Embed Size (px)
Citation preview

Class III β-tubulin, a novel biomarker in the human melanocyte lineage
Heiko Locher a,n, Karien E. de Rooij b,e, John C.M.J. de Groot a, Remco van Doorn c,Nelleke A. Gruis c, Clemens W.G.M. Löwik b, Susana M. Chuva de Sousa Lopes d,Johan H.M. Frijns a, Margriet A. Huisman a
a Department of Otorhinolaryngology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandsb Department of Radiology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandsc Department of Dermatology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandsd Department of Anatomy and Embryology, Leiden University Medical Center, PO Box 9600, 2300 RC Leiden, The Netherlandse Percuros B.V., Plesmanlaan 1, 2333 BZ Leiden, The Netherlands
a r t i c l e i n f o
Article history:Received 7 September 2012Received in revised form16 May 2013Accepted 23 May 2013
Keywords:Class III beta-tubulinHumansMelanocytesMelanoblastsNeurons
a b s t r a c t
It is generally thought that class III β-tubulin expression is limited to cells of the neural lineage and istherefore often used to identify neurons amongst other cell types, both in vivo and in vitro. Melanocytesare derived from the neural crest and share both morphological features and functional characteristicswith peripheral neurons. Here, we show that these similarities extend to class III β-tubulin (TUBB3)expression, and that human melanocytes express this protein both in vivo and in vitro. In addition, westudied the expression of class III β-tubulin in two murine melanogenic cell lines and show thatexpression of this protein starts as melanoblasts mature into melanocytes. Melanin bleaching experi-ments revealed close proximity between melanin and TUBB3 proteins. In vitro stimulation of primaryhuman melanocytes by α-MSH indicated separate regulatory mechanisms for melanogenesis and toTUBB3 expression. Together, these observations imply that human melanocytes express TUBB3 and thatthis protein should be recognized as a wider marker for multiple neural crest-derived cells.
& 2013 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.
1. Introduction
Over the last several years, there has been a notable increase ininterest in the culture of skin and skin stem cells, in part for thegeneration of skin substitutes, in part with view of understandingcutaneous pathologies. It is of great importance to be able toidentify the different skin cell subtypes accurately in culture inorder to fully understand their biology. A widely used biomarker inthis context is class III β-tubulin (TUBB3), which is generallythought to be expressed only in cells of the neural lineage.
Microtubules are one of the main classes of cytoskeletalfilaments involved in cellular morphology, motility, division andintracellular transport. They are composed of assemblies of het-erodimers one α- and one β-tubulin subunit (Bryan and Wilson,1971; Downing and Nogales, 1998). There are multiple β-tubulin
isotypes and their expression is tissue-specific and depends on thedevelopmental stage (Ludueña, 1998). Isotype TUBB3 was firstdescribed in avian and mammalian species (Sullivan andCleveland, 1986). Antibodies directed against TUBB3 epitopesdistinguished between neurons and glial cells (Geisert andFrankfurter, 1989; Moody et al., 1989), so that it became a standardbiomarker for neuronal identification. TUBB3 expression is oftenused to identify neuronal cells in skin-derived and (epidermal)stem cell cultures. In vivo it is used to trace the dendritic processesof neurons in the dermis and epidermis, of which the cell bodiesare located elsewhere.
TUBB3 is not exclusively neuron-specific. It is, albeit weakly,expressed in testicular and a few other tissue types (Burgoyneet al., 1988; Ludueña, 1998). Recently, it was shown to be presentin cultures of mesenchymal and epithelial cells (Jouhilahti et al.,2008). A variety of solid tumors such as ovarian tumors, lungtumors and malignant melanomas also express TUBB3 and it hasbeen suggested that this tubulin isotype is associated with cellulardrug resistance (Mariani et al., 2011). Tubb3 has also been foundexpressed in developing melanoblasts in mouse and chickenembryo's (Adameyko et al., 2012, 2009). Interestingly, in a recentstudy positive immunoreactivity for the TUBB3 protein wasreported in cells in the basal layer of normal human adultepidermis (K. Akasaka et al., 2009). Although it is suggestive thatthese cells could be melanocytes, no confirmation has been
Contents lists available at SciVerse ScienceDirect
journal homepage: www.elsevier.com/locate/diff
Differentiation
0301-4681/$ - see front matter & 2013 International Society of Differentiation. Published by Elsevier B.V. All rights reserved.Join the International Society for Differentiation (www.isdifferentiation.org)http://dx.doi.org/10.1016/j.diff.2013.05.003
Abbreviations: TUBB3, class III beta-tubulin; DCT, dopachrome tautomerase;TYRP1, tyrosinase-related protein 1; MITF, microphthalmia-associated transcrip-tion factor; α-MSH, alpha-melanocyte-stimulating hormone
n Corresponding author. Tel.: +31 71 526 1179; fax: +31 71 524 8201.E-mail addresses: [email protected] (H. Locher),
[email protected] (K.E. de Rooij), [email protected] (J.C.M.J. de Groot),[email protected] (R.van Doorn), [email protected] (N.A. Gruis), [email protected] (C.W.G.M. Löwik),[email protected] (S.M. Chuva de Sousa Lopes), [email protected] (J.H.M. Frijns), [email protected] (M.A. Huisman).
Differentiation 85 (2013) 173–181

provided. This is of importance as an identical epidermal immu-nostaining pattern can be found in a study on human epidermalinnervation (Lauria et al., 2004). A potential TUBB3 expression bymelanocytes could therefore mislead neuronal cell identificationwhen using antibodies directed against TUBB3.
In the present study, we have resolved this issue by showingthat TUBB3 is expressed in human epidermal and hair folliclemelanocytes, both in vivo and in vitro, and therefore have identi-fied TUBB3 as a novel marker of melanocytes. In addition, we haveexplored its relation to melanocyte differentiation and melano-genesis. Because melanocytes do express TUBB3, this has majorimplications in interpreting results of cultured skin and skin stemcells, as well as in studies on skin innervation.
2. Materials and methods
2.1. Tissue samples
Human adult retroauricular tissue was obtained during surgeryon a 70-year-old female patient. Fetal scalp skin was collected fromtissue of 20 weeks gestational age obtained by elective abortion.Both cases were approved by the Medical Ethical Committee of theLeiden University Medical Center (fetal tissue: P08.087) andinformed consent was obtained in accordance with the Declarationof Helsinki guidelines. Skin specimens were fixed in 4% formalde-hyde in PBS overnight at 4 1C followed by ethanol dehydration andembedding in paraffin wax using standard procedures.
2.2. Cell culture
Fetal scalp skin was mechanically dissociated and cultured forseven days in RPMI 1640 (Life Technologies, Carlsbad, CA, USA)containing 10% fetal bovine serum (FBS) GOLD (PAA LaboratoriesGmbH, Pasching, Austria), 200 nM 12-O-tetradecanoylphorbol 13-acetate (TPA, Sigma-Aldrich, Saint Louis, MO, USA), 200 pM cho-lera toxin (CT, Sigma-Aldrich), 10 ng/ml stem cell factor (SCF, R&DSystems, Inc., MN, USA), GlutaMAX (Life Technologies) and anti-biotic antimycotic solution (Sigma-Aldrich). Cultures were main-tained in a humidified incubator with 5% CO2 at 37 1C. Mediumwas changed every third day.
Melb-a cells were cultured in RPMI with 10% FBS, 20 ng/ml SCF,10 ng/ml bFGF (R&D Systems, Inc.), GlutaMAX and antibioticantimycotic solution. Melan-Ink4a2 cells were cultured in RPMIcontaining 10% FBS, 200 nM TPA, 200 pM CT, 10 ng/ml SCF,GlutaMAX and antibiotic antimycotic solution. Both cell lines weremaintained in a humidified incubator with 10% CO2 at 37 1C.Medium was changed every other day, and cells were passagedusing TrypLE Express (Life Technologies) upon reaching 90%confluency.
Primary normal human melanocytes were cultured in Ham'sF10 supplemented with 100 U/ml penicillin, 100 U/ml streptomy-cin and 292 mg/ml L-glutamine (Invitrogen, Breda, the Nether-lands), 1% Ultroser G (Biosepra, Fremont, CA, USA), 5 ng/mlendothelin-1 (Sigma-Aldrich), 5 ng/ml basic-FGF (Sigma-Aldrich),30 mg/ml CT, 33 mM IBMX (Sigma-Aldrich) and 8 nM TPA. Melano-cytes were cultured in a humidified incubator with 7.5% CO2 at37 1C. Medium was changed every other day, and cells werepassaged using 0.01% Trypsin (Life Technologies) upon reaching90% confluency. Melanocytes were stimulated with 109 U alpha-MSH (Sigma-Aldrich) for 6, 24 and 48 h.
2.3. Immunochemistry and image processing
Paraffin sections (5 μm) of skin specimen were deparaffinizedin xylene, rehydrated in a descending ethanol series (100%, 90%,
80%, 70%), and rinsed in distilled water. Antigen unmasking wasperformed in 0.01 M sodium citrate buffer (pH 6.0) for 12 min at97 1C using a microwave oven. Sections were blocked with 1%bovine serum albumin (BSA, Sigma-Aldrich) in PBS containing0.05% Tween-20 (Promega, Madison, WI) for 30 min, and incu-bated with primary antibodies (appropriately diluted in BSA/Tween-20/PBS) overnight at room temperature in a humidifiedchamber. Secondary antibodies were diluted in 1% normal goatserum (NGS, DAKO, Glostrup, Denmark) in Tween-20/PBS, andsections were incubated for 1 hour at room temperature. Nucleiwere stained with 4',6-diamidino-2-phenylindole (DAPI) and sec-tions were mounted in ProLong Gold (Molecular Probes, LifeTechnologies). Cultures were fixed in 1% formaldehyde in PBSovernight at 4 1C and, when necessary, stored in PBS. Bleachingwas performed in 2% potassium permanganate solution for 30 minfollowed by 0.5% oxalic acid solution for 10 min. Cells werepermeabilized with ice-cold methanol for 10 min, blocked in 5%NGS and 0.05% Tween-20 in PBS, and incubated with primaryantibodies (appropriately diluted in NGS/Tween-20/PBS) overnightat 4 1C. Next, the cells were incubated with fluorochrome-conjugated secondary antibodies (diluted in NGS/Tween-20/PBS)for 1 h at room temperature and followed by nuclear staining withDAPI added to Vectashield mounting medium (Vector Labora-tories, Burlingame, CA). Primary and secondary antibodies, andtheir dilutions, are listed in Table S1. In all double stainings withmelanocytic markers, monoclonal anti-TUBB3 2G10 was used. Dueto technical limitations, DCT was used to identify the pigmentedmelanocytes in deparaffinized sections whereas TYRP1 was usedto identify these cells in culture. These enzymes catalyze subse-quent steps in melanin biosysthesis. Human dermal fibroblastcultures (HDF-a, ScienCell, Carlsbad, CA, USA) were used as anegative tissue control whereas in specificity controls the primaryantibody was omitted. Brightfield, phase contrast and fluorescentimages were taken with either a DM5500 microscope and camera(Leica Microsystems, Rijswijk, the Netherlands) or an OlympusIX70 microscope (Olympus, Zoeterwoude, the Netherlands)equipped with a Leica DFC340 FX camera (Leica) using LAS AFsoftware (Leica). Brightness and contrast adjustments and cellcounting was performed using ImageJ 1.45s (National Institutes ofHealth, http://imagej.nih.gov/ij).
2.4. Western blot analysis
For Western blotting, p34 melb-a and p19 melan-Ink4a2 cellswere lysed in ice cold lysis buffer (50 mM Tris–HCl (pH 7.5),100 mM NaCl, 0.2% Tween-20, supplemented with 1 mini proteaseinhibitor cocktail tablet (Roche, Basel, Switzerland) per 10 ml).Human melanocytes were harvested for Western analysis in lysisbuffer M-per (Thermo Scientific, Rockford, IL), supplemented with1 tablet PhosSTOP and 1 tablet protease inhibitor (Roche) per10 ml. Protein concentration was determined using a Pierce BCAProtein assay kit (Thermo Scientific). Equal amounts of proteinwere loaded on a 10% SDS-PAGE gel, separated for 90 min at 100 Vand blotted onto polyvinylidene difluoride membranes (Immobi-lon-P, Millipore, Bedford, MA) for 90 min at 300 mA. Membraneswere blocked with 3% non-fat dry milk (Elk skimmed instant milkpowder; FrieslandCampina, the Netherlands) in TBS containing0.01% Tween-20 (TBST, pH 7.5) for 1 h at room temperaturefollowed by incubation with the primary antibodies TUJ1(1:2000, Covance, Princeton, NJ), 2G10 (1:400, Abcam, Cambridge,UK) and DCT (1:1000, Abcam) overnight at 4 1C. GAPDH (1:2000,Santa Cruz Biotechnology Inc, Santa Cruz, CA) and β-actin (1:5000,Sigma-Aldrich, St. Louis, MO) were used as loading controls.Subsequently, the membranes were inubated with horseradishperoxidase-conjugated secondary antibodies (Promega) for 1 h atroom temperature in TBST. Protein was detected with SuperSignal
H. Locher et al. / Differentiation 85 (2013) 173–181174

West Pico Chemiluminescent Substrate (Thermo Scientific) using aGel Doc XR System (Bio-Rad, Hercules, CA). Quantity One 1-DAnalysis software (Bio-Rad) was used to determine band intensityfor analysis of protein expression levels in the human melanocytes.
2.5. qRT-PCR analysis
RNA from p34 melb-a and p19 melan-Ink4a2 cell cultures wasisolated using RNA-Bee (Tel-Test Inc., Friendswood, TX) accordingto the manufacturer's instructions. Primers against Mitf, Dct andTubb3 were designed with Primer-BLAST (http://www.ncbi.nlm.nih.gov/tools/primer-blast). A complete list of the primers usedcan be found in Table S2. cDNA was generated by incubating 1 mgRNA with 500 ng Random Hexamers (Promega), supplementedwith H2O to 10 ml, for 10 min at 70 1C. Per sample, 100 unitsM-MLV reverse transcriptase, 1.25 ml 10 mM dNTP mix, 20 unitsRNAsin ribonuclease inhibitor and 1! buffer (all from Promega)were mixed, incubated at room temperature for 10 min, andheated for 50 minutes at 45 1C and 10 min at 70 1C. Real-timePCR was performed using the IQ5 Real Time PCR Detection System(Bio-Rad) with an initial denaturation step of 15 min at 95 1Cfollowed by 40 cycles of 15 s at 95 1C, 30 s at 60 1C and 30 s at72 1C. Cycle threshold values were attained and relative expressionlevels of mRNA were normalized to the housekeeping genes Hprtand Gapdh and calculated by the ΔΔCT method. qRT-PCRs wereperformed on two independent samples of each cell line and threetechnical replicas were performed for each sample.
2.6. Statistical analysis
Results are expressed as the mean7standard deviation. Two-tailed, paired Student's t-tests were used to determine statisticalsignificance. A value of po0.05 was considered to be statisticallysignificant. Analysis was performed using Microsoft Excel 2010(Microsoft, Redmond, WA).
3. Results
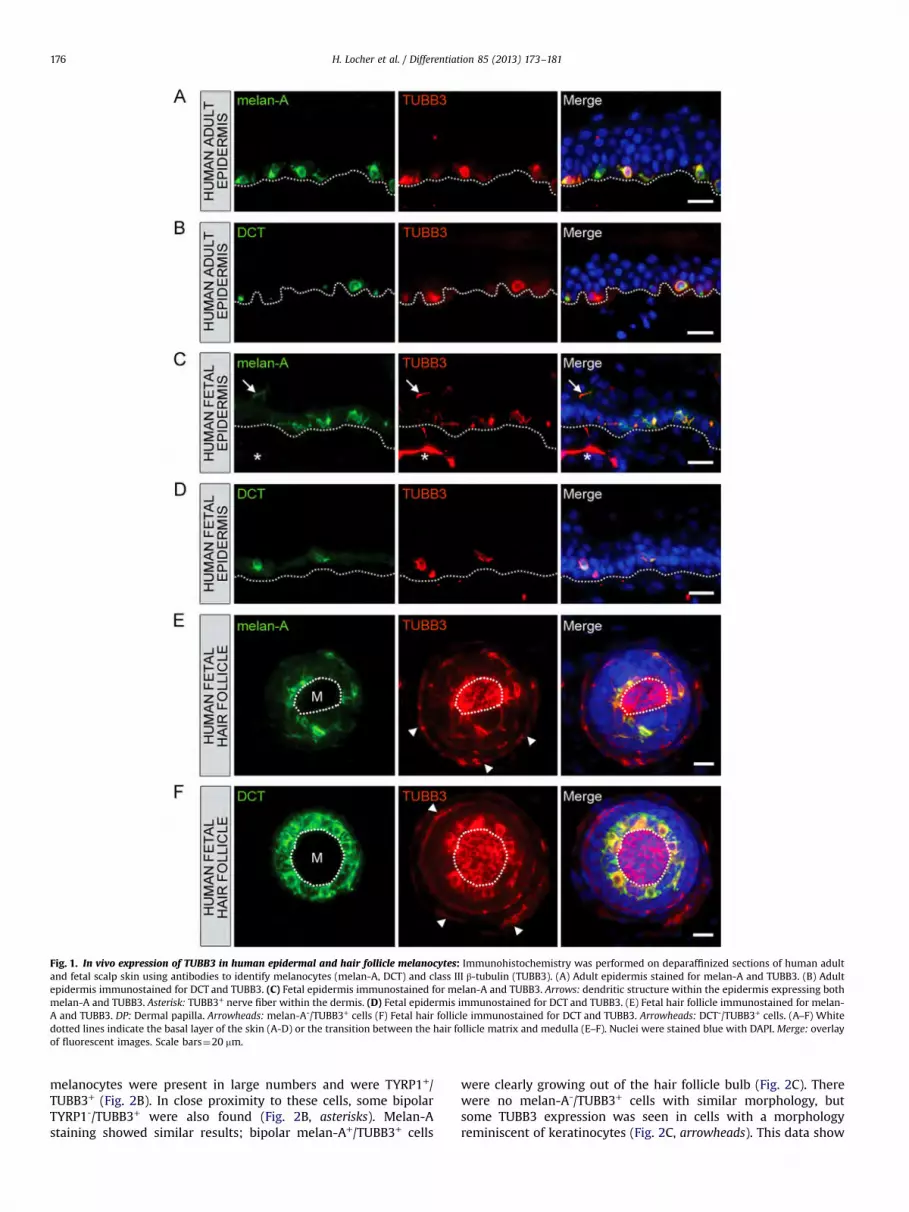
3.1. TUBB3 is expressed in human epidermal melanocytes in vivo
To confirm the TUBB3 expression reported by Akasaka et al.(2009) in the basal layer of human epidermis and to investigate ifthis was indeed localized to melanocytes, we analyzed humanepidermal skin sections by immunohistochemistry for both TUBB3and two melanocytic markers, melanoma-specific antigen 1(melan-A) and dopachrome tautomerase (DCT). Melan-A (alsoknown as MART-1) is a protein present both in non-pigmenedand in pigmented melanocytes. DCT (also known as tyrosinase-related protein 2, TYRP2) is an enzyme involved in melanogenesisand is a direct target of microphthalmia-associated transcriptionfactor (MITF) and therefore specifically marks the early andterminally differentiated pigment producing melanocytes. In adulthuman epidermis, most melan-A+ and DCT+ cells in the basal layerof the epidermis immunostained for TUBB3 (Fig. 1A and B). TUBB3expression was seen in 83.7%75.4 (n"49) of the melan-A+ cellsand in all of the DCT+ cells (n"21). There were also TUBB3+ cells inthe basal layer that did not immunostain for melan-A (6.8%75.0,n"44) nor for DCT (43.2%719.3, n"37). Fetal scalp skin at 20weeks of gestational age showed a similar staining pattern.Already, melan-A+/TUBB3+ and DCT+/TUBB3+ melanocyteswere found in the basal layer (Fig. 1C and D) and the majority ofmelan-A+ and DCT+ cells expressed TUBB3 (98.0%73.0 (n"50)and 100.0% (n"27), respectively). Of the TUBB3+ cells, 3.9%73.4(n"51) did not immunostain for melan-A, whereas 42.6%75.5(n"47) were negative for DCT. Taken together, these data show
that normal human epidermal non-pigmented and terminallydifferentiated pigmented melanocytes express TUBB3 in vivo.
We also observed TUBB3+ dendritic-like structures in theepidermis (Fig. 1C, arrow) which we assumed to be epidermalnerve endings and therefore be of neuronal origin. Interestingly,many of these structures were also melan-A+ (Fig. 1C, arrow). Sincethe dermal nerve fibers did not stain for melan-A (Fig. 1C, asterisk),we conclude that these intraepidermal melan-A+/TUBB3+ struc-tures are melanocytic dendrites rather than free nerve endings.
3.2. Human hair follicle melanocytes express TUBB3 in vivo
Our observations in epidermal melanocytes raised the questionof whether hair follicle melanocytes also expressed TUBB3. In thehair follicle, non-pigmented melanocytes reside within the bulge/sub-bulge area and the outer root sheet of the infundibulum(Botchkareva et al., 2001; Nishimura, 2011; Tobin et al., 1999).They are thought to be the source of the mature melanocyteslocated around the dermal papilla and lower portion of themedulla, which provide pigment to the hair shaft keratinocytes.In fetal scalp hair follicles, melan-A+ and DCT+ melanocytes wereseen in the hair matrix surrounding the basal area of the medulla(Fig. 1E and F). As epidermal melanocytes, these hair folliclemelanocytes immunostained for TUBB3 (Fig. 1E and F). In addition,TUBB3 expression was observed in cells of the outer root sheath(Fig. 1E and F, arrowheads). These TUBB3+ cells, however, werenegative for melan-A and DCT and it could be surmised that thesecells are non-pigmented melanocytes which are known to residein this area. TUBB3 immunostaining was also seen in the hair shaftmedulla keratinocytes (Fig. 1E and F, M). In summary, in thehuman hair follicle melan-A+/TUBB3+ and DCT+/TUBB3+ melano-cytes were observed in the hair matrix, whereas melan-A-/TUBB3+
and DCT-/TUBB3+ cells were found in the outer root sheath of thehair follicle bulb.
3.3. Fetal epidermal and hair follicle melanocytes express TUBB3in vitro
Based on the in vivo expression of TUBB3 in both epidermal andhair follicle melanocytes, we anticipated that TUBB3 would also beexpressed ex vivo/in vitro in these cells. For this purpose, humanfetal scalp skin which contained an abundance of pigmented hairfollicles was mechanically dissociated and cultured for seven daysin standard culture medium containing additional factors (SCF, TPAand CT, see (Materials and methods) to support the whole range ofdifferentiated melanocytes, including the non-pigmented, early-pigmented and the terminally differentiated pigmented melano-cytes. This resulted in heterogeneous cultures containing kerati-nocytes, fibroblasts and melanocytes. The melanocytic cells wereidentified by means of different types of light microscopy. Usingphase-contrast microscopy, melanocytes could be recognized bytheir bipolar or multipolar dendrites and the halo surroundingtheir cell body (Fig. 2). Bright-field microscopy was used to detectpigment inclusions (Fig. 2). Using both approaches, epidermal andhair follicle melanocytes could be distinguished on basis of thecriteria formulated by Tobin et al. (Tobin and Bystryn, 1996) whoshowed that cultured epidermal melanocytes have 3–6 dendrites,whereas cultured hair follicle melanocytes are mostly bipolar.Definite confirmation was provided by immunofluorescent stain-ing for tyrosine-related protein 1 (TYRP1) and melan-A.
As expected, we were able to identify TYRP1+/TUBB3+ pigmen-ted epidermal melanocytes in the dissociated skin cultures(Fig. 2A). Interestingly, TYRP1-/TUBB3+ cells were also observed(Fig. 2A, asterisks). Morphologically, these non- or early-pigmentedmelanocytes were similar to the TYRP1+/TUBB3+ cells, but theywere less dark under bright-field microscopy. Bipolar hair follicle
H. Locher et al. / Differentiation 85 (2013) 173–181 175
melanocytes were present in large numbers and were TYRP1+/TUBB3+ (Fig. 2B). In close proximity to these cells, some bipolarTYRP1-/TUBB3+ were also found (Fig. 2B, asterisks). Melan-Astaining showed similar results; bipolar melan-A+/TUBB3+ cells
were clearly growing out of the hair follicle bulb (Fig. 2C). Therewere no melan-A-/TUBB3+ cells with similar morphology, butsome TUBB3 expression was seen in cells with a morphologyreminiscent of keratinocytes (Fig. 2C, arrowheads). This data show
Fig. 1. In vivo expression of TUBB3 in human epidermal and hair follicle melanocytes: Immunohistochemistry was performed on deparaffinized sections of human adultand fetal scalp skin using antibodies to identify melanocytes (melan-A, DCT) and class III β-tubulin (TUBB3). (A) Adult epidermis stained for melan-A and TUBB3. (B) Adultepidermis immunostained for DCT and TUBB3. (C) Fetal epidermis immunostained for melan-A and TUBB3. Arrows: dendritic structure within the epidermis expressing bothmelan-A and TUBB3. Asterisk: TUBB3+ nerve fiber within the dermis. (D) Fetal epidermis immunostained for DCT and TUBB3. (E) Fetal hair follicle immunostained for melan-A and TUBB3. DP: Dermal papilla. Arrowheads: melan-A-/TUBB3+ cells (F) Fetal hair follicle immunostained for DCT and TUBB3. Arrowheads: DCT-/TUBB3+ cells. (A–F) Whitedotted lines indicate the basal layer of the skin (A-D) or the transition between the hair follicle matrix and medulla (E–F). Nuclei were stained blue with DAPI. Merge: overlayof fluorescent images. Scale bars"20 mm.
H. Locher et al. / Differentiation 85 (2013) 173–181176

that, as in vivo, both human epidermal and hair follicle melano-cytes express TUBB3 in vitro and that expression starts beforeinitiation of melanogenesis.
3.4. Expression of TUBB3 starts upon differentiation of melanoblastsinto mature melanocytes
In neurons of the central nervous system TUBB3 expressiondecreases as the cells mature and can therefore be used to identifyyoung developing neurons. In the peripheral nervous system, how-ever, the opposite is found: TUBB3 levels increase with neuronal cellmaturation in dorsal root ganglia (Jiang and Oblinger, 1992). Ourexperiments with human melanocytes show that TUBB3 is expressedin the end stages of the melanocytic lineage (non-pigmented andpigmented melanocytes). However, expression in the undifferen-tiated, melanoblast stage remains inconclusive. To further examine
this relationship of TUBB3 expressionwithin the melanocytic lineage,we analyzed two widely-used immortalized mouse melanocytic celllines derived from the C57BL/6J strain. Melb-a is a clonal line ofmelanoblasts, which do not produce melanin (Sviderskaya et al.,1995). Using bright-field microscopy, cells could not be seen due tothe obvious lack of pigment inclusions (data not shown), confirmingtheir immature state. Although most cells were negative for TUBB3(Fig. 3A), rare TUBB3+ cells could be observed. Interestingly, theseTUBB3+ cells demonstrated a more differentiated cell morphology bythe presence of dendrites and a halo surrounding their cell bodies.TUBB3- cells were generally flat with few and short dendrites, ornone at all (Fig. 3A).
To analyze TUBB3 expression in mature murine melanocytes,we used melan-Ink4a2 cells (Ha et al., 2007). Cells of this lineproduce different amounts of melanin, so both non-pigmentedand pigmented melanocytes are present which can be visualized
Fig. 2. In vitro expression of TUBB3 in human epidermal and hair follicle melanocytes: Immunostaining was performed on cultures of human fetal scalp skin usingantibodies to identify melanocytes (TYRP1 and melan-A) and class III β-tubulin (TUBB3). (A) Cultured epidermal melanocytes immunostained for TYRP1 and TUBB3. Asterisksidentify TYRP1-/TUBB3+ cells. (B) Cultured hair follicle melanocytes immunostained for TYRP1 and TUBB3. Asterisks identify TYRP1-/TUBB3+ cells. (C) Cultured cellsimmunostained for melan-A and TUBB3. White dotted lines outline hair follicle bulb (B). (A–C) Nuclei are stained blue with DAPI. Merge: overlay of fluorescent images. Phasecontrast: phase contrast microscopy. Bright field: bright field microscopy. Scale bars"20 mm.
H. Locher et al. / Differentiation 85 (2013) 173–181 177
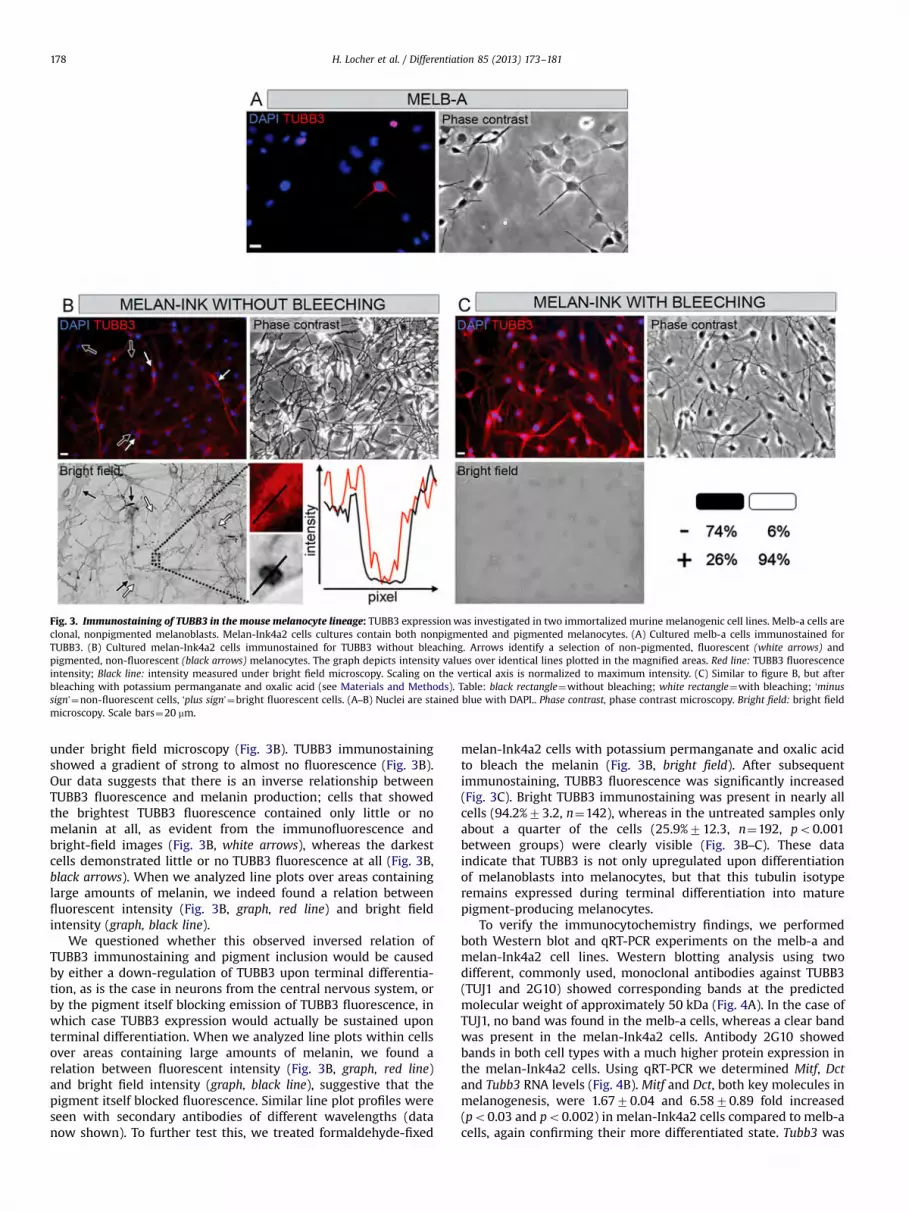
under bright field microscopy (Fig. 3B). TUBB3 immunostainingshowed a gradient of strong to almost no fluorescence (Fig. 3B).Our data suggests that there is an inverse relationship betweenTUBB3 fluorescence and melanin production; cells that showedthe brightest TUBB3 fluorescence contained only little or nomelanin at all, as evident from the immunofluorescence andbright-field images (Fig. 3B, white arrows), whereas the darkestcells demonstrated little or no TUBB3 fluorescence at all (Fig. 3B,black arrows). When we analyzed line plots over areas containinglarge amounts of melanin, we indeed found a relation betweenfluorescent intensity (Fig. 3B, graph, red line) and bright fieldintensity (graph, black line).
We questioned whether this observed inversed relation ofTUBB3 immunostaining and pigment inclusion would be causedby either a down-regulation of TUBB3 upon terminal differentia-tion, as is the case in neurons from the central nervous system, orby the pigment itself blocking emission of TUBB3 fluorescence, inwhich case TUBB3 expression would actually be sustained uponterminal differentiation. When we analyzed line plots within cellsover areas containing large amounts of melanin, we found arelation between fluorescent intensity (Fig. 3B, graph, red line)and bright field intensity (graph, black line), suggestive that thepigment itself blocked fluorescence. Similar line plot profiles wereseen with secondary antibodies of different wavelengths (datanow shown). To further test this, we treated formaldehyde-fixed
melan-Ink4a2 cells with potassium permanganate and oxalic acidto bleach the melanin (Fig. 3B, bright field). After subsequentimmunostaining, TUBB3 fluorescence was significantly increased(Fig. 3C). Bright TUBB3 immunostaining was present in nearly allcells (94.2%73.2, n"142), whereas in the untreated samples onlyabout a quarter of the cells (25.9%712.3, n"192, po0.001between groups) were clearly visible (Fig. 3B–C). These dataindicate that TUBB3 is not only upregulated upon differentiationof melanoblasts into melanocytes, but that this tubulin isotyperemains expressed during terminal differentiation into maturepigment-producing melanocytes.
To verify the immunocytochemistry findings, we performedboth Western blot and qRT-PCR experiments on the melb-a andmelan-Ink4a2 cell lines. Western blotting analysis using twodifferent, commonly used, monoclonal antibodies against TUBB3(TUJ1 and 2G10) showed corresponding bands at the predictedmolecular weight of approximately 50 kDa (Fig. 4A). In the case ofTUJ1, no band was found in the melb-a cells, whereas a clear bandwas present in the melan-Ink4a2 cells. Antibody 2G10 showedbands in both cell types with a much higher protein expression inthe melan-Ink4a2 cells. Using qRT-PCR we determined Mitf, Dctand Tubb3 RNA levels (Fig. 4B). Mitf and Dct, both key molecules inmelanogenesis, were 1.6770.04 and 6.5870.89 fold increased(po0.03 and po0.002) in melan-Ink4a2 cells compared to melb-acells, again confirming their more differentiated state. Tubb3 was
Fig. 3. Immunostaining of TUBB3 in the mouse melanocyte lineage: TUBB3 expressionwas investigated in two immortalized murine melanogenic cell lines. Melb-a cells areclonal, nonpigmented melanoblasts. Melan-Ink4a2 cells cultures contain both nonpigmented and pigmented melanocytes. (A) Cultured melb-a cells immunostained forTUBB3. (B) Cultured melan-Ink4a2 cells immunostained for TUBB3 without bleaching. Arrows identify a selection of non-pigmented, fluorescent (white arrows) andpigmented, non-fluorescent (black arrows) melanocytes. The graph depicts intensity values over identical lines plotted in the magnified areas. Red line: TUBB3 fluorescenceintensity; Black line: intensity measured under bright field microscopy. Scaling on the vertical axis is normalized to maximum intensity. (C) Similar to figure B, but afterbleaching with potassium permanganate and oxalic acid (see Materials and Methods). Table: black rectangle"without bleaching; white rectangle"with bleaching; ‘minussign’"non-fluorescent cells, ‘plus sign’"bright fluorescent cells. (A–B) Nuclei are stained blue with DAPI.. Phase contrast, phase contrast microscopy. Bright field: bright fieldmicroscopy. Scale bars"20 mm.
H. Locher et al. / Differentiation 85 (2013) 173–181178

also found in both cell lines, with a 4.1270.32-fold higherexpression level in the melan-Ink4a2 cells (po0.02). In summary,immunocytochemistry, Western blot and qRT-PCR data indicatedthat TUBB3 expression starts when melanoblasts commit to themelanocyte lineage and differentiate into pigment producingmelanocytes.
3.5. !-MSH stimulation does not increase TUBB3 expressionin cultured human melanocytes
As TUBB3 expression in the melanocytic lineage coincides withthe induction of melanin, we questioned if the melanogenesispathway could also regulate TUBB3 expression. To assess this, wecultured primary human melanocytes and stimulated the cellswith alpha-melanocyte-stimulating hormone (α-MSH) for 6 h,24 h or 48 h. After exposure, we performed quantitative westernblot analysis for DCT and TUBB3 expression on the cell lysates. Asα-MSH binds to the melanocortin 1 receptor (MC1R), which in turnpromotes transcription of the MITF transcription factor, anobserved increase in expression of the MITF target gene DCT
would indicate a successful stimulation of the melanogenesispathway. DCT expression was found to be increased (232%, 175%and 166% after 6 h, 24 h and 48 h, respectively) confirmingsuccessful stimulation (Fig. 5). In contrast, TUBB3 showed noconcomitant increase in expression (108%, 105% and 91%, respec-tively (Fig. 5)). Combined, no association between TUBB3 expres-sion and induction of melanogenesis could be found.
4. Discussion
In this study, we show that TUBB3 is not only expressed withinperipheral neurons but also in the melanocytic lineage. In mela-nocytes, TUBB3 expression was found both in human epidermaland hair follicle melanocytes. Using skin sections, we showed thatnearly all melan-A+ and DCT+ melanocytes express TUBB3. Con-versely, TUBB3+ cells in the basal layer of the epidermis werealmost always melan-A+, although many did not express DCT.Similar results were found in cultured human fetal epidermal andhair follicle melanocytes. In the melanocytic lineage, melan-A isexpressed both by non-pigmented and pigmented melanocytes.However, DCT and TYRP1 are not expressed before terminaldifferentiation into pigment producing melanocytes. This indicatesthat TUBB3 is widely expressed within the melanocyte lineage,with expression starting before pigment production. Indeed, whenwe investigated TUBB3 expression in mouse melanoblasts andmelanocytes, most melanoblasts did not (as yet) express TUBB3whereas both non-pigmented and pigmented melanocytes didexpress TUBB3. This implicates that expression of TUBB3 startsaround the point of time where melanoblasts commit to themelanocyte lineage (Fig. 5).
Like peripheral neurons, melanocytes are derived from theneural crest. There are also other similarities, both in morphologyand functional characteristics. Both cell types have small, roundcell bodies and multiple dendritic processes. Both can expresssimilar receptors (e.g. p75NTR and c-Kit) and therefore respond toidentical cytokines and neurotrophins (Yaar and Park, 2012).Furthermore, both melanins (produced by melanocytes) andcatecholamines (e.g. dopamine, produced by neurons) are derivedfrom tyrosine, and both melanosomes and synaptic vesicle
Fig. 4. Western blot and quantitative RT-PCR analysis of TUBB3 expression in themelanocyte lineage: (A) Representative Western blots of TUBB3 expression inmelb-a and melan-Ink4a2 cells using two different monoclonal antibodies, TUJ1and 2G10. Both antibodies recognize the C-terminal end of the protein. TUBB3 has amolecular weight of #50 kDa. GAPDH was used as loading control. (B) QuantitativeRT-PCR analysis of melb-a and melan-Ink4a2 cell lysates. The figure shows theexpression levels of RNA from two genes regulating melanogenesis, Mitf and DCT,as well as the RNA levels for TUBB3. All PCR products are normalized againstaverages of Hprt and Gapdh controls, and shown as relative expression levelsagainst melb-a RNA. *po0.05; **po0.01.
Fig. 5. Western blot analysis of DCT and TUBB3 expression after !-MSH stimula-tion in normal human melanocytes: (A) Representative Western blots showingDCT and TUBB3 expression in human melanocytes after stimulation with α-MSH for6, 24 and 48 h. The same blots were incubated with a monoclonal antibody againstβ-actin to correct for equal loading. (B) The intensity of the bands in (A) wasmeasured using Quantity One 1-D analysis and the resulting change in expressionis given in percentage of the unstimulated control, corrected for loading.
H. Locher et al. / Differentiation 85 (2013) 173–181 179

precursors are transported over microtubules by kinesins. In turn,melanosomes and neurotransmitters are both released from theirdendrites or axons. Presently, there is an emerging concept thatmelanocytes could potentially be used to represent neuronal cellsin disease model systems. In this light, the expression of neuron-specific TUBB3 in melanocytes might not be so surprising. In theskin, TUBB3 expression is considered to be of neuronal origin,whether it be intraepidermal free nerve endings or nerves pro-jecting to Merkel cells, Langerhans cells or even melanocytes(Chateau and Misery, 2004; Hara et al., 1996; Hosoi et al., 1993).As we show that at least a part of these endings belong tomelanocytes, it is of great interest to further investigate the extentof TUBB3 expression by skin melanocytes, examine their relation-ship with surrounding cells and investigate expression of otherneuronal biomarkers.
TUBB3 has now been found expressed in chicken, mouse andhuman melanocytes. This evolutionary conservation is suggestiveof an important role of TUBB3 in these cells. We observedinitiation of TUBB3 expression in the melanocytic lineage aroundthe time of differentiation into pigment producing melanocytes.Melanocyte development as well as the UV-induced pigmentationresponse are regulated by the MITF transcription factor thatmodulates multiple differentiation and melanin biosynthesisgenes. We questioned whether TUBB3 expression could be regu-lated by the same pathway and stimulated cultured humanmelanocytes with α-MSH, known to induce MITF expression.Whereas expression of the MITF target gene DCT was induced byα-MSH stimulation, there was no associated increase in TUBB3expression. Therefore, TUBB3 does not seem to be regulated by theMITF-pathway which regulates melanocyte development andpigmentation response. This is in line with results of studiesexamining MITF target genes, which consistently found thatTUBB3 was not among the targets of this transcription (Cheliet al., 2010; Hoek et al., 2008; Levy et al., 2006).
Using a well-established bleaching protocol, we show thatfluorescent signals emitted by TUBB3 stainings are blocked bymelanin. This was in contrast to control experiments with melan-A immunostaining which did not show this clear relationship (Fig.S1), indicating that the melanin specifically interfered with theTUBB3 fluorescence. Therefore, we conclude that the diminishedTUBB3 fluorescence was caused by specific co-localization ofmelanin. This could indicate a novel role of TUBB3 in melanosomaltransport. It is known that mature melanosomes are transportedby kinesin and dynein motor proteins (Marks & Seabra, 2001).These proteins use microtubule tracks to travel in a bi-directionalway (Wu et al., 1998). Also, by specifically disrupting microtubules,it has been shown that melanosome transport is microtubuledependent (Wu et al., 1998). Potentially, TUBB3 could be thecorresponding β-unit isotype (Fig. S2). However, its role in cellularprocesses remains to be investigated.
A recent study by Dalziel et al. (2011) showed that TUBB3 isphysically located immediately downstream of MC1R (Fig. S3).This receptor binds to melanocortins and plays an important rolein melanogenesis through its ability to influence MITF transcrip-tion. Aside from showing that TUBB3 is located immediatelydownstream of MC1R, in itself suggesting a role within melano-cytes, Dalziel and colleagues found that chimeric proteins aretranscribed by intergenic splicing between these two genes. Themonoclonal TUBB3 antibodies used in our study, TUJ1 and 2G10,both recognize the C-terminus of the TUBB3 protein (epitopesEAQGPK (Lee et al., 1990) and MYEDDEEESEAQGPK (Spano andFrankfurter, 2010)) and immunostain the same structures (Fig. S4).In chick and mice, Tubb3 has only one protein product. In humans,aside from the chimeric proteins, there are four additional TUBB3isoforms (Fig. S3). Both antibodies will recognize at least isoforms1 and 2 and the chimeric transcript RP11-566K11.2-001. Currently,
these antibodies react both with neuronal and melanocytic cells.However, it could be that both cell types express different isoformsof the TUBB3 protein. It will be interesting to investigate the exactexpression of these isoforms in normal human neurons andmelanocytes to find out whether different and/or identical iso-forms are present in these cell types. Together, our results andthese findings suggest a dynamic role for TUBB3 in multipleaspects of melanocytic function.
Finally, in addition to TUBB3 expression in melanocytes, wehave also observed its presence in hair follicle medulla keratino-cytes in vivo. Expression of TUBB3 in the mitotic spindle ofcultured keratinocytes and fibroblasts has been previouslyreported (Jouhilahti et al., 2008), something we have observed aswell in our cultured melanocytes (Fig. S5). This indicates that thisbiomarker, especially in the skin, cannot be reliably used todifferentiate between neuronal cells and other neural crest andectodermal cell types. Primary skin cultures and differentiationstudies on skin stem cells or even (induced) pluripotent stem cellsmight be prone to this identification issue. Aside from identifyingneurons, TUBB3 can be used as a much wider marker of neuralcrest derived and epithelial cells.
Conflict of Interest
The authors state no conflict of interest.
Acknowledgments
We thank L. van Iperen for technical support and Prof. dr. C.L.Mummery for critical reading of the manuscript. Cell lines melb-a(Sviderskaya et al., 1995) and melan-Ink4a2 (Ha et al., 2007) werekindly provided by the Wellcome Trust Functional Genomics CellBank (St George's, University of London, UK). This research wassupported by the Heinsius Houbolt Fund, the Netherlands Orga-nization for Scientific Research (NWO Aspasia 015.007.037) andthe Netherlands Institute for Regenerative Medicine (NIRM,FES0908).
Appendix A. Supporting information
Supplementary data associated with this article can be found inthe online version at http://dx.doi.org/10.1016/j.diff.2013.05.003.
References
Adameyko, I., Lallemend, F., Aquino, J.B., Pereira, J.A., Topilko, P., Müller, T., Fritz, N.,Beljajeva, A., Mochii, M., Liste, I., Usoskin, D., Suter, U., Birchmeier, C., Ernfors, P.,2009. Schwann cell precursors from nerve innervation are a cellular origin ofmelanocytes in skin. Cell 139, 366–379.
Adameyko, I., Lallemend, F., Furlan, A., Zinin, N., Aranda, S., Kitambi, S.S., Blanchart,A., Favaro, R., Nicolis, S., Lübke, M., Müller, T., Birchmeier, C., Suter, U., Zaitoun,I., Takahashi, Y., Ernfors, P., 2012. Sox2 and Mitf cross-regulatory interactionsconsolidate progenitor and melanocyte lineages in the cranial neural crest.Development 139, 397–410.
Akasaka, K., Maesawa, C., Shibazaki, M., Maeda, F., Takahashi, K., Akasaka, T.,Masuda, T., 2009. Loss of class III beta-tubulin induced by histone deacetylationis associated with chemosensitivity to paclitaxel in malignant melanoma cells.J. Invest. Dermatol. 129, 1516–1526.
Botchkareva, N.V, Khlgatian, M., Longley, B.J., Botchkarev, V.A., Gilchrest, B.A., 2001.SCF/c-kit signaling is required for cyclic regeneration of the hair pigmentationunit. FASEB J.: Off. Publ. Fed. Am. Soc. Exp. Biol. 15, 645–658.
Bryan, J., Wilson, L., 1971. Are cytoplasmic microtubules heteropolymers? Proc.Natl. Acad. Sci. USA 68, 1762–1766.
Burgoyne, R.D., Cambray-Deakin, M.A., Lewis, S.A., Sarkar, S., Cowan, N.J., 1988.Differential distribution of beta-tubulin isotypes in cerebellum. EMBO J. 7,2311–2319.
Chateau, Y., Misery, L, 2004. Connections between nerve endings and epidermalcells: are they synapses? Exp. Dermatol. 13, 2–4.
H. Locher et al. / Differentiation 85 (2013) 173–181180

Cheli, Y., Ohanna, M., Ballotti, R., Bertolotto, C., 2010. Fifteen-year quest formicrophthalmia-associated transcription factor target genes. Pigm. Cell Mela-noma Res. 23, 27–40.
Dalziel, M., Kolesnichenko, M., Pires das Neves, R., Iborra, F., Goding, C., Furger, A.,2011. Alpha-MSH regulates intergenic splicing of MC1R and TUBB3 in humanmelanocytes. Nucleic Acids Res. 39, 2378–2392.
Downing, K.H., Nogales, E., 1998. Tubulin and microtubule structure. Curr. Opin.Cell. Biol. 10, 16–22.
Geisert, E.E., Frankfurter, A., 1989. The neuronal response to injury as visualizedby immunostaining of class III beta-tubulin in the rat. Neurosci. Lett. 102,137–141.
Ha, L., Ichikawa, T., Anver, M., Dickins, R., Lowe, S., Sharpless, N.E., Krimpenfort, P.,Depinho, R.A., Bennett, D.C., Sviderskaya, E.V, Merlino, G., 2007. ARF functionsas a melanoma tumor suppressor by inducing p53-independent senescence.Proc. Natl. Acad. Sci. USA 104, 10968–10973.
Hara, M., Toyoda, M., Yaar, M., Bhawan, J., Avila, E.M., Penner, I.R., Gilchrest, B.A.,1996. Innervation of melanocytes in human skin. J. Exp. Med. 184, 1385–1395.
Hoek, K.S., Schlegel, N.C., Eichhoff, O.M., Widmer, D.S., Praetorius, C., Einarsson, S.O.,Valgeirsdottir, S., Bergsteinsdottir, K., Schepsky, A., Dummer, R., Steingrimsson,E., 2008. Novel MITF targets identified using a two-step DNA microarraystrategy. Pigm. Cell Melanoma Res. 21, 665–676.
Hosoi, J., Murphy, G.F., Egan, C.L., Lerner, E.A., Grabbe, S., Asahina, A., Granstein, R.D.,1993. Regulation of Langerhans cell function by nerves containing calcitoningene-related peptide. Nature 363, 159–163.
Jiang, Y.Q., Oblinger, M.M., 1992. Differential regulation of beta III and other tubulingenes during peripheral and central neuron development. J. Cell Sci. 103 (Pt 3),643–651.
Jouhilahti, E.-M., Peltonen, S., Peltonen, J., 2008. Class III beta-tubulin is acomponent of the mitotic spindle in multiple cell types. J. Histochem.Cytochem. 56, 1113–1119.
Lauria, G., Borgna, M., Morbin, M., Lombardi, R., Mazzoleni, G., Sghirlanzoni, A.,Pareyson, D., 2004. Tubule and neurofilament immunoreactivity in humanhairy skin: markers for intraepidermal nerve fibers. Muscle Nerve 30, 310–316.
Lee, M.K., Tuttle, J.B., Rebhun, L.I., Cleveland, D.W., Frankfurter, A., 1990. Theexpression and posttranslational modification of a neuron-specific beta-tubulinisotype during chick embryogenesis. Cell. Motil. Cytoskeleton 17, 118–132.
Levy, C., Khaled, M., Fisher, D.E., 2006. MITF: master regulator of melanocytedevelopment and melanoma oncogene. Trends Mol. Med. 12, 406–414.
Ludueña, R.F., 1998. Multiple forms of tubulin: different gene products and covalentmodifications. Int. Rev. Cytol. 178, 207–275.
Mariani, M., Shahabi, S., Sieber, S., Scambia, G., Ferlini, C., 2011. Class III β-tubulin(TUBB3): more than a biomarker in solid tumors? Curr. Mol. Med. 11, 726–731.
Marks, M.S., Seabra, M.C., 2001. The melanosome: membrane dynamics in blackand white. Nat. Rev. Mol. Cell. Biol. 2, 738–748.
Moody, S.A., Quigg, M.S., Frankfurter, A., 1989. Development of the peripheraltrigeminal system in the chick revealed by an isotype-specific anti-beta-tubulinmonoclonal antibody. J. Comp. Neurol. 279, 567–580.
Nishimura, E.K., 2011. Melanocyte stem cells: a melanocyte reservoir in hairfollicles for hair and skin pigmentation. Pigm. Cell Melanoma Res. 24, 401–410.
Spano, A.J., Frankfurter, A., 2010. Characterization of anti-beta-tubulin antibodies.Methods Cell Biol. 95, 33–46.
Sullivan, K.F., Cleveland, D.W., 1986. Identification of conserved isotype-definingvariable region sequences for four vertebrate beta tubulin polypeptide classes.Proc. Natl. Acad. Sci. USA 83, 4327–4331.
Sviderskaya, E.V, Wakeling, W.F., Bennett, D.C., 1995. A cloned, immortal line ofmurine melanoblasts inducible to differentiate to melanocytes. Development121, 1547–1557.
Tobin, D.J., Bystryn, J.C., 1996. Different populations of melanocytes are present inhair follicles and epidermis. Pigm. Cell Res. 9, 304–310.
Tobin, D.J., Slominski, A., Botchkarev, Vladimir, A., Paus, R., 1999. The fate of hairfollicle melanocytes during the hair growth cycle. J. Invest. Dermatol. Symp.Proc. 4, 323–332.
Wu, X., Bowers, B., Rao, K., Wei, Q., Hammer, JA, 1998. Visualization of melanosomedynamics within wild-type and dilute melanocytes suggests a paradigm formyosin V function In vivo. J. Cell Biol. 143, 1899–1918.
Yaar, M., Park, H.-Y., 2012. Melanocytes: a window into the nervous system. J.Invest. Dermatol. 132, 835–845.
H. Locher et al. / Differentiation 85 (2013) 173–181 181





























