Embed Size (px)
Citation preview

Chemical Geology, 95 (1992) 347-360 347 Elsevier Science Publishers B.V., Amsterdam
[1]
Biogeochemistry of hot spring environments 3. Apolar and polar lipids in the biologically active layers of
a cyanobacterial mat
Y. Bing Zeng a,~, David M. Ward b, Simon C. Brassell c and Geoffrey Eglinton a aOrganic Geochemistry Unit, School of Chemistry, University of Bristol, Bristol BS8 1TS, UK
bDepartment of Microbiology, Montana State University, Bozeman, MT 59717, USA CDepartrnent of Geology, Stanford University, Stanford, CA 94305-2115, USA
(Received August 14, 1990; revised and accepted August 2, 1991 )
ABSTRACT
Zeng, Y.B., Ward, D.M., Brassell, S.C. and Eglinton, G., 1992. Biogeochemistry of hot spring environments, 3. Apolar and polar lipids in the biologically active layers ofa cyanobacterial mat. Chem. Geol., 95: 347-360.
The apolar lipid, glycolipid and phospholipid components of the biologically active top 5-mm surface layers of a hot spring cyanobacterial mat were investigated. Most of the major components could be associated with bacteria isolated from the mat; the vertical distribution of lipids followed the known or presumed vertical distribution of these organisms. For example, hydrocarbons (e.g., 7-methylheptadecane), phytadienes (methanolysis products from chlorophyll a) and polar lipid fatty acids typical of mat-forming cyanobacteria maximize in the top 0-1 mm and decrease in concentration with depth. Wax esters and octadecanol (produced upon methanolysis of bacteriochlorophyll cs) typical of Chloroflexus aurantiacus maximized in the 1-2- and 2-4-mm intervals. Long-chain diols derived mainly from glycolipids and typical of the aerobic heterotroph Thermomicrobium roseum maximize in the 1-2- and 2-4-mm intervals. 1-O-Alkylglycerols derived from polar lipids and typical of anaerobic fermentative or sulphate-reducing bacteria, increase in concentration with depth and maximize in deeper layers. The relative abundances of lipids appear to reflect the trophic structure of the microbial community.
I. Introduction
This paper is a continuation of our collabo- rative investigation of lipid biomarkers in hot spring microbial mats as model systems in which community composition is simplified and relatively well defined (Ward et al., 1985, 1989 ). In earlier papers of the series we inves- tigated the apolar lipids of the cyanobacterial mat in Octopus Spring, Yellowstone National Park (Dobson et al., 1988), as well as polar lipids of this and other hot spring microbial mats of varying degrees of community com-
"Present address: Institute for Water Sciences, Western Michigan University, Kalamazoo, MI 49008-5150, USA.
plexity (Zeng et al., 1992 in this issue; in this paper referred to as Part 2). Much of our pre- vious work has compared the composition of extractable lipids or polar lipid components in bulk mat samples. Here, we concentrate on the Octopus Spring cyanobacterial mat, well-char- acterized with respect to many of the microor- ganisms which are thought to be involved in photosynthetic formation and subsequent de- composition of the mat. Many of these micro- organisms have been obtained in pure culture and their lipid compositions have been inves- tigated (Ward et al., 1989). The relationship of cultivated to uncultivated mat inhabitants of similar phylogeny is becoming increasingly understood (Ward et al., 1990, 1992). Pro-
0009-2541/92/$05.00 © 1992 Elsevier Science Publishers B.V. All rights reserved.

348 Y.B. ZENG ET AL.
duction and decomposition of this mat occur principally within the top 5 mm (Ward et al., 1987 ). We investigated this zone of biological activity at depth intervals relevant to the dis- tribution of microorganisms and the reactions they catalyze in order to learn whether the ver- tical distribution of mat inhabitants was re- flected in the distribution of the lipids they are likely to synthesize.
2. Methods
Octopus Spring is located in the Lower Gey- ser Basin of Yellowstone National Park, ~ 150 m SSE of Great Fountain Geyser. Samples were removed from a 52-55°C site along the south- ernmost effluent channel using a stainless-steel coring tube (44-mm diameter). Using a spat- ula one core was immediately sectioned along natural laminae into the top green layer ( ~ 0- 1 mm; 160 mg dry weight), a reddish under- layer ( ~ 1-2 mm; 84 mg dry weight), a deep- red coloured layer ( ~ 2 - 4 mm; 95 mg dry weight) and a brown-green layer ( ~ 4-5 mm; 41 mg dry weight). Samples were immediately frozen on dry ice for transit, lyophilized upon return to the laboratory and kept frozen except for a few days in transit to the U.K.
All solvents were redistilled and all glass- ware and materials (including sampling mate- rials and containers) were solvent-rinsed be- fore use. The samples were ground to powder with a mortar and pestle before extraction us- ing a modification of the Bligh and Dyer (1959) method (see Part 2). The total lipid extract was separated into apolar lipid, glyco- lipid and phospholipid fractions by column chromatography (see Part 2 ) and their weights were determined after solvent evaporation. Following addition of internal standards, gly- colipid and phospholipid fractions were sub- jected to methanolysis, derivatized with N,O- bis (trimethylsilyl)trifluoroacetamide (BSTFA) and analyzed by gas chromato- graphy (GC) and gas chromatography-mass spectrometry (GC-MS) as previously de- scribed (see Part 2 ).
3. Results
3. I. Lipid class composition
Concentrations of the various fractions, and the total concentrations of wax ester compo- nents and polar lipid fatty acid methyl ester (FAME) methanolysis products (estimated
TABLE 1
Compound classes of extractable lipids
Compound class Concentration in #g g - 1 dry mat (% of total extracts )
0-1 mm 1-2 mm 2 - 4 m m 4-5 mm
Apolarl ipids *~ 10,959 ( 1 8 . 0 % ) 26,027 (36 .5% ) 28,235 ( 4 1 . 3 % ) 19,231 (27 .8% ) - Wax esters .2 7,462 20,294 16,254 8,600
Glycolipids *~ 31,507 ( 5 1 . 7 % ) 28,767 ( 4 0 . 4 % ) 27,059 (39 .7% ) 23,077 (33 .3% ) - FAME's .2 15,389 7,806 6,795 3,629
Phospholipids .1 18,493 ( 3 0 . 3 % ) 16,438 (23 .1% ) 12,941 (19 .0% ) 26,923 (38 .9% ) - FAME's .2 8.325 4,219 3,177 4,597
Total extracts .3 60,959 (100%) 71,233 (100%) 68,235 (100%) 69,231 (100%)
* ~Concentration determined by gravimetric method. *:Concentration obtained by summation of GC quantitation of individual methanolysis products (Tables 2-4 ). *3Sum of apolar lipids, glycolipids and phospholipids.

BIOCHEMISTRY OF HOT SPRING ENVIRONMENTS, 3 349
0 - 1 m m
1 . 3 2 Y 1 - 2 m m
1 8
3 , x . A
2 - 4 m m
4 - 5 mrn
11 8 ._ , 1 . | . _ .,
' I ' ' ' ' 1 ' ' ' '
10 20 30
. . . . . 1 I I I I ~ I I
40
20
lB
2O
18 19
320
5O 60
RETENTION TIME (minutes)
Fig. 1. Gas chromatograms of apolar lipid fractions of the biologically active layers of the 52-55°C Octopus Spring cy- anobacterial mat. Assignments and abundances of major components are given in Table 2. Minor constituents include: 15, 23, 24 = n, n-C29, -C37 and -C38 wax esters, respectively; il, i2 = internal standards (n-C23 aikane and 5a (H)-choles- tane, respectively). All carboxyl and hydroxyl groups were present as the TMS esters and ethers, respectively. Unlabelled peaks represent components which could not be unambiguously assigned from their mass spectra.
from GC analysis), are reported in Table 1. The glycolipid fractions (and their component FAME's) were most abundant in the top layer and decreased in concentration with depth. The phospholipid fractions (and their component FAME's) showed a similar pattern, with the exception that concentrations were higher in the 4 -5-mm layer. Apolar lipids and their principal components, wax esters, maximized in the 1-2- and 2 -4-mm subsurface intervals,
where they comprised ~ 4 0 % o f t h e total lipid extracts.
3.2. A p o ~ r l ~ i d s
Gas chromatograms of apolar lipid fractions are shown in Fig. 1 and principal components are quantif ied in Table 2. In all samples wax esters were the dominant components. Several series of wax ester homologs were detected,

350 Y.B. ZENG ET AL.
TABLE 2
Concentration of major compounds in the neutral lipid fractions
Peak label (Fig. 1 )
Compound Concentration (gg g-~ dry mat)
0-1 mm 1-2 mm 2-4 mm 4-5 mm
Hydrocarbons: I 2 3 4
Alcohols: 8 n-Cl7:o
I 1 i-C 17:o
Wax esters: n,n-Chain: 16 C3o 17 C31 18 C32 19 C33 20 C34 21 C35 22 C36
/,n-Chain: 26 C31 27 C32 28 C33 29 C34 30 C35
/,/-Chain: 31 C32 33 C34
n-CtTalkane 248 281 142 200 n-Ctsalkane 136 110 42 97 7Me heptadecane 270 37 12 7 phyt-l-ene 51 43 194 131
30 144 161 82 13 54 54 83
80 204 197 111 255 714 542 317
1,326 3,650 3,165 1,523 1,194 3,491 2,627 1,190 2,633 6,220 4,822 1,639
682 1,659 1,035 322 329 683 517 193
29 124 112 164 112 517 524 651 157 658 519 514 223 867 825 793
91 312 211 126
tr. 47 73 128 11 101 142 239
tr.=trace.
with straight-chain (n,n-) ester components predominating over branched ones. Most in- dividual wax esters showed peak concentra- tion in the 1-2- or 2-4-mm depth interval.
Hydrocarbon fractions contained predomi- nantly n-C17 and n-C18 alkanes and 7Me-hep- tadecane in the 0-1-mm top layer, n-C17 w a s also predominant in deeper layers, whereas 7Me-heptadecane decreased dramatically. Phyt-l-ene, also a major hydrocarbon compo- nent, increased with depth.
Free alcohols ranging from C~5 to C,8 (max- imizing at C~7) were present as minor compo- nents, n-Alkan-1-01s predominated over iso-al- kan-l-ols. Phytol, which was previously
detected in a whole mat sample (see Part 2 ), was below detection in the upper layers we in- vestigated. Similarly, bishomohopan-32-ol, a minor component in the whole mat sample (see Part 2), was not detected in the individ- ual layers.
3.3. Glycolipid fraction constituents
Gas chromatograms of the glycolipid frac- tion methanolysis products are presented in Fig. 2. Major components of this fraction are quantified in Table 3.
FAME's were abundant in the methanolysis products of the glycolipid fractions in all lay-

BIOCHEMISTRY OF HOT SPRING ENVIRONMENTS, 3 351
2 6 7
2
2
6 11
33 i 5 i 20
20
¢, 30 5 ? •
2O
I 26 8 2
• 16 17 1O
~7
2 21
O - l m m
1 -2mm
2-4 ram
28 .L.
26
' I ' ' ' I ' ' ' I ' ' ' I I ' '
10 20 30 40
RETENTION TIME (minutes)
4 -5 ram
1 1 1 1 1 1 1 1 1
50 60
Fig. 2. Gas chromatograms of methanolysis products of glycolipid fractions obtained from the biologically active layers of the 52-55 °C Octopus Spring cyanobacterial mat. Assignments and abundances of major components are given in Table 3. Minor constituents include: 22 = br-C22 alkane- 1,2-diol; 23, 24 = n-C16 and -Cl 7 l-O-alkylglycerol, respectively; 28 = C, 5,C~ 5 1, 2-di-O-dialkylglycerol; 30 = n-C ~ 7:0 alcohol; 33 = phytadienes; il, i2 = internal standards ( t/-C23 alkane and 5a (H)-cholestane, respectively ). All hydroxyl groups were analyzed as the TMS derivatives.
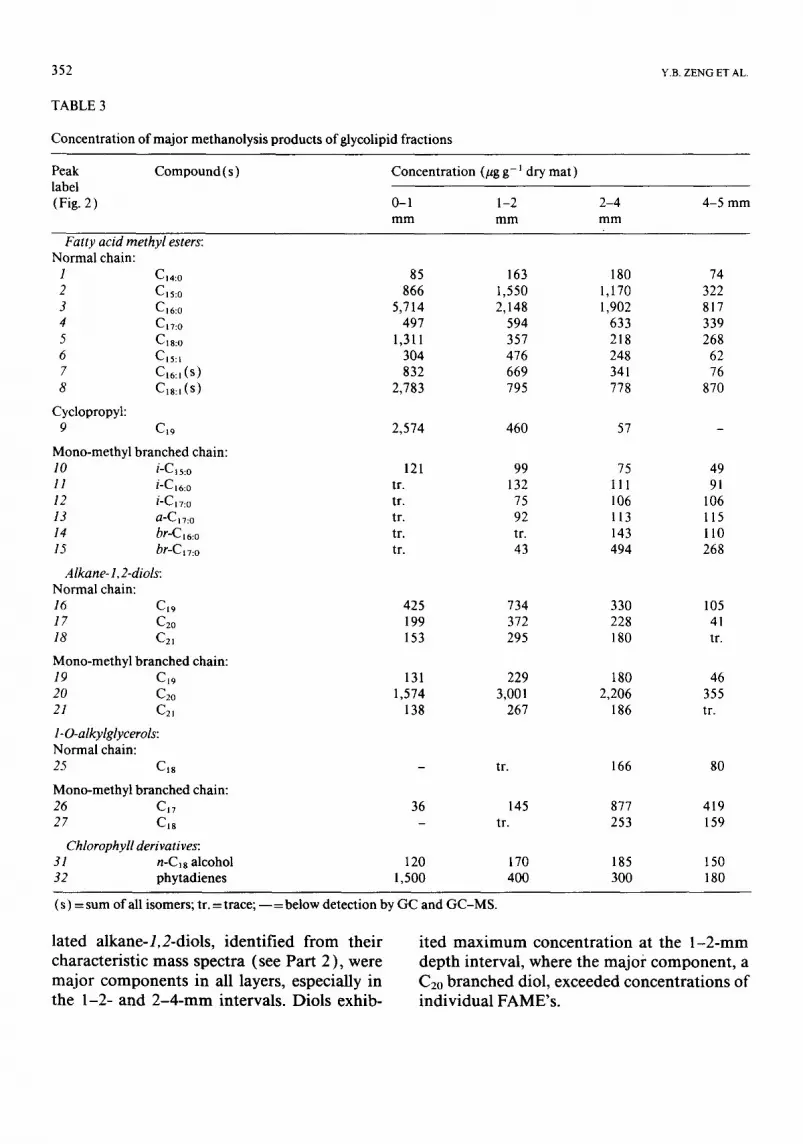
ers. Ind iv idua l F A ME' s showed di f ferent ver- t ical dis t r ibut ions. The top layer was domi-
na ted by C16:0 , C18:0 , Cl6:l, C18:1 and cyclopropyl-C~9 FAME's , and these decreased in concen t ra t ion with depth. Othe r FAME' s present in relat ively high concen t ra t ion in the top layer, such as Cis:0, C~7:0 and Cls:l FAME's ,
showed m a x i m u m concent ra t ions in the 1-2- or 2 - 4 - m m layers, Branched FAME's were rel- a t ively low in concen t ra t ion in all cases, except for i-Cls:O which occurred only in traces in the top layer, and increased in concen t ra t ion in subsurface layers.
CI9-C21 s t raight-chain and m o n o m e t h y -
352 Y.B. ZENG ET AL.
TABLE 3
Concentration of major methanolysis products of glycolipid fractions
Peak label (Fig. 2)
Compound(s) Concentration (#g g - 1 dry mat)
0-1 1-2 2-4 mm mm mm
4-5 mm
Fatty acid methyl esters: Normal chain:
I Ci4:o 85 163 2 C15:o 866 1,550 3 Ct6:o 5,714 2,148 4 C|7:o 497 594 5 Ct8:o 1,311 357 6 Cls:l 304 476 7 Cl6:l(S) 832 669 8 C18:~(s) 2,783 795
Cyclopropyl: 9 Cl9 2,574 460
Mono-methyl branched chain: 10 i-Cis:o 121 99 I 1 i-Cl6:o tr. 132 12 i-Ci7:o tr. 75 13 a-Cl7:o tr. 92 14 br-C 16:0 tr. tr. 15 br-C~ 7:0 tr. 43
Alkane- 1,2-diols: Normal chain: 16 Cl9 425 734 17 C2o 199 372 18 C21 153 295
Mono-methyl branched chain: 19 Cl9 131 229 20 C2o 1,574 3,001 21 C21 138 267
1-O-alkylglycerols: Normal chain: 25 C~8 - tr.
Mono-methyl branched chain: 26 Cl7 36 145 27 C18 - tr.
Chlorophyll derivatives: 31 n-C~8 alcohol 120 170 32 phytadienes 1,500 400
180 1,170 1,902
633 218 248 341 778
57
75 I11 106 113 143 494
330 228 180
180 2,206
186
166
877 253
185 300
74 322 817 339 268
62 76
870
49 91
106 115 110 268
105 41 tr.
46 355 tr.
80
419 159
150 180
(s) = sum of all isomers; tr. = trace; - - = below detection by GC and GC-MS.
l a t e d alkane- l ,2-diols , i d e n t i f i e d f r o m t h e i r
c h a r a c t e r i s t i c m a s s s p e c t r a ( s e e P a r t 2 ) , w e r e
m a j o r c o m p o n e n t s i n a l l l a y e r s , e s p e c i a l l y in
t h e 1 - 2 - a n d 2 - 4 - r a m i n t e r v a l s . D i o l s e x h i b -
i t e d m a x i m u m c o n c e n t r a t i o n a t t h e 1 - 2 - m m
d e p t h i n t e r v a l , w h e r e t h e m a j o r c o m p o n e n t , a
C2o b r a n c h e d d i o l , e x c e e d e d c o n c e n t r a t i o n s o f
i n d i v i d u a l F A M E ' s .

BIOCHEMISTRY OF HOT SPRING ENVIRONMENTS, 3 3 5 3
$
1 2 5
0 - 1 mm
1 - 2 m m
, . ~ ~ I 11 ] 8 9 16 IT Lr/1825
7
IIlll
10
2 - 4 m m
4 - S m m t?
' ' ' I ' ' ' ' I ' '
30 40
R E T E N T I O N T I M E ( m i n u t e s )
I ' ' ' I ' ' ' ' I ' 2O 50 60
Fig. 3. Gas chromatograms of methanolysis products of phospholipid fractions obtained from the biologically active layers of the 52-55°C cyanobacterial mat in Octopus Spring. Assignments and abundances of major components are given in Tables 3 and 4. Minor constituents are assigned in Fig. 2. All hydroxyl groups were analyzed as the TMS derivatives.
Similarly, 1-O-alkylglycerols were also iden- tified from their characteristic mass spectra (see Part 2 ). A l-O-alkylglycerol with a possi- ble methyl branched CI7 alkyl group was pres- ent in all samples and increased with depth to become a major product of methanolysis of the glycolipid fraction in the 2-4-ram interval. Other CI7 and CIS straight-chain or branched
1-O-alkylglycerols were absent or present in only trace amounts in the 0-1- and 1-2-ram layers, but they also increased in deeper layers.
Other products of methanolysis of the gly- colipid fraction may have been derived from chlorophyll pigments which coeluted with the glycolipid fraction on column separation. These included phytadienes, present mainly in

354
TABLE 4
Concentration of major methanolysis products of phospholipid fractions
Y.B. ZENG ET AL.
Peak label (Fig. 3)
Compound(s) Concentration (#g g- 1 dry mat)
0-1 mm 1-2 mm 2-4 mm 4-5 mm
Fatty acid methyl esters: Normal chain:
I C14:o 49 2 Cls:o 273 3 C|6:o 2,800 4 ClT:O 230 5 Cls:o 1,429 7 C16:1 (s) 199 8 CIs:I(S) 1,075
Cyclopropyl: 9 Ci9 872
Mono-methyl branched chain: I0 i - e l 5:0 771 11 i-C16:o 258 12 i-CI7:o 205 I5 br-C 17:o tr.
Alkane- I, 2-diols: Normal chain: 16 Ct9 24 17 C2o 15 18 C21 17
Mono-methyl branched chain: 20 C2o 94 21 C21 7
1-O-alkylglycerols: 25 n-C~s tr. 26 br-C l 7 tr.
52 54 75 399 250 197
1,361 996 1,459 300 289 340 567 503 1,171 135 58 77 308 136 219
341 108 211
345 184 123 105 89 84 193 183 230 30 184 239
90 41 29 54 35 26 53 36 28
306 180 180 24 20 19
15 72 99 46 183 248
(s) =sum of all isomers; tr. =trace.
the top layer and decreasing with depth, and n- C i 7 and n-C~8 alcohols, present mainly in the 1-2-, 2-4- and 4-5-mm layers and more abun- dant in subsurface layers.
A C~5,C~5 1,2-di-O-dialkylglycerol, identi- fied from its mass spectrum (see Part 2), was detected as a minor component in the 0-1-, 1- 2- and 2-4-mm samples.
3.4. Phospholipid fraction constituents
Apart from the low abundance or absence of alcohols and phytadienes, the products of
methanolysis of the phospholipid fraction were similar in composition and depth distribution to those of methanolysis of the glycolipid frac- tion (Fig. 3; Table 4). The predominant prod- ucts were FAME's. In comparison to the gly- colipid FAME's there was a lower relative concentration of C16:l FAME and cyclopropyl- C19 FAME and a higher relative concentration of i-Cls:o FAME. Also, monomethyl FAME's were more abundant in the 0-1-mm layer and decreased in concentration with depth. Diols were much less abundant in the phospholipid fraction than in the glycolipid fraction, but

BIOCHEMISTRY OF HOT SPRING ENVIRONMENTS, 3 3 5 5
showed a vertical profile similar to that of diols derived from the glycolipid fraction, maximiz- ing in the l -2-mm layer, l-O-Alkylglycerol ethers were also less abundant in the phospho- lipid fraction, but, as in the glycolipid fraction, maximized in the deeper layers.
4. Discussion
4.1. Correlations between vertical distribution of lipids and bacteria
There appear to be three distinct classes of vertical distribution of lipids in the Octopus Spring cyanobacterial mat bioactive zone, as illustrated in Fig. 4. These distributions can be interpreted in light of the many component bacteria whose lipids have been studied, tak- ing into account what is known of the vertical distributions of these organisms and the turn- over oflipids which is likely to occur upon bur- ial in the mat.
The major polar lipid FAME's, C16:0 , C18:o
and C18:1, maximize in the 0-1-mm uppermost layer and their concentrations decrease with depth. These are the major polar lipid fatty acids of the cyanobacterial isolate which is thought to play a role in formation of hot spring mats, Synechococcus lividus, when grown at 55°C (Miller, 1976; Fork et al., 1979). These are not particularly distinctive polar lipid fatty acids. They are, for instance, the major total cellular fatty acids of the other cultivated mat phototroph, the photosynthetic green nonsul- fur bacterium Chloroflexus aurantiacus (Ken- yon and Gray, 1974; Knudsen et al., 1982 ), and the aerobic heterotrophic mat isolate Iso- phaera pallida (Giovannoni et al.,1987 ). Orga- nisms such as these are thought to inhabit the 0-1-mm interval (Doemel and Brock, 1977 ). Based on the vertical distributions of chloro- phyll a (Bauld and Brock, 1973), oxygenic photosynthesis (Revsbech and Ward, 1984) and S. lividus-shaped cells (Doemel and Brock, 1977 ), cyanobacteria are restricted to the top 0-1-mm interval. Presumably they consume all
consume all the light available for oxygenic photosynthesis very close to the mat surface. which may be more diagnostic of cyanobac- teria also maximize in the 0-1-mm interval and decrease in concentration with depth below the 0-1-mm layer. Phytadienes are presumably derived during the methanolysis of cyanobac- terial chlorophyll a in the glycolipid fraction. This inference is supported by the compara- tive prominence of phytadienes and n-octade- canol as methanolysis products of glycolipids from cyanobacterial- and Chloroflexus-domi- nated mats, respectively (see Part 2). 7Me- heptadecane is also often attributed to cyano- bacterial sources (Han et al., 1968; Gelpi et al., 1970; Blumer et al., 1971; Shiea et al., 1990).
Based on the vertical distribution of bacte- riochlorophylls (Bauld and Brock, 1973), Chloroflexus aurantiacus is thought to be more abundant in the 1- or 2-mm undermat beneath the cyanobacteria-dominated top layer, where it receives infrared light suitable for its photo- synthesis. Perhaps the most diagnostic lipid biomarkers for this organism are the C28-C38 wax esters it produces (Edmunds, 1982; Knudsen et al., 1982; Shiea et al., 1991 ) as a significant proportion of its lipids ( Beyer et al., 1983 ). Compounds of this type maximized in the 1-2- and 2-4-mm depth intervals, coinci- dent with the distribution of C. aurantiacus. This bacterium produces a unique bacterio- chlorophyll which esterifies mainly n-octade- canol (Gloe and Risch, 1978). It is thus con- sistent that the n-octadecanol released during methanolysis of the glycolipid fraction, possi- bly derived from this bacteriochlorophyll, is most abundant in subsurface layers.
Long-chain diols of the type found in this study have only been reported in the hot spring isolate Thermomicrobium roseum, an aerobic heterotrophic bacterium (Pond et al., 1986). The mat diols were dominated by the branched C2o component, the most abundant diol of T. roseum cultured at 60 °C (Pond and Langwor- thy, 1987). It is certainly possible that other mat inhabitants might also produce diols, but

356
A !1-C16:0 F A M E rng/g TO<:;
o ~ ~ ~
i
_n-Cl8:l FAME mg/g TOC:
o;
phytadienes mcj/g TO<~
Y.B. ZENG ET AL
7MeC17 ug/g TOG
|
i i !
-!
3 c.-
o i
3 ~_
_n,_n-C~ wax ester rng/g TO<3
--m
br-C2o dioi rncj/g TOC
--m
n-Cog alcohol ug/g TOG
o ~
L
c~
L 5" 3
C
5" c~-
~-%7 FAME ug/g "[OC
°i ' ? ?
br-Cl7 monoether
ug/g TO(]
il l I I I
=
Fig. 4. Vertical distributions of lipids in the biologically active layers of the 52-55°C cyanobacterial mat of Octopus Spring: (A) compounds which maximize at the surface and decrease in concentration with depth; (B) compounds which maximize in intermediate depths; and (C) compounds which increase in concentration with depth and maximize in deeper layers.

BIOCHEMISTRY OF H O T S P R I N G E N V I R O N M E N T S , 3 3 5 7
their presence in the 0-1-mm layer and max- imization in the l -2-mm layer is consistent with the aerobic physiology of T. roseum and the greater abundance of oxygen in the layers near the active zone of oxygenic photosyn- thesis (Revsbech and Ward, 1984).
Branched-chain FAME components of the glycolipid and phospholipid fractions showed two distribution patterns with depth. The ma- jor phospholipid branched FAME's, i-Cls:0, - Cl6:O and -Cl7:O FAME's, were most abundant in the 0 - l - m m layer and decreased in concen- tration with depth. These are characteristic components of the phospholipids of two mat heterotrophic isolates which are aerobic, Ther- mus aquaticus (Ray et al., 1971a, b), or facul- tatively aerobic, Bacillus stearothermophilus (Card et al., 1969; Card, 1973). A branched C17:o phospholipid FAME and most of the branched glycolipid FAME's showed an in- crease in concentration with depth, with high- est concentrations in the 2-4- or 4-5-ram lay- ers. This might indicate a source organism of different, possibly anaerobic, physiology. In this regard, it is interesting that the anaerobic fermentative mat isolate, Clostridium thermo- sulfurogenes, is known to produce i-C17 and - C~5 fatty acyl chains (Langworthy and Pond, 1986). Interestingly, the C3o dicarboxylic acid with "head-to-head" condensed iso-C~5 fatty acids, which comprises a major proportion of this organism's lipids (Langworthy and Pond, 1986), was not detected in the mat.
1-O-alkylglycerols maximized in the deeper layers of the mat. Two mat isolates are known to produce glycerol monoethers with alkyl moieties comparable to those of the major mat monoethers. One of these is the anaerobic fer- menter, C. thermosulfurogenes (Langworthy and Pond, 1986); the other is the anaerobic sulphate reducer Thermodesulfobacterium commune (Langworthy et al., 1983). The lat- ter produces mainly glycerol diethers which were not found in abundance in the mat layers, implying that the former organism is the more likely source of the monoethers found.
1,2-Di-O-dialkylglycerols typical of sul- phate-reducing and methanogenic bacteria, which were observed in a bulk mat sample (see Part 2 ), were not detected in the individual mat layers. This is presumably due to the high trophic status and, thus, very low abundance of these organisms and their distinctive lipids. The vertical distribution of ether-linked iso- prenoid lipids in the Octopus Spring 55 °C cy- anobacterial mat was investigated previously (Ward et al., 1985, 1987). Phytanyl and bi- phytanyl ethers characteristic of the only methanogenic bacterium isolated from the mat, Methanobacterium thermoautotrophicum (Tornabene and Langworthy, 1979; Torna- bene et al., 1978), were low in the 0-3-mm in- terval, but maximized in the 3-6-mm and deeper layers, correlating with the obligately anaerobic nature of this organism.
Our analysis was done on samples collected during a mid-day period of high light intensity. As this mat undergoes diurnal change in light and oxygen distribution (Revsbech and Ward, 1984) which might influence repositioning of organisms in the mat, the lipid distribution might be more dynamic than indicated by our results. For instance, Doemel and Brock (1977) have suggested that Chloroflexus mi- grates upward by positive aerotaxis at night - - a possible mechanism driving upward mat ac- cretion. Thus, the vertical distribution of Chloroflexus lipids (e.g., wax esters, octade- canol derived from bacteriochlorophyll c~) might maximize in the 0-1-mm interval after a period of darkness.
4.2. Lipid abundance and trophic structure
As we have previously observed (Ward et al., 1989; and Part 2) there seems to be a correla- tion between lipid patterns and abundances and the expected abundance of organisms oc- cupying different trophic levels in this com- munity. The vertical distribution of lipids rep- resentative of organisms occupying different trophic levels also follows the expected distri-

358 Y.B. ZENG ET AL.
bution of such organisms relative to light and oxygen gradients in the mat. Lipids in upper layers of the mat which are likely to represent inputs of phototrophic microorganisms (base of the food chain, e.g., major polar lipid FAME's, wax esters) are in greatest abun- dance. Lipids which occur in upper or middle mat layers and which may reflect the inputs of aerobic heterotrophic mat decomposers (e.g., diols, some branched polar lipid FAME's), and lipids which maximize in deeper layers and characterize anaerobic fermentative bacteria (e.g., 1-O-alkylglycerols, some branched FAME's) (middle of the food chain) are sec- ondary in abundance. The least abundant lip- ids are those maximizing in the deepest layers and which reflect the inputs of methanogenic bacteria ( ~ 10 and ~25 pg g-1 biphytane and phytane, respectively, released during ether cleavage of the 3-6-mm layer, see Ward et al., 1987). This type of organism terminates the consortium of microorganisms which carries out anaerobic decomposit ion of the mat (top of the food chain).
nonisoprenoid glycerol diethers. Its presence in top layers of the mat suggests that it may orig- inate from a phototrophic or aerobic microor- ganism. Isopentadecane has been released dur- ing ether cleavage of Messel oil shale kerogen of Germany (Chappe et al., 1980) and from polymeric organic matter subfractions of a sea- loch sediment in Scotland (Eglinton, 1983). At present, it is not known whether the diether pentadecyl groups found in the mat are branched.
Lipids of unassigned origin might reflect in- puts of known mat isolates which are not ex- pressed in pure cultures due to some difference between culture and natural environment. However, there is evidence from other lipid analyses (Ward et al., 1985), and more re- cently from 16S rRNA sequence analysis (Ward et al., 1990, 1992) that this mat con- tains numerous uncultivated communi ty members which could be the sources of some of these lipid components.
5. Conclusions
4.3. Lipids of unknown origin
Several lipids we observed are not known to be produced by the many bacteria which have been isolated from the Octopus Spring cyano- bacterial mat. Cyclopropyl-Cl9 FAME, de- rived from both glycolipid and phospholipid fractions, is a major component of the 0-1-mm layer and decreases in concentration with depth, suggesting a possible link with source organisms having either phototrophic or aero- bic metabolism.
Branched-chain wax esters are not known to be produced by C. aurantiacus. However, their similarity in carbon number and depth distri- bution to the straight-chain wax esters proba- bly synthesized by this organism suggests a common origin.
The C l ~,C 15 1,2-di-O-dialkylglyceryl diether is not known to be produced by T. commune, the only mat inhabitant known to synthesize
The major lipid components of the 52-55 ° C Octopus Spring cyanobacterial mat are typical of many bacteria which have been isolated from this communi ty and seem to reflect the known or predicted distribution of these types of microorganisms within the mat vertical pro- file. 7-Methylheptadecane, phytadienes de- rived from glycolipid fraction components (probably chlorophyll a ) and major polar lipid FAME's are typical of mat-forming cyanobac- teria. These show a maximum in the top 0-1 m m and decrease in concentration with depth. Wax esters and octadecanol (presumably de- rived from bacteriochlorophyll cs in the glyco- lipid fraction) are characteristic of the photo- synthetic green nonsulfur bacterium, C. aurantiacus, and maximize in the 1-2- and 2- 4-mm intervals. Long-chain diols derived mainly from the glycolipid fraction are typical of the aerobic heterotrophic thermophile T. ro- scum and maximize in the 1-2-mm interval.

BIOCHEMISTRY OF HOT SPRING ENVIRONMENTS, 3 359
Some b r a n c h e d fa t ty acids and 1-O-alkylgly- cerols de r ived f r om pola r l ipid f rac t ions are
typical o f anaerob ic f e rmen ta t ive isolates, such as C. thermosulfurogenes. These increase in c o n c e n t r a t i o n wi th dep th and m a x i m i z e in deepe r layers. 1,2-Di-O-dialkylglycerols typi- cal o f the un ique su lphate reducer , T. com- mune , or the m e t h a n o g e n , M. thermoautotro- ph icum, i sola ted f rom this ma t are below de tec t ion in po la r l ipid f rac t ions o f ind iv idua l layers. Howe ve r , the ver t ica l d i s t r ibu t ion o f p h y t a n e and b i p h y t a n e de r i ved f rom an e ther c leavage reac t ion suggests tha t m e t h a n o g e n i c bac te r ia also increase in a b u n d a n c e in deepe r layers.
T h e a b u n d a n c e o f the var ious l ipid c o m p o - nents seems to reflect the t roph ic s t ruc ture o f the c o m m u n i t y wi th l ipids charac te r i s t i c o f p h o t o t r o p h s p r e d o m i n a t i n g ove r l ipids char- acter is t ic o f he t e ro t rophs , which in tu rn pre- d o m i n a t e ove r l ipids charac te r i s t ic o f bac te r ia t e rmina t ing the anae rob i c food chain.
Some lipids, notably , cyclopropyl-C19 FAME and C ls ,Cls-di-O-dialkylglycerol ( m e t h a n o - lysis p r oduc t s o f po la r l ipid f rac t ions ) and b r a n c h e d wax esters ( in apo la r l ipid frac- t i ons ) , are no t k n o w n to be p r o d u c e d by bac- ter ia i so la ted f rom this ma t and r ema in o f un- cer ta in origin.
Acknowledgements
Y.B.Z. was s u p p o r t e d by S E D C and the Royal Socie ty o f L o n d o n . G C - M S facili t ies were p r o v i d e d by the U.K. Na tu ra l Env i ron - men t Resea rch Counc i l ( G R C / 2 9 5 1 and G R 3 / 3 7 4 8 ). We t h a n k the U.S. Na t i ona l Sci- ence F o u n d a t i o n (gran t B S R - 8 5 0 6 6 0 2 ) for suppor t ing sample col lec t ion and travel , and the N a t i ona l P a rk Service for grant ing pe rmis - sion to collect samples in Yel lowstone Na- t ional Park.
References
Bauld, J., and Brock, T.D., 1973. Ecological studies of Chloroflexus, a gliding photosynthetic bacterium. Arch. Mikrobiol., 92: 267-284.
Beyer, P., Falk, H. and Kleinig, H., 1983. Particulate frac- tions from Chloroflexus aurantiacus and distribution oflipids and polyprenoid forming activities. Arch. Mi- crobiol., 134: 60-63.
Bligh, E.G. and Dyer, W.J., 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37:911-917.
Blumer, M., Guillard, R.R.L. and Chase, T., 1971. Hy- drocarbons of marine phytoplankton. Mar. Biol., 8: 183-189.
Card, G.L., 1973. Metabolism of phosphatidylglycerol, phosphatidylethanolamine, and cardiolipin of of Ba- cillus stearotkermopkilus. J. Bacteriol., 114:1125-1137.
Card, G.L., Georgi, C.E. and Militzer, W.E., 1969. Phos- pholipids from Bacillus stearothermophilus. J. Bacte- riol., 97: 186-192.
Chappe, B., Michaelis, W. and Albrecht, P., 1980. Molec- ular fossils of archaebacteria as selective degradation products of kerogen. In: A. Douglas and J.R. Maxwell (Editors), Advances in Organic Geochemistry. 1979. Pergamon, Oxford, pp. 265-274.
Dobson, G., Ward, D.M., Robinson, N. and Eglinton, G., 1988. Biogeochemistry of hot spring environments: extractable lipids of a cyanobacterial mat. Chem. Geol., 68: 155-179.
Doemel, W.N. and Brock, T.D., 1977. Structure, growth, and decomposition of laminated algal-bacterial mats in alkaline hot springs. Appl. Environ. Microbiol., 34: 433-452.
Edmunds, K.L.H., 1982. Organic geochemistry of lipids and carotenoids in the Solar Lake microbial mat se- quence. Ph.D. Thesis, University of Bristol, Bristol, (unpublished).
Eglinton, T., 1983. The distribution of lipids in organic matter subfractions from a Scottish sea-loch sedi- ment. M.Sc. Thesis. University of Newcastle-upon- Tyne, Newcastle-upon-Tyne, (unpublished).
Fork, D.C., Murata, N. and Sato, N., 1979. Effect of growth temperature on the lipid and fatty acid com- position, and the dependence on temperature of light- induced redox reactions of cytochrome l and of light energy redistribution in the thermophilic blue-green alga Synechococcus lividus. Plant Physiol., 63: 524-530.
Gelpi, E., Schneider, H., Mann, J. and Oro. J., 1970. Hy- drocarbons of geochemical significance in micro- scopic algae. Phytochemistry, 9:603-612.
Giovannoni, S.J., Godchaux III, W., Schabtach, E. and Castenholz, R.W., 1987. Cell wall and lipid composi- tion of lsosphaera pallida, a budding eubacterium from hot springs. J. Bacteriol.. 169: 2702-2707.
Gloe, A. and Risch, N., 1978. Bacteriochlorophyll cs, a new bacteriochlorophyll from ChlorolTexus aurantiacus. Arch. Microbiol., 102: 103-109.
Han, J., McCarthy, E.D. and Calvin, M., 1968. Hydrocar- bon constituents of the blue-green algae Nostoc mus- corum, Anacystis nidulans, Phormidium luridum and

360 Y.B. ZENG ET AL.
Chlorogloea fritschii. J. Chem. Soc., Sect. C, 1968: 2785-2791.
Kenyon, C.N. and Gray, A.M., 1974. Preliminary analy- sis of lipids and fatty acids of green bacteria and Chlo- roflexus aurantiacus. J. Bacteriol., 120:131-138.
Knudsen, E., Jantzen, E., Bryn, K., Ormerod, J.G. and Sirevag, R., 1982. Quantitative and structural charac- teristics oflipids in Chlorobium and Chloroflexus. Arch. Microbiol., 132:149-154.
Langworthy, T.A. and Pond, J.L., 1986. Membranes and lipids ofthermophiles. In: T.D. Brock (Editor), Ther- mophiles: General, Molecular and Applied Microbi- ology. Wiley, New York, N.Y., pp. 107-135.
Langworthy, T.A., Holzer, G., Zeikus, J.G. and Torna- bene, T.G., 1983. lso- and anteiso-branched glycerol diethers of the thermophilic anaerobe Thermodesulfo- tobacterium commune. Syst. Appl. Microbiol., 4: 1-17.
Miller, L.S., 1976. Effect of carbon dioxide on pigment and membrane content in the thermophilic blue-green alga, Synechococcus lividus. Diss. Abstr. Int., B, 37: 1561-1562.
Pond, J.L. and Langworthy, T.A., 1987. Effect of growth temperature on the long-chain diols and fatty acids of Thermomicrobium roseum. J. Bacteriol., 169: 1328- 1330.
Pond, J.L., Langworthy, T.A. and Holzer, G., 1986. Long- chain diols: a new class of membrane lipids from a thermophilic bacterium. Science, 231: 1134-1136.
Ray, P.H., White, D.C. and Brock, T.D., 1971a. Effect of temperature on the fatty acid composition of Thermus aquaticus. J. Bacteriol., 106: 25-30.
Ray, P.H., White, D.C. and Brock, T.D., 1971b. Effect of growth temperature on the lipid composition of Ther- mus aquaticus. J. Bacteriol., 108: 227-235.
Revsbech, N.P. and Ward, D.M., 1984. Microelectrode studies of interstitial water chemistry and photosyn- thetic activity in a hot spring microbial mat. Appl. En- viron. Microbiol., 48: 270-275.
Shiea, J., Brassell, S.C. and Ward, D.M., 1990. Mid-chain branched mono- and dimethyl alkanes in hot spring cyanobaeterial mats: a direct biogenic source for
branched alkanes in ancient sediments? Org. Geo- chem., 15: 223-231.
Shiea, J., Brassell, S.C. and Ward, D.M., 1991. Compar- ative analysis of free lipids in hot spring cyanobacte- rial and photosynthetic bacterial mats and their com- ponent photosynthetic bacteria. Org. Geochem., 17: 309-319.
Tornabene, T.G. and Langworthy, T.A., 1979. Diphy- tanyl and dibiphytanyl glycerol ether lipids of meth- anogenic archaebacteria. Science, 203:51-53.
Tornabene, T.G., Wolfe, R.S., Balch, W.E., Holzer, G., Fox, G.E. and Oro, J., 1978. Phytanyl-glycerol ethers and squalenes in the archaebacterium Methanobacte- rium thermoautotrophicum. J. Molec. Evol., 11: 259- 266.
Ward, D.M., Brassell, S.C. and Eglinton, G., 1985. Ar- chaebacterial lipids in hot-spring microbial mats. Na- ture (London), 318: 656-659.
Ward, D.M., Tayne, T.A., Anderson, K.L. and Bateson, M.M., 1987. Community structure and interactions among community members in hot spring cyanobac- terial mats. Symp. Soc. Gen. Microbiol., 41: 179-210.
Ward, D.M., Shiea, J., Zeng, Y.B., Dobson, G., Brassell, S.C. and Eglinton, G., 1989. Lipid biochemical mark- ers and the composition of microbial mats. In: Y. Cohen and E. Rosenberg (Editors), Microbial Mats: Physiological Ecology of Benthic Microbial Commu- nities. Am. Soc. Microbiol., Washington, D.C., pp. 439-454.
Ward, D.M., Weller, R. and Bateson, M.M., 1990. 16S rRNA sequences reveal numerous uncultured micro- organisms in a natural community. Nature (London), 345: 63-65.
Ward, D.M., Bateson, M.M., Weller, R. and Ruff, A.L., 1992. Ribosomal RNA analysis of microorganisms as they occur in nature. Adv. Microbial Ecol. (in press).
Zeng, Y.B., Ward, D.M., Brassell, S.C. and Eglinton, G., 1992. Biogeochemistry of hot spring environments, 2. Lipid compositions of Yellowstone (Wyoming, U.S.A. ) cyanobacterial and Chloroflexus mats. Chem. Geol., 95:327-345 (this issue; in this paper referred to as Part 2).





























