Embed Size (px)
Citation preview

Hindawi Publishing CorporationClinical and Developmental ImmunologyVolume 2013 Article ID 959650 11 pageshttpdxdoiorg1011552013959650
Research ArticleAn Elucidation of Neutrophil Functions againstMycobacterium tuberculosis Infection
Devin Morris1 Thien Nguyen1 John Kim2 Christine Kassissa1 Melissa Khurasany2
Jennifer Luong1 Sarah Kasko1 Shalin Pandya1 Michael Chu1 Po-Ting Chi3 Judy Ly1
Minette Lagman3 and Vishwanath Venketaraman13
1 College of Osteopathic Medicine of the Pacific Western University of Health Sciences Pomona CA 91766 USA2College of Dental Medicine Western University of Health Sciences Pomona CA 91766 USA3Graduate College of Biomedical Sciences Western University of Health Sciences Pomona CA 91766 USA
Correspondence should be addressed to Vishwanath Venketaraman vvenketaramanwesternuedu
Received 28 August 2013 Accepted 1 October 2013
Academic Editor Masha Fridkis-Hareli
Copyright copy 2013 Devin Morris et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
We characterized the functions of neutrophils in response toMycobacterium tuberculosis (M tb) infection with particular referenceto glutathione (GSH)We examined the effects of GSH in improving the ability of neutrophils to control intracellularM tb infectionOur findings indicate that increasing the intracellular levels of GSH with a liposomal formulation of GSH (L-GSH) resulted inreduction in the levels of free radicals and increased acidification ofM tb containing phagosomes leading to the inhibition in thegrowth of M tb This inhibitory mechanism is dependent on the presence of TNF-120572 and IL-6 Our studies demonstrate a novelregulatory mechanism adapted by the neutrophils to controlM tb infection
1 Introduction
M tb a causative agent for tuberculosis (TB) is known tobe a highly successful intracellular pathogen and is estimatedto be harbored in one third of the worldrsquos population [1]Left untreated TB is known to have a 70 mortality rate[2] Increasing drug resistance strains of M tb and poorprecautionarymeasurements for containing the spread espe-cially in ldquohigh burdenrdquo countries have kept the disease fromglobal eradication [1] M tb infection is mostly confined tolungs and can be transmitted by inhalation of droplets fromindividuals with active TB M tb has the ability to evadethe host killing mechanisms and can successfully persistinside the phagocytic cells Specifically M tb is capable ofsurviving within the host cells by preventing the fusion ofthe phagosomes containing bacteria with lysosomes therebyavoiding exposure to the toxic lysosomal hydrolases [3]
The terminally differentiated polymorphonuclear leuko-cytes (PMN) or neutrophils are essential component ofthe human innate immune system and act as a first-line
defense against invading microorganisms Neutrophils aremajor inhabitants of the white blood cell population (WBC)since they constitute sim70 of the total WBC population [4]Neutrophils form the bulk of early recruited leukocyte popu-lation that mediate and elicit immune responses againstM tb[4] Neutrophils after encountering themycobacteria exhibitinnate immune responses such as phagocytosis which in turntriggers signaling events resulting in secretion of cytokinessuch as TNF-120572 and IL-1 that mediate early inflammatoryresponses [4ndash6]
The tripeptide glutathione (GSH) protects all cellsagainst oxidizing agents free radicals and reactive oxygenintermediates (ROI) either directly or through enzymaticaction of GSH peroxidases and GSH-transferases [7 8]GSH is also important for cellular homeostasis as well asmany different cellular functions such as protein synthesisenzyme catalysis transmembrane transport receptor actionintermediary metabolism and cell maturation [9ndash11] GSHis produced by nearly all cell types and exists in two formsreduced or free GSH (rGSH) and oxidized (GSSG) rGSH is
2 Clinical and Developmental Immunology
responsible for the antioxidant functions ofGSHwhileGSSGis the byproduct of the free radical scavenging activity of GSHand lacks antioxidant function [9ndash11]
We have previously reported that GSH has antimycobac-terial effects and furthermore enhancing the levels of GSHin human macrophages resulted in inhibition in the growthof intracellular M tb [12] Additionally increasing the levelsof GSH in immune cells such as natural killer (NK) cellsand T cells resulted in improved control of M tb infectioninside humanmonocytes indicating that GSH has both directantimycobacterial effects as well as immune enhancing effects[13 14]
Though many studies have highlighted the roles ofmonocytes and macrophages in innate immune responsesagainst M tb infection very few studies have demonstratedthe importance of neutrophils during M tb infection [5 6]The role of GSH in regards to the neutrophil innate responseand pathophysiology has been poorly defined In this studyan attempt was made to investigate the role of neutrophils ininnate defense againstM tb infection
We chose to examine the functions of neutrophils againstM tb infection for three chief reasons first they mirrormacrophage mechanisms mainly by phagocytosing bacteriasecond they are one of the first responders to bacterialinfections and third they are closely tied to free radical andinflammatory responses We also characterized the under-lying mechanisms by which GSH-enhanced neutrophils areable to successfully inhibit the growth ofM tb We hypothe-sized that augmenting the levels of GSH in neutrophils willincrease the fusion of phagosomes containing M tb withlysosomes leading to inhibition in the growth of M tb Theultimate purpose of this research is to determine the abilityof GSH-enhanced neutrophils to effectively control M tbinfection By better characterizing the role of neutrophilswhich form a major population of WBC we can furtheradvance the crusade againstM tb control and eradication
We tested our hypothesis by examining the effects oftwo different GSH-enhancing agents a GSH precursor N-acetyl cysteine (NAC) and L-GSH a liposomal formulationof GSH in mediating the control of M tb infection insideneutrophils We correlated the inhibition in the growth ofMtb inside neutrophils with the levels of GSH and free radicalsproduction of inflammatory cytokines such as IL-6 and TNF-120572 and the extent of phagosome lysosome fusion Our resultsindicate that treatment of neutrophils with L-GSH resultedin inhibition in the growth of M tb and this inhibitionwas accompanied by increased fusion between phagosomescontaining M tb with lysosomes Our findings unveiled oneof the important mechanisms by which GSH improves theability of neutrophils to controlM tb infection
2 Materials and Methods
21 Statement of Ethics All studies were approved by both theInstitutional Review Board and the Institutional bio-safetycommittee of the Western University of Health Sciences Allstudy participants were above the legal age of consent atthe time of participation and written informed consent was
obtained from all volunteers prior to participation in thestudy
22 Subjects Healthy subjects without a history of TB wererecruited from the students faculty and staff of WesternUniversity of Health Sciences Forty milliliters of blood wasdrawn once from each healthy volunteer after obtainingsigned informed consent
23 Isolation of Neutrophils Neutrophils were isolated fromthe whole blood of healthy individuals using density gradientcentrifugation with dextran and Ficoll histopaque a highdensity pH neutral polysaccharide solution (Sigma) Briefly40 milliliters of whole blood was diluted in equal volumeof 09 NaCl saline solution (1 1) mixed gently layered ontop of the Ficoll-hypaque and centrifuged for duration of40 minutes at 1400 rpm The neutrophils rich RBC layer wasdiluted to a total volume of 25mL with hanks balanced saltsolution mixed with equal volume of 6 dextran (specificaverage molecular weight of 250000 g) and incubated for2 hours to allow for gravitational sedimentation of RBCsFollowing incubation the semitransparent supernatant wasaspirated off and centrifuged for 10minutes at 1400 rpm Sub-sequent to centrifugation the soft pellet containing PMNrsquosand RBCrsquos was gently dispersed and resuspended in PBSCells were washed once in PBS and the contaminatingRBCrsquos were eliminated by treating the cell suspension withRBC lysis buffer Purified neutrophils were resuspended inRoswell ParkMemorial Institute (RPMI)medium containing5 AB serum 1 times 104 neutrophils were added to eachwell of the polylysine coated 96-well tissue culture platesand incubated for 12 hours at 37∘C 5 CO
2incubator for
adherence Following overnight adherence nonadherent cellswere removed by washing and the adherent neutrophilswere resuspended in fresh media and used for variousassays Purity of the cultures was determined by microscopicexamination of nuclear morphology after staining with 41015840 6-diamidino-2-phenylindole (DAPI)
24 Preparation of Bacterial Cells for Neutrophil Infection Allinfection studies were performed using the virulent labora-tory strain ofM tb H37Rv inside the biosafety level 3 (BSL-3)facility M tb H37Rv was processed for infection as followsstatic cultures of H37Rv at their peak logarithmic phase ofgrowth (between 05 and 08 at A600) were used for infectionof neutrophils The bacterial suspension was washed and re-suspended in RPMI (Sigma) containing AB serum (Sigma)Bacterial clumps were disaggregated by vortexing five timeswith 3mm sterile glass beads The bacterial suspension waspassed through a 5 120583m syringe filter (Millipore) to removeany existing clumps The total number of organisms in thesuspension was ascertained by plating Processed H37Rvwere frozen as stocks at minus80∘C At the time of infectionfrozen stocks of processed H37Rv were thawed and used forinfection of neutrophils
25 Infection of Neutrophils with H37Rv To examine therole of neutrophils and GSH in innate defense against
Clinical and Developmental Immunology 3
M tb infection we determined the intracellular survival ofM tb inside neutrophils that were cultured in the presenceand absence of GSH-enhancing agents Neutrophils wereinfected with processed H37Rv at a low dose multiplicityof infection of 1 10 (1 bacterium for every 10 neutrophils)and incubated for 2 hours to allow for phagocytosis Un-phagocytosed bacteria were removed by washing the infectedneutrophil cultures three times with warm sterile PBS (basedon the results from our standardization studies this low dosemultiplicity of infection usually resulted in approximately10of the neutrophils infectedwithH37Rv and each infectedcell had between 1 and 5 bacteria) Infected neutrophils werecultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in
the presence and absence of NAC (10mM) or L-GSH (10and 20120583M Your Energy Systems LLC Palo Alto CA) Theliposomal formulation of GSH (L-GSH) used in these studiesis a proprietary product of Your Energy Systems (YES) LLC(Palo Alto CA) called ReadiSorb Infected neutrophils wereterminated at 1 hour and 24 hours after infection to determinethe intracellular survival of H37Rv
26 Termination of Infected Neutrophils and Measurementof Colony Forming Units (CFU) Termination of H37Rv-infected neutrophils was performed by the addition of 200120583Lof sterile distilled water to each culture well The collectedneutrophil lysates were diluted in sterile water and plated on7H11 medium (Hi Media) enriched with albumin dextrosecomplex (ADC) to estimate the extent of H37Rv growth inneutrophils by counting the CFUs
27 Neutrophil Viability Assay Theeffects of GSH-enhancingagents in altering the viability of H37Rv-infected neutrophilswere determined by fluorescent DAPI staining Neutrophils(1 times 104well) were distributed in sterile glass cover slipspositioned in 24-well tissue culture plates and incubated for12 hours for adherence Adhered neutrophils were infectedH37Rv at a multiplicity of infection of 1 10 (1 bacteria to 10neutrophils) Infected neutrophils were cultured in RPMI +5 AB serum at 37∘C + 5 CO
2in the presence and absence
of NAC (10mM) or L-GSH (10 and 20120583M) for 24 hoursInfected neutrophils were fixed with 38 paraformaldehydein PBS for 30minutes Fixed neutrophils were washed twicefor 5minutes in PBS A single drop of mounting mediacontaining DAPI was placed per glass slide The stainedneutrophils were then attached to glass cover slips andinverted onto the glass slide The cover slips were thensealed to the glass slide by smearing nail polish around theedges of the cover slips The stained slides were then viewedusing an inverted fluorescent microscope Intact cells withDAPI staining were counted under 20x magnification in7 randomly selected fields for each category A neutrophilsurvival index was generated by dividing the number ofsurviving neutrophils in each treatment category by thenumber of surviving neutrophils in the untreated controlcategory
28 Assay of GSH Levels in Neutrophils GSH levels inneutrophils were measured by spectrophotometry using an
assay kit from Arbor Assays Briefly uninfected and H37Rv-infected neutrophils (1 times 104) were lysed by the additionof ice cold 5 5-sulfosalicylic acid dehydrate solution (MPBiomedicals) Lysates were centrifuged and supernatantscollected after centrifugation were analyzed for total GSHandoxidizedGSH (GSSG) as permanufacturerrsquos instructionsLevels of free or reduced GSH (rGSH) were calculatedby subtracting measured GSSG concentrations from themeasured total GSH concentrations All GSH measurementswere normalized with total protein levels
29 Assay of Total Protein Levels in Lysates Proteins in theisolated cell lysates were measured by Bradfordrsquos methodusing a Coomassie protein assay reagent (Thermo Scientific)This assay helped to determine the amount of protein in eachwell By dividing the total protein value from the GSH assayit helped to confirm that the variation of GSH is due to thetreatments added
210 Assay of IL-6 and TNF-120572 in Neutrophil SupernatantsLevels of IL-6 and TNF-120572 in supernatants from uninfectedand H37Rv-infected neutrophils were measured by enzymelinked immunosorbent assay (ELISA) (eBioscience)
211 Assay of Free Radicals in Neutrophil Lysates Free radicallevels in neutrophil lysates were determined bymeasuring thelevels of malondialdehyde (MDA) using a colorimetric assaykit from Cayman The MDA levels in the infected groups(M tb-infectedM tb-infected and treated with NAC andMtb-infected and treated with L-GSH) were normalized to thevalues in uninfected neutrophils
212 Fluorescent Microscopic Analysis of M tb PhagosomeAcidification in Neutrophils The effects of GSH-enhancingagents in inducing acidification of M tb-containing phago-somes were determined by quantifying the colocalizationof Fluorescein Isothiocyanate (FITC) labeled H37Rv in thelysotracker labeled acidified compartments by fluorescentmicroscope Lysotracker Red DND99 (Molecular Probes)an acidotropic dye is a weak base conjugated to a redfluorophore Lysotracker freely permeates cell membranesand gets trapped upon protonation in an acidified compart-ment The excitation and emission maxima of lysotrackerare 577 and 592 nm respectively Neutrophils (1 times 104well)were distributed in polylysine coated sterile glass cover slipspositioned in 24-well tissue culture plates and incubated for12 hours for adherence Adhered neutrophils were treatedwith lysotracker (1103 dilution) for 1 hour H37Rvwas labeledwith FITC as follows bacterial cells were incubated with08mgmL FITC for 1 hour The labeled cells were thencentrifuged and the pellet was resuspended in PBS and usedfor infecting the lysotracker-labeled neutrophils Lysotracker-labeled neutrophils were incubatedwith FITC-labeledH37Rvfor 15 hours to allow phagocytosis Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5CO2in the presence and absence of NAC (10mM) or L-
GSH (10 and 20120583M) for 24 hours Infected neutrophils werefixed with 38 paraformaldehyde in PBS for 30 minutes
4 Clinical and Developmental Immunology
0
2
4
6
M tb
Fold
incr
ease
d in
M t
b gr
owth
M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(a)
0
01
02
03
04
05
06
07
Neu
troph
il su
rviv
al in
dex
M tb M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(b)
Figure 1 Intracellular survival of H37Rv in neutrophils treated with NAC and L-GSH (119899 = 5)Neutrophils were infected with processedH37Rvat a low dose multiplicity of infection of 1 10 (1 bacterium for every 10 neutrophils) and incubated for 2 hours to allow for phagocytosisUnphagocytosed mycobacteria were removed by washing the infected neutrophils three times with warm sterile PBS Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in the presence and absence of NAC (10mM) or L-GSH (10 and 20120583M) Infected
neutrophils were terminated at 1 hour and 24 hours after infection to determine the intracellular survival of H37Rv Graph shows mean plusmnstandard error(a) Represents foldincrease in intracellular growth of H37Rv in neutrophils treated with NAC and L-GSH (b) Determinationof neutrophil viability following infection with H37Rv and treatment with NAC and L-GSH by quantifying DAPI stained cells (119899 = 5)
Fixed neutrophils were washed twice for 5 minutes in PBSA single drop of mounting media containing DAPI wasplaced per glass slide The stained neutrophils were thenattached to glass cover slips and inverted onto the glassslide The cover slips were then sealed to the glass slide bysmearing nail polish around the edges of the cover slips Thestained slides were then viewed using an inverted fluorescentmicroscope Images were obtained using an integrated digitalcamera Images were subsequently analyzed using ImageJ afree software program available from the National Institutesof Health (httprsbwebnihgovij) Following backgroundfluorescence correction the average fluorescent intensity wasmeasured
213 Statistical Analysis All statistical analysis was doneusing Graph Pad Prism6 software on the mean plusmn stan-dard error for 119899 = 5 individuals For all assays theinfected-untreated category was compared to the uninfected-untreated control category using a two-tailed Studentrsquos 119905-testwith 95 confidence intervals All infected-treated categorieswere compared to the infected-untreated category using aKruskal-Wallis 1-way ANOVA with Dunnrsquosmultiple compar-isons posttest Results were considered significant for 119875 le005
3 Results
31 Intracellular Survival of H37Rv inside Human NeutrophilsWe tested the effects of GSH-enhancing agents in improv-ing the functions of neutrophils to inhibit the growth ofM tb This was achieved by determining the intracellularsurvival of H37Rv a virulent laboratory strain ofM tb insideNACL-GSH-treated human neutrophils We observed a 4-fold increase in the intracellular growth of H37Rv inside
unstimulated neutrophils (Figure 1(a)) Treatment of neu-trophils with L-GSH at 10 and 20 120583M concentrations resultedin a decrease in the foldgrowth of M tb (Figure 1(a)) Infact we observed maximum inhibition in the growth ofH37Rv in neutrophils that were treated with L-GSH andthis is in contrast to untreated neutrophils in which therewas a 4-fold increase in the growth of intracellular M tb(Figure 1(a)) Interestingly treatment of neutrophils with theGSH-precursor NAC at 50x higher concentration comparedto L-GSH did not reduce the growth of M tb but insteadwe observed a 6-fold increase in the intracellular growth ofH37Rv (Figure 1(a)) These results indicate that an optimalincrease in the intracellular levels of GSH in neutrophilswill lead to inhibition in the growth of M tb Treatment ofH37Rv-infected neutrophils with GSH-enhancing agents didnot result in any loss in the viability of neutrophils as evidentfrom the quantification of DAPI stained cells (Figure 1(b))
32 Assay of GSH Levels in Neutrophils We determined theability of NAC and L-GSH to enhance the intracellular levelsof GSH in uninfected neutrophils and restore the levels ofGSH in H37Rv-infected neutrophils by spectrophotometryWe used an assay kit from Arbor Assays We observed thattreatment of uninfected neutrophils with NAC resulted in3-fold increase in the levels of both total GSH and rGSH(Figure 2) L-GSH treatment did not result in any increasein the levels of both total GSH and rGSH in uninfectedneutrophils (Figure 2) Importantly H37Rv infection resultedin a 20 decrease in the levels of total GSH in neutrophils(Figure 3(a)) The percentages of rGSH versus GSSG inH37Rv-infected neutrophils was 56 and 44 respectively(Figure 3(d)) Whereas the percentage of rGSH and GSSGin uninfected neutrophils were 76 and 24 respectively(Figure 2(d)) Our results signify that M tb infection leads
Clinical and Developmental Immunology 5
Control NAC 10120583M LGSH 20120583M LGSH000
002
004
006
008
010To
tal G
SH120583
g to
tal p
rote
in
(a)
000
002
004
006
008
Control 10mM NAC 10120583M LGSH 20120583M LGSH
120583M
rGSH
120583g
tota
l pro
tein
(b)
0000
0002
0004
0006
0008
00010
Tota
l GSS
G120583
g to
tal p
rote
in
Control 10mM NAC 10120583M LGSH 20120583M LGSH
(c)
24
76
6
94
14
86
24
76
rGSHGSSG
rGSHGSSG
Control 10mM NAC
10120583M LGSH 20120583M LGSH
(d)
Figure 2 GSH measurements in uninfected neutrophils cultured in the presence and absence of NAC or L-GSH (119899 = 5) GSH levelswere measured in isolated neutrophils from healthy subjects that were treated with GSH-enhancing agents such as NAC and L-GSH byspectrophotometry using an assay kit Briefly neutrophils (3 times 105) were lysed at 24 hours after treatment with ice cold 5 5-sulfosalicylicacid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH (a) and GSSG (c) as per manufacturerrsquosinstructions rGSH was calculated by subtracting measured GSSG concentrations from the measured total GSH concentrations (b) All GSHmeasurements were normalized with total protein levels Proteins in the cell lysates of neutrophils were measured by Bradfordrsquos method usinga Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressed as percentage of total GSH in neutrophil lysates Graphshows mean plusmn standard error
to decreased levels of rGSH the functional form of GSHTreatment of H37Rv-infected neutrophils with 10mM NACresulted in a 10-fold increase in the intracellular levels oftotal GSH (Figure 3(a)) Treatment of H37Rv-infected neu-trophils with L-GSH (10 and 20 120583M concentration) resultedin restoration in the intracellular levels of both total GSHand rGSH to similar levels as uninfected neutrophils (Figures3(a) and 3(b)) Treatment ofH37Rv-infected neutrophils withGSH-enhancing agents (NAC and L-GSH) also resulted inreversal of GSSG and rGSH ratio similar to the percentagein uninfected neutrophils (Figure 3(d)) Our results indicatethatM tb infection decreases the levels of GSH in neutrophilspossibly by inducing oxidative stress (as evident from theincreased percentage of GSSG) and treatment of M tb-infected neutrophils with GSH-enhancing agents restores theintracellular levels of both total GSH and rGSH
33 Determination of Free Radicals Levels in Neutrophils Wedetermined the inverse correlation of GSH levels and ROIby quantifying the levels of free radicals in H37Rv-infected
neutrophils We observed that infection of neutrophils withH37Rv resulted in a 3-fold increase in the production offree radicals compared to the uninfected neutrophils (Fig-ure 4(a)) NAC and L-GSH treatments led to a noticeabledecrease in the levels of free radicals The decreased levelswere similar to the uninfected neutrophils (Figure 4(a))Maximum decrease in the levels of free radicals was observedwhen H37Rv-infected neutrophils were treated with 20120583ML-GSH (Figure 4(a)) Our results highlight the ability of Mtb infection to induce neutrophils to produce free radicalsleading to a decrease in the levels of rGSH
34 TNF-120572 Assay in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 28-fold increase inthe production of TNF-120572 compared to uninfected neutrophils(Figure 4(b)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a dramatic decrease in the productionof TNF-120572 In fact the TNF-120572 levels were undetectable insupernatants collected from H37Rv-infected +NAC-treatedneutrophils (Figure 4(b)) These results indicate that NAC is
6 Clinical and Developmental Immunology
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
005
010
015120583
M to
tal G
SH120583
g to
tal p
rote
in
lowast
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
004
002
006
008
010
120583M
tota
l GSH
120583g
tota
l pro
tein lowast
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
002
001
003
004
005
120583M
tota
l GSS
G120583
g to
tal p
rote
in
(c)
56
20
80
26
74
26
74
rGSHGSSG
rGSHGSSG
44
M tb
10120583M LGSH 20120583M LGSH
M tb +
M tb + M tb +
10 mM NAC
(d)
Figure 3 GSH measurements in neutrophils infected with H37Rv and treated with NAC and L-GSH (119899 = 5) GSH levels were measured inisolated neutrophils from healthy subjects that were infected in vitro with H37Rv and treated with GSH-enhancing agents such as NAC andL-GSH by spectrophotometry using an assay kit Briefly H37Rv-infected neutrophils (3 times 105) were lysed at 24 hours after infection withice cold 5 5-sulfosalicylic acid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH ((a) lowast119875 le 005)and GSSG (c) as per manufacturerrsquos instructions rGSH ((b) lowast119875 le 005) was calculated by subtracting measured GSSG concentrations fromthe measured total GSH concentrations All GSH measurements were normalized with total protein levels Proteins in the cell lysates ofneutrophils were measured by Bradfordrsquos method using a Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressedas percentage of total GSH in neutrophil lysates Graph shows mean plusmn standard error
capable of completely diminishing the production of TNF-120572byH37Rv-infected neutrophils Treatment ofH37Rv-infectedneutrophils with 10 and 20 120583M L-GSH did not result in anydecrease in the production of TNF-120572 compared to untreatedH37Rv-infected neutrophils (Figure 4(b))
35 Assay of IL-6 in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 38-fold increase inthe production of IL-6 compared to uninfected neutrophils(Figure 4(c)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a striking decrease in the productionof IL-6 (Figure 4(c)) IL-6 levels were undetectable in super-natants collected from H37Rv-infected + NAC-treated neu-trophils Treatment of H37Rv-infected neutrophils with L-GSH resulted did not result in any significant decrease in theproduction of IL-6 compared to untreated H37Rv-infected
neutrophils (Figure 4(c)) These results indicate that NACtreatment at millimolar concentrations is capable of com-pletely diminishing the production of both IL-6 and TNF-120572by H37Rv-infected neutrophils (Figures 4(b) and 4(c))
36 Examining the Acidification of M tb Containing Phago-somes by Quantifying the Colocalization of FITC-LabeledH37Rv with Lysotracker We examined the effects of GSHon increasing the acidification of phagosomes containingMtb by quantifying the colocalization of FITC-labeled H37Rvwith lysotracker trapped in the acidified compartments inthe neutrophils (Figure 5) We observed that treatment ofH37Rv-infected neutrophils with 10120583M L-GSH resulted in40 increase in the colocalization of FITC-labeled H37Rvwith lysotracker (Figure 4(d)) Interestingly treatment ofH37Rv-infected neutrophils with 20120583M L-GSH resulted in a
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010

2 Clinical and Developmental Immunology
responsible for the antioxidant functions ofGSHwhileGSSGis the byproduct of the free radical scavenging activity of GSHand lacks antioxidant function [9ndash11]
We have previously reported that GSH has antimycobac-terial effects and furthermore enhancing the levels of GSHin human macrophages resulted in inhibition in the growthof intracellular M tb [12] Additionally increasing the levelsof GSH in immune cells such as natural killer (NK) cellsand T cells resulted in improved control of M tb infectioninside humanmonocytes indicating that GSH has both directantimycobacterial effects as well as immune enhancing effects[13 14]
Though many studies have highlighted the roles ofmonocytes and macrophages in innate immune responsesagainst M tb infection very few studies have demonstratedthe importance of neutrophils during M tb infection [5 6]The role of GSH in regards to the neutrophil innate responseand pathophysiology has been poorly defined In this studyan attempt was made to investigate the role of neutrophils ininnate defense againstM tb infection
We chose to examine the functions of neutrophils againstM tb infection for three chief reasons first they mirrormacrophage mechanisms mainly by phagocytosing bacteriasecond they are one of the first responders to bacterialinfections and third they are closely tied to free radical andinflammatory responses We also characterized the under-lying mechanisms by which GSH-enhanced neutrophils areable to successfully inhibit the growth ofM tb We hypothe-sized that augmenting the levels of GSH in neutrophils willincrease the fusion of phagosomes containing M tb withlysosomes leading to inhibition in the growth of M tb Theultimate purpose of this research is to determine the abilityof GSH-enhanced neutrophils to effectively control M tbinfection By better characterizing the role of neutrophilswhich form a major population of WBC we can furtheradvance the crusade againstM tb control and eradication
We tested our hypothesis by examining the effects oftwo different GSH-enhancing agents a GSH precursor N-acetyl cysteine (NAC) and L-GSH a liposomal formulationof GSH in mediating the control of M tb infection insideneutrophils We correlated the inhibition in the growth ofMtb inside neutrophils with the levels of GSH and free radicalsproduction of inflammatory cytokines such as IL-6 and TNF-120572 and the extent of phagosome lysosome fusion Our resultsindicate that treatment of neutrophils with L-GSH resultedin inhibition in the growth of M tb and this inhibitionwas accompanied by increased fusion between phagosomescontaining M tb with lysosomes Our findings unveiled oneof the important mechanisms by which GSH improves theability of neutrophils to controlM tb infection
2 Materials and Methods
21 Statement of Ethics All studies were approved by both theInstitutional Review Board and the Institutional bio-safetycommittee of the Western University of Health Sciences Allstudy participants were above the legal age of consent atthe time of participation and written informed consent was
obtained from all volunteers prior to participation in thestudy
22 Subjects Healthy subjects without a history of TB wererecruited from the students faculty and staff of WesternUniversity of Health Sciences Forty milliliters of blood wasdrawn once from each healthy volunteer after obtainingsigned informed consent
23 Isolation of Neutrophils Neutrophils were isolated fromthe whole blood of healthy individuals using density gradientcentrifugation with dextran and Ficoll histopaque a highdensity pH neutral polysaccharide solution (Sigma) Briefly40 milliliters of whole blood was diluted in equal volumeof 09 NaCl saline solution (1 1) mixed gently layered ontop of the Ficoll-hypaque and centrifuged for duration of40 minutes at 1400 rpm The neutrophils rich RBC layer wasdiluted to a total volume of 25mL with hanks balanced saltsolution mixed with equal volume of 6 dextran (specificaverage molecular weight of 250000 g) and incubated for2 hours to allow for gravitational sedimentation of RBCsFollowing incubation the semitransparent supernatant wasaspirated off and centrifuged for 10minutes at 1400 rpm Sub-sequent to centrifugation the soft pellet containing PMNrsquosand RBCrsquos was gently dispersed and resuspended in PBSCells were washed once in PBS and the contaminatingRBCrsquos were eliminated by treating the cell suspension withRBC lysis buffer Purified neutrophils were resuspended inRoswell ParkMemorial Institute (RPMI)medium containing5 AB serum 1 times 104 neutrophils were added to eachwell of the polylysine coated 96-well tissue culture platesand incubated for 12 hours at 37∘C 5 CO
2incubator for
adherence Following overnight adherence nonadherent cellswere removed by washing and the adherent neutrophilswere resuspended in fresh media and used for variousassays Purity of the cultures was determined by microscopicexamination of nuclear morphology after staining with 41015840 6-diamidino-2-phenylindole (DAPI)
24 Preparation of Bacterial Cells for Neutrophil Infection Allinfection studies were performed using the virulent labora-tory strain ofM tb H37Rv inside the biosafety level 3 (BSL-3)facility M tb H37Rv was processed for infection as followsstatic cultures of H37Rv at their peak logarithmic phase ofgrowth (between 05 and 08 at A600) were used for infectionof neutrophils The bacterial suspension was washed and re-suspended in RPMI (Sigma) containing AB serum (Sigma)Bacterial clumps were disaggregated by vortexing five timeswith 3mm sterile glass beads The bacterial suspension waspassed through a 5 120583m syringe filter (Millipore) to removeany existing clumps The total number of organisms in thesuspension was ascertained by plating Processed H37Rvwere frozen as stocks at minus80∘C At the time of infectionfrozen stocks of processed H37Rv were thawed and used forinfection of neutrophils
25 Infection of Neutrophils with H37Rv To examine therole of neutrophils and GSH in innate defense against
Clinical and Developmental Immunology 3
M tb infection we determined the intracellular survival ofM tb inside neutrophils that were cultured in the presenceand absence of GSH-enhancing agents Neutrophils wereinfected with processed H37Rv at a low dose multiplicityof infection of 1 10 (1 bacterium for every 10 neutrophils)and incubated for 2 hours to allow for phagocytosis Un-phagocytosed bacteria were removed by washing the infectedneutrophil cultures three times with warm sterile PBS (basedon the results from our standardization studies this low dosemultiplicity of infection usually resulted in approximately10of the neutrophils infectedwithH37Rv and each infectedcell had between 1 and 5 bacteria) Infected neutrophils werecultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in
the presence and absence of NAC (10mM) or L-GSH (10and 20120583M Your Energy Systems LLC Palo Alto CA) Theliposomal formulation of GSH (L-GSH) used in these studiesis a proprietary product of Your Energy Systems (YES) LLC(Palo Alto CA) called ReadiSorb Infected neutrophils wereterminated at 1 hour and 24 hours after infection to determinethe intracellular survival of H37Rv
26 Termination of Infected Neutrophils and Measurementof Colony Forming Units (CFU) Termination of H37Rv-infected neutrophils was performed by the addition of 200120583Lof sterile distilled water to each culture well The collectedneutrophil lysates were diluted in sterile water and plated on7H11 medium (Hi Media) enriched with albumin dextrosecomplex (ADC) to estimate the extent of H37Rv growth inneutrophils by counting the CFUs
27 Neutrophil Viability Assay Theeffects of GSH-enhancingagents in altering the viability of H37Rv-infected neutrophilswere determined by fluorescent DAPI staining Neutrophils(1 times 104well) were distributed in sterile glass cover slipspositioned in 24-well tissue culture plates and incubated for12 hours for adherence Adhered neutrophils were infectedH37Rv at a multiplicity of infection of 1 10 (1 bacteria to 10neutrophils) Infected neutrophils were cultured in RPMI +5 AB serum at 37∘C + 5 CO
2in the presence and absence
of NAC (10mM) or L-GSH (10 and 20120583M) for 24 hoursInfected neutrophils were fixed with 38 paraformaldehydein PBS for 30minutes Fixed neutrophils were washed twicefor 5minutes in PBS A single drop of mounting mediacontaining DAPI was placed per glass slide The stainedneutrophils were then attached to glass cover slips andinverted onto the glass slide The cover slips were thensealed to the glass slide by smearing nail polish around theedges of the cover slips The stained slides were then viewedusing an inverted fluorescent microscope Intact cells withDAPI staining were counted under 20x magnification in7 randomly selected fields for each category A neutrophilsurvival index was generated by dividing the number ofsurviving neutrophils in each treatment category by thenumber of surviving neutrophils in the untreated controlcategory
28 Assay of GSH Levels in Neutrophils GSH levels inneutrophils were measured by spectrophotometry using an
assay kit from Arbor Assays Briefly uninfected and H37Rv-infected neutrophils (1 times 104) were lysed by the additionof ice cold 5 5-sulfosalicylic acid dehydrate solution (MPBiomedicals) Lysates were centrifuged and supernatantscollected after centrifugation were analyzed for total GSHandoxidizedGSH (GSSG) as permanufacturerrsquos instructionsLevels of free or reduced GSH (rGSH) were calculatedby subtracting measured GSSG concentrations from themeasured total GSH concentrations All GSH measurementswere normalized with total protein levels
29 Assay of Total Protein Levels in Lysates Proteins in theisolated cell lysates were measured by Bradfordrsquos methodusing a Coomassie protein assay reagent (Thermo Scientific)This assay helped to determine the amount of protein in eachwell By dividing the total protein value from the GSH assayit helped to confirm that the variation of GSH is due to thetreatments added
210 Assay of IL-6 and TNF-120572 in Neutrophil SupernatantsLevels of IL-6 and TNF-120572 in supernatants from uninfectedand H37Rv-infected neutrophils were measured by enzymelinked immunosorbent assay (ELISA) (eBioscience)
211 Assay of Free Radicals in Neutrophil Lysates Free radicallevels in neutrophil lysates were determined bymeasuring thelevels of malondialdehyde (MDA) using a colorimetric assaykit from Cayman The MDA levels in the infected groups(M tb-infectedM tb-infected and treated with NAC andMtb-infected and treated with L-GSH) were normalized to thevalues in uninfected neutrophils
212 Fluorescent Microscopic Analysis of M tb PhagosomeAcidification in Neutrophils The effects of GSH-enhancingagents in inducing acidification of M tb-containing phago-somes were determined by quantifying the colocalizationof Fluorescein Isothiocyanate (FITC) labeled H37Rv in thelysotracker labeled acidified compartments by fluorescentmicroscope Lysotracker Red DND99 (Molecular Probes)an acidotropic dye is a weak base conjugated to a redfluorophore Lysotracker freely permeates cell membranesand gets trapped upon protonation in an acidified compart-ment The excitation and emission maxima of lysotrackerare 577 and 592 nm respectively Neutrophils (1 times 104well)were distributed in polylysine coated sterile glass cover slipspositioned in 24-well tissue culture plates and incubated for12 hours for adherence Adhered neutrophils were treatedwith lysotracker (1103 dilution) for 1 hour H37Rvwas labeledwith FITC as follows bacterial cells were incubated with08mgmL FITC for 1 hour The labeled cells were thencentrifuged and the pellet was resuspended in PBS and usedfor infecting the lysotracker-labeled neutrophils Lysotracker-labeled neutrophils were incubatedwith FITC-labeledH37Rvfor 15 hours to allow phagocytosis Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5CO2in the presence and absence of NAC (10mM) or L-
GSH (10 and 20120583M) for 24 hours Infected neutrophils werefixed with 38 paraformaldehyde in PBS for 30 minutes
4 Clinical and Developmental Immunology
0
2
4
6
M tb
Fold
incr
ease
d in
M t
b gr
owth
M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(a)
0
01
02
03
04
05
06
07
Neu
troph
il su
rviv
al in
dex
M tb M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(b)
Figure 1 Intracellular survival of H37Rv in neutrophils treated with NAC and L-GSH (119899 = 5)Neutrophils were infected with processedH37Rvat a low dose multiplicity of infection of 1 10 (1 bacterium for every 10 neutrophils) and incubated for 2 hours to allow for phagocytosisUnphagocytosed mycobacteria were removed by washing the infected neutrophils three times with warm sterile PBS Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in the presence and absence of NAC (10mM) or L-GSH (10 and 20120583M) Infected
neutrophils were terminated at 1 hour and 24 hours after infection to determine the intracellular survival of H37Rv Graph shows mean plusmnstandard error(a) Represents foldincrease in intracellular growth of H37Rv in neutrophils treated with NAC and L-GSH (b) Determinationof neutrophil viability following infection with H37Rv and treatment with NAC and L-GSH by quantifying DAPI stained cells (119899 = 5)
Fixed neutrophils were washed twice for 5 minutes in PBSA single drop of mounting media containing DAPI wasplaced per glass slide The stained neutrophils were thenattached to glass cover slips and inverted onto the glassslide The cover slips were then sealed to the glass slide bysmearing nail polish around the edges of the cover slips Thestained slides were then viewed using an inverted fluorescentmicroscope Images were obtained using an integrated digitalcamera Images were subsequently analyzed using ImageJ afree software program available from the National Institutesof Health (httprsbwebnihgovij) Following backgroundfluorescence correction the average fluorescent intensity wasmeasured
213 Statistical Analysis All statistical analysis was doneusing Graph Pad Prism6 software on the mean plusmn stan-dard error for 119899 = 5 individuals For all assays theinfected-untreated category was compared to the uninfected-untreated control category using a two-tailed Studentrsquos 119905-testwith 95 confidence intervals All infected-treated categorieswere compared to the infected-untreated category using aKruskal-Wallis 1-way ANOVA with Dunnrsquosmultiple compar-isons posttest Results were considered significant for 119875 le005
3 Results
31 Intracellular Survival of H37Rv inside Human NeutrophilsWe tested the effects of GSH-enhancing agents in improv-ing the functions of neutrophils to inhibit the growth ofM tb This was achieved by determining the intracellularsurvival of H37Rv a virulent laboratory strain ofM tb insideNACL-GSH-treated human neutrophils We observed a 4-fold increase in the intracellular growth of H37Rv inside
unstimulated neutrophils (Figure 1(a)) Treatment of neu-trophils with L-GSH at 10 and 20 120583M concentrations resultedin a decrease in the foldgrowth of M tb (Figure 1(a)) Infact we observed maximum inhibition in the growth ofH37Rv in neutrophils that were treated with L-GSH andthis is in contrast to untreated neutrophils in which therewas a 4-fold increase in the growth of intracellular M tb(Figure 1(a)) Interestingly treatment of neutrophils with theGSH-precursor NAC at 50x higher concentration comparedto L-GSH did not reduce the growth of M tb but insteadwe observed a 6-fold increase in the intracellular growth ofH37Rv (Figure 1(a)) These results indicate that an optimalincrease in the intracellular levels of GSH in neutrophilswill lead to inhibition in the growth of M tb Treatment ofH37Rv-infected neutrophils with GSH-enhancing agents didnot result in any loss in the viability of neutrophils as evidentfrom the quantification of DAPI stained cells (Figure 1(b))
32 Assay of GSH Levels in Neutrophils We determined theability of NAC and L-GSH to enhance the intracellular levelsof GSH in uninfected neutrophils and restore the levels ofGSH in H37Rv-infected neutrophils by spectrophotometryWe used an assay kit from Arbor Assays We observed thattreatment of uninfected neutrophils with NAC resulted in3-fold increase in the levels of both total GSH and rGSH(Figure 2) L-GSH treatment did not result in any increasein the levels of both total GSH and rGSH in uninfectedneutrophils (Figure 2) Importantly H37Rv infection resultedin a 20 decrease in the levels of total GSH in neutrophils(Figure 3(a)) The percentages of rGSH versus GSSG inH37Rv-infected neutrophils was 56 and 44 respectively(Figure 3(d)) Whereas the percentage of rGSH and GSSGin uninfected neutrophils were 76 and 24 respectively(Figure 2(d)) Our results signify that M tb infection leads
Clinical and Developmental Immunology 5
Control NAC 10120583M LGSH 20120583M LGSH000
002
004
006
008
010To
tal G
SH120583
g to
tal p
rote
in
(a)
000
002
004
006
008
Control 10mM NAC 10120583M LGSH 20120583M LGSH
120583M
rGSH
120583g
tota
l pro
tein
(b)
0000
0002
0004
0006
0008
00010
Tota
l GSS
G120583
g to
tal p
rote
in
Control 10mM NAC 10120583M LGSH 20120583M LGSH
(c)
24
76
6
94
14
86
24
76
rGSHGSSG
rGSHGSSG
Control 10mM NAC
10120583M LGSH 20120583M LGSH
(d)
Figure 2 GSH measurements in uninfected neutrophils cultured in the presence and absence of NAC or L-GSH (119899 = 5) GSH levelswere measured in isolated neutrophils from healthy subjects that were treated with GSH-enhancing agents such as NAC and L-GSH byspectrophotometry using an assay kit Briefly neutrophils (3 times 105) were lysed at 24 hours after treatment with ice cold 5 5-sulfosalicylicacid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH (a) and GSSG (c) as per manufacturerrsquosinstructions rGSH was calculated by subtracting measured GSSG concentrations from the measured total GSH concentrations (b) All GSHmeasurements were normalized with total protein levels Proteins in the cell lysates of neutrophils were measured by Bradfordrsquos method usinga Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressed as percentage of total GSH in neutrophil lysates Graphshows mean plusmn standard error
to decreased levels of rGSH the functional form of GSHTreatment of H37Rv-infected neutrophils with 10mM NACresulted in a 10-fold increase in the intracellular levels oftotal GSH (Figure 3(a)) Treatment of H37Rv-infected neu-trophils with L-GSH (10 and 20 120583M concentration) resultedin restoration in the intracellular levels of both total GSHand rGSH to similar levels as uninfected neutrophils (Figures3(a) and 3(b)) Treatment ofH37Rv-infected neutrophils withGSH-enhancing agents (NAC and L-GSH) also resulted inreversal of GSSG and rGSH ratio similar to the percentagein uninfected neutrophils (Figure 3(d)) Our results indicatethatM tb infection decreases the levels of GSH in neutrophilspossibly by inducing oxidative stress (as evident from theincreased percentage of GSSG) and treatment of M tb-infected neutrophils with GSH-enhancing agents restores theintracellular levels of both total GSH and rGSH
33 Determination of Free Radicals Levels in Neutrophils Wedetermined the inverse correlation of GSH levels and ROIby quantifying the levels of free radicals in H37Rv-infected
neutrophils We observed that infection of neutrophils withH37Rv resulted in a 3-fold increase in the production offree radicals compared to the uninfected neutrophils (Fig-ure 4(a)) NAC and L-GSH treatments led to a noticeabledecrease in the levels of free radicals The decreased levelswere similar to the uninfected neutrophils (Figure 4(a))Maximum decrease in the levels of free radicals was observedwhen H37Rv-infected neutrophils were treated with 20120583ML-GSH (Figure 4(a)) Our results highlight the ability of Mtb infection to induce neutrophils to produce free radicalsleading to a decrease in the levels of rGSH
34 TNF-120572 Assay in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 28-fold increase inthe production of TNF-120572 compared to uninfected neutrophils(Figure 4(b)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a dramatic decrease in the productionof TNF-120572 In fact the TNF-120572 levels were undetectable insupernatants collected from H37Rv-infected +NAC-treatedneutrophils (Figure 4(b)) These results indicate that NAC is
6 Clinical and Developmental Immunology
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
005
010
015120583
M to
tal G
SH120583
g to
tal p
rote
in
lowast
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
004
002
006
008
010
120583M
tota
l GSH
120583g
tota
l pro
tein lowast
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
002
001
003
004
005
120583M
tota
l GSS
G120583
g to
tal p
rote
in
(c)
56
20
80
26
74
26
74
rGSHGSSG
rGSHGSSG
44
M tb
10120583M LGSH 20120583M LGSH
M tb +
M tb + M tb +
10 mM NAC
(d)
Figure 3 GSH measurements in neutrophils infected with H37Rv and treated with NAC and L-GSH (119899 = 5) GSH levels were measured inisolated neutrophils from healthy subjects that were infected in vitro with H37Rv and treated with GSH-enhancing agents such as NAC andL-GSH by spectrophotometry using an assay kit Briefly H37Rv-infected neutrophils (3 times 105) were lysed at 24 hours after infection withice cold 5 5-sulfosalicylic acid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH ((a) lowast119875 le 005)and GSSG (c) as per manufacturerrsquos instructions rGSH ((b) lowast119875 le 005) was calculated by subtracting measured GSSG concentrations fromthe measured total GSH concentrations All GSH measurements were normalized with total protein levels Proteins in the cell lysates ofneutrophils were measured by Bradfordrsquos method using a Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressedas percentage of total GSH in neutrophil lysates Graph shows mean plusmn standard error
capable of completely diminishing the production of TNF-120572byH37Rv-infected neutrophils Treatment ofH37Rv-infectedneutrophils with 10 and 20 120583M L-GSH did not result in anydecrease in the production of TNF-120572 compared to untreatedH37Rv-infected neutrophils (Figure 4(b))
35 Assay of IL-6 in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 38-fold increase inthe production of IL-6 compared to uninfected neutrophils(Figure 4(c)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a striking decrease in the productionof IL-6 (Figure 4(c)) IL-6 levels were undetectable in super-natants collected from H37Rv-infected + NAC-treated neu-trophils Treatment of H37Rv-infected neutrophils with L-GSH resulted did not result in any significant decrease in theproduction of IL-6 compared to untreated H37Rv-infected
neutrophils (Figure 4(c)) These results indicate that NACtreatment at millimolar concentrations is capable of com-pletely diminishing the production of both IL-6 and TNF-120572by H37Rv-infected neutrophils (Figures 4(b) and 4(c))
36 Examining the Acidification of M tb Containing Phago-somes by Quantifying the Colocalization of FITC-LabeledH37Rv with Lysotracker We examined the effects of GSHon increasing the acidification of phagosomes containingMtb by quantifying the colocalization of FITC-labeled H37Rvwith lysotracker trapped in the acidified compartments inthe neutrophils (Figure 5) We observed that treatment ofH37Rv-infected neutrophils with 10120583M L-GSH resulted in40 increase in the colocalization of FITC-labeled H37Rvwith lysotracker (Figure 4(d)) Interestingly treatment ofH37Rv-infected neutrophils with 20120583M L-GSH resulted in a
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010

Clinical and Developmental Immunology 3
M tb infection we determined the intracellular survival ofM tb inside neutrophils that were cultured in the presenceand absence of GSH-enhancing agents Neutrophils wereinfected with processed H37Rv at a low dose multiplicityof infection of 1 10 (1 bacterium for every 10 neutrophils)and incubated for 2 hours to allow for phagocytosis Un-phagocytosed bacteria were removed by washing the infectedneutrophil cultures three times with warm sterile PBS (basedon the results from our standardization studies this low dosemultiplicity of infection usually resulted in approximately10of the neutrophils infectedwithH37Rv and each infectedcell had between 1 and 5 bacteria) Infected neutrophils werecultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in
the presence and absence of NAC (10mM) or L-GSH (10and 20120583M Your Energy Systems LLC Palo Alto CA) Theliposomal formulation of GSH (L-GSH) used in these studiesis a proprietary product of Your Energy Systems (YES) LLC(Palo Alto CA) called ReadiSorb Infected neutrophils wereterminated at 1 hour and 24 hours after infection to determinethe intracellular survival of H37Rv
26 Termination of Infected Neutrophils and Measurementof Colony Forming Units (CFU) Termination of H37Rv-infected neutrophils was performed by the addition of 200120583Lof sterile distilled water to each culture well The collectedneutrophil lysates were diluted in sterile water and plated on7H11 medium (Hi Media) enriched with albumin dextrosecomplex (ADC) to estimate the extent of H37Rv growth inneutrophils by counting the CFUs
27 Neutrophil Viability Assay Theeffects of GSH-enhancingagents in altering the viability of H37Rv-infected neutrophilswere determined by fluorescent DAPI staining Neutrophils(1 times 104well) were distributed in sterile glass cover slipspositioned in 24-well tissue culture plates and incubated for12 hours for adherence Adhered neutrophils were infectedH37Rv at a multiplicity of infection of 1 10 (1 bacteria to 10neutrophils) Infected neutrophils were cultured in RPMI +5 AB serum at 37∘C + 5 CO
2in the presence and absence
of NAC (10mM) or L-GSH (10 and 20120583M) for 24 hoursInfected neutrophils were fixed with 38 paraformaldehydein PBS for 30minutes Fixed neutrophils were washed twicefor 5minutes in PBS A single drop of mounting mediacontaining DAPI was placed per glass slide The stainedneutrophils were then attached to glass cover slips andinverted onto the glass slide The cover slips were thensealed to the glass slide by smearing nail polish around theedges of the cover slips The stained slides were then viewedusing an inverted fluorescent microscope Intact cells withDAPI staining were counted under 20x magnification in7 randomly selected fields for each category A neutrophilsurvival index was generated by dividing the number ofsurviving neutrophils in each treatment category by thenumber of surviving neutrophils in the untreated controlcategory
28 Assay of GSH Levels in Neutrophils GSH levels inneutrophils were measured by spectrophotometry using an
assay kit from Arbor Assays Briefly uninfected and H37Rv-infected neutrophils (1 times 104) were lysed by the additionof ice cold 5 5-sulfosalicylic acid dehydrate solution (MPBiomedicals) Lysates were centrifuged and supernatantscollected after centrifugation were analyzed for total GSHandoxidizedGSH (GSSG) as permanufacturerrsquos instructionsLevels of free or reduced GSH (rGSH) were calculatedby subtracting measured GSSG concentrations from themeasured total GSH concentrations All GSH measurementswere normalized with total protein levels
29 Assay of Total Protein Levels in Lysates Proteins in theisolated cell lysates were measured by Bradfordrsquos methodusing a Coomassie protein assay reagent (Thermo Scientific)This assay helped to determine the amount of protein in eachwell By dividing the total protein value from the GSH assayit helped to confirm that the variation of GSH is due to thetreatments added
210 Assay of IL-6 and TNF-120572 in Neutrophil SupernatantsLevels of IL-6 and TNF-120572 in supernatants from uninfectedand H37Rv-infected neutrophils were measured by enzymelinked immunosorbent assay (ELISA) (eBioscience)
211 Assay of Free Radicals in Neutrophil Lysates Free radicallevels in neutrophil lysates were determined bymeasuring thelevels of malondialdehyde (MDA) using a colorimetric assaykit from Cayman The MDA levels in the infected groups(M tb-infectedM tb-infected and treated with NAC andMtb-infected and treated with L-GSH) were normalized to thevalues in uninfected neutrophils
212 Fluorescent Microscopic Analysis of M tb PhagosomeAcidification in Neutrophils The effects of GSH-enhancingagents in inducing acidification of M tb-containing phago-somes were determined by quantifying the colocalizationof Fluorescein Isothiocyanate (FITC) labeled H37Rv in thelysotracker labeled acidified compartments by fluorescentmicroscope Lysotracker Red DND99 (Molecular Probes)an acidotropic dye is a weak base conjugated to a redfluorophore Lysotracker freely permeates cell membranesand gets trapped upon protonation in an acidified compart-ment The excitation and emission maxima of lysotrackerare 577 and 592 nm respectively Neutrophils (1 times 104well)were distributed in polylysine coated sterile glass cover slipspositioned in 24-well tissue culture plates and incubated for12 hours for adherence Adhered neutrophils were treatedwith lysotracker (1103 dilution) for 1 hour H37Rvwas labeledwith FITC as follows bacterial cells were incubated with08mgmL FITC for 1 hour The labeled cells were thencentrifuged and the pellet was resuspended in PBS and usedfor infecting the lysotracker-labeled neutrophils Lysotracker-labeled neutrophils were incubatedwith FITC-labeledH37Rvfor 15 hours to allow phagocytosis Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5CO2in the presence and absence of NAC (10mM) or L-
GSH (10 and 20120583M) for 24 hours Infected neutrophils werefixed with 38 paraformaldehyde in PBS for 30 minutes
4 Clinical and Developmental Immunology
0
2
4
6
M tb
Fold
incr
ease
d in
M t
b gr
owth
M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(a)
0
01
02
03
04
05
06
07
Neu
troph
il su
rviv
al in
dex
M tb M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(b)
Figure 1 Intracellular survival of H37Rv in neutrophils treated with NAC and L-GSH (119899 = 5)Neutrophils were infected with processedH37Rvat a low dose multiplicity of infection of 1 10 (1 bacterium for every 10 neutrophils) and incubated for 2 hours to allow for phagocytosisUnphagocytosed mycobacteria were removed by washing the infected neutrophils three times with warm sterile PBS Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in the presence and absence of NAC (10mM) or L-GSH (10 and 20120583M) Infected
neutrophils were terminated at 1 hour and 24 hours after infection to determine the intracellular survival of H37Rv Graph shows mean plusmnstandard error(a) Represents foldincrease in intracellular growth of H37Rv in neutrophils treated with NAC and L-GSH (b) Determinationof neutrophil viability following infection with H37Rv and treatment with NAC and L-GSH by quantifying DAPI stained cells (119899 = 5)
Fixed neutrophils were washed twice for 5 minutes in PBSA single drop of mounting media containing DAPI wasplaced per glass slide The stained neutrophils were thenattached to glass cover slips and inverted onto the glassslide The cover slips were then sealed to the glass slide bysmearing nail polish around the edges of the cover slips Thestained slides were then viewed using an inverted fluorescentmicroscope Images were obtained using an integrated digitalcamera Images were subsequently analyzed using ImageJ afree software program available from the National Institutesof Health (httprsbwebnihgovij) Following backgroundfluorescence correction the average fluorescent intensity wasmeasured
213 Statistical Analysis All statistical analysis was doneusing Graph Pad Prism6 software on the mean plusmn stan-dard error for 119899 = 5 individuals For all assays theinfected-untreated category was compared to the uninfected-untreated control category using a two-tailed Studentrsquos 119905-testwith 95 confidence intervals All infected-treated categorieswere compared to the infected-untreated category using aKruskal-Wallis 1-way ANOVA with Dunnrsquosmultiple compar-isons posttest Results were considered significant for 119875 le005
3 Results
31 Intracellular Survival of H37Rv inside Human NeutrophilsWe tested the effects of GSH-enhancing agents in improv-ing the functions of neutrophils to inhibit the growth ofM tb This was achieved by determining the intracellularsurvival of H37Rv a virulent laboratory strain ofM tb insideNACL-GSH-treated human neutrophils We observed a 4-fold increase in the intracellular growth of H37Rv inside
unstimulated neutrophils (Figure 1(a)) Treatment of neu-trophils with L-GSH at 10 and 20 120583M concentrations resultedin a decrease in the foldgrowth of M tb (Figure 1(a)) Infact we observed maximum inhibition in the growth ofH37Rv in neutrophils that were treated with L-GSH andthis is in contrast to untreated neutrophils in which therewas a 4-fold increase in the growth of intracellular M tb(Figure 1(a)) Interestingly treatment of neutrophils with theGSH-precursor NAC at 50x higher concentration comparedto L-GSH did not reduce the growth of M tb but insteadwe observed a 6-fold increase in the intracellular growth ofH37Rv (Figure 1(a)) These results indicate that an optimalincrease in the intracellular levels of GSH in neutrophilswill lead to inhibition in the growth of M tb Treatment ofH37Rv-infected neutrophils with GSH-enhancing agents didnot result in any loss in the viability of neutrophils as evidentfrom the quantification of DAPI stained cells (Figure 1(b))
32 Assay of GSH Levels in Neutrophils We determined theability of NAC and L-GSH to enhance the intracellular levelsof GSH in uninfected neutrophils and restore the levels ofGSH in H37Rv-infected neutrophils by spectrophotometryWe used an assay kit from Arbor Assays We observed thattreatment of uninfected neutrophils with NAC resulted in3-fold increase in the levels of both total GSH and rGSH(Figure 2) L-GSH treatment did not result in any increasein the levels of both total GSH and rGSH in uninfectedneutrophils (Figure 2) Importantly H37Rv infection resultedin a 20 decrease in the levels of total GSH in neutrophils(Figure 3(a)) The percentages of rGSH versus GSSG inH37Rv-infected neutrophils was 56 and 44 respectively(Figure 3(d)) Whereas the percentage of rGSH and GSSGin uninfected neutrophils were 76 and 24 respectively(Figure 2(d)) Our results signify that M tb infection leads
Clinical and Developmental Immunology 5
Control NAC 10120583M LGSH 20120583M LGSH000
002
004
006
008
010To
tal G
SH120583
g to
tal p
rote
in
(a)
000
002
004
006
008
Control 10mM NAC 10120583M LGSH 20120583M LGSH
120583M
rGSH
120583g
tota
l pro
tein
(b)
0000
0002
0004
0006
0008
00010
Tota
l GSS
G120583
g to
tal p
rote
in
Control 10mM NAC 10120583M LGSH 20120583M LGSH
(c)
24
76
6
94
14
86
24
76
rGSHGSSG
rGSHGSSG
Control 10mM NAC
10120583M LGSH 20120583M LGSH
(d)
Figure 2 GSH measurements in uninfected neutrophils cultured in the presence and absence of NAC or L-GSH (119899 = 5) GSH levelswere measured in isolated neutrophils from healthy subjects that were treated with GSH-enhancing agents such as NAC and L-GSH byspectrophotometry using an assay kit Briefly neutrophils (3 times 105) were lysed at 24 hours after treatment with ice cold 5 5-sulfosalicylicacid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH (a) and GSSG (c) as per manufacturerrsquosinstructions rGSH was calculated by subtracting measured GSSG concentrations from the measured total GSH concentrations (b) All GSHmeasurements were normalized with total protein levels Proteins in the cell lysates of neutrophils were measured by Bradfordrsquos method usinga Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressed as percentage of total GSH in neutrophil lysates Graphshows mean plusmn standard error
to decreased levels of rGSH the functional form of GSHTreatment of H37Rv-infected neutrophils with 10mM NACresulted in a 10-fold increase in the intracellular levels oftotal GSH (Figure 3(a)) Treatment of H37Rv-infected neu-trophils with L-GSH (10 and 20 120583M concentration) resultedin restoration in the intracellular levels of both total GSHand rGSH to similar levels as uninfected neutrophils (Figures3(a) and 3(b)) Treatment ofH37Rv-infected neutrophils withGSH-enhancing agents (NAC and L-GSH) also resulted inreversal of GSSG and rGSH ratio similar to the percentagein uninfected neutrophils (Figure 3(d)) Our results indicatethatM tb infection decreases the levels of GSH in neutrophilspossibly by inducing oxidative stress (as evident from theincreased percentage of GSSG) and treatment of M tb-infected neutrophils with GSH-enhancing agents restores theintracellular levels of both total GSH and rGSH
33 Determination of Free Radicals Levels in Neutrophils Wedetermined the inverse correlation of GSH levels and ROIby quantifying the levels of free radicals in H37Rv-infected
neutrophils We observed that infection of neutrophils withH37Rv resulted in a 3-fold increase in the production offree radicals compared to the uninfected neutrophils (Fig-ure 4(a)) NAC and L-GSH treatments led to a noticeabledecrease in the levels of free radicals The decreased levelswere similar to the uninfected neutrophils (Figure 4(a))Maximum decrease in the levels of free radicals was observedwhen H37Rv-infected neutrophils were treated with 20120583ML-GSH (Figure 4(a)) Our results highlight the ability of Mtb infection to induce neutrophils to produce free radicalsleading to a decrease in the levels of rGSH
34 TNF-120572 Assay in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 28-fold increase inthe production of TNF-120572 compared to uninfected neutrophils(Figure 4(b)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a dramatic decrease in the productionof TNF-120572 In fact the TNF-120572 levels were undetectable insupernatants collected from H37Rv-infected +NAC-treatedneutrophils (Figure 4(b)) These results indicate that NAC is
6 Clinical and Developmental Immunology
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
005
010
015120583
M to
tal G
SH120583
g to
tal p
rote
in
lowast
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
004
002
006
008
010
120583M
tota
l GSH
120583g
tota
l pro
tein lowast
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
002
001
003
004
005
120583M
tota
l GSS
G120583
g to
tal p
rote
in
(c)
56
20
80
26
74
26
74
rGSHGSSG
rGSHGSSG
44
M tb
10120583M LGSH 20120583M LGSH
M tb +
M tb + M tb +
10 mM NAC
(d)
Figure 3 GSH measurements in neutrophils infected with H37Rv and treated with NAC and L-GSH (119899 = 5) GSH levels were measured inisolated neutrophils from healthy subjects that were infected in vitro with H37Rv and treated with GSH-enhancing agents such as NAC andL-GSH by spectrophotometry using an assay kit Briefly H37Rv-infected neutrophils (3 times 105) were lysed at 24 hours after infection withice cold 5 5-sulfosalicylic acid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH ((a) lowast119875 le 005)and GSSG (c) as per manufacturerrsquos instructions rGSH ((b) lowast119875 le 005) was calculated by subtracting measured GSSG concentrations fromthe measured total GSH concentrations All GSH measurements were normalized with total protein levels Proteins in the cell lysates ofneutrophils were measured by Bradfordrsquos method using a Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressedas percentage of total GSH in neutrophil lysates Graph shows mean plusmn standard error
capable of completely diminishing the production of TNF-120572byH37Rv-infected neutrophils Treatment ofH37Rv-infectedneutrophils with 10 and 20 120583M L-GSH did not result in anydecrease in the production of TNF-120572 compared to untreatedH37Rv-infected neutrophils (Figure 4(b))
35 Assay of IL-6 in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 38-fold increase inthe production of IL-6 compared to uninfected neutrophils(Figure 4(c)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a striking decrease in the productionof IL-6 (Figure 4(c)) IL-6 levels were undetectable in super-natants collected from H37Rv-infected + NAC-treated neu-trophils Treatment of H37Rv-infected neutrophils with L-GSH resulted did not result in any significant decrease in theproduction of IL-6 compared to untreated H37Rv-infected
neutrophils (Figure 4(c)) These results indicate that NACtreatment at millimolar concentrations is capable of com-pletely diminishing the production of both IL-6 and TNF-120572by H37Rv-infected neutrophils (Figures 4(b) and 4(c))
36 Examining the Acidification of M tb Containing Phago-somes by Quantifying the Colocalization of FITC-LabeledH37Rv with Lysotracker We examined the effects of GSHon increasing the acidification of phagosomes containingMtb by quantifying the colocalization of FITC-labeled H37Rvwith lysotracker trapped in the acidified compartments inthe neutrophils (Figure 5) We observed that treatment ofH37Rv-infected neutrophils with 10120583M L-GSH resulted in40 increase in the colocalization of FITC-labeled H37Rvwith lysotracker (Figure 4(d)) Interestingly treatment ofH37Rv-infected neutrophils with 20120583M L-GSH resulted in a
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010
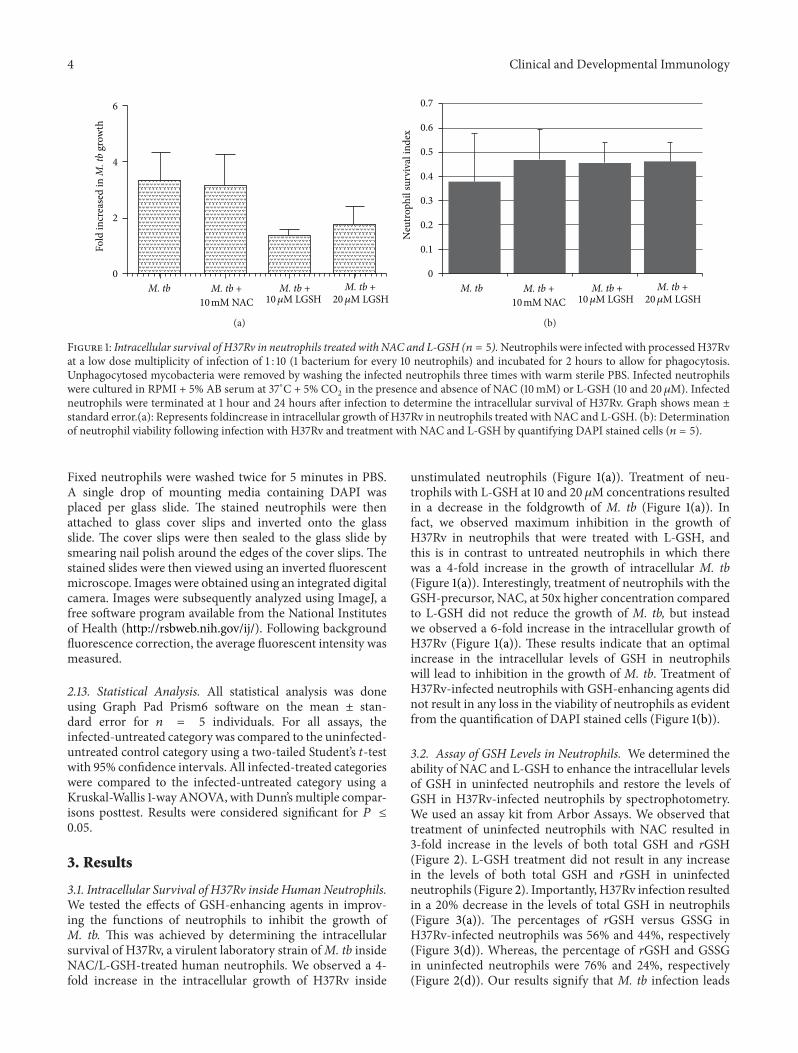
4 Clinical and Developmental Immunology
0
2
4
6
M tb
Fold
incr
ease
d in
M t
b gr
owth
M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(a)
0
01
02
03
04
05
06
07
Neu
troph
il su
rviv
al in
dex
M tb M tb +10120583M LGSH
M tb +20120583M LGSH
M tb +10 mM NAC
(b)
Figure 1 Intracellular survival of H37Rv in neutrophils treated with NAC and L-GSH (119899 = 5)Neutrophils were infected with processedH37Rvat a low dose multiplicity of infection of 1 10 (1 bacterium for every 10 neutrophils) and incubated for 2 hours to allow for phagocytosisUnphagocytosed mycobacteria were removed by washing the infected neutrophils three times with warm sterile PBS Infected neutrophilswere cultured in RPMI + 5 AB serum at 37∘C + 5 CO
2in the presence and absence of NAC (10mM) or L-GSH (10 and 20120583M) Infected
neutrophils were terminated at 1 hour and 24 hours after infection to determine the intracellular survival of H37Rv Graph shows mean plusmnstandard error(a) Represents foldincrease in intracellular growth of H37Rv in neutrophils treated with NAC and L-GSH (b) Determinationof neutrophil viability following infection with H37Rv and treatment with NAC and L-GSH by quantifying DAPI stained cells (119899 = 5)
Fixed neutrophils were washed twice for 5 minutes in PBSA single drop of mounting media containing DAPI wasplaced per glass slide The stained neutrophils were thenattached to glass cover slips and inverted onto the glassslide The cover slips were then sealed to the glass slide bysmearing nail polish around the edges of the cover slips Thestained slides were then viewed using an inverted fluorescentmicroscope Images were obtained using an integrated digitalcamera Images were subsequently analyzed using ImageJ afree software program available from the National Institutesof Health (httprsbwebnihgovij) Following backgroundfluorescence correction the average fluorescent intensity wasmeasured
213 Statistical Analysis All statistical analysis was doneusing Graph Pad Prism6 software on the mean plusmn stan-dard error for 119899 = 5 individuals For all assays theinfected-untreated category was compared to the uninfected-untreated control category using a two-tailed Studentrsquos 119905-testwith 95 confidence intervals All infected-treated categorieswere compared to the infected-untreated category using aKruskal-Wallis 1-way ANOVA with Dunnrsquosmultiple compar-isons posttest Results were considered significant for 119875 le005
3 Results
31 Intracellular Survival of H37Rv inside Human NeutrophilsWe tested the effects of GSH-enhancing agents in improv-ing the functions of neutrophils to inhibit the growth ofM tb This was achieved by determining the intracellularsurvival of H37Rv a virulent laboratory strain ofM tb insideNACL-GSH-treated human neutrophils We observed a 4-fold increase in the intracellular growth of H37Rv inside
unstimulated neutrophils (Figure 1(a)) Treatment of neu-trophils with L-GSH at 10 and 20 120583M concentrations resultedin a decrease in the foldgrowth of M tb (Figure 1(a)) Infact we observed maximum inhibition in the growth ofH37Rv in neutrophils that were treated with L-GSH andthis is in contrast to untreated neutrophils in which therewas a 4-fold increase in the growth of intracellular M tb(Figure 1(a)) Interestingly treatment of neutrophils with theGSH-precursor NAC at 50x higher concentration comparedto L-GSH did not reduce the growth of M tb but insteadwe observed a 6-fold increase in the intracellular growth ofH37Rv (Figure 1(a)) These results indicate that an optimalincrease in the intracellular levels of GSH in neutrophilswill lead to inhibition in the growth of M tb Treatment ofH37Rv-infected neutrophils with GSH-enhancing agents didnot result in any loss in the viability of neutrophils as evidentfrom the quantification of DAPI stained cells (Figure 1(b))
32 Assay of GSH Levels in Neutrophils We determined theability of NAC and L-GSH to enhance the intracellular levelsof GSH in uninfected neutrophils and restore the levels ofGSH in H37Rv-infected neutrophils by spectrophotometryWe used an assay kit from Arbor Assays We observed thattreatment of uninfected neutrophils with NAC resulted in3-fold increase in the levels of both total GSH and rGSH(Figure 2) L-GSH treatment did not result in any increasein the levels of both total GSH and rGSH in uninfectedneutrophils (Figure 2) Importantly H37Rv infection resultedin a 20 decrease in the levels of total GSH in neutrophils(Figure 3(a)) The percentages of rGSH versus GSSG inH37Rv-infected neutrophils was 56 and 44 respectively(Figure 3(d)) Whereas the percentage of rGSH and GSSGin uninfected neutrophils were 76 and 24 respectively(Figure 2(d)) Our results signify that M tb infection leads
Clinical and Developmental Immunology 5
Control NAC 10120583M LGSH 20120583M LGSH000
002
004
006
008
010To
tal G
SH120583
g to
tal p
rote
in
(a)
000
002
004
006
008
Control 10mM NAC 10120583M LGSH 20120583M LGSH
120583M
rGSH
120583g
tota
l pro
tein
(b)
0000
0002
0004
0006
0008
00010
Tota
l GSS
G120583
g to
tal p
rote
in
Control 10mM NAC 10120583M LGSH 20120583M LGSH
(c)
24
76
6
94
14
86
24
76
rGSHGSSG
rGSHGSSG
Control 10mM NAC
10120583M LGSH 20120583M LGSH
(d)
Figure 2 GSH measurements in uninfected neutrophils cultured in the presence and absence of NAC or L-GSH (119899 = 5) GSH levelswere measured in isolated neutrophils from healthy subjects that were treated with GSH-enhancing agents such as NAC and L-GSH byspectrophotometry using an assay kit Briefly neutrophils (3 times 105) were lysed at 24 hours after treatment with ice cold 5 5-sulfosalicylicacid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH (a) and GSSG (c) as per manufacturerrsquosinstructions rGSH was calculated by subtracting measured GSSG concentrations from the measured total GSH concentrations (b) All GSHmeasurements were normalized with total protein levels Proteins in the cell lysates of neutrophils were measured by Bradfordrsquos method usinga Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressed as percentage of total GSH in neutrophil lysates Graphshows mean plusmn standard error
to decreased levels of rGSH the functional form of GSHTreatment of H37Rv-infected neutrophils with 10mM NACresulted in a 10-fold increase in the intracellular levels oftotal GSH (Figure 3(a)) Treatment of H37Rv-infected neu-trophils with L-GSH (10 and 20 120583M concentration) resultedin restoration in the intracellular levels of both total GSHand rGSH to similar levels as uninfected neutrophils (Figures3(a) and 3(b)) Treatment ofH37Rv-infected neutrophils withGSH-enhancing agents (NAC and L-GSH) also resulted inreversal of GSSG and rGSH ratio similar to the percentagein uninfected neutrophils (Figure 3(d)) Our results indicatethatM tb infection decreases the levels of GSH in neutrophilspossibly by inducing oxidative stress (as evident from theincreased percentage of GSSG) and treatment of M tb-infected neutrophils with GSH-enhancing agents restores theintracellular levels of both total GSH and rGSH
33 Determination of Free Radicals Levels in Neutrophils Wedetermined the inverse correlation of GSH levels and ROIby quantifying the levels of free radicals in H37Rv-infected
neutrophils We observed that infection of neutrophils withH37Rv resulted in a 3-fold increase in the production offree radicals compared to the uninfected neutrophils (Fig-ure 4(a)) NAC and L-GSH treatments led to a noticeabledecrease in the levels of free radicals The decreased levelswere similar to the uninfected neutrophils (Figure 4(a))Maximum decrease in the levels of free radicals was observedwhen H37Rv-infected neutrophils were treated with 20120583ML-GSH (Figure 4(a)) Our results highlight the ability of Mtb infection to induce neutrophils to produce free radicalsleading to a decrease in the levels of rGSH
34 TNF-120572 Assay in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 28-fold increase inthe production of TNF-120572 compared to uninfected neutrophils(Figure 4(b)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a dramatic decrease in the productionof TNF-120572 In fact the TNF-120572 levels were undetectable insupernatants collected from H37Rv-infected +NAC-treatedneutrophils (Figure 4(b)) These results indicate that NAC is
6 Clinical and Developmental Immunology
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
005
010
015120583
M to
tal G
SH120583
g to
tal p
rote
in
lowast
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
004
002
006
008
010
120583M
tota
l GSH
120583g
tota
l pro
tein lowast
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
002
001
003
004
005
120583M
tota
l GSS
G120583
g to
tal p
rote
in
(c)
56
20
80
26
74
26
74
rGSHGSSG
rGSHGSSG
44
M tb
10120583M LGSH 20120583M LGSH
M tb +
M tb + M tb +
10 mM NAC
(d)
Figure 3 GSH measurements in neutrophils infected with H37Rv and treated with NAC and L-GSH (119899 = 5) GSH levels were measured inisolated neutrophils from healthy subjects that were infected in vitro with H37Rv and treated with GSH-enhancing agents such as NAC andL-GSH by spectrophotometry using an assay kit Briefly H37Rv-infected neutrophils (3 times 105) were lysed at 24 hours after infection withice cold 5 5-sulfosalicylic acid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH ((a) lowast119875 le 005)and GSSG (c) as per manufacturerrsquos instructions rGSH ((b) lowast119875 le 005) was calculated by subtracting measured GSSG concentrations fromthe measured total GSH concentrations All GSH measurements were normalized with total protein levels Proteins in the cell lysates ofneutrophils were measured by Bradfordrsquos method using a Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressedas percentage of total GSH in neutrophil lysates Graph shows mean plusmn standard error
capable of completely diminishing the production of TNF-120572byH37Rv-infected neutrophils Treatment ofH37Rv-infectedneutrophils with 10 and 20 120583M L-GSH did not result in anydecrease in the production of TNF-120572 compared to untreatedH37Rv-infected neutrophils (Figure 4(b))
35 Assay of IL-6 in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 38-fold increase inthe production of IL-6 compared to uninfected neutrophils(Figure 4(c)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a striking decrease in the productionof IL-6 (Figure 4(c)) IL-6 levels were undetectable in super-natants collected from H37Rv-infected + NAC-treated neu-trophils Treatment of H37Rv-infected neutrophils with L-GSH resulted did not result in any significant decrease in theproduction of IL-6 compared to untreated H37Rv-infected
neutrophils (Figure 4(c)) These results indicate that NACtreatment at millimolar concentrations is capable of com-pletely diminishing the production of both IL-6 and TNF-120572by H37Rv-infected neutrophils (Figures 4(b) and 4(c))
36 Examining the Acidification of M tb Containing Phago-somes by Quantifying the Colocalization of FITC-LabeledH37Rv with Lysotracker We examined the effects of GSHon increasing the acidification of phagosomes containingMtb by quantifying the colocalization of FITC-labeled H37Rvwith lysotracker trapped in the acidified compartments inthe neutrophils (Figure 5) We observed that treatment ofH37Rv-infected neutrophils with 10120583M L-GSH resulted in40 increase in the colocalization of FITC-labeled H37Rvwith lysotracker (Figure 4(d)) Interestingly treatment ofH37Rv-infected neutrophils with 20120583M L-GSH resulted in a
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010
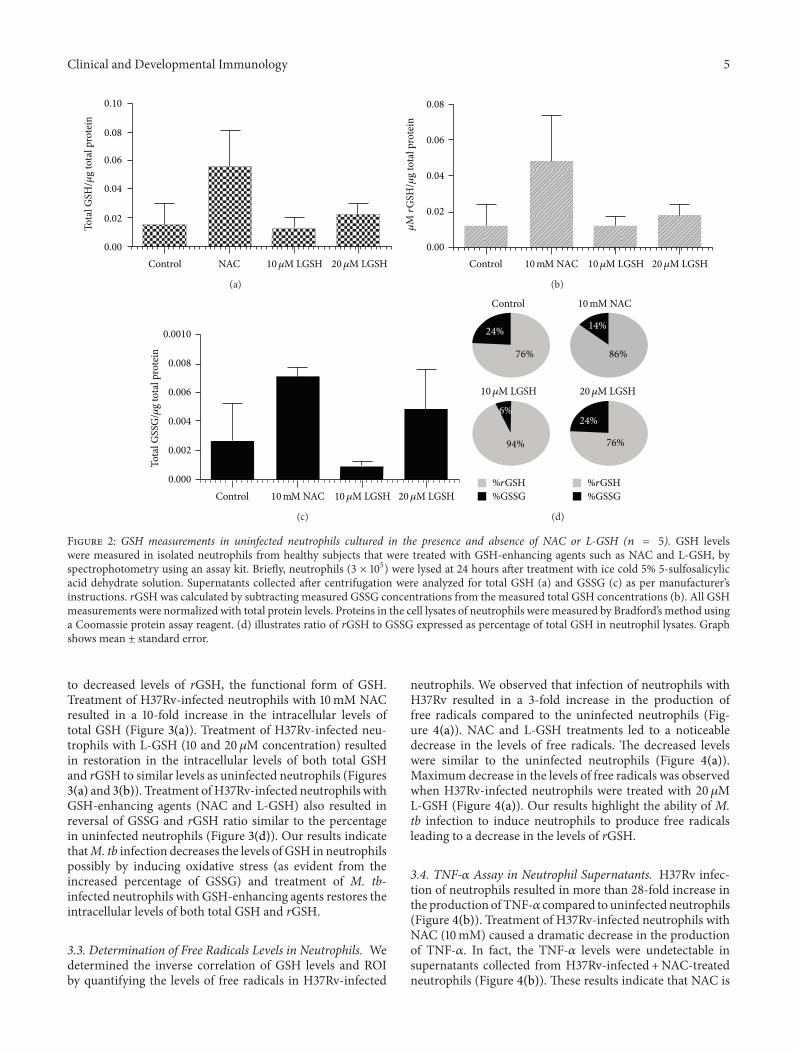
Clinical and Developmental Immunology 5
Control NAC 10120583M LGSH 20120583M LGSH000
002
004
006
008
010To
tal G
SH120583
g to
tal p
rote
in
(a)
000
002
004
006
008
Control 10mM NAC 10120583M LGSH 20120583M LGSH
120583M
rGSH
120583g
tota
l pro
tein
(b)
0000
0002
0004
0006
0008
00010
Tota
l GSS
G120583
g to
tal p
rote
in
Control 10mM NAC 10120583M LGSH 20120583M LGSH
(c)
24
76
6
94
14
86
24
76
rGSHGSSG
rGSHGSSG
Control 10mM NAC
10120583M LGSH 20120583M LGSH
(d)
Figure 2 GSH measurements in uninfected neutrophils cultured in the presence and absence of NAC or L-GSH (119899 = 5) GSH levelswere measured in isolated neutrophils from healthy subjects that were treated with GSH-enhancing agents such as NAC and L-GSH byspectrophotometry using an assay kit Briefly neutrophils (3 times 105) were lysed at 24 hours after treatment with ice cold 5 5-sulfosalicylicacid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH (a) and GSSG (c) as per manufacturerrsquosinstructions rGSH was calculated by subtracting measured GSSG concentrations from the measured total GSH concentrations (b) All GSHmeasurements were normalized with total protein levels Proteins in the cell lysates of neutrophils were measured by Bradfordrsquos method usinga Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressed as percentage of total GSH in neutrophil lysates Graphshows mean plusmn standard error
to decreased levels of rGSH the functional form of GSHTreatment of H37Rv-infected neutrophils with 10mM NACresulted in a 10-fold increase in the intracellular levels oftotal GSH (Figure 3(a)) Treatment of H37Rv-infected neu-trophils with L-GSH (10 and 20 120583M concentration) resultedin restoration in the intracellular levels of both total GSHand rGSH to similar levels as uninfected neutrophils (Figures3(a) and 3(b)) Treatment ofH37Rv-infected neutrophils withGSH-enhancing agents (NAC and L-GSH) also resulted inreversal of GSSG and rGSH ratio similar to the percentagein uninfected neutrophils (Figure 3(d)) Our results indicatethatM tb infection decreases the levels of GSH in neutrophilspossibly by inducing oxidative stress (as evident from theincreased percentage of GSSG) and treatment of M tb-infected neutrophils with GSH-enhancing agents restores theintracellular levels of both total GSH and rGSH
33 Determination of Free Radicals Levels in Neutrophils Wedetermined the inverse correlation of GSH levels and ROIby quantifying the levels of free radicals in H37Rv-infected
neutrophils We observed that infection of neutrophils withH37Rv resulted in a 3-fold increase in the production offree radicals compared to the uninfected neutrophils (Fig-ure 4(a)) NAC and L-GSH treatments led to a noticeabledecrease in the levels of free radicals The decreased levelswere similar to the uninfected neutrophils (Figure 4(a))Maximum decrease in the levels of free radicals was observedwhen H37Rv-infected neutrophils were treated with 20120583ML-GSH (Figure 4(a)) Our results highlight the ability of Mtb infection to induce neutrophils to produce free radicalsleading to a decrease in the levels of rGSH
34 TNF-120572 Assay in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 28-fold increase inthe production of TNF-120572 compared to uninfected neutrophils(Figure 4(b)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a dramatic decrease in the productionof TNF-120572 In fact the TNF-120572 levels were undetectable insupernatants collected from H37Rv-infected +NAC-treatedneutrophils (Figure 4(b)) These results indicate that NAC is
6 Clinical and Developmental Immunology
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
005
010
015120583
M to
tal G
SH120583
g to
tal p
rote
in
lowast
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
004
002
006
008
010
120583M
tota
l GSH
120583g
tota
l pro
tein lowast
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
002
001
003
004
005
120583M
tota
l GSS
G120583
g to
tal p
rote
in
(c)
56
20
80
26
74
26
74
rGSHGSSG
rGSHGSSG
44
M tb
10120583M LGSH 20120583M LGSH
M tb +
M tb + M tb +
10 mM NAC
(d)
Figure 3 GSH measurements in neutrophils infected with H37Rv and treated with NAC and L-GSH (119899 = 5) GSH levels were measured inisolated neutrophils from healthy subjects that were infected in vitro with H37Rv and treated with GSH-enhancing agents such as NAC andL-GSH by spectrophotometry using an assay kit Briefly H37Rv-infected neutrophils (3 times 105) were lysed at 24 hours after infection withice cold 5 5-sulfosalicylic acid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH ((a) lowast119875 le 005)and GSSG (c) as per manufacturerrsquos instructions rGSH ((b) lowast119875 le 005) was calculated by subtracting measured GSSG concentrations fromthe measured total GSH concentrations All GSH measurements were normalized with total protein levels Proteins in the cell lysates ofneutrophils were measured by Bradfordrsquos method using a Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressedas percentage of total GSH in neutrophil lysates Graph shows mean plusmn standard error
capable of completely diminishing the production of TNF-120572byH37Rv-infected neutrophils Treatment ofH37Rv-infectedneutrophils with 10 and 20 120583M L-GSH did not result in anydecrease in the production of TNF-120572 compared to untreatedH37Rv-infected neutrophils (Figure 4(b))
35 Assay of IL-6 in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 38-fold increase inthe production of IL-6 compared to uninfected neutrophils(Figure 4(c)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a striking decrease in the productionof IL-6 (Figure 4(c)) IL-6 levels were undetectable in super-natants collected from H37Rv-infected + NAC-treated neu-trophils Treatment of H37Rv-infected neutrophils with L-GSH resulted did not result in any significant decrease in theproduction of IL-6 compared to untreated H37Rv-infected
neutrophils (Figure 4(c)) These results indicate that NACtreatment at millimolar concentrations is capable of com-pletely diminishing the production of both IL-6 and TNF-120572by H37Rv-infected neutrophils (Figures 4(b) and 4(c))
36 Examining the Acidification of M tb Containing Phago-somes by Quantifying the Colocalization of FITC-LabeledH37Rv with Lysotracker We examined the effects of GSHon increasing the acidification of phagosomes containingMtb by quantifying the colocalization of FITC-labeled H37Rvwith lysotracker trapped in the acidified compartments inthe neutrophils (Figure 5) We observed that treatment ofH37Rv-infected neutrophils with 10120583M L-GSH resulted in40 increase in the colocalization of FITC-labeled H37Rvwith lysotracker (Figure 4(d)) Interestingly treatment ofH37Rv-infected neutrophils with 20120583M L-GSH resulted in a
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010
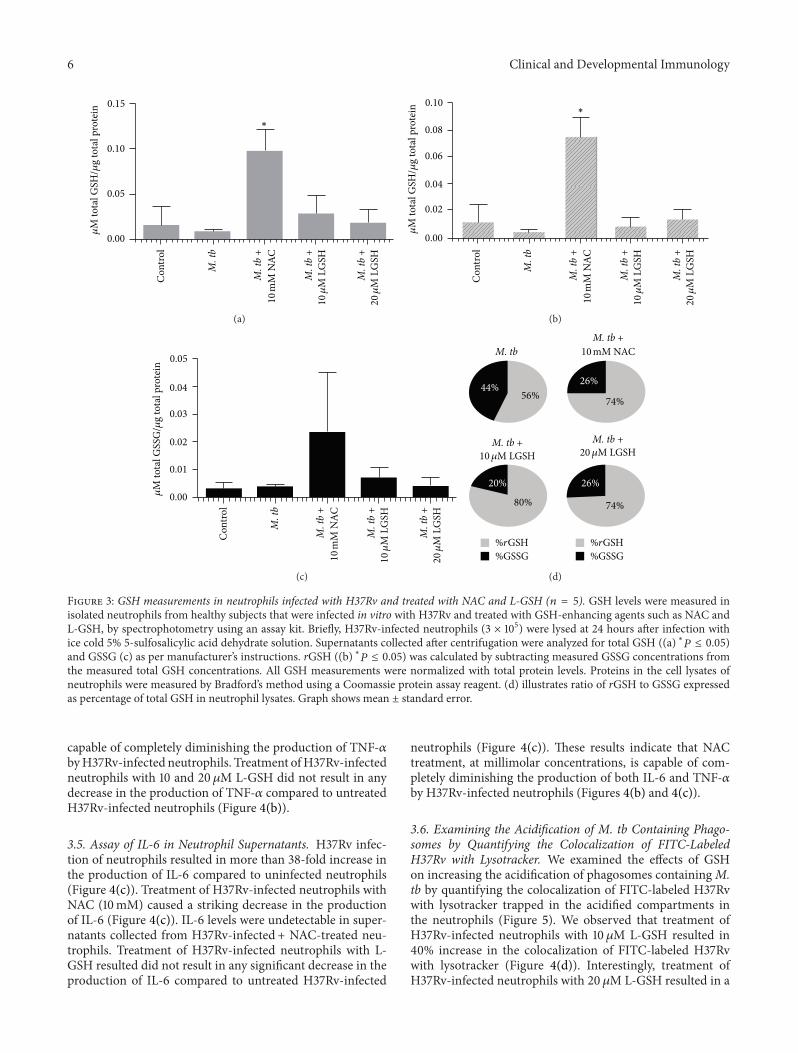
6 Clinical and Developmental Immunology
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
005
010
015120583
M to
tal G
SH120583
g to
tal p
rote
in
lowast
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
004
002
006
008
010
120583M
tota
l GSH
120583g
tota
l pro
tein lowast
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
000
002
001
003
004
005
120583M
tota
l GSS
G120583
g to
tal p
rote
in
(c)
56
20
80
26
74
26
74
rGSHGSSG
rGSHGSSG
44
M tb
10120583M LGSH 20120583M LGSH
M tb +
M tb + M tb +
10 mM NAC
(d)
Figure 3 GSH measurements in neutrophils infected with H37Rv and treated with NAC and L-GSH (119899 = 5) GSH levels were measured inisolated neutrophils from healthy subjects that were infected in vitro with H37Rv and treated with GSH-enhancing agents such as NAC andL-GSH by spectrophotometry using an assay kit Briefly H37Rv-infected neutrophils (3 times 105) were lysed at 24 hours after infection withice cold 5 5-sulfosalicylic acid dehydrate solution Supernatants collected after centrifugation were analyzed for total GSH ((a) lowast119875 le 005)and GSSG (c) as per manufacturerrsquos instructions rGSH ((b) lowast119875 le 005) was calculated by subtracting measured GSSG concentrations fromthe measured total GSH concentrations All GSH measurements were normalized with total protein levels Proteins in the cell lysates ofneutrophils were measured by Bradfordrsquos method using a Coomassie protein assay reagent (d) illustrates ratio of rGSH to GSSG expressedas percentage of total GSH in neutrophil lysates Graph shows mean plusmn standard error
capable of completely diminishing the production of TNF-120572byH37Rv-infected neutrophils Treatment ofH37Rv-infectedneutrophils with 10 and 20 120583M L-GSH did not result in anydecrease in the production of TNF-120572 compared to untreatedH37Rv-infected neutrophils (Figure 4(b))
35 Assay of IL-6 in Neutrophil Supernatants H37Rv infec-tion of neutrophils resulted in more than 38-fold increase inthe production of IL-6 compared to uninfected neutrophils(Figure 4(c)) Treatment of H37Rv-infected neutrophils withNAC (10mM) caused a striking decrease in the productionof IL-6 (Figure 4(c)) IL-6 levels were undetectable in super-natants collected from H37Rv-infected + NAC-treated neu-trophils Treatment of H37Rv-infected neutrophils with L-GSH resulted did not result in any significant decrease in theproduction of IL-6 compared to untreated H37Rv-infected
neutrophils (Figure 4(c)) These results indicate that NACtreatment at millimolar concentrations is capable of com-pletely diminishing the production of both IL-6 and TNF-120572by H37Rv-infected neutrophils (Figures 4(b) and 4(c))
36 Examining the Acidification of M tb Containing Phago-somes by Quantifying the Colocalization of FITC-LabeledH37Rv with Lysotracker We examined the effects of GSHon increasing the acidification of phagosomes containingMtb by quantifying the colocalization of FITC-labeled H37Rvwith lysotracker trapped in the acidified compartments inthe neutrophils (Figure 5) We observed that treatment ofH37Rv-infected neutrophils with 10120583M L-GSH resulted in40 increase in the colocalization of FITC-labeled H37Rvwith lysotracker (Figure 4(d)) Interestingly treatment ofH37Rv-infected neutrophils with 20120583M L-GSH resulted in a
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010
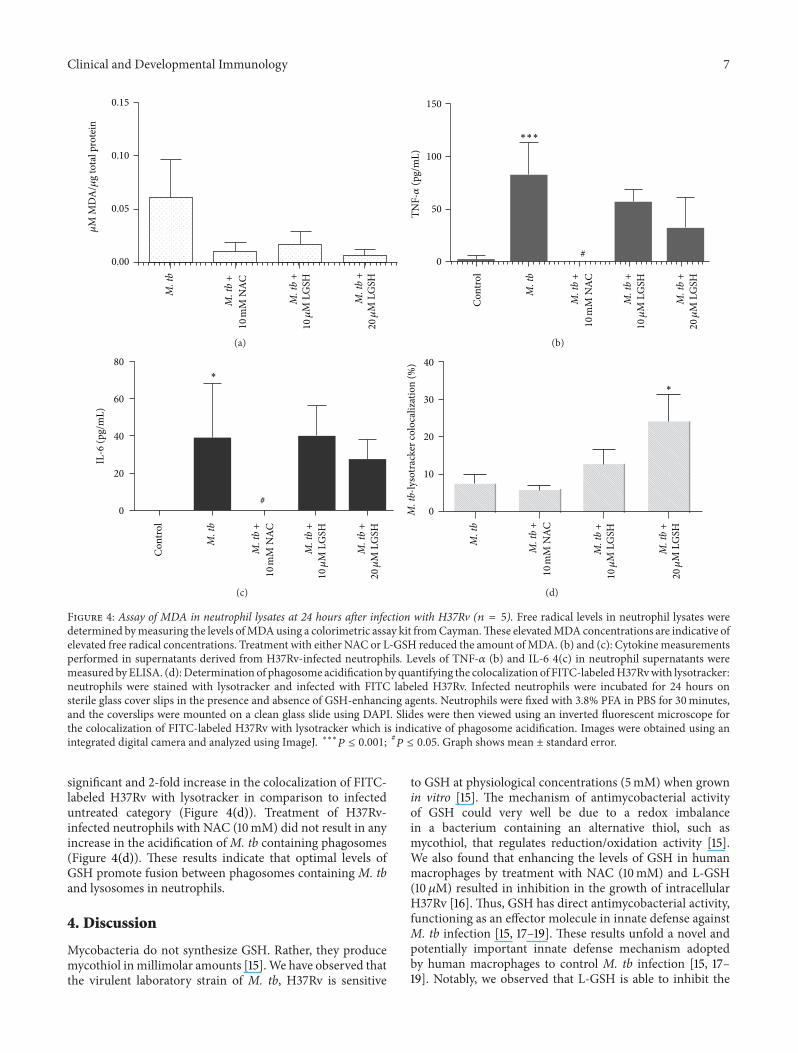
Clinical and Developmental Immunology 7
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
000
005
010
015120583
M M
DA
120583g
tota
l pro
tein
M t
b +
10 m
M N
AC
(a)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
50
100
150
lowastlowastlowast
TNF-120572
(pg
mL)
(b)
Con
trol
M t
b
10120583
M L
GSH
20120583
M L
GSH
M t
b +
M t
b +
M t
b +
10 m
M N
AC
0
20
60
40
80lowast
IL-6
(pg
mL)
(c)
0
10
30
20
40
M t
b-ly
sotr
acke
r col
ocal
izat
ion
()
lowast
M t
b
M t
b +
10120583
M L
GSH
M t
b +
20120583
M L
GSH
M t
b +
10 m
M N
AC
(d)
Figure 4 Assay of MDA in neutrophil lysates at 24 hours after infection with H37Rv (119899 = 5) Free radical levels in neutrophil lysates weredetermined bymeasuring the levels ofMDAusing a colorimetric assay kit fromCaymanThese elevatedMDAconcentrations are indicative ofelevated free radical concentrations Treatment with either NAC or L-GSH reduced the amount ofMDA (b) and (c) Cytokinemeasurementsperformed in supernatants derived from H37Rv-infected neutrophils Levels of TNF-120572 (b) and IL-6 4(c) in neutrophil supernatants weremeasured byELISA (d) Determination of phagosome acidification by quantifying the colocalization of FITC-labeledH37Rvwith lysotrackerneutrophils were stained with lysotracker and infected with FITC labeled H37Rv Infected neutrophils were incubated for 24 hours onsterile glass cover slips in the presence and absence of GSH-enhancing agents Neutrophils were fixed with 38 PFA in PBS for 30minutesand the coverslips were mounted on a clean glass slide using DAPI Slides were then viewed using an inverted fluorescent microscope forthe colocalization of FITC-labeled H37Rv with lysotracker which is indicative of phagosome acidification Images were obtained using anintegrated digital camera and analyzed using ImageJ lowastlowastlowast119875 le 0001
119875 le 005 Graph shows mean plusmn standard error
significant and 2-fold increase in the colocalization of FITC-labeled H37Rv with lysotracker in comparison to infecteduntreated category (Figure 4(d)) Treatment of H37Rv-infected neutrophils with NAC (10mM) did not result in anyincrease in the acidification ofM tb containing phagosomes(Figure 4(d)) These results indicate that optimal levels ofGSH promote fusion between phagosomes containing M tband lysosomes in neutrophils
4 Discussion
Mycobacteria do not synthesize GSH Rather they producemycothiol inmillimolar amounts [15]We have observed thatthe virulent laboratory strain of M tb H37Rv is sensitive
to GSH at physiological concentrations (5mM) when grownin vitro [15] The mechanism of antimycobacterial activityof GSH could very well be due to a redox imbalancein a bacterium containing an alternative thiol such asmycothiol that regulates reductionoxidation activity [15]We also found that enhancing the levels of GSH in humanmacrophages by treatment with NAC (10mM) and L-GSH(10 120583M) resulted in inhibition in the growth of intracellularH37Rv [16] Thus GSH has direct antimycobacterial activityfunctioning as an effector molecule in innate defense againstM tb infection [15 17ndash19] These results unfold a novel andpotentially important innate defense mechanism adoptedby human macrophages to control M tb infection [15 17ndash19] Notably we observed that L-GSH is able to inhibit the
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010
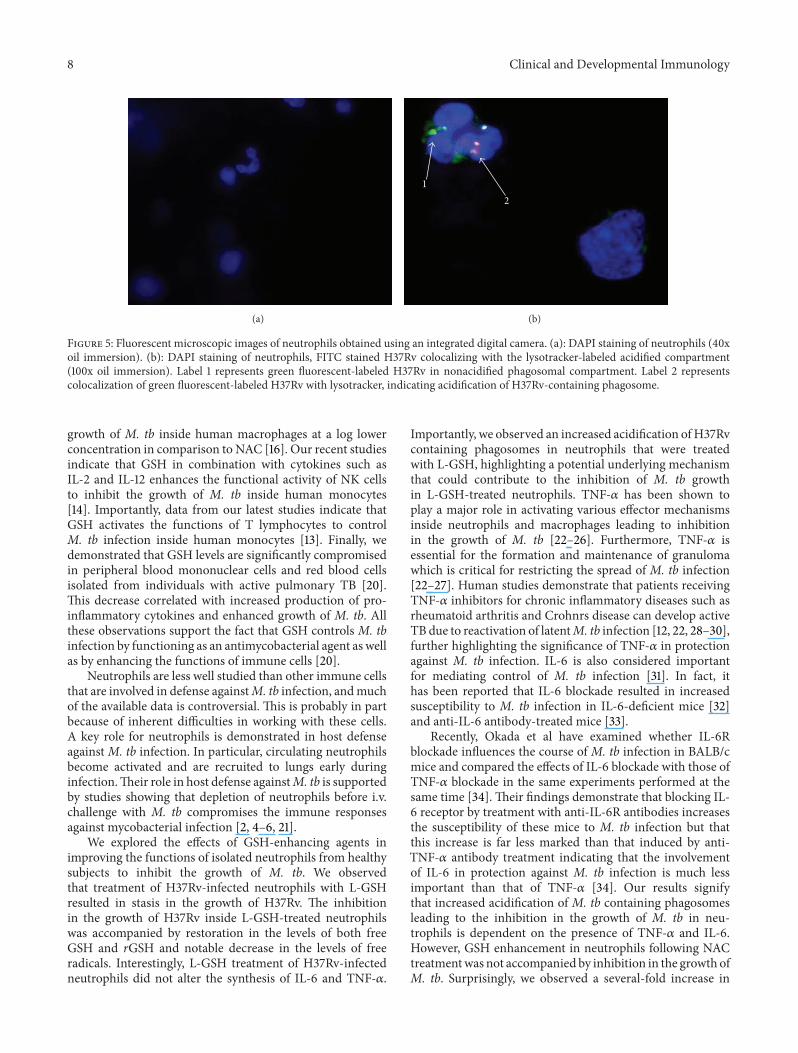
8 Clinical and Developmental Immunology
(a)
12
(b)
Figure 5 Fluorescent microscopic images of neutrophils obtained using an integrated digital camera (a) DAPI staining of neutrophils (40xoil immersion) (b) DAPI staining of neutrophils FITC stained H37Rv colocalizing with the lysotracker-labeled acidified compartment(100x oil immersion) Label 1 represents green fluorescent-labeled H37Rv in nonacidified phagosomal compartment Label 2 representscolocalization of green fluorescent-labeled H37Rv with lysotracker indicating acidification of H37Rv-containing phagosome
growth of M tb inside human macrophages at a log lowerconcentration in comparison to NAC [16] Our recent studiesindicate that GSH in combination with cytokines such asIL-2 and IL-12 enhances the functional activity of NK cellsto inhibit the growth of M tb inside human monocytes[14] Importantly data from our latest studies indicate thatGSH activates the functions of T lymphocytes to controlM tb infection inside human monocytes [13] Finally wedemonstrated that GSH levels are significantly compromisedin peripheral blood mononuclear cells and red blood cellsisolated from individuals with active pulmonary TB [20]This decrease correlated with increased production of pro-inflammatory cytokines and enhanced growth of M tb Allthese observations support the fact that GSH controls M tbinfection by functioning as an antimycobacterial agent as wellas by enhancing the functions of immune cells [20]
Neutrophils are less well studied than other immune cellsthat are involved in defense againstM tb infection andmuchof the available data is controversial This is probably in partbecause of inherent difficulties in working with these cellsA key role for neutrophils is demonstrated in host defenseagainst M tb infection In particular circulating neutrophilsbecome activated and are recruited to lungs early duringinfectionTheir role in host defense againstM tb is supportedby studies showing that depletion of neutrophils before ivchallenge with M tb compromises the immune responsesagainst mycobacterial infection [2 4ndash6 21]
We explored the effects of GSH-enhancing agents inimproving the functions of isolated neutrophils from healthysubjects to inhibit the growth of M tb We observedthat treatment of H37Rv-infected neutrophils with L-GSHresulted in stasis in the growth of H37Rv The inhibitionin the growth of H37Rv inside L-GSH-treated neutrophilswas accompanied by restoration in the levels of both freeGSH and rGSH and notable decrease in the levels of freeradicals Interestingly L-GSH treatment of H37Rv-infectedneutrophils did not alter the synthesis of IL-6 and TNF-120572
Importantly we observed an increased acidification ofH37Rvcontaining phagosomes in neutrophils that were treatedwith L-GSH highlighting a potential underlying mechanismthat could contribute to the inhibition of M tb growthin L-GSH-treated neutrophils TNF-120572 has been shown toplay a major role in activating various effector mechanismsinside neutrophils and macrophages leading to inhibitionin the growth of M tb [22ndash26] Furthermore TNF-120572 isessential for the formation and maintenance of granulomawhich is critical for restricting the spread of M tb infection[22ndash27] Human studies demonstrate that patients receivingTNF-120572 inhibitors for chronic inflammatory diseases such asrheumatoid arthritis and Crohnrs disease can develop activeTB due to reactivation of latentM tb infection [12 22 28ndash30]further highlighting the significance of TNF-120572 in protectionagainst M tb infection IL-6 is also considered importantfor mediating control of M tb infection [31] In fact ithas been reported that IL-6 blockade resulted in increasedsusceptibility to M tb infection in IL-6-deficient mice [32]and anti-IL-6 antibody-treated mice [33]
Recently Okada et al have examined whether IL-6Rblockade influences the course of M tb infection in BALBcmice and compared the effects of IL-6 blockade with those ofTNF-120572 blockade in the same experiments performed at thesame time [34] Their findings demonstrate that blocking IL-6 receptor by treatment with anti-IL-6R antibodies increasesthe susceptibility of these mice to M tb infection but thatthis increase is far less marked than that induced by anti-TNF-120572 antibody treatment indicating that the involvementof IL-6 in protection against M tb infection is much lessimportant than that of TNF-120572 [34] Our results signifythat increased acidification of M tb containing phagosomesleading to the inhibition in the growth of M tb in neu-trophils is dependent on the presence of TNF-120572 and IL-6However GSH enhancement in neutrophils following NACtreatmentwas not accompanied by inhibition in the growth ofM tb Surprisingly we observed a several-fold increase in
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010
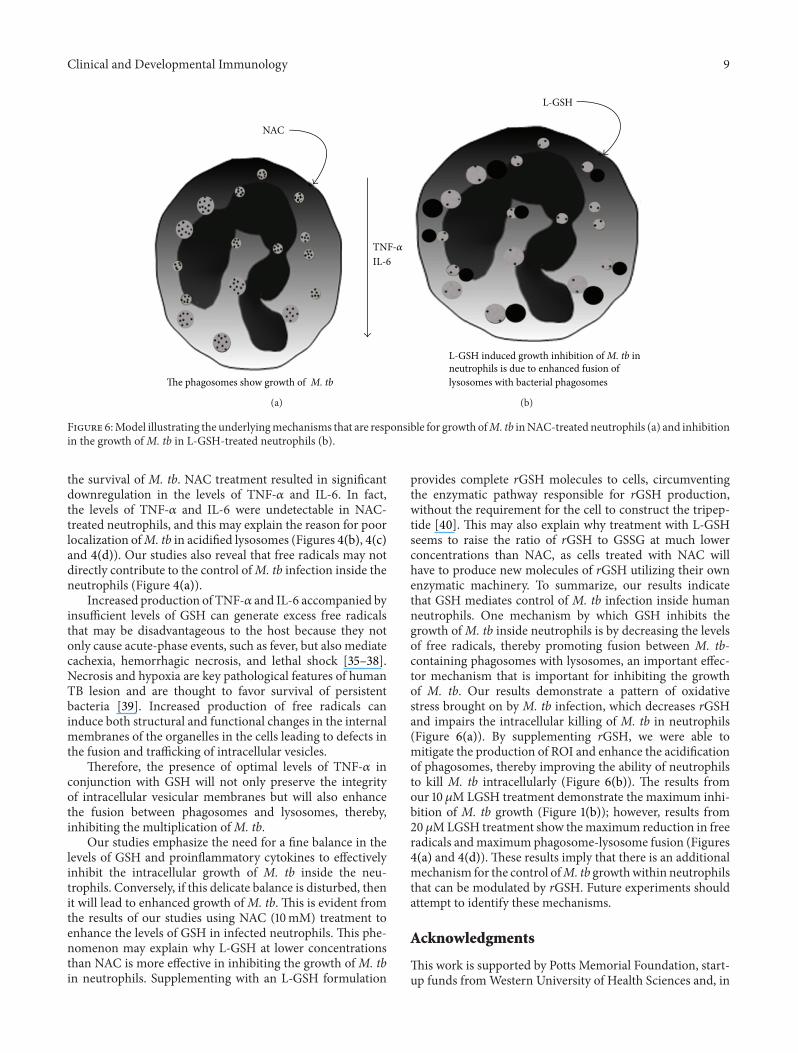
Clinical and Developmental Immunology 9
NAC
The phagosomes show growth of M tb
TNF-120572IL-6
(a)
L-GSH
L-GSH induced growth inhibition of M tb inneutrophils is due to enhanced fusion oflysosomes with bacterial phagosomes
(b)
Figure 6Model illustrating the underlyingmechanisms that are responsible for growth ofM tb inNAC-treated neutrophils (a) and inhibitionin the growth ofM tb in L-GSH-treated neutrophils (b)
the survival of M tb NAC treatment resulted in significantdownregulation in the levels of TNF-120572 and IL-6 In factthe levels of TNF-120572 and IL-6 were undetectable in NAC-treated neutrophils and this may explain the reason for poorlocalization ofM tb in acidified lysosomes (Figures 4(b) 4(c)and 4(d)) Our studies also reveal that free radicals may notdirectly contribute to the control ofM tb infection inside theneutrophils (Figure 4(a))
Increased production of TNF-120572 and IL-6 accompanied byinsufficient levels of GSH can generate excess free radicalsthat may be disadvantageous to the host because they notonly cause acute-phase events such as fever but also mediatecachexia hemorrhagic necrosis and lethal shock [35ndash38]Necrosis and hypoxia are key pathological features of humanTB lesion and are thought to favor survival of persistentbacteria [39] Increased production of free radicals caninduce both structural and functional changes in the internalmembranes of the organelles in the cells leading to defects inthe fusion and trafficking of intracellular vesicles
Therefore the presence of optimal levels of TNF-120572 inconjunction with GSH will not only preserve the integrityof intracellular vesicular membranes but will also enhancethe fusion between phagosomes and lysosomes therebyinhibiting the multiplication ofM tb
Our studies emphasize the need for a fine balance in thelevels of GSH and proinflammatory cytokines to effectivelyinhibit the intracellular growth of M tb inside the neu-trophils Conversely if this delicate balance is disturbed thenit will lead to enhanced growth ofM tb This is evident fromthe results of our studies using NAC (10mM) treatment toenhance the levels of GSH in infected neutrophils This phe-nomenon may explain why L-GSH at lower concentrationsthan NAC is more effective in inhibiting the growth ofM tbin neutrophils Supplementing with an L-GSH formulation
provides complete rGSH molecules to cells circumventingthe enzymatic pathway responsible for rGSH productionwithout the requirement for the cell to construct the tripep-tide [40] This may also explain why treatment with L-GSHseems to raise the ratio of rGSH to GSSG at much lowerconcentrations than NAC as cells treated with NAC willhave to produce new molecules of rGSH utilizing their ownenzymatic machinery To summarize our results indicatethat GSH mediates control of M tb infection inside humanneutrophils One mechanism by which GSH inhibits thegrowth ofM tb inside neutrophils is by decreasing the levelsof free radicals thereby promoting fusion between M tb-containing phagosomes with lysosomes an important effec-tor mechanism that is important for inhibiting the growthof M tb Our results demonstrate a pattern of oxidativestress brought on by M tb infection which decreases rGSHand impairs the intracellular killing of M tb in neutrophils(Figure 6(a)) By supplementing rGSH we were able tomitigate the production of ROI and enhance the acidificationof phagosomes thereby improving the ability of neutrophilsto kill M tb intracellularly (Figure 6(b)) The results fromour 10 120583M LGSH treatment demonstrate the maximum inhi-bition of M tb growth (Figure 1(b)) however results from20120583MLGSH treatment show themaximum reduction in freeradicals andmaximum phagosome-lysosome fusion (Figures4(a) and 4(d)) These results imply that there is an additionalmechanism for the control ofM tb growthwithin neutrophilsthat can be modulated by rGSH Future experiments shouldattempt to identify these mechanisms
Acknowledgments
This work is supported by Potts Memorial Foundation start-up funds fromWestern University of Health Sciences and in
10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010

10 Clinical and Developmental Immunology
part by Your Energy Systems The authors thank Ms TrinaRich and Dr Caesar Ochoa at the Patient Care Center ofWestern University of Health Sciences for their help withdrawing blood from the volunteers they thank Dr FrederickT Guilford for the useful discussions and the volunteers fortheir participation in this study
References
[1] World Health Organization ldquoTuberculosisrdquo httpwwwwhointtbpublicationsfactsheet globalpd
[2] E W Tiemersma M J van der Werf M W Borgdorff BG Williams and N J D Nagelkerke ldquoNatural history oftuberculosis duration and fatality of untreated pulmonarytuberculosis in HIV negative patients a systematic reviewrdquoPLoS ONE vol 6 no 4 Article ID e17601 2011
[3] M B Goren P DrsquoArcy Hart M R Young and J A Arm-strong ldquoPrevention of phagosome lysosome fusion in culturedmacrophages by sulfatides ofMycobacterium tuberculosisrdquo Pro-ceedings of the National Academy of Sciences of the United Statesof America vol 73 no 7 pp 2510ndash2514 1976
[4] S-Y Eum J-H Kong M-S Hong et al ldquoNeutrophils are thepredominant infected phagocytic cells in the airways of patientswith active pulmonary TBrdquo Chest vol 137 no 1 pp 122ndash1282010
[5] I Sugawara T Udagawa and H Yamada ldquoRat neutrophils pre-vent the development of tuberculosisrdquo Infection and Immunityvol 72 no 3 pp 1804ndash1806 2004
[6] P Seiler P Aichele B Raupach B Odermatt U Steinhoffand S H E Kaufmann ldquoRapid neutrophil response controlsfast-replicating intracellular bacteria but not slow-replicatingMycobacterium tuberculosisrdquo Journal of Infectious Diseases vol181 no 2 pp 671ndash680 2000
[7] O W Griffith ldquoBiologic and pharmacologic regulation ofmammalian glutathione synthesisrdquo Free Radical Biology andMedicine vol 27 no 9-10 pp 922ndash935 1999
[8] R Buhl H A Jaffe K J Holroyd et al ldquoSystemic glutathionedeficiency in symptom-free HIV-seropositive individualsrdquo TheLancet vol 2 no 8675 pp 1294ndash1298 1989
[9] B de Quay R Malinverni and B H Lauterburg ldquoGlutathionedepletion in HIV-infected patients role of cysteine deficiencyand effect of oral N-acetylcysteinerdquo AIDS vol 6 no 8 pp 815ndash819 1992
[10] H-P Eck H Gmunder M Hartmann D Petzoldt V Danieland W Droge ldquoLow concentrations of acid-soluble thiol (cys-teine) in the blood plasma ofHIV-1-infected patientsrdquoBiologicalChemistry Hoppe-Seyler vol 370 no 2 pp 101ndash108 1989
[11] B Helbling J von Overbeck and B H Lauterburg ldquoDecreasedrelease of glutathione into the systemic circulation of patientswith HIV infectionrdquo European Journal of Clinical Investigationvol 26 no 1 pp 38ndash44 1996
[12] M Klein K Jarosova S Forejtova et al ldquoGold and tuberculinskin tests for the detection of latent tuberculosis infection inpatients treated with tumour necrosis factor alpha blockingagentsrdquo Clinical and Experimental Rheumatology vol 31 no 1pp 111ndash117 2013
[13] C Guerra D Morris A Sipin et al ldquoGlutathione and adaptiveimmune responses against Mycobacterium tuberculosis infec-tion in healthy and HIV infected individualsrdquo PLoS ONE vol6 no 12 Article ID e28378 2011
[14] C Guerra K Johal D Morris et al ldquoControl ofMycobacteriumtuberculosis growth by activated natural killer cellsrdquoClinical andExperimental Immunology vol 168 no 1 pp 142ndash152 2012
[15] V Venketaraman Y K Dayaram M T Talaue and N DConnell ldquoGlutathione and nitrosoglutathione in macrophagedefense against Mycobacterium tuberculosisrdquo Infection andImmunity vol 73 no 3 pp 1886ndash1889 2005
[16] D Morris C Guerra M Khurasany et al ldquoGlutathione supple-mentation improves macrophage functions in HIVrdquo Journal ofInterferon amp Cytokine Research vol 33 no 5 pp 270ndash279 2013
[17] V Venketaraman Y K Dayaram A G Amin et al ldquoRole ofglutathione in macrophage control of mycobacteriardquo Infectionand Immunity vol 71 no 4 pp 1864ndash1871 2003
[18] Y K Dayaram M T Talaue N D Connell and V Venketara-man ldquoCharacterization of a glutathione metabolic mutant ofMycobacterium tuberculosis and its resistance to glutathione andnitrosoglutathionerdquo Journal of Bacteriology vol 188 no 4 pp1364ndash1372 2006
[19] D Morris C Guerra C Donohue H Oh M Khurasany andV Venketaraman ldquoUnveiling the mechanisms for decreasedglutathione in individuals with HIV infectionrdquo Clinical andDevelopmental Immunology vol 2012 Article ID 734125 2012
[20] V Venketaraman A Millman M Salman et al ldquoGlutathionelevels and immune responses in tuberculosis patientsrdquo Micro-bial Pathogenesis vol 44 no 3 pp 255ndash261 2008
[21] DG Russell C E Barry III and J L Flynn ldquoTuberculosis whatwe donrsquot know can and does hurt usrdquo Science vol 328 no 5980pp 852ndash856 2010
[22] J Keane S Gershon R P Wise et al ldquoTuberculosis associatedwith infliximab a tumor necrosis factor 120572-neutralizing agentrdquoTheNew England Journal of Medicine vol 345 no 15 pp 1098ndash1104 2001
[23] J J Gomez-Reino L Carmona V Rodrıguez Valverde EM Mola and M D Montero ldquoTreatment of rheumatoidarthritis with tumor necrosis factor inhibitors may predisposeto significant increase in tuberculosis risk a multicenter active-surveillance reportrdquo Arthritis and Rheumatism vol 48 no 8pp 2122ndash2127 2003
[24] T Botha and B Ryffel ldquoReactivation of latent tuberculosisinfection in TNF-deficient micerdquo Journal of Immunology vol171 no 6 pp 3110ndash3118 2003
[25] S Ehlers ldquoWhy does tumor necrosis factor targeted therapyreactivate tuberculosisrdquo Journal of Rheumatology vol 32 no74 pp 35ndash39 2005
[26] J L Flynn M M Goldstein J Chan et al ldquoTumor necrosisfactor-120572 is required in the protective immune response againstMycobacterium tuberculosis inmicerdquo Immunity vol 2 no 6 pp561ndash572 1995
[27] K O Kisich M Higgins G Diamond and L Heifets ldquoTumornecrosis factor alpha stimulates killing ofMycobacterium tuber-culosis by human neutrophilsrdquo Infection and Immunity vol 70no 8 pp 4591ndash4599 2002
[28] J Askling C M Fored L Brandt et al ldquoRisk and case char-acteristics of tuberculosis in rheumatoid arthritis associatedwith tumor necrosis factor antagonists in SwedenrdquoArthritis andRheumatism vol 52 no 7 pp 1986ndash1992 2005
[29] MMWR ldquoTuberculosis associated with blocking agents againsttumor necrosis factor-alpha-California 2002-2003rdquo Morbidityand Mortality Weekly Report vol 53 p 68 2004
[30] R S Wallis M S Broder J Y Wong M E Hanson and DO Beenhouwer ldquoGranulomatous infectious diseases associated
Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010

Clinical and Developmental Immunology 11
with tumor necrosis factor antagonistsrdquo Clinical InfectiousDiseases vol 38 no 9 pp 1261ndash1265 2004
[31] I S Leal M Florido P Andersen and R AppelbergldquoInterleukin-6 regulates the phenotype of the immune responseto a tuberculosis subunit vaccinerdquo Immunology vol 103 no 3pp 375ndash381 2001
[32] C H Ladel C Blum A Dreher K Reifenberg M Kopf and SH E Kaufmann ldquoLethal tuberculosis in interleukin-6-deficientmutant micerdquo Infection and Immunity vol 65 no 11 pp 4843ndash4849 1997
[33] R Appelberg A G Castro J Pedrosa and P Minoprio ldquoRoleof interleukin-6 in the induction of protective T cells duringmycobacterial infections in micerdquo Immunology vol 82 no 3pp 361ndash364 1994
[34] M Okada Y Kita N Kanamaru et al ldquoAnti-IL-6 receptorantibody causes less promotion of tuberculosis infection thanAnti-TNF-120572 antibody in micerdquo Clinical and DevelopmentalImmunology vol 2011 Article ID 404929 2011
[35] J L Rothstein T F Lint and H Schreiber ldquoTumor necrosisfactorcachectin Induction of hemorrhagic necrosis in normaltissue requires the fifth component of complement (C5)rdquoJournal of Experimental Medicine vol 168 no 6 pp 2007ndash20211988
[36] R M Strieter S L Kunkel and R C Bone ldquoRole of tumornecrosis factor-120572 in disease states and inflammationrdquo CriticalCare Medicine vol 21 no 10 pp S447ndashS463 1993
[37] R Buhl C Vogelmeier M Critenden et al ldquoAugmentationof glutathione in the fluid lining the epithelium of the lowerrespiratory tract by directly administering glutathione aerosolrdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 87 no 11 pp 4063ndash4067 1990
[38] J H Roum Z Borok N G McElvaney et al ldquoGlutathioneaerosol suppresses lung epithelial surface inflammatory cell-derived oxidants in cystic fibrosisrdquo Journal of Applied Physiol-ogy vol 87 no 1 pp 438ndash443 1999
[39] C Skerry J Harper M Klunk W R Bishai and S K JainldquoAdjunctive TNF inhibition with standard treatment enhancesbacterial clearance in a murine model of necrotic TB granulo-masrdquo PLoS ONE vol 7 no 6 Article ID e39680 2012
[40] G D Zeevalk L P Bernard and F T Guilford ldquoLiposomal-glutathione provides maintenance of intracellular glutathioneand neuroprotection in mesencephalic neuronal cellsrdquo Neuro-chemical Research vol 35 no 10 pp 1575ndash1587 2010





























