Embed Size (px)
Citation preview

Cretaceous Research 29 (2008) 9e26www.elsevier.com/locate/CretRes
Barremian ammonite faunas from the western High Atlas,Morocco e biostratigraphy and palaeobiogeography
Miguel Company a,*, Jose Sandoval a, Jose M. Tavera a,Mohamed Aoutem b, Mohssine Ettachfini c
a Departamento de Estratigrafıa y Paleontologıa, Universidad de Granada, E-18002 Granada, Spainb Departement de Geologie, Faculte des Sciences, Universite Ibn Zohr, BP28/S, Agadir, Morocco
c Departement de Geologie, Faculte des Sciences Semlalia, Universite Cadi Ayyad, B.P. 2390, Marrakech, Morocco
Received 5 December 2006; accepted in revised form 14 March 2007
Available online 22 October 2007
Abstract
An analysis of the stratigraphic distribution of ammonite faunas collected from several sections in the Essaouira-Agadir area (western HighAtlas, Morocco) has enabled the establishment of a detailed succession of assemblages for the uppermost Hauterivian to lower Upper Barremianinterval. All Mediterranean standard zones of this interval have been identified. The correlation potential of bioevents which define and char-acterise these units is discussed. Two significant regional hiatuses, one at the Hauterivian/Barremian boundary, the other involving the lower partof the Kotetishvilia nicklesi Zone, are related to third-order sea level falls. Moreover, the upper part of the succession corresponding to the upperToxancyloceras vandenheckii Zone and the entire Heinzia sartousiana Zone is markedly condensed. The taxonomic composition of the ammo-nite assemblage reveals a strong Mediterranean character, yet, it shows some peculiar features as well.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Barremian; Lower Cretaceous; Ammonites; Biostratigraphy; Palaeobiogeography; Sequence stratigraphy; High Atlas; Morocco
1. Introduction
Offshore marine sediments of Barremian age in the westernHigh Atlas of Morocco comprise marls and bioclastic marlylimestones of the Taboulouart Formation, which yield richand well-preserved ammonite faunas. The earliest referencesto these faunas date back to the early twentieth century; sincethen, Barremian ammonites have been frequently noted in theregional stratigraphic literature.
However, there has never been a detailed biostratigraphicstudy of these ammonite faunas, which contrasts stronglywith appreciable progress made over the last two decades inour knowledge of Barremian ammonite faunas from elsewherein the Tethyan realm. To fill this gap, we have undertaken ananalysis of the stratigraphic distribution of ammonites from
* Corresponding author.
E-mail address: [email protected] (M. Company).
0195-6671/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.cretres.2007.03.001
ten systematically sampled sections located throughoutthe coastal strip between Essaouira and Agadir (Fig. 1). Ourresults, presented in this paper (see also Company et al.,2005b), allow correlation of the Barremian ammonite succes-sion in the western High Atlas with the current standard zona-tion for the Lower Cretaceous (Hoedemaeker et al., 2003;Reboulet et al., 2006), and complement several recent contri-butions on ammonites from older Cretaceous strata in the area(Ettachfini, 1991, 2004; Ettachfini et al., 1998; Wippich, 2001,2003).
2. Geological setting and sections studied
Lower Cretaceous strata which crop out in the western HighAtlas were deposited in a marine setting previously referred toas the Haha Basin (Roch, 1930; Choubert and Faure-Muret,1962), the south-western Moroccan Basin (Duffaud et al.,1966) or the Atlas Gulf (Behrens et al., 1978). This basin

Fig. 1. Location of the sections studied.
10 M. Company et al. / Cretaceous Research 29 (2008) 9e26
developed during the Mesozoic on the north-west African pas-sive margin, opened like a funnel towards the Atlantic Ocean,and was bordered by the emergent area of the Moroccan Me-seta to the north and by the Anti-Atlas uplands to the south.
The sections studied comprise green to yellow marls, irregu-larly alternating with sandy and marly limestone beds. Limestonesare wavy- to nodular-bedded biomicrites and biomicrosparites(mudstones, wackestones and packstones), which may containabundant remains of ammonites, bivalves, gastropods, brachio-pods, echinoids, serpulids, ostracods and benthic forams. Thesesediments constitute the Taboulouart Formation (Duffaud et al.,1966) and are interpreted as distal shelf deposits (Rey et al.,1986, 1988; Witam, 1998).
The base of the Taboulouart Formation, of Late Hauterivianage, is generally separated from the sandy sediments of the un-derlying Talmest Formation by a discontinuity related to rapidrelative sea level rise (Rey et al., 1988). The age of its topvaries from latest Early Barremian to early Late Barremian(see below), being invariably capped by marginal marinesandy sediments which mark the base of the BouzergounFormation.
The ammonite-bearing strata of the Taboulouart Formationonly crop out in the western part of the basin, along a coastalstrip that is 25e30 km wide. To the east, towards the basinmargin, ammonites become increasingly rare and lithologies
gradually change to marginal claystones and sandstones(Tazought Formation).
In the present study, >1500 ammonites were collected bybed-by-bed sampling in ten selected sections. Specimens are,in general, moderately well preserved and not deformed, ex-cept for the inner whorls which, during diagenesis, either re-mained empty or were infilled by calcite, and are stronglyflattened or even lost. The ammonite material is housed inthe palaeontological collections of the University of Granada.
The sections sampled are as follows (Fig. 1):Takouwit (co-ordinates 31�17006’’N 9�38028’’W), 10 km
NNE of Smimou; on the northern limb of Jbel Mradma,near the southern edge of the Tidzi diapiric structure. This sec-tion has previously been described by Rey et al. (1986, 1988)and by Witam (1998).
Akoui Griz (co-ordinates 31�17044’’N 9�43025’’W), 9.5 kmN of Smimou; small outcrop along a gentle slope along theroad from Essaouira to Agadir, 1 km N of the bridge overthe Oued Tidzi. Witam (1998) analysed another Barremiansection in the same area.
Sidi M’Bark (co-ordinates 31�15040’’N 9�48013’’W), 11 kmNW of Smimou; excellent exposure on a hill on the left bankof the Oued Aghbalou, between where it joins the AtlanticOcean and the marabou of Sidi M’Bark. This locality hasformerly been reported upon by Lemoine (1905) and byRoch (1930) and, more recently, by Witam (1998). The strati-graphic distribution of ammonites from this section is shownin Fig. 2.
Tafadna (co-ordinates 31�05046’’N 9�48022’’W), 16 kmNW of Tamanar; outcrop on the right bank of the Oued Igou-zouln, some 2 km before it joins the Atlantic Ocean onTafadna beach.
Imsouane (co-ordinates 30�52007’’N 9�48037’’W), 19 kmSW of Tamanar; hill on the right side of the road leading tothe Port of Imsouane. This locality exposes a highly completesuccession of the Taboulouart Formation, corresponding to thedeepest part of the basin. It has previously been studied byWitam (1998). The stratigraphic distribution of ammonitesfrom this section is shown in Fig. 3.
Assaka (co-ordinates 30�48056’’N 9�46036’’W), 22 km SWof Tamanar; steep cliff on the left bank of the Oued Tamkour-imt, 1 km W of the hamlet of Assaka. This section is the sameas that referred to as Igueni Ouram by Roch (1930). It was alsoanalysed by Witam (1998).
Tamri (co-ordinates 30�42059’’N 9�48007’’W), 3 km NE ofTamri; small gully on the right side of the Oued A€ıt Amer. Thesuccession at this locality, which corresponds to a distal swellin the basin, is rather incomplete, showing reduced thickness,condensed levels and many stratigraphic discontinuities. Thestratigraphic distribution of ammonites from this section isshown in Fig. 4.
Addar (co-ordinates 30�36028’’N 9�43043’’W), 16 km SWof Tamri; very good exposure on a saddle at the summit ofIghourad Hill, 1.5 km NW of the hamlet of Addar. This local-ity was mentioned by Witam et al. (1993) and later describedby Witam (1998). The stratigraphic distribution of ammonitesfrom this section is shown in Fig. 5.

Fig. 2. Lithology, ammonite distribution and zonation of the Sidi M’Bark
section.
11M. Company et al. / Cretaceous Research 29 (2008) 9e26
A€ıt Hamouch (co-ordinates 30�36016’’N 9�41008’’W), 4 km Eof the Addar section, in the same hill range, near the dirt road link-ing Imi Waddar with Allagh ou Awrga. A section in the same areahas previously been analysed by Ambroggi (1963).
Sidi Yahya ou Sa€ıd (co-ordinates 30�35046’’N 9�39019’’W),2.5 km E of the A€ıt Hamouch section, on a steep slope oppo-site the marabou of Sidi Yahya ou Sa€ıd. This section was
mentioned by Witam et al. (1993) and later described by Wi-tam (1998).
3. Previous work
The first mention of Barremian ammonites from the west-ern High Atlas is that by Lemoine (1905), who recognisedthis stage in the cliffs of the Oued Aghbalou on the basis ofthe ammonites Pulchellia dumasiana and Crioceras (Ancylo-ceras) aff. zitteli. Shortly afterwards, Kilian and Gentil(1907) recorded a rich and diverse ammonite fauna collectedby the latter author in the Agadir area. They listed morethan thirty ammonite species, mainly from the Lower Barre-mian, and noted the absence of holcodiscids and the predom-inance of crioceratitids.
Roch (1930) was the first to provide a comprehensive surveyof the stratigraphy of the region. Among sediments attributed tothe Barremian Stage he identified, in the deeper westerly part ofthe basin, a cephalopod-bearing marly limestone facies, equiv-alent to the material studied here. The abundant ammonite faunacollected, inclusive of some new forms, allowed him to recog-nise two successive assemblages, a lower one characterised byPulchellia and Desmoceras difficile, and an upper featuring Het-eroceras astieri. He considered these two levels to be the equiv-alent of the two classic horizons in the French Barremian(Combe Petite and Morteyron) introduced several decades ear-lier by Kilian (1888). The same twofold division was main-tained in a much later publication by Roch (Roch, 1950).
Ambroggi and Breistroffer (1959) recognised a Lower Bar-remian substage, with several species of Nicklesia and Barre-mites, a Middle Barremian one with Heinzia provincialis andan Upper Barremian with Heteroceras moutonianum and Mel-chiorites. Later, Ambroggi (1963) modified this scheme byidentifying a Paraspiticeras gr. percevali horizon at the baseof the Barremian and by eliminating any reference to the levelwith H. provincialis.
Duffaud et al. (1966), when introducing their synoptic lithos-tratigraphic chart for the basin, interpreted the Taboulouart For-mation as corresponding to the entire Barremian, with Nicklesiagr. pulchella and Barremites difficilis in its lower part, and Het-eroceras gr. moutonianum in its upper. Those authors attributedthe entire Bouzergoun Formation to the Lower Aptian.
Wiedmann et al. (1978, 1982) presented two large-scale,synthetic stratigraphic logs for the Cretaceous in the Essaouiraand Agadir areas. Their data on Barremian ammonites werescant and ambiguous but, in any case, did not substantially al-ter previous interpretations.
More significant new data were introduced by Rey et al. (1986,1988) and Canerot et al. (1986), who analysed Lower Cretaceousdeposits in the northern part of the basin. They reported, amongother species, Paraspiticeras percevali and Torcapella? barre-mensis from the lower half of the Taboulouart Formation andNicklesia pulchella, Pulchellia compressissima and Moutonicerasmoutonianum from its upper part. This fauna indicated an EarlyBarremian age for the entire Taboulouart Formation. Althoughlacking in palaeontological evidence, the Bouzergoun Formationwas assigned to the Upper Barremian by these authors.
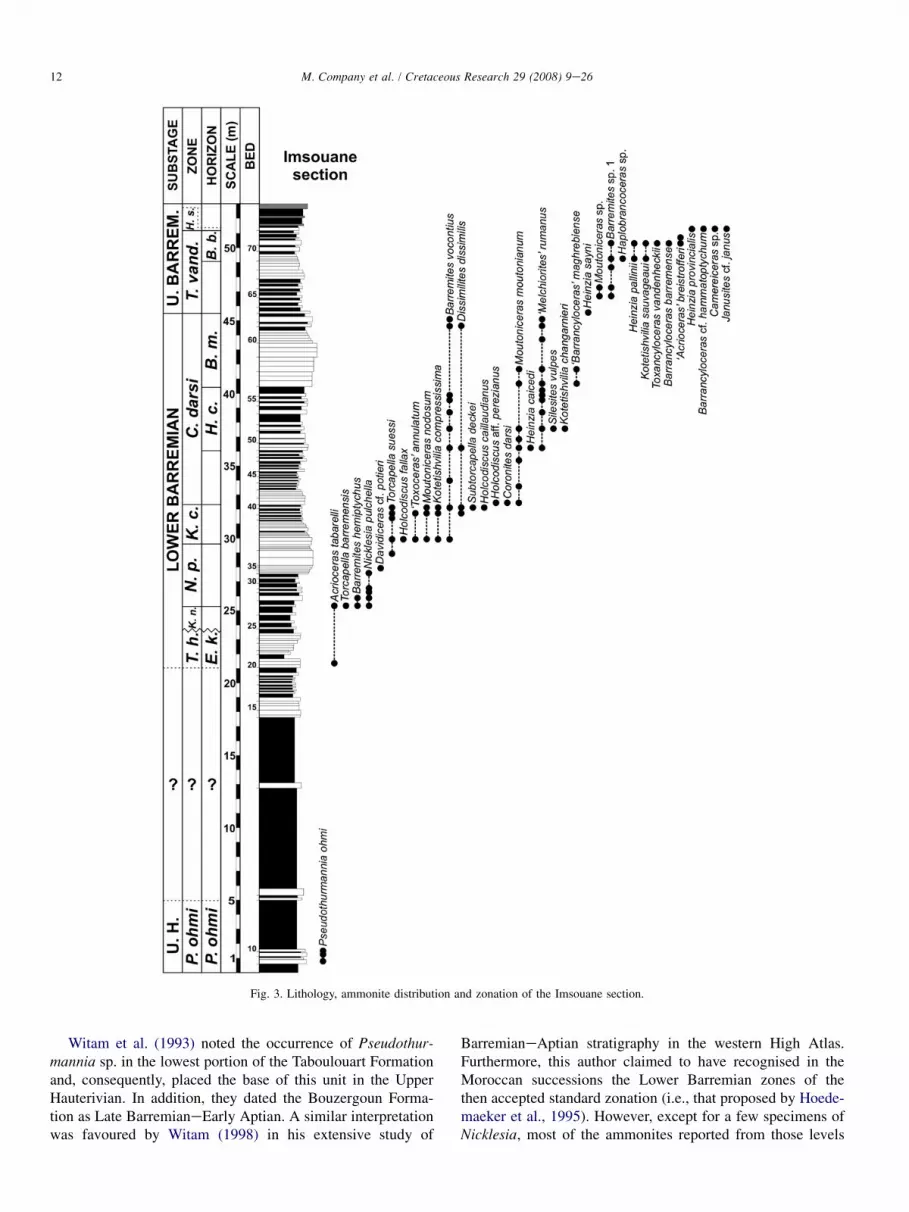
Fig. 3. Lithology, ammonite distribution and zonation of the Imsouane section.
12 M. Company et al. / Cretaceous Research 29 (2008) 9e26
Witam et al. (1993) noted the occurrence of Pseudothur-mannia sp. in the lowest portion of the Taboulouart Formationand, consequently, placed the base of this unit in the UpperHauterivian. In addition, they dated the Bouzergoun Forma-tion as Late BarremianeEarly Aptian. A similar interpretationwas favoured by Witam (1998) in his extensive study of
BarremianeAptian stratigraphy in the western High Atlas.Furthermore, this author claimed to have recognised in theMoroccan successions the Lower Barremian zones of thethen accepted standard zonation (i.e., that proposed by Hoede-maeker et al., 1995). However, except for a few specimens ofNicklesia, most of the ammonites reported from those levels

Fig. 4. Lithology, ammonite distribution and zonation of the Tamri section.
13M. Company et al. / Cretaceous Research 29 (2008) 9e26
by Witam have little diagnostic value and, hence, his strati-graphic attributions do not appear to be fully justified.
4. Ammonite succession and zonation
Our analysis of the stratigraphic distribution of ammonitespecies identified in the Taboulouart Formation has led us toestablish a detailed faunal succession that can be easily corre-lated with the current standard zonation (Hoedemaeker et al.,2003; Reboulet et al., 2006) (Fig. 6).
The stratigraphic interval analysed extends from the upper-most Hauterivian (Pseudothurmannia ohmi Zone) to the lowerUpper Barremian (Heinzia sartousiana Zone). All standardzones of this interval have been recognised. Nevertheless,more or less important hiatuses and condensations, either oflocal or regional extent, which can obviously distort the actualranges of taxa, have been detected.
4.1. Pseudothurmannia ohmi Zone
This zone corresponds to the interval comprised betweenthe first appearance datum of the index species and thelower boundary of the Barremian Stage. It was introduced
by Hoedemaeker and Leereveld (1995) to replace the Pseu-dothurmannia angulicostata Zone. In our opinion (Companyet al., 2002, 2003, 2005a), the Ps. ohmi Zone can be subdi-vided into three subzones characterised by the successivefirst appearance datums of Pseudothurmannia ohmi, Ps.mortilleti - here considered to be a senior synonym of P.catulloi - and Ps. picteti. This subdivision was also incorpo-rated into the latest versions of the Lower Cretaceous stan-dard zonation (Hoedemaeker et al., 2003; Reboulet et al.,2006).
In the western High Atlas we have identified this zone atthe bottom of several of the sections sampled. Although am-monites are rare, we did collect Ps. ohmi (Fig. 7A) at thebase of the interval and, slightly higher, a characteristic asso-ciation composed of Ps. mortilleti and Ps. pseudomalbosi(Fig. 7B). Accompanying these species of Pseudothurmanniaare a few specimens of Plesiospitidiscus subdifficilis, Abrytu-sites neumayri and Paraspiticeras sp. (Fig. 7C) which occurthroughout the interval.
In sections where the Ps. mortilleti beds have been identi-fied, these are directly overlain by deposits of Barremianage, there being no record of strata corresponding to the Ps.picteti Subzone (see below).

Fig. 5. Lithology, ammonite distribution and zonation of the Addar section.
14 M. Company et al. / Cretaceous Research 29 (2008) 9e26
4.2. Taveraidiscus hugii Zone and the Hauterivian/Barremian boundary
The base of the Barremian Stage is currently defined(Rawson, 1996) by the first occurrence of Taveraidiscus hu-gii, which is also the index of the lowest zone of the stage.
The suitability of T. hugii as a marker has recently been ques-tioned by Vermeulen (2003), who regarded this species ashaving been generally misinterpreted (see also Hoedemaekeret al., 2003). Nevertheless, we consider differences betweenthe lectotype of T. hugii (see Ooster, 1860, pl. 24, fig. 10)and other specimens originally attributed to this species by

Fig. 6. Stratigraphic distribution of the most significant ammonite species in the Barremian of the western High Atlas. Mediterranean standard zonation after Hoe-
demaeker et al. (2003); sequence stratigraphy after Hardenbol et al. (1998), as modified by Arnaud (2005).
15M. Company et al. / Cretaceous Research 29 (2008) 9e26
Ooster not to be so marked as maintained by Vermeulen, anddo not agree with a separate taxonomic treatment. Moreover,as implicitly shown by Busnardo et al. (2003), all of Ooster’soriginal syntypes must have come from the same stratigraphicinterval, e.g., a few beds, corresponding to the basal Barre-mian, in the highest part of the Veveyse de Chatel-St-Denissection.
In any case, neither T. hugii nor any other holcodiscid hasbeen found in the lowest beds of Barremian age in the westernHigh Atlas. The assemblages collected from these beds aremainly composed of Emericiceras koechlini (Fig. 7K), Acrio-ceras tabarelli and Torcapella barremensis, accompanied byrare specimens of Discoidellia favrei, Barremites dimbovicior-ensis, ‘Barremites’ boutini, Lytoceras densifimbriatum, Para-spiticeras sp. and Hamulinites sp.
According to our unpublished data from the Betic Cordil-lera, E. koechlini and A. tabarelli, which might in fact consti-tute the dimorphic pair of a single biological species, do notappear at the very base of the T. hugii Zone, but somewhathigher, slightly below the middle of the zone. As these twospecies occur in most of the Moroccan sections immediatelyabove the Ps. mortilleti beds, the existence of a regional hiatusspanning the uppermost Hauterivian (Ps. picteti Subzone) andthe lower part of the T. hugii Zone can be inferred. Only in theImsouane section, which represents the most distal and deep-est part of the basin, are there sediments that might correspond
to this age interval, but unfortunately so far they have yieldedno ammonites (Fig. 3).
Torcapella barremensis appears to be an endemic species,which could be the earliest member of the genus. In fact, ex-cept for a single record from the uppermost T. hugii Zone insouth-east France (Vermeulen, 2003), all other species ofthis genus have been recorded from higher levels.
Rare specimens of D. favrei and B. dimboviciorensis havebeen collected from the lowest beds of this interval, and donot range into its upper part. This agrees with their range inthe Mediterranean region, where they disappear at the baseof the Psilotissotia colombiana Subzone (sensu Vermeulen,2003), i.e., well below the top of the T. hugii Zone.
4.3. Kotetishvilia nicklesi Zone
The base of this unit is defined by the first appearance da-tum of the index species (Fig. 7D). This species, although notvery abundant, has been consistently recorded from most ofthe sections studied. In all cases, the thickness of the strati-graphic interval attributable to this zone is quite reduced(only two or three beds). As there is no sign of condensation,it can be concluded that this zone is only partially representedin the western High Atlas.
Co-occurring with the index species in this interval aresome taxa already present in the previous zone. Among these,

16 M. Company et al. / Cretaceous Research 29 (2008) 9e26

17M. Company et al. / Cretaceous Research 29 (2008) 9e26
T. barremensis is still common, whereas E. koechlini, A. tab-arelli, Hamulinites sp. and Paraspiticeras sp. are much rarer.Other species found at these levels are Subpulchellia oehlerti,Holcodiscus thomeli, Almohadites camelinus (Fig. 7E. F), Bar-remites hemiptychus, Davidiceras cf. potieri, Phylloceras pon-ticuli and Lytoceras sp. Except for B. hemiptychus, which isrelatively common in some of the more southerly sections,these taxa are only represented by very few specimens. Never-theless, the presence of A. camelinus and D. cf. potieri ishighly significant because, in the Mediterranean region, thesespecies have been recorded only from the upper part of the K.nicklesi Zone, or from higher levels (Vermeulen, 2003, 2004;Company et al., 2006). This implies that, taking into accountthe incompleteness of the stratigraphic record as mentionedabove, only the upper part of the zone would be representedin the western High Atlas.
4.4. Nicklesia pulchella Zone
The lower boundary of this zone is defined by the firstr ap-pearance datum of Nicklesia pulchella (Fig. 7GeJ). This inter-val is one of the most characteristic in the Barremian of thewestern High Atlas, being well developed throughout the areaand yielding abundant and well-preserved ammonite faunas.
Assemblages from this zone are largely dominated by theindex species, which represents more than 80% of the totalfauna. Most of the other components are holdover taxa fromlower levels, which become extinct in this zone, such as K.nicklesi, B. hemiptychus, D. cf. potieri, T. barremensis andA. tabarelli, together with some long-ranging forms such asPh. ponticuli, Hamulinites sp. and Paraspiticeras sp. All thesespecies are represented by very few specimens, except for T.barremensis, which is still common in the lower beds of thisunit, but is replaced by Torcapella suessi (Simionescu) to-wards the top. Apart from the latter and the first specimensof Moutoniceras nodosum (Fig. 7N), also appearing at thesame level, the upper part of the N. pulchella Zone which usu-ally corresponds to a distinct, thick-bedded limestone interval,is virtually barren of ammonites.
4.5. Kotetishvilia compressissima Zone
The appearance of T. suessi and M. nodosum at the top of theN. pulchella Zone marks the beginning of a major faunal turnoverwhich continues into the overlying K. compressissima Zone. Thisaffects practically all ammonite groups present in the westernHigh Atlas. Thus, among the pulchelliids, Kotetishvilia compres-sissima (Fig. 7M), the first appearance datum of which definesthe base of this zone, completely replaces rich populations ofN. pulchella from the previous zone. In this respect, it should
Fig. 7. Late Hauterivian-Early Barremian ammonites from the western High Atla
Zone. B, Pseudothurmannia pseudomalbosi, X.AS.15.1, Assaka, Ps. ohmi Zone. C,
X.AD.92.1, Addar, basal N. pulchella Zone. E, F, Almohadites camelinus, X.AH.45.
basal N. pulchella Zone. I, J, Nicklesia pulchella, X.SB.21.17, Sidi M’Bark, N. pu
‘Melchiorites’ rumanus, X.TN.18.2, Tamri, C. darsi Zone. M, Kotetishvilia comp
nodosum, X.TN.10.52, Tamri, K. compressissima Zone. All are natural size, excep
be pointed out that specimens recorded by Vermeulen (2003,pl. 36, figs. 2e6, 9, 10) as coming from the base of the K. com-pressissima Zone and attributed by him to N. pulchella are hereregarded as quite typical morphotypes of K. compressissima.Other pulchelliids occurring in this zone are Kotetishvilia di-dayana (Fig. 8C, D), which characterises a level towards the mid-dle of the zone recognisable in several of the sections studied,and Heinzia communis (Fig. 8J, K) and Kotetishvilia changar-nieri (Fig. 8I), which appear in the upper part of the zone.
Coinciding with the base of the K. compressissima Zone,holcodiscids show a conspicuous taxonomic diversification(Company et al., 2006). Although representatives of this fam-ily are very rare in the western High Atlas, some specimens ofHolcodiscus fallax, H. perezianus (Fig. 8A, B) and Avramidis-cus gastaldianus have been found near this level. Holcodiscuscaillaudianus appears slightly higher, in association with Kdidayana.
Desmoceratids are mainly represented by T. suessi and Bar-remites vocontius, which also appear at the base of the zone. Inaddition, some specimens of Subtorcapella deckei and ‘Mel-chiorites’ rumanus (Fig. 7L) occur in the upper part of the unit.
Among the heteromorphs, M. nodosum, Dissimilites dissi-milis, ‘Toxoceras’ annulatum and Hamulinites varusensis arerelatively common throughout this interval.
4.6. Coronites darsi Zone
Originally defined (Vermeulen, 1996) as the interval be-tween the first appearance datums of Coronites darsi and ofHeinzia sartousiana, its extent was subsequently restricted(Vermeulen, 1998) to include only the lower part of that inter-val, i.e., between the first appearance datums of C. darsi and ofH. sayni. This modified C. darsi Zone was incorporated intothe Lyon version of the standard zonation (Hoedemaekeret al., 2003) to replace the M. moutonianum Zone.
More recently, Vermeulen (2003) further separated thehighest portion of the C. darsi Zone and named it the Holco-discus uhligi Zone, and equated the Lower/Upper Barremianboundary with the base of this new unit. Both proposalswere adopted by the Kilian Group for the latest version ofthe standard zonation (Reboulet et al., 2006). In our opinion,however, these changes are not fully justified. First, Holcodis-cus uhligi is not a well-known species; it forms part of a com-plex group of finely ribbed holcodiscids developing betweenthe K. compressissima Zone up to the lower part of theT. vandenheckii Zone. Moreover, the stratigraphic provenanceof the holotype is uncertain, as it comes from condensed levelsin Crimea. Finally, the ammonite assemblage which character-ises the H. uhligi Zone has so far only been identified in theAngles section (Vermeulen, 2003, 2005). These circumstances
s, Morocco. A, Pseudothurmannia ohmi, X.AH.32.1, A€ıt Hamouch, Ps. ohmi
Paraspiticeras sp., X.TN.2.14, Tamri, T. hugii Zone. D, Kotetishvilia nicklesi,4, A€ıt Hamouch, K. nicklesi Zone. G, H, Nicklesia pulchella, X.TN.7.18, Tamri,
lchella Zone. K, Emericiceras koechlini, X.TN.3.8, Tamri, T. hugii Zone. L,
ressissima, X.MS.40.1, Imsouane, K. compressissima Zone. N, Moutoniceras
t for E, F which are� 2.

18 M. Company et al. / Cretaceous Research 29 (2008) 9e26

19M. Company et al. / Cretaceous Research 29 (2008) 9e26
severely restrict the use of this zone as a standard unit, and forthis reason we prefer not to use it, but retain the C. darsi Zoneas in the Lyon version of the standard zonation, i.e., corre-sponding to the interval between the first appearance datumsof C. darsi and of Toxancyloceras vandenheckii.
Coronites darsi (Fig. 8E) is a very rare species in the west-ern High Atlas. As an alternative, we use Moutoniceras mou-tonianum (Fig. 8F) to characterise this zone. Typicalspecimens of this taxon are relatively common in the sectionsstudied and, as shown by Company et al. (2004) and Vermeu-len (2005), the first appearance datum of M. moutonianum vir-tually coincides with that of C. darsi.
Moutoniceras moutonianum is accompanied in the lowerand middle part of the zone by numerous specimens of D. dis-similis, B. vocontius and ‘M.’ rumanus and by much rarer C.darsi, K. changarnieri, T. suessi, Parasaynoceras horridum,Holcodiscus gr. diversecostatus, Holcodiscus aff. perezianusand Silesites vulpes. Heinzia caicedi (Fig. 8G, H) is quite com-mon in the middle part of the zone, characterising a level iden-tified in most of the sections studied.
Moutoniceras moutonianum becomes very rare in the upperpart of the zone, where it is replaced by ‘Barrancyloceras’maghrebiense (Fig. 8L, M). This is an endemic species whichwas held to be related to boreal crioceratitids by Immel(1978), but which we tentatively consider to be one of the ear-liest hemihoplitids (sensu Vermeulen, 2000, 2001). Ammo-nites are abundant in this interval, but assemblages are oflow diversity. Apart from ‘B.’ maghrebiense and the last spec-imens of M. moutonianum, only D. dissimilis, B. vocontius and‘M.’ rumanus occur. The presence of the two last-named spe-cies, which in the Betic Cordillera do not range up into the T.vandenheckii Zone (Company et al., 2004), enables us to attri-bute this interval unambiguously to the M. moutonianum Zone.
4.7. Toxancyloceras vandenheckii Zone
This zone corresponds to the interval between the first ap-pearance datum of Toxancyloceras vandenheckii (Fig. 8N),which also marks the base of the Upper Barremian, and thatof Heinzia sartousiana. In the current standard zonation(Reboulet et al., 2006), this unit has been subdivided intoa lower Heinzia sayni Subzone and an upper Barrancylocerasbarremense Subzone.
The lower part of the zone is poorly represented in thewestern High Atlas. In fact, it has only been identified in theImsouane section on the basis of the occurrence of an impov-erished ammonite assemblage with a single specimen of Hein-zia sayni accompanied by the last representatives of the genusMoutoniceras and some desmoceratids. In other sections,strata corresponding to this stratigraphic interval have eitheryielded no ammonites or are entirely missing.
Fig. 8. Barremian ammonites from the western High Atlas, Morocco. A, B, HolcoKotetishvilia didayana, X.TN.10.33, Tamri, K. compressissima Zone. E, Coronites d
X.SB.46.1, Sidi M’Bark, C. darsi Zone. G, H, Heinzia caicedi, X.MS.49.17, Imsoua
pressissima Zone. J, K, Heinzia communis, X.TK.45.5, Takouwit, K. compressissi
Zone. M, ‘Barrancyloceras’ maghrebiense, X.OG.R.23, Tafadna, C. darsi Zone
Zone. All are natural size, except for E which is� 2.
The B. barremense Subzone has, in contrast, been identified inseveral of the sections studied as a more or less condensed intervalcharacterised by the co-occurrence of T. vandenheckii and Barran-cyloceras barremense (Fig. 9A). Other taxa common at this levelare ‘Acrioceras’ breistrofferi, Heinzia pallinii (Fig. 9B, C),Kotetishvilia sauvageaui and Barremites sp., whereas Acantholy-toceras longispinum and Haplobrancoceras sp. are much rarer.
4.8. Heinzia sartousiana Zone
The base of this zone is defined by the first appearancedatum of the index species and its upper boundary coincideswith the first appearance datum of Hemihoplites feraudianus.Following a proposal by Vermeulen (1996), this zone hasbeen subdivided in the current standard zonation into two sub-zones: a lower Heinzia sartousiana Subzone and an upperHeinzia provincialis Subzone.
Hoedemaeker et al. (1993) and Delanoy (1995, 1997, 1998)placed within this zone the Camereiceras limentinum Horizon,a guide level first identified at the base of the Urgonian lime-stones in the Vercors Massif (Arnaud-Vanneau et al., 1976)and subsequently recognised at other localities in south-eastFrance (Thieuloy, 1979; Delanoy, 1990, 1995). More recently,Vermeulen (2003) questioned this interpretation and assignedthe C. limentinum Horizon to the upper part of his H. sayniZone (i.e., equivalent to the T. vandenheckii Zone), arguingthat the first occurrence of C. limentinum preceded that of H.sartousiana at some of those localities. However, as Vermeulenhimself acknowledged, the first local occurrence of H. sartousi-ana could correspond to the beginning of its acme rather than toits real appearance. In the Betic Cordillera, Camereiceras lime-ntinum is a very rare species, but Janusites janus, another typicalelement of the C. limentinum Horizon, is common and has in-variably been found in association with Heinzia sartousiana(Company et al., 2004, unpublished data). Therefore, it can beconcluded that the C. limentinum Horizon very likely correlateswith the lower part of the H. sartousiana Zone.
In the western High Atlas, deposits assigned to this zonehave only been recognised in some of the most distal sections(i.e., Imsouane, Tamri and Addar). They occur as a thin inter-val between the B. barremense beds and the sandy sedimentsof the Bouzergoun Formation. Fossils present at this level arepoorly preserved, showing clear signs of reworking and con-densation. In fact, the ammonite assemblage recorded is com-posed of a mixture of taxa belonging to both the H. sartousianaand H. provincialis Subzones. Thus, according to the data of Ver-meulen (2003, 2005) and Company et al. (2004), Kotetishviliabrevicostata, Janusites cf. janus and Camereiceras sp. would becharacteristic of the lower half of the zone, whereas Heinziaprovincialis (Fig. 9D), Pachyhemihoplites contei (Fig. 9E) andP. gerthi (Fig. 9F) would be typical of its upper part. Other less
discus perezianus, X.AH.51.1, A€ıt Hamouch, K. compressissima Zone. C, D,
arsi, X.AH.55.1, A€ıt Hamouch, C. darsi Zone. F, Moutoniceras moutonianum,
ne, C. darsi Zone. I, Kotetishvilia changarnieri, X.TK.45.1, Takouwit, K. com-
ma Zone. L, ‘Barrancyloceras’ maghrebiense, X.MS.R.9, Imsouane, C. darsi
. N, Toxancyloceras vandenheckii, X.MS.71.30, Imsouane, T. vandenheckii

Fig. 9. Late Barremian ammonites from western High Atlas, Morocco. A, Barrancyloceras barremense, X.MS.71.35, Imsouane, T. vandenheckii Zone. B, C, Hein-zia pallinii, X.MS.71.4, Imsouane, T. vandenheckii Zone. D, Heinzia provincialis, X.TN.25.6, Tamri, H. sartousiana Zone. E, Pachyhemihoplites contei,
X.TN.25.22, Tamri, H. sartousiana Zone. F, Pachyhemihoplites gerthi, X.TS.23.1, Tamri, H. sartousiana Zone. All are natural size.

21M. Company et al. / Cretaceous Research 29 (2008) 9e26
diagnostic taxa present at this level are Melchiorites cassidoidesand Barrancyloceras cf. hammatoptychum.
4.9. The uppermost Barremian
The H. sartousiana Zone is the highest one recorded fromthe Taboulouart Formation; no ammonites have been found inthe overlying Bouzergoun Formation. Therefore, it has notbeen possible to identify the three uppermost Barremian am-monite zones, i.e., those of Hemihoplites feraudianus, Imeritesgiraudi and Martelites sarasini. Records of ammonites charac-teristic of these zones in earlier literature may be due to mis-identification. In particular, this is the case with relativelyfrequent records of species of Heteroceras (Kilian and Gentil,1907; Roch, 1930; Ambroggi, 1963; Wiedmann et al., 1978,1982); these presumably refer to specimens of Moutoniceras.Similar misinterpretations led Witam (1998) to define the Im-souane Formation, which he regarded as being the lateralequivalent of the Bouzergoun Formation in the central part ofthe basin, and which he dated as Late Barremianeearliest Ap-tian on the basis of the occurrence of Melchiorites falloti andPseudohaploceras aff. matheroni in the middle part of thisunit, and Toxoceratoides sp. in its upper part. In fact, these am-monites could be reinterpreted as ‘Melchiorites’ rumanus, Da-vidiceras sp. and ‘Acrioceras’ breistrofferi and/or related forms,respectively. This would imply that the Imsouane Formation ac-tually corresponded to the interval between the K. compressis-sima and T. vandenheckii zones, and that it can not bedifferentiated from the Taboulouart Formation. As a furtherconsequence, the Tankourimt Formation, created by Witam(1998) for sandy sediments overlying the Imsouane Formation,would merely be an equivalent of the Bouzergoun Formation.
5. Taxonomic composition e palaeobiogeograpic andpalaeoecologic considerations
As stated above, the Essaouira-Agadir Basin was situatedalong the southern margin of the Mesozoic Central Atlantic,very close to the western end of the Mediterranean Tethys.
Table 1
Faunal composition (number of specimens, plus percentages) of ammonite assemb
Taveraidiscushugii Zone
Kotetishvilianicklesi Zone
Nicklesiapulchella
Zone
Kotetishcompres
Zone
Lytoceratina 1 (0.84%) 1 (1.41%) d d
Phylloceratina d 1 (1.41%) 1 (0.21%) 2 (0.66
Ancyloceratidae 73 (61.34%) 12 (16.90%) 20 (4.23%) 84 (27.7
Hemihoplitidae d d d d
Leptoceratoididae 2 (1.68%) 5 (7.04%) 8 (1.69%) 25 (8.25
Paraspiticeras 3 (2.52%) 1 (1.41%) 2 (0.42%) 1 (0.33
Hamulinidae d d d d
Ptychoceratidae d d d d
Macroscaphitidae d d d d
Desmoceratidae 39 (32.77%) 30 (42.25%) 54 (11.42%) 86 (28.3
Pulchelliidae 1 (0.84%) 18 (25.35%) 388 (82.03%) 94 (31.0
Holcodiscidae d 3 (4.23%) d 10 (3.30
Silesitidae d d d 1 (0.33
Total 119 71 473 303
According to this palaeogeographic location, the ammonite as-semblages show a distinctly Mediterranean character. In fact,only two species (out of about 65) recorded from our samplingof the Barremian of the western High Atlas are unknown fromthe Mediterranean area, namely T. barremensis and ‘B.’ magh-rebiense, which appear to be endemic. Nevertheless, despitetheir patent affinity with Mediterranean faunas, the Moroccanassemblages display certain peculiar features. They show, ingeneral, a lesser diversity than assemblages from the westernMediterranean (Subbetic domain and Vocontian Basin), eventhough they all developed in the same latitudinal belt. More-over, some ammonite groups (such as phylloceratids, lytocer-atids, leptoceratoids, hamulinids, ptychoceratids,macroscaphitids, holcodiscids and silesitids), which may con-stitute a substantial part of Barremian Mediterranean assem-blages (Cecca, 1998) are either very rare or absent in thewestern High Atlas (Table 1).
These differences can, in part, be explained by the fact thatthe Essaouira-Agadir Basin corresponded to a platform area,whereas most of the Mediterranean faunas have been recordedfrom distal, pelagic habitats. In particular, this is the case withthe extreme paucity of Phylloceratina and Lytoceratina(<0.5% of total ammonite fauna), usually considered tohave been inhabitants of mesopelagic, deep-water environ-ments, where they can be the main components of ammoniteassemblages (Cecca, 1998; Company et al., 2005a).
In contrast Ancyloceratina are very common in the sectionsstudied, being by far the dominant group at certain strati-graphic levels. They are mostly represented by the familiesAncyloceratidae and Hemihoplitidae. To ancyloceratids, weassign the large-sized genera Emericiceras, Moutonicerasand Toxancyloceras, together with the small Acrioceras andDissimilites, which presumably represent their microconchcounterparts. All of them are strongly ornamented ancylo-cones with a U-shaped adult body chamber; they have been in-terpreted either as epipelagic vertical migrants (Seilacher andGunji, 1993) or planktonic drifters (Westermann, 1996). In anycase, they seem to have been eurytopic organisms, frequentboth on platforms and in deep basins (although the very large
lages for each of the zones and for the Barremian Stage as a whole
viliasissima
Coronitesdarsi Zone
Toxancylocerasvandenheckii
Zone
Heinziasartousiana
Zone
Barremian
1 (0.28%) d d 3 (0.19%)
%) d d d 4 (0.25%)
2%) 64 (17.70%) 33 (24.63%) 11 (10.28%) 297 (18.93%)
100 (27.62%) 41 (30.60%) 74 (69.15%) 215 (13.70%)
%) d d d 40 (2.55%)
%) d d d 7 (0.44%)
d d d d
d d d d
d 1 (0.75%) d 1 (0.06%)
8%) 159 (43.92%) 34 (25.37%) 3 (2.80%) 405 (25.81%)
2%) 33 (9.12%) 25 (18.66%) 19 (17.76%) 578 (36.84%)
%) 4 (1.10%) d d 17 (1.08%)
%) 1 (0.28%) d d 2 (0.13%)
362 134 107 1569

22 M. Company et al. / Cretaceous Research 29 (2008) 9e26
adult macroconchs, common in the western High Atlas, aregenerally confined to shallow environments) and they have,as a group, a very wide palaeobiogeographic distribution.
The above-mentioned ancyloceratids range from the T. hu-gii Zone up to the T. vandenheckii Zone. They are progres-sively replaced in the lower part of the Upper Barremian byhemihoplitids, represented in the western High Atlas by thegenera Barrancyloceras, Camereiceras, Janusites and Pachy-hemihoplites. We have provisionally referred the endemicspecies ‘B.’ maghrebiense to this family on account of its crio-conic coiling, but its affinities remain unclear. Several speciesattributed to the genus Hemihoplites have been recorded frommany different palaeogeographic areas (Kakabadze, 1994;Kakabadze et al., 2004), but we follow Cecca (1998) in con-sidering hemihoplitids as a group to be confined to the Medi-terranean Tethys and adjacent regions, where they arecommon both on platforms and in deep basins.
Leptoceratoids are relatively rare in our sections, wherethey are represented only by the genus Hamulinites. Thesesmall, loosely coiled, heteromorph ammonites seem to becommoner in the Mediterranean region, especially in its pe-lagic areas, where they may be the predominant componentsof the fauna in certain organic-rich beds (Rieber, 1977; Vası-�cek and Wiedmann, 1994; Lukeneder, 2003; Company et al.,2005a). Nevertheless, a biostratinomic bias as a result of pref-erential destruction of their weak shells cannot be completelyruled out to explain the low frequency of leptoceratoids in theshallower western Moroccan environments.
Paraspiticeras is a minor element of the Barremian faunasas well. This peculiar ammonite, with advolute coiling, de-pressed and rapidly enlarging whorls, and blunt ribs with lat-eral tubercles, markedly differs from other Ancyloceratina.Although it has often been linked to the Aptian chelonicera-tids, its real phylogenetic relationships have yet to be resolved.Paraspiticeras has also been reported from southern Europeand Argentina, and interpreted as having had a passive plank-tonic mode of life (Company et al., 2005a). The large size ofsome Moroccan specimens (up to 80 cm in diameter) suggestsa preference in adults for proximal environments.
The families Hamulinidae, Ptychoceratidae and Macrosca-phitidae constitute a group of related heteromorph ammonitescharacterised by a quadrilobate suture with a bifid lateral lobe.Except for a single specimen of Acantholytoceras longispinumfrom the T. vandenheckii Zone (Tamri section), this groupseems to be absent from the Barremian of the western HighAtlas. These families are, in contrast, relatively common, al-though never dominant, on the platforms and in deep basinsof the Mediterranean Tethys and have also been reportedfrom areas as distant as California and Japan.
The suborder Ammonitina is represented in the sectionsstudied by the families Desmoceratidae, Pulchelliidae, Holco-discidae and Silesitidae. Desmoceratids make up, similar toMediterranean assemblages, a considerable percentage of thetotal ammonite fauna, being common throughout the intervalstudied, except for the H. sartousiana Zone, where they arecomparatively rarer. However, the genus Barremites, whichcan be predominant in outer neritic and pelagic settings of
the Mediterranean Tethys, plays only a secondary role in theMoroccan associations. In contrast, the oxyconic, simple-sutured genus Torcapella, which is rare in European pelagic sec-tions, becomes the most characteristic desmoceratid in theshallower western Moroccan Barremian, being by far the com-monest one throughout the interval between the T. hugii andK. compressissima zones. By analogy, in the C. darsi Zone, ‘ Mel-chiorites’ rumanus is commoner than Barremites vocontius.
Pulchelliids have a very wide, near-Pantethyan distribution.In fact, their presence or absence was used by Hoedemaeker(1990) to define the boundaries between the Boreal andTethyan domains during the Barremian. They are also verycommon in the Essaouira-Agadir Basin (>35% of total ammo-nite fauna), but their stratigraphic distribution is uneven. Thus,they are barely represented in the T. hugii Zone (only onespecimen of Discoidellia favrei), whereas assemblages fromthe N. pulchella Zone are overwhelmingly dominated by theindex species. Most of the species in this family are eurytopic,which, together with their discoconic to oxyconic shells andsimple sutures, points to a nektonic habitat in the upper partof the water column (Company et al., 2005a). Nevertheless,some species, such as K. didayana, H. communis and H. cai-cedi - all of them present in our sections - are nearly restrictedto shallow environments (see also Vermeulen, 1998, 2003). Incontrast, some of the early members of the group, e.g., thegenera Discoidellia and Psilotissotia, which have more com-plex sutures, seem to have preferred deeper waters, as substan-tiated by their rarity in the Essaouira-Agadir Basin and otherplatform areas and their relatively high frequency in epio-ceanic settings, such as the Umbria-Marche Apennines (Ceccaet al., 1994; Cecca, 1998).
Some previous authors (Kilian and Gentil, 1907; Roch,1930) drew attention to the absence of Holcodiscidae andSilesitidae in the Barremian of the western High Atlas. Eventhough we managed to collect some specimens belonging tothese groups, they really are uncommon; only seventeen hol-codiscid and two silesitid specimens were found. Silesitids ap-pear to be invariably rare in shallow environments (Cecca,1998). However, the low frequency of holcodiscids stronglycontrasts with their abundance in the Mediterranean Tethys(Vası�cek, 2006), especially in the outer neritic areas of itsnorthern margin (Prebetic, Provence platform, Fore-Balkan,Crimea, etc.), but also well represented along the African mar-gin (Algeria, Tunisia). Moreover, some Barremian holcodis-cids have sporadically been reported from as far afield asColombia, Mexico and Japan.
In summary, despite its evidently Mediterranean nature,the Barremian ammonite fauna from the western High Atlasshows some peculiar features. Some of these, such as the pau-city of certain groups (lytoceratids, phylloceratids, lepto-ceratoids and silesitids), the high frequency of large-sizedancyloceratids, the occurrence of certain coarsely ornamentedpulchelliids, and the predominance of the genus Torcapellaamong desmoceratids, are most probably linked to the neriticcharacter of the Moroccan successions. This explanation,however, is not sufficient to account for the rarity of holcodis-cids and the near-absence of hamulinids, ptychoceratids and

23M. Company et al. / Cretaceous Research 29 (2008) 9e26
macroscaphitids, which are common, and even frequent, inplatform environments of the Mediterranean Tethys. As nogeographic barrier can be invoked, only strictly ecologicalfactors which are difficult to evaluate (currents, water temper-ature, food supply, etc.) might have prevented massive disper-sion into and colonization of the Essaouira-Agadir Basin bythese ammonite groups.
6. Sequence stratigraphy interpretation
As noted above, the stratigraphic successions of the westernHigh Atlas Barremian are, in general, rather incomplete, in-cluding discontinuities, hiatuses and condensed levels. These,and other features of the stratigraphic record, can be inter-preted in terms of sequence stratigraphy when comparingour biostratigraphic succession with the sequence-chronostra-tigraphic framework drawn by Hardenbol et al. (1998), and re-cently modified by Arnaud (2005) (see Fig. 6). Thus, the Ps.ohmi and Ps. mortilleti subzones, recognised in several sec-tions at the base of the Taboulouart Formation, would corre-spond to the TST and early HST of sequence Ha6, since themaximum flooding surface of this sequence - represented inmany pelagic settings of the western Mediterranean Tethysby the anoxic Faraoni Level - has been accurately correlatedwith the base of the Ps. mortilleti Subzone (Cecca et al.,1994; Baudin et al., 1999; Company et al., 2003). These levelsare, in most cases, directly overlain by sediments dated to themiddle and upper portions of the T. hugii Zone, which corre-late with the TST and HST of sequence Ha7. As sequenceboundary Ha7 is located within the Ps. picteti Subzone, thisimplies that the late HST of sequence Ha6 and the LST of se-quence Ha7 are absent from most of the Essaouira-Agadir Ba-sin. However, this interval could be present, in the most distalpart of the basin (Imsouane section), corresponding to theammonite-barren interval mentioned above.
Sequence Barr1 is also incomplete. Its LST, which corre-lates roughly with the top of the T. hugii Zone and the lowerpart of the K. nicklesi Zone, is missing. In contrast, the TSTand HST of this sequence, which correspond to the upperpart of the K. nicklesi Zone and the lower part of the N. pul-chella Zone, are very well documented, coinciding with a pe-riod of maximum flooding in the basin during the Barremian,as shown by the record of N. pulchella at localities such asAzour, Takoucht and Askouti, situated far to the east of thestudy area.
The thick-bedded limestone interval of the upper part of theN. pulchella Zone, recognised throughout the study area (seeabove), is interpreted as representing the LST of sequenceBarr2. The K. compressissima Zone and the lower and middleportion of the C. darsi Zone would correspond to the TST andHST of sequence Barr2 and the whole of sequence Barr3. Itshould be pointed out, however, that these two sequences can-not be easily individualised in the sections studied. In fact,Arnaud (2005) acknowledged that the significance of sequenceBarr3 (BA2 in his terminology) was unclear.
Sequence boundary Barr4 is drawn at the base of a thicklimestone bed set, nearly coinciding with the first occurrence
of ‘B.’ maghrebiense. The LST of this sequence, which wouldcorrespond to the ‘ B’. maghrebiense horizon and the lowerpart of the T. vandenheckii Zone, has been documented, moreor less completely, in all the sections studied, except for Tamri,where it is missing. Its correlative sea level fall would havecaused a certain isolation of the basin, as suggested by the oc-currence of endemic forms and the poorly diversified assem-blages recorded from this interval. The TST and maximumflooding surface of this sequence are represented by thestrongly condensed deposits of the B. barremense horizon andthe H. sartousiana Zone. Nouidar and Chellai (2002) consid-ered the lower member of the Bouzergoun Formation to repre-sent wave-dominated deltaic sediments associated with theHST of the same sequence. However, the base of the Bouzer-goun Formation clearly represents an erosive unconformity,as the age of the immediately underlying levels at the top ofthe Taboulouart Formation varies between latest Early Barre-mian (‘B’. maghrebiense horizon, at Sidi M’Bark) and earlyLate Barremian (H. sartousiana Zone) elsewhere.
7. Conclusions
Our analysis of the ammonite distribution in ten selected,bed-by-bed sampled sections of the Taboulouart Formationhas enabled the establishment of a detailed faunal successioncovering the stratigraphic interval between the uppermostHauterivian to the lower Upper Barremian. All standard zonescomprised in this interval (i.e., from base to top, the Ps. ohmi,T. hugii, K. nicklesi, N. pulchella, K. compressissima, C. darsi,T. vandenheckii and H. sartousiana zones) have been identi-fied, thus confirming the correlation potential of most ofthe events defining and characterising these zones. More-over, some biohorizons of regional interest have also beenrecognised.
Two significant hiatuses and a condensed level of regionalextent have been detected for the first time. The first hiatusspans the uppermost Hauterivian (Ps. picteti Subzone) andthe lowermost Barremian (base of the T. hugii Zone). Theother affects the top of the T. hugii Zone and the base of theK. nicklesi Zone. Both are correlated with third-order sea levelfalls. The condensed level, encompassing the B. barremensehorizon (upper part of the T. vandenheckii Zone) and the entireH. sartousiana Zone, corresponds to a transgressive/maximumflooding interval.
Finally, concerning taxonomic composition, the ammonitefauna displays a clearly Mediterranean character, being dom-inated largely by pulchelliids, desmoceratids, ancyloceratidsand hemihoplitids (Table 2). The scarcity of ‘pelagic’ ammo-nites (phylloceratids, lytoceratids, silesitids and leptocera-toids), and the relatively high frequency of large-sizedancyloceratids, coarsely ribbed pulchelliids and the genus Tor-capella are features in common with shallow Mediterraneansuccessions. In contrast, the paucity of holcodiscids and thenear-absence of certain heteromorph groups (hamulinids, pty-choceratids and macroscaphitids), together with the occur-rence of a few endemic taxa, are peculiar features that areproof of the existence of specific environmental conditions.

Table 2
List of species mentioned in the text
Abrytusites neumayri (Haug)
Acantholytoceras longispinum (Uhlig)
‘Acrioceras’ breistrofferi Sarkar
Acrioceras tabarelli (Astier)
Almohadites camelinus (d’Orbigny)
Audouliceras sp.
Avramidiscus gastaldianus (d’Orbigny)
Barrancyloceras barremense (Kilian)
Barrancyloceas cf. hammatoptychum (Uhlig)
‘Barrancyloceras’ maghrebiense (Immel)
‘Barremites’ boutini (Matheron)
Barremites difficilis (d’Orbigny)
Barremites dimboviciorensis Breskovski
Barremites hemiptychus (Kilian)
Barremites vocontius (Lory and Sayn)
Camereiceras limentinum (Thieuloy)
Coronites darsi Vermeulen
Crioceras (Ancyloceras) aff. zitteli Uhlig
Davidiceras cf. potieri (Matheron)
Discoidellia favrei (Ooster)
Dissimilites dissimilis (d’Orbigny)
Emericiceras koechlini (Astier)
Hamulinites sp.
Hamulinites varusensis (d’Orbigny)
Haplobrancoceras sp.
Heinzia caicedi (Karsten)
Heinzia communis (Burgl)
Heinzia pallinii (Cecca)
Heinzia provincialis (d’Orbigny)
Heinzia sartousiana (d’Orbigny)
Heinzia sayni Hyatt
Hemihoplites feraudianus (d’Orbigny)
Heteroceras astierianum (d’Orbigny)
Holcodiscus caillaudianus (d’Orbigny)
Holcodiscus gr. diversecostatus (Coquand)
Holcodiscus fallax (Matheron)
Holcodiscus perezianus (d’Orbigny)
Holcodiscus thomeli Company, FTzy, Sandoval and Tavera
Holcodiscus uhligi (Karakasch)
Imerites giraudi (Kilian)
Janusites janus (Thieuloy)
Kotetishvilia brevicostata (Kotetishivili)
Kotetishvilia changarnieri (Sayn)
Kotetishvilia compressissima (d’Orbigny)
Kotetishvilia didayana (d’Orbigny)
Kotetishvilia nicklesi (Hyatt)
Kotetishvilia sauvageaui (Hermite)
Lytoceras sp.
Lytoceras densifimbriatum (Uhlig)
Martelites sarasini (Rouchadze)
Melchiorites cassidoides (Uhlig)
Melchiorites falloti (Kilian)
‘Melchiorites’ rumanus (Kilian)
Moutoniceras moutonianum (d’Orbigny)
Moutoniceras nodosum (d’Orbigny)
Nicklesia pulchella (d’Orbigny)
Pachyhemihoplites contei Delanoy
Pachyhemihoplites gerthi (Sarkar)
Paraspiticeras gr. percevali (Uhlig)
Paraspiticeras sp.
Parasaynoceras horridum (d’Orbigny)
Phylloceras ponticuli (Rousseau)
Plesiospitidiscus subdifficilis (Karakasch)
Pseudohaploceras aff. matheroni (d’Orbigny)
Pseudothurmannia angulicostata (d’Orbigny)
Pseudothurmannia catulloi (Parona)
Table 2 (continued )
Pseudothurmannia mortilleti (Pictet and de Loriol)
Pseudothurmannia ohmi (Winkler)
Pseudothurmannia picteti (Sarkar)
Pseudothurmannia pseudomalbosi (Sarasin and Schondelmayer)
Pulchellia dumasiana (d’Orbigny)
Silesites vulpes (Matheron)
Subpulchellia oehlerti (Nickles)
Subtorcapella deckei (Kilian and Reboul)
Taveraidiscus hugii (Ooster)
Torcapella barremensis (Roch)
Torcapella suessi (Simionescu)
Toxancyloceras vandenheckii (Astier)
‘Toxoceras’ annulatum Sarkar
24 M. Company et al. / Cretaceous Research 29 (2008) 9e26
Acknowledgements
We thank Dr Ph. J. Hoedemaeker and an anonymous ref-eree, as well as Associate Editor Dr J.W.M Jagt, for reviewingthe manuscript and for their valuable comments. We are alsograteful to our colleagues Antonio P. Jimenez and LuisO’Dogherty for photographic assistance. This work has beenco-financed by Project CGL2005-02500 (Spanish Ministry ofEducation and Science) and Research Group RNM-178 (Juntade Andalucıa).
References
Ambroggi, R., 1963. Etude geologique du versant meridional du Haut Atlas
occidental et de la plaine du Souss. Notes et Memoires du Service Geolo-
gique 157, 1e321.
Ambroggi, R., Breistroffer, M., 1959. Stratigraphie du Cretace du Haut Atlas
occidental (Sud marocain). In: Kellum, L.B. (Ed.), El sistema Cretacico:
un symposium sobre el Cretacico en el hemisferio occidental y su correla-
cion mundial. Congreso Geologico Internacional, XX Sesion, Mexico,
1956, vol. 2, pp. 33e39.
Arnaud, H., 2005. Sequence stratigraphy interpretation. Geologie Alpine,
Serie Colloques et excursions 7, 174e179.
Arnaud-Vanneau, A., Arnaud, H., Thieuloy, J.P., 1976. Bases nouvelles pour la
stratigraphie des calcaires urgoniens du Vercors (Massifs subalpins septen-
trionaux, France). Newsletters on Stratigraphy 5, 143e159.
Baudin, F., Bulot, L.G., Cecca, F., Coccioni, R., Gardin, S., Renard, M., 1999.
Un equivalent du ‘‘Niveau Faraoni’’ dans le Bassin du Sud-Est de la
France, indice possible d’un evenement anoxique fini-hauterivien etendu
a la Tethys mediterraneenne. Bulletin de la Societe geologique de France
170, 487e498.
Behrens, M., Krumsiek, K., Meyer, D.E., Schaffer, A., Siehl, A., Stets, J.,
Thein, J., Wurster, P., 1978. Sedimentationsablaufe im Atlas-Golf (Kreide
Kustenbecken Marocco). Geologische Rundschau 67, 424e453.
Busnardo, R., Charollais, J., Weidmann, M., Clavel, B., 2003. Le Cretace in-
ferieur de la Veveyse de Chatel (Ultrahelvetique des Prealpes externes;
canton de Fribourg, Suisse). Revue de Paleobiologie 22, 1e174.
Canerot, J., Cugny, P., Peybernes, B., Rahhali, I., Rey, J., Thieuloy, J.P., 1986.
Comparative study of the Lower and Mid-Cretaceous sequences on differ-
ent Maghrebian shelves and basinsdtheir place in the evolution of the
North African Atlantic and Neotethysian margins. Palaeogeography Palae-
oclimatology Palaeoecology 53, 213e232.
Cecca, F., 1998. Early Cretaceous (pre-Aptian) ammonites of the Mediterra-
nean Tethys: palaeoecology and palaeobiogeography. Palaeogeography,
Palaeoclimatology, Palaeoecology 138, 305e323.
Cecca, F., Marini, A., Pallini, G., Baudin, F., Begouen, V., 1994. A guide-level
of the uppermost Hauterivian (Lower Cretaceous) in the pelagic succes-
sions of Umbria-Marche Apennines (Central Italy): the Faraoni Level.
Rivista Italiana di Paleontologia e Stratigrafia 99 (for 1993), 551e568.

25M. Company et al. / Cretaceous Research 29 (2008) 9e26
Choubert, G., Faure-Muret, A., 1962. Evolution du domaine Atlasique depuis
les temps paleozo€ıques. In: Durand-Delga, M. (Ed.), Livre a la memoire du
Professeur Paul Fallot. Societe geologique de France, Paris, pp. 447e527.
Company, M., Sandoval, J., Tavera, J.M., 2002. Ammonite bioevents and zo-
nation of the uppermost Hauterivian in the Betic Cordillera (SE Spain).
Documents des Laboratoires de Geologie de Lyon 156, 83e84.
Company, M., Sandoval, J., Tavera, J.M., 2003. Ammonite biostratigraphy of
the uppermost Hauterivian in the Betic Cordillera (SE Spain). Geobios 36,
685e694.
Company, M., Sandoval, J., Tavera, J.M., 2004. El Barremiense de Fontcalent
y L’Alcoraia. Geo-Temas 7, 217e221.
Company, M., Aguado, R., Sandoval, J., Tavera, J.M., Jimenez de Cisneros, C.,
Vera, J.A., 2005a. Biotic changes linked to a minor anoxic event (Faraoni
Level, latest Hauterivian, Early Cretaceous). Palaeogeography, Palaeocli-
matology, Palaeoecology 224, 186e199.
Company, M., FTzy, I., Sandoval, J., Tavera, J.M., 2006. Deitanites n. g. and
other related ammonites. Their significance within the family Holcodisci-
dae (Lower Cretaceous, Mediterranean region). Neues Jahrbuch fur Geolo-
gie und Palaontologie, Monatshefte 2006 (1), 1e14.
Company, M., Sandoval, J., Tavera, J.M., 2005b. Barremian ammonite biostra-
tigraphy in the western High Atlas (Morocco). In: Godet, A., Mort, H.,
Linder, P., Bodin, S. (Eds.), Scientific Program and Abstracts of the 7th In-
ternational Symposium on the Cretaceous, Neuchatel, 2005, pp. 61e62.
Delanoy, G., 1990. Camereiceras nov.gen. (Ammonoidea, Ancyloceratina) du
Barremien superieur du Sud-Est de la France. Geobios 23, 71e93.
Delanoy, G., 1995. Les zones a Feraudianus, Giraudi et Sarasini du Barremien
superieur de la region stratotypique d’Angles-Barreme-Castellane (Sud-Est
de la France). Geologie Alpine, Memoire Hors-Serie 20 (for 1994),
279e319.
Delanoy, G., 1997. Biostratigraphie haute resolution du Barremien superieur
du Sud-Est de la France. Comptes Rendus de l’Academie des Sciences
de Paris, Sciences de la Terre et des Planetes 325, 689e694.
Delanoy, G., 1998. Biostratigraphie des faunes d’ammonites a la limite
Barremien-Aptien dans la region d’Angles-Barreme-Castellane. Etude par-
ticuliere de la famille des Heteroceratidae Spath, 1922 (Ancyloceratina,
Ammonoidea). Annales du Museum d’Histoire Naturelle de Nice 12 (for
1997), 1e270.
Duffaud, F., Brun, L., Plauchut, B., 1966. Le Bassin du Sud-ouest Marocain.
In: Reyre, D. (Ed.), Bassins sedimentaires du littoral africain. Association
des Services geologiques africains, Paris, pp. 5e12.
Ettachfini, M., 1991. Le Valanginien de l’Atlas atlantique (Maroc): stratigra-
phie et ammonitofaune. Strata, Memoire 15, 1e177.
Ettachfini, M., 2004. Les ammonites neocomiennes dans l’Atlas atlantique
(Maroc): biostratigraphie, paleontologie, paleobiogeographie et paleoeco-
logie. Strata, Memoire 43, 1e223.
Ettachfini, M., Company, M., Rey, J., Taj-Eddine, K., Tavera, J.M., 1998. Le
Valanginien du bassin de Safi (Maroc atlantique) et sa faune d’ammonites.
Implications paleobiogeographiques. Comptes Rendus de l’Academie des
Sciences de Paris, Sciences de la Terre et des Planetes 327, 319e325.
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., de Graciansky, P.C.,
Vail, P.R., 1998. Mesozoic and Cenozoic sequence chronostratigraphic
framework of European basins. In: de Graciansky, P.C., Hardenbol, J.,
Jacquin, T., Vail, P.R. (Eds.), Mesozoic and Cenozoic sequence stratigra-
phy of European Basins. Society of Economic Paleontologists and Miner-
alogists Special Publication 60, 3e13.
Hoedemaeker, P.J., 1990. The Neocomian boundaries of the Tethyan Realm
based on the distribution of ammonites. Cretaceous Research 11,
331e342.
Hoedemaeker, P.J., Cecca, F., Avram, E., Company, M., Delanoy, G., Erba, E.,
Ettachfini, M., Faraoni, P., Kakabadze, M.V., Landra, G., Marini, A.,
Memmi, L., Pallini, G., Rawson, P.F., Ropolo, P., Sandoval, J.,
Tavera, J.M., Vasicek, Z., 1995. Report on the 3rd international Workshop
on the standard Lower Cretaceous ammonite zonation of the Mediterra-
nean region. Memorie descrittive della Carta geologica d’Italia 51,
213e215.
Hoedemaeker, P.J., Company, M., Aguirre-Urreta, M.B., Avram, E.,
Bogdanova, T.N., Bujtor, L., Bulot, L., Cecca, F., Delanoy, G.,
Ettachfini, M., Memmi, L., Owen, H.G., Rawson, P.F., Sandoval, J.,
Tavera, J.M., Thieuloy, J.P., Tovbina, S.Z., Vasicek, Z., 1993. Ammonite
zonation for the Lower Cretaceous of the Mediterranean region; basis
for the stratigraphic correlations within IGCP-Project 262. Revista espa-~nola de Paleontologıa 8, 117e120.
Hoedemaeker, P.J., Leereveld, H., 1995. Biostratigraphy and sequence stratig-
raphy of the Berriasianelowest Aptian (Lower Cretaceous) of the Rıo
Argos succession, Caravaca, SE Spain. Cretaceous Research 16, 195e230.
Hoedemaeker, P.J., Reboulet, S., Aguirre-Urreta, M.B., Alsen, P., Aoutem, M.,
Atrops, F., Barragan, R., Company, M., Gonzalez-Arreola, C., Klein, J.,
Lukeneder, A., Ploch, I., Raisossadat, N., Rawson, P.F., Ropolo, P.,
Vasicek, Z., Vermeulen, J., Wippich, M.G.E., 2003. Report on the 1st
International Workshop of the IUGS Lower Cretaceous Ammonite Work-
ing Group, the ‘Kilian Group’ (Lyon, 11 July 2002). Cretaceous Research
24, 89e94.
Immel, H., 1978. Die Crioceratiten (Ancyloceratina, Ammonoidea) des med-
iterranen und borealen Hauterive-Barreme (Unterkreide). Palaeontograph-
ica A163, 1e185.
Kakabadze, M.V., 1994. On biogeography of some Lower Cretaceous ammo-
nites. In: Pallini, G. (Ed.), Proceedings of the 3rd International Symposium
‘‘Fossili, Evoluzione, Ambiente’’, Pergola, 1990. Palaeopelagos, Special
Publication 1, 203e208.
Kakabadze, M.V., Hoedemaeker, P.J., Bogdanova, T.N., Sharikadze, M.Z.,
2004. On the Barremian-Early Albian biogeography (by ammonites) of
Colombia. Scripta Geologica 128, 515e558.
Kilian, W., 1888. Description geologique de la Montagne de Lure (Basses-
Alpes). Masson, Paris, 458 pp.
Kilian, W., Gentil, L., 1907. Sur les terrains cretaces de l’Atlas occidental mar-
ocain. Comptes Rendus hebdomadaires des Seances de l’Academie des
Sciences de Paris 144, 49e51.
Lemoine, P., 1905. Quelques resultats d’une mission dans le Maroc occidental.
Comite du Maroc, Paris, 223 pp.
Lukeneder, A., 2003. The Karsteniceras level: dysoxic ammonoid beds within
the Early Cretaceous (Barremian, northern Calcareous Alps, Austria).
Facies 49, 87e100.
Nouidar, M., Chellai, E.H., 2002. Facies and sequence stratigraphy of a Late
Barremian wave-dominated deltaic deposit, Agadir Basin, Morocco.
Sedimentary Geology 150, 375e384.
Ooster, W.A., 1860. Catalogue des cephalopodes fossiles des Alpes suisses.
IVeme partie. Nouveaux Memoires de la Societe helvetique des Sciences
naturelles 18, 1e160.
Rawson, P.F., 1996. The Barremian Stage. Bulletin de l’Institut Royal des
Sciences Naturelles de Belgique, Sciences de la Terre 66 (Supplement),
25e30.
Reboulet, S., Hoedemaeker, P.J., Aguirre-Urreta, M.B., Alsen, P., Atrops, F.,
Baraboshkin, E.Y., Company, M., Delanoy, G., Dutour, Y., Klein, J.,
Latil, J.L., Lukeneder, A., Mitta, V., Mourgues, F.A., Ploch, I.,
Raisossadat, N., Ropolo, P., Sandoval, J., Tavera, J.M., Vasicek, Z.,
Vermeulen, J., 2006. Report on the 2nd International meeting of the
IUGS Lower Cretaceous Ammonite Working Group, the ‘‘Kilian Group’’
(Neuchatel, Switzerland, 8 September 2005). Cretaceous Research 27,
712e715.
Rey, J., Canerot, J., Peybernes, B., Taj-Eddine, K., Rahhali, I., Thieuloy, J.P.,
1986. Le Cretace inferieur de la region d’Essaouira: donnees biostratigra-
phiques et evolutions sedimentaires. Revue de la Faculte des Sciences de
Marrakech, Numero special 2, 413e439.
Rey, J., Canerot, J., Peybernes, B., Taj-Eddine, K., Thieuloy, J.P., 1988. Lith-
ostratigraphy, biostratigraphy and sedimentary dynamics of the Lower Cre-
taceous deposits on the northern side of the western High Atlas (Morocco).
Cretaceous Research 9, 141e158.
Rieber, H., 1977. Eine Ammonitenfauna aus der oberen Maiolica der Breggia-
Schlucht (Tessin/Schweiz). Eclogae Geologicae Helvetiae 70, 777e787.
Roch, E., 1930. Etudes geologiques dans la region meridionale du Maroc
occidental. Notes et Memoires du Service geologique 9, 1e542.
Roch, E., 1950. Histoire stratigraphique du Maroc. Notes et Memoires du Ser-
vice geologique 80, 1e437.
Seilacher, A., Gunji, P.Y., 1993. Morphogenetic countdowns in heteromorph
shells. Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen
190, 237e265.

26 M. Company et al. / Cretaceous Research 29 (2008) 9e26
Thieuloy, J.P., 1979. Matheronites limentinus n. sp. (Ammonoidea) espece-
type d’un horizon-repere barremien superieur du Vercors meridional
(Massif subalpin francais). Geobios, Memoire special 3, 305e317.
Vası�cek, Z., 2006. A remarkable assemblage of Early Barremian ammonites in
the Central Western Carpathians (Butkow Quarry, Slovakia). Acta Geolog-
ica Polonica 56, 421e440.
Vası�cek, Z., Wiedmann, J., 1994. The Leptoceratoidinae: small heteromorph
ammonites from the Barremian. Palaeontology 37, 203e239.
Vermeulen, J., 1996. Nouvelle biozonation du Barremien basee sur la famille
des Pulchelliidae (Ammonoidea). Geologie Alpine 71 (for 1995),
119e211.
Vermeulen, J., 1998. Biohorizons ammonitiques dans le Barremien du Sud-Est
de la France (de la zone a Hugii a la zone a Sartousiana). Geologie Alpine
73 (for 1997), 99e117.
Vermeulen, J., 2000. Nouvelles donnees sur les repartitions stratigraphiques,
les evolutions et les classifications de trois familles d’ammonites du
Cretace inferieur. Geologie Alpine 75 (for 1999), 123e132.
Vermeulen, J., 2001. Nouvelles donnees sur l’origine, l’evolution et la classi-
fication des Hemihoplitidae Spath, 1924 (Ammonoidea, Ancylocerata-
ceae). Annales du Museum d’Histoire Naturelle de Nice 15 (for 2000),
91e101.
Vermeulen, J., 2003. Etude stratigraphique et paleontologique de la famille des
Pulchelliidae (Ammonoidea, Ammonitina, Endemocerataceae). Geologie
Alpine, Memoire H. S. 42 (for 2002), 1e333.
Vermeulen, J., 2004. Ammonites nouvelles ou peu connues du Barremien med-
iterraneen. Annales du Museum d’Histoire Naturelle de Nice 18 (for 2003),
57e109.
Vermeulen, J., 2005. Boundaries, ammonite fauna and main subdivisions of the
stratotype of the Barremian. Geologie Alpine, Serie Colloques et excursions
7, 147e173.
Westermann, G.E.G., 1996. Ammonoid life and habitat. In: Landman, N.,
Tanabe, K., Davis, R.A. (Eds.), Ammonoid Paleobiology. Plenum Press,
New York, pp. 607e707.
Wiedmann, J., Butt, A., Einsele, G., 1978. Vergleich von marokkanischen
Kreide-Kustenaufschlussen und Tiefseebohrungen (DSDP): Stratigraphie,
Palaoenvironment und Subsidenz an einem passiven Kontinentalrand.
Geologische Rundschau 67, 454e508.
Wiedmann, J., Butt, A., Einsele, G., 1982. Cretaceous stratigraphy, environ-
ment, and subsidence history at the Moroccan continental margin. In:
von Rad, U., Hinz, K., Seibold, E. (Eds.), Geology of the Northwest
African continental margin. Springer, Berlin, pp. 366e395.
Wippich, M.G.E., 2001. Die tiefe Unter-Kreide (Berrias bis Unter-Hauterive)
im Sudwestmarokkanischen Becken: Ammomnitofauna, Bio- und Sequenz-
stratigraphie. Unpublished PhD thesis, Ruhr-Universitat, Bochum, 142 pp.
Wippich, M.G.E., 2003. Valanginian (Early Cretaceous) ammonite faunas
from the western High Atlas, Morocco, and the recognition of western
Mediterranean ‘standard’ zones. Cretaceous Research 24, 357e374.
Witam, O., 1998. Le Barremien-Aptien de l’Atlas Atlantique (Maroc): lithos-
tratigraphie, biostratigraphie, sedimentologie, stratigraphie sequentielle,
geodynamique et paleontologie. Strata, Memoire 30, 1e421.
Witam, O., Rey, J., Aadjour, M., Magniez-Jannin, F., Delanoy, G., 1993. Nou-
velles donnees biostratigraphiques et sequentielles sur la serie barremienne
et aptienne du bassin d’Agadir (Maroc). Revue de Paleobiologie 12,
193e202.





























