Embed Size (px)
Citation preview

Characterization of the myostatin gene and a
linked microsatellite marker in shi drum
(Umbrina cirrosa, Sciaenidae)
L. Maccatrozzo a, L. Bargelloni b, P. Patarnello c, G. Radaelli a,F. Mascarello a, T. Patarnello a,b,*
aFacolta di Medicina Veterinaria-Agripolis, Universita di Padova, Via Romea 16 I-35020 Legnaro, ItalybDipartimento di Biologia, Universita di Padova, Via Ugo Bassi 58/B, I-35121 Padua, ItalycDipartimento di Scienze della Produzione Animale, Universita di Udine, Via S. Mauro 2,
33100 Pagnacco, Italy
Received 21 January 2001; accepted 1 May 2001
Abstract
We report the characterisation of the gene coding for myostatin (or growth differentiation factor
8, GDF8) in the shi drum, Umbrina cirrosa. Because of its relevant role in muscle growth
regulation, GDF8 is an important candidate locus for improvement of animal production.
RT-PCR and RACE protocols were used to obtain the full-length GDF8 cDNA sequence. The
complete cDNA was 2086-bp long, containing an open reading frame of 376 amino acids.
Similarity searches on protein databases give highest values with zebrafish GDF8 (291 identical
residues over 376 compared amino acid positions), and other vertebrate GDF8s. This evidence
suggests that the putative protein isolated is the GDF8 homologue in the shi drum. At the genomic
level, the position and complete sequence was determined for two introns. While intron–exon
boundaries were conserved compared to mammalian GDF8 genes, both introns were considerably
smaller, of 342 and 735 bp, respectively. In the 3Vnoncoding region of the cDNA, a microsatellite
repeat (AC29) was present. This repeat region was examined at the genomic level, and its size
polymorphism assessed in a preliminary way, revealing four distinct alleles. This microsatellite
locus might represent a useful polymorphic marker for studying phenotype/genotype association.
We also examined the pattern of expression of sdGDF8, which is wider in the shi drum
compared to other vertebrates, suggesting possible additional functions of myostatin in the teleost
fish. These results should be taken into account if null mutants for GDF8 in this species are to be
0044-8486/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0044-8486 (01 )00659 -7
* Corresponding author. Dipartimento di Biologia, Universita di Padova, Via Ugo Bassi 58/B, I-35121 Padua,
Italy. Tel.: +39-49-8276218; fax: +39-49-8276209.
E-mail address: [email protected] (T. Patarnello).
www.elsevier.com/locate/aqua-online
Aquaculture 205 (2002) 49–60

produced, as they might have a more severe phenotype than observed in mammals. D 2002
Elsevier Science B.V. All rights reserved.
Keywords: Myostatin; GDF8; Muscle growth; Sciaenids; cDNA; Microsatellite
1. Introduction
Growth-differentiation factor 8 (GDF8) or myostatin (MSTN) is a member of the
transforming growth factor b (TGF-b) superfamily, which includes proteins that mediate
key events in cell growth and development through signal transduction. GDF8 seems to be
a negative regulator of skeletal muscle growth as mice knock-out for GDF8 display
increased muscle mass (McPherron et al., 1997) and cattle with null mutations of GDF8
gene present muscle hypertrophy (McPherron and Lee, 1997; Grobet et al., 1997).
Because of its role in muscle growth, GDF8 is an extremely interesting locus for genetic
improvement of farmed animals. Such potential applications in animal husbandry have
prompted the sequencing of GDF8 cDNA from several vertebrate species (man, baboon,
cattle, pig, sheep, chicken, turkey and zebrafish) (McPherron and Lee, 1997). However,
information on GDF8 expression is confined to a few mammalian species (McPherron et
al., 1997; Gonzalez-Cadavid et al., 1998; Ji et al., 1998; Sharma et al., 1999). Comparative
analysis of expression patterns might be helpful in understanding how function of specific
proteins evolved, and therefore, to which extent results and applications from higher
vertebrate taxa might be transferred to teleost fish.
Here, we report the isolation and characterisation of expression profile of the GDF8
gene in the shi drum, Umbrina cirrosa, a teleost fish belonging to the perciform family
Sciaenidae, which account for about 270 species and 70 genera worldwide. Sciaenids are
generally demersal fishes living in shallow waters on the continental shelf, though a few
marine species enter into brackish waters, while others are restricted to freshwater habitats.
Some sciaenids are important foodfishes, and in the Mediterranean area, at least five
species are exploited in bottom fisheries. In particular, the shi drum, because of its meat
quality and large size, is highly appreciated and commercially valuable. Experiments of
captive breeding have been already carried out, and this species is now a promising new
candidate for extensive marine aquaculture, in response to the increasing need of
diversification of marketed fish products.
The aim of the present work is to contribute to the study of shi drum biology, and in
particular, toward the understanding of mechanisms of muscle growth. This might in
turn lead to important practical applications for improvement of its aquaculture produc-
tion.
2. Materials and methods
Samples were collected from a farmed population, at the experimental fish farm
‘‘Impianto di Acquacoltura di Pellestrina, Veneto Agricoltura’’, Italy. Analysed tissues
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–6050
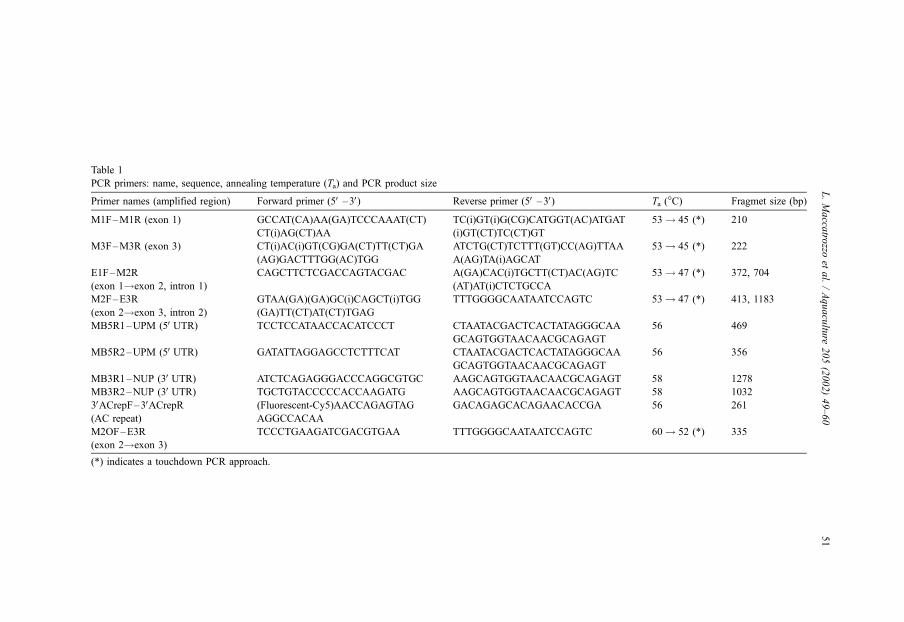
Table 1
PCR primers: name, sequence, annealing temperature (Ta) and PCR product size
Primer names (amplified region) Forward primer (5V–3V) Reverse primer (5V–3V) Ta (�C) Fragmet size (bp)
M1F–M1R (exon 1) GCCAT(CA)AA(GA)TCCCAAAT(CT)
CT(i)AG(CT)AA
TC(i)GT(i)G(CG)CATGGT(AC)ATGAT
(i)GT(CT)TC(CT)GT
53! 45 (*) 210
M3F–M3R (exon 3) CT(i)AC(i)GT(CG)GA(CT)TT(CT)GA
(AG)GACTTTGG(AC)TGG
ATCTG(CT)TCTTT(GT)CC(AG)TTAA
A(AG)TA(i)AGCAT
53! 45 (*) 222
E1F–M2R
(exon 1!exon 2, intron 1)
CAGCTTCTCGACCAGTACGAC A(GA)CAC(i)TGCTT(CT)AC(AG)TC
(AT)AT(i)CTCTGCCA
53! 47 (*) 372, 704
M2F–E3R
(exon 2!exon 3, intron 2)
GTAA(GA)(GA)GC(i)CAGCT(i)TGG
(GA)TT(CT)AT(CT)TGAG
TTTGGGGCAATAATCCAGTC 53! 47 (*) 413, 1183
MB5R1–UPM (5VUTR) TCCTCCATAACCACATCCCT CTAATACGACTCACTATAGGGCAA
GCAGTGGTAACAACGCAGAGT
56 469
MB5R2–UPM (5VUTR) GATATTAGGAGCCTCTTTCAT CTAATACGACTCACTATAGGGCAA
GCAGTGGTAACAACGCAGAGT
56 356
MB3R1–NUP (3VUTR) ATCTCAGAGGGACCCAGGCGTGC AAGCAGTGGTAACAACGCAGAGT 58 1278
MB3R2–NUP (3VUTR) TGCTGTACCCCCACCAAGATG AAGCAGTGGTAACAACGCAGAGT 58 1032
3VACrepF–3VACrepR(AC repeat)
(Fluorescent-Cy5)AACCAGAGTAG
AGGCCACAA
GACAGAGCACAGAACACCGA 56 261
M2OF–E3R
(exon 2!exon 3)
TCCCTGAAGATCGACGTGAA TTTGGGGCAATAATCCAGTC 60! 52 (*) 335
(*) indicates a touchdown PCR approach.
L.Macca
trozzo
etal./Aquacultu
re205(2002)49–60
51

(adipose tissue, brain, eye, gonad, heart, intestine, kidney, liver, skeletal lateral muscle and
spleen) were dissected from 2-year-old anaesthetized shi drums, rapidly frozen in liquid
nitrogen and then stored at �80 �C.An RT-PCR approach together with the Rapid Amplification of CDNA Ends (RACE)
technique (Frohman et al., 1988) was used to isolate the complete GDF8 cDNA. Total
RNA was extracted from skeletal muscle tissue (50–100 mg) using Trizol Reagent
(Gibco, Gaithersburg, MD), and following the manufacturer’s instructions. The protocol
is a modification of the single-step RNA isolation method by Chomczynski and Sacchi
(1987). An aliquot of extracted RNA was tested spectrophotometrically and run on
agarose gel under denaturing conditions to confirm yield and quality. One microgram of
RNA was reverse-transcribed using a reverse transcriptase (Superscript II, Gibco) and
random hexamers to obtain first strand cDNA. The obtained cDNA was then used as
template for the subsequent PCR reactions (experimental conditions are given in Table
1). All PCR were performed with 32 cycles using GeneAmp PCR System 9700 (Perkin
Elmer, Branchburg, NJ, USA) in 20 ml of PCR mix [1� Promega PCR buffer, 1.8 mM
MgCl2, 0.1 mM of each dNTPs, 0.5 mM of each primer, 0.5 U Taq DNA Polymerase
(Promega, Madison, USA), 1 ml cDNA]. Initially, two fragments were amplified using
degenerate primers, designed on the basis of GDF8 amino acid sequences in other
species (M1F–M1R, M3F–M3R, Table 1). PCR products were cloned into a plasmid
vector (pCR-II, Invitrogen, Carlsbad, CA) using a TOPO-TA cloning kit (Invitrogen) and
sequenced using the Thermosequenase pre-mixed cycle sequencing kit (Amersham-
Pharmacia Biotech, Uppsala, Sweden). At least three independent clones were
sequenced. Based on the sequenced fragments, specific primers were designed to amplify
other two cDNA regions (E1F–M2R, M2F–E3R, Table 1), that were cloned and
sequenced as described above.
The SMART RACE cDNA Amplification kit (Clontech, Palo Alto, CA) was used
according to the manufacturer’s instructions to obtain the 5V and 3V unknown regions.
RACE reactions were carried out using shi drum specific primers (MB5R1, MB5R2,
MB3R1, MB3R2, Table 1), designed on the basis of the already obtained cDNA
sequence. PCR products of 5V and 3V RACE were cloned and sequenced as described
above.
In addition to the cDNA sequence, two introns were completely sequenced. Total
genomic DNA was extracted using a salting-out protocol (Patwary et al., 1994) from 200
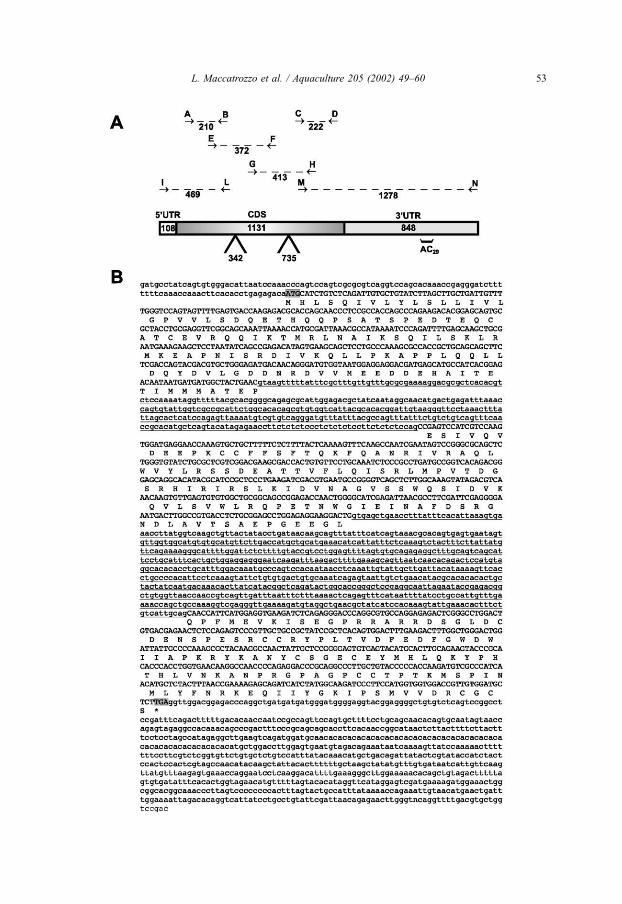
Fig. 1. (A) Experimental strategy used to obtain complete sdGDF8 cDNA. Dotted lines indicate amplified
fragment with the corresponding primers: A–B (M1F–M1R), C–D (M3F–M3R), E–F (E1F–M2R), G–H
(M2F–E3R), I–L (MB5R1–UPM), M–N (MB3R1–NUP). The fragment size is indicated in base pair. The grey
rectangles represent the full-length sdGDF8 cDNA; 5VUTR, 3VUTR and the coding sequence (CDS) are
differently shaded. The position and size of both introns and microsatellite (AC29) region are indicated on the
cDNA. (B) Complete nucleotide sequence of sdMSTN cDNA (2086 bp) (GenBank AF316881), and sequence of
intron 1 (342 bp) and intron 2 (735 bp) (underlined) as determined by PCR amplification and sequencing of
genomic DNA (GenBank AF316882). The inferred protein sequence is reported below the corresponding coding
region (upper case), which is delimited by the putative starting and stop codons (in grey). The noncoding
sequences are typed in lower case.
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–6052
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–60 53

mg of muscle tissue. Hundred nanograms of extracted DNA were subjected to PCR
amplification, using four primers (E1F–M2R, M2F–E3R, Table 1), presumably encom-
passing the position of the two introns, based on information from human (Gonzalez-
Cadavid et al., 1998) and porcine GDF8 gene (Stratil and Kopecny, 1999). Both PCR
products obtained were cloned and completely sequenced as described above.
Similarity searches were performed using the BLASTP program (http://www.
ncbi.nlm.nih.gov/BLAST/) with default settings on the complete, non-redundant Gen-
Bank database of translated coding sequences (release 2.0.11, Jan-20-2000).
To test the polymorphism of the identified microsatellite (see Results), two specific
primers (3VACrepF–3VACrepR, Table 1) were designed on the sequences flanking the
microsatellite region. Genomic DNA (50 ng) was obtained as described above, from 12
shi drum specimens, collected at the earlier mentioned location. Individual DNA
samples were amplified and PCR products were resolved on 6% denaturing polyacry-
lamide gels run onto an ALFexpressII automatic sequencer (Amersham Pharmacia
Biotech).
RT-PCR expression analysis was carried out on total RNA that was extracted using
the same method described above, from different tissues and organs of adult shi drum
(adipose tissue, brain, eye, gonad, heart, intestine, kidney, liver, skeletal lateral muscle
and spleen). For each sample, the same amount of RNA (1.5 mg) was reverse-tran-
scribed, using as primers either a (T)30 oligo or a mixture of random hexamers. RT
products were amplified with two specific primers encompassing intron 2 (M2OF–E3R,
Table 1). PCR products from each tissue were electrophoresed on a 1.8% agarose gel. To
confirm the specificity of PCR products, the agarose gel was blotted onto a nylon
membrane and hybridised with a specific probe (M2OF–E3R, Table 1) that was
digoxigenin-labelled (DIG Nucleic Acid Detection Kit, Roche, Mannheim, Germany).
DIG-labelled probe was detected, after hybridisation, by enzyme-linked immunoassay
using an antibody anti-DIG alkaline phosphatase conjugate. A subsequent enzyme-cata-
lysed colour reaction with 5-bromo-4-chloro-3-indolyl phosphate (BCIP) and nitro blue
tetrazolium salt (NBT) produced an insoluble blue precipitate, which visualised hybrid
molecules.
3. Results
Initially, two fragments of 210 and 222 base pairs (bp), respectively, were amplified and
sequenced by means of RT-PCR (A–B, C–D, Fig. 1A). Subsequently, other two
Fig. 2. Multiple alignment of GDF8 amino acid sequences. Shi drum myostatin (sdMSTN), Atlantic salmon
(asMSTN, GenBank AJ297267), brook trout (btMSTN, GenBank AF247650), zebrafish (zMSTN, GenBank
AF019626), mouse (mMSTN, GenBank U84005) and human (hMSTN, GenBank AF019627). Dashes indicate
insertion–deletions, shading refers to different degree of overall conservation for each site (black—100%, dark
grey—80%, light grey—60%). Other four fish GDF8 cDNA can be found in GenBank (yellow perch, mahi-
mahi, little tunny and king mackerel). As these are only partial sequences, they are not included in the
alignment.
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–6054

L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–60 55

fragments, 372- and 413-bp long (E–F, G–H), were amplified using shi drum specific-
primers. To characterise the 5V and 3V unknown regions, two PCR products of 469 and
1278 bp were obtained using a RACE protocol (I–J, K–L, Fig. 1A), and completely
Fig. 3. Results of electrophoretic separation of microsatellite alleles on ALFexpressII sequencer. Each lane
corresponds to a different screened individual (1–12). The uppermost and lowermost lanes (C) represent the
amplified product from the original clone containing the microsatellite repeat (262 bp, 29 AC repeats). In the
chromatogram, peaks represent microsatellite alleles. For each individual, the allele dimension is indicated in
base pairs.
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–6056

sequenced. Overlapping sequences of all fragments allowed to achieve a full-length
cDNA (GenBank accession number AF316881), which consisted of 2086 bp. A search
for open reading frames (ORF) revealed an ORF of 1131 bp, with the first possible start
codon located at position 109. This ORF encoded a peptide sequence of 376 amino
acids. The putative 5V untranslated region (UTR) was 108 bp, while the 3VUTR was
847 bp in length (Fig. 1B). The latter region is characterised by an ‘‘imperfect’’ AC29
microsatellite repeat (Fig. 1A,B). The entire inferred amino acid sequence (376 residues)
was used as a query in a BLASTP search with default parameters (Fig. 2). The highest
similarity values were obtained for the Atlantic salmon and brook trout myostatin with
301–300 identical residues over 376 compared amino acid positions, and the zebrafish
GDF8 (292/376), followed by other vertebrate GDF8s (226–230/372). Analysis of the
shi drum protein sequence showed a potential proteolytic processing site (RARR,
matching the RXXR consensus site) and seven cysteine residues in the carboxy-terminal
portion of the coding sequence, which corresponds to the mature processed protein
(McPherron and Lee, 1996), as observed in all TGF-b-like factors. The C-terminal
region, which is the most conserved, also contained two additional cysteines shared
among all vertebrate GDF8s. Based on the above results, the isolated sequence was
assumed to be the shi drum homologue of GDF8 (sdGDF8).
A partial characterisation of sdGDF8 gene was carried out through genomic amplifi-
cation of two regions (E–F, G–H), putatively encompassing two intronic sequences.
The complete sequence of the two amplified products revealed two introns, one (342-bp
long) in the first region (E–F), and a second one (735 bp) within fragment G–H
(GenBank accession number AF316882). Comparison between shi drum and mam-
malian GDF8 genes showed that intron–exon boundaries are conserved. However, both
introns are considerably smaller in the sdGDF8 gene than in its mammalian counter-
parts, because size of the first intron is 1808 bp in porcine GDF8 gene (Gonzalez-Cadavid
et al., 1998) and 1789 bp in human GDF8 (Stratil and Kopecny, 1999), while the second
intron spans 1977 and about 2400 bp, respectively.
An additional feature of the sdGDF8 cDNA was the presence of a repeat region
(AC29) in the 3VUTR. As this was a potential microsatellite marker, the same region was
investigated also at genomic level to evaluate its degree of polymorphism. Although
based on a limited sample size (12 individuals), results of this preliminary screening
revealed that the microsatellite is polymorphic, showing four alleles with size ranging
from 260 to 272 bp (Fig. 3). This microsatellite represents, therefore, a polymorphic
marker in tight linkage with the GDF8 locus.
To further characterise the sdGDF8 gene, its expression pattern was analysed by
means of RT-PCR. RT-PCR results for different organs and tissues were visualised on
agarose gel. A clearly visible PCR product was observed in the brain and muscle tissue,
while a much weaker band was obtained for the eye, gonad and heart. No band was
present in adipose tissue, intestine, kidney, liver and spleen (Fig. 4A). Hybridisation
with a DIG-labelled specific-probe confirmed the specificity of the RT-PCR assay,
yielding a strong signal for those tissue sample which were already positive on agarose
gel (Fig. 4B). In addition, this method revealed the presence of a faint band for other
tissues/samples (adipose tissue, kidney and spleen). No signal was present in the
intestine and liver even after Southern hybridisation. As a positive control, a b-actin
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–60 57
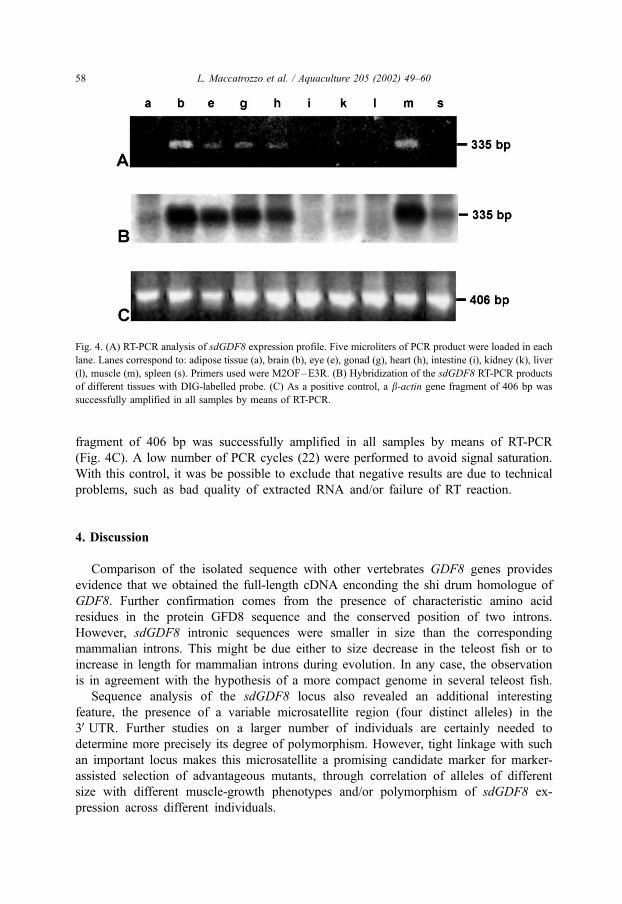
fragment of 406 bp was successfully amplified in all samples by means of RT-PCR
(Fig. 4C). A low number of PCR cycles (22) were performed to avoid signal saturation.
With this control, it was be possible to exclude that negative results are due to technical
problems, such as bad quality of extracted RNA and/or failure of RT reaction.
4. Discussion
Comparison of the isolated sequence with other vertebrates GDF8 genes provides
evidence that we obtained the full-length cDNA enconding the shi drum homologue of
GDF8. Further confirmation comes from the presence of characteristic amino acid
residues in the protein GFD8 sequence and the conserved position of two introns.
However, sdGDF8 intronic sequences were smaller in size than the corresponding
mammalian introns. This might be due either to size decrease in the teleost fish or to
increase in length for mammalian introns during evolution. In any case, the observation
is in agreement with the hypothesis of a more compact genome in several teleost fish.
Sequence analysis of the sdGDF8 locus also revealed an additional interesting
feature, the presence of a variable microsatellite region (four distinct alleles) in the
3VUTR. Further studies on a larger number of individuals are certainly needed to
determine more precisely its degree of polymorphism. However, tight linkage with such
an important locus makes this microsatellite a promising candidate marker for marker-
assisted selection of advantageous mutants, through correlation of alleles of different
size with different muscle-growth phenotypes and/or polymorphism of sdGDF8 ex-
pression across different individuals.
Fig. 4. (A) RT-PCR analysis of sdGDF8 expression profile. Five microliters of PCR product were loaded in each
lane. Lanes correspond to: adipose tissue (a), brain (b), eye (e), gonad (g), heart (h), intestine (i), kidney (k), liver
(l), muscle (m), spleen (s). Primers used were M2OF–E3R. (B) Hybridization of the sdGDF8 RT-PCR products
of different tissues with DIG-labelled probe. (C) As a positive control, a b-actin gene fragment of 406 bp was
successfully amplified in all samples by means of RT-PCR.
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–6058

The most striking finding of the present work, however, was obtained when RT-PCR
analysis of sdGDF8 expression was carried out on different organs and tissues. Positive
signal observed in the skeletal and heart muscle is in keeping with results for mammalian
GDF8s (McPherron et al., 1997; Gonzalez-Cadavid et al., 1998; Sharma et al., 1999),
thereby suggesting that its role in regulating muscle growth might be conserved across
vertebrate taxa. On the other hand, GDF8 is expressed also in other tissues/organs in the
shi drum, especially in the brain. Except for the adipose tissue, these results are at odd
with the expression profile in mammals. Since most expression data on mammalian
GDF8s were obtained using Northern blot analysis, it might be possible that expression in
a larger number of tissues could be revealed in other species by means of RT-PCR.
Alternatively, the wider expression profile of sdGDF8 might suggest that GDF8 have
additional functions in U. cirrosa. Further studies are thus important to better understand
the role of GDF8 in lower vertebrates.
In conclusion, sequence conservation and expression in muscle tissue indicate a
conserved role for GDF8 in the shi drum, thereby confirming the potential of this gene
for practical applications also in aquacultured fish species. Moreover, the associated
microsatellite locus might be extremely useful for marker-assisted selection of advanta-
geous mutants. At present, no technology is available for homologous recombination in
fish, to achieve specific gene knock-out. However, null mutants might be obtained for
instance by means of production of dominant negative transgenic animals. If such GDF8
mutants are to be produced for aquaculture purposes, the wide expression profile of
GDF8 observed in the shi drum indicates that inactivation of GDF8 might be lethal or
highly detrimental, unless experimental suppression or reduction of GDF8 function is
limited to the muscle tissue.
Acknowledgements
We acknowledge the ‘‘Impianto di Acquacoltura di Pellestrina, Veneto Agricoltura’’ for
providing access to samples. This work was supported by ‘‘cofinanziamento Ministero
Italiano per le Politiche Agricole-Universita di Padova’’ to F.M. and by ‘‘ex-40%
MURST’’ to T.P.
References
Chomczynski, P., Sacchi, N., 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate–
phenol–chloroform extraction. Anal. Biochem. 162 (1), 156–159.
Frohman, M.A., Dush, M.K., Martin, G.R., 1988. Rapid production of full-length cDNAs from rare transcripts:
amplification using a single gene-specific oligonucleotide primer. Proc. Natl. Acad. Sci. U. S. A. 85, 8998–
9000.
Gonzalez-Cadavid, N.F., Taylor, W.E., Yarasheski, K., Sinha, H.I., Ma, K., Ezzat, S., Shen, R., Lalani, R., Asa, S.,
Mamita, M., Nair, G., Arver, S., Bhasin, S., 1998. Organization of the human myostatin gene and expression
in healthy men and HIV-infected men with muscle wasting. Proc. Natl. Acad. Sci. U. S. A. 95, 14938–14943.
Grobet, L., Martin, L.J., Poncelet, D., Pirottin, D., Brouwers, B., Riquet, J., Schoeberlein, A., Dunner, S.,
Menissier, F., Massabanda, J., Fries, R., Hanset, R., Georges, M., 1997. A deletion in the bovine myostatin
gene causes the double-muscled phenotype in cattle. Nat. Genet. 17, 71–74.
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–60 59

Ji, S.Q., Losinski, R.L., Cornelius, S.G., Frank, G.R., Willis, G.M., Gerrard, D.E., Depreux, F.F.S., Spurlock,
M.E., 1998. Myostatin expression in porcine tissues: tissue specificity and developmental and postnatal
regulation. Am. J. Physiol.: Regul., Integr. Comp. Physiol. 44, R1265–R1273.
McPherron, A.C., Lee, S.J., 1996. The transforming growth factor-beta superfamily. Growth Factors Cytokines
Health Dis. 1B, 357–393.
McPherron, A.C., Lee, S.J., 1997. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl.
Acad. Sci. U. S. A. 94, 12457–12461.
McPherron, A.C., Lawler, A.M., Lee, S.J., 1997. Regulation of skeletal muscle mass in mice by a new TGF-beta
superfamily member. Nature 387, 83–90.
Patwary, M.U., Kenchington, E.L., Bird, C.J., Zouros, E., 1994. The use of random amplified polymorphic DNA
markers in genetic studies of the sea scallop Plactopecten magellanicus (GMELLIN, 1791). J. Shellfish Res.
13, 547–553.
Sharma, M., Kambadur, R., Matthews, K.G., Somers, W.G., Devlin, G.P., Conaglen, J.V., Fowke, P.J., Bass, J.J.,
1999. Myostatin, a transforming growth factor-beta superfamily member, is expressed in heart muscle and is
upregulated in cardiomyocytes after infarct. J. Cell. Physiol. 180, 1–9.
Stratil, A., Kopecny, M., 1999. Genomic organization, sequence and polymorphism of the porcine myostatin
(GDF8; MSTN) gene. Anim. Genet. 30, 468–470.
L. Maccatrozzo et al. / Aquaculture 205 (2002) 49–6060





























