Embed Size (px)
Citation preview

Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394www.elsevier.nl/locate/palaeo
Bivalve palaeobiogeography and the Hispanic Corridor:time of opening and effectiveness of a proto-Atlantic seaway
Martin Aberhan *Museum fur Naturkunde, Institut fur Palaontologie, Invalidenstr. 43, D-10115 Berlin, Germany
Received 8 December 1999; accepted for publication 19 July 2000
Abstract
The Hispanic Corridor is an embryonic seaway between the eastern Pacific and western Tethyan oceans that hasbeen postulated to have preceded Middle Jurassic drifting and the birth of the Atlantic Ocean by many millions ofyears. In this study the distribution of pectinoid bivalve morphotypes is analysed to examine the origin of the HispanicCorridor and its effectiveness during Early Jurassic times. A comparison of pectinoid faunal similarities, based onsimilarity coefficients, between various regions at opposite ends of the Corridor suggests that it opened progressivelyduring Early Jurassic times. However, with this approach it is difficult to exclude alternative dispersal routes, and todetermine when faunal interchange began. Analysis of the percentage of pectinoid morphotypes that were (1) presentat opposite sides of the Corridor, (2) simultaneously absent in the western Pacific/eastern Tethys and (3) confined torelatively low palaeolatitudes provides evidence that the Hispanic Corridor developed from an effective barrier inearlier Early Jurassic (Hettangian to Sinemurian) times into a filter, allowing the passage of a few morphotypes,during later Early Jurassic (Pliensbachian and Toarcian) times. The apparently two-way faunal exchange through theHispanic Corridor is consistent with the establishment of a megamonsoonal circulation for Pangaea, which may havecaused seasonally changing directions of oceanic surface currents through the Corridor. © 2001 Elsevier Science B.V.All rights reserved.
Keywords: bivalves; Hispanic Corridor; Jurassic; palaeobiogeography; palaeoceanography
1. Introduction Atlantic Ocean a shallow marine connection wasestablished across rifting continental crust linkingthe western end of the Tethyan Ocean with theOne of the most prominent palaeogeographiceastern Pacific (Fig. 1). The time of formationchanges in earth history was initiated by the Earlyof this narrow passageway, called the HispanicMesozoic breakup of the supercontinent Pangaea.Corridor (Smith, 1983), and its subsequent effec-Although there is no direct sedimentological ortiveness for the equatorial dispersal of marinegeophysical evidence, palaeontological data sug-organisms is the subject of continuous debate andgest that prior to drifting and the birth of thewas certainly controlled by both plate-tectonicactivity and eustatic sea-level changes (e.g. Hallam,1983; Smith and Tipper, 1986; Smith et al., 1990;* Fax: +49 30 20938868.
E-mail address: [email protected] (M. Aberhan) Westermann, 1993; Stanley, 1994).
0031-0182/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.PII: S0031-0182 ( 00 ) 00172-3

376 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Fig. 1. Early Jurassic (Pliensbachian) palaeogeography (modified from Barron et al., 1981; Damborenea, 1987) and possible dispersalroutes for marine organisms (modified from Hallam, 1983, 1994). C1=Hispanic Corridor; C2=Viking Corridor; P1, P2=routesaround Pangaea margins; T=trans-Pacific route.
The Early Jurassic, divided into four stages 2. Early Jurassic palaeogeography and thedispersal of marine organisms(Hettangian, Sinemurian, Pliensbachian,
Toarcian), and estimated to span about 26 millionyears (Gradstein et al., 1994), is a critical time During the Early Jurassic, the supercontinent
Pangaea, surrounded by the superoceaninterval in the geological history of the HispanicCorridor. Based on the biogeography of the Early Panthalassa or palaeo-Pacific, was nearly symmet-
rical about the equator. About 56% of the landJurassic bivalve genus Weyla, Damborenea andMancenido (1979) postulated the presence of a was in the northern hemisphere (Parrish, 1993;
Fig. 1). As the supercontinent began to break upshallow seaway connecting the western Tethyanand eastern Pacific oceans as early as Pliensbachian and sea level rose during the Jurassic, several
epeiric seaways became established, separating thetimes. Biogeographical evidence from a variety offossil groups offers support for this hypothesis (see areas of exposed land (e.g. Hallam, 1994). Apart
from the Hispanic Corridor, another seaway,below). In recent years, our knowledge of EarlyJurassic bivalves along the eastern palaeo-Pacific termed the Viking Corridor by Westermann
(1993), opened in Late Pliensbachian time betweenmargin has increased substantially for both westernSouth America (e.g. Damborenea, 1987, 1996; Greenland and Norway and connected the Arctic
and Tethys oceans.Aberhan, 1994) and western North America(Aberhan, 1998a). In the present paper, new data Possible migration routes of shelf faunas
between the eastern Pacific and western Tethyson the distribution of a particular group ofbivalves, the pectinoids, are used to: (1) analyse are indicated in Fig. 1. Apart from utilizing the
Hispanic or Viking corridors, organisms couldthe permeability of the Hispanic Corridor duringthe various stages of the Early Jurassic; (2) investi- have migrated around the periphery of Pangaea,
either by the north or the south, or could havegate if dispersal was preferentially one-way (west–east or east–west) or if a two-way faunal exchange crossed the palaeo-Pacific, either as long-range
dispersal of larvae or via island hopping. Finally,prevailed; and (3) consider Early Jurassic palaeo-ceanography in the light of the results obtained it has been proposed that fossil biotas could have
been transported across the palaeo-Pacific onfrom (2).

377M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
various displaced terranes, which now form large adaptations and are obviously closely related.parts of the North American Cordillera, but origi- Although a classification of taxa in morphotypesnated in more remote positions in the ancient does not reach the acuity of a species-level dataPacific. However, for the bivalve bearing Early set, a much finer taxonomic (and biogeographic)Jurassic terranes a location close to the western resolution is obtained than from genera alone. Itmargin of Pangaea is most plausible (e.g. Aberhan, avoids taxonomic biases introduced by misidenti-1998b and references cited therein). fications and contrasting species concepts of
different authors, and yields a uniform taxonomicconcept in a manageable amount of time withouthaving to revise a whole fauna at the species level.3. Materials and methodsA definition of pectinoid morphotypes, which arenamed after a characteristic representative, is pre-3.1. Early Jurassic pectinoid bivalvessented in Appendix A.
The present analysis is based on the distributionof bivalves of the order Pectinida. Representatives
3.3. The studied areas and temporal frameworkof this sessile group of organisms are fairlycommon and well preserved in the studied areas
The geographic areas selected for the study werewhere they occur in a wide range of environments.lumped into five major geographic units. TheseTheir diversity is well documented in the literature,are: (1) northwestern Europe (Great Britain,and a uniform taxonomic concept can be applied.Sweden, Denmark, Belgium, Luxemburg,What little is known about the larval developmentGermany, Switzerland, northern France); (2) west-of Jurassic pectinoids (Palmer, 1989) and theirern Tethys (SW-France, Italy, Spain, Portugal,modern counterparts (Waller, 1991) suggests thatMorocco, Algeria); (3) western South Americathey possessed planktotrophic larvae, and there-(Colombia, Peru, Chile, Argentina); (4) westernfore depended on oceanic currents for their dis-North American craton (Huayacocotla trough ofpersal (but see Waller, 1993 for an example of aeastern Mexico, western Nevada, northernlecithotrophic-type larval shell in a reef-adaptedCalifornia, cratonal areas of western Canada); andpectinid from the Caribbean). Recently, distribu-(5) western North American terranes (Antimoniotional patterns of pectinoid bivalves proved usefulterrane of NW-Mexico, Wrangellia, Stikinia,in constraining the Early Jurassic tectonic evolu-Quesnellia, Cadwallader terrane). The areas of thetion of western Canadian terranes (Aberhan,last category are crustal fragments that are alloch-1998b, 1999), thus testifying to their high potentialthonous relative to the autochthonous craton, butin geologically orientated palaeobiogeographic
analyses. palaeogeographic evidence suggests proximity tothe North American craton throughout the EarlyJurassic (e.g. Smith and Tipper, 1986; Aberhan,3.2. The morphotype concept1998b). Data for comparison with the westernPacific were included where deemed necessary.Studies on Jurassic bivalve biogeography are
The data (see Appendix B and references citedusually carried out at the generic/subgeneric level.therein) were analysed separately for four timeCompared with species-based data sets, this clearlyintervals corresponding to the four stages ofresults in the loss of potentially useful information.the Early Jurassic: Hettangian, Sinemurian,On the other hand, a sound taxonomic databasePliensbachian and Toarcian. Only records withat species level is not feasible in most cases, especi-illustrations entered the database, provided thatally if large data sets are involved. The morphotypethe specimens were well preserved enough to allowconcept, as applied in the present study, forms aassignment to a morphotype and the age wascompromise. A morphotype groups together
species that exhibit very similar morphological determinable at the stage level. Faunal affinities of

378 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
the various areas to each other are established bycalculating Dice (D) and Jaccard (J ) similaritycoefficients. These coefficients are defined as
D=[2C/(N1 +N2 )]×100
and
J=[C/(N1 +N2−C )]×100
where N1 is the number of taxa in sample 1, N2 isthe number of taxa in sample 2, and C is thenumber of taxa common to both samples.
4. Results
4.1. Comparison of faunal similarities
Dice and Jaccard similarity coefficients showexactly the same temporal trends (e.g. Fig. 2A),and therefore only Dice coefficients are reproducedin Fig. 2B and C.
In order to gain an overall impression of thefaunal similarities between the eastern Pacific andthe western Tethys and adjacent epicontinentalseas of NW-Europe, data for the three easternPacific areas and those for the two European/NW-African areas were combined (Fig. 2A). Thesimilarity of pectinoid bivalves increases steadilyfrom comparatively low values in the Hettangian
Fig. 2. Early Jurassic similarities of pectinoid bivalve morpho-and Sinemurian to intermediate values in thetypes between western Tethys, NW-Europe and various easternPliensbachian and highest values in the Toarcian. Pacific regions. (A) Increasing Dice and Jaccard coefficients
If the data from the various eastern Pacific areas between eastern Pacific and Europe/NW-Africa suggest pro-are compared with the western Tethyan Ocean gressive opening of the Hispanic Corridor. (B) Dice coefficients
between western Tethys and various eastern Pacific areas. (C)(Fig. 2B), pectinoids from the North AmericanDice coefficients between NW-Europe and various easterncraton and those from the terranes show a trendPacific areas.of increasing similarity to the western Tethys with
time. Dice coefficients between South America andthe western Tethys are higher than those for theother two Pacific areas. Values fluctuate around a Hettangian to lower values in the Sinemurian and
Pliensbachian and reaches a peak in the Toarcian.level of 45 from Hettangian to Pliensbachian times,and show a marked increase in the Toarcian. In avery similar way, similarity values between both 4.2. Dispersal of selected morphotypesthe North American craton and the terranes andthose from NW-Europe (Fig. 2C) increase through As a next step we analyse how the distribution
of morphotypes can contribute to the reconstruc-time. Similarity between the South Americancraton and NW-Europe decreases from the tion of the early development of the Hispanic

379M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Corridor. In order to preclude alternative dispersal European areas, this morphotype does not provideany clues concerning the opening of the Hispanicroutes, a taxon must fulfil three criteria, at leastCorridor. Western Pacific records of the other fourduring the early phases of its duration: (1) presencemorphotypes are of the same ages as first occur-on either end of the Hispanic Corridor; (2) simul-rences on opposite sides of the Hispanic Corridor:taneous absence in western Pacific regions; andOxytoma cygnipes (Hettangian of New Zealand,(3) confinement to relatively low palaeolatitudes.Damborenea and Mancenido, 1992); EntoliumOf the 48 morphotypes considered in this study,corneolum (Toarcian of North-East Russia,15 meet criterion (1) (see Appendix A). In orderMilova, 1988); Entolium lunare (Sinemurian ofto evaluate criterion (2), the possible presence ofJapan, Hayami, 1975); and Chlamys valoniensisthese morphotypes in the North-East and Far East(Hettangian of North-East Russia, Milova, 1976;of Russia, in Japan and in New Zealand wasHettangian/Early Sinemurian of New Zealand,explored. A survey of the relevant literature revealsDamborenea and Mancenido, 1992). Because athat seven morphotypes appear to be absent fromdispersal route other than the Hispanic Corridorwestern Pacific regions, at least throughout Earlycannot be excluded, these morphotypes are notJurassic times, and therefore meet criterion (2):considered further here.Placunopsis striatula, Pseudopecten equivalvis,
For the 10 morphotypes that fulfil criteria (1)Camptonectes auritus, Camptonectes subulatus,and (2), the known Early Jurassic palaeolatitudi-Eopecten abjectus, Eopecten velatus and Weylanal ranges are presented in Table 1. Three morpho-meeki. To these may be added, Oxytoma inequi-types (Oxytoma inequivalvis, Placunopsis striatulavalvis, Propeamussium pumilum and Chlamys texto-and Chlamys textoria) were spread over a relativelyria. Although known from western Pacific areasbroad palaeolatitudinal range during the Earlyduring Early Jurassic times (see Efimova et al.,Jurassic, and may have used a variety of alternative1968; Milova, 1976, 1985, 1988; Hayami, 1975;migration routes. The remaining seven morpho-Sey, 1984; Sey and Polubotko in Damboreneatypes, however, seem to be confined to a belt
et al., 1992), all these records are younger thanextending from about 40° north to 45° south of
their first occurrences at opposite ends of theHispanic Corridor. The difference in age is smallestfor Propeamussium pumilum. Its first appearance
Table 1in the Pliensbachian of Europe (Johnson, 1984) isSelected morphotypes of pectinoid bivalves and their Early
followed by apparently contemporaneous occur- Jurassic palaeolatitudinal ranges. N=northern palaeolatitude;rences in the Toarcian of South America and S=southern palaeolatitude. Records from the North-East and
Far East of Russia are not considered as they are commonlyNorth-East Russia. However, as records fromfrom terranes, the Early Jurassic palaeolatitudinal positions ofSouth America are from the Early Toarcian (e.g.which are poorly constrained. For discussion see text. DataHillebrandt and Schmidt-Effing, 1981; Aberhan,based on Aberhan (1994, 1998a,b), Damborenea (1996),
1994), they predate the mid-Toarcian occurrence Damborenea and Mancenido (1979), Johnson (1984),in eastern Russia (Polubotko and Repin, 1988), Trechmann (1923) and unpublished information
and consequently this morphotype is potentiallyMorphotype Palaeolatitudinal rangeuseful in constraining the early history of the
Hispanic Corridor. Oxytoma inequivalvis 60°N–75°SPlacunopsis striatula 70°N–42°SFrom the five morphotypes that do not meetPropeamussium pumilum 35°N–45°Scriterion (2), one (Meleagrinella substriata)Pseudopecten equivalvis 40°N–40°Sappears to have occurred first during the Camptonectes auritus 30°N–35°S
Hettangian in North-East Russia where it is repre- Camptonectes subulatus 40°N–35°SChlamys textoria 70°N–45°Ssented by the species Meleagrinella subolifexEopecten abjectus 35°N–38°SPolubotko (Efimova et al., 1968; Sey andEopecten velatus 35°N–35°SPolubotko in Damborenea et al., 1992). As this isWeyla meeki 40°N–30°S
earlier than oldest records from eastern Pacific and

380 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
the palaeoequator. Their restriction to relatively origination rates of endemics were very high fromLate Sinemurian to Late Pliensbachian timeslow palaeolatitudes, simultaneous presence in
Europe and along the eastern Pacific margin, and (Aberhan and Fursich, 1997, 2000). However, agenerally high percentage of Andean endemicsabsence, or at least stratigraphical restriction to
younger stages, from western Pacific and eastern cannot explain the relatively low similarity ofpectinoid morphotypes in mid-Early Jurassic timesTethys suggest dispersal via the Hispanic Corridor
at various times during the Early Jurassic. between South America and NW-Europe. In fact,the percentages of pectinoid morphotypes that areendemic to either NW-Europe or South Americaare by no means significantly higher during the5. DiscussionSinemurian and Pliensbachian than during theHettangian and Toarcian (Fig. 3).5.1. Interpretation of faunal similarity patterns
This suggests that it is high Hettangian ratherthan low Sinemurian and Pliensbachian similarityThe general trend of pectinoid bivalves is a
progressive increase in similarity between the west- that deviates from the normal trend. Examinationof the six Hettangian morphotypes common toern Americas and Europe during the Early Jurassic
(Fig. 2). The most marked exceptions are South America and NW-Europe reveals that onlytwo, Camptonectes subulatus and Eopecten velatus,Sinemurian and Pliensbachian similarity values
between South America and NW-Europe, which are considered as candidates for dispersal throughthe Hispanic Corridor (see Section 4.2). Since theyare relatively low in comparison with the
Hettangian value (Fig. 2C). A diversity analysis are first recorded from the pre-planorbis bedsand the planorbis Zone respectively, their originsof bivalves from the Andean basins showed that
Fig. 3. Degree of endemism of pectinoid bivalve morphotypes in Early Jurassic time. Data based on Appendix B; absolute age datesfrom Gradstein et al. (1994).

381M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
probably lie in the Triassic (Johnson, 1984). Their data set. By analogy, the pectinoid data presentedherein are compatible with the concept of theoccurrence in South America and Europe mayHispanic Corridor opening progressively duringbe the result of dispersal around southernEarly Jurassic time. The major shortcomings ofGondwanaland or along a circum-Laurasian routeHallam’s approach, however, are that the latitudi-during the Late Triassic ‘hothouse’ climate system.nal range of taxa as well as data from the westernFor these reasons, relatively high Hettangian DicePacific and eastern Tethys are not taken intocoefficients between South America and Europeanaccount. As a consequence, it is impossible toareas are unlikely to reflect migration via thedisentangle the relative importance of high versusHispanic Corridor in earliest Jurassic time.low palaeolatitude migration routes, and to deter-Applying a similar approach, Hallam (1983)mine when the interchange of organisms began.has analysed the similarity of Jurassic bivalve
genera between Europe, North America and SouthAmerica. Rising Simpson similarity coefficients
5.2. Excluding alternative dispersal routesbetween Europe and the western Americas (Fig. 4)are in good agreement with similarity curves of
Whilst a similarity study with only a few regionspectinoid morphotypes (Fig. 2). Hallam’s datafails to test if and to what degree the Hispanicwere interpreted to indicate that some degree ofCorridor was utilized for migration by organisms,isolation persisted between the Americas andanalysis of distributional patterns holds muchEurope as at first only a few organisms filteredmore promise. Because of ample data, establish-through the Hispanic Corridor, and that relativelyment of the presence/absence of pectionoidfree intermigration was established by the Middlemorphotypes on opposite sides of the HispanicJurassic (Hallam, 1983; Smith and Tipper, 1986;Corridor and in the western Pacific was fairlySmith, 1988). These conclusions were essentiallystraightforward. Less information is available fromconfirmed in a recent analysis by Damboreneathe eastern Tethys, the pre-Toarcian bivalve record(2000), who used the same method as Hallam,of which is rather poor or lacking, and from highalthough within a temporally and spatially morepalaeolatitude areas.finely resolved framework and with an updated
Data from high northern and southern areasare considered as particularly important for evalu-ating high latitude routes for dispersal. Althoughthe concept of climatic equability in the Jurassichas been challenged recently (Crowley and North,1991), there is no doubt that temperature gradientsfrom the equator to the poles were distinctly lowercompared with the present (e.g. Hallam, 1998 andreferences cited therein). Still, the geographicdifferentiation of Early Jurassic bivalves intoboreal, austral, bipolar and low latitude forms(Damborenea, 1993, 1996; Aberhan, 1998b, Liuet al., 1998), and the well-known Tethyan–Borealprovinciality among the ammonites (e.g. Hallam,1994), suggest that environmental conditionsvaried enough to produce latitudinally distinctfaunas. Although documented less intensely, theabsence of a pectinoid morphotype from such
Fig. 4. Simpson similarity coefficients of bivalve genera betweenrelatively high palaeolatitude areas as the southernthe western Americas and Europe through time as calculatedAndes and New Zealand in the southern hemi-by Hallam (1983). Absolute age dates from Gradstein et al.
(1994). sphere, and northwestern Canada, Arctic Canada
382 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
and East Greenland in the northern hemisphere, With respect to low palaeolatitude routes, analternative to the Hispanic Corridor is long-rangeis therefore regarded as adequate evidence to
render unlikely its dispersal around southern larval dispersal across the ancestral Pacific. ForPangaean times, tropical trans-oceanic faunal dis-Gondwanaland, northern Laurasia or via the
Viking Corridor. persal was hypothesized both from east to west(e.g. Kristan-Tollmann and Tollmann, 1982), andThe palaeolatitudinal range from 40°N to 45°S,
inferred for those five morphotypes that have from west to east (e.g. the pantropical model ofNewton, 1988; see comments by Smith et al.,apparently utilized the Hispanic Corridor (see
Sections 4.2 and 5.1, Table 1), seems to be fairly 1990). For the five pectinoid morphotypes underdiscussion, the apparent absence in the westernlarge to be called ‘low latitude’, but is compatible
with palaeoclimatic data. In the Early Jurassic Pacific and eastern Tethys renders a trans-Pacificdispersal more unlikely than migration throughterrestrial realm, phytogeographic data suggest
that the transition from warm to cool-temperate the Hispanic Corridor. In addition, it is knownthat open oceans act as effective barriers to dis-conditions in the northern hemisphere took place
at about 45° (Ziegler et al., 1993), and growth persal of many marine invertebrates, and duringEarly Jurassic times the central palaeo-Pacificring studies of fossil wood indicate a broad equato-
rial zone ranging in latitude from approximately ocean was at its widest (Smith and Tipper, 1986;Smith et al., 1990).30°N to 30°S (Creber and Chaloner, 1984).
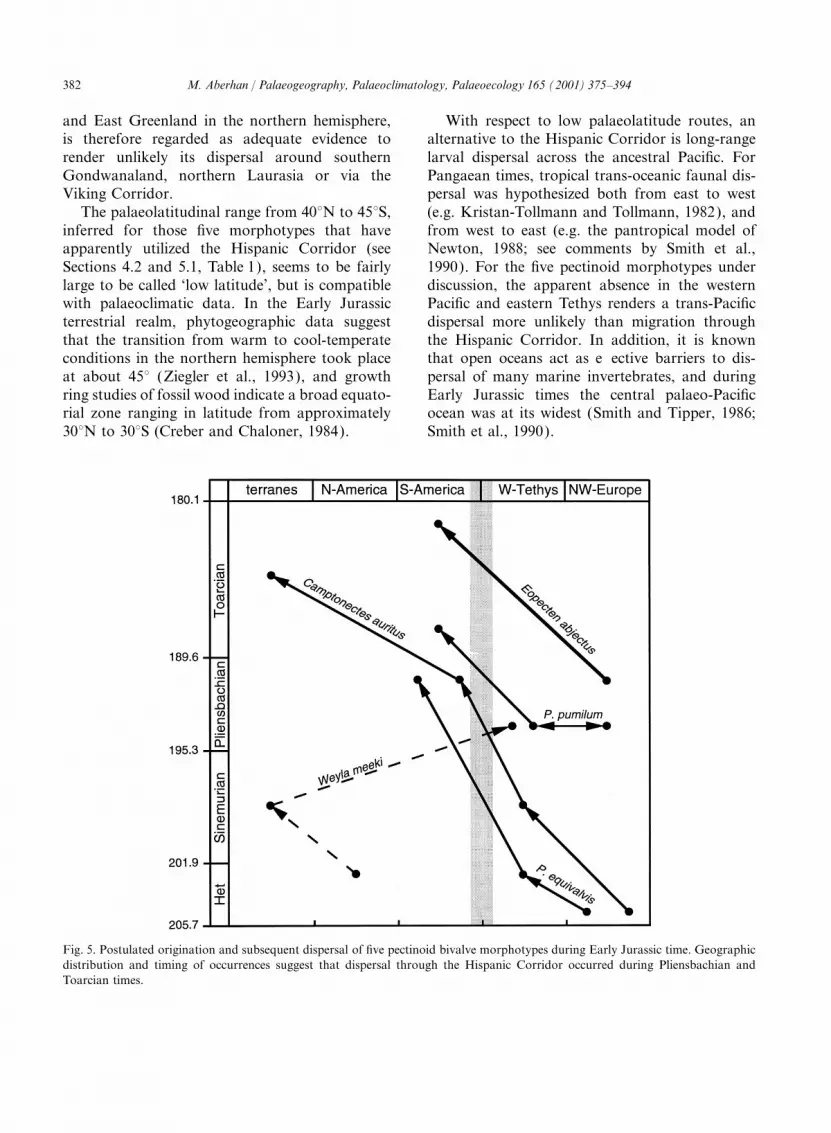
Fig. 5. Postulated origination and subsequent dispersal of five pectinoid bivalve morphotypes during Early Jurassic time. Geographicdistribution and timing of occurrences suggest that dispersal through the Hispanic Corridor occurred during Pliensbachian andToarcian times.

383M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
5.3. Origin of the Hispanic Corridor To put these results in perspective, literaturedata on time and direction of dispersal of variousfossil groups via the Hispanic Corridor, includingThe European and eastern Pacific stratigraphic
ranges of the five morphotypes considered to have calcareous sponges, corals, bivalves, ammonites,belemnites and brachiopods, are summarized inused the Hispanic Corridor indicate that three of
them migrated during Pliensbachian and two Table 2. These data are consistent with the resultsof the present study, and point at an initial openingduring Toarcian times (Fig. 5, Table 2). Even if
it cannot be excluded from the available data in early Pliensbachian time. Only in one casehas an earlier origin been inferred (Stanley andthat a few morphotypes migrated prior to the
Pliensbachian, there is no unequivocal support for Beauvais, 1994). Based on the distribution ofcorals, these authors conclude that the Hispanicthis hypothesis. This argues for an opening of the
Corridor in the Pliensbachian, and suggests that it Corridor may have played an important roleduring Sinemurian time. Their data, however,was not a viable dispersal route for pectinoid
bivalves during earliest Jurassic times. The sim- imply east–west dispersal of two coral species inlate Hettangian time and west–east dispersal ofilarity curve (Fig. 2A) is consistent with a
Pliensbachian origin of the Hispanic Corridor, but two other species later in the early Pliensbachian(Stanley and Beauvais, 1994, fig. 7). Whilst theof course does not prove it (see discussion above).
Table 2List of marine invertebrates bearing on the Early Jurassic origin of the Hispanic Corridor. Het=Hettangian; Plb=Pliensbachian;Toa=Toarcian; E–W=from east to west; W–E=from west to east
Taxon Group Presumed time Direction Referenceof dispersal
Stylothalamia calcareous sponge early Plb E–W Hillebrandt, 1981Phacelostylophyllum rugosum, corals late Het E–W Stanley and Beauvais, 1994Actinastrea planaStylophyllopsis victoriae, corals early Plb W–E Stanley and Beauvais, 1994Actinastrea minimaCamptonectes auritus, bivalve morphotypes Plb E–W this studyPseudopecten equivalvisEopecten abjectus, bivalve morphotypes Toa E–W this studyPropeamussium pumilumWeyla meeki bivalve morphotype Plb W–E this studyWeyla bivalve early Plb W–E Damborenea and Mancenido, 1979, 1988;
Hallam, 1983Lithiotis (=Plicatostylus) bivalve Plb – Loriga and Neri, 1976; Hillebrandt, 1981;
Hallam, 1983; Nauss and Smith, 1988Opisoma bivalve mid Toa E–W Hillebrandt, 1981; Hallam, 1983;
Aberhan and Hillebrandt, 1999Bouleiceras ammonite early Toa – Hillebrandt, 1973Dayiceras/Dubariceras ammonites early Plb – Smith, 1983; Smith and Tipper, 1986Frechiella ammonite mid Toa – Hillebrandt, 1973Leukadiella, Paroniceras ammonites mid Toa – Hillebrandt and Schmidt-Effing, 1981;
Jakobs, 1995Oregonites ammonite Plb – Smith and Tipper, 1986hastitid ancestors of Dicoelitidae belemnites latest Plb or E–W Jeletzky, 1980
earliest ToaThecideaceans brachiopods late Plb E–W Mancenido and Damborenea, 1990;
Baker and Mancenido, 1997Squamiplana (Cuersithyris) brachiopod early Plb – Mancenido and Dagys, 1992

384 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
latter event is consistent with all the other datapresented in Table 2, a late Hettangian age seemssurprisingly old. Even more extreme is the hypoth-esis of a corridor being in existence possibly asearly as the Late Triassic, which was based on thepalaeobiogeography of brachiopods and corals(Sandy and Stanley, 1993; Stanley, 1994 and refer-ences cited therein). Fluctuations in eustatic sea-level and tectonic activity along the corridor are
Fig. 6. Percentage of pectinoid morphotypes that apparentlyregarded as the main control of its effectiveness.utilized the Hispanic Corridor for the first time during the fourDuring Hettangian time, Jurassic eustatic sea-levelstages of the Early Jurassic. Numbers in the diagram indicatewas at its lowest stand, and Late Triassic sea-level the total number of morphotypes per time interval.
was characterized by a major regression (e.g.Hallam, 1992). This is in accord with the LateTriassic to earliest Jurassic stratigraphic record ofthe numerous rift basins along the postulated that also extend into high palaeolatitudes and
western Pacific/eastern Tethys areas obviouslycorridor, which consists of continental sequencescomprising lacustrine and fluvial clastic rocks, could tolerate a broad range of environmental
conditions. As eurytopic taxa they are also theevaporites, coals and basalts (Olsen, 1997). Thesalt deposits apparently formed in evaporitic lakes most likely to spread through a filter, in which
strong fluctuations in environmental parametersand continental playa or sabka environments fedby non-marine waters. Marine sediments, includ- such as temperature and salinity can be expected.
Restriction to low palaeolatitude taxa excludesing salt deposits of a probably marine source, arelimited to the opposite ends of the rift in Spain, these potential travellers, and the number of those
that actually passed through the corridor may bewest of Morocco and in Mexico (Smith andTipper, 1986; Stanley, 1994 and references cited somewhat higher. Nevertheless, such a low percen-
tage of pectinoid morphotypes travelling along thetherein). For these reasons, the idea of a pre-Jurassic to earliest Jurassic marine connection Corridor suggests that it acted as a filter during
Pliensbachian and Toarcian times. This view isseems to be very speculative, and needs to betested rigorously against other models such as supported by the observation that, in contrast to
some shallow water bivalves, Early Jurassic ‘off-trans-Pacific dispersal.shore’ genera, such as Posidonotis and Otapiria,were unable to pass the Corridor (Damborenea,2000, but see discussion on the distribution pattern5.4. Effectiveness of the Hispanic Corridorof Posidonotis in Aberhan and Palfy, 1996).
Studies comparable with the present one, whichReflecting its effectiveness a migration routemay be classified as a corridor if it allows relatively determine the percentage of a fauna that migrated
via the Hispanic Corridor, have not been carriedfree migration, or as a filter if it allows the passageof some organisms but not others. A very effective out so far. But again, faunal similarities of bivalves
(Hallam, 1983; Damborenea, 2000; Figs. 2 and 4)barrier that could only be overcome by chanceevents is called a sweepstake route (Simpson, 1940; are compatible with the hypothesis of a pro-
gressively opening marine connection from EarlyMcKenna, 1973). During Pliensbachian times, 35pectinoid morphotypes are recognised, 8% of to Middle Jurassic times. Both palaeobiogeo-
graphic and geological evidence indicate that awhich apparently passed through the HispanicCorridor (Fig. 6). In the Toarcian (28 morpho- relatively open corridor was in operation, at least
intermittently, in Middle and Late Jurassic timestypes) this number is slightly less (7%). Due to therigorous criteria applied to identify corridor voyag- (e.g. Hallam, 1977, 1983; Westermann, 1977, 1993;
Riccardi, 1991; Boomer and Ballent, 1996).ers, these values may be too low. Widespread taxa

385M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Development from a filter into a corridor is paral- ammonites, transport of eggs may be more impor-tant factors than the ecological requirements ofleled by an overall rise in Hallam’s (1988) eustaticadults.sea-level curve of the Jurassic. Obviously, this
increase in sea-level was not compensated by tec-tonic activity along the rift, and probably both 5.5. A two-way migration routeprocesses have reinforced each other.
The ecological diversity of bivalves, charac- Stratigraphic age relationships provide clues toterized by various life habits and feeding modes, the direction of migrations. From the five morpho-makes this group a potential source of information types considered to have used the Hispanicon the physical conditions existing along the route. Corridor for dispersal, four originated in EuropePectinoid bivalves are an epifaunal suspension- (Fig. 5; Table 2). Only Weyla meeki occurred firstfeeding group, and individual taxa belong to the in the eastern palaeo-Pacific. This indicates thatfree-lying, byssate and cemented guilds. Whilst among pectinoids east–west migration prevailed.Eopecten abjectus shows a strong preference In contrast, for late Toarcian–Aalenian times,for condensed ferruginous oolites, the species Hallam (1983) concluded that bivalve traffic wasPropeamussium pumilum, Camptonectes auritus, largely one-way, as eastern Pacific taxa moved
eastward to occupy habitat space which had beenPseudopecten equivalvis and Weyla meeki werevacated during the Early Toarcian mass extinctionremarkably eurytopic with respect to the substrateof bivalves in Europe. Examination of Table 2,(Johnson, 1984; Aberhan, 1998b). P. pumilum washowever, shows that Early Jurassic migrationalso able to live under variable conditions offrom east to west was at least as common as inoxygen tension and turbulence, and probably hadthe opposite direction. This view of a two-wayan opportunistic adaptive strategy (Johnson, 1984;migration route is corroborated by several bivalveAberhan, 1993); C. auritus could cope with highgenera (including oysters, bakevelliids andphysical stress, oxygen deficiency and various tem-trigoniids), which, according to their knownperatures (Johnson, 1984), and was also reporteddistribution through time, probably migratedfrom the upper brachyhaline salinity regime, closealong the Corridor (Damborenea, 2000). Similarly,to fully marine conditions (e.g. Fursich, 1993);
Westermann and Riccardi (1985) did not recogniseand P. equivalvis was able to tolerate high levelsa preferred direction in the migration of Middleof water energy (Johnson, 1984). Such broadJurassic ammonites through the Corridor.environmental tolerances are additional evidence
that the Corridor was an ecological barrier thatcould only be overcome by eurytopic taxa.Considering the large amounts of evaporites that 6. Biotic dispersal through the Hispanic Corridorhave been deposited in the rifts, it seems likely and Early Jurassic palaeoceanographythat extreme salinities or strongly fluctuating salin-ity values were limiting biotic exchange. This may On an evolutionary time scale, organisms withexplain the low percentage of pectinoids, which a holopelagic life cycle have the potential to occurare at best subordinate faunal components in virtually everywhere, provided their ecologicalsalinity-controlled Mesozoic environments. On requirements are met. Sessile, benthic organismsthe other hand, ammonites apparently used the such as pectinoid bivalves depend more stronglyCorridor from Pliensbachian times onwards on oceanic currents for their dispersal during the(Table 2), although they could not cope with larval stages. Given the apparently two-way faunalpalaeosalinities which deviated from normal exchange through the Hispanic Corridor, oceanicmarine values. In this respect it would be interes- circulation must have been favourable for larvalting to analyse the dispersal of other bivalve groups transport in both directions. In principal, oceanicsuch as the relatively euryhaline oysters. Anyway, circulation is driven by wind stress and by density
variations of seawater. In the following Ithe ecology of bivalve larvae and, in the case of

386 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
speculate on the possible influence of both factors northern summer. As a consequence, the southeaston oceanic circulation through the Hispanic trade winds would have blown across the palaeo-Corridor. equator and, having been diverted to the right by
the Coriolis force, would give rise to a currentflowing from the eastern Pacific to western Tethys.
6.1. Wind-driven circulation Current flow through the Corridor in oppositedirections would have happened semi-annually.
Present-day equatorial current systems in the Such a reversal of the normal trade wind patternopen oceans are a direct result of the trade winds. across the continent is in agreement with changesThese blow at about 45° to the equator and induce of surface wind vectors obtained in numericalwestward flows in the surface water (the north and model simulations of Pangaean climates (seesouth equatorial currents). An eastward flowing models of Kutzbach and Gallimore, 1989;equatorial countercurrent is driven by the hori- Chandler et al., 1992).zontal pressure gradient that is set up when west-ward flowing surface water is blocked by alandmass and the sea-surface slopes up to the 6.2. Density-driven circulationwest.
Predicted equatorial ocean currents of the Early The density of seawater is controlled by varia-Jurassic palaeo-Pacific are similar to those encoun- tions in temperature and salinity. The Mesozoic istered in the modern Pacific (Parrish, 1992). With generally conceived as a time interval when thethe opening of the Hispanic Corridor some temperature distribution was more even thanthroughflow of the equatorial current may have today, so that salinity becomes an important factorintroduced warm Tethyan surface water into the in the density of seawater. It is therefore possiblePacific. However, it is unlikely that during this that vertical circulation in the Mesozoic oceansembryonic stage of the Atlantic Ocean a circum- was driven largely by salinity differences ratherglobal, westward flowing equatorial current was than primarily by temperature as today (e.g. vanfully established.
Andel, 1994). In the case of the Hispanic Corridor,An Early Jurassic equatorial countercurrent,
it is conceivable that a thermohaline circulationflowing eastward through the Hispanic Corridor,was established similar to that between theis not a viable hypothesis. In the case of westwardpresentday Mediterranean and Atlantic Ocean.throughflow of the equatorial current a counter-Evaporation in the Mediterranean Sea in summercurrent would not have existed at all. In contrast,exceeds the influx of freshwater, and high-salinitywith a barrier in place across western Tethys, thesurface water is formed. When this surface waterequatorial countercurrent may have been strongcools in winter it sinks, and flows out at depth(Parrish, 1992). However, at the western end ofinto the Atlantic Ocean, and relatively low-the Hispanic Corridor its driving force, the west–salinity water flows into the Mediterranean ateast difference in sea level across the palaeo-Pacific,the surface.was no longer effective.
While the western end of the Hispanic CorridorBecause of its size and the distribution of landwas close to the equator, its eastern end wasin both hemispheres, a strong monsoonal circula-located in the subtropics at about 25°N during thetion should have existed for Pangaea (e.g.Pliensbachian (Fig. 1). Considering this differenceKutzbach and Gallimore, 1989; Parrish, 1992,in palaeolatitudes and the extension of the1993). During the northern winter, northeast tradeCorridor from one margin of a supercontinent towinds would have generated a westward flowingthe other, it can be expected that palaeoclimatolog-current through the Hispanic Corridor. In analogyical parameters differed at opposite ends of theto the present-day summer monsoonal circulationseaway. Information on palaeoclimate and onin Asia, the intertropical convergence zone would
have shifted considerably northward during the the kind of water masses can be obtained from

387M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
geological data and from conceptual and numerical currents flowing through the Hispanic Corridor,and deserves further testing and modelling. Theclimate models. During the Late Triassic and earli-
est Jurassic, salt deposits in the eastern and central densities of water masses in the equatorial easternPacific and subtropical western Tethys, and theirpart of the rift suggest extreme arid conditions,
which contrast with synchronous lake, playa and seasonal variability, are insufficiently known atpresent, and it is difficult to evaluate how theycoal swamp deposits in basins in the western part
of the rift complex (Hay et al., 1982). The affected oceanic currents within the Corridor.From the available evidence it seems possible thatonly Lower Jurassic marine sediments known at
the western end of the Corridor are in the a density-driven current reinforced a wind-driven,eastward flowing current that was in operationHuayacocotla trough of eastern Mexico, which
Schmidt-Effing (1980) has interpreted as an aulao- during the northern summer.cogen associated with the opening of the Gulf ofMexico. The marine sedimentary fill consists ofshales, siltstones and sandstones of Sinemurianage, which suggest relatively humid conditions. Incontrast, the Early Jurassic carbonate platform 7. Conclusionsdeposits of western Tethys and the abundant EarlyJurassic evaporites in northern Africa and north- Palaeobiogeographic patterns of pectinoid
bivalve morphotypes provide further evidence thateastern North America apparently formed undermore arid conditions. the Hispanic Corridor was in existence during
Early Jurassic times. In order to determine its timeFor Pliensbachian time, conceptual models ofrainfall patterns predict relatively high rainfall in of formation and subsequent effectiveness, the
current method of calculating similarity coefficientsthe equatorial eastern Pacific and moderately lowrainfall in the western Tethys (Parrish et al., 1982; alone seems to be insufficient. The presence of
common taxa in a few areas at opposite sides ofParrish, 1992). Numerical modelling of Pangaeanclimate during the Early Jurassic yielded very high the Corridor does not, in itself, imply a proto-
Atlantic marine connection because several alter-evaporation rates in western Tethys, particularlyduring the northern winter (Chandler et al., 1992). native dispersal routes cannot be ruled out. A
more rigorous approach is to identify the percen-In this model, however, the annually averagedprecipitation minus evaporation values are nega- tage of taxa per time interval that were (1) present
at opposite sides of the Corridor, (2) simulta-tive at both ends of the Corridor, and it is difficultto assess their seasonal variability. Nevertheless, it neously absent in the western Pacific/eastern
Tethys and (3) confined to relatively low palaeolat-is conceivable that, during the Early Jurassic,supersaline surface water formed by evaporation itudes. The results suggest that during Hettangian
and Sinemurian times the future site of thein the western Tethys and sank into deep basins,thereby causing a flow of eastern Pacific surface Hispanic Corridor most likely was an effective
barrier to the spread of marine organisms. Thewater through the Hispanic Corridor into westernTethys. The Corridor was relatively shallow and Hispanic Corridor first opened in the
Pliensbachian and acted as a filter until Toarcianthe floor of the rift system probably had a markedrelief. Because vertical barriers could hinder the times, providing a restricted level of faunal
exchange between the eastern Pacific and westernmovement of saline bottom currents, it is unclearto what extent a bottom return flow of relatively Tethys oceans. Apparently, development into a
marine corridor was not before the Middlehigh salinity carried water from the western Tethysback to the eastern Pacific. Jurassic. Bidirectional faunal exchange through
the Corridor is in agreement with the establishmentIn conclusion, the establishment of a megamon-soonal circulation for Pangaea (cf. Kutzbach and of a megamonsoonal circulation, which may have
caused seasonal alternation of flow directionsGallimore, 1989; Parrish, 1993) offers a plausiblemechanism for seasonally changing directions of within the Corridor.

388 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Acknowledgements Meleagrinella papyria (Quenstedt): shellorthocline, ribs relatively widely spaced.
I wish to thank Bill Hay, Axel von Hillebrandt, 1Meleagrinella substriata (Munster): shell proso-Wolfgang Kiessling and Dave Lazarus for critically cline, with numerous strong radial ribs.
Terquemia arietis (Quenstedt): attachment areareading the manuscript, and Gerd Westermannof right valve relatively large.and an anonymous reviewer for their constructiveTerquemia pectiniformis (Eudes-Deslongchamps):reviews. This study was financially supported by aattachment area of right valve relatively small.grant from the Deutsche Forschungsgemeinschaft1Placunopsis striatula (Oppel ): includes all radi-(Ab 109/1-1), which is acknowledged withally ribbed nominal species of Early Jurassicgratitude.Placunopsis.Propeamussium laeviradiatum ( Waagen): dorsalmargins of right valve extended into horn-like
Appendix A: Definition of morphotypes of Early processes, left valve coarsely ribbed.Jurassic pectinoid bivalves occurring in NW-Europe,
1Propeamussium pumilum (de Lamarck): dorsalthe western Tethys and the eastern palaeo-Pacific margins of right valve extended slightly beyondmargin hinge line, left valve finely ribbed.
Propeamussium patriciae Poulton: shell large, withaxa marked with an asterisk are known from both radial ornament present on both valves.ends of the Hispanic Corridor. Note that the Kolymonectes carlottensis ( Whiteaves): rightdefinition of morphotypes of European Jurassic valve with relatively broad ribs.pectinids and propeamussiids corresponds to the Kolymonectes coloradoensis ( Weaver): right valvespecies concept of Johnson (1984). with very fine radial striae.Otapiria inaequicostata Geyer: shell sub-orbicu- 1Entolium lunare (Roemer): small byssal notchlar, left valve ornamented with ribs of different present in juveniles.orders of strength. 1Entolium corneolum (Young and Bird): byssalOtapiria neuquensis Damborenea: shell sub-ovate, notch absent.radial ornament similar on both valves. Posidonotis semiplicata (Hyatt): includes all mate-Otapiria pacifica Covacevich and Escobar: shell rial referable to this genus.elongated, ornament stronger on right valve than Agerchlamys wunschae (Marwick): ornament ofon left valve. radial riblets and commarginal lamellae, whichOtapiria tailleuri Imlay: shell elongated, radial give rise to a reticulate pattern.ornament more or less equally developed on 1Camptonectes auritus (Schlotheim): sub-orbicu-both valves. lar disc, fine divaricate striae on all parts of disc.Lupherella boechiformis (Hyatt): includes all 1Camptonectes subulatus (Munster): sub-orbicu-material referable to this genus. lar disc, fine divaricate striae restricted to anteriorAnningella carixensis (Cox): includes all material and posterior margins of disc.referable to this genus. Camptonectes obscurus (J. Sowerby): ornamentOxytoma calva (Schlonbach): shell smooth or consisting of radial striae and commarginal lamel-very weakly ribbed. lae, striae restricted to within a few centimetres1Oxytoma inequivalvis (J. Sowerby): left valve of the umbo.with moderately high number of well-developed Camptonectes (Costicamptonectes): refers toprimary ribs and intercalated secondary ribs. Camptonectes (Costicamptonectes) sp. A in1Oxytoma cygnipes (Young and Bird): left valve Aberhan (1998a).with small number of spinose ribs. Canadonectites paucicostatus Aberhan: right andArctotis? frenguellii Damborenea: includes this left valve smooth except for fine radial riblets on
antero-dorsal part of right valve.species and Arctotis sp. A in Aberhan (1998a).

389M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
1Chlamys textoria (Schlotheim): imbricate lamel- 1Weyla meeki Damborenea: shell plano-convex tolae present on the plicae. feebly biconvex, ribs and interspaces of right and1Chlamys valoniensis (Defrance): ornament lack- left valve triangular in cross-section.ing on the plicae. Weyla unca (Philippi): shell clearly biconvex, ribsChlamys pollux (d’Orbigny): presence of long and interspaces of right and left valve triangulartubular spines. in cross-section.1Eopecten abjectus (Phillips): intercalary costae Weyla yukonensis Aberhan: shell biconvex, ribsrapidly gain the same height as original costae, of right valve with more or less flat crests andtwo median costae greatly enlarged and bearing triangular interspaces, ribs of left valve narrow,tubercles. triangular in cross-section and with concaveEopecten hartzi (Rosenkrantz): ornament on left interspaces.valve differentiated into costae and striae, a fewcostae are enlarged.Eopecten spondyloides (Roemer): original costae
Appendix B: Stratigraphic and geographic distri-similar in height, intercalary costae rapidly gainbution of Early Jurassic pectinioid bivalvethe same size as original costae.morphotypes1Eopecten velatus (Goldfuss): ornament on left
valve differentiated into costae and striae, no1=NW-Europe; 2=western Tethys; 3=Southenlarged costae.
America; 4=North American craton; 5=westernOchotochlamys aequistriata Aberhan: ornamentNorth American terranes. [ ]=occurrenceof radial riblets and commarginal lamellae andassumed. Data based on Aberhan (1994 and refer-folds equally developed on right and left valves.ences cited therein), Aberhan (1998a,b and refer-Ochotochlamys bureiensis Sey: style of ribbing onences cited therein), Arkell (1933), Behmel andleft and right valves different.Geyer (1966), Benecke (1905), Berini (1957),Pseudopecten barbatus (J. Sowerby): long spinesBertuletti (1962), Brauns (1871), Bronn (1836),present on right valve.Chapuis and Dewalque (1853), Cossmann (1903),Pseudopecten dentatus (J. de C. Sowerby): discCox (1928, 1936), Damborenea (1987 andflanks high and vertically striated, down-sulcalreferences cited therein), Damborenea (1996),tongueing of commarginal striae, modal numberDamborenea and Gonzalez-Leon (1998), Daresteof plicae 17/18.de la Chavanne (1920), Dechaseaux (1936), Dubar1Pseudopecten equivalvis (J. Sowerby): disc flanks(1948), Fucini (1920), Gardet and Gerard (1946),low, commarginal striae curvilinear.Gemmellaro (1872–1882), Geyer (1973), GoldfussPseudopecten veyrasensis (Dumortier): vertically(1833, 1835), Hallam (1987), Holder (1978, 1990),striated disc flanks, down-sulcal tongueing ofJohnson (1984), Joly (1936), Kuhn (1935, 1936,commarginal striae, modal number of plicae 14.1938), de Lamarck (1819), Lanquine (1929),Radulonectites sosneadoensis (Weaver): includesLentini (1973), Leymerie (1881), Lissajous (1907–all eastern Pacific forms referable to this genus.1912), Oppel (1853, 1856), Pedersen (1986),Weyla alata (von Buch): shell concave–convex toPhillips (1829, 1871), Quenstedt (1851–1852,plano-convex, ribs of right valve with almost flat1856–1857, 1865–1866, 1882–1885), Riccardi et al.crests and concave interspaces, ribs of left valve(1990), Rollier (1915), Schafle (1929), Schlonbachnarrow and with concave interspaces.(1863), Staesche (1926), Terquem (1855),Weyla ayarti (Dubar): right valve small, withTerquem and Piette (1865), Troedsson (1951),only up to six subtriangular main ribs.Vacek (1886), de Verneuil and Collomb (1853),Weyla bodenbenderi (Behrendsen): shell concave–Zieten (1833) and unpublished information fromconvex, ribs of right and left valve relativelythe Huayacocotla trough in eastern Mexico andbroad, number of ribs increasing by intercalation
and branching. the Antimonio terrane of NW-Mexico.

390 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Morphotype Hettangian Sinemurian Pliensbachian Toarcian
Otapiria inaequicostata 3Otapiria neuquensis 3 3Otapiria pacifica 3 3Otapiria tailleuri 4Lupherella boechiformis 4, 5Anningella carixensis 1Oxytoma calva 1 1Oxytoma inequivalvis 1, 2, 3, 4, 5 1, 2, 3, 4, 5 1, [2], 3, [4], 5 1, 2, 3, 4, 5Oxytoma cygnipes 1, 3 1, 4 1, 4, 5 5Arctotis? frenguellii 4 3Meleagrinella papyria 1 1Meleagrinella substriata 1, 5 1, 5 1Terquemia arietis 1 1 2Terquemia pectiniformis 1Placunopsis striatula 1, 2 1, [2], 3, 5 1, [2], 3, 5 1, 2, 3, 4Propeamussium laeviradiatum 1 1, 2Propeamussium pumilum 1, 2 1, 2, 3Propeamussium patriciae 4, 5Kolymonectes carlottensis 5Kolymonectes coloradoensis 4 3Entolium lunare 1, 2 1, 2, 3 1, 2 1, 5Entolium corneolum 3, 5 3, 4, 5 3, [4], 5 1, 2, 3, 4, 5Posidonotis semiplicata 4, 5 3, 5 3, 5Agerchlamys wunschae 3, 4, 5 4, 5 3, 5 5Camptonectes auritus 1 1, 2 [1], 2, 3 1, 2, 3, 5Camptonectes subulatus 1, 3 1, [2], 3, 4 1, 2, [4] 1, 2, 4Camptonectes obscurus 1Camptonectes (Costicamptonectes) 5Canadonectites paucicostatus 5 5Chlamys textoria 1, 2, 3 1, 2, 3, 4, 5 1, 2, 3, 4, 5 1, 2, 3, 5Chlamys valoniensis 1, 2, 3 1, 3, 4, 5Chlamys pollux 1, 2 1, 2Eopecten abjectus 1 1, 3Eopecten hartzi 4, 5 4, 5 3, 4Eopecten spondyloides 1Eopecten velatus 1, 2, 3 1, 2, [3] 1, 2, 3 1, 2, 3Ochotochlamys aequistriata 5 4, 5 4, 5Ochotochlamys bureiensis 4 4Pseudopecten barbatus 1, 2Pseudopecten dentatus 1, 2 [1], 2 1, 2 1, [2]Pseudopecten equivalvis 1, 2 1, 2 1, 2, 3 1, 2, 3Pseudopecten veyrasensis 1, 2 [1], [2] 1, 2Radulonectites sosneadoensis 3, 4, 5Weyla alata 4 3, 4, 5 3, 4, 5 3Weyla ayarti 2Weyla bodenbenderi 3, 5 3, 5 3, 5Weyla meeki 4 4, 5 2, 5 2Weyla unca 4 3, 4, 5 3, 5 3Weyla yukonensis 5 4, 5

391M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Brauns, D., 1871. Der untere Jura im nordwestlichen Deutsch-Referencesland von der Grenze der Trias bis zu den Amaltheenthonenmit besonderer Berucksichtigung seiner Molluskenfauna.
Aberhan, M., 1993. Benthic macroinvertebrate associations on Vieweg & Sohn Braunschweig. 493 pp.a carbonate-clastic ramp in segments of the Early Jurassic Bronn, H.G., 1836. In: Lethaea geognostica. Schweizerbart,back-arc basin of northern Chile (26–29°S). Rev. Geol. Stuttgart pp. 225–480.Chile 20, 105–136. Chandler, M.A., Rind, D., Ruedy, R., 1992. Pangaean climate
Aberhan, M., 1994. Early Jurassic Bivalvia of northern Chile.during the Early Jurassic: GCM simulations and the sedi-
Part I. Subclasses Palaeotaxodonta, Pteriomorphia andmentary record of paleoclimate. Geol. Soc. Am. Bull. 104,
Isofilibranchia. Beringeria 13, 3–115.543–559.
Aberhan, M., 1998a. Early Jurassic Bivalvia of western Canada.Chapuis, F., Dewalque, C., 1853. Description des fossiles des
Part I. Subclasses Palaeotaxodonta, Pteriomorphia andterraines secondaires de la province de Luxembourg. Mem.
Isofilibranchia. Beringeria 21, 57–150.Acad. R. Belge 35 303 pp.
Aberhan, M., 1998b. Paleobiogeographic patterns of pectinoidCossmann, M., 1903. Note sur L’Infralias de la Vendee et des
bivalves and the Early Jurassic tectonic evolution of westernDeux Serres. II. Pelecypodes. Bull. Soc. Geol. Fr., Ser. 4
Canadian terranes. Palaios 13, 129–148.3, 497–545.Aberhan, M., 1999. Terrane history of the Canadian Cordillera:
Cox, L.R., 1928. The Belemnite Marls of Charmouth, a seriesestimating amounts of latitudinal displacement and rotationin the Lias of the Dorset Coast. VI. Gastropod and Lamelli-of Wrangellia and Stikinia. Geol. Mag. 136, 481–492.branch Molluscs. Q. J. Geol. Soc. London 84, 233–245.Aberhan, M., Fursich, F.T., 1997. Diversity analysis of Lower
Cox, L.R., 1936. The Gastropoda and Lamellibranchia of theJurassic bivalves of the Andean Basin and the Pliensbachian-Green Ammonite Beds of Dorset. Q. J. Geol. Soc. London–Toarcian mass extinction. Lethaia 29, 181–195.92, 456–471.Aberhan, M., Fursich, F.T., 2000. Mass origination versus mass
Creber, G.T., Chaloner, W.G., 1984. Influence of environmen-extinction: the biological contribution to the Pliensbachian-tal factors on the wood structure of living and fossil trees.–Toarcian extinction event. J. Geol. Soc. London 157,Bot. Rev. 50, 357–448.55–60.
Crowley, T.J., North, G.R., 1991. Paleoclimatology. OxfordAberhan, M., Hillebrandt, A.v., 1999. The bivalve Opisoma inMonogr. Geol. Geophys. 18, 1–349.the Lower Jurassic of northern Chile. Profil 16, 149–164.
Damborenea, S.E., 1987. Early Jurassic Bivalvia of Argentina.Aberhan, M., Palfy, J., 1996. A low oxygen tolerant East PacificPart 2. Superfamilies Pteriacea, Buchiacea and part of Pecti-flat clam (Posidonotis semiplicata) from the Lower Jurassicnacea. Palaeontographica (A) 199, 113–216.of the Canadian Cordillera. Can. J. Earth Sci. 33, 993–1006.
Damborenea, S.E., 1993. Early Jurassic South American pecti-Arkell, W.J., 1933. A monograph of British Corallian Lamelli-naceans and circum-Pacific palaeobiogeography. Palaeo-branchia. Monogr. Palaeontogr. Soc. London 5, 181–228.geogr., Palaeoclimatol., Palaeoecol. 100, 109–123.Baker, P.G., Mancenido, M.O., 1997. The morphology and
Damborenea, S.E., 1996. Palaeobiogeography of Early Jurassicshell microstructure of the thecideidine brachiopod Ancorel-bivalves along the southeastern Pacific margin. In: XIIIlina ageri from the Lower Jurassic of Argentina. Palaeon-Congreso Geologico Argentino and III Congreso de Explor-tology 40, 191–200.acion de Hidrocarburos Actas 5, 151–167.Barron, E.J., Harrison, C.G.A., Sloan II, J.L., Hay, W.W.,
Damborenea, S.E., 2000. Hispanic Corridor: its evolution and1981. Paleogeography, 180 million years ago to present.the biogeography of bivalve molluscs. In: Hall, R.L., Smith,Eclog. Geol. Helv. 74, 443–470.P.L. (Eds.), Advances in Jurassic Research 2000, GeoRes.Behmel, H., Geyer, O.F., 1966. Beitrage zur Stratigraphie undForum 6, 369–380.Palaontologie des Juras von Ostspanien. III. Stratigraphie
Damborenea, S.E., Gonzalaz-Leon, C.M., 1998. Late Triassicund Fossilfuhrung im Unterjura von Albarracin (Provinzand Early Jurassic bivalves from Sonora, Mexico. Rev. Mex.Teruel ). N. Jb. Geol. Palaont. Abh. 124, 1–52.Cienc. Geol. 14, 178–201.Benecke, E.W., 1905. Die Versteinerungen der Eisenerzforma-
Damborenea, S.E., Mancenido, M.O., 1979. On the palaeogeo-tion von Deutsch-Lothringen und Luxemburg. Abh. Geol.graphical distribution of the pectinid genus Weyla (Bivalvia,Spez.-Karte von Elsaß-Lothringen, N.F. 6, 1–598.Lower Jurassic). Palaeogeogr., Palaeoclimatol., Palaeoecol.Berini, L., 1957. Studi paleontologici sul Lias del Monte27, 85–102.Albenza (Bergamo). Lamellibranchi e gastropodi del Lias
Damborenea, S.E., Mancenido, M.O., 1988. Weyla: Semblanzainferiore. Riv. Ital. Paleont. Stratigr. 63, 31–64.de un bivalvo Jurasico Andino. In: V Congreso GeologicoBertuletti, C., 1962. Studi paleontologici sul Lias del MonteChileno Tomo 2., C13–C15.Albenza (Bergamo). I lamellibranchi dell’ Hettangiano. Riv.
Damborenea, S.E., Mancenido, M.O., 1992. A comparison ofItal. Paleont. Stratigr. 68, 169–192.Jurassic marine benthonic faunas from South America andBoomer, I., Ballent, S., 1996. Early–Middle Jurassic ostracodNew Zealand. J. R. Soc. N.Z. 22, 131–152.migration between the northern and southern hemispheres:
Damborenea, S.E., Polubotko, I.V., Sey, I.I., Paraketsov, K.V.,further evidence for a proto Atlantic–Central America con-1992. Bivalve zones and assemblages of the circum-Pacificnection. Palaeogeogr., Palaeoclimatol., Palaeoecol. 121,
53–64. region. In: Westermann, G.E.G. (Ed.), The Jurassic of the

392 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Circum-Pacific. Cambridge University Press, Cambridge, Hallam, A., 1994. An outline of phanerozoic biogeography.Oxford Biogeogr. Ser. 10, 1–246.pp. 300–307.
Dareste de la Chavanne, J., 1920. Fossiles liasiques de la region Hallam, A., 1998. The determination of Jurassic environmentsusing palaeoecological methods. Bull. Soc. Geol. Fr. 169,de Guelma. Mater. Carte Geol. Alger., Ser. 1 (Paleont.)
5 72 pp. 681–687.Hay, W.W., Behensky, J.F., Barron, E.J., Sloan II, J.L., 1982.Dechaseaux, C., 1936. Pectinides jurassiques de l’est du Bassin
de Paris. Revision et biogeographie. Ann. Paleont. 25, Late Triassic–Liassic paleoclimatology of the proto-centralNorth Atlantic rift system. Palaeogeogr., Palaeoclimatol.,1–148.
de Lamarck, J.B, 1819. Histoire naturelle des animaux sans Palaeoecol. 40, 13–30.Hayami, I., 1975. A systematic survey of the Mesozoic Bivalviavertebres Tome 6. Verdiere. 343 pp.
de Verneuil, M., Collomb, E., 1853. Coup d’oeil sur la constitu- from Japan. Univ. Mus., Univ. Tokyo, Bull. 10, 1–249.Hillebrandt, A.v., 1973. Die Ammonitengattungen Bouleicerastion geologique de quelques provinces de l’Espagne. Bull.
Soc. Geol. Fr. Paris, Ser. 2 10, 61–147, 162–166. und Frechiella im Jura von Chile und Argentinien. Eclog.Geol. Helv. 66, 351–363.Dubar, G., 1948. Etudes paleontologiques sur le Lias du Maroc.
La faune Domerienne du Djebel Bou-Dahar. Notes Mem. Hillebrandt, A.v., 1981. Kontinentalverschiebung und diepalaozoogeographischen Beziehungen des sudamerikan-Serv. Geol. Maroc 68, 1–250.
Efimova, A.F., Kinasov, V.P., Paraketsov, K.V., Polubotko, ischen Lias. Geol. Rdsch. 70, 570–582.Hillebrandt, A.v., Schmidt-Effing, R., 1981. Ammoniten ausI.V., Repin, Yu.S., Dagis, A.S., 1968. Field atlas of the Jur-
assic fauna and flora of the northeastern part of the USSR. dem Toarcium (Jura) von Chile (Sudamerika). Zitteliana6, 1–74.Ministry of Geology RSFSR, Severo-vostochnoye ordena
trydovogo krasnogo Znameni geologicheskoe upravlenie, Holder, H., 1978. Uber die Pectiniden-Gattung Parvamussiumim Jura. Stutt. Beitr. Naturk., Ser. B 38 37 pp.Magadanskoe Knizhnoe-Izdatel’stvo, 378 pp. (in Russian).
Fucini, A., 1920. Fossili domeriani dei dintorni di Taormina. Holder, H., 1990. Uber die Muschelgattung Placunopsis (Pecti-nacea, Placunopsidae) in Trias und Jura. Stutt. Beitr.Parte I. Palaeontogr. Ital. 26, 75–116.
Fursich, F.T., 1993. Palaeoecology and evolution of Mesozoic Naturk., Ser. B 165 63 pp.Jakobs, G.K., 1995. New occurrences of Leukadiella and Paro-salinity-controlled benthic macroinvertebrate associations.
Lethaia 26, 327–346. niceras (Ammonoidea) from the Toarcian (Lower Jurassic)of the Canadian Cordillera. J. Paleont. 69, 89–98.Gardet, G., Gerard, C., 1946. Contribution a l’etude paleon-
tologique du Moyen-Atlas septentrional. Notes Mem. Serv. Jeletzky, J.A., 1980. Dicoelitid belemnites from the Toarcian–Middle Bajocian of western and Arctic Canada. Geol. Surv.Geol. Maroc 64, 1–88.
Gemmellaro, G.G., 1872–1882. Sopra alcune faune giuresi e Can. Bull. 338, 1–71.Johnson, A.L.A., 1984. The palaeobiology of the bivalve fami-liasiche della Sicilia. Studi paleontologici. Lao. Palermo,
434 pp. lies Pectinidae and Propeamussiidae in the Jurassic ofEurope. Zitteliana 11, 1–235.Geyer, O.F., 1973. Das prakretazische Mesozoikum von
Kolumbien. Geol. Jb. B 5 155 pp. Joly, H., 1936. Les fossiles du Jurassique de la Belgique avecdescription stratigraphique de chaque etage. DeuxiemeGoldfuss, G.A., 1833. In: Petrefacta Germaniae. II (1). Arnz,
Dusseldorf, pp. 1–68. partie. Lias inferieur. Mem. Mus. R. Hist. Nat. Belg. 79,1–244.Goldfuss, G.A., 1835. In: Petrefacta Germaniae. II (2). Arnz,
Dusseldorf, pp. 69–140. Kristan-Tollmann, E., Tollmann, A., 1982. Die Entwicklungder Tethystrias und Herkunft ihrer Fauna. Geol. Rdsch.Gradstein, F.M., Agterberg, F.P., Ogg, J.G., Hardenbol, J., van
Veen, P., Thierry, J., Huang, Z., 1994. A Mesozoic time 71, 987–1019.Kuhn, O., 1935. Weitere Beitrage zur Fauna des untersten Liasscale. J. Geophys. Res. 99, 24,051–24,074.
Hallam, A., 1977. Jurassic bivalve biogeography. Paleobiology in Schwaben und Franken. Jb. Ver. Vaterland. Naturk.Wurtt. 91, 2–18.3, 58–73.
Hallam, A., 1983. Early and mid-Jurassic molluscan biogeogra- Kuhn, O., 1936. Die Fauna des Amaltheentons (Lias ∂) in Fran-ken. N. Jb. Min. Geol. Palaont. B 75, 231–311.phy and the establishment of the central Atlantic seaway.
Palaeogeogr., Palaeoclimatol., Palaeoecol. 43, 181–193. Kuhn, O., 1938. Die Fauna des Dogger ∂ der Frankenalb (MitNachtragen zum ubrigen Jura). Nova Acta Leopold. N.F.Hallam, A., 1987. Radiations and extinctions in relation to envi-
ronmental change in the marine Lower Jurassic of northwest 6, 125–170.Kutzbach, J.E., Gallimore, R.G., 1989. Pangaean climates:Europe. Paleobiology 13, 152–168.
Hallam, A., 1988. A reevaluation of Jurassic eustasy in the light megamonsoons of the megacontinent. J. Geophys. Res. 94,3341–3357.of new data and the revised Exxon curve. In: Wilgus, C.K.,
Hastings, B.S.Posamentier, H., Wagoner, J.V., Ross, C.A., Lanquine, A., 1929. Le Lias et le Jurassique des chainesprovencales. I. Le Lias et le Jurassique Inferieur. Bull. Serv.Kendall, C.G.S.C. (Eds.), Sea-level Changes — An Integ-
rated Approach, SEPM Spec. Publ. 42, 261–273. Carte Geol. Fr. 32, 41–425.Lentini, F., 1973. I molluschi del Lias inferiore di Lorigi (SiciliaHallam, A., 1992. Phanerozoic Sea-Level Changes. Columbia
University Press, Columbia. nordorientale). Boll. Soc. Paleont. Ital. 12, 23–75.

393M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Leymerie, A., 1881. Description geologique et paleontologique of the Circum-Pacific. Cambridge University Press, Cam-bridge, pp. 365–379.des Pyrenees de la Haute-Garonne. Edouard Privat.
Toulouse 1010 pp. Parrish, J.T., 1993. Climate of the supercontinent Pangea.J. Geol. 101, 215–233.Lissajous, M., 1907–1912. Jurassique Maconnais. Description
des fossiles caracteristiques et des especes les plus commu- Parrish, J.T., Ziegler, A.M., Scotese, C.R., 1982. Rainfall pat-terns and the distribution of coals and evaporites in thenes. Bull. Soc. Hist. Nat. Macon 3 496 pp.
Liu, C., Heinze, M., Fursich, F.T., 1998. Bivalve provinces Mesozoic and Cenozoic. Palaeogeogr., Palaeoclimatol.,Palaeoecol. 40, 67–101.in the Proto-Atlantic and along the southern margin of
the Tethys in the Jurassic. Palaeogeogr., Palaeoclimatol., Pedersen, G.K., 1986. Changes in the bivalve assemblage of anEarly Jurassic mudstone sequence (the Fjerritslev Forma-Palaeoecol. 137, 127–151.
Loriga, C., Neri, C., 1976. Aspetti paleobiologici e paleogeo- tion in the Gassum 1 Well, Denmark). Palaeogeogr., Palaeo-climat., Palaeoecol. 53, 139–160.grafici della facies a ‘Lithiotis’ (Giurese Inf.). Riv. Ital.
Paleont. Stratigr. 82, 651–706. Phillips, J., 1829. Illustrations of the Geology of Yorkshire.Wilson and Sons, York. 199 pp.Mancenido, M.O., Dagys, A.S., 1992. Brachiopods of the
circum-Pacific region. In: Westermann, G.E.G. (Ed.), The Phillips, J., 1871. Geology of Oxford and the Valley of theThames. Clarendon Press, Oxford. 523 pp.Jurassic of the Circum-Pacific. Cambridge University Press,
Cambridge, pp. 328–333. Polubotko, I.V., Repin, Yu.S., 1988. Lower and Middle Jurassicof the North-East. In: Westermann, G.E.G., Riccardi, A.C.Mancenido, M.O., Damborenea, S.E., 1990. Corallophilous
micromorphic brachiopods from the Lower Jurassic of west (Eds.), Jurassic Taxa Ranges and Correlation Charts forthe Circum Pacific. 1. Soviet Union, Newslett. Stratigr. 19,central Argentina. In: MacKinnon, D.I., Lee, D.E., Camp-
bell, J.D. (Eds.), Brachiopods through Time. Balkema, Rot- 1–17.Quenstedt, F.A., 1851–1852. Handbuch der Petrefaktenkunde.terdam, pp. 89–96.
McKenna, M.C., 1973. Sweepstakes, filters, corridors, Noah’s first ed., Laupp and Siebeck. Tubingen 792 pp.Quenstedt, F.A., 1856–1857. Der Jura. Tubingen Laupp.arks, and beached Viking funeral ships in palaeogeography.
In: Tarling, D.H., Runcorn, S.K. (Eds.), Implications of 842 pp.Quenstedt, F.A., 1865–1866. Handbuch der Petrefaktenkunde.Continental Drift to the Earth Sciences. Academic Press,
New York, pp. 295–308. second ed., Laupp and Siebeck. Tubingen 982 pp.Quenstedt, F.A., 1882–1885. Handbuch der Petrefaktenkunde.Milova, L.V., 1976. Stratigraphy and bivalve molluscs of the
Triassic–Jurassic deposits of northern Priokhotya. Akade- third ed., Laupp. Tubingen 1239 pp.Riccardi, A.C., 1991. Jurassic and Cretaceous marine connec-miya Nauk SSSR, Dal’nevostochnyy Nauchnyy Tsentr,
Trudy Severo-Vostochnyy Kompleksnyy Nauchno-Issledo- tions between the Southeast Pacific and Tethys. Palaeo-geogr., Palaeoclimatol., Palaeoecol. 87, 155–189.vatel’-skiy Instituta 65, pp. 3–110 (in Russian).
Milova, L.V., 1985. New Pliensbachian bivalve molluscs of Riccardi, A.C., Damborenea, S.E., Mancenido, M.O., 1990.Lower Jurassic of South America and Antarctic Peninsula.North Priokhotya. In: Bivalve and Cephalopod Mollusks
of the North-East of the USSR. Akademiya Nauk SSSR, Newslett. Stratigr. 21, 75–103.Rollier, L., 1915. Fossiles nouveaux ou peu connus des terrainspp. 42–56. in Russian.
Milova, L.V., 1988. Early Jurassic bivalve molluscs of north- secondaires (mesozoiques) du Jura et des contrees envi-ronnantes. Mem. Soc. Paleont. Suisse 41, 345–500.eastern USSR. Akademiya Nauk SSSR, Dal’nevostochnyy
Otdeleniye, Trudy Severo-Vostochnyy Kompleksnyy Sandy, M.R., Stanley Jr., G.D., 1993. Late Triassic brachiopodsfrom the Luning Formation, Nevada, and their palaeobio-Nauchno-Issledovatel’-skiy Instituta, 152 pp. (in Russian).
Nauss, A.L., Smith, P.L., 1988. Lithiotis (Bivalvia) bioherms in geographical significance. Palaeontology 36, 439–480.Schafle, L., 1929. Uber Lias- und Doggeraustern. Geol. Palaont.the Lower Jurassic of east-central Oregon, USA. Palaeo-
geogr., Palaeoclimatol., Palaeoecol. 65, 253–268. Abh. N.F. 17, 63–150.Schlonbach, U., 1863. Ueber den Eisenstein des mittleren LiasNewton, C.R., 1988. Significance of ‘Tethyan’ fossils in the
American Cordillera. Science 242, 385–391. im nord-westlichen Deutschland, mit Berucksichtigung deralteren und jungeren Lias-Schichten. Zeitschr. dt. Geol. Ges.Olsen, P.E., 1997. Stratigraphic record of the Early Mesozoic
breakup of Pangea in the Laurasia–Gondwana rift system. 15, 465–566.Schmidt-Effing, R., 1980. The Huayacocotla aulacogen inAnnu. Rev. Earth Planet. Sci. 25, 337–401.
Oppel, A., 1853. Der mittlere Lias Schwabens. Jh. Ver. Vater- Mexico (Lower Jurassic) and the origin of the Gulfof Mexico. In: Pilger, R.H. (Ed.), The Origin of the Gulf ofland. Naturk. Wurtt. 10, 39–136.
Oppel, A., 1856. Die Juraformation Englands, Frankreichs und Mexico and the Early Opening of the Central North AtlanticOcean. Louisiana State University, Baton Rouge, pp. 79–86.des sudwestlichen Deutschlands. Wurtt. Naturwiss. Jh. 12,
1–438. Sey, I.I., 1984. The Late Pliensbachian bivalves of BureyaTrough (Far East of Russia). In: Poyarkova, Z.N., Zonova-Palmer, C.P., 1989. Larval shells of four Jurassic bivalve mol-
luscs. Bull. Br. Mus. Nat. Hist. (Geol.) 45, 57–69. lov, V.P. (Eds.), New Data on the Detailed Biostratigraphyof the Phanerozoic of the Far East. Akademiya Nauk SSSR,Parrish, J.T., 1992. Jurassic climate and oceanography of the
Pacific region. In: Westermann, G.E.G. (Ed.), The Jurassic pp. 86–97. in Russian.

394 M. Aberhan / Palaeogeography, Palaeoclimatology, Palaeoecology 165 (2001) 375–394
Simpson, G.G., 1940. Mammals and land bridges. J. Wash- (Rhaeto-Lias). Lunds Univ. Arsskrift, N.F. Avd. 2 (47),1–268.ington Acad. Sci. 30, 137–163.
Smith, P.L., 1983. The Pliensbachian ammonite Dayiceras day- Vacek, M., 1886. Uber die Fauna der Oolithe von Cap. S. Vigi-lio verbunden mit einer Studie uber die obere Liasgrenze.iceroides and Early Jurassic paleogeography. Can. J. Earth
Sci. 20, 86–91. Abh. k. k. Geol. Reichsanst. 12, 1–212.van Andel, T.H., 1994. New Views on an Old Planet. Cam-Smith, P.L., 1988. Paleobiogeography and plate tectonics.
Geosci. Can. 15, 261–279. bridge Univerity Press, Cambridge. 439 pp.Waller, T.R., 1991. Evolutionary relationships among commer-Smith, P.L., Tipper, H.W., 1986. Plate tectonics and paleobioge-
ography: Early Jurassic (Pliensbachian) endemism and cial scallops (Mollusca: Bivalvia: Pectinidae). In: Shumway,S.E. (Ed.), Scallops: Biology, Ecology and Aquaculture.diversity. Palaios 1, 399–412.
Smith, P.L., Westermann, G.E.G., Stanley, G.D., Yancey, T.E., Elsevier, Amsterdam, pp. 1–73.Waller, T.R., 1993. The evolution of ‘Chlamys’ (Mollusca:1990. Paleobiogeography of the ancient Pacific. Science
249, 680–681. Bivalvia: Pectinidae) in the tropical western Atlantic andeastern Pacific. Am. Malacol. Bull. 10, 195–249.Staesche, K.v., 1926. Die Pectiniden des Schwabischen Jura.
Geol. Palaont. Abh. N.S. 15 136 pp. Westermann, G.E.G., 1977. Comments to Hallam’s conclusionregarding the first marine connection between the easternStanley Jr., G.D., 1994. Late Paleozoic and early Mesozoic reef-
building organisms and paleogeography: the Tethyan– Pacific and western Tethys. In: West, R.M. (Ed.), Paleontol-ogy and Plate Tectonics, Milwaukee Publ. Mus. Spec. Publ.North American connection. Cour. Forsch.-Inst. Senckenb-
erg 172, 69–75. Biol. Geol. 2, 35–38.Westermann, G.E.G., 1993. Global bio-events in mid-JurassicStanley Jr., G.D., Beauvais, L., 1994. Corals from an Early
Jurassic coral reef in British Columbia refuge on an oceanic ammonites controlled by seaways. In: House, M.R. (Ed.),The Ammonoidea: Environment, Ecology and Evolutionaryisland reef. Lethaia 27, 35–47.
Terquem, O., 1855. Paleontologie de l’etage inferieur de la for- Change, System. Assoc. Spec. Vol. 47, 187–226.Westermann, G.E.G., Riccardi, A., 1985. Middle Jurassicmation liasique de la province de Luxembourg, Grand-
Duche (Hollande) et de Hettange, du Departement de la ammonite evolution in the Andean Province and emigrationto Tethys. In: Bayer, U., Seilacher, A. (Eds.), SedimentaryMoselle. Mem. Soc. Geol. Fr., Ser. 2 5, 219–343.
Terquem, O., Piette, E., 1865. Le Lias inferieur de l’est de la and Evolutionary Cycles. Springer, Berlin, pp. 4–34.Ziegler, A.M., Parrish, J.M., Yao, J., Gyllenhaal, E.D., Rowley,France comprenant la Meurthe, la Moselle, le Grand-Duche
de Luxembourg, la Belgique et la Meuse. Mem. Soc. Geol. D.B., Parrish, J.T., Nie, S., Bekker, A., Hulver, M.L., 1993.Early Mesozoic phytogeography and climate. Philos. Trans.Fr., Ser. 2 8, 1–175.
Trechmann, C.T., 1923. The Jurassic rocks of New Zealand. Q. R. Soc. London, Ser. B: 341, 297–305.Zieten, C.H.v., 1833. In: Die Versteinerungen Wurttembergs.J. Geol. Soc. London 79, 246–312.
Troedsson, G., 1951. On the Hoganas Series of Sweden Schweizerbart Stuttgant, pp. 65–102.





























