Embed Size (px)
Citation preview

Neurochemistry International 38 (2001) 453–461
Cultured astrocytes react to LPS with increased cyclooxygenaseactivity and phagocytosis
B. Kalmar a, A. Kittel b, R. Lemmens c, Zs. Kornyei b, E. Madarasz b,*a Department of Electrophysiology and Cellular Biology, Gedeon Richter Ltd. Chemical Works, PO Box 27, H-1475 Budapest, Hungary
b Institute of Experimental Medicine, Hungarian Academy of Sciences, Szigony u. 43, H-1083 Budapest, Hungaryc MBW Department, Limburgs Uni6ersitair Centrum, B-3590 Diepenbeek, Belgium
Received 28 March 2000; received in revised form 10 July 2000; accepted 18 July 2000
Abstract
Phagocytosis and prostaglandin E2 production were investigated in purified cultures of perinatal rat forebrain astrocytes. Lightand electron microscopic data indicated that astrocytes respond to bacterial endotoxin, lipopolysaccharide (LPS) by increasedphagocytosis and by activating the cyclooxygenase enzyme-pathway. LPS-inducible phagocytosis of astrocytes was demonstratedby electron microscopic studies on colloidal gold uptake and by photometric determination of fluorescent bead ingestion. Theinternalisation of fragments of the plasma membrane was shown by histochemical detection of membrane-bound ecto-ATPaseactivity within intracellular vesicles. Activation of the cyclooxygenase pathway, a characteristic reaction of immune cells underinflammatory conditions, was also detected in astroglial cells upon treatment with LPS. The increased prostaglandin E2 (PGE2)production by astrocytes in response to LPS was reduced by the non-steroid anti-inflammatory drug, indomethacin. Our dataindicate that astrocytes display some tissue-protective reactions in response to inflammation inducing factors, even in the absenceof peripheral immune cells or central microglia. The role of inducible astrocytic phagocytosis in a non-immune protection-path-way is discussed. © 2001 Elsevier Science Ltd. All rights reserved.
Keywords: Astrocytes; Lipopolysaccharide (LPS); Cyclooxygenase activity; Phagocytosis
www.elsevier.com/locate/neuint
1. Introduction
The extracellular milieu of the central nervous systemhas to be continuously cleaned of decaying cells, celldebris or foreign particles generated not only underpathophysiological conditions but also in the course ofnormal functioning. While the peripheral tissues arepermanently ‘cleaned’ by blood-derived micro- andmacrophages, the tissue-protective functions should beexecuted by astrocytes or microglial cells in the centralnervous system. Although phagocytotic activity is usu-ally attributed to activated microglial cells (Giulian etal., 1989; Rinaman et al., 1991; Smith et al., 1998),some in vivo studies demonstrated that astrocyticphagocytosis is also activated in the traumatised brain(Bechmann and Nitsch, 1997; Tornquist et al., 1997). In
vitro studies on uptake of latex beads have demon-strated that the phagocytotic activity of astrocytes de-pends on the age and physiological stage of the cells(Noske et al., 1982; Watabe et al., 1989). In spite of thelarge proportion of astrocytes in the central nervoustissue, the importance of astroglial phagocytosis hasnot been thoroughly investigated.
Bacterial toxins were shown to elicit several appar-ently pro-inflammatory reactions in astrocytes. In re-sponse to bacterial toxins, the protein kinase C activityincreases in astroglial cells resulting in enhanced NOand prostaglandin production (Ben-Menachem et al.,1998). In response to lipopolysaccharide (LPS) (Wu etal., 1998), to cytokines or interleukins (Nam et al.,1995; Dayton and Mayor, 1996; Busija and Thore,1997; Luo et al., 1998), astrocytes were shown to pro-duce cyclooxygenase(COX)-2 enzyme protein and toincrease the COX enzyme-activity. Major histocompati-bility complex (MHC)-class-II molecules, CD4+ ,CD21, CD24, and also the LPS receptor CD14 (Ennas et
* Corresponding author. Tel.: +36-1-210-0819; fax: +36-1-210-0813.
E-mail address: [email protected] (E. Madarasz).
0197-0186/01/$ - see front matter © 2001 Elsevier Science Ltd. All rights reserved.PII: S 0 1 9 7 -0186 (00 )00090 -5

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461454
al., 1992; Galea et al., 1996; Guillemin et al., 1997) havebeen detected on the surface of stimulated astrocytes.Production of several pro-inflammatory mediators byastrocytes (Kimberlin et al., 1995; Freyer et al. 1996;Glabinski et al., 1996; Kong et al., 1997; Aschner, 1998;Guo et al., 1998) was also demonstrated indicating arole for astrocytes in neuro-inflammatory processes.
While some evidence points to the pro-inflammatoryroles of activated astroglia, data have been accumulat-ing on some inflammation-deregulating functions ofastrocytes as well. Astrocytes might lessen some symp-toms of neural inflammation (Brodie et al., 1998; Haileret al., 1998). Co-culture of macrophages or monocyteswith primary glial cells or with astrocytoma resulted inreduced expression of several inflammatory molecules,such as leukocyte function antigen (LFA-1) and ICAM-1. Treatment of LPS-stimulated macrophages with as-trocyte culture supernatant resulted in an effectivedown-regulation of MHC-II molecules and astrocyte-derived soluble factors could deactivate invadingmacrophages (Hailer et al., 1998). Astrocyte-condi-tioned medium or co-culturing with astrocytes reducedthe IL-12 production by activated microglia (Aloisi etal., 1997).
The aim of our studies was to analyse the responsesof astrocytes to LPS, in the absence of ‘immune’ cells.The inducibility of phagocytosis, the changes in ecto-ATPase and cyclooxygenase enzyme activities were in-vestigated in cultures highly enriched in rat forebrainastrocytes.
2. Materials and methods
2.1. Astroglia cultures
Cultures of astrocytes were obtained from forebrainsof newborn rats as previously described (Madarasz etal., 1991). Briefly, after removal of the meninges andblood vessels, forebrain tissue of newborn (P0–P1) ratswere chopped and incubated in 0.05% trypsin (in phos-phate buffered saline) for 10 min. The enzyme solutionwas aspirated, and the predigested tissue was trituratedin Eagle’s Minimal Essential Medium (MEM) contain-ing 10% FCS. Single cells were separated from non-dis-sociated tissue fragments by sieving through a nylonmesh (pore diameter: 45 mm). Single cells were seededonto 24- or 96-well plates coated with poly-L-lysine atcell densities of 2×105 or 1×104 cells/well,respectively.
Cells were grown in MEM (Sigma), supplementedwith 10% FCS (Gibco), 4 mM glutamine, and 40 mg/mlgentamycin. Fungizone (amphotericin-B, Sigma; 0.5 mg/ml) was added to the medium for the first 2 days afterplating. Cells were grown at 37°C, in humidified atmo-sphere containing 5% CO2 and 95% air. Media werechanged twice a week.
In order to reduce the potential microglial contami-nation, the cultures were split and transferred intosecondary or tertiary cultures. The cell composition ofthe cultures was checked by immunocytochemical stain-ing for GFAP, MHC-II and ED1. Cultures containingmore than 95–98% GFAP-positive cells were involvedin the studies.
2.2. Treatment of the cultures
The media of 15–22-day-old cultures were changed24 h prior to the start of the experiments. LPS (1–100mg/ml), IL-1b (1–10 ng/ml) and occasionally in-domethacin (1 mM) were added to the culture medium,and the cells were grown with the agents for 24 h. Afterthe 24-h incubation, culture media were removed andstored at −70°C. The cells were fixed and processedfor light or electron microscopic investigations.
2.3. Cell 6iability test
Viability was determined according to the method ofMosmann (1983). Cells grown in 24-well plates wereincubated with 0.25 mg/ml 3-(4,5 dimethylthiasole-2-il)-2,5-diphenyltetrasolium (MTT, Sigma) in the culturemedium, for 1.5 h. Cells and reaction products weredissolved in acidic (0.08 M HCl) isopropyl-alcohol.Aliquots of the solutions were transferred to microtiterplates and the optical densities were determined by anSLT-210 ELISA reader at dual wavelengths of 570 and630 nm. Viability assays were carried out on four to sixidentically treated sister cultures, and averages andstandard deviations were calculated. The relative viabil-ity of treated cultures was determined as the percentageof the average of OD (570–630 nm) values measured innon-treated sister cultures.
2.4. Immunocytochemical identification
Cells grown on poly-L-lysine coated glass coverslipswere fixed with 4% paraformaldehyde (w/v in phos-phate buffered saline, PBS) for 20 min, at room temper-ature. The cell membranes were permeabilised with0.1% Triton X-100 in PBS for 5 min and incubated with5% FCS in PBS (60 min) in order to block non-specificantibody binding. Primary antibodies against GFAP(Sigma), MHC-II (OX-42) and ED1 (Serotec) wereapplied at 4°C, overnight in dilutions proposed by themanufacturers. After three washes with FCS-PBS, bi-otin-conjugated second-layer antibodies (Sigma, 1:2000)were applied. After a 1-h incubation the preparationswere washed three times with PBS, and avidin perox-idase (Sigma, 1:1000) was added. The immunoreactivitywas visualised with diaminobenzidine in the presence of0.1% H2O2. For negative controls the primary antibod-ies were omitted.

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461 455
2.5. Measurement of cyclooxygenase acti6ity ofcultured astrocytes
The prostaglandin E2 (PGE2) content of the culturesupernatants was determined by radioimmunoassay(RIA). PGE2 RIA kits were purchased from the Insti-tute of Isotopes, Hungary. The determination wasbased on the competition between the prostaglandinspresent in the sample and the I125 prostaglandin tracerfor the antibodies specific for prostaglandin E2. Theminimum detection limit of the kit was 30 pg/ml. Finalprostaglandin levels were corrected to the prostanoidcontent in the fresh, serum-containing culture medium.
2.6. Measurement of fluorescent bead uptake
Primary astroglial cultures grown on 96-well platesfor 15 or 22 days were treated with LPS (10 mg/ml) orIL-1b (10 ng/ml) for 24 h in the presence or absence of1 mM indomethacin, or were used as non-treated con-trols. For the last 4 h of the treatments, 1.6×1012 latexbeads of 30-nm average diameter and with UV-in-ducible green fluorescence (Sigma; L 2528) were addedto each well. At the end of the incubation, the mediumand the excess beads were removed, the cells werewashed four times with PBS and dissolved in 100 ml 0.1N NaOH per well. Fluorescence was measured at 485-nm excitation and 538-nm emission wavelengths. Thefluorescence intensity of the samples was compared tothe emission values measured in bead preparationscontaining known amounts of beads (1 intensity-unitcorresponds to a bead concentration of 0.68×1010
beads/ml).
2.7. Studies on colloidal gold uptake
Colloidal gold was prepared by adding 4 ml 1%sodium citrate to 100 ml boiling HAuCl4 (0.01% w/v inbidistilled water). When the colour turned deep red, 5ml of the solution was cooled and centrifuged at 15 000rpm, for 10 min. The supernatant was removed and thepellet was re-suspended in 5 ml distilled water. Theprocedure was repeated three times. Finally, the pelletwas resuspended in 1 ml distilled water and the colloidsolution was filtered through a 0.2-mm sterile filter.
A total of 60 ml of gold solution was added tocultures grown on 24-well plates for 15–22 days. Thecultures of each experimental series were either preincu-bated with LPS (10 mg/ml) for 24 h (n=6) or weretreated with LPS and gold simultaneously (n=6).Some of the cultures were not treated with LPS andwere used as controls (n=6). The cells were incubatedwith colloidal gold for 4 h either at 37°C (n=3 for eachtreatment) or at 4°C (n=3 for each treatment).
2.8. Localisation of ecto-ATPase acti6ity by electronmicroscopy
Ecto-ATPase activity was determined as described byKittel (1999). Briefly, half of the culture medium wasremoved and cold fixative (3% paraformaldehyde, 0.5%glutaraldehyde, 2 mM CaCl2, and 0.25 M sucrose in0.05 M cacodylate buffer, pH 7.4.) was added for 2min. The fluid was removed, and fresh ice-cold fixativewas added for 30 min, at 4°C. The samples werewashed three times with cacodylate-sucrose buffer (0.25M sucrose in 0.05 M cacodylate, pH 7.4). The prepara-tions were incubated in a reaction mixture containing 3mM CeCl3, 5 mM MnCl2, 2 mM CaCl2, 1 mMlevamisol, 1 mM ouabain, 50 mM a,b-methylene ADPand 1 mM ATP in 0.07 M Tris-maleate buffer (pH 7.4),at 37°C, for 30 min. After washing with Tris-maleatebuffer, the preparations were postfixed in 1% OsO4 for30 min. After washing with distilled water, the sampleswere dehydrated in graded ethanol, block-stained with2% uranyl acetate in 70% ethanol for 1 h and embeddedin Epon 812. Ultrathin sections were examined in aHitachi 2001 transmission electron microscope.
Some of the fixed preparations were incubated for 45min at room temperature in the above reaction mixturebut containing 5 mM PbCl2 instead of CeCl3 (pH 7.5).After washing with Tris-maleate buffer, the prepara-tions were rinsed in 1% (NH4)2S for 1 min. Afterwashing with distilled water the cultures were eithermounted with Depex for light microscopic studies orwere postfixed with OsO4, dehydrated and embedded asabove for electron microscopic (EM) investigations.
2.9. Morphometry
Morphometric analyses were carried out on ran-domly chosen fields of EM photomicrographs taken at8000× magnification from cytoplasmic areas (n=25–30 for each treatment group). The number of phagocy-tosed Au particles was counted on each field and thearea of the cytoplasm was calculated. The number ofgold particles was corrected to the cytoplasmic area oneach field and presented as NAu.
The area of endosomes and lysosomes (involvingmultilamellar bodies) (ELS) was determined and wasrelated to the area of the cytoplasm (C) on each field(ELS/C). The characteristic proportion of the endo-some/lysosome bound areas (ELS/C) was determinedfor each treatment group by calculating the averagesand standard deviations of ratio values obtained on25–30 fields from the same treatment group.
2.10. Biochemical determination of ecto-ATPaseacti6ity
The amount of the inorganic phosphate produced by

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461456
ecto-ATPases was measured both in fixed and non-fixed cultures incubated with normal assay medium(see below) or in solutions lacking Ca2+ or Mg2+ orboth. The inorganic phosphate, produced during theincubation, was determined according to the methodof Lebel et al. (1978).
Astrocytes grown in 96-well plates were treatedwith LPS for 2 and 24 h or were grown as non-treated controls. Living cells were washed three timeswith TBS and incubated with the reaction buffer (1mM ouabain, 0.1 mg/ml concanavalin-A, 1 mM N-ethylmaleimide, 5 mg/ml oligomycin, 1 mMlevamisole, 100 mM NaCl, 5 mM KCl, 2 mM ATP,30 mM Tris–HCl, pH 7.5) for 2 h, at 37°C. For thedetermination of Ca- and Mg-dependent ATPase ac-tivity, one of the following supplements was added: 2mM Ca2+ and 0.1 mM EDTA; 2 mM Mg2+ and 0.1mM EGTA or 0.5 mM EDTA. The reaction wasstopped by removing the reaction buffer. After wash-ing three times with TBS, some of the cultures werefixed with cold fixative (3% paraformaldehyde, 0.5%glutaraldehyde, 2 mM CaCl2, 0.25 M sucrose in 0.05M cacodylate buffer, pH 7.4) for 30 min. Fixed cul-tures were incubated in reaction buffers as above. Af-ter the incubation, the inorganic phosphateconcentration in the supernatants was monitored byadding 200 ml ‘stop’ solution (0.25% CuSO4 · 5H2O,4.6% Na-acetate · 3H2O, 1% SDS, 5%(NH4)6Mo7O24 · 4H2O, 2% 4-methylamino-phenolsul-phate, 5% Na2SO3, pH 4). After a 15-min incubation,the light absorption was measured at 650 nm. Thephosphate content was determined by using a stan-dard curve obtained by measuring the absorbance ofknown amounts of phosphate.
Two controls have been used for correction. Theautohydrolysis of ATP was determined by measuring
the phosphate content in the reaction mixture incu-bated with ATP but without cells. The amount ofecto-ATPase independent inorganic phosphate produc-tion was evaluated by measuring the phosphate con-tent in the reaction mixture incubated with cells butwithout exogeneous ATP.
3. Results
Monolayer cultures of astrocytes were preparedfrom the forebrain tissue of neonatal rats. On the 2ndweek after plating, the vast majority (95–98%) of thecells showed GFAP immunoreactivity. The proportionof GFAP-negative cells was further reduced by sub-cultivating the cells. In order to check microglial con-tamination, the cultures were stained for themacrophage-microglial marker, ED1. Microglial con-tamination was found to be less than 2% in primarycultures and was decreased below 1% in secondarypreparations. The cultures were treated with increas-ing concentrations of LPS from 1 to 100 mg/ml for 24h and the viabilities were determined by measuringthe reductive capacity of the cultures using the tetra-zolium-reduction assay (Mosmann, 1983). The viabil-ity tests failed to show any toxic effects of LPS onastrocytes (Fig. 1).
3.1. Cyclooxygenase acti6ity of primary astroglial cells
Cyclooxygenase enzyme activity was determined bymeasuring the enzyme product, PGE2 by radioim-munoassay. The results showed that treatment with 10mg/ml LPS or 10 ng/ml IL-1b resulted in a similar,approximately twofold increase in the PGE2 produc-tion by astrocytes. In both cases, the PGE2 produc-tion was reduced in the presence of 1 mMindomethacin (IND), a non-specific cyclooxygenase in-hibitor. The LPS-induced PGE2 production, however,was prevented by IND, while the IL-1b induced PGE2
production was only reduced by the treatment (Fig.2).
3.2. Uptake of fluorescent beads
Assays on fluorescent bead uptake by astrocytes in-dicated that treatment with 10 mg/ml LPS or 10 ng/mlIL-1b resulted in a similar, slight increase in beadingestion (Fig. 3). Transforming the emitted light in-tensities to bead numbers showed that the number ofphagocytosed bead particles increased from 0.8490.65×1010 in control cultures to 3.992.2×1010 incultures treated with LPS. The bead uptake was ap-parently not influenced by the presence of in-domethacine.
Fig. 1. Relative viability of astrocytes was determined by measuringthe reductive capacity of whole cultures (see text). The data obtainedafter treatment with various concentrations of LPS (n=9–12 foreach concentration) were compared to those obtained on non-treatedsister cultures (100%). Averages and standard deviations from threeindependent series of experiments are presented.

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461 457
Fig. 2. Relative PGE2 production by cultured astrocytes in thepresence or absence of 10 mg/ml LPS, 10 ng/ml IL-1b and 1 mMindomethacin (IND). The results are presented as percentages of thePGE2 production by non-treated control cultures (100%=18839443pg/mg protein in 24 h; * PB0.05). Each column represents theaverage and standard deviation of nine to 12 data obtained fromthree independent experiments.
Fig. 5. Morphometric evaluation of electron microscopic data. (A)The area occupied by endo-lysosomes increased significantly in re-sponse to the treatment with 10 mg/ml LPS (24 h). ELS/C ratiostands for the ratio of the area of the endo-lysosomal system com-pared to the total area of the cytoplasm (100%) on each electronmicroscopic frame (* PB0.05). (B) The number of phagocytosed Auparticles was normalised to the area of the cytoplasm (NAu). The dataindicate that the number of internalised gold particles increasedsignificantly as a consequence of LPS treatment. The results areaverages and standard deviations of data obtained from 2×10individual frames from two independent series of experiments (* PB0.05).
Fig. 3. Effects of LPS (10 mg/ml), IL-1b (10 mg/ml) and indomethacin(IND; 1 mM) on the uptake of fluorescent beads by cultured astro-cytes were evaluated by measuring the fluorescence-intensity in thesolubilized cell-material. The averages and standard deviations werecalculated from data (n=9–12) obtained from three independentseries of experiments (1 intensity-unit corresponds to a bead concen-tration of 0.68×1010 beads/ml; * PB0.05 in comparison to thecontrol).
3.3. Electron microscopic studies on the phagocyticacti6ity of cultured astrocytes
In order to further analyse the effect of LPS on thephagocytotic activity of astrocytes, changes in the ex-tent of the endosomal-lysosomal system (ELS) wereinvestigated at the electron microscopic level. Vacuoleswith various shapes were regarded as constituents ofELS. The area occupied by endosomal-lysosomal parti-cles was corrected to the area of the cytoplasm (C)resulting in ELS/C ratio values for each electron micro-scopic frame. The ELS/C values were compared be-tween the control and LPS-treated cultures. In bothcontrol and LPS-treated astrocytes, well-developedendo-lysosomal systems were found. A 24-h treatmentwith LPS (10 mg/ml) resulted in an increase in the sizeand frequency of the vacuoles (Fig. 4). The ELS/Cvalue increased significantly as a consequence of treat-
Fig. 4. Electron microscopic images of cells in control (A) and LPS-treated (B) cultures after a 4-h incubation with colloidal gold. LPS causedremarkable changes in the morphology and a significant increase in the size of the endo-lysosomes. Scale bar=0.9 mm.
B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461458
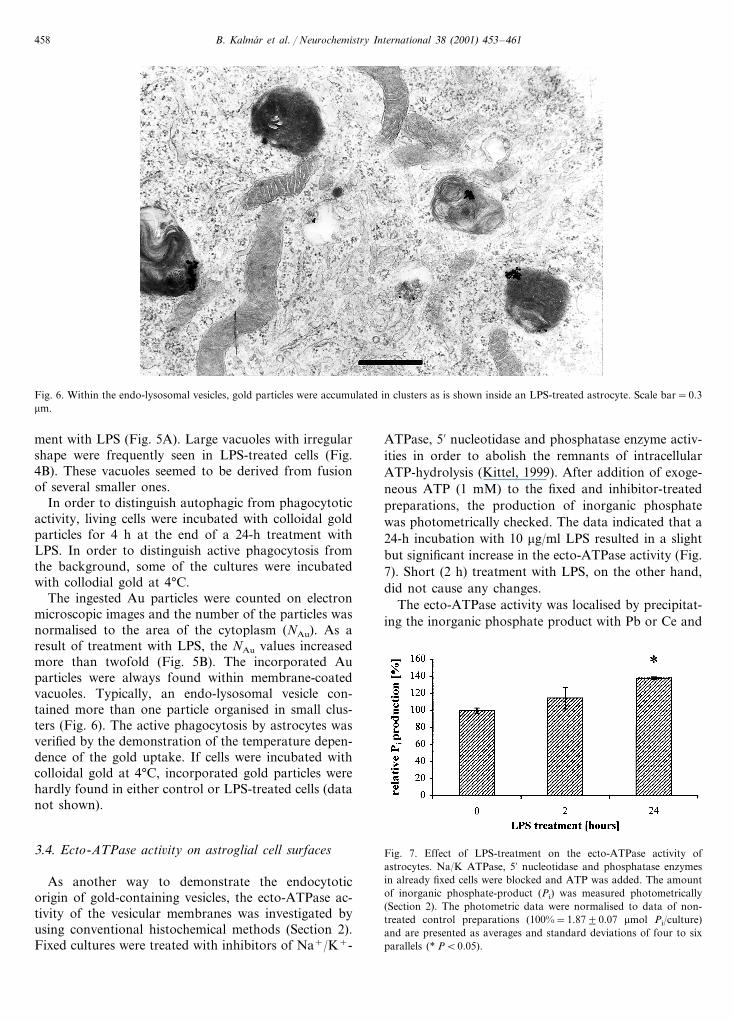
Fig. 6. Within the endo-lysosomal vesicles, gold particles were accumulated in clusters as is shown inside an LPS-treated astrocyte. Scale bar=0.3mm.
ment with LPS (Fig. 5A). Large vacuoles with irregularshape were frequently seen in LPS-treated cells (Fig.4B). These vacuoles seemed to be derived from fusionof several smaller ones.
In order to distinguish autophagic from phagocytoticactivity, living cells were incubated with colloidal goldparticles for 4 h at the end of a 24-h treatment withLPS. In order to distinguish active phagocytosis fromthe background, some of the cultures were incubatedwith collodial gold at 4°C.
The ingested Au particles were counted on electronmicroscopic images and the number of the particles wasnormalised to the area of the cytoplasm (NAu). As aresult of treatment with LPS, the NAu values increasedmore than twofold (Fig. 5B). The incorporated Auparticles were always found within membrane-coatedvacuoles. Typically, an endo-lysosomal vesicle con-tained more than one particle organised in small clus-ters (Fig. 6). The active phagocytosis by astrocytes wasverified by the demonstration of the temperature depen-dence of the gold uptake. If cells were incubated withcolloidal gold at 4°C, incorporated gold particles werehardly found in either control or LPS-treated cells (datanot shown).
3.4. Ecto-ATPase acti6ity on astroglial cell surfaces
As another way to demonstrate the endocytoticorigin of gold-containing vesicles, the ecto-ATPase ac-tivity of the vesicular membranes was investigated byusing conventional histochemical methods (Section 2).Fixed cultures were treated with inhibitors of Na+/K+-
ATPase, 5% nucleotidase and phosphatase enzyme activ-ities in order to abolish the remnants of intracellularATP-hydrolysis (Kittel, 1999). After addition of exoge-neous ATP (1 mM) to the fixed and inhibitor-treatedpreparations, the production of inorganic phosphatewas photometrically checked. The data indicated that a24-h incubation with 10 mg/ml LPS resulted in a slightbut significant increase in the ecto-ATPase activity (Fig.7). Short (2 h) treatment with LPS, on the other hand,did not cause any changes.
The ecto-ATPase activity was localised by precipitat-ing the inorganic phosphate product with Pb or Ce and
Fig. 7. Effect of LPS-treatment on the ecto-ATPase activity ofastrocytes. Na/K ATPase, 5% nucleotidase and phosphatase enzymesin already fixed cells were blocked and ATP was added. The amountof inorganic phosphate-product (Pi) was measured photometrically(Section 2). The photometric data were normalised to data of non-treated control preparations (100%=1.8790.07 mmol Pi/culture)and are presented as averages and standard deviations of four to sixparallels (* PB0.05).

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461 459
Fig. 8. Electron microscopic localisation of ecto-ATPase enzymeactivity in control (A) and in LPS-treated cells (B) using Ce precipita-tion method. Ce precipitates are indicated by arrows on the cellsurfaces and by arrowheads within the intracellular vacuoles. Scalebars: 0.6 mm in A and 0.3 mm in B.
Under physiological conditions, the astroglial phagocy-tosis does not induce inflammatory reactions. Inflam-mation and reactive gliosis can be induced and/oramplified by microglia, the resident macrophage-typecell of the CNS. Microglia is known to produce several‘pro-inflammatory’ cytokines and interleukins, such asIL-1b, IFNg, TNFa, (Gonzalez-Scarano and Baltuch,1999) and NO (Matsuoka et al., 1999) in response tovarious tissue damage or bacterial toxins. Data accu-mulated in the last decade (Nam et al., 1995; Daytonand Mayor, 1996; Busija and Thore, 1997; Luo et al.,1998), however, indicate that astrocytes also respond toseveral damaging stimuli by producing both inflamma-tion promoting and inhibiting compounds.
Our results showed that cultured astroglial cells in-creased their PGE2 production, ecto-ATPase activityand phagocytotic activity in response to a bacterialendotoxin, LPS. Purified cultures highly enriched inastrocytes were used, but the contribution of contami-nating microglia (less than 1% of total cells) to theinitiation of the astrocytic reactions could not be prop-erly ruled out. The reactions, however, were notchanged if the microglial contamination was reduced bysubcultivation. In vivo, the main inducer of the as-troglial ‘activation’ is IL-1b, a pro-inflammatory inter-leukin which is produced mainly by microglia in theCNS. Microglial cells respond to LPS with increasedIL-1b production. In our astroglial cultures, the PGE2
production increased similarly in response to LPS andIL-1b, but displayed different sensitivity to the in-hibitory effect of indomethacin. The finding indicatedthat the arachidonic acid cascade was activated ondistinct routes by LPS and by IL-1b. The distinctresponse to LPS suggested that at least a part of thereactions was not initiated by microglia-derived IL-1b.Specific LPS receptor(s), such as CD14 demonstratedrecently on astroglial cells (Galea et al., 1996), mayserve as molecular basis for the immediate astrocyticresponses to LPS.
In response to treatment with LPS, the volume of theendosome/lysosome-like structures and the number ofingested gold particles increased. The internalised parti-cles were always enclosed in membranous vacuoles. Thedata indicated that long-term LPS treatment resulted inan activation of endocytotic processes. The enlargedsize and the irregular shape of the vacuoles, on theother hand, suggested that they were derived from thefusion of several smaller vesicles. In order to distinguishbetween phagocytotic and autophagic vacuoles, the in-ternalization of the plasma membrane was investigatedby localizing the activity of ecto-ATPases, a group ofouter surface-bound enzymes. The translocation of theecto-ATPase activity from the cell surfaces to someintracellular vesicular compartments was previouslyshown in stimulated human neutrophils (Kobayashi et
detecting the precipitate by standard electron micro-scopic methods (Kittel, 1999). In the absence of ATP,precipitate was not formed. In the presence of exoge-neous ATP, the precipitated phosphate was evenly dis-tributed throughout the cell surfaces (Fig. 8A). Incontrol cells, intracellular precipitate was only sporadi-cally detected. In LPS-treated cells, on the other hand,the membranes of several vacuoles displayed heavyecto-ATPase activity (Fig. 8B), suggesting that theirmembranes derived from the internalisation of the sur-face membrane.
4. Discussion
Phagocytotic activity is one of the physiological func-tions of astroglial cells. During development or in thecourse of tissue rearrangements, astrocytes contributeto the removal of cell debris derived from decaying cellsor withdrawn neuronal processes (Dalmau et al., 1998).

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461460
al., 1997). In non-treated control cultures of astrocytes,the ecto-ATPase activity was evenly distributed overthe cell surfaces and was very rarely seen inside thecells. If the cultures were exposed to LPS, the mem-branes of several intracellular vacuoles displayedmarked ecto-ATPase activity. Because of the lack ofsimultaneous visualization of gold particles and ecto-ATPase activity, the origin of the enzyme-containingvacuoles was not clearly defined. They might be trans-port vesicles carrying freshly produced ecto-enzymes tothe surface membrane. Alternatively, they might beendocytotic vesicles importing material from the cellsurface. The relatively large size and the irregular shapeof the ecto-ATPase-positive vacuoles, however, arguedfor their endocytotic origin. For all cases, the enhancedintracellular level of ecto-enzyme activity indicated thatastrocytes responded to LPS with some acceleration ofthe membrane metabolism.
Biochemical evaluation of the inorganic phosphateproduction showed that ecto-ATPase activity was up-regulated in response to LPS. In the CNS, both neu-rons and astrocytes express ecto-ATPases on theirsurfaces (Kegel et al., 1997; Zimmermann et al., 1998).The role of these ecto-enzymes has not been properlyunderstood. In the CNS, there are several receptorsbinding adenosine or adenosine-tri-, di- or mono-phos-phates (Buell et al., 1996). The ecto-ATPases serve bothas neurotransmitter producing and degrading ectoen-zymes (Juranyi et al., 1997). In damaged neural tissue,a marked upregulation of the ecto-ATPase activity wasdemonstrated (Braun et al., 1998). The significance ofpurinergic signalling in the neuro-inflammatory pro-cesses or in other self-protective reactions of the neuraltissue needs further clarification. In endothelial cells,adenosine, the end product of the combined action ofecto-ATPase and 5% nucleotidase enzymes, down-regu-lates the vascular inflammation in conjunction with theproduction of prostaglandins and nitric oxide (NO)(Robson et al., 1997). Adenosine was also shown toreduce the TNFa production by murine macrophagesboth in vitro and in endotoxemic mice (Hasko et al.,1996).
In case of astrocytes, further in vitro studies shoulddecide on the correlation between the upregulation ofthe ecto-ATPase activity and the production of inflam-matory mediators. The increased PGE2 level, however,has been shown to result in a reduced TNFa produc-tion (Aggarwal and Natarajan, 1996). The results ofour experiments suggest that LPS-induced astrocytesproduce TNFa level reducing compounds and ingestforeign particles from their environment. Functioningin that way, early astroglial reactions to damagingstimuli might postpone the onset of the activation ofthe immunocompetent cells and prevent the brain fromnon-necessary overreactions.
References
Aggarwal, B.B., Natarajan, K., 1996. Tumor necrosis factors: devel-opments during the last decade. Eur. Cytokine Netw. 7 (2),93–124.
Aloisi, F., Penna, G., Cerase, J., Menendez-Iglesias, B., Adorini, L.,1997. IL-12 production by central nervous system microglia isinhibited by astrocytes. J. Immunol. 159 (4), 1604–1612.
Aschner, M., 1998. Astrocytes as mediators of immune and inflam-matory responses in the CNS. Neurotoxicology 19 (2), 269–281.
Bechmann, I., Nitsch, R., 1997. Identification of phagocytic glial cellsafter lesion induced anterograde degeneration using double-fluorescence labelling: combination of axonal tracing and lectin orimmunostaining. Histochem. Cell Biol. 107, 391–397.
Ben-Menachem, G., Rottem, S., Tarshis, M., Barash, V., Brenner, T.,1998. Mycoplasma fermentans glycolipid triggers inflammatoryresponse in rat astrocytes. Brain Res. 803, 34–38.
Braun, N., Zhu, Y., Krieglstein, J., Culmsee, C., Zimmermann, H.,1998. Upregulation of the enzyme chain hydrolysing extracellularATP after transient forebrain ischemia in the rat. J. Neurosci. 18(13), 4891–4900.
Brodie, C., Goldreich, N., Haiman, T., Kazimirsky, G., 1998. Func-tional IL-4 receptors on mouse astrocytes: IL-4 inhibits astrocyteactivation and induces NGF secretion. J. Neuroimmunol. 81(1–2), 20–30.
Buell, G., Collo, G., Rassendren, F., 1996. P2X receptors: an emerg-ing channel family. Eur. J. Neurosci. 8, 2221–2228.
Busija, D.W., Thore, C., 1997. Modulation of prostaglandin produc-tion by nitric oxide in astroglia. Prostaglandins LeucotrienesEssent. Fatty Acids 56 (5), 355–359.
Dalmau, I., Finsen, B., Zimmer, J., Gonzalez, B., Castellano, B.,1998. Development of microglia in the postnatal rat hippocam-pus. Hippocampus 8 (5), 458–474.
Dayton, E.D., Mayor, E.O., 1996. Recombinant human interleukin1b induces production of prostaglandins in primary human fetalastrocytes and immortalised human fetal astrocyte cultures. J.Neuroimmunol. 71, 11–18.
Ennas, M.G., Cocchia, D., Silvetti, E., Sogos, V., Riva, A., Torelli,S., Gremo, F., 1992. Immunocompetent cell markers in humanfetal astrocytes and neurons in culture. J. Neurosci. Res. 32,424–436.
Freyer, D., Weih, M., Weber, J.R., Burger, W., Scholz, P., Manz, R.,Ziegenhorn, A., Angestwurm, K., Dirnagl, U., 1996. Pneumococ-cal cell wall components induce nitric oxide synthase and TNF-al-pha in astroglial-enriched cultures. Glia 16 (1), 1–6.
Galea, E., Reis, D.J., Fox, E.S., Xu, H., Feinstein, D.L., 1996. CD14mediate endotoxin induction of nitric oxide synthase in culturedbrain glial cells. J. Neuroimmunol. 64 (1), 19–28.
Giulian, D., Chen, J., Ingeman, J.E., George, J.K., Noponen, M.,1989. The role of mononuclear phagocytes in wound healing aftertraumatic injury to adult mammalian brain. J. Neurosci. 9 (12),4416–4429.
Glabinski, A.R., Balasingam, V., Tani, M., Kunkel, S.L., Strieter,R.M., Yong, V.W., Ransohoff, R.M., 1996. Chemokine monocytechemoattractant protein-1 is expressed by astrocytes after me-chanical injury to the brain. J. Immunol. 156 (11), 4363–4368.
Gonzalez-Scarano, F., Baltuch, G., 1999. Microglia as mediators ofinflammatory and degenerative diseases. Annu. Rev. Neurosci. 22,219–240.
Guillemin, G., Boussin, F.D., Croitoru, J., Franck-Duchenne, M.,Le-Grand, R., Lazarini, F., Dormont, D., 1997. Obtention andcharacterisation of primary astrocyte and microglial cultures fromadult monkey brains. J. Neurosci. Res. 49 (5), 576–591.
Guo, H., Jin, Y.X., Ishikawa, M., Huang, Y.M., van-der-Meide,P.H., Link, H., Xiao, B.G., 1998. Regulation of beta-chemokinemRNA expression in adult rat astrocytes by lipopolysaccharide,

B. Kalmar et al. / Neurochemistry International 38 (2001) 453–461 461
proinflammatory and immunoregulatory cytokines. Scand. J. Im-munol. 48 (5), 502–508.
Hailer, N.P., Heppner, F.L., Haas, D., Nitsch, R., 1998. Astrocyticfactors deactivate antigen presenting cells that invade the centralnervous system. Brain Pathol. 8 (3), 459–474.
Hasko, G., Szabo, C., Nemeth, Z.H., Kvetan, V., Pastores, S.M.,Vizi, E.S., 1996. Adenosine receptor agonists differentially regu-late IL-10, TNF-alpha, and nitric oxide production in RAW 264.7macrophages and in endotoxemic mice. J. Immunol. 157 (10),4634–4640.
Juranyi, Z., Orso, E., Janossy, A., Szalay, K.S., Sperlagh, B.,Windisch, K., Vinson, G.P., Vizi, E.S., 1997. ATP and[3H]noradrenaline release and the presence of ecto-Ca(2+ )-AT-Pases in the capsule-glomerulosa fraction of the rat adrenal gland.J. Endocrinol. 153 (1), 105–114.
Kegel, B., Braun, N., Heine, P., Maliszewski, C.R., Zimmermann, H.,1997. An ecto-ATPase and an ecto-ATP diphosphohydrolase areexpressed in rat brain. Neuropharmacology 36, 1189–1200.
Kimberlin, D.W., Velasco, S., Paris, M.M., Hickey, S.M., Mc-Cracken, G.H., Jr, Nisen, P.D., 1995. Modulation of expressionof genes involved in the inflammatory response by lipopolysaccha-ride and temperature in cultured human astroglial cells. Immunol.Invest. 24 (5), 775–785.
Kittel, A., 1999. Lipopolysaccharide treatment modifies pH- andcation-dependent ecto-ATPase activity of endothelial cells. J. His-tochem. Cytochem. 47 (3), 393–399.
Kobayashi, T., Okada, T., del Saz, E.G., Seguchi, H., 1997. Internal-isation of ecto-ATPase activity in human neutrophils upon stimu-lation with phorbol ester and formyl peptide. Histochem. CellBiol. 107, 353–363.
Kong, L.Y., Lai, C., Wilson, B.C., Simpson, J.N., Hong, J.S., 1997.Protein tyrosine kinase inhibitors decrease lipopolysaccharide-in-duced proinflammatory cytokine production in mixed glia, mi-croglia-enriched or astrocyte-enriched cultures. Neurochem. Int.30 (4–5), 491–497.
Lebel, D., Poirier, G.G., Beaudoin, A.R., 1978. A convenient methodfor the ATPase assay. Anal. Biochem. 85, 86–89.
Luo, J., Lang, J.A., Miller, M.W., 1998. Transforming growth factorb regulates the expression of cyclooxygenase in cultured corticalastrocytes and neurons. J. Neurochem. 71, 526–534.
Madarasz, E., Theodosis, D.T., Poulain, D.A., 1991. In vitro forma-tion of type 2 astrocytes derived from postnatal rat hypothalamusor cerebral cortex. Neuroscience 43, 211–221.
Matsuoka, Y., Kitamura, Y., Takahashi, H., Tooyama, I., Kimura,H., Gebicke-Haerter, P.J., Nomura, Y., Taniguchi, T., 1999.Interferon-gamma plus lipopolysaccharide induction of delayedneuronal apoptosis in rat hippocampus. Neurochem. Int. 34,91–99.
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxic assays. J.Immunol. Methods 65, 55–63.
Nam, M.J., Thore, C., Busija, D.W., 1995. Rapid induction ofprostaglandin synthesis in piglet astroglial cells by interleukin 1a.Brain Res. Bull. 36 (3), 215–218.
Noske, W., Lentzen, H., Lange, K., Keller, K., 1982. Phagocytoticactivity of glial cells in culture. Exp. Cell Res. 142, 437–445.
Rinaman, L., Milligan, C.E., Levitt, P., 1991. Persistence of fluoro-gold following degeneration of labelled motoneurons is due tophagocytosis by microglia and macrophages. Neuroscience 44 (3),765–776.
Robson, S.C., Kaczmarek, E., Siegel, J.B., Candinas, D., Koziak, K.,Millan, M., Hancock, W.W., Bach, F.H., 1997. Loss of ATPdiphosphohydrolase activity with endothelial cell activation. J.Exp. Med. 185, 153–163.
Smith, M.E., van der Maesen, K., Somera, F.P., 1998. Macrophageand microglial responses to cytokines in vitro: phagocytic activity,proteolytic enzyme release and free radical production. J. Neu-rosci. Res. 54, 68–78.
Tornquist, E., Liu, L., Mattsson, P., Svensson, M., 1997. Response ofglial cells and activation of complement following motoneurondegeneration induced by toxic ricin. Neurosci. Res. 28, 167–175.
Watabe, K., Osborne, D., Kim, S.U., 1989. Phagocytic activity ofhuman adult astrocytes and oligodendrocytes in culture. J. Neu-ropathol. Exp. Neurol. 48, 499–506.
Wu, V.W., Nishiyama, N., Schwartz, J.P., 1998. A culture model ofreactive astrocytes: increased nerve growth factor synthesis andreexpression of cytokine responsiveness. J. Neurochem. 71, 749–756.
Zimmermann, H., Braun, N., Kegel, B., Heine, P., 1998. New insightsinto molecular structure and function of ectonucleotidases in thenervous system. Neurochem. Int. 32, 421–425.
.





























