Embed Size (px)
Citation preview

Biotechnol. Appl. Biochem. (2000) 31, 135–152 (Printed in Great Britain) 135
REVIEWAdvances in microbial amylases
Ashok Pandey*1, Poonam Nigam†, Carlos R. Soccol*, Vanete T. Soccol‡, Dalel Singh¶and Radjiskumar Mohan*
*Laboratorio de Processos Biotecnologicos, Departamento de Engenharia Quimica, Universidade Federal do Parana (UFPR),CEP 81531-970 Curitiba-PR, Brazil, †School of Applied Biological and Chemical Sciences, University of Ulster,Coleraine BT52 1AS, Northern Ireland, U.K., ‡Departamento de Patologia BaU sica, Universidade Federal do Parana (UFPR),CEP 81531-970 Curitiba-PR, Brazil, and ¶Department of Microbiology, Haryana Agriculture University, Hisar-125 004, India
This review makes a comprehensive survey of microbialamylases, i.e. α-amylase, β-amylase and glucoamylase.Amylases are among the most important enzymes andare of great significance in present-day biotechnology.Although they can be derived from several sources,such as plants, animals and micro-organisms, theenzymes from microbial sources generally meetindustrial demands. Microbial amylases could be po-tentially useful in the pharmaceutical and fine-chemicalindustries if enzymes with suitable properties could beprepared. With the advent of new frontiers in bio-technology, the spectrum of amylase application haswidened in many other fields, such as clinical, medicinaland analytical chemistries, as well as their widespreadapplication in starch saccharification and in the textile,food, brewing and distilling industries. In this review,after a brief description of the sources of amylases, wediscuss the molecular biology of amylases, describingstructures, cloning, sequences, and protoplast fusionand mutagenesis. This is followed by sections on theirproduction and finally the properties of variousamylases.
Introduction
Amylases [a term that refers here α-amylase, β-amylase andglucoamylase (GA)] are among the most important enzymesin present-day biotechnology. The amylase family ofenzymes is of great significance due to its wide area ofpotential application. Interestingly, the first enzyme pro-duced industrially was an amylase from a fungal source in1894, which was used as a pharmaceutical aid for the treat-ment of digestive disorders [1]. Amylases find potentialapplication in a number of industrial processes such as inthe food, fermentation, textiles and paper industries.Microbial amylases have successfully replaced the chemicalhydrolysis of starch in starch-processing industries. Theywould be potentially useful in the pharmaceutical and fine-
chemicals industries if enzymes with suitable propertiescould be prepared [2].
With the advent of new frontiers in biotechnology, thespectrum of amylase application has expanded into manyother fields, such as clinical, medicinal and analyticalchemistries. Recently Witczak [3] presented a review on thebiological relevance of thio-sugars as potential new thera-peutics, which are gaining substantial attention. The newdevelopments, especially in the synthetic and medicinalchemistry of thio-sugars are critically important for carbo-hydrate drug design. Enzyme-controlled mechanisms, in-volving enzymes such as amylase and lipase, have contributedto the understanding of the biological processes. There areseveral processes in the medicinal and clinical areas thatinvolve the application of amylases [4–14]. Sutton et al. [4]assessed 13 analytes, including amylase, for serum evaluation.Lepp et al. [5] developed a liquid-stable reagent containing α-amylase for the new Olympus AU6004 chemistry analyser.Chiu and Chandler [6] described a revised amylase methodfor the DuPont Dimension2 clinical chemistry system. Theapplication of a liquid-stable reagent, based on for the CibaCorning Express clinical chemistry system has been de-scribed by Becks et al. [7]. Giri et al. [8] developed a processfor the detection of higher oligosaccharides, which involvedapplication of α-amylase. The method was claimed to bemore efficient than the silver nitrate test, a method that islimited in its sensitivity. Menzel et al. [9] developedbiosensors with an electrolyte isolator semiconductorcapacitor (EIS-CAP) transducer for process monitoring.Amongst other enzymes this contained β-amylase and GAand was used for sugar syrup analysis. α-Amylase was used toprepare a hybrid membrane using chitosan as a dispersantin the sol–gel process [10]. α-Amylase was also used as anenzyme thermistor for the biochemical analysis of cyclo-dextrins [11]. GA has been used as chiral selector inchromatography [13].
Abbreviation used : GA, glucoamylase.1 Present address and address for correspondence : Biotechnology Division,
Regional Research Laboratory, Trivandrum-695 019, India.
# 2000 Portland Press Ltd

136 A. Pandey and others
Table 1 Microbial sources of α-amylase, β-amylase and GA
Source Reference
α-AmylaseAeromonas caviae [15]Alicyclobacillus acidocaldarius [16]Alteromonas haloplanetis [17]Anaerobic bacterium [18]Archaeobacterium pyrococcus woesei [19]Aspergillus sp. [20]
A awamorei [21,22]A. flavus [23]A. fumigatus [24]A. kawachi [25]A. niger [26,27]A. oryzae [28,29]A. usanii [30]
Bacillus sp. [31–33]B. acidocoldarius [34]B. amyloliquefaciens [35–38]B. brevis [39–41]B. circulans [42]B. coagulans [43]B. flavothermus [44]B. globisporus [45]B. licheniformis [39,42,46–49]B. megaterium [50]B. stearothermophilus [51]B. subtilis [52–54]
Chloroflexus aurantiacus [55]Clostridium acetobutylicum [56]C. butricum [57]C. thermohydrosulfuricum [19]C. thermosulfurogenes [58,63]Eubacterium sp. [59]Filobasidium capsuligenum [60]Halobacterium halobium [61]H. salinarium [49]Humicola insolens [66]H. lanuginosa [66]H. stellata [66]Lactobacillus brevis [64]L. cellobiosus [65]Malbrachea pulchella var. sulfurea [66]Micrococcus luteus [67]M. varians [68]Micromonospora vulgaris [69]Mucor pusillus [66]Myceliophthora thermophila [70]Myxococcus coralloides [71]Nocardia asteroides [72]Penicillium brunneum [73]Pseudomonsa stutzeri [74]Pycnoporus sanguineus [75]Pyrococcus woesei [76]Rhizopus sp. [77]Scytalidium sp. [78]Schizophyllum commune [79]Talaromyces thermophilus [66]Thermus sp. [81]Thermoactinomyces sp. [78]
T. vulgaricus [81]Thermococcus profundus [82]Thermomonospora viridis [83]Thermonospora curvata [84]T. vulgaris [85]Thermomyces lanuginosus [86]Thermotoga maritima [87]
β-AmylaseBacillus cereus [88]
B. circulans [89]B. megatarium [78,90]B. polymyxa [88]Clostridium thermosulfurogenes [91,92]C. thermocellum [93]Pseudomonas sp. [88]Rhizopus japonicus [88]Streptomyces sp. [88]
GAAcremonium zonatum [94]Amylomyces rouxii [94]Arxula adeninivorans [95]Aspergillus sp. [96]
A. awamori [94,97–100]A. candidus [94]A. foetidus [94]A. niger [94,101–111]A. oryzae [112,113]A. phoenicus [114]A. saitri [94]A. terreus [115,116]
Bacillus firmus/lentus [117]B. stearothermophillus [94]Candida famata [94]C. fennica [94]Cephalosporium eichhorniae [94]C. charticola [94]Chalara paradoxa [94]Clostridium sp. [94]
C. acetobutylicum [94]C. thermohydrosulfuricum [118]C. thermosaccharolyticum [119]C. thermosulfurogenes [94]
Collectotrichum gloesporoides [120]Coniophora cerebella [94]Endomyces sp. [94]Endomycopsis capsularis [94]E. fibuligera [121,122]Flavobacterium sp. [94]Fusidium sp. [123]Halobacter sodamense [94]Humicola lanuginosa [94]Lactobacillus brevis [124]Monscus kaoling [94]Mucor rouxianus [94]M. javanicus [94]Neurospora crassa [94]N. sitophila [125]Paecilomyces globosus [94]P. varioti [94]Penicillium italicum [94]P. oxalicum [94]Piricularia oryzae [94]Rhizoctania solani [94]Rhizopus sp. [121,126]
R. delemar [127,128]R. javanicus [94]R. niveus [35,94]R. oligospora [94]R. oryzae [127,128]
Rhizooctonia solani [109]Saccharomyces diastaticus [129]Schizophyllum commune [94]Schwanniomyces alluvius [94]S. occidentalis [130]Thermomyces lanuginosus [94]Torula thermophilla [94]Trichoderma reesei [131]T. viride [94]
# 2000 Portland Press Ltd

Microbial amylases 137
Sources of amylases
Amylases have most widely been reported to occur inmicro-organisms, although they are also found in plants andanimals. Two major classes of amylases have been identifiedin micro-organisms, namely α-amylase and GA. In addition,β-amylase, which is generally of plant origin, has also beenreported from a few microbial sources. α-Amylases (endo-1,4-α-D-glucan glucohydrolase, EC 3.2.1.1) are extracellularenzymes that randomly cleave the 1,4-α-D-glucosidic linkagesbetween adjacent glucose units in the linear amylose chain.These are endoenzymes that split the substrate in the in-terior of the molecules and are classified according to theiraction and properties. For example, amylases that producefree sugars are termed ‘saccharogenic ’ and those thatliquefy starch without producing free sugars are known as‘ starch-liquefying ’. β-Amylase (α-1,4-glucan maltohydrolase,EC 3.2.1.2) is usually of plant origin, but a few microbialstrains are also known to produce it. It is an exo-actingenzyme that cleaves non-reducing chain ends of amylose,amylopectin and glycogen molecules. It hydrolyses alternateglycosidic linkages, yielding maltose (β-anomeric form). Sinceβ-amylase is unable to by-pass α-1,6-glycosidic linkagesin amylopectin, it results in incomplete degradation of themolecule, yielding 50–60% maltose and a β-limit dextrin. GA(synonyms amyloglucosidase, ‘ glucogenic enzyme’, ‘ starchglucogenase ’ and ‘γ-amylase ’ ; exo-1,4-α-D-glucan glucano-hydrolase, EC 3.2.1.3) hydrolyses single glucose units fromthe non-reducing ends of amylose and amylopectin in astepwise manner. Unlike α-amylase, most glucoamylases arealso able to hydrolyse the 1,6-α-linkages at the branchingpoints of amylopectin, although at a lower rate than 1,4-linkages. Thus glucose, maltose and limit dextrins are theend products of GA action.
α-Amylaseα-Amylase may be derived from several bacteria, yeasts andfungi. Bacterial amylase, however, is generally preferred overfungal amylase due to several characteristic advantages thatit offers. Table 1 enumerates important microbial sources ofα-amylase [15–87]. Strains of Aspergillus sp. and Bacillus sp.,mainly Bacillus amyloliquefaciens and B. licheniformis, areemployed for commercial applications. Thermostable α-amylases are generally preferred as their application mini-mizes contamination risk and reduces reaction time, thusprovidingconsiderableenwergysaving.Hydrolysiscarriedoutat higher temperatures also minimizes polymerization of D-glucose to iso-maltose.
β-AmylaseUnlike other members of the amylase family, only a fewattempts have been made to study β-amylases of microbial
origin, as β-amylases have generally been obtained from plantsources. As is evident from the Table 1, bacterial strainsbelonging to Bacillus, Pseudomonas (aerobic) and Clostridium(anaerobic) sp., actinomycete strains belonging to Strepto-myces sp. and fungal strains belonging to Rhizopus sp. havebeen reported to synthesize β-amylase [88–93].
GAPandey [94] has reviewed the main features of GA research,providing insight into various aspects, including the sourcesof the enzyme. GA can be derived from a number of sources,such as plants, animals and micro-organisms. Commercialneed, however, is met by GA obtained from microbialsources. Filamentous fungi apparently constitute themajor source of GA among all microbes. Table 1 lists somemicro-organisms used as the sources of GA [95–131].
Molecular biology of amylases
Gene structure, cloning and sequencingGenetic engineering has been used extensively for cloning ofamylase-producing strains, mainly α-amylase and GA, inorder to achieve desirable characteristics in the cloned host.The purpose of gene cloning can be, amongst others, theexpression of thermostable enzymes, higher enzyme pro-ductivity and co-expression of two enzymes by the sameorganism.
A great deal of work has been done on the cloning of α-amylase genes in different microbes, mostly in Escherichia colior Saccharomyces cerevisiae (Table 2). Liebl et al. [87]described the gene structure of the α-amylase from Thermo-toga maritima MSB8. It is a chromosomal α-amylase gene,designated amyA, and was predicted to code for a 553-aminoacid preprotein with significant amino acid sequence. The T.maritima α-amylase appeared to be the first known exampleof a lipoprotein α-amylase. Following the signal peptide, a 25-residue putative linker sequence rich in serine and threonineresidues was found. The amylase gene was expressed in E.coli. Suganuma et al. [30] studied the N-terminal sequence ofthe amino acids of the α-amylase from Aspergillus usanii. Thesequence of the first 20 amino acids was identical to the α-amylase from A. niger. Kim et al. [132] described a geneencoding a new α-amylase of Bacillus licheniformis, which wascloned and expressed in E. coli. The genomic DNA of B.licheniformis was double-digested with EcoRI and BamHI andligated the pBR322. The transformed E. coli carried therecombinant plasmid pIJ322 containing a 3.5-kb fragment ofB. licheniformis DNA. The purified enzyme encoded by pIJ322was capable of hydrolysing pullulan and cyclodextrin as wellas starch. Iefuji et al. [133] described the cloning andsequencing of a raw-starch-digesting and thermostable α-
# 2000 Portland Press Ltd

138 A. Pandey and others
Table 2 Expression of cloned genes for microbial amylases : some examples
Source of gene Recombinant host References
α-AmylaseThermotoga maritima Escherichia coli [87]Alicyclobacillus acidocaldarius E. coli [16]Bacillus lichenformis E. coli [132]B. subtilis Saccharomyces
cerevisiae[138]
B. amyloliquefaciens S. cerevisiae [140]B. stearothermophilus E. coli [141]B. amyloliquefaciens S. cerevisiae [140]B. subtilis Xanthomonas
campestris[142]
B. subtilis E. coli [134]B. lichenformisl E. coli [41]B. lichenformisl B. brevis [41]B. stearothermophilus E. coli [164]B. stearothermophilus E. coli [165]S. cerevisiae S. cerevisiae [166]Saccharomycopsis fibuligera S. cerevisiae [167]
GAAspergillus awamori S. cerevisiae [138]A. awamori S. cerevisiae [152]Schwanniomyces occidentalis S. cerevisiae [130]S. cerevisiae S. cerevisiae [154]Arxula adeninivorans S. cerevisiae [95]Saccharomyces diastaticus S. cerevisiae [155]A. awamori S. cerevisiae [156]A. awamori S. cerevisiae [157]S. diastaticus S. cerevisiae [140]Rhizopus S. cerevisiae [168]A. niger S. cerevisiae [169]A.oryzae S. cerevisiae [140]A. terreus E. coli [171]S. diastaticus Schizosaccharomyces
pombe[172]
S. cerevisiae Saccharomyces pombe [173]S. fibuligera S. cerevisiae [173]Clostridium sp. E. coli [174]Lactobacillus amylovorus E. coli [145]
amylase from Cryptococcus sp. S2. An open reading frame ofthe cDNA specified 611 amino acids, including a putativesignal peptide of 20 amino acids. The N-terminal region ofthe enzyme (from the N-terminus to position 496) shared49.7% similarity with that of an α-amylase from A. oryzaewhile the C-terminus had a sequence similar to the C-terminal region of GA from A. niger. Marco et al. [134]inserted a B. subtilis α-amylase gene into a plasmid, which wastransferred to E. coli. During the cloning, a 3« region encoding171 C-terminal amino acids was replaced by a nucleotidesequence that encoded 33 amino acid residues not presentin the indigenous protein. The transformed proteinproduced substantial amylolytic activity. Matsuura et al.[135] and Tada et al. [136] have described the amino acidsequence and three-dimensional structure of α-amylase.
To develop a yeast strain that could produce ethanoldirectly from starch, α-amylase cDNA was introduced intothe haploid Saccharomyces diastitcus secreting GA by using alinearized integrating vector [137]. The integrating vectorcontained a LEU2 gene and KpnI was used to cut the LEU2
gene to make the linearized vector. One of the transformantsexhibited 100% mitotic stability after 100 generations of cellmultiplication. To improve its ethanol fermentability, thehaploid transformant was rare-mated with a polyploidindustrial strain with no amylase activity. The resultinghybrid RH51 gave high ethanol yields. Birol et al. [138]cloned three S. cerevisiae strains (YPG}AB, YPG}MM andYPB-G). YPB-G secreted a bifunctional fusion protein thatcontained the B. subtilis α-amylase and Aspergillus awamoriGA. When these strains were used for ethanol productionfrom different substrates such as glucose and starch,YPG}AB showed the most efficient utilization of starch forethanol production. The superior performance of theYPG}AB as compared with YPB-G was found to correlatewith its higher level of α-amylase activity. Shiba et al. [139]investigated the effect of ethanol concentration on cloned α-amylase gene expression in recombinant S. cerevisiae strain20B-12 containing one of the two plasmids, pNA3 andpNA7. Both the plasmids contained α-amylase gene underthe control of the SUC2 and PGK promoters, respectively.When the ethanol concentration was 2–5 g}l, the geneexpression was twice as high as when ethanol concentrationwas 20 g}l.
Steyn and Pretorius [140] cloned an α-amylase-encoding gene (AMY) from B. amyloliquefaciens and aGA-encoding gene (STA2) from S. diastaticus into a yeast-integrating shuttle vector (Yip5), generating recombinantplasmids pSP1 and pSP2. The STA2 and AMY genes werejointly cloned into Yip5, generating plasmid pSP3. Subse-quently, the dominant selectable marker APH1, encodingresistance to Geneticin 418, was cloned into pSP3 resultingpSP4. For enhanced expression of Geneticin 418, the APH1gene was fused to the GAL10 promoter and terminated bythe URA3 terminator, resulting in pSP5. Plasmid pSP5 wasconverted to a circular minichromosome (pSP6) by theaddition of ARS1 and CEN4 sequences. Laboratory strains ofS. cerevisiae transformed with plasmids pSP1–pSP6 stablyproduced several α-amylases and GAs.
Miranda and Berglund [141] described a B. stearo-thermophilus α-amylase expressed in E. coli y the use of afood-grade polymer. Stripecke et al. [142] studied the role ofa cloned α-amylase gene in fermentation by Xanthomonascampestris, which was prepared by transforming a hybridplasmid pAP1 containing the α-amylase gene from B. subtilisinto amylolytic and non-amylolytic X. campestris cells. Juge etal. [143] described α-amylase production by heterologousgene expression in A. niger. The cDNA encoding α-amylaseisozyme 1 (AMY1) and its signal peptide was placed underthe control of the A. nidulans glyceraldehyde-3-phosphatedehydrogenase (GPD) promoter and the A. nidulans trpCgene terminator. The recombinant AMY1 was purified tohomogeneity. α-Amylase was from a plant source (barley)and this has been claimed to the first plant protein efficiently
# 2000 Portland Press Ltd

Microbial amylases 139
secreted and correctly possessed by A. niger using its ownsignal sequence.
Apparently, no attention has been paid to the cloning ofgenes for β-amylase of microbial origin, although a fewreports are available on its expression in E. coli, mostly fromBacillus sp. and at least one from Thermoanaerobacterium[144–148]. To express the cloned β-amylase cDNA in E. coliunder control of the tac promoter, Yoshigi et al. [149]constructed a plasmid pBETA92, which consisted of 6312 bp.Later, these authors [150] constructed plasmid pB927, whichwas used for the expression of a 7-fold increase inproduction of β-amylase with increased thermostability inE. coli. The intragenic amino acid replacements were foundto have simple additive effects on the thermostability of theenzyme.
Like α-amylase, the number of GA-encoding genes alsovaries between strains. In a recent work, James and Lee[151] reviewed the GA multigene family. A. awamori and A.niger, the two most frequently used strains for GA synthe-sis, generally produced two forms of GA but from onlyone structural gene. Rhizopus oryzae too possesses only onestructural gene. As is evident from Table 2, most of the workon GA cloning has been done in S. cerevisiae. Many of suchstudies were intended to develop an improved (cloned)yeast strain for industrial production of ethanol. Zhang et al.[152] developed a mathematical model for the aerobicculture of a recombinant yeast, which was formulated tostimulate plasmid loss and recombinant protein productionamongst other things. The yeast strain was S. cerevisiaecontaining the GA gene from A. awamori.
Erratt and Nasim [153] described a protocol for thegenetic engineering of a yeast strain (brewer’s yeast), whichcarried the S. cerevisiae var. diastaticus gene. These authorscloned the STA1 gene from S. cerevisiae using a modifiedYep13 plasmid vector and obtained the first patent for thegene-cloning procedure. Nakamura et al. [154] constructeda starch-fermenting yeast, S. cerevisiae SR93, by integrating aGA-producing gene (STA1) into the chromosome of S.cerevisiae SH1089. The GA was constitutively produced bythe recombinant yeast. The GA gene of the yeast Arxulaadeninvorans Ls3 was cloned from a genomic library andsequenced. The gene was localized to chromosome 2 fromAr. adeninvorans and comprised 1875 bp. The first 16 N-terminal amino acids represented the signal sequence forentering the endomembrane system. When the amino acidsequence of this GA was compared with GA from otherfungal sources, it showed homology with the GA from R.oryzae (32.6%), Sachharomycopsis fibuligera (23.1%), A. niger(22.1%) and S. diastaticus (15.4%). No homology wasdetected with the GA of Schwaninomyces occidentalis. Byusing the GAL1 promoter from S. cerevisiae within anautonomously replicating plasmid, it was possible to expressthe isolated Arxula GA gene in S. cerevisiae.
When a new allelic variant of the STA2 gene from S.diastaticus, designated as STA2(K) and coding for a secretedGA, was cloned, differences were revealed both in thestructural gene and in the promoter region (as comparedwith other STA genes). STA2(K) showed peculiar charac-teristics, such as a 1.1-kb natural deletion in its promoterlocated 189 nucleotides upstream of the translation startcodon and an Asn ! Asp single amino acid change within theputative active site of the encoded GA [155]. Chen et al.[156] expressed Asn-182 ! Ala A. awamori GA in S.cerevisiae, which showed increased thermostability.Suominen et al. [157] described the fusion of poly(asparticacid) tails of different lengths (5, 7 and 10 Asp residues) tothe GA of A. awamori into the N-terminus of the full-lengthmature GA (amino acids 1–616). Fusion proteins weredesignated as GAND5, GAND7 and GAND10. Three otherfusion proteins, designated GACD0, GACD5 and GACD7(0, 5 and 7 Asp residues, respectively) were also constructed.All of the charged tails showed the general sequence Met-Ala-Aspn-Tyr, where n was 0, 5, 7 or 10. The modified geneswere expressed in S. cerevisiae and the proteins weresecreted in the culture medium.
A few attempts have also been made to clone thebacterial GA gene. The GA gene from Clostridium sp. G0005was apparently the first bacterial GA gene to be cloned[151]. James et al. [158] cloned the GA gene fromLactobacillus amylovorus in E. coli. GA from the wild andrecombinant sources showed similar characters.
Protoplast fusion and mutagenesisProtoplast fusion and mutagenesis have been used widely byseveral workers as a tool of protein engineering to achievestrains with higher enzyme productivity or desiredcharacters. Svensson and Sogaard [159] reviewed the effectsof mutation on the structure and function of GA and relatedenzymes. Linardi et al. [160] described an intraspecificprotoplast fusion of amylase-producing strains of Candidafennica. Although protoplast fusion did not significantlystimulate the synthesis of GA in the fusants, the productionof α-amylase was increased (32%) by one of the auxotrophicmutants of the yeast.
Mutagenesis has mainly been employed to obtain GAhyper-producing strains. It can be effected with the help ofchemicals such as N-methyl-N«-nitro-N-nitrosoguanidine(NTG), or by radiation, such as UV radition. Suntornsuk andHang [161] reported strain improvement of R. oryzae for GAproduction using NTG as well as UV radiation. One mutant,3N4, produced more GA than the parent strain. Themutagenesis of a GA-producing strain of A. awamori resultedin a mutant that exhibited an 80% increase in enzymeproductivity [162]. Flory et al. [163] carried out randommutagenesis of A. awamori to generate thermosensitive
# 2000 Portland Press Ltd

140 A. Pandey and others
mutants, which then were expressed in S. cerevisiae.Hulseweh et al. [130] carried out site-directed mutagenesisto define the active site of the Sch. occidentalis GA. Themutated GAM1 genes were expressed in S. cerevisiae andtransformants were evaluated. Mutants were transcribedand translated similarly to wild-type GA. Therefore, alleffects on enzymic activity could be traced to single aminoacid substitutions. Asp-470 was shown to be essential forenzyme activity. Replacement of Asp-470 by glycine led to acomplete loss of activity. Substitution of Trp-468 by Alaaffected predominantly the α-1,6 activity and not α-1,4activity of the enzyme. Chen et al. [156] also reported thatsite-directed mutagenesis obtained thermostable GA. Table2 shows some other studies of expression of cloned genesfor amylase production [164–174].
Production of amylases
Although amylases can be produced by several micro-organisms, it remains a challenging task to obtain a straincapable of producing commercially acceptable yields. Selec-tion of a suitable strain is the most significant factor in theamylase production process. Sometimes a single strain canproduce more than one enzyme, i.e. α-amylase as well asGA. For example, the strains of A. niger can produce as manyas 19 enzymes, whereas α-amylase can be produced inreasonably good titres by as many as 28 strains [175].Commercial production of amylases is carried out in varioussteps, essentially because the environmental factors requiredfor the optimum growth of the micro-organism beingemployed for production may differ from those required forthe production of enzymes. These parameters includenutrient supplementation, pH of the medium, osmoticrelationship, degree of aeration, temperature and thecontrol of contamination during fermentation. Maintainingthe purity of the medium is also a very important factor,especially when the fermentation is carried out underaerobic conditions. Although the details of the specificfermentation processes adopted by different manufacturersvary, there remain two main methods for amylase pro-duction, submerged fermentation and solid-state fermen-tation. Solid-state fermentation has gained renewed interestfrom researchers for the production of these enzymes inview of its several economic and engineering advantagesand has been often employed to produce amylases[110,175–179]. Selvakumar et al. [180] reviewed microbialsynthesis of starch-saccharifying enzymes in solid cultures.
Since thermostability is a feature of most of the enzymessold in bulk for industrial application, thermophilic micro-organisms are of special interest for the production ofthermophilic amylases. Recent research on thermostableamylases, especially α-amylase, has concentrated on the
enzymes of thermophiles and extreme thermophiles. Be-tween the thermophilic and mesophilic groups lies the muchless common and largely unexplored facultative thermo-philes. Facultative thermophiles generally grow in themesophilic temperature range. Whereas their growth maybe optimum at 45 °C, they are capable of growing well athigher temperatures, thus covering both the mesophilic andthermophilic ranges. However, not much is known about theprocesses for enzyme production involving such organisms.Comparative analysis of the structure and physico-chemicalproperties of these two groups (mesophilic and thermo-philic) may provide some information about the molecularbasis of the high-temperature tolerance ability of thermo-stable amylases.
Production of α-amylaseBacillus species are considered to be the most importantsources of α-amylase and have been used for enzymeproduction using SSF [43,52,181] or SmF [35–37,44,51,182,183]. Lonsane and Ramesh [184] reviewed theproduction of bacterial thermostable α-amylase in SSF by B.amyloliquefaciens and B. licheniformis. They referred to theSSF process as the potential tool for achieving economy inenzyme production and starch hydrolysis.
Bajpai et al. [182] and Omidiji et al. [51] developedsimple and cheap media based on cheese whey, corn steepliquor and soya bean meal for α-amylase production. It wasclaimed that the medium could be exploited for the industrialproduction of α-amylase. Salva and Moraes [183] studied theeffects of different carbon sources on α-amylase production.Whereas lactose, dextran and soluble starch were foundsuitable for enzyme production, the highest enzyme yieldwas obtained when glucose was used. El Helow and ElGazaerly [54] compared α-amylase production in threedifferent nutritional media. Different patterns of enzymeinduction were obtained when beet pulp, corn cob, ricehusk, wheat bran and wheat straw were used separately topartially replace the nutrient contents of the selectedmedium. α-Amylase was maximally expressed in the pres-ence of corn cob or wheat bran. Syu and Chen [36] alsoinvestigated the effects of different carbon sources (glucose,maltose, xylose and starch) on α-amylase production. Highercell density and higher specific growth rate were obtainedfrom glucose but higher enzyme activity and higher specificenzyme activity were obtained from starch. Using a definedsynthetic medium, Hiller et al. [37] demonstrated the effect oflactose and nitrogen on cell physiology and α-amylase pro-duction. Results showed cell-growth and α-amylase-production patterns to be similar regardless of the limitingnutrient and suggested stationary phase gene control of α-amylase production as opposed to a direct response tonutrient limitation. Kelly et al. [44] described the production
# 2000 Portland Press Ltd

Microbial amylases 141
of α-amylase in a bi-phasic process in which high α-amylaseactivities were achieved. Oxygen-transfer conditions, andespecially the dissolved oxygen tension, were reported asvital factors for α-amylase production [35]. High aerationrates were found to be essential for good yields of enzyme.Control of dissolved oxygen tension, however, was notfound to be advantageous. Babu and Satyanarayana [43] alsoachieved maximum α-amylase yield in aerated reactors.
In view of the apparent advantages offered by cellrecycling in bioprocesses, Gron et al. [185] investigated α-amylase production in a cell-recycling bioreactor incor-porating a membrane-filtration module for cell separation.The reactor gave increased enzyme yield and volumetricproductivity compared with conventional continuous fer-mentation. Coupling of fermentation and microfiltration forα-amylase production was also studied by Morcel andBiedermann [186]. Compared with a batch process, con-tinuous fermentation with cell recycling led to a reduction inα-amylase concentrations but to a doubling of volumetricproductivities. Tari et al. [187] also demonstrated operationstrategies of a two-stage bioreactor for α-amylase pro-duction. They claimed that this would improve enzymeproduction.
During laboratory studies on enzyme production, as apractice, single-stage inoculum is used for fermentationprocesses. Generally, it is carried out in a routine waywithout being given critical attention. Keeping this in mind,Milner et al. [35] studied one-stage and two-stage inocula forα-amylase production. Significantly better results wereobtained using two-stage inoculum.
Since filamentous fungi are generally considered to bethe most prolific producers of extracellular enzymes,attempts have been made to study α-amylase productionusing them. The thermophilic fungus Thermomyces lanuginosawas reported to be an excellent producer of α-amylase[86,188]. Increased production of the enzyme could beobtained by manipulating the growth conditions and mediumcomposition. Sudo et al. [189] compared acid-stable α-amylase production in SmF and SSF and examined the reasonwhy A. kawachii IFO 4308 produced larger amounts of acid-stable α-amylase in SSF than in SmF. Some of the SSFcharacteristics were given as the major reasons for higherenzyme production in SSF. A new source of α-amylase wasidentified in Pycnoporus sanguineus. Cultivation in SSF resultedin 4-fold-higher enzyme production than in SmF [15,75]. deSouza et al. [77] tested more than 800 Rhizopus strains for α-amylase production. One of the strains showed great abilityto produce a thermostable α-amylase in SSF. Torrado et al.[29] studied pH regulation in solid-state cultures using theinitial ratio between oxidized and reduced sources ofnitrogen. On the basis of these, they suggested a model forα-amylase production. Krishna and Chandrasekaran [52]cultivated Aeromonas caviae (CBTK 185) on banana waste.
The results indicated the excellent scope for utilizing thisstrain and banana waste for commercial production of α-amylase in SSF.
The cell-immobilization technique has also beenemployed for α-amylase production. Ivanova et al. [190]compared various immobilization techniques including en-trapment in gels such as calcium alginate, κ-carrageenan, agarand their combinations with polyethylene oxide, adsorptionon cut disks of polymerized polyethylene oxide and fixationon formaldehyde-activated acrylonitrile}acrylamide mem-branes for α-amylase production. Among these, agar, κ-carrageenan, agar}polyethylene oxide gels and the mem-branes were found to be suitable. Membrane-immobilizedcells of B. licheniformis produced 176% more enzyme thanfree cells. Ariga et al. [41] studied continuous production ofα-amylase by PVA-encapsulated recombinant E. coli and B.brevis. In all the experiments with B. brevis, the enzyme wasreleased from the encapsulated cells without cell leakage andcell growth outside the capsules. In the case of immobilizedE. coli cells, it was necessary to add glycine to the medium forenzyme to be released. Stefanova et al. [40] used agar-gel-immobilized cells of B. brevis for the production of thermo-stable α-amylase. Dobreva et al. [46] used polymer mem-brane to immobilize B. licheniformis cells for the productionof α-amylase. The enzyme yields were affected by thereactive chemical groups of the carriers and the spacer size.Formaldehyde-activated polysulphone membranes were themost suitable for effective immobilization. A 62% increaseover the control experiment was obtained in repeated batch-mode operation. Lin et al. [38] described the design of acontinuously rotating bioreactor for immobilized cells of B.amyloliquefaciens to produce α-amylase. They claimed thatthe volumetric productivity of the enzyme in this bioreactorcould be much higher than in a shake flask, if the reactor wasoperated properly. The cells of a bacterial strain of Halo-bacterium salinarium were immobilized in calcium alginatebeads and on a polyvinyl alcohol film with an aim to producehalophilic α-amylase [62].
Production of β-amylaseAs mentioned previously, β-amylases are usually of plantorigin, and not much work has been done on the productionof β-amylase using micro-organisms. Some of the micro-organisms reported to produce β-amylases include Bacilluspolymyxa, B. cereus, B. megatarium, Streptomyces sp., Pseudo-monas sp. and Rhizopus japonicus [1,88].
Ray et al. [90] compared the production of β-amylasefrom starch waste by a hyper-amylolytic strain of B.megaterium B6 mutant UN12 in SmF and SSF. The starchywastes used as substrates were from arrowroot, arum,maize, potato, pulse, rice, rice husk, tamarind, kernel,cassava, water chestnut, wheat and wheat bran. Arum andwheat bran gave the highest yields. They also used
# 2000 Portland Press Ltd

142 A. Pandey and others
immobilized cells of B. megatarium B6 for enzyme pro-duction. Among the tested methods, ionotropic gelation wasfound to be most suitable for β-amylase production [191].
Production of GAExtensive work has been carried out on the productionof GA in solid cultures using A. niger [94,101–111,180,191–195]. The study included screening of a numberof agro-industrial residues such as wheat bran, rice bran,rice husk, gram flour, wheat flour, corn flour, tea waste,copra waste etc. [101,107,177,192]. Apart from the sub-strate particle size, which showed profound impact onfungal growth and activity, substrate moisture and wateractivity also influenced the enzyme yield significantly[94,105,111,191].
Strains of A. awamori too have been used frequently forGA production [100,196,197]. Queiroz et al. [100] con-ducted a study on GA production by A. awamori in order todefine the rheological changes of broth. They proposed twokinds of mathematical correlation ; one between the biomassconcentration and the rheological parameter consistencyindex (K) from the ‘Power law’, and another between eitherthe specific growth rate or the specific GA production rateand K. Wang and Webb [197] studied the effect of cellconcentration on the rheology of GA fermentation broth ofA. awamori using a Bohlin CS rheometer. By controlling thecell morphology, spore aggregation was prevented com-pletely. They also described the experimental results interms of the Power-law model.
Elegado and Fujio [198] screened 39 Rhizopus newisolates and nine authentic Rhizopus strains (grown on wheatbran in SSF) for their soluble-starch-digestive GA (SSGA)and raw-starch-digestive GA (RSGA) activities. Resultsshowed that these strains could be classified into fourgroups, based on their SSGA and RSGA production and ratioof SSGA to RSGA. Chiarello et al. [128] compared raw-starch-degrading GA production by the strains of R. oryzaeMUCL 28627, R. oryzae MUCL 28168 and R. delemar ATCC34612. R. oryzae MUCL 28627 presented the highestspecific activity values. Soccol et al. [126] screened 19Rhizopus strains for their ability to grow on raw cassava.Only three strains grew significantly and GA production washigher on raw cassava than on cooked cassava.
GA has also been produced using some bacterial andyeast cultures such as Lactobacillus brevis [124], B. coagulans[199], S. cerevisiae [154], Saccharomycopsis fibuligera [200], C.fennica, C. famata [201] and Endymycopsis fibuligera [202].
Design and configuration of fermenter, its mode ofoperation and fermentation influence the enzymeproduction strongly. Different types of bioreactor wereused to evaluate GA production. These included flasks,trays, rotary reactors and columns (vertical and horizontal)[103,108,192,195]. Fermentation techniques included SmF,
SSF, semi-solid and an aqueous two-phase system [203–205].Enzyme production in trays occurred in maximum quantitiesin 36 h compared with 96 h typically required in flasks [192].Ariff and Webb [98] studied GA production using freshlysuspended cells of A. awamori in batch and continuousoperations. They used an air-lift fermenter and a stirred-tankfermenter. In comparison with freely suspended cell fermen-tations, neither batch nor continuous fermentation ofimmobilized cells improved GA production significantly.Interestingly, contrary to the general findings, Fujio andMorita [204] reported 4.6-fold lower GA yield by Rhizopussp. A-11 from conventional SSF on wheat-bran medium thanthat in SmF, which used metal-ion-supplemented medium.Ramadas et al. [203] also noted a similar trend when a fungalstrain of A. niger produced higher GA titres in SmF (102units}ml) than in SSF (66 units}ml) in shorter period (66 h incomparison with 96 h). The production in the fed-batchprocess was higher in comparison with the batch process. Inan aqueous two-phase system of poly(ethylene glycol) andsalt, however, both the GA productivity and yield weretwice as high as those in the control fermentation.
Purification and properties of amylases
Enzyme application in pharmaceutical and clinical sectorsrequires high-purity amylases. Thus, it is significant todevelop economic processes for their purification to obtainchemically pure enzymes with maximum specific activity.Traditionally the purification of amylases from fermentationmedia has been done in several steps, which includecentrifugation of the culture (a step of extraction may berequired for solid media), selective concentration of thesupernatant, usually by ultrafiltration, and selective pre-cipitation of the enzyme by ammonium sulphate or organicsolvents such as ethanol in the cold. Then the crude enzymeis subjected to chromatography (usually affinity or ion-exchange chromatography) and gel filtration.
α-AmylaseBacterial α-amylase As bacterial α-amylases have generallybeen produced from the strains belonging to genus Bacillus,several attempts have been made at their purification andcharacterization, from both mesophilic and thermophilicstrains. Uguru et al. [206] described a thermostable extra-cellular α-amylase from B. subtilis, which was purified 24-foldto a specific activity of 2200 units}mg per litre. Bolton et al.[207] purified an α-amylase to homogeneity using a com-bination of ammonium sulphate precipitation, ion-exchangechromatography and gel filtration. The enzyme was inhibitedby diethyl pyrocarbonate. An extracellular thermostable α-amylase produced by B. licheniformis was purified by two-phase separation in a poly(ethylene glycol)}dextran system
# 2000 Portland Press Ltd

Microbial amylases 143
followed by gel filtration and ion-exchange chromatography[208]. Thermostability was Ca2+-dependent. The purifiedenzyme was inhibited strongly by N-bromosuccinimide andEDTA. Marco et al. [134] described a genetically modified α-amylase. Its molecular mass (48000 Da) was lower than themolecular mass calculated from the derived amino acidsequences of the parent strain (57700 Da). The enzyme wasinhibited by Hg2+, Fe3+ and Al3+ and stimulated by Mn2+ andCo2+. Katoh et al. [209] purified an α-amylase, produced byrecombinant cells, by specific elution from anti-peptideantibodies.
The fundamental characteristics of extracellularamylases are expected to reflect the pH and temperature ofthe environment in which they are grown [210]. This wasreflected with B. flavothermus grown at pH 6.0 and 55 °Cwhose α-amylase showed pH and temperature optima at5.5–6.0 and 60 °C [210]. Another example was an alkalophi-lic strain of Bacillus sp. GM8901, which best grew at pH 10.5and 50 °C and produced an α-amylase that showedpH and temperature optima at 11.0–12.0 and 60 °C [211].Another strain of Bacillus sp. grew best at pH 8.5 and the α-amylase produced by it had a pH optimum of 9.0 [212].
Industrial autofocusing has also been used as a tech-nological purification technique to obtain chemically pureamylases. Dobransky et al. [213] used this technique topurify α-amylase produced by different bacterial strains of B.subtilis. A natural pH gradient in autofocusing is usuallycreated automatically by the purified supernatant. Irrespec-tive of the undulations or the steps in the pH gradient, theproteins reach an appropriate pI along the pH gradient.The peaks are sufficiently separated and the void volume isfar enough from the peak containing most of the α-amylaseactivity. In this method, large amounts of the starting material(up to 1000 litres with a protein concentration of about3 g}100 ml) can be employed. It must be emphasized thatthis method should not be used in cases where the highestactivity of the enzyme corresponds with the main peak of theproteins [213].
Satoh et al. [214] purified an intracellular α-amylasefrom Streptococcus bovis [148], which was induced by maltoseand soluble starch and produced about 80% maltotriosefrom soluble starch. The enzyme showed different charac-teristics when compared with the extracellular enzyme fromthe same strain and there was no homology between them.An extracellular α-amylase from a different strain of S. bovisJB1 showed 40 and 27% homologies with α-amylases ofBacillus sp. and Clostridium acetobutylicum, respectively, andproduced maltose, maltotriose and maltotetraose fromamylose [215]. Ilori et al. [64,67] purified extracellular α-amylase from Micrococcus luteus and Lactobacillus brevis.Purification was by 28- and 70-fold by gel filtration and ion-exchange chromatographies, respectively. Enzyme fromboth the sources was inhibited by EDTA, KCN and citric
acid but was activated by Mg2+, Ca2+, Na+ and K+. ABifidobacterium adolescentis extracellular α-amylase wasinhibited significantly by EDTA, glucose, maltose, Cu2+, Zn2+
and N-bromosuccinimide but was activated by β-mercapto-ethanol [216].
A few reports also deal with the properties of theamylases produced by facultative thermophiles [207]. Itwould be interesting to know if they offer any technologicaladvantages in their application. In the first report of its kindon the characterization of α-amylase from a thermophilicstrain of the genus Thermus, Egas et al. [217] purified theenzyme by ion-exchange chromatography. Ca2+ enhancedthe thermostability at temperatures above 80 °C. EDTAinhibited the enzyme activity, which was reversed by theaddition of Ca2+ or Sr2+ ions. The enzyme was also inhibitedby Hg2+, Cu2+ and p-chloromercuribenzoic acid. Shaw et al.[80] also purified the α-amylase from Thermus sp. by affinityadsorption on starch granules. The enzyme activity wasinhibited strongly by Cu2+ and Fe2+ but Ca2+ did not stimulateits activity. A thermostable α-amylase from an archae-bacterium Thermococccus profundus was purified to hom-ogeneity by ammonium sulphate precipitation, DEAE–Toyopearl chromatography and gel filtration on Superdex200 HR. The thermostability of the enzyme was enhanced byCa2+ [82]. However, the thermostability or activity of α-amylase from another archaebacterium, Pyrococcus furiosus,was not dependent on Ca2+ [218]. The α-amylase fromAlicyclobacillus acidocaldarius showed an acidophilic nature[16]. A maltotriose- and maltotetraose-producing α-amylase, which was stable at alkaline pH (up to 12) and hightemperatures (up to 55 °C) was purified from a strain ofChloroflexux aurantiacus, a thermophilic, green, photo-synthetic bacterium [55].
Fungal α-amylase In fungi, detailed studies on α-amylasepurification have largely been limited to a few species ofmesophilic fungi [219–223]. Although the growth andamylase production from several thermophilic fungi havebeen described, extended data concerning their physico-chemical properties have been published with regard to onlya few species, such as Mucor pusilus [224], Talaromycesemersonii [225] and Thermomyces lanuginosa [86]. Thermo-stable α-amylase from T. lanuginosa [86] had a pH optimumabout 1 pH unit lower than the α-amylase from A. oryzae[226] and 1 pH unit higher than that from M. pusilus [222],but it resembled α-amylases from T. emersonii [223] and A.awamori [227].
Yeast α-amylase There are only a very few reportsdescribing the purification of α-amylase from yeasts. A raw-starch-digestive thermostable α-amylase from yeast Crypto-coccus sp. was purified in just one step by using an α-cyclodextrin–Sepharose 6B column [133]. The enzyme
# 2000 Portland Press Ltd
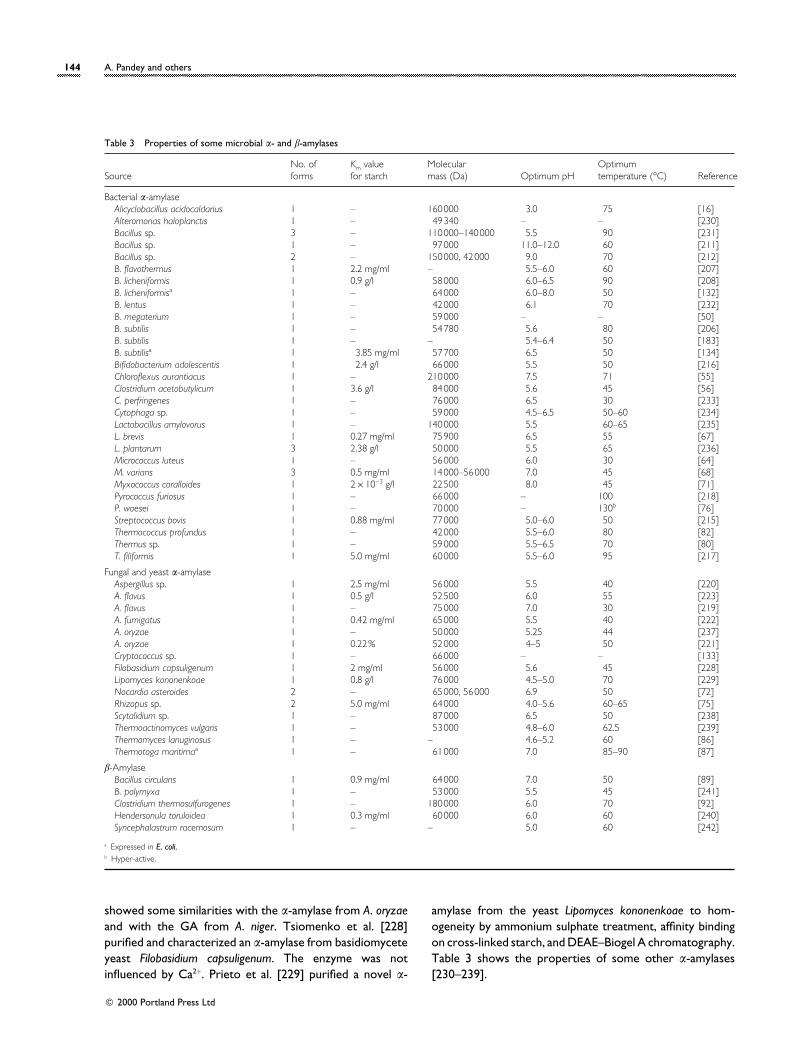
144 A. Pandey and others
Table 3 Properties of some microbial α- and β-amylases
SourceNo. offorms
Km valuefor starch
Molecularmass (Da) Optimum pH
Optimumtemperature (°C) Reference
Bacterial α-amylaseAlicyclobacillus acidocaldarius 1 – 160000 3.0 75 [16]Alteromonas haloplanctis 1 – 49340 – – [230]Bacillus sp. 3 – 110000–140000 5.5 90 [231]Bacillus sp. 1 – 97000 11.0–12.0 60 [211]Bacillus sp. 2 – 150000, 42000 9.0 70 [212]B. flavothermus 1 2.2 mg/ml – 5.5–6.0 60 [207]B. licheniformis 1 0.9 g/l 58000 6.0–6.5 90 [208]B. licheniformisa 1 – 64000 6.0–8.0 50 [132]B. lentus 1 – 42000 6.1 70 [232]B. megaterium 1 – 59000 – – [50]B. subtilis 1 – 54780 5.6 80 [206]B. subtilis 1 – – 5.4–6.4 50 [183]B. subtilisa 1 3.85 mg/ml 57700 6.5 50 [134]Bifidobacterium adolescentis 1 2.4 g/l 66000 5.5 50 [216]Chloroflexus aurantiacus 1 – 210000 7.5 71 [55]Clostridium acetobutylicum 1 3.6 g/l 84000 5.6 45 [56]C. perfringenes 1 – 76000 6.5 30 [233]Cytophaga sp. 1 – 59000 4.5–6.5 50–60 [234]Lactobacillus amylovorus 1 – 140000 5.5 60–65 [235]L. brevis 1 0.27 mg/ml 75900 6.5 55 [67]L. plantarum 3 2.38 g/l 50000 5.5 65 [236]Micrococcus luteus 1 – 56000 6.0 30 [64]M. varians 3 0.5 mg/ml 14000–56000 7.0 45 [68]Myxococcus coralloides 1 2¬10−3 g/l 22500 8.0 45 [71]Pyrococcus furiosus 1 – 66000 – 100 [218]P. woesei 1 – 70000 – 130b [76]Streptococcus bovis 1 0.88 mg/ml 77000 5.0–6.0 50 [215]Thermococcus profundus 1 – 42000 5.5–6.0 80 [82]Thermus sp. 1 – 59000 5.5–6.5 70 [80]T. filiformis 1 5.0 mg/ml 60000 5.5–6.0 95 [217]
Fungal and yeast α-amylaseAspergillus sp. 1 2.5 mg/ml 56000 5.5 40 [220]A. flavus 1 0.5 g/l 52500 6.0 55 [223]A. flavus 1 – 75000 7.0 30 [219]A. fumigatus 1 0.42 mg/ml 65000 5.5 40 [222]A. oryzae 1 – 50000 5.25 44 [237]A. oryzae 1 0.22% 52000 4–5 50 [221]Cryptococcus sp. 1 – 66000 – – [133]Filobasidium capsuligenum 1 2 mg/ml 56000 5.6 45 [228]Lipomyces kononenkoae 1 0.8 g/l 76000 4.5–5.0 70 [229]Nocardia asteroides 2 – 65000, 56000 6.9 50 [72]Rhizopus sp. 2 5.0 mg/ml 64000 4.0–5.6 60–65 [75]Scytalidium sp. 1 – 87000 6.5 50 [238]Thermoactinomyces vulgaris 1 – 53000 4.8–6.0 62.5 [239]Thermomyces lanuginosus 1 – – 4.6–5.2 60 [86]Thermotoga maritimaa 1 – 61000 7.0 85–90 [87]
β-AmylaseBacillus circulans 1 0.9 mg/ml 64000 7.0 50 [89]B. polymyxa 1 – 53000 5.5 45 [241]Clostridium thermosulfurogenes 1 – 180000 6.0 70 [92]Hendersonula toruloidea 1 0.3 mg/ml 60000 6.0 60 [240]Syncephalastrum racemosum 1 – – 5.0 60 [242]
a Expressed in E. coli.b Hyper-active.
showed some similarities with the α-amylase from A. oryzaeand with the GA from A. niger. Tsiomenko et al. [228]purified and characterized an α-amylase from basidiomyceteyeast Filobasidium capsuligenum. The enzyme was notinfluenced by Ca2+. Prieto et al. [229] purified a novel α-
amylase from the yeast Lipomyces kononenkoae to hom-ogeneity by ammonium sulphate treatment, affinity bindingon cross-linked starch, and DEAE–Biogel A chromatography.Table 3 shows the properties of some other α-amylases[230–239].
# 2000 Portland Press Ltd

Microbial amylases 145
Table 4 Properties of some microbial GAs
Source No. of forms Molecular mass (Da) Optimum pHOptimumtemperature (°C)
Raw starchdigestion Reference
Aspergillus awamori 1 83700 4.5 60 + [246]A. awamori 2 110000, 86000 – – - [99]A. awamori var. kawachi 3 57000–90000 3.8–4.5 – + [256]A. niger 2 74000, 96000 4.2, 4.5 60, 65 + [257]A. niger 4 61000–112000 4.4 60 + [100]A .niger 6 58400–99500 3.5–5.0 – +a [247]A. niger 2 99000–112000 4.5–5.0 – 0 [247]A. niger 3 70000–90000 3.5–4.0 50–60 0 [258]A. oryzae 3 38000–76000 4.5 50, 60 +a [259]A. phoenicus 1 – 4.5 60 0 [260]A. saitri 1 90000 4.5 – 0 [261]A. terreus 1 70000 5.0 60 0 [262]Cephalosporium charicola 1 – 5.4 60 0 [263]Coniphora cerebella 1 – 4.0–4.5 – 0 [244]Corticum rolfsii 1 – 4.5 40–50 0 [245]Clostridium 1 77000 4.5 65 0 [264]C. thermosaccharolyticum 1 75000 5.0 70 + [119]Endomyces sp. – 55000 – – 0 [265]Endomycopsis sp. – 53000 – – 0 [266]E. capsularis – – 4.5 40–50 0 [267]Humicola lanuginosa 2 – 4.9, 6.6 65, 70 0 [243]Monascus kaoligang 2 48000, 68000 4.5 50 0 [252]Mucor rouxianus 2 49000, 59000 4.7 55 + [246]Paecilomyces varioti 1 69000 5.0 – 0 [253]Penicillium oxalicum 2 84000, 86000 4.6 55–60 + [246]Piricularia oryzae 1 94000 4.5 50–55 0 [268]Rhizopus sp. 3 58600–74000 4.5–50 – + [269]R. delemer 1 100000 4.5 40 0 [270]R. niveus 5 – 4.5–6.0 – + [251]Schizophyllum commune 1 66000 5.0 40 + [255]Termomyces lanuginosa 1 37000 – 70 0 [254]
a Raw-starch digestion only by one form.
β-Amylaseβ-Amylases from microbial sources generally possess greaterthermal resistance than plant β-amylases. β-Amylases havehigher pH optima than α-amylases and they do not requireCa2+ for stabilization or enhancement of activity. Odibo et al.[240] purified and characterized a β-amylase fromHendersonula toruloidea by ammonium sulphate fractionation,ion-exchange chromatography on DEAE–cellulose and gelfiltration on Sephadex G-75. The enzyme activity wasinhibited strongly by Hg2+, Zn2+ and Cu2+ but was activatedby Na+. An extracellular β-amylase was purified by ad-sorption on raw corn starch to 22.5-fold from a new isolateof B. polymyxa [241]. The degradation of raw starch by theenzyme was greatly stimulated by pullulanase addition. Athermostable β-amylase was purified with ammonium sul-phate, DEAE–cellulose column chromatography and gelfiltration using Sephadex G-200 [92]. Maltose was identifiedas the major hydrolysis product of starch and it had β-anomeric form. Another thermotolerant β-amylase waspurified from a strain of Bacillus circulans S31 that was notactivated by any metal ions [89]. Ray and Chakraverty [242]partially purified a β-amylase from Syncephalastrum race-mosum. The enzyme activity remained unaffected in the
presence of metal ions and was not increased in the presenceof exogenous thiols, indicating the absence of thiols at theactive site of the enzyme.
GAPandey presented a comprehensive review on the puri-fication and properties of GA [94]. Table 4 cites someexamples. It is evident that GAs are generally active on theacidic side of neutrality, with optimum pH values of around4.5–5.0. The optimum temperature for GA activity frommost sources has generally been found to be between 40 and60 °C. However, there are a few reports describing theextreme nature of GA. GA I and GA II from a thermophilicfungus Humicola lanuginosa showed pH optima of 4.9 and 6.6,respectively [243]. GA II was stable up to pH 11.0. The GAsfrom Coniophora cerebella [244] and Corticum rolfsii [245]were stable up to pH 9.0 whereas that of Mucor rouxianusshowed pH stability up to 8.0 [246].
A GA preparation from solid cultures of A. niger waspurified 32.4-fold with a final specific activity of 49.25units}mg of protein, which contained four different forms(GA-I, GA-I«, GA-II and GA-II«), each with different charac-
# 2000 Portland Press Ltd

146 A. Pandey and others
teristics. This was the first report on four forms of GAproduced by A. niger in SSF [102]. The enzymes wereglycoproteins with carbohydrate contents of 16%. Enzymeactivity was inhibited strongly by Hg2+ whereas Mn2+ and Fe2+
were stimulatory. Hata et al. [205] compared the propertiesof two GAs produced in SmF and SSF by A. oryzae. Enzymeproduced in SSF could digest raw starch but that in SmFcould not. GAs obtained in SmF and SSF exhibited differentcharacteristics. Medda et al. [96] studied the raw-starchadsorption and elution behaviour of GA I of black Aspergillussp. The enzyme was completely adsorbed on to raw starchat pH 3.4, which was the isoelectric point of the enzyme. GAof A. awamori var. kawachi had three forms, GA I, GA I«and GA II, with molecular masses of 90000, 83000 and57000 Da, respectively [256]. GA I and GA I« were optimallyactive at pH 3.8, whereas for GA II the optimum was atpH 4.0. The pH-stability ranges were 2–10, 2.5–6.5 and 4–7,respectively, for isoforms. The isoelectric points were 3.55,3.45 and 3.28, and, at 60 °C, GA I was stable, while GA I« andGA II were unstable. GA I could hydrolyse raw starchwhereas the other two could not. The N-terminal aminoacids were Ala for GA I, Ala+Ile for GA I« and Ile for GA II.
A commercial preparation of GA from A. niger consistedof at least six species, five forms of which were characterizedfully [247]. They were apparently different in molecularmass. The optimum pH varied between 3.5 and 5.0, andwhen enzymological characteristics were compared, one ofthe species was peculiar and quite different from others.Lineback et al. [248] reported two forms of GA from A. niger.The molecular mass of GA I was 99000 Da and that of GAII was 112000 Da. Both forms of GA contained covalentlylinked carbohydrate (containing D-mannose, D-glucose andD-galactose residues) and were therefore glycoenzymes.Both showed identical amino acid composition. However,the two GAs differed in carbohydrate content, with GA IIcontaining as much as double the number of carbohydrateunits per molecule as GA I. The carbohydrate–proteinlinkage in the GA was essentially glycosidic to the hydroxylgroup of L-serine and L-threonine residues. There are severalother reports that have described the characterization oftwo forms of GA from A. niger (e.g. [94]).
Ueda and Kano [249] purified the GA system ofRhizopus sp. By fractionation, two kinds of GA wereobtained ; one with strong debranching activity and the otherwith weak debranching activity. Moriyam et al. [250] studiedthe influence of dielectric constants and ligand binding on thethermostability of GA from Rhizopus niveus. Investigationswere carried out (mainly at 60 °C and pH 4.5) in relation tokinetics, in both the presence and absence of various ligands.With substrate analogues, such as glucose, lactose andgluconolactones, and with glycerol, the thermostability ofthe enzyme greatly increased, whereas with dioxane, ethanoland glycol it decreased. From the model equations, binding
constants of the ligands were estimated in order to confirmthe validity of the assumptions. GA of R. niveus was alsostudied by Saha and Ueda [251] for raw-starch adsorption,elution and digestion behaviour. The preparation consistedof five forms, which were separated by preparative iso-electric focusing. All of them were strong in debranchingactivity, raw-starch adsorption and digestion. Among them,two were major, GA C and GA D, having isoelectric pointsof 8.45 and 9.1, and the carbohydrate contents were 14.9and 12.7%, respectively.
Two forms of GA were isolated and purified from aculture of Monascus kaofiang nov. sp. F-I, which werehomogeneous in nature. GA I and GA II exhibited pH optimaof 4.5 and 4.7, respectively. The molecular mass of GA I was48000 Da and that of GA II was 68000 Da [252]. Takeda etal. [253] studied the purification and substrate specificity ofGA of Paecifomyces varioti AHU 9417. The enzyme washomogeneous ; its molecular mass was estimated to be69000 Da and the pH optimum was 4.5. The GA fromPiricularia oryzae, the pathogenic fungus of rice blast disease,was optimally active at pH 6.5 and at temperatures of50–60 °C. Its molecular mass was 94000 Da [246]. The GAof Thermomyces lanuginosus was established to be hom-ogenous [254]. This was a glycoprotein with an averagemolecular mass of about 57000 Da and a carbohydratecontent of 10–12%. Shimazaki et al. [255] purified anextracellular GA from Schizophyllum commune by ammoniumsulphate precipitation, acid-clay treatment and Sephadex G-100 gel filtration, which gave a single band.
A bacterial GA was purified and characterized from theanaerobic thermophilic bacterium Clostridium thermo-saccharolyticum by Specka et al. [119]. The enzyme, whichconsisted of a single subunit, had an apparent molecular massof 75000 Da. The purified enzyme attacked α-1,4 as well asα-1,6 glycosidic linkages in various α-glucans. The enzymeshowed a pH optimum of 5.0 and a temperature optimum of70 °C, and attacked polysaccharides preferentially. Table 3lists some properties of GAs from some other sources[256–270].
Conclusions
Amylases are among the most important enzymes used forindustrial purposes, and now in the light of biotechnologythey are considered useful for biopharmaceutical appli-cations. They are useful tools in medicinal and clinicalchemistry. Although amylases can be derived from a numberof sources, microbial sources, especially for α-amylase andGA, are significant for commercial production. Since starchis the only natural substrate to be hydrolysed by amylases, itis desirable to isolate efficient microbial strains producinghigh amylase titres active on raw starch. Starch-hydrolysis
# 2000 Portland Press Ltd

Microbial amylases 147
processes may be improved by the use of new thermostableGA that is mutually compatible with α-amylase, which wouldlead to the whole process being performed in a single step,providing economic benefits. Microbial strains with efficientdual activity, i.e. liquefication and saccharification, e.g.amylolytic yeasts, need to be developed. Microbial strainswith efficient β-amylase activity should be developed as theenzyme could be used to produce maltose syrup. Utilizationof low-value agro-industrial residues as substrates should befocused upon for enzyme production, as this would reducethe cost of production and help to solve their disposal andpollution problems. Since thermostability is considered auseful and important feature of amylases for industrialapplication, attempts should be made to develop enzymesfrom thermophilic and extremely thermophilic micro-organisms. Finally, it is hoped that amylases will continue toprovide new opportunities in biotechnology as biocatalystsand that new applications will emerge in the bio-pharmaceutical sector.
References
1 Crueger, W. and Crueger, A. (eds.) (1989) Industrial Micro-
biology, pp. 189–218, Sinauer Associates, Sunderland, MA
2 Fogarty, W. M. and Kelly, C. T. (1980) in Economic Micro-
biology, Microbial Enzymes and Bioconversions, vol. 5 (Rose,
A. H., ed.), pp. 115–170, Academic Press, London
3 Witczak, Z. J. (1999) Curr. Med. Chem. 6, 165–178
4 Sutton, A., Dawson, H., Hoff, B., Grift, E. and Shoukri, M. (1999)
Can. Vet. J. 40, 255–260
5 Lepp, C. A., Gilligan, S., Tobin, J. J., Gunzer, G., Conway, P.,
Sugrue, M., Oleary, D., Ilchmann, D. and Mahon, G. (1996) Clin.
Chem. 42, SS, 23
6 Chiu, L. Y. and Chandler, W. L. (1995) Clin. Chem. 41,
S238–S239
7 Becks, S., Bielawaski, C., Henton, D., Padala, R., Burrows, K. and
Slaby, R. (1995) Clin. Chem. 41, S186
8 Giri, N. Y., Mohan, A. R., Rao, L. V. and Rao, C. P. (1990)
Curr. Sci. 59, 1339–1340
9 Menzel, C., Lerch, T., Schneider, K., Weidemann, R., Tollnick, C.,
Kretzmer, G., Scheper, T. and Schugerl, K. (1998) Process
Biochem. 33, 175–180
10 Ch, G. J., Moon, I. S. and Lee, J. S. (1997) Chem. Lett. 6,
577–578
11 Kolb, M., Zentgraf, B., Mattisson, B., Arvidson, P. and Danielsson,
B. (1996) Termochim. Acta 277, 1–6
12 Chelly, M., Dahan, R., Labyod, M., Dupont, F., Lacoste, C. and
Larue, F. (1996) Ann. Pediatr. 43, 392–402
13 Strandberg, A., Nystrom, A., Behr, S. and Karlsson, A. (1999)
Chromatography 50, 215–222
14 Lebenthal, E., Khinmaungu., Rolston, D. D. K., Khinmyattun,
T. T., Tinnuswe, Theintheinmyint, Jirapinyo, P., Visitsuntron,
N., Firmsansyah, A., Sunoto, S. et al. (1995) Pediatrics 95,
198–202
15 De-Almeida, S. E. M., Mizuta, K. and Giglio, R. (1997) Mycol.
Res. 101, 188–190
16 Schwermann, B., Pfau, K., Liliensiek, B., Schleyer, M., Fischer, T.
and Bakker, E. P. (1994) Eur. J. Biochem. 226, 981–991
17 Feller, G., Narinx, E., Arpigny, J. L., Zekhini, Z., Swings, J. and
Gerday, C. (1994) Appl. Microbiol. Biotechnol. 41, 477–479
18 Sugita, H., Kuruma, A. and Deguchi, Y. (1997) Biosci. Biotechnol.
Biochem. 61, 1757–1759
19 Melasniemi, H. (1988) Biochem. J. 250, 813–818
20 Abe, J., Nakajima, K., Nagano, H., Hizukuri, S. and Obata, K.
(1988) Carbohydr. Res. 175, 85–92
21 Bhella, R. S. and Tosaar, I. (1985) Can. J. Microbiol. 31, 149–153
22 Siedenberg, D., Gerlach, S. R., Czwalinna, A. and Schugerl, K.
(1997) J. Biotechnol. 56, 205–216
23 Ali, S. and Abdelmoneim, A. A. (1989) Zentral Mikrobiol. 144,
615–621
24 Goto, C. E. (1998) FEMS Microbiol. Lett. 167, 139–142
25 Ohnishi, M., Iwata, K., Tomita, T., Nishikawa, U. and Hiromi, K.
(1990) Starch/Sta4 rke 42, 486–489
26 Iqbal, M. and Zafar, S. I. (1994) Biotechnol. Tech. 10, 755–758
27 Hoshino, K., Katagiri, M., Taniguchi, M., Sasakura, T. and Fujii, M.
(1994) J. Ferment. Bioeng. 77, 407–412
28 Murado, M. A., Gonzalez, M. P., Torrado, A. and Pastrana, L. M.
(1997) Process Biochem. 32, 35–42
29 Torrado, A., Gonzalez, P., Pintado, J., Montemayor, I., Miron, J.
and Murado, M. (1994) Meded Fac Landbouwwet Rijksuniv
Gent 59, 4b, 2471–2473
30 Suganuma, T., Tahara, N., Kitahara, K., Nagahama, T. and
Inuzuka, K. (1996) Biosci. Biotechnol. Biochem. 60, 177–179
31 Kikuchi, S., Shibayama, J., Yasui, M., Takeuchi, T., Yamamoto, I.
and Sato, M. (1993) Biosci. Biotechnol. Biochem. 57, 845–846
32 Outtrup, H. and Norman, B. E. (1984) Starch/Sta4 rke 36,
247–251
33 Mamo, G. and Gessesse, A. (1997) Biotechnol. Tech. 11,
447–450
34 Boyer, E. W., Ingle, M. B. and Mercer, G. D. (1979) Starch/
Sta4 rke 31, 166–170
35 Milner, J. A., Martin, J. D. and Smith, A. (1997) Enz. Microb.
Technol. 21, 382–386
36 Syu, M. J. and Chen, Y. H. (1997) Chem. Eng. J. 65, 237–247
37 Hillier, P., Wase, D. A. J., Emery, A. N. and Solomon, G. L.
(1997) Process Biochem. 32, 51–59
38 Lin, C. S., Hong, S. C., Cheng, C. Y. and Chen, C. L. (1997)
J. Chem. Technol. Biotechnol. 69, 433–437
39 Tonkova, A., Ivanova, V., Dobreva, E., Stefanova, M. and
Spassova, D. (1994) Appl. Microbiol. Biotechnol. 41, 517–522
40 Stefanova, M., Tonkova, A., Dobreva, E. and Spassova, D.
(1998) Folia Microbiol. 43, 42–44
41 Ariga, O., Tpyofuku, H., Minegishi, I., Hattori, T., Ano, Y. and
Nagura, M. (1997) J. Ferment. Bioeng. 84, 553–557
# 2000 Portland Press Ltd

148 A. Pandey and others
42 Ivanova, V. and Dobreva, E. (1994) Process Biochem. 29,
607–612
43 Babu, K. R. and Satyanarayana, T. (1995) Process Biochem. 30,
305–309
44 Kelly, C. T., Bolton, D. J. and Fogarty, W. M. (1997) Biotechnol.
Lett. 19, 675–677
45 Bandopadhyay, A., Sen, S. K. and Pal, S. C. (1994) Acta
Biotechnol. 14, 97–104
46 Dobreva, E., Tonkova, A., Ivanova, V., Stefanova, M. and
Kabaivanova, L. (1998) J. Ind. Microbiol. Biotechnol. 20,
166–170
47 Decordt, S., Vanhoof, K., Hu, J., Maesmans, G., Hendrickx, M.
and Tobback, P. (1992) Biotechnol. Bioeng. 40, 396–402
48 Varlan, A. R., Sansen, W., VanLoey, A. and Hendrickx, M.
(1996) Biosens. Bioelectr. 11, 443–448
49 Ramesh, M. V. and Lonsane, B. K. (1989) Process Biochem. 24,
176–178
50 Brumm, P. J., Hebeda, R. E. and Teague, W. M. (1991) Starch/
Sta4 rke 43, 319–323
51 Omidiji, O., Amund, O. O., Braimoh, A. A. and Ilori, M. O.
(1997) Microbios 90, 155–162
52 Krishna, C. and Chandrasekaran, M. (1996) Appl. Microbiol.
Biotechnol. 46, 106–111
53 Nishimura, T., Kometani, T., Takii, H., Terada, Y. and Okada, S.
(1994) J. Ferment. Bioeng. 78, 31–36
54 El Helow, E. R. and El Gazaerly, M. A. (1996) J. Basic Microbiol.
36, 75–81
55 Ratanakhanokchai, K., Kaneko, J., Kamio, Y. and Izaki, K. (1992)
Appl. Environ. Microbiol. 58, 2490–2494
56 Paquet, V., Croux, C., Goma, G. and Soucaille, P. (1991) Appl.
Environ. Microbiol. 57, 212–218
57 Tanaka, T., Ishimoto, E., Shimomura, Y., Taniguchi, M. and Oi, S.
(1987) Agric. Biol. Chem. 51, 399–405
58 Swamy, M. V. and Seenayya, G. (1996) Process Biochem. 31,
157–162
59 Delahaye, E. P., Foglietti, M. J., Andrieux, C., Chardonloriaux, I.,
Szylit, O. and Raibaud, P. (1991) Comp. Biochem. Physiol. A.
Physiol. 98, 351–354
60 Tsiomenko, A. B., Musaev, D. S., Lupashin, V. V. and Kulaev, I. S.
(1992) Biochemistry (Moscow) 57, 287–303
61 Patel, S., Bagai, R. and Madamwar, D. (1996) Biocat. Biotrans.
14, 147–155
62 Bagai, R. and Madamwar, D. (1997) Appl. Biochem. Biotechnol.
62, 213–218
63 Swamy, M. V. and Seenayya, G. (1996) Indian J. Microbiol. 36,
181–184
64 Ilori, M. O., Amund, O. O. and Omidiji, O. (1995) World J.
Microbiol. Biotechnol. 11, 595–596
65 Sen, S. and Chakraborty, S. L. (1987) Enz. Microb. Technol. 9,
112–116
66 Fergus, C. L. (1969) Mycologia 61, 1171–1177
67 Ilori, M. O., Amund, O. O. and Omidiji, O. (1997) Folia
Microbiol. 42, 445–449
68 Adeleye, A. I. (1990) J. Basic Microbiol. 30, 717–722
69 Loginova, L. G., Tsaplina, I. A., Guzhova, E. P., Boltyanskaya,
E. V., Kovolevskaya, I. D. and Seregina, L. M. (1970) Mikrobio-
logiya 39, 784–789
70 Sadhukhan, R., Roy, S. K. and Chakraborty, S. L. (1993) Enz.
Microb. Technol. 15, 801–804
71 Farezvidal, M. E., Fernandezvivas, A., Gonzalez, F. and Arias, J. M.
(1995) J. Appl. Bacteriol. 78, 14–19
72 Stevens, A., Gunasekaran, M. and Sangan, P. (1994) J. Gen. Appl.
Microbiol. 40, 319–329
73 Haska, R. and Ohta, Y. (1994) Starch/Sta4 rke 46, 480–485
74 Kimura, T., Ogata, M., Yoshida, M. and Nakakui, T. (1989)
Biotechnol. Bioeng. 33, 845–855
75 Siqueira, E. M. D., Mizuta, K. and Giglio, J. R. (1997) Mycol. Res.
101, 188–190
76 Koch, R., Spreinat, A., Lemke, K. and Antranikian, G. (1991)
Arch. Microbiol. 155, 572–578
77 de Souza, E. L., Hoffmann, E. H. E., Castilho, V. M., de Lima,
V. A. and Bellini, M. Z. (1996) Arq. Biol. Technol. 39, 831–839
78 Ray, R. R., Jana, D. C. and Nanda, G. (1995) J. Appl. Bacteriol.
79, 157–162
79 Shimazaki, T., Hara, S. and Sato, M. (1984) J. Ferment. Technol.
62, 165–170
80 Shaw, J. F., Lin, F. P., Chen, S. C. and Chen, H. C. (1995)
Bot. Bull. Acad. Sinica 36, 195–200
81 Abramov, Z. T., Tsaplina, I. A., Stepanov, V. M. and Loginova,
L. G. (1986) Biochemistry (Moscow) 51, 100–106
82 Chung, Y. C., Kobayashi, T., Kanai, H., Akiba, T. and Kudo, T.
(1995) Appl. Environ. Microbiol. 61, 1502–1506
83 Muramatsu, Y., Takahashi, K. and Nakamura, N. (1993)
Starch/Sta4 rke 45, 99–104
84 Snutzenberger, F. and Carnell, R. (1977) Appl. Environ.
Microbiol. 34, 231–235
85 Allam, A. M., Hussein, A. M. and Ragab, A. M. (1975) Z. Allg.
Mikrobiol. 15, 393–399
86 Jensen, B. and Olsen, J. (1992) Enz. Microb. Technol. 14,
112–116
87 Liebl, W., Stemplinger, I. and Ruile, P. (1997) J. Bacteriol. 179,
941–948
88 Fogarty, W. M. and Kelly, C. T. (1990) in Microbial Enzymes and
Biotechnology, 2nd edn. (Fogarty, W. M. and Kelly, C. T., eds.),
pp. 71–132, Elsevier Science Publishers, London
89 Kwan, H. S., So, K. H., Chan, K. Y. and Cheng, S. C. (1994)
World J. Microbiol. Biotechnol. 10, 597–598
90 Ray, R. R., Jana, S. C. and Nanda, G. (1997) Indian J. Exp. Biol. 35,
285–288
91 Hyun, H. H. and Zeikus, J. G. (1985) Appl. Environ. Microbiol.
49, 1162–1167
92 Reddy, P. R. M., Swamy, M. V. and Seenayya, G. (1998) World
J. Microbiol. Biotechnol. 14, 89–94
93 Swamy, M. V., Sai Ram, M. and Seenayya, G. (1994) Lett. Appl.
Microbiol. 18, 301–304
94 Pandey, A. (1995) Starch/Sta4 rke 42, 439–445
# 2000 Portland Press Ltd

Microbial amylases 149
95 Bui, D. M., Kunze, I., Forster, S., Wartmann, T. and Horstmann,
C. (1996) Appl. Microbiol. Biotechnol. 44, 610–619
96 Medda, S., Saha, B. C. and Ueda, S. (1982) J. Ferment. Technol.
60, 261–263
97 Lucia, C. M., Facciotti, M. C. R., Kilkian, B. V. and Schmidell, W.
(1992) Biotechnol. Lett. 14, 465–468
98 Ariff, A. B. and Webb, C. (1996) Asia-Pacific J. Mol. Biol. 4,
183–195
99 Silva, J. G., Nascimento, H. J., Soares, V. F. and Bon, E. P. S.
(1997) Appl. Biochem. Biotechnol. 63–65, 87–96
100 Queiroz, M. C. R., Faxxiotti, M. C. R. and Schmidell, W. (1997)
Biotechnol. Lett. 19, 167–170
101 Selvakumar, P., Ashakumary, L. and Pandey, A. (1998) Biores.
Technol. 65, 83–85
102 Selvakumar, P., Ashakumary, L., Helen, A. and Pandey, A.
(1996) Lett. Appl. Microbiol. 23, 403–406
103 Pandey, A., Selvakumar, P. and Ashakumary, L. (1996) Process
Biochem. 31, 43–46
104 Pandey, A., Ashakumary, L. and Selvakumar, P. (1995) Biores.
Technol. 51, 217–220
105 Pandey, A., Ashakumary, L., Selvakumar, P. and Vijaylakshmi,
K. S. (1994) World J. Microbiol. Biotechnol. 10, 485–486
106 Pandey, A., Selvakumar, P. and Ashakumary, L. (1994) World
J. Microbiol. Biotechnol. 10, 348–349
107 Pandey, A. and Radhakrishnan, S. (1993) Process Biochem. 28,
305–309
108 Pandey, A. and Radhakrishnan, S. (1992) Enz. Microb. Technol.
14, 486–488
109 Singh, D., Dahiya, J. S. and Nigam, P. (1995) J. Basic Microbiol.
35, 117–121
110 Pandey, A. (1992) in Industrial Biotechnology (Malik, V. S. and
Sridhar, P., eds.), pp. 525–537, IBH and Oxford Publishing Co.,
New Delhi
111 Pandey, A. (1992) Starch/Sta4 rke 39, 75–77
112 Kobayashi, T., Nagamure, T. and Endo, I. (1990) Ann. NY
Acad. Sci. 613, 846
113 Imai, Y., Suzuki, M., Masamoto, M., Nagayasu, K. and Kishimoto,
M. (1994) J. Ferment. Bioeng. 78, 310–314
114 Kneck, C. (1991) Appl. Microbiol. Biotechnol. 35, 466–468
115 Ghosh, A. B., Chaterjee, B. and Das, A. (1991) Indian J.
Technol. 29, 529–533
116 Ali, S., Hossain, Z., Mahmood, S. and Alam, R. (1990) World.
J. Mirobiol. Biotechnol. 6, 19–21
117 Wijbenga, D. J., Beldman, G., Veen, A. and Binnema, D. J.
(1991) Appl. Microbiol. Biotechnol. 35, 180–184
118 Hyun, H. H. and Zeikus, J. G. (1985) Appl. Environ. Microbiol.
49, 1168–1173
119 Specka, U., Meyer, F. and Antranikian, G. (1991) Appl. Environ.
Microbiol. 57, 2317
120 Krause, D. R., Wood, C. J. and Maclean, D. J. (1991) J. Gen.
Microbiol. 137, 2463–2466
121 Gonzalez, C., Delgado, O., Baigori, M., Abate, C., De Fagueroa,
L. I. C. and Callieri, D. A. (1998) Acta Biotechnol. 18, 149–155
122 Reddy, O. V. S. and Basappa, S. C. (1996) Biotechnol. Lett. 18,
1315–1318
123 Ohno, N., Juln, T., Song, S., Uchiyama, S., Shinoyoma, H., Ando,
A. and Fujii, T. (1992) Biosci. Biochem. Biotechnol. 56,
465–469
124 Ilori, M. O., Amund, O. O. and Omidjii, O. (1996) Folia
Microbiol. 41, 339–340
125 Fogarty, M. M. and Dixon, M. M. (1979) Prog. Ind. Microbiol.
15, 118
126 Soccol, C. R., Marin, B., Raimbault, M. and Lebeault, J. M.
(1994) Appl. Microbiol. Biotechnol. 41, 330–336
127 Soccol, C. R., Iloki, I., Marin, B. and Raimbault, M. (1994)
J. Food Sci. Technol. 31, 320–323
128 Chiarello, M. D., Soccol, C. R., Stertz, S. C., Furlanetto, L.,
Fontana, J. D. and Krieger, N. (1997) Arq. Biol. Technol. 40,
771–785
129 Furuta, H., Arai, T., Hama, H., Shiomi, N., Kondo, A. and
Fukuda, H. (1997) J. Ferment. Bioeng. 84, 169–171
130 Hulseweh, B., Dahlems, U. M., Dohmen, J., Strasser, A. W. M.
and Hollenberg, C. P. (1997) Eur. J. Biochem. 244, 128–133
131 Joutsjoki, V. V., Parkkinen, E. E. M. and Torkkeli, T. K. (1993)
Biotechnol. Lett. 15, 277–282
132 Kim, I. C., Cha, J. H., Kim, J. R., Jang, S. Y., Seo, B. C., Cheong,
T. K. and Lee, D. S. (1992) J. Biol. Chem. 267, 22108–22114
133 Iefuji, H., Chino, M., Kato, M. and Iimura, Y. (1996) Biochem. J.
318, 989–996
134 Marco, J. L., Bataus, L. A., Valencia, F. F., Ulhora, C. J., Astolfi, S.
and Felix, C. R. (1996) Appl. Microbiol. Biotechnol. 44,
746–752
135 Matsuura, Y., Kusunoki, M., Harada, W. and Kakaso, M. (1984)
J. Biochem. (Tokyo) 95, 697–702
136 Tada, S., Iamira, Y., Gomi, K., Takahashi, K., Hara, S. and
Yoshizawa, S. (1989) Agric. Biol. Chem. 53, 593–599
137 Kim, T. G. and Kim, K. (1996) Appl. Biochem. Biotechnol. 59,
39–51
138 Birol, G., Onsan, Z. I., Kirdar, B. and Oliver, S. G. (1998) Enz.
Microb. Technol. 22, 672–677
139 Shiba, S., Nishida, Y., Park, Y. S., Iijima, S. and Kobayashi, T.
(1994) Biotechnol. Bioeng. 44, 1055–1063
140 Steyn, A. J. C. and Pretorius, I. S. (1991) Gene 100, 85–93
141 Miranda, E. A. and Berglund, K. A. (1993) Biotechnol. Progress
9, 411–420
142 Stripecke, R., Rosato, Y. B. and Astolfifilho, S. (1990) Rev.
Brasil Genetica 13, 661–668
143 Juge, N., Svensson, B. and Williamson, G. (1998) Appl.
Microbiol. Biotechnol. 48, 385–392
144 Kitamoto, N., Yamagata, H., Kato, T., Tsukagoshi, N. and
Udaka, S. (1988) J. Bacteriol. 170, 5848–5854
145 Siggens, K. W. (1987) Mol. Microbiol. 1, 86–91
146 Nitta, Y., Shirakawa, M. and Takasaki, Y. (1996) Biosci.
Biotechnol. Biochem. 60, 823–827
147 Nammori, T., Nagai, M., Shimizu, Y., Shinke, R. and Mikami, B.
(1993) Appl. Environ. Microbiol. 59, 623–627
# 2000 Portland Press Ltd

150 A. Pandey and others
148 Uozumi, N., Sakurai, K., Sasaki, T., Takekawa, S., Yamagata, H.,
Tsukagoshi, N. and Udaka, S. (1989) J. Bacteriol. 171, 375–382
149 Yoshigi, N., Okada, Y., Sahara, H. and Koshino, S. (1994) Biosci.
Biotechnol. Biochem. 58, 1080–1086
150 Yoshigi, N., Okada, Y., Maeba, H., Sahara, H. and Tamaki, T.
(1995) J. Biochem. 118, 562–567
151 James, J. A. and Lee, B. H. (1997) J. Food Biochem. 21, 1–52
152 Zhang, Z., Scharer, J. M. and Mooyoung, M. (1997) Bioprocess
Eng. 17, 235–240
153 Erratt, JA and Nasim, A., US Patent 4870014 (1989)
154 Nakamura, Y., Kobayashi, F., Ohnaga, M. and Sawada, T.
(1997) Biotechnol. Bioeng. 53, 21–25
155 Kim, K., Bajszar, G., Lee, S. Y., Knudsen, F. and Mattoon, J. R.
(1994) Appl. Biochem. Biotechnol. 44, 161–185
156 Chen, M. M., Bakir, U., Reilly, P. J. and Ford, C. (1994)
Biotechnol. Bioeng. 43, 101–105
157 Suominen, I., Ford, C., Stavhon, D., Heimo, H., Niederauer, M.,
Nurmela, H. and Glatz, C. (1993) Enz. Microb. Technol. 15,
593–600
158 James, J. A., Robert, N. and Lee, B. H. (1996) Biotechnol. Lett.
18, 1407–1412
159 Svensson, B. and Sogaard, M. (1993) J. Biotechnol. 29, 1–37
160 Linardi, V. R., Carvalho, C. M. M. C. and Dias, A. A. M. (1993)
World J. Microbiol. Biotechnol. 9, 601–602
161 Suntornsuk, W. and Hang, Y. D. (1994) Lett. Appl. Microbiol.
19, 249–252
162 Witkowskagwiazdowska, A., Ilnickaolejniczak, O., Kostka, G.
and Czajkowska, D. (1991) Acta Biotechnol. 11, 235–239
163 Flory, N., Gorman, M., Coutinho, P. M., Ford, C. and Reilly,
P. J. (1994) Protein Eng. 7, 1005–1012
164 Ariga, O., Toyofuku, H., Matsudaria, K., Sano, Y. and Kuroiwa,
I. (1995) J. Ferment. Bioeng. 80, 616–618
165 Ariga, O., Andoh, Y., Fujishita, Y., Watari, T. and Sano, Y.
(1991) J. Ferment. Bioeng. 71, 397–402
166 Walls, E. L. and Gainer, J. L. (1991) Biotechnol. Bioeng. 37,
1029–1036
167 Bigelis, R. (1989) in Biotechnology, vol. 7b (Jacobson, G. K. and
Jolly, S. O., eds.), pp. 229–260, VCH, Weinheim
168 Ashikari, T., Kunisaki, S., Matsumoto, N., Amachi, T. and
Yoshizumi, H. (1989) Appl. Microbiol. Biotechnol. 32,
129–133
169 Innis, MA. (1985) Science 228, 21–26
170 Hata, Y., Kitamoto, K., Gomi, K., Kumagai, C., Tamura, G. and
Hara, S. (1991) Agric. Biol. Chem. 55, 941–949
171 Ventura, L., Gonzalez-Candelas, L., Perez-Gonzalez, A. and
Ramon, D. (1995) Appl. Environ. Microbiol. 61, 399–402
172 Yamashita, I. and Fukui, S. (1984) Agric. Biol. Chem. 48,
137–142
173 Hostinova, E., Balanova, J. and Zelinka, J. (1990) Biologia 45,
301–306
174 Ohnishi, H., Kitamura, H., Sakai, H. and Ohta, T. (1992) Eur. J.
Biochem. 207, 413–418
175 Pandey, A. (1992) Process Biochem. 27, 109–117
176 Pandey, A. (1991) Process Biochem. 26, 355–361
177 Pandey, A. and Radhakrishnan, S. (1990) Proc. Kerala Acad.
Sci. 1, 69–79
178 Pandey, A. and Soccol, C. R. (1998) Brazilian Arch. Biol.
Technol. 41, 379–390
179 Pandey, A., Selvakumar, P., Soccol, C. R. and Nigam, P. (1999)
Curr. Sci. 77, 149–162
180 Selvakumar, P., Ashakumary, L. and Pandey, A. (1996) J. Sci.
Ind. Res. 55, 443–449
181 Ramesh, M. V. and Lonsane, B. K. (1987) Biotechnol. Lett. 9,
501–504
182 Bajpai, P., Gera, R. K. and Bajpai, P. K. (1992) Enz. Microb.
Technol. 14, 670–683
183 Salva, T. J. G. and Moraes, I. O. (1995) Rev. Microbiol. 26,
46–51
184 Lonsane, B. K. and Ramesh, M. V. (1990) Adv. Appl. Biochem.
35, 1–56
185 Gron, S., Morcel, C., Emborg, C. and Biedermann, K. (1995)
Bioprocess Eng. 14, 23–41
186 Morcel, C. and Biedermann, K. (1994) FEMS Microbiol. Rev.
14, 57–61
187 Tari, C., Parulekar, S. J., Stark, B. C. and Webster, D. A. (1998)
Biotechnol. Bioeng. 59, 673–678
188 Arnesen, S., Eriksen, S. H., Olsen, J. and Jensen, B. (1998) Enz.
Microb. Technol. 23, 249–252
189 Sudo, S., Ishikawa, T., Sato, T. and Oba, T. (1994) J. Ferment.
Bioeng. 77, 483–489
190 Ivanova, V., Stefanova, M., Tonkova, A., Dobreva, E. and
Spassova, D. (1995) Appl. Biochem. Biotechnol. 50, 305–316
191 Pandey, A. (1991) Biores. Technol. 37, 169–172
192 Pandey, A. (1990) Biol. Wastes 34, 11–19
193 Ashakumary, L., Selvakumar, P. and Pandey, A. (1994) in Solid
State Fermentation (Pandey, A., ed.), pp. 33–37, Wiley Eastern
Publishers, New Delhi
194 Mukherjee, P. S., Pandey, A., Selvakumar, P., Ashakumary, L.
and Gurusamy, P. (1998) J. Sci. Ind. Res. 57, 583–586
195 Pandey, A (1994) in Solid State Fermentation (Pandey, A.,
ed.), p. 188, Wiley Eastern Publishers, New Delhi
196 Kilkian, B. V. (1996) Rev. Microbiol. 27, 10–12
197 Wang, R. H. and Webb, C. (1995) Biotechnol. Tech. 9, 55–58
198 Elegado, F. B. and Fujio, Y. (1993) J. Gen. Appl. Microbiol. 39,
541–546
199 Rajagopalan, S., Rockstraw, D. A. and Munson-McGee, S. H.
(1997) Biores. Technol. 61, 175–183
200 Shimada, A., Koda, T. and Nakamura, I. (1998) J. Ferment.
Bioeng. 85, 542–545
201 Linardi, V. R., Andrade, C. M. M. C., Figueira, M. M., Andrad,
M. C. and Souza, A. A. V. (1993) Folia Microbiol. 38, 281–284
202 Reddy, O. V. S. and Basappa, S. C. (1993) Starch/Sta4 rke 45,
187–194
203 Ramadas, M., Holst, O. and Mattiasson, B. (1996) World J.
Microbiol. Biotechnol. 12, 267–271
204 Fujio, Y. and Morita, H. (1996) J. Ferment. Bioeng. 82, 554–557
# 2000 Portland Press Ltd

Microbial amylases 151
205 Hata, Y., Ishida, H., Kojima, Y., Ichikawa, E. K. A., Suginami, K.
and Imayasu, S. (1997) J. Ferment. Bioeng. 84, 532–537
206 Uguru, G. C., Robb, D., Akinyanju, J. A. and Sani, A. (1997)
J. Ind. Microbiol. Biotechnol. 19, 273–279
207 Bolton, D. J., Kelly, C. T. and Fogarty, W. M. (1997) Enz.
Microb. Technol. 20, 340–343
208 Ivanova, V. N., Dobreva, E. P. and Emanuilova, E. I. (1993)
J. Biotechnol. 28, 277–289
209 Katoh, S., Terashima, M. and Miyaoku, K. (1997) Appl.
Microbiol. Biotechnol. 47, 521–524
210 McTigue, M. A., Kelly, C. T., Fogarty, W. M. and Doyle, E. M.
(1994) Biotechnol. Lett. 16, 569–574
211 Kim, T. U., Gu, B. G., Jeong, J. Y., Byun, S. M. and Shin, Y. C.
(1995) Appl. Environ. Microbiol. 61, 3105–3112
212 Lin, L. L., Chyau, C. C. and Hsu, W. H. (1998) Biotechnol.
Appl. Biochem. 28, 61–68
213 Dobransky, T., Polivka, L., Haas, I., Sova, O. and Petrvasky, E.
(1987) J. Chromatogr. 411, 486–489
214 Satoh, K., Uchimura, T., Kudo, T. and Komagata, K. (1997)
Appl. Environ. Microbiol. 63, 4941–4944
215 Freer, S. N. (1993) Appl. Environ. Microbiol. 59, 1398–1402
216 Lee, S. K., Kim, Y. B. and Ji, G. E. (1997) J. Appl. Microbiol. 83,
267
217 Egas, M. C. V., da Costa, M. S., Cowan, D. A. and Pires,
E. M. V. (1998) Exothermophiles 2, 23–32
218 Laderman, K. A., Davis, B. R., Krutzsch, H. C., Lewis, M. S.,
Griko, Y. V., Privalov, P. L. and Anfinsen, C. B. (1993) J. Biol.
Chem. 268, 24394–24401
219 AbouZeid, A. M. (1997) Microbios 89, 55–66
220 Park, H. N., Yoo, J. C. and Yang, Y. K. (1995) J. Microbiol. 33,
80–84
221 Chang, C. T., Tang, M. S. and Lin, C. F. (1995) Biochem. Mol.
Biol. Int. 36, 185–193
222 Planchot, V. and Colonna, P. (1995) Carbohydr. Res. 272,
97–109
223 Khoo, S. L., Amirul, A. A., Kamaruzaman, M., Nazalan, N. and
Azizan, M. A. (1994) Folia Microbiol. 39, 392–398
224 Somkutti, G. A. and Steinberg, D. H. (1980) Devel. Ind.
Microbiol. 21, 327–337
225 Bunni, L., McHale, L. and McHale, A. P. (1989) Enz. Microb.
Technol. 11, 370–375
226 Sills, A. M., Saunders, M. E. and Stewart, G. G. (1983) Devel.
Ind. Microbiol. 24, 295–303
227 Bhella, R. and Altosaar, I. (1985) Can. J. Microbiol. 31, 149–153
228 Tsiomenko, A. B., Musaev, D. S., Lupashin, V. V. and Kulaev,
I. S. (1992) Biochemistry (Moscow) 57, 297–303
229 Prieto, J. A., Bort, B. R., Martinez, J., Randezgil, F., Buesa, C. and
Sanz, P. (1995) Biochem. Cell. Biol. 73, 41–49
230 Feller, G., Lohnienne, T., Deroanne, C., Libioulle, C.,
Vanbeeumen, J. and Gerday, C. (1992) J. Biol. Chem. 267,
5217–5221
231 Zhang, X. Z., Xie, S. Y., Wu, X. X., Jin, F. X. and Li, X. Z. (1994)
Enz. Microb. Technol. 16, 985–990
232 Elaassar, S. A., Omar, S. H., Gouda, M. K., Ismail, A. M. and
Abdelfattah, A. F. (1992) Appl. Microbiol. Biotechnol. 38,
312–314
233 Shih, N. J. and Labbe, R. G. (1995) Appl. Environ. Microbiol.
61, 1776–1779
234 Jeang, C. L., Lee, Y. H. and Chang, L. W. (1995) Biochem. Mol.
Biol. Int. 35, 549–557
235 Burgesscassler, A. and Imam, S. (1991) Curr. Microbiol. 23,
207–213
236 Giraud, E., Gosselin, L., Marin, B., Parada, J. L. and Raimbault, M.
(1993) J. Appl. Bacteriol. 75, 276–282
237 Juhasz, O., Lukacova, V., Vollek, V. and Skarka, B. (1991)
Biologia 46, 627–634
238 Odibo, F. J. C., Okafor, N. and Okafor, B. U. (1992) J. Gen.
Appl. Microbiol. 38, 1–11
239 Hesse, O., Hansen, G., Hohne, W. E. and Korner, D. (1991)
Biomed. Biochim. Acta 50, 225–232
240 Odibo, F. J. C., Okafor, N., Tom, M. U. and Oyeka, C. A.,
Starch/Sta4 rke (1992) 44, 192–195
241 Sohn, C. B., Lee, S. M., Kim, M. H., Ko, J. H., Kim, J. S., Chang,
J. E., Ahn, Y. K. and Kim, C. H. (1996) J. Food Sci. 61, 230–234
242 Ray, R. R. and Chakraverty, R. (1998) Mycol. Res. 102,
1563–1567
243 Taylor, P. M., Napier, E. J. and Fleming, L. D. (1978) Carbohydr.
Res. 61, 301–304
244 King, J. (1967) Biochem. J. 105, 577
245 Kaji, A., Sato, M., Koyabashi, M. and Murao, T. (1976) J. Agric.
Chem. Soc. Jpn. 50, 509–511
246 Yamasaki, Y., Suzuki, Y. and Ozawa, J. (1977) Agric. Biol.
Chem. 41, 765–771
247 Ono, K., Shintani, K. and Shikata, S. (1988) Agric. Biol. Chem.
52, 1699
248 Lineback, D. R., Russel, L. J. and Rasmnseen, C. (1969) Arch.
Biochem. Biophys. 134, 529–531
249 Ueda, S. and Kano, S. (1975) Starch/Starke 27, 123–126
250 Moriyam, S., Matsuno, R. and Kamikubo, T. (1977) Agric. Biol.
Chem. 41, 1985–1987
251 Saha, B. C. and Ueda, S. (1983) J. Ferment. Technol. 61, 67
252 Iizuka, H. and Minek, S. (1977) J. Gen. Appl. Microbiol. 23,
217–221
253 Takeda, Y., Matsui, H., Tanida, M., Tani, S. and Chiba, S. (1985)
Agric. Biol. Chem. 49, 1633
254 Rao, V. B., Sastry, N. V. S. and Rao, P. V. S. (1981) Biochem. J.
193, 379–382
255 Shimazaki, T., Hara, S. and Sato, M. (1984) J. Ferment. Technol.
62, 165–168
256 Yoshino, E. and Hayashida, S. (1978) J. Ferment. Technol. 56,
289–294
257 Amirul, A. A., Khoo, S. L., Nazalan, M. N., Razip, M. S. and
Azizan, M. N. (1996) Folia Microbiol. 41, 165–174
258 Venkataraman, K., Manjunath, P. and Rao, M. R. R. (1975)
Indian J. Biochem. Biophys. 12, 107–110
259 Miah, M. N. and Ueda, S. (1977) Starch/Sta4 rke 29, 235–239
# 2000 Portland Press Ltd

152 A. Pandey and others
260 Lineback, D. R. and Bumann, W. E. (1970) Carbohydr. Res. 14,
341–343
261 Takhashi, T., Inokuchi, N. and Irie, M. (1981) J. Biochem.
(Tokyo) 89, 125–129
262 Ghosh, A., Chatterjee, B. and Das, A. (1991) J. Appl. Bacteriol.
71, 162–164
263 Krzechowsk, M. and Urbanek, H. (1975) Appl. Microbiol. 30,
163–167
264 Ohnishi, H., Sakai, H. and Ohta, T. (1991) Agric. Biol. Chem.
55, 1901–1903
265 Fukui, T. and Nikuni, Z. (1969) Agric. Biol. Chem. 33, 884–885
266 Gracheve, I. M., Lushchik, T. A., Tyrsin, Y. A. and Pinchukova,
E. E. (1977) Biokhimiya 42, 1803–1805
267 Evertova, H. (1966) Folia Microbiol. 11, 422–426
268 Yukhi, A., Watanabe, T. and Matsuda, K. (1977) Starch/Sta4 rke
29, 265–269
269 Takahasi, T., Kato, K., Ikegami, Y. and Irie, M. (1985) J. Biochem.
(Tokyo) 98, 663–665
270 Pazur, J. and Okada, S. (1967) Carbohydr. Res. 4, 371–376
Received 23 July 1999}7 November 1999; accepted 21 January 2000
# 2000 Portland Press Ltd





























