Embed Size (px)
Citation preview

LYNN EL HADDAD
CARACTERISATION DES PHAGES DE STAPHYLOCOCCUS A UREUS
Mémoire présenté à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en Microbiologie pour l'obtention du grade de Maîtrise es Sciences (M. Sc.)
DEPARTEMENT DE BIOCHIMIE, DE MICROBIOLOGIE ET DE BIO-INFORMATIQUE
FACULTÉ DES SCIENCES ET DE GÉNIE UNIVERSITÉ LAVAL
QUÉBEC
2010
Lynn El Haddad, 2010

Résumé Staphylococcus aureus est une bactérie pathogène des animaux et de l'homme causant
plusieurs symptômes chez ceux-ci. De plus, elle est responsable d'intoxications
alimentaires liées à l'ingestion de nourritures contaminées par des entérotoxines
staphylococciques. Avec l'émergence de souches staphylococciques résistantes aux
.antibiotiques, l'utilisation de phages virulents est maintenant envisagée comme alternative
pour lutter contre les souches pathogènes de S. aureus.
Au cours de ce projet, un nouveau phage virulent (LH1) a été isolé à partir d'échantillons
de lait cru. L'utilisation de ce dernier et du phage K dans du lait UHT a montré qu'ils sont
capables d'éliminer les souches de S. aureus mais que leur activité lytique est inhibée dans
le lait cru par un composé encore inconnu. La présence de deux gènes codant pour la
toxine leucocidine de Panton-Valentin dans le génome du phage LH1 compromet son
utilisation comme agent de biocontrôle dans les applications alimentaires et médicales.
Finalement, un arbre phylogénétique a été créé en comparant et regroupant 54 génomes de
phages différents de S. aureus disponibles dans GenBank en plus de deux nouveaux phages
isolés incluant LH1.
L'avancement des connaissances sur des phages de S. aureus pourrait favoriser leur
utilisation dans la prévention ou le contrôle des intoxications alimentaires.

11
Abstract Staphylococcus aureus can cause a range of illnesses in animals and humans. In cases of
food poisoning, this is mostly due to the ingestion of heat-labile enterotoxins produced by
S. aureus, leading to nausea, diarrhea, vomiting, and abdominal cramps. Virulent phages
could be used to eliminate these bacteria in milk products thereby reducing the risks of
food contamination and improving food safety.
In this work, a new virulent S. aureus phage, named LH1, was isolated from raw milk
samples. The use of phages LH1 and K in UHT milk showed that they were effective in
reducing S. aureus counts. However, both phages were inhibited in raw milk. Furthermore,
the analysis of the genome of LH1 showed the presence of two genes coding for the
Panton-Valentine leukocidin toxin compromising its use as a biocontrol agent. In parallel,
54 S. aureus complete phage genomes available in GenBank in addition to two newly
isolated phages were analyzed and classified using the MEGA 4.1 and the DOTTER
softwares.
The advances in the field of S. aureus bacteriophages may lead to the reduction of the
occurrence of infections and with this, control food poisoning.

Ill
Avant-Propos En août 2008, j 'ai quitté mon pays, le Liban. J'ai dit au revoir à ma famille, à mes amis et à
la vie que j'avais. Voyageant vers un pays inconnu, j'avais peur; peur de ce qui m'attend,
de ce que je vais endurer. Et c'est au début du mois de septembre que je me suis
familiarisée avec la vie étudiante et la vie sociale au Canada. L'accueil, l'hospitalité et la
gentillesse de tout le monde m'a donné de l'énergie pour réussir. Je voudrais remercier du
fond du cœur tous ceux qui m'ont aidé, de loin ou de près, à la réalisation de mon projet de
maîtrise.
Merci Dr Sylvain Moineau, vous avez mis toute votre confiance en moi et vous m'avez
encouragé tout au long de ma maîtrise. Sans votre soutien et vos conseils, je ne serai pas ce
que je suis devenue aujourd'hui.
Merci aux membres du comité aviseur; Dr Michel Frenette et Dr Steve Labrie. Vos
suggestions et vos commentaires m'ont été précieux.
Je voudrais de même remercier tous mes collègues au laboratoire : Denise, Manuela,
Geneviève, Mourad, Josiane, Julie, Simon, Alfonso, Jeannot, Sih.am, Marie-Ève, Audrey,
Maxim et Rym. Je vous embêtais chaque jour avec mes nombreuses questions! Mais
sachez qu'elles m'ont aidé à avcincer mon projet et ont élargi mon sens de l'apprentissage
et mon esprit scientifique.
Merci au Programme Canadien de Bourse de la Francophonie et à l'ACDI pour leur
soutien financier, leur présence ainsi que le suivi fourni tout au long des études et de
l'intégration.
Je remercie aussi mes plus fidèles amies, Rana et Geneviève. Geneviève, tu es toujours
restée à mes côtés. Et c'est grâce à ta gentillesse, ta bonté, ta merveilleuse amitié et ton
adorable famille que j 'ai pu réussir. Rana, tu représentes pour moi la grande sœur que je
n'ai jamais eue. Notre amitié s'est renforcée durant ces deux années et a fait naître un
sentiment de fraternité incontournable et indestructible.

IV
Je voudrais aussi remercier mes frères Nicolas et Alain et ma sœur Léa. Nous avons
partagé pleins de moments de joie et de succès ensembles qui seront gravés dans ma
mémoire à jamais. Sachez que rien, même pas les distances qui nous séparent, ne pourront
remplacer ou effacer ces souvenirs merveilleux.
Je remercie du fond du cœur mes parents, Paula et Sami. Vous êtes toujours restés à mes
côtés et dans mes pensées que ce soit au Liban ou à Québec. Vous m'avez immergé de
bonté, de confiance et d'amour. Vous m'avez appris à être aimable, modeste, généreuse,
confiante, forte et toujours positive. C'est grâce à vous que je suis devenue la femme que je
suis aujourd'hui. Je vous dédis ce mémoire qui représente une très petite portion de ma
gratitude envers vous.
Je remercie de tout mon cœur mon copain Georges. Tu as cru en moi et tu m'as encouragé
tout au long de ma maîtrise. Tu m'as donné la force et la confiance pour l'achever avec
progrès et succès. Je te dédis ce mémoire accompagné de trois mots : Forever and Always.
Et enfin, j'offre ce mémoire à toute ma famille et mes amis. C'est grâce à vous tous que je
termine ma maîtrise avec plein d'inoubliables moments de joie, de bonheur, de réussite et
d'amour.
Bonne lecture!

Progress lies not in enhancing what is, but in advancing toward what will be.
-Kahlil Gibran-
A toutes les personnes que j'aime et A mon pays le Liban,

VI
Table des matières Résumé i Abstract ii Avant-Propos iii Liste des figures viii Liste des tableaux x Liste des abréviations xi 1. Introduction 1 1.1. Staphylococcus aureus 1
1.1.1. Caractéristiques générales 1 1.1.2. Milieux de culture pour l'isolement et l'identification de S. aureus 3 1.1.3. Problématique de S. aureus 5 1.1.4. Facteurs de virulence de S. aureus et leur régulation 6 1.1.5. Intoxications alimentaires staphylococciques 9 1.1.6. Solutions envisagées pour la prévention et le contrôle de S. aureus 11
1.2. Les bacteriophages 13 1.2.1. Historique 13 1.2.2. Classification des phages 14 1.2.3. Phages de S. aureus 17
1.2.3.1. Gasification génomique des phages de S. aureus 17 1.2.4. Cycle de multiplication des phages 21 1.3. Applications des phages de S. aureus 25
1.3.1. Thérapie par phages 26 1.3.2. Applications alimentaires des phages de S. aureus 27 1.3.2.1. Test de l'activité des phages dans le lait 27 1.3.2.2. Utilisation de phages isolés du lait cru et production de cocktails de phages 31
1.4. Problématique à l'étude et objectifs de recherche 32 2. Matériel et méthodes 34 2.1. Objectif 1: Isolement de phages de S. aureus à partir d'échantillons de lait cru 34 2.2. Objectif 2: Caractérisation du nouveau phage isolé 35
2.2.1. Observation des phages en microscopie électronique 35 2.2.2. Spectre lytique du nouveau phage 36 2.2.3. Séquençage d'ADN 37
2.3. Objectif 3: Comparaison du phage isolé aux autres phages de S. aureus 38 2.3.1. Profils de restriction du génome de phages de S. aureus 38 2.3.2. Spectre lytique des phages 39 2.3.3. Test d'adsorption des phages à des cellules de S. aureus 39 2.3.4. Analyse bioinformatique des génomes de phages 40
2.4. Objectif 4: Utilisation des phages pour éliminer les souches de S. aureus dans différents types de laits 41
2.4.1. Test de l'inhibition de S. aureus par des phages dans le lait UHT 41 2.4.2. Test de l'inhibition de S. aureus par des phages dans le lait cru 43
3. Résultats 44 3.1. Objectif 1: Isolement de phages de S. aureus à partir d'échantillons de lait cru 44 3.2. Objectif 2: Caractérisation du nouveau phage isolé 44
3.2.1. Observation des phages en microscopie électronique 44

vu
3.2.2. Spectre lytique du phage LH1 45 3.2.3. Génome du phage LH1 46
3.3. Objectif 3: Comparaison du phage isolé aux autres phages de S. aureus 50 3.3.1. Profils de restriction du génome de phages de S. aureus 50 3.3.2. Spectre lytique des phages 50 3.3.3. Test d'adsorption des phages à des cellules de S. aureus 52 3.3.4. Analyse bioinformatique des génomes de phages 52
3.4. Objectif 4: Utilisation des phages pour éliminer les souches de S. aureus dans différents types de laits 58
3.4.1. Test de l'inhibition de S. aureus par des phages dans le lait UHT 58 3.4.2. Test de l'inhibition de S. aureus par des phages dans le lait cru 61
4. Discussion 62 4.1. Caractérisation du phage LH1 62 4.2. Comparaison des phages de S. aureus au phage LH1 63 4.3. Activité du phage LH1 dans différents types de lait contaminés par S. aureus 66 5. Conclusion et perspectives 68 6. Bibliographie 70

vin
Liste des figures
Figure 1. Photographie de Staphylococcus aureus observée en microscopie électronique à
balayage 2
Figure 2. Milieu Baird Parker inoculé avec une souche de S. aureus et comparaison avec
un milieu non inoculé 4
Figure 3. Milieu Mannitol Salt Agar inoculé avec une souche de S. aureus et comparaison
aux colonies de S. epidermidis et à un milieu non inoculé 5
Figure 4. Structure d'une cellule de S. aureus 7
Figure 5. Représentation schématique du système agr 8
Figure 6. Représentation schématique des principales familles de bacteriophages 14
Figure 7. Caractéristiques morphologiques et génomiques des phages caudés 15
Figure 8. Analyse comparative des séquences nucléotidiques des génomes de phages de S.
aureus 18
Figure 9. Arbre protéomique des phages appartenant à la famille des Siphoviridae 19
Figure 10. Étapes des cycles des bacteriophages : cycles lytique et lysogénie 21
Figure 11. Structure des acides téichoïques 22
Figure 12. Voie d'assemblage des protéines structurales des phages caudés 24
Figure 13. Micrographies prises par microscopie électronique à balayage 28
Figure 14. Micrographies de lait cru et de lait traité à haute température 30
Figure 15. Représentation graphique de la concentration bactérienne de S. aureus en
fonction de la densité optique 42
Figure 16. Image en microscopie électronique du phage LH1 45
Figure 17. Alignement du génome des phages LH1, 3A, 42e et 47 49
Figure 18. Profils de restriction obtenus avec EcoRV du génome de onze phages
caractérisés 50
Figure 19. Image en microscopie électronique du phage QW 54
Figure 20. Arbre phylogénétique comparant les séquences nucléotidiques des génomes de
phages de S. aureus 55
Figure 21. Analyse comparative des séquences nucléotidiques des génomes de phages de
S. aureus 56

IX
Figure 22. Analyse comparative des séquences nucléotidiques des génomes de phages de
S. aureus appartenant à la famille des Podoviridae 57
Figure 23. Représentation graphique du logarithme de la concentration bactérienne de
S. aureus en fonction du temps dans du lait à UHT 58
Figure 24. Représentation graphique du logarithme de la concentration des phages K et
LH1 en fonction du temps dans du lait à UHT 59
Figure 25. Représentation graphique du logarithme de la concentration du phage K et de
S. aureus 01 fonction du temps dans du lait à UHT 60
Figure 26. Représentation graphique du logarithme de la concentration du phage LH1 et de
S. aureus 01 en fonction du temps dans du lait à UHT 60
Figure 27. Représentation graphique de la concentration phagique de K et LH1 en fonction
du temps dans les laits UHT et cru 61
Figure 28. Identification d'au moins neuf groupes génétiquement différents de phages de
S. aureus 65

Liste des tableaux
Tableau 1. Classification des phages de S. aureus selon les bases de données de NCBI... 16
Tableau 2. Les différentes souches de S. aureus disponibles au laboratoire et leur
provenance 36
Tableau 3. Liste d'amorces utilisées pour l'assemblage des contigs de l'ADN du nouveau
phage isolé 37
Tableau 4. Onze phages caractérisés dans cette étude, leur provenance et leur souche hôte
de S. aureus 38
Tableau 5. Les génomes de phages de staphylocoques disponibles dans GenBank 40
Tableau 6. Caractéristiques des cadres de lecture ouverts du génome du phage LH1 47
Tableau 7. Spectre lytique des onze phages étudiés sur 32 souches de S. aureus 51
Tableau 8. Pourcentage d'adsorption des phages aux souches mutantes de S. aureus
01-P68et01-QW 52
Tableau 9. Caractéristiques des cadres de lecture ouverts du génome du phage QW 53

XI
Liste des °c ADN
agr
AIP
ARN
ARNm
ATP
BIM
CMRSA
D.O.
db
Dig
DNase
EDTA
FDA
GC
gP HCI
ICTV
kb
kDa
Lan
LTA
MAPAQ
MDa
MeLan
ml
abréviations Degré Celsius
Acide DésoxyriboNucléique
Gène régulateur accessoire (accessory gene regulator)
Peptide autoinduit (Auto Inducing Peptide)
Acide RiboNucléique
Acide RiboNucléique messager
Adenosine Triphosphate
Mutant résistant aux bacteriophages (Bacteriophage-Insensitive Mutants)
Souche canadienne de Staphylococcus aureus résistante à la méthicilline
(Canadian Methicillin-Resistant Staphylococcus aureus)
Densité Optique
Double brin
Digoxigénine
DésoxyriboNucléase
Acide ethylene diamine tétr.aacétique (EthyleneDiamineTetraacetic Acid)
L'administration américaine des denrées alimentaires et des médicaments
(Food and Drug Administration)
Ratio des nucleotides G et C par rapport aux nucleotides A et T
Produit de gène (Gene Product)
Acide chlorhydrique
Le comité international de la taxonomie des virus (International Committee
on Taxonomy of Viruses)
Kilob.ase
KiloDalton
Lanthionine
Acide lipotéichoïque (LipoTeichoic Acid)
Ministère de l'Agriculture, des Pêcheries et de l'Alimentation du Québec
MégaDalton
Méthyllanthionine
Millilitre

Xll
mm Millimètre
MM Masse Moléculaire
mmol (mM) Millimolaire
MOI Multiplicité d'infection (Multiplicity Of Infection)
MRSA Methicillin-Resistant Staphylococcus aureus
MSSA Souche de Staphylococcus aureus sensible à la méthicilline (Methicillin-
Sensitive Staphylococcus aureus)
NaCl Chlorure de Sodium
NAG N-Acétyl-Glucosamine
NAM Acide N-Acétyl-Muramique
NCBI National Center Biotechnology Information
nm Nanometre
OatA O-acétyltransférase
ORF Cadre de lecture ouvert (Open Reading Frame)
pb Paire de bases
PBP2a Protéine 2a liée à la pénicilline (Penicillin-Binding Protein 2a)
PCR Réaction en chaîne par polymerase (Polymerase Chain Reaction)
PEG PolyEthylene Glycol
pH Potentiel d'Hydrogène
pi Point isoélectrique
PLR Protéine de Liaison au Récepteur
PTA Acide phosphotungstique (PhosphoTungstic Acid)
P/V Poids/volume
RNase RiboNucléase
SARM Souche de Staphylococcus aureus résistante à la Méthicilline
sb Simple brin
SD Shine-Dalgarno
TAE Tampon de Tris-acétate contenant de l'EDTA
TSB Bouillon tryptone au soja (Trypticase Soy Broth)
UA Acétate d'uracyle (Uracyl Acetate)
UFC/UFP Unités Formatrices de Colonies/Unités Formées de Plages de lyse

Xlll
UHT Ultra Haute Température
Ml Microlitre
UV Ultraviolet V Volt

1. Introduction 1.1. Staphylococcus aureus
1.1.1. Caractéristiques générales
Staphylococcus aureus est une bactérie omniprésente chez l'homme, les animaux ainsi que
dans l'aliment. Le nom de « Staphylococcus » a été attribué en 1882 à un regroupement de
cocci causant un grand nombre de maladies pyogenes chez l'humain. Ces bactéries ont été
isolées et analysées par le chirurgien écossais Sir Alexander Ogston en 1880. Il fut
démontré que les microorganismes dérivés de pus prélevés d'un abcès humain
provoquaient les mêmes symptômes et des lésions pyogenes chez une souris (82). De plus,
il a remédié à la maladie en inactivant les staphylocoques par des traitements thermiques et
au phénol. Le genre Staphylococcus a été séparé de celui de Micrococcus par Evans en
1957. Ces deux organismes donnent des colonies opaques, blanches, orange ou jaune sur
gélose au sang. Tout deux sont des bactéries à Gram positif et catalase positive (82).
Cependant, ils sont physiologiquement distincts de par leur capacité à croître en condition
anaérobique et à fermenter le glucose (16). En effet, le genre Staphylococcus est capable de
fermenter le glucose et de produire de l'acide lactique sous conditions anaérobiques
contrairement au métabolisme respiratoire aérobique strict du genre Micrococcus (43, 82,
97).
Le genre Staphylococcus regroupe plus de 40 espèces classées, entre autres, en fonction de
leur capacité à coaguler le plasma de lapin (50). L'espèce aureus est coagulase positive et
comprend deux sous-espèces : aureus et anaerobius. Celles-ci présentent des différences
au test de la catalase, de la (3-glucosidase, de la pigmentation des colonies, de la présence
du monosaccharide N-acétylglucosamine de la surface bactérienne et de l'utilisation du
mannose, lactose et trehalose (92).
S. aureus est l'espèce de staphylocoque la plus pathogène aussi bien chez l'homme que
chez les animaux (111). Ce staphylocoque est non-motile et se présente en coques de 0,5 à
1,5 pm de diamètre associés en paires ou en grappes. Sa morphologie peut être observée à
la figure 1.
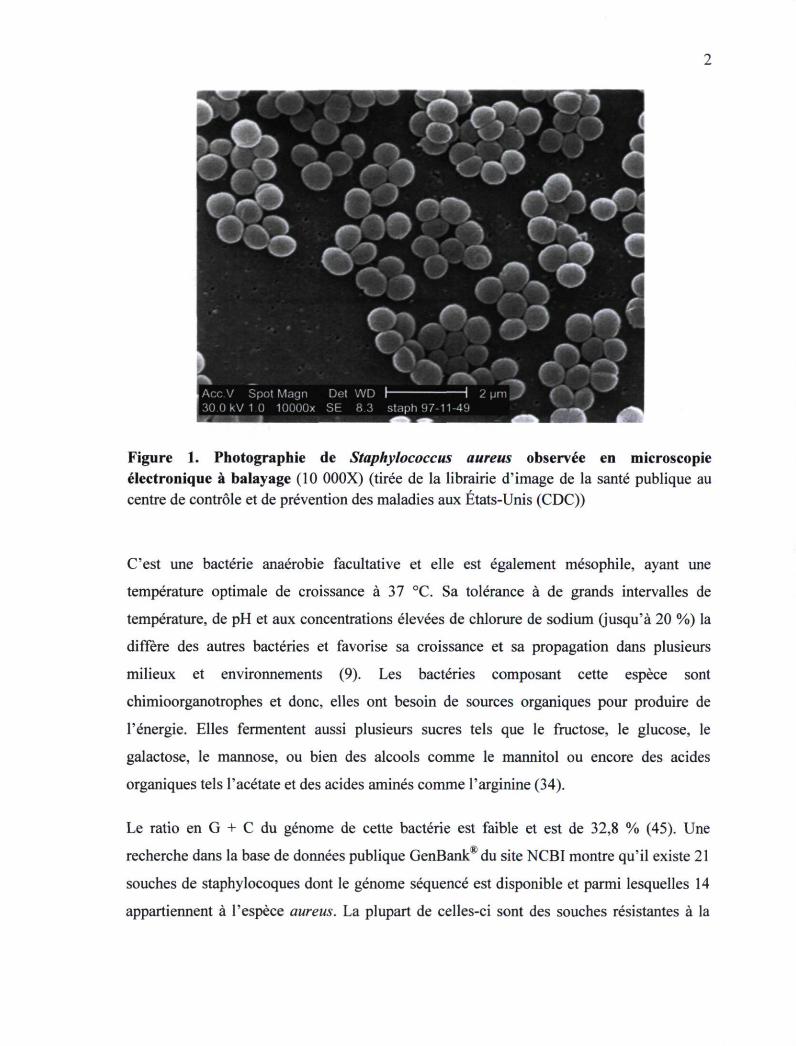
Figure 1. Photographie de Staphylococcus aureus observée en microscopie électronique à balayage (10 000X) (tirée de la librairie d'image de la santé publique au centre de contrôle et de prévention des maladies aux États-Unis (CDC))
C'est une bactérie anaérobie facultative et elle est également mésophile, ayant une
température optimale de croissance à 37 °C. Sa tolérance à de grands intervalles de
température, de pH et aux concentrations élevées de chlorure de sodium (jusqu'à 20 %) la
diffère des autres bactéries et favorise sa croissance et sa propagation dans plusieurs
milieux et environnements (9). Les bactéries compos.ant cette espèce sont
chimioorganotrophes et donc, elles ont besoin de sources organiques pour produire de
l'énergie. Elles fermentent aussi plusieurs sucres tels que le fructose, le glucose, le
galactose, le mannose, ou bien des alcools comme le mannitol ou encore des acides
organiques tels l'acétate et des acides aminés comme l'arginine (34).
Le ratio en G + C du génome de cette bactérie est faible et est de 32,8 % (45). Une
recherche dans la base de données publique GenBank® du site NCBI montre qu'il existe 21
souches de staphylocoques dont le génome séquence est disponible et parmi lesquelles 14
appartiennent à l'espèce aureus. La plupart de celles-ci sont des souches résistantes à la

méthicilline qui sont considérées importantes à sequencer et analyser. La taille de ces
génomes est environ de 2000 kb.
1.1.2. Milieux de culture pour l'isolement et l'identification de S. aureus
Afin d'identifier et d'isoler S. aureus, des milieux de culture sélectifs et différentiels sont
souvent utilisés. Un milieu sélectif encourage la croissance des organismes désirés en
inhibant les autres alors qu'un milieu différentiel permet une discrimination basée sur une
propriété physiologique comme la fermentation des sucres.
Il existe un milieu de culture sélectif pour S. aureus et il se nomme Baird-Parker. Ce milieu
a été développé par Baird Parker en 1962 après avoir trouvé que S. aureus est capable
d'hydrolyser 5 % du jaune d'œuf dans un milieu de gélose au sang (Difco). Les colonies de
S. aureus contrairement aux autres espèces de staphylocoques, étaient entourées de zones
claires (15). Les colonies de S. aureus obtenues sur ce milieu sont noires, brillantes et
convexes, de 1 à 5 mm de diamètre. Elles possèdent également une mince bordure blanche
entourée d'un halo clair de 2 à 5 mm de large. Le milieu Baird-Parker contient du jaune
d'œuf, du pyruvate ainsi que des agents sélectifs dont la tellurite, la glycine et le chlorure
de lithium qui jouent un rôle dans l'inhibition de la flore secondaire (9, 16). L'hydrolyse de
la lipovitelline (la protéine du jaune d'œuf la plus abondante) donne naissance à une zone
claire autour de la colonie. De plus, S. aureus peut réduire la tellurite contenue dans le
milieu Baird-Parker en tellure qui donne un précipité noir observable au centre des
colonies et permet leur sélection. Un halo opaque apparaît dans la zone claire après 48
heures d'incubation et est due à l'activité de la lécithinase (Figure 2).

Figure 2. Milieu Baird Parker inoculé avec une souche de S. aureus et comparaison avec un milieu non inoculé (Difco™).
Un deuxième milieu est utilisé pour isoler d'une façon sélective et différentielle les
souches de S. aureus. Il s'agit du milieu Mannitol Salt Agar ou MSA. Celui-ci contient
7,5 % de NaCl inhibant la croissance des bactéries à l'exception des staphylocoques, ce qui
le rend sélectif pour les staphylocoques. D'autre part, alors que la plupart des espèces de
staphylocoques sont capables de croître à de hautes concentrations de sels, seul S. aureus
peut fermenter le mannitol. Le milieu MSA est également différentiel puisqu'il contient un
indicateur de pH nommé rouge de phénol. À bas pH, ce dernier colore en jaune les colonies
qui fermentent le mannitol comme celles de S. aureus et garde en orange ou rouge les
autres colonies (33, 104) (Figure 3).

Figure 3. Milieu Mannitol Salt Agar inoculé avec une souche de S. aureus (colonies jaunes) et comparaison aux colonies de S. epidermidis (colonies rouges) et à un milieu non inoculé (Difco™).
1.1.3. Problématique de S. aureus
S. aureus est l'une des rares bactéries présentant une résistance à l'action du lysozyme.
Ceci contribue fortement à sa persistance et sa capacité à coloniser la peau et les
muqueuses que ce soit chez l'homme ou l'animal (20). Le lysozyme est une muramidase
qui clive la liaison entre les résidus d'acide N-Acétyl-Muramique (NAM) et N-Acétyl-
Glucosamine (NAG). Il est produit par plusieurs tissus humains (muqueuses, tractus
respiratoire et intestinal) et est présent dans le sérum, la salive, la sueur, ainsi que dans les
larmes. C'est une enzyme de défense importante produite suite à l'augmentation de son
titre par infection et à son induction par les cellules phagocytaires et épithéliales (19, 20).
La résistance de S. aureus à cette enzyme est causée par deux facteurs; le haut degré de
reticulation de la couche de peptidoglycane (liaisons transversales donnant un polymère
tridimensionnel) et la production de la protéine OatA ou O-acétyltransférase. Cette dernière
est responsable de l'acétylation du carbone situé à la position 6 du NAM portant l'acide
téichoïque(18, 19).
S. aureus appartient à la microflore normale des narines, du vagin, du pharynx, des
aisselles et de la peau (73). Cependant, lorsque les staphylocoques adhèrent aux tissus ou

traversent dans la circulation sanguine de l'hôte, des infections apparaissent conduisant à
des pathologies diverses comme des infections de la peau, des muqueuses et des viscères
ainsi que les septicémies. Celles-ci sont dues à la production de facteurs de virulence par S.
aureus (voir section 1.1.4). D'autres pathologies sont causées par un seul type de toxine
comme le syndrome de la peau ébouillantée dû à l'exfoliatine, le syndrome de choc toxique
dû à la toxine TSST-1 et les toxi-infections alimentaires dues aux entérotoxines (111). Ces
dernières, contrairement aux autres toxines et facteurs de virulence causent
l'empoisonnement de l'individu ingérant un aliment contaminé par celles-ci. Le
mécanisme par lequel les entérotoxines pénètrent le corps via l'intestin et causent les
symptômes d'intoxication alimentaire est cependant peu connu jusqu'à présent (17).
1.1.4. Facteurs de virulence de S. aureus et leur régulation
La virulence de S. aureus provient de l'action combinée de différentes composantes de la
surface bactérienne ainsi que de la sécrétion de toxines et d'enzymes multiples (59). Les
facteurs de virulence de S. aureus peuvent être répartis en trois catégories fonctionnelles :
ceux qui aident à l'adhésion de la bactérie aux cellules et tissus, ceux qui contribuent aux
lésions tissulaires et à la propagation de l'infection et enfin, ceux qui protègent la bactérie
contre le système immunitaire de l'hôte (13). En tout, il existe plus de 40 protéines de
surface et extracellulaires identifiées dans les souches de S. aureus. Quelques unes sont
schématisées à la figure 4 (73). Les protéines associées à la surface, dont la protéine A et
les adhésines, sont illustrées dans la figure de la page suivante. Ces protéines ont la
capacité de fixer les anticorps, la fibronectine, le collagène et l'élastine et donc de conférer
à la bactérie une résistance à la phagocytose et d'améliorer son adhésion aux cellules de
l'hôte. De plus, la coagulase (Figure 4) cause une coagulation du plasma sanguin et donc la
formation de thrombus obturant le vaisseau sanguin (111). Une autre enzyme, la 0-
lactamase est à la base de la résistance de S. aureus aux antibiotiques appartenant à la
famille des P-lactames (Figure 4). L'enzyme hydrolyse l'anneau P-lactame et inactive la
molécule de pénicilline (72). S. aureus peut produire des toxines protéiques dont la toxine
a, la toxine du choc toxique TSST-1 et les entérotoxines dont l'entérotoxine B
schématisées dans la figure de la page suivante (Figure 4).
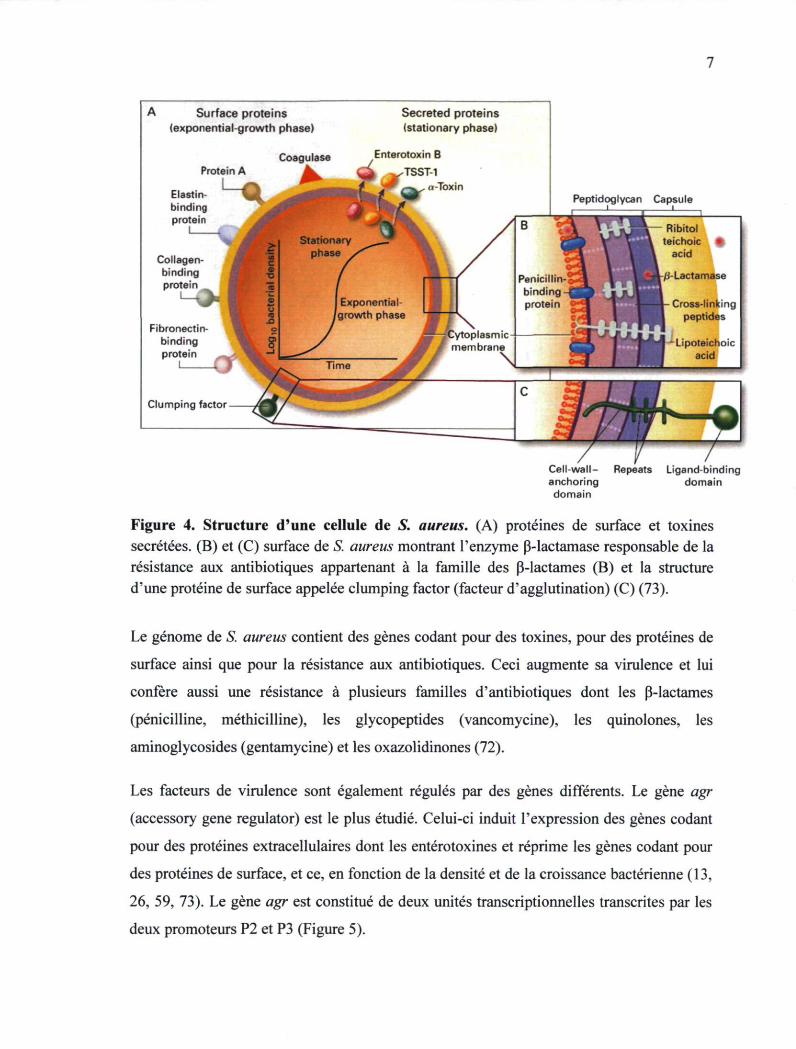
Cell-wall- Repeats Ligand-binding anchoring domain domain
Figure 4. Structure d'une cellule de S. aureus. (A) protéines de surface et toxines sécrétées. (B) et (C) surface de S. aureus montrant l'enzyme P-lactamase responsable de la résistance aux antibiotiques appartenant à la famille des P-lactames (B) et la structure d'une protéine de surface appelée dumping factor (facteur d'agglutination) (C) (73).
Le génome de S. aureus contient des gènes codant pour des toxines, pour des protéines de
surface ainsi que pour la résistance aux antibiotiques. Ceci augmente sa virulence et lui
confère aussi une résistance à plusieurs familles d'antibiotiques dont les P-lactames
(pénicilline, méthicilline), les glycopeptides (vancomycine), les quinolones, les
aminoglycosides (gentamycine) et les oxazolidinones (72).
Les facteurs de virulence sont également régulés par des gènes différents. Le gène agr
(accessory gene regulator) est le plus étudié. Celui-ci induit l'expression des gènes codant
pour des protéines extracellulaires dont les entérotoxines et réprime les gènes codant pour
des protéines de surface, et ce, en fonction de la densité et de la croissance bactérienne (13,
26, 59, 73). Le gène agr est constitué de deux unités transcriptionnelles transcrites par les
deux promoteurs P2 et P3 (Figure 5).

EXTRACELLULAR FLUID S Agr inducing peptide
Activation of P2 and P3
ïid
I agr transcript. RNAII
Regulator of virulence gene expression
Figure 5. Représentation schématique du système agr. Figure tirée d'Arvidson et al, 2001 (13).
Le transcrit généré à partir de P2 contient quatre gènes appelés agrB, D, CctA. Les gènes
agrB et agrD produisent l'AIP (Auto Induced Peptide). Ce dernier va se lier au domaine
cytoplasmique de la protéine AgrC qui va se phosphoryler et transférer un groupe
phosphate à la protéine AgrA. Celle-ci va activer la transcription des promoteurs P2 et P3.
Le transcrit produit à partir du promoteur P3 est l'ARN III, la molécule effectrice du
régulon agr. Elle active la transcription des gènes encodant pour les protéines sécrétées et
inhibe celle des gènes encodant pour les protéines de surface et coagulase. Ceci semble être
en relation avec la concentration bactérienne. En effet, lorsque le nombre de bactéries est
moindre comme dans la circulation sanguine, la transcription du gène agr est inactivée
donc la bactérie aura un phénotype adhésif dû à l'expression élevée des protéines de
surface. D'un autre coté, dans un abcès par exemple, la concentration importante de
bactéries va activer la transcription du gène agr conduisant à la production d'enzymes et de
toxines (13, 26, 59).
Un deuxième opéron contrôle de la même façon l'expression des facteurs de virulence. Il
s'agit de l'opéron sarA qui code pour la protéine SarA. Cette dernière est une activatrice de
l'opéron agr. C'est une protéine qui se lie à l'ADN au niveau de régions conservées riches

en bases azotées A et T. Elle se lie aux régions des promoteurs P2 et P3 de l'opéron agr et
augmente par ce fait le taux d'ARN III modifiant donc la synthèse des différents facteurs
de virulence. Elle peut se lier directement aux régions conservées des promoteurs des
différentes protéines de surface et exoprotéines (26).
1.1.5. Intoxications alimentaires staphylococciques
Par la diversité et la quantité de ses gènes de virulence, 5. aureus est l'une des causes
majeures des intoxications alimentaires. Cette bactérie peut croître dans plusieurs aliments
dont le lait, la crème, le beurre, les salades, le fromage, le jambon ainsi que d.ans les
saucisses (68). La présence de cette bactérie dans les différents types d'aliments est, entre
autres, le résultat d'une mauvaise manipulation de ceux-ci durant le processus de
fabrication, de stockage, de transport et de distribution (2). C'est à partir d'une
concentration de IO5 UFC/g d'aliment (80) que certaines souches de S. aureus peuvent
produire une concentration significative d'entérotoxines. Celles-ci causent des nausées,
vomissements, diarrhées, crampes abdominales, des maux de tête, des crampes
musculaires, des variations de la pression artérielle et même dans de très rares cas, la mort
de la personne par entérotoxicose (17). Les symptômes apparaissent 1 à 6 heures après
l'ingestion d'à peine 100 nanogrammes d'entérotoxines (17, 48).
Les intoxications alimentaires staphylococciques font l'objet d'une charge sociale
considérable en termes de coûts d'hospitalisations, de baisses d'heures de travail, de
productivité, etc. (83). De ce fait, les coûts de toutes les toxi-infections atteignent 152
milliards de dollars annuellement aux États-Unis (103). Au Canada, on note une perte
annuelle de 3,8 milliards de dollars suite aux intoxications alimentaires dont 171 millions
sont reliées aux produits laitiers (108). On suspecte 76 millions de malades, 325 000
hospitalisations et 5 200 morts chaque année dû à des maladies d'origine alimentaire aux
États-Unis (CFSAN, 2002). S. aureus est responsable de 16 millions de ces maladies et de
17 100 hospitalisations en France (68) tout comme en Amérique du Nord (CDC, 2007).
Dans l'industrie laitière, le lait cru constitue l'une des sources de contamination par S.
aureus. Dans une étude faite en 2006, 50 % à 60 % des élevages canadiens ont obtenus des
résultats positifs concernant la présence de S. aureus dans leur lait et 12 % des mammites
(infections des mamelles chez les vaches) au Canada sont causées par S. aureus (89). En

10
effet, selon l'étude de Normanno et a l , 38,4 % des échantillons de lait cru testés sont
contaminés par des staphylocoques à coagulase positive en Italie (83). De plus,
l'Organisation Mondiale de la Santé (OMS, 2000) a attribué, en 1999, 25 des 149
intoxications alimentaires staphylococciques à la consommation de fromages de lait cru en
France ainsi que 3 de 13 ont été reportées en Italie (52).
Les entérotoxines sont des protéines solubles dans l'eau et dans les solutions salines. Elles
appartiennent à la famille des exotoxines pyrogènes staphylococciques et streptococciques.
Ces toxines comprennent, entre autres, les toxines du syndrome de choc toxique (TSST),
les exfoliatines A et B, un groupe d'exotoxines streptococciques pyrogènes ainsi que les
entérotoxines staphylococciques (68). Ces toxines gastrointestinales puissantes et ces
superantigènes stimulent la prolifération des cellules T non spécifiques. Les gènes codant
pour les entérotoxines sont encodes par des prophages ou un phage défectif (phage tempéré
mutant ne pouvant plus s'exciser), le chromosome, des plasmides et des îlots de
pathogénicité (localisés sur le chromosome bactérien ou sur un plasmide) (68).
Présentement, on compte 11 entérotoxines, nommées SEA à SEJ avec trois sous-types de
SEC (9, 17).
Les caractéristiques les plus dangereuses des entérotoxines proviennent du fait qu'elles
sont fortement stables et résistantes à la chaleur ainsi qu'à la plupart des enzymes
protéolytiques comme la pepsine et la trypsine (68). Selon Anderson et al. (11),
l'entérotoxine staphylococcique A (SEA) demeure active après un traitement de 28
minutes à 121 °C et de 15 minutes à 127 °C. SEA est la toxine la plus fréquemment
impliquée dans les intoxications alimentaires (14, 32). D'un autre côté, SEB peut être
détruite par digestion de la pepsine à pH 2 indiquant que la réponse des toxines aux divers
traitements peut varier (21). La stabilité des entérotoxines à la chaleur semble être
dépendante du milieu, du pH, de la concentration de sels et de d'autres facteurs
environnementaux (11). Toutefois, les entérotoxines peuvent être éliminées après
stérilisation si elles sont présentes en petites quantités (68). Lors de la production de
fromages par exemple, elles peuvent même être soutirées avec le lactosérum (80).

11
1.1.6. Solutions envisagées pour la prévention et le contrôle de S. aureus
Ne pouvant pas g.arantir que les aliments, et plus particulièrement le lait et ses dérivés,
soient dépourvus d'entérotoxines, le nombre de S. aureus doit être contrôlé. Cette bactérie
est capable de croître dans le lait puisqu'elle possède, entre autres, des systèmes de
transport phosphotransferase et permeases lui permettant d'utiliser le lactose (34). Afin
d'inhiber la croissance de cette bactérie dans le lait et ses dérivés tout en maintenant la
biodiversité et l'écologie microbienne de l'environnement, plusieurs options ont été
examinées.
L'une des solutions envisagées est l'utilisation de cultures bactériennes protectrices dont
les bactéries lactiques. Ces dernières peuvent influencer la croissance de S. aureus en
modifiant physiquement et chimiquement l'environnement. Elles peuvent sécréter des
bactériocines comme la nisine et la pédiocine ou du peroxyde d'hydrogène ayant un effet
bactéricide à une concentration de l'ordre de 0,6 mmol. La compétition pour les nutriments
et le ratio (bactéries lactiques/S. aureus) peuvent aussi affecter la croissance de cette
dernière dans le milieu laitier (34). De plus, selon Even et a l (47), les bactéries lactiques
ont la possibilité de réduire le potentiel pathogène de S. aureus en modulant l'expression
de ses facteurs de virulence. Mais ceci n'est pas un fait général puisque l'expression de
l'entérotoxine A a augmenté avec l'ajout de bactéries lactiques (47). Cependant,
l'utilisation de culture bactérienne protectrice n'est pas souhaitable dans tout type
d'aliment.
Une autre solution est le traitement thermique des aliments lors de leur fabrication. En
effet, la pasteurisation du lait est efficace pour éliminer S. aureus (69) mais ceci n'empêche
pas une recontamination de l'aliment par des porteurs sains de S. aureus (106).
D'un autre côté, l'utilisation des huiles essentielles s'est avérée efficace pour lutter contre
les souches de S. aureus. En effet, la méthode de l'aromatogramme a révélé que la
croissance de S. aureus est inhibée par presque 40 huiles essentielles issues de plusieurs
plantes dont Cinnamomum cassia, Cinnamosma fragrans, Eucalyptus citriodora,
Eucalyptus dives, Eugenia caryophyllata et autres (42). Cependant, plusieurs huiles
essentielles sont à l'origine de réactions allergiques, d'irritation et de toxicité. De plus,

12
l'utilisation d'huiles essentielles change les propriétés organoleptiques des aliments (28).
Pour ce qui est de leur utilisation dans le lait, leur mécanisme d'action et leurs effets dans
celui-ci ne sont pas encore compris et peuvent résulter en un usage inapproprié des huiles
essentielles (29).
L'utilisation d'endolysines est également documentée dans la littérature scientifique (24,
85, 88). Celles-ci sont des enzymes muréolytiques encodées par le génome de
bacteriophages et synthétisées à la fin du cycle lytique pour lyser la cellule hôte et libérer
de nouveaux virions. Elles ciblent les liaisons de peptidoglycanes à la surface bactérienne.
Les endolysines sembleraient être utiles dans le biocontrôle des produits laitiers et avoir un
bon potentiel antimicrobien dans la prévention et le traitement d'infections causées par S.
aureus. En effet, l'endolysine a une activité antibactérienne rapide et puissante in vitro et in
vivo (24). Elle est active contre une bactérie (Gram positif seulement) indifféremment de sa
sensibilité aux antibiotiques. La probabilité de développer une résistance aux endolysines
est faible. L'endolysine d'un phage de S. aureus est uniquement active contre une même
espèce ou des espèces apparentées. Ainsi, elle a un spectre restreint approprié pour son
usage dans la biopréservation des produits laitiers. De plus, les endolysines dérivées de
phages isolés à partir d'un environnement laitier co-évoluent avec leur hôte et peuvent
donc spécifiquement cibler les souches de S. aureus trouvées dans ce même
environnement. Cependant, la possibilité de libération de différents composants pro-
inflammatoires pendant la lyse bactérienne, les coûts élevés liés à la production et à la
purification de l'endolysine ainsi que sa faible stabilité dans une matrice alimentaire, rend
leur utilisation limitée pour l'instant (24).
Les endolysines purifiées du phage H5 (appartenant à la famille des Siphoviridae) isolé du
lait cru (51, 88) et du phage K (appartenant à la fiamille des Myoviridae) (85), ont été
obtenues et testées contre différentes espèces de staphylocoques. L'endolysine du phage
H5 s'est avérée capable d'inhiber la croissance de S. aureus dans le lait pasteurisé. Son
activité lytique est efficace à pH 7,0 et à 37 °C Celle-ci est perdue à un pH inférieur à 5 et
à de hautes températures arrivant jusqu'à 72 °C (88). Pour ce qui est de l'endolysine du
phage K, celle-ci présente un spectre large à l'intérieur du genre Staphylococcus. Elle a

13
inhibée une variété d'espèces de staphylocoques isolées d'infections bovines et humaines y
compris les souches de Staphylococcus aureus résistantes à la méthicilline (85).
Finalement, une autre option est maintenant considérée, celle de l'utilisation de phages
complets comme d'éventuels agents de biocontrôle dans les denrées alimentaires.
1.2. Les bacteriophages
1.2.1. Historique
Les bacteriophages sont des virus infectant uniquement les bactéries. Ce sont des parasites
intracellulaires obligatoires. Ils ont été co-découverts par Frederick William Twort et Félix-
Hubert d'Hérelle. En 1915, Twort a constaté que les colonies de microcoques devenaient
vitreuses et transparentes à cause d'un agent filtrable et infectieux. Puis, en 1917, d'Hérelle
a montré qu'après incubation toute la nuit, la turbidité de la culture bactérienne avait
disparu laissant place à une solution limpide. De plus, il a constaté une absence de
croissance bactérienne sur boîte de Pétri déduisant ainsi que la cause derrière ceci était
probablement un virus filtrable, parasite de bactéries. Il lui donna le nom de bacteriophage
(46).
C'est en 1922 que les premiers travaux sur le traitement des infections à staphylocoques
avec des phages ont été rapportés. Ces travaux ont été publiés par Gratia lors d'une étude à
Bruxelles (56). Cependant, la phagothérapie a été abandonnée dans le monde occidental
avec la découverte de la pénicilline en 1945 (72). Mais c'est avec l'apparition des souches
bactériennes staphylococciques résistantes aux antibiotiques dans les années 1960 que
l'antibiothérapie a été remise en question (77).
Les phages sont présents partout sur notre planète. Ils se retrouvent dans les océans, le sol,
l'eau potable et la nourriture (63). En 1989, un groupe norvégien a surpris la communauté
scientifique lors de la publication d'un rapport révélant une haute concentration de phages
dans les eaux côtières, les lacs et les océans (22). En effet, les phages seraient les entités
biologiques les plus abondantes sur Terre avec une population totale estimée à plus de 1030
phages (5, 36, 63, 120). L'avènement du microscope électronique a favorisé la mise en

14
place de la première classification de phages en 1943 par Ernst Ruska (6). La classification
de phages contemporaine est dérivée de celle de Bradley proposée en 1967 (25). À ce jour,
plus de 5 500 phages ont été examinés en microscopie électronique (6) et les plus
importants sont conservés au centre de référence Félix d'Hérelle (http://www.phage,
ulaval.ca/).
1.2.2. Classification des phages
La classification de phages, mise à jour par 1TCTV (International Committee on
Taxonomy of Viruses), inclut présentement un ordre, celui des Caudovirales (qui
comprend tous les phages caudés ou portant une queue), 14 familles officiellement
reconnues (6) et 37 genres (49). Ces 14 familles en plus de 5 autres non classées jusqu'à
présent sont illustrées à la figure 6.
ssDNA
O dsDNA
99 9 Myovialdaa)
Cor tkov i r id i» Ttet iv i r ic l i i Podov.rida» SH1
Siphoviridat
P l U f i u v i r i d M Gtobulovirtdae'
O o Fi»a>lk>virld»a- Sd t t r p rov t ru *
Lipothrlxvlrldae
GutUvt- idM Ampu l l i v I r ldM
■Blc iud jv i r .d i» '
ssRNA dsRMA
O Cy&tovir fdK
Figure 6. Représentation schématique des principales familles de bacteriophages (6).
Les phages sont principalement classés d'après leur morphologie et la nature de leurs
acides nucléiques (3, 7, 98). La plupart des phages sont constitués d'ADN double brin. Une

15
minorité possède de l'ADN simple brin, de l'ARN double brin ainsi que de l'ARN simple
brin. Les phages peuvent être à symétrie cubique, filamenteuse, pléomorphique ou binaire.
Ceux ayant une symétrie binaire et possédant une queue prennent le nom de phages caudés
et appartiennent donc à l'ordre des Caudovirales. Ce dernier est très abondant et diversifié.
Il contient 96 % de tous les phages connus jusqu'à présent (6). Les phages caudés
proviennent de toutes les régions et environnements du monde. La plupart ont été isolés
dans les cultures bactériennes d'importance industrielle ou médicale comme les
entérobactéries, les lactocoques, les streptocoques et les staphylocoques (4). L'ordre des
Caudovirales est divisé en trois familles : Myoviridae, Podoviridae et Siphoviridae. Celles-
ci ont des propriétés morphologiques et génomiques différentes (6) (Figure 7).
O
Al
Myoviridae Siphoviridae 3 morphotypes
Cl cz
Podoviridae 2 morphotypes
Morphologie de la queue et de la
capside
Longue queue contractile,
capside et queue séparés par un
cou
Longue queue non contractile (capside
icosaédnqueou allongée dépendant
du morphotype)
Petite queue non contractile (capside
icosaédnqueou allongée dependant
du morphotype) Taille du génome 100 000 pb 50 000 pb 10 000pb
Figure 7. Caractéristiques morphologiques et génomiques des phages caudés.
Ces phages ont 3,5 milliards d'années comparativement aux virus d'animaux et de
végétaux qui dateraient de 700 millions d'années (8). Donc, les phages caudés ont
apparemment eu plus de temps pour se diversifier et se différencier. Cette diversification
peut être causée par l'accumulation des mutations ponctuelles, les réarrangements intra
génomiques (deletions, duplications, inversions et transpositions), les recombinaisons et les
co-infections avec d'autres phages (échange de segments génétiques) (8).

16
1.2.3. Phages de 5. aureus
Les phages de Staphylococcus aureus appartiennent tous à l'ordre des Caudovirales. Il
existe présentement 54 génomes de phages de S. aureus séquences dans la banque de
données publique GenBank du « National Center for Biotechnology Information » (NCBI)
(Tableau 1).
Tableau 1. Classification des phages de S. aureus selon les bases de données de NCBI
Myoviridae Podoviridae Siphoviridae Non classifies
K,G1, 44AHJD, phil 1, phi 12, phi 13,187,2638 A, 37,3 A, P954, PT1028,
Twort 66, P68, 42E, 55,77, 80alpha, phiPV83, CNPH82, ROSA,
SAP-2 EW, PHI5, PVL, phiETA, phiETA2, phi2958PVL,
phiETA3, phiMRl 1, phiMR25, phiN315, phiPVL-CN125,
phiNM, phiNM3, phiPVL108, phiSLT, tp310-l,tp310-2,
phiSauS-IPLA35, phiSauS-IPLA88, 53, 29,
47, 52A, 69, 71, 85, 88,92,96, X2
tp310-3
Toutefois, trois de ces génomes ne sont pas des phages de S. aureus. En effet, les phages
PHI5 et CNPH82 sont des phages de Staphylococcus epidermidis (40) et le phage Twort
est un phage de S. hyicus (113). Cependant ces phages partagent une identité génomique
élevée avec les phages de S. aureus. Le phage Twort partage 51 à 52 % d'identité
génomique avec les phages GI et K de S. aureus respectivement (64).
Les séquences nucléotidiques complètes des phages lyriques P68 et 44AHJD sont les
premières de S. aureus à avoir été publiées (116). En se basant sur leur morphologie, la
taille de leur génome, la similarité des produits de leurs gènes et le type d'infection, les
phages caudés P68 et 44AHJD sont regroupés dans la famille des Podoviridae. L'analyse
comparative des génomes complets des phages P68 (18 227 pb) et 44AHJD (16 784 pb) a
montré qu'une deletion de 1 440 nucleotides semble s'être produite dans le génome du
phage 44AHJD. Alternativement, une insertion pourrait avoir eu lieu aux positions 10 091
et 11 531 du génome de P68 (116).

17
D'autre part, une autre étude comparative a été réalisée sur le génome des phages tempérés
phil 1, phi 12 et phi 13 (60). Ceux-ci ont un génome variant de 42 000 à 45 000 pb et
contiennent 90 cadres de lecture dont environ 50 partagent de l'homologie avec des gènes
connus ou des gènes de d'autres phages de S. aureus. Récemment, les génomes de deux
nouveaux phages mutants, phiIPLA35 et phiIPLA88, ont été examinés et comparés à ceux
des phages phil 1 et phi 12. Ces premiers sont les dérivés virulents de deux phages isolés du
lait cru. L'analyse bioinformatique de ces deux nouveaux mutants a révélé que la structure
des génomes des phages phiIPLA35 (45 344 pb) et phiIPLA88 (42 526 pb) ressemble à
celle des phages phil2 et phil 1 respectivement (53). Une colinéarité de ces séquences a été
observée presque tout au long du génome. En effet, le phage phiIPLA35 est un
Siphoviridae de morphotype B2 possédant une longue queue non contractile et une capside
allongée tout comme les phages phi 12, 47, 3A, 42E, phiSLT, entre autres. Une plus faible
identité génomique est observée en comparant les génomes des phages phiIPLA88 et
phill. De manière générale, la séquence nucléotidique des gènes tardifs de ces deux
phages est plus conservée que celle des gènes précoces (53).
Le génome du phage virulent et polyvalent K a également été analysé en détails (87).
Celui-ci appartient à la famille des Myoviridae avec un génome de 127 395 pb et 118
cadres de lecture. Il est intéressant de souligner que le génome de ce phage ne contient
aucun site GATC le rendi t donc insensible aux enzymes de restriction qui reconnaissent
et clivent cette séquence. De plus, ses gènes structuraux sont hautement homologues à ceux
du phage A511 de la bactérie Listeria suggérant l'existence d'un transfert horizontal entre
les genres de bactéries. L'analyse du génome du phage K montre que celui-ci n'appartient
à aucun groupe déjà défini. Il est très faiblement apparenté aux Podoviridae PZA-like et
aux prophages plasmidiques de Borrelia burgdorferi (87). Ceci permet de le mettre dans un
groupe taxonomique distinct des autres (87).
1.2.3.1 Classification génomique des phages de S. aureus
Dans une étude réalisée par Kwan et a l , 27 phages de S. aureus ont été regroupés suivant
leur identité génomique en utilisant, entre autres, le logiciel DOTTER (Figure 8). Les
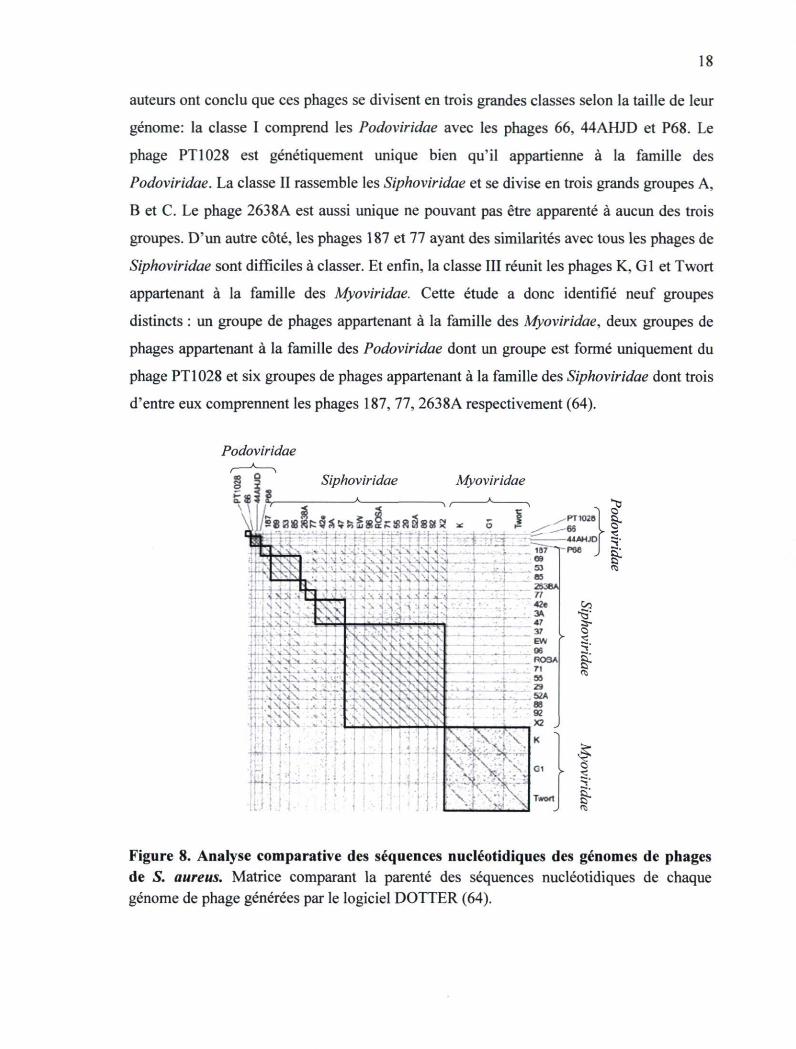
18
auteurs ont conclu que ces phages se divisent en trois grandes classes selon la taille de leur
génome: la classe I comprend les Podoviridae avec les phages 66, 44AHJD et P68. Le
phage PT1028 est génétiquement unique bien qu'il appartienne à la famille des
Podoviridae. La classe II rassemble les Siphoviridae et se divise en trois grands groupes A,
B et C. Le phage 2638A est aussi unique ne pouvant pas être apparenté à aucun des trois
groupes. D'un autre côté, les phages 187 et 77 ayant des similarités avec tous les phages de
Siphoviridae sont difficiles à classer. Et enfin, la classe III réunit les phages K, GI et Twort
appartenant à la famille des Myoviridae. Cette étude a donc identifié neuf groupes
distincts : un groupe de phages appartenant à la famille des Myoviridae, deux groupes de
phages appartenant à la famille des Podoviridae dont un groupe est formé uniquement du
phage PT1028 et six groupes de phages appartenant à la famille des Siphoviridae dont trois
d'entre eux comprennent les phages 187, 77, 2638A respectivement (64).
Podoviridae
Siphoviridae Myoviridae
1 / S î?
T "1 : ■
LL- . . . - :___•_ L _ — -: 45 -1
187 09 M K ■tfïmtirK rr 42e 3A 47 V EW 06 ROSA 71 M 29 52A » 92 X2
1 S3
-PTW28 O .
'« V$ -aUARIDl 5j*
-TO J S;
T*wt
I'm
S" ca <
I re
Figure 8. Analyse comparative des séquences nucléotidiques des génomes de phages de S. aureus. Matrice comparant la parenté des séquences nucléotidiques de chaque génome de phage générées par le logiciel DOTTER (64).

19
Une étude subséquente a ajouté à cette classification un nouveau grand groupe (D) de la
classe II combinant les différents phages appartenant à la famille des Siphoviridae (38).
Celui-ci a réunit les phages phi 13, PV83, PVL, N315 et 77; ce dernier était difficile à
classifier dans les études précédentes à cause de sa haute similarité avec les membres de
tous les groupes de la classe II. Les phages tempérés de S. epidermidis, CNPH82 et PHI5
ont été classés dans le groupe C de la classe II partageant une haute identité génomique
avec le phage phiETA. De plus, un arbre phylogénétique basé sur les distances
protéomiques a été conçu. Celui-ci compare tous les phages connus de Siphoviridae
appartenant à plusieurs genres bactériens (Figure 9). Il confirme les regroupements réalisés
après l'analyse génomique de phages staphylococciques (40).
Gram negative bacteria
>-* 1
i \ \ \
^ \ \ \ \
\\\ V .
. <m±. . - c \ ^
* N \ \ i ■
, LK - 4 ,
Lax-tocaccj and Streptococci
Figure 9. Arbre protéomique des phages appartenant à la famille des Siphoviridae. Figure tirée de Daniel et al, 2007 (40).

20
Une autre étude a permis de classer les prophages de S. aureus appartenant à la famille des
Siphoviridae (55). Cependant, cette classification est basée sur le polymorphisme du gène
de l'intégrase et non sur le génome complet des phages (55). De leur coté, Canchaya et al
ont pu assembler les 12 prophages de S. aureus, disponibles à ce moment, appartenant à la
famille des Siphoviridae et les diviser en 5 groupes distincts de modules structuraux grâce
à des matrices de type dot-plot. Trois groupes du genre Sfi21-like ayant des extrémités
génomique de type cos ont été identifiés: [1] PVL, PV83, phi 13, phi Sa3, [2] SLT, phiSa2,
phi 12 et [3] phiMu50A, phiN315 ainsi que deux groupes du genre Sfill-like ayant des
sites de types pac : [1] phiMu50B, phil 1 et [2] phiETA (30). D'autre part, Pantucek et al.
(90) ont réalisé le premier test moléculaire basé sur la méthode de la réaction en chaîne de
la polymerase (ou PCR) pour identifier et classifier les phages staphylococciques et
détecter les prophages dans le génome des souches lysogéniques de S. aureus. Après
l'alignement des séquences d'ADN de ces derniers, quatre régions conservées d'ADN ont
été identifiées dans tous les phages et prophages staphylococciques par les phages de types
de 3 A (représentant du sérogroupe A), 11 (sérogroupe B) et 77 (sérogroupe F) (90).
Cependant, un débat sur la taxonomie des phages est présentement en cours. Plusieurs
pensent que le type de classification établi par l'ICTV il y a plus de 30 ans, doit être révisé
avec la venue de l'âge post-génomique (81). Une classification alternative basée sur la
comparaison génomique et protéomique est envisagée (37, 98). La classification basée
surtout sur la morphologie a plusieurs inconvénients. En effet, les prophages ne sont pas
reconnus par l'ICTV bien que les phages lysogéniques existent. De plus, cette taxonomie
n'est pas en mesure de classifier tous les phages puisqu'ils n'ont pas tous été observés en
microscopie électronique, alors que le génome complet de beaucoup de ces phages non
classifies est déjà séquence. Contrairement à la bactérie et à l'ARN ribosomal, un phage ne
possède pas de gènes ou de séquences protéiques conservés et communs à tous les phages.
Une approche proposée par Rohwer et al. a permis de regrouper les phages et de construire
un arbre protéomique (98). Les solutions proposées sont loin d'être sans faille. La première
proposition se base sur la division du génome des phages en plusieurs modules, la
classification des modules par similarité des séquences, et la description des phages à l'aide
d'un code de chiffres qui représente chaque allele spécifique (67). Cependant, cette
méthode deviendrait complexe lorsque le nombre des différents alleles pour un module

21
spécifique est grand et que la taille du génome de ce phage est grande. Une autre méthode
ne prend pas en considération la structure des modules, et intègre le génome complet dans
un logiciel qui produit un .arbre protéomique de phages (98). La troisième méthode
proposée par Proux et al. compare les gènes structuraux entre eux (94).
1.2.4. Cycle de multiplication des phages
Comme il a déjà été mentionné plus haut, l'utilisation de phages pour la lutte contre les
souches de S. aureus est présentement à l'étude. Les phages les plus prometteurs sont
virulents. Ils suivent donc un cycle lytique menant à la lyse de l'hôte suite à leur
replication. Ce cycle comporte plusieurs grandes étapes dont l'infection de l'hôte, la
transcription des gènes de phages, la traduction de l'ARNm en protéines phagiques, la
replication de l'ADN de phage, l'encapsidation de ce dernier, l'assemblage des particules
de phage et enfin la libération des nouveaux virions (lyse) (Figure 10).
I yje cellulaire el ^ - ^ libération des *• irions J A
Adsorption rl éjection «Je l'ADN du phage
Sous conditions de stress, excision du prophage
l)u|>U aiiam bactérienne et transmission du prophage aux cellules filles
Synttiese drs protéines de structure cl encapsulation de l'ADN du phage
Lysogénie Replication de ^ ^ .— l'ADN du phage f f ^ | Intégration du génome du phage
dam le chromosome hocténen
Figure 10. Étapes des cycles des bacteriophages : cycles lytique et lysogénie. Figure
tirée et adaptée de Labrie (2004) (66).

22
Contrairement aux virus de cellules eucaryotes ayant un cycle lytique d'une durée d'une
heure, celui des bacteriophages peut s'achever en moins de 20 minutes (4).
L'infection débute par la reconnaissance et l'adsorption du phage à un récepteur spécifique
de la surface bactérienne. Dans le cas de S. aureus, les récepteurs de phages à la surface
des bactéries sont peu connus et le mécanisme de fixation du phage à son hôte est peu
caractérisé. De plus, il y a peu d'information sur les protéines phagiques jouant un rôle
dans la reconnaissance et la fixation du phage à S. aureus (61). Des récepteurs
hypothétiques ont été identifiées dont notamment la présence d'acides téichoïques (Figure
11) liés de façon covalente à la couche de peptidoglycane et traversant la paroi bactérienne
(118). La présence des résidus D-glucose et NAG (N-AcétylGlucosamine) ou même de 4-
0-p-(N-acétyl-D-glucosaminyl) dans la structure de l'acide téichoïque semble avoir un rôle
très important dans la reconnaissance du phage à son hôte (35, 39, 70, 115).
Peptidoglyea n- anc hored CWGs (P-CWGs)
Wall teichoic acid [Staphylococcus oireus]
-Peptidog!yc.3n
Membraneanchored CWGs (MCWGs)
Lipoteichotc *ad (S aurtui)
!8 SO
Figure 11. Structure des acides téichoïques. Les trioses, peptoses et hexoses sont représentés par des triangles, pentagones et hexagones respectivement. Les sucres, les alcools dérivés de sucres et les acides dérivés de sucres sont représentés en jaune, orange et violet respectivement. Les acides gras sont indiqués par des lignes en zigzag. N-acétylglucosamine (GlcNAc); M-CWG: glycopolymères de la surface bactérienne ancrés dans la membrane cytoplasmique; P-CWG: glycopolymères de la surface bactérienne ancrés dans la couche de peptidoglycanes. Résidus non-glycosylés: P, phosphate ; Rto, ribitol ; A, D-alanine ; Gro, glycerol ; ManNAc, jV-acétylmannosamine et Glc, glucose (118).

23
Le phage s'adsorbe de son côté à la surface bactérienne grâce à une protéine de liaison
(PLR) située à l'extrémité de sa queue (115). Cependant, celle-ci n'est pas encore
caractérisée chez les phages de S. aureus (61). Puis, la paroi cellulaire bactérienne est
digérée par une enzyme spécialisée du phage, présente au bout de sa queue (4). Moak et al.
ont noté dans leur étude la présence d'hydrolases à peptidoglycane chez quelques phages
de S. aureus appartenant à la famille des Siphoviridae : phil 1 et 85 (78). Plus récemment,
une étude a été réalisée par Rashel et al. (95) qui ont montré pour la première fois qu'un
phage de S. aureus, nommé OMR11, possède un gène (gp 61) codant pour ce type
d'hydrolase à peptidoglycane situé à l'extrémité de sa queue. En effet, cette dernière
possède deux domaines catalytiques dont le domaine amidase et le domaine lysozyme
situés dans les régions N-terminale et C-terminale respectivement. Ceux-ci ont une activité
lytique contre S. aureus et donc agissent en formant des trous dans la couche de
peptidoglycane de la bactérie permettant ainsi la pénétration de l'ADN phagique dans le
cytoplasme de S. aureus (95). L'ADN viral se circularise grâce à ses extrémités cohésives
ou soit par la recombinaison des bouts redondants (4, 115). Arrivé à cette étape, le phage
peut, s'il possède les gènes appropriés, poursuivre le mode lytique ou prendre le mode
lysogénique(115).
Dans ce dernier cas, l'ADN de phage s'intègre à l'ADN chromosomique bactérien et se
réplique passivement avec la cellule. Ce sont les phages virulents et les phages tempérés
devenus lyriques (suite à une induction) qui poursuivent le mode lytique. Ceux-ci débutent
p.ar la transcription de leurs gènes assurée par l'ARN polymerase de phage ou soit une de
l'hôte et se divisant en trois phases : précoce, intermédiaire et tardive (115). Les modules
des gènes précoces sont les premiers à être transcrits. Les produits de ces gènes ont une
fonction dans l'arrêt ou le contrôle du métabolisme de la bactérie, la dégradation de l'ADN
de l'hôte, la protection contre les enzymes de restriction, l'initiation de la replication de
l'ADN du phage et la synthèse des protéines de régulation essentielles pour la suite du
cycle lytique (4, 74). Pour plusieurs phages, la phase de transcription des gènes
intermédiaires et tardifs se déroule pendant plus de 15 minutes après l'infection (4). Le
produit des gènes intermédiaires joue un rôle dans la synthèse de l'ADN du phage et code
pour l'ADN polymerase nécessaire à l'étape de replication du phage. Cette dernière résulte
en l'obtention de concatémères soit de longues molécules d'ADN composées de multiples

24
copies d'un même génome (93). L'étape suivante est celle de la morphogénèse au cours de
laquelle les protéines structurales sont produites et assemblées pour former la capside et la
queue du phage et l'ADN est encapsidé (Figure 12). Ces protéines sont encodées par les
gènes compris dans le module des gènes tardifs (4).
» 1
î @ O dp
Ll
Q 4- Tail protwnt
i
0 -Q
Figure 12. Voie d'assemblage des protéines structurales des phages caudés (4).
L'encapsidation représente la dernière étape dans la maturation de l'ADN de phages
caudés et elle est reliée à l'expansion de la procapside formée d'une tête protéique vide.
C'est un processus complexe au cours duquel l'ADN est introduit à haute vitesse dans la
procapside. Cette étape est réalisée grâce à une enzyme appelée terminase qui, dotée d'une
activité endonucléase, ATP dépendante, se lie au concatémère par sa petite sous-unité,
coupe l'ADN et le transporte dans la capside préformée grâce à sa grande sous-unité au
prix de l'hydrolyse d'ATP (115). Le site de clivage de l'ADN diffère d'un phage à l'autre
(4). En effet, chez les phages de type pac, l'ADN encapsidé commence à un site pac et
continue jusqu'à ce que la procapside soit remplie d'ADN suivant le mécanisme de la tête
pleine. Pour le phage T4, environ 2 % du génome est additionné au génome entier dans la
procapside (93). En effet, la terminase coupe successivement le concatémère à des
distances toujours un peu plus loin que le site pac entraînant ainsi l'obtention d'extrémités
redondantes. Cependant, pour les phages de type cos (extrémités cohésives), l'ADN
commence et se termine par un site cos reconnu par la terminase (63, 101). Ainsi, une seule

25
copie linéaire et complète du génome est retrouvée à l'intérieur de la capside de ces
phages.
Enfin, le cycle lytique s'achève par la lyse cellulaire de l'hôte suite à l'action de la holine
et de l'endolysine ou aussi appelée hydrolase à peptidoglycane. Leurs gènes sont aussi
retrouvés sur le module tardif des gènes viraux (4). La holine cause des lésions non
spécifiques dans la membrane cytoplasmique de l'hôte, facilitant ainsi le passage de
l'endolysine vers la couche de peptidoglycane ce qui entraîne sa dégradation. La
dégradation se fait grâce à quatre classes d'endolysines : les muramidases, les amidases, les
peptidases et les transglycosylases. Habituellement, on retrouve un seul gène codant pour
une holine ou endolysine par génome de phage. À la suite de ce stade, la lyse de la cellule
hôte permet le relargage de nouveaux virions (4, 101, 122).
La plupart des phages tempérés passent par un cycle lytique suite à une induction (UV ou
autre). Ceci est manifesté par l'excision et la libération de l'ADN de phage grâce à une
excisionase (4, 93, 115). Cependant, d'autres phages suivent le mode lysogénique (Figure
10). Cette étape consiste en l'intégration de leur génome dans le chromosome bactérien
grâce à l'intégrase, une enzyme de type recombinase (115). Le maintien du génome viral
dans le chromosome bactérien est assuré par un répresseur. Le répresseur se lie à des
opérateurs phagiques et bloque l'activité de l'ARN polymerase, inhibe la synthèse
d'ARNm de d'autres phages qui tentent d'infecter l'hôte et assure une immunité de la
cellule infectée contre ceux-ci (93). Le phage entre donc en un stade de latence (prophage),
se protège de l'environnement et se réplique comme étant une partie du génome bactérien.
1.3. Applications des phages de S. aureus
Les phages commencent à être utilisés pour le contrôle des bactéries pathogènes dans
l'aliment ainsi que dans des sujets animaux. Les phages virulents ont plusieurs avantages
comparés aux phages tempérés (77). Une souche lysogène (contenant un prophage) est
également immune à une infection par un phage tempéré similaire au prophage. De plus, la
fréquence d'apparition de BIM (Bacteriophage Insensitive Mutant) est moindre avec les
phages virulents (51). Les BIM sont des bactéries qui acquièrent naturellement une

26
résistance à un phage. L'émergence de BIM est présumément associée à des mutations
ponctuelles dans les gènes codant pour les récepteurs de phage à la surface bactérienne.
D'un autre côté, les phages tempérés, devenus prophages, peuvent aussi transférer, entre
autres, des gènes codant pour la production de toxines dans les populations bactériennes
par conversion lysogénique (53). De plus, les phages virulents ayant un large spectre
d'action ou dits polyvalents sont ceux utilisés lors des applications de biocontrôle et de la
thérapie. Ils ont la possibilité de cibler plusieurs souches bactériennes de la même espèce
ce qui représente un net avantage (12, 77, 87, 91). Par conséquent, ce sont les phages
virulents qui sont utilisés comme additifs alimentaires et pour la prévention ou le
traitement (1,91).
1.3.1. Thérapie par les phages
Les phages ont été testés chez des animaux pour montrer leur capacité à lyser des cellules
pathogènes et donc à protéger l'organisme contre les infections. Pour ce qui est de
l'application des phages de S. aureus, au début du 20ieme siècle, le laboratoire parisien
commercial de d'Hérelle a commencé à produire au moins cinq préparations de phages
contre des infections bactériennes multiples dont celles causées par S. aureus. Ces
dernières ont pris le nom de Bacté-staphy-phage et ont été commercialisées par la
compagnie qui a pris le nom récent de L'Oréal. De même, dans les années 1940, la
compagnie américaine Eli Lilly d'Indianapolis a produit sept préparations différentes de
phages pour usage humain contre les staphylocoques sous le nom de Staphylo-lysate ou
Staphylo-jel. Celles-ci ont été utilisées pour remédier aux infections comme les abcès, les
plais suppuratives et autres maladies associées à cet organisme (109). Plus récemment,
l'efficacité de trois phages virulents a été testée contre les infections à S. aureus chez les
animaux. Le phage 9812 est un membre de la famille des Myoviridae et possède un large
spectre lytique. Pantucek et al. ont montré que ce phage peut lyser 95 % des 782 souches
de 5. aureus testées; ces souches provenant de divers environnements dont des hôpitaux
(91). D'autre part, 39 sur 53 souches de S. aureus résistantes à la méthicilline sont
sensibles au phage K, un autre phage appartenant à la famille des Myoviridae. Celui-ci est
également efficace contre les souches de S. aureus résistantes à la vancomycine et à la
téicoplanine (86, 96). Enfin, le phage 44AHJD est également efficace contre les infections
de S. aureus. C'est un phage virulent appartenant à la famille des Podoviridae (116).

27
1.3.2. Applications alimentaires des phages de 5. aureus
L'utilisation des phages comme additifs alimentaires a été approuvé par la FDA pour
contrer Listeria monocytogenes (114, 117). Ces phages n'affectent que les bactéries non
désirées dans les aliments. De plus, ils ne changent pas les propriétés organoleptiques des
produits alimentaires (goût, structure, couleur, odeur) (57). Les phages sont omniprésents
autant dans l'environnement, les aliments ainsi que dans le système digestif. Jusqu'à
présent, aucun mélange de phages n'a été produit et reconnu comme sécuritaire pour lutter
contre S. aureus au niveau d'une industrie alimentaire et donc comme agent de biocontrôle.
Néanmoins, différentes méthodes d'utilisation de phages dans du lait et d'autres aliments
contaminés par des S. aureus ont fait l'objet de recherches spécifiques. La suite de la
présente section sera focalisée sur les applications faites d.ans le lait.
1.3.2.1. Test de l'activité des phages d.ans le lait pour remédier à S. aureus
Des études ont révélé le potentiel lytique des phages contre S. aureus dans du lait traité à la
chaleur, du lait cru ainsi que du lactosérum (41, 54, 84). Dans la plupart de ces applications
laitières contre S. aureus, le phage K a été utilisé. Celui-ci est un phage polyvalent
appartenant à la famille des Myoviridae et, comme mentionné auparavant, ayant un large
spectre lytique (57). Il utiliserait les NAG (N-AcétylGlucosamine) à la surface des acides
téichoïques pour l'adsorption (87). Selon Chatterjee et a l (35), les cellules bactériennes
testées, résistantes au phage K et dépourvues de NAG n'ont pas été capables d'inactiver ce
phage. Dans ces études, les auteurs ont constaté que le phage K est fortement inhibé dans le
lait cru entier. Gill et al. ont tenté d'identifier la cause de cette inhibition en étudiant l'effet
du lactosérum obtenu à partir de lait cru (54). Le lactosérum est la fraction protéique
soluble résiduelle du lait après avoir retiré le gras et la caséine. Elle contient plusieurs
protéines dont la P-lactoglobuline, l'a-lactalbumine, la sérumalbumine, les
immunoglobulines et les protéose-peptones. Des photos de cellules de S. aureus incubées
dans le lactosérum de lait cru et dans du t.ampon ont été obtenues par microscopie à
épifluorescence et par microscopie électronique à balayage (Figure 13).

28
Figure 13. Micrographies prises par microscopie électronique à balayage, (a) Cellules incubées dans du lait cru entier pendant 10 minutes et lavées à deux reprises, (b) Cellules incubées dans du tampon contenant 10 mg/ml d'albumine de sérum bovin et lavées à deux reprises (54).
Ces dernières suggèrent qu'une composante du lait pourrait masquer le récepteur et être à
la base de cette inhibition. Lorsque le lactosérum a été traité par la chaleur, par la
proteinase K ou par ultrafiltration, l'adsorption des phages a augmenté et est devenue
comparable à celle obtenue dans du milieu nutritif TSB. Ceci montre donc que le composé
inhibiteur serait une protéine ou un groupe de protéines qui s'attachent à la surface de S.
aureus ou modifie cette dernière de façon à empêcher les phages de venir s'y adsorber. Des
cellules bactériennes ont ensuite été exposées au lactosérum puis remises dans du milieu
nutritif et testée contre le phage K. Le nombre de colonies formées a diminué de presque
10 % en comparaison avec le nombre obtenu dans du lactosérum cru. Ceci montre que le
phage n'a pas été dégradé, mais que son activité a été inhibée en présence d'un composant
du lactosérum cru. D'autre part, une chromatographie des fractions de lactosérum a révélée
que la taille des protéines inhibitrices est de 1 500 KDa, ce qui suggère que
l'immunoglobuline majeure IgGl (180 kDa) de lait bovin n'est pas responsable de cette
inhibition (54).
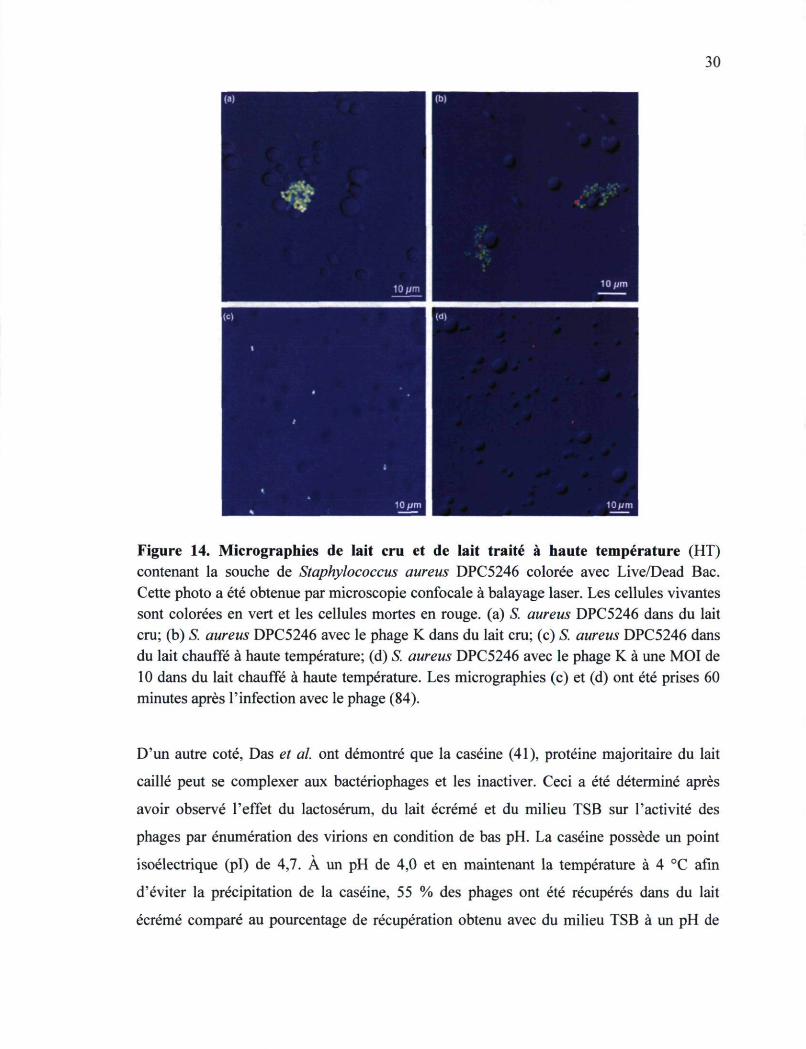
Dans une deuxième étude menée par O'Flaherty et al, l'activité lytique du phage K a été
testée contre une souche de S. aureus résistante à la méthicilline poussée pendant une durée
de quatre heures dans du lait chauffé à haute température et dans du lait cru (84). Les
résultats ont également indiqué que l'adsorption du phage K est inhibée dans le lait cru

29
compté au lait chauffé où l'adsorption phagique est restaurée. Pour mieux visualiser la
cause de cette inhibition, des photos de cellules d'une souche de S. aureus isolée de
mammite bovine prises une heure après l'addition du phage K par microscopie confocale
ont été obtenues (Figure 14). Le système de coloration Live/Dead Bac a été utilisé où les
cellules bactériennes vivantes sont vertes et les cellules bactériennes mortes ou lysées par
le phage K sont rouges. Les photos obtenues ont dévoilé l'apparition d'une agglutination
des cellules bactériennes dans le lait cru, ces dernières étant toujours vivantes une heure
après l'ajout du phage K (Figure 14). Cette même constatation a été faite auparavant par
Mamo et al. (75).
Cet agglomérat de cellules a masqué la surface des cellules la rendant inaccessible aux
phages. À l'inverse, les cellules bactériennes de S. aureus ne sont pas agglomérées dans du
lait chauffé à haute température. Après une heure d'infection avec le phage K à une MOI
(multiplicité d'infection) de 10, presque aucune cellule n'est restée vivante (84). Donc, le
composé causant cette agrégation est thermosensible et favorise l'agrégation des cellules.
Les auteurs ont suggéré qu'il pourrait probablement être une agglutinine ou appartiendrait
à la famille des immunoglobulines (62, 71).
30
I I I J H
* » i. * H
Figure 14. Micrographies de lait cru et de lait traité à haute température (HT) contenant la souche de Staphylococcus aureus DPC5246 colorée avec Live/Dead Bac. Cette photo a été obtenue p.ar microscopie confocale à balayage laser. Les cellules vivantes sont colorées en vert et les cellules mortes en rouge, (a) S. aureus DPC5246 dans du lait cru; (b) S. aureus DPC5246 avec le phage K dans du lait cru; (c) S. aureus DPC5246 dans du lait chauffé à haute température; (d) S. aureus DPC5246 avec le phage K à une MOI de 10 dans du lait chauffé à haute température. Les micrographies (c) et (d) ont été prises 60 minutes après l'infection avec le phage (84).
D'un autre coté, Das et al. ont démontré que la caséine (41), protéine majoritaire du lait
caillé peut se complexer aux bacteriophages et les inactiver. Ceci a été déterminé après
avoir observé l'effet du lactosérum, du lait écrémé et du milieu TSB sur l'activité des
phages par enumeration des virions en condition de bas pH. La caséine possède un point
isoélectrique (pi) de 4,7. À un pH de 4,0 et en maintenant la température à 4 °C afin
d'éviter la précipitation de la caséine, 55 % des phages ont été récupérés d.ans du lait
écrémé comparé au pourcentage de récupération obtenu avec du milieu TSB à un pH de

31
6,5. Il est donc possible qu'à un pH plus bas que le pi, ces derniers soient libérés de leur
complexe avec la caséine (41).
Une autre explication serait la formation d'une microcapsule à la surface des souches de S.
aureus (isolées de mammite bovine) après une incubation de 2 heures dans du lactosérum.
Incubées dans ces conditions, ces bactéries induisent la production d'une structure anti-
phagocytaire à leur surface. Celle-ci est hydrophile et résiste au traitement à la chaleur et à
la protease. Cependant, elle est altérée par un traitement au Périodate IO4" (composé
modifiant les cycles saccharidiques) et est donc probablement de nature saccharidique (76).
Ce matériel capsulaire hydrophile serait à l'origine du camouflage de l'organisme et donc
de sa persistance dans le milieu. Il serait attaché à la surface membranaire via une structure
sensible au Périodate comme les acides téichoïques (75, 76, 102). Cette structure pourrait
donc se comporter comme une barrière et empêcher l'interaction entre les phages et les
récepteurs à la surface de S. aureus (119).
1.3.2.2. Utilisation de phages isolés du lait cru et production de cocktails de phages
Certains ont émis l'hypothèse que les phages isolés du lait cru seraient plus efficaces que
d'autres dans le contrôle des bactéries pathogènes isolées du même milieu (27).
Récemment, deux phages tempérés ont été isolés à partir d'échantillons de lait cru : OH5 et
OA72 (51). Ils appartiennent à la famille des Siphoviridae et leur génome d'ADN double
brin consiste en une molécule linéaire. Les dérivés lyriques de ces derniers, nommés
phiIPLA35 et phiIPLA88, ont été testés contre les souches de S. aureus Sa9 dans différents
types de lait. L'expérience a été produite en testant l'efficacité de chaque phage séparément
puis en testant le mélange des deux, et ce, dans les laits cru, pasteurisé et UHT (Ultra Haute
Température), entiers et écrémés et ainsi qu'à différentes températures et MOI (52). Ces
phages ont infecté les souches de S. aureus d'origine laitière et bovine alors que les isolats
cliniques semblent y être résistants. Les auteurs ont ensuite démontré que le mélange de
phages a une plus grande efficacité sur l'inhibition des souches bactériennes de S. aureus
que l'action d'un seul phage. Le cocktail de phages est capable de couvrir un spectre
d'hôtes plus large et pourrait éviter l'apparition de résistance bactérienne. Les deux phages
se sont bien adaptés à l'environnement laitier et ont efficacement inhibé la croissance de S.

32
aureus dans le lait ainsi que dans le lait caillé en cours de fabrication. Cela a été efficace à
des températures de 4 °C, 22 °C et 37 °C (51, 52) confirmant l'utilisation potentielle des
phages comme des agents de biocontrôle dans cette matrice alimentaire. Cependant, leur
biosécurité n'a pas été caractérisée puisque ces phages contiennent des gènes
potentiellement toxiques pour l'humain (53). En effet, un motif du nom de VirE trouvé
dans la protéine bactérienne de virulence E a été détecté dans le produit de gène gp32 du
phage phiIPLA35. Ce motif a été décrit dans plusieurs protéines de replication d'ADN de
phage et pourrait avoir un rôle particulier dans le métabolisme de l'ADN. Cependant, ceci
n'exclu pas le fait qu'il pourrait aussi avoir un rôle putatif dans la pathogénicité de S.
aureus (53).
La question de pH a aussi été traitée par Garcia et al. (51). L'activité lytique du cocktail de
phages a été testée contre des souches de S. aureus à différents pH dans du lait caillé à
25 °C pendant une incubation de 12 heures. À bas pH, le mélange de phages a été
partiellement inactivé entre 4 heures et 6 heures d'incubation à 25 °C. De plus, une
différence de concentration de phages a été observée entre le lait caillé obtenu après l'ajout
d'acide et celui obtenu après l'ajout enzymatique de présure (chymotrypsine). La
concentration phagique, utilisée dans le lait caillé obtenu après l'ajout d'acide, a diminué
de presque 2 unités de log pend.ant une période de 12 heures tandis que celle trouvée par le
lait caillé obtenu après l'ajout de chymotrypsine est restée stable tout au long de cette
même période d'incubation. Le pH semblerait être donc un paramètre clé dans le processus
d'inactivation des phages dans le lait (51). Aucune référence traitant de l'effet du pH dans
le lactosérum n'a été trouvée dans la littérature.
1.4. Problématique à l'étude et objectifs de recherche
L'utilisation des phages de S. aureus et les conséquences reliées à leur utilisation ne sont
pas encore claires. Cependant puisque les phages ont déjà été utilisés comme agents de
biocontrôle, et que le phage K, d'origine inconnue, n'a pas pu lyser les souches de S.
aureus dans le lait cru, il serait intéressant de tester l'activité de phages isolés du lait cru
dans l'élimination des souches de S. aureus. Dans cette étude, un nouveau phage a été isolé

33
du lait cru. Il a ensuite été caractérisé et comparé aux autres phages de S. aureus et ce, aux
niveaux microbiologique et génomique. Et enfin, deux phages ont été choisis pour
l'application alimentaire et leur activité contre une souche de S. aureus isolée de fromage
de lait cru a été testée dans des laits UHT et cru.

34
2. Matériel et méthodes
2.1. Objectif 1. Isolement de nouveaux phages de S. aureus à partir
d'échantillons de lait cru
La présence de phages de S. aureus a été testée dans six échantillons de lait cru provenant
d'autant de fermes différentes :
B-l: Lait cru d'un troupeau de suisse brune de la région de Chaudière-Appalaches
B-2: Lait cru d'un troupeau de Holstein de la région de Chaudière-Appalaches
B-3: Lait cru d'un troupeau de Holstein de la région de Chaudière-Appalaches
B-4: Lait cru d'un troupeau de Holstein de la région de Chaudière-Appalaches
B-5: Lait cru d'un troupeau de Holstein de la région de Chaudière-Appalaches
CRDA: Lait cru de consommation, industriel, de la région de la Montérégie
Ceux-ci ont été obtenus via le laboratoire du Professeur Steve Labrie (Université Laval).
Les éch.antillons ont été centrifugés pour éliminer une quantité plus ou moins importante de
gras. Par la suite, deux amplifications des phages potentiellement présents ont été réalisées
en utilisant 5 ml d'échantillons dans 5 ml de milieu TSB deux fois concentré et en utilis.ant
un isolât bactérien mutant d'une souche isolée de fromage de lait cru procurée par le
MAPAQ (numéro de référence # 44635101). Cette souche mutante est résistante au phage
QW de S. aureus. Pour la suite de cet objectif, le nom BIM 01-QW sera adopté. Ce BIM a
été choisi afin d'élargir le spectre lytique de notre collection de phages. Le contrôle
d'amplification de phages utilisé dans cette expérience a été d'ajouter intentionnellement le
phage staphylococcique cp812 lors des amplifications afin de s'assurer du bon
fonctionnement des étapes du protocole. Après une filtration de la deuxième amplification,
la présence de phages a été testée à l'aide d'une titration : 100 pi de la solution bactérienne
du mutant BIM 01-QW ainsi que 100 pi du filtrat ont été ajoutés à 3 ml de gélose molle
contenant 0,75 % d'agar. Le mélange a été versé sur une boîte de Pétri et incubé pour la
nuit à 37 °C. Ensuite, des plages de lyse ont été isolées et amplifiées avec la souche hôte
BIM 01-QW. L'amplification de la plage de lyse isolée a été par la suite titrée afin
d'obtenir le compte du nombre de phages. Un titre supérieur à IO8 UFP/ml permet de

35
visualiser les phages en microscopie électronique et d'obtenir une concentration d'ADN
suffisamment élevée pour poursuivre la caractérisation génomique du phage.
2.2. Objectif 2. Caractérisation du nouveau phage isolé
Une observation microscopique du phage a permis de connaître son morphotype et de le
classer dans une des familles connues de phages. De plus, le spectre lytique de ce phage
ainsi que l'analyse de son génome ont permis de caractériser le phage aux niveaux
microbiologique et génomique.
2.2.1. Observation des phages en microscopie électronique
Le lysat du phage à observer a été centrifugé à température ambiante et à vitesse maximale
pendant 10 minutes puis filtré à l'aide d'un filtre de 0,45 pm. Pour l'observation du phage
en microscopie électronique, une étape de lavage du filtrat de phages a été réalisée. Elle
consiste à centrifuger 1,5 ml du filtrat de phages à une concentration d'au moins IO8
UFP/ml à 4 °C, à 17 000 rpm pendant 1 heure. Ensuite, le surnageant a été retiré
délicatement en gardant 100 pi du filtrat au fond du tube. Le lavage des 100 pi restants a
été effectué avec de l'acétate d'ammonium 0,1 M (pH 7,0) à deux reprises à la température
de la pièce. Un volume de 100 pi a été conservé afin de préparer les grilles pour
l'observation des phages en microscopie électronique (44). Par la suite, une aliquote de 7,5
pi de celui-ci a été déposé sur une grille en cuivre et recouverte d'une couche de
Formvar/Carbone (Compagnie Ted Pella). Une coloration au PTA (2 %) ou au UA (2 %) a
été faite et, après élimination du liquide à l'aide d'un papier buvard, la grille a été
introduite dans le microscope électronique à transmission JEOL JEM-1230 pour
l'observation (44). Ce dernier est situé à l'Institut de Biologie Integrative et des Systèmes
(IBIS) de l'Université Laval. Les observations ont été effectuées à 80 kV et la prise de
photo a été faite avec une caméra numérique.

36
2.2.2. Spectre lytique du nouveau phage
Pour caractériser ce nouveau phage au niveau microbiologique, son spectre lytique a été
réalisé contre les différentes souches et isolats de S. aureus disponibles dans notre
laboratoire (Tableau 2).
Tableau 2. Les différentes souches de S. aureus disponibles au laboratoire et leur provenance
PROVENANCE NOM DE LA SOUCHE Collection Félix d'Hérelle HER 1101, HER 1049, HER 1225, SA812, A170,
HER 1226, HER 1238 MAPAQ 01,09 Hôpitaux canadiens Dix souches de S. aureus résistantes à la méthicilline
(SARM) (38) Laboratoire 13 souches résistantes aux trois familles de phages
Les différentes souches mutantes obtenues et isolées au laboratoire à la suite d'un test de
dépôt de la goutte, sont résistantes à plusieurs phages de S. aureus disponibles dans la
collection Félix d'Hérelle. Celles-ci ont pris le nom de 01-QW, 09-QW, 01-P68, 09-P68,
01-44AHJD, 09-44AHJD, 09-Msa, 01-K, 01-LH1, 1049-Pyophage, 1049-Team 2, 1049-
(p812etA170-Team2.
Le filtrat du phage a été dilué (1/100) pour éliminer une possible activité de l'endolysine.
La souche cible a été inoculée dans du milieu TSB et incubée à 37 °C pour la nuit. Ensuite,
10 UFC/ml de bactéries ont été ajoutées à 1 ml de TSB et pipetées sur une boîte de Pétri
contenant 1,5 % d'agar. Une aliquote de 5 pi de cette dilution a été déposée sur le tapis
bactérien de la souche cible. Cette expérience a été répétée au moins trois fois pour
confirmer les résultats obtenus. Après environ 6 heures d'incubation à 37 °C, la présence
d'une plage de lyse à l'endroit où l'aliquote a été déposée indique que le phage est capable
de lyser la souche cible. L'absence de celle-ci signifie que la souche testée est résistante au
phage ou bien que le phage n'est pas capable de terminer son cycle lytique dans cette
souche.

37
2.2.3. Séquençage d'ADN
L'ADN du nouveau phage (nommé LH1) a été isolé en utilisant la trousse «Qiagen
Lambda Maxi Kit » (Qiagen). À l'aide d'un spectrophotomètre de type NanoDrop 2000
(Thermo Scientific), la concentration de cet ADN a été estimée. Une digestion avec
l'endonucléase EcoRV a été effectuée afin de confirmer l'identité de l'ADN isolé. Pour ce
faire, environ 500 ng d'ADN ont été mélangés à 10 U d'enzyme de restriction (Roche) et
du tampon B [10 mM Tris-HCl, 0,1 M NaCl, 5 mM MgCl2, 1 mM 2-mercaptoéthanol, pH
8,0]. Huit microgrammes de l'ADN du phage LH1 ont été envoyés sous forme suspendue
dans 50 pi de tampon EB [10 mM TrisHCl (pH 8,0)] (Qiagen) au service de séquençage du
centre d'innovation de Génome Québec et de l'Université de McGill situé à Montréal. Ce
dernier utilise l'appareil Genome Sequencer FLX Titanium (Roche) pour le séquençage.
Par la suite, une marche sur le génome a été effectuée avec des amorces synthétiques
(Invitrogen) afin de rassembler les différents contigs obtenus du service de séquençage et
obtenir une séquence génomique complète (Tableau 3).
Tableau 3. Liste d'amorces utilisées pour l'assemblage des contigs de l'ADN du nouveau phage isolé
amorce 1 (5'- 3') amorce 2 (5'- 3') contig 1 AAGTTCATCTCTTAACCGAC ACAACTGCAACTGTAATGGC contig 2 TTTTAGAACAGGAGAGAGTG TGCAGAAGTGTCCTCTATTG
■ contig 3 CCATTGCTTTCGCACGATCC GCTGGTCAAGTCATGAATCG contig 4 GTCAGTTTGTTATAGCTGTC CACTAAACGCAGTCTTGGAC
Les séquences ont été assemblées à l'aide des logiciels Staden
(http://staden.sourceforge.net/) et BioEdit 7.0.9.0
(http://www.mbio.ncsu.edu/BioEdit/bioedit.html) (58). Les cadres de lecture ouverts ont
été détectés avec l'outil « ORF Finder » disponible sur le site Internet de NCBI
(http://www.ncbi.nlm.nih.gov/) ainsi qu'avec le logiciel GeneMark™ 2.5
(http://exon.biology.gatech.edu/) (23). La fonction potentielle de chacune des protéines a
été déduite par homologie à l'aide du programme BLAST 2.2.23 disponible sur le site de
NCBI (10). Et enfin, le point isoélectrique (pi) et la masse moléculaire (MM) de chacune
des protéines de phage déduites ont été obtenus en utilisant Compute pI/Mw
(http://ca.expasy.org/tools/pi_tool.html).

38
2.3. Objectif 3. Comparaison du nouveau phage isolé aux autres phages
de S. aureus
Le génome du nouveau phage LH1 a été comparé aux génomes de phages de S. aureus
déjà disponibles dans la base de données publiques GenBank. La caractérisation et
l'analyse bioinformatique ont permis le regroupement des phages selon la similarité des
profils de restriction et leur identité génomique. Ceci permettra de faire un choix éclairé
des phages à utiliser dans le lait. De plus, le phage ayant le spectre lytique le plus large sera
considéré comme un agent de biocontrôle potentiel et sera utilisé ultérieurement dans
l'application alimentaire.
2.3.1. Profils de restriction du génome des phages de S. aureus
Dix autres phages de S. aureus, disponibles au Centre de référence Félix d'Hérelle pour
virus bactériens, ont été comparés au nouveau phage isolé (Tableau 4).
Tableau 4. Onze phages caractérisés dans cette étude, leur provenance et leur souche hôte de S. aureus
PHAGES PROVENANCE SOUCHES HÔTES
mj
■S 1 t
Pyophage Géorgie
SA812 mj
■S 1 t
Team 1 Géorgie SA812 mj
■S 1 t
Team 2 Géorgie
SA812
mj
■S 1 t phi812 République Tchèque
(91) SA812
mj
■S 1 t
K Belgique (56) SA812
|
1 P68 France (99) 68 (HER 1049)
|
1 QW Eaux Usées du Québec 68 (HER 1049)
|
1 44AHJD États-Unis (99) 44A (HER 1101)
1 :s s a-03
3A Canada HER 1225 1 :s s a-03
Msa Italie (31) Al 70

39
Dans le tableau 4, les dix phages disponibles dans notre laboratoire sont énumérés ainsi
que leur provenance et leurs souches hôtes respectives. Tout d'abord, chaque phage a été
amplifié et titré sur sa souche hôte dans du milieu TSB. Le titre doit être supérieur à IO8
UFP/ml pour obtenir une préparation assez concentrée afin d'isoler l'ADN phagique. Puis,
l'ADN a été extrait suivant le guide accompagnant la trousse lambda de Qiagen. Le dosage
de l'ADN obtenu a été effectué à l'aide du NanoDrop 2000. Par la suite et afin d'établir le
profil de restriction de l'ADN de chaque phage, celui-ci a été digéré avec l'enzyme
EcoRV. Une migration sur gel d'agarose 0,8 % (P/V) a été effectuée pendant environ une
heure à 100 V. Enfin, les bandes d'ADN obtenues ont été colorées au bromure d'éthidium
0,5 pg/ml et visualisées aux UV avec l'appareil Gel Doc (BioRad). Les profils de
restriction de chaque phage ont été comparés entre eux afin de pouvoir regrouper les
phages ayant un profil similaire.
2.3.2. Spectre lytique des phages
Le spectre lytique de chaque phage a été réalisé contre toutes les souches de S. aureus
disponibles, la méthodologie étant décrite plus haut. Les spectres ont été comparés entre
eux et avec celui du nouveau phage LH1 pour identifier le phage capable de lyser le plus
de souches de S. aureus tout en étant compatible avec LH1.
2.3.3. Test d'adsorption des phages à des cellules de S. aureus
Certaines souches testées dans le spectre lytique sont des souches mutantes résistantes aux
phages qui sont des dérivés naturels de souches sauvages sensibles aux phages. La
résistance peut être due à plusieurs facteurs liés au cycle de multiplication du phage. S'il
s'agit de BIMs d'adsorption, c'est l'interaction du phage avec le récepteur bactérien qui
serait bloquée et le phage n'arriverait plus à s'adsorber à la surface de la bactérie. Afin de
déterminer si les BIMs obtenus étaient des BIMs d'adsorption, un test d'adsorption des
phages a été réalisé. Un lysat de phages à une concentration de 103 UFP/ml a été ajouté à
différentes cultures bactériennes inoculées dans du milieu TSB ayant une D.O.600 située
entre 0,6 et 0,8 (correspondant à une concentration bactérienne de plus de IO8 UFC/ml). Le
surnageant a été recueilli après 15 minutes d'incubation à 37 °C et centrifugé pour une

40
minute à vitesse maximale à la température de la pièce. Le pourcentage d'adsorption s'est
fait par ce calcul :
titre du surnageant contrôle tampon - titre du surnageant cellules titre du surnageant contrôle tampon xlOO
Plus le pourcentage d'adsorption est élevé, plus le phage s'adsorbe efficacement à la
cellule et donc dans ce cas, la souche mutante ne serait pas un mutant d'adsorption.
2.3.4. Analyse bioinformatique des génomes de phages
Il existe jusqu'à présent 54 génomes complets de phages de staphylocoques disponibles
dans les bases de données publiques (GenBank). Plusieurs de ces phages, dont le génome
est à ADN double brin, ne sont pas classés dans les familles connues (Tableau 5). Les
phages soulignés dans le tableau 5 ne sont pas des phages de S. aureus. Cependant, ceux-ci
partagent une identité génomique élevée avec les phages de S. aureus, d'où leur inclusion
dans cette étude (40, 64).
Tableau 5. Les génomes de phages de staphylocoques disponibles dans GenBank
Myoviridae Podoviridae Siphoviridae Non classés
K,G1, 44AHJD, phil 1, phil2, phil3,187,2638A, 37, 3A, P954,PT1028,
Twort 66, P68, 42E, 55,77, 80alpha, phiPV83, CNPH82, ROSA,
SAP-2 EW. PHI 5. PVL. phiETA. DhiETAZ phi2958PVL,
phiETA3, phiMRl 1, phiMR25, phiN315, phiPVL-CN125,
phiNM, phiNM3, phiPVL108, phiSLT, tp310-1, tp310-2,
phiSauS-IPLA35, phiSauS-IPLA88, 53,29, tp310-3
47, 52A, 69, 71, 85, 88,92, 96, X2
Des analyses bioinformatiques de ces génomes ont été réalisées afin de comparer et de
regrouper les différents phages ayant un pourcentage élevé d'identité nucléotidique. Ceci a

41
été effectué à l'aide des logiciels MEGA 4.1 (contenant ClustalW et Tree Explorer) en
utilisant la méthode du Neighbor-Joining (112) et à l'aide du logiciel DOTTER (107).
Le génome du nouveau phage LH1 a été comparé aux autres afin de le classer dans un
groupe de phages. De même, un autre phage de S. aureus a été isolé des eaux usées du
Québec par le médecin résident Dr Jeannot Dumaresq (données non publiées). Ce phage
nommé QW appartient à la famille des Podoviridae. Son génome a aussi été séquence et
comparé aux autres phages de S. aureus afin de le classer.
2.4. Objectif 4. Utilisation des phages pour éliminer la présence des S.
aureus dans différents types de laits
Suite à leur caractérisation et leur regroupement, les phages K et LH1 ont été choisis pour
une application alimentaire. Il a déjà été montré que l'activité du phage polyvalent K est
inhibée dans le lait cru (54, 84). Il sera donc intéressant de le tester dans le lait et confirmer
les résultats des articles précédents. Dans cet objectif, l'activité du phage K a été comparée
à celle du nouveau phage isolé à partir du lait cru. L'interaction phage-bactérie dans le lait
chauffé à ultra haute température (UHT) et le lait cru ainsi que l'inhibition de la croissance
bactérienne par les phages en fonction du temps ont été vérifiées.
2.4.1. Test de l'inhibition de S. aureus par des phages dans le lait UHT
L'élimination de S. aureus par l'ajout de phages a été réalisée dans du lait UHT et du lait
cru. La souche bactérienne utilisée dans cet objectif est la souche 01 obtenue du MAPAQ
et isolée d'un fromage au lait cru québécois. La croissance bactérienne a été testée sur une
période de dix heures à une température d'incubation de 37 °C. Les trois premiers tubes
sont les contrôles : contrôle lait contenant seulement du lait, contrôle lait contenant une
quantité précise du phage K et un contrôle lait contenant le nouveau phage LH1. Ensuite,
un quatrième tube contenait du lait et la culture bactérienne de S. aureus, et dans les
cinquième et sixième tubes, les deux phages ont été rajoutés à la solution bactérienne. Le
dénombrement des plages de lyse s'est fait sur du milieu TSB alors que le milieu Baird-
Parker a été employé pour l'énumération des colonies bactériennes.
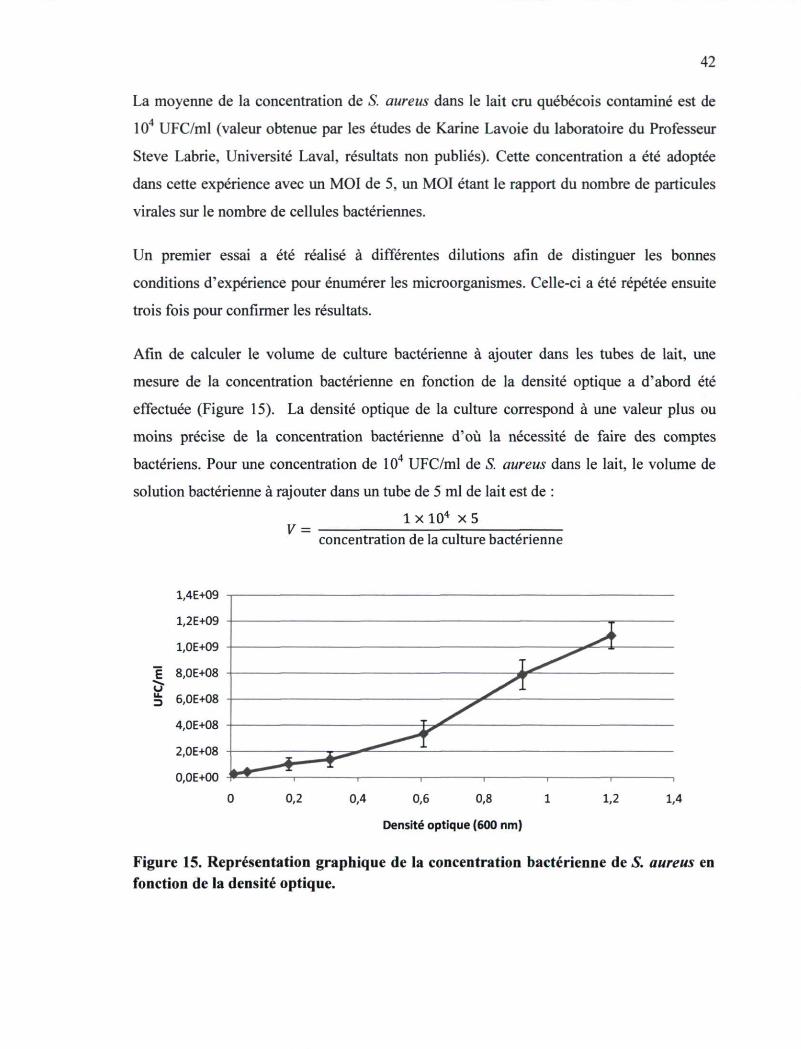
42
La moyenne de la concentration de S. aureus dans le lait cru québécois contaminé est de
104 UFC/ml (valeur obtenue par les études de Karine Lavoie du laboratoire du Professeur
Steve Labrie, Université Laval, résultats non publiés). Cette concentration a été adoptée
dans cette expérience avec un MOI de 5, un MOI étant le rapport du nombre de particules
virales sur le nombre de cellules bactériennes.
Un premier essai a été réalisé à différentes dilutions afin de distinguer les bonnes
conditions d'expérience pour énumérer les microorganismes. Celle-ci a été répétée ensuite
trois fois pour confirmer les résultats.
Afin de calculer le volume de culture bactérienne à ajouter dans les tubes de lait, une
mesure de la concentration bactérienne en fonction de la densité optique a d'abord été
effectuée (Figure 15). La densité optique de la culture correspond à une valeur plus ou
moins précise de la concentration bactérienne d'où la nécessité de faire des comptes
bactériens. Pour une concentration de IO4 UFC/ml de S. aureus dans le lait, le volume de
solution bactérienne à rajouter dans un tube de 5 ml de lait est de :
1 x IO4 x 5 V = concentration de la culture bactérienne
l,4E+09
1.2E+09
l,0E+09
Ê 8,0E+08
5 6,0E+08
4.0E+08
2,0E+08
0,0E+00
0,2 0,4 0,6 0,8
Densité optique (600 nm)
1,2 1,4
Figure 15. Représentation graphique de la concentration bactérienne de S. aureus en fonction de la densité optique.

43
2.4.2. Test de l'inhibition de S. aureus par des phages dans le lait cru
L'action des phages seuls (sans l'ajout de la culture bactérienne dans le lait) a été testée
dans différents types de lait. Le lait cru a été chauffé selon les conditions standards de
pasteurisation (63 °C pendant 30 minutes puis refroidi à 13 °C) (69). Un autre échantillon
de lait cru a été maintenu dans de l'eau bouillante (100 °C) pendant environ une minute
pour se rapprocher des conditions de production du lait UHT. Du lait UHT commercial
ainsi que du lait cru ont été utilisés comme contrôles. Le dénombrement des plages de lyse
des deux phages a été réalisé dans un milieu TSB. La présence de S. aureus dans les
différents types de laits a été suivie sur milieu Baird-Parker en tant que contrôle tout au
long de l'application alimentaire en fonction du temps.

44
3. Résultats
3.1. Objectif 1. Isolement de nouveaux phages de S. aureus à partir
d'échantillons de lait cru
Deux amplifications utilisant la souche BIM 01-QW ont été nécessaires afin d'obtenir des
plages de lyse dans les six différents échantillons de lait cru. L'isolement d'une plage de
lyse de chaque échantillon testé a été effectué. Ensuite, trois amplifications de celles-ci ont
été réalisées afin d'obtenir un titre de phage élevé permettant une extraction de l'ADN de
phages des six échantillons. Un nouveau phage a été isolé et a été nommé LH1. La
connaissance de sa morphologie et de sa séquence permettra de le classer dans un groupe
de phages et de le comparer aux autres phages de S. aureus pour lesquels le génome est
disponible.
3.2. Objectif 2. Caractérisation du nouveau phage isolé
3.2.1. Observation des phages en microscopie électronique
Observé sous microscope électronique, LH1 a une capside allongée d'une longueur de 83,6
nm ± 0,5 et d'une largeur de 44,3 nm ± 0,2. Le phage LH1 a une queue non contractile
d'une longueur de 286,1 nm ± 10, 7 et d'une épaisseur moyenne de 5,3 nm ± 0,6. Donc,
celui-ci appartient à la famille des Siphoviridae et est de morphotype B2 (Figure 16).

45
te
-f
■0 100 nm I I
Figure 16. Image en microscopie électronique du phage LH1. Échelle : 100 nm. Cette photographie a été prise par la caméra numérique fixée au microscope électronique de marque JEOL JEM-1230.
Peu de phages de S. aureus caractérisés possèdent ce morphotype B2. Il existe seulement
les phages 3A, 47 et 42e (64). L'analyse bioinformatique du génome du phage LH1 nous
permettra de déterminer s'il est vraiment similaire à ces phages.
3.2.2. Spectre lytique du phage LH1
Le phage LH1 a été testé contre 32 souches différentes de S. aureus disponibles au
laboratoire. Ceci a permis l'obtention du spectre lytique de LH1. Il s'est avéré que 7 des 32
souches testées sont sensibles à LH1 soient les souches 01, 09, SARM-2 (canadien),
SARM-7, BIMs 01-QW, 01-44AHJD et 01-P68. Ces mêmes souches sont également
lysées par le phage Msa qui appartient aussi à la famille des Siphoviridae. Cependant, en
plus de ceux-ci, le phage Msa est capable de lyser les souches A170, SARM-1, S ARM-10
et les BIMs 01-LH1 et A170-Pyophage. D'autre part, les phages apprenant à la famille
des Myoviridae sont capables de lyser 26 souches de S. aureus testées et de lyser
partiellement les six autres souches restantes. Ceci suggère donc que le spectre lytique du
nouveau phage LH1 est restreint par rapport à certains autres phages de S. aureus.

46
3.2.3. Génome du phage LH1
Le génome du phage LH1 a été séquence afin de le classer dans un groupe connu de
phages de S. aureus. De plus, il est important de s'assurer de l'absence de gènes de
virulence dans le génome de LH1 avant de l'utiliser dans une application alimentaire
comme agent de biocontrôle. Le génome a une taille de 46 050 paires de bases. La totalité
du génome du phage LH1 a été couverte 36 fois par séquençage. Il contient 58 cadres de
lecture ouverts ayant plus de 40 codons. Après l'assemblage du génome de LH1 avec le
logiciel Staden, et l'analyse des chromatogrammes obtenus, une chute de signal a été
observée aux deux extrémités du génome du phage LH1. Ceci indique que ce phage
possède des extrémités cohésives ou sites cos. L'alignement du génome du phage LH1
ainsi que celui de trois autres génomes de phages appartenant à la famille des Siphoviridae
et partageant un niveau d'identité génomique élevé avec le phage LH1 ont été obtenus, et
ce, à l'aide du logiciel BioEdit. L'édition graphique a été réalisée à l'aide du logiciel
Adobe Illustrator (Figure 17). La fonction putative de chacune des séquences en acides
aminés de ces derniers a été déterminée grâce au programme pBLAST disponible sur le
site de NCBI. Les codons de départ (ATG, TTG, CTG, ATA et GTG) ainsi que les codons
d'arrêt (TAG, TAA et TGA) ont été utilisés (64). Le RBS ou site de fixation du ribosome
constitue la séquence nucléotidique conservée Shine-Dalgarno (SD) (105). Il est situé entre
8 à 12 nucleotides en amont du codon de départ. Celle de S. aureus (5'-AGGAGG-3') a été
utilisée pour identifier les séquences de RBS dans les génomes de bacteriophages infectant
cette bactérie (64). Toutes ces données sont affichées dans le tableau 6 à la page suivante.

II lllll MINI II III •»* "ft C l <"*. n < * , o c T ^ O
1 «ft ^ . K •> «-ft «ft T> f i f - r -
o* — _ _
v . t N n r t f . ^ — oo -c © 3 - i - ^ i - g * — so — -ft CT* ? A r-, (O V . N C
. . "ft 0 \ 3s » ^ W ^ N C
O r - c n r*~, r * o © r - © - r
5:1 £ ? £ z I i £ >
I - . «r, r - l r - , 0 0 aj- N r - , O ( N l f t •a» — af* ar. aO ij-, a/* </-, r * -C CO «- -■Oi o r*i o to <-*-, m x o Q r - rt -*". O -—, O -3 !> . "J-. «a*-oo, o c4, <N O , ra se oc o o t s
pa. - ft. - 1 . p . - ' f t . ' - i . ' f t . ' s . ' f t ,1
-H SX 1
3 ■a
'mi
E o
B
WC
3 -a n
r CU > 3 e eu cu
_aj cu
T3 n
9 M CU
■o n cu 3 O"
-CU -w CU •JS
u
u sO 3 S CU
s H H
99 -
r-i i - . r t © sO vO <ft *ft «o rW v-tf m' ' H rsf
O r - ^ o v ô s o o ^ c c oo" a ' i o «r* V Tt T* a-'
<N CT «ft — VO oo oo I N O O r n ~+ oc * f
m «.r, i f t
^ 1 —, oo r-r «*> i c ï <**,' c ? «ft vc? r-* •■* un
S — — Ç —- — *Q en Ç - ^ - - « r ^
B v , n r i «3 t sq f^ — •*•-< s> ^ oo «in ift ■•* -rr vtf o-' 0s •--> Os* CT oT «ft
= 3 S g s - ^ - ^ - a l S F 3 S S I Ï R R ? 8 2 S 2 s sas a* rN ^ 0 * " - r a _ « r N — — " - g M " " , s* .C r-l — — ^ ' — — *♦ c-, en a-*- » . r
o
"? o es o ( N f l * f
. Oa
' ( S
II Ô a/*. Ô «N ar, *
O I - ar, an £ r - — <N <s P <—i OS r**ï a n L . I - t - » o o 2
M ir, ara — I-- o* 00 »r, © oc fn SO "r, -a* r - . t * * r n N M ( a l \ O r a ) m — *<r r-, - » H n i >© a n r - - r * - c j * ' 0 0 - ^ — i -—a — " - _ N N N ( N i
— - rN m Cal fN c s |
Cr. CL hn — O 0 O C f > — Cr. — f > ***a — f**a f N -—a f N ^—
f l 00 fN *af 00 mm w . o o — —a — r-i
—■ — a f N C J f N C - l f N C - l
a ^ a o oo r - a*3K -> rai i r , ■-.--*• OS fN ara — 00 oc Oa — o r - « ? » S S ^ £ ?, en r~ r , -o m — n rs
* ar, an >r, ad .c I-- r- 00 ce 2 — — — S Î S S f N I
l <--r r^i
S3
00 oc
r - O rm-. OC C— -O •ar 00 <r, i— r» o rN r - l fN
r f •<-. Oa •<-. —. CJ t ÇJ SS 8? è>3 — e-; 5 o i**-" -d ari* *-f oo |
os fN * * oc r» P - 0> OS fN fN a/ï ar, c*ï <L hn OS O —a — <N r-i r-i m r*-, m
n x o o o r i Oa O- — —■ —
O f-*" fa^ r*-, f n
— a r N t * 5 * - f * a - > -O t ~ O C O a O —a rN a*a-| -"T «*> N r s r i N N rN «s r s P i r - , en m en m en

X - 1 -
s Wl t i i ^ O s C T ^ C T ~ ! ^ ~ C T O l ~ ~ ! ~ « " * " i < ~ . . 0 ^
-ft f "*CT^r -©r*- . © — © C T r - >-*. © -t r- ■-*. vo n f-» x o\ r r-vorsir«-sCif tCTT*-rtf*>rj v O ^ . d C T v O C T V p ^ Q ^ s C Ô ^ r CT cl — O s r - i O o O O t O s — (S C r*"r, H -> O r j o. rj os ;. ( N O C N C N n M i N N c © ( N i n e ©, r^ «rs, o «N, o N, rsi û - W " Pu C L ' A . û-'Ot ^ W " ' cu c-'au ' a . ' pu '.a-'-pu 2- pu'û- pu ' a .
« N s C CT 3t rJ Os rn a A o -J A -J o N — - - ,S T •*
- 111 J, g r - o o x n c 2 * c « S S =
é>— - ^ ^ s ? s r o o — o o — 8 s *— O 0 0 — 2 — "-'-> W S O C — S rS oc en rn t PJ g OC Si S
j — . ,—, ^ ^ . . - ^
— O mO 00 3 - OO
- i f o o o » * , o r- © rN oo oc an ss -" ~ c; ~ -S * t ^ ^ J N * f oc © — 00 oc
~ — en —
S c* ^-SJ cr ">
<ilTfSS
*s ^ Z _
J <■ <3 fN r ^ _ o o _ • „
â
_] an
Sis •r, g -g
o.co. '**- ,** o-m era*» -r
U O U U U U U U H P H B H H H H g ^ ro y. où 3 ™
h p H H h P h < < < a < < <
Tt vo oc m CT *n —* o <s" m' *J - rf"*. 2 * oo ^ © C ^ O O I f t f N O ^ T t ^ i f t ^ - T t CT' VO T t Ift -ft, SO Tf' — SO' V Tt
oo os **■ m ■-<■ © sc © - oo - *1 ^ ï - —" *n K <N § 3; os 3: s§ c •< — Ç 5 ; r -
^ - O rs VO (N «sj T, - ^ so ^ o V *r, «ft rt 'rt so ift CT Vîf CT OS oo'
S S 8 8 8 - ï = a S " f S - r o o o c m ^ 2 * c f e - c 2 S — rorNSO* 0 — o o a o * D — ' T ' ' - r - - n " * ô o c > < * - l r ' - — 3
T r r * . o c m — © — *ftocooCT «ft © s C — OCOO — OS «ft © r--N C » — 0«v- , © r - - * o i f t C T
_j oo oo oo d s ^ O s o o o n n - t - r , m-ft i m n n en n c i n c i r N C i ^ < t n
w' * ' t
,t t « j t
C •« fN M O O — <N 0C •-• «ft ■r, i*. n vj <r, n rs ft i - r» sc
_.-ft. S 0 C T « T t « « T j - ^ C T © s C ( n ^ t l O r s - l N S C O O X 0 0 C T C T « C T © O © r v - ) O T t » f t i r ,
n c i r N C i ^ n n c i n c i n M r N f n < s - i - ^ ' t T f T r ' ^ t j * < t t r
s O t - * o o O s o — r s | f * , T t » f t s o I « n CJ TT •♦ r f -»t ^t •♦ t t t f A l f l '
\ «m Tf «m -o r - c > I f t «ft I f t iy-, I f t «-

LH1 46050 bp
3A 43095 bp
42e 45861 bp
47 44777 bp
49
/ S
/ / / J? s .£
sf f / f J ^
! 1 1 1 ! ! 1 1 1 TEp 1 5000 10000 1S0OO 20000 2S000 30000 35000 40000 4S000
■ 11 2 1 2 4 32 39 44 47 50 S3 SS 7 10 13 20 23 31 34 34 38 43 46 49 52 SS S7
I 2 3 4 S 6 9 12 14 15 16 17 18 19 22 25 26 27 2» 29 30 33 35 37 40 41 42 45 48 SI 54 56
Aiài i i^à JMftilU J i i TTTTIWT m m 1 "TT
36 17 41 130 S3 » 31 67 38 JO 47 SS 59 9 40 IS I I 4 6 l p J f * 1 22 13 '4 6 44 4 7 - j *35 8 21 7aJ
,6-$%3
7Sp4
333
U 19 3 T i - F s i ' j V 2 7 ? 137*
43 3 8 IS 172-0 2V41 I J I 14 4 52 2 S 36 87 HI '«1 12 11 2» I» 13 7 1 » 32 40 7 3 ^ 3 9 24, „23» 15S
7»%
9 » * • & * ] . ? 1 IS 6 43 4 45^7 J 4 71 .1 i t ^ n ' O W w 22 3 J t f M t f 7 l e 8 2S»
!#♦ HH» -I R Figure 17. Alignement du génome des phages LH1, 3A, 42e et 47. Chaque flèche
représente un cadre de lecture ouvert. Ceux partageant plus de 80 % d'identité en acides
aminés sont indiqués par la même couleur et sont reliés par des ombrages gris. Les
pourcentages d'identité inférieurs à 80% sont affichés sur la figure.
L'analyse bioinformatique du génome du nouveau phage LH1 montre qu'il se rapproche le
plus des phages 3A, 47, phiSLT, et phi2958PVL. Cependant, il n'est pas clairement montré
quel phage parmi ceux-ci représente le plus haut pourcentage d'identité génomique avec le
phage LH1. La comparaison des résultats avec d'autres logiciels bioinformatiques
confirmera cette identité.
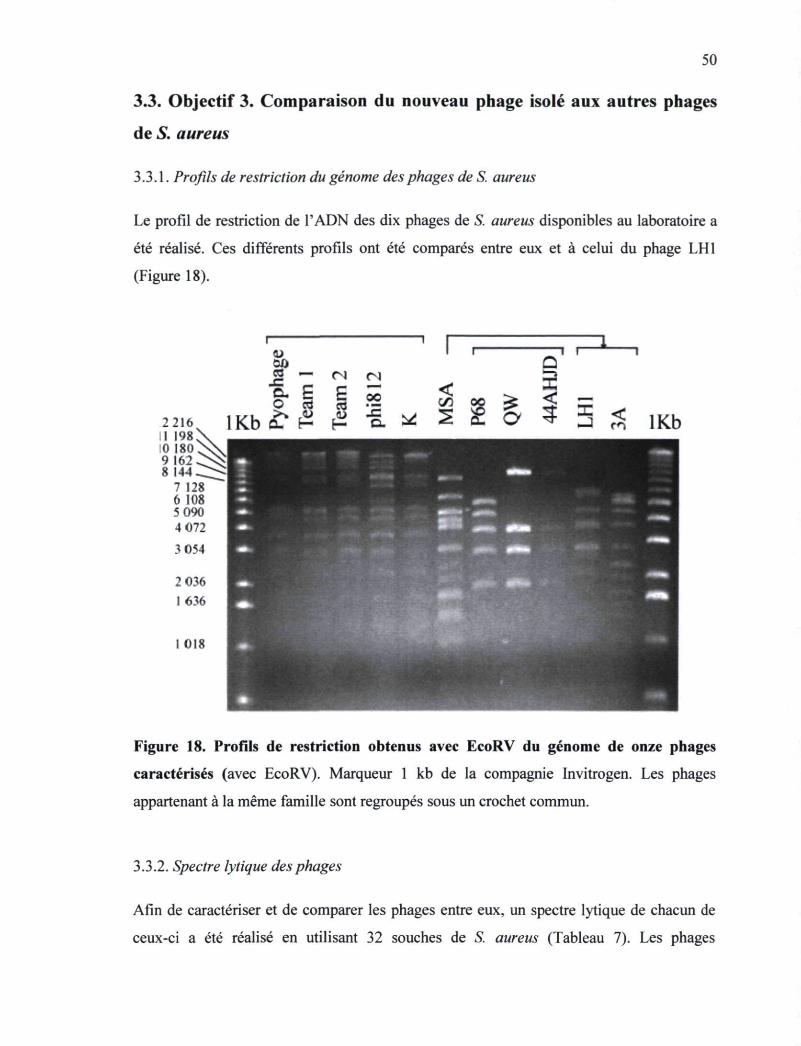
50
3.3. Objectif 3. Comparaison du nouveau phage isolé aux autres phages
de S. aureus
3.3.1. Profils de restriction du génome des phages de S. aureus
Le profil de restriction de l'ADN des dix phages de 5. aureus disponibles au laboratoire a
été réalisé. Ces différents profils ont été comparés entre eux et à celui du phage LH1
(Figure 18).
2216 Il 198 10 180 9 162 8 144
7 128 6 108 5 090 4 072
Figure 18. Profils de restriction obtenus avec EcoRV du génome de onze phages
caractérisés (avec EcoRV). Marqueur 1 kb de la compagnie Invitrogen. Les phages
appartenant à la même famille sont regroupés sous un crochet commun.
3.3.2. Spectre lytique des phages
Afin de caractériser et de comparer les phages entre eux, un spectre lytique de chacun de
ceux-ci a été réalisé en utilisant 32 souches de S. aureus (Tableau 7). Les phages

51
appartenant à la famille des Myoviridae ont le plus large spectre lytique. Seules deux
souches de S. aureus résistantes à la méthicilline, CMRSA-2 et CMRSA-6, sont
p.artiellement sensibles aux phages Pyophage, Team 1, Te.am 2 et phi812 et K
respectivement. Les isolats 01-K, A170-Team 2, 1049-Pyophage, 1049-Team 2 et 1049-
phi812 sont partiellement résistants à ces cinq phages. Jusqu'à présent, aucun isolât de S.
aureus n'est complètement résistant aux phages appartenant à la famille des Myoviridae.
Les phages P68, QW et 44AHJD appartenant à la famille des Podoviridae ont un
spectre lytique plus restreint que celui des phages appartenant à la famille des Myoviridae.
Ces trois phages ont un niveau d'identité génomique dépassant les 99%. Cela s'illustre
aussi p.ar leur capacité à lyser les différentes souches de S. aureus. Enfin, les phages
appartenant à la famille des Siphoviridae possèdent un spectre lytique restreint par rapport
aux deux autres familles. Les phages Msa, LH1 et 3A sont capables de lyser 13, 7 et 1 sur
32 souches de S. aureus testées respectivement. Il serait donc intéressant de choisir un
phage appartenant à la famille des Myoviridae dont le phage K pour comparer son activité
à celle du phage LH1 dans le lait.
Tableau 7. Spectre lytique des onze phages étudiés sur 32 souches de S. aureus
Collection Félix d'Hérelle MAPAC Hôpitaux canadiens Laboratoire
o - m
- m _ J X
en
O
ce
_ o r-.
<
ra co
J") OJ
rN
ce I
-D I N r-j
CC
X
co m IN
cc r en
$ ce
(N
Si m
Si ce
Si CC
LTV
rr:
'■r
Si CC
--
Si CC
00
Si ce
s cc
\> u
o --H
Si r r 5 o
O
o
5 a o
Q X <
o
ï < «T "•T
O
a. 00 B £ ai o
m o
I
o o
E a»
ô r-.
<
i a o >■
CL
n O
fN E f-} a.
H-
cr. «3-O
(N
00
e Cl o
S *o 'C ■s o
f
pyophage -
S *o 'C ■s o
f
t e a m l --S *o 'C ■s o
f team 2 -
S *o 'C ■s o
f phi812 -
S *o 'C ■s o
f K -
c, 8 'fa t l
£ 1
P68 c, 8
'fa t l
£ 1 QW
c, 8 'fa t l
£ 1 44AHJD
, h, o o -c *o S* s
<j"i S
M " , h, o o -c *o S* s
<j"i S
LH1 -1 II , h, o o -c *o S* s
<j"i S 3A
] Q partiellement sensible ] Πsensible
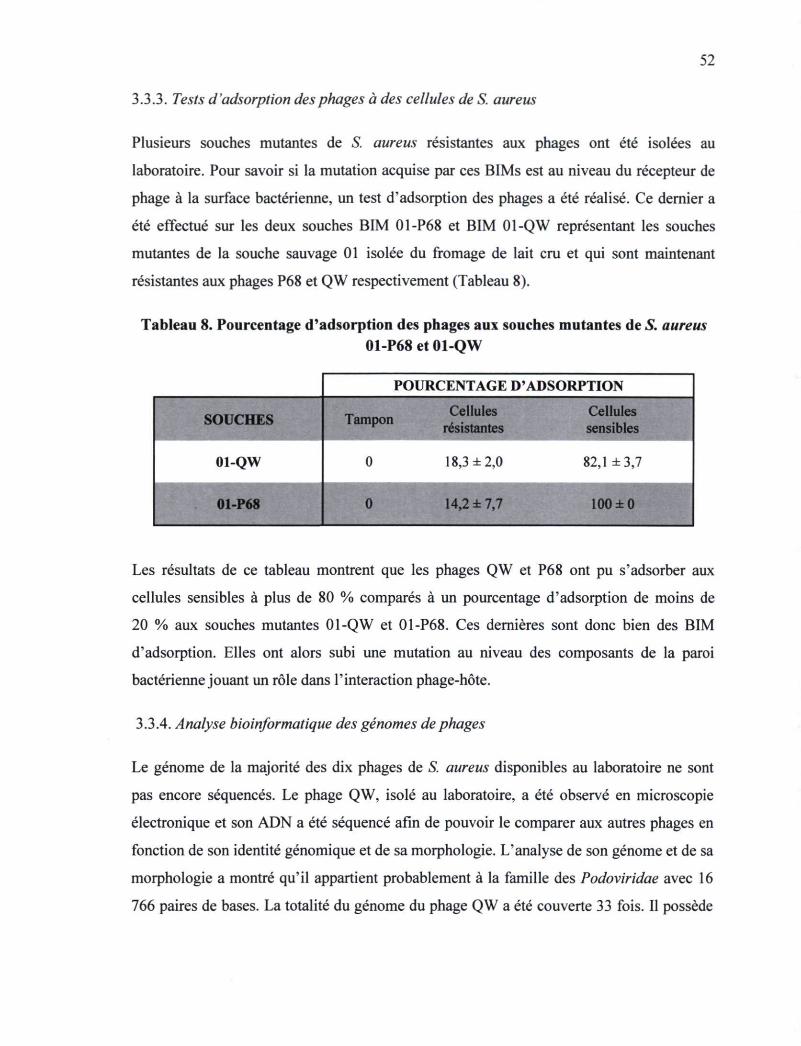
52
3.3.3. Tests d'adsorption des phages à des cellules de S. aureus
Plusieurs souches mutantes de S. aureus résistantes aux phages ont été isolées au
laboratoire. Pour savoir si la mutation acquise par ces BIMs est au niveau du récepteur de
phage à la surface bactérienne, un test d'adsorption des phages a été réalisé. Ce dernier a
été effectué sur les deux souches BIM 01-P68 et BIM 01-QW représentant les souches
mutantes de la souche sauvage 01 isolée du fromage de lait cru et qui sont maintenant
résistantes aux phages P68 et QW respectivement (Tableau 8).
Tableau 8. Pourcentage d'adsorption des phages aux souches mutantes de S. aureus 01-P68 et 01-QW
POURCENTAGE D'ADSORPTION
SOUCHES T Cellules Cellules résistantes sensibles
01-QW 0 18,3 ±2,0 82,1 ±3,7
01-P68 0 14,2 ±7,7 100 ± 0
Les résultats de ce tableau montrent que les phages QW et P68 ont pu s'adsorber aux
cellules sensibles à plus de 80 % comparés à un pourcentage d'adsorption de moins de
20 % aux souches mutantes 01-QW et 01-P68. Ces dernières sont donc bien des BIM
d'adsorption. Elles ont alors subi une mutation au niveau des composants de la paroi
bactérienne jouant un rôle dans l'interaction phage-hôte.
3.3.4. Analyse bioinformatique des génomes de phages
Le génome de la majorité des dix phages de S. aureus disponibles au laboratoire ne sont
pas encore séquences. Le phage QW, isolé au laboratoire, a été observé en microscopie
électronique et son ADN a été séquence afin de pouvoir le comparer aux autres phages en
fonction de son identité génomique et de sa morphologie. L'analyse de son génome et de sa
morphologie a montré qu'il appartient probablement à la famille des Podoviridae avec 16
766 paires de bases. La totalité du génome du phage QW a été couverte 33 fois. Il possède
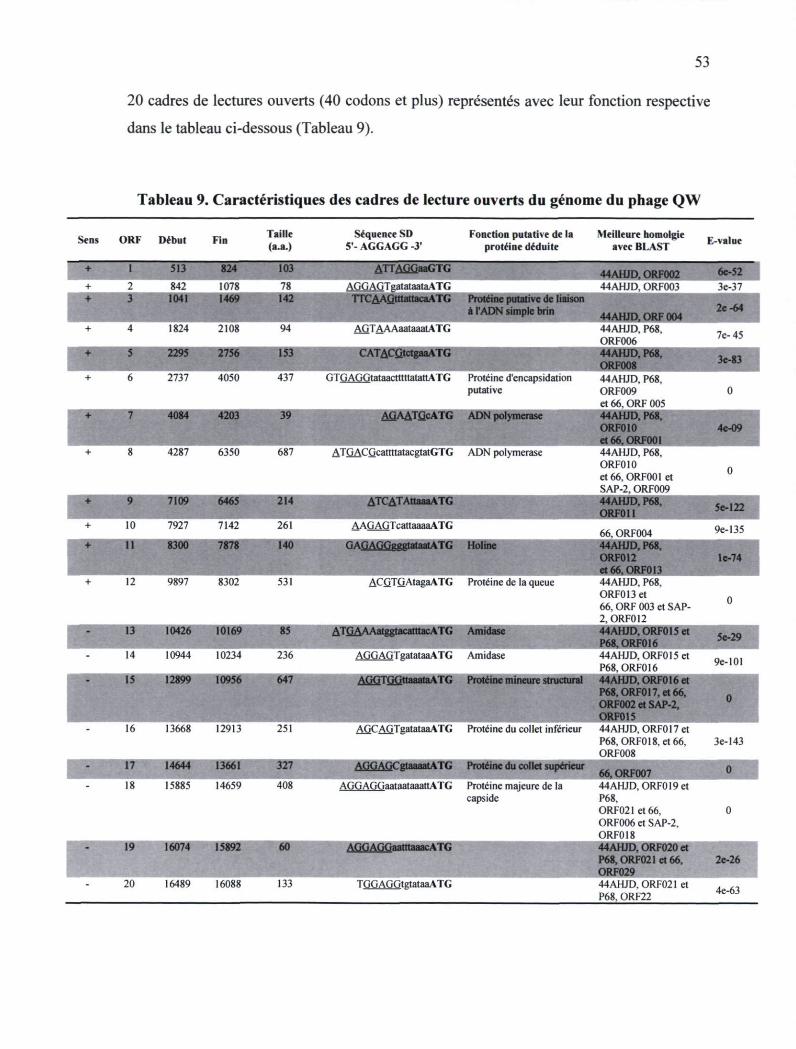
53
20 cadres de lectures ouverts (40 codons et plus) représentés avec leur fonction respective dans le tableau ci-dessous (Tableau 9).
Tableau 9. Caractéristiques des cadres de lecture ouverts du génome du phage QW
Sens ORF Début Fin
19
20
Taille (a.a.)
Séquence SD 5'- AGGAGG -3'
Fonction putative de la protéine déduite
Meilleure homolgie avec BLAST
513 842 1041
824
1078 1469
103
78 142
ATTAGGaaGTG AGGAGTgatataataATG
TTCAAfitttattacaATG
1824 2108 94
2295 2756 153
2737 4050 437
AGTAAAaataaatATG
:ATACfitctgaaATG
GTGAGGtataactttttatattATG
Protéine putative de liaison à l'ADN simple brin
Protéine d'encapsidation putative
4084
4287
4203
6350
ADN polymerase
687 ATGACGcattttatacgtatGTG ADN polymerase
7142
7878
261
140
AAGAGTcattaaaaATG
GAQAGiïgggtataatATG Holine
9897 8302 531
10426 10169 85
10944 10234 236 12899 10956 647
ACGTGAtagaATG Protéine de la queue
ATQAAAatggtacatttacATG Amidase
AGGAGTgatataaATG Amidase
AGGTGGttaaataATG Protéine mineure structural
13668 12913 251 AGCAGTeatataaATG Protéine du collet inférieur
14644 13661 327
15885 14659 408
16074 15892
16489 16088
60
133
AGGAGCgtaaaatATG Protéine
AGGAGGaataataaattATG Protéine majeure de la capside
AGGAGGaatttaaacATG
TGGAGGtgtataaATG
44AHJD, ORF002 44AHJD. ORF003
44AHJD.ORF004 44AHJD. P68. OR F006 44AHJD, Pi ORF008 •14AHJD, P68, ORF009 et 66, ORF 005 44AHJD, P68, ORF010 et66,ORF001 •MAHJD,P68, ORF010 et 66, ORF001 et SAP-2, ORF009 44AHJD, P68, ORF011
66, ORF004 44AHJD, P68, ORF012 et66,ORF013
E-value
6e-52 3e-37
2e -
7e-45
le-74
0
44AHJD, P68, ORF013 et 66, ORF 003 et SAP-2, ORF012 44AHJD,ORF015et P68.ORF016 44AHJDORF015et P68. ORF016 44AJUD,ORF016et P68,ORF017,et66, ORF002 et SAP-2, ORF015 44AHJD, ORF017et P68,ORF018,et66, 3e-143 ORF008
5e-;
9e-101
66, ORF007 44AHJD. ORF019et P68, ORF021et66, ORF006 et SAP-2, ORF018 44AHJD, ORF020 et P68,ORF021et66, ORF029 44AHJD,ORF021 et P68, ORF22
2e-26
4e-63
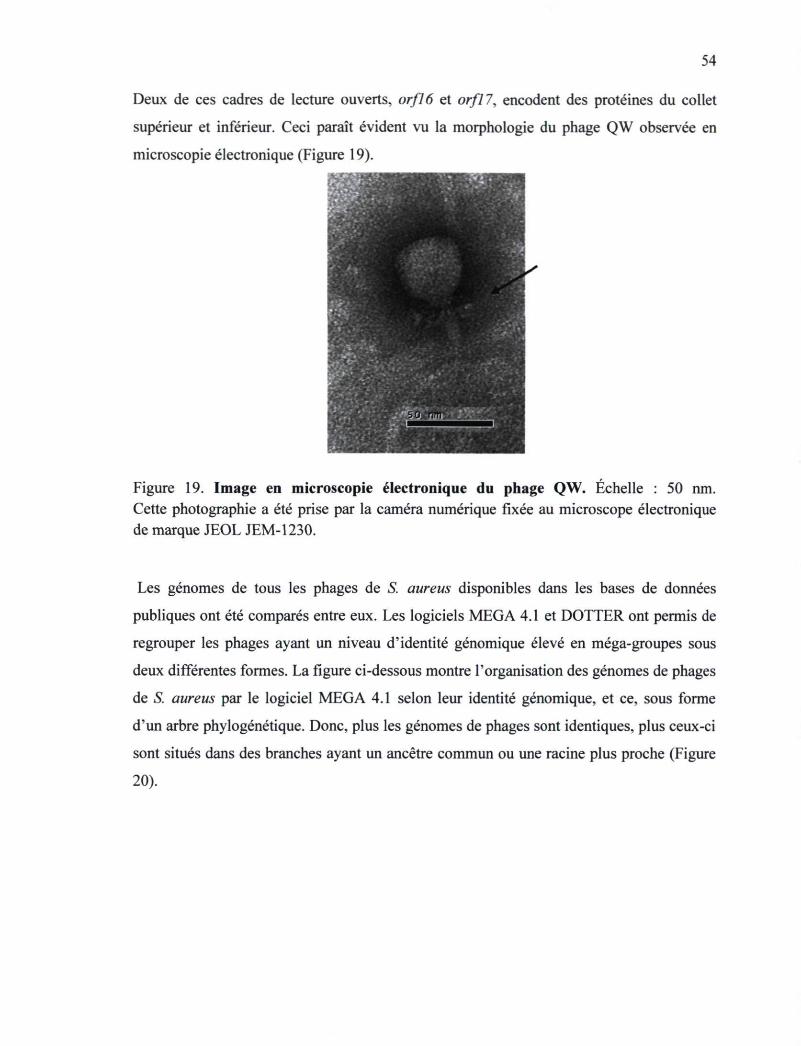
54
Deux de ces cadres de lecture ouverts, or/76 et orfl7, encodent des protéines du collet
supérieur et inférieur. Ceci paraît évident vu la morphologie du phage QW observée en
microscopie électronique (Figure 19).
Figure 19. Image en microscopie électronique du phage QW. Échelle : 50 nm. Cette photographie a été prise par la caméra numérique fixée au microscope électronique de marque JEOL JEM-1230.
Les génomes de tous les phages de S. aureus disponibles dans les bases de données
publiques ont été comparés entre eux. Les logiciels MEGA 4.1 et DOTTER ont permis de
regrouper les phages ayant un niveau d'identité génomique élevé en méga-groupes sous
deux différentes formes. La figure ci-dessous montre l'organisation des génomes de phages
de S. aureus par le logiciel MEGA 4.1 selon leur identité génomique, et ce, sous forme
d'un arbre phylogénétique. Donc, plus les génomes de phages sont identiques, plus ceux-ci
sont situés dans des branches ayant un ancêtre commun ou une racine plus proche (Figure
20).

Myoviridae 55 Podoviridae
Siphoviridae
Figure 20. Arbre phylogénétique comparant les séquences nucléotidiques des génomes de phages de S. aureus. Cet arbre est généré par le logiciel MEGA 4.1 en utilisant la méthode Neighbor-Joining. Échelle: 200 paires de nucleotides.
Une autre analyse de type « dot plot » a également permis de regrouper les phages selon
leur niveau d'identité génomique et a confirmé les résultats obtenus avec le premier
logiciel utilisé. La matrice obtenue rassemble tous les génomes de phages et permet
d'identifier les différences nucléotidiques entre deux phages. La comparaison des génomes
se fait en examinant la continuité de la droite de comparaison entre deux génomes qui
constitue l'élément primordial; plus la droite est continue et passe par la diagonale du
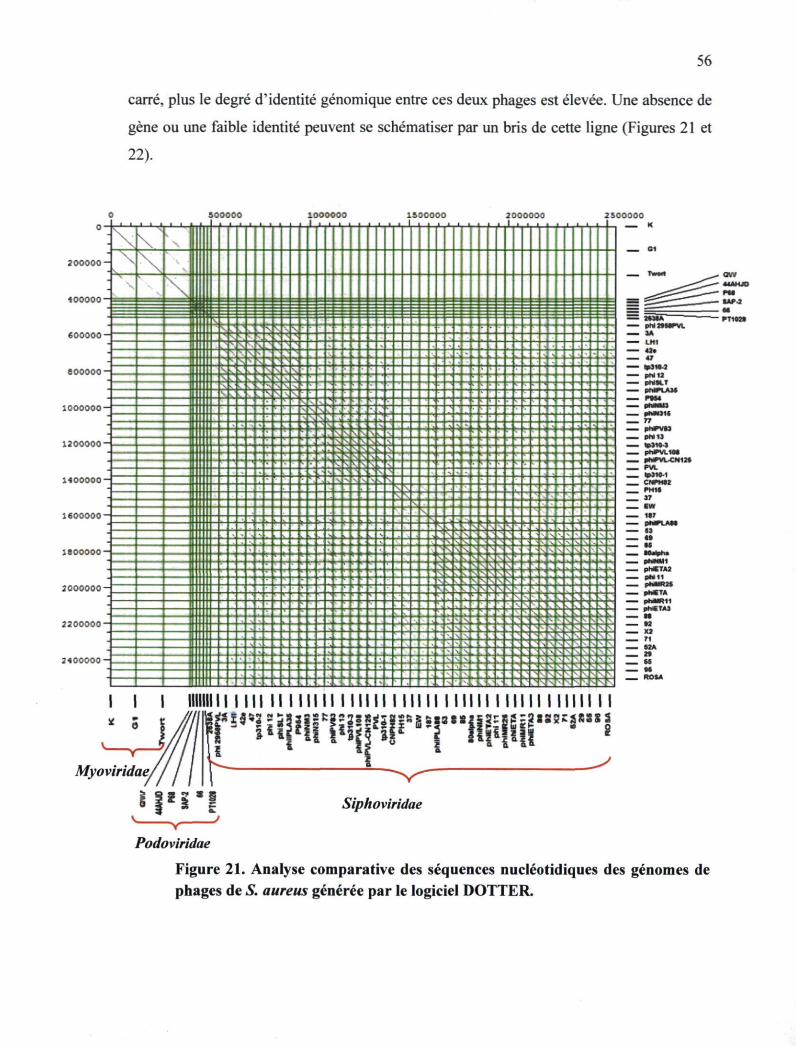
56
carré, plus le degré d'identité génomique entre ces deux phages est élevée. Une absence de
gène ou une faible identité peuvent se schématiser par un bris de cette ligne (Figures 21 et
22).
aOOOMM 1 . . . a . i ■ t 1 . . a
\ 2 0 0 0 0 0 -
.... : — a-. — — ,
; — l l l l l l l l l l l l ' ^\ s «s ■
- - - ' € 0 0 0 0 0 — l l l l l l l l l l l l
■
1 . 1 a '■ v l l l l l l l l l l l l . «, * ■ - .
. • J -■ . a -. . l l l l l l l l l l l l
~ —■ "» a
N >a| -_ . A _ - »
-. ta » » — i -J ^ 4 _ -.. !.. . J3L _J J i . à - - J J
8 0 0 0 0 0 — l l l l l l l - - " ^ -, N ■ - ï - î - r ■
l ima - .i>| - T ~ ^ r r z - r - - r -r 1 N P P p l - — — — r- - -, ■ " ^ ■ ^ ^
- - - - r — , ^ * ' v i l S' V, ■ .
1 0 0 0 0 0 0 ^ - - : - - * ; j - i | | - - - ^ . . Z \ X " " . . . . - J .
■ ' H
- r i - iss^i - - » ■ ■ ■ » P > 4 > * H * - ( - ! - • X « i ^ . 1 4 4 - 4 4
m ■ 1 , . ? N "*■• ' >Ë3o f
Mil»! . P r , J P « J J { S s s S s ; - ■ ■ - . * ■
. P r , J P « J J { S s s S s ; - ■ ■ - . * - r
1 r> "a -J t ; -
" . ^ i ] ] . ., -1400000 — f Î ^ I ï ï s s i s i - -ZZ s : :
s -tZZZ.
*\A -^ - * - U- -*- « . ^ >_ _; - f_ > - a- ^ _ p _ i ■ ^ S - 5 - 5 -
■al- M . L k .i b- L. » , ta m i* — ta f . -i m. .1 _ _ i l - i - i - i
k^ J i . -
~ - — c- T S ■ «- r ««" t - r r - — -r * c- r r
■ r r » - " -- r- If - — < :-t^i? iHF<* l
h-4». a*-t. -1 h, h * h,
2 0 0 0 0 0 0 - i ■ i ^ ' r ""T
-1"
T r *«■ r* r r -*-*---- J: - a — a - a - - - -: > ^ L ■j ; | ' •
' J^ ' - J ' : ■ s T -
, *- " *" T î ? * J a m . h l h l h -~^\*^ 5*.!!. S. SSSS ■ j ^ : 1 S » à. Ha. V k C C « SÏSS
. ï à _ _ . . . 12 2 2 » a» ,
i ï S i " • V i 5» SL> . i . j SiSSs
2 4 0 0 0 0 0 - * i _ k. i L h l h « > - • * ■ * ! -
.jit j l à . . ! , . . . .__-__ I ■ v S T * ^ ! s s S S
—--. - . j ^ ï - i . • La a i » _ ! . _>- S hi J<l \ J x ™ \ ' - r r p r
2 5 C 0 D D 0
l l l l MM II l l l l l l l l l l l l l l l l l l MINI l l l l l l MINI I I
Myoviridae L
[fil! I t C h
Siphoviridae
Podoviridae
53
PMETA2 - p 4 - . 1 1
plaaUR25 p * > * TA ptaaURU phlETAJ tt
- «a X2 71 S2A »
_ SS
Figure 21. Analyse comparative des séquences nucléotidiques des génomes de phages de S. aureus générée par le logiciel DOTTER.
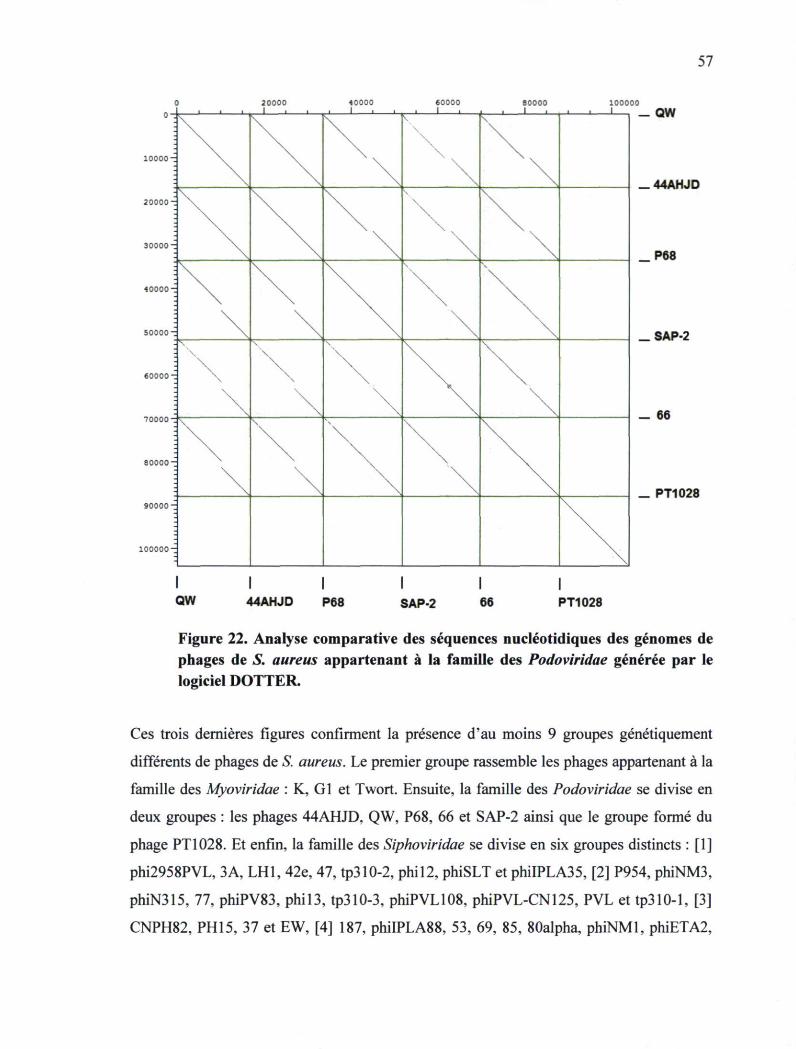
57
' 20000 1 1
e:::: l
80000 10001
\ : \ \ \ \
10000 **: \ x \
- -\ \ ^ 20000 -\
30000-i
40000-: \ \
50000-i \ \ : \ \ \ \
«0000-; \ \ \ \
E \ 70000 — \
\ 80000 *.
90000 *.
ooooo -j
QW I 44AHJD P68 SAP-2 66
_ QW
_ 44AHJD
_ P 6 8
_ SAP-2
_ 66
_ PT1028
I PT1028
Figure 22. Analyse comparative des séquences nucléotidiques des génomes de phages de S. aureus appartenant à la famille des Podoviridae générée par le logiciel DOTTER.
Ces trois dernières figures confirment la présence d'au moins 9 groupes génétiquement
différents de phages de S. aureus. Le premier groupe rassemble les phages appartenant à la
famille des Myoviridae : K, GI et Twort. Ensuite, la famille des Podoviridae se divise en
deux groupes : les phages 44AHJD, QW, P68, 66 et SAP-2 ainsi que le groupe formé du
phage PT1028. Et enfin, la famille des Siphoviridae se divise en six groupes distincts : [1]
phi2958PVL, 3A, LH1, 42e, 47, tp310-2, phi 12, phiSLT et phiIPLA35, [2] P954, phiNM3,
phiN315, 77, phiPV83, phil3, tp310-3, phiPVL108, phiPVL-CN125, PVL et tp310-l, [3]
CNPH82, PH15, 37 et EW, [4] 187, phiIPLA88, 53, 69, 85, 80alpha, phiNMl, phiETA2,
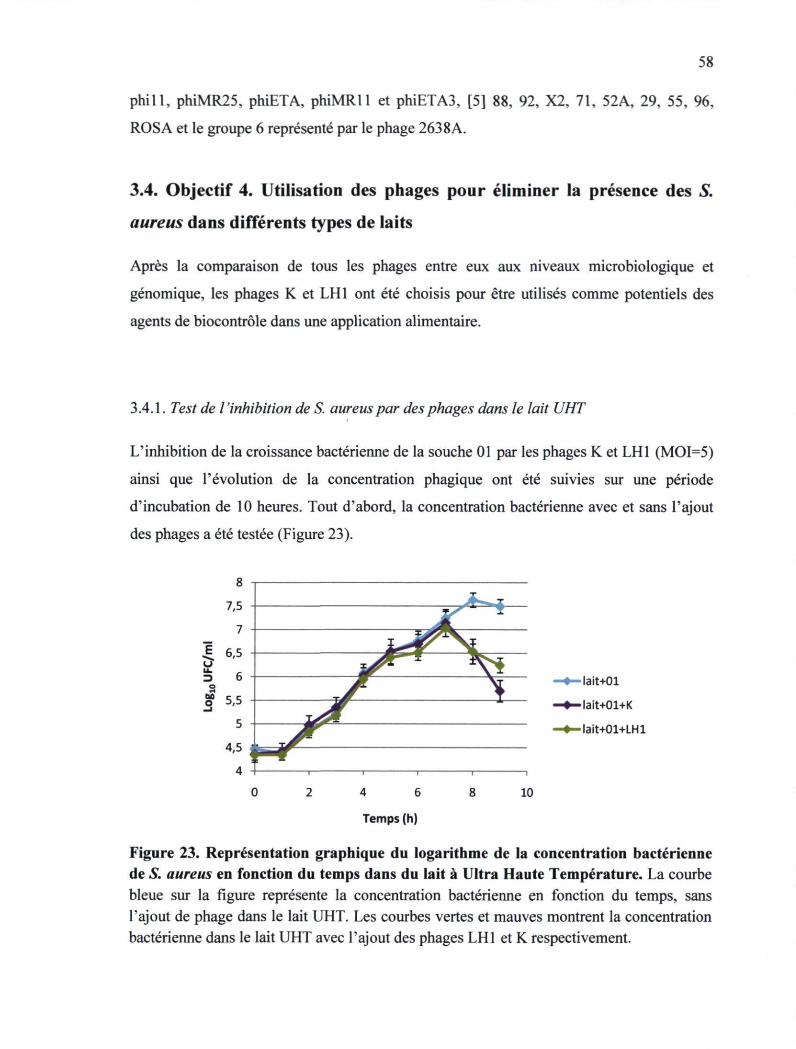
58
phill, phiMR25, phiETA, phiMRll et phiETA3, [5] 88, 92, X2, 71, 52A, 29, 55, 96,
ROSA et le groupe 6 représenté par le phage 2638A.
3.4. Objectif 4. Utilisation des phages pour éliminer la présence des S.
aureus dans différents types de laits
Après la comparaison de tous les phages entre eux aux niveaux microbiologique et
génomique, les phages K et LH1 ont été choisis pour être utilisés comme potentiels des
agents de biocontrôle dans une application alimentaire.
3.4.1. Test de l'inhibition de S. aureus par des phages dans le lait UHT
L'inhibition de la croissance bactérienne de la souche 01 par les phages K et LH1 (MOI=5)
ainsi que l'évolution de la concentration phagique ont été suivies sur une période
d'incubation de 10 heures. Tout d'abord, la concentration bactérienne avec et sans l'ajout
des phages a été testée (Figure 23).
u =J
o
i ■lait+01
■lait+01+K
■lait+01+LHl
4 6
Temps (h)
10
Figure 23. Représentation graphique du logarithme de la concentration bactérienne de S. aureus en fonction du temps dans du lait à Ultra Haute Température. La courbe bleue sur la figure représente la concentration bactérienne en fonction du temps, sans l'ajout de phage dans le lait UHT. Les courbes vertes et mauves montrent la concentration bactérienne dans le lait UHT avec l'ajout des phages LH1 et K respectivement.
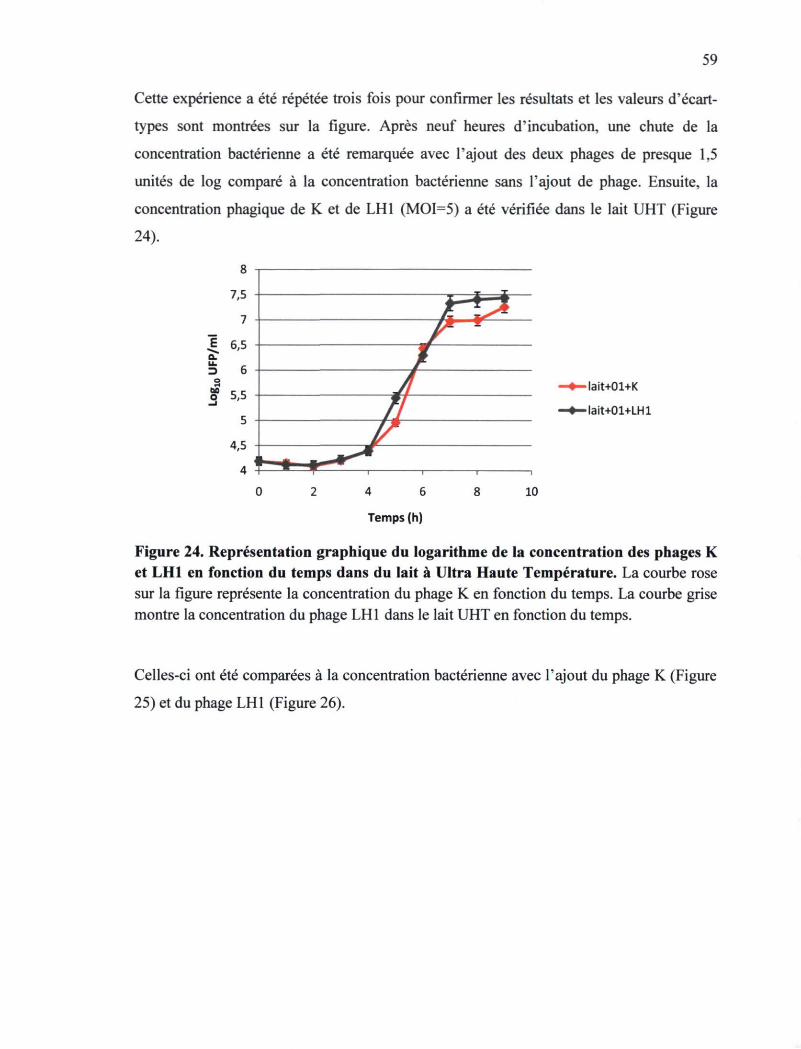
59
Cette expérience a été répétée trois fois pour confirmer les résultats et les valeurs d'écart-
types sont montrées sur la figure. Après neuf heures d'incubation, une chute de la
concentration bactérienne a été remarquée avec l'ajout des deux phages de presque 1,5
unités de log comparé à la concentration bactérienne sans l'ajout de phage. Ensuite, la
concentration phagique de K et de LH1 (MOI=5) a été vérifiée dans le lait UHT (Figure
24).
E LL. D
S? ■lait+01+K
•lait+01+LHl
Figure 24. Représentation graphique du logarithme de la concentration des phages K et LH1 en fonction du temps dans du lait à Ultra Haute Température. La courbe rose sur la figure représente la concentration du phage K en fonction du temps. La courbe grise montre la concentration du phage LH1 dans le lait UHT en fonction du temps.
Celles-ci ont été comparées à la concentration bactérienne avec l'ajout du phage K (Figure
25) et du phage LH1 (Figure 26).
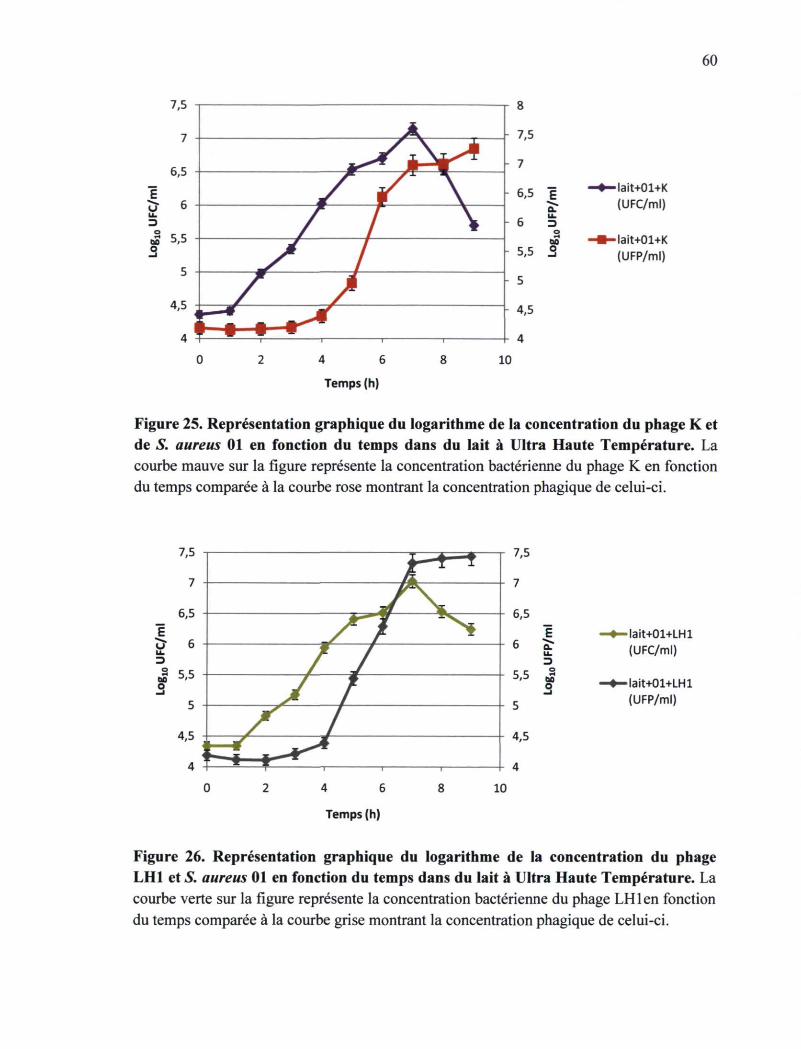
60
4 6
Temps (h)
10
LL.
ao O
•lait+01+K (UFC/ml)
■lait+01+K (UFP/ml)
Figure 25. Représentation graphique du logarithme de la concentration du phage K et de S. aureus 01 en fonction du temps dans du lait à Ultra Haute Température. La courbe mauve sur la figure représente la concentration bactérienne du phage K en fonction du temps comparée à la courbe rose montrant la concentration phagique de celui-ci.
7,5
7
6,5
6
5,5
I o" LL. Z)
o m,
M q
—♦—lait+01+LHl (UFC/ml)
•^^ la i t+01+LHl
5 (UFP/ml)
4,5
4
Temps (h)
Figure 26. Représentation graphique du logarithme de la concentration du phage LH1 et S. aureus 01 en fonction du temps dans du lait à Ultra Haute Température. La courbe verte sur la figure représente la concentration bactérienne du phage LHlen fonction du temps comparée à la courbe grise montrant la concentration phagique de celui-ci.
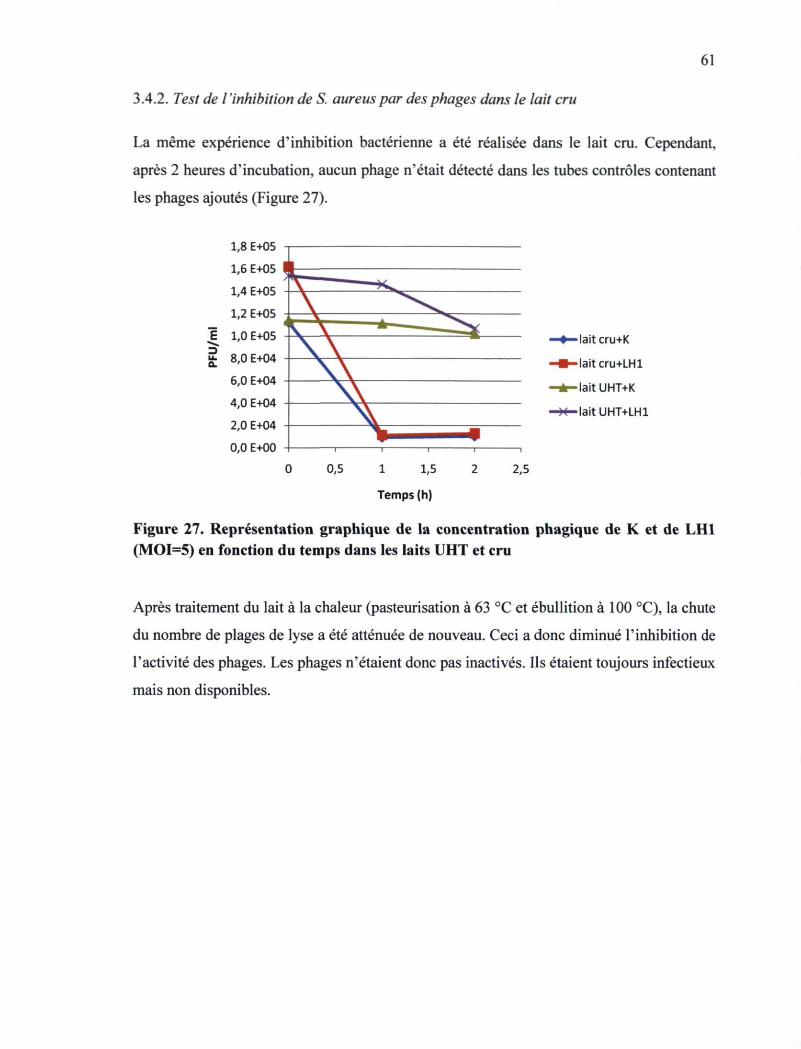
61
3.4.2. Test de l'inhibition de S. aureus par des phages dans le lait cru
La même expérience d'inhibition bactérienne a été réalisée dans le lait cru. Cependant,
après 2 heures d'incubation, aucun phage n'était détecté dans les tubes contrôles contenant
les phages ajoutés (Figure 27).
1,8 E+05 1,6 E+05 1,4 E+05 1,2 E+05
E 1,0 E+05 _ > LL. a. 8,0 E+04
6,0 E+04 4,0 E+04 2,0 E+04 0,0 E+00
•lait cru+K
■laitcru+LHl
■laitUHT+K
•laitUHT+LHl
Figure 27. Représentation graphique de la concentration phagique de K et de LH1 (MOI=5) en fonction du temps dans les laits UHT et cru
Après traitement du lait à la chaleur (pasteurisation à 63 °C et ebullition à 100 °C), la chute
du nombre de plages de lyse a été atténuée de nouveau. Ceci a donc diminué l'inhibition de
l'activité des phages. Les phages n'étaient donc pas inactivés. Ils étaient toujours infectieux
mais non disponibles.

62
4. Discussion
Il est reconnu que les phages évoluent rapidement avec leur souche hôte et leur
environnement (27). Ainsi, l'activité des phages isolés d'un produit laitier par exemple
devrait être plus efficace que celle de phages isolés d'un environnement différent contre
des souches pathogènes isolées de produits laitiers. Une fois qu'un phage de S. aureus a été
isolé du lait cru, il a été caractérisé et comparé à d'autres phages de staphylocoques et
enfin, son activité a été testée dans du lait UHT ainsi que du lait cru contre une souche de
S. aureus isolée d'un fromage québécois de lait cru. Les différents résultats obtenus pour
chaque objectif ont été analysés et rassemblés afin de bien répondre à l'hypothèse de
travail exposée.
4.1. Caractérisation du phage LH1
Le phage LH1 a été isolé d'un échantillon de lait cru. Le protocole utilisé est donc efficace
pour l'isolement de nouveaux phages de staphylocoques. Grâce à la caractérisation de ce
phage, il s'est avéré que LH1 est un membre de la famille des Siphoviridae de morphotype
B2 puisqu'il possède un génome d'ADN double brin, une capside allongée et une longue
queue non contractile. Ce phage a un spectre lytique relativement restreint pouvant lyser 7
sur 32 souches de S. aureus testées. Étonnement, le phage LH1 peut lyser partiellement
quelques souches de SARM (SARM-2 et SARM-7). Il pourrait être ajouté ultérieurement,
dans un autre projet, à un mélange de phages destinés à la thérapie phagique. L'analyse de
son génome a permis de mettre en évidence plusieurs points. Tout d'abord, le génome de
LH1 a des extrémités cohésives vu qu'une chute de signal a été observée après
l'assemblage du génome. Ensuite, il possède les deux modules lysogénique et lytique. La
présence de cadres de lecture codant pour une holine et un anti-répresseur de même que
pour une intégrase, un répresseur et le régulateur transcriptionnel rinA montre que le phage
LH1 était probablement un prophage et est devenu virulent suite à une induction et une ou
des mutations. Le régulateur transcriptionnel rinA a été révélé comme nécessaire dans
l'activation de l'expression du gène de l'intégrase du phage (pli de S. aureus (121).

63
Ensuite, TORF053 encode la protéine E associée à la virulence. Cette dernière a déjà été
décrite dans le processus de replication de l'ADN du phage phiIPLA35 appartenant à la
famille des Siphoviridae et semble jouer un rôle particulier dans le métabolisme de l'ADN
(110). Cependant, ceci n'efface pas le fait qu'il pourrait être impliqué dans la pathogénicité
de l'hôte bactérien (53). Et enfin, deux gènes du phage LH1 encodent pour une toxine, la
leucocidine de Panton-Valentine. Celle-ci est une cytotoxine produite et sécrétée par des
souches de S. aureus résistantes à la méthicilline associées à la communauté. Elle est
capable de détruire les cellules du système immunitaire, tels que les neutrophiles
polymorphonucléaires, en formant des pores dans celles-ci ainsi que de causer une
pneumonie nécrosante létale. Cette leucocidine est codée par deux gènes co-transcrits lukS-
PV et lukF-PV habituellement localisés sur des prophages. La forme active de la
leucocidine de Panton-Valentine requiert l'assemblage de deux polypeptides, LukS-PV et
LukF-PV (65). Ces derniers sont codés par les deux ORF 26 et ORF 27 respectivement
chez le phage LH1. Ce phage ne pourra donc pas être utilisé comme un agent de
biocontrôle en dehors de ce projet de maîtrise. Par contre, il serait probablement possible
d'enlever ces deux gènes par génie génétique, mais il n'est p.as clair comment les autorités
réglementaires seraient réceptives à l'utilisation de phages génétiquement modifiés.
4.2. Comparaison des phages de S. aureus au phage LH1
Différents phages de S. aureus disponibles ont été comparés entre eux afin de bien choisir
les phages à utiliser dans une application alimentaire. Le profil de restriction de dix phages
de S. aureus disponibles au laboratoire ainsi que celui du phage LH1 ont été réalisés.
L'analyse des résultats a montré qu'il existe une grande similarité de profils entre les
phages Pyophage, Team 1 et Te.am 2, cp812 et K ainsi qu'entre les phages P68 et QW. En
effet, Pyophage, Team 1, Te.am 2, cp812 et K infectent la même souche bactérienne de S.
aureus SA 812, alors que les phages P68 et QW lysent la souche S. aureus 68. Cependant,
les profils des phages QW et 44AHJD semblent être identiques. De plus, l'analyse du
génome du phage QW montre une grande similarité entre les cadres de lecture ouverts de
QW et 44AHJD. D'autre part, le profil de restriction de LH1 est similaire à celui du phage

64
3A. Effectivement, ce dernier est aussi un Siphoviridae de morphotype B2, ce qui justifie
ce rapprochement.
Ensuite, le spectre lytique de chacun de ces onze phages a été obtenu en utilisant 32
souches de S. aureus. En comparant ceux-ci, il est clair que les phages appartenant à la
famille des Myoviridae ont un spectre lytique plus large que les autres phages. Cette
observation a également été notée par d'autres chercheurs (87, 100). Aucune souche n'a
résisté complètement à ces phages. Même des souches mutantes résistantes aux autres
phages ont été testées dont 01-P68 et 01-QW. Ces deux souches mutantes ont été
confirmées comme étant des BIMs d'adsorption puisque le phage s'adsorbait à des taux de
18,35% et 14,21% respectivement. L'isolement d'un phage pouvant infecter ces BIMs
d'adsorption serait souhaitable dans les applications alimentaires et la thérapie par phages.
Une comparaison de tous les génomes des phages de S. aureus disponibles dans les bases
de données publiques a été réalisée avec les logiciels MEGA 4.1 et DOTTER. En
regroupant les résultats de ces deux logiciels, les différents phages peuvent être divisés en
au moins neuf groupes génétiquement distincts dont un groupe pour la famille des
Myoviridae et deux pour la famille des Podoviridae et six groupes de phages appartenant à
la famille des Siphoviridae (Figure 28).

65
•
Myoviridae
rPodoviridae
I I I l l l l l l l II l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l II
i , l j ,H il?y a!4 j Hpfiiî 8
Siphoviridae
Figure 28. Identification d'au moins neufs groupes génétiquement différents de
phages de S. aureus
De plus, d'après cette analyse bioinformatique, le phage LH1 semble se rapprocher le plus
des phages de type 3A. Ceci a aussi été démontré par la similarité du profil de restriction de
l'ADN de ces deux phages. L'analyse du génome du phage LH1 a bien montré un haut
niveau d'identité génomique entre le phage LH1 et 3A ainsi que les phages 47, phiSLT,
phi2958PVLetphil2.
D'autre part, le phage QW dont le génome a aussi été séquence au cours de ce projet, s'est
rapproché le plus du phage 44AHJD. Cependant, en comparant le spectre lytique de ces

66
deux phages et en analysant le génome de QW, une faible distinction est observée entre ces
deux phages permettant ainsi de les différencier.
Cette analyse bioinformatique a permis de trouver une éventuelle corrélation entre les
groupes de phages, leur identité morphologique et génomique. Ainsi, les phages différents
au niveau de leur génome et de leur spectre lytique pourraient être choisis pour la
production de cocktails de phages afin de cibler le plus grand nombre de souches de S.
aureus possibles.
4.3. Activité du phage LH1 dans différents types de lait contaminés par S.
aureus
L'activité du phage LH1 a d'abord été comparée à celle du phage K dans le lait UHT. Le
suivi des concentrations bactérienne et phagique dans le temps a montré qu'avec l'ajout de
LH1 et K (MOI=5), la concentration bactérienne a diminué de plus de 1,5 unités de log
après neuf heures d'incubation à 37 °C. Cette diminution a été compensée par une
augmentation d'environ 1,7 unités de log de la concentration des phages LH1 et K après le
même intervalle de temps. Ceci montre donc que les phages sont bien à l'origine de cette
inhibition puisqu'ils se sont multipliés.
D'un autre coté, en comparant l'activité et l'efficacité des phages LH1 et K sur la
croissance bactérienne, celles-ci sont quasiment similaires. Cependant, le phage K semble
inhiber légèrement davantage la croissance bactérienne. En effet, les résultats montrent une
inhibition d'environ 1,8 unités de log avec l'ajout du phage K avec une MOI de 5 comparé
à une diminution d'à peu près 1,3 unités de log avec l'ajout du phage LH1. On peut donc
déduire que le phage LH1 isolé du lait cru n'est pas plus efficace que le phage K dans
l'élimination de S. aureus d.ans le lait UHT.
D'autre part, il a été démontré que les deux phages ne peuvent infecter la souche 01 dans le
lait cru. En effet, l'activité des phages a disparu après une heure d'incubation à 37 °C.
Cependant, après un traitement du lait cru à la chaleur, le nombre de phages a augmenté de

67
nouveau. Ceci indique qu'un composé inconnu thermosensible du lait cru bloque l'action
des phages. De plus, la présence naturelle de S. aureus dans le lait cru a été suivie. Sa
concentration approximative de 150 UFC/ml est trop faible pour être la cause de
l'inactivation des phages K et LH1 (5 x 104 UFP/ml). Donc, le composé qui est à l'origine
de cette inactivation provient du lait lui-même et est thermosensible. Il pourrait s'agir
d'une protéine du lactosérum (54) ou bien d'une protéine du caillé qui vient se complexer
au phage (41). Les particules de gras pourraient de même jouer un rôle dans cette
inactivation. Cette même expérience devrait être réalisée dans le lactosérum obtenu après
l'ajout d'un volume précis d'acide pour précipiter les caséines du lait caillé et d'un volume
de base afin de rétablir le pH de la solution à sa valeur d'origine. Ceci va permettre de
savoir si ce composé provient précisément du lactosérum ou du lait caillé (milieu acide) et
donc d'avoir une idée de l'effet du pH sur l'activité des phages.

68
5. Conclusions et perspectives Ce projet a fait appel à des recherches fondamentales ainsi qu'appliquées. Des phages de S.
aureus ont été isolés, caractérisés et testés comme d'éventuels agents de biocontrôle dans
le lait. Les différents phages de S. aureus ont été regroupés selon leur niveau d'identité
génomique. Ceci a aidé dans le choix des phages à utiliser dans l'application alimentaire. Il
est important de choisir des phages ayant un spectre lytique et un génome différent pour la
préparation des cocktails de phages afin de cibler le plus grand nombre de souches de S.
aureus différentes pouvant se retrouver dans le lait et le contaminer. De plus, cela peut
prévenir le développement de souches résistantes aux phages.
Le génome du nouveau phage LH1 a été séquence afin de s'assurer de son innocuité avant
son utilisation dans l'application alimentaire. Malheureusement, dans le cadre de ce projet,
le séquençage du génome du phage LH1 a été effectué vers la fin des travaux. Ainsi, nous
avons réalisé tard que ce phage portait des gènes codant pour des toxines. Donc, il serait
préférable de rapidement déterminer la séquence génomique des nouveaux phages isolés
ou aussi de vérifier la présence de gènes de toxines par PCR avant de passer aux travaux
appliqués. Ceci étant dit, nous avons quand même poursuivi les travaux puisque le phage
LH1 nous permettait de vérifier l'hypothèse à savoir si les phages de S. aureus isolés du
lait cru sont plus efficaces que ceux isolés de d'autres environnements.
Le phage LH1 ainsi que le phage K déjà disponible, ont été testés dans le lait UHT et le lait
cru pour vérifier leur efficacité à éliminer la présence de cellules de S. aureus. Cette
dernière expérience a montré que le lait cru contiendrait un composé thermosensible qui
inhiberait l'action des phages sur S. aureus. Ceci a aussi été rapporté dans la littérature (54,
84). Il est quand même intéressant de constater que le lait cru contient des phages de S.
aureus mais que ceux-ci ne sont pas efficaces. Il serait souhaitable d'identifier avec
certitude le composé inhibiteur pour connaître s'il serait possible de l'inactiver facilement
dans le lait cru et ainsi relâcher ce potentiel anti-staph. De plus, il est amplement connu que
les phages des bactéries lactiques se multiplient rapidement dans le lait (79). Une autre
question à résoudre serait de comprendre pourquoi les phages de staphylocoques sont
inactivés dans le lait cru alors que les phages des bactéries lactiques ne le sont pas. Ceci
nous pousse également à nous demander si le mélange K-LH1 serait plus efficace. Un autre

69
point serait de se questionner si tous les phages isolés du lait ne sont pas plus efficaces
contre les souches de S. aureus laitières que ceux isolés d'ailleurs ou bien si ceci est un fait
particulier pour LH1 seulement. Ces réflexions nous mènent aussi à nous demander si ces
phages peuvent éliminer la présence de S. aureus dans le fromage, un milieu dans lequel
les phages pourront facilement être dégradés ou inactivés et où le lait peut avoir été
chauffé.
De plus amples recherches se poursuivront afin d'isoler du lait cru d'autres phages
virulents de S. aureus résistants au bas pH et insensibles aux composés du lait. Ceci devrait
être suivi par des recherches sur la combinaison bactériophage-pathogène-aliment afin
d'empêcher une recontamination de la matrice alimentaire par des bactéries mutantes. De
plus, il est intéressant de comprendre la nature et le mode d'action du composé laitier ou de
la constante physico-chimique jouant un rôle dans l'inhibition de l'interaction de S. aureus
et de ses phages et de déterminer s'il provient du lactosérum ou du lait caillé. Finalement,
l'application alimentaire devrait se faire à la température utilisée dans les industries
laitières et non à la température optimale de croissance de S. aureus. Elle devrait aussi se
réaliser avec d'autres phages et d'autres souches de S. aureus.
Toutes ces réflexions et perspectives sont suggérées afin d'optimiser les conditions pour
l'industrie alimentaire et minimiser les risques de contamination et d'intoxication par S.
aureus.

70
6. Bibliographie 1. Abuladze, T., M. Li, M. Y. Menetrez, T. Dean, A. Senecal et A. Sulakvelidze.
2008. Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli 0157:H7. Appl. Environ. Microbiol. 74:6230-6238.
2. Acco, M., F. S. Ferreira, J. A. P. Henriques et E. C. Tondo. 2003. Identification of multiple strains of Staphylococcus aureus colonizing nasal mucosa of food handlers. Food Microbiol. 20:489-493.
3. Ackermann, H.-W. 1975. La classification des bacteriophages des cocci gram-positifs: Micrococcus, Staphylococcus et Streptococcus. Pathologie-Biologie 23:247-253.
4. Ackermann, H.-W. 1999. Tailed bacteriophages: The Order Caudovirales. Adv. Virus Res. 51:135-201.
5. Ackermann, H. -W. 2001. Frequency of morphological phage descriptions in the year 2000. Arch. Virol. 146:843-857.
6. Ackermann, H.-W. 2009. Phage classification and characterization. Tiré de Bacteriophages: Methods .and Protocols. Volume 1: Isolation, Characterization, and Interactions, vol. 501, Humana Press.
7. Ackermann, H. -W., M. S. DuBow, A. W. Jarvis, L. A. Jones, V. N. Krylov, J. Maniloff, J. Rocourt, R. S. Safferman, J. Schneider, L. Seldin, T. Sozzi, P. R. Stewart, M. Werquin et L. Wiinsche. 1992. The species concept and its application to tailed phages. Arch. Virol. 124:69-82.
8. Ackermann, H. -W., A. Elzanowski, G. Fobo et G. Stewart. 1995. Relationships of tailed phages: a survey of protein sequence identity. Arch. Virol. 140:1871-1884.
9. Adams, M. R. et M. O. Moss. 2008. Food Microbiology, 3rd ed, vol. Royal Society of Chemistry Publishing, Cambridge.
10. Altschul, S. F., W. Gish, W. Miller, E. W. Myers et D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410.
11. Anderson, J. E., R. R. Beelman et S. Doores. 1996. Persistence of serological and biological activities of staphylococcal enterotoxin A in canned mushrooms. J. Food Protect. 59:1292-1299.

71
12. Andreoletti, O., H. Budka, S. Buncic, P. Colin, J. D. Collins, A. De Koeijer, J. Griffin, A. Havelaar, J. Hope, G. Klein, H. Kruse, S. Magnino, A. M. Lopez, J. McLauchlin, Ch. Nguyen-The, K. Noeckler, B. Noerrung, M. P. Maradona, T. Roberts, I. Vagsholm et E. Vanopdenbosch. 2009. The use and mode of action of bacteriophages in food production. Scientific opinion of the panel on biological hazards. EFSA J. 1076:1-26.
13. Arvidson, S. et K. Tegmark. 2001. Regulation of virulence determinants in Staphylococcus aureus. 291:159-170.
14. Asao, T., Y. Kumeda, T. Kawai, T. Shibata, H. Oda, K. Haruki, H. Nakazawa et S. Kozaki. 2003. An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 130:33-40.
15. Baird-Parker, A. C. 1962. An improved diagnostic and selective medium for isolating coagulase positive staphylococci. J. Appl. Microbiol. 25:12-19.
16. Baird-Parker, A. C. 1990. The staphylococci: an introduction. Soc. Appl. Bacteriol. Symp. Ser. 19:1S-8S.
17. Balaban, N. et A. Rasooly. 2000. Staphylococcal enterotoxins. Int. J. Food Microbiol. 61:1-10.
18. Bera, A., R. Biswas, S. Herbert et F. Gotz. 2006. The presence of peptidoglycan O-acetyltransferase in various staphylococcal species correlates with lysozyme resistance and pathogenicity. Infect. Immun. 74:4598-4604.
19. Bera, A., R. Biswas, S. Herbert, E. Kulauzovic, C. Weidenmaier, A. Peschel et F. Gotz. 2007. Influence of wall teichoic acid on lysozyme resistance in Staphylococcus aureus. J. Bacteriol. 189:280-283.
20. Bera, A., S. Herbert, A. Jakob, W. Vollmer et F. Gotz. 2005. Why are pathogenic staphylococci so lysozyme resist.ant? The peptidoglycan O-acetyltransferase OatA is the major determinant for lysozyme resistance of Staphylococcus aureus. Mol. Microbiol. 55:778-787.
21. Bergdoll, M. S. 1983. Staphylococci and staphylococcal infections, Easman, C.S.F. and Adlam, C. ed, vol. Academic Press, London, UK.
22. Bergh, O., K. Y. Borsheim, G. Bratbak et M. Heldal. 1989. High abundance of viruses found in aquatic environments. Nature 340:467-468.

72
23. Borodovsky, M. et J. Mclninch. 1993. GENMARK: Parallel gene recognition for both DNA strands. Comp. Chem. 17:123-133.
24. Borysowski, J., B. Weber-Dabrowska et A. Gorski. 2006. Bacteriophage endolysins as a novel class of antibacterial agents. Exp. Biol. Med. 231:366-377.
25. Bradley, D. E. 1967. Ultrastructure of bacteriophage and bacteriocins. Microbiol. Mol. Biol. Rev. 31:230-314.
26. Bronner, S., H. Monteil et G. Prévost. 2004. Regulation of virulence determinants in Staphylococcus aureus: complexity .and applications. FEMS Microbiol. Rev. 28:183-200.
27. Brûssow, H., C. Canchaya et W.-D. Hardt. 2004. Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogénie conversion. Microbiol. Mol. Biol. Rev. 68:560-602.
28. Burt, S. 2004. Essential oils: their antibacterial properties and potential applications in foods-a review. Int. J. Food Microbiol. 94:223-253.
29. Calsamiglia, S., M. Busquet, P. W. Cardozo, L. Castillejos et A. Ferret. 2007. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 90:2580-2595.
30. Canchaya, C , C. Proux, G. Fournous, A. Bruttin et H. Brûssow. 2003. Prophage genomics. Microbiol. Mol. Biol. Rev. 67:238-276.
31. Capparelli, R., M. Parlato, G. Borriello, P. Salvatore et D. Iannelli. 2007. Experimental phage therapy against Staphylococcus aureus in mice. Antimicrob. Agents Chemother. 51:2765-2773.
32. Casman, E. P. 1965. Staphylococcal enterotoxin. Ann. NY Acad. Sci. 128:124-131.
33. Chapman, G. H. 1945. The significance of sodium chloride in studies of staphylococci. J. Bacteriol. 50:201-203.
34. Charlier, C , M. Cretenet, S. Even et Y. Le Loir. 2009. Interactions between Staphylococcus aureus and lactic acid bacteria: An old story with new perspectives. Int. J. Food Microbiol. 31:30-39.

73
35. Chatterjee, A. N. 1969. Use of bacteriophage-resistant mutants to study the nature of the bacteriophage receptor site of Staphylococcus aureus. J. Bacteriol. 98:519-527.
36. Chibani-Chennoufi, S., A. Bruttin, M.-L. Dillmann et H. Brûssow. 2004. Phage-host interaction: an ecological perspective. J. Bacteriol. 186:3677-3686.
37. Chibani-Chennoufi, S., M.-L. Dillmann, L. Marvin-Guy, S. Rami-Shojaei et H. Brûssow. 2004. Lactobacillus plantarum bacteriophage LP65: a new member of the SPOl-like genus of the family Myoviridae. J. Bacteriol. 186:7069-7083.
38. Christianson, S., G. R. Golding, J. Campbell, the Canadian Nosocomial Infection Surveillance Program et M. R. Mulvey. 2007. Comparative genomics of Canadian epidemic lineages of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 45:1904-1911.
39. Coyette, J. et J. M. Ghuysen. 1968. Structure of the cell wall of Staphylococcus aureus, strain Copenhagen. IX. Teichoic acid and phage adsorption. Biochemistry 7:2385-2389.
40. Daniel, A., P. E. Bonnen et V. A. Fischetti. 2007. First complete genome sequence of two Staphylococcus epidermidis bacteriophages. J. Bacteriol. 189:2086-2100.
41. Das, N. K. et R. T. Marshall. 1967. Adsorption of staphylococcal bacteriophage by milk proteins. Appl. Environ. Microbiol. 15:1095-1098.
42. De Billerbeck, V. 2007. Huiles essentielles et bactéries résistantes aux antibiotiques. Phytothérapie 5:249-253.
43. De La Fuente, R., G. Suarez et K. H. Schleifer. 1985. Staphylococcus aureus subsp. anaerobius subsp. nov., the causal agent of abscess disease of sheep. Int. J. Syst. Bacteriol. 35:99-102.
44. Deveau, H., S. J. Labrie, M.-C. Chopin et S. Moineau. 2006. Biodiversity and classification of lactococcal phages. Appl. Environ. Microbiol. 72:4338-4346.
45. Dlawer Ala'Aldeen et K. Hiramatsu. 2004. Staphylococcus aureus: molecular and clinical aspects, vol. Horwood Publishing Limited, Japan. 267 pp.
46. Duckworth, D. H. 1976. "Who discovered bacteriophage?". Bacteriol. Rev. 40:793-802.

74
47. Even, S., C. Charlier, S. Nouaille, N. L. Ben Zakour, M. Cretenet, F. J. Cousin, M. Gautier, M. Cocaign-Bousquet, P. Loubiere et Y. Le Loir. 2009. Staphylococcus aureus virulence expression is impaired by Lactococcus lactis in mixed cultures. Appl. Environ. Microbiol. 75:4459-4472.
48. Evenson, M. L., M. W. Hinds, R. S. Bernstein et M. S. Bergdoll. 1988. Estimation of human dose of staphylococcal enterotoxin A from a large outbreak of staphylococcal food poisoning involving chocolate milk. Int. J. Food Microbiol. 7:311-316.
49. Fauquet, C. M., M. A. Mayo, J. Maniloff, U. Desselberger et L. A. Ball. 2005. Virus taxonomy: VHIth report of the International Committee on Taxonomy of Viruses, vol. Academic Press, Elsevier, London.
50. Fitzgerald, J. R. 2008. Staphylococci of animals. Dans J. A. Lindsay (ed.), Staphylococcus: molecular genetics. Caister Academic Press, Norfolk, UK.
51. Garcia, P., C. Madera, B. Martinez et A. Rodriguez. 2007. Biocontrol of Staphylococcus aureus in curd manufacturing processes using bacteriophages. Int. Dairy J. 17:1232-1239.
52. Garcia, P., C. Madera, B. Martinez, A. Rodriguez et J. E. Suarez. 2009. Prevalence of bacteriophages infecting Staphylococcus aureus in dairy samples and their potential as biocontrol agents. J. Dairy Sci. 92:3019-3026.
53. Garcia, P., B. Martinez, J. M. Obeso, R. Lavigne, R. Lurz, et A. Rodriguez. 2009. Functional genomic analysis of two Staphylococcus aureus phages isolated from the dairy environment. Appl. Environ. Microbiol.75: 7663-7673.
54. Gill J. J., P. M. S., K. E. Leslie et M. W. Griffiths. 2006. Bovine whey proteins inhibit the interaction of Staphylococcus aureus and bacteriophage K. J. Appl. Microbiol. 101:377-386.
55. Goerke, C , R. Pantucek, S. Holtfreter, B. Schulte, M. /ink, D. Grumann, B. M. Broker, J. Doskar et C. Wolz. 2009. Diversity of prophages in dominât Staphylococcus aureus clonal lineages. J. Bacteriol. 191: 3462-3468.
56. Gratia, A. 1922. The Twort-D'Hérelle phenomenon. J. Exp. Med. 35:287-302.
57. Hagens, S. et M. L. Offerhaus. 2008. Bacteriophages - New weapons for food safety. Food Technol. 62:46-54.

75
58. Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41:95-98.
59. Iandolo, J. J. 1989. Genetic analysis of extracellular toxins of Staphylococcus aureus. Annu. Rev. Microbiol. 43:375-402.
60. Iandolo, J. J., V. Worrell, K. H. Groicher, Y. Qian, R. Tian, S. Kenton, A. Dorman, H. Ji, S. Lin, P. Loh, S. Qi, H. Zhu et B. A. Roe. 2002. Comparative analysis of the genomes of the temperate bacteriophages phil 1, phi 12, and phi 13 of Staphylococcus aureus 8325. Gene 289:109-118.
61. Kaneko, J., S. Narita-Yamada, Y. Wakabayashi et Y. Kamio. 2009. Identification of ORF636 in phage phi SLT carrying Panton-Valentine Leukocidin genes, acting as an adhesion protein for a poly(Glycerophosphate) chain of lipoteichoic acid on the cell surface of Staphylococcus aureus. J. Bacteriol. 191:4674-4680.
62. Korhonen, H., P. Marnila et H. S. Gill. 2000. Milk immunoglobulins and complement factors. Br. J. Nutr. 84:75-80.
63. Kutter, E. et A. Sulakvelidze. 2005. Bacteriophages: Biology and Applications, E. Kutter & A. Sulakvelidze ed, vol. CRC Press, Washington.
64. Kwan, T., J. Liu, M. DuBow, P. Gros et J. Pelletier. 2005. The complete genomes and proteomes of 27 Staphylococcus aureus bacteriophages. Proc. Nat. Acad. Sci. U.S.A. 102:5174-5179.
65. Labandeira-Rey, M., F. Couzon, S. Boisset, E. L. Brown, M. Bes, Y. Benito, E. M. Barbu, V. Vazquez, M. Hook, J. Etienne, F. Vandenesch et M. G. Bowden. 2007. Staphylococcus aureus Panton-Valentine Leukocidin causes necrotizing pneumonia. Science 315:1130-1133.
66. Labrie, S. 2004. Étude des bacteriophages de Lactococcus lactis: Isolement, détection et génomique. Dans Département de biochimie et de microbiologie, thèse de doctorat, 256 pp., Québec: Université Laval.
67. Lawrence, J. G., G. F. Hatfull et R. W. Hendrix. 2002. Imbroglios of viral taxonomy: genetic exchange and failings of phenetic approaches. J. Bacteriol. 184:4891^1905
68. Le Loir, Y., F. Baron et M. Gautier. 2003. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2:63-76.

76
69. LeJeune, J. T. et P. J. Rajala-Schultz. 2009. Food safety: unpasteurized milk: a continued public health threat. Clin. Infect.Dis. 48:93-100.
70. Lindberg, A. A. 1973. Bacteriophage receptors. Annu. Rev. Microbiol. 27:205-241.
71. Loimarante, V., A. Carlen, J. Olsson, J. Tenovuo, S. Eeva-Liisa et H. Korhonen. 1998. Concentrated bovine colostral whey proteins from Streptococcus mutansi'Strep, sobrinus immunized cows inhibit the adherence of Strep, mutans and promote the aggregation of mutans streptococci. J. Dairy Res. 65:599-607.
72. Lowy, F. D. 2003. Antimicrobial resistance: the example of Staphylococcus aureus. J. Clin. Invest. 111:1265-1273.
73. Lowy, F. D. 1998. Staphylococcus aureus infections. N. Engl. J. Med. 339:520-532.
74. Madigan, M. T., J. M., Martinko, P. V., Dunlap et D. P., Clark. 2009. Brock biology of microorganisms, twelfth ed, vol. Pearson Benjamin Cummings, San Francisco. 1168 pp.
75. Mamo, W. et G. Froman. 1994. In v/vo-like antigenic surface properties of Staphylococcus aureus from bovine mastitis induced upon growth in milk whey. Microbiol. Immunol. 38:801-804.
76. Mamo, W., F. Rozgonyi, S. Hjertén et T. Wadstrôm. 1987. Effect of milk on surface properties of Staphylococcus aureus from bovine mastitis. FEMS Microbiol. Lett. 48:195-200.
77. Mann, N. H. 2008. The potential of phages to prevent MRSA infections. Res. Microbiol. 159:400-405.
78. Moak, M. et I. J. Molineux. 2004. Peptidoglycan hydrolytic activities associated with bacteriophage virions. Mol. Microbiol. 51:1169-1183.
79. Moineau, S., D. Tremblay et S. Labrie. 2002. Phages of lactic acid bacteria: from genomics to industrial applications. ASM News. 68:388-393
V
80. Necidovâ L., Z. S., M. Pospisilovâ, B. Jans tovâ, J. Strejcek, M. Duskovâ et R. Karpiskovâ. 2009. Influence of soft cheese technology on the growth and enterotoxin production of Staphylococcus aureus. Czech J. Food Sci. 27:127-133.
81. Nelson, D. 2004. Phage taxonomy: We agree to disagree. J. Bacteriol. 186:7029-7031.

77
82. NHS. 2007. Identification of Staphylococcus species, Micrococcus species and Rothia species. Standards Unit, Evaluations and Standards Laboratory 2.1:1-17.
83. Norman no, G., A. Firinu, S. Virgilio, G. Mula, A. Dambrosio, A. Poggiu, L. Decastelli, R. Mioni, S. Scuota, G. Bolzoni, E. Di Giannatale, A. P. Salinetti, G. La Salandra, M. Bartoli, F. Zuccon, T. Pirino, S. Sias, A. Parisi, N. C. Quaglia et G. V. Celano. 2005. Coagulase-positive staphylococci and Staphylococcus aureus in food products marketed in Italy. Int. J. Food Microbiol. 98:73-79.
84. O'Flaherty, S., A. Coffey, W. J. Meaney, G. F. Fitzgerald et R. P. Ross. 2005. Inhibition of bacteriophage K proliferation on Staphylococcus aureus in raw bovine milk. Lett. Appl. Microbiol. 41:274-279.
85. O'Flaherty, S., A. Coffey, W. J. Meaney, G. F. Fitzgerald et R. P. Ross. 2005. The recombinant phage lysin lysK has a broad spectrum of lytic activity against clinically relevant staphylococci, including methicillin-resistant Staphylococcus aureus. J. Bacteriol. 187:7161-7164.
86. O'Flaherty, S., R. P. Ross, J. Flynn, W. J. Meaney, G. F. Fitzgerald et A. Coffey. 2005. Isolation and characterization of two anti-staphylococcal bacteriophages specific for pathogenic Staphylococcus aureus associated with bovine infections. Lett. Appl. Microbiol. 41:482-486.
87. O'Flaherty, S., A. Coffey, R. Edwards, W. J. Meaney, G. F. Fitzgerald et R. P. Ross. 2004. Genome of staphylococcal phage K: a new lineage of Myoviridae infecting gram-positive bacteria with a low G+C content. J. Bacteriol. 186:2862-2871.
88. Obeso, J. M., B. Martinez, A. Rodriguez et P. Garcia. 2008. Lytic activity of the recombinant staphylococcal bacteriophage PhiH5 endolysin active against Staphylococcus aureus in milk. Int. J. Food Microbiol. 128:212-218.
89. Olde Riekerink R. et H. Barkema. 2006. Mastitis: The Canadian perspective. WCDS Adv. Dairy Technol. 18:275-283.
90. Pantucek, R., J. Doskar, V. Rûzickovâ, P. Kaspârek, E. Orâcovâ, V. Kvardovâ et S. Rosypal. 2004. Identification of bacteriophage types and their carriage in Staphylococcus aureus. Arch.Virol. 149:1689-1703.
91. Pantucek, R., A. Rosy pa lova, J. Doskar, J. Kailerovâ, V. Rûzickovâ, P. Boreckâ, S. Snopkovâ, R. Horvâth, F. Gôtz et S. Rosypal. 1998. The polyvalent staphylococcal phage phi812: its host-range mutants and related phages. Virology. 246:241-252.

78
92. Peake, S. L., J. V. Peter, L. Chan, R. P. Wise, A. R. Butcher et D. I. Grove. 2006. First report of septicemia caused by an obligately anaerobic Staphylococcus aureus infection in a human. J. Clin. Microbiol. 44:2311-2313.
93. Prescott, L. M., J. P. Harley et D. A. Klein. 2005. Microbiology, Sixth ed, vol. McGraw-Hill New York.
94. Proux, C , D. van Sinderen, J. Suarez, P. Garcia, V. Ladero, G. F. Fitzgerald, F. Desiere et H. Brûssow. 2002. The dilemma of phage taxonomy illustrated by comparative genomics of Sfi21-like Siphoviridae in lactic acid bacteria. J. Bacteriol. 184:6026-6036.
95. Rashel, M., J. Uchiyama, I. Takemura, H. Hoshiba, T. Ujihara, H. Takatsuji, K. Honke et S. Matsuzaki. 2008. Tail-associated structural protein gp61 of Staphylococcus aureus phage phiMRl 1 has bifunctional lytic activity. FEMS Microbiol. Lett. 284:9-16.
96. Rees, P. J. et B. A. Fry. 1981. The morphology of staphylococcal bacteriophage K and DNA metabolism in infected Staphylococcus aureus. J. Gen. Virol. 53:293-307.
97. Rheinbaben, K. E. et R. M. Hadlok. 1981. Rapid distinction between micrococci and staphylococci with furazolidone agars. Ant. van Leeuwenhoek 47:41-51.
98. Rohwer, F. et R. Edwards. 2002. The phage proteomic tree: a genome-based taxonomy for phage. J. Bacteriol. 184:4529-4535.
99. RosenblumE. D. et S. Tyrone. 1964. Serology, density, and morphology of staphylococcal phages. J. Bacteriol. 88:1737-1742.
100. Rountree, P. M. 1949. The serological differentiation of staphylococcal bacteriophages. J. Gen. Microbiol. 3:164-173.
101. Rousseau, G. 2007. Étude de l'évolution des phages de l'espèce 936 de Lactococcus lactis. Université Laval, Québec.
102. Sandgren, C. H., W. Mamo, I. Larsson, M. Lindahl et I. Bjôrk. 1991. A periodate-sensitive anti-phagocytic surface structure, induced by growth in milk whey, on Staphylococcus aureus isolated from bovine mastitis. Microb. Pathog. 11:211-220.
103. Scharff, R. L. 2010. Health related costs from foodborne illness in the United States. Produce safety project. 28 pp.

79
104. Sharp, S. E. et C. Searcy. 2006. Comparison of mannitol salt agar and blood agar plates for identification and susceptibility testing of Staphylococcus aureus in specimens from cystic fibrosis patients. J. Clin. Microbiol 44:4545-4546.
105. Shine, J. et L. Dalgarno. 1975. Determinant of cistron specificity in bacterial ribosomes. Nature. 254:34-38.
106. Sinell, H.-J. 1995. Control of food-borne infections and intoxications. Int. J. Food Microbiol. 25:209-217.
107. Sonnhammer, E. L. L. et R. Durbin. 1995. A dot-matrix program with dynamic threshold control suited for genomic DNA and protein sequence analysis. Gene 167:GC1-GC10.
108. St-Yves, M.-A. 2007-2008. Étude sur l'évolution de Staphylococcus aureus lors de la transformation fromagère au lait cru CILQ. 45 pp.
109. Sulakvelidze, A., Z. Alavidze et J. G. Morris, Jr. 2001. Bacteriophage therapy. Antimicrob. Agents Chemother. 45:649-659.
110. Summer, E. J., J. J. Gill, C. Upton, C. F. Gonzalez et R. Young. 2007. Role of phages in the pathogenesis of Burkholderia, or 'Where are the toxin genes in Burkholderia phages?'. Curr. Opin. Microbiol. 10:410-417.
111. Sutra, L., M. Frederighi, J. L. Jouve. 1998. Manuel de bactériologie alimentaire, vol. Polytechnica, Paris. 308 pp.
112. Tamura, K., J. Dudley, M. Nei et S. Kumar. 2007. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24:1596-1599.
113. Twort, F. W. 1915. An investigation on the nature of ultra-microscopic viruses. The Lancet. 186:1241-1243.
114. USFDA, C. 2006. Agency response letter GRAS notice no. GRN 000198 Disponible sur: http://www.cfsan.fda.gov/~rdb/opa-gl98.html. Consulté le 3 mars 2010.
115. Voet, D. et J. G. Voet. 2005. Biochimie, vol. De Boeck Université, Bruxelles. 1600 pp.
116. Vybiral, D., M. Takac, M. Loessner, A. Witte, U. von Ahsen et U. Blasi. 2003. Complete nucleotide sequence and molecular characterization of two lytic

80
Staphylococcus aureus phages: 44AHJD and P68. FEMS Microbiol. Lett. 219:275-283.
117. Walker, K. 2006. Use of bacteriophages as novel food additives. Michigan State University. Michigan State University. Food Regulation in the United States. FS06 ANR 490/811.
118. Weidenmaier, C. et A. Peschel. 2008. Teichoic acids and related cell-wall glycopolymers in Gram-positive physiology and host interactions. Nat. Rev. Micro. 6:276-287.
119. Wilkinson, B. J. et K. M. Holmes. 1979. Staphylococcus aureus cell surface: capsule as a barrier to bacteriophage adsorption. Infect. Immun. 23:549-552.
120. Wommack, K. E. et R. R. Colwell. 2000. Virioplankton: viruses in aquatic ecosystems. Microbiol. Mol. Biol. Rev. 64:69-114.
121. Ye, Z. H. et C. Y. Lee. 1993. Cloning, sequencing, and genetic characterization of regulatory genes, rinA and rinB, required for the activation of staphylococcal phage phi 11 int expression. J. Bacteriol. 175:1095-1102.
122. Young, R. 1992. Bacteriophage lysis: mechanism and regulation. Microbiol. Rev. 56:430-81.





























