Embed Size (px)
Citation preview

ARTICLE IN PRESS
1040-6182/$ - se
doi:10.1016/j.qu
�CorrespondE-mail addr
Quaternary International 191 (2008) 170–183
Animal use at the Postclassic Maya center of Mayapan
Marilyn A. Massona,�, Carlos Peraza Lopeb
aDepartment of Anthropology AS 237, University at Albany—SUNY, Albany, NY 12222, USAbCentro INAH—Yucatan, Km 65 Carretera Progreso, Merida, Yucatan, Mexico 97000, Mexico
Available online 14 February 2008
Abstract
This paper presents archeological evidence for animal use at Mayapan, the largest capital city of the Postclassic period Maya lowlands.
The most commonly consumed species were white-tailed deer, turkey, and iguana, and other important but less frequent animals in the
assemblage were dog, peccary, and brocket deer. A wide variety of local and non-local fauna were also recovered. Our analysis of 97,416
faunal bones is based on two distinct samples that are compared in this study—the site’s monumental center buildings (temples, halls,
shrines, and nearby houses) and the outlying domestic settlement zones. Four arguments are presented regarding Mayapan’s animal use
in this paper. First, certain rare mammals and marine species were likely obtained through trade. Second, evidence suggests that white-
tailed deer were either raised in captivity or were carefully managed in habitats surrounding the city. Surplus deer meat and skeletal
elements were major commodities for exchange and local consumption. Third, dog, exotic animals, and specific deer elements were
preferentially utilized for monumental center activities. Fourth, culturally prescribed methods of ritually discarding deer skulls were
practiced at Mayapan. Mayapan’s faunal exploitation patterns were embedded in important ways within the city’s larger regional,
coastal-inland economic system. Key similarities and differences with other Maya sites are identified.
r 2008 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
This paper investigates some strategies of animal use atthe ancient Maya city of Mayapan. In what ways werevarious animal products important to economic institu-tions such as local food production, hunting, subsistenceexchange, and ritual or social events? Mayapan, the largestpolitical capital of the Late Postclassic period in the Mayaarea, was founded during the 12th century A.D. and wasabandoned by A.D. 1441–1461, only decades before thearrival of the Spanish in the early 1500s (Peraza Lope et al.,2006). Ethnohistorical accounts of faunal use duringMayapan’s occupation and into the Contact period (e.g.,Landa, 1941) are potentially biased, but provide usefulmodels that can be tested with empirical archeologicaldata.
The range and quantities of wild, tamed, and domes-ticated animals found in consumption contexts at the sitereveal diverse methods for animal acquisition and con-sumption, including exchange, fishing, hunting, and
e front matter r 2008 Elsevier Ltd and INQUA. All rights re
aint.2008.02.002
ing author. Tel.: +1518 442 5199.
ess: [email protected] (M.A. Masson).
husbandry or game management. This paper documentsselected aspects of this complex subsistence and economicsystem and contrasts patterns observed in the monumentalcenter versus ordinary domestic contexts. A more detailedanalysis of the spatial distribution of faunal remains byspecific contexts within the monumental center and atvarious elite and commoner houselots throughout the cityis forthcoming.
2. Regional setting: Mayapan
Mayapan is located in the west-central portion of theMexican state of Yucatan, at the northern end of theYucatan peninsula. The city was the largest of its timewithin the Maya area, with a population of around 15,000(Smith, 2005, pp. 411, 419; Russell, 2008). The walledportion of the city itself is 4.2 km2 (Fig. 1), and regulardomestic settlement occurs outside of the city wall to adistance of 500m (Russell, 2008). Ethnohistorical docu-ments suggest that the city’s council of confederatedtownship leaders was likely ruled by a paramount (Ringleand Bey, 2001), and obligations of tribute, corvee labor,and military service were imposed on subject towns (Roys,
served.

ARTICLE IN PRESS
Fig. 1. Top: Location of site monumental center (INAH sample), outlying
milpa areas (domestic zone/PEMY sample) and Mayapan’s city wall.
Bottom: A typical Mayapan commoner house group from Milpa 9
showing walled yard space and a pen.
1Of the 86 PEMY groups, 21 are represented only from surface
collection materials and 65 were sampled in test pits and surface
collections. Of the 30,309 PEMY bones, 2604 are from surface collections
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 171
1962, p. 50). The political nucleus of the city is representedby epicentral buildings surrounding the main plazas wherethe majority of the site’s temples, colonnaded meetinghalls, and specialized ritual buildings are concentrated(Proskouriakoff, 1962). This monumental center is foundin Square Q (Fig. 1), a 500� 500m segment of the site map(Jones, 1962). Surrounding the monumental center is theepicenter including the largest elite dwellings at Mayapanwhich likely served as palaces for the city’s most prominentofficials (Masson et al., n.d.). Outside of the epicenter andwithin the city wall, Mayapan’s domestic zone consists ofdensely packed houselots; over 4000 dwellings and out-buildings have been documented in this outlying settlementzone (Smith, 1962). A typical commoner houselot is shownin Fig. 1. Residential groups were delineated by stone walls(albarradas), as shown in the example (Fig. 1), whichindicate specific social units (Bullard, 1952; Brown, 1999).The domestic zone was probably divided into barrios orwards (kuchteel), as Colonial period sources claim (Roys,1957); these neighborhoods, distributed across through the
city, are thought to be marked by temples, halls, raisedroads, major waterholes (cenotes), a large marketplace,stone lanes, and city gates (Hare et al., 2006). Earlyzooarcheological investigations were conducted by Pollockand Ray (1957, pp. 636–637) who provided a taxonomiclist of 5472 remains from 145 excavation lots at structuresin or near the monumental center (Palace Group R-85-90,temple/hall Q-152/151, Cenote Ch’en Mul, and TempleQ-95).Two zooarcheological samples are compared in this
study. The first is from the site’s monumental center andsome nearby houses (Fig. 2), excavated by the InstitutoNacional de Antropologıa e Historia (INAH—Yucatan),and the second is from the domestic settlement zonelocated outside of this monumental center (Fig. 1),excavated by the authors’ joint project, the ProyectoEconomico de Mayapan (PEMY). The monumental centersample reflects consumption activities of the city’s highest-ranking political and religious officials and includesmaterials that are primarily from the monumental orpolitical center and the surrounding elite palaces. Thesample from the domestic zone includes a majority ofcommoner dwellings and a small number of lesser,secondary elite dwellings and associated buildings. Thebulk of this latter sample reflects the remains of animalused in every day life as foods and other products.
3. Materials and methods
3.1. Collection and analysis
Fig. 1 shows the location of contexts from which thedomestic zone (PEMY) faunal material derives. Clearedmilpas (swidden agricultural plots) represented our sampleunits for mapping, surface collection, and test-pitting inthese non-epicentral locations at Mayapan. Test pitmaterials from these domestic zone contexts were collectedwith the use of a 1
4in screen. The sample from the site
monumental center and nearby houses (Fig. 2) was hand-collected, quite systematically and meticulously by theINAH project. Due to differences in collection strategy,these samples are reported separately in the tables of thisarticle. Analysis occurred in project laboratories in Tecoh,Yucatan, and Albany, New York, using comparativeskeletal collections housed at both localities and standardillustration reference works (e.g., Olsen, 1982; Gilbert,1993). One-hundred percent of the domestic zone samplewas analyzed (from the 2001–2003 seasons), and 85% ofthe monumental center sample was analyzed (from the1996 to 2004 seasons). Altogether, we analyzed 97,416pieces of fauna (67,107 INAH from 94 buildings, 30,309PEMY from 86 building groups).1 Faunal taxonomy

ARTICLE IN PRESS
Fig. 2. Map of monumental center (left), published by Jones (1962). The Temple of Kukulkan (Q-162), adjacent buildings, and location of units discussed
in the text on the right (modified from Delgado Ku, 2004, Fig. 29; Peraza Lope et al., 2007, Fig. 16).
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183172
presented here was verified with www.itis.gov on January7, 2007. All scientific names are presented in the tables.
3.2. Animal products and exchange
The relative frequencies of identified fish skeletalelements are evaluated here to assess evidence for differentmeans of fish acquisition at Mayapan. Comparisons ofmarine catfish to marine non-catfish taxa are offered(following Masson, 2004). An over-representation of fishcranial elements at coastal sites and a similar disproportio-nately high representation of postcranial spines andvertebrae at inland sites are criteria that have been usedto suggest that fish heads were removed at coastal sitesprior to the preparation of salted postcranial portionsdestined for exchange (Carr, 1985; Rackham, 1994, p. 52).Alternatively, if fish were captured, prepared, and con-sumed at a single site, we might expect cranial andpostcranial elements to approximate those proportionspresent in the body. This set of expectations has been testedarcheologically and is supported by data from a faunalsample from at the coastal salt-making site of NorthernRiver Lagoon, Belize (Mock, 1994; Masson, 2004, p. 113),which had a disproportionate quantity of catfish crania, incontrast to other marine fish where cranial and postcranialelements were balanced. A complementary industry in salt-making and salted catfish postcranial ‘‘salt fish’’ produc-tion was identified at Northern River Lagoon. Few inlandsite samples have been examined with the goal of testingthis model of exchange.
(footnote continued)
and the remainder (91%) are from test pits. Only test pit data are used
here.
3.3. Identifying deer management in the archeological
record
Age composition of deer remains is used here in theMayapan samples to investigate the possibility of deerhusbandry or management. In ancient Mesoamerica, deer,unlike dog, were not domesticated, and thus husbanded ormanaged individuals would exhibit no diagnostic morpho-logical differences from wild individuals. The age composi-tion of an archeological faunal assemblage provides theprimary key to identifying whether human managementstrategies were in effect. This approach is commonlyadopted in archeological studies of animal husbandryelsewhere in the world (Ducos, 1978, p. 55; Rackham,1994, pp. 44, 47), and traditionally involves comparing theage profile of an archeological sample of a species to thatexpected from a natural population. Our assessment ofwhite-tailed deer management at Mayapan focuses on oldersubadults; specifically, animals that have reached full size butdo not yet have fully fused long bones (late-fusing elementsthat fuse close to or after sexual maturity). Fusion rates arebased on Reitz andWing (1999, Table 3.5) and Brothwell andHiggs (1963, pp. 252–253, Table A). Ideally, the study of ageprofiles should be performed using complete mandibles todocument tooth eruption and wear. This was not possible dueto the near absence of teeth in the sample. It was also notpossible to ascertain sex at this point in the analysis althoughthis is theoretically possible for husbanded and wild deerpopulations (Rackham, 1994).Techniques of husbandry often include the slaughter of a
high proportion of animals in late subadulthood. At thispoint in the life cycle, animals attain adult size and providethe maximum amount of meat. Butchering animals soonafter they reach full size is an efficient strategy, as keeping
ARTICLE IN PRESSM.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 173
animals longer would involve more feeding but no increasein meat return (Davis, 1987, p. 150). We infer that anexceptionally high proportion of older subadult white-tailed deer (rather than full adults), at Mayapan wouldsuggest that deer were raised and probably bred incaptivity. Alternatively, a sophisticated form of forestgame management was in place.
4. Results
4.1. Identified taxa
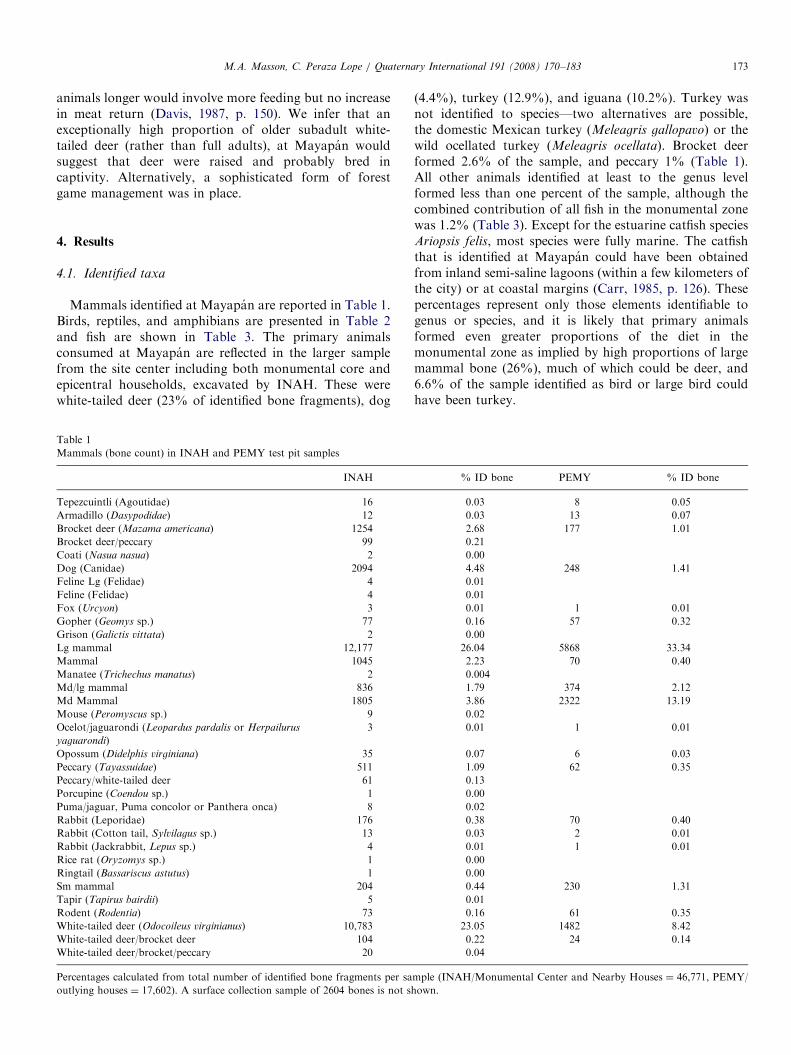
Mammals identified at Mayapan are reported in Table 1.Birds, reptiles, and amphibians are presented in Table 2and fish are shown in Table 3. The primary animalsconsumed at Mayapan are reflected in the larger samplefrom the site center including both monumental core andepicentral households, excavated by INAH. These werewhite-tailed deer (23% of identified bone fragments), dog
Table 1
Mammals (bone count) in INAH and PEMY test pit samples
INAH
Tepezcuintli (Agoutidae) 16
Armadillo (Dasypodidae) 12
Brocket deer (Mazama americana) 1254
Brocket deer/peccary 99
Coati (Nasua nasua) 2
Dog (Canidae) 2094
Feline Lg (Felidae) 4
Feline (Felidae) 4
Fox (Urcyon) 3
Gopher (Geomys sp.) 77
Grison (Galictis vittata) 2
Lg mammal 12,177
Mammal 1045
Manatee (Trichechus manatus) 2
Md/lg mammal 836
Md Mammal 1805
Mouse (Peromyscus sp.) 9
Ocelot/jaguarondi (Leopardus pardalis or Herpailurus
yaguarondi)
3
Opossum (Didelphis virginiana) 35
Peccary (Tayassuidae) 511
Peccary/white-tailed deer 61
Porcupine (Coendou sp.) 1
Puma/jaguar, Puma concolor or Panthera onca) 8
Rabbit (Leporidae) 176
Rabbit (Cotton tail, Sylvilagus sp.) 13
Rabbit (Jackrabbit, Lepus sp.) 4
Rice rat (Oryzomys sp.) 1
Ringtail (Bassariscus astutus) 1
Sm mammal 204
Tapir (Tapirus bairdii) 5
Rodent (Rodentia) 73
White-tailed deer (Odocoileus virginianus) 10,783
White-tailed deer/brocket deer 104
White-tailed deer/brocket/peccary 20
Percentages calculated from total number of identified bone fragments per sa
outlying houses ¼ 17,602). A surface collection sample of 2604 bones is not sh
(4.4%), turkey (12.9%), and iguana (10.2%). Turkey wasnot identified to species—two alternatives are possible,the domestic Mexican turkey (Meleagris gallopavo) or thewild ocellated turkey (Meleagris ocellata). Brocket deerformed 2.6% of the sample, and peccary 1% (Table 1).All other animals identified at least to the genus levelformed less than one percent of the sample, although thecombined contribution of all fish in the monumental zonewas 1.2% (Table 3). Except for the estuarine catfish speciesAriopsis felis, most species were fully marine. The catfishthat is identified at Mayapan could have been obtainedfrom inland semi-saline lagoons (within a few kilometers ofthe city) or at coastal margins (Carr, 1985, p. 126). Thesepercentages represent only those elements identifiable togenus or species, and it is likely that primary animalsformed even greater proportions of the diet in themonumental zone as implied by high proportions of largemammal bone (26%), much of which could be deer, and6.6% of the sample identified as bird or large bird couldhave been turkey.
% ID bone PEMY % ID bone
0.03 8 0.05
0.03 13 0.07
2.68 177 1.01
0.21
0.00
4.48 248 1.41
0.01
0.01
0.01 1 0.01
0.16 57 0.32
0.00
26.04 5868 33.34
2.23 70 0.40
0.004
1.79 374 2.12
3.86 2322 13.19
0.02
0.01 1 0.01
0.07 6 0.03
1.09 62 0.35
0.13
0.00
0.02
0.38 70 0.40
0.03 2 0.01
0.01 1 0.01
0.00
0.00
0.44 230 1.31
0.01
0.16 61 0.35
23.05 1482 8.42
0.22 24 0.14
0.04
mple (INAH/Monumental Center and Nearby Houses ¼ 46,771, PEMY/
own.
ARTICLE IN PRESS
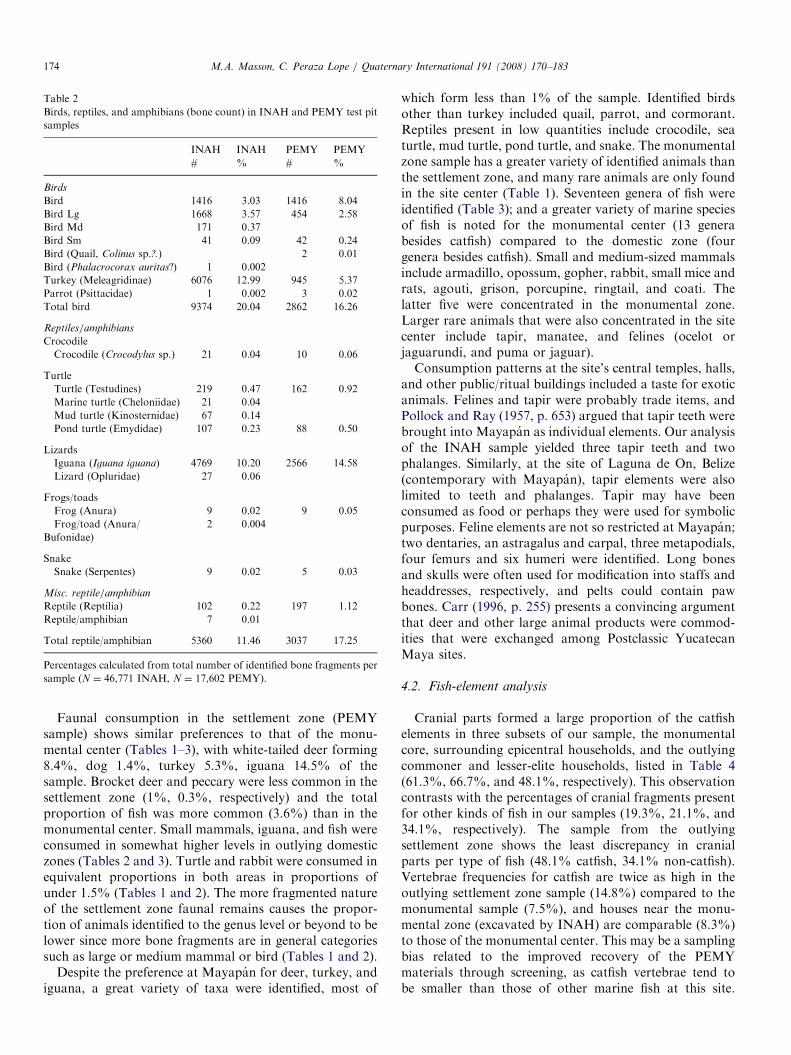
Table 2
Birds, reptiles, and amphibians (bone count) in INAH and PEMY test pit
samples
INAH
#
INAH
%
PEMY
#
PEMY
%
Birds
Bird 1416 3.03 1416 8.04
Bird Lg 1668 3.57 454 2.58
Bird Md 171 0.37
Bird Sm 41 0.09 42 0.24
Bird (Quail, Colinus sp.?.) 2 0.01
Bird (Phalacrocorax auritas?) 1 0.002
Turkey (Meleagridinae) 6076 12.99 945 5.37
Parrot (Psittacidae) 1 0.002 3 0.02
Total bird 9374 20.04 2862 16.26
Reptiles/amphibians
Crocodile
Crocodile (Crocodylus sp.) 21 0.04 10 0.06
Turtle
Turtle (Testudines) 219 0.47 162 0.92
Marine turtle (Cheloniidae) 21 0.04
Mud turtle (Kinosternidae) 67 0.14
Pond turtle (Emydidae) 107 0.23 88 0.50
Lizards
Iguana (Iguana iguana) 4769 10.20 2566 14.58
Lizard (Opluridae) 27 0.06
Frogs/toads
Frog (Anura) 9 0.02 9 0.05
Frog/toad (Anura/
Bufonidae)
2 0.004
Snake
Snake (Serpentes) 9 0.02 5 0.03
Misc. reptile/amphibian
Reptile (Reptilia) 102 0.22 197 1.12
Reptile/amphibian 7 0.01
Total reptile/amphibian 5360 11.46 3037 17.25
Percentages calculated from total number of identified bone fragments per
sample (N ¼ 46,771 INAH, N ¼ 17,602 PEMY).
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183174
Faunal consumption in the settlement zone (PEMYsample) shows similar preferences to that of the monu-mental center (Tables 1–3), with white-tailed deer forming8.4%, dog 1.4%, turkey 5.3%, iguana 14.5% of thesample. Brocket deer and peccary were less common in thesettlement zone (1%, 0.3%, respectively) and the totalproportion of fish was more common (3.6%) than in themonumental center. Small mammals, iguana, and fish wereconsumed in somewhat higher levels in outlying domesticzones (Tables 2 and 3). Turtle and rabbit were consumed inequivalent proportions in both areas in proportions ofunder 1.5% (Tables 1 and 2). The more fragmented natureof the settlement zone faunal remains causes the propor-tion of animals identified to the genus level or beyond to belower since more bone fragments are in general categoriessuch as large or medium mammal or bird (Tables 1 and 2).
Despite the preference at Mayapan for deer, turkey, andiguana, a great variety of taxa were identified, most of
which form less than 1% of the sample. Identified birdsother than turkey included quail, parrot, and cormorant.Reptiles present in low quantities include crocodile, seaturtle, mud turtle, pond turtle, and snake. The monumentalzone sample has a greater variety of identified animals thanthe settlement zone, and many rare animals are only foundin the site center (Table 1). Seventeen genera of fish wereidentified (Table 3); and a greater variety of marine speciesof fish is noted for the monumental center (13 generabesides catfish) compared to the domestic zone (fourgenera besides catfish). Small and medium-sized mammalsinclude armadillo, opossum, gopher, rabbit, small mice andrats, agouti, grison, porcupine, ringtail, and coati. Thelatter five were concentrated in the monumental zone.Larger rare animals that were also concentrated in the sitecenter include tapir, manatee, and felines (ocelot orjaguarundi, and puma or jaguar).Consumption patterns at the site’s central temples, halls,
and other public/ritual buildings included a taste for exoticanimals. Felines and tapir were probably trade items, andPollock and Ray (1957, p. 653) argued that tapir teeth werebrought into Mayapan as individual elements. Our analysisof the INAH sample yielded three tapir teeth and twophalanges. Similarly, at the site of Laguna de On, Belize(contemporary with Mayapan), tapir elements were alsolimited to teeth and phalanges. Tapir may have beenconsumed as food or perhaps they were used for symbolicpurposes. Feline elements are not so restricted at Mayapan;two dentaries, an astragalus and carpal, three metapodials,four femurs and six humeri were identified. Long bonesand skulls were often used for modification into staffs andheaddresses, respectively, and pelts could contain pawbones. Carr (1996, p. 255) presents a convincing argumentthat deer and other large animal products were commod-ities that were exchanged among Postclassic YucatecanMaya sites.
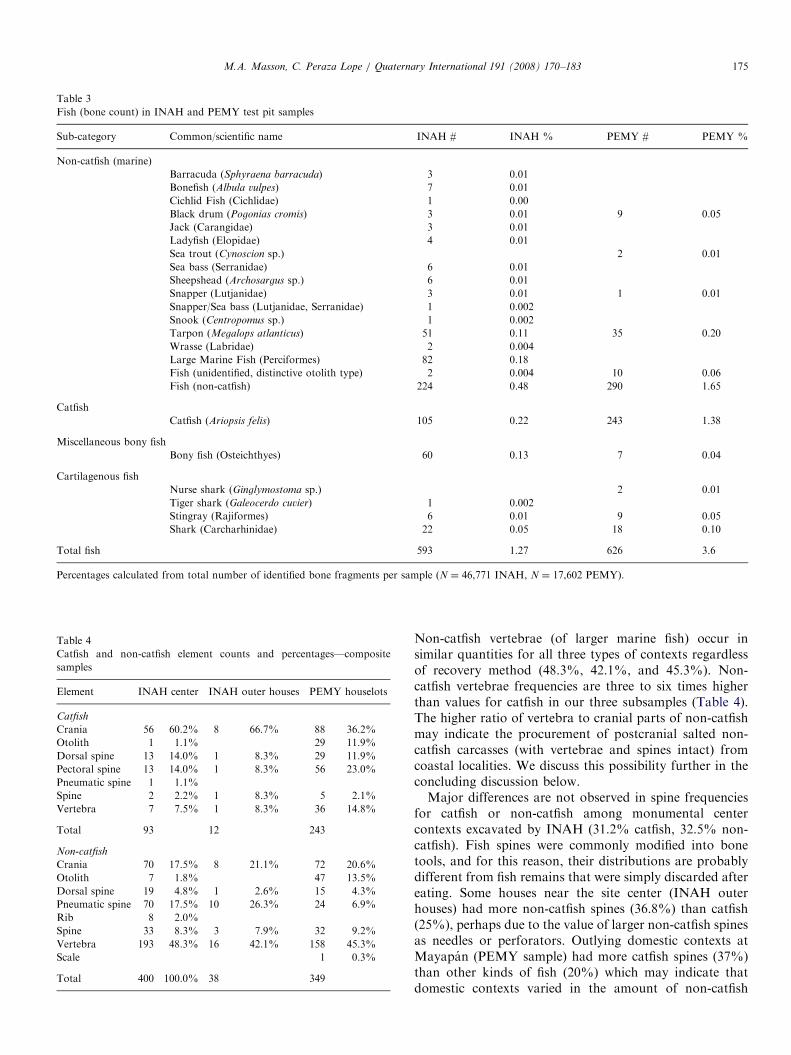
4.2. Fish-element analysis
Cranial parts formed a large proportion of the catfishelements in three subsets of our sample, the monumentalcore, surrounding epicentral households, and the outlyingcommoner and lesser-elite households, listed in Table 4(61.3%, 66.7%, and 48.1%, respectively). This observationcontrasts with the percentages of cranial fragments presentfor other kinds of fish in our samples (19.3%, 21.1%, and34.1%, respectively). The sample from the outlyingsettlement zone shows the least discrepancy in cranialparts per type of fish (48.1% catfish, 34.1% non-catfish).Vertebrae frequencies for catfish are twice as high in theoutlying settlement zone sample (14.8%) compared to themonumental sample (7.5%), and houses near the monu-mental zone (excavated by INAH) are comparable (8.3%)to those of the monumental center. This may be a samplingbias related to the improved recovery of the PEMYmaterials through screening, as catfish vertebrae tend tobe smaller than those of other marine fish at this site.
ARTICLE IN PRESS
Table 3
Fish (bone count) in INAH and PEMY test pit samples
Sub-category Common/scientific name INAH # INAH % PEMY # PEMY %
Non-catfish (marine)
Barracuda (Sphyraena barracuda) 3 0.01
Bonefish (Albula vulpes) 7 0.01
Cichlid Fish (Cichlidae) 1 0.00
Black drum (Pogonias cromis) 3 0.01 9 0.05
Jack (Carangidae) 3 0.01
Ladyfish (Elopidae) 4 0.01
Sea trout (Cynoscion sp.) 2 0.01
Sea bass (Serranidae) 6 0.01
Sheepshead (Archosargus sp.) 6 0.01
Snapper (Lutjanidae) 3 0.01 1 0.01
Snapper/Sea bass (Lutjanidae, Serranidae) 1 0.002
Snook (Centropomus sp.) 1 0.002
Tarpon (Megalops atlanticus) 51 0.11 35 0.20
Wrasse (Labridae) 2 0.004
Large Marine Fish (Perciformes) 82 0.18
Fish (unidentified, distinctive otolith type) 2 0.004 10 0.06
Fish (non-catfish) 224 0.48 290 1.65
Catfish
Catfish (Ariopsis felis) 105 0.22 243 1.38
Miscellaneous bony fish
Bony fish (Osteichthyes) 60 0.13 7 0.04
Cartilagenous fish
Nurse shark (Ginglymostoma sp.) 2 0.01
Tiger shark (Galeocerdo cuvier) 1 0.002
Stingray (Rajiformes) 6 0.01 9 0.05
Shark (Carcharhinidae) 22 0.05 18 0.10
Total fish 593 1.27 626 3.6
Percentages calculated from total number of identified bone fragments per sample (N ¼ 46,771 INAH, N ¼ 17,602 PEMY).
Table 4
Catfish and non-catfish element counts and percentages—composite
samples
Element INAH center INAH outer houses PEMY houselots
Catfish
Crania 56 60.2% 8 66.7% 88 36.2%
Otolith 1 1.1% 29 11.9%
Dorsal spine 13 14.0% 1 8.3% 29 11.9%
Pectoral spine 13 14.0% 1 8.3% 56 23.0%
Pneumatic spine 1 1.1%
Spine 2 2.2% 1 8.3% 5 2.1%
Vertebra 7 7.5% 1 8.3% 36 14.8%
Total 93 12 243
Non-catfish
Crania 70 17.5% 8 21.1% 72 20.6%
Otolith 7 1.8% 47 13.5%
Dorsal spine 19 4.8% 1 2.6% 15 4.3%
Pneumatic spine 70 17.5% 10 26.3% 24 6.9%
Rib 8 2.0%
Spine 33 8.3% 3 7.9% 32 9.2%
Vertebra 193 48.3% 16 42.1% 158 45.3%
Scale 1 0.3%
Total 400 100.0% 38 349
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 175
Non-catfish vertebrae (of larger marine fish) occur insimilar quantities for all three types of contexts regardlessof recovery method (48.3%, 42.1%, and 45.3%). Non-catfish vertebrae frequencies are three to six times higherthan values for catfish in our three subsamples (Table 4).The higher ratio of vertebra to cranial parts of non-catfishmay indicate the procurement of postcranial salted non-catfish carcasses (with vertebrae and spines intact) fromcoastal localities. We discuss this possibility further in theconcluding discussion below.Major differences are not observed in spine frequencies
for catfish or non-catfish among monumental centercontexts excavated by INAH (31.2% catfish, 32.5% non-catfish). Fish spines were commonly modified into bonetools, and for this reason, their distributions are probablydifferent from fish remains that were simply discarded aftereating. Some houses near the site center (INAH outerhouses) had more non-catfish spines (36.8%) than catfish(25%), perhaps due to the value of larger non-catfish spinesas needles or perforators. Outlying domestic contexts atMayapan (PEMY sample) had more catfish spines (37%)than other kinds of fish (20%) which may indicate thatdomestic contexts varied in the amount of non-catfish
ARTICLE IN PRESSM.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183176
spines they were able to obtain. Alternatively, screening ofPEMY contexts may have improved the recovery of catfishspines.
4.3. Deer skull vs. postcranial elements
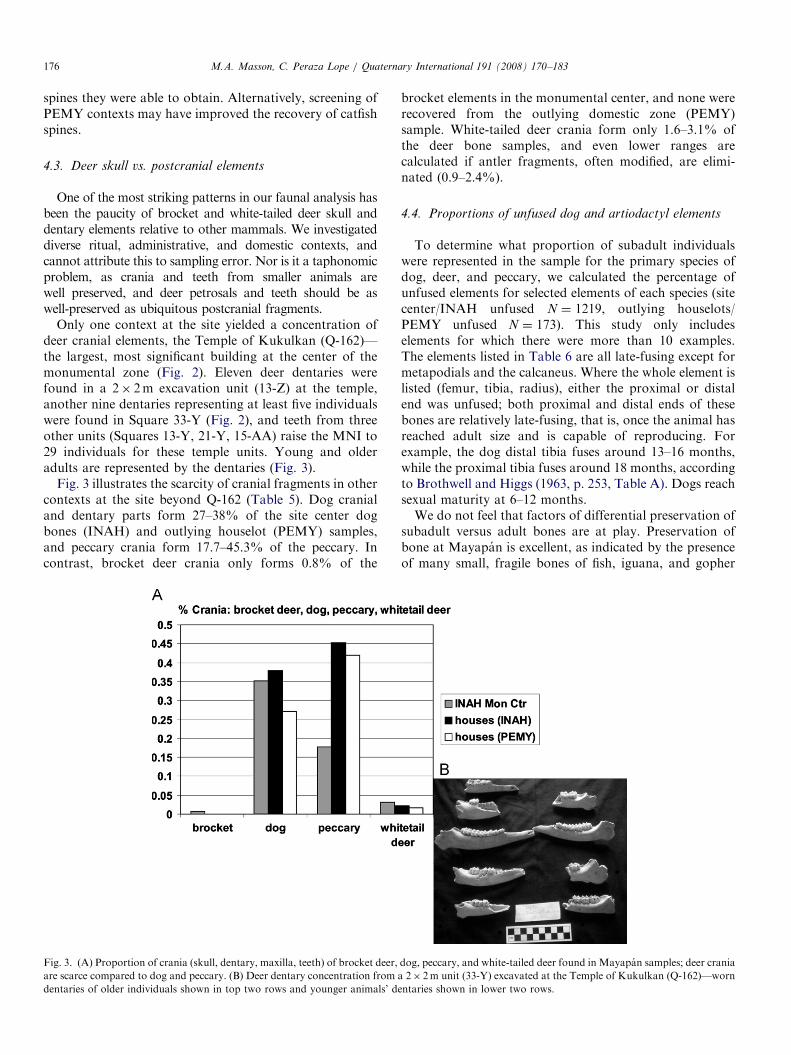
One of the most striking patterns in our faunal analysis hasbeen the paucity of brocket and white-tailed deer skull anddentary elements relative to other mammals. We investigateddiverse ritual, administrative, and domestic contexts, andcannot attribute this to sampling error. Nor is it a taphonomicproblem, as crania and teeth from smaller animals arewell preserved, and deer petrosals and teeth should be aswell-preserved as ubiquitous postcranial fragments.
Only one context at the site yielded a concentration ofdeer cranial elements, the Temple of Kukulkan (Q-162)—the largest, most significant building at the center of themonumental zone (Fig. 2). Eleven deer dentaries werefound in a 2� 2m excavation unit (13-Z) at the temple,another nine dentaries representing at least five individualswere found in Square 33-Y (Fig. 2), and teeth from threeother units (Squares 13-Y, 21-Y, 15-AA) raise the MNI to29 individuals for these temple units. Young and olderadults are represented by the dentaries (Fig. 3).
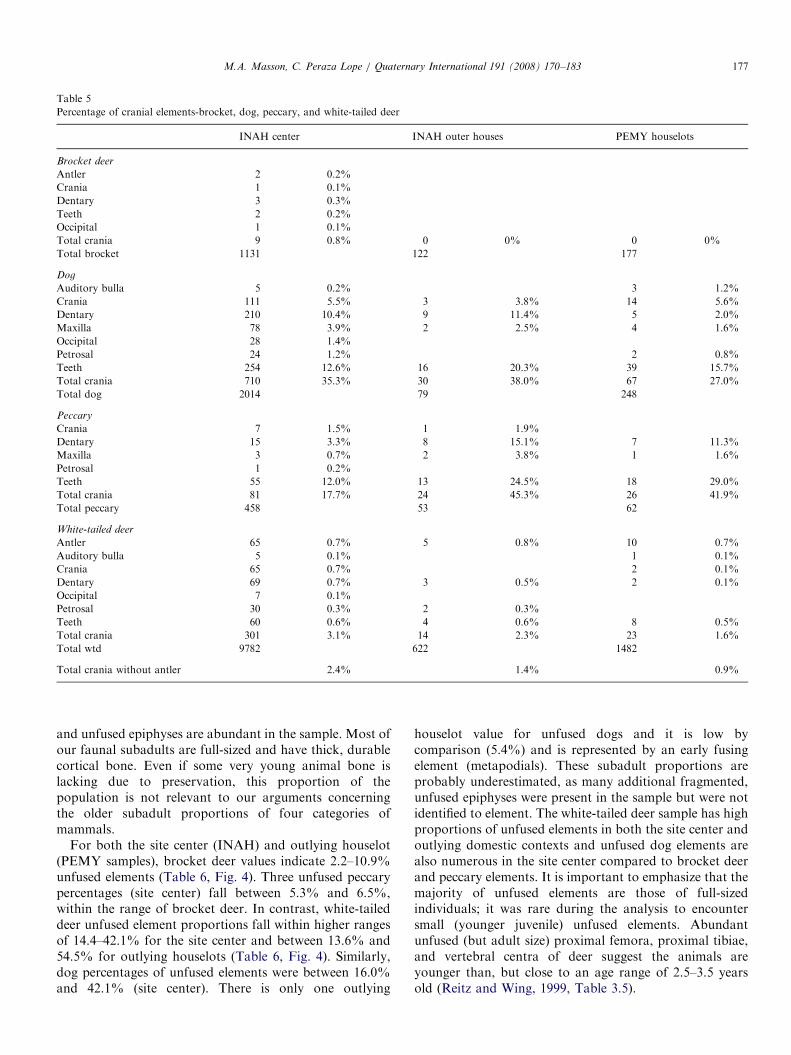
Fig. 3 illustrates the scarcity of cranial fragments in othercontexts at the site beyond Q-162 (Table 5). Dog cranialand dentary parts form 27–38% of the site center dogbones (INAH) and outlying houselot (PEMY) samples,and peccary crania form 17.7–45.3% of the peccary. Incontrast, brocket deer crania only forms 0.8% of the
Fig. 3. (A) Proportion of crania (skull, dentary, maxilla, teeth) of brocket deer,
are scarce compared to dog and peccary. (B) Deer dentary concentration from
dentaries of older individuals shown in top two rows and younger animals’ de
brocket elements in the monumental center, and none wererecovered from the outlying domestic zone (PEMY)sample. White-tailed deer crania form only 1.6–3.1% ofthe deer bone samples, and even lower ranges arecalculated if antler fragments, often modified, are elimi-nated (0.9–2.4%).
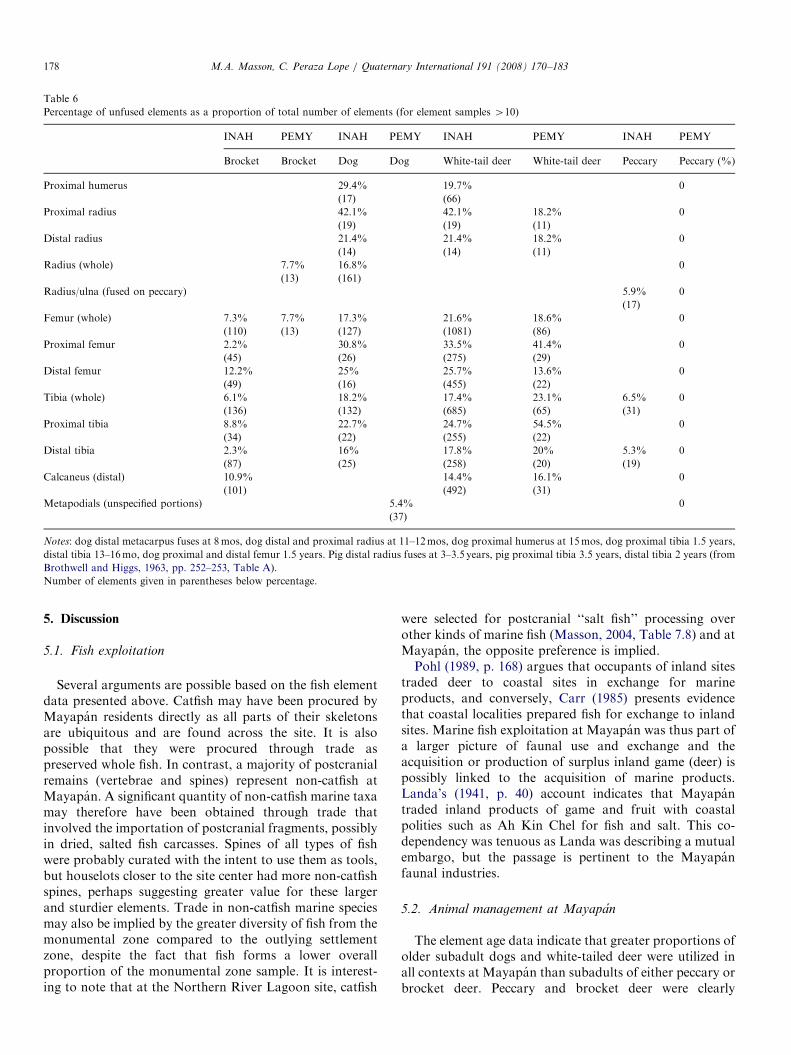
4.4. Proportions of unfused dog and artiodactyl elements
To determine what proportion of subadult individualswere represented in the sample for the primary species ofdog, deer, and peccary, we calculated the percentage ofunfused elements for selected elements of each species (sitecenter/INAH unfused N ¼ 1219, outlying houselots/PEMY unfused N ¼ 173). This study only includeselements for which there were more than 10 examples.The elements listed in Table 6 are all late-fusing except formetapodials and the calcaneus. Where the whole element islisted (femur, tibia, radius), either the proximal or distalend was unfused; both proximal and distal ends of thesebones are relatively late-fusing, that is, once the animal hasreached adult size and is capable of reproducing. Forexample, the dog distal tibia fuses around 13–16 months,while the proximal tibia fuses around 18 months, accordingto Brothwell and Higgs (1963, p. 253, Table A). Dogs reachsexual maturity at 6–12 months.We do not feel that factors of differential preservation of
subadult versus adult bones are at play. Preservation ofbone at Mayapan is excellent, as indicated by the presenceof many small, fragile bones of fish, iguana, and gopher
dog, peccary, and white-tailed deer found in Mayapan samples; deer crania
a 2� 2m unit (33-Y) excavated at the Temple of Kukulkan (Q-162)—worn
ntaries shown in lower two rows.
ARTICLE IN PRESS
Table 5
Percentage of cranial elements-brocket, dog, peccary, and white-tailed deer
INAH center INAH outer houses PEMY houselots
Brocket deer
Antler 2 0.2%
Crania 1 0.1%
Dentary 3 0.3%
Teeth 2 0.2%
Occipital 1 0.1%
Total crania 9 0.8% 0 0% 0 0%
Total brocket 1131 122 177
Dog
Auditory bulla 5 0.2% 3 1.2%
Crania 111 5.5% 3 3.8% 14 5.6%
Dentary 210 10.4% 9 11.4% 5 2.0%
Maxilla 78 3.9% 2 2.5% 4 1.6%
Occipital 28 1.4%
Petrosal 24 1.2% 2 0.8%
Teeth 254 12.6% 16 20.3% 39 15.7%
Total crania 710 35.3% 30 38.0% 67 27.0%
Total dog 2014 79 248
Peccary
Crania 7 1.5% 1 1.9%
Dentary 15 3.3% 8 15.1% 7 11.3%
Maxilla 3 0.7% 2 3.8% 1 1.6%
Petrosal 1 0.2%
Teeth 55 12.0% 13 24.5% 18 29.0%
Total crania 81 17.7% 24 45.3% 26 41.9%
Total peccary 458 53 62
White-tailed deer
Antler 65 0.7% 5 0.8% 10 0.7%
Auditory bulla 5 0.1% 1 0.1%
Crania 65 0.7% 2 0.1%
Dentary 69 0.7% 3 0.5% 2 0.1%
Occipital 7 0.1%
Petrosal 30 0.3% 2 0.3%
Teeth 60 0.6% 4 0.6% 8 0.5%
Total crania 301 3.1% 14 2.3% 23 1.6%
Total wtd 9782 622 1482
Total crania without antler 2.4% 1.4% 0.9%
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 177
and unfused epiphyses are abundant in the sample. Most ofour faunal subadults are full-sized and have thick, durablecortical bone. Even if some very young animal bone islacking due to preservation, this proportion of thepopulation is not relevant to our arguments concerningthe older subadult proportions of four categories ofmammals.
For both the site center (INAH) and outlying houselot(PEMY samples), brocket deer values indicate 2.2–10.9%unfused elements (Table 6, Fig. 4). Three unfused peccarypercentages (site center) fall between 5.3% and 6.5%,within the range of brocket deer. In contrast, white-taileddeer unfused element proportions fall within higher rangesof 14.4–42.1% for the site center and between 13.6% and54.5% for outlying houselots (Table 6, Fig. 4). Similarly,dog percentages of unfused elements were between 16.0%and 42.1% (site center). There is only one outlying
houselot value for unfused dogs and it is low bycomparison (5.4%) and is represented by an early fusingelement (metapodials). These subadult proportions areprobably underestimated, as many additional fragmented,unfused epiphyses were present in the sample but were notidentified to element. The white-tailed deer sample has highproportions of unfused elements in both the site center andoutlying domestic contexts and unfused dog elements arealso numerous in the site center compared to brocket deerand peccary elements. It is important to emphasize that themajority of unfused elements are those of full-sizedindividuals; it was rare during the analysis to encountersmall (younger juvenile) unfused elements. Abundantunfused (but adult size) proximal femora, proximal tibiae,and vertebral centra of deer suggest the animals areyounger than, but close to an age range of 2.5–3.5 yearsold (Reitz and Wing, 1999, Table 3.5).
ARTICLE IN PRESS
Table 6
Percentage of unfused elements as a proportion of total number of elements (for element samples 410)
INAH PEMY INAH PEMY INAH PEMY INAH PEMY
Brocket Brocket Dog Dog White-tail deer White-tail deer Peccary Peccary (%)
Proximal humerus 29.4% 19.7% 0
(17) (66)
Proximal radius 42.1% 42.1% 18.2% 0
(19) (19) (11)
Distal radius 21.4% 21.4% 18.2% 0
(14) (14) (11)
Radius (whole) 7.7% 16.8% 0
(13) (161)
Radius/ulna (fused on peccary) 5.9% 0
(17)
Femur (whole) 7.3% 7.7% 17.3% 21.6% 18.6% 0
(110) (13) (127) (1081) (86)
Proximal femur 2.2% 30.8% 33.5% 41.4% 0
(45) (26) (275) (29)
Distal femur 12.2% 25% 25.7% 13.6% 0
(49) (16) (455) (22)
Tibia (whole) 6.1% 18.2% 17.4% 23.1% 6.5% 0
(136) (132) (685) (65) (31)
Proximal tibia 8.8% 22.7% 24.7% 54.5% 0
(34) (22) (255) (22)
Distal tibia 2.3% 16% 17.8% 20% 5.3% 0
(87) (25) (258) (20) (19)
Calcaneus (distal) 10.9% 14.4% 16.1% 0
(101) (492) (31)
Metapodials (unspecified portions) 5.4% 0
(37)
Notes: dog distal metacarpus fuses at 8mos, dog distal and proximal radius at 11–12mos, dog proximal humerus at 15mos, dog proximal tibia 1.5 years,
distal tibia 13–16mo, dog proximal and distal femur 1.5 years. Pig distal radius fuses at 3–3.5 years, pig proximal tibia 3.5 years, distal tibia 2 years (from
Brothwell and Higgs, 1963, pp. 252–253, Table A).
Number of elements given in parentheses below percentage.
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183178
5. Discussion
5.1. Fish exploitation
Several arguments are possible based on the fish elementdata presented above. Catfish may have been procured byMayapan residents directly as all parts of their skeletonsare ubiquitous and are found across the site. It is alsopossible that they were procured through trade aspreserved whole fish. In contrast, a majority of postcranialremains (vertebrae and spines) represent non-catfish atMayapan. A significant quantity of non-catfish marine taxamay therefore have been obtained through trade thatinvolved the importation of postcranial fragments, possiblyin dried, salted fish carcasses. Spines of all types of fishwere probably curated with the intent to use them as tools,but houselots closer to the site center had more non-catfishspines, perhaps suggesting greater value for these largerand sturdier elements. Trade in non-catfish marine speciesmay also be implied by the greater diversity of fish from themonumental zone compared to the outlying settlementzone, despite the fact that fish forms a lower overallproportion of the monumental zone sample. It is interest-ing to note that at the Northern River Lagoon site, catfish
were selected for postcranial ‘‘salt fish’’ processing overother kinds of marine fish (Masson, 2004, Table 7.8) and atMayapan, the opposite preference is implied.Pohl (1989, p. 168) argues that occupants of inland sites
traded deer to coastal sites in exchange for marineproducts, and conversely, Carr (1985) presents evidencethat coastal localities prepared fish for exchange to inlandsites. Marine fish exploitation at Mayapan was thus part ofa larger picture of faunal use and exchange and theacquisition or production of surplus inland game (deer) ispossibly linked to the acquisition of marine products.Landa’s (1941, p. 40) account indicates that Mayapantraded inland products of game and fruit with coastalpolities such as Ah Kin Chel for fish and salt. This co-dependency was tenuous as Landa was describing a mutualembargo, but the passage is pertinent to the Mayapanfaunal industries.
5.2. Animal management at Mayapan
The element age data indicate that greater proportions ofolder subadult dogs and white-tailed deer were utilized inall contexts at Mayapan than subadults of either peccary orbrocket deer. Peccary and brocket deer were clearly

ARTICLE IN PRESS
Fig. 4. The upper graph (A) illustrates that the proportion of juvenile elements for brocket deer and peccary is much lower than the values for white-tailed
deer and dog shown in the lower graph (B). This pattern suggests that white-tailed deer and dog were commonly selected for butchering and consumption
at a young age.
M.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 179
acquired and utilized differently than white-tails and dogs.Pollock and Ray (1957, p. 653) were the first to identify theimportance of dogs in activities of Mayapan’s site center,and the city is hardly unique in this regard. Older subadultdogs (19%) were common in ritual contexts at PostclassicCozumel Island (Hamblin, 1984, Table 7.9), where dog wasmost often used for sacrifice and other rituals (Hamblin,1984, pp. 116–117). Dogs were an important resourceduring the Late Preclassic period at Colha (Shaw, 1991,1999, p. 94), Cerros (Carr, 1985, p. 131), and otherPreclassic settlements (Pohl, 1985b, p. 109).
The high proportions of older (full-size) subadult white-tailed deer in both the PEMY and INAH samples suggeststhe practice of animal husbandry, or alternatively, asophisticated system of game management in forestscontrolled by the city. This pattern suggests an efficientstrategy of game production in which deer and dog wereallowed to grow until they reached full size before theywere consumed, which would have maximized the meatyield from the animals and minimized the time invested infeeding them. The fact that the proportions of oldersubadult white-tailed deer resemble the proportions ofsubadult dog, a known domesticate, and do not resemble
the proportions of either brocket deer or peccary strength-ens the argument that white-tailed deer, like dog, were bredand raised in captivity or were carefully harvested atMayapan. These data independently support Landa’s(1941, p. 127) statement that deer were raised by womenin Contact period Yucatan.The Mayapan data match a key criterion of animal
management—the age structure is different from a normalpopulation (Davis, 1987, p. 150). In Old World cases whereanimal management is suspected, ‘‘many animals are killedwhen fairly large but still immature’’ (Rackham, 1994,p. 46, also, Carr, 1996, p. 256).As initial claims for animal husbandry at Mayapan made
by the authors in earlier versions of this paper were metwith sharp skepticism by reviewers, it is helpful to eliminatealternative explanations and list additional corroboratingdata regarding the prevalence of older subadults in theMayapan deer sample. First, immature deer were notsimply preferred by site center elites as they also occur inlarge proportions in outlying houselots. Second, the highproportion of subadults at Mayapan is unlikely due toselective hunting practices as there is no ethnographicevidence that fully grown adults were ever excluded as a

ARTICLE IN PRESSM.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183180
food source in Mesoamerican deer hunts. Third, infra-structural features suitable for game confinement arepresent at Mayapan in the form of pens and corrals thatare located adjacent to domestic houselot walls (Fig. 1).Similar, smaller features at Cozumel were used as birdpens, according to soil test results (Hamblin, 1985, p. 188).The larger enclosures at Mayapan could have corralledturkeys, peccaries, dogs, or tethered deer. A fifth alter-native explanation for Mayapan’s abundant young deer,high levels of predation, merits further consideration.
Unmanaged high levels of predation can have the effectof reducing the age structure of a population, and shortlythereafter, the availability of the population. There is noevidence for depletion of deer at Mayapan, where it ispresent in great abundance throughout the city’s occupa-tion. However, if game reserves were carefully managed inMayapan’s territory, older subadults could have beenallowed to reproduce prior to being hunted—such astrategy would have replenished the population but wouldhave inhibited the numbers of older adults in the sample.Although this alternative may seem far-fetched, both gamereserves (Pohl, 1989, pp. 153–154) and selective hunting(Teeter and Chase, 2004, p. 165) have been proposed forthe Maya sites of Seibal and Caracol. As modern hunters inthe US sometimes kill large proportions of older subadultsin a managed setting (Cahalane, 1931), the idea of gamereserves and high, but balanced predation managementmay be worth considering for sites like Mayapan. Mann(2005, pp. 273–274, 348–350) eloquently summarizes theextensive new evidence for the management of anthropo-genic landscapes and their botanical and faunal resourcesby indigenous populations across the New World. TheMaya were likely no exception (e.g., Fedick, 1996). Morework is needed to determine whether forest game manage-ment or domestic houselot husbandry accounts for the agestructure of Mayapan’s deer.
High proportions of subadult deer are known from otherMaya sites, which may suggest that deer management (andpossibly husbandry) was practiced at other settlements.Late Classic Altar de Sacrificios had 36% juveniles, LateClassic/Early Postclassic Macanche had 33%, and Post-classic Flores had 33%, according to Pohl (1989, Table 3).It is not known whether these proportions were primarilyolder subadults or whether they represent juveniles ofyounger ages as well. Caracol is another site with a highproportion of subadults—Teeter and Chase (2004, p. 165)report that 84% of the individuals for which age could bedetermined (only 15% of the total sample of deer) were notfully mature.
In the southern Maya lowlands, studies of deer skeletalisotope samples from the Preclassic and Classic periodsshow no evidence of intentional corn-feeding of deer, eitherwild, tame, free-ranging, or houselot-raised (Emery et al.,2000, p. 545; White et al., 2001). Emery et al. (2000, p. 546)report one example of a probable tamed deer (with a healedinjury and found as part of a cache) that did not show anyevidence of maize feeding, suggesting that even tamed deer
were likely fed wild fodder in addition to maize by theirkeepers. Isotopic studies are currently being conducted onMayapan deer (by Lori Wright) that may help to determinetheir maize consumption levels and dependency on foodprovided by humans—patterns may be quite different inthe Postclassic northern lowlands from those of earlierperiods.Despite the fact that archeological evidence for raising or
breeding deer in captivity is scarce, documentary accountsare explicit on this point (Ximenez, 1967, p. 57; Pohl andFeldman, 1982, p. 295; Pohl, 1985a, p. 143; Carr, 1996,p. 251). Pohl (1985a, p. 140) observes that the Maya had aterm for a deer that was raised in a dwelling (ah may),according to the Motul dictionary. Perhaps this practicewas more characteristic in areas under direct Spanishobservation, i.e., northwest Yucatan in which Mayapan issituated. A striking passage in Landa’s (1941, p. 127)account claims that the women of 16th century Yucatan‘‘ylet the deer suck their breasts, by which means theyraise them and make them so tame that they never will gointo the woodsy’’. This quote is perhaps an overlymetaphoric description of the practice of deer-raising.Comparative data are needed from consumer sites that
were directly engaged in exchange with Mayapan. ThePostclassic Maya sites of Cozumel Island are contemporarywith Mayapan and have been the target of an exhaustivefaunal study (Hamblin, 1984). Fish were a staple food,and peccary was abundant on Cozumel (Hamblin, 1984,Table 9.3). These plentiful animals at Cozumel are rare atMayapan and Mayapan’s abundant deer was scarce atCozumel. Whether or not this site traded animal resourcesdirectly with Mayapan, the patterns there do supportcoastal-inland system of animal exchange (Hamblin, 1984,pp. 138, 141).
5.3. Deer Crania (lack thereof)
The paucity of crania elements of white-tailed deersuggests that specialized processing and disposal occurred.The deer skull proportions for Mayapan are low by anystandards. Skull proportions reported at other Maya sitesby Carr (1996, Table 15.1) are higher, around 14% atCozumel and Dzibilchaltun, 5% at Isla Cerritos where deerare thought to have been imported, and higher at ChichenItza (38%). If antler frequencies are included in theproportion of skull fragments from these other sites, theyare even higher: 46% for Chichen Itza, 35% for Cozumel,15% for Dzibilchaltun, and 19% for Isla Cerritos. Clearly,Mayapan’s proportion of white-tail deer skull, even withantler, at 1.6–3.1%, is the lowest of all reported sites.The lack of deer skulls in most monumental and
domestic contexts suggests that commoner and nobleconsumers obtained or processed their deer through similarmechanisms. Deer skulls, including those of brocket andwhite-tail, were likely disposed of in culturally prescribedways that resulted in their removal from almost all of ourcontexts. Dentaries were were placed at the Temple of

ARTICLE IN PRESSM.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 181
Kukulkan (Q-162) for specific ceremonies. However, theamount of deer at this temple (29 individuals) does notaccount for all of the missing deer crania from Mayapan,as the MNI for the combined INAH-PEMY samples was241 white-tailed deer.2
The dentaries at Q-162 were placed in special locations.Quadripartite (four staircase) temples like this structure aresignificant markers for central, sacred points on thelandscape at Maya sites (Fox, 1987; Freidel et al., 1993),including Mayapan (Pugh, 2001). The squares with dentaryconcentrations are located along an east-west axis thatcrosses the south base of the temple (Fig. 2); units 13-Z/13-Y/15-AA and 33-Y are in the vicinity of the southeast andsouthwest corners of the pyramid, suggesting the boneswere part of a dedicatory offering linked to establishing (orre-establishing) the temple’s status as a sacred place.Square 21-Y is at the base of the temple’s south staircase,which is also a meaningful location for dedicationofferings.
There are probably other specific contexts, perhaps inthe forest or in outlying caves or cenotes, where deer craniabones might be found. For example, Maya hunters in theGuatemalan highlands currently dispose of animal skulls incaves (Brown, 2005). Pohl (1985a, p. 136) cites an accountof a scientist (A.W. Anthony, quoted in Goodwin, 1934, p. 2)who observed small thatch-enclosed altars in Guatemalawhere animal skulls had been deposited and blackened byritual fires for many years. These small structures werelocated near corn fields and away from settlement zones,and such features may exist for Mayapan. Like the Castilloat Mayapan, a cache at Cuello also consisted of deerdentaries (Pohl, 1985a, p. 140), many of which weresubadults. At Laguna de On, a contemporary of Mayapanlocated in northeastern Belize, skulls of large game (deer,peccary, crocodile, and tapir) were disposed of at a singlebuilding, along with incense burners, as part of a ritual(Masson, 1999). The deer skulls found at the Temple ofKukulkan may have been deposited in the context ofperiod ending rituals, perhaps dedication or terminationevents associated with calendrical celebrations as proposedfor Laguna de On (Masson, 1999). Skulls may also havebeen a commodity for exchange. Deer metapodials andantlers were imported to coastal sites for bone toolmanufacture (Carr, 1996, p. 255) and Postclassic Mayamurals at the site site of Santa Rita Corozal, Belizeillustrate a jaguar and a peccary skull in the headdresses oftwo individuals (Masson, 2000, Figs. 6.16, #7, 6.18, #18).
6. Conclusions
The evidence described above suggests that the produc-tion and exchange of animal commodities within andbeyond the city of Mayapan was an integral component of
2MNI based on 18L distal humeri for the PEMY sample, 18 proximal R
tibia for the INAH-house sample, and 205L astragali and 205L calcanei
for the INAH-center sample.
the city’s economy. Surplus meat and bone products mayhave been highly significant as one of the few subsistenceproducts that Mayapan merchants offered to coastalpolities from whom they obtained salt for more distantexchange voyages (Masson et al., n.d.). The likelihood thatmarine fish other than catfish were obtained at the sitethrough trade is implied by lower proportions of cranialelements of these taxa and a diverse list of species from thesite center. We argue that white-tailed deer-raising ormanagement was a major production industry for the sitebased on the high proportions of full-sized subadults—these deer provided a staple food source for the city’sresidents and surplus meat and bone products forexchange. Ethnohistoric evidence suggests that womenwere heavily involved in many aspects of animal exchange,including selling dogs in the marketplace, providing birdsfor tribute, and delivering peccaries and deer destined forsacrifice (Pohl and Feldman, 1982, pp. 295, 305). Deer-raising/management at Mayapan was probably the respon-sibility of women (Landa, 1941, p. 127).The selective disposal of deer crania implied by the
paucity of such elements at all contexts other than theTemple of Kukulkan attests to another probable dimen-sion of ritual practice involving animal resources. Ethno-historic documents suggest that consumption activities inthe political center of the site included animal sacrifices andmeals associated with political and ritual gatherings(Landa, 1941, pp. 92, 106, 141, 158); these claims aresometimes supported in the archeological record ofMayapan and a number of other Postclassic Maya sites(Masson, 1999).Our initial analyses of Mayapan faunal remains reflect
that fact that animal use was incorporated into multipledimensions of daily life. Animal products were criticalcommodities that were utilized in the construction andnegotiation of class relationships and ritual and economicactivities that linked the residents of the city to governingofficials and to a greater regional context of exchange andinteraction. Diverse techniques for acquiring fauna havebeen documented across the Maya area (Carr, 1985,p. 126), and adaptations and diet can be vary considerablyat specific locales (Pohl, 1985a, p. 137, b, p. 109; Masson,1999, 2004). Mayapan’s patterns contribute valuableinformation from a large urban settlement of the Post-classic period for whose occupants faunal resources wereessential.
Acknowledgments
This analysis was made possible with the dedicatedassistance of UAlbany graduate students Amanda Schrei-ner, Juliana Novic, and Elizabeth Paris, as well as twoclasses of undergraduates in Albany in 2004 and 2005.Marilyn Masson checked all identifications and assumesfull responsibility for any errors. Elizabeth France, Beatrizdel Pilar Espinoza Geracitano, Megumi Orima, SarahHampson, Shauna McKown, Patrick Rodriguez, Carlos

ARTICLE IN PRESSM.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183182
Ariraga Mejıa, and Ismene Cowoh devoted many hours todata entry. Immense thanks are due to Kitty Emery andtwo anonymous reviewers for helpful comments. Researchat Mayapan is supported by the National ScienceFoundation (SBR-1018919), FAMSI, Inc., with the gra-cious permission of the Consejo de Arqueologıa of theInstituto Nacional de Antropologıa e Historia in MexicoCity.
References
Brothwell, D., Higgs, E., 1963. Science in Archaeology: A Comprehensive
Survey of Progress and Research. Basic Books, New York.
Brown, C.T., 1999. Mayapan Society and Ancient Maya Social
Organization. Ph.D. Dissertation, Department of Anthropology,
Tulane University, New Orleans.
Brown, L.A., 2005. Planting the bones: hunting ceremonialism at
contemporary and nineteenth-century shrines in the Guatemalan
highlands. Latin American Antiquity 16, 131–146.
Bullard, W.R., Jr., 1952. Residential property walls at Mayapan.
Department of Archaeology, Carnegie Institute of Washington,
Washington, DC, pp. 36–44 [Current Report 3].
Cahalane, V.H., 1931. Age classes of whitetail bucks killed in northern
Michigan in 1929. Journal of Mammology 12, 285–291.
Carr, H.S., 1985. Subsistence and ceremony: faunal utilization in a Late
Preclassic community at Cerros, Belize. In: Pohl, M. (Ed.), Prehistoric
Lowland Maya Environment and Subsistence Economy. Papers of the
Peabody Museum of Archaeology and Ethnology, vol. 77. Harvard
University, Cambridge, pp. 115–132.
Carr, H.S., 1996. Precolumbian Maya exploitation and management of
deer populations. In: Fedick, S.L. (Ed.), The Managed Mosaic:
Ancient Maya Agriculture and Resource Use. University of Utah
Press, Salt Lake City, pp. 251–261.
Davis, S.J.M., 1987. The Archaeology of Animals. B.T. Batsford, Ltd.,
London.
Delgado Ku, P.C., 2004. Estudio de la Arquitectura Publica del Nucleo
Principal de Mayapan, Yucatan. Tesis Profesional, Universidad
Autonoma de Yucatan, Merida, Mexico.
Ducos, P., 1978. ‘‘Domestication’’ defined and methodological approaches
to its recognition in faunal assemblages. In: Meadow, R.H., Zeder,
M.A. (Eds.), Approaches to Faunal Analysis in the Middle East.
Peabody Museum Bulletin, vol. 2. Peabody Museum of Archaeology
and Ethnology, Harvard University, Cambridge, MA, pp. 53–56.
Emery, K.F., Wright, L.E., Schwarcz, H., 2000. Isotopic analysis of
ancient deer bone: biotic stability in collapse period Maya land-use.
Journal of Archaeological Science 27, 537–550.
Fedick, S.L. (Ed.), 1996. The Managed Mosaic: Ancient Maya Agriculture
and Resource Use. University of Utah Press, Salt Lake City.
Fox, J.W., 1987. Late Postclassic State Formation. Cambridge University
Press, Cambridge.
Freidel, D.A., Schele, L., Parker, J., 1993. The Maya Cosmos: Three
Thousand Years on the Shaman’s Path. William Morrow and Co.,
New York.
Gilbert, B.M., 1993. Mammalian Osteology. Missouri Archaeological
Society, Columbia, MO.
Goodwin, G.G., 1934. Mammals collected by A.W. Anthony in
Guatemala, 1924–1928. Bulletin of the American Museum of Natural
History 68, 1–60.
Hamblin, N.L., 1984. Animal Use by the Cozumel Maya. The University
of Arizona Press, Tucson.
Hamblin, N.L., 1985. Isla Cozumel archaeological avifauna. In: Pohl, M.
(Ed.), Prehistoric Lowland Maya Environment and Subsistence
Economy. Papers of the Peabody Museum of Archaeology and
Ethnology, vol. 77. Harvard University, Cambridge, pp. 175–192.
Hare, T. S., Masson, M. A., Peraza Lope, C., 2006. The spatial and social
organization of Mayapan. Society for American Archaeology Meet-
ings, San Juan, Puerto Rico.
Jones, M., 1962. Map of Mayapan. In: H.E.D. Pollock, Roys, R.L.,
Proskouriakoff, T., Smith, A.L., Mayapan, Yucatan, Mexico, Rear
Inset. Carnegie Institute of Washington Publication No. 619,
Washington, DC.
Landa, D., 1941. Landa’s Relaciones de las Cosas de Yucatan (A. Tozzer,
Trans.). Papers of the Peabody Museum of Archaeology and
Ethnology, vol. 18. Harvard University Press, Cambridge.
Mann, C.C., 2005. 1491: New Revelations of the Americas Before
Columbus. Alfred A. Knopf, New York.
Masson, M.A., 1999. Animal resource manipulation in ritual and
domestic contexts at Postclassic Maya communities. World Archae-
ology 31, 93–120.
Masson, M.A., 2000. In the Realm of Nachan Kan: Postclassic Maya
Archaeology at Laguna de On, Belize. University Press of Colorado,
Boulder.
Masson, M.A., 2004. Fauna exploitation from the Preclassic to the
Postclassic periods at four Maya settlements in morthern Belize. In:
Emery, K. (Ed.), Maya Zooarchaeology: New Directions in Method
and Theory. The Cotsen Institute of Archaeology Press, University of
California, Los Angeles, pp. 97–122.
Masson, M.A., Peraza Lope, C., Hare, T.S., n.d. Postclassic Maya
economic heterogeneity at Mayapan. In: Sanders, W., Cobean, R.
(Eds.), El Urbanismo en Mesoamerica/Urbanism in Mesoamerica, vol.
3. Instituto Nacional de Antropologıa e Historia/Penn State
University, Mexico, D.F., and University Park.
Mock, S.B., 1994. The Northern River Lagoon site (NRL): Late to
Terminal Classic Maya settlement, saltmaking, and survival on the
northern Belize coast. Ph.D. Dissertation, Department of Anthropol-
ogy, University of Texas at Austin.
Olsen, S.J., 1982. An Osteology of Some Maya Mammals. Papers of the
Peabody Museum of Archaeology and Ethnology, vol. 73. Harvard
University, Cambridge.
Peraza Lope, C., Masson, M.A., Hare, T.S., Pedro, C., Delgado Ku, P.C.,
2006. The Late Postclassic chronology of Mayapan: new radiocarbon
evidence. Ancient Mesoamerica 17, 153–176.
Peraza Lope, C., Euan, M.E., Delgado Ku, P.C., Escamilla Ojeda, B.,
Chim, M.L., Garcıa Ayala, C., 2007. Trabajos de Mantenimiento y
Conservacion Arquitectonica en Mayapan, Yucatan. Informe de la
Temporada 1996 para el Consejo De Arqueologıa, Instituto Nacional
de Antropologıa e Historia. Centro INAH—Yucatan, Mexico.
Pohl, M.D., 1985a. The privileges of Maya elites: prehistoric vertebrate
fauna from Seibal. In: Pohl, M. (Ed.), Prehistoric Lowland Maya
Environment and Subsistence Economy. Papers of the Peabody
Museum of Archaeology and Ethnology, vol. 77. Harvard University,
Cambridge, pp. 133–145.
Pohl, M.D., 1985b. Osteological evidence for subsistence and status. In:
Pohl, M. (Ed.), Prehistoric Lowland Maya Environment and
Subsistence Economy. Papers of the Peabody Museum of Archaeology
and Ethnology, vol. 77. Harvard University, Cambridge, pp. 109–113.
Pohl, M.D., 1989. Ethnozoology of the Maya: faunal remains from five
sites in the Peten, Guatemala. In: Graham, J.A., Tourtellot, III, G.,
Pohl, M.D., Willey, G.R. (Eds.), Excavations at Seibal, Department of
Peten, Guatemala. Memoirs of the Peabody Museum, vol. 17(3).
Harvard University, Cambridge.
Pohl, M.D., Feldman, L.H., 1982. The traditional role of women and
animals in lowland Maya economy. In: Flannery, K.V. (Ed.), Maya
Subsistence: Studies in Memory of Dennis E. Puleston. Academic
Press, New York, pp. 295–311.
Pollock, H.E.D., Ray, C.E., 1957. Notes on vertebrate animal remains
from Mayapan. Carnegie Institute of Washington, Department of
Archaeology, Washington, DC, Current Reports 41, pp. 633–656.
Proskouriakoff, T., 1962. Civic and religious structures of Mayapan. In:
Pollock, H.E.D., Roys, R.L., Proskouriakoff, T., Smith, A.L. (Eds.),
Mayapan, Yucatan, Mexico, Publication No. 619, Carnegie Institute
of Washington, Washington, DC, pp. 87–164.

ARTICLE IN PRESSM.A. Masson, C. Peraza Lope / Quaternary International 191 (2008) 170–183 183
Pugh, T., 2001. Flood reptiles, serpent temples and the quadripartite
universe: the imajo mundi of Late Postclassic Mayapan. Ancient
Mesoamerica 12, 247–258.
Rackham, J., 1994. Animal Bones. University of California Press/British
Museum, Berkeley and Los Angeles.
Reitz, E.J., Wing, E.S., 1999. Zooarchaeology. Cambridge University
Press, Cambridge.
Ringle, W.M., Bey, G.J., 2001. Post-classic and terminal classic courts of
the northern Maya lowlands. In: Inomata, T., Houston, S. (Eds.),
Royal Courts of the Maya, Volume Two: Data and Case Studies.
Westview Press, Boulder, pp. 266–307.
Roys, R.L., 1957. The Political Geography of the Yucatan Maya.
Publication 613, Carnegie Institute of Washington, Washington, DC.
Roys, R.L., 1962. Literary sources for the history of Mayapan. In:
Pollock, H.E.D., Roys, R.L., Proskouriakoff, T., Smith, A.L. (Eds.),
Mayapan, Yucatan, Mexico, Publication No. 619, Carnegie Institute
of Washington, Washington, DC, pp. 25–86.
Russell, B.W., 2008. Postclassic Maya Settlement on the rural–urban
Fringe of Mayapan, Yucatan, Mexico. Ph.D. Dissertation. On file at
the Department of Anthropology, The University at Albany—SUNY,
Albany, New York.
Shaw, L.C., 1991. The articulation of social inequality and faunal resource
use in the Preclassic community of Colha, northern Belize. Ph.D.
Dissertation, University of Massachusetts, Amherst.
Shaw, L.C., 1999. Social and ecological aspects of preclassic Maya meat
consumption at Colha, Belize. In: White, C.D. (Ed.), Reconstructing
Ancient Maya Diet. University of Utah Press, Salt Lake City,
pp. 83–102.
Smith, A.L., 1962. Residential and associated structures at Mayapan. In:
Pollock, H.E.D., Roys, R.L., Proskouriakoff, T., Smith, A.L. (Eds.),
Mayapan, Yucatan, Mexico, Publication No. 619, Carnegie Institute
of Washington, Washington, DC, pp. 165–320.
Smith, M.E., 2005. City size in Late Postclassic Mesoamerica. Journal of
Urban History 31, 403–434.
Teeter, W.G., Chase, A.F., 2004. Adding flesh to the bones: using
zooarchaeology research to answer the big-picture questions. Archae-
ofauna 13, 155–172.
White, C.D., Pohl, M.D., Schwarcz, H., Longstaffe, F.J., 2001. Preclassic
Maya patterns of deer and dog exploitation: the isotopic evidence from
Colha, Belize. Journal of Archaeological Science 28, 89–107.
Ximenez, F., 1967. Historia natural del reino de Guatemala. Sociedad de
Geografıa e Historia de Guatemala, Special Publication 14.





























