Embed Size (px)
Citation preview

Comp. by:bala Date:27/5/05 Time:20:26:44 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Cellular Functions of EndoplasmicReticulum Chaperones Calreticulin,Calnexin, and Erp57
Karen Bedard,* Eva Szabo,{ Marek Michalak,* and Michal Opas{
*Membrane Protein Research Group and Department of Biochemistry,
University of Alberta, Edmonton, Alberta, Canada T6G 2H7{Department of Laboratory Medicine and Pathobiology,
University of Toronto, Toronto, Ontario, Canada M5S 1A8
Glycosylated proteins destined for the cell surface or to be secreted from the
cell are trafficked trough the endoplasmic reticulum during synthesis and folding.
Correct folding is determined in large part by the sequence of the protein, but it is
also assisted by interaction with enzymes and chaperones of the endoplasmic
reticulum. Calreticulin, calnexin, and ER57 are among the endoplasmic
chaperones that interact with partially folded glycoproteins and determine if the
proteins are to be released from the endoplasmic reticulum to be expressed, or
alternatively, if they are to be sent to the proteosome for degradation. Studies on
the effect of alterations in the expression and function of these proteins are
providing information about the importance of this quality control system, as well
as uncovering other important functions these proteins play outside of the
endoplasmic reticulum.
KEY WORDS: Adhesion, Calreticulin, Calnexin, ERp57, Protein folding,
Calcium homeostasis. � 2005 Elsevier Inc.
I. Introduction
Attaining the correct conformation is essential for the proper functioning
and expression of proteins. For surface and secreted proteins, which pass
through the endoplasmic reticulum (ER) during assembly, this folding
process is aided by interaction with chaperone proteins, which stabilize
International Review of Cytology, Vol. 245 91 0074-7696/05 $35.00Copyright 2005, Elsevier Inc. All rights reserved. DOI: 10.1016/S0074-7696(05)45004-4

Comp. by:bala Date:27/5/05 Time:20:26:44 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
intermediate conformations allowing time to achieve the correct folding
structure and preventing aggregations. There is an interest in understanding
this process, as defects in folding and processing have been associated with a
number of diseases including cystic fibrosis, prion diseases, and Alzheimer’s.
Furthermore, while these ER chaperones are primarily identified for their
role in protein folding, they are also responsible for a number of other critical
functions, including regulation of calcium homeostasis, activation of specific
transcription factors, and oxidative stress, to name a few. The multifunction-
al nature and differential expression of these unique chaperones may have
relevance to a wide range of conditions, including cardiovascular develop-
ment and function, cancer and neurodegenerative conditions, metabolic
problems, and others. This review will focus on the function of ER chaper-
ones calreticulin, calnexin, and ERp57, and the consequences of their altered
expression and function.
II. Endoplasmic Reticulum and Chaperone Proteins
A. Endoplasmic Reticulum
The ER is a network of membrane‐bound tubules continuous with the
nuclear envelope and found throughout the cytoplasm. The lumen of ER is
a distinctly different environment from the rest of the cell, suited to perform
its functions of xenobiotic metabolism, phospholipid and steroid synthesis,
calcium sequestration, and the synthesis of membrane‐bound and secreted
proteins. As these surface and secreted proteins are transcribed, they are
translocated to the lumen of the ER, where they interact with chaperone
proteins, including Grp78, Grp94, protein disulfide isomerase (PDI),
PDI‐like proteins, calreticulin, calnexin, and ERp57. These chaperones assist
the newly formed protein in reaching its correct folding formation by stabi-
lizing intermediate forms, slowing the folding process, and preventing
misfolding and aggregations.
ER is also an important ion storage organelle. The ion concentrations in
the lumen of the ER resemble the extracellular environment, which may be
important in the synthesis of membrane surface and secreted proteins. The
reported calcium content of the ER ranges from the high micromolar to
millimolar range, orders of magnitude higher than 100 nM calcium con-
centrations found the cytosol. Release of calcium from the ER into the
cytosol leads to large increases in the cytoplasmic calcium concentration,
and is an important component of intracellular signaling. Numerous cyto-
plasmic transcription factors, phosphatases, enzymes, and channels are
sensitive to changes in calcium concentration modulator of their activity.
92 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:44 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
The sarcoplasmic/endoplasmic calcium ATPase (SERCA) acts to transport
calcium from the cytosol into the ER, and maintain this concentration
gradient, while channels in the ER allow for stimulated release of calcium.
The high concentration of calcium in the ER, combined with the volume of
the cell that the ER can occupy, allows for the potential of calcium released
from the ER to reach toxic levels in the cell if its release is not controlled.
Calcium‐binding proteins within the lumen of the ER, including calreticulin
and calnexin (Baksh andMichalak, 1991; Tjoelker et al., 1994), act to further
regulate the amount of free versus bound calcium within the ER. This free
calcium concentration is important for controlling the amount of calcium
that can be released, as well as for regulating calcium‐dependent processes inthe ER, such as chaperone interactions with each other and with their
substrates. The binding of glycoproteins to calnexin is calcium dependent
(Capps and Zuniga, 1994; Le et al., 1994).
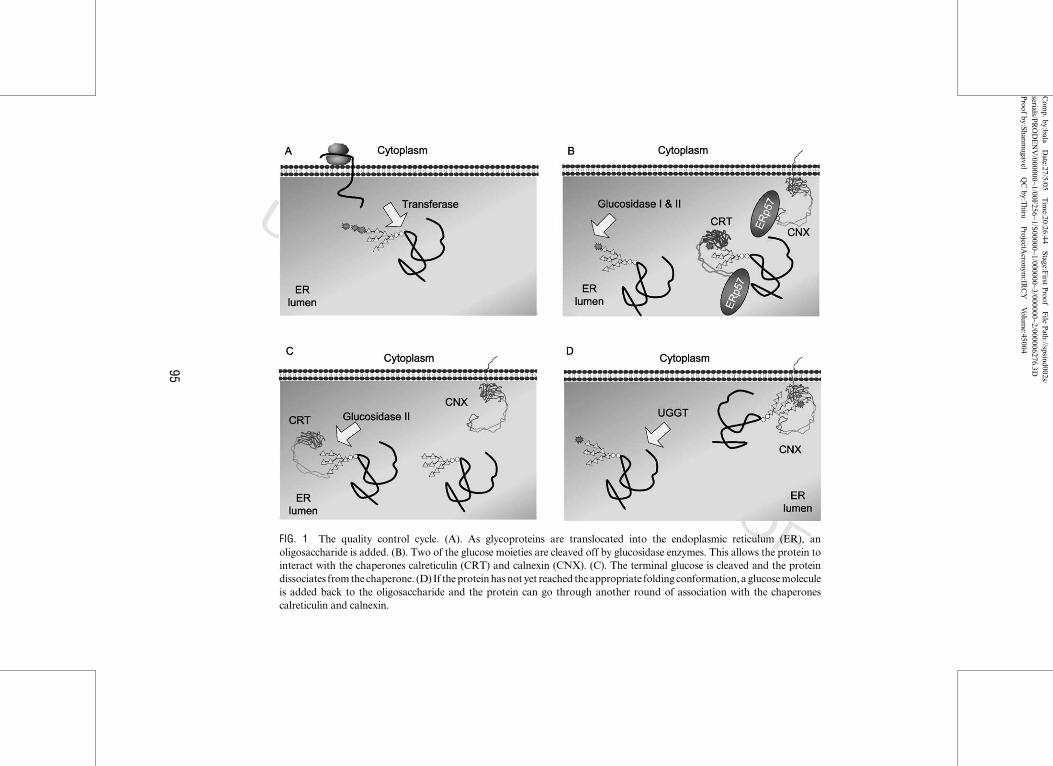
B. Chaperoning of N‐Glycosylated Proteins through ER
As proteins are translocated into the ER, the leader sequence is cleaved by
protease. This is followed by the addition of an oligosaccharide by a trans-
ferase. This addition of sugar can act to stabilize the protein, increase
its solubility (Drickamer and Taylor, 1998; Dwek, 1996; O’Connor and
Imperiali, 1996; Wormald and Dwek, 1999; Wormald et al., 2002), and assist
in trafficking the protein. Although mature glycoproteins have very hetero-
geneous N‐linked glycans, all glycoproteins go through similar trimming in
the ER, and acquire their final glycoprotein structure as they pass through
the Golgi. TwoN‐acetylglucosamines and nine mannoses with three terminal
glucose residues are assembled onto a core carbohydrate, which is then
transferred to asparagine residues of the nascent polypeptide chain. As
soon as the oligosaccharide is added, trimming begins. The three terminal
glucoses are trimmed by glucosidase I and II, and a terminal mannose is
trimmed by one or more ER mannosidases (Helenius and Aebi, 2004). Both
calreticulin and calnexin bind transiently to a newly synthesized glycoprotein
intermediate, which still contains one terminal glucose. This serves to prevent
aggregation, protect proteins from premature degradation, and ensure the
correct folding status of proteins before continuing in the intracellular traf-
ficking pathway. The binding of calreticulin and calnexin to their substrates
is mediated, at least in part, by a lectin site that recognizes the N‐linkedoligosaccharide processing intermediate Glc1Man9GlcNAc2 (Chen et al.,
1995; Helenius and Aebi, 2001). Both chaperones require the presence of
calcium (Chevet et al., 1999; Vassilakos et al., 1998) and both require the
terminal glucose to be present for the binding of the majority of their
substrates (Di Jeso et al., 2003; Hammond et al., 1994; Nauseef et al., 1995;
CELLULAR FUNCTIONS OF ER CHAPERONES 93

Comp. by:bala Date:27/5/05 Time:20:26:44 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Ou et al., 1993; Peterson et al., 1995; Rodan et al., 1996), there is evidence
that calnexin and calreticulin can recognize the polypeptide segments of
the glycoproteins as well (Arunachalam and Cresswell, 1995; Baksh et al.,
1995; Carreno et al., 1995; Ihara et al., 1999; Margolese et al., 1993; Nigam
et al., 1994; Rojiani et al., 1991; van Leeuwen and Kearse, 1996; Ware et al.,
1995; Zhang et al., 1995; Zapun et al., 1997). The binding of proteins to
calreticulin or calnexin is terminated by removal of the third glucose by
glucosidase II. Inhibition of glucose trimming may lead to accelerated deg-
radation (Moore et al., 1993; Saito et al., 1999) or delayed secretion (Kearse
et al., 1994; Lodish and Kong, 1984; Sasak et al., 1985). If the protein is not
correctly folded, it can be reglycosylated by UDP‐glucose:glycoprotein glu-
cosyltransferase (UGGT), and reassociate with calreticulin and calnexin. In
this model, a sensor of glycoprotein folding, UGGT, recognizes incompletely
folded proteins and reglucosylates them, allowing them to reassociate with
the ER chaperones (Rabouille and Spiro, 1992). This cycle of association
with the chaperone, glucosidase II trimming and release from the chaperone,
assessment of foldedness, and glucosylation if necessary by UGGT followed
by reassociation with the chaperones acts as a quality control mechanism
(Fig. 1).
C. ER Quality Control Proteins
Calreticulin is a soluble protein found within the lumen of the ER. It has
three domains, the compact globular N‐domain encompassing the first 200
residues that does not bind calcium, the extended arm formed by the hairpin
loop of the P‐domain encompassing residues 187–317, enriched in proline
residues, and binding calcium with low capacity, high affinity (�1 mol/mol
protein, K¼ 10 mM), and finally the carboxy‐terminal C domain that encom-
passes 310–401 and binds calcium with high capacity, low affinity
(�18 mol/mol protein, K ¼ 2 mM) (Parodi, 2000). The primary function
ascribed to calreticulin has been as a chaperone protein where it binds to
newly synthesized glycoproteins, preventing aggregation and allowing the
proteins to attain their correct folding conformation. Calreticulin also has an
important role in calcium regulation.
Although calreticulin is found primarily in the ER (Fig. 2A), the protein
has also been detected on the cell surface and circulating outside of the cell
(Johnson et al., 2001). The function of this extra‐ER calreticulin is not clear,
but its presence in these locations may have implications in some autoim-
mune diseases (Eggleton, 2003). Adverse drug reactions with circulating
antibodies to calreticulin have also been reported (Murphy‐Ullrich, 2001;
Nair et al., 1999). In addition to these locations calreticulin was also detected
with specific antibodies in the nucleus of some cells, such as the nucleus of
94 BEDARD ET AL.
Com
p.by:balaDate:27/5/05
Tim
e:20:26:44Stage:F
irstProof
File
Path://spsind002s/
serials/PRODENV/000000~1/00F
256~1/S00000~1/000000~3/000000~2/000006276.3D
Proof
by:Shanm
ugavelQCby:T
hiruProjectA
cronym:IR
CY
Volum
e:45004
UNCORRECTEDPROOFFIG. 1 The quality control cycle. (A). As glycoproteins are translocated into the endoplasmic reticulum (ER), an
oligosaccharide is added. (B). Two of the glucose moieties are cleaved off by glucosidase enzymes. This allows the protein to
interact with the chaperones calreticulin (CRT) and calnexin (CNX). (C). The terminal glucose is cleaved and the protein
dissociates fromthe chaperone. (D) If theproteinhasnot yet reached theappropriate folding conformation, aglucosemolecule
is added back to the oligosaccharide and the protein can go through another round of association with the chaperones
calreticulin and calnexin.
95
Comp. by:bala Date:27/5/05 Time:20:26:45 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
squamous carcinoma cells in response to ionizing radiation (Ramsamooj
et al., 1995) and in the nucleus of dexamethasone‐treated LM cells (Roderick
et al., 1997), but evidence contrary to these findings indicated that the nuclear
localization of calreticulin was an artifact of immunostaining (Michalak
et al., 1996). However, calreticulin is clearly present in the nuclear structures
throughout early human development. It is not present intranuclearly, in-
stead it appears to be enriched in the nuclear envelope of human oocytes and
embryos (Balakier et al., 2002). The nuclear envelope functions as a calcium
store continuous with the ER, thus calcium‐mediated events have been
implicated in a variety of nuclear activities, including modulation of chro-
matin structure and function, gene expression, DNA synthesis, nucleocyto-
plasmic transport, and changes in nuclear architecture (Ashby and Tepikin,
2001; Bachs et al., 1992; Berridge et al., 2000; Petersen et al., 1998; Santella
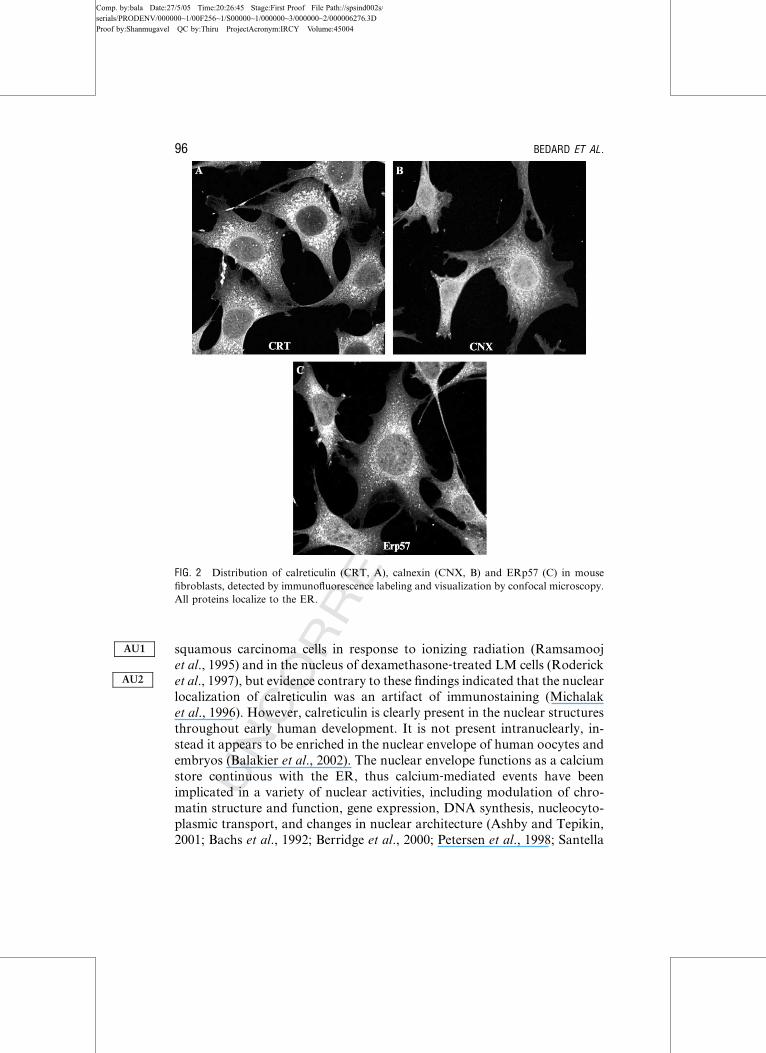
FIG. 2 Distribution of calreticulin (CRT, A), calnexin (CNX, B) and ERp57 (C) in mouse
fibroblasts, detected by immunofluorescence labeling and visualization by confocal microscopy.
All proteins localize to the ER.
AU1
AU2
96 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:46 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
and Carafoli, 1997). Regulation of gene expression seems to be capable
of differentiating between nuclear and cytoplasmic calcium signaling
(Hardingham and Bading, 1999; Hardingham et al., 1997). Therefore, the
presence of the calcium‐binding chaperones (including sometimes calnexin)
within nuclear structures of human oocytes and embryos might also be
crucial for similar nuclear activities. There is also evidence that calreticulin
can be found in the cytoplasm (Baksh and Michalak, 1991; Jethmalani et al.,
1997; Macias et al., 2003; Yoon et al., 2000). Calreticulin has also been found
in the Golgi where it may associate with endomannosidase. This may play a
role in calreticulin–substrate dissociation (Holaska et al., 2001).
Calnexin is a type I membrane protein composed of a luminal domain
highly homolgous to calreticulin and a cytoplasmic domain of 88 amino
acids. Calnexin shares 42–78% sequence identity with calreticulin (Spiro
et al., 1996), with the highest homology in the proline‐rich extended arm of
the high‐affinity calcium‐binding P‐domain (Baksh and Michalak, 1991;
Wada et al., 1991). Calnexin acts to prevent export of incorrectly or incom-
pletely folded proteins (Jackson et al., 1994; Rajagopalan et al., 1994;
Tjoelker et al., 1994). The protein also helps to prevent rapid degradation
(Jackson et al., 1994; Kearse et al., 1994; Rajagopalan and Brenner, 1994).
Calreticulin and calnexin exhibit prolonged interaction with mutant glyco-
proteins that fail to fold (Helenius, 1994; Moore et al., 1993; Nauseef et al.,
1995; Peterson et al., 1995). Prolonged interaction with calnexin results in the
substrate being directed to the proteosome for degradation (Jakob et al.,
1998; Otteken and Moss, 1996). Misfolded proteins, but not proteins under-
going productive folding, are extracted from calnexin by the a‐mannosidase
I‐like protein EDEM (Liu et al., 1999). EDEM does not interact with
calreticulin (Molinari et al., 2003).
Although calnexin lacks the high capacity calcium‐binding domain, it has
been reported that the phosphorylation status of the cytoplasmic tail can
regulate calcium through an interaction with SERCA2B (Oda et al., 2003;
Roderick et al., 2000). Like calreticulin, calnexin is predominantly located in
the ER (Fig. 2B), but it has also been identified at the cell surface of a number
of cells (Schrag et al., 2001). What role calnexin plays at the cell surface is not
clear, but again, circulating autoantibodies to calnexin have been found in
patients with autoimmune diseases. Calnexin has not been found in the
cytoplasm or nucleus, unlike calreticulin. On the other hand, it appears
that in mammalian somatic cells, calnexin and calreticulin are always
expressed in an identical pattern within the ER. The heterogeneity in com-
partmentalization of calnexin and calreticulin is, however, evident during
development. In human oocytes the two chaperones calnexin and calreticulin
are nonuniformly distributed (Balakier et al., 2002). Even though both are
localized to the cortex region of the oocyte, their distribution in the region
differs; calnexin has an interesting trilaminar arrangement, while calreticulin
CELLULAR FUNCTIONS OF ER CHAPERONES 97
Comp. by:bala Date:27/5/05 Time:20:26:46 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
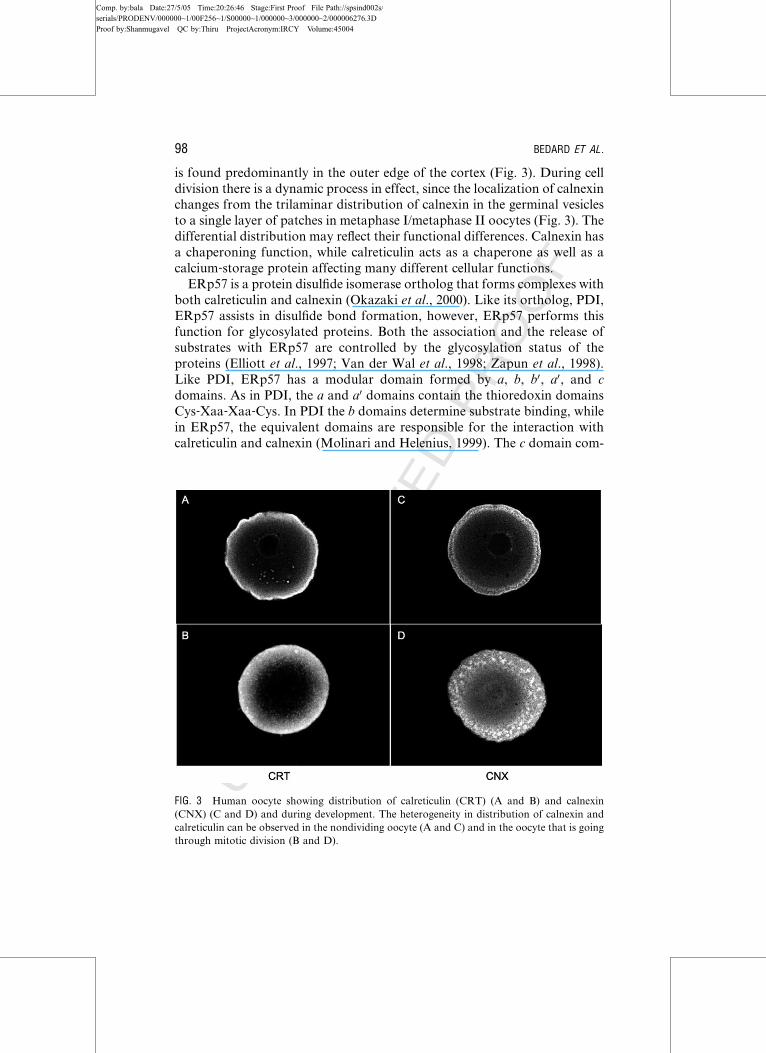
is found predominantly in the outer edge of the cortex (Fig. 3). During cell
division there is a dynamic process in effect, since the localization of calnexin
changes from the trilaminar distribution of calnexin in the germinal vesicles
to a single layer of patches in metaphase I/metaphase II oocytes (Fig. 3). The
differential distribution may reflect their functional differences. Calnexin has
a chaperoning function, while calreticulin acts as a chaperone as well as a
calcium‐storage protein affecting many different cellular functions.
ERp57 is a protein disulfide isomerase ortholog that forms complexes with
both calreticulin and calnexin (Okazaki et al., 2000). Like its ortholog, PDI,
ERp57 assists in disulfide bond formation, however, ERp57 performs this
function for glycosylated proteins. Both the association and the release of
substrates with ERp57 are controlled by the glycosylation status of the
proteins (Elliott et al., 1997; Van der Wal et al., 1998; Zapun et al., 1998).
Like PDI, ERp57 has a modular domain formed by a, b, b0, a0, and c
domains. As in PDI, the a and a0 domains contain the thioredoxin domains
Cys‐Xaa‐Xaa‐Cys. In PDI the b domains determine substrate binding, while
in ERp57, the equivalent domains are responsible for the interaction with
calreticulin and calnexin (Molinari and Helenius, 1999). The c domain com-
FIG. 3 Human oocyte showing distribution of calreticulin (CRT) (A and B) and calnexin
(CNX) (C and D) and during development. The heterogeneity in distribution of calnexin and
calreticulin can be observed in the nondividing oocyte (A and C) and in the oocyte that is going
through mitotic division (B and D).
98 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:47 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
prises the carboxy‐terminus. Mapping of interaction sites indicates that
ERp57 will interact with calreticulin and calnexin through association with
the proline‐rich P‐domain arm of the chaperones. ERp57 has also been
shown to regulate the redox status of the luminal face of SERCA2b,
providing dynamic control over ER calcium (Russell et al., 2004). So far
exclusive localization of ERp57 has been to the ER (Fig. 2C).
D. Importance of the Components of the QualityControl Cycle
The importance of the components involved in the synthesis and quality
control of glycosylated proteins is revealed through the effects of deficits in
these components. Mice with inactivated N‐acetlyglucosamine transferase I
die at mid‐gestation age. Patients with mutations in the glucosyltransferase
involved in the synthesis of the oligosaccharide suffer from severe abnorm-
alities including psychomotor retardation and seizures (Li and Camacho,
2004). In yeast, UDP‐Glc:glycoprotein glucosyltransferase is essential for
Schizosaccharomyces pombe viability under conditions of extreme ER stress
(Stanley and Ioffe, 1995). Calreticulin‐deficient mice die mid‐gestationally(Westphal et al., 2003). Calnexin‐deficient mice are viable but with a
pronounced ataxic phenotype (Fanchiotti et al., 1998).
The absence of calreticulin is lethal to the developing mouse embryo.
Calreticulin‐deficient mice display defects in the developing heart (Mesaeli
et al., 1999) and 16% displayed exencephaly (brain outside of skull) (Denzel
et al., 2002). Examination of the expression of the calreticulin gene in the
developing embryo shows little expression in most tissue, but strong expres-
sion in the heart, liver, and in some central nervous system (CNS) tissue at
the stage of development when calreticulin deficiency is lethal. Calreticulin‐deficient mice are ‘‘rescued’’ by a cardiac‐specific constitutively active form
of calcineurin (Mesaeli et al., 1999). The ‘‘rescued’’ mouse is by no means
completely normal, however, with a reduced body size and problems with
lipid regulation, among other things. This may be related to the high level of
expression of calreticulin in adipose tissue. The rescue mouse, along with
further in vitro studies, did however indicate that an important function of
calreticulin is its role as an upstream regulator of calcineurin (Rauch et al.,
2000).
Just as the absence of calreticulin is lethal during embryonic devel-
opment, where its expression in the heart is normally high, overexpression
of calreticulin leads to heart problems after birth, when the expression of
calreticulin is normally down‐regulated. Mice overexpressing calreticulin
in the heart experience complete and sudden heart block after birth (Guo
et al., 2002).
CELLULAR FUNCTIONS OF ER CHAPERONES 99

Comp. by:bala Date:27/5/05 Time:20:26:47 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
The absence of calnexin leads to a very different phenotype. The mice are
viable, but with a reduced survivability. They are smaller in size than their
wild‐type littermates. They display an abnormal gait and appear to have
reduced limb coordination (Lynch and Michalak, 2003). The mice are viable
with no histological evidence of cardiovascular defects or changes in cardio-
vascular functional parameters (K. Bedard and M. Michalak, unpublished).
Neurologically, there is a reduction in the number of large myelinated nerve
fibers (Nakamura et al., 2001), which may account for the motor defect. No
difference was observed in the total expression of a number of proteins
chaperoned by calnexin (Denzel et al., 2002).
The cardiovascular phenotype in the calreticulin embryo and neurological
phenotype in the calnexin embryo correlate with the pattern of gene expres-
sion. Calreticulin expression in the developing embryo is low in the CNS but
high in the heart. The opposite pattern is observed for calnexin expression
(K. Bedard, unpublished), suggesting the need for two similar proteins may
relate to their expression patterns.
A striking feature revealed by these studies is that calreticulin and calnexin
are unable to compensate for the loss of each other, therefore suggesting
unique and nonoverlapping functions (Denzel et al., 2002; Mesaeli et al.,
1999; Nakamura et al., 2001). One function of calreticulin that cannot be
compensated by calnexin is its role in modulation of calcium homeostasis
(Denzel et al., 2002; Knee et al., 2003) We created viable crt� and cnx�/� cell
lines indicating that in mammalian cell culture calreticulin and calnexin (and
the calreticulin/calnexin cycle) are not essential for cell survival (Arnaudeau
et al., 2002; Nakamura et al., 2001). Other studies support this idea, for
example, Saccharomyces cervisiae lacks most of the calnexin/calreticulin
components (Mesaeli et al., 1999). Deletion of glucosidase II in mammalian
cells and glucosidase II and UGGT, which are key components of the
calreticulin/calnexin cycle, in S. pombe has no consequences on cellular
function (Scott and Dawson, 1995). Yet, calnexin deficiency is lethal in
S. pombe (Parlati et al., 1995). In Dictyostelium and Caenorhabditis elegans
calnexin and calreticulin deficiency is not lethal but it affects phagocytosis in
Dictyostelium (D’Alessio et al., 1999) and promotes necrotic cell death in
C. elegans (Parlati et al., 1995). In summary, these findings support our
hypothesis that calreticulin and calnexin are multifunctional proteins. The
molecular chaperone function of calreticulin and calnexin may only partially
explain phenotypes of cnx�/� and crt�/� mice.
E. Need for Two Similar Chaperone Proteins
ERp57 is homologous with PDI and calreticulin is homologous with cal-
nexin. The existence of such homologous proteins in structure and function
may seem redundant. However, there are important differences between
100 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:47 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
these and other chaperones. ERp57 tends to catalyze the rearrangement of
disulfide bonds in glycosylated proteins while PDI handles nonglycosylated
proteins. Calreticulin is a soluble luminal protein, while calnexin is bound to
the membrane. This may lead to differences in the substrates with which each
interacts. For example, there are a large number of substrates that have been
demonstrated to interact with both calreticulin and calnexin. However, for
many substrates, interaction has been reported for only one or the other, and
in a few cases, selectivity for one over the other has been observed (Table I).
Even among the substrates that are able to interact with either chaperone,
there may be differences in the stage of folding at which the substrate
interacts with each chaperone (Cresswell, 2000; Hebert et al., 1996; Muller‐Taubenberger et al., 2001; Xu et al., 2001). Although physical interactions
with either substrate may be possible, the relevance of those interactions may
be affected by the relative abundance of each chaperone in a given tissue or at
a specific stage of development. This is an area that has not been fully
explored, but there are changes in the transcriptional activation of calreticu-
lin and calnexin in various tissues through development from embryo to
adult. Further, there are differences in the relative protein expression of
calreticulin and calnexin in adult tissues. Finally, the requirement of two
seemingly similar proteins may be related to unique functions of one or both
proteins outside of the quality control process. This is well documented by
gene knockout results.
III. Nonchaperone Functions of the Quality ControlCycle Components
The additional functions of the components of the glycosylation and quality
control cycle have been elucidated by genetic manipulation of the proteins
involved. In calreticulin‐deficient cells, there is acceleration of protein fold-
ing, but quality control is impaired (Guo et al., 2002; Knee et al., 2003;
Molinari et al., 2004). Substrate interaction with calnexin is reduced and the
accumulation of unfolded proteins leads to the triggering of an unfolded
protein response (Diedrich et al., 2001). Depletion of calreticulin accelerates
viral glycoprotein maturation, with only a small decrease in folding efficien-
cy. Similarly, depletion of calnexin had little effect on the maturation of
many viral proteins. Only when both were depleted was a large decrease in
ER quality control observed (Sadasivan et al., 1996). From the whole animal
studies, the depletion of these chaperones has made it clear that these
proteins are very important. However, cell culture studies, and the fact
that the embryos or mice developed to the extent that they did, also make
it clear that chaperoning is not required for the expression of surface
and secreted glycoproteins. These ER chaperones not only have roles in
CELLULAR FUNCTIONS OF ER CHAPERONES 101

Comp. by:bala Date:27/5/05 Time:20:26:47 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
TABLE I
Proteins Known to Be Chaperoned by Calreticulin, Calnexin, or Both Chaperones
Calreticulin Calnexin Both
Myeloperoxidaseb Pmp‐22c,d MHC class Ie
Myelin proteolipid
protein f
Type I and III IP3Rc,g Influenza HAh
Integrin a‐chaini VSV G‐proteinc,h Tyrosinasej
MHC class II a and bk Glut Il
NMDA subunit NR1m Thromboglobulinn
Naþ, Kþ‐ATPasek Thyrotropin receptoro
a‐Antitrypsinp Meprin Ag
Integrin b‐chainr SERTs
AMPA receptort von Willebrand factoru
Nicotinic acetylcholine
receptorvTransferrinw
a‐Fetoproteinw
HIV envelope protein
gp160x
aMHC, major histocompatibility complex; IP3R, inositol 1,4,5‐trisphosphate receptor; VSV
¼ vesicular stomatitis virus; NMDA ¼ N‐methyl‐D‐aspartate; SERT ¼ serotonin transporter;
AMPA ¼ a‐amino‐3‐hydroxy‐5‐methyl‐4‐isoxazolepropionate.bNauseef et al. (1995).cDoes not interact with calreticulin.dDickson et al. (2002).eBouvier, M. (2003). Accessory proteins and the assembly of human class I MHC molecules:
A molecular and structural perspective. Mol. Immunol. 39, 697–706.fGudz, T. I., Schneider, T. E., Haas, T. A., and Macklin, W. B. (2002). Myelin proteolipid
protein forms a complex with integrins and may participate in integrin receptor signaling in
oligodendrocytes. J. Neurosci. 22, 7398–7407.gJoseph, S. K., Boehning, D., Bokkala, S., Watkins, R., and Widjaja, J. (1999). Biosynthesis
of inositol trisphosphate receptors: Selective association with the molecular chaperone calnexin.
Biochem. J. 342(Pt. 1), 153–161.hPeterson et al. (1995).iKwon, M. S., Park, C. S., Choi, K., Ahnn, J., Kim, J. I., Eom, S. H., Kaufman, S. J., and
Song, W. K. (2000). Calreticulin couples calcium release and calcium influx in integrin‐mediated
calcium signaling. Mol. Biol. Cell 11, 1433–1443.jHalaban et al. (2000).kArunachalam and Cresswell (1995).lOliver, J. D., Hresko, R. C., Mueckler, M., and High, S. (1996). The glut 1 glucose
transporter interacts with calnexin and calreticulin. J. Biol. Chem. 271, 13691–13696.mHughes, P. D., Wilson, W. R., and Leslie, S. W. (2001). Effect of gestational ethanol
exposure on the NMDA receptor complex in rat forebrain: From gene transcription to cell
surface. Brain Res. Dev. Brain Res. 129, 135–145.nDi Jeso et al. (2003).
102 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:47 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
regulating protein folding, but also calcium homeostasis, cell adhesion,
cancer, apoptosis, oxidative stress, mitochondrial function, phagocytosis,
and gene transcription.
Calreticulin also plays an important role in control of calcium homeosta-
sis. Calreticulin‐deficient cells display impaired agonist‐stimulated calcium
release from ER stores by bradykinin, which may be in part due to a failure
of the bradykinin to interact with its surface receptor and increase inositol
1,4,5‐triphosphate (InsP3) (Knee et al., 2003). Calreticulin‐deficient cells alsolack the transient rise in calcium from outside the cell that normally accom-
panies engagement of integrins during cell adhesion (Molinari et al., 2004).
However, there are conflicting reports on the effect of calreticulin on ER
calcium storage. Calreticulin‐deficient cells have a reduced ER calcium stor-
age, (Coppolino et al., 1997; Nakamura et al., 2001). Cells overexpressing
calreticulin had higher levels of ER calcium and a larger release of calcium
leading to larger cytosolic calcium levels. The mitochondrial calcium re-
sponse was shorter. There was no difference in calcium response in calnexin
overexpressing cells (Nakamura et al., 2001). Some studies have found no
effect of calreticulin deficiency on the amount of thapsigargin or InsP3-
sensitive ER stored calcium (Arnaudeau et al., 2002; Opas et al., 1996).
Cell shape, adhesion, and motility are controlled by a variety of pathways,
many of them calcium regulated. Alterations in the level of expression of
calreticulin indeed affect all the aforementioned cell properties (Fadel et al.,
1999, 2001; Opas et al., 1996). Calreticulin‐deficient cells have impaired
adhesion (Coppolino et al., 1997; Liu et al., 1994). It has been suggested
that this may be mediated by direct interaction between calreticulin and
oSiffroi‐Fernandez, S., Giraud, A., Lanet, J., and Franc, J. L. (2002). Association of the
thyrotropin receptor with calnexin, calreticulin and BiP. Effects on the maturation of the
receptor. Eur. J. Biochem. 269, 4930–4937.pLe et al. (1994).qTsukuba et al. (2002).rLenter and Vestweber (1994).sTate, C. G., Whiteley, E., and Betenbaugh, M. J. (1999). Molecular chaperones stimulate
the functional expression of the cocaine‐sensitive serotonin transporter. J. Biol. Chem. 274,
17551–17558.tRubio, M. E., and Wenthold, R. J. (1999). Calnexin and the immunoglobulin binding
protein (BiP) coimmunoprecipitate with AMPA receptors. J. Neurochem. 73, 942–948.uAllen et al. (2001).vChang, W., Gelman, M. S., and Prives, J. M. N. (1997). Calnexin‐dependent enhancement
of nicotinic acetylcholine receptor assembly and surface expression. J. Biol. Chem. 272, 28925–
28932.wWada, I., Imai, S., Kai, M., Sakane, F., and Kanoh, H. (1995). Chaperone function of
calreticulin when expressed in the endoplasmic reticulum as the membrane‐anchored and
soluble forms. J. Biol. Chem. 270, 20298–20304.xOtteken and Moss (1996).
CELLULAR FUNCTIONS OF ER CHAPERONES 103
Comp. by:bala Date:27/5/05 Time:20:26:48 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
104

Comp. by:bala Date:27/5/05 Time:20:26:48 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
KxGFFKR sequence of a‐integrins (Coppolino et al., 1995; Dedhar, 1994).
Consequently, to functionally affect integrins clustered in focal contacts,
calreticulin should be present in the cytoplasm, but there is no direct evidence
for this. Thus, calreticulin‐modulated changes in cell adhesiveness have to be
correlated with up‐regulation of adhesion‐specific proteins. Overexpression
of calreticulin increases both cell‐to‐substratum and cell‐to‐cell adhesiveness,and establishes vinculin‐rich cell‐to‐cell junctions by increasing overall vin-
culin levels in cells (Opas et al., 1996). Thus the adhesion‐related effects of
differential expression of calreticulin are vinculin mediated and the absence
from focal contacts or cytoplasm indicates that, in vivo, the adhesion‐relatedfunctions are performed from within the ER lumen. Also, overexpressed
cytoplasmically targeted GFP‐calreticulin did not localize in focal contacts
(M. Opas, unpublished data). Furthermore, targeting of calreticulin to the
cytoplasm either by microinjection or by expression of a leaderless calreticu-
lin had no effect on cell morphology, cytoskeleton, or cell adhesion
(M. Opas, unpublished data). Leung‐Hagesteijn et al. (1994) reported colo-
calization of antibody‐clustered integrins with calreticulin at the cytoplasmic
surface of carcinoma cells, but in normal cells no such colocalization was
found. Hence, it can be concluded that cytoplasmic calreticulin is both not
detectable and nonfunctional in terms of regulating cell adhesion. Most
importantly, studies show that both transcriptional activation by steroid
receptors and cell adhesion in vivo are affected only by the full‐length,ER‐targeted form of calreticulin but not by a truncated, cytosolically tar-
geted mutant protein (Fadel et al., 1999; Michalak et al., 1996; Opas et al.,
1996). A more recent report from Dedhar’s group postulates that calreticulin
may be involved in integrin‐dependent Ca signaling rather than direct regu-
lation of integrin activity (Coppolino et al., 1997). While this observation
requires further investigation, it is not inconsistent with the hypothesis that
calreticulin may function in adhesion as a ‘‘signaling’’ molecule from within
the ER lumen (Michalak et al., 2002; Papp et al., 2003).
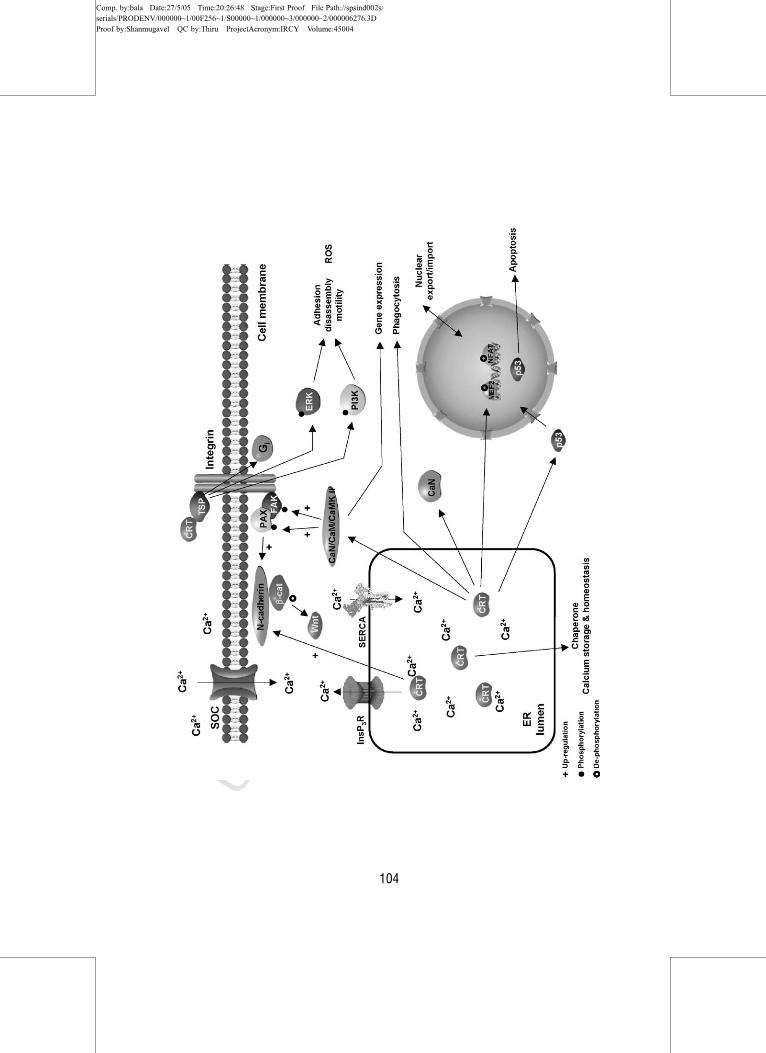
Overexpression of calreticulin also increases N‐cadherin levels and
decreases tyrosine phosphorylation of cellular proteins, such as b‐catenin(Fig. 4) (Fadel et al., 2001). b‐Catenin is a component of the cadherin‐mediated adhesion complexes and is also part of the Wnt signaling pathway
(Hutzfeld, 1999). Calreticulin from the ER influences tyrosine phosphoryla-
tion of b‐catenin but not its expression; b‐catenin is underphosphorylated in
calreticulin overexpressor cells, but protein and mRNA levels stay the same
FIG. 4 Calreticulin influences a number of adhesion‐related pathways, such as cadherin/b‐catenin, calmodulin/CaMK II, ERK, and PI3K pathways and steroid receptor‐mediated
pathways. It can also influence nuclear trafficking. TSP, thrombospondin; PAX, paxillin; FAK,
focal adhesion kinase; CaM, calmodulin; CRT, calreticulin; SOC, store‐operated channel.
CELLULAR FUNCTIONS OF ER CHAPERONES 105

Comp. by:bala Date:27/5/05 Time:20:26:48 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
in the cells (Fadel et al., 2001). Calreticulin can also affect cell adhesion
through the calmodulin and calcium‐mediated kinase (CaMK) II pathway.
Inhibition of calmodulin or CaMK II rescued the calreticulin under‐expressor phenotype by increased spreading (over 3‐fold) and increased
paxillin and focal adhesion kinase (FAK) phosphorylation and protein levels
(E. Szabo, unpublished data). FAK is a regulatory molecule that binds to
paxillin and in turn paxillin provides docking sites for FAK, src, and vinculin
targeting them to the focal contacts (Burridge and Chrzanowska‐Wodnicka,
1996; Turner, 1998). Paxillin levels are elevated when calreticulin is under-
expressed, which indicates that calreticulin influences paxillin levels, suggest-
ing that calreticulin function in the case of this focal adhesion protein is as a
chaperone (Fig. 4) (E. Szabo, unpublished data). Fibronectin levels are also
regulated by differential calreticulin levels. Overexpression of calreticulin
leads to increased fibronectin protein levels and fibronectin deposition,
explaining the increased spreading observed when calreticulin is overex-
pressed; the converse is observed for calreticulin underexpressing cells.
When calmodulin and the CaMK II pathway are inhibited, fibronectin
level and deposition increase and the calreticulin underexpressing cell pheno-
type is rescued (E. Szabo, unpublished data). When the calreticulin under-
expressor cells were plated on fibronectin‐coated substrata spreading of the
cells was induced. The cells overcame their poorly adhesive phenotype by
induction of many tensin‐rich fibrillar adhesions, thus compensating for
the paucity of vinculin in these cells. The calreticulin overexpressing cells
form vinculin‐rich focal contacts as opposed to tensin‐rich adhesions, since
vinculin levels are elevated in these cells (Fadel et al., submitted).
Extracellular calreticulin is involved in cellular adhesion and migration,
but its role is unclear, since it does not possess a transmembrane domain.
Extracellular calreticulin is a C1q coreceptor (Ghiran et al., 2003; McGreal
and Gasque, 2001), which complexes with CD91 on phagocytes for apoptotic
cell ingestion (Basu et al., 2001; Ogden et al., 2001; Vandivier et al., 2002), has
antithrombotic effects (Dai et al., 1997; Kuwabara et al., 1995), inhibits
melanoma cell spreading (White et al., 1995; Zhu et al., 1997), and inhibits
angiogenesis (Pike et al., 1998, 1999); however, the mechanisms behind these
phenomena are unknown. It was recently shown that thrombospondin med-
iates the disassembly of focal contacts by interacting with cell surface calre-
ticulin (Fig. 4) (Goicoechea et al., 2000). The thrombospondin‐binding site
was mapped to the N‐domain of calreticulin (Goicoechea et al., 2002) and
biochemical evidence indicates the presence of a calreticulin complex with
low‐density lipoprotein receptor‐related protein at the cell surface (Orr et al.,
2003). Thrombospondin stimulates focal adhesion disassembly and motility
through the heparin‐binding domain, hep I, which binds to the calreticulin
and leads to phosphoinositide 3‐kinase (PI3K) activation, and stimulation of
extracellular signal‐regulated kinase (ERK) and Gi protein systems (Fig. 4)
AU3
AU4
AU5
106 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:48 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
(Orr et al., 2002, 2003). The sources of extracellular calreticulin have been
subject to speculation (Eggleton et al., 1997); nevertheless the serum level
of calreticulin in patients with systemic lupus erythematosus averages
4.44 mg/ml versus 0.42 mg/ml in control sera (Fukazawa, 1994).
Another indirect way that calreticulin may influence cell adhesion is
through its affects on the transmembrane influx of calcium via store‐operatedcalcium channels (SOCs), conceivably by controlling InsP3‐releasable calci-
um from the ER (Bastianutto et al., 1995; Fasolato et al., 1998; Mery et al.,
1996; Xu et al., 2000). Calreticulin also affects the function of SERCA2b and
the InsP3R (Camacho and Lechleiter, 1995; Jouaville et al., 1995), both of
which may be structurally coupled to SOCs (Lockwich et al., 2001). The
structural relationship between the SR/ER calcium release channels and
SOCs of the plasma membrane has been a matter of controversy, however,
substantial evidence points to structural coupling between the two channels
(Putney, 1999). In view of the heterogeneity of the SR/ER (Meldolesi and
Pozzan, 1998; Petersen et al., 2001), it is intuitive that not all of the ER is
coupled to the plasma membrane, however, morphological data supporting
this notion are scarce. Trp (a putative SOC component) reportedly associates
with InsP3R, SERCA, and caveolin in caveolar calcium signaling complexes
(Lockwich et al., 2000, 2001). Calreticulin was shown to coimmunoprecipi-
tate with caveolin (Darby et al., 2000). Caveolin coclusters with a1 integrinsand its down‐regulation inhibits a1 integrin‐mediated adhesion to fibronectin
(Wei et al., 1999). Furthermore, tyrosine phosphorylated caveolin has been
localized to focal contacts (Volonte et al., 2001). Finally, in fibroblasts, the
InsP3R was localized to focal contacts (Sugiyama et al., 2000) and pilot
TIRFM data showed localization of InsP3Rs to a subset of focal contacts
(M. Opas, unpublished data).
Although calreticulin mechanism(s) of action are still elusive, it is conceiv-
able that the affects observed on focal adhesion may be due to calreticulin
effects on multiple signaling pathways, which include cadherin/vinculin pro-
tein system, changes in tyrosine kinases and phosphatases, interaction with
the Wnt pathway, direct affect on the calmodulin/CaMK II pathway, inter-
action with InsP3 receptor‐mediated signaling, and steroid receptors on the
cell surface (Fig. 4).
Recent studies implicated that calreticulin functions in nucleocytoplasmic
transport, but it is not actually localized to the nucleus. Immunogold labeling
indicated that calreticulin is localized to the ER reticulum and to the nuclear
envelope (Huh and Yoo, 2003). Importantly, the nuclear envelope is contin-
uous with the ER and calcium‐mediated events have been implicated in
nucleocytoplasmic transport (Ashby and Tepikin, 2001; Bachs et al., 1992;
Berridge et al., 2000; Petersen et al., 1998; Santella and Carafoli, 1997).
Hence, it can be deduced that calreticulin would have a function in these
calcium‐mediated events. Calreticulin has been shown to be involved in
AU6
AU7
CELLULAR FUNCTIONS OF ER CHAPERONES 107

Comp. by:bala Date:27/5/05 Time:20:26:49 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
regulation of nuclear export and import of NFAT3 (Mesaeli et al., 1999) and
MEF2C (Li et al., 2002). Calreticulin has also been shown to be involved in
nuclear localization of p53. In calreticulin‐null cells p53 localization to the
nucleus and apoptosis rate are greatly reduced, indicating that calreticulin is
needed for p53 nuclear localization and for proper apoptotic function
(Mesaeli and Phillipson, 2004). It has been shown in vitro and in hetrokaryon
fusion assays that nuclear export of glucocorticoid receptors is mediated
through direct contact between calreticulin and the receptor DNA‐bindingdomain; moreover, the export of glucocorticoid receptors to the cytoplasm is
compromised in calreticulin‐deficient cells (Holaska et al., 2001, 2002).
Calnexin expression has also been linked to adhesion. Integrins associate
during their synthesis with calnexin (Coppolino et al., 1997). Adhesion of
cells stimulates expression of adhesion‐related proteins, including calnexin
(Opas et al., 1996). Breast cancer cells grown in suspension express less
calnexin compared to cells as a monolayer, where adhesion is possible
(Lenter and Vestweber, 1994). Expression of adhesion molecules CD44 and
LFA‐1 is lower in cells lacking calnexin (Lam et al., 2001).
Changes in cell adhesion properties are extremely important in the pro-
gression, invasion, and metastasis of cancerous tumors. The importance of
calreticulin and calnexin in the expression and function of adhesion proteins
may therefore make these chaperone proteins important players in cancer.
On the one hand, to be released to metastasize, the adhesion must be reduced.
On the other, to invade the new target, adhesions must be formed between
the circulating cancerous cell and the invaded tissue. Metastasis of tumors is
responsible for the vast majority of deaths associated with cancer. Calreti-
culin influences cell adhesion. Calreticulin is also strongly induced in colon
cancer, where it is found in the nuclear matrix (Yeates and Powis, 1997).
Abnormalities in the expression and functional activity of cell adhesion
molecules are implicated in the development and progression of the majority
of colorectal cancers (Malyguine et al., 1998). Similarly, calnexin, which can
also influence adhesion, is increased in the progressive stages of breast cancer
(Brunagel et al., 2003). On the other hand, calnexin expression is decreased in
metastatic stages of melanoma compared to primary stages (Buda and
Pignatelli, 2004). Calnexin expression is also decreased in colon and breast
cancer cells grown in suspension compared to those adhered to a plate (Li
et al., 2001). This is consistent with the idea that calreticulin and calnexin
expression can influence adhesion and thus affect tumor invasion and metas-
tasis. Extracellular calreticulin may be especially important in affecting
adhesive phenotype by contributing to deadhesion or, in other words, main-
tenance of the intermediate adhesive state of a cell (Murphy‐Ullrich, 2001)
(Fig. 4).
One of the ways the body protects itself from developing cancers is to
initiate apoptosis in cells with abnormalities. The tumor suppressor protein
108 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:49 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
p53 is a transcription factor involved in inducing apoptosis in potentially
damaged cells. p53 induces expression of genes that contain the p53 binding
site and represses those that do not. Calreticulin‐deficient cells had a reduced
level of p53, and a reduced ability to induce p53 in response to DNA damage
(Dissemond et al., 2004). This is consistent with the finding that overexpres-
sion of calreticulin results in an increased release of cytochrome c from the
mitochondria and an increased sensitivity to thapsigargin‐ and staurosporin‐induced apoptosis, while cells deficient in calreticulin had a decrease in
cytochrome c release and caspase 3 activity, and were more resistant to
apoptosis (Yeates and Powis, 1997). Overexpression of calnexin had no effect
on drug‐induced apoptosis (Mesaeli and Phillipson, 2004). Calnexin‐deficientcells, however, are relatively resistant to apoptosis (Nakamura et al., 2000).
Oxidative stress can lead to either apoptotic or necrotic cell death. Reac-
tive oxygen species (ROS) are produced normally during the production of
ATP and are managed effectively by antioxidant systems such as glutathione,
superoxide dismutase, catalase, and other cellular components. However,
excessive generation of ROS, or impaired protective mechanisms, can lead
to toxicity. ROS are thought to play an important role in a wide range of
pathologies including Alzheimer’s disease, Parkinson’s disease, stroke, and
aging. Oxidative stress can occur from sustained ER stress (Nakamura et al.,
2000). Calnexin‐ and calreticulin‐deficient cells show evidence of existing in a
state of sustained ER stress, and further showed an impaired ability to
respond to further ER stress by inducing protective chaperones such as
GRP78. Oxidative stress from ER stress can derive from two sources, the
oxidative process of bond formation as the ER deals with accumulated
proteins, and also through mitochondrial reactive oxygen species (ROS)
production (Zuppini et al., 2002). The lumen of the ER has an oxidizing
environment with a reduced glutathione (GSH) to glutathione disulfide
(GSSG) ratio of 2:1 compared to 30:1 to 100:1 found in the cytosol (Haynes
et al., 2004). Disruption of the ER luminal environment therefore can alter
the oxidative state of the cell. The mitochondrial contribution of ROS after
ER stress could be either due to depletion of GSH due to the demand of the
ER, thereby reducing the antioxidant capacity of the cell, or alternatively by
signaling from the ER to the mitochondria (Haynes et al., 2004). ER stress
has been shown to impact on mitochondrial‐associated proteins (Noiva,
1999). Another mechanism by which calnexin and calreticulin can alter the
level of oxidative stress is through their influence on the production of
protective proteins. Tyrosinase is an enzyme associated with the production
of the protein melanin. Mutations in tyrosinase lead to the prolonged inter-
action of the enzymes with calnexin and calreticulin, and results in albinism
(Merad‐Boudia et al., 1998). Abnormal calnexin association with tyrosinase‐related protein can also lead to increased sensitivity to oxidative stress (Hori
et al., 2002). It is further hypothesized that calnexin may be involved in
CELLULAR FUNCTIONS OF ER CHAPERONES 109

Comp. by:bala Date:27/5/05 Time:20:26:49 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
signaling to increase production of oxidative stress relief proteins (Halaban
et al., 2000).
Phagocytosis involves the uptake of microorganisms, damaged or dead
cells, cell debris, or insoluble circulating particles. It involves attraction of
phagocytic cells, adhesion, then protrusion of the membrane to encompass
and take in the particle. In macrophages, ER chaperones including calreti-
culin and calnexin were targeted to the phagocytic cup (Jimbow et al., 2001).
The results do not indicate whether the phagocytosis is dependent on these
chaperones or if they just mark the proximity of the ER to the phagocytic
cup. However, in Dictyostelium, the only microorganism known to contain
both calnexin and calreticulin, gene replacement of both of these genes led to
an impairment in phagocytic ability. Immunofluorescence demonstrated that
the ER comes in direct contact with the phagocytic cup (Vinayagamoorthy
and Rajakumar, 1996).
The main role of calnexin appears not to be related to assisting in the
expression of proteins, as calnexin‐deficient mice and cells appear to have
normal protein expression. Rather the literature suggests that its main func-
tion is binding to and retaining mutant proteins and directing them for
degradation. Examples of mutations in proteins leading to their prolonged
association with calnexin include mutations in the von Willebrand factor
resulting in a bleeding disorder (Muller‐Taubenberger et al., 2001), muta-
tions in the peripheral myelin protein pmp‐22 resulting in neuropathy
(Winrow et al., 1995), and mutations in tyrosinase resulting in albinism
(Allen et al., 2001).
Degradation of proteins involves communication from the ER to the
cytosolically located proteosome. It is possible mutations in membrane
spanning proteins may be detected in the cytosol, but many proteins synthe-
sized in the ER lack a membrane spanning domain. The cytoplasmic tail of
calnexin may provide communication about defective proteins from the ER
to the cytoplasm. Mutation of the secretory protein a1‐antitrypsin leads to its
retention in the ER associated with calnexin, followed by polyubiquitination
of calnexin (Dickson et al., 2002). Similar observations were made for the
mutations in the secretory protein meprin (Halaban et al., 2000).
A clinically significant example of calnexin retaining mutant proteins is
the �F508 mutation of the CFTR chloride channel responsible for 70%
of clinical cases of cystic fibrosis. In this disease, the deletion of an amino
acid leads to a decreased expression of the chloride channel at the cell surface,
as the channel is retained in the ER bound to calnexin. Interestingly, in
in vitro studies where the channel is permitted to move to the surface
by altering glycogen content (Qu et al., 1996), or temperature (Tsukuba
et al., 2002), the channel has some chloride flux activity. This opens the
possibility that alteration in calnexin function may have potential therapeutic
benefits.
110 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:49 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
There are other examples in which mutant proteins are retained in the ER
by calnexin. Mutations in cartilage oligomeric matrix protein (COMP) result
in early osteoarthritis. This COMP mutation results in retention of the
protein in the ER, and an increased expression of calnexin, suggesting the
mutated substrate may be retained by calnexin (Sato et al., 1996). There is a
prolonged interaction between calnexin and the ER‐retained mutated vaso-
pressin receptor, one of many mutations involved in nephrogenic diabetes
insipidus (Denning et al., 1992). Mutations in lysozyme enzymes can be
associated with systemic amyloidosis, a condition in which protein deposits
form in tissues from proteolytic fragments of serum amyloid. In calnexin
disrupted S. cerevisiae, the secretion of a mutant amyloidogenic lysozyme
was increased, while the mutant enzyme was retained by calnexin in wild‐type S. cerevisiae (Morello et al., 2001; Vranka et al., 2001). Amyloids can
be formed not only from deposits of serum amyloid fragments, but may be
made up of a number of plasma proteins, which have been transformed from
soluble proteins into insoluble fibrils. These formations occur in a number of
diseases including Alzheimer’s and prion diseases.
Calnexin appears to play a role in prion diseases. Prion proteins are N‐glycosolated proteins found abundantly in the brain, and normally transported
to the surface. In prion disease the prion protein accumulates in a different
physical state with reduced solubility and protease susceptibility. The disease
form of the prion protein has been shown to coimmunoprecipitate with
calnexin and to be retained in the ER.
IV. Concluding Remarks
The presence of each of the components involved in the processing of glycosy-
lated proteins through the ER is essential for the normal development of a
healthy animal. However, it is remarkable, given the large and diverse group of
surface and secreted proteins handled by this pathway, that development pro-
ceeds to the extent that it does. Studies on calreticulin have revealed that it is
involved in a wide variety of cellular functions outside of its role as a chaperone
(Johnson et al., 2001), and that these, and not protein folding, may be the
essential functions of this ER protein. Indeed, calreticulin affects important cell
functions such as adhesion via regulation of expression of proteins important in
adhesion, as well as via its effects on intracellular signaling pathways (Fig. 4).
Consequently, calreticulin knockout yields a lethal phenotype. The primary
function of calnexin, it appears, is in recognizing and retaining defective pro-
teins, but it is not required for the synthesis and expression of proteins. This is
supported by the finding that the absence of calnexin, a chaperone with fewer of
these extra functions, has a comparatively mild phenotype.
CELLULAR FUNCTIONS OF ER CHAPERONES 111

Comp. by:bala Date:27/5/05 Time:20:26:49 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Acknowledgments
This work was supported by grants from the CIHR (to M.M. and M.O.) and from the Heart
and Stroke Foundations of Ontario (to M.O.). M.M. is a CIHR Senior Investigator. M.O. is a
member of the Heart & Stroke/Richard Lewar Centre of Excellence.
References
Allen, S., Goodeve, A. C., Peake, I. R., and Daly, M. E. N. (2001). Endoplasmic reticulum
retention and prolonged association of a von Willebrand’s disease‐causing von Willebrand
factor variant with ERp57 and calnexin. Biochem. Biophys. Res. Commun. 280, 448–453.
Arnaudeau, S., Frieden, M., Nakamura, K., Castelbou, C., Michalak, M., and Demaurex, N.
(2002). Calreticulin differentially modulates calcium uptake and release in the endoplasmic
reticulum and mitochondria. J. Biol. Chem. 277, 46696–46705.
Arunachalam, B., and Cresswell, P. (1995). Molecular requirements for the interaction of class
II major histocompatibility complex molecules and invariant chain with calnexin. J. Biol.
Chem. 270, 2784–2790.
Ashby, M. C., and Tepikin, A. V. (2001). ER calcium and the functions of intracellular
organelles. Semin. Cell Dev. Biol. 12, 11–17.
Bachs, O., Agell, N., and Carafoli, E. (1992). Calcium and calmodulin function in the cell
nucleus. Biochim. Biophys. Acta Rev. Biomembr. 1113, 259–270.
Baksh, S., and Michalak, M. (1991). Expression of calreticulin in Escherichia coli and
identification of its Ca2þ binding domains. J. Biol. Chem. 266, 21458–21465.
Baksh, S., Burns, K., Andrin, C., and Michalak, M. (1995). Interaction of calreticulin with
protein disulfide isomerase. J. Biol. Chem. 270, 31338–31344.
Balakier, H., Dziak, E., Sojecki, A., Librach, C., Michalak, M., and Opas, M. (2002). Calcium‐binding proteins and calcium‐release channels in human maturing oocytes, pronuclear
zygotes and early preimplantation embryos. Hum. Reprod. 11, 2938–2947.
Bastianutto, C., Clementi, E., Codazzi, F., Podini, P., De Giorgi, F., Rizzuto, R., Meldolesi, J.,
and Pozzan, T. (1995). Overexpression of calreticulin increases the Ca2þ capacity of rapidly
exchanging Ca2þ stores and reveals aspects of their lumenal microenvironment and function.
J. Cell Biol. 130, 847–855.
Basu, S., Binder, R. J., Ramalingam, T., and Srivastava, P. K. (2001). CD91 is a common
receptor for heat shock proteins gp96, hsp90, hsp70, and calreticulin. Immunity 14, 303–313.
Berridge, M. J., Lipp, P., and Bootman, M. D. (2000). The versatility and universality of
calcium signalling. Nat. Rev. Mol. Cell. Biol. 1, 11–21.
Brunagel, G., Shah, U., Schoen, R. E., and Getzenberg, R. H. (2003). Identification of
calreticulin as a nuclear matrix protein associated with human colon cancer. J. Cell. Biochem.
89, 238–243.
Buda, A., and Pignatelli, M. (2004). Cytoskeletal network in colon cancer: From genes to
clinical application. Int. J. Biochem. Cell Biol. 36, 759–765.
Camacho, P., and Lechleiter, J. D. (1995). Calreticulin inhibits repetitive intracellular Ca2þwaves. Cell 82, 765–771.
Capps, G. G., and Zuniga, M. C. (1994). Class I histocompatibility molecule association with
phosphorylated calnexin. Implications for rates of intracellular transport. J. Biol. Chem. 269,
11634–11639.
Carreno, B. M., Schreiber, K. L., McKean, D. J., Stroynowski, I., and Hansen, T. H. (1995).
Aglycosylated and phosphatidylinositol‐anchored MHC class I molecules are associated with
112 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:50 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
calnexin. Evidence implicating the class I‐connecting peptide segment in calnexin association.
J. Immunol. 154, 5173–5180.
Chen, W., Helenius, J., Braakman, I., and Helenius, A. (1995). Cotranslational folding and
calnexin binding during glycoprotein synthesis. Proc. Natl. Acad. Sci. USA 92, 6229–6233.
Chevet, E., Wong, H. N., Gerber, D., Cochet, C., Fazel, A., Cameron, P. H., Gushue, J. N.,
Thomas, D. Y., and Bergeron, J. J. (1999). Phosphorylation by CK2 and MAPK enhances
calnexin association with ribosomes. EMBO J. 18, 3655–3666.
Coppolino, M., Leung‐Hagesteijn, C., Dedhar, S., and Wilkins, J. (1995). Inducible interaction
of integrin a2b1 with calreticulin‐dependence on the activation state of the integrin. J. Biol.
Chem. 270, 23132–23138.
Coppolino, M. G., Woodside, M. J., Demaurex, N., Grinstein, S., St‐Arnaud, R., and Dedhar,
S. (1997). Calreticulin is essential for integrin‐mediated calcium signalling and cell adhesion.
Nature 386, 843–847.
Cresswell, P. (2000). Intracellular surveillance: Controlling the assembly of MHC class I‐peptidecomplexes. Traffic 1, 301–305.
Dai, E., Stewart, M., Ritchie, B., Mesaeli, N., Raha, S., Kolodziejczyk, D., Hobman, M. L.,
Liu, L. Y., Etches, W., Nation, N., Michalak, M., and Lucas, A. (1997). Calreticulin, a
potential vascular regulatory protein, reduces intimal hyperplasia after arterial injury.
Arterioscler. Thromb. Vasc. Biol. 17, 2359–2368.
D’Alessio, C., Fernandez, F., Trombetta, E. S., and Parodi, A. J. (1999). Genetic evidence for
the heterodimeric structure of glucosidase II. The effect of disrupting the subunit‐encodinggenes on glycoprotein folding. J. Biol. Chem. 274, 25899–25905.
Darby, P. J., Kwan, C. Y., and Daniel, E. E. (2000). Caveolae from canine airway smooth
muscle contain the necessary components for a role in Ca(2þ) handling. Am. J. Physiol. Lung
Cell Mol. Physiol. 279, L1226–L1235.
Dedhar, S. (1994). Novel functions of calreticulin: Interaction with integrins and modulation of
gene expression. Trends Biochem. Sci. 19, 269–271.
Denning, G. M., Anderson, M. P., Amara, J. F., Marshall, J., Smith, A. E., and Welsh, M. J.
(1992). Processing of mutant cystic fibrosis transmembrane conductance regulator is
temperature‐sensitive. Nature 358, 761–764.
Denzel, A., Molinari, M., Trigueros, C., Martin, J. E., Velmurgan, S., Brown, S., Stamp, G.,
and Owen, M. J. (2002). Early postnatal death and motor disorders in mice congenitally
deficient in calnexin expression. Mol. Cell. Biol. 22, 7398–7404.
Dickson, K. M., Bergeron, J. J., Shames, I., Colby, J., Nguyen, D. T., Chevet, E., Thomas,
D. Y., and Snipes, G. J. (2002). Association of calnexin with mutant peripheral myelin
protein‐22 ex vivo: A basis for ‘‘gain‐of‐function’’ ER diseases. Proc. Natl. Acad. Sci. USA 99,
9852–9857.
Diedrich, G., Bangia, N., Pan, M., and Cresswell, P. (2001). A role for calnexin in the assembly
of the MHC class I loading complex in the endoplasmic reticulum. J. Immunol. 166,
1703–1709.
Di Jeso, B., Ulianich, L., Pacifico, F., Leonardi, A., Vito, P., Consiglio, A., Formisano, S., and
Arvan, P. (2003). Folding of thyroglobulin in the clanexin/calreticulin pathway and its
alteration by loss of Ca2þ from the endoplasmic reticulum. Biochem. J. 370, 449–458.
Dissemond, J., Busch, M., Kothen, T., Mors, J., Weimann, T. K., Lindeke, A., Goos, M., and
Wagner, S. N. (2004). Differential downregulation of endoplasmic reticulum‐residingchaperones calnexin and calreticulin in human metastatic melanoma. Cancer Lett. 203,
225–231.
Drickamer, K., and Taylor, M. E. (1998). Evolving views of protein glycosylation. Trends
Biochem. Sci. 23, 321–324.
Dwek, R. A. (1996). Glycobiology: Toward understanding the function of sugars. Chem. Rev.
96, 683–720.
CELLULAR FUNCTIONS OF ER CHAPERONES 113

Comp. by:bala Date:27/5/05 Time:20:26:50 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Eggleton, P. (2003). Stress protein‐polypeptide complexes acting as autoimmune triggers. Clin.
Exp. Immunol. 134, 6–8.
Eggleton, P., Reid, K. B. M., Kishore, U., and Sontheimer, R. D. (1997). Clinical relevance of
calreticulin in systemic lupus erythematosus. Lupus 6, 564–571.
Elliott, J. G., Oliver, J. D., and High, S. (1997). The thiol‐dependent reductase ERp57 interacts
specifically with N‐glycosylated integral membrane proteins. J. Biol. Chem. 272,
13849–13855.
Fadel, M. P., Dziak, E., Lo, C. M., Ferrier, J., Mesaeli, N., Michalak, M., and Opas, M. (1999).
Calreticulin affects focal contact‐dependent but not close contact‐dependent cell‐substratumadhesion. J. Biol. Chem. 274, 15085–15094.
Fadel, M. P., Szewczenko‐Pawlikowski, M., Leclerc, P., Dziak, E., Symonds, J. M., Blaschuk,
O., Michalak, M., and Opas, M. (2001). Calreticulin affects beta‐catenin associated
pathways. J. Biol. Chem. 276, 27083–27089.
Fadel, M. P., Szewczenki‐Pawlirowski, M., Papp, S., Michalak, M., and Opas, M. (2004).
Calreticulin affects expression and deposition of fibrone ctin.
Fanchiotti, S., Fernandez, F., D’Alessio, C., and Parodi, A. J. (1998). The UDP‐Glc:
Glycoprotein glucosyltransferase is essential for Schizosaccharomyces pombe viability under
conditions of extreme endoplasmic reticulum stress. J. Cell. Biol. 143, 625–635.
Fasolato, C., Pizzo, P., and Pozzan, T. (1998). Delayed activation of the store‐operated calcium
current induced by calreticulin overexpression in RBL‐1 cells. Mol. Biol. Cell 9, 1513–1522.
Ghiran, I., Klickstein, L. B., and Nicholson‐Weller, A. (2003). Calreticulin is at the surface of
circulating neutrophils and uses CD59 as an adaptor molecule. J. Biol. Chem. 278,
21024–21031.
Goicoechea, S., Orr, A. W., Pallero, M. A., Eggleton, P., and Murphy‐Ullrich, J. E. (2000).
Thrombospondin mediates focal adhesion disassembly through interactions with cell surface
calreticulin. J. Biol. Chem. 275, 36358–36368.
Goicoechea, S., Pallero, M. A., Eggleton, P., Michalak, M., and Murphy‐Ullrich, J. E. (2002).
The anti‐adhesive activity of thrombospondin is mediated by the N‐terminal domain of cell
surface calreticulin. J. Biol. Chem. 277, 37219–37228.
Guo, L., Nakamura, K., Lynch, J., Opas, M., Olson, E. N., Agellon, L. B., and Michalak, M.
(2002). Cardiac‐specific expression of calcineurin reverses embryonic lethality in calreticulin‐deficient mouse. J. Biol. Chem. 277, 50776–50779.
Halaban, R., Svedine, S., Cheng, E., Smicun, Y., Aron, R., and Hebert, D. N. (2000).
Endoplasmic reticulum retention is a common defect associated with tyrosinase‐negativealbinism. Proc. Natl. Acad. Sci. USA 97, 5889–5894.
Hammond, C., Braakman, I., and Helenius, A. (1994). Role of N‐linked oligosaccharide
recognition, glucose trimming, and calnexin in glycoprotein folding and quality control. Proc.
Natl. Acad. Sci. USA 91, 913–917.
Hardingham, G. E., and Bading, H. (1999). Calcium as a versatile second messenger in the
control of gene expression. Microsc. Res. Tech. 46, 348–355.
Hardingham, G. E., Chawla, S., and Johnson, C. M. (1997). Distinct functions of nuclear and
cytoplasmic calcium in the control of gene expression. Nature 385, 260–265.
Haynes, C. M., Titus, E. A., and Cooper, A. A. (2004). Degradation of misfolded proteins
prevents ER‐derived oxidative stress and cell death. Mol. Cell 15, 767–776.
Hebert, D. N., Foellmer, B., and Helenius, A. (1996). Calnexin and calreticulin promote
folding, delay oligomerization and suppress degradation of influenza hemaglutinin in
microsomes. EMBO J. 15, 2961–2968.
Helenius, A. (1994). How N‐linked oligosaccharides affect glycoprotein folding in the
endoplasmic reticulum. Mol. Biol. Cell. 5, 253–265.
Helenius, A., and Aebi, M. N. (2001). Intracellular functions of N‐linked glycans. Science 291,
2364–2369.
AU8
114 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:50 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Helenius, A., and Aebi, M. (2004). Roles of N‐linked glycans in the endoplasmic reticulum.
Annu. Rev. Biochem. 73, 1019–1049.
Holaska, J. M., Black, B. E., Love, D. C., Hanover, J. A., Leszyk, J., and Paschal, B. M. (2001).
Calreticulin is a receptor for nuclear export. J. Cell. Biol. 152, 127–140.
Holaska, J. M., Black, B. E., Rastinejad, F., and Paschal, B. M. (2002). Ca‐dependent nuclearexport mediated by calreticulin. Mol. Cell. Biol. 22, 6286–6297.
Hori, O., Ichinoda, F., Tamatani, T., Yamaguchi, A., Sato, N., Ozawa, K., Kitao, Y.,
Miyazaki, M., Harding, H. P., Ron, D., Tohyama, M., M. Stern, D., and Ogawa, S. (2002).
Transmission of cell stress from endoplasmic reticulum to mitochondria: Enhanced
expression of Lon protease. J. Cell. Biol. 157, 1151–1160.
Huh, Y. H., and Yoo, S. H. (2003). Presence of the inositol 1,4,5‐triphosphate receptor isoforms
in the nucleoplasm. FEBS Lett. 555, 411–418.
Hutzfeld, M. (1999). The armadillo family of structural proteins. Int. Rev. Cytol. 186, 179–224.
Ihara, Y., Cohen‐Doyle, M. F., Saito, Y., and Williams, D. B. (1999). Calnexin discriminates
between protein conformational states and functions as a molecular chaperone in vitro. Mol.
Cell. 4, 331–341.
Jackson, M. R., Cohen‐Doyle, M. F., Peterson, P. A., and Williams, D. B. (1994). Regulation
of MHC class I transport by the molecular chaperone, calnexin (p88, IP90). Science 263,
384–387.
Jakob, C. A., Burda, P., Roth, J., and Aebi, M. (1998). Degradation of misfolded endoplasmic
reticulum glycoproteins in Saccharomyces cerevisiae is determined by a specific oligosaccha-
ride structure. J. Cell. Biol. 142, 1223–1233.
Jethmalani, S. M., Henle, K. J., Gazitt, Y., Walker, P. D., and Wang, S. Y. (1997). Intracellular
distribution of heat‐induced stress glycoproteins. J. Cell. Biochem. 66, 98–111.
Jimbow, K., Chen, H., Park, J. S., and Thomas, P. D. (2001). Increased sensitivity of
melanocytes to oxidative stress and abnormal expression of tyrosinase‐related protein in
vitiligo. Br. J. Dermatol. 144, 55–65.
Johnson, S., Michalak, M., Opas, M., and Eggleton, P. (2001). The ins and outs of calreticulin:
From the ER lumen to the extracellular space. Trends Cell Biol. 11, 122–129.
Jouaville, L. S., Ichas, F., Holmuhamedov, E. L., Camacho, P., and Lechleiter, J. D. (1995).
Synchronization of calcium waves by mitochondrial substrates in Xenopus laevis oocytes.
Nature 377, 438–441.
Kearse, K. P., Williams, D. B., and Singer, A. (1994). Persistence of glucose residues on core
oligosaccharides prevents association of TCR alpha and TCR beta proteins with calnexin and
results specifically in accelerated degradation of nascent TCR alpha proteins within the
endoplasmic reticulum. EMBO. J. 13, 3678–3686.
Kishore, U., Sontheimer, R. D., Sastry, K. N., Zappi, E. G., Hughes, G. R. V., Khamashta,
M. A., Reid, K. B. M., and Eggleton, P. (1997). The systemic lupus erythematosus (SLE)
disease autoantigen—Calreticulin can inhibit C1q association with immune complexes. Clin.
Exp. Immunol. 108, 181–190.
Knee, R., Ahsan, I., Mesaeli, N., Kaufman, R. J., and Michalak, M. (2003). Compromised
calnexin function in calreticulin‐deficient cells. Biochem. Biophys. Res. Commun. 304,
661–666.
Kuwabara, K., Pinsky, D. J., Schmidt, A. M., Benedict, C., Brett, J., Ogawa, S., Broekman,
M. J., Marcus, A. J., Sciacca, R. R., Michalak,M., Wang, F., Pan, Y. C., Grunfeld, S., Patton,
S., Malinski, T., Stern, D. M., and Ryan, J. (1995). Calreticulin, an antithrombotic agent
which binds to vitamin K‐dependent coagulation factors, stimulates endothelial nitric oxide
production, and limits thrombosis in canine coronary arteries. J. Biol. Chem. 270, 8179–8187.
Lam, K., Zhang, L., Yamada, K. M., and Lafrenie, R. M. (2001). Adhesion of epithelial cells to
fibronectin or collagen I induces alterations in gene expression via a protein kinase
C‐dependent mechanism. J. Cell Physiol. 189, 79–90.
CELLULAR FUNCTIONS OF ER CHAPERONES 115

Comp. by:bala Date:27/5/05 Time:20:26:50 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Le, A., Steiner, J. L., Ferrell, G. A., Shaker, J. C., and Sifers, R. N. (1994). Association between
calnexin and a secretion‐incompetent variant of human alpha 1‐antitrypsin. J. Biol. Chem.
269, 7514–7519.
Lenter, M., and Vestweber, D. (1994). The integrin chains beta 1 and alpha 6 associate with the
chaperone calnexin prior to integrin assembly. J. Biol. Chem. 269, 12263–12268.
Leung‐Hagesteijn, C. Y., Milankov, K., Michalak, M., Wilkins, J., and Dedhar, S. (1994). Cell
attachment to extracellular matrix substrates is inhibited upon downregulation of expression
of calreticulin, an intracellular integrin‐subunit‐binding protein. J. Cell Sci. 107, 589–600.
Li, F., Mandal, M., Barnes, C. J., Vadlamudi, R. K., and Kumar, R. (2001). Growth factor
regulation of the molecular chaperone calnexin. Biochem. Biophys. Res. Commmun. 289,
725–732.
Li, J., Puceat, M., Perez‐Terzic, C., Merry, A., Nakamura, K., Michalak, M., Krause, K. H.,
and Jaconi, M. E. (2002). Calreticulin reveals a critical Ca2þ checkpoint in cardiac
myofibrillogenesis. J. Cell Biol. 158, 103–113.
Li, Y., and Camacho, P. (2004). Ca2þ‐dependent redox modulation of SERCA 2b by ERp57.
J. Cell Biol. 164, 35–46.
Liu, N., Fine, R. E., Simons, E., and Johnson, R. J. (1994). Decreasing calreticulin expression
lowers the Ca2þ response to bradykinin and increases sensitivity to ionomycin in NG‐108–15cells. J. Biol. Chem. 269, 28635–28639.
Liu, Y., Choudhury, P., Cabral, C. M., and Sifers, R. N. (1999). Oligosaccharide modification
in the early secretory pathway directs the selection of a misfolded glycoprotein for
degradation by the proteasome. J. Biol. Chem. 274, 5861–5867.
Lockwich, T. P., Liu, X., Singh, B. B., Jadlowiec, J., Weiland, S., and Ambudkar, I. S. (2000).
Assembly of Trp1 in a signaling complex associated with caveolin‐scaffolding lipid raft
domains. J. Biol. Chem. 275, 11934–11942.
Lockwich, T. P., Singh, B. B., Liu, X., and Ambudkar, I. S. (2001). Stabilization of cortical
actin induces internalization of Trp3–associated caveolar Ca2þ signaling complex and loss of
Ca2þ influx without disruption of Trp3–IP3R association. J. Biol. Chem. 276, 42401–42408.
Lodish, H. F., and Kong, N. (1984). Glucose removal from N‐linked oligosaccharides is
required for efficient maturation of certain secretory glycoproteins from the rough
endoplasmic reticulum to the Golgi complex. J. Cell Biol. 98, 1720–1729.
Lynch, J., and Michalak, M. (2003). Calreticulin is an upstream regulator of calcineurin.
Biochem. Biophys. Res. Commun. 311, 1173–1179.
Macias, M., Escames, G., Leon, J., Coto, A., Sbihi, Y., Osuna, A., and Acuna‐Castroviejo, D.
(2003). Calreticulin‐melatonin. An unexpected relationship. Eur. J. Biochem. 270, 832–840.
Malyguine, A. M., Scott, J. E., and Dawson, J. R. (1998). The role of calnexin in NK‐target cellinteraction. Immunol. Lett. 61, 67–71.
Margolese, L., Waneck, G. L., Suzuki, C. K., Degen, E., Flavell, R. A., and Williams, D. B.
(1993). Identification of the region on the class I histocompatibility molecule that interacts
with the molecular chaperone, p88 (calnexin, IP90). J. Biol. Chem. 268, 17959–17966.
McGreal, E., and Gasque, P. (2001). Structure‐function studies of the receptors for complement
C1q. Biochem. Soc. Trans 30, 1010–1014.
Meldolesi, J., and Pozzan, T. (1998). The heterogeneity of ER Ca2þ stores has a key role in
nonmuscle cell signaling and function. J. Cell Biol. 142, 1395–1398.
Merad‐Boudia, M., Nicole, A., Santiard‐Baron, D., Saille, C., and Ceballos‐Picot, I. (1998).Mitochondrial impairment as an early event in the process of apoptosis induced by
glutathione depletion in neuronal cells: Relevance to Parkinson’s disease. Biochem.
Pharmacol. 56, 645–655.
Mery, L., Mesaeli, N., Michalak, M., Opas, M., Lew, D. P., and Krause, K.‐H. (1996).
Overexpression of calreticulin increases intracellular Ca2þ‐storage and decreases store‐operated Ca2þ influx. J. Biol. Chem. 271, 9332–9339.
116 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:51 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Mesaeli, N., and Phillipson, C. (2004). Impaired p53 expression, function, and nuclear
localization in calreticulin‐deficient cells. Mol. Biol. Cell 15, 1862–1870.
Mesaeli, N., Nakamura, K., Zvaritch, E., Dickie, P., Dziak, E., Krause, K.‐H., Opas, M.,
MacLennan, D. H., and Michalak, M. (1999). Calreticulin is essential for cardiac
development. J. Cell Biol. 144, 857–868.
Michalak, M., Burns, K., Mesaeli, N., Andrin, C., Busaan, L. J., Jass, G. H., and Opas, M.
(1996). Endoplasmic reticulum form of calreticulin modulates glucocorticoid‐sensitive gene
expression. J. Biol. Chem. 271, 29436–26445.
Michalak, M., Parker, J. M. R., and Opas, M. (2002). Ca2þ signaling and calcium binding
chaperones of the endoplasmic reticulum. Cell Calcium 32, 269–278.
Molinari, M., and Helenius, A. (1999). Glycoproteins form mixed disulphides with
oxidoreductases during folding in living cells. Nature 402, 90–93.
Molinari, M., Calanca, V., Galli, C., Lucca, P., and Paganetti, P. (2003). Role of EDEM in the
release of misfolded glycoproteins from the calnexin cycle. Science 299, 1397–1400.
Molinari, M., Eriksson, K. K., Calanca, V., Galli, C., Cresswell, P., Michalak, M., and
Helenius, A. (2004). Contrasting functions of calreticulin and calnexin in glycoprotein folding
and ER quality control. Mol. Cell 13, 125–135.
Moore, E. D., Etter, E. F., Philipson, K. D., Carrington, W. A., Fogarty, K. E., Lifshitz, L. M.,
and Fay, F. S. (1993). Coupling of the Naþ/Ca2þ exchanger, Naþ/Kþ pump and sarcoplasmic
reticulum in smooth muscle. Nature 365, 657–660.
Morello, J. P., Salahpour, A., Petaja‐Repo, U. E., Laperriere, A., Lonergan, M., Arthus, I. R.,
Nabi, M. F., Bichet, D. G., and Bouvier, M. (2001). Association of calnexin with wild type
and mutant AVPR2 that causes nephrogenic diabetes insipidus. Biochemistry 40, 6766–6775.
Muller‐Taubenberger, A., Lupas, A. N., Li, H., Ecke, M., Simmeth, E., and Gerisch, G. (2001).
Calreticulin and calnexin in the endoplasmic reticulum are important for phagocytosis.
EMBO J. 20, 6772–6782.
Murphy‐Ullrich, J. E. (2001). The de‐adhesive activity of matricellular proteins: Is intermediate
cell adhesion an adaptive state? J. Clin. Invest. 107, 785–790.
Nair, S., Wearsch, P. A., Mitchell, D. A., Wassenberg, J. J., Gilboa, E., and Nicchitta, C. V.
(1999). Calreticulin displays in vivo peptide‐binding activity and can elicit CTL response
against bound peptide. J. Immunol. 162, 6426–6432.
Nakamura, K., Bossy‐Wetzel, E., Burns, K., Fadel, M. P., Lozyk, M., Goping, I. S., Opas, M.,
Bleackley, R. C., Green, D. R., and Michalak, M. (2000). Changes in endoplasmic reticulum
luminal environment affect cell sensitivity to apoptosis. J. Cell Biol. 150, 731–740.
Nakamura, K., Robertson, M., Liu, G., Dickie, P., Guo, J. Q., Duff, H. J., Opas, M.,
Kavanagh, K., and Michalak, M. (2001a). Complete heart block and sudden death in mice
overexpressing calreticulin. J. Clin. Invest. 107, 1245–1253.
Nakamura, K., Zuppini, A., Arnaudeau, S., Lynch, J., Ahsan, I., Krause, R., Papp, S., De
Smedt, H., Parys, J. B., Muller‐Esterl, W., Lew, D. P., Krause, K.‐H., Demaurex, N., Opas,
M., and Michalak, M. (2001b). Functional specialization of calreticulin domains. J. Cell Biol.
154, 961–972.
Nauseef, W. M., McCormick, S. J., and Clark, R. A. (1995). Calreticulin functions
as a molecular chaperone in the biosynthesis of myeloperoxidase. J. Biol. Chem. 270,
4741–4747.
Nigam, S. K., Goldberg, A. L., Ho, S., Rohde, M. F., Bush, K. T., and Sherman, M. (1994). A
set of endoplasmic reticulum proteins possessing properties of molecular chaperones includes
Ca(2þ)‐binding proteins and members of the thioredoxin superfamily. J. Biol. Chem. 269,
1744–1749.
Noiva, R. (1999). Protein disulfide isomerase: The multifunctional redox chaperone of the
endoplasmic reticulum. Semin. Cell Dev. Biol. 10, 481–493.
CELLULAR FUNCTIONS OF ER CHAPERONES 117

Comp. by:bala Date:27/5/05 Time:20:26:51 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
O’Connor, S. E., and Imperiali, B. (1996). Modulation of protein structure and function by
asparagine‐linked glycosylation. Chem. Biol. 3, 803–812.
Oda, Y., Hosokawa, N., Wada, I., and Nagata, K. (2003). EDEM as an acceptor of terminally
misfolded glycoproteins released from calnexin. Science 299, 1394–1397.
Ogden, C. A., deCathelineau, A., Hoffmann, P. R., Bratton, D., Ghebrehiwet, B., Fadok,
V. A., and Henson, P. M. (2001). C1q and mannose binding lectin engagement of cell surface
calreticulin and cd91 initiates macropinocytosis and uptake of apoptotic cells. J. Exp. Med
194, 781–796.
Okazaki, Y., Ohno, H., Takase, K., Ochiai, T., and Saito, T. (2000). Cell surface expression of
calnexin, a molecular chaperone in the endoplasmic reticulum. J. Biol. Chem. 275,
35751–35758.
Opas, M., Szewczenko‐Pawlikowski, M., Jass, G. K., Mesaeli, N., and Michalak, M. (1996).
Calreticulin modulates cell adhesiveness via regulation of vinculin expression. J. Cell Biol.
135, 1913–1923.
Orr, A. W., Pallero, M. A., and Murphy‐Ullrich, J. E. (2002). Thrombospondin stimulates focal
adhesion disassembly through Gi‐ and phosphoinositide 3–kinase‐dependent ERK activa-
tion. J. Biol. Chem. 277, 20453–20460.
Orr, A. W., Pedraza, C. E., Pallero, M. A., Elzie, C. A., Goicoechea, S., Strickland, D. K., and
Murphy‐Ullrich, J. E. (2003a). Low density lipoprotein receptor‐related protein is a
calreticulin coreceptor that signals focal adhesion disassembly. J. Cell Biol. 161, 1179–1189.
Orr, A. W., Elzie, C. A., Kucik, D. F., and Murphy‐Ullrich, J. E. (2003b). Thrombospondin
signaling through the calreticulin/LDL receptor‐related protein co‐complex stimulates
random and directed cell migration. J. Cell Sci. 116, 2917–2927.
Otteken, A., and Moss, B. (1996). Calreticulin interacts with newly synthesized human
immunodeficiency virus type 1 envelope glycoprotein, suggesting a chaperone function
similar to that of calnexin. J. Biol. Chem. 271, 97–103.
Ou, W. J., Cameron, P. H., Thomas, D. Y., and Bergeron, J. J. (1993). Association of folding
intermediates of glycoproteins with calnexin during protein maturation. Nature 364, 771–776.
Papp, S., Dziak, E., Michalak, M., and Opas, M. (2003). Is all of the endoplasmic reticulum
created equal? The effect of the heterogeneous distribution of endoplasmic reticulum Ca2þ‐handling protein. J. Cell. Biol. 160, 475–479.
Parlati, F., Dignard, D., Bergeron, J. J. M., and Thomas, D. Y. (1995a). The calnexin
homologue cnx1þ in Schizosaccharomyces pombe, is an essential gene which can be
complemented by its soluble ER domain. EMBO J. 14, 3064–3072.
Parlati, F., Dominguez, M., Bergeron, J. J. M., and Thomas, D. Y. (1995b). Saccharomyces
cerevisiae CNE1 encodes an endoplasmic reticulum (ER) membrane protein with sequence
similarity to calnexin and calreticulin and functions as a constituent of the ER quality control
apparatus. J. Biol. Chem. 270, 244–253.
Parodi, A. J. (2000). Role of N‐oligosaccharide endoplasmic reticulum processing reactions in
glycoprotein folding and degradation. Biochem. J. 348, 1–13.
Peterson, J. R., Ora, A., Van, P. N., and Helenius, A. (1995). Transient, lectin‐like associationof calreticulin with folding intermediates of cellular and viral glycoproteins. Mol. Biol. Cell 6,
1173–1184.
Petersen, O. H., Gerasimenko, O. V., and Gerasimenko, J. V. (1998). The calcium store in the
nuclear envelope. Cell Calcium 23, 87–90.
Petersen, O. H., Tepikin, A., and Park, M. K. (2001). The endoplasmic reticulum: One
continuous or several separate Ca2þ stores? Trends Neurosci. 24, 271–276.
Pike, S. E., Yao, L., Jones, K. D., Cherney, B., Appella, E., Sakaguchi, K., Nakhasi, H.,
Teruya‐Feldstein, J., Wirth, P., Gupta, G., and Tosato, G. (1998). Vasostatin, a calreticu-
lin fragment, inhibits angiogenesis and suppresses tumor growth. J. Exp. Med. 188,
2349–2356.
118 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:52 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Pike, S. E., Yao, L., Setsuda, J., Jones, K. D., Cherney, B., Appella, E., Sakaguchi, K.,
Nakhasi, H., Atreya, C. D., Teruya‐Feldstein, J., Wirth, P., Gupta, G., and Tosato, G.
(1999). Calreticulin and calreticulin fragments are endothelial cell inhibitors that suppress
tumor growth. Blood 94, 2461–2468.
Putney, J. W., Jr. (1999). ‘‘Kissin’ cousins’’: Intimate plasma membrane‐ER interactions
underlie capacitative calcium entry. Cell 99, 5–8.
Qu, D., Teckman, J. H., Omura, S., and Perlmutter, D. H. (1996). Degradation of a mutant
secretory protein, alpha1–antitrypsin Z, in the endoplasmic reticulum requires proteasome
activity. J. Biol. Chem. 271, 22791–22795.
Rabouille, C., and Spiro, R. G. (1992). Nonselective utilization of the endomannosidase
pathway for processing glycoproteins by human hepatoma (HepG2) cells. J. Biol. Chem. 267,
11573–11578.
Rajagopalan, S., and Brenner, M. B. (1994). Calnexin retains unassembled major histocompat-
ibility complex class I free heavy chains in the endoplasmic reticulum. J. Exp. Med. 180,
407–412.
Rajagopalan, S., Xu, Y., and Brenner, M. B. (1994). Retention of unassembled components of
integral membrane proteins by calnexin. Science 263, 387–390.
Rauch, F., Prud’homme, J., Arabian, A., Dedhar, S., and St‐Arnaud, R. (2000). Heart, brain,
and body wall defects in mice lacking calreticulin. Exp. Cell Res. 256, 105–111.
Rodan, A. R., Simons, J. F., Trombetta, E. S., and Helenius, A. (1996). N‐linkedoligosaccharides are necessary and sufficient for association of glycosylated forms of bovine
RNAse with calnexin and calreticulin. EMBO J. 15, 6921–6930.
Roderick, H. L., Lechleiter, J. D., and Camacho, P. N. (2000). Cytosolic phosphorylation of
calnexin controls intracellular Ca(2þ) oscillations via an interaction with SERCA2b. J. Cell
Biol. 149, 1235–1248.
Rojiani, M. V., Finlay, B. B., Gray, V., and Dedhar, S. (1991). In vitro interaction of a
polypeptide homologous to human Ro/SS‐A antigen (calreticulin) with a highly conserved
amino acid sequence in the cytoplasmic domain of integrin alpha subunits. Biochemistry 30,
9859–9866.
Russell, S. J., Ruddock, L. W., Salo, K. E., Oliver, J. D., Roebuck, Q. P., Llewellyn, D. H.,
Roderick, H. L., Koivunen, P., Myllyharju, J., and High, S. (2004). The primary substrate
binding site in the b’ domain of ERp57 is adapted for endoplasmic reticulum lectin
association. J. Biol. Chem. 279, 18861–18869.
Sadasivan, B., Lehner, P. J., Ortmann, B., Spies, T., and Cresswell, P. (1996). Roles for
calreticulin and a novel glycoprotein, tapasin, in the interaction of MHC class I molecules
with TAP. Immunity 5, 103–114.
Saito, Y., Ihara, Y., Leach, M. R., Cohen‐Doyle, M. F., and Williams, D. B. N. (1999).
Calreticulin functions in vitro as a molecular chaperone for both glycosylated and non‐glycosylated proteins. EMBO J. 18, 6718–6729.
Santella, L., and Carafoli, E. (1997). Calcium signaling in the cell nucleus. FASEB J. 11,
1091–1109.
Sasak, V. W., Ordovas, J. M., Elbein, A. D., and Berninger, R. W. (1985). Castanospermine
inhibits glucosidase I and glycoprotein secretion in human hepatoma cells. Biochem. J. 232,
759–766.
Sato, S., Ward, C. L., Krouse, M. E., Wine, J. J., and Kopito, R. R. (1996). Glycerol reverses
the misfolding phenotype of the most common cystic fibrosis mutation. J. Biol. Chem. 271,
635–638.
Schrag, J. D., Bergeron, J. J., Li, Y., Borisova, S., Hahn, M., Thomas, D. Y., and Cygler, M.
(2001). The structure of calnexin, an ER chaperone involved in quality control of protein
folding. Mol. Cell 8, 633–644.
CELLULAR FUNCTIONS OF ER CHAPERONES 119

Comp. by:bala Date:27/5/05 Time:20:26:52 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Scott, J. E., and Dawson, J. R. (1995). MHC class I expression and transport in a calnexin‐deficient cell line. J. Immunol. 155, 143–148.
Spiro, R. G., Zhu, Q., Bhoyroo, V., and Soling, H. D. (1996). Definition of the lectin‐likeproperties of the molecular chaperone, calreticulin, and demonstration of its copurification
with endomannosidase from rat liver Golgi. J. Biol. Chem. 271, 11588–11594.
Stanley, P., and Ioffe, E. (1995). Glycosyltransferase mutants: Key to new insights in
glycobiology. FASEB J. 9, 1436–1444.
Sugiyama, T., Matsuda, Y., and Mikoshiba, K. (2000). Inositol 1,4,5–trisphosphate receptor
associated with focal contact cytoskeletal proteins. FEBS Lett. 466, 29–34.
Tjoelker, L. W., Seyfried, C. E., Eddy, R. L., Byers, M. G., Shows, T. B., Calderon, J.,
Schreiber, R. B., and Gray, P. W. (1994). Human, mouse, and rat calnexin cDNA cloning:
Identification of potential calcium binding motifs and gene localization to human
chromosome 5. Biochemistry 33, 3229–3236.
Tsukuba, T., Kadowaki, T., Hengst, J. A., and Bond, J. S. (2002). Chaperone interactions of the
metalloproteinase meprin A in the secretory or proteasomal‐degradative pathway. Arch.
Biochem. Biophys. 397, 191–198.
Van der Wal, F. J., Oliver, J. D., and High, S. (1998). The transient association of
ERp57 with N‐glycosylated proteins is regulated by glucose trimming. Eur. J. Biochem. 256,
51–59.
Vandivier, R. W., Ogden, C. A., Fadok, V. A., Hoffmann, P. R., Brown, K. K., Botto, M.,
Walport, M. J., Fisher, J. H., Henson, P. M., and Greene, K. E. (2002). Role of surfactant
proteins A, D, and C1q in the clearance of apoptotic cells in vivo and in vitro: Calreticulin and
CD91 as a common collectin receptor complex. J. Immunol. 169, 3978–3986.
van Leeuwen, J. E., and Kearse, K. P. (1996). Calnexin associates exclusively with individual
CD3 delta and T cell antigen receptor (TCR) alpha proteins containing incompletely trimmed
glycans that are not assembled into multisubunit TCR complexes. J. Biol. Chem. 271,
9660–9665.
Vassilakos, A., Michalak, M., Lehrman, M. A., and Williams, D. B. (1998). Oligosaccharide
binding characteristics of the molecular chaperones calnexin and calreticulin. Biochemistry
37, 3480–3490.
Vinayagamoorthy, T., and Rajakumar, A. R. (1996). Stress relief protein modulation by
calnexin. Ann. NY Acad. Sci. 793, 479–484.
Volonte, D., Galbiati, F., Pestell, R. G., and Lisanti, M. P. (2001). Cellular stress induces the
tyrosine phosphorylation of caveolin‐1 (Tyr14) via activation of p38 mitogen‐activatedprotein kinase and c‐Src kinase—Evidence for caveolae, the actin cytoskeleton, and focal
adhesions as mechanical sensors of osmotic stress. J. Biol. Chem. 276, 8094–8103.
Vranka, J., Mokashi, A., Keene, D. R., Tufa, S., Corson, G., Sussman, M., Horton, W. A.,
Maddox, K., Sakai, L., and Bachinger, H. P. (2001). Selective intracellular retention of
extracellular matrix proteins and chaperones associated with pseudoachondroplasia. Matrix
Biol. 20, 439–450.
Wada, I., Rindress, D., Cameron, P. H., Ou, W. J., Doherty, J. J., Louvard, D., Bell, A. W.,
Dignard, D., Thomas, D. Y., and Bergeron, J. J. (1991). SSR alpha and associated calnexin
are major calcium binding proteins of the endoplasmic reticulum membrane. J. Biol. Chem.
266, 19599–19610.
Ware, F. E., Vassilakos, A., Peterson, P. A., Jackson, M. R., Lehrman, M. A., and Williams,
D. B. (1995). The molecular chaperone calnexin binds Glc1Man9GlcNAc2 oligosaccharide as
an initial step in recognizing unfolded glycoproteins. J. Biol. Chem. 270, 4697–4704.
Wei, Y., Yang, X. W., Liu, Q. M., Wilkins, J. A., and Chapman, H. A. (1999). A role for
caveolin and the urokinase receptor in integrin‐mediated adhesion and signaling. J. Cell Biol.
144, 1285–1294.
120 BEDARD ET AL.

Comp. by:bala Date:27/5/05 Time:20:26:52 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
UNCORRECTEDPROOF
Westphal, V., Xiao, M., Kwok, P. Y., and Freeze, H. H. (2003). Identification of a frequent
variant in ALG6, the cause of congenital disorder of glycosylation‐Ic. Hum. Mutat. 22,
420–421.
White, T. K., Zhu, Q., and Tanzer, M. L. (1995). Cell surface calreticulin is a putative
mannoside lectin which triggers mouse melanoma cell spreading. J. Biol. Chem. 270,
15926–15929.
Winrow, C. J., Miyata, K. S., Marcus, S. L., Burns, K., Michalak, M., Capone, J. P., and
Rachubinski, R. A. (1995). Calreticulin modulates the in vitro DNA binding but not the
in vivo transcriptional activation by peroxisome proliferator‐activated receptor/retinoid X
receptor heterodimers. Mol. Cell. Endocrinol. 111, 175–179.
Wormald, M. R., and Dwek, R. A. (1999). Glycoproteins: Glycan presentation and protein‐foldstability. Structure Fold Des. 7, R155–R160.
Wormald, M. R., Petrescu, A. J., Pao, Y. L., Glithero, A., Elliott, T., and Dwek, R. A. (2002).
Conformational studies of oligosaccharides and glycopeptides: Complementarity of NMR,
X‐ray crystallography, and molecular modelling. Chem. Rev. 102, 371–386.
Xu, K., Tavernarakis, N., and Driscoll, M. (2001). Necrotic cell death in C. elegans requires the
function of calreticulin and regulators of Ca2þ release from the endoplasmic reticulum.
Neuron 31, 957–971.
Xu, W., Longo, F. J., Wintermantel, M. R., Jiang, X. Y., Clark, R. A., and DeLisle, S. (2000).
Calreticulin modulates capacitative Ca2þ influx by controlling the extent of inositol 1,4,5–
trisphosphate‐induced Ca2þ store depletion. J. Biol. Chem. 275, 36676–36682.
Yeates, L. C., and Powis, G. (1997). The expression of the molecular chaperone calnexin is
decreased in cancer cells grown as colonies compared to monolayer. Biochem. Biophys. Res.
Commun. 238, 66–70.
Yoon, G. S., Lee, H., Jung, Y., Yu, E., Moon, H. B., Song, K., and Lee, I. (2000). Nuclear
matrix of calreticulin in hepatocellular carcinoma. Cancer Res. 60, 1117–1120.
Zapun, A., Petrescu, S. M., Rudd, P. M., Dwek, R. A., Thomas, D. Y., and Bergeron, J. J.
(1997). Conformation‐independent binding of monoglucosylated ribonuclease B to calnexin.
Cell 88, 29–38.
Zapun, A., Darby, N. J., Tessier, D. C., Michalak, M., Bergeron, J. J., and Thomas, D. Y.
(1998). Enhanced catalysis of ribonuclease B folding by the interaction of calnexin or
calreticulin with ERp57. J. Biol. Chem. 273, 6009–6012.
Zhang, Q., Tector, M., and Salter, R. D. (1995). Calnexin recognizes carbohydrate and protein
determinants of class I major histocompatibility complex molecules. J. Biol. Chem. 270,
3944–3948.
Zhu, Q., Zelinka, P., White, T., and Tanzer, M. L. (1997). Calreticulin‐integrin bidirectional
signaling complex. Biochem. Biophys. Res. Commun. 232, 354–358.
Zuppini, A., Groenendyk, J., Cormack, L. A., Shore, G., Opas, M., Bleackley, R. C., and
Michalak, M. (2002). Calnexin deficiency and endoplasmic reticulum stress‐inducedapoptosis. Biochemistry 41, 2850–2858.
CELLULAR FUNCTIONS OF ER CHAPERONES 121
Comp. by:bala Date:27/5/05 Time:20:26:52 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
Author Query Form
International Review of Cytology, 245
Chapter 4
_____________________________________________________Dear Author,
During the preparation of your manuscript for typesetting some questions have arisen.These are listed below. Please check your typeset proof carefully and mark anycorrections in the margin of the proof or compile them as a separate list. This formshould then be returned with your marked proof/list of corrections to Elsevier Science.
Disk use
In some instances we may be unable to process the electronic file of your article and/orartwork. In that case we have, for efficiency reasons, proceeded by using the hard copyof your manuscript. If this is the case the reasons are indicated below:
Disk damaged Incompatible file format LaTeX file for non-LaTeX journal
Virus infected Discrepancies between electronic file and (peer-reviewed,therefore definitive) hard copy.
Other: ...................................................
We have proceeded as follows:
Manuscript scanned Manuscript keyed in Artwork scanned
Files only partly used (parts processed differently:................................................)
Bibliography
If discrepancies were noted between the literature list and the text references, thefollowing may apply:
The references listed below were noted in the text but appear to be missing from yourliterature list. Please complete the list or remove the references from the text.
» Uncited references: This section comprises references which occur in the referencelist but not in the body of the text. Please position each reference in the text or,alternatively, delete it. Any reference not dealt with will be retained in this section.
Query Refs. Details Required Author’s response
AU1 add to refs
AU2 add to refs
AU3 add to refs
AU4 add to refs
AU5 update
AU6 a or b
AU7 add to refs
AU8 Update.
Comp. by:bala Date:27/5/05 Time:20:26:52 Stage:First Proof File Path://spsind002s/serials/PRODENV/000000~1/00F256~1/S00000~1/000000~3/000000~2/000006276.3DProof by:Shanmugavel QC by:Thiru ProjectAcronym:IRCY Volume:45004
Typesetter Query
Attention authors: Please address every typesetter query below. Failure to do so mayresult in references being deleted from your proofs. Thanks for your cooperation.
Query Refs. Details Required Author’s response
TS1 Please provide the subparts a orb for the reference parlati et al.(1995) in text.
TS2 The following references are notincluded in the reference list.Nakamura et al. (2001)Volonte et al. (2001).
TS3 The foot note a, q are not cross-refered in the body part of Table1. please provide.





























