Embed Size (px)
Citation preview

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
1
Diversity and Abundance of Seaweed at Satang Besar Island, Sarawak.
Farah Adibah Esa
*, Mohd. Nasarudin Harith & Ruhana Hassan
Faculty of Resource Science and Technology, Universiti Malaysia Sarawak, 94300, Kota Samarahan,
Sarawak, Malaysia.
*E-mail: [email protected]
Abstract
Satang Besar Island (N 01°46.820’ E 110°09.908’) is one of the islands in Talang-
Satang National Park, Sarawak. Diversity and abundance of seaweed on the intertidal
area of Satang Besar Island were studied with aim to provide baseline data for seaweed
in Sarawak. The study was carried out in 10 stations using transect line method and
quadrate sampling techniques. Results revealed that there were 17 species of seaweeds
(9 families, 11 genera) at Satang Besar Island. Chlorophyta in Satang Besar Island was
represented by Caulerpa serrulata, Halimeda tuna, H. discoidea, H. opuntia, H.
macroloba, Avrainvillea erecta and Cladophoropsis membranaceae. Phaeophyta was
represented by Dictyota dichotoma, D. bartayresii, Padina boryana, P.tetrasomatica,
Sargassum polycystum and Colpomenia sinuosa. Rhodophyta was presented by
Acanthopora spicifera, A. muscoides, Laurencia majuscula and Amphiroa fragillisima.
Colpomenia sinuosa was the most abundance and dominant species at Satang Besar
Island followed by Padina tetrasomatica, Dictyota bartayresii and P.boryana, whereas
the least abundance was Avrainvillea erecta. Bray-Curtis similarity analysis showed that
species abundance clustered according to habitat preferences.
Keywords: Satang Besar Island, diversity, abundance, seaweed, Sarawak.
Introduction
Satang Besar Island (SBI) (N 01°46.820’ E 110°09.908’), is one of the islands
in Talang-Satang National Park located at the western part of Sarawak (Figure 1). This
island supports a diverse flora and fauna including seaweed (Fisheries Research
Institute, 1998). In addition itpossesses a small coral reef area and also known to be one
of the main nesting and grazing grounds for green turtle, Chelonia mydas and hawksbill
turtle, Eretmochelys imbricata.
Seaweeds are known as marine macroalgae (Thomas, 2002; Phang et al., 2008)
and are categorized into three groups based on pigmentation namely Chlorophyta,

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
2
Rhodophyta and Phaeophyta. They play an important roles in the ecology of coral reefs
which include contributions in construction and cementation of reef framework
(Thomas, 2002), besides providing food sources and nursery grounds for marine
organism (Prathep et al., 2011). In addition, seaweeds assemblages are important for
small fishes which provide refuge from predators (Ranjitham et al., 2008). Other than
that, seaweeds contribute to the primary productions in the oceans, as well as carbon
sequestration, thus aid in the reduction of global warming (Phang et al., 2008).
The livelihood of seaweed communities in the tropical areas received less
attention compared to those in the temperate countries thus the information concerning
the diversity of these communities is still limited (Wong et al., 2012), and especially in
Sarawak (Nurridan, 2007). Recently, there is an increase interest in seaweed research as
several studies had been conducted to assess the diversity and population of seaweed in
Malaysia, for example in Similajau National Park (Zakaria et al, 2006), Port Dickson
(Uddin et al., 2006), Teluk Kemang and Teluk Pelanduk in Port Dickson (Norashikin et
al., 2012), Kuala Similajau (Wong et al., 2010) and Tanjung Batu and Kampung Kuala
Nyalau at Bintulu (Wong et al., 2012). Seaweed assemblages in SBI are least known in
terms of species diversity and abundance. Therefore, this study was designed to identify
and quantify seaweed in SBI. The findings in this study will serve as additional
information on the current status of seaweed in Sarawak coastal waters.
Materials and Methods
During low tide period, seaweeds were collected in ten selected stations within
the intertidal area of SBI (Figure 1). The locations of the sites were recorded using
Global Positioning System (GPS) GARMIN 76csx and the brief information of each
location as in Table 1. Three 100 m line transects were laid perpendicular to the shore at
every station. For every 5 m of the transect line, four 50 cm×50 cm quadrat was placed
randomly following method by Dhargalkar and Kavlekar (2004), Uddin et al. (2007)
and Wong et al. (2012).

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
3
Figure 1: Map of Sarawak showing SBI and the ten selected stations. (Picture obtained from
Google Earth).
Table 1: Brief description of the locations and GPS coordinates
Station GPS Coordinate Location Description
1 N 01°46.820’;
E 110°09.908’
Shallow beaches with rocks and sandy
substrate. Sandy beach with intertidal zone is
about 70 m. Subtidal area is approximately 115
m during low tide.
2 N 01°46.772’;
E 110°09.895’
Rocky and sandy substrate for intertidal and
subtidal. Intertidal area is about 100 m and
subtidal area is approximately 150 m during
low tide. Patches of sea urchin were observed.

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
4
Table 1 continued.
3 N 01°46.740’;
E 110°09.822’
Sandy beaches with shallow coral reefs.
Intertidal zone extending up to 100 m and
subtidal area is approximately 155 m during
low tide. Patches of sea urchin were observed.
4 N 01°46.796’;
E 110°09.667’
Rocky shore with intertidal zone of 200 m and
subtidal area is approximately 135 m during
low tide.
5 N 01°46.796’;
E 110°09.501’
Rocky area with intertidal zone of 200 m.
Subtidal area is approximately 135 m during
low tide.
6 N 01°47.060’
E 110°09.507’
Rocky shore (very sharp and steep rocks), very
strong wave action, intertidal zone less than
1m.
7 N 01 ° 47.242’
E 110°09.562’
Rocky shore with intertidal zone of 50 m, very
strong wave action.
8 N 01°47.372’
E 110°10.150’
Rocky shore with intertidal zone of 80 m.
9 N 01°47.259’
E 110° 10.185’
Sandy beach and rocky shore with intertidal
zone of 100 m.
10 N 01°47.085’
E 110° 10.134’
Sandy beach and rocky shore with intertidal
zone of 100 m.
Counting of seaweed was conducted to compile data on the abundance by
calculating the total number of individuals of each species in all quadrates, divided by
the total number of quadrates in which the species occurred (Curtis & McIntosh, 1950).
The seaweed specimens were washed and cleaned to remove any foreign material such
as epiphytes, sand and soil, kept cool and transported back to laboratory. Photograph of
each specimen was captured and kept as record. Specimens were identified up to species
level following standard species identification keys of Abbot and Dawson (1956),
Menez and Calumpong (1982), Dawes (1981), Bandeira-Pedrosa et al. (2004) Nurridan
(2007), Aisha and Shameel (2010).
The cluster analysis was performed using the Bray-Curtis similarities measure
(Bray-Curtis, 1957). The relationship was based on the comparison of similarity

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
5
matrices, and was displayed using hierarchical agglomerative clustering (group average)
and multi-dimensional scaling (MDS) Bray-Curtis similarity analyses were done on
seaweed assemblages to reveal the similarities between the stations for the spatial
differences, using Plymouth Routines in Multivariate Ecological Research (PRIMER
v6.0) (Clarke & Green, 2006).
Results and Discussions
A total of 17 species of seaweed (9 families, 11 genera) was present in SBI.
Based on Figure 2, Chlorophyta has the highest percentage of seaweed found in SBI
(41.2%). This may be due to high number of species from Chlorophyta are mainly
found in shallow tropical waters (Phang 2006). Chlorophyta in SBI was represented by
Caulerpa serrulata, Halimeda tuna, H. discoidea, H. opuntia, H. macroloba,
Avrainvillea erecta and Cladophoropsis membranaceae.
Besides Chlorophyta, Division Phaeophyta was represented by Dictyota
dichotoma, D. bartayresii, Padina boryana, P.tetrasomatica, Sargassum polycystum
and Colpomenia sinuosa. Furthermore, Rhodophyta was presented by Acanthopora
spicifera, A. muscoides, Laurencia majuscula and Amphiroa fragillisima. Details of
species found according to stations are in Table 2.
Figure 2: Percentage of seaweed found in all stations, based on Divisions.
Rhodophyta has the least number of species in SBI. In contrast, Phang (1995)
reported that Cape Rachado supported many species of Rhodophyta compared to other
Chlorophyta 41.2%
Phaeophyta 35.3%
Rhodophyta 23.5%

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
6
seaweed divisions. This could be due to the sampling that was done in deeper water area
of Cape Rachado where Rhodophyta are most likely to occur. Rhodophyta species are
able to withstand limited sunlight because they could absorb light at low intensity
compared to the species from the division Chlorophyta that need to absorb light at the
highest intensity due to Chlorophyta ability to photosynthesize in the same way as
higher plant (Diez et al., 2003). Since the present study mainly focuses on the intertidal
area, less of Rhodophyta was recorded.
Table 2: Seaweed distribution according to species and stations in SBI.
Division Family/Species Station
1 2 3 4 5 6 7 8 9 10
Chlorophyta Family Caulerpaceae
Caulerpa serrulata
Family Halimedaceae
Halimeda tuna
H.discoidea
H. macroloba
H.opuntia
Family Udoteaceae
Avrainvillea erecta
Family Siphonocladaceae
Cladophoropsis
membranaceae
+
+
+
+
+
+
-
+
+
+
+
+
+
-
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
-
-
-
-
-
+
-
-
+
+
+
+
+
-
-
+
+
+
+
+
-
Phaeophyta Family Dictyoceae
Dictyota dichotoma
D.bartayresii
Padina boryana
P.tetrasomatica
Family Sargassaceae
Sargassum polycystum
Family Scytosiphonaceae
Colpomenia sinuosa
-
+
+
+
-
+
+
-
+
+
-
+
+
-
+
+
-
-
-
-
-
+
-
-
-
-
-
+
-
+
-
-
-
-
-
-
+
+
+
-
+
+
+
+
+
+
+
+
-
-
+
+
-
+
-
-
+
+
-
+
Rhodophyta Family Rhodomelaceae
Acanthopora spicifera
A.muscoides
Laurencia majuscula
Family Corallinales
Amphiroa fragillisima
+
+
-
+
+
+
-
+
+
+
-
+
-
-
-
-
-
-
-
-
-
-
+
-
+
+
+
-
+
-
-
+
+
+
-
+
Total number
of species
13 13 12 1 2 0 7 10 10 11

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
7
+ = presence of seaweed, - = absence of seaweed
Most of the Chlorophyta species namely C. serrulata, H. tuna, H. discoidea, H.
macroloba, H. opuntia and Avrainvillea erecta that were found in SBI were siphonous
seaweeds that produce penetrating, rootlike holdfast in sandy substratum for attachment
(Lobban & Harrison, 1994). In addition this character also involve in nutrient uptake
(Lobban & Harrison, 1994). Genus Halimeda was mainly found in ST1, 2, 3, 9 and 10
(Table 2) because those stationscomprised sandy, rocky and coral reef/rubble substrates
(Table 1). In addition, the abundance of Halimeda in these stations was coherent with
the characteristics of the stations as Halimeda also contributes in building up the coral
rocks (Sundararajan et al., 1999).
Phaeophyta was found inhabiting the rocky and sandy substrates in SBI. The
composition of seaweeds at rocky shore in these islands was dominated by family
Dictyotaceae. Colpomenia sinuosa was the most abundant and dominant species at SBI
with values of 27.55 followed by Padina tetrasomatica (19.11), Dictyota bartayresii
(12.33) and P. boryana (11.32) as shown in Table 3. The lowest abundance value was
recorded by Avrainvillea erecta of 2.80 (Table 3) Another factor which contributes to
low number of seaweed species may be due that this island is vulnerable to grazing
especially by sea urchins, for example in Station 2 and 3. Combined feeding activities
by aggregations of sea urchins can remove all large plant material which can impact the
seaweed abundance.
Table 3: Species abundance of seaweed in Satang Besar Island.
Name of species Abundance
CHLOROPHYTA
Caulerpa serrulata 4.42
Halimeda tuna 6.32
Halimeda discoidea 5.50
Halimeda opuntia 4.26
Halimeda macroloba 3.87
Avrainvillea erecta 2.80
Cladophoropsis membranaceae 4.00
PHAEOPHYTA
Padina tetrastomatica 19.11
Padina boryana 11.32
Dictyota dichotoma 5.64

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
8
Dictyota bartayresii 12.33
Sargassum polycystum 3.73
Colpomenia sinuosa 27.55
Table 3 continued.
RHODOPHYTA
Acanthophora spicifera 5.68
Acanthopora muscoides 8.20
Laurencia majuscula 4.22
Amphiroa fragilissima 5.00
In the intertidal habitats, changing tides had made this area susceptible to stress
such as desiccation. The stress could also be due to overexposed to rainfall or direct
sunlight. A small specific surface is the best way to reduce desiccation such as in the
voluminous thallus of Codium, as it can retain more water and have a smaller relative
circumference resulting in reduced water loss (Churchill, 2009). During low water level,
most of the seaweed has little chance to survive unless they have special adaptations to
cope with harsh environment. In this case, C.sinuosa has spherical to lobed shaped and
its surface was approximately 2.5 cm across.This characters could be regarded as
adaptation to stress thus may explain the abundance of this species in this SBI. Padina
inhabits a variety of substratum namely sandy areas, coral reefs and rocky shores. In
this study, Padina tetrastomatica was well represented in all stations and formed their
own dense patches. This is due to the ability of spore dispersal of genus Padina that are
limited only nearby to the parent cells. It is also possible due to the lower water
velocity around stations, thus thisgenus is being recruited and grew mainly next to the
parent’s plant forming patches (Mayakun & Prathep, 2005).
There is no seaweed found in ST6 in Satang Besar Island (Table 2) probably
due to the characteristics of the location which was rocky shore with many big rocks,
strong wave and exposed to sunlight. As the island experienced semi diurnal tides, low
tides may occur in the middle of the day in which heating and desiccation can be
extreme (Lobban & Harrison, 1994) thus, affect the ability of seaweed to colonize this
area. According to Lobban and Harrison (1994), seaweeds that are out of the water more
than half an hour are more exposed to the atmosphere, which could lead to stress.
Similarly, Japar Sidik et al. (2012) stated that areas of low tides create a harsh
environment for seaweed due to dessication and strong solar radiation.
The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
9
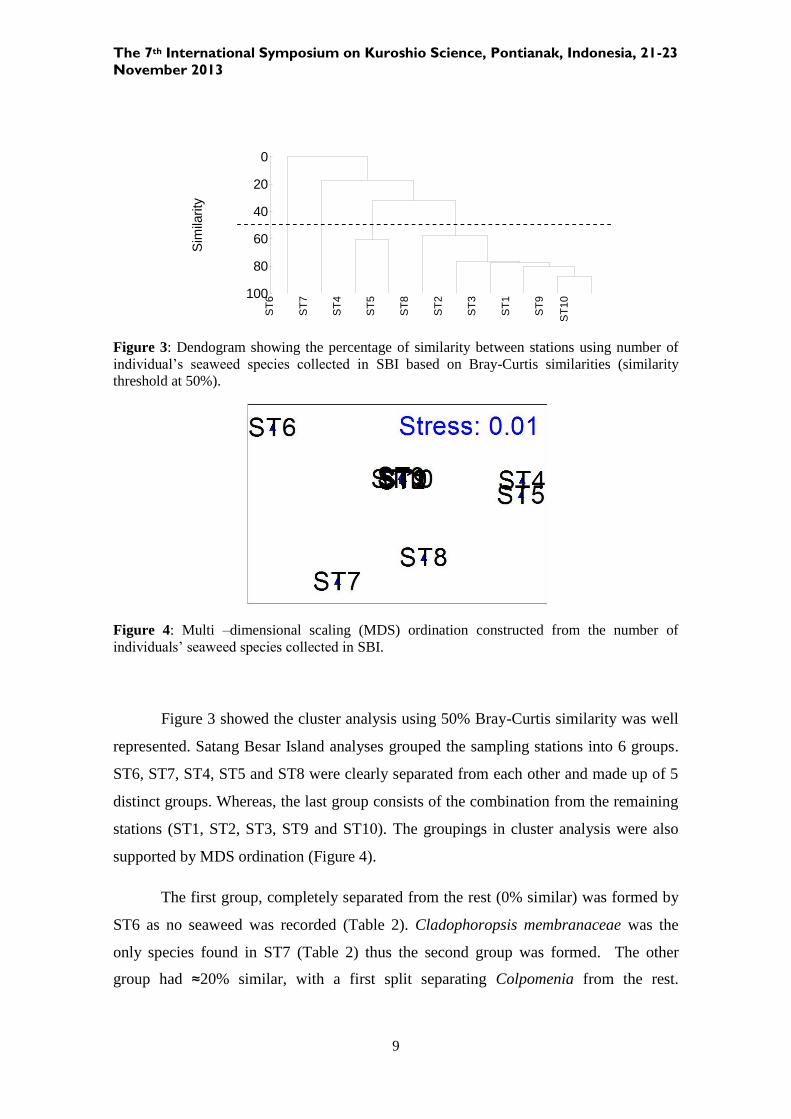
Figure 3: Dendogram showing the percentage of similarity between stations using number of
individual’s seaweed species collected in SBI based on Bray-Curtis similarities (similarity
threshold at 50%).
Figure 4: Multi –dimensional scaling (MDS) ordination constructed from the number of
individuals’ seaweed species collected in SBI.
Figure 3 showed the cluster analysis using 50% Bray-Curtis similarity was well
represented. Satang Besar Island analyses grouped the sampling stations into 6 groups.
ST6, ST7, ST4, ST5 and ST8 were clearly separated from each other and made up of 5
distinct groups. Whereas, the last group consists of the combination from the remaining
stations (ST1, ST2, ST3, ST9 and ST10). The groupings in cluster analysis were also
supported by MDS ordination (Figure 4).
The first group, completely separated from the rest (0% similar) was formed by
ST6 as no seaweed was recorded (Table 2). Cladophoropsis membranaceae was the
only species found in ST7 (Table 2) thus the second group was formed. The other
group had ≈20% similar, with a first split separating Colpomenia from the rest.
ST
6
ST
7
ST
4
ST
5
ST
8
ST
2
ST
3
ST
1
ST
9
ST
10100
80
60
40
20
0
Sim
ilarity

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
10
Colpomenia is a genus that has an almost consistent distribution in the sampling
stations. The next group to fall out at >20% similarity comprised of Padina
tetrasomatica, which was frequently found in harsh environment since they could
tolerate to it. In addition, also noticeable was the clustering of ST4 and ST5 in species
generates patches distribution of seaweed in the area (rocky areas). These results reflect
that the substrate property is possibly a factor in determining the species for recruitment
and growth, and consequently, the pattern of algal species and abundance.
The final group with 50% similarity comprised of Caulerpa, Halimeda and
Avrainvillea. The pattern of habitat preferences can be observed in this study as the last
remaining groups (ST1, ST2, ST3, ST9 and ST10) shared similar topography and
seaweed species. MDS analysis confirms the seaweed communities in these stations
were closer to each other. Most of the siphonous green algae (Bryopsidales), which
have extensive systems of rhizoids and colonized the sandy substrates, were mainly
found in these stations.
Conclusion
Overall, a total of 17 species of seaweed (9 families, 11 genera) was present in
Satang Besar Island . Phaeophyta dominates the intertidal areas with Padina as the most
abundant. Heterogeneity of habitats had promoted suitable niches for seaweed to
colonize, leading to high seaweed diversity in the island.
References
Abbot, I. A. & Dawson, Y. E. (1956). How to know the seaweed. Dubuque, Iowa : Wm. C.
Brown Company Publishers.
Aisha, K. & Shameel, M. (2010). Occurrence of the genus Padina (Dictyophyceae, Phaeophyta)
in the coastal waters of Karachi. Pakistan Journal Botany (S.I. Ali Festschrift), 42, 319-340.
Bandeira-Pedrosa, M. E., Pereira, S. M. B. & Oliveira, E. C. (2004). Taxonomy and distribution
of the green algae genus Halimeda (Bryopsidales, Chlorophyta) in Brazil. Revista Brasil
Botany, 27, 363-377.

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
11
Buckland, S. T., Magurran, A. E., Green, R. E. and Fewster, R. M. (2005). Monitoring change
in biodiversity through composite indices. Philosophoval Transactions of the Royal Society B:
Biological, 360, 243-254.
Bray, J. R. & Curtis, J. T. (1957). An ordination of the upland forest communities of Southern
Wisconsin. Ecological Monograph, 27: 325-349.
Clarke, K. R. and Green, R. N. (2006). PRIMER v6: User Manual/ Tutorial. PRIMER-E,
Plymouth.
Churchill, H. R. (2009). The role of macroalgal morphology and community structure on the
accumulation of sediment and the subsequent effects on the dynamics of marine intertidal
communities. Master dissertations for The University of Hull.
Curtis, J. T. & McIntosh, R. P. (1950). The interrelations of certain analytic and synthetic
phytosociological characters. Ecology, 31, 438-455.
Dawes, C. J. (1974). Marine algae of the west coast of Florida. United States of America :
University of Miami Press.
Dhalgalkar, V.K. and Kavlekar, D. (2004). Seaweed - A Field Manual. National Institute of
Oceanography, Dona Paula, Goa.
Diez, I., Santolaria, A. & Gorostiaga, J. (2003). The relationship of environmental factors to the
structure and distribution of subtidal seaweed vegetation of the western Basque coast.Estuarine,
Coastal and Shelf Science, 56, 1041-1054.
Japar Sidik, B., Muta Harah, Z. & Kawaguchi, S. (2012). Historical review of seaweed research
in Malaysia before 2001. Coastal Marine Science, 35 (1), 169-177.
Konar, B., Iken, K., Cruz-Motta, J. J., Benedetti-Cecchi, L., Knowlton, A., et al. (2010). Current
Patterns of Macroalgal Diversity and Biomass in Northern Hemisphere Rocky Shores, PLoS
ONE 5(10), 1-8.
Likens (2005). Ecology: Population Ecology. In Campbell N. A., & Reece, J. B. (Eds), Biology
(7th Edition), (pp1137-1180). Pearson-Benjamin Cummings.
Lobban, C. S. & Harrison, P.J. (1994). Seaweed ecology and physiology. United Kingdom:
Cambridge University Press.
Mayakun, J. & Prathep, A. (2005). Seasonal variation in diversity and abundance of macroalgae
at Samui Island, Surat Thani Province, Thailand. Songklanakarin Journal of Science and
Technology, 27(3), 653-663.
Menez, E. G. & Calumpong, H. P. (1982). The genus Caulerpa from Central Visayas,
Philippines. City of Washington: Smithsonian Institution Press.
Muchlisin, Z. A. & Siti Azizah, M. N. (2009). Diversity and distributions of freshwater fishes in
Aceh Water, Northern-Sumatra, Indonesia. International Journal of Zoological Research. 1-18.
Norashikin, A., Muta Harah, Z. & Japar Sidik, B. (2013). Intertidal seaweed and their multi-
forms. Journal of Fisheries and Aquatic Science, 8 (3): 452-461.

The 7th International Symposium on Kuroshio Science, Pontianak, Indonesia, 21-23
November 2013
12
Nurridan, A. H. (2007). Seaweeds of Sarawak Malaysia Borneo. Fisheries Research Institute
Kuching Sarawak, Malien Press.
Phang, S.M., Lim, P. E., Ooi, J. L. S., Yeong, H. Y., Ng, W. S. & Küpper, F. C. (2008). Marine
algae of Perak Island, Jarak Island and the Sembilan Group of Islands in the Straits of Malacca.
Malaysian Journal of Science, 27 (3), 47-60.
Prathep, A., Pongparadon, S., Darakrai, A., Wichachucherd, B & Sinutok, S. (2011). Diversity
and distribution of seaweed at Khanom-Mu Ko Thale Tai National Park, Nakhon Si Thammarat
Province, Thailand. Songklanakarin Journal of Science and Technology, 33(6), 633-640.
Ranjitham, N. S., Thirumaran, G., Anantharaman, P., Nightingale, C. D. R. & Balasubramanian,
R. (2008). Associated fauna of seaweed and seagrass in Vellar Estuary. American-Eurasian
Journal of Botany, 1(1), 9-16.
Sundararajan, M., Rajendran, M., Kaliaperumal, N. & Kalimuthu, S. (1999). Studies on species
of Halimeda from Lakshadweep. Seaweed Resource Ulitization, 21 (1&2), 161-169.
Thomas, D. (2002). Seaweeds. Washington, D. C.: Smithsonian Institution Press.
Uddin, S. M. M., Ismail, A. & Ismail, A. (2007). Species composition of seaweeds in Port
Dickson, Peninsular Malaysia. Malaysia Applied Biology, 36 (2), 69-73.
Wong, S. C., Muta Harah, Z., Japar Sidik, B. & Arshad, A. (2012b). Comparison of seaweed
communities of the two rockt shores in Sarawak, Malaysia. Coastal Marine Science, 35 (1), 78-
84.
Zakaria, M. H., Bujang, J. S., Amit, R., Awing, S. A. & Ogawa, H. (2006). Marine
macrophytes: Macroalgae species and life forms from Golden Beach Similajau National Park,
Bintulu, Sarawak, Malaysia. Coastal Marine Science, 30 (1), 243-246





























