Embed Size (px)
Citation preview

Volume **, ** ** TRANSFUSION 1
Blackwell Publishing IncMalden, USATRFTransfusion0041-11322006 American Association of Blood Banks** ****8••••Original ArticlePLT-MACROPHAGE INTERACTIONBADLOU ET AL.
ABBREVIATIONS: BAPTA = 1,2-bis-(2-aminophenoxy)ethane-
NNN′N′-tetraacetic acid; GlcNAc = N-acetylglucosamine;
GP = glycoprotein; MSP(s) = metabolic suppressed platelet(s);
PS = phosphatidylserine; PGI2 = prostacyclin; PMA = phorbol 12-
myristate 13-acetate; PSGL-1 = P-selectin glycoprotein ligand 1.
From the Thrombosis and Hemostasis Laboratory, Department
of Hematology, University Medical Center Utrecht, the Sanquin
Blood Bank Region North-West, Amsterdam; and The Institute for
Biomembranes, Utrecht University, Utrecht, the Netherlands.
Address reprint requests to: J.W.N. Akkerman, PhD,
Thrombosis and Haemostasis Laboratory, Department of
Haematology, UMCU, PO Box 85500, 3508 GA, Utrecht, the
Netherlands; e-mail: [email protected].
Supported by the Sanquin Blood Supply Foundation (Grant
A.02.002) and the Netherlands Thrombosis Foundation (JWNA).
Received for publication October 6, 2005; revision received
December 30, 2005, and accepted December 31, 2005.
doi: 10.1111/j.1537-2995.2006.00913.x
TRANSFUSION **;**:**-**.
H E M O S T A S I S
Platelet binding and phagocytosis by macrophages
Bahram A. Badlou, Ya Ping Wu, W. Martin Smid, and Jan-Willem N. Akkerman
BACKGROUND: Earlier it was reported that metabolic arrest followed by incubation at 4°C reduces the platelet (PLT) storage defect. Here it is reported that this treatment also reduces binding and phagocytosis by macrophages.STUDY DESIGN AND METHODS: Phagocytosis of mepacrine-labeled PLTs by macrophages changes the latter into bright fluorescent particles easily detected by fluorescence-activated cell sorting.RESULTS: In combination with conventional binding analysis it was found that binding to phorbol 12-myristate 13-acetate–matured THP-1 cells is primarily regulated by PLT P-selectin expression and phagocytosis by combined phosphatidylserine (PS) exposure and glycoprotein (GP) Ibα clustering. It was found that trapping of PLT Ca2+ and raising cAMP reduces phagocytosis by lowering PS exposure. Chilling of PLTs leads to an increase in binding and PS- and GPIbα-mediated phagocytosis. Prior depletion of PLT energy stores prevents this increase by preserving low Ca2+ concentration, PS exposure, and PS-mediated phagocytosis.CONCLUSION: These data characterize the individual factors that control PLT binding and phagocytosis and might help to define conditions that improve the survival of stored PLTs after transfusion.
he storage of platelet (PLT) concentrates at roomtemperature facilitates bacterial growth andintroduces changes in PLTs indicative for activa-tion and initiation of apoptosis. Improvements
have been sought in lowering the storage temperature butthis treatment severely reduces the survival of transfusedPLTs. A main feature of PLT storage is the gradual increasein surface expression of P-selectin (CD62P), a componentof α-granule membranes. The counter receptor for P-selectin, P-selectin glycoprotein (GP) ligand 1 (PSGL-1), ispresent on white blood cells (WBCs) and the inverse cor-relation between P-selectin expression and PLT recoveryafter transfusion is sought to reflect PLT destructionthrough coupling of these receptors. The contribution ofP-selectin expression to PLT destruction, however, canonly be small because transfused human PLTs lose P-selectin in the circulation1 and P-selectin–deficient micePLTs survive normally.2,3 A second feature of PLT storage isthe membrane appearance of negatively charged phos-pholipids such as phosphatidylserine (PS), which is aproperty of activated PLTs4,5 and cells going into apopto-sis.3,6 PS exposure may form recognition sites for destruc-tion of senescent PLTs, and a similar mechanism mightremove stored PLTs from the circulation.6
T
trf_00913.fm Page 1 Wednesday, June 14, 2006 4:44 PM

BADLOU ET AL.
2 TRANSFUSION Volume **, ** **
Attempts to reduce the surface expression of recogni-tion sites for PLT destruction were based on prostacyclin,which inhibits secretion and PS exposure;7 arrest of glyc-olytic and oxidative energy generation, which inhibits ATPresynthesis required for PLT functions;8,9 and storage atlow temperature, which slows down PLT metabolism andin addition suppresses bacterial growth. Chilling of PLTshas long been thought to cause irreversible cell damagereflected by loss of the discoid shape and an increasein cytosolic Ca2+ and causing poor posttransfusion sur-vival.10,11 Treatments that preserve PLT shape and a lowCa2+ level, however, failed to improve in vivo survival.12,13
Recently, Hoffmeister and colleagues14 showed that thechilling of PLTs triggers the rearrangement of GPIbα(CD42b) into clusters that are recognized by αMβ2 (CR3/Mac-1) receptors on liver macrophages triggering PLTdestruction. A resting PLT contains approximately 20,000GPIbα molecules bound to GPV and GPIX in a 2:1:2stoichiometry randomly distributed over the plasmamembrane.11,15,16 GPIbα serves in the binding of PLTs toactivated von Willebrand factor at sites of vascular damageslowing down their velocity in flowing blood and enablingother receptors to firmly attach PLTs to the wound.16-19
Interestingly, cold-induced formation of GPIbα clustersleaves the hemostatic functions of GPIbα undisturbed.20
An alternative means to prolong the storage of PLTs isby metabolic suppression before cold storage. We showedearlier that PLTs sustain a period of metabolic arrest with-out losing their adhesive properties and their capacity toaggregate and secrete granule contents.21 During meta-bolic arrest, PLTs fail to respond to PLT-activating agentssuch in agreement with the energy requirement of aggre-gation and secretion.
In this study, we investigated the mechanisms thatmediate binding of PLTs to macrophages and initiate theirdestruction. The results reveal major roles for surface P-selectin in binding and exposed PS and clustered GPIbαin phagocytosis. We also show that chilling of PLTs inducesbinding and phagocytosis and that prior metabolic arrestprotects PLTs against these cold-induced changes. Thisprotective effect can be explained by suppression of PSexposure providing a means to interfere with surfaceexpression of phagocytotic signals through metabolicintervention.
MATERIALS AND METHODS
Materials
We obtained antimycin A, phorbol 12-myristate 13-acetate (PMA), cytochalasin B, 1,2-bis-(2-aminophe-noxy)ethane-NNN′N′-tetraacetic acid (BAPTA)-AM,N-acetylglucosamine (GlcNAc), mepacrine (Quinacrine),and Fura-2 AM from Sigma Chemical Co. (Mannheim,Germany); the anti-human CD42b (GPIbα-PE [R7014]),anti-human CD14-fluorescein isothiocyanate (FITC) anti-
body, annexin V-PE, annexin V-FITC, anti-human CD62P(P-selectin), and FITC-labeled immunoglobulin G as neg-ative control from Dako A/S (Glusdorp, Denmark); serum-free cell culture medium RPMI 1640 from Corning Inc.(Corning, NY); fetal calf serum (FCS) from Cambrex (Viers,Belgium); penicillin, streptomycin sulfate, and trypsinfrom Gibco Invitrogen Corp. (Grand Island, NY); prostacy-clin (PGI2) from Cayman Chemical Co. (Ann Arbor, MI);and the stable prostacyclin analog iloprost from ScheringA.G. (Berlin, Germany). Annexin V was a gift from W.L. vanHeerde, Department of Hematology, Radboud UniversityNijmegen.
PLT isolation and storage
Freshly drawn venous blood from healthy, medication-free volunteers (40 mL) was collected with informed con-sent into 1:10 (vol/vol) 130 mmol per L trisodium citrate.PLT-rich plasma was prepared by centrifugation (200 × g,15 min, 20°C). ACD (0.1 volume of 2.5 g trisodium citrate,1.5 g citric acid, and 2.0 g D-glucose in 100 mL of distilledwater) was added to lower the pH value to 6.0 and preventPLT activation during isolation. The suspension (10 mL)was washed by centrifugation (330 × g, 15 min, 22°C) andresuspended in glucose-free Hepes-Tyrode (137 mmol/LNaCl, 2.68 mmol/L KCl, 0.42 mmol/L NaH2PO4, 1.7 mmol/L MgCl2, and 11.9 mmol/L NaHCO3, pH 7.2) to a final con-centration of 2 × 108 PLTs per mL. PLT count was measuredon a cell counter (Model AL871, Molab, Hilden, Germany).PLTs in glucose (5 mmol/L)-containing Hepes-Tyrodewere stored at 22°C, a storage temperature currently usedin blood banks, and at 0°C, a condition known to induceclustering of GPIbα, binding to macrophage αMβ2, and PLTdestruction after transfusion.22 Metabolic suppressed PLTs(MSPs) were prepared by incubation in glucose-freeHepes-Tyrode solution containing 20 μmol per L antimy-cin A for 40 minutes at 37°C to deplete energy stores.23
MSPs were stored at 4°C for the indicated periods. Suspen-sions were stored in closed tubes impermeable for gasexchange without agitation. Before each measurement,MSPs and PLTs stored at 22°C were incubated with20 mmol per L glucose for 1 hour at 37°C to optimallyrestore energy generation; the PLTs stored at 0°C werewarmed for 15 minutes at 37°C.23
Preparation of mepacrine-labeled PLTs and PMA-matured THP-1 cells
Immediately before the binding and phagocytosis experi-ments, 100-μL PLT suspensions were labeled with 1 μmolper L mepacrine in Hepes-Tyrode (pH 7.2, 5 min, 22°C).Mepacrine is a fluorescent polyphenolic compound thatemits at 519 nm, which is in the range of the emission ofFITC (530 ± 15 nm). Free mepacrine was removed (5 min,350 × g, 22°C, with soft mode) while preventing PLT acti-
trf_00913.fm Page 2 Wednesday, June 14, 2006 4:44 PM

PLT-MACROPHAGE INTERACTION
Volume **, ** ** TRANSFUSION 3
vation with 10 ng per mL PGI2. Monocytoid THP-1 celllines were cultured to a density of 2 × 105-4 × 105 cells permL in RPMI 1640 containing 10 percent FCS, 2 mmol perL glutamine, penicillin (0.1 U/L), and streptomycin (1 μg/L) at 37°C. THP-1 cells were counted in a Burker-Turkchamber and a 1-mL suspension containing 1 × 106 cellswas added to a well of a 48-well plate (Corning Inc., Corn-ing, NY). Maturation was induced by incubation with500 nmol per L PMA for 24 hours at 37°C unless statedotherwise. In a few experiments free PMA was removedfrom the THP-1 cells before incubation with PLTs to pre-vent PLT activation, as indicated.
PLT-macrophage interaction
The binding of PLTs to macrophages was a modificationof the procedure described by Hoffmeister and cowork-ers.14 In short, 2 × 106 PLTs in Hepes-Tyrode buffer (pH 7.2)were added to a well containing 1 × 106 PMA-maturedTHP-1 cells in 1 mL of RPMI 1640, and the mixed suspen-sions were incubated at 37°C without agitation. Wells weregently washed with HBSS buffer (0.3 mmol/L KH2PO4,13.7 mmol/L NaCl, 417 mmol/L NaHCO3, 31 mmol/LNa2HPO4, and 0.5 mmol/L KCl in aqua dest) and free PLTscollected in the wash medium were isolated by centrifu-gation-resuspension under protection of PGI2. Wells wereincubated with 200 mL HBSS buffer containing 5 mmol/Lethylenediaminetetraacetate for 15 minutes at 0°C, recon-stituted with free PLTs removed during the wash step, andthereafter a 100-μL suspension was incubated with 2 μgper mL anti-human CD42b-PE and 2 μg per mL anti-human CD14-FITC antibody for 15 minutes at 37°C. HBSSbuffer was then added and 20,000 particles were mea-sured by flow cytometry (FACSCalibur, Becton-Dickinson,San Jose, CA). Fluorescence-activated cell sorting (FACS)data were analyzed with computer software (WinMDI,The Scripps Research Institute, La Jolla, CA). Binding ofPLTs to macrophages was expressed as the percentageof CD42b/CD14-positive particles of the total number ofCD42b- and/or CD14-positive particles. Phagocytosis ofPLTs by PMA-matured THP-1 cells was measured by FACSanalysis of mepacrine-positive CD14 cells that wereinaccessible to the CD42b-PE antibody and expressed aspercentage of total number of CD14-positive, CD42b-negative particles.
In a few experiments THP-1 cells (1 × 106 cells) wereallowed to attach to glass, mildly matured by incubationwith 100 nmol per L PMA (15 min, 22°C), and incubatedat 22°C with PLTs stored for 48 hours at 22°C. Binding andphagocytosis were measured by real-time analysis with afluorescence microscope (Orthoplan, Leica, Heidelberg,Germany) with a 100× objective connected to a charge-coupled device (CCD) camera system (Jai, Copenhagen,Denmark) coupled to a personal computer with software(Optimas 6.0 DVS, Breda, the Netherlands).
Interference with binding and phagocytosis
PLTs stored at the indicated conditions were incubatedwith P-selectin antibody (17 μg/mL), annexin V (17 μg/mL), cytochalasin B (10 μmol/L), BAPTA-AM (10 μmol/L),and iloprost (10 μmol/L) for 1 hour at 22°C. Additionswere removed by a wash step before mixing with THP-1cells and analysis of binding and phagocytosis. The role ofGPIbα clusters was evaluated by measuring phagocytosisof the cold PLTs in the presence of GlcNAc (100 mmol/L).
Measurement of P-selectin and PS expression
PLTs were incubated with glucose, cytochalasin B, BAPTA-AM, or iloprost for 1 hour at 37°C and P-selectin and PSexpression were measured on a FACScalibur.23,24 Sampleswere fixed with 2 percent paraformaldehyde (30 min,22°C) and washed with 500 μL of phosphate-bufferedsaline (PBS; 500 × g, 5 min, 22°C). Pellets were resus-pended in 50 mL PBS containing 1 percent BSA and0.01 percent Tween 20 (5 min, 22°C) and PLT suspensionswere incubated with annexin V-FITC and CD62P-PE (1 μg/mL each) for 1 hour at 22°C in the dark. Samples werewashed in PBS, and 10,000 particles were analyzed withWinMDI software.
Measurement of [Ca2+]i
PLTs were incubated with 0.2 μmol per L Fura-2-AM for45 minutes at 37°C (light-protected), and free Fura-2 wasremoved by a wash step under protection of PGI2
(10 nmol/L). Fura-2 fluorescence was recorded at 37°C ina fluorescence spectrophotometer (F-4500, Hitachi Ltd.,Tokyo, Japan) with excitation wavelengths of 340 and380 nm and emission at 510 nm. Basal [Ca2+]i was calcu-lated according to the method of Grynkiewicz andcoworkers.25
Confocal microscopy
Human washed PLTs (10 × 106) were incubated with mon-oclonal antibody CD42b-PE (2 μg/mL; AN51-PE, Dako A/S) and mepacrine (50 μg/mL) for 15 minutes at 37°C. THP-1 cells (5 × 106) were stimulated with PMA for 15 minutesat 37°C on coverslip. PLTs were then mixed with THP-1cells and incubated for 30 minutes at 22°C. Samples werefirst identified with a confocal laser scanning fluorescencemicroscope (TCS 4D, Leica) as defined.26 A 63× objective,0.75 NA oil immersion lens (Zeiss, Thornwood, NY) wasused to illuminate the sample and zoom 4 (31 × 31 μm)images were collected.
Statistical analysis
Data are expressed as means ± SEM with number ofobservations. Statistical analysis was based on a paired
trf_00913.fm Page 3 Wednesday, June 14, 2006 4:44 PM
BADLOU ET AL.
4 TRANSFUSION Volume **, ** **
t test or one-way analysis of variance (with post t test) forcomparison between two and more groups. Differenceswere considered significant at a p value of less than 0.05.
RESULTS
Binding and uptake of mepacrine-labeled PLTs by macrophages
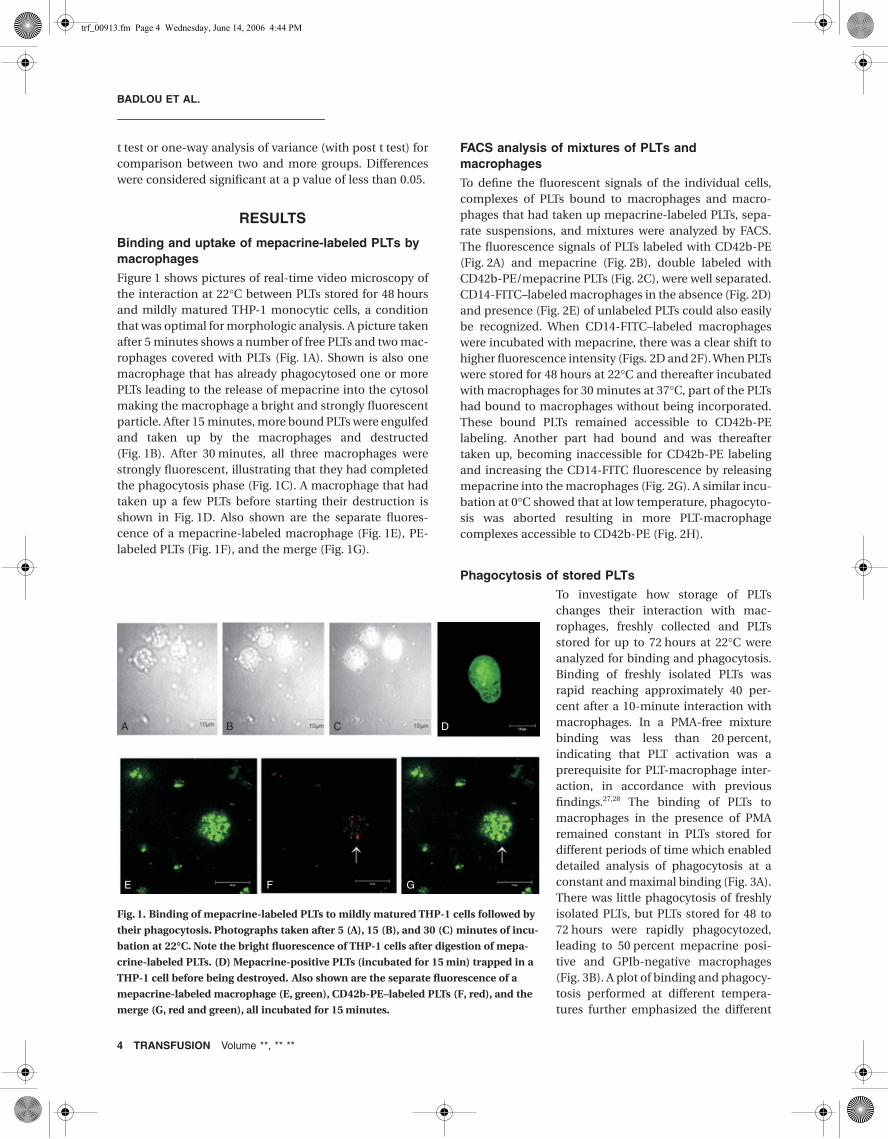
Figure 1 shows pictures of real-time video microscopy ofthe interaction at 22°C between PLTs stored for 48 hoursand mildly matured THP-1 monocytic cells, a conditionthat was optimal for morphologic analysis. A picture takenafter 5 minutes shows a number of free PLTs and two mac-rophages covered with PLTs (Fig. 1A). Shown is also onemacrophage that has already phagocytosed one or morePLTs leading to the release of mepacrine into the cytosolmaking the macrophage a bright and strongly fluorescentparticle. After 15 minutes, more bound PLTs were engulfedand taken up by the macrophages and destructed(Fig. 1B). After 30 minutes, all three macrophages werestrongly fluorescent, illustrating that they had completedthe phagocytosis phase (Fig. 1C). A macrophage that hadtaken up a few PLTs before starting their destruction isshown in Fig. 1D. Also shown are the separate fluores-cence of a mepacrine-labeled macrophage (Fig. 1E), PE-labeled PLTs (Fig. 1F), and the merge (Fig. 1G).
FACS analysis of mixtures of PLTs and macrophages
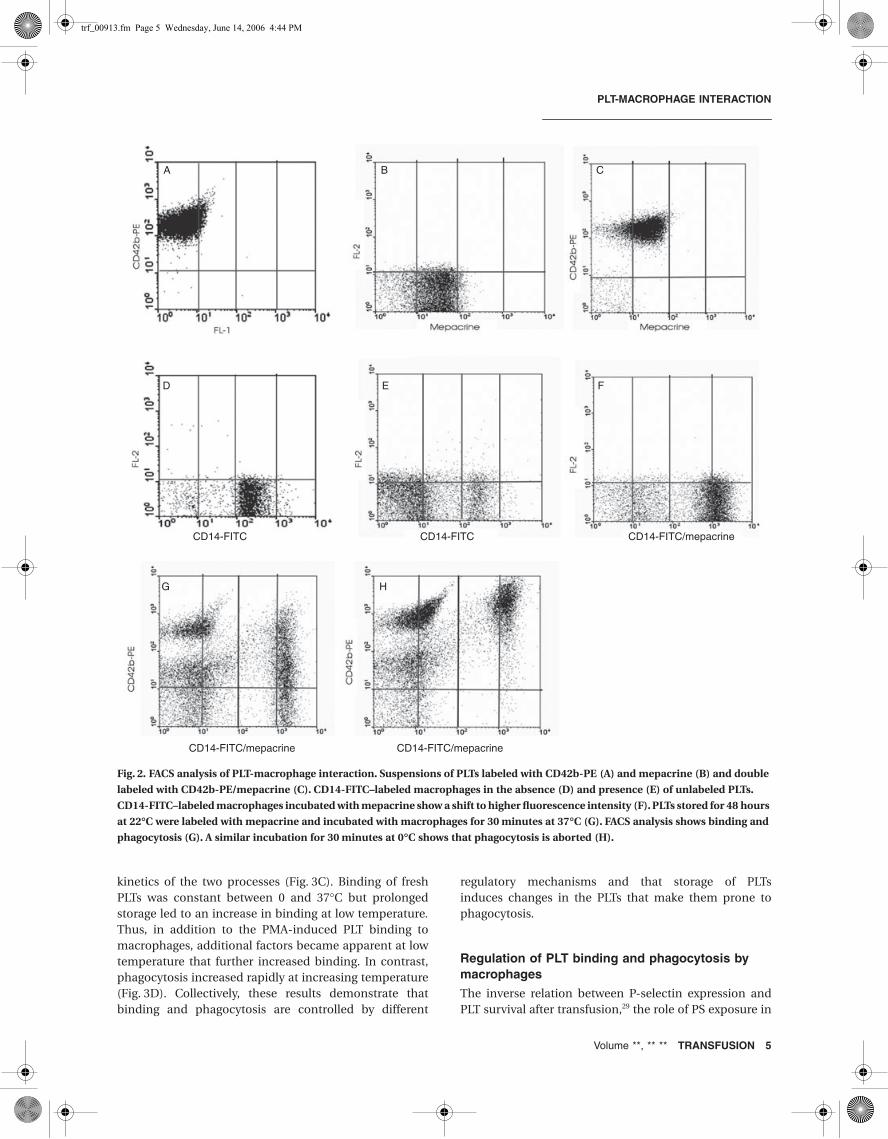
To define the fluorescent signals of the individual cells,complexes of PLTs bound to macrophages and macro-phages that had taken up mepacrine-labeled PLTs, sepa-rate suspensions, and mixtures were analyzed by FACS.The fluorescence signals of PLTs labeled with CD42b-PE(Fig. 2A) and mepacrine (Fig. 2B), double labeled withCD42b-PE/mepacrine PLTs (Fig. 2C), were well separated.CD14-FITC–labeled macrophages in the absence (Fig. 2D)and presence (Fig. 2E) of unlabeled PLTs could also easilybe recognized. When CD14-FITC–labeled macrophageswere incubated with mepacrine, there was a clear shift tohigher fluorescence intensity (Figs. 2D and 2F). When PLTswere stored for 48 hours at 22°C and thereafter incubatedwith macrophages for 30 minutes at 37°C, part of the PLTshad bound to macrophages without being incorporated.These bound PLTs remained accessible to CD42b-PElabeling. Another part had bound and was thereaftertaken up, becoming inaccessible for CD42b-PE labelingand increasing the CD14-FITC fluorescence by releasingmepacrine into the macrophages (Fig. 2G). A similar incu-bation at 0°C showed that at low temperature, phagocyto-sis was aborted resulting in more PLT-macrophagecomplexes accessible to CD42b-PE (Fig. 2H).
Phagocytosis of stored PLTs
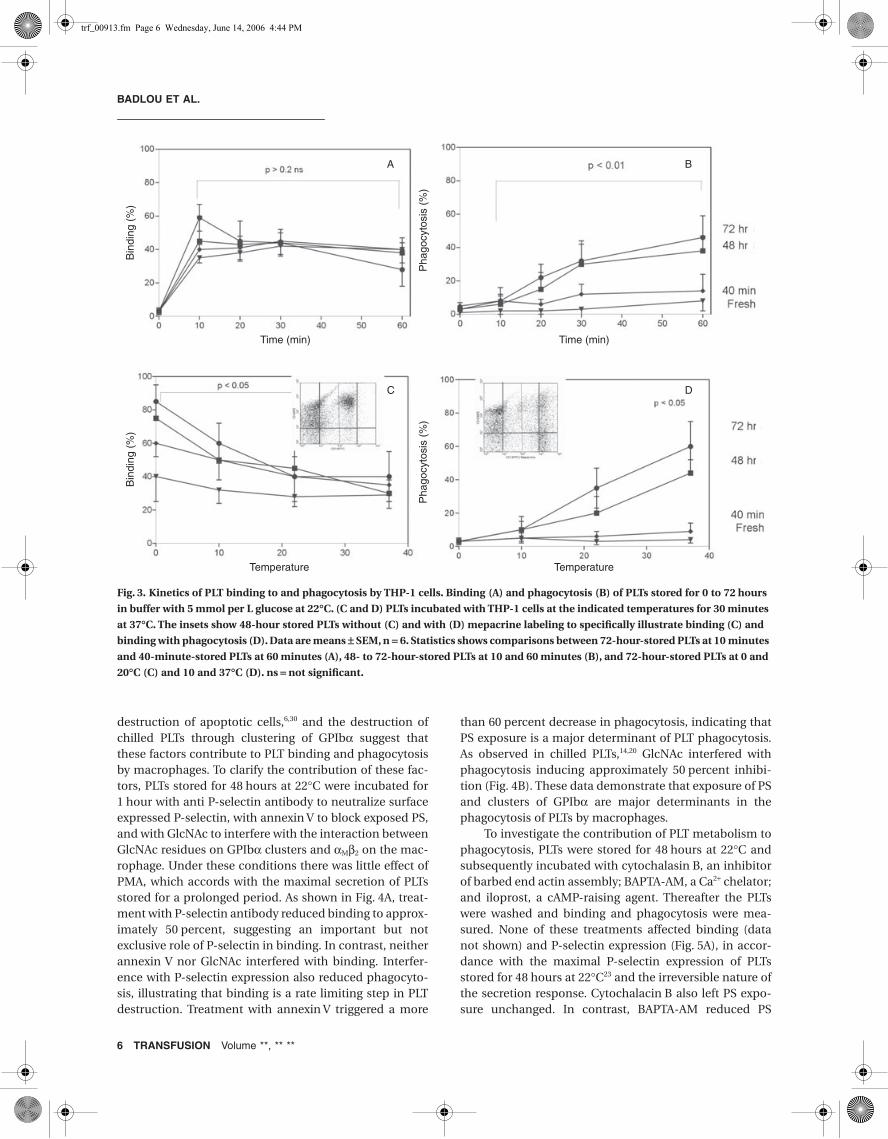
To investigate how storage of PLTschanges their interaction with mac-rophages, freshly collected and PLTsstored for up to 72 hours at 22°C wereanalyzed for binding and phagocytosis.Binding of freshly isolated PLTs wasrapid reaching approximately 40 per-cent after a 10-minute interaction withmacrophages. In a PMA-free mixturebinding was less than 20 percent,indicating that PLT activation was aprerequisite for PLT-macrophage inter-action, in accordance with previousfindings.27,28 The binding of PLTs tomacrophages in the presence of PMAremained constant in PLTs stored fordifferent periods of time which enableddetailed analysis of phagocytosis at aconstant and maximal binding (Fig. 3A).There was little phagocytosis of freshlyisolated PLTs, but PLTs stored for 48 to72 hours were rapidly phagocytozed,leading to 50 percent mepacrine posi-tive and GPIb-negative macrophages(Fig. 3B). A plot of binding and phagocy-tosis performed at different tempera-tures further emphasized the different
Fig. 1. Binding of mepacrine-labeled PLTs to mildly matured THP-1 cells followed by
their phagocytosis. Photographs taken after 5 (A), 15 (B), and 30 (C) minutes of incu-
bation at 22∞C. Note the bright fluorescence of THP-1 cells after digestion of mepa-
crine-labeled PLTs. (D) Mepacrine-positive PLTs (incubated for 15 min) trapped in a
THP-1 cell before being destroyed. Also shown are the separate fluorescence of a
mepacrine-labeled macrophage (E, green), CD42b-PE–labeled PLTs (F, red), and the
merge (G, red and green), all incubated for 15 minutes.
A B C
E F G
D
trf_00913.fm Page 4 Wednesday, June 14, 2006 4:44 PM
PLT-MACROPHAGE INTERACTION
Volume **, ** ** TRANSFUSION 5
kinetics of the two processes (Fig. 3C). Binding of freshPLTs was constant between 0 and 37°C but prolongedstorage led to an increase in binding at low temperature.Thus, in addition to the PMA-induced PLT binding tomacrophages, additional factors became apparent at lowtemperature that further increased binding. In contrast,phagocytosis increased rapidly at increasing temperature(Fig. 3D). Collectively, these results demonstrate thatbinding and phagocytosis are controlled by different
regulatory mechanisms and that storage of PLTsinduces changes in the PLTs that make them prone tophagocytosis.
Regulation of PLT binding and phagocytosis by macrophages
The inverse relation between P-selectin expression andPLT survival after transfusion,29 the role of PS exposure in
Fig. 2. FACS analysis of PLT-macrophage interaction. Suspensions of PLTs labeled with CD42b-PE (A) and mepacrine (B) and double
labeled with CD42b-PE/mepacrine (C). CD14-FITC–labeled macrophages in the absence (D) and presence (E) of unlabeled PLTs.
CD14-FITC–labeled macrophages incubated with mepacrine show a shift to higher fluorescence intensity (F). PLTs stored for 48 hours
at 22∞C were labeled with mepacrine and incubated with macrophages for 30 minutes at 37∞C (G). FACS analysis shows binding and
phagocytosis (G). A similar incubation for 30 minutes at 0∞C shows that phagocytosis is aborted (H).
A B C
D
CD14-FITC
CD14-FITC/mepacrine CD14-FITC/mepacrine
CD14-FITC CD14-FITC/mepacrine
E
G H
F
trf_00913.fm Page 5 Wednesday, June 14, 2006 4:44 PM
BADLOU ET AL.
6 TRANSFUSION Volume **, ** **
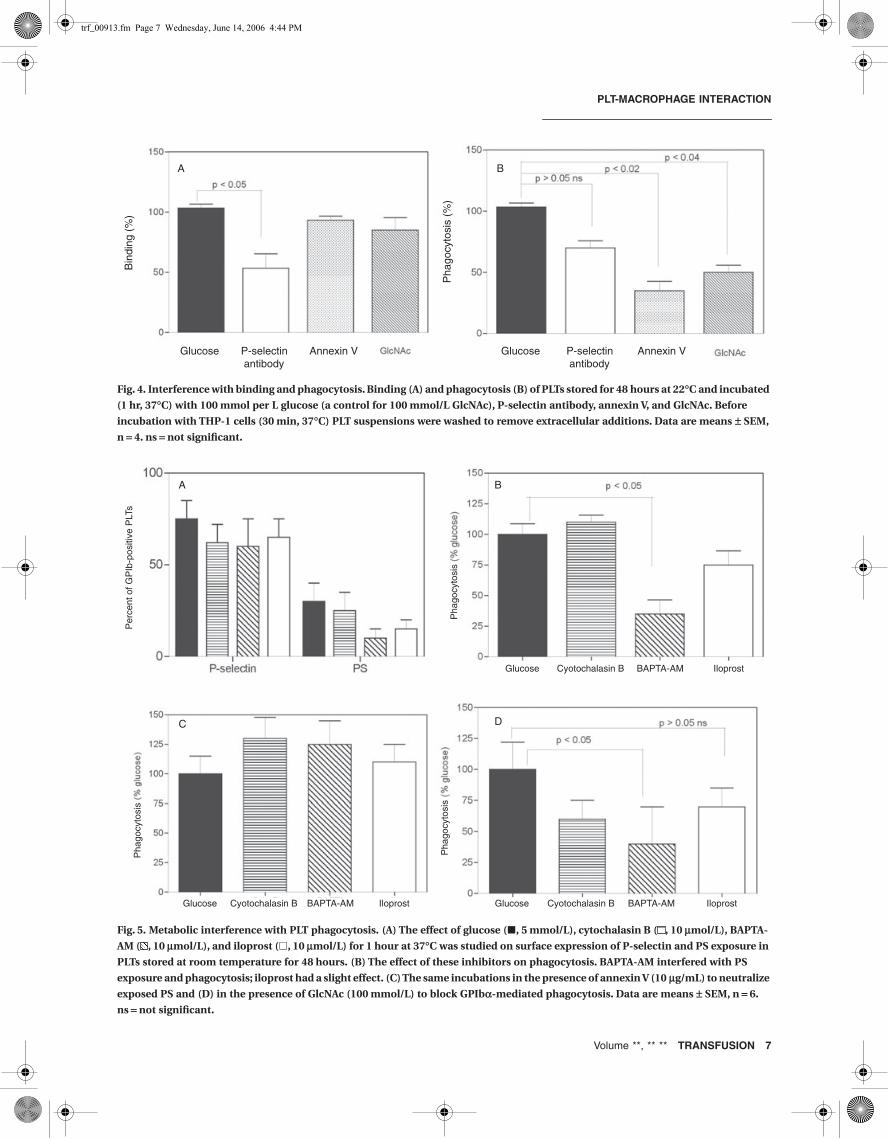
destruction of apoptotic cells,6,30 and the destruction ofchilled PLTs through clustering of GPIbα suggest thatthese factors contribute to PLT binding and phagocytosisby macrophages. To clarify the contribution of these fac-tors, PLTs stored for 48 hours at 22°C were incubated for1 hour with anti P-selectin antibody to neutralize surfaceexpressed P-selectin, with annexin V to block exposed PS,and with GlcNAc to interfere with the interaction betweenGlcNAc residues on GPIbα clusters and αMβ2 on the mac-rophage. Under these conditions there was little effect ofPMA, which accords with the maximal secretion of PLTsstored for a prolonged period. As shown in Fig. 4A, treat-ment with P-selectin antibody reduced binding to approx-imately 50 percent, suggesting an important but notexclusive role of P-selectin in binding. In contrast, neitherannexin V nor GlcNAc interfered with binding. Interfer-ence with P-selectin expression also reduced phagocyto-sis, illustrating that binding is a rate limiting step in PLTdestruction. Treatment with annexin V triggered a more
than 60 percent decrease in phagocytosis, indicating thatPS exposure is a major determinant of PLT phagocytosis.As observed in chilled PLTs,14,20 GlcNAc interfered withphagocytosis inducing approximately 50 percent inhibi-tion (Fig. 4B). These data demonstrate that exposure of PSand clusters of GPIbα are major determinants in thephagocytosis of PLTs by macrophages.
To investigate the contribution of PLT metabolism tophagocytosis, PLTs were stored for 48 hours at 22°C andsubsequently incubated with cytochalasin B, an inhibitorof barbed end actin assembly; BAPTA-AM, a Ca2+ chelator;and iloprost, a cAMP-raising agent. Thereafter the PLTswere washed and binding and phagocytosis were mea-sured. None of these treatments affected binding (datanot shown) and P-selectin expression (Fig. 5A), in accor-dance with the maximal P-selectin expression of PLTsstored for 48 hours at 22°C23 and the irreversible nature ofthe secretion response. Cytochalacin B also left PS expo-sure unchanged. In contrast, BAPTA-AM reduced PS
Fig. 3. Kinetics of PLT binding to and phagocytosis by THP-1 cells. Binding (A) and phagocytosis (B) of PLTs stored for 0 to 72 hours
in buffer with 5 mmol per L glucose at 22∞C. (C and D) PLTs incubated with THP-1 cells at the indicated temperatures for 30 minutes
at 37∞C. The insets show 48-hour stored PLTs without (C) and with (D) mepacrine labeling to specifically illustrate binding (C) and
binding with phagocytosis (D). Data are means ± SEM, n = 6. Statistics shows comparisons between 72-hour-stored PLTs at 10 minutes
and 40-minute-stored PLTs at 60 minutes (A), 48- to 72-hour-stored PLTs at 10 and 60 minutes (B), and 72-hour-stored PLTs at 0 and
20∞C (C) and 10 and 37∞C (D). ns = not significant.
Bin
ding
(%
)B
indi
ng (
%)
Pha
gocy
tosi
s (%
)P
hago
cyto
sis
(%)
A
Time (min)
Temperature Temperature
Time (min)
B
C D
trf_00913.fm Page 6 Wednesday, June 14, 2006 4:44 PM
PLT-MACROPHAGE INTERACTION
Volume **, ** ** TRANSFUSION 7
Fig. 4. Interference with binding and phagocytosis. Binding (A) and phagocytosis (B) of PLTs stored for 48 hours at 22∞C and incubated
(1 hr, 37∞C) with 100 mmol per L glucose (a control for 100 mmol/L GlcNAc), P-selectin antibody, annexin V, and GlcNAc. Before
incubation with THP-1 cells (30 min, 37∞C) PLT suspensions were washed to remove extracellular additions. Data are means ± SEM,
n = 4. ns = not significant.
Bin
ding
(%
)
Pha
gocy
tosi
s (%
)
Glucose P-selectin antibody
Annexin V Glucose P-selectin antibody
Annexin V
A B
Fig. 5. Metabolic interference with PLT phagocytosis. (A) The effect of glucose (�, 5 mmol/L), cytochalasin B ( , 10 mmol/L), BAPTA-
AM ( , 10 mmol/L), and iloprost (�, 10 mmol/L) for 1 hour at 37∞C was studied on surface expression of P-selectin and PS exposure in
PLTs stored at room temperature for 48 hours. (B) The effect of these inhibitors on phagocytosis. BAPTA-AM interfered with PS
exposure and phagocytosis; iloprost had a slight effect. (C) The same incubations in the presence of annexin V (10 mg/mL) to neutralize
exposed PS and (D) in the presence of GlcNAc (100 mmol/L) to block GPIba-mediated phagocytosis. Data are means ± SEM, n = 6.
ns = not significant.
Pha
gocy
tosi
sP
erce
nt o
f GP
Ib-p
ositi
ve P
LTs
Pha
gocy
tosi
s
Glucose Cyotochalasin B BAPTA-AM Iloprost Glucose Cyotochalasin B BAPTA-AM Iloprost
Glucose Cyotochalasin B BAPTA-AM Iloprost
Pha
gocy
tosi
s
A B
C D
trf_00913.fm Page 7 Wednesday, June 14, 2006 4:44 PM

BADLOU ET AL.
8 TRANSFUSION Volume **, ** **
exposure by approximately 70 percent (Fig. 5B). Also ilo-prost slightly reduced phagocytosis but the difference wasnot significant. Phagocytosis was not affected by cytocha-lasin B treatment, which accords with observations inchilled PLTs.14,22 These data indicate that cytosolic Ca2+
and cAMP contribute to the regulation of phagocytosis ofPLTs. To investigate whether the lower phagocytosis in thepresence of these inhibitors was the result of interferencewith GPIbα clustering or PS expression, studies wererepeated in the presence of annexin V to block exposed PSand with GlcNAc to interfere with phagocytosis throughclusters of GPIbα. The presence of annexin V rescuedphagocytosis of PLTs pretreated with BAPTA-AM and ilo-prost (Fig. 5C). In the presence of GlcNAc, the effect ofthe inhibitors remained the same (Fig. 5D). In addition,cytochalasin B reduced phagocytosis by 55 percent.Because this inhibitor failed to interfere with PS expres-sion, this points to the involvement of other factors thatcontribute to phagocytosis regulation under conditionsthat suppress the clustering of GPIbα. Together with thereduced surface expression of PS in the presence of theseinhibitors, these findings indicate that BAPTA-AM andiloprost reduce phagocytosis by lowering the exposureof PS without interfering with the formation of GPIbαclusters.
Optimal preservation of PLTs
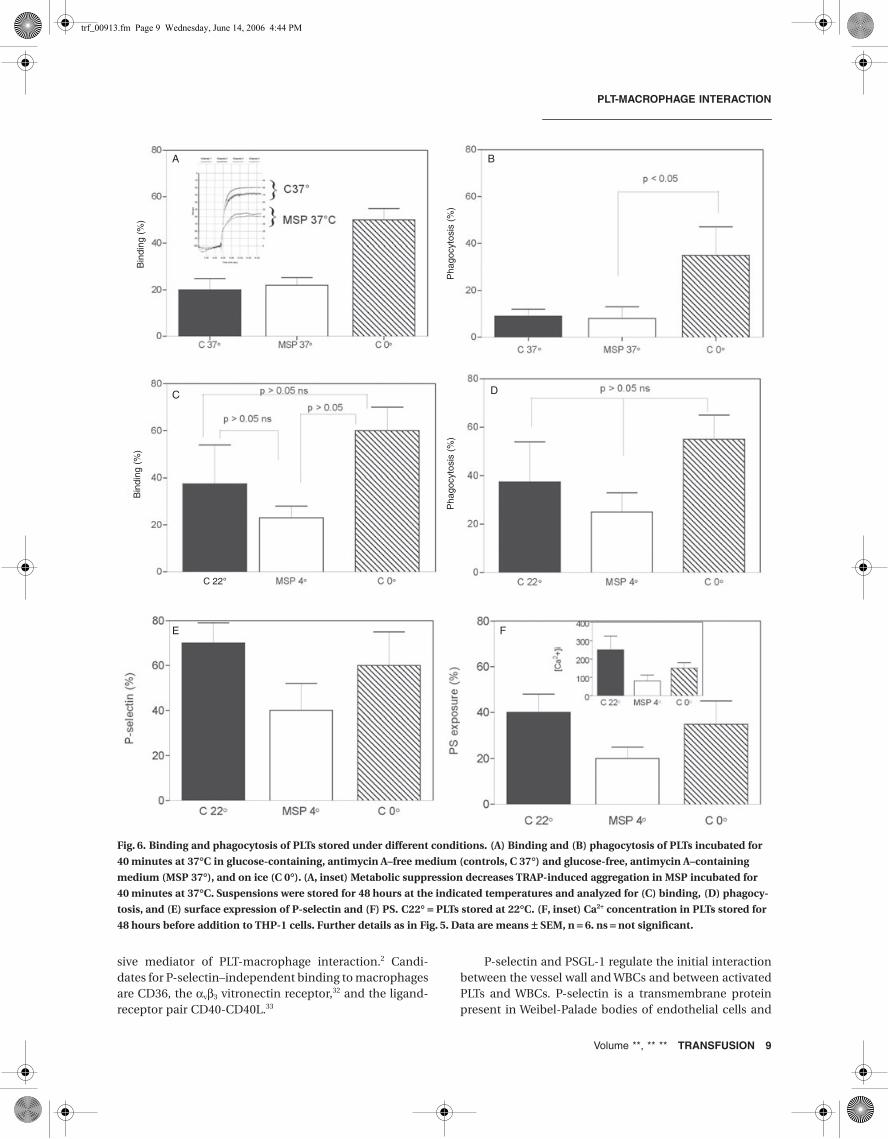
Our earlier observation23 that metabolic arrest followed bystorage at low temperature and recovery with glucose at37°C better preserves PLT adhesion and aggregation thanPLTs stored at 22 and 0°C led to the question whether thistreatment also reduced binding and phagocytosis. MSPswere prepared by incubating washed PLTs in glucose-free,antimycin A–containing medium for 40 minutes at 37°Cto reduce glycolytic energy supply and abort mitochon-drial resynthesis of metabolic ATP. The PLTs were thenstored at 4°C for 48 hours, and thereafter energy genera-tion was restored by incubation with glucose at 37°C. Aftera 40-minute incubation in energy-rich medium, PLTsshowed 20 percent binding to macrophages in a PMA-richmilieu. A similar incubation in energy-free mediuminduced a similar binding, indicating that under theseconditions lowered energy content does not affect bindingalthough it reduced aggregation (Fig. 6A and inset). Incontrast, PLTs chilled on ice without metabolic interfer-ence showed a 2.5-fold increase in binding compared withcontrols. When phagocytosis of the three PLT preparationswas analyzed, similar differences were observed. PLTsstored at 22°C and MSPs showed approximately10 percent phagocytosis (Fig. 6B). Chilling induced almostthree times more, illustrating the induction of phagocyto-sis by a temperature fall as described earlier.14 Subsequentstorage at 22°C increased binding but not at 4 and 0°C(Fig. 6C).
In contrast, phagocytosis increased during prolongedstorage in all three storage conditions (Fig. 6D). These dif-ferences were accompanied by similar changes in P-selec-tin expression and PS exposure. Compared with roomtemperature–stored and chilled PLTs, MSPs appear betterprotected against the storage-induced changes that trig-ger PS expression, probably because of their capacity topreserve a low [Ca2+]i (Fig. 6F, inset). Together these datashow that differences in binding explain a major part ofthe differences in phagocytosis observed between thethree suspensions. Similar studies in the absence of PMAshowed lower binding and phagocytosis after 40 minutesof storage but the differences between the three suspen-sions remained the same. After 48 hours of storage,removal of PMA had little effect (data not shown).
To investigate the contribution of PS exposure andGPIbα clustering in MSPs and chilled PLTs in more detail,phagocytosis was measured in the presence of annexin Vto block the role of expressed PS and GlcNAc to blockphagocytosis through clusters of GPIbα. In addition,MSPs were analyzed before and after the recovery phasewith glucose to address the role of metabolic energy(Fig. 7). MSPs that were still in the energy-depleted phasewere phagocytosed by macrophages as seen with MSPsafter the recovery phase. Neither annexin V nor GlcNAcinterfered with phagocytosis of these cells, indicatingthat other processes than PS exposure and GPIbα cluster-ing were responsible for destruction. Recovery withglucose induced a 30 percent lower phagocytosis. Thisnumber was unchanged in the presence of annexin V butthe GlcNAc induced a fall in phagocytosis as observed inPLTs stored at 22C. PLTs chilled to 0°C and stored for48 hours showed lower phagocytosis in the presence ofGlcNAc, which is similar as seen with PLTs stored at roomtemperature.
DISCUSSION
This study shows that binding of PLTs to macrophages andthe phagocytosis that follows are regulated by differentmechanisms. Freshly collected PLTs in the absence of PLTactivators show little binding, and PMA treatment isrequired to induce rapid and complete interaction withmacrophages. The PMA-induced binding remains con-stant when PLTs are stored at 22°C, which makes it pos-sible to investigate the regulation of phagocytosis. Thebinding appears for a major part mediated through P-selectin expressed on the PLT surface because 1) it isabsent in PLTs with blocked secretion by prostacyclintreatment,7 2) present in PLTs that express P-selectin,29,31
and 3) inhibited by a P-selectin antibody (this study). Inhi-bition by anti–P-selectin is incomplete, suggesting that anadditional mechanism might be involved that has yet tobe identified. In fact, PLTs survive normally in P-selectin–deficient mice indicating that P-selectin is not an exclu-
trf_00913.fm Page 8 Wednesday, June 14, 2006 4:44 PM
PLT-MACROPHAGE INTERACTION
Volume **, ** ** TRANSFUSION 9
sive mediator of PLT-macrophage interaction.2 Candi-dates for P-selectin–independent binding to macrophagesare CD36, the αvβ3 vitronectin receptor,32 and the ligand-receptor pair CD40-CD40L.33
P-selectin and PSGL-1 regulate the initial interactionbetween the vessel wall and WBCs and between activatedPLTs and WBCs. P-selectin is a transmembrane proteinpresent in Weibel-Palade bodies of endothelial cells and
Fig. 6. Binding and phagocytosis of PLTs stored under different conditions. (A) Binding and (B) phagocytosis of PLTs incubated for
40 minutes at 37∞C in glucose-containing, antimycin A–free medium (controls, C 37∞) and glucose-free, antimycin A–containing
medium (MSP 37∞), and on ice (C 0∞). (A, inset) Metabolic suppression decreases TRAP-induced aggregation in MSP incubated for
40 minutes at 37∞C. Suspensions were stored for 48 hours at the indicated temperatures and analyzed for (C) binding, (D) phagocy-
tosis, and (E) surface expression of P-selectin and (F) PS. C22∞ = PLTs stored at 22∞C. (F, inset) Ca2+ concentration in PLTs stored for
48 hours before addition to THP-1 cells. Further details as in Fig. 5. Data are means ± SEM, n = 6. ns = not significant.
Bin
ding
(%
)
Pha
gocy
tosi
s (%
)
A B
Bin
ding
(%
)
C 22°
Pha
gocy
tosi
s (%
)
C D
E F
trf_00913.fm Page 9 Wednesday, June 14, 2006 4:44 PM

BADLOU ET AL.
10 TRANSFUSION Volume **, ** **
α-granules of PLTs and rapidly translocates to the cell sur-face after cell activation.34 Together with αMβ2 integrin,PSGL-1 regulates WBC adhesion to the endothelium.35
PSGL-1 is also present on matured THP-1 cells,36 and thecoupling with P-selectin might be a first step in the pro-cess that lead to PLT destruction. PSGL-1–mediatedmonocyte activation leads to conformational activation ofαMβ2; synthesis and release of various cytokines, chemo-kines, and reactive oxygen species; and tissue factorexpression.37 It also triggers surface expression of PScontributing to PS-mediated activation of the coagulationsystem.37 If a similar mechanism is operational in mac-rophages, stimulation of PSGL-1 would initiate bothrelease of chemokines and phagocytosis of PLTs.
Phagocytosis of PLTs depends on surface exposure ofPS and clustering of GPIbα, and neither one appearsinvolved in binding. The binding of lymphocytes, Jurkatcells, and neutrophils to macrophages is also independentof PS.38 Hoffmann and associates6 postulated that regard-less of the receptors engaged on the phagocyte, ingestiondoes not occur in the absence of exposed PS. This studyshows that neutralization of exposed PS by annexin Vleads to more than 50 percent inhibition of PLT phagocy-tosis, indicating that exposed PS is a crucial component inthe phagocytotic pathway such in agreement with find-ings in lymphocytes or neutrophils.3,30,38 The asymmetricphospholipid distribution in plasma membranes is nor-mally maintained by energy-dependent lipid transportersthat translocate different phospholipids from one mono-layer to the other against their respective concentration
gradients.30 When PLTs are activated or enter apoptosis,lipid asymmetry might be perturbed by scramblases thatshuttle phospholipids nonspecifically between the twomonolayers30 in a process depending on ATP and [Ca2+]i.In stored PLTs PS exposure may function in the same waygenerating a signal for ingestion.6,14 Recognition of PSdepends on PS receptor6 on the macrophage. The scaven-ger receptors LOX-1,39 CD36,40 SRB-1,41 and integrin αvβ3
may serve a role in PS recognition. The signaling cascadesinitiated by the PS counter receptor may involve the smallGTPases Rac42 and cdc42,43 which are implicated in mem-brane ruffling.6
GPIbα clustering through sugar linkages has beendemonstrated in PLTs that were rapidly chilled on ice andis considered a main cause for the short survival of cold-stored PLTs after transfusion. The present data show thatGPIbα-mediated phagocytosis by macrophages alsooccurs during prolonged storage at room temperature,which is daily practice in blood banks. Clustering of GPIbαis independent of barbed end actin assembly because itoccurs in the presence of cytochalasin B in agreementwith earlier observations.14,22 Interference with PLTmetabolism after prolonged storage did not change thebinding properties but reduced phagocytosis. BecauseGPIbα clustering appears not to be affected by intracellu-lar mediators,20 it is likely that the reduced phagocytosisin the presence of BAPTA-AM and iloprost reflect interfer-ence with PS exposure. BAPTA binds cytosolic Ca2+ andsuppresses Ca2+ increases after PLT activation44 and stor-age. BAPTA left binding unchanged but reduced phago-
Fig. 7. PS- and GPIba-mediated phagocytosis of MSPs and chilled PLTs. (A) Phagocytosis of MSPs stored at 4∞C (MSP 4∞) for 48 hours
before (0 mmol/L glucose)and after (20 mmol/L glucose) in the presence of annexin V and GlcNAc. Recovery reduces phagocytosis
and restores the inhibition by GlcNAc but not by annexin V. (B) Chilled PLTs (C 0∞) stored on ice for 48 hours and treated with 100 mmol
per L glucose, annexin V, and GlcNAc. PLTs stored at 0∞C remain sensitive to GlcNAc. Data are means ± SEM, n = 4. ns = not significant.
Pha
gocy
tosi
s (%
)
Control Annexin V Glucose Glucose Annexin VAnnexin V
Pha
gocy
tosi
s (%
)
A B
trf_00913.fm Page 10 Wednesday, June 14, 2006 4:44 PM

PLT-MACROPHAGE INTERACTION
Volume **, ** ** TRANSFUSION 11
cytosis by more than 50 percent. Earlier studies havedemonstrated that PS exposure is a reversible processalthough the mechanism that restores the positioning ofPS on the inner leaflet of the plasma membrane is unex-plained.45 Iloprost is a stable prostacyclin derivative andinduced a slight inhibition of phagocytosis but did notinterfere with binding. Prostacyclin is known to increasecAMP, which through protein kinase A inhibits many stepsin the PLT activation cascades. It also suppresses anincrease in [Ca2+]i, and the decrease in phagocytosisinduced by iloprost might well reflect the inhibition ofCa2+ signaling thereby lowering PS expression.46
Storage at room temperature also led to a decrease inCD42b-PE fluorescence. A similar decrease was found inPLTs stored at 4°C in the presence of proteolysis inhibitors(data not shown). Whether this change in antibody bind-ing reflects changes in GPIbα involved in phagocytosis hasyet to be investigated.
To investigate how different storage conditionsinduced binding and phagocytosis, PLTs stored at 22°Cwere compared with PLTs stored at 0°C and PLTs with met-abolic arrest (MSP). At the end of the 40-minute starvationperiod at 37°C, binding and phagocytosis of MSP were notdifferent from PLTs incubated in a normal milieu. In con-trast, chilling of PLTs to 0°C increased binding and phago-cytosis. Chilling-induced phagocytosis has been reportedearlier,14 and the present findings suggest that increasedbinding contributes to the high phagocytosis of thesecells. Subsequent storage for 48 hours further increasedbinding of PLTs stored at 22°C but not of PLTs stored at0°C. Analysis of P-selectin expression confirmed the dif-ferences in binding between these suspensions. In con-trast, after 48 hours of storage all suspensions showedmore phagocytosis. Again, differences in phagocytosisbetween suspensions were accompanied with similar dif-ferences in PS exposure confirming the role of PS as amajor phagocytotic signal. Thus, the advantage of low-temperature storage is suppression of binding and phago-cytosis signals during storage. The advantage of metabolicsuppression lies in the protection against cell alterationsinflicted during the fall in temperature. Separate analysisof PS- and GPIbα-mediated phagocytosis learned thatmetabolic suppression attenuated the increase in [Ca2+]i
during PLT storage, resulting in a lower PS expression andPS-mediated phagocytosis compared with PLTs storedat 22 and 0°C. GPIbα-mediated phagocytosis remainedpresent provided that energy metabolism was restored. Inaddition to GPIbα-mediated phagocytosis, chilled pre-served PS mediated phagocytosis in accordance with thehigher level of [Ca2+]i and exposed PS seen in these cells.A limitation of this study that should be acknowledged isits restriction to in vitro conditions. Thus, although therelevance of our findings for PLT survival in vivo remainsunanswered, they may help to define conditions thatimprove the survival of PLTs after transfusion.
ACKNOWLEDGMENTS
The authors thank A. Martens PhD and H. Rozenmuller PhD,
Department of Haematology, UMCU, for helpful discussions.
REFERENCES
1. Rinder HM, Bonan JL, Rinder CS, Ault KA, Smith BR.
Dynamics of leukocyte-platelet adhesion in whole blood.
Blood 1991;78:1730-7.
2. Berger G, Hartwell DW, Wagner DD. P-selectin and platelet
clearance. Blood 1998;92:4446-52.
3. Brown SB, Clarke MC, Magowan L, Sanderson H, Savill J.
Constitutive death of platelets leading to scavenger
receptor-mediated phagocytosis. A caspase-independent
cell clearance program. J Biol Chem 2000;275:5987-96.
4. Stuart MC, Bevers EM, Comfurius P, et al. Ultrastructural
detection of surface exposed phosphatidylserine on
activated blood platelets. Thromb Haemost 1995;74:1145-
51.
5. Watala C, Waczulikova I, Wieclawska B, et al. Merocyanine
540 as a fluorescent probe of altered membrane
phospholipid asymmetry in activated whole blood platelets.
Cytometry 2002;49:119-33.
6. Hoffmann PR, deCathelineau AM, Ogden CA, et al.
Phosphatidylserine (PS) induces PS receptor-mediated
macropinocytosis and promotes clearance of apoptotic
cells. J Cell Biol 2001;155:649-59.
7. Elias M, Heethuis A, Weggemans M, et al. Stabilization of
standard platelet concentrates and minimization of the
platelet storage lesion by a prostacyclin analogue. Ann
Hematol 1992;64:292-8.
8. Oury C, Sticker E, De Cornelissen HV, Vermylen J, Hoylaerts
MF. ATP augments von Willebrand factor-dependent shear-
induced platelet aggregation through Ca2+-calmodulin and
myosin light chain kinase activation. J Biol Chem 2004;279:
26266-73.
9. Verhoeven AJ, Mommersteeg ME, Akkerman JW. Kinetics of
energy consumption in human platelets with blocked ATP
regeneration. Int J Biochem 1986;18:985-90.
10. Solum NO. Procoagulant expression in platelets and defects
leading to clinical disorders. Arterioscler Thromb Vasc Biol
1999;19:2841-6.
11. Andrews RK, Berndt MC. Platelet physiology: in cold blood.
Curr Biol 2003;13:R282-4.
12. Winokur R, Hartwig JH. Mechanism of shape change in
chilled human platelets. Blood 1995;85:1796-804.
13. Hoffmeister KM, Falet H, Toker A, et al. Mechanisms of cold-
induced platelet actin assembly. J Biol Chem 2001;276:
24751-9.
14. Hoffmeister KM, Felbinger TW, Falet H, et al. The clearance
mechanism of chilled blood platelets. Cell 2003;112:87-97.
15. Andrews RK, Gardiner EE, Shen Y, Whisstock JC, Berndt MC.
Glycoprotein Ib-IX-V. Int J Biochem Cell Biol 2003;35:1170-
4.
trf_00913.fm Page 11 Wednesday, June 14, 2006 4:44 PM

BADLOU ET AL.
12 TRANSFUSION Volume **, ** **
16. Huizinga EG, Tsuji S, Romijn RA, et al. Structures of
glycoprotein Ibalpha and its complex with von Willebrand
factor A1 domain. Science 2002;297:1176-9.
17. Andrews RK, Lopez JA, Berndt MC. Molecular mechanisms
of platelet adhesion and activation. Int J Biochem Cell Biol
1997;29:91-105.
18. Kovacsovics TJ, Hartwig JH. Thrombin-induced GPIb-IX
centralization on the platelet surface requires actin
assembly and myosin II activation. Blood 1996;87:618-
29.
19. Wu YP, Vink T, Schiphorst M, et al. Platelet thrombus
formation on collagen at high shear rates is mediated by von
Willebrand factor-glycoprotein Ib interaction and inhibited
by von Willebrand factor-glycoprotein IIb/IIIa interaction.
Arterioscler Thromb Vasc Biol 2000;20:1661-7.
20. Hoffmeister KM, Josefsson EC, Isaac NA, et al. Glycosylation
restores survival of chilled blood platelets. Science 2003;
301:1531-4.
21. Akkerman JW, Gorter G. Relation between energy
production and adenine nucleotide metabolism in human
blood platelets. Biochim Biophys Acta 1980;590:107-16.
22. Josefsson EC, Gebhard HH, Stossel TP, Hartwig JH,
Hoffmeister KM. The macrophage alphaMbeta2 integrin
alphaM lectin domain mediates the phagocytosis of chilled
platelets. J Biol Chem 2005;280:18025-32.
23. Badlou BA, Ijseldijk MJ, Smid WM, Akkerman JW. Prolonged
platelet preservation by transient metabolic suppression.
Transfusion 2005;45:214-22.
24. Tibbles HE, Navara CS, Hupke MA, Vassilev AO, Uckun FM.
Thrombopoietin induces P-selectin expression on platelets
and subsequent platelet/leukocyte interactions. Biochem
Biophys Res Commun 2002;292:987-91.
25. Grynkiewicz G, Poenie M, Tsien RY. A new generation of
Ca2+ indicators with greatly improved fluorescence
properties. J Biol Chem 1985;260:3440-50.
26. Wu YP, Bloemendal HJ, Voest EE, et al. Fibrin-incorporated
vitronectin is involved in platelet adhesion and thrombus
formation through homotypic interactions with platelet-
associated vitronectin. Blood 2004;104:1034-41.
27. Caron A, Theoret JF, Mousa SA, Merhi Y. Anti-platelet effects
of GPIIb/IIIa and P-selectin antagonism, platelet activation,
and binding to neutrophils. J Cardiovasc Pharmacol
2002;40:296-306.
28. Gutensohn K, Geidel K, Brockmann M, et al. Binding of
activated platelets to WBCs in vivo after transfusion.
Transfusion 2002;42:1373-80.
29. Leytin V, Allen DJ, Gwozdz A, Garvey B, Freedman J. Role of
platelet surface glycoprotein Ibalpha and P-selectin in the
clearance of transfused platelet concentrates. Transfusion
2004;44:1487-95.
30. Fadok VA, Henson PM. Apoptosis: giving phosphatidylserine
recognition an assist—with a twist. Curr Biol 2003;13:R655-
R657.
31. Fijnheer R, Modderman PW, Veldman H, et al. Detection of
platelet activation with monoclonal antibodies and flow
cytometry: changes during platelet storage. Transfusion
1990;30:20-5.
32. Monks J, Rosner D, Geske FJ, et al. Epithelial cells as
phagocytes: apoptotic epithelial cells are engulfed by
mammary alveolar epithelial cells and repress inflammatory
mediator release. Cell Death Differ 2005;12:107-14.
33. Langer F, Ingersoll SB, Amirkhosravi A, et al. The role of
CD40 in CD40L- and antibody-mediated platelet activation.
Thromb Haemost 2005;93:1137-46.
34. Merten M, Thiagarajan P. P-selectin in arterial thrombosis.
Z Kardiol 2004;93:855-63.
35. Vandendries ER, Furie BC, Furie B. Role of P-selectin and
PSGL-1 in coagulation and thrombosis. Thromb Haemost
2004;92:459-66.
36. del CI, Nabi F, Tonda R, Thiagarajan P, Lopez JA, Kleiman
NS. Effect of P-selectin on phosphatidylserine exposure and
surface-dependent thrombin generation on monocytes.
Arterioscler Thromb Vasc Biol 2005;25:1065-70.
37. Del Conde I, Shrimpton CN, Thiagarajan P, Lopez JA. Tissue
factor-bearing microvesicles arise from lipid rafts and fuse
with activated platelets to initiate coagulation. Blood
2005;106:1604-11.
38. Savill J, Smith J, Sarraf C, et al. Glomerular mesangial cells
and inflammatory macrophages ingest neutrophils
undergoing apoptosis. Kidney Int 1992;42:924-36.
39. Oka K, Sawamura T, Kikuta K, et al. Lectin-like oxidized low-
density lipoprotein receptor 1 mediates phagocytosis of
aged/apoptotic cells in endothelial cells. Proc Natl Acad Sci
U S A 1998;95:9535-40.
40. Tait JF, Smith C. Phosphatidylserine receptors. role of CD36
in binding of anionic phospholipid vesicles to monocytic
cells. J Biol Chem 1999;274:3048-54.
41. Linton MF, Fazio S. Macrophages, inflammation, and
atherosclerosis. Int J Obes Relat Metab Disord 2003;27(Suppl
3):S35-40.
42. Park JB. Phagocytosis induces superoxide formation and
apoptosis in macrophages. Exp Mol Med 2003;35:325-35.
43. Sumita C, Yamane M, Matsuda T, et al. Platelet activating
factor induces cytoskeletal reorganization through Rho
family pathway in THP-1 macrophages. FEBS Lett 2005;
579:4038-42.
44. Haslam RJ, Coorssen JR. Evidence that activation of
phospholipase D can mediate secretion from permeabilized
platelets. Adv Exp Med 1993;344:149-64. 149-64.
45. Comfurius P, Senden JM, Tilly RH, et al. Loss of membrane
phospholipid asymmetry in platelets and red cells may be
associated with calcium-induced shedding of plasma
membrane and inhibition of aminophospholipid
translocase. Biochim Biophys Acta 1990;1026:153-60.
46. Perrotta PL, Perrotta CL, Snyder EL. Apoptotic activity in
stored human platelets. Transfusion 2003;43:526-35.
trf_00913.fm Page 12 Wednesday, June 14, 2006 4:44 PM





























