Embed Size (px)
Citation preview

Age-associated tyrosine nitration of rat skeletal muscle glycogen
phosphorylase b: characterization by HPLC–nanoelectrospray–Tandem
mass spectrometry
Victor S. Sharov a, Nadezhda A. Galeva b, Jaroslaw Kanski a,
Todd D. Williams b, Christian Schoneich a,*
a Department of Pharmaceutical Chemistry, University of Kansas, 2095 Constant Avenue, Lawrence, KS 66047, USAb Mass Spectrometry Laboratory, University of Kansas, Lawrence, KS 66047, USA
Received 9 December 2005; received in revised form 23 February 2006; accepted 28 February 2006
Abstract
We identified age-dependent post-translational modifications of skeletal muscle glycogen phosphorylase b (Ph-b), isolated from F1 hybrids of
Fisher 344!Brown Norway rats. Ph-b isolated from 34 months old rats showed a statistically significant decrease in specific activity compared to
6 months old animals: 13.8G0.7 vs. 20.6G0.8 UmgK1 protein, respectively. Western blot analysis of the purified Ph-b with anti-3-NT antibodies
revealed an age-dependent accumulation of 3-nitrotyrosine (3-NT), quantified by reverse-phase HPLC–UV analysis to increase from 0.05G0.03
to 0.34G0.11 (mol 3-NT/mol Ph-b) for 6 vs. 34 months old rats, respectively. HPLC–nanoelectrospray ionization—tandem mass spectrometry
revealed the accumulation of 3-NT on Tyr113, Tyr161 and Tyr573. While nitration of Tyr113 was detected for both young and old rats, 3-NT at
positions 161 and 573 was identified only for Ph-b isolated from 34 months old rats. The sequence of the rat muscle Ph-b was corrected based on
our protein sequence mapping and a custom rat PHS2 sequence containing 17 differently located amino acid residues was used instead of the
database sequence. The in vitro reaction of peroxynitrite with Ph-b resulted in the nitration of multiple Tyr residues at positions 51, 52, 113, 155,
185, 203, 262, 280, 404, 473, 731, and 732. Thus, the in vitro nitration conditions only mimic the nitration of a single Tyr residue observed in vivo
suggesting alternative pathways controlling the accumulation of 3-NT in vivo. Our data show a correlation of age-dependent 3-NT accumulation
with Ph-b inactivation.
q 2006 Elsevier Inc. All rights reserved.
Keywords: Aging; Rat skeletal muscle; Glycogen phosphorylase b; Oxidative posttranslational modifications; 3-Nitrotyrosine; Mass spectrometry
1. Introduction
The post-translational oxidative modification of proteins has
been implicated in the age-dependent loss of physiological
function (Starke-Reed and Oliver, 1989; Stadtman, 1992;
Sohal et al., 1993). The accumulation of modified proteins is
species-and tissue-specific, controlled by a variety of factors
such as nature of the oxidant, antioxidant protection, protein
repair and protein turnover (Ji et al., 1990; Lawler et al., 1993;
Larsen, 1993; Reiter, 1995; Parkes et al., 1998; Sun and Tower,
1999; Arking et al., 2000; Leeuwenburgh et al., 1994;
Vigouroux et al., 2004; Mary et al., 2004; Grune et al.,
2004). Although, elevated tissue levels of protein carbonyls
0531-5565/$ - see front matter q 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.exger.2006.02.012
* Corresponding author. Tel.: C1 785 864 4880; fax: C1 785 864 5736.
E-mail address: [email protected] (C. Schoneich).
(Yan et al., 1997, Yan and Sohal 1998) and 3-nitrotyrosine (3-
NT) (van der Loo et al., 2000; Paik et al., 2001; Drew and
Leeuwenburgh, 2002) as a result of aging and age-associated
pathology have been reported, the physiological significance of
these modifications must be addressed via the detailed
characterization of individual modified proteins. In fact, only
a limited number of papers specifically analyzed the age-
dependent posttranslational oxidative modifications of purified
proteins and their functional significance. In a few studies, Cys
oxidation of phosphoglycerate kynase (Gafni, 1981) or
sarcoplasmic reticulum Ca-ATPase (SERCA) (Viner et al.
1997, 1999), Met sulfoxidation of calmodulin (Gao et al.,
1998), Tyr nitration of SERCA2a (Viner et al., 1996, 1999;
Knyushko et al., 2005), flotillin-1 and a-tubulin (Dremina
et al., 2005) and actin (Aslan et al., 2003), or carbonylation of
mitochondrial aconitase (Das et al., 2001), adenine nucleotide
translocase (Yan and Sohal, 1998) and carbonic anhydrase III
(Cabiscol and Levine, 1995) have been characterized. An
Experimental Gerontology 41 (2006) 407–416
www.elsevier.com/locate/expgero

V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416408
important message from these studies is a remarkable
selectivity of the age-dependent oxidative protein modifi-
cation. More recently, larger scale proteomic analyses
identified a number of proteins sensitive to oxidative/nitrative
modifications in vivo as a result of biological aging
(Schoneich, 2003; Kanski et al., 2003, 2005a,b; Castegna
et al., 2002; Poggioli et al., 2004; Levine, 2002). In our
laboratory, we characterized the sequence-specific age-depen-
dent tyrosine nitration of several rat skeletal muscle proteins,
such as b-enolase, a-fructose aldolase, creatine kinase and
glycogen phosphorylase b (Ph-b) (Kanski et al., 2005b), which
perform important functions in energy metabolism, suggesting
that the nitration of such key proteins can be, in part,
responsible for the well-described age-associated decline of
muscle motor function (Larsson and Ansved, 1995). The
detection of such age-associated site-specific protein tyrosine
nitration on a proteomic scale serves as a first step before a
more complete characterization of these protein modifications
and their physiological consequences on individual purified
proteins. Therefore, in the current paper, we analyzed the age-
dependent accumulation of 3-NT, oxidative modification of
Cys residues, and the enzymatic activity of Ph-b isolated from
the skeletal muscle of 6 and 34 months old F1 hybrids of Fisher
344 and Brown Norway rats (F344!BN F1), an established
animal model of longevity (Fernandes et al., 1997). This
protein plays a key role in myophosporylase deficiency
(McArdle’s disease), a common glycogenosis affecting skeletal
muscle function. The latter is caused by a number of mutations
or splicing of the muscle glycogen phosphorylase gene
(PYGM), which affect protein activity (Vorgerd et al., 1998;
Di Mauro et al., 2002). Given that aging is generally
characterized by a progressive loss of skeletal muscle
performance, the accumulation of oxidative and/or nitrative
modifications on muscle Ph-b may represent a potential
molecular basis for this phenomenon.
2. Materials and methods
2.1. Chemicals
TPCK-treated sequence grade trypsin was from Promega
(Madison, WI). Rabbit muscle Ph-b, bovine serum albumin
(BSA), dithiothreitol (DTT), SDS, 5,5 0-dithio-bis-(2-nitroben-
zoic acid) (DTNB), urea, NADP, rabbit liver glycogen,
adenosine-5 0-monophosphate, glucose-6-phosphate dehydro-
genase and sodium iodoacetate were purchased from Sigma
(St. Louis, MO). Phosphoglucomutase was obtained from
Boeringer Mannheim (Indianapolis, IN). Pre-cast Novexw tris–
glycine-SDS gels, molecular weight standard Mark12, running
and sample buffers were from Invitrogen (Carlsbad, CA).
Monoclonal antibodies against protein-bound 3-NT (clone
1A6) were from Upstate Biotechnology (Lake Placid, NY). A
maleimide-based fluorescence dye, ThioGlo1, was obtained
from Covalent Associates (Woburn, MA). All other chemicals
of highest commercially available grade were obtained from
Fisher (Pittsburgh, PA).
2.2. Animals
The research protocol outlined in this manuscript was
approved by the University of Kansas Animal Care Facility.
Young (6 months) and old (34 months) Fisher 344!Brown
Norway F1 hybrid rats were purchased from the National
Institute of Aging colonies maintained at Harlan Sprague-
Dawley, Inc. (Indianapolis, IN, USA). The rats were allowed to
adapt for 2 weeks after arrival in a 12 h light/dark cycle and
were provided with water and food ad libitum. The animals
were sacrificed by decapitation, and the muscle tissue rapidly
removed, immediately frozen in liquid nitrogen, and stored at
K80 8C.
2.3. Purification of rat skeletal muscle phosphorylase b
Isolation of Ph-b was performed essentially as described
earlier (Fisher and Krebs, 1962) with minor modifications as
outlined below. Briefly, frozen muscle tissue (5 g) was
thawed in 2.5 volumes of ice-cold isolation buffer (IB)
consisting of 20 mM Tris–HCl (pH 7.4), 1 mM EDTA,
0.5 mM DTT and 0.1 mM freshly added PMSF, homogen-
ized for 2 min at maximal speed in an ice-cooled Waring
blender LB10 (Waring, Torrington, CT) followed by 3!15 s sonication with a probe sonicator (Sonic Dismembrator
500, Fisher Scientific, Pittsburg, PA) at 10% power. The
homogenate was centrifuged at 8000g for 10 min; the
supernatant was saved and pooled with the supernatant of a
second extraction from the pellet with the same volume of
IB. Protein extracts were filtered through four layers of
cheesecloth and kept for 30 min at 37 8C for converting
phosphorylase a (Ph-a) to Ph-b and denaturing some
thermolabile proteins. Solid (NH4)2SO4 was slowly added
to precipitate proteins at 4 8C under stirring; the fractions
between 25 and 50% saturation after 6 h incubation were
collected by centrifugation at 20,000g for 20 min. Pellets
were dissolved in a small volume of IB and exhaustively
dialyzed against IB at 4 8C to remove the excess of
ammonium sulfate. The dialyzate was centrifuged at
20,000!g for 1 h to remove insoluble proteins, and applied
to a self-packed DEAE-Sephacel column (4.6!100 mm),
equilibrated with IB (in the cold cabinet). The column was
washed with this buffer until complete removal of unbound
proteins, and eluted by a linear gradient of NaCl from 0 to
0.3 M in ca. 10 bed volumes of IB at a flow rate of 1 ml/
min using an Acta Prime FPLC system (Amersham
Biosciences, Piscataway, NJ), equipped with absorbance
(monitoring at 280 nm) and conductivity detectors. Fractions
containing protein and Ph-b activity were pooled and further
separated/analyzed by SDS-PAGE and reverse-phase HPLC.
Protein concentration was determined by the bicinchoninic
acid (BCA) assay using BSA as a standard according to the
manufacturer’s instructions (Pierce, Rockford, IL). In some
experiments, where indicated, Ph-b isolation media did not
contain any reducing agents in order to maintain the
oxidation state of the protein isolated from muscle tissue.

V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416 409
2.4. Gel electrophoresis
Samples of Ph-b containing 10–100 mg protein were mixed
with an equal volume of tris–glycine-SDS sample buffer
(containing 5% b-mercaptoethanol for reducing conditions),
heated for 2 min at 95 8C, and loaded into 1.5 mm thick 10-
well Novex 4–12% Tris–glycine gradient gels. After running
the SDS-polyacrylamide gel electrophoresis (SDS-PAGE) at
200 V for 90 min, the gels were stained with 0.2% Coomassie
R250 in 10% acetic acid/30% methanol/60% H2O (v/v/v) for
2 h followed by destaining in 7.5% acetic acid/40%
methanol/52.5% H2O (v/v/v) until the bands were visible
and the background was clear.
2.5. Reverse-phase HPLC of proteins
For reverse-phase HPLC separation of Ph-b, we used a
procedure described by us earlier (Sharov et al., 2002). Protein
samples were dissolved in neat TFA and injected onto Vydac
C4 column (250!4.6 mm id.), protected by a guard column
(10!4.6 mm id.), equilibrated with 40% (v/v) ethanol in 0.1%
aqueous TFA, and eluted at a flow rate of 0.5 ml/min by a
linear gradient increasing the ethanol content by 1.3%/min.
Chromatograms were monitored by absorbance in the spectral
range of 200–600 nm using a photodiode array (PDA) detector
(SPD-M10A) or by fluorescence using a fluorescence detector,
RF-10A (both from Shimadzu Corp., Japan).
The 3-NT content in Ph-b was estimated using the following
equation (Sharov et al., 2002)
3 � NT=Ph � bðmol=molÞZA360=A280!3280=3360;
where A280 and A360 are the Ph-b chromatographic peak areas
quantified at 280 (for protein) and 360 nm (for 3-NT), 3280Z116,720 MK1 cmK1 and 3360Z2100 MK1 cmK1 are the
respective molar extinction coefficients for Ph-b (calculated
from the protein sequence using the GPMAW 3.0 software
from ChemSW Inc., Fairfield, CA), and 3-NT at acidic pH
(Sharov et al., 2002). Similarly, the amount of ThioGlo1 (TG1)
attached to Ph-b Cys residues was quantified using the Ph-b
chromatographic peak areas detected at 379 and 280 nm
according to the following equation
TG1 � Cys=Ph � bðmol=molÞZA379=A280!3280=3379;
where A379 and 3379 are the Ph-b peak area and the extinction
coefficient (3379Z14,454 MK1 cmK1 according to the sup-
plier) for TG1-Cys adducts, respectively; A280 is the Ph-b
chromatographic peak area detected at 280 nm and 3280Z116,720 MK1 cmK1 (see above). The absolute amount of rat
skeletal muscle Ph-b was determined by both gel band
densitometry and RP-HPLC peak area using commercial
rabbit skeletal muscle Ph-b (Sigma, St Louis, MO) as an
authentic standard.
2.6. Assay of phosphorylase b activity
Ph-b activity was assayed at 21 8C in the direction
of glycogen degradation by coupling the production of
glucose-1-phosphate to NADP reduction by phosphoglucomu-
tase and glucose-6-phosphate dehydrogenase, essentially as
described earlier (Bergmeyer and Gawehn, 1974). The assay
medium contained 0.1 mM EDTA, 3 mM MgCl2, 0.3 mg mlK1
NADP, 0.2 mg mlK1 glycogen, 30 mM adenosine-5 0-mono-
phosphate, 0.7 U mlK1 phosphoglucomutase, and 3 U mlK1
glucose-6-phosphate dehydrogenase in 0.05 M potassium
phosphate buffer (pH 6.8). After recording a blank rate for
2 min, the reaction was initiated by the addition of Ph-b
samples, and the linear increase in absorbance at 340 nm was
monitored for 8 min. The activity was determined as DA340/
min and calibrated using the commercially rabbit skeletal
muscle Ph-b of known activity. To calculate the specific
activity of Ph-b in samples from young and old rats, data were
normalized for the protein quantity estimated by RP-HPLC of
respective samples, as described above.
2.7. Western Blot (WB) analysis
Proteins were transferred from gels to PVDF membranes by
electroblotting (at 100 V for 2 h). Primary antibodies
(mouse anti-3NT clone 1A6) were obtained from Upstate
Biotechnology (Lake Placid, NY). Peroxidase conjugated anti-
mouse IgG was purchased from Pierce (Rockford, IL), and the
blots were visualized with the ECL-plus WB detection kit from
Amersham Pharmacia Biotech (Piskataway, NY) as described
by the manufacturer. To assess specificity of protein 3-NT
detection with antibodies, two important controls were
employed, treatment with dithionite to reduce 3-NT on the
membrane prior to WB analysis, and blocking of the antibodies
with free 3-NT, as described elsewhere (Ye et al., 1996). Both
controls showed a negligible non-specific affinity of the 3-NT
antibodies to Ph-b bands.
2.8. In-gel digestion of Ph-b
Protein bands with apparent mass of 96 kDa were excised
from the gel, cut into pieces of about 1 mm3 size, and processed
as described elsewhere (Sharov et al., 2002). In brief, gel slices
were washed twice for 45 min at 37 8C in 0.5 ml of 200 mM
NH4HCO3/50% (v/v) acetonitrile with agitation. For protein
alkylation, the gel slices were incubated with 2 mM DTT in
200 mM NH4HCO3 for 30 min at 50 8C, followed by the
reaction with 5 mM iodoacetic acid for 30 min at room
temperature. After removal of the solvent the gel slices were
additionally washed for 1 h in 0.5 ml of 200 mM NH4HCO3/
50% acetonitrile (v/v) with agitation, and shrunk in pure
acetonitrile for 15 min. After removal of acetonitrile and
drying under vacuum, the samples were re-swollen with a
buffer containing 40 mM NH4HCO3, 1 mM CaCl2, 10% (v/v)
acetonitrile, and trypsin at a ca. 10:1 molar ratio of the protein
to trypsin (usually, 2 mg trypsin per gel band containing 20 mg
protein). The volume of buffer was ca. 1.5-times that of the
excised gel band. After the adsorption of trypsin, additional
buffer (30–50 mL) was used to cover the gel pieces during
overnight digestion (16–18 h) at 37 8C. For the extraction of
peptides from a gel, the overlay of the in-gel digest was

V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416410
obtained after sonication for 30 min in a water bath of an
Ultrasonic Cleaner ME 4.6 (Mettler Electronics Corp.,
Anaheim, CA, USA) and short centrifugation, and used for
MS analysis.
2.9. Mass spectrometry
In-gel digests of Ph-b were analyzed by nano-HPLC with
electrospray ionization-tandem mass spectrometry (NESI-
MS/MS) using either a ThermoElectron LCQ Duo or a
ThermoElectron Classic (San Jose, CA) mass spectrometer
equipped with a nanoelectrospray source (Thermo Electron).
Separation of tryptic peptides was achieved on-line prior to
MS/MS analysis on in-house packed BioBasic C18 stationary
phase (Thermo Electron) nanoflow columns (300 A, 10 cm!75 mm, 15 mm tip size) (New Objective, Woburn, MA) with the
following chromatographic conditions: mobile phase A: 0.1%
formic acid in water, mobile phase B: 0.1% formic acid in
MeCN. The flow rate was 0.5 mL minK1, delivered by a
MicroTech Scientific Ultra Plus II pump (after 1:20 split), or by
a MicroTech Xtreme Simple nano-flow pump (direct flow).
The following gradient profile was used to increase mobile
phase B linearly to the following fractions: from 0 to 5 min
gradient held at 10% B, then increased to 60% B within
105 min, and continued at 60% B for additional 5 min. After
each run, the column was washed by a short gradient (0–60% B
for 20 min) and allowed to re-equilibrate to the initial
conditions for 15 min. The following instrumental conditions
were used for mass spectrometric analysis: number of
microscansZ3, length of microscansZ200 ms, capillary
temperatureZ160 8C, spray voltageZ1.9 kV, capillary volta-
geZ35 V, tube lens offsetZK14 V. The mass spectrometer
was tuned using the static nanospray setup with a 5 mM
solution of angiotensin I (MW 1296.5) infused by a pico-tip
emitter (New Objective). Data acquisition was performed in
the data-dependent fashion, i.e. an MS scan followed by 3 or 4
MS/MS scans of the 3 or 4 most intense peaks with the
normalized collision energy for MS/MS set at 35% and the
isolation width of 2.0 mzK1. A minimal signal for MS/MS
acquisition was set to 2!106. Additionally, the dynamic
exclusion option was enabled and set with the following
parameters: repeat countZ3, repeat duration 5 min, exclusion
list sizeZ25, exclusion durationZ5, and exclusion mass
widthZ3.
MS/MS data were analyzed using the Sequest algorithm
(Ducret et al., 1998) searching the most current non-redundant
NCBI protein database downloaded from the ftp.ncbi.nlm.nift-
gov/blast/db. The filter used for peptide identification included
correlation factors (XCorr) being greater than 1.5, 2.0, and 2.5
for the charge states C1,C2 and C3, respectively. In order to
identify post-translational modifications of rat Ph-b, database
search was performed against a custom database created of
only rat (or later mouse) Ph-b sequences. The following
modifications were accounted for during the search: oxidation
of Met (C16 amu; amuZatomic mass units), carboxymethyla-
tion (C58 amu) or oxidation of Cys (C32 and C48 amu), and
nitration of Tyr (C45 amu). Additionally, MS/MS spectra of
interest were examined manually for the presence 3-NT-
containing peptides (Kanski et al., 2005b). Analysis of the MS/
MS spectra was based on a search for the major sequence-
indicating ions resulting from the cleavage of the parent ion at
specific locations relative to the peptide bond. The Roepstorf-
Fohlman nomenclature (Roepstoff and Fohlman, 1984) was
used for the annotations of N-terminal (b) and C-terminal (y 00)
fragments.
2.10. Mapping rat Ph-b sequence using Q-TOF-MS/MS data
Rat Ph-b samples were subjected to in detail sequence
mapping using Waters CapLC XE system, Q-TOF mass
spectrometer 2 (Micromass, UK) and Protein Lynx Global
Server (PLGS 2.0.5) (Waters, Milford, MA). HPLC separation
of peptides derived from digestion of rat Ph-b with trypsin was
performed on RP-HPLC column (0.32!150 mm Symmetry
C18) at a flow rate of 8 ml minK1 with a linear gradient raising
from 25 to 75% (v/v) methanol in 0.08% (v/v) aqueous formic
acid over a period of 50 min. Ions of intensity greater than 4
counts in a survey scan were selected as precursors for MS/MS.
The obtained data were challenged against SWISS-PROT
databank using PLGS 2.0.5. The MS/MS data files were
processed using slow deisotopic deconvoluting parameters.
The initial PLGS workflow was amended by adding AutoMod
which allows non-specific cleavages and amino acid
substitutions.
MS/MS data obtained on both Q-TOF and LCQ instruments
were searched against a protein database using PLGS and
Sequest, respectively. Mouse skeletal muscle Ph-b
(PHS2_MOUSE, accession code Q9WUB3) was shown to be
the top hit returned by a simple databank search with a
coverage of up to 68%, while for rat skeletal muscle Ph-b
(PHS2_RAT, Swiss-Prot protein database accession code
P09812) only a 58% sequence coverage was obtained. To
resolve this discrepancy and improve mapping coverage,
multiple Ph-b in-gel digests were analyzed using CapLC and
Q-TOF instrumentation.
The databank search by PLGS with the enabled AutoMod
feature allowed us searching without enzymatic specificity and
to consider amino acid substitutions. The results demonstrated
that the experimentally obtained sequence of purified rat
skeletal muscle Ph-b differs from that published in the protein
database: we found 17 amino acid residue deviations at
sequence coverage of 67.8% (Fig. 1). These 17 experimentally
determined amino acid deviations from the published rat
sequence actually match the amino acids in the published
mouse Ph-b sequence. CLUSTAL W (1.82) multiple sequence
alignment (website http://www.ch.embnet.org/cgi-bin/clus-
talw_parser) of our experimentally obtained partial sequence
(496 out of 841 amino acids) with other database PHS2
sequences revealed that there were 492 identities for mouse,
485 identities for human and rabbit, 475 for bovine and sheep
sequences comparative to 479 identities for the rat sequence.
Because the experiment for rat Ph-b demonstrated a better
overlap with the published mouse than the rat sequence, the
mouse sequence was used in subsequent studies.

Fig. 1. Alignment of rat muscle Ph-b (PHS2_RAT) sequence from the Swiss-Prot database and sequence fragments obtained in our experiments. Highlighted italic
letters show differences in the rat Ph-b sequence relative to experimental MS/MS data. The dotted lines indicate peptides, which were not detected by MS/MS.
V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416 411
2.11. Determination of DTNB-reactive Cys residues in Ph-b
For the quantitative analysis of reduced Cys residues in
Ph-b, the reaction with dithio-bis-dinitrobenzoic acid (DTNB)
was employed according to a well-established method
(Ellman, 1959) with minor modifications. Briefly, 100 mg of
dialyzed protein in 20 mM tris–HCl (pH 8.5) containing 1%
SDS and 0.2 mM DTNB was incubated for 30 min at 37 8C
and the absorbance at 412 nm was measured. A standard
curve was obtained using known concentrations of gluta-
thione (GSH) under the same conditions.
2.12. In vitro nitration of Ph-b
Purified Ph-b (1 mg mlK1 protein in 30 mM NH4HCO3, pH
7.8) was subjected to peroxynitrite at desired final concen-
tration up to 1 mM, as described earlier (Sharov et al., 2002;
Kanski et al., 2005b). The protein was then immediately
separated by SDS-PAGE and submitted to in-gel trypsin
digestion prior to NESI-MS/MS analysis, as described above.
2.13. Statistical analysis
Quantitative results were obtained from the data of at least
three independent experiments involving material isolated
from individual animals (nZ4 for each age group).
Values are presented as meanGstandard deviation. Signifi-
cance of a difference between two averages was assessed by
probability (P) associated with a Student’s t-test, calculated
using a two-sample unequal variance and two-tailed dis-
tribution with Microsoft Excel XP Pro software.
3. Results
3.1. Biochemical characterization of Ph-b Isolated from young
and old rat skeletal muscle
The purity of isolated Ph-b was assessed by SDS-PAGE
showing that anion-exchange chromatography yields pre-
parations significantly enriched with Ph-b (Fig. 2A). No
significant differences in apparent protein molecular weight
were observed between young (6 months old) and old (34
months old) samples. These preparations were used for the
analysis of enzyme activity. Further fractionation of the
enriched Ph-b samples by RP-HPLC (chromatograms are not
presented) and collection of the fractions eluted at 23–26 min.
yielded even more homogenous Ph-b preparations (Fig. 2B).
For quantitation of both Ph-b and 3-NT/Ph-b molar ratios, we
analyzed the RP-HPLC peak eluting at ca. 24.5 min., which
contains predominantly Ph-b (Fig. 2B). Analysis of 3-NT
molar content was done as described in the section 2.5. Based
on the independent isolation of Ph-b from 5 young and 5 old
animals, this analysis yields a statistically significant (P!0.05)
age-dependent accumulation of 3-NT on rat skeletal muscle
Ph-b. The 3-NT content rises from 0.05G0.03 mol 3-NT/mol
Ph-b in young to 0.34G0.11 mol 3-NT/mol Ph-b in old rats,
i.e. showing a ca. seven-fold increase (Fig. 3A). At the same
time, biological aging resulted in a ca. 33% decrease in the

Fig. 2. Characterization of the purified Ph-b from skeletal muscle of 6 (Y) and
34 (O) months old rats. (A) and (B): SDS-PAGE separation of Ph-b samples
after ion-exchange chromatography and after additional RP-HPLC purification,
respectively. (C) WB analysis of Ph-b immunoreactivity with anti-3-NT
antibodies. (D) Densitometry analysis of the immunoblots from panel C.
Protein load w 20 mg/lane in (A) and (B) or 4 mg/lane in (C). Data are
representative from at least three independent Ph-b purifications involving
muscle tissue from different rats.
V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416412
specific activity of Ph-b (Fig. 3B) isolated from 34 months old
rats (13.8G0.7 U/mg protein) compared to 6 months old rats
(20.6G0.8 U/mg protein).
3.2. WB analysis of 3-NT content of Ph-b
Equal amounts of the purified protein from young and old
animals were resolved by SDS-PAGE. WB analysis demon-
strated a ca. 40% increase in 3-NT immunoreactivity for the
Ph-b bands obtained from 34 vs. 6 months old animals (Fig. 2C
and D). Reduction of the blots with sodium dithionite prior to
WB analysis abolished the 3-NT immunoreactivity (data not
shown), demonstrating the specificity of the anti-3-NT
antibody. The results of 3-NT quantitation on Ph-b by WB
analysis (Fig. 2D) are different from the data obtained by RP-
HPLC (Fig. 3A) largely due to higher background 3-NT
immunoreactivity in samples from young animals.
Fig. 3. Age-associated changes in Ph-b specific activity (A) and 3-NTcontent of
Ph-b assayed by HPLC–UV (B).
This apparent discrepancy may also be attributed to the semi-
quantitative nature of WB analysis; nevertheless, the WB
analysis qualitatively confirms the age-dependent increase in
the accumulation of 3-NT on rat skeletal muscle Ph-b.
3.3. MS analysis of Ph-b in-gel digests: identification of
sequences nitrated in vivo
Unambiguous proof for the 3-NT accumulation on Ph-b was
demonstrated by using NESI-MS/MS analysis. In three
independent experiments involving Ph-b isolated from three
different young and three different old rats, we identified 3
nitropeptides containing 3-NT at positions 113, 161 and 573.
MS/MS spectra of these nitropeptides are shown in Fig. 4A–C.
Peptides showing nitration of Tyr161, IHEY(NO2)KRQ
LLNC(O3)LHIITLYNR and Tyr573, Y(NO2)EFGIFNQ
KIC(O3)GGWQMEEADDWLR, were detected only in Ph-b
digests from old rats, whereas sequences containing nitrated
Tyr113, TLQNTMVNLALENACDEATY(NO2)QLDMEEL
EEIEEDAGLGNGGLGR, were identified in Ph-b from both
young and old animals at sufficiently high scores. For the latter
nitropeptide, multiple isoforms containing oxidized Met and/or
Cys residues were also observed although no age-dependent
tendencies in the accumulation of these oxidized peptides were
detected. For comparison and the generation of authentic
standards, we performed the reaction of purified rat Ph-b with
peroxynitrite in vitro.
3.4. Comparison of in vivo with in vitro nitration
Incubation of Ph-b isolated from 6 months old rat with
1 mM peroxynitrite resulted in an average formation of ca.
4.5 mol 3-NT/mol Ph-b, as monitored by RP-HPLC coupled to
UV detection (chromatograms not presented). At the same
time, multiple 3-NT-conaining peptides were detected from in-
gel tryptic digests. We found that the Xcorr values for
matching MS/MS data are useful to determine nitration site
selectivity. The ‘XCorr’ value represents a score calculated by
the Sequest algorithm, which cross-correlates experimental
MS/MS spectra from peptides with the theoretical mass spectra
produced from sequences generated from a protein database;
the higher the XCorr value, the higher the probability that a
given MS/MS spectrum belongs to a candidate peptide. Fig. 5
displays the XCorr values obtained for both Tyr and 3-NT-
containing Ph-b peptides showing protein tyrosine nitration at
multiple sites. For a qualitative analysis of the differential
sensitivity of individual Tyr residues towards nitration we used
only MS/MS spectra of XCorr values O1.5, O2.0, and O2.5
for Tyr- and 3-NT-containing peptides of the charge states
C1,C2 and C3, respectively. To compare the in vitro
nitration levels for individual Tyr residues, we classified
them into 3 groups: (1) significant levels of nitration, with
significant scores for the 3-NT-containing peptides (Tyr at
positions 51, 52, 113, 185, 203, 262, 280, 404, 473, 731, and
732), (2) insignificant nitration levels, where XCorr values for
the 3-NT-containing peptides are insignificant whereas the
XCorr values for respective native Tyr-containing peptides are

Fig. 4. 3-NT-containing sequences, detected by NESI-MS/MS, on Ph-b isolated
from 34 months old rats. (A)–(C) Representative tandem MS spectra for 3 Tyr
nitration sites in the sequence of rat skeletal muscle Ph-b. Inserts show the
respective sequences and decoding of the spectra.
V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416 413
reliable (Tyr at positions 83, 84, 90, 155, 157, 161, 297, 511,
524, 548, 648, 726, 777, and 780), and (3) nitration levels that
cannot be resolved by our method, where the XCorr values for
both native and nitrated peptides were insignificant (Tyr at
positions 74, 75, 233, 374, 553, 573, 587, 613, 791, and 820).
Interestingly, all three of the 3-NT-containing peptides
detected in vivo did not fit to any single group classified
above. The Ph-b peptide containing Tyr113 can be significantly
nitrated in vitro. Peptides containing Tyr161 and Tyr537 were
not found in the nitrated form in vitro and not resolved at a
reliable score, respectively, although nitration in vitro yielded a
ca. 10 times higher levels of total 3-NT on Ph-b than those
observed in vivo (4.5 vs. 0.3 mol 3-NT per mol protein,
respectively). Most of the in vitro nitration-sensitive peptides
from group 1, except Tyr113-containing peptide, displayed no
significant accumulation of 3-NT in vivo. These data show that
the reactivity and access of peroxynitrite do not explain the
nitration of a given protein in vivo.
3.5. Analysis of reduced Cys residues in Ph-b
Determination of DTNB-reactive Cys residues did not
show a statistically significant age-dependent difference
between Ph-b purified from 6 and 34 months old rats (55.3G1.7 and 57.6G2.5 nmol SH/mg total protein, respectively).
Taking into account the Ph-b content in the fractions collected
after ion-exchange chromatography (ca. 60% based on gel
densitometry; Fig. 2A), we conclude that the Cys residues
in the purified Ph-b (ca. 8.9G0.3 and 9.3G0.4 mol Cys/mol
Ph-b, respectively) are virtually all in the reduced state. Our
values are close to the mouse muscle Ph-b content of 8 Cys per
protein molecule. The slight deviation of the experimental
values from the theoretical Cys number may be the result of
uncertainties in the gel densitometry and small level of
contaminating proteins. The HPLC analysis of ThioGlo1-
reactive Cys residues in Ph-B confirms these data (not shown).
This result is not surprising as the successful isolation of Ph-b
required the presence of a reductant (0.5 mM DTT), which
reduces disulfides and sulfenic acid. However, these data also
suggest that any potential oxidation of Ph-b in vivo does not
lead to a significant accumulation of Cys sulfinic and sulfonic
acid. Unfortunately, the isolation of Ph-b under non-reducing
conditions was accompanied by a significant drop in the protein
yields.
4. Discussion
This study confirms our previous observation derived from
proteomic experiments that the skeletal muscle glycogen
phosphorylase isoform PHS2 (encoded by the gene PYGM) is
a target for Tyr nitration in vivo and, furthermore, characterizes
the accumulation of 3-NT in the purified protein during
biological aging in an effort to correlate protein modification
with age-associated loss of protein function. Our earlier
proteomic study detected Ph-b nitration on Tyr113 in rat
skeletal muscle homogenate (Kanski et al., 2005b). The present
paper confirms the nitration of Tyr113 in the purified Ph-b and
presents MS/MS evidence for the in vivo nitration of Ph-b Tyr
residues at two additional locations, Tyr161 and Tyr573. Taking
into account that our MS method covered the in vitro 3-NT
formation on peptides containing 26 out of a total 35 Tyr
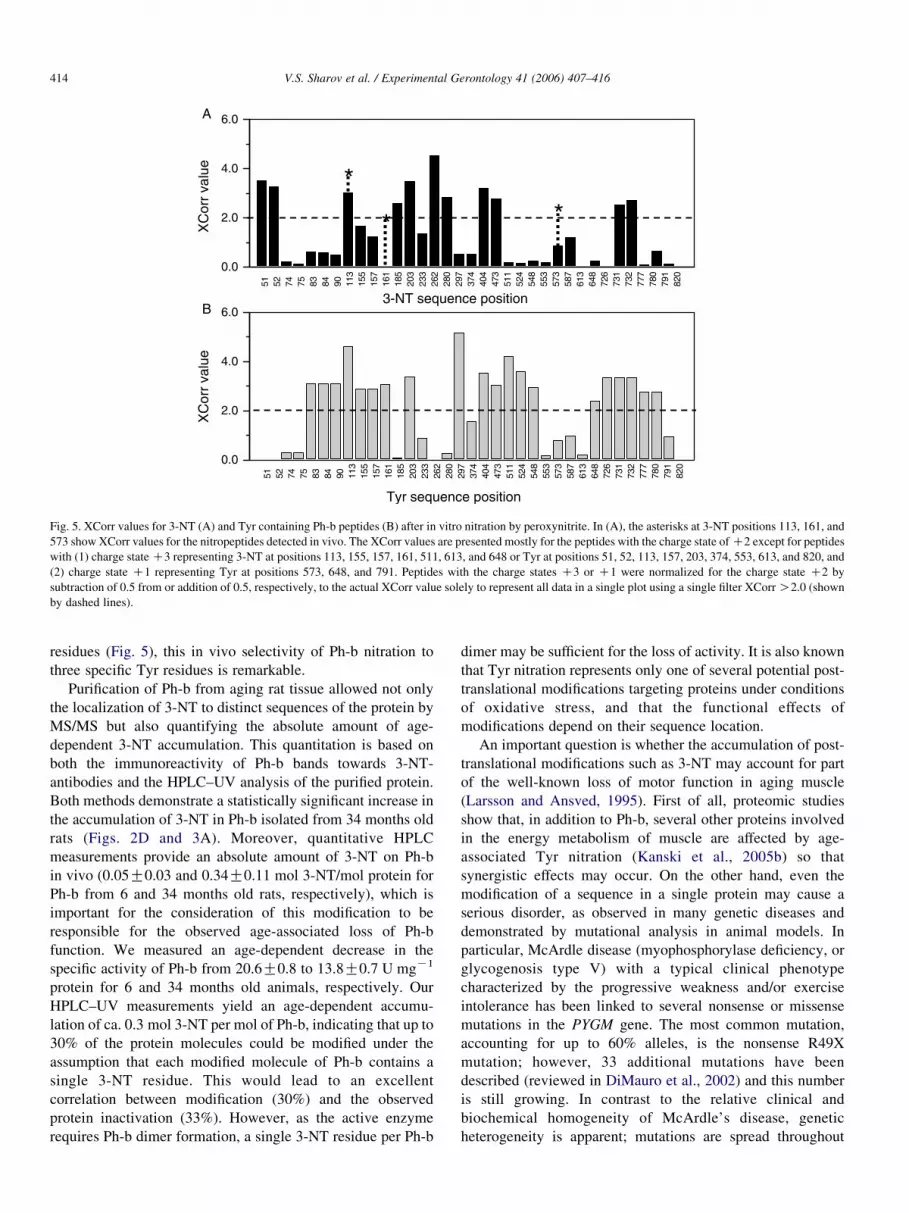
Fig. 5. XCorr values for 3-NT (A) and Tyr containing Ph-b peptides (B) after in vitro nitration by peroxynitrite. In (A), the asterisks at 3-NT positions 113, 161, and
573 show XCorr values for the nitropeptides detected in vivo. The XCorr values are presented mostly for the peptides with the charge state of C2 except for peptides
with (1) charge state C3 representing 3-NT at positions 113, 155, 157, 161, 511, 613, and 648 or Tyr at positions 51, 52, 113, 157, 203, 374, 553, 613, and 820, and
(2) charge state C1 representing Tyr at positions 573, 648, and 791. Peptides with the charge states C3 or C1 were normalized for the charge state C2 by
subtraction of 0.5 from or addition of 0.5, respectively, to the actual XCorr value solely to represent all data in a single plot using a single filter XCorr O2.0 (shown
by dashed lines).
V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416414
residues (Fig. 5), this in vivo selectivity of Ph-b nitration to
three specific Tyr residues is remarkable.
Purification of Ph-b from aging rat tissue allowed not only
the localization of 3-NT to distinct sequences of the protein by
MS/MS but also quantifying the absolute amount of age-
dependent 3-NT accumulation. This quantitation is based on
both the immunoreactivity of Ph-b bands towards 3-NT-
antibodies and the HPLC–UV analysis of the purified protein.
Both methods demonstrate a statistically significant increase in
the accumulation of 3-NT in Ph-b isolated from 34 months old
rats (Figs. 2D and 3A). Moreover, quantitative HPLC
measurements provide an absolute amount of 3-NT on Ph-b
in vivo (0.05G0.03 and 0.34G0.11 mol 3-NT/mol protein for
Ph-b from 6 and 34 months old rats, respectively), which is
important for the consideration of this modification to be
responsible for the observed age-associated loss of Ph-b
function. We measured an age-dependent decrease in the
specific activity of Ph-b from 20.6G0.8 to 13.8G0.7 U mgK1
protein for 6 and 34 months old animals, respectively. Our
HPLC–UV measurements yield an age-dependent accumu-
lation of ca. 0.3 mol 3-NT per mol of Ph-b, indicating that up to
30% of the protein molecules could be modified under the
assumption that each modified molecule of Ph-b contains a
single 3-NT residue. This would lead to an excellent
correlation between modification (30%) and the observed
protein inactivation (33%). However, as the active enzyme
requires Ph-b dimer formation, a single 3-NT residue per Ph-b
dimer may be sufficient for the loss of activity. It is also known
that Tyr nitration represents only one of several potential post-
translational modifications targeting proteins under conditions
of oxidative stress, and that the functional effects of
modifications depend on their sequence location.
An important question is whether the accumulation of post-
translational modifications such as 3-NT may account for part
of the well-known loss of motor function in aging muscle
(Larsson and Ansved, 1995). First of all, proteomic studies
show that, in addition to Ph-b, several other proteins involved
in the energy metabolism of muscle are affected by age-
associated Tyr nitration (Kanski et al., 2005b) so that
synergistic effects may occur. On the other hand, even the
modification of a sequence in a single protein may cause a
serious disorder, as observed in many genetic diseases and
demonstrated by mutational analysis in animal models. In
particular, McArdle disease (myophosphorylase deficiency, or
glycogenosis type V) with a typical clinical phenotype
characterized by the progressive weakness and/or exercise
intolerance has been linked to several nonsense or missense
mutations in the PYGM gene. The most common mutation,
accounting for up to 60% alleles, is the nonsense R49X
mutation; however, 33 additional mutations have been
described (reviewed in DiMauro et al., 2002) and this number
is still growing. In contrast to the relative clinical and
biochemical homogeneity of McArdle’s disease, genetic
heterogeneity is apparent; mutations are spread throughout

V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416 415
the gene and there is no clear genotype-phenotype correlation
(Martin et al., 2001). An important message from the genetic
studies is that, besides nonsense mutations generating
truncated protein variants (R49X, E124X, and Q754X), there
are different missense mutations of single amino acids, which
affect Ph-b function, e.g. L115P, P193W, G204S, L291P,
E348K, L396P, T487N, K542T, R601W, E654K, A659D,
Q665E, N684Y, G685R, A686P, A703V, W797R, and the
deletion of F708 (cf. SwissProt database entry P11217,
webaddress: http://ca.expasy.org/uniprot/PHS2_HUMAN).
Based on structural studies, it is well-recognized that some
Ph-b domains play a more important role in the regulation of
the protein activity, dependent on their proximity to (i) Ser14
(phosphorylation site responsible for the transformation of
homodimeric Ph-b to Ph-a, the tetrameric form of higher
specific activity); (ii) Tyr75 and, probably, Tyr155 (putative
AMP binding sites involved in the allosteric control of Ph-b
function); (iii) Cys108 and Cys142 (Involved in the association
of Ph-b homodimer subunits); and (iv) Lys680 (covalent
binding site for the cofactor pyridoxal phosphate, PLP)
(Barford et al., 1991; Titani et al., 1977). From our data, we
cannot decide, which of the three in vivo nitrated Tyr residues
may be functionally more important. Two of them may
potentially disturb local interactions within Ph-b domains
involved in the allosteric control and homodimerization due to
sequence location close to Tyr155 (for 3-NT at position 161)
and Cys108 (for 3-NT at position 113), respectively. The 3-NT
residue at position 573 is located far from functionally
important sites based on the protein sequence. However,
inspection of the 3D structure of Ph-b in the T-state PDB code:
1GPB; Barford et al., 1991) shows a close spatial proximity of
Tyr573 and Lys680-PLP (the distance between Tyr and
phosphate group amounts to only 5.7 A). Hence, the formation
of the three 3-NT residues on Ph-b may significantly affect the
enzyme activity. The vulnerability of Ph-b to Tyr nitration may
partially originate from the high abundance of Tyr residues in
the sequence (37 out of total 841 amino acids, or 4.4% for rat
muscle Ph-b). In addition, the accumulation of posttransla-
tional modifications on Ph-b may be favored by its relatively
long half-life of up to 11 days (Butler et al., 1985), and further
promoted by an even lower rate of the Ph-b turnover under
conditions of muscle dystrophy and aging (Leyland and
Beynon, 1991; Flannery et al., 1992).
Interestingly, the in vitro nitration by peroxynitrite does not
completely mimic the selectivity of nitration in vivo. In vitro,
multiple nitration sites were detected, among which was only
one out of the three nitration sites detected in vivo, Tyr113. A
similar observation was recently reported for skeletal muscle
cytosolic creatine kinase (Kanski et al., 2005b). These data
indicate the possibility of alternative nitration pathways in vivo
(Greenacre and lschiropoulos, 2001; Gow et al., 2004) and a
potentially site-specific 3-NT repair by the putative ‘denitrase’
(Irie et al., 2003). In addition, only some nitrated Ph-b isoforms
may be selected for turnover, potentially leading to the
accumulation of nitrated Ph-b isoforms, which are not subject
to protein turnover. At last, the apparent discrepancy between
ex vivo and in vitro tyrosine nitration profiles may originate
from different conformational states and/or protein interactions
that are likely to occur in vivo where Ph-b structural properties
can be affected by effector molecules and phosphorylation state
(Barford et al., 1991).
Acknowledgements
This work was supported by grants from the NIH (AG12993
and AG23551). The Q-TOF-2TM was purchased with support
from NSF EPSCoR and the University of Kansas. The Waters
CapLC and ProteinLynx Global Server were purchased for KU
by the Kansas City Area Life Sciences Institute.
References
Arking, R., Burde, V., Graves, K., Hari, R., Feldman, E., Zeevi, A., Soliman, S.,
Saraiya, A., Buck, S., Vettraino, J., Sathrasala, K., Wehr, N., Levine, R.L.,
2000. Forward and reverse selection for longevity in Drosophila is
characterized by alteration of antioxidant gene expression and oxidative
damage patterns. Exp. Gerontol. 35, 167–185.
Aslan, M., Ryan, T.M., Townes, T.M., Coward, L., Kirk, M.C., Barnes, S.,
Alexander, C.B., Rosenfeld, S.S., Freeman, B.A., 2003. Nitric oxide-
dependent generation of reactive species in sickle cell disease. Actin
tyrosine induces defective cytoskeletal polymerization. J. Biol. Chem. 278,
4194–4204.
Barford, D., Hu, S.H., Johnson, L.N., 1991. Structural mechanism for glycogen
phosphorylase control by phosphorylation and AMP. J. Mol. Biol. 218,
233–260.
Bergmeyer, H.U., Gawehn, K., 1974. Methods of enzymatic analysis, vol. 1.
Academic Press, New York pp. 505–506.
Butler, P.E., Cookson, E.J., Beynon, R.J., 1985. The turnover of skeletal muscle
glycogen phosphorylase studied using the cofactor, pyridoxal phosphate, as
a specific label. Biochim. Biophys. Acta 847, 316–323.
Cabiscol, E., Levine, R.L., 1995. Carbonic anhydrase III. Oxidative
modification in vivo and loss of phosphatase activity during aging.
J. Biol. Chem. 270, 14742–14747.
Castegna, A., Aksenov, M., Aksenova, M., Thongboonkerd, V., Klein, J.B.,
Pierce, W.M., Booze, R., Markesbery, W.R., Butterfield, D.A., 2002.
Proteomic identification of oxidatively modified proteins in Alzheimer’s
disease brain. Part I: creatine kinase BB, glutamine synthase, and ubiquitin
carboxy-terminal hydrolase L-1. Free Radic. Biol. Med. 33, 562–571.
Das, N., Levine, R.L., Orr, W.C., Sohal, R.S., 2001. Selectivity of protein
oxidative damage during aging in Drosophila melanogasterSelectivity of
protein oxidative damage during aging in Drosophila melanogaster.
Biochem. J. 360, 209–216.
DiMauro, S., Andreu, A.L., Bruno, C., Hadjigeorgiou, G.M., 2002. Myopho-
sphorylase deficiency (glycogenosis type V; McArdle disease). Curr. Mol.
Med. 2, 189–196.
Dremina, E.S., Sharov, V.S., Schoneich, C., 2005. Protein tyrosine nitration in
rat brain is associated with raft proteins, flotillin-1 and alpha-tubulin: effect
of biological aging. J. Neurochem. 93, 1262–1271.
Drew, B., Leeuwenburgh, C., 2002. Aging and the role of reactive nitrogen
species. Ann. NY Acad. Sci. 959, 66–81.
Ducret, A., Van Ooxtveen, I., Eng, J.K., Yates III.., J.R., Aebersold, R.,
1998. High throughput protein characterization by automated reverse-
phase chromatography/electrospray tandem mass spectrometry. Prot. Sci.
7, 706–719.
Ellman, G.L., 1959. Tissue sulfhdryl groups. Arch. Biochem. Biophys. 82,
70–77.
Fernandes, G., Venkatraman, J.T., Turturro, A., Attwood, V.G., Hart, R.W.,
1997. Effect of food restriction on life span and immune functions in long-
lived Fischer-344 ! Brown Norway F1 rats. J. Clin. Immunol. 17, 85–95.
Fisher, E.H., Krebs, E.G., 1962. Muscle phosphorylase b. Methods Enzymol. 5,
369–373.

V.S. Sharov et al. / Experimental Gerontology 41 (2006) 407–416416
Flannery, A.V., Easterby, J.C., Beynon, R.J., 1992. Turnover of glycogen
phosphorylase in the pectoralis muscle of broiler and layer chickens.
Biochem. J. 286, 915–922.
Gafni, A., 1981. Purification and comparative study of glyceraldehyde-3-
phosphate dehydrogenase from the muscles of young and old rats.
Biochemistry 20, 6035–6040.
Greenacre, S.A., Ischiropoulos, H., 2001. Tyrosine nitration: localization,
quantification, consequences for protein function and signal transduction.
Free Rad. Biol. Med. 34, 541–581.
Gao, J., Yin, D.H., Yao, Y., Sun, H., Qin, Z., Schoneich, C., Williams, T.D.,
Squier, T.C., 1998. Loss of conformational stability in calmodulin upon
methionine oxidation. Biophys. J. 74, 1115–1134.
Gow, A.J., Farkouth, C.R., Munson, D.A., Posencheg, M.A., Ischiropoulos, H.,
2004. Biological significance of nitric oxide-mediated protein modifi-
cations. Am. J. Physiol. Lung, Cell Mol. Physiol. 287, L262–L268.
Grune, T., Jung, T., Merker, K., Davies, K.J., 2004. Decreased proteolysis
caused by protein aggregates, inclusion bodies, plaques, lipofuscin, ceroid,
and ‘aggresomes’ during oxidative stress, aging, and disease. Int.
J. Biochem. Cell Biol. 36, 2519–2530.
Irie, Y., Saeki, M., Kamisaki, Y., Martin, E., Murad, F., 2003. Histone H1.2 is a
substrate for denitrase, an activity that reduces nitrotyrosine immunor-
eactivity in proteins. Proc. Natl Acad. Sci. USA 100, 5634–5639.
Ji, L.L., Dillon, D., Wu, E., 1990. Alteration of antioxidant enzymes with aging
in rat skeletal muscle and liver. Am. J. Physiol. 258, R918–R923.
Kanski, J., Alterman, M.A., Schoneich, C., 2003. Proteomic identification of
age-dependent protein nitration in rat skeletal muscle. Free Radic. Biol.
Med. 35, 1229–1239.
Kanski, J., Behring, A., Pelling, J., Schoneich, C., 2005a. Proteomic
identification of 3-nitrotyrosine-containing rat cardiac proteins: effects of
biological aging. Am. J. Physiol. Heart Circ. Physiol. 288, H371–H381.
Kanski, J., Hong, S.J., Schoneich, C., 2005b. Proteomic analysis of protein
nitration in aging skeletal muscle and identification of nitrotyrosine-
containing sequences in vivo by nanoelectrospray ionization tandem mass
spectrometry. J. Biol. Chem. 280, 24261–24266.
Knyushko, T.V., Sharov, V.S., Williams, T.D., Schoneich, C., Bigelow, D.J.,
2005. 3-Nitrotyrosine-modification of SERCA2a in the aging heart: a
distinct signature of cellular redox environment. Biochemistry 44, 13071–
13081.
Larsen, P.L., 1993. Aging and resistance to oxidative damage in
Caenorhabditis elegans. Proc. Natl Acad. Sci. USA 90, 8905–8909.
Larsson, L., Ansved, T., 1995. Effects of ageing on the motor unit. Prog.
Neurobiol. 45, 397–458.
Lawler, J.M., Powers, S.K., Visser, T., van Dijk, H., Kordus, M.J., Ji, L.L.,
1993. Acute exercise and skeletal muscle antioxidant and methabolic
enzymes: effect of fiber type and age. Am. J. Physiol. 265, R1344–R1350.
Leeuwenburgh, C., Fiebig, R., Chandwaney, R., Ji, L.L., 1994. Aging and
exercise training in skeletal muscle: responses of glutathione and
antioxidant enzyme systems. Am. J. Physiol. 267, R439–R445.
Levine, R.L., 2002. Carbonyl modified proteins in cellular regulation, aging,
and disease. Free Radic. Biol. Med. 32, 790–796.
Leyland, D.M., Beynon, R.J., 1991. The expression of glycogen phosphorylase
in normal and dystrophic muscle. Biochem. J. 278, 113–117.
Martin, M.A., Rubio, J.C., Buchbinder, J., Fernandez-Hojas, R., del Hoyo, P.,
Teijeira, S., Gamez, J., Navarro, C., Fernandez, J.M., Cabello, A.,
Campos, Y., Cervera, C., Culebras, J.M., Andreu, A.L., Fletterick, R.,
Arenas, J., 2001. Molecular heterogeneity of myophosphorylase deficiency
(McArdle’s disease): a genotype–phenotype correlation study. Ann.
Neurol. 50, 574–581.
Mary, J., Vougier, S., Picot, C.R., Perichon, M., Petropoulos, I., Friguet, B.,
2004. Enzymatic reactions involved in the repair of oxidized proteins. Exp.
Gerontol. 39, 1117–1123.
Paik, D.C., Dillon, J., Galicia, E., Tilson, M.D., 2001. The nitrite/collagen
reaction: non-enzymatic nitration as a model system for age-related
damage. Connect. Tissue Res. 42, 111–122.
Parkes, T.L., Elia, A.J., Dickinson, D., Hilliker, A.J., Phillips, J.P.,
Boulianne, G.L., 1998. Extension of Drosophila lifespan by overexpression
of human SOD1 in motoneurons. Nature Genet. 19, 171–174.
Poggioli, S., Mary, J., Bakala, H., Friguet, B., 2004. Evidence of preferential
protein targets for age-related modifications in peripheral blood lympho-
cytes. Ann. NY Acad. Sci. 1019, 211–214.
Reiter, R.J., 1995. Oxidative processes and antioxidative defense mechanisms
in the aging brain. FASEB J. 9, 526–533.
Roepstoff, P., Folman, J., 1984. Proposal for a common nomenclature for
sequence ions in mass spectra of peptides. Biomed. Mass Spectrom. 11,
601.
Schoneich, C., 2003. Proteomics in gerontological research. Exp. Gerontol. 38,
473–481.
Sharov, V.S., Galeva, N.A., Knyushko, T.V., Bigelow, D.J., Williams, T.D.,
Schoneich, C., 2002. Two-dimensional separation of the membrane protein
sarcoplasmic reticulum Ca-ATPase for high-performance liquid chroma-
tography-tandem mass spectrometry analysis of posttranslational protein
modifications. Anal. Biochem. 308, 328–335.
Sohal, P.S., Agarwal, S., Dubey, A., Orr, W.C., 1993. Protein oxidative damage
is associated with life expectancy of houseflies. Proc. Natl Acad. Sci. USA
90, 7255–7259.
Stadtman, E.R., 1992. Protein oxidation and aging. Science 257, 1220–1224.
Starke-Reed, P.E., Oliver, C.N., 1989. Protein oxidation and proteolysis
during aging and oxidative stress. Arch. Biochem. Biophys. 275, 559–
567.
Sun, J., Tower, J., 1999. FLP Recombinase-mediated induction of Cu/Zn-
superoxide dismutase transgene expression can extend the life span of adult
Drosophila melanogaster flies. Mol. Cell Biol. 19, 216–228.
Titani, K., Koide, A., Hermann, J., Ericsson, L.H., Kumar, S., Wade, R.D.,
Walsh, K.A., Neurath, H., Fischer, E.H., 1977. Complete amino acid
sequence of rabbit muscle glycogen phosphorylase. Proc. Natl Acad. Sci.
USA 74, 4762–4766.
van der Loo, B., Labugger, R., Skepper, J.N., Bachschmid, M., Kilo, J.,
Powell, J.M., Palacios-Callender, M., Erusalimsky, J.D., Quaschning, T.,
Malinski, T., Gygi, D., Ullrich, V., Luscher, T.F., 2000. Enhanced
peroxynitrite formation is associated with vascular aging. J. Exp. Med.
192, 1731–1744.
Vigouroux, S., Briand, M., Briand, Y., 2004. Linkage between the proteasome
pathway and neurodegenerative diseases and aging. Mol. Neurobiol. 30,
201–221.
Viner, R.I., Ferrington, D.A., Huhmer, A.F., Bigelow, D.J., Schoneich, C.,
1996. Accumulation of nitrotyrosine on the SERCA2a isoform of SR Ca-
ATPase of rat skeletal muscle during aging: a peroxynitrite-mediated
process? FEBS Lett. 379, 286–290.
Viner, R.I., Ferrington, D.A., Aced, G.I., Miller-Schlyer, M., Bigelow, D.J.,
Schoneich, C., 1997. In vivo aging of rat skeletal muscle sarcoplasmic
reticulum Ca-ATPase. Chemical analysis and quantitative simulation by
exposure to low levels of peroxyl radicals. Biochim. Biophys. Acta 1329,
321–335.
Viner, R.I., Ferrington, D.A., Williams, T.D., Bigelow, D.J., Schoneich, C.,
1999. Protein modification during biological aging: selective tyrosine
nitration of the SERCA2a isoform of the sarcoplasmic reticulum Ca2C-
ATPase in skeletal muscle. Biochem. J. 340, 657–669.
Vorgerd, M., Kubisch, C., Burwinkel, B., Reichmann, H., Mortier, W.,
Tettenborn, B., Pongratz, D., Lindemuth, R., Tegenthoff, M., Malin, J.P.,
Kilimann, M.W., 1998. Mutation analysis in myophosphorylase deficiency
(McArdle’s disease). Ann. Neurol. 43, 326–331.
Yan, L., Levine, R.L., Sohal, R.S., 1997. Oxidative damage during aging
targets mitochondrial aconitase. Proc. Natl Acad. Sci. USA 94, 11168–
11172.
Yan, L., Sohal, R.S., 1998. Mitochondrial adenine nucleotide translocase is
modified oxidatively during aging. Proc. Natl Acad. Sci. USA 95, 12896–
12901.
Ye, Y.Z., Strong, M., Huang, Z.Q., Beckman, J.S., 1996. Antibodies that
recognize nitrotyrosine. Meth. Enzymol. 269, 201–209.





























