Embed Size (px)
Citation preview

lable at ScienceDirect
Biochimie 100 (2014) 151e158
Contents lists avai
Biochimie
journal homepage: www.elsevier .com/locate/biochi
Research paper
Biological evidence for the world’s smallest tRNAs
Sandra Wende a, Edward G. Platzer b, Frank Jühling d, Joern Pütz c, Catherine Florentz c,Peter F. Stadler d,e,f,g,h,i, Mario Mörl a,*aUniversity of Leipzig, Institute for Biochemistry, Leipzig, GermanybUniversity of California, Riverside, Department of Nematology, Riverside, CA 92521, USAcArchitecture et Réactivité de l’ARN, Université de Strasbourg, CNRS, IBMC, 67084 Strasbourg, FrancedUniversity of Leipzig, Bioinformatics Group, Department of Computer Science and Interdisciplinary Center for Bioinformatics, Leipzig, GermanyeMax Planck Institute for Mathematics in the Sciences, Leipzig, Germanyf Fraunhofer Institut für Zelltherapie und Immunologie e IZI, Leipzig, GermanygDepartment of Theoretical Chemistry, University of Vienna, Vienna, AustriahCenter for Non-coding RNA in Technology and Health, University of Copenhagen, Frederiksberg C, Denmarki Santa Fe Institute, Santa Fe, NM, USA
a r t i c l e i n f o
Article history:Received 26 June 2013Accepted 24 July 2013Available online 17 August 2013
Keywords:EnopleaMitochondriaArmless tRNAProcessingtRNA structureCCA addition
* Corresponding author. Tel.: þ49 (0) 341 9736 911E-mail address: [email protected] (M. Mörl).
0300-9084/$ e see front matter � 2013 Elsevier Mashttp://dx.doi.org/10.1016/j.biochi.2013.07.034
a b s t r a c t
Due to their function as adapters in translation, tRNA molecules share a common structural organizationin all kingdoms and organelles with ribosomal protein biosynthesis. A typical tRNA has a cloverleaf-likesecondary structure, consisting of acceptor stem, D-arm, anticodon arm, a variable region, and T-arm,with an average length of 73 nucleotides. In several mitochondrial genomes, however, tRNA genesencode transcripts that show a considerable deviation of this standard, having reduced D- or T-arms oreven completely lack one of these elements, resulting in tRNAs as small as 66 nts. An extreme case ofsuch truncations is found in the mitochondria of Enoplea. Here, several tRNA genes are annotated thatlack both the D- and the T-arm, suggesting even shorter transcripts with a length of only 42 nts.However, direct evidence for these exceptional tRNAs, which were predicted by purely computationalmeans, has been lacking so far. Here, we demonstrate that several of these miniaturized armless tRNAsconsisting only of acceptor- and anticodon-arms are indeed transcribed and correctly processed by non-encoded CCA addition in the mermithid Romanomermis culicivorax. This is the first direct evidence for theexistence and functionality of the smallest tRNAs ever identified so far. It opens new possibilities towardsexploration/assessment of minimal structural motifs defining a functional tRNA and their evolution.
� 2013 Elsevier Masson SAS. All rights reserved.
1. Introduction
To allow an efficient protein biosynthesis in vertebrate mito-chondria, the circular genome of these organelles encodes a mini-mal set of structural RNAs essential for translation. Besides genesfor the two ribosomal RNAs, typical animal mitogenomes contain22 tRNA genes that are required and sufficient for translation in theorganelle [1]. In comparison to the nuclear encoded translationalmachinery, the mitochondrial system is functional not only withsuch a reduced set of individual tRNAs for 20 different amino acids,but also with smaller tRNA transcripts that have simplified struc-tural properties or lack specific features. Posttranscriptional editingof such tRNA sequences is a frequently observed strategy to obtain acomplete and functional set of mitochondrial tRNAs. In these cases,
; fax: þ49 (0) 341 9736 919.
son SAS. All rights reserved.
50- or 30-terminally truncated tRNA transcripts, resulting fromoverlapping gene organization, are completed by the addition ofthe missing residues, ranging from single positions to stretches ofseveral nucleotides [2e4]. Other editing events alter the identity ofindividual bases by deamination reactions, changing the aminoacid identity of the complete tRNA [5].
Due to these facts, the mitochondrially encoded tRNAsfrequently show strong deviations from their cytosolic counter-parts and their genes are sometimes hard to identify so that specifictRNA search algorithms were developed [6]. In marsupial mito-chondria, this is further complicated by processing place holdersthat are required to ensure a proper release of the neighboringmRNA and rRNAs out of the huge primary transcript that is syn-thesized. A degenerate tRNA for lysine has lost its original tRNAfunction but still acts as a processing signal [7]. The most extremestructural deviation and miniaturization of tRNAs, however, isfound in the mitochondrial genome of nematodes like Caeno-rhabditis elegans or Ascaris suum, where either the D- or the T-arm

S. Wende et al. / Biochimie 100 (2014) 151e158152
is replaced by a simple small loop consisting of five to eight nu-cleotides [8,9].
Recently, in the genome of nematode and mite mitochondria,the first genes for even smaller tRNA transcripts have beendescribed. In the mite Dermatophagoides farina, the mitochondrialgenome harbors genes for tRNAs lacking the D-arm and having onlya small and unstable T-arm [10]. In addition, in the nematode groupof Enoplea, containing the orders Mermithidia (arthropod para-sites), Dorylaimida (plant parasites), Trichocephalida (vertebrateparasites), mitochondrial tRNA gene sets with predicted transcriptslacking both D- or T-domains were detected, leading to furtherreduced tRNA structures [6]. As an example, the mermithidRomanomermis culicivorax carries two mitochondrial tRNA geneswhere the D-arm is missing, 11 with a deletion of the T-arm, andnine with a deletion of both arms. These armless tRNAs wereinferred from the genomic sequence by purely computationalmeans. Nevertheless, there are several arguments why thesetranscripts very likely represent functional molecules. First, thegene sequence is highly conserved between the different Enopleaspecies. Second, the observed base replacements are compensatoryand maintain the proposed secondary structure of the transcripts.Third, the observed genome arrangements do not disturb theintegrity of the individual tRNA gene sequences [6]. It remainshowever unclear to what extent they are functional. Here we pro-vide direct experimental evidence for the existence of the corre-sponding processed transcripts. Intriguingly, these tRNAs carry thehighly conserved nucleotide triplet CCA at the 30-terminus, which isan absolute prerequisite for becoming charged by the cognateaminoacyl tRNA synthetases [11]. As these CCA sequences are notencoded genomically but added in a posttranscriptional reaction bythe CCA-adding enzyme, their presence is compelling evidence thatthese transcripts indeed represent functional tRNAs.
2. Materials and methods
2.1. Rearing and cultivation of R. culicivorax
The isogenic female strain of R. culicivorax 3B4 [12] was reared inmass cultures using the procedures of Stirling & Platzer [13].Autogenous Culex pipiens was employed as the host and the mos-quito larvae were fed a mixture of finely ground laboratory rodentdiet (LabDiet 5001, meal form; 2 parts) and brewer’s yeast (MPBiochemicals 903312; 1 part). Post-parasitic nematodes werecollected in emergence chambers [14] and washed by sedimenta-tion with deionized water and kept at �80 �C until use.
2.2. Preparation of RNA
Total RNA was isolated from 100 mg R. culicivorax 3B4 cells ac-cording to Chomczynski et al. [15]. Worms were homogenized us-ing a 1.4 mm precellys ceramic matrix (Peqlab) for 2 � 40 s at 6 m/sin a FastPrep-24 homogenizer (MP). Isolated total RNAwas DNase I(NEB) digested (2 units) and rRNA was depleted using Ribo-Zero�magnetic gold kit (human/mouse/rat) (Epicentre) according to themanufacturers.
2.3. cDNA synthesis
Using T4 RNA ligase, isolated total RNA was fused to a taggingDNA oligonucleotide that carried a ribonucleotide at the phos-phorylated 50-end and a 30-end blocked by a 30-30- linked T [16].cDNAwas synthesized by hybridizing DNA primer P1 to the taggingoligonucleotide and subsequent primer elongation with RevertAidreverse transcriptase according to the manufacturer (Fermentas).Subsequently, the enzyme was inactivated by heat (70 �C, 10 min).
2.4. tRNA 30-end sequence determination
cDNA of individual mitochondrial tRNAs was amplified in a 50 mlPCR reaction consisting of 35 cycles with 1 min. 95 �C, 1 min. 39 �C,1 min. 60 �C. Due to the extreme AT-content of the sequences, sucha reduced temperature profile was required for efficient amplifi-cation. For polymerization, 1 U Phusion High-Fidelity DNA poly-merase (ThermoScientific) was incubated in the presence of the RTprimer and an upstream primer specific for the 50-part of an indi-vidual tRNA. PCR products were cloned using the Clone Jet CloningKit (Fermentas Molecular Biology Tools; Thermo Scientific) andused for transformation of E. coli. Individual cDNA clones wereamplified by colony PCR and sequenced on an ABI Prism 3700automated sequencer (Amersham Pharmacia Biotech, Freiburg,Germany). Obtained sequences were analyzed using the softwareSeqMan.
2.5. tRNA 50-end sequence determination
The cDNA preparation described above was used in a tailingreaction with terminal deoxynucleotidyl-transferase and dCTP ac-cording to the manufacturer (Invitrogen). The reaction product wasamplified by PCR as described above, using a tail-specific primerand a second oligonucleotide annealing to the 30-part of the indi-vidual tRNA. PCR products were cloned and sequenced as describedabove.
2.6. Oligonucleotides used as primers
Tagging oligonucleotide:50-pUGG ATC GCG TAG CTC ATA CGA GT(inverse)T-30.RT primer:50-AAC TCG TAT GAG CTA CGC GAT C-30.C-tail primer (according to invitrogen):50-GGC CAC GCG TCG ACT AGT ACG GGI IGG GII GGG IIG-30.Upstream amplification primer:50-GGC CAC GCG TCG ACT AGT AC-30.tRNAArg primers:30-end analysis:50-AAA CTT TTA GCA GGA TTT CG-30.50-end analysis:50-AAA ACT TAT ATA AAT TAG GAT TC-30.tRNACys primers:30-end analysis:50-TAA AGA AGA AAC TTC ATT TG-30.50-end analysis:50-CAA AAG AGA TTT TTT TTC ATT TTG-30.tRNAHis primers:30-end analysis:50-TAA TAA ATT AAA TAT AAA TTG TG-30.50-end analysis:50-ATA ATA AAA TTT TTT ATA AAT TCA C-30.tRNAIle primers:30-end analysis:50-TCT TAA TAG AGA ACA ATT TAA ATT G-30.50-end analysis:50-ATC TTA AAT TTT AAT TTA AAT TAT C-30.tRNAPhe primers:30-end analysis:50-GCT TAC AAA AAA GAT TAA ATTe30.50-end analysis:50-ATG CTT ACT TAT TAA TTA AAT TTT C-30.tRNAThr primers:30-end analysis:50-TTC CTG CTG ACT ATG TTT TAT TTT G-30.

S. Wende et al. / Biochimie 100 (2014) 151e158 153
50-end analysis:50-TTC CTT AGT AAT TTT ATT TAC-30.
2.7. tRNA structure prediction
tRNA secondary structures were predicted using the RNAfoldprogram of the Vienna RNA package [17], output structureswere manually curated. Structure presentations were done usingVARNA [18]. 3D structure predictions were calculated using RNAComposer [19].
3. Results
Due to several peculiarities of mitochondrial tRNAs in meta-zoans, including structural deviations and editing, the identifica-tion of unusual tRNA genes in Enoplea is not a direct proof for theexistence and the functionality of the corresponding transcripts. Toinvestigate whether such unexpected armless tRNAs are indeedexpressed in these nematodes, total RNA was isolated fromR. culicivorax material and depleted of ribosomal RNA. This totalRNA preparationwas 30-terminally ligated to a DNA oligonucleotide
Fig. 1. Analysis of tRNA 50- and 30-ends. A. Strategy: The RNA preparation (black) was ligatedsynthesized. For 50-RACE, the cDNA was tailed with C residues (gray). Individual tRNA seque30-part of the individual tRNA. For 30-RACE, the PCR was performed with RT primer and a pshown as an example. Left: The 50-part resulted in a PCR product of about 90 bp, dependingtRNA, a PCR product of about 70e80 bp could be detected, corresponding to the expected
and transcribed into cDNA using a primer complementary to thefused oligonucleotide. The resulting cDNA was subjected to 30-C-tailing and subsequently used as template for the specific amplifi-cation of 50- and 30-ends of the mitochondrial armless tRNAs forArg, Cys, Ile, His, Thr, and Phe (Fig. 1A). A tailing reactionwith A or Tresidues was avoided, as the tRNA sequences are extremely AU-richand would easily lead to frequent mispriming in the subsequentPCR reaction. Due to the high AU content of these transcripts, thePCR elongation step had to be carried out at a reduced temperatureof 60 �C in order to ensure stable primer binding and efficientpolymerization. An example of the resulting amplification productsis shown for 50- and 30-parts of tRNAIle in Fig. 1B. The PCR productswere cloned and individual clones were analyzed by sequencing,yielding between six and 29 sequences for the individual tRNAs.
The analysis of the 50-ends of the tRNAs was successful for threecandidates, tRNAArg, tRNAIle and tRNAThr (Fig. 2A, B; SupplementaryTable). A possible reason for this inefficient 50-end amplificationmight be the considerable difference in the annealing temperaturesof the C-tail primer and the tRNA-specific AT-rich oligonucleotide.In addition, most of the sequences show non-encoded heteroge-neous extra nucleotides between the G-tail and the 50-end of the
to the tagging oligonucleotide (gray). Using the complementary RT primer, cDNA wasnces were amplified using a C-tail primer in combination with a primer specific for therimer specific for the 50-part of the tRNA. B. The amplification products for tRNAIle areon the C-tailing and annealing position of the C-tail primer. Right: For the 30-part of thesize of 76 bp.

Fig. 2. Mini tRNAs. A. As an example, the 50- and 30-sequences of tRNAIle are shown. Primer binding sites are indicated by black arrows. The predicted sequences are indicated inblack characters, identified sequences in color. Lower case characters represent deviations (missing bases) in the predictions, indicated by asterisks. Due to the extreme AU contentof the tRNA sequence, both tRNA specific primers ended at the same G position in the tRNA to ensure proper hybridization of the primer 30-ends. Hence, this individual position isthe only one that was not sequenced. B. Presentation of validated tRNA sequences. Black: predicted sequences. Green: identified sequences, gray: missing 50-parts. Red: deviationsfrom prediction, leading to truncated 50- and 30-end positions, or representing CCA-additions that indicate the functionality of these transcripts. Putative acceptor- and anticodon-stem pairings are indicated in pink and blue.
S. Wende et al. / Biochimie 100 (2014) 151e158154
tRNA sequence which are probably the result of cDNA synthesisartifact (see discussion). Nevertheless, the 50-termini of these threetRNAs are a convincing evidence for the correct expression andprocessing of the transcripts. While tRNAArg and tRNAThr have a 50-end as predicted, tRNAIle is truncated for one position. Hence, thistRNA seems to be even smaller than expected. In contrast to the 50-end determination, the 30-end analysis was successful for all tRNAcandidates. Here, all of the investigated tRNA sequences carried anon-encoded posttranscriptionally added CCA triplet at the 30-ter-minus (Fig. 2B)While most of the clones carried a complete versionof the triplet, several single sequences also showed partial CCAaddition, ending with one or two C-residues. Hence, these productsobviously represent either reaction intermediates of the CCAaddition process or indicate some degradation during preparation,as it is probably also the case in two 50-end sequences of tRNAIle,
where 2 nucleotides are missing (Supplementary Table). In addi-tion, in the tRNAs for Ile, Cys, His, and Phe, the actual 30-end of theencoded tRNA differs from the predicted one, as the expecteddiscriminator position is not present in these transcripts. Instead,the CCA-end is attached to the position upstream of the predicteddiscriminator nucleotide, indicating that the actual tRNA is at leastone nucleotide shorter than predicted originally. In the case oftRNAIle, this truncation corresponds to the situation at the 50-ter-minus and generates an acceptor stem with the conventionalsingle-stranded discriminator position upstream of the CCA-end. Itis highly likely that similar truncations exist at the 50-termini of theother identified tRNAs with shorter 30-ends.
As these transcripts show strong deviations of a standard tRNAsecondary structure, usual tRNA structure prediction algorithmslike tRNAScanSE [20] fail to recognize and fold these sequences,

S. Wende et al. / Biochimie 100 (2014) 151e158 155
even in a mode optimized for nematode mitochondria. Hence,several of the sequences were analyzed using RNAfold from theVienna RNA package [17] and the predicted structures were sub-sequently curated by hand and illustrated using the VARNA tool[18]. For the fully sequenced tRNAArg and tRNAIle, structures withacceptor- and anticodon-like stem elements can be formed (Fig. 3),although the numbers of base pairs in these stems differ from thestandard. D- and T-arms, however, are completely missing and arereplaced by bulges between 5 and 9 nucleotides. Yet, in the onlineRNA Composer program [19], these secondary structures can befolded into small L-shaped tertiary structures that show a consid-erable similarity to conventional tRNA shapes.
4. Discussion
The first complete sequence of a tRNA was identified in 1965and led immediately to the suggestion of several alternative sec-ondary structures [21]. Among these predictions, a cloverleaf-likeform of this transcript was proposed which later turned out to becorrect. This structure represents the basis for the three-dimensional organization of tRNA, resulting in an L-shapedarrangement, where acceptor- and T-stem as well as anticodon-and D-stem are stacked to form elongated helices [22,23]. Thisarrangement turned out to represent the canonical three-dimensional structure of tRNAs in all kingdoms as demonstratedby a large number of crystallographic data (reviewed in Giegé et al.[24],), with a conserved distance of about 70 �A between theanticodon and the CCA terminus, carrying the amino acid. Sur-prisingly, metazoan mitochondrial tRNAs show considerable de-viations from the cloverleaf secondary structure, with sizereductions in loop as well as stem elements, with the mostextreme case of lacking the D- or T-stem. Yet, these tRNAs with asize down to 59 nucleotides (human mt tRNASer) still can fold intothe canonical L shape with conserved dimensions [24].
Hence, it is very surprising that in several recently analyzednematode and mite species, even smaller mitochondrial tRNAswere predicted [10,25]. While these annotations are predictionsbased on DNA sequences, the results presented here are the firstexperimental proof for the actual existence of these minimalizedtRNAs and confirm the correctness of the prediction algorithms.The additional nucleotides located between the 50-end and the C-tail added during the 50-RACE analysis do not represent post-transcriptional editing events but are very likely the result of apreparation artifact. First, the added nucleotides differ from cloneto clone and are not identical within the sequence analysis of anindividual tRNA. For a functional editing reaction, a specific tRNAwould have identical nucleotides added. Second, the most likelyexplanation for the origin of these extra nucleotides is that aremaining excess of dNTPs from the cDNA synthesis was not effi-ciently removed before the C-tailing reaction was started, resultingin the incorporation of these nucleotides by terminal nucleotidyltransferase. Correspondingly, the extensive use of size exclusionchromatography strongly reduced the amount of the additionalnucleotides. Hence, the tRNA 50-ends are very likely to start at theindicated positions, without carrying extra nucleotides.
Nevertheless, in the identified tRNAs for Ile, Cys, His and Phe, theexact 50- and/or 30-ends differ for one position compared to thepredictions, resulting in even shorter transcripts (Fig. 2B). Thiscorrelated recession of the 50- and 30-termini restores the typicalacceptor stem end in tRNAs, where the 30-terminal residue (position73 according to the standard numbering [26]) is unpaired and rep-resents in many transcripts an essential identity element for ami-noacylation, referred to as discriminator position [27]. A special caseis tRNAHis, where this position 73 (usually a C or anA residue) is base-paired with a G residue at position �1 that e in the case of animal
mitochondria e is posttranscriptionally added, leading to an addi-tional G-1/C73 or G-1/A73 base pair [25,28,29]. As it was not possibleto identify the 50-end of the R. culicivoraxmitochondrial tRNAHis, theexact situation in this tRNA remains unclear. Nevertheless, theidentified 30-end is one nucleotide shorter than predicted (Fig. 2B).Hence, if the actual 50-end corresponds to the proposed one, thediscriminator position A73 would be base-paired with a U residue atthe 50-end. In other tRNAHis transcripts, the G residue base-pairedwith the discriminator represents a major identity element for thecorresponding synthetase [30e33]. A deviation of this conservedfeature was recently identified in the cytosolic tRNAHis of Acantha-moeba castellanii, where the discriminator position remains unpaired[34]. It is possible that in R. culicivorax, a similar situation exists in thecorresponding mitochondrial tRNA.
The presented data indicate that these armless tRNAs are indeedcorrectly processed by mitochondrial RNase P and tRNase Z activ-ities and released from the mitochondrial precursor transcripts. Inaddition, and most significantly, the obtained sequences show thatthe released tRNAs represent bona-fide substrates for the CCA-adding enzyme (Fig. 4). Most of the processing reactions might becarried out by specialized mitochondrial enzyme versions. For theCCA-adding enzyme, however, eukaryotic genomes carry only asingle gene, encoding the nuclear/cytosolic as well as the mito-chondrial form of this activity [35e37]. Being responsible for CCAincorporation in both cytosolic and mitochondrial sets of tRNAs,this enzyme must have undergone a specific adaptation in order torecognize such a diverse set of tRNA substrates.
The fact that all analyzed transcripts carry a complete or (insome rare cases) partial CCA-end at the 30-termini is a strong evi-dence that the transcripts do not only represent reduced processingsignals for precursor RNA maturation in order to release the adja-cent mRNAs according to the mitochondrial punctuation model[38]. Instead, these transcripts seem to be functional tRNAs that arecompleted by the essential site for aminoacylation, although tRNAcharging per se and a direct involvement of the tRNAs in mito-chondrial protein synthesis could not be analyzed due to limitedmaterial. In agreement with this, the armless tRNAs have the abilityto fold into three-dimensional structures that resemble the ca-nonical L-shape as anticipated from folding programs, although thedistance between anticodon and aminoacylation site differsstrongly from the standard (w63�A for tRNAArg, w46�A for tRNAIle).Such a structure might be further optimized and stabilized by theintroduction of modifications, as it was shown to be a requirementfor the proper folding of the human mitochondrial tRNALys thatadopts an L shape only if an adenosine residue at position 9 ismodified to m1A [39]. For the armless tRNAs with such reduceddimensions, two solutions are conceivable. Either the tRNA struc-ture is so flexible that it can extend the distance between the twofunctional ends in order to be recognized by synthetases and ri-bosomes, or the mitochondrial translational machinery ofR. culicivorax (and of other Enoplea) has a size very different of whatis known so far.
The sequence data also rule out the possibility that the struc-tural deviations of these minimal transcripts are corrected bymassive insertional editing events as they are described for mito-chondrial transcripts in trypanosomes [40]. Obviously, the corre-sponding mitochondrial maturation, aminoacylation andtranslational machinery co-evolved with the minimization and isadapted to recognize such a peculiar set of tRNAs even in theabsence of otherwise highly conserved structural features. Inter-action with the synthetase may not be hampered by the size of thetRNA. Indeed, it is well established that synthetases from variousorigins are capable of aminoacylating much shorter RNAs, such asmini- or micro-helices mimicking the aminoacyl-accepting arm, oronly part of it [41]. These findings indicate that the synthetase does
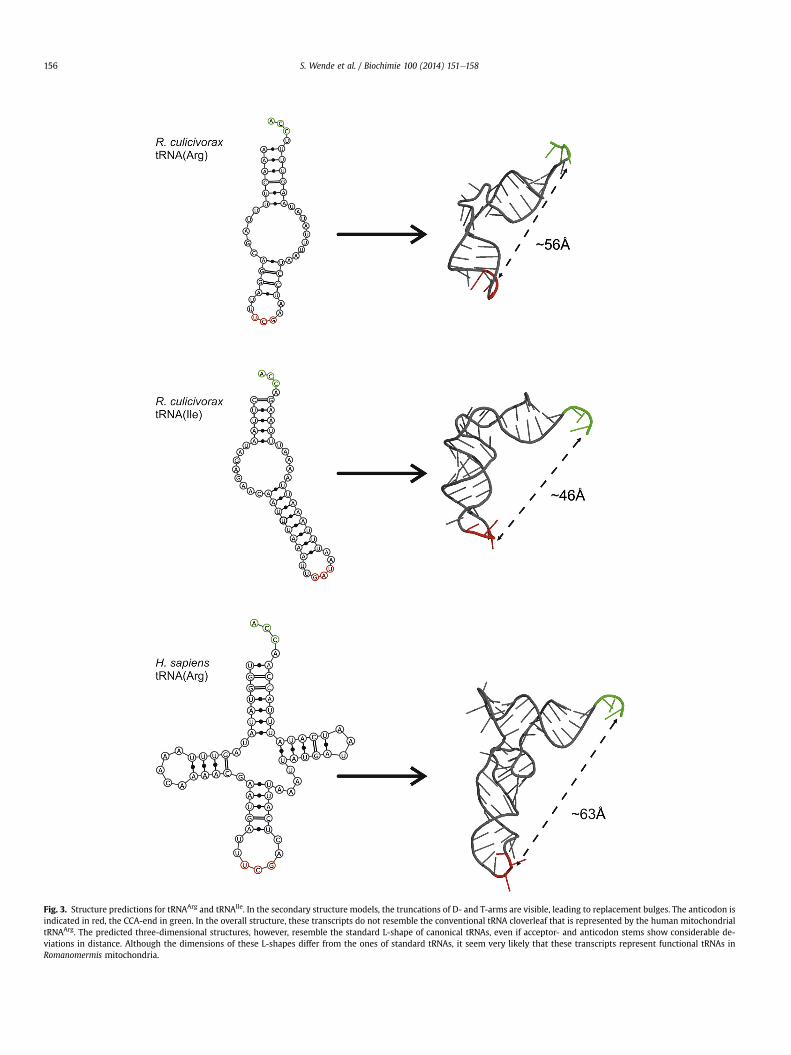
Fig. 3. Structure predictions for tRNAArg and tRNAIle. In the secondary structure models, the truncations of D- and T-arms are visible, leading to replacement bulges. The anticodon isindicated in red, the CCA-end in green. In the overall structure, these transcripts do not resemble the conventional tRNA cloverleaf that is represented by the human mitochondrialtRNAArg. The predicted three-dimensional structures, however, resemble the standard L-shape of canonical tRNAs, even if acceptor- and anticodon stems show considerable de-viations in distance. Although the dimensions of these L-shapes differ from the ones of standard tRNAs, it seem very likely that these transcripts represent functional tRNAs inRomanomermis mitochondria.
S. Wende et al. / Biochimie 100 (2014) 151e158156

Fig. 4. tRNA processing reactions. For both conventional as well as bizarre minimizedtRNAs, the same set of maturation steps, including aminoacylations, is required. Thereactions that were identified in Romanomermis mitochondria are ticked off in green,while the missing reactions are indicated by the red question marks.
S. Wende et al. / Biochimie 100 (2014) 151e158 157
not need to interact with both the anticodon and the acceptingCCA-end. Additional examples for aminoacylation of unusuallystructured tRNA substrates include mimics formed by an anticodonhairpin linked to a CCA-end by a single stranded RNA stretch [42].In mitochondrial ribosomes, a dramatic size reduction in rRNAcomponents was described, leading to ribosomes that obviouslyhave lost the tRNA exit site [43]. Furthermore, the decrease in rRNAsize seems to be compensated by an increase in the number of ri-bosomal proteins, leading to an RNA content of 25e30% inmammalian mitochondria compared to 60e70% in bacteria [44]. InC. elegans, this trend is continued, where the mitochondrial ribo-somes have an even higher protein composition [45]. However, asthe nematode mitochondrial ribosomes have a size larger than thebacterial counterparts [45], a size reduction compensating the tRNAminiaturization is excluded. It is more likely that specific features ofribosomal RNAs or proteins evolved in order to tolerate suchreduced tRNAs as translational adapters. One such co-evolutionmight concern the A-site finger element that in cytosolic ribo-somes interacts with D- and T-arms of an incoming tRNA. Inmitochondrial ribosomes, this element is missing so that no sensingfor D- and T-loop structures occurs [46]. Despite such likely adap-tations in the mitochondrial ribosomes accepting armless tRNAs, itis possible that the efficiency of translation is reduced in compar-ison to protein synthesis with canonical tRNAs, as it was shown thatthe bovine mt tRNASer
GCU, lacking the D-arm, leads to much lowerprotein synthesis [47].
In other components of the mitochondrial translation machin-ery, adaptations to recognize non-canonical tRNA structures werealready identified. For the conventional interaction of EF-Tu,recognition of the T-arm of tRNAs is required. In C. elegans, this isnot possible for most of the 20 mitochondrial tRNAs, as thisstructural element is deleted. A gene duplication of EF-Tu thenallowed a co-evolution of one copy of the factor with the bizarremitochondrial tRNA genes [48]. To recognize these transcripts, thenew version of EF-Tu obtained an additional C-terminal domain,allowing a specific interaction with the D-loop of these tRNAs [49].Whether similar adaptations occurred in R. culicivorax in the genesfor RNase P, tRNase Z, CCA-adding enzyme and aminoacyl tRNAsynthetases currently remains to be clarified.
5. Conclusions
Despite the remarkable conservation of tRNAs in most trans-lation systems there is growing evidence for dramatic deviationsfrom the norm in particular in mitochondria. Aberrant geneticcodes as well as bizarre tRNA architectures lacking either the T-arm
or the D-arm apparently have evolved independently in severalclades [6,10,50]. The armless tRNAs of Enoplea emphasize theflexibility of the protein synthesis machinery despite the stringentselection pressures acting on it. We have provided here the firstevidence that tRNAs with a size of 42 nts and a structure reduced totwo base-paired stems connected by a variable bulge element areprocessed like conventional tRNAs, leading to transcripts endingwith a single unpaired nucleotide at the 30-end, followed by aposttranscriptionally added CCA-sequence.
It will be interesting to see whether there are signs of co-evolution of RNA structures and interacting protein partners, asalready observed in nematodes for the unique elongation factor EF-Tu and the T-armless tRNAs in Chromadorea such as C. elegans.There, an extension of the protein recognized the D-loop instead[49]. The drastically reduced structure and the unusual lack of thediscriminator position suggests that other steps in tRNA processing,including the cleavage by RNase P RNA and tRNase Z, as well as CCAaddition may be adapted to the unusual shape of these tRNAs. Theinvestigation of this co-evolution as well as the interplay of thesemolecules will expand our understanding of functionality andmechanism of protein synthesis.
Acknowledgments
We thank Sonja Bonin and Tobias Friedrich for expert technicalassistance and Heike Betat for valuable discussion. This work wassupported by the Deutsche Forschungsgemeinschaft DFG (Mo 634/8-1, Sta 850-3), by CNRS/Université de Strasbourg and the FrenchExcellence Program (Labex MitCross; ANR-10-IDEX-002-02).
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.biochi.2013.07.034.
References
[1] C.G. Kurland, Problems and paradigns. Evolution of mitochondrial genomesand the genetic code, Bioessays 14 (1992) 709e714.
[2] M.G. Abad, Y. Long, A. Willcox, J.M. Gott, M.W. Gray, J.E. Jackman, A role fortRNA(His) guanylyltransferase (Thg1)-like proteins from Dictyostelium dis-coideum in mitochondrial 50-tRNA editing, RNA 17 (2011) 613e623.
[3] J.M. Gott, B.H. Somerlot, M.W. Gray, Two forms of RNA editing are required fortRNA maturation in Physarum mitochondria, RNA 16 (2010) 482e488.
[4] A. Reichert, U. Rothbauer, M. Mörl, Processing and editing of overlappingtRNAs in human mitochondria, J. Biol. Chem. 273 (1998) 31977e31984.
[5] G.V. Börner, M. Mörl, A. Janke, S. Pääbo, RNA editing changes the identity of amitochondrial tRNA in marsupials, EMBO J. 15 (1996) 5949e5957.
[6] F. Jühling, J. Pütz, C. Florentz, P.F. Stadler, Armless mitochondrial tRNAs inenoplea (nematoda), RNA Biol. 9 (2012) 1161e1166.
[7] M. Dörner, M. Altmann, S. Pääbo, M. Mörl, Evidence for import of a lysyl-tRNAinto marsupial mitochondria, Mol. Biol. Cell 12 (2001) 2688e2698.
[8] R. Okimoto, D.R. Wolstenholme, A set of tRNAs that lack either the T psi C armor the dihydrouridine arm: towards a minimal tRNA adaptor, EMBO J. 9 (1990)3405e3411.
[9] D.R. Wolstenholme, J.L. Macfarlane, R. Okimoto, D.O. Clary, J.A. Wahleithner,Bizarre tRNAs inferred from DNA sequences of mitochondrial genomes ofnematode worms, Proc. Natl. Acad. Sci. USA 84 (1987) 1324e1328.
[10] P.B. Klimov, B.M. Oconnor, Improved tRNA prediction in the American housedust mite reveals widespread occurrence of extremely short minimal tRNAs inacariform mites, BMC Genomics 10 (2009) 598.
[11] M. Sprinzl, F. Cramer, The -C-C-A end of tRNA and its role in proteinbiosynthesis, Prog. Nucleic Acid Res. Mol. Biol. 22 (1979) 1e69.
[12] T.O. Powers, E.G. Platzer, B.C. Hyman, Large mitochondrial genome andmitochondrial DNA size polymorphism in the mosquito parasite, Romano-mermis culicivorax, Curr. Genet. 11 (1986) 71e77.
[13] A.M. Stirling, E.G. Platzer, Catenaria anguillulae in the mermithid nematodeRomanomermis culicivorax, J. Invertebr. Pathol. 32 (1978) 348e354.
[14] J.J. Petersen, O.R. Willis, Procedures for the mass rearing of a mermithidparasite of mosquitoes, Mosquito News 32 (1972) 226e230.
[15] P. Chomczynski, N. Sacchi, Single-step method of RNA isolation by acid gua-nidinium thiocyanate-phenol-chloroform extraction, Anal. Biochem. 162(1987) 156e159.

S. Wende et al. / Biochimie 100 (2014) 151e158158
[16] H. Dickinson, S. Tretbar, H. Betat, M. Mörl, The TRAMP complex shows tRNAediting activity in S. cerevisiae, Mol. Biol. Evol. 29 (2012) 1451e1459.
[17] R. Lorenz, S.H. Bernhart, C. Höner Zu Siederdissen, H. Tafer, C. Flamm,P.F. Stadler, I.L. Hofacker, ViennaRNA package 2.0, Algorithms Mol. Biol. 6(2011) 26.
[18] K. Darty, A. Denise, Y. Ponty, VARNA: interactive drawing and editing of theRNA secondary structure, Bioinformatics 25 (2009) 1974e1975.
[19] M. Popenda, M. Szachniuk, M. Antczak, K.J. Purzycka, P. Lukasiak, N. Bartol,J. Blazewicz, R.W. Adamiak, Automated 3D structure composition for largeRNAs, Nucleic Acids Res. 40 (2012) e112.
[20] P. Schattner, A.N. Brooks, T.M. Lowe, The tRNAscan-SE, snoscan and snoGPSweb servers for the detection of tRNAs and snoRNAs, Nucleic Acids Res. 33(2005) W686eW689.
[21] R.W. Holley, J. Apgar, G.A. Everett, J.T. Madison, M. Marquisee, S.H. Merrill,J.R. Penswick, A. Zamir, Structure of a Ribonucleic acid, Science 147 (1965)1462e1465.
[22] S.H. Kim, F.L. Suddath, G.J. Quigley, A. McPherson, J.L. Sussman, A.H. Wang,N.C. Seeman, A. Rich, Three-dimensional tertiary structure of yeast phenyl-alanine transfer RNA, Science 185 (1974) 435e440.
[23] H. Shi, P.B. Moore, The crystal structure of yeast phenylalanine tRNA at 1.93 Aresolution: a classic structure revisited, RNA 6 (2000) 1091e1105.
[24] R. Giegé, F. Jühling, J. Pütz, P. Stadler, C. Sauter, C. Florentz, Structure oftransfer RNAs: similarity and variability, Wiley Interdiscip. Rev. RNA 3 (2012)37e61.
[25] F. Jühling, M. Mörl, R.K. Hartmann, M. Sprinzl, P.F. Stadler, J. Pütz, tRNAdb2009: compilation of tRNA sequences and tRNA genes, Nucleic Acids Res. 37(2009) D159eD162.
[26] M. Sprinzl, F. Grueter, A. Spelzhaus, D.H. Gauss, Compilation of tRNA se-quences, Nucleic Acids Res. 8 (1980) r1er22.
[27] D.M. Crothers, T. Seno, G. Söll, Is there a discriminator site in transfer RNA?Proc. Natl. Acad. Sci. USA 69 (1972) 3063e3067.
[28] D. L’Abbé, B.F. Lang, P. Desjardins, R. Morais, Histidine tRNA from chickenmitochondria has an uncoded 5’-terminal guanylate residue, J. Biol. Chem. 265(1990) 2988e2992.
[29] C. Francklyn, J. Arnez, Histidyl-tRNA synthetases, in: M. Ibba, C. Francklyn,S. Cusack (Eds.), The Aminoacyl-tRNA Synthetases, 2005, pp. 265e277.
[30] H. Himeno, T. Hasegawa, T. Ueda, K. Watanabe, K. Miura, M. Shimizu, Role ofthe extra G-C pair at the end of the acceptor stem of tRNA(His) in amino-acylation, Nucleic Acids Res. 17 (1989) 7855e7863.
[31] C. Francklyn, P. Schimmel, Enzymatic aminoacylation of an eight-base-pairmicrohelix with histidine, Proc. Natl. Acad. Sci. USA 87 (1990) 8655e8659.
[32] J. Rudinger, C. Florentz, R. Giegé, Histidylation by yeast HisRS of tRNA or tRNA-like structure relies on residues -1 and 73 but is dependent on the RNAcontext, Nucleic Acids Res. 22 (1994) 5031e5037.
[33] I.U. Heinemann, A. Nakamura, P. O’Donoghue, D. Eiler, D. Soll, tRNAHis-gua-nylyltransferase establishes tRNAHis identity, Nucleic Acids Res. 40 (2011)333e344.
[34] B.S. Rao, F. Mohammad, M.W. Gray, J.E. Jackman, Absence of a universalelement for tRNAHis identity in Acanthamoeba castellanii, Nucleic Acids Res.41 (2013) 1885e1894.
[35] J.Y. Chen, P.B. Joyce, C.L. Wolfe, M.C. Steffen, N.C. Martin, Cytoplasmic andmitochondrial tRNA nucleotidyltransferase activities are derived from thesame gene in the yeast Saccharomyces cerevisiae, J. Biol. Chem. 267 (1992)14879e14883.
[36] A.S. Reichert, D.L. Thurlow, M. Mörl, A eubacterial origin for the human tRNAnucleotidyltransferase? Biol. Chem. 382 (2001) 1431e1438.
[37] T. Nagaike, T. Suzuki, Y. Tomari, C. Takemoto-Hori, F. Negayama, K. Watanabe,T. Ueda, Identification and characterization of mammalian mitochondrialtRNA nucleotidyltransferases, J. Biol. Chem. 276 (2001) 40041e40049.
[38] D. Ojala, J. Montoya, G. Attardi, tRNA punctuation model for RNA processing inhuman mitochondria, Nature 290 (1981) 470e474.
[39] M. Helm, R. Giegé, C. Florentz, A WatsoneCrick base-pair-disrupting methylgroup (m1A9) is sufficient for cloverleaf folding of human mitochondrialtRNALys, Biochemistry 38 (1999) 13338e13346.
[40] A.M. Estévez, L. Simpson, Uridine insertion/deletion RNA editing in trypano-some mitochondria e a review, Gene 240 (1999) 247e260.
[41] P. Schimmel, R. Giegé, D. Moras, S. Yokoyama, An operational RNA code foramino acids and possible relationship to genetic code, Proc. Natl. Acad. Sci.USA 90 (1993) 8763e8768.
[42] A.D. Wolfson, A.M. Khvorova, C. Sauter, C. Florentz, R. Giegé, Mimics of yeasttRNAAsp and their recognition by aspartyl-tRNA synthetase, Biochemistry 38(1999) 11926e11932.
[43] R.K. Agrawal, M.R. Sharma, Structural aspects of mitochondrial translationalapparatus, Curr. Opin. Struct. Biol. 22 (2012) 797e803.
[44] B.E. Christian, L.L. Spremulli, Mechanism of protein biosynthesis in mamma-lian mitochondria, Biochim. Biophys. Acta 1819 (2012) 1035e1054.
[45] F. Zhao, T. Ohtsuki, K. Yamada, S. Yoshinari, K. Kita, Y. Watanabe, K. Watanabe,Isolation and physiochemical properties of protein-rich nematode mito-chondrial ribosomes, Biochemistry 44 (2005) 9232e9237.
[46] J.A. Mears, M.R. Sharma, R.R. Gutell, A.S. McCook, P.E. Richardson,T.R. Caulfield, R.K. Agrawal, S.C. Harvey, A structural model for the largesubunit of the mammalian mitochondrial ribosome, J. Mol. Biol. 358 (2006)193e212.
[47] K. Watanabe, Unique features of animal mitochondrial translation systems.The non-universal genetic code, unusual features of the translational appa-ratus and their relevance to human mitochondrial diseases, Proc. Jpn. Acad.Ser. B Phys. Biol. Sci. 86 (2010) 11e39.
[48] T. Ohtsuki, Y. Watanabe, T-armless tRNAs and elongated elongation factor Tu,TBMB 59 (2007) 68e75.
[49] M. Sakurai, Y. Watanabe, K. Watanabe, T. Ohtsuki, A protein extension toshorten RNA: elongated elongation factor-Tu recognizes the D-arm of T-armless tRNAs in nematode mitochondria, Biochem. J. 399 (2006) 249e256.
[50] K. Watanabe, S. Yokobori, tRNA modification and genetic code variations inanimal mitochondria, J. Nucleic Acids 2011 (2011) 623095.





























