents
to oceans. J. Geol. Soc. Lond. 137, 1171–1188. Soc. 21, 299–320.Roth, P.H., 1994. Distribution of coccoliths in oceanic sedi-Kemper, E., Rawson, P.F., Thieuloy, J.-P., 1981. Ammonites
of Tethyan ancestry in the early Lower Cretaceous of north- ments. In: Winter, A., Siesser, W.G. (Eds.), Coccolitho-phores. Cambridge University Press, Cambridge, pp. 39–50.west Europe. Palaeontology 24, 251–311.
Kemper, E., 1987. Das Klima der Kreide-Zeit. Geol. Jahrb. A Roth, P.H., Bowdler, J.L., 1981. Middle Cretaceous calcareousnannoplankton biogeography and oceanography of the96, 5–185.
Kominz, M.A., 1984. Oceanic ridge volumes and sea-level Atlantic Ocean. Soc. Econ. Paleontol. Mineral. Spec. Publ.32, 517–546.change—an error analysis. In: Schlee, J., Schlee, S. (Eds.),
Interregional Unconformities and Hydrocarbon Accumula- Roth, P.H., Krumbach, K.R., 1986. Middle Cretaceous calcare-ous nannofossil biogeography and preservation in the Atlan-tion. Am. Assoc. Petrol. Geol, pp. 109–127.

372 J. Mutterlose, K. Kessels / Palaeogeography, Palaeoclimatology, Palaeoecology 160 (2000) 347–372
tic and Indian oceans: implications for paleoceanography. of Eurasia and the paleofloristic provinces of this period.Trans. Geol. Inst. Moscow 102, 1–263, (in Russian).Mar. Micropaleontol. 10, 235–266.
Ruffell, A., Batten, D.J., 1990. The Barremian–Aptian arid Vakhrameev, V.A., 1991. Jurassic and Cretaceous floras andclimates of the Earth. Cambridge University Press,phase in northern Europe. Palaeogeogr. Palaeoclimatol.
Palaeoecol. 80, 197–212. Cambridge.Watkins, D.K., 1986. Calcareous nannofossil paleoceanographyRuffell, A., 1991. Sea-level events during the Early Cretaceous
in western Europe. Cretaceous Res. 12, 527–551. of the Cretaceous Greenhorn Sea. Geol. Soc. Am. Bull. 97,1239–1249.Samtleben, C., Baumann, K.-H., Schroder-Ritzrau, A., 1995.
Distribution, composition and seasonal variation of coccli- Watkins, D.K., 1989. Nannoplankton productivity fluctuationsand rhythmically-bedded pelagic carbonates of the Green-thophore communities in the northern Atlantic. In: Flores,
J.A., Sierro, F.J. (Eds.), Proc. 5th INA Conf. Salamanca, horn Limestone (Upper Cretaceous). Palaeogeogr. Palaeo-climatol. Palaeoecol. 74, 75–86.219–235.
Schlanger, S.O., Jenkyns, H.C., 1976. Cretaceous anoxic events: Weissert, H., Lini, A., 1991. Ice age interludes during the timeof Cretaceous greenhouse climate? In: Mueller, D.W.,causes and consequences. Geol. Mijnbouw 55, 79–184.
Skarbø, O., Bakke, S., Jacobsen, T., Krokstad, W., Lundschien, Mckenzie, J.A., Weissert, H. (Eds.), Controversies inModern Geology. Academic Press, London, pp. 173–191.B., Myhr, M.B., Rise, L., Schou, L., Smelror, M., Verdenius,
J., Vigran, J., Arhus, N., 1988. Shallow Drilling off Williams, J.R., Bralower, T.J., 1995. Nannofossil assemblages,fine fraction stable isotopes, and the paleoceanography ofMøre–Trøndelag. IKU-Report. Trondheim.
Smelror, M., Mørk, A., Monteil, E., Rutledge, D., Leereveld, the Valanginian–Barremian (Early Cretaceous) North SeaBasin. Paleoceanography 10, 815–839.H., 1998. The Klippfisk Formation—a new lithostrati-
graphic unit of Lower Cretaceous platform carbonates on Winterer, E.L., 1991. The Tethyan Pacific during Late Jurassicand Cretaceous times. Palaeogeogr. Palaeoclimatol. Palaeo-the Western Barents Shelf. Polar Res. 17, 181–202.
Stoll, H.M., Schrag, D.P., 1996. Evidence for glacial control of ecol. 87, 253–265.Ziegler, P.A., 1988. Geological Atlas of Western and Centralrapid sea-level changes in the Early Cretaceous. Science 272,
1771–1774. Europe. Shell Internationale Petroleum Maatschappij, DenHaag. 239 pp.Vakhrameev, V.A., 1964. Jurasic and Early Cretaceous floras





























