Embed Size (px)
Citation preview

Hindawi Publishing CorporationJournal of Parasitology ResearchVolume 2013 Article ID 591520 9 pageshttpdxdoiorg1011552013591520
Research ArticleDevelopment of Eimeria nieschulzi (Coccidia Apicomplexa)Gamonts and Oocysts in Primary Fetal Rat Cells
Hong Chen Stefanie Wiedmer Sacha Hanig Rolf Entzeroth and Michael Kurth
Institute of Zoology Technische Universitat Dresden Helmholtzstraszlige 10 01062 Dresden Germany
Correspondence should be addressed to Michael Kurth kurthmichaelgooglemailcom
Received 18 April 2013 Accepted 3 June 2013
Academic Editor Renato A Mortara
Copyright copy 2013 Hong Chen et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The in vitro production of gametocytes and oocysts of the apicomplexan parasite genus Eimeria is still a challenge in coccidiosisresearch Until today an in vitro development of gametocytes or oocysts had only been shown in some Eimeria species For severalmammalian Eimeria species partial developments could be achieved in different cell types but a development up to gametocytes oroocysts is still lackingThis study compares several permanent cell lines with primary fetal cells of the black rat (Rattus norvegicus)concerning the qualitative in vitro development of the rat parasite Eimeria nieschulzi With the help of transgenic parasites thedevelopmental progress was documented The selected Eimeria nieschulzi strain constitutively expresses the yellow fluorescentprotein and a macrogamont specific upregulated red tandem dimer tomato In the majority of all investigated host cells thedevelopment stopped at the second merozoite stage In a mixed culture of cells derived from inner fetal organs the developmentof schizont generations I-IV macrogamonts and oocysts were observed in crypt-like organoid structures Microgamonts andmicrogametes could not be observed and oocysts did not sporulate under air supply By immunohistology we could confirmthat wild-type E nieschulzi stages can be found in the crypts of the small intestine The results of this study may be helpful forcharacterization of native host cells and for development of an in vitro cultivation system for Eimeria species
1 Introduction
The parasites of the genus Eimeria can be found in almost allvertebrates and some species are of high economical rele-vance as they can cause coccidiosis in livestock
Eimeria parasites are known to have a stringent stageprogression in their life cycles in the host animals Theydevelop mainly in the intestinal tract but some species arealso found in the liver (rabbit) or kidney (goose) To studyall stages of the parasites it is helpful to have the completedevelopment in vitro In primary cells only a few Eimeriaspecies are able to form gametocytes or complete their lifecycles in vitro [1] Often this is only possible if merozoites areused for infection [2ndash4]
For rodent Eimeria species which are frequently used asmodel organisms a gametocyte or oocyst development wasnot achieved thus far In previous studies with the rat parasiteEimeria nieschulzi the stage development in vitro did notexceed the secondmerozoite stage [5ndash7]Whereas up to four
schizont generations are found in the host before an in vivogamont development occurs [8 9] In a study by Tilley andUpton [10] the development of four schizont generations ofEimeria nieschulzi in a permanent cell line under reducingconditions was described Their results are discussed in thiswork
In this study the progress of the in vitro development ofEimeria nieschulzi was investigated using primary tissue cellsfrom fetal rats and compared to permanent cell lines Forbetter documentation a transgenic (yellow fluorescent pro-tein YFP expressing) Eimeria nieschulzi strain with an addi-tional gamogony specific reporter (tandem dimeric tomato(TDT)) gene expression [11] was used The TDT expres-sion served as an indicator for successful development upto the gametocyte stage and beyond Furthermore we con-firmed the localization of wild-type E nieschulzi parasites inthe crypts of the small intestine by immunohistology andcompared our in vitro data in detail with the data in theliterature
2 Journal of Parasitology Research
2 Material and Methods
21 Cell Culture Pregnant rats (Crl CD (SD)) were eutha-nized at day 16ndash18 of gestation The fetuses were extractedand decapitated prior to further usage The decapitatedfetuses werewashed in cold (4∘C)DulbeccorsquosModified EaglersquosMedium (DMEM) to remove bloodThe fetuses were openedventrally and the inner organs were extracted Liver kidneyand intestine were separately collected and one assay wasperformed with all inner organs together as fetal mix (FM)All explanted tissues were washed twice with 37∘C preheatedDMEM minced into small pieces and ground througha sieve (mesh size 05mm) with a plunger The cellcellconglomerate suspension was transferred to a centrifugetube and washed once with completed DMEM (37∘C) andtransferred into completed preheated DMEM (with 10 FBS2 L-glutamine 2 penicillinstreptomycin 1 HEPESand 1 sodium pyruvate) The FM-suspension was seededin IBIDI 120583-slides with fibronectin (IBIDI) gelatin [12]Matrigel-coating (60 120583Lwell according to themanufacturerrsquosrecommendations) as well as in uncoated slides For primaryliver and kidney cells fibronectin coated IBIDI 120583-slides wereused Primary intestinal cells were seeded into IBIDI 120583-slides coated with fibronectin or gelatine as well as uncoatedslides Quality evaluation of the parasite development in vitrowas performed by comparing the following permanent celllines IEC6 (rat intestinal) VERO (green monkey kidney)ST (swine testicular) and IPEC-J2 (porcine small intestinalepithelial cell) which were routinely cultivated in completedDMEM For infection with parasites 100000ndash150000 ofthese cells were seeded in IBIDI 120583-slides per well Prior toinoculation with parasites permanent cells were incubatedabout 3-4 h and primary cell about 24 h at 37∘C and 5 CO
2
atmosphere
22 Parasites and Infection of Cells Oocysts of the passagesP2 and P3 of the previously described transgenic Eimerianieschulzi [11] were used in this study Sporozoites wereexcysted and purified as previously described [13] and resus-pended in cell culture medium (DMEM completed) Forinfection of the cells 20000ndash50000 sporozoites were incu-bated togetherwith the cells in the120583-slide at 37∘Cand 5CO
2
atmosphere Every day the cell culturemedium (DMEM)waspartially (50) removed and replaced with fresh mediumtwice
23 Immunohistology and Observation of Living IntestinalParasite Stages Rats (CD)were infected orally by gavagewith500000ndash1000000 sporulated oocysts of E nieschulzi andeuthanized at different time points after inoculation with theoocystsThe exact time points can be found in the descriptionof Figure 3 Living parasite stages were observed in smallbowel tissue smears in a 120583-slide
For histology the small intestine was cut into pieces of05 cm length and fixed in 4 paraformaldehyde1xPBS (pH74) for 24 h at 4∘C After washing in 1xPBS and dehydrationin a graded series of ethanol the tissue pieces were embeddedin paraffin Sections of 5120583mwere cutwith a rotarymicrotome(Leica RM2125RT) The sections were transferred to slides
and the paraffin was removed by Roti-Histol (Co Roth) Theantigen retrieval was performed with 01M sodium citratebuffer pH 60 for 30min at 90∘C If HRP-linked secondaryantibodies were used endogenous peroxidase was blocked byincubation with 1H
2O2in 1xPBS about 15min After block-
ing of unspecific antigenswith 20normal goat serum (NGS)and 005 Tween in 1xPBS about 1 h at room temperature thesections were incubated with hybridoma supernatant of theB1C4 clone [14] for about 2 h at room temperature (1 10dilution in 20NGS005Tween1xPBS) After the removalof the primary antibody the slides were incubated for 1 h atroom temperature with the secondary antibody diluted in20NGS005 Tween1xPBS (FITC conjugated anti-mouseIgG Co Sigma Aldrich F6257 1 100 or HRP conjugated anti-mouse IgG 1 200 Co Sigma Aldrich A9044) Fluorescenceprobed sections were washed counterstained with DAPI(1 120583gmL) about 10min and washed three times about 10minprior to mounting The sections labelled with Peroxidase(HRP) probes were equilibrated in 005M Tris buffer pH 76about 5min and further incubated in DABH
2O2staining
solution (5mg 331015840-Diaminobenzidine and 5 120583LH2O2in
10mLH2O) The staining was performed about 20 minutes
After removing all staining solutions the sections weremounted in MowiolDABCO according to the manufac-turerrsquos instructions Control experiments were performedwith infected tissue but 1xPBS instead the B1C4 antibody
24 Microscopy Images were taken using the Zeiss Axiovert100 microscope combined with the Olympus F-viewIImonochrome CCD-camera and the image processing soft-ware Cell F (Olympus) from 24 hndash270 h inoculation (pi)The fluorescence of intracellular as well as extracellular para-sitic stages was observed directly in the 120583-slides with FITCfilter set (Ex D48030) for the yellow fluorescent protein(YFP) and with a rhodamine filter set (Ex 56040) fortandem dimer tomato (TDT) For the visualization of theautofluorescence of the oocyst wall the DAPI filter set(excitation 365 nm emission long pass 420 nm)was usedThereproduction of images was performed comparable to themicroscopic view according to the filter set dependent emis-sionwavelength Capturing of the peroxidase probed sectionswas performed with the Olympus C7070 camera combinedwith the Olympus C5060-ADU ocular adapter or with themonochrome FviewII and CellF software
3 Results
In the permanent cell lines IEC6 VERO IPEC-J2 and STas well in primary liver kidney and intestinal cells thedevelopment stopped after formation of the second schizontsrespectively merozoite generation (not shown)We could notobserve any further development until 268 h pi in thesementioned cells and thereby we confirm the results ofprevious studies
Developmental stages beyond the second-generationmerozoites were observed in the fetal cell mix (FM) derivedfrom the inner organs on the coated slides but not in theuncoated control slide These results were confirmed by two
Journal of Parasitology Research 3
YFP
(a) 145 h pi
YFP
(b) 122 h pi
TL
(c) 122 h pi
YFP
(d) 216 h pi
YFP
(e) 164 h pi
YFP
(f) 180 h pi
YFP
(g) 218 h pi
TL
(h) 245 h pi
TDT
(i) 245 h pi
TL
(j) 245 h pi
TL
(k) 245 h pi
Figure 1 Development of the schizonts II and IV as well as clusters of macrogametocytes in organoid crypt-like structures in the fetal mixcell assay (a) Cluster of second-generation schizonts (YFP) which can be found early from 68 h pi and also late at 145 h pi (b) and (c)probably large schizont of the second-generation YFP (b) and transmitted light (c) (d) Probably merozoite of the second-generation (YFP)with a length of more than 20 120583m (22 120583m) Marquardt 1966 [9] and Roudabush 1937 [8] described a maximum length of 16 120583m (e) Youngfourth-generation schizonts (YFP) after few nuclear divisions (f) Clustered fourth generation schizonts (YFP) after multiple cell divisions aswells as young schizonts (g) Fourth generation merozoites (YFP) (h) Large cluster of around 100 gamonts (transmitted light micrograph)(i) Corresponding (to (h)) fluorescence micrograph of the gamonts with macrogamont specific expressed TDT signal YFP not shown (j)Transmitted light micrograph with gamonts (arrows) within cells of a crypt-like structure (k) Uninfected crypt-like structure (a) to (g) and(j) bars = 20120583m (h) (i) and (k) bars = 100 120583m
experimenters in separate experiments We have to remarkthat only in half of the performed experiments with coatedslides crypt-like organoid structures were observed Thesuccess of development of macrogamonts and oocysts wasdirectly connected with occurrence of these crypt-like struc-tures The particular experiments and the results of parasitedevelopment are listed in Table 1
In the fetal cell mix we could identify schizonts of thefirst- (not shown) second- (Figure 1(a)) and fourth-gener-ation (Figures 1(e) and 1(f)) as well as free merozoites ofthe second (Figure 1(d)) and fourth generation (Figure 1(g))
and also gametocytes and unsporulated oocysts (Figures1(h)ndash1(j) and 2)
The third-generation schizonts and merozoites are diffi-cult to distinguish from the second generation Consideringthe literature the large schizont in Figures 1(b) and 1(c) maybelong to the second or third generation Despite the size the22120583m long merozoite in Figure 1(d) would belong to thesecond generation This issue is explained in the followingdiscussion paragraph
Schizonts of the fourth generation were found clus-tered with up to 15 schizonts (picture details Figures 1(e)
4 Journal of Parasitology Research
Table 1 Number and type of experiments performed in this study
Cells Coating Cell growth Max parasitedevelopment
IPEC-J2 [2] Uncoated Monolayer Sch IIMz IIST [2] Uncoated Monolayer Sch IIMz IIVERO [gt2] Uncoated Monolayer later multilayer Sch IIMz IIIEC6 [gt2] Uncoated Monolayer Sch IIMz IIPrimintestine [3]Primintestine [1]Primintestine [1]
FibronectinGelatinUncoated
Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
Primary liver [2] Fibronectin Monolayer Sch IIMz IIPrimary kidney [2] Fibronectin Monolayer Sch IIMz II
Primary fetal mix [3] Fibronectin Monolayer and cell conglomerates in two experiments (1 2) no growingorganoids in one experiments (3) growing crypt-like organoids
(1) Sch IIMz II(2) Sch IIMz II(3) MagOoc
Primary fetal mix [2] Gelatin Monolayer and cell conglomerates in one experiment no organoids (1) inone experiment growing crypt-like organoids (2)
(1) Sch IIMz II(2) MagOoc
Primary fetal mix [1] Matrigel Monolayer and growing organoids MagOoc
Primary fetal mix (3) Uncoated Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
[number of experiments] (experiment number) Sch II schizont 2nd generation Mz II merozoite 2nd generation Mag macrogamont Ooc oocyst
and 1(f)) from 164 h pi Free merozoites IV were observed(Figure 1(g)) separately or close to the gametocytes (Figures2(a)ndash2(e) 2(j)ndash2(m)) Gametocytes show an additional redfluorescent tandem dimeric tomato signal The TDT-proteinis expressed under the microgametocyte-specific promoterof the gam56 gene found in Eimeria tenella [11 15] Thered fluorescent signal was found the earliest at 206 h piThe gametocytes often occurred in clusters with up to 100cells and grew in cells which were situated in a crypt-likeformation (Figures 1(h) 1(i) and 1(j)) However gametocyteswere not found in all crypt-like structures (Figure 1(k))
If crypt-like organoids had developed up to four clustersof macrogamonts in 25ndash50 of the 120583-slide wells could beidentified The best results were obtained in one experimentwith FM-cells on a fibronectin coated 120583-slide
Microgamonts or free microgametes were not identifiedwith certainty in the cell culture Notwithstanding unsporu-lated oocysts with autofluorescent oocyst walls could beobserved but these oocysts did not sporulate in the cell cul-ture slide or outside under air supply (Figures 2(n)ndash2(r))In summary the FM-cells provide the qualitatively potentialto develop Eimeria nieschulzi sporozoites over four asexualgenerations and the gametocyte stage to the point of oocystdevelopment in vitro The presence of macrogametocytes inorganoid crypt-like structure indicates that these cells havesimilar characteristics to the native host cells in the rat
In a selected number of performed experiments welocalized the wild type E nieschulzi in situ in the crypt of thesmall intestine by immunohistology at 72 120 and 144 h pi(Figure 3) The antibody B1C4 originally produced against Etenella sporozoites and described 1995 byGreif and Entzeroth[14] was used for immunohistology The antibody showedcross-reactivity to E nieschulzi schizonts and gamonts
At 72 h pi schizonts were detected in the middle parts ofthe crypts (Figures 3(a) and 3(b)) At higher magnificationsit was not possible to distinguish which schizont generation islabelled only the visible multiple nuclei indicated schizontsAt 117 h pi longmerozoites with a length ofmore than 25120583mand straight cell shape in the intestine (Figure 3(c)) wereidentified These merozoites belong to the third generationAt 120 h pi parasite stages were labelled in the middle andlower part of the crypts (Figure 3(d)) No parasites weredetected in the villi The schizonts shown in Figure 3(e)containing long merozoites belong to the third generationand were found in middle parts of the crypts Figure 3(f)shows schizonts of different sizes in the lower part ofthe crypts Marquardt [9] described large schizonts with amaximum of 12 merozoites as second or third generationHowever corresponding to the observed in vitro stages wewould assign them to the second-schizont-generation
At 144 h pi the parasite stageswere foundpredominantlyin the middle and upper part of the crypts as well as atthe base of the villi (Figure 3(g)) At higher magnificationsit was possible to identify mono- and multinuclear stagesThe mononuclear stages are probably young macrogamontsand the multinuclear stages fourth generation schizonts(Figure 3(h)) The control experiments without the primaryantibodies did not show any specific signal derived by thesecondary antibodies (not shown)
4 Discussion
In primary fetal cells the in vitro development of a rodentEimeria species up to the oocyst stage could be shown Asaffirmed by various cell lines in this study and in previous
Journal of Parasitology Research 5
YFP
(a) 206 h pi bar 20120583m
TDT
(b) 206 h pi bar 20120583m
YFP + TDT
(c) 206 h pi bar 20120583m
TL
(d) 206 h pi bar 20120583m
OL
(e) 206 h pi bar 20120583m
YFP
(f) 230 h pi bar 50120583m
TDT
(g) 230 h pi bar 50120583m
TL
(h) 230 h pi bar 50120583m
OL
(i) 230 h pi bar 50120583m
YFP
(j) 245 h pi bar 10120583m
TDT
(k) 245 h pi bar 10120583m
TL
(l) 245 h pi bar 10120583m
OL
(m) 245 h pi bar 10120583m
YFP
(n) 230 h pi bar 10120583m
TDT
(o) 230 h pi bar 10120583m
UV
(p) 230 h pi bar 10120583m
TL
(q) 230 h pi bar 10120583m
OL
(r) 230 h pi bar 10120583m
Figure 2 Development of macrogamonts and oocysts in primary fetal rat cells in vitro (a)ndash(e) Transgenic schizonts IV merozoites IV andmacrogamonts (a) YFP is expressed in all parasite stages (b) TDT is exclusively expressed in the macrogamonts (c) Overlay of (a) and (b)(d) Corresponding transmitted light micrograph with low contrasted parasite stages (e) Overlay of (a) (b) and (d) helps to identify the lowcontrasted gametocytes in the transmitted light micrograph and differentiating the asexual stages from the macrogamont (f)ndash(i) Cluster ofgamonts (f) YFP signal (g) TDT signal (h) Transmission light micrograph shows location of developed gamonts mainly in the periphery ofa crypt like structure An overlay of (f)ndash(h) is shown in panel (i) (j)ndash(m)Macrogamont stages adjacent to a mature schizont IV the YFP (j) isexpressed in all parasitic stages the TDT (k) in the macrogamonts The colocalization with the corresponding transmitted light micrograph(l) is shown in (m) (n)ndash(r) Formation of oocysts in vitro (n) YFP signals in unsporulated oocysts and gametocytes (o) CorrespondingTDT signals (p) Developed oocysts excited by UV light showing typical autofluorescent oocyst walls (q) Corresponding transmitted lightmicrograph (r) Overlay of (n)ndash(q)
studies the in vitro development of Eimeria nieschulzi islimited to the second-generation merozoite [5ndash7] Tilley andUpton [10] reported a development until the fourth gen-eration in vitro under reducing conditions However Eime-ria nieschulzi can have in vitro variations concerning themorphology of the schizont stages There are large schizontswith more than 20 merozoites [11] but also small schizontsless than 10 merozoites (Figure 1(a)) Thus it is difficult todistinguish the second schizont stage from the third schizontstage with certainty Because in vivo first- and second-generation schizonts can have delayed development up to96 h pi [8 9] the duration of the development cannot lead
to concrete conclusions Only the size of explicitly visiblemerozoites in or out of schizonts should be consulted fordetermination between second (MzII) and third-generationmerozoite (MzIII) but there are further challenges con-cerning the interpretation of literature data Marquardt [9]described the occurrence of MzIII beginning from 48 h piwith an average length of 208 120583m 255120583m at 72 h pi and269 120583m at 120 h pi At 120 h pi the merozoites showed ahigh motility while the previous smaller merozoites werenearly inactive The schizont II shown by Marquardt 1966(compare Figure 8 in Marquardt 1966 [9]) is similar to invitro stages at 48 h pi [6 11] and we would assign them to
6 Journal of Parasitology Research
Muc
MuscSubmuc
V VV
V
C
FITCDAPI
FITCDAPI
FITCDAPI
FITCDAPITL TL TL
TL
lowast
lowast
lowastlowast
lowast
(a)
(b) (c)
(d)
(e) (f)
(g)
(h)
Figure 3 In situ localization of intestinal stages and third-generation merozoites (a) FITCDAPI overlay of B1C4 antibody probed section at72 h pi Fluorescence signals are limited to the middle and upper part of the crypts but were absent in the villi Bar 200 120583m (b) FITCDAPIoverlay of B1C4 antibody probed section at 72 h pi at higher magnification shows multinuclear schizonts Bar 25120583m (c) Transmitted light(TL) micrograph of third-generation merozoites At this time of infection they are characterized with a length of more than 25120583m and astraight cell shape and were found in the intestine at 117 h pi Bar 25120583m (d) TLmicrograph of B1C4 probed section at 120 h pi the enzymaticreaction of the secondary antibody linked peroxidase can be found in the lower and middle parts of the crypts (c) Signals are absent in thevilli (v) musc muscularis submuc submucosa muc mucosa Bar 200 120583m (e) TLmicrograph shows antibody derived peroxidase reaction inschizonts of the third-generation in the middle part of the crypts at 120 h pi The schizonts contain approximately 12 merozoites Bar 50120583m(f) TL micrograph shows schizonts of different shapes and sizes at 120 h pi The asterisk designates a schizont which contained more than12 merozoites and probably belongs to the second-generation schizont considering the stages occurred in vitro Bar 25 120583m (g) FITC labelledparasite stages at 144 h pi are visible in larger clusters in the middle and upper parts of the crypts and at the base of the atrophied villi Bar200 120583m (h) Higher magnifications revealed mono- and multinuclear stages (FITCDAPI overlay) The mononuclear stages (hash key) areprobably early macrogamonts and similar to them observed in vitro The multinuclear stages (asterisk) are probably schizonts of the fourthgeneration because the localization of nuclei is not limited to the periphery The corresponding DAPI image can be found in SupplementaryFigure 2 (see Supplementary Material available online at httpdxdoiorg1011552013591520) Bar 50 120583m
the first-generation schizont stage The schizont III in Figure11 shown in the publication by Marquardt 1966 [9] is similarto that we assign to the schizont II stages (Figure 1(a)) whichare visible in permanent cell lines beginning from 68 h pi(Figure 1(a)) A large schizont II shown by Tilley and Upton(compare Figure 7 in Tilley and Upton 1988 [10]) we wouldassign to schizont I stage A schizont III shown by Tilley andUpton 1988 (compare with Figure 8 in Tilley and Upton [10])we would assign to the schizont II stage Rick et al [6] alsodescribed large schizonts and long merozoites (21120583m) at 73 hpi and assigned them to the second schizont generationThe residual tandem dimeric tomato signal described byHanig et al [11] additionally supported our interpretationof the first- and second-generation stages occurring in thepermanent cell culture
Our observations around 117 h pi and 120 h pi (Figure 3)were conclusive to those described by Marquardt [9] Wecould also identify large merozoites about 25120583m and longer
with a high motility (Figure 3(c) Supplementary Figure 3)The merozoites moved forward and backward as previouslydescribed byMarquardt [9] Additionally we observedmove-ments ofmerozoites III with distances ofmore than their owncell length (Supplementary Figure 3) The model of glidingmotility would prefer the forward gliding [16] and in mostapicomplexan parasites this is observed in connection withbanana or arch-shaped parasite cells Backward moving isalso described in Plasmodium in a similar dimension [17]We think that the E nieschulzi merozoites perform this byrotating on their length axis during the gliding Otherwiseit would contradict the existing gliding motility model Thestraight cell shape would support a rotation hypothesis andwe observed that the merozoites almost used the same trailsduring their dislocation It is conceivable that through thehigh motility of this stage a further expansion of the parasitecan be accomplished In vitro we could not observe suchhigh motile stages with more than 25 120583m length in vitro
Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

2 Journal of Parasitology Research
2 Material and Methods
21 Cell Culture Pregnant rats (Crl CD (SD)) were eutha-nized at day 16ndash18 of gestation The fetuses were extractedand decapitated prior to further usage The decapitatedfetuses werewashed in cold (4∘C)DulbeccorsquosModified EaglersquosMedium (DMEM) to remove bloodThe fetuses were openedventrally and the inner organs were extracted Liver kidneyand intestine were separately collected and one assay wasperformed with all inner organs together as fetal mix (FM)All explanted tissues were washed twice with 37∘C preheatedDMEM minced into small pieces and ground througha sieve (mesh size 05mm) with a plunger The cellcellconglomerate suspension was transferred to a centrifugetube and washed once with completed DMEM (37∘C) andtransferred into completed preheated DMEM (with 10 FBS2 L-glutamine 2 penicillinstreptomycin 1 HEPESand 1 sodium pyruvate) The FM-suspension was seededin IBIDI 120583-slides with fibronectin (IBIDI) gelatin [12]Matrigel-coating (60 120583Lwell according to themanufacturerrsquosrecommendations) as well as in uncoated slides For primaryliver and kidney cells fibronectin coated IBIDI 120583-slides wereused Primary intestinal cells were seeded into IBIDI 120583-slides coated with fibronectin or gelatine as well as uncoatedslides Quality evaluation of the parasite development in vitrowas performed by comparing the following permanent celllines IEC6 (rat intestinal) VERO (green monkey kidney)ST (swine testicular) and IPEC-J2 (porcine small intestinalepithelial cell) which were routinely cultivated in completedDMEM For infection with parasites 100000ndash150000 ofthese cells were seeded in IBIDI 120583-slides per well Prior toinoculation with parasites permanent cells were incubatedabout 3-4 h and primary cell about 24 h at 37∘C and 5 CO
2
atmosphere
22 Parasites and Infection of Cells Oocysts of the passagesP2 and P3 of the previously described transgenic Eimerianieschulzi [11] were used in this study Sporozoites wereexcysted and purified as previously described [13] and resus-pended in cell culture medium (DMEM completed) Forinfection of the cells 20000ndash50000 sporozoites were incu-bated togetherwith the cells in the120583-slide at 37∘Cand 5CO
2
atmosphere Every day the cell culturemedium (DMEM)waspartially (50) removed and replaced with fresh mediumtwice
23 Immunohistology and Observation of Living IntestinalParasite Stages Rats (CD)were infected orally by gavagewith500000ndash1000000 sporulated oocysts of E nieschulzi andeuthanized at different time points after inoculation with theoocystsThe exact time points can be found in the descriptionof Figure 3 Living parasite stages were observed in smallbowel tissue smears in a 120583-slide
For histology the small intestine was cut into pieces of05 cm length and fixed in 4 paraformaldehyde1xPBS (pH74) for 24 h at 4∘C After washing in 1xPBS and dehydrationin a graded series of ethanol the tissue pieces were embeddedin paraffin Sections of 5120583mwere cutwith a rotarymicrotome(Leica RM2125RT) The sections were transferred to slides
and the paraffin was removed by Roti-Histol (Co Roth) Theantigen retrieval was performed with 01M sodium citratebuffer pH 60 for 30min at 90∘C If HRP-linked secondaryantibodies were used endogenous peroxidase was blocked byincubation with 1H
2O2in 1xPBS about 15min After block-
ing of unspecific antigenswith 20normal goat serum (NGS)and 005 Tween in 1xPBS about 1 h at room temperature thesections were incubated with hybridoma supernatant of theB1C4 clone [14] for about 2 h at room temperature (1 10dilution in 20NGS005Tween1xPBS) After the removalof the primary antibody the slides were incubated for 1 h atroom temperature with the secondary antibody diluted in20NGS005 Tween1xPBS (FITC conjugated anti-mouseIgG Co Sigma Aldrich F6257 1 100 or HRP conjugated anti-mouse IgG 1 200 Co Sigma Aldrich A9044) Fluorescenceprobed sections were washed counterstained with DAPI(1 120583gmL) about 10min and washed three times about 10minprior to mounting The sections labelled with Peroxidase(HRP) probes were equilibrated in 005M Tris buffer pH 76about 5min and further incubated in DABH
2O2staining
solution (5mg 331015840-Diaminobenzidine and 5 120583LH2O2in
10mLH2O) The staining was performed about 20 minutes
After removing all staining solutions the sections weremounted in MowiolDABCO according to the manufac-turerrsquos instructions Control experiments were performedwith infected tissue but 1xPBS instead the B1C4 antibody
24 Microscopy Images were taken using the Zeiss Axiovert100 microscope combined with the Olympus F-viewIImonochrome CCD-camera and the image processing soft-ware Cell F (Olympus) from 24 hndash270 h inoculation (pi)The fluorescence of intracellular as well as extracellular para-sitic stages was observed directly in the 120583-slides with FITCfilter set (Ex D48030) for the yellow fluorescent protein(YFP) and with a rhodamine filter set (Ex 56040) fortandem dimer tomato (TDT) For the visualization of theautofluorescence of the oocyst wall the DAPI filter set(excitation 365 nm emission long pass 420 nm)was usedThereproduction of images was performed comparable to themicroscopic view according to the filter set dependent emis-sionwavelength Capturing of the peroxidase probed sectionswas performed with the Olympus C7070 camera combinedwith the Olympus C5060-ADU ocular adapter or with themonochrome FviewII and CellF software
3 Results
In the permanent cell lines IEC6 VERO IPEC-J2 and STas well in primary liver kidney and intestinal cells thedevelopment stopped after formation of the second schizontsrespectively merozoite generation (not shown)We could notobserve any further development until 268 h pi in thesementioned cells and thereby we confirm the results ofprevious studies
Developmental stages beyond the second-generationmerozoites were observed in the fetal cell mix (FM) derivedfrom the inner organs on the coated slides but not in theuncoated control slide These results were confirmed by two
Journal of Parasitology Research 3
YFP
(a) 145 h pi
YFP
(b) 122 h pi
TL
(c) 122 h pi
YFP
(d) 216 h pi
YFP
(e) 164 h pi
YFP
(f) 180 h pi
YFP
(g) 218 h pi
TL
(h) 245 h pi
TDT
(i) 245 h pi
TL
(j) 245 h pi
TL
(k) 245 h pi
Figure 1 Development of the schizonts II and IV as well as clusters of macrogametocytes in organoid crypt-like structures in the fetal mixcell assay (a) Cluster of second-generation schizonts (YFP) which can be found early from 68 h pi and also late at 145 h pi (b) and (c)probably large schizont of the second-generation YFP (b) and transmitted light (c) (d) Probably merozoite of the second-generation (YFP)with a length of more than 20 120583m (22 120583m) Marquardt 1966 [9] and Roudabush 1937 [8] described a maximum length of 16 120583m (e) Youngfourth-generation schizonts (YFP) after few nuclear divisions (f) Clustered fourth generation schizonts (YFP) after multiple cell divisions aswells as young schizonts (g) Fourth generation merozoites (YFP) (h) Large cluster of around 100 gamonts (transmitted light micrograph)(i) Corresponding (to (h)) fluorescence micrograph of the gamonts with macrogamont specific expressed TDT signal YFP not shown (j)Transmitted light micrograph with gamonts (arrows) within cells of a crypt-like structure (k) Uninfected crypt-like structure (a) to (g) and(j) bars = 20120583m (h) (i) and (k) bars = 100 120583m
experimenters in separate experiments We have to remarkthat only in half of the performed experiments with coatedslides crypt-like organoid structures were observed Thesuccess of development of macrogamonts and oocysts wasdirectly connected with occurrence of these crypt-like struc-tures The particular experiments and the results of parasitedevelopment are listed in Table 1
In the fetal cell mix we could identify schizonts of thefirst- (not shown) second- (Figure 1(a)) and fourth-gener-ation (Figures 1(e) and 1(f)) as well as free merozoites ofthe second (Figure 1(d)) and fourth generation (Figure 1(g))
and also gametocytes and unsporulated oocysts (Figures1(h)ndash1(j) and 2)
The third-generation schizonts and merozoites are diffi-cult to distinguish from the second generation Consideringthe literature the large schizont in Figures 1(b) and 1(c) maybelong to the second or third generation Despite the size the22120583m long merozoite in Figure 1(d) would belong to thesecond generation This issue is explained in the followingdiscussion paragraph
Schizonts of the fourth generation were found clus-tered with up to 15 schizonts (picture details Figures 1(e)
4 Journal of Parasitology Research
Table 1 Number and type of experiments performed in this study
Cells Coating Cell growth Max parasitedevelopment
IPEC-J2 [2] Uncoated Monolayer Sch IIMz IIST [2] Uncoated Monolayer Sch IIMz IIVERO [gt2] Uncoated Monolayer later multilayer Sch IIMz IIIEC6 [gt2] Uncoated Monolayer Sch IIMz IIPrimintestine [3]Primintestine [1]Primintestine [1]
FibronectinGelatinUncoated
Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
Primary liver [2] Fibronectin Monolayer Sch IIMz IIPrimary kidney [2] Fibronectin Monolayer Sch IIMz II
Primary fetal mix [3] Fibronectin Monolayer and cell conglomerates in two experiments (1 2) no growingorganoids in one experiments (3) growing crypt-like organoids
(1) Sch IIMz II(2) Sch IIMz II(3) MagOoc
Primary fetal mix [2] Gelatin Monolayer and cell conglomerates in one experiment no organoids (1) inone experiment growing crypt-like organoids (2)
(1) Sch IIMz II(2) MagOoc
Primary fetal mix [1] Matrigel Monolayer and growing organoids MagOoc
Primary fetal mix (3) Uncoated Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
[number of experiments] (experiment number) Sch II schizont 2nd generation Mz II merozoite 2nd generation Mag macrogamont Ooc oocyst
and 1(f)) from 164 h pi Free merozoites IV were observed(Figure 1(g)) separately or close to the gametocytes (Figures2(a)ndash2(e) 2(j)ndash2(m)) Gametocytes show an additional redfluorescent tandem dimeric tomato signal The TDT-proteinis expressed under the microgametocyte-specific promoterof the gam56 gene found in Eimeria tenella [11 15] Thered fluorescent signal was found the earliest at 206 h piThe gametocytes often occurred in clusters with up to 100cells and grew in cells which were situated in a crypt-likeformation (Figures 1(h) 1(i) and 1(j)) However gametocyteswere not found in all crypt-like structures (Figure 1(k))
If crypt-like organoids had developed up to four clustersof macrogamonts in 25ndash50 of the 120583-slide wells could beidentified The best results were obtained in one experimentwith FM-cells on a fibronectin coated 120583-slide
Microgamonts or free microgametes were not identifiedwith certainty in the cell culture Notwithstanding unsporu-lated oocysts with autofluorescent oocyst walls could beobserved but these oocysts did not sporulate in the cell cul-ture slide or outside under air supply (Figures 2(n)ndash2(r))In summary the FM-cells provide the qualitatively potentialto develop Eimeria nieschulzi sporozoites over four asexualgenerations and the gametocyte stage to the point of oocystdevelopment in vitro The presence of macrogametocytes inorganoid crypt-like structure indicates that these cells havesimilar characteristics to the native host cells in the rat
In a selected number of performed experiments welocalized the wild type E nieschulzi in situ in the crypt of thesmall intestine by immunohistology at 72 120 and 144 h pi(Figure 3) The antibody B1C4 originally produced against Etenella sporozoites and described 1995 byGreif and Entzeroth[14] was used for immunohistology The antibody showedcross-reactivity to E nieschulzi schizonts and gamonts
At 72 h pi schizonts were detected in the middle parts ofthe crypts (Figures 3(a) and 3(b)) At higher magnificationsit was not possible to distinguish which schizont generation islabelled only the visible multiple nuclei indicated schizontsAt 117 h pi longmerozoites with a length ofmore than 25120583mand straight cell shape in the intestine (Figure 3(c)) wereidentified These merozoites belong to the third generationAt 120 h pi parasite stages were labelled in the middle andlower part of the crypts (Figure 3(d)) No parasites weredetected in the villi The schizonts shown in Figure 3(e)containing long merozoites belong to the third generationand were found in middle parts of the crypts Figure 3(f)shows schizonts of different sizes in the lower part ofthe crypts Marquardt [9] described large schizonts with amaximum of 12 merozoites as second or third generationHowever corresponding to the observed in vitro stages wewould assign them to the second-schizont-generation
At 144 h pi the parasite stageswere foundpredominantlyin the middle and upper part of the crypts as well as atthe base of the villi (Figure 3(g)) At higher magnificationsit was possible to identify mono- and multinuclear stagesThe mononuclear stages are probably young macrogamontsand the multinuclear stages fourth generation schizonts(Figure 3(h)) The control experiments without the primaryantibodies did not show any specific signal derived by thesecondary antibodies (not shown)
4 Discussion
In primary fetal cells the in vitro development of a rodentEimeria species up to the oocyst stage could be shown Asaffirmed by various cell lines in this study and in previous
Journal of Parasitology Research 5
YFP
(a) 206 h pi bar 20120583m
TDT
(b) 206 h pi bar 20120583m
YFP + TDT
(c) 206 h pi bar 20120583m
TL
(d) 206 h pi bar 20120583m
OL
(e) 206 h pi bar 20120583m
YFP
(f) 230 h pi bar 50120583m
TDT
(g) 230 h pi bar 50120583m
TL
(h) 230 h pi bar 50120583m
OL
(i) 230 h pi bar 50120583m
YFP
(j) 245 h pi bar 10120583m
TDT
(k) 245 h pi bar 10120583m
TL
(l) 245 h pi bar 10120583m
OL
(m) 245 h pi bar 10120583m
YFP
(n) 230 h pi bar 10120583m
TDT
(o) 230 h pi bar 10120583m
UV
(p) 230 h pi bar 10120583m
TL
(q) 230 h pi bar 10120583m
OL
(r) 230 h pi bar 10120583m
Figure 2 Development of macrogamonts and oocysts in primary fetal rat cells in vitro (a)ndash(e) Transgenic schizonts IV merozoites IV andmacrogamonts (a) YFP is expressed in all parasite stages (b) TDT is exclusively expressed in the macrogamonts (c) Overlay of (a) and (b)(d) Corresponding transmitted light micrograph with low contrasted parasite stages (e) Overlay of (a) (b) and (d) helps to identify the lowcontrasted gametocytes in the transmitted light micrograph and differentiating the asexual stages from the macrogamont (f)ndash(i) Cluster ofgamonts (f) YFP signal (g) TDT signal (h) Transmission light micrograph shows location of developed gamonts mainly in the periphery ofa crypt like structure An overlay of (f)ndash(h) is shown in panel (i) (j)ndash(m)Macrogamont stages adjacent to a mature schizont IV the YFP (j) isexpressed in all parasitic stages the TDT (k) in the macrogamonts The colocalization with the corresponding transmitted light micrograph(l) is shown in (m) (n)ndash(r) Formation of oocysts in vitro (n) YFP signals in unsporulated oocysts and gametocytes (o) CorrespondingTDT signals (p) Developed oocysts excited by UV light showing typical autofluorescent oocyst walls (q) Corresponding transmitted lightmicrograph (r) Overlay of (n)ndash(q)
studies the in vitro development of Eimeria nieschulzi islimited to the second-generation merozoite [5ndash7] Tilley andUpton [10] reported a development until the fourth gen-eration in vitro under reducing conditions However Eime-ria nieschulzi can have in vitro variations concerning themorphology of the schizont stages There are large schizontswith more than 20 merozoites [11] but also small schizontsless than 10 merozoites (Figure 1(a)) Thus it is difficult todistinguish the second schizont stage from the third schizontstage with certainty Because in vivo first- and second-generation schizonts can have delayed development up to96 h pi [8 9] the duration of the development cannot lead
to concrete conclusions Only the size of explicitly visiblemerozoites in or out of schizonts should be consulted fordetermination between second (MzII) and third-generationmerozoite (MzIII) but there are further challenges con-cerning the interpretation of literature data Marquardt [9]described the occurrence of MzIII beginning from 48 h piwith an average length of 208 120583m 255120583m at 72 h pi and269 120583m at 120 h pi At 120 h pi the merozoites showed ahigh motility while the previous smaller merozoites werenearly inactive The schizont II shown by Marquardt 1966(compare Figure 8 in Marquardt 1966 [9]) is similar to invitro stages at 48 h pi [6 11] and we would assign them to
6 Journal of Parasitology Research
Muc
MuscSubmuc
V VV
V
C
FITCDAPI
FITCDAPI
FITCDAPI
FITCDAPITL TL TL
TL
lowast
lowast
lowastlowast
lowast
(a)
(b) (c)
(d)
(e) (f)
(g)
(h)
Figure 3 In situ localization of intestinal stages and third-generation merozoites (a) FITCDAPI overlay of B1C4 antibody probed section at72 h pi Fluorescence signals are limited to the middle and upper part of the crypts but were absent in the villi Bar 200 120583m (b) FITCDAPIoverlay of B1C4 antibody probed section at 72 h pi at higher magnification shows multinuclear schizonts Bar 25120583m (c) Transmitted light(TL) micrograph of third-generation merozoites At this time of infection they are characterized with a length of more than 25120583m and astraight cell shape and were found in the intestine at 117 h pi Bar 25120583m (d) TLmicrograph of B1C4 probed section at 120 h pi the enzymaticreaction of the secondary antibody linked peroxidase can be found in the lower and middle parts of the crypts (c) Signals are absent in thevilli (v) musc muscularis submuc submucosa muc mucosa Bar 200 120583m (e) TLmicrograph shows antibody derived peroxidase reaction inschizonts of the third-generation in the middle part of the crypts at 120 h pi The schizonts contain approximately 12 merozoites Bar 50120583m(f) TL micrograph shows schizonts of different shapes and sizes at 120 h pi The asterisk designates a schizont which contained more than12 merozoites and probably belongs to the second-generation schizont considering the stages occurred in vitro Bar 25 120583m (g) FITC labelledparasite stages at 144 h pi are visible in larger clusters in the middle and upper parts of the crypts and at the base of the atrophied villi Bar200 120583m (h) Higher magnifications revealed mono- and multinuclear stages (FITCDAPI overlay) The mononuclear stages (hash key) areprobably early macrogamonts and similar to them observed in vitro The multinuclear stages (asterisk) are probably schizonts of the fourthgeneration because the localization of nuclei is not limited to the periphery The corresponding DAPI image can be found in SupplementaryFigure 2 (see Supplementary Material available online at httpdxdoiorg1011552013591520) Bar 50 120583m
the first-generation schizont stage The schizont III in Figure11 shown in the publication by Marquardt 1966 [9] is similarto that we assign to the schizont II stages (Figure 1(a)) whichare visible in permanent cell lines beginning from 68 h pi(Figure 1(a)) A large schizont II shown by Tilley and Upton(compare Figure 7 in Tilley and Upton 1988 [10]) we wouldassign to schizont I stage A schizont III shown by Tilley andUpton 1988 (compare with Figure 8 in Tilley and Upton [10])we would assign to the schizont II stage Rick et al [6] alsodescribed large schizonts and long merozoites (21120583m) at 73 hpi and assigned them to the second schizont generationThe residual tandem dimeric tomato signal described byHanig et al [11] additionally supported our interpretationof the first- and second-generation stages occurring in thepermanent cell culture
Our observations around 117 h pi and 120 h pi (Figure 3)were conclusive to those described by Marquardt [9] Wecould also identify large merozoites about 25120583m and longer
with a high motility (Figure 3(c) Supplementary Figure 3)The merozoites moved forward and backward as previouslydescribed byMarquardt [9] Additionally we observedmove-ments ofmerozoites III with distances ofmore than their owncell length (Supplementary Figure 3) The model of glidingmotility would prefer the forward gliding [16] and in mostapicomplexan parasites this is observed in connection withbanana or arch-shaped parasite cells Backward moving isalso described in Plasmodium in a similar dimension [17]We think that the E nieschulzi merozoites perform this byrotating on their length axis during the gliding Otherwiseit would contradict the existing gliding motility model Thestraight cell shape would support a rotation hypothesis andwe observed that the merozoites almost used the same trailsduring their dislocation It is conceivable that through thehigh motility of this stage a further expansion of the parasitecan be accomplished In vitro we could not observe suchhigh motile stages with more than 25 120583m length in vitro
Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

Journal of Parasitology Research 3
YFP
(a) 145 h pi
YFP
(b) 122 h pi
TL
(c) 122 h pi
YFP
(d) 216 h pi
YFP
(e) 164 h pi
YFP
(f) 180 h pi
YFP
(g) 218 h pi
TL
(h) 245 h pi
TDT
(i) 245 h pi
TL
(j) 245 h pi
TL
(k) 245 h pi
Figure 1 Development of the schizonts II and IV as well as clusters of macrogametocytes in organoid crypt-like structures in the fetal mixcell assay (a) Cluster of second-generation schizonts (YFP) which can be found early from 68 h pi and also late at 145 h pi (b) and (c)probably large schizont of the second-generation YFP (b) and transmitted light (c) (d) Probably merozoite of the second-generation (YFP)with a length of more than 20 120583m (22 120583m) Marquardt 1966 [9] and Roudabush 1937 [8] described a maximum length of 16 120583m (e) Youngfourth-generation schizonts (YFP) after few nuclear divisions (f) Clustered fourth generation schizonts (YFP) after multiple cell divisions aswells as young schizonts (g) Fourth generation merozoites (YFP) (h) Large cluster of around 100 gamonts (transmitted light micrograph)(i) Corresponding (to (h)) fluorescence micrograph of the gamonts with macrogamont specific expressed TDT signal YFP not shown (j)Transmitted light micrograph with gamonts (arrows) within cells of a crypt-like structure (k) Uninfected crypt-like structure (a) to (g) and(j) bars = 20120583m (h) (i) and (k) bars = 100 120583m
experimenters in separate experiments We have to remarkthat only in half of the performed experiments with coatedslides crypt-like organoid structures were observed Thesuccess of development of macrogamonts and oocysts wasdirectly connected with occurrence of these crypt-like struc-tures The particular experiments and the results of parasitedevelopment are listed in Table 1
In the fetal cell mix we could identify schizonts of thefirst- (not shown) second- (Figure 1(a)) and fourth-gener-ation (Figures 1(e) and 1(f)) as well as free merozoites ofthe second (Figure 1(d)) and fourth generation (Figure 1(g))
and also gametocytes and unsporulated oocysts (Figures1(h)ndash1(j) and 2)
The third-generation schizonts and merozoites are diffi-cult to distinguish from the second generation Consideringthe literature the large schizont in Figures 1(b) and 1(c) maybelong to the second or third generation Despite the size the22120583m long merozoite in Figure 1(d) would belong to thesecond generation This issue is explained in the followingdiscussion paragraph
Schizonts of the fourth generation were found clus-tered with up to 15 schizonts (picture details Figures 1(e)
4 Journal of Parasitology Research
Table 1 Number and type of experiments performed in this study
Cells Coating Cell growth Max parasitedevelopment
IPEC-J2 [2] Uncoated Monolayer Sch IIMz IIST [2] Uncoated Monolayer Sch IIMz IIVERO [gt2] Uncoated Monolayer later multilayer Sch IIMz IIIEC6 [gt2] Uncoated Monolayer Sch IIMz IIPrimintestine [3]Primintestine [1]Primintestine [1]
FibronectinGelatinUncoated
Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
Primary liver [2] Fibronectin Monolayer Sch IIMz IIPrimary kidney [2] Fibronectin Monolayer Sch IIMz II
Primary fetal mix [3] Fibronectin Monolayer and cell conglomerates in two experiments (1 2) no growingorganoids in one experiments (3) growing crypt-like organoids
(1) Sch IIMz II(2) Sch IIMz II(3) MagOoc
Primary fetal mix [2] Gelatin Monolayer and cell conglomerates in one experiment no organoids (1) inone experiment growing crypt-like organoids (2)
(1) Sch IIMz II(2) MagOoc
Primary fetal mix [1] Matrigel Monolayer and growing organoids MagOoc
Primary fetal mix (3) Uncoated Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
[number of experiments] (experiment number) Sch II schizont 2nd generation Mz II merozoite 2nd generation Mag macrogamont Ooc oocyst
and 1(f)) from 164 h pi Free merozoites IV were observed(Figure 1(g)) separately or close to the gametocytes (Figures2(a)ndash2(e) 2(j)ndash2(m)) Gametocytes show an additional redfluorescent tandem dimeric tomato signal The TDT-proteinis expressed under the microgametocyte-specific promoterof the gam56 gene found in Eimeria tenella [11 15] Thered fluorescent signal was found the earliest at 206 h piThe gametocytes often occurred in clusters with up to 100cells and grew in cells which were situated in a crypt-likeformation (Figures 1(h) 1(i) and 1(j)) However gametocyteswere not found in all crypt-like structures (Figure 1(k))
If crypt-like organoids had developed up to four clustersof macrogamonts in 25ndash50 of the 120583-slide wells could beidentified The best results were obtained in one experimentwith FM-cells on a fibronectin coated 120583-slide
Microgamonts or free microgametes were not identifiedwith certainty in the cell culture Notwithstanding unsporu-lated oocysts with autofluorescent oocyst walls could beobserved but these oocysts did not sporulate in the cell cul-ture slide or outside under air supply (Figures 2(n)ndash2(r))In summary the FM-cells provide the qualitatively potentialto develop Eimeria nieschulzi sporozoites over four asexualgenerations and the gametocyte stage to the point of oocystdevelopment in vitro The presence of macrogametocytes inorganoid crypt-like structure indicates that these cells havesimilar characteristics to the native host cells in the rat
In a selected number of performed experiments welocalized the wild type E nieschulzi in situ in the crypt of thesmall intestine by immunohistology at 72 120 and 144 h pi(Figure 3) The antibody B1C4 originally produced against Etenella sporozoites and described 1995 byGreif and Entzeroth[14] was used for immunohistology The antibody showedcross-reactivity to E nieschulzi schizonts and gamonts
At 72 h pi schizonts were detected in the middle parts ofthe crypts (Figures 3(a) and 3(b)) At higher magnificationsit was not possible to distinguish which schizont generation islabelled only the visible multiple nuclei indicated schizontsAt 117 h pi longmerozoites with a length ofmore than 25120583mand straight cell shape in the intestine (Figure 3(c)) wereidentified These merozoites belong to the third generationAt 120 h pi parasite stages were labelled in the middle andlower part of the crypts (Figure 3(d)) No parasites weredetected in the villi The schizonts shown in Figure 3(e)containing long merozoites belong to the third generationand were found in middle parts of the crypts Figure 3(f)shows schizonts of different sizes in the lower part ofthe crypts Marquardt [9] described large schizonts with amaximum of 12 merozoites as second or third generationHowever corresponding to the observed in vitro stages wewould assign them to the second-schizont-generation
At 144 h pi the parasite stageswere foundpredominantlyin the middle and upper part of the crypts as well as atthe base of the villi (Figure 3(g)) At higher magnificationsit was possible to identify mono- and multinuclear stagesThe mononuclear stages are probably young macrogamontsand the multinuclear stages fourth generation schizonts(Figure 3(h)) The control experiments without the primaryantibodies did not show any specific signal derived by thesecondary antibodies (not shown)
4 Discussion
In primary fetal cells the in vitro development of a rodentEimeria species up to the oocyst stage could be shown Asaffirmed by various cell lines in this study and in previous
Journal of Parasitology Research 5
YFP
(a) 206 h pi bar 20120583m
TDT
(b) 206 h pi bar 20120583m
YFP + TDT
(c) 206 h pi bar 20120583m
TL
(d) 206 h pi bar 20120583m
OL
(e) 206 h pi bar 20120583m
YFP
(f) 230 h pi bar 50120583m
TDT
(g) 230 h pi bar 50120583m
TL
(h) 230 h pi bar 50120583m
OL
(i) 230 h pi bar 50120583m
YFP
(j) 245 h pi bar 10120583m
TDT
(k) 245 h pi bar 10120583m
TL
(l) 245 h pi bar 10120583m
OL
(m) 245 h pi bar 10120583m
YFP
(n) 230 h pi bar 10120583m
TDT
(o) 230 h pi bar 10120583m
UV
(p) 230 h pi bar 10120583m
TL
(q) 230 h pi bar 10120583m
OL
(r) 230 h pi bar 10120583m
Figure 2 Development of macrogamonts and oocysts in primary fetal rat cells in vitro (a)ndash(e) Transgenic schizonts IV merozoites IV andmacrogamonts (a) YFP is expressed in all parasite stages (b) TDT is exclusively expressed in the macrogamonts (c) Overlay of (a) and (b)(d) Corresponding transmitted light micrograph with low contrasted parasite stages (e) Overlay of (a) (b) and (d) helps to identify the lowcontrasted gametocytes in the transmitted light micrograph and differentiating the asexual stages from the macrogamont (f)ndash(i) Cluster ofgamonts (f) YFP signal (g) TDT signal (h) Transmission light micrograph shows location of developed gamonts mainly in the periphery ofa crypt like structure An overlay of (f)ndash(h) is shown in panel (i) (j)ndash(m)Macrogamont stages adjacent to a mature schizont IV the YFP (j) isexpressed in all parasitic stages the TDT (k) in the macrogamonts The colocalization with the corresponding transmitted light micrograph(l) is shown in (m) (n)ndash(r) Formation of oocysts in vitro (n) YFP signals in unsporulated oocysts and gametocytes (o) CorrespondingTDT signals (p) Developed oocysts excited by UV light showing typical autofluorescent oocyst walls (q) Corresponding transmitted lightmicrograph (r) Overlay of (n)ndash(q)
studies the in vitro development of Eimeria nieschulzi islimited to the second-generation merozoite [5ndash7] Tilley andUpton [10] reported a development until the fourth gen-eration in vitro under reducing conditions However Eime-ria nieschulzi can have in vitro variations concerning themorphology of the schizont stages There are large schizontswith more than 20 merozoites [11] but also small schizontsless than 10 merozoites (Figure 1(a)) Thus it is difficult todistinguish the second schizont stage from the third schizontstage with certainty Because in vivo first- and second-generation schizonts can have delayed development up to96 h pi [8 9] the duration of the development cannot lead
to concrete conclusions Only the size of explicitly visiblemerozoites in or out of schizonts should be consulted fordetermination between second (MzII) and third-generationmerozoite (MzIII) but there are further challenges con-cerning the interpretation of literature data Marquardt [9]described the occurrence of MzIII beginning from 48 h piwith an average length of 208 120583m 255120583m at 72 h pi and269 120583m at 120 h pi At 120 h pi the merozoites showed ahigh motility while the previous smaller merozoites werenearly inactive The schizont II shown by Marquardt 1966(compare Figure 8 in Marquardt 1966 [9]) is similar to invitro stages at 48 h pi [6 11] and we would assign them to
6 Journal of Parasitology Research
Muc
MuscSubmuc
V VV
V
C
FITCDAPI
FITCDAPI
FITCDAPI
FITCDAPITL TL TL
TL
lowast
lowast
lowastlowast
lowast
(a)
(b) (c)
(d)
(e) (f)
(g)
(h)
Figure 3 In situ localization of intestinal stages and third-generation merozoites (a) FITCDAPI overlay of B1C4 antibody probed section at72 h pi Fluorescence signals are limited to the middle and upper part of the crypts but were absent in the villi Bar 200 120583m (b) FITCDAPIoverlay of B1C4 antibody probed section at 72 h pi at higher magnification shows multinuclear schizonts Bar 25120583m (c) Transmitted light(TL) micrograph of third-generation merozoites At this time of infection they are characterized with a length of more than 25120583m and astraight cell shape and were found in the intestine at 117 h pi Bar 25120583m (d) TLmicrograph of B1C4 probed section at 120 h pi the enzymaticreaction of the secondary antibody linked peroxidase can be found in the lower and middle parts of the crypts (c) Signals are absent in thevilli (v) musc muscularis submuc submucosa muc mucosa Bar 200 120583m (e) TLmicrograph shows antibody derived peroxidase reaction inschizonts of the third-generation in the middle part of the crypts at 120 h pi The schizonts contain approximately 12 merozoites Bar 50120583m(f) TL micrograph shows schizonts of different shapes and sizes at 120 h pi The asterisk designates a schizont which contained more than12 merozoites and probably belongs to the second-generation schizont considering the stages occurred in vitro Bar 25 120583m (g) FITC labelledparasite stages at 144 h pi are visible in larger clusters in the middle and upper parts of the crypts and at the base of the atrophied villi Bar200 120583m (h) Higher magnifications revealed mono- and multinuclear stages (FITCDAPI overlay) The mononuclear stages (hash key) areprobably early macrogamonts and similar to them observed in vitro The multinuclear stages (asterisk) are probably schizonts of the fourthgeneration because the localization of nuclei is not limited to the periphery The corresponding DAPI image can be found in SupplementaryFigure 2 (see Supplementary Material available online at httpdxdoiorg1011552013591520) Bar 50 120583m
the first-generation schizont stage The schizont III in Figure11 shown in the publication by Marquardt 1966 [9] is similarto that we assign to the schizont II stages (Figure 1(a)) whichare visible in permanent cell lines beginning from 68 h pi(Figure 1(a)) A large schizont II shown by Tilley and Upton(compare Figure 7 in Tilley and Upton 1988 [10]) we wouldassign to schizont I stage A schizont III shown by Tilley andUpton 1988 (compare with Figure 8 in Tilley and Upton [10])we would assign to the schizont II stage Rick et al [6] alsodescribed large schizonts and long merozoites (21120583m) at 73 hpi and assigned them to the second schizont generationThe residual tandem dimeric tomato signal described byHanig et al [11] additionally supported our interpretationof the first- and second-generation stages occurring in thepermanent cell culture
Our observations around 117 h pi and 120 h pi (Figure 3)were conclusive to those described by Marquardt [9] Wecould also identify large merozoites about 25120583m and longer
with a high motility (Figure 3(c) Supplementary Figure 3)The merozoites moved forward and backward as previouslydescribed byMarquardt [9] Additionally we observedmove-ments ofmerozoites III with distances ofmore than their owncell length (Supplementary Figure 3) The model of glidingmotility would prefer the forward gliding [16] and in mostapicomplexan parasites this is observed in connection withbanana or arch-shaped parasite cells Backward moving isalso described in Plasmodium in a similar dimension [17]We think that the E nieschulzi merozoites perform this byrotating on their length axis during the gliding Otherwiseit would contradict the existing gliding motility model Thestraight cell shape would support a rotation hypothesis andwe observed that the merozoites almost used the same trailsduring their dislocation It is conceivable that through thehigh motility of this stage a further expansion of the parasitecan be accomplished In vitro we could not observe suchhigh motile stages with more than 25 120583m length in vitro
Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

4 Journal of Parasitology Research
Table 1 Number and type of experiments performed in this study
Cells Coating Cell growth Max parasitedevelopment
IPEC-J2 [2] Uncoated Monolayer Sch IIMz IIST [2] Uncoated Monolayer Sch IIMz IIVERO [gt2] Uncoated Monolayer later multilayer Sch IIMz IIIEC6 [gt2] Uncoated Monolayer Sch IIMz IIPrimintestine [3]Primintestine [1]Primintestine [1]
FibronectinGelatinUncoated
Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
Primary liver [2] Fibronectin Monolayer Sch IIMz IIPrimary kidney [2] Fibronectin Monolayer Sch IIMz II
Primary fetal mix [3] Fibronectin Monolayer and cell conglomerates in two experiments (1 2) no growingorganoids in one experiments (3) growing crypt-like organoids
(1) Sch IIMz II(2) Sch IIMz II(3) MagOoc
Primary fetal mix [2] Gelatin Monolayer and cell conglomerates in one experiment no organoids (1) inone experiment growing crypt-like organoids (2)
(1) Sch IIMz II(2) MagOoc
Primary fetal mix [1] Matrigel Monolayer and growing organoids MagOoc
Primary fetal mix (3) Uncoated Monolayered fibroblast degrading cell conglomerates no growingorganoids Sch IIMz II
[number of experiments] (experiment number) Sch II schizont 2nd generation Mz II merozoite 2nd generation Mag macrogamont Ooc oocyst
and 1(f)) from 164 h pi Free merozoites IV were observed(Figure 1(g)) separately or close to the gametocytes (Figures2(a)ndash2(e) 2(j)ndash2(m)) Gametocytes show an additional redfluorescent tandem dimeric tomato signal The TDT-proteinis expressed under the microgametocyte-specific promoterof the gam56 gene found in Eimeria tenella [11 15] Thered fluorescent signal was found the earliest at 206 h piThe gametocytes often occurred in clusters with up to 100cells and grew in cells which were situated in a crypt-likeformation (Figures 1(h) 1(i) and 1(j)) However gametocyteswere not found in all crypt-like structures (Figure 1(k))
If crypt-like organoids had developed up to four clustersof macrogamonts in 25ndash50 of the 120583-slide wells could beidentified The best results were obtained in one experimentwith FM-cells on a fibronectin coated 120583-slide
Microgamonts or free microgametes were not identifiedwith certainty in the cell culture Notwithstanding unsporu-lated oocysts with autofluorescent oocyst walls could beobserved but these oocysts did not sporulate in the cell cul-ture slide or outside under air supply (Figures 2(n)ndash2(r))In summary the FM-cells provide the qualitatively potentialto develop Eimeria nieschulzi sporozoites over four asexualgenerations and the gametocyte stage to the point of oocystdevelopment in vitro The presence of macrogametocytes inorganoid crypt-like structure indicates that these cells havesimilar characteristics to the native host cells in the rat
In a selected number of performed experiments welocalized the wild type E nieschulzi in situ in the crypt of thesmall intestine by immunohistology at 72 120 and 144 h pi(Figure 3) The antibody B1C4 originally produced against Etenella sporozoites and described 1995 byGreif and Entzeroth[14] was used for immunohistology The antibody showedcross-reactivity to E nieschulzi schizonts and gamonts
At 72 h pi schizonts were detected in the middle parts ofthe crypts (Figures 3(a) and 3(b)) At higher magnificationsit was not possible to distinguish which schizont generation islabelled only the visible multiple nuclei indicated schizontsAt 117 h pi longmerozoites with a length ofmore than 25120583mand straight cell shape in the intestine (Figure 3(c)) wereidentified These merozoites belong to the third generationAt 120 h pi parasite stages were labelled in the middle andlower part of the crypts (Figure 3(d)) No parasites weredetected in the villi The schizonts shown in Figure 3(e)containing long merozoites belong to the third generationand were found in middle parts of the crypts Figure 3(f)shows schizonts of different sizes in the lower part ofthe crypts Marquardt [9] described large schizonts with amaximum of 12 merozoites as second or third generationHowever corresponding to the observed in vitro stages wewould assign them to the second-schizont-generation
At 144 h pi the parasite stageswere foundpredominantlyin the middle and upper part of the crypts as well as atthe base of the villi (Figure 3(g)) At higher magnificationsit was possible to identify mono- and multinuclear stagesThe mononuclear stages are probably young macrogamontsand the multinuclear stages fourth generation schizonts(Figure 3(h)) The control experiments without the primaryantibodies did not show any specific signal derived by thesecondary antibodies (not shown)
4 Discussion
In primary fetal cells the in vitro development of a rodentEimeria species up to the oocyst stage could be shown Asaffirmed by various cell lines in this study and in previous
Journal of Parasitology Research 5
YFP
(a) 206 h pi bar 20120583m
TDT
(b) 206 h pi bar 20120583m
YFP + TDT
(c) 206 h pi bar 20120583m
TL
(d) 206 h pi bar 20120583m
OL
(e) 206 h pi bar 20120583m
YFP
(f) 230 h pi bar 50120583m
TDT
(g) 230 h pi bar 50120583m
TL
(h) 230 h pi bar 50120583m
OL
(i) 230 h pi bar 50120583m
YFP
(j) 245 h pi bar 10120583m
TDT
(k) 245 h pi bar 10120583m
TL
(l) 245 h pi bar 10120583m
OL
(m) 245 h pi bar 10120583m
YFP
(n) 230 h pi bar 10120583m
TDT
(o) 230 h pi bar 10120583m
UV
(p) 230 h pi bar 10120583m
TL
(q) 230 h pi bar 10120583m
OL
(r) 230 h pi bar 10120583m
Figure 2 Development of macrogamonts and oocysts in primary fetal rat cells in vitro (a)ndash(e) Transgenic schizonts IV merozoites IV andmacrogamonts (a) YFP is expressed in all parasite stages (b) TDT is exclusively expressed in the macrogamonts (c) Overlay of (a) and (b)(d) Corresponding transmitted light micrograph with low contrasted parasite stages (e) Overlay of (a) (b) and (d) helps to identify the lowcontrasted gametocytes in the transmitted light micrograph and differentiating the asexual stages from the macrogamont (f)ndash(i) Cluster ofgamonts (f) YFP signal (g) TDT signal (h) Transmission light micrograph shows location of developed gamonts mainly in the periphery ofa crypt like structure An overlay of (f)ndash(h) is shown in panel (i) (j)ndash(m)Macrogamont stages adjacent to a mature schizont IV the YFP (j) isexpressed in all parasitic stages the TDT (k) in the macrogamonts The colocalization with the corresponding transmitted light micrograph(l) is shown in (m) (n)ndash(r) Formation of oocysts in vitro (n) YFP signals in unsporulated oocysts and gametocytes (o) CorrespondingTDT signals (p) Developed oocysts excited by UV light showing typical autofluorescent oocyst walls (q) Corresponding transmitted lightmicrograph (r) Overlay of (n)ndash(q)
studies the in vitro development of Eimeria nieschulzi islimited to the second-generation merozoite [5ndash7] Tilley andUpton [10] reported a development until the fourth gen-eration in vitro under reducing conditions However Eime-ria nieschulzi can have in vitro variations concerning themorphology of the schizont stages There are large schizontswith more than 20 merozoites [11] but also small schizontsless than 10 merozoites (Figure 1(a)) Thus it is difficult todistinguish the second schizont stage from the third schizontstage with certainty Because in vivo first- and second-generation schizonts can have delayed development up to96 h pi [8 9] the duration of the development cannot lead
to concrete conclusions Only the size of explicitly visiblemerozoites in or out of schizonts should be consulted fordetermination between second (MzII) and third-generationmerozoite (MzIII) but there are further challenges con-cerning the interpretation of literature data Marquardt [9]described the occurrence of MzIII beginning from 48 h piwith an average length of 208 120583m 255120583m at 72 h pi and269 120583m at 120 h pi At 120 h pi the merozoites showed ahigh motility while the previous smaller merozoites werenearly inactive The schizont II shown by Marquardt 1966(compare Figure 8 in Marquardt 1966 [9]) is similar to invitro stages at 48 h pi [6 11] and we would assign them to
6 Journal of Parasitology Research
Muc
MuscSubmuc
V VV
V
C
FITCDAPI
FITCDAPI
FITCDAPI
FITCDAPITL TL TL
TL
lowast
lowast
lowastlowast
lowast
(a)
(b) (c)
(d)
(e) (f)
(g)
(h)
Figure 3 In situ localization of intestinal stages and third-generation merozoites (a) FITCDAPI overlay of B1C4 antibody probed section at72 h pi Fluorescence signals are limited to the middle and upper part of the crypts but were absent in the villi Bar 200 120583m (b) FITCDAPIoverlay of B1C4 antibody probed section at 72 h pi at higher magnification shows multinuclear schizonts Bar 25120583m (c) Transmitted light(TL) micrograph of third-generation merozoites At this time of infection they are characterized with a length of more than 25120583m and astraight cell shape and were found in the intestine at 117 h pi Bar 25120583m (d) TLmicrograph of B1C4 probed section at 120 h pi the enzymaticreaction of the secondary antibody linked peroxidase can be found in the lower and middle parts of the crypts (c) Signals are absent in thevilli (v) musc muscularis submuc submucosa muc mucosa Bar 200 120583m (e) TLmicrograph shows antibody derived peroxidase reaction inschizonts of the third-generation in the middle part of the crypts at 120 h pi The schizonts contain approximately 12 merozoites Bar 50120583m(f) TL micrograph shows schizonts of different shapes and sizes at 120 h pi The asterisk designates a schizont which contained more than12 merozoites and probably belongs to the second-generation schizont considering the stages occurred in vitro Bar 25 120583m (g) FITC labelledparasite stages at 144 h pi are visible in larger clusters in the middle and upper parts of the crypts and at the base of the atrophied villi Bar200 120583m (h) Higher magnifications revealed mono- and multinuclear stages (FITCDAPI overlay) The mononuclear stages (hash key) areprobably early macrogamonts and similar to them observed in vitro The multinuclear stages (asterisk) are probably schizonts of the fourthgeneration because the localization of nuclei is not limited to the periphery The corresponding DAPI image can be found in SupplementaryFigure 2 (see Supplementary Material available online at httpdxdoiorg1011552013591520) Bar 50 120583m
the first-generation schizont stage The schizont III in Figure11 shown in the publication by Marquardt 1966 [9] is similarto that we assign to the schizont II stages (Figure 1(a)) whichare visible in permanent cell lines beginning from 68 h pi(Figure 1(a)) A large schizont II shown by Tilley and Upton(compare Figure 7 in Tilley and Upton 1988 [10]) we wouldassign to schizont I stage A schizont III shown by Tilley andUpton 1988 (compare with Figure 8 in Tilley and Upton [10])we would assign to the schizont II stage Rick et al [6] alsodescribed large schizonts and long merozoites (21120583m) at 73 hpi and assigned them to the second schizont generationThe residual tandem dimeric tomato signal described byHanig et al [11] additionally supported our interpretationof the first- and second-generation stages occurring in thepermanent cell culture
Our observations around 117 h pi and 120 h pi (Figure 3)were conclusive to those described by Marquardt [9] Wecould also identify large merozoites about 25120583m and longer
with a high motility (Figure 3(c) Supplementary Figure 3)The merozoites moved forward and backward as previouslydescribed byMarquardt [9] Additionally we observedmove-ments ofmerozoites III with distances ofmore than their owncell length (Supplementary Figure 3) The model of glidingmotility would prefer the forward gliding [16] and in mostapicomplexan parasites this is observed in connection withbanana or arch-shaped parasite cells Backward moving isalso described in Plasmodium in a similar dimension [17]We think that the E nieschulzi merozoites perform this byrotating on their length axis during the gliding Otherwiseit would contradict the existing gliding motility model Thestraight cell shape would support a rotation hypothesis andwe observed that the merozoites almost used the same trailsduring their dislocation It is conceivable that through thehigh motility of this stage a further expansion of the parasitecan be accomplished In vitro we could not observe suchhigh motile stages with more than 25 120583m length in vitro
Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
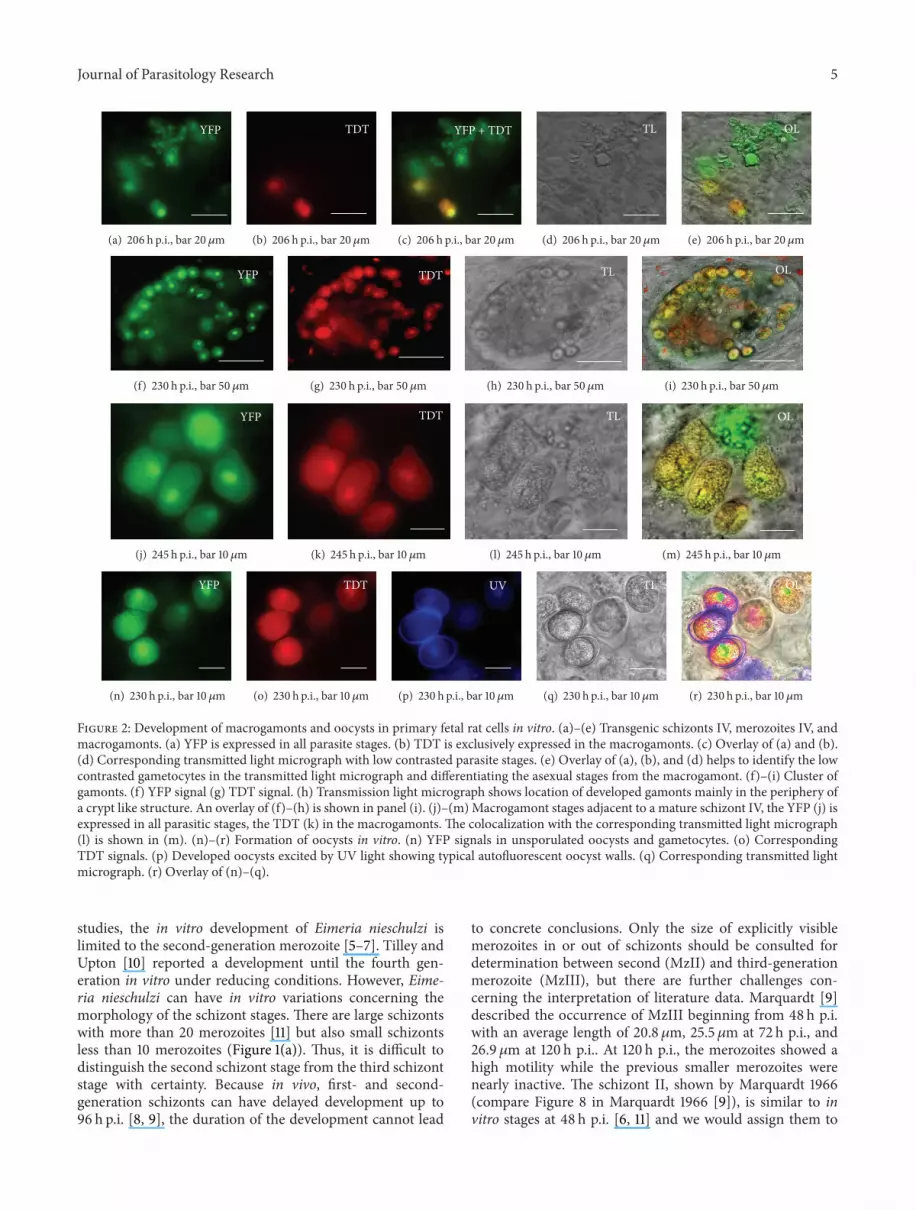
Journal of Parasitology Research 5
YFP
(a) 206 h pi bar 20120583m
TDT
(b) 206 h pi bar 20120583m
YFP + TDT
(c) 206 h pi bar 20120583m
TL
(d) 206 h pi bar 20120583m
OL
(e) 206 h pi bar 20120583m
YFP
(f) 230 h pi bar 50120583m
TDT
(g) 230 h pi bar 50120583m
TL
(h) 230 h pi bar 50120583m
OL
(i) 230 h pi bar 50120583m
YFP
(j) 245 h pi bar 10120583m
TDT
(k) 245 h pi bar 10120583m
TL
(l) 245 h pi bar 10120583m
OL
(m) 245 h pi bar 10120583m
YFP
(n) 230 h pi bar 10120583m
TDT
(o) 230 h pi bar 10120583m
UV
(p) 230 h pi bar 10120583m
TL
(q) 230 h pi bar 10120583m
OL
(r) 230 h pi bar 10120583m
Figure 2 Development of macrogamonts and oocysts in primary fetal rat cells in vitro (a)ndash(e) Transgenic schizonts IV merozoites IV andmacrogamonts (a) YFP is expressed in all parasite stages (b) TDT is exclusively expressed in the macrogamonts (c) Overlay of (a) and (b)(d) Corresponding transmitted light micrograph with low contrasted parasite stages (e) Overlay of (a) (b) and (d) helps to identify the lowcontrasted gametocytes in the transmitted light micrograph and differentiating the asexual stages from the macrogamont (f)ndash(i) Cluster ofgamonts (f) YFP signal (g) TDT signal (h) Transmission light micrograph shows location of developed gamonts mainly in the periphery ofa crypt like structure An overlay of (f)ndash(h) is shown in panel (i) (j)ndash(m)Macrogamont stages adjacent to a mature schizont IV the YFP (j) isexpressed in all parasitic stages the TDT (k) in the macrogamonts The colocalization with the corresponding transmitted light micrograph(l) is shown in (m) (n)ndash(r) Formation of oocysts in vitro (n) YFP signals in unsporulated oocysts and gametocytes (o) CorrespondingTDT signals (p) Developed oocysts excited by UV light showing typical autofluorescent oocyst walls (q) Corresponding transmitted lightmicrograph (r) Overlay of (n)ndash(q)
studies the in vitro development of Eimeria nieschulzi islimited to the second-generation merozoite [5ndash7] Tilley andUpton [10] reported a development until the fourth gen-eration in vitro under reducing conditions However Eime-ria nieschulzi can have in vitro variations concerning themorphology of the schizont stages There are large schizontswith more than 20 merozoites [11] but also small schizontsless than 10 merozoites (Figure 1(a)) Thus it is difficult todistinguish the second schizont stage from the third schizontstage with certainty Because in vivo first- and second-generation schizonts can have delayed development up to96 h pi [8 9] the duration of the development cannot lead
to concrete conclusions Only the size of explicitly visiblemerozoites in or out of schizonts should be consulted fordetermination between second (MzII) and third-generationmerozoite (MzIII) but there are further challenges con-cerning the interpretation of literature data Marquardt [9]described the occurrence of MzIII beginning from 48 h piwith an average length of 208 120583m 255120583m at 72 h pi and269 120583m at 120 h pi At 120 h pi the merozoites showed ahigh motility while the previous smaller merozoites werenearly inactive The schizont II shown by Marquardt 1966(compare Figure 8 in Marquardt 1966 [9]) is similar to invitro stages at 48 h pi [6 11] and we would assign them to
6 Journal of Parasitology Research
Muc
MuscSubmuc
V VV
V
C
FITCDAPI
FITCDAPI
FITCDAPI
FITCDAPITL TL TL
TL
lowast
lowast
lowastlowast
lowast
(a)
(b) (c)
(d)
(e) (f)
(g)
(h)
Figure 3 In situ localization of intestinal stages and third-generation merozoites (a) FITCDAPI overlay of B1C4 antibody probed section at72 h pi Fluorescence signals are limited to the middle and upper part of the crypts but were absent in the villi Bar 200 120583m (b) FITCDAPIoverlay of B1C4 antibody probed section at 72 h pi at higher magnification shows multinuclear schizonts Bar 25120583m (c) Transmitted light(TL) micrograph of third-generation merozoites At this time of infection they are characterized with a length of more than 25120583m and astraight cell shape and were found in the intestine at 117 h pi Bar 25120583m (d) TLmicrograph of B1C4 probed section at 120 h pi the enzymaticreaction of the secondary antibody linked peroxidase can be found in the lower and middle parts of the crypts (c) Signals are absent in thevilli (v) musc muscularis submuc submucosa muc mucosa Bar 200 120583m (e) TLmicrograph shows antibody derived peroxidase reaction inschizonts of the third-generation in the middle part of the crypts at 120 h pi The schizonts contain approximately 12 merozoites Bar 50120583m(f) TL micrograph shows schizonts of different shapes and sizes at 120 h pi The asterisk designates a schizont which contained more than12 merozoites and probably belongs to the second-generation schizont considering the stages occurred in vitro Bar 25 120583m (g) FITC labelledparasite stages at 144 h pi are visible in larger clusters in the middle and upper parts of the crypts and at the base of the atrophied villi Bar200 120583m (h) Higher magnifications revealed mono- and multinuclear stages (FITCDAPI overlay) The mononuclear stages (hash key) areprobably early macrogamonts and similar to them observed in vitro The multinuclear stages (asterisk) are probably schizonts of the fourthgeneration because the localization of nuclei is not limited to the periphery The corresponding DAPI image can be found in SupplementaryFigure 2 (see Supplementary Material available online at httpdxdoiorg1011552013591520) Bar 50 120583m
the first-generation schizont stage The schizont III in Figure11 shown in the publication by Marquardt 1966 [9] is similarto that we assign to the schizont II stages (Figure 1(a)) whichare visible in permanent cell lines beginning from 68 h pi(Figure 1(a)) A large schizont II shown by Tilley and Upton(compare Figure 7 in Tilley and Upton 1988 [10]) we wouldassign to schizont I stage A schizont III shown by Tilley andUpton 1988 (compare with Figure 8 in Tilley and Upton [10])we would assign to the schizont II stage Rick et al [6] alsodescribed large schizonts and long merozoites (21120583m) at 73 hpi and assigned them to the second schizont generationThe residual tandem dimeric tomato signal described byHanig et al [11] additionally supported our interpretationof the first- and second-generation stages occurring in thepermanent cell culture
Our observations around 117 h pi and 120 h pi (Figure 3)were conclusive to those described by Marquardt [9] Wecould also identify large merozoites about 25120583m and longer
with a high motility (Figure 3(c) Supplementary Figure 3)The merozoites moved forward and backward as previouslydescribed byMarquardt [9] Additionally we observedmove-ments ofmerozoites III with distances ofmore than their owncell length (Supplementary Figure 3) The model of glidingmotility would prefer the forward gliding [16] and in mostapicomplexan parasites this is observed in connection withbanana or arch-shaped parasite cells Backward moving isalso described in Plasmodium in a similar dimension [17]We think that the E nieschulzi merozoites perform this byrotating on their length axis during the gliding Otherwiseit would contradict the existing gliding motility model Thestraight cell shape would support a rotation hypothesis andwe observed that the merozoites almost used the same trailsduring their dislocation It is conceivable that through thehigh motility of this stage a further expansion of the parasitecan be accomplished In vitro we could not observe suchhigh motile stages with more than 25 120583m length in vitro
Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
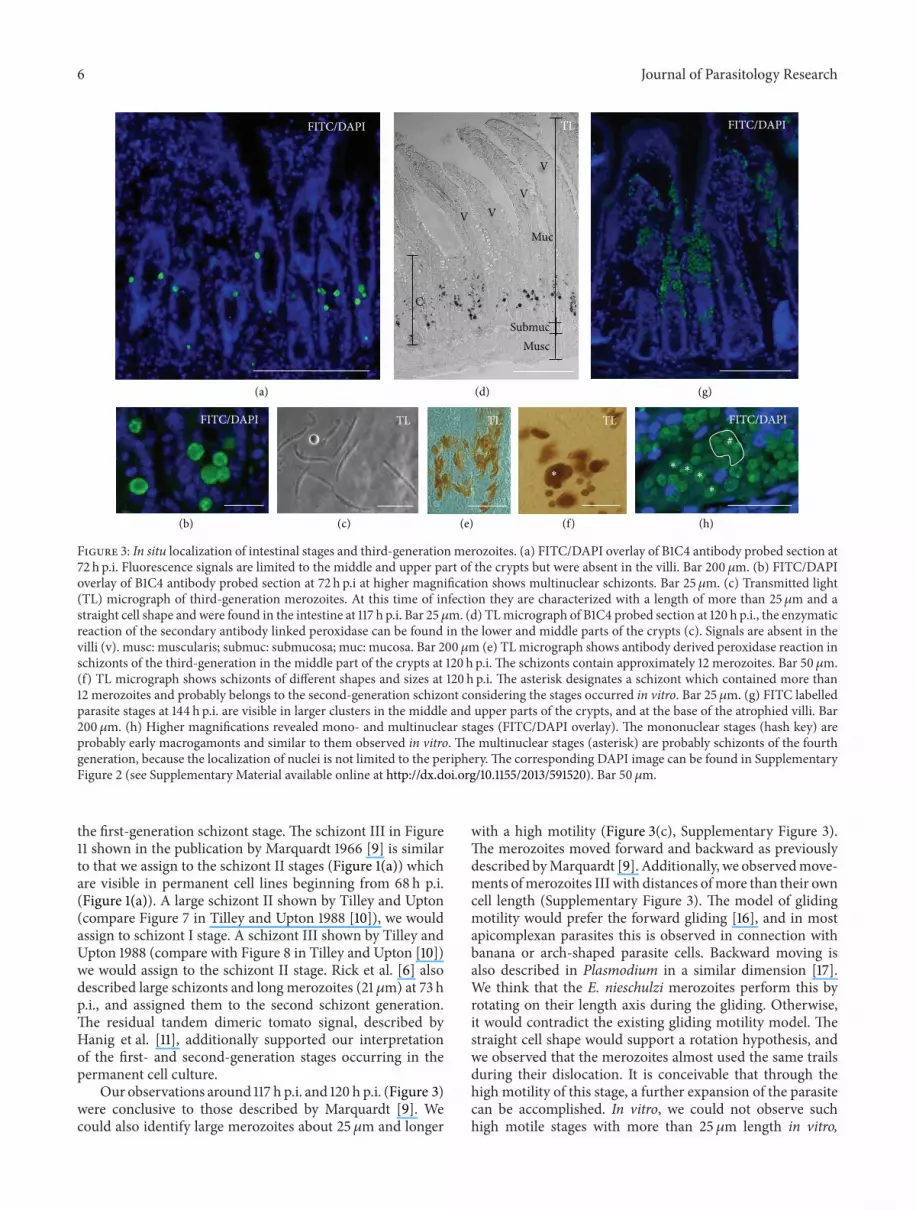
6 Journal of Parasitology Research
Muc
MuscSubmuc
V VV
V
C
FITCDAPI
FITCDAPI
FITCDAPI
FITCDAPITL TL TL
TL
lowast
lowast
lowastlowast
lowast
(a)
(b) (c)
(d)
(e) (f)
(g)
(h)
Figure 3 In situ localization of intestinal stages and third-generation merozoites (a) FITCDAPI overlay of B1C4 antibody probed section at72 h pi Fluorescence signals are limited to the middle and upper part of the crypts but were absent in the villi Bar 200 120583m (b) FITCDAPIoverlay of B1C4 antibody probed section at 72 h pi at higher magnification shows multinuclear schizonts Bar 25120583m (c) Transmitted light(TL) micrograph of third-generation merozoites At this time of infection they are characterized with a length of more than 25120583m and astraight cell shape and were found in the intestine at 117 h pi Bar 25120583m (d) TLmicrograph of B1C4 probed section at 120 h pi the enzymaticreaction of the secondary antibody linked peroxidase can be found in the lower and middle parts of the crypts (c) Signals are absent in thevilli (v) musc muscularis submuc submucosa muc mucosa Bar 200 120583m (e) TLmicrograph shows antibody derived peroxidase reaction inschizonts of the third-generation in the middle part of the crypts at 120 h pi The schizonts contain approximately 12 merozoites Bar 50120583m(f) TL micrograph shows schizonts of different shapes and sizes at 120 h pi The asterisk designates a schizont which contained more than12 merozoites and probably belongs to the second-generation schizont considering the stages occurred in vitro Bar 25 120583m (g) FITC labelledparasite stages at 144 h pi are visible in larger clusters in the middle and upper parts of the crypts and at the base of the atrophied villi Bar200 120583m (h) Higher magnifications revealed mono- and multinuclear stages (FITCDAPI overlay) The mononuclear stages (hash key) areprobably early macrogamonts and similar to them observed in vitro The multinuclear stages (asterisk) are probably schizonts of the fourthgeneration because the localization of nuclei is not limited to the periphery The corresponding DAPI image can be found in SupplementaryFigure 2 (see Supplementary Material available online at httpdxdoiorg1011552013591520) Bar 50 120583m
the first-generation schizont stage The schizont III in Figure11 shown in the publication by Marquardt 1966 [9] is similarto that we assign to the schizont II stages (Figure 1(a)) whichare visible in permanent cell lines beginning from 68 h pi(Figure 1(a)) A large schizont II shown by Tilley and Upton(compare Figure 7 in Tilley and Upton 1988 [10]) we wouldassign to schizont I stage A schizont III shown by Tilley andUpton 1988 (compare with Figure 8 in Tilley and Upton [10])we would assign to the schizont II stage Rick et al [6] alsodescribed large schizonts and long merozoites (21120583m) at 73 hpi and assigned them to the second schizont generationThe residual tandem dimeric tomato signal described byHanig et al [11] additionally supported our interpretationof the first- and second-generation stages occurring in thepermanent cell culture
Our observations around 117 h pi and 120 h pi (Figure 3)were conclusive to those described by Marquardt [9] Wecould also identify large merozoites about 25120583m and longer
with a high motility (Figure 3(c) Supplementary Figure 3)The merozoites moved forward and backward as previouslydescribed byMarquardt [9] Additionally we observedmove-ments ofmerozoites III with distances ofmore than their owncell length (Supplementary Figure 3) The model of glidingmotility would prefer the forward gliding [16] and in mostapicomplexan parasites this is observed in connection withbanana or arch-shaped parasite cells Backward moving isalso described in Plasmodium in a similar dimension [17]We think that the E nieschulzi merozoites perform this byrotating on their length axis during the gliding Otherwiseit would contradict the existing gliding motility model Thestraight cell shape would support a rotation hypothesis andwe observed that the merozoites almost used the same trailsduring their dislocation It is conceivable that through thehigh motility of this stage a further expansion of the parasitecan be accomplished In vitro we could not observe suchhigh motile stages with more than 25 120583m length in vitro
Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

Journal of Parasitology Research 7
and we are not sure if very large schizonts in Figures 1(b)and 1(c) belong to the third-generation Third generationschizonts probably occur in very low numbers in our cellculture which is discussed below Marquardt [9] observedthe fourth generation only at day six and described a very fasttransition frommerozoite III tomerozoite IV In combinationwith its description of merozoites III at 48 and 72 h pi themerozoites IV would be expected earlier Hence we haveadditional doubts concerning the early merozoites III whichare probably merozoites II To shed light on that issuefurther studies with modern molecular methods should beperformed
In size and shape the schizonts of the fourth-generationare comparable to the first-generation schizonts originated bysingle sporozoites The merozoites IV are smaller than themerozoites I [8 9] In contrast to the isolated appearanceof the first schizont stage the fourth-generation schizontsoccurred in a cluster observed from 164 h pi This is causedby infection of merozoites III close to their original host cellThe assumed fourth-generation schizont shown by Tilley andUpton [10] is probably a first or second-generation schizontin a delaminated host cell Our experiments suggest that areduced environment is not necessary for the development ofEimeria nieschulzi in vitro It is assumed that the developmentof the third-generation schizont stage along with the invasionof the second-generation merozoites is a crucial point for thein vitro development In order to invade and further developinto schizonts of the third-generation at least the merozoitesII obviously need special host cell requirements In perma-nent cell lines and primary liver and kidney cells no furtherdevelopment beyond the second-generation merozoite wasobserved The second-generation schizont forms clusters too(Figure 1(a)) and this would also be expected for the thirdgeneration In the nonhomogenous fetal cell mix used in thisstudy only a few of the merozoites II seem to find a properhost cell for invasion
This would explain the nonoccurrence of clusters inthe third-generation schizonts Thus parasite expansionwithin the third-generation schizont in cell culture would berepressed This interpretation is supported by the occurrenceof macrogamonts in only a part of the cell culture slide wells
However once in a proper cellular environment (crypt-like organoid) the majority of the developed merozoites III(derived from a single schizont III) can infect a new hostcell An expansion of parasitic stages occurred once moreand merozoites III developed into large clusters of schizontsIV (Figures 1(e) and 1(f)) If merozoites IV are infecting newproper host cells they would form even larger clusters ofmacrogamonts and this is shown in this study
In the natural host Eimeria nieschulzi directly infectscrypt cells of the small intestine and can be also found incells at the basis of the villi [8] In other parts of the epithe-lium they are absent We could confirm these original find-ings (Figure 3) The majority of crypt cells are known to beundifferentiated or partly differentiated They are so-calledtransit-amplifying cells (TA-cells) proliferative progenitorcells derived from stem cells at the base of the crypt Inside thevilli these formerTA-cells are fully differentiated into absorp-tive cells and goblet cells [18] These cells are not infected
by Eimeria nieschulzi but the TA-cells obviously harbor anattractor for the parasite In the in vitro experiments shownhere the majority of the developed macrogamonts could beobserved in crypt-like structures This suggests that thesecells have similar properties to the intestinal crypt cells in thehost These crypt-like structures were only observed in theFM-cell assay andmay be based on the necessity of epithelial-mesenchymal interactions for the development and prolifer-ation of crypt cells [19] These mesenchymal cells were foundin the FM-cell assay but not in the assay with only primaryintestinal cells This is probably the reason why intestinalcells solely do not form crypt-like organoids Cell cultureslides coated with Matrigel fibronectin or gelatine may benecessary for the growth of the FM-cells and the formationof crypt-like structures while the cell culture medium seemsto be less important Nevertheless the formation of crypt-like organoid occurred not absolutely reliable (Table 1) TheFM cells are a variable undefined cell mixture with differentcell content in every preparation and this can influence thecell growth Furthermore a high content of red blood cellsseems to negatively affect the growth of the primary cells andthis content alternates also in the cell preparations Thus infurther experiments the growth of the crypt-like organoidsshould be aspired in a more defined manner
The possibility to build crypt-villous structures in vitrofrom single Lgr5+ stem cells without a mesenchymal nicheby stimulating the cell proliferation with external factors wasshown 2009 by Sato et al [20] in the mouse model Theirresults could give further perspectives for the in vitro culti-vation of potentially crypt cell affine Eimeria species
The molecules involved in invasion of Eimeria parasitesmore precisely Eimeria tenella are relatively well known [2122] However there is a little characterization of the surfacemolecules and gene expression of the natural host cellsEimeria species can infect multiple cell lines but stop theirdevelopment finally after one or two schizont generationsEstablished cell lines seem to be only partly sufficient toexplore the involved molecules for infection of host cellsbecause they only harbour a minimum of qualities for infec-tion anddevelopment of the parasites It is striking that in vivoE nieschulzi directly infects the cell of the crypts and passesthe villous cells whereas in vitro they infect several types ofepithelial cells Evidently there are factors of the villous cellswhich prevent an infection and factors of the crypt cells whichtrigger an infection in vivo Sialylated glycans on the host cellsurface and MIC3 protein on the parasites side interact andmay play an important role in host cell tropism of Eimeriatenella [23] However a colocalization of E tenella stages andthe lectin MAAII (a lectin binding to 1205722ndash6 or 1205722-3 sialyllinked glycan) positive cells as well as the existence of suchsialylated glycans in primary chicken kidney cells where acomplete development of E tenella in vitro is possible wasnot shown Therefore it is still unclear if sialylated glycansare the most important mediators for host cell recognitionThus the characterization and isolation of natural host cellshave to be improved to designate the host cell ligands neededfor infection and development Genetically modified Eimeriaspecies are helpful for these studies
8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

8 Journal of Parasitology Research
Furthermore it was remarkable that no microgamonts ormicrogametes in FM-cell culture were definitively observedwhereas macrogametocytes and oocysts were clearly devel-opedThere are several explanations possible First nomicro-gamonts developed and no furthermicrogametes developedSecond microgamonts developed but microgametogenesisof the microgametes was disturbed for example by serumIt would be also possible that through the three-dimensionalcharacter of the crypt-like organoid a detection of microg-amonts could not be performed by transmitted light micro-scopy Only the macrogamonts have a specific marker andmicrogamonts are supposed to express only the yfp fluo-rochrome Our observation in small bowel smears showedonly weak fluorescence signals in microgametes (not shown)and no micrographs could be taken through the rapid move-ment of the microgametes Shi et al [24] described the devel-opment of yfp-expressing microgametes of Eimeria tenella invitro but did not show any transmitted light micrographsTheymentioned that the nuclei inmicrogametes in immaturemicrogamonts were not clear We could also see only yfpexpressing stages between TDT-expressing macrogamontswith no distinct nuclear signals (Supplementary Figure 1)whichwere regularly observed in schizont stages (Figures 1(e)and 1(f)) Also the macrogamont typical central nuclear(Figure 1(f)) signal was absent The question still remains ifthese stages are microgamonts
Since we had not observed free microgametes and theoocysts did not sporulate we assume no fertilization of themacrogamete took place As the oocyst walls were formedin vitro the wall formation process seems to be indepen-dent from the fertilization process Ferguson [25] assumedthat even fertilization is not obligatory for sporulation inToxoplasma gondii and other Apicomplexa Our observa-tions do not support this speculation More studies needto be done Crossing experiments with different geneticallymodified parasites could shed light in these processes Inthat manner the genus Eimeria could even acts as a modelfor Toxoplasma gondii because such experiments are moredifficult to accomplish in cats than in rats or chicken Thetransfection technology is already established for Eimeriaspecies infecting these host animals [7 11 26 27] and can givethe opportunity to deepen the knowledge of sexual processesin oocyst-forming Apicomplexa
Disclosure
The authors declare that the experiments comply with thecurrent laws of Germany where they were performed
Conflict of Interests
The authors declare that they have no conflict of interests
Authorsrsquo Contribution
H Chen and M Kurth equally distributed to the experimen-tal work
Acknowledgments
The authors gratefully acknowledge the financial supportprovided by Sachsische Aufbaubank through the EuropeanSocial Fund relating to higher education and research in theFree State of SaxonyThey also gratefully acknowledge AaronPegram and Annie Griffin (Dresden Germany) for paperproofreading andNicoleWeidenbacher (TUDresden) for herhelp in the immunohistology experiments
References
[1] D J Doran ldquoEimeria tenella from sporozoites to oocystsin cell culturerdquo Proceedings of the Helminthological Society ofWashington vol 70 pp 84ndash92 1970
[2] C A Speer and D M Hammond ldquoDevelopment of gameto-cytes and oocysts of Eimeriamagna from rabbits in cell culturerdquoProceedings of the Helminthological Society of Washington vol39 pp 114ndash118 1972
[3] P Bdernik ldquoDevelopment of sexual stages and oocysts from the2nd generation merozoites in tissue culturesrdquo Folia Parasitolog-ica vol 14 p 364 1967
[4] T A Shibalova ldquoCultivation of the endogenous stages ofchicken coccidian in embryos and tissue culturesrdquo Journal ofParasitology vol 56 no 4 pp 315ndash316 1970
[5] C A Speer D M Hammond and G L Kelley ldquoStimulationof motility in merozoites of five Eimeria species by bile saltsrdquoJournal of Parasitology vol 56 no 5 pp 927ndash929 1970
[6] B Rick J-F Dubremetz and R Entzeroth ldquoA merozoite-specific 22-kDa rhoptry protein of the coccidium Eimerianieschulzi (Sporozoa Coccidia) is exocytosed in the para-sitophorous vacuole upon host cell invasionrdquo ParasitologyResearch vol 84 no 4 pp 291ndash296 1998
[7] M Kurth and R Entzeroth ldquoReporter gene expression in cellculture stages and oocysts of Eimeria nieschulzi (CoccidiaApicomplexa)rdquo Parasitology Research vol 104 no 2 pp 303ndash310 2009
[8] R L Roudabush ldquoThe endogenous phases of the life cycles ofEimeria nieschulzi Eimeria separata and Eimeria miyairii coc-cidian parasites of the ratrdquo Iowa State College Journal of Sciencevol 2 pp 135ndash163 1937
[9] W C Marquardt ldquoThe living endogenous stages of the rat coc-cidium Eimeria nieschulzirdquo Journal of Protozoology vol 13 no3 pp 509ndash514 1966
[10] M Tilley and S J Upton ldquoA comparative study of the develop-ment of Eimeria nieschulzi in vitro under aerobic and reducingconditionsrdquo Journal of Parasitology vol 74 no 6 pp 1042ndash10451988
[11] S Hanig R Entzeroth and M Kurth ldquoChimeric fluorescentreporter as a tool for generation of transgenic Eimeria (Api-complexa Coccidia) strains with stage specific reporter geneexpressionrdquo Parasitology International vol 61 no 3 pp 391ndash398 2012
[12] T Lindl Zell- Und Gewebekultur Spektrum Akademischer 4thedition 2000
[13] M Kurth and R Entzeroth ldquoImproved excystation protocolfor Eimeria nieschulzi (Apikomplexa Coccidia)rdquo ParasitologyResearch vol 102 no 4 pp 819ndash822 2008
[14] GGreif andR Entzeroth ldquoEimeria tenella localisation of rhop-try antigens during parasite-host cell interactions by a rhoptry-specific monoclonal antibody in PCKC culturerdquo Applied Para-sitology vol 37 no 4 pp 253ndash259 1996
Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

Journal of Parasitology Research 9
[15] J Krucken R J Hosse A N Mouafo et al ldquoExcystation ofEimeria tenella sporozoites impaired by antibody recognizinggametocyteoocyst antigens GAM22 and GAM56rdquo EukaryoticCell vol 7 no 2 pp 202ndash211 2008
[16] J Baum A T Papenfuss B Baum T P Speed and A F Cow-man ldquoRegulation of apicomplexan actin-basedmotilityrdquoNatureReviews Microbiology vol 4 no 8 pp 621ndash628 2006
[17] F Frischknecht P Baldacci B Martin et al ldquoImaging move-ment of malaria parasites during transmission by Anophelesmosquitoesrdquo Cellular Microbiology vol 6 no 7 pp 687ndash6942004
[18] C Crosnier D Stamataki and J Lewis ldquoOrganizing cell renewalin the intestine stem cells signals and combinatorial controlrdquoNature Reviews Genetics vol 7 no 5 pp 349ndash359 2006
[19] A Shaker and D C Rubin ldquoIntestinal stem cells and epithelial-mesenchymal interactions in the crypt and stem cell nicherdquoTranslational Research vol 156 no 3 pp 180ndash187 2010
[20] T Sato R G Vries H J Snippert et al ldquoSingle Lgr5 stem cellsbuild crypt-villus structures in vitro without a mesenchymalnicherdquo Nature vol 459 no 7244 pp 262ndash265 2009
[21] F M Tomley ldquoCharacterization of rhoptry proteins of Eimeriatenella sporozoites antigenic diversity of rhoptry epitopeswithin species of the genus Eimeria and among three asexualgenerations of a single species E tenellardquo Infection and Immu-nity vol 62 no 10 pp 4656ndash4658 1994
[22] V B Carruthers and F M Tomley ldquoMicroneme proteins inapicomplexansrdquo Sub-Cellular Biochemistry vol 47 pp 33ndash452008
[23] L Lai J Bumstead Y Liu et al ldquoThe role of sialyl glycan recog-nition in host tissue tropism of the avian parasite Eimeria ten-ellardquo PLoS Pathogens vol 7 no 10 Article ID e1002296 2011
[24] T Y Shi X Y Liu L L Hao et al ldquoTransfected Eimeria tenellacould complete its endogenous development in vitrordquo Journal ofParasitology vol 94 no 4 pp 978ndash980 2008
[25] D J P Ferguson ldquoToxoplasma gondii and sex essential oroptional extrardquo Trends in Parasitology vol 18 no 8 pp 355ndash359 2002
[26] J D Clark K Billington J M Bumstead et al ldquoA toolboxfacilitating stable transfection of Eimeria speciesrdquo Molecularand Biochemical Parasitology vol 162 no 1 pp 77ndash86 2008
[27] W Yan X Liu T Shi L Hao F M Tomley and X Suo ldquoStabletransfection of Eimeria tenella constitutive expression of theYFP-YFP molecule throughout the life cyclerdquo InternationalJournal for Parasitology vol 39 no 1 pp 109ndash117 2009
Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013

Submit your manuscripts athttpwwwhindawicom
Enzyme Research
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Advances in
Virolog y
ISRN Biotechnology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioinformaticsAdvances in
Hindawi Publishing Corporationhttpwwwhindawicom
GenomicsInternational Journal of
Volume 2013
ISRN Microbiology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
PeptidesInternational Journal of
ISRN Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Stem CellsInternational
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
ISRN Cell Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
BioMed Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Biochemistry Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
Journal of
Marine Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013Hindawi Publishing Corporation httpwwwhindawicom Volume 2013
The Scientific World Journal
ISRN Molecular Biology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2013
International Journal of
Evolutionary Biology
Hindawi Publishing Corporation httpwwwhindawicom Volume 2013





























