Embed Size (px)
Citation preview

古生物学报,48(3):364-374(2009年9月)
ActaPalaeontologicaSinica,48(3):364-374(September,2009)
收稿日期:20090630国家自然科学基金(40572070)、中国科学院、国家外国专家局创新团队国际伙伴合作项目和现代古生物学与地层学国家重点实验室课
题(20081107)联合资助。
谱系年代研究进展
杨 群1) 丛培允1
,2) 孙晓燕1
) 马俊业1
) 盖永华1
) 李春香1
) 郝家胜1
,3) 夏旭华4
)
1)现代古生物学与地层学国家重点实验室,中国科学院南京地质古生物研究所,南京210008,qunyang@nigpas.a.cn;
2)云南大学云南省古生物研究重点实验室,昆明650091;
3)安徽师范大学生命科学学院,安徽芜湖241000;
4)渥太华大学生物系,安大略K1N6N5,加拿大
提要 谱系年代学是结合化石记录和分子钟方法计算“生命树”(TreeofLife)上各分歧点时间的一个新兴交
叉学术领域。由于化石记录的不完整性,各类生物的起源年代和支系分化时间的确定可借助于部分化石记录和通
过计算现生生物类群之间的遗传距离转换得出的相对分歧时间相结合的办法进行讨论。化石记录可代表部分生
物类群起源时间的保守估计值,而分子钟方法可为那些不易保存为化石的生物类群与其姊妹群的分化时间提供依
据。如果使用得当的话,两者可进行相互验证。在分子谱系年代分析中,正确使用化石校准方案是获得准确分歧
时间的关键,这需要:1)正确确定化石物种在谱系树上的位置;2)正确解释化石记录所代表的时间含义(最小值、确
定值、最大值及其标准差)。分子钟估算分歧时间的技术在不断改进,目前常用的分子分歧时间估算法(宽松分子
钟法,如贝叶斯法、补偿性似然法等)包容分子演化速率在谱系间和随时间的变化。随着谱系年代研究的不断深
入,长期困扰人们的化石记录时间与分子钟计算结果悬殊的问题正在逐渐趋于和谐并得到正确诠释。文中还讨论
了有关动物起源与早期分化时间以及早期陆生节肢动物的谱系年代学研究进展。我们强调,化石记录和分子钟分
析可以优势互补,两者的整合无疑将提高生物演化历史时间格架的准确度和精度,以利更好地将生命演化事件置
于地球系统科学及地球环境演化史之中。
关键词 谱系年代学 分子古生物学 宽松分子钟 生命之时间树 早期动物登陆
1 前 言
“生命树”(TreeofLife)是当今全球生命科学
领域研究者致力于建立的一个重大科学计划,它旨
在回答有关地球生物各类群之间的亲缘关系问题
(TreeofLifeWebProject,1997)。而如何确定这
棵庞大的生命树上各支系的年代问题则是演化生物
学关注的又一热点科学问题,可称之为谱系年代学
(Yang犲狋犪犾.,2003,2007),其直接任务是采用古生
物学和分子系统学相结合的方法为“生命树”内部节
点定年 (BentonandAyala,2003;HedgesandKu
mar,2009)。准确测定谱系发生时间对于探索生物
演化史的意义是显而易见的,这不仅关系到大家熟
知的演化速率和式样(tempoandmodeofevolu
tion)问题,而且关系到生物谱系演化与地球环境变
化的协同机理的讨论。例如,寒武纪大爆发是否与
大气氧含量、气候或海洋化学组分的变化关联?哺
乳动物的早期分化是否和板块构造区系的演化相
关?早期后生动物的登陆与辐射演化是否有原始陆
地生态系统的形成作为先导?这些科学问题必须置
于一个确定的时间框架之中加以讨论。
我们长期依赖于化石记录作为讨论生物演化时
间的唯一证据。然而,分子钟的发现(Zuckerkandl
andPauling,1962;Runnegar,1982)宣告了生物演
化年代研究的一种新方法的诞生,即通过计算现生
生物类群之间的分子(DNA或蛋白质序列)距离来
推算相关生物类群的分化时间。Runnegar(1986)
在首次阐述分子古生物学时,将存在于现生生物细
胞内的基因分子称为一类特殊的“分子化石”。早在

20世纪80年代初,Runnegar(1982)便大胆地应用
分子钟方法推测动物门类的起源时间。
采用分子钟测定谱系分化时间的主要争议在于
分子演化速率的不恒定性,即同源的基因分子在不
同生物类群中的演化速率可变、在同一演化谱系中
可能随时间发生变化,基因分子序列内部不同位点
的演化速率也不恒定。针对这些问题,分子生物信
息学领域展开了大规模探索,并建立了一系列统计
模型以适应速率不恒定的分子数据的分析,发展到
目前普遍采用的“宽松分子钟”概念(“relaxedmo
lecularclock”concept)(Thorne犲狋犪犾.,1998)。例
如,研究者采用相对速率检验法(relativeratetest,
Tajima,1993)来鉴别谱系树的不同支系中具有明
显速率差异的分支;采用局域分子钟法(localclock
method)(YoderandYang,2000;Kumar,2005)将
谱系树的不同支系分为一系列速率组;采用贝叶斯
方法为可变速率拟合最佳统计模型(Kishino犲狋犪犾.,
2001;Aris-BrosouandYang,2002);采用补偿性
似然法(penalizedlikelihood)进行速率修匀(rate
smoothing)(Sanderson,1997,2002)。应该说,分
子系统学进展为探索早期生物演化时间表奠定了良
好的基础,后者在当今生物演化领域已成为一个前
沿研究方向。基因测年分析也可作为检验某些古生
物学 假 说 的 方 法。例 如,Waggoner 和 Collins
(2004)的谱系年代分析显示,曾被研究者归入水螅
动物纲Chondrophorina亚目的埃迪卡拉动物犗狏犪
狋狅狊犮狌狋狌犿,犆犺狅狀犱狉狅狆犾狅狀, 犓犪犻狊犪犾犻犪, 犓狌犾犾犻狀犵犻犪,
犈狅狆狅狉狆犻狋犪等实际上不可能属于该现代水螅动物分
类单元;原因是,根据谱系年代分析结果推测,如果
Chondrophorina亚目出现于新元古代或古生代,那
么腔肠动物的起源时间就相应地被前推至16—18
亿年前,与事实不符。
化石记录无疑是探索生命演化历史的唯一直接
证据,它在谱系年代研究中发挥基础作用。但是,由
于化石记录的不完整性,许多生物类群缺乏化石记
录,更多生物类群可能缺失起源初期的记录。因此,
分子钟推测谱系时间的方法便成为生物演化年代研
究中的一个重要补充。化石记录和其它地质事件一
方面构成生物演化史的基本(但是可能极为粗略的)
时间框架,另一方面,前者为分子钟测定谱系发育年
代提供校准点(标尺),即充当将分子距离转换为年
代的参照系。
本文旨在回顾近年来谱系发育年代学领域取得
的重要进展,重点介绍研究方法及概念、早期动物起
源及陆生无脊椎动物登陆研究现状。谱系发育年代
研究正在向不同生物门类渗透,其宏伟目标是构建
完整的生命年代树(theTimetreeofLife)。
2 分子钟与宽松分子钟
Zuckerkandl和Pauling(1962)观察到,在不
同生物的血红蛋白分子中,氨基酸位点差异的数目
大致与时间呈正比,这个时间就相当于化石记录的
时间,从而提出了物种间遗传距离随时间推移而增
大的分 子 钟 的 概 念 (ZuckerkandlandPauling,
1965)。Margoliash(1963)比较分析了细胞色素c
的数据,表明马和猪只有3个氨基酸差别,马和金枪
鱼差19个氨基酸,而马和酵母的差别是44个氨基
酸。显然,亲缘关系越近的物种之间遗传距离小,而
亲缘关系疏远的物种间的遗传距离大。这些经验数
据为分子钟理论奠定了基础(插图1)。分子钟也称
“基因钟”或“演化钟”。
但是,前面已经提到,更多研究表明分子钟不是
匀速的 (WelchandBromham,2005)。为此,研究
者已建立一系列针对可变速率进行测年的宽松分子
钟方法(表Ⅰ)。
最简单的方法是对谱系树进行相对速率检验
(Tajima,1993),剔除演化速率过快或过慢的分支,
将剩余支系构成的谱系树作为恒速处理(Hedges
andKumar,2003)。这个方法的主要局限性是:1)
相对速率检验方法的有效性不高,通常只能有效排
除演化速率差异极大的类群,而差异较大的类群不
易排除。如果提高速率检验门槛的话,则易于导致
接下来的问题,即2)由于演化速率不恒定是普遍
的,速率检验后剩余类群所构成的谱系树经常不足
以分析特定科学问题。
局域分子钟方法(localmolecularclockmeth
od)将谱系树分成若干个不同速率的支系(速率群),
然后分别对于各个支系运用恒速分子钟进行谱系年
代分析。如何划分速率群的方法有若干种,比如表
Ⅰ中的 Qdate(四元法)将4个物种作为一组分析,
然后分成两个速率群;Yoder和Yang(2000)根据相
对速率检验或枝长估计将所研究的谱系树分成速率
群。划分若干速率群的缺陷是其人为性(如何确定
划分界线和速率群数目);此外,研究者无法用同一
组数据估计所用划分方案是否最优。
最复杂的宽松分子钟方法是允许速率无限制变
化。如果局域分子钟方法可称作“手动变速”技术的
563 第3期 杨 群等:谱系年代研究进展
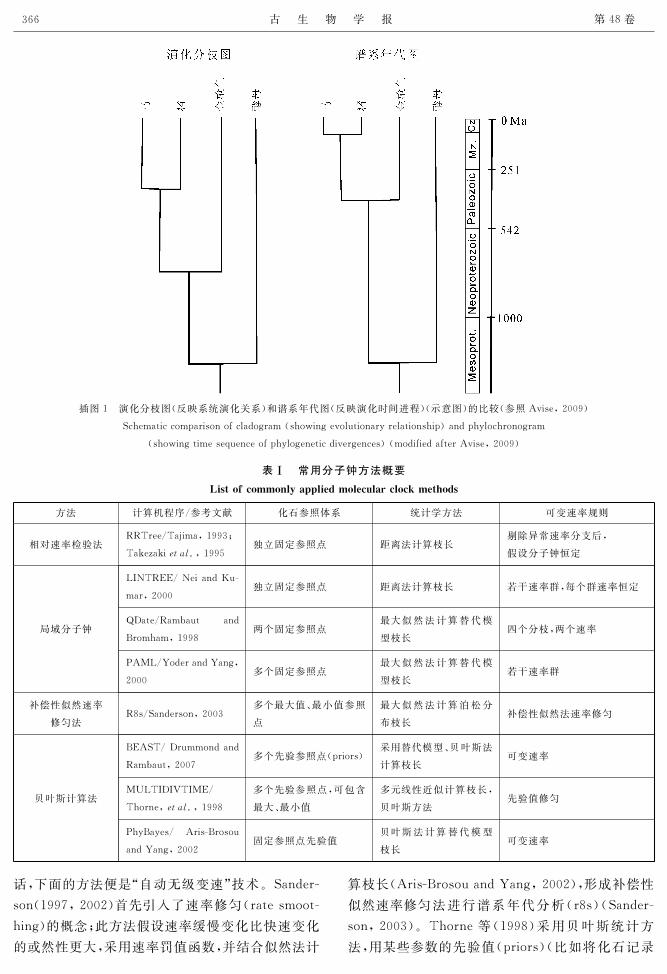
插图1 演化分枝图(反映系统演化关系)和谱系年代图(反映演化时间进程)(示意图)的比较(参照Avise,2009)
Schematiccomparisonofcladogram(showingevolutionaryrelationship)andphylochronogram
(showingtimesequenceofphylogeneticdivergences)(modifiedafterAvise,2009)
表Ⅰ 常用分子钟方法概要
犔犻狊狋狅犳犮狅犿犿狅狀犾狔犪狆狆犾犻犲犱犿狅犾犲犮狌犾犪狉犮犾狅犮犽犿犲狋犺狅犱狊
方法 计算机程序/参考文献 化石参照体系 统计学方法 可变速率规则
相对速率检验法RRTree/Tajima,1993;
Takezaki犲狋犪犾.,1995独立固定参照点 距离法计算枝长
剔除异常速率分支后,
假设分子钟恒定
局域分子钟
LINTREE/NeiandKu
mar,2000独立固定参照点 距离法计算枝长 若干速率群,每个群速率恒定
QDate/Rambaut and
Bromham,1998两个固定参照点
最大似然法计算替代模
型枝长四个分枝,两个速率
PAML/YoderandYang,
2000多个固定参照点
最大似然法计算替代模
型枝长若干速率群
补偿性似然速率
修匀法R8s/Sanderson,2003
多个最大值、最小值参照
点
最大似然法计算泊松分
布枝长补偿性似然法速率修匀
贝叶斯计算法
BEAST/Drummondand
Rambaut,2007多个先验参照点(priors)
采用替代模型、贝叶斯法
计算枝长可变速率
MULTIDIVTIME/
Thorne,犲狋犪犾.,1998
多个先验参照点,可包含
最大、最小值
多元线性近似计算枝长,
贝叶斯方法先验值修匀
PhyBayes/ ArisBrosou
andYang,2002固定参照点先验值
贝叶斯法计算替代模型
枝长可变速率
话,下面的方法便是“自动无级变速”技术。Sander
son(1997,2002)首先引入了速率修匀(ratesmoot
hing)的概念;此方法假设速率缓慢变化比快速变化
的或然性更大,采用速率罚值函数,并结合似然法计
算枝长(ArisBrosouandYang,2002),形成补偿性
似然速率修匀法进行谱系年代分析(r8s)(Sander
son,2003)。Thorne等(1998)采用贝叶斯统计方
法,用某些参数的先验值(priors)(比如将化石记录
663 古 生 物 学 报 第48卷
作为某些支系分歧点的估算值)来建立这些参数的
概率分布,并采用速率修匀法,通过计算相关参数的
后验概率分布来估计谱系分歧时间。
一般认为,宽松分子钟方法优于恒定分子钟方
法。但是,由于各种方法处理数据的具体算法不同
(比如计算枝长法、校准方法以及处理速率变化的统
计分析技术),其结果很难判定方法上的优劣
(WelchandBromham,2005)。许多谱系年代学分
析结果与数据集构成和化石校准方法有关。
3 化石记录作为分子钟的时间尺度和
限定系统
正确使用化石记录是谱系年代研究中最为关键
的一个环节。事实上,化石参照点的选择和处理方
法可能对分子钟运算结果产生极大影响(Hugand
Roger,2007)。在分子钟分析中,至少需要一个化
石(或导致生物区系形成的地质事件,例如陆地裂
解、地峡形成等)作为时间参照点(校准点,calibra
tionpoint),才能将遗传距离转换为时间,也就是为
谱系演化提供一个时间标尺,可简单表示为(Nei
andKumar,2000):
狉=犱2犜
上式中,狉=演化速率;犱=遗传距离;犜=分歧时间
(通常就是化石参照点)。这样,通过参照体系中的
遗传距离和化石时间得到演化速率狉,然后将这个
速率推广到其它分歧点,便可以获得谱系树上其它
未知分歧点的时间犜,即
犜=犱2狉
应用化石参照点的首要任务是将化石类群和具
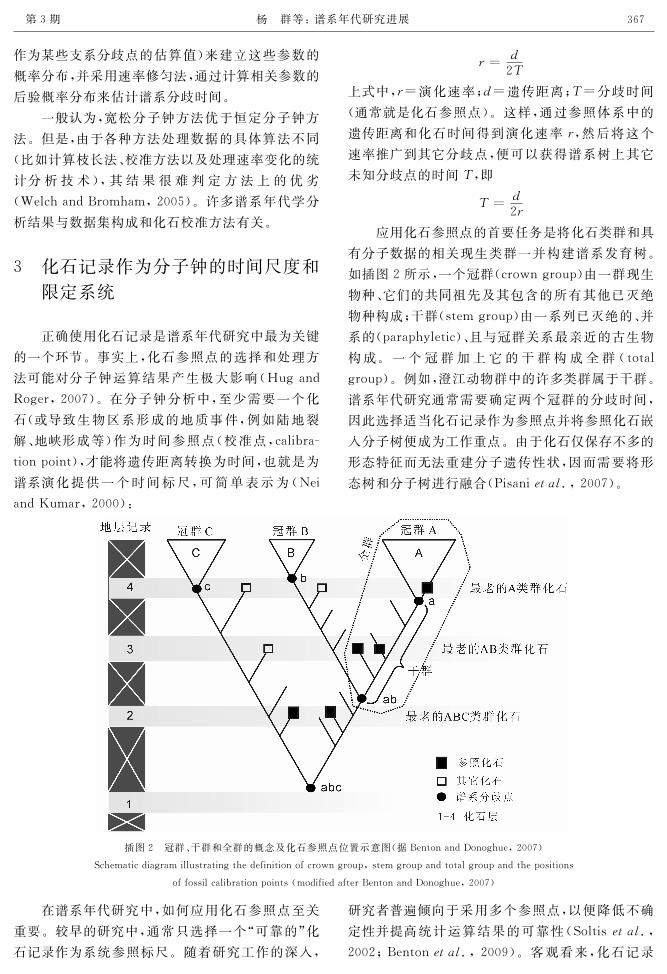
有分子数据的相关现生类群一并构建谱系发育树。
如插图2所示,一个冠群(crowngroup)由一群现生
物种、它们的共同祖先及其包含的所有其他已灭绝
物种构成;干群(stemgroup)由一系列已灭绝的、并
系的(paraphyletic)、且与冠群关系最亲近的古生物
构成。一个冠群加上它的干群构成全群(total
group)。例如,澄江动物群中的许多类群属于干群。
谱系年代研究通常需要确定两个冠群的分歧时间,
因此选择适当化石记录作为参照点并将参照化石嵌
入分子树便成为工作重点。由于化石仅保存不多的
形态特征而无法重建分子遗传性状,因而需要将形
态树和分子树进行融合(Pisani犲狋犪犾.,2007)。
插图2 冠群、干群和全群的概念及化石参照点位置示意图(据BentonandDonoghue,2007)
Schematicdiagramillustratingthedefinitionofcrowngroup,stemgroupandtotalgroupandthepositions
offossilcalibrationpoints(modifiedafterBentonandDonoghue,2007)
在谱系年代研究中,如何应用化石参照点至关
重要。较早的研究中,通常只选择一个“可靠的”化
石记录作为系统参照标尺。随着研究工作的深入,
研究者普遍倾向于采用多个参照点,以便降低不确
定性并提高统计运算结果的可靠性(Soltis犲狋犪犾.,
2002;Benton犲狋犪犾.,2009)。客观看来,化石记录
763 第3期 杨 群等:谱系年代研究进展

仅仅代表有关生物起源时间的最小估计值,而两个
类群的实际分歧时间总是早于它们的最早化石记
录。在实际应用中,研究者将化石记录作为时间尺
度的方式可概括为3种:1)作为确定的谱系分化时
间点(HedgesandKumar2004;PetersonandBut
terfield,2005);2)作为最小分歧时间(下限)(Glaz
ko犲狋犪犾.,2005;Near犲狋犪犾.,2005);3)作为最大分
歧时间(上限)(Kishino犲狋犪犾.,2001;Bentonand
Donoghue,2007)。研究者还引入了“刚性控制点”
(hardcalibration)和“柔性控制点”(softcalibra
tion)的概念(HedgesandKumar,2004;Yangand
Rannada,2006);所谓“刚性点”指不可逾越的界线
(最早化石记录时间常常被用作刚性下限),而“柔性
点”是可以超出的界线(化石参照点的估计上限常被
作为柔性控制点),其超出概率可以用概率密度模型
表示 (Benton犲狋犪犾.,2009)。例 如,Benton 和
Donoghue(2007)采用刚性下限(最早化石记录)和
柔性上限(采用较老姊妹群的最早化石记录,nea
restrelative)来限定一个参照时间点。
运用多个化石参照点作为谱系年代系统的时间
标尺也存在一些问题,比如这样可能混合有用时间
信息和错误时间信息。因此,不同化石参照点的结
果应当进行交叉验证(NearandSanderson,2004;
Near犲狋犪犾.,2005),检验结果明显不一致的须剔
除。明显偏小的化石参照点可能由于观察到的化石
记录太少(距起源时间太远),而明显偏老的参照点
则可能由于系统鉴定错误。只有通过交叉检验后相
互匹配的化石参照点才能构成一个多点时间校准组
(HugandRoger,2007)。
4 研究实例
近年来有关谱系年代学方面的工作在不断深
化,真核生物的起源 (HugandRoger,2006)、后生
动物的起源与早期分化、被子植物起源(Magallon
andSanderson,2005)、哺乳动物(vanRheede犲狋
犪犾.,2006)和鸟类起源(PereiraandBaker,2006)以
及人类起源时间(Kumar犲狋犪犾.,2005)等课题已成
为该领域的热点。国际上已诞生两部这个领域的专
著:《TellingtheEvolutionaryTime》(Donoghue
andSmith,2004)和 《The TimetreeofLife》
(HedgesandKumar,2009)。前者较系统地叙述了
理论和方法上的进展,而后者分述了从原核生物到
真核生物一些生物门类的谱系年代学研究现状,具
有一定的参考价值。由于篇幅限制,本文仅就后生
动物起源和早期分化以及无脊椎动物登陆时间问题
的研究现状进行讨论。
4.1 后生动物起源与早期分歧时间
关于后生动物起源和早期分化年代问题是当前
谱系年代学领域最为热门也是争议最多的研究课题
(BlairandHedges,2005;HugandRoger,2007;
Peterson犲狋犪犾.,2008)。Yang等 (2007)概述了早
期后生动物谱系年代分析结果(插图3)。结合近期
研究新进展,有关研究成果可简要归纳为如下几点:
1)分子谱系年代研究表明,后生动物的共同祖
先与其近邻———领鞭毛虫类的分歧时间大约为
849—863Ma(Douzery犲狋犪犾.,2004;Berneyand
Pawlowski,2006)或更早 (ca.1020 Ma,Bhat
takarya犲狋犪犾.,2009);也就是说,真核生物从单细
胞向多细胞动物演化这一重要事件很可能根植于新
元古代成冰纪(Cryogenian,635—850Ma)之前的拉
伸纪(Tonian,850—1000Ma),该时期正值地球上大
气氧含量逐渐升高的阶段 ((Anbarand Knoll
2002))。尽管多细胞动物的形态化石证据到目前为
止仅发现于新元古代埃迪卡拉纪(542—635Ma),最
新分子古生物研究成果显示,海绵动物的分子化石出
现于埃迪卡拉纪之前的成冰纪(Love犲狋犪犾.,2009)。
关于后生动物起源时间的谱系发育年代学成果对于
进一步探索地质生物具有一定的指导意义。
2)最原始的后生动物—多孔动物门的实体化石
出现于新元古代晚期埃迪卡拉纪地层中(Gehling
andRigby,1996;Li犲狋犪犾.,1998;ca.580Ma)、分
子化石出现在新元古代成冰期(>635Ma,Love犲狋
犪犾.,2009)。分子年代分析显示,多孔动物与其它
后生动物的分歧时间大约为764—839Ma(Berney
andPawlowski,2006,SupplementaryTable2);多
孔动物冠部类群(crowngroups)的分歧时间约为
733M Ma(BerneyandPawlowski,2006,Supple
mentaryTable3)。综合分子化石、实体化石和基因
测年数据推测,海绵动物起源于成冰期或者更早,但
是由于沉积环境或早期海绵生物的数量或特性的问
题而难以保存为化石。显然,在新元古代成冰纪之
后,海绵动物和其他后生动物经历了演化辐射事件,
而这次动物辐射演化之前或前期存在一次显著的生
物和环境事件(ca.630—640Ma):大量大型疑源类
在华南被发现(Xiao,2004)伴随明显的碳同位素负
异常及大冰期的结束(插图3)。
863 古 生 物 学 报 第48卷
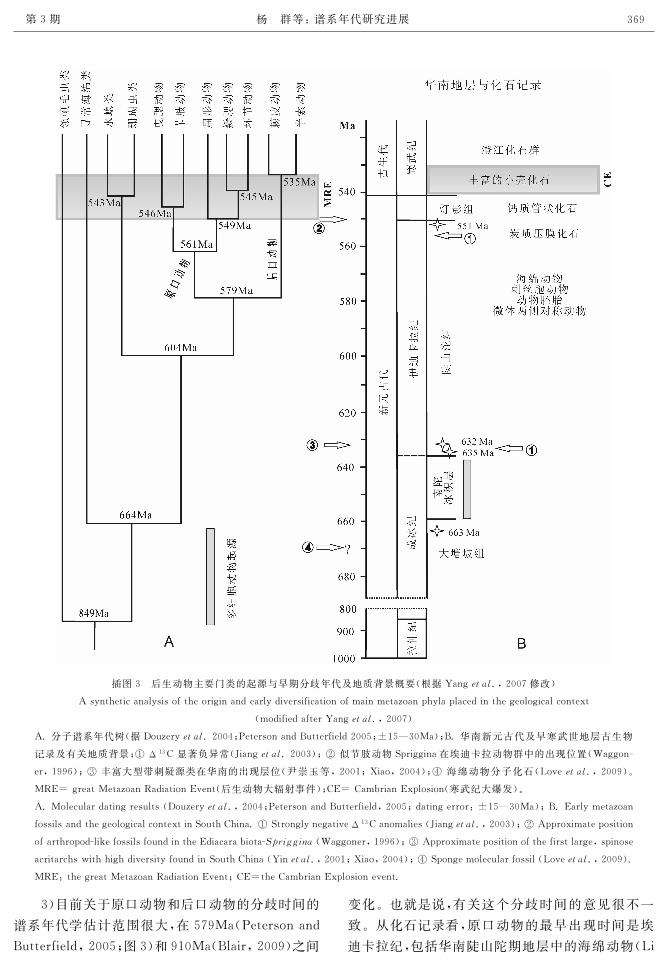
插图3 后生动物主要门类的起源与早期分歧年代及地质背景概要(根据Yang犲狋犪犾.,2007修改)
Asyntheticanalysisoftheoriginandearlydiversificationofmainmetazoanphylaplacedinthegeologicalcontext
(modifiedafterYang犲狋犪犾.,2007)
A.分子谱系年代树(据Douzery犲狋犪犾.2004;PetersonandButterfield2005;±15—30Ma);B.华南新元古代及早寒武世地层古生物
记录及有关地质背景;① Δ13C显著负异常(Jiang犲狋犪犾.2003);② 似节肢动物Spriggina在埃迪卡拉动物群中的出现位置(Waggon
er,1996);③ 丰富大型带刺疑源类在华南的出现层位(尹崇玉等,2001;Xiao,2004);④ 海绵动物分子化石(Love犲狋犪犾.,2009)。
MRE=greatMetazoanRadiationEvent(后生动物大辐射事件);CE=CambrianExplosion(寒武纪大爆发)。
A.Moleculardatingresults(Douzery犲狋犪犾.,2004;PetersonandButterfield,2005;datingerror:±15—30Ma);B.Earlymetazoan
fossilsandthegeologicalcontextinSouthChina.①StronglynegativeΔ13Canomalies(Jiang犲狋犪犾.,2003);② Approximateposition
ofarthropodlikefossilsfoundintheEdiacarabiota犛狆狉犻犵犵犻狀犪(Waggoner,1996);③ Approximatepositionofthefirstlarge,spinose
acritarchswithhighdiversityfoundinSouthChina(Yin犲狋犪犾.,2001;Xiao,2004);④Spongemolecularfossil(Love犲狋犪犾.,2009).
MRE:thegreatMetazoanRadiationEvent;CE=theCambrianExplosionevent.
3)目前关于原口动物和后口动物的分歧时间的
谱系年代学估计范围很大,在579Ma(Petersonand
Butterfield,2005;图3)和910Ma(Blair,2009)之间
变化。也就是说,有关这个分歧时间的意见很不一
致。从化石记录看,原口动物的最早出现时间是埃
迪卡拉纪,包括华南陡山陀期地层中的海绵动物(Li
963 第3期 杨 群等:谱系年代研究进展

书书书
犲狋犪犾.,1998)、!"#$
(Xiao犲狋犪犾.,2000)、%&
'()*#$
(Chen犲狋犪犾.,2004)+,-./01
23#$
(Spriggina,Waggoner,1996)。4#$5
6789
,:;<=#$>?@ABCDE
,FGH
#$IJ>?@ABCKLM
;NOP&@ABC
DQRSTUVWXY
(>850Ma)Z[\]^_`
abcde
。)@fg56hiQjklXm
,n
oDpXQqrstQRabuvHwxyz
,{
|}D~�����
:��L��D6����T
;
�������QRab
;+,��c�56��
���D|��
。
4):��3A(� ¡
)¢£
,¤b¥w¦§D
QRUVWX
(PetersonandButterfield,2005),G
H#$¨©ªD>?kl«¬549—535Ma®
l
,̄*°
“#$x±²hi
”(MRE)(Yang犲狋犪犾.,
2007),V³´©µ
、©µ#$ª¶DQ6
;\6�·
¸D¹ºCx»¼
(530—540Ma;��3-B)k
l�]½¾
,¿ÀgQÁÂ
,GÃV³wĵQªÅ
DƲ56
。Ç�ÈÉ
:① FGH#$©ªDQj
kl
(MRE)Ê&@wĵQªÅDƲQ6kl
(CE);②¹ºCx»¼®oDFGH#$aËw
Ì
,ÍÎ6�Qª�Ï@¨#$©DÐÅ
(stem
groups);③ FGH#$56ÑÒ¤Ó@ÔÕÖV
,
×FGH#$¨©ªDQ6ÎaIJ¼HÔÕÖ
VABC®G
,Ø-./0C
。¹ºCx»¼V³
wĵQªÅDxË>?
,IJ\ÙÚHÛÜÝD
Þßàá
。
1SunXiaoyan(âãä
),2004.AMolecularPalaeobiologicalAnalysisonOriginandEvolutoinofConchostracaandrelatedGroups.Master’sthesis,
GraduateSchooloftheChineseAcademyofSciences.1—172(inChinesewithextendedEnglishsummary)
2CongPeiyun(åæç
),2008.Phylogenyandphylochronologyofmajordiplopodgroups.Dissertation,GraduateSchooloftheChineseAcademyof
Sciences.1—167(inChinesewithextendedEnglishsummary)
4.2 !"#$%&'()!*+
èéê#$ë<ìíîïðDhiñH${6
Ò�|òÁóôõö
。ë@&÷ïH#$6�,ï
àðø·¸à)ùÌ
,èéê#$úïklDûü
ýþÿ�6�·¸tQRSTUV`!b
,°"#
&÷ïðHÛTpD$%,&\ðÚÜÝ®lD'
�56()_`!b
。
ÖH$~�³*
,+,%&D-.Ó$ð/6
�I01263ÎUoD-./0C
(Yuan犲狋
犪犾.,2005),×ñ2no°4
,%&IJÏ@ïH#
$D6�·¸56@78x¹ºC9:C®;
(470—500Ma)Dðø¬
(MacNaughton犲狋犪犾.,
2002),ئ<=A>?¬D23#$@A6�
,
IJV³%&DïH#$BCK#$DE<FÜÝ
D
“EGÃ
”;HI9:CM÷DJ<àKLMN$
¬O Î P ª 2 3 # $
(P A
)(Johnson犲狋犪犾.,
1994)。QR
,6�·¸Z£
,ïH#$IJ9:
C]S56
,�T4<H23#$UB56NV
。
��4³£4STUVQqW2D&÷23#
$¨XóªÅQj56DklYZ
,&¬a,bQ¶
V³'ª&÷23#$
—ÎP#$t[P\P#
$úïhi
,ÇP&÷ïH
(]^_`H
,a:
,[P
ª
)23#$IJñÿò#$bíîïðHÝD&
÷V³
。QR
,ûücdD>?klefüg&÷
ïðhiHÛTp$%Dkljk
。��l5
,#
$ë<ìmïðHÝD56hi+,GVëïðn
o<ì
(a:
,pª
、<qr
;Proche,2008)D56
hiH$56Ò�sÎt¼H
。ÇôXóuv&
֕H23#$D>?klwx
,�+XóHy
z{ÜÝDÎP#$
(myriapods)t_`HD[P
ª
(branchiopods)°|í}
。
Pisanir
(2004)~°
,ÎP#$¬D�P�
(Chilopoda)\�P�
(Diplopoda)DQjkl
(442
±50Ma)V³&÷23#$D
(%M
)úïkl
,I
J°9:CÀ��C÷l
。Ç|�}�v\6�·
¸�c��
:%&DïH�Pª6�56M9:
�
(Johnson犲狋犪犾.,1994),N%&D�Pª6�5
6M���
(Jeram犲狋犪犾.,1990)。�dD%Ô~
�³*
:Hy@_`ÜÝD[P�
(Branchiopoda)
23#$D>?klx�¹ºC¬M÷
(�gª
ÅDQjkl
:508±15Ma)(âãä
,2004)1。ÎP
#$©�P����ï��\���ï��DQj
klIJ�¹ºC
(512.2±26.2Ma)(åæç
,
2008)2。Robison(1990)~°
,¹ºC¬÷D�3�
<ì23#$�IJñ���3�DÎP#$DU
BªÅ
;N��¬¹º�¼6DV³ïHÓ$D
��R6�
(Strother犲狋犪犾.,2004)�*
,ïðÜÝ
(�T<ì23#$D567�
)]S°&÷èéê
#$úïfüg��
。
��6�·¸tQRSTUVQqab
,I+
��
,&÷23#$�IJ¹ºCM÷oG��
mïðHÛÜÝ�
,gQA¡úïDªÅ]^¢
073 Ö H $ ` £ ¤48¥
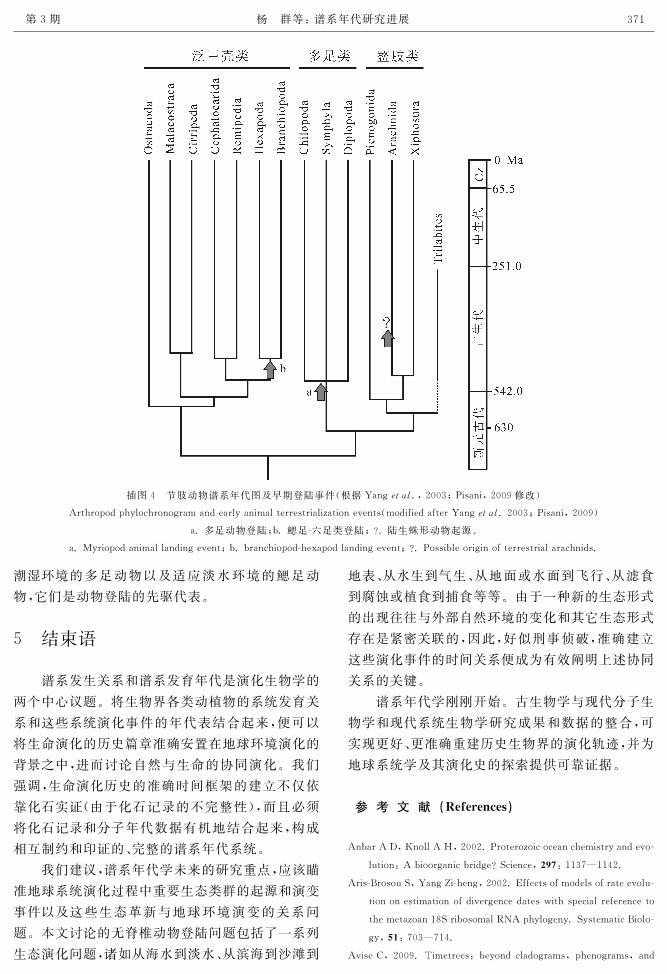
插图4 节肢动物谱系年代图及早期登陆事件(根据Yang犲狋犪犾.,2003;Pisani,2009修改)
Arthropodphylochronogramandearlyanimalterrestrializationevents(modifiedafterYang犲狋犪犾.2003;Pisani,2009)
a.多足动物登陆;b.鳃足六足类登陆;?.陆生蛛形动物起源。
a.Myriopodanimallandingevent;b.branchiopodhexapodlandingevent;?.Possibleoriginofterrestrialarachnids.
潮湿环境的多足动物以及适应淡水环境的鳃足动
物,它们是动物登陆的先驱代表。
5 结束语
谱系发生关系和谱系发育年代是演化生物学的
两个中心议题。将生物界各类动植物的系统发育关
系和这些系统演化事件的年代表结合起来,便可以
将生命演化的历史篇章准确安置在地球环境演化的
背景之中,进而讨论自然与生命的协同演化。我们
强调,生命演化历史的准确时间框架的建立不仅依
靠化石实证(由于化石记录的不完整性),而且必须
将化石记录和分子年代数据有机地结合起来,构成
相互制约和印证的、完整的谱系年代系统。
我们建议,谱系年代学未来的研究重点,应该瞄
准地球系统演化过程中重要生态类群的起源和演变
事件以及这些生态革新与地球环境演变的关系问
题。本文讨论的无脊椎动物登陆问题包括了一系列
生态演化问题,诸如从海水到淡水、从滨海到沙滩到
地表、从水生到气生、从地面或水面到飞行、从滤食
到腐蚀或植食到捕食等等。由于一种新的生态形式
的出现往往与外部自然环境的变化和其它生态形式
存在是紧密关联的,因此,好似刑事侦破,准确建立
这些演化事件的时间关系便成为有效阐明上述协同
关系的关键。
谱系年代学刚刚开始。古生物学与现代分子生
物学和现代系统生物学研究成果和数据的整合,可
实现更好、更准确重建历史生物界的演化轨迹,并为
地球系统学及其演化史的探索提供可靠证据。
参 考 文 献 (犚犲犳犲狉犲狀犮犲狊)
AnbarAD,KnollAH,2002.Proterozoicoceanchemistryandevo
lution:Abioorganicbridge?Science,297:1137—1142.
ArisBrosouS,YangZiheng,2002.Effectsofmodelsofrateevolu
tiononestimationofdivergencedateswithspecialreferenceto
themetazoan18SribosomalRNAphylogeny.SystematicBiolo
gy,51:703—714.
AviseC,2009.Timetrees:beyondcladograms,phenograms,and
173 第3期 杨 群等:谱系年代研究进展

phylograms.犐狀:HedgesB,KumarS(eds.),TheTimetreeof
Life.Oxford:OxfordUniversityPress.19—25.
BentonMJ,AyalaFJ,2003.Datingthetreeoflife.Science,300:
1698—700.
BentonMJ,DonoghuePCJ,2007.PaleontologicalEvidenceto
DatetheTreeofLife.MolecularBiologyandEvolution,24(1):
26—53.
BentonMJ,DonoghuePCJ,AsherRJ,2009.Calibratingand
constrainingmolecularclocks.犐狀:HedgesB,KumarS(eds.),
TheTimetreeofLife.Oxford:OxfordUniversityPress.35—
86.
BerneyC,PawlowskiJ,2006.Amoleculartimescaleforeukaryote
evolutionrecalibratedwiththecontinuousmicrofossilrecord.
ProceedingsoftheRoyalSocietyofLondonB,273:1867—
1872.
BhattacharyaD,YoonHS,HedgesSB,HackettJD,2009.Eu
karyotes(Eukaryota).犐狀:HedgesSB,KumarS(eds.),The
TimetreeofLife.Oxford:OxfordUniversityPress.116—120.
BlairJE,2009.Animals(Metazoa).犐狀:HedgesB,KumarS
(eds.),The TimetreeofLife.Oxford:Oxford University
Press.223—230.
BlairJE,HedgesSB,2005.Molecularclocksdonotsupportthe
Cambrianexplosion.MolecularBiologyand Evolution,22:
387—390.
ChenJunyuan,BottjerDJ,OliveriP,DornbosSQ,GaoFeng,
RuffinsS,ChiHuimei,LiChiawei,DavidsonE H,2004.
Smallbilaterianfossilsfrom40to55millionyearsbeforethe
Cambrian.Science,305:218—222.
DonoghuePCJ,SmithMP,2004.TellingtheEvolutionaryTime.
London:TaylorandFrancis.1—268.
DouzeryEJP,SnellEA,BaptesteE,DelsucF,PhilippeH,2004.
Thetimingofeukaryoticevolution:doesarelaxedmolecular
clockreconcileproteinsandfossils.ProceedingsoftheNational
AcademyofSciencesofUSA,101:15386—15391.
DrummondAJ,RambautA,2007.“BEAST:Bayesianevolution
aryanalysisbysamplingtrees.”BMCEvolutionaryBiology,7:
214.
GehlingJG,RigbyJK,1996.Longexpectedspongesfromthe
NeoproterozoicEdiacarafaunaofSouthAustralia.Journalof
Paleontology,70:185—195.
GlazkoGV,KooninEV,RogozinIB,2005.Moleculardating:ape
bonesagreewithchickenentrails.TrendsinGenetics,21:89—
92.
HedgesSB,KumarS,2009.TheTimetreeofLife.Oxford:Ox
fordUniversityPress.1—576.
HedgesSB,KumarS,2003.Genomicclocksandevolutionarytime
scales.TrendsinGenetics,19:200—206.
HedgesSB,KumarS,2004.Precisionofmoleculartimeestimates.
TrendsinGenetics,20:242—247.
HugLA,RogerAJ,2006.Theoriginanddiversificationofeu
karyotes:problemswithmolecularphylogeneticsandmolecular
clockestimation.PhilosophicalTransactionsofRoyalSociety
LondonB,361:1039—1054.
HugLA,RogerAJ,2007.Theimpactoffossilsandtaxonsam
plingonancientmoleculardatinganalyses.MolecularBiology
andEvolution,24:1889—1897.
JeramAJ,SeldenPA,EdwardsD,1990.LandanimalsintheSilu
rian:arachnidsandmyriapodsfromShropshire,England.Sci
ence,250:658—661.
JiangGanqing,KennedyMJ,ChristieBlickN,2003.Stableiso
topicevidenceformethaneseepsinNeoproterozoicpostglacial
capcarbonates.Nature,426:822—826.
JohnsonEW,BriggsDEG,SuthrenRJ,WrightJL,TunnicliffS
P,1994.NonmarinearthropodtracesfromthesubaerialOrdo
vicianBorrowdalevolcanicgroup,EnglishLakeDistrict.Geo
logicalMagazine,131:395—406.
KishinoH,ThorneJL,BrunoWJ,2001.Performanceofadiver
gencetimeestimationmethodunderaprobabilisticmodelofrate
evolution.MolecularBiologyandEvolution,18:352—361.
KumarS,FilipskiA,SwarnaV,WalkerA,HedgesSB,2005.
Placingconfidencelimitsonthemolecularageofthehuman
chimpanzeedivergence.ProceedingsoftheNationalAcademyof
SciencesofUSA,102:18842—18847.
KumarS,2005.Molecularclocks:fourdecadesofevolution.Na
tureReviewsGenetics,6:654662.
LiChiawei,ChenJunyuan,HuaTzuen,1998.Precambrianspon
geswithcellularstructures.Science,279:879—882.
LoveGD,GrosjeanE,StalviesC,FikeDA,GrotzingerJP,Brad
leyAS,KellyAE,BhatiaM,MeredithW,SnapeCE,Bow
ringSA,CondonDJ,SummonsRE,2009.
FossilsteroidsrecordtheappearanceofDemospongiaeduringthe
Cryogenianperiod.Nature,457:18—21.
MacNaughtonRB,ColeJM,DalrympleRW,BraddySJ,Briggs
DEG,LukieTD,2002.Firststepsonland:Arthropodtrack
waysinCambrianOrdovicianeoliansandstone,southeastern
Ontario,Canada.Geology,30:391—394.
MagallonSA,SandersonMJ,2005.Angiospermdivergencetimes:
Theeffectofgenes,codonpositions,andtimeconstraints.E
volution,59:1653—1670.
MargoliashE,1963.Primarystructureandevolutionofcytochrome
c.ProceedingsofNationalAcademyofSciencesofUSA,50:
672—679.
NearTJ,SandersonMJ,2004.Assessingthequalityofmolecular
divergencetimeestimatesbyfossilcalibrationsandfossilbased
modelselection.PhilosophicalTransactionsofRoyalSociety
LondonB,359:1477—1483.
NearTJ,MeylanPA,ShafferHB,2005.Assessingconcordance
offossilcalibrationpointsinmolecularclockstudies:anexam
pleusingturtles.AmericanNaturalist,165:137—146.
NeiM,KumarS,2000.MolecularEvolutionandPhylogenetics.
Oxford:OxfordUniversityPress.1—336.
PereiraSL,BakerAJ,2006.Amitogenomicstimescaleforbirds
detectsvariablephylogeneticratesofmolecularevolutionand
refutesthestandardmolecularclock.MolecularBiologyandE
273 古 生 物 学 报 第48卷

volution,23:1731—1740.
PetersonKJ,ButterfieldNJ,2005.OriginoftheEumetazoa:Tes
tingecologicalpredictionsofmolecularclocksagainsttheProt
erozoicfossilrecord.ProceedingsoftheNationalAcademyof
SciencesofUSA,102:9547—9552.
PetersonKJ,CottonJA,GehlingJG,PisaniD,2008.TheEdia
caranemergenceofbilaterians:congruencebetweenthegenetic
andthegeologicalfossilrecords.PhilosophicalTransactionsof
theRoyalSocietySeriesB,363:1435—1443.
PisaniD,2009.Arthropods(Arthropoda).犐狀:HedgesSB,Kumar
S(eds.),TheTimetreeofLife.Oxford:OxfordUniversity
Press.251—254.
PisaniD,BentonMJ,WilkinsonW,2007.Congruenceofmorpho
logicaland molecularPhylogenies.ActaBiotheoretica,55:
269—281.
ProcheE,2008.Backtothesea:secondarymarineorganismsfrom
abiogeographicalperspective.BiologicalJournaloftheLinnean
Society,74:197—203.
RambautA,BromhamL,1998.Estimatingdivergencedatesfrom
molecularsequences.MolecularBiologyandEvolution,15:
442—448.
RobisonRA,1990.Earliestknownuniramousarthropod.Nature,
343:163,164.
RunnegarB,1986.Molecularpalaeontology.Palaeontology,29:
1—24.
RunnegarB,1982.Amolecularclockdatefortheoriginofanimal
phyla.Lethaia,15:199—205.
SandersonMJ,1997.Anonparametricapproachtoestimatingdi
vergencetimesintheabsenceofrateconstancy.MolecularBiol
ogyandEvolution,14:1218—1231.
SandersonMJ,2002.Estimatingabsoluteratesofmolecularevolu
tionanddivergencetimes:apenalizedlikelihoodapproach.Mo
lecularBiologyandEvolution,19:101—109.
SandersonMJ,2003.r8s:inferringabsoluteratesofmolecularevo
lutionanddivergencetimesintheabsenceofamolecularclock.
Bioinformatics,19:301,302.
SoltisPS,SoltisDE,SavolainenV,CranePR,BarracloughTG,
2002.Rateheterogeneityamonglineagesoftracheophytes:in
tegrationofmolecularandfossildataandevidenceformolecular
livingfossils.ProceedingsoftheNationalAcademyofSciences
ofUSA,99:4430—4435.
StrotherPK,WoodGD,TaylorW A,BeckJH,2004.Middle
Cambriancryptosporesandtheoriginoflandplants.Memoirs
oftheAssociationofAustralasianPalaeontologists,29:99—
113.
TajimaF,1993.Simplemethodsfortestingmolecularclockhypoth
esis.Genetics,135:599—607.
TakezakiN,RzhetskyA,NeiM,1995.Phylogenetictestofthe
molecularclockandlinearizedtrees.MolecularBiologyandE
volution,12:823—833.
ThorneJL,KishinoH,PainterIS,1998.Estimatingtherateofe
volutionoftherateofmolecularevolution.MolecularBiology
andEvolution,15:1647—1657.
TreeofLifeWebProject,1997.LifeonEarth.http://tolweb.org/
vanRheedeT,BastiaansT,BooneDN,HedgesSB,deJong
W W,MadsenO,2006.Theplatypusisinitsplace:nuclear
genesandindelsconfirmthesistergrouprelationofmonotremes
andtherians.MolecularBiologyandEvolution,23:587—597.
WaggonerBM,1996.Phylogenetichypothesesoftherelationships
ofarthropodstoPrecambrianandCambrianproblematicfossil
taxa.SystematicBiology,45:190—222.
WaggonerBM,CollinsAG,2004.Reductioadabsurdum:testing
theevolutionaryrelationshipsofEdiacaranandPaleozoicprob
lematicfossilsusingmoleculardivergencedates.JournalofPa
leontology,78:51—61.
WelchJJ,BromhamL,2005.Moleculardatingwhenratesvary.
TrendsinEcologyandEvolution,20:320327.
XiaoShuhai,2004.Neoproterozoicglaciationsandthefossilre
cord.犐狀:Jenkins G,McMenamin M,SohlL,McKay C
(eds.),TheExtremeProterozoic:Geology,Geochemistry,
andClimate.WashingtonDC:AmericanGeophysicalUnion.
199—214.
YangQun,MaJunye,SunXiaoyan,CongPeiyun,2007.Phylo
chronologyofearlymetazoans:combinedevidencefrommolecu
larandfossildata.GeologicalMagazine,42(3/4):281—295.
YangQun,SunXiaoyan,ZhangKeyun,WuPing,2003.Arthro
podphylogeneticchronology.JournalofGeneticsandMolecular
Biology,14(3):145—158.
YangZiheng,RannalaB,2006.Bayesianestimationofspeciesdi
vergencetimesunderamolecularclockusingmultiplefossilcali
brationswithsoftbounds.MolecularBiologyandEvolution,
23:212—226.
YinChongyu(尹崇玉),GaoLinzhi(高林志),XingYusheng(邢裕
盛).2001.DiscoveryofTianzhushaniainDoushantuophos
phorites,inWeng’an,GuizhouProvince.ActaPalaeontologica
Sinica,40(4):497—504(inChinesewithEnglishabstract).
YoderAD,YangZiheng,2000.Estimationofprimatespeciation
datesusinglocalmolecularclocks.MolecularBiologyandEvo
lution,17:1081—1090.
YuanXunlai,XiaoShuhai,TaylorTN,2005.Lichenlikesymbi
osis600millionyearsago.Science,308:1017—1020.
ZuckerkandlE,PaulingL,1962.Moleculardisease,evolution,and
genicheterogeneity.犐狀:KashaM,PullmanB(eds.),Hori
zonsinBiochemistry.NewYork:AcademicPress.189—225.
ZuckerkandlE,PaulingL,1965.Evolutionarydivergenceandcon
vergenceinproteins.犐狀:BrysonV,VogelHJ(eds),Evolving
GenesandProteins.NewYork:AcademicPress.97—166.
373 第3期 杨 群等:谱系年代研究进展

犃犇犞犃犖犆犈犛犐犖犘犎犢犔犗犌犈犖犈犜犐犆犆犎犚犗犖犗犔犗犌犢
YANGQun1),CONGPeiyun
1,2),SUNXiaoyan1),MAJunye
1),GAIYonghua1),
LIChunxiang1),HAOJiasheng
1,3)andXIAXuhua4)
1)犛狋犪狋犲犓犲狔犔犪犫狅狉犪狋狅狉狔狅犳犘犪犾犪犲狅犫犻狅犾狅犵狔犪狀犱犛狋狉犪狋犻犵狉犪狆犺狔,犖犪狀犼犻狀犵犐狀狊狋犻狋狌狋犲狅犳犌犲狅犾狅犵狔犪狀犱犘犪犾犪犲狅狀狋狅犾狅犵狔,
犆犺犻狀犲狊犲犃犮犪犱犲犿狔狅犳犛犮犻犲狀犮犲狊,犖犪狀犼犻狀犵210008,犆犺犻狀犪,狇狌狀狔犪狀犵@狀犻犵狆犪狊.犪.犮狀;
2)犢狌狀狀犪狀犓犲狔犔犪犫狅狉犪狋狅狉狔犳狅狉犘犪犾犪犲狅犫犻狅犾狅犵狔,犢狌狀狀犪狀犝狀犻狏犲狉狊犻狋狔,犓狌狀犿犻狀犵650091,犆犺犻狀犪;
3)犛犮犺狅狅犾狅犳犔犻犳犲犛犮犻犲狀犮犲,犃狀犺狌犻犖狅狉犿犪犾犝狀犻狏犲狉狊犻狋狔,犠狌犺狌241000,犃狀犺狌犻,犆犺犻狀犪;
4)犇犲狆犪狉狋犿犲狀狋狅犳犅犻狅犾狅犵狔,犝狀犻狏犲狉狊犻狋狔狅犳犗狋狋犪狑犪,犗犖犓1犖6犖5,犆犪狀犪犱犪
犓犲狔狑狅狉犱狊 Phylochronolgy,molecularpaleontology,relaxedmolecularclock,TimetreeofLife,animalterrestrial
ization
犃犫狊狋狉犪犮狋
Phylogeneticchronology,orphylochronolo
gy,hasbecomeanactiveresearchareainevolu
tionary biology. With phylogenetic divergence
datesestablishedfortheevolutionaryhistory,the
bioticevolutionandthephylogeneticstepsmaybe
comprehendedinthecontextofthephysicalenvi
ronmentalchangesduringtheearthhistory.Itis
emphasizedhereinthattheintegrativeapproachof
fossildatesandtherelaxedmolecularclocktech
niqueswouldyieldabetterpictureoftheevolu
tionarytimescaleoramorepreciseandmoreaccu
ratephylochronogramsfororganismgroupsthanif
onlyfossildatesoronlymoleculartimescalesare
considered.Aconsensustimescalewithbothmo
leculardatesandfossilconstraintsispreferred
and,furthermore,aninterpretationfortheevolu
tionaryschemesthusestablishedshouldbedonein
thecontextofthegeologicalbackgrounds;inother
words,theevolutionarytimescaleislaidoutasthe
timeframeforthecoevolutionaryhistorybetween
lifeandtheEarth’sphysicalenvironment.We
presentrecentdevelopmentin moleculardating
techniquesbasedontherelaxed molecularclock
conceptandstrategiesinfossilcalibration.Exam
plesinrecentdevelopmentofanimalphylochronol
ogyandestimationofthedatesofearlyinverte
brateanimallandingarediscussedwiththeintegra
tionoffossildates,molecularchronology,andge
ologicalbackgrounds.Westresstheimportanceof
fossildatesasthefoundationforphylochronology
andgeologicalbackgroundasprobablemechanisms
forphylogeneticdivergencesorradiations.
473 古 生 物 学 报 第48卷





























