Embed Size (px)
Citation preview

Vol. 2, 503-510, October 1991 Cell Growth & Differentiation 503
A Developmentally Regulated Octamer-binding Activityin Embryonal Carcinoma Cells Which Represses/3-Interferon Expression’
Allison Haggarty,2 Rino Camato, Gary Paterno,Lucie Cohen, John Hiscott, and Daniel Skup3
Institut du Cancer de Montr#{233}al,Montr#{233}al,Quebec H2L 4M1 [A. H.,R. C., D. S.]; Lady Davis Institute for Medical Research, Sir Mortimer B.Davis-Jewish General Hospital and Department of Microbiology andImmunology, McGill University, Montreal, Quebec H3T 1E2 EL. C.,J. H.]; and Faculty of Medicine, Memorial University, Saint John’s,Newfoundland [G. P.], Canada
AbstradWe have previously described ECIF-1, a DNA-bindingfador present in nuclear extracts of murine embryonalcarcinoma cells which specifically recognizes a regionwithin the human a-interferon promoter. We show thatthe promoter region located between -1 1 2 and -93 issufficient for this binding adivity, which is not due tobinding of interferon-regulatory fador 1 or 2. Bymutational analysis of the ECIF-1 site, it wasdetermined that the central nucleotides which arecritical for binding contain an odameric motif:ATTTACAT. The binding adivity of ECIF-1 with itscognate site within the a-interferon promoter decreasesupon differentiation concurrently with the onset ofinterferon inducibility. Furthermore, by using an invitro transcription assay with deleted promoterelements of the $-interferon gene, we show thatundifferentiated P19 nuclear extrads contain arepressing adivity which depends on the presence ofthe ECIF-1 site. This repression is not observed usingnuclear extrads from differentiated P19 cells.Comparison of the binding adivity of this odamer sitewith others previously shown to be adive in embryonalcarcinoma cells reveals similarities and differences inthe spedrum of proteins binding there.
Introdudion
EC4 cells serve as a model system for investigating theprocesses involved during cell differentiation. MurineP19 EC cells (1) will differentiate in vitro in response toretinoic acid or DMSO to form neuronal or muscle cells,
Received 11/12/90.1 This work was supported by grants from the Medical Research Councilofdanadato D. S. andJ. H.2 A. H. holds a postdoctoral fellowship from the Fonds de Ia rechercheen sante de Qu#{233}bec.Present address: Centre for Research in Neurosci-ence, McGill University, Montreal General Hospital Research Institute,Montreal, Quebec, Canada.3 To whom requests for reprints should be addressed, at Institut duCancer de Montr#{233}al,1560 Sherbrooke est, Montr#{233}al,Qu#{233}bec,H2L 4Mi,Canada.4 The abbreviations used are: Ed, embryonal carcinoma; DMSO, dimethylsulfoxide; IFN, interferon; IRE, interferon-responsive element; PRD andNRD, positive and negative regulatory domains; IRF, interferon-regulatoryfactor; bp, base pair(s); R�G, rabbit �-globin; Kd, kilodalton(s); BrdUrd,5-bromodeoxyuridine; SDS, sodium dodecyl sulfate.
respectively. During the course of differentiation, themorphology of the cells is altered, and levels of differ-entiation markers are seen to change (2). One marker ofdifferentiation is the inducibility of the IFN genes. IFNsare not made in response to inducers (virus or doublestranded RNA) in undifferentiated cells (3), but differen-tiating cells rapidly acquire the ability to respond toinducers by producing IFNs (4, 5).
The IFN-fi promoter contains a region known as theIRE (-77 to -37 upstream ofthe cap site), which containstwo positive (PRD-l and PRD-lI) and one negative (NRD-I) regulatory domain (6, 7). A study using deletion mutantsgave an indication of a negative control region furtherupstream of the IRE (-210 to -107) (8). A number ofpositive factors bind to the IRE, including IRF-1, NF-KB,and PRD-ll-BF-1 (9-11), as well as at least one negativefactor, IRF-2 (12). IRF-1 and IRF-2 bind to a hexamericmotif in the PRD-II region (consensus = AA(G/T)(T/G)GA), seven variations of which are found within 110bp upstream of the cap site (1 3). Up until now, analysisof the regulation of the IFN-fl gene has focused primarilyon the events surrounding induction of the gene indifferentiated cells, and developmentally regulated fac-tors have not been fully investigated except for a recentpaper showing that IRF-1 is an activator and IRF-2 arepressor of type 1 IFN expression in EC cells (14).
Many proteins that have been implicated in the regu-lation of development and differentiation contain a re-gion of homology called the homeodomain, 60 aminoacids long (reviewed in Refs. 15 and 16). To date, allhomeodomain-containing proteins have been localizedto the nucleus, and some have been shown to act astranscription factors. Proteins in the #{176}POU”subfamilycontain a homeodomain in donjunction with a 75-aminoacid POU-specific domain. Two members ofthis subfam-ily are Oct-i (17), a ubiquitous factor, and Oct-2 (18, 19),a B-cell-specific factor. They bind to an octameric se-quende, ATTTGCAT, found in the promoters of a varietyofgenes (15).
Octamer-binding proteins have also been describedin EC cells. Lenardo et a/. (20) have described NF-A3, afactor from F9 EC cells that binds to the ATTTGCAToctameric sequence and whose activity decreases whenthe cells are differentiated. NF-A3 has been cloned (21)and appears to be derived from the same gene as Oct-4[cloned from F9 cells by Sch#{244}leret a!. (22-25)] and Oct-3 [cloned by Okamoto et a/. (26)]. Oct-3, identified bythe enhancer trap method in P19 cells (27), was shownto bind to the ATTAGCAT octamer, which has one nu-cleotide changed from the original octameric sequence.All three of these transcription factors interact with en-hancers on reporter genes in undifferentiated cells. How-ever, their target genes in vivo have not yet beenidentified.
In order to study the process of gene regulation duringdifferentiation in more detail, we looked for nuclear
504 Octamer-binding Protein Recognizing the Human IFN-fl Promoter
I 23
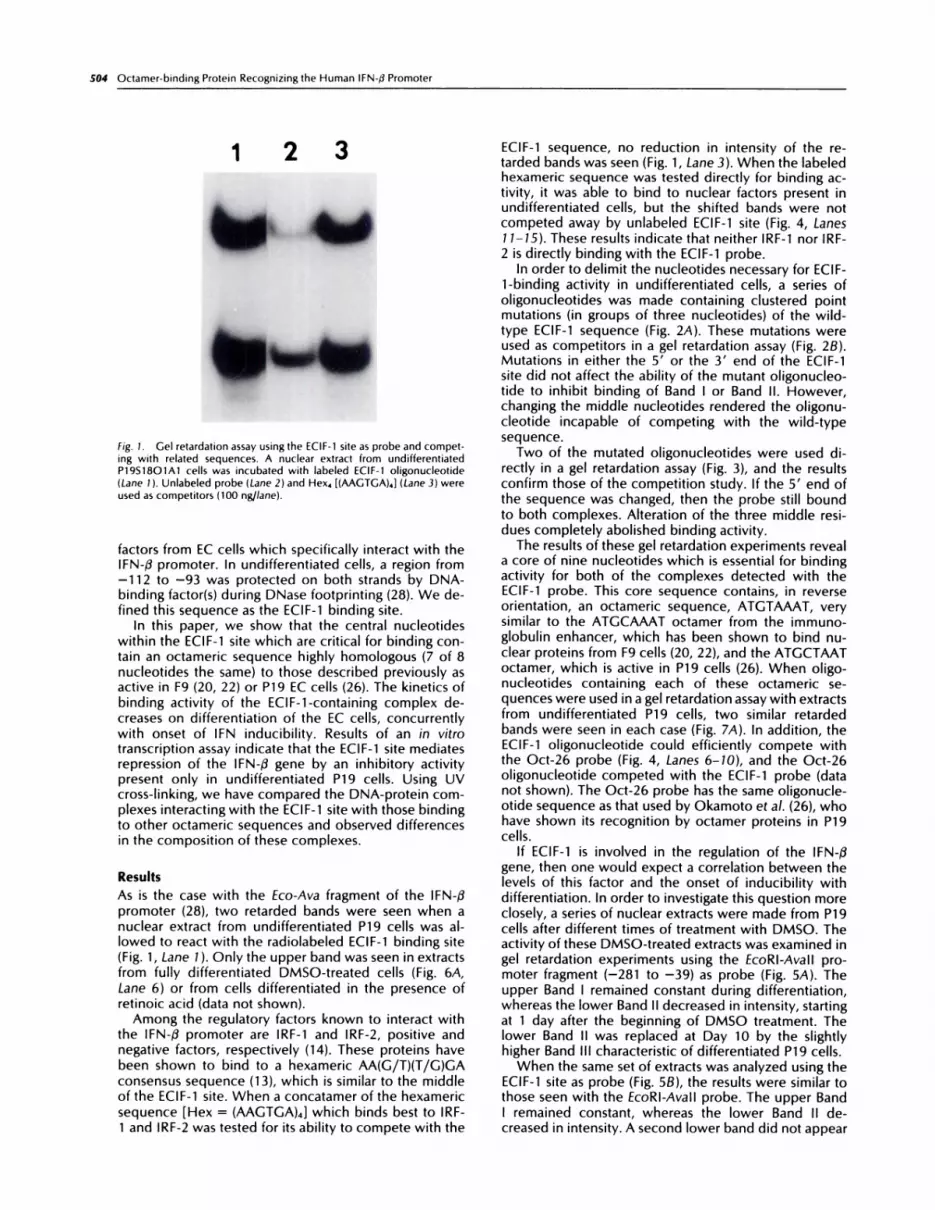
fig. 1. Gel retardation assay using the ECIF-1 site as probe and compet-ing with related sequences. A nuclear extract from undifferentiatedP19S18O1A1 cells was incubated with labeled ECIF-1 oligonucleotide
(Lane 1 ). Unlabeled probe (Lane 2) and Hex4 [(AAGTGA)4} (Lane 3) wereused as competitors (100 ng/lane).
factors from EC cells which specifically interact with theIFN-f3 promoter. In undifferentiated cells, a region from-112 to -93 was protected on both strands by DNA-binding factor(s) during DNase footprinting (28). We de-fined this sequence as the ECIF-1 binding site.
In this paper, we show that the central nucleotideswithin the ECIF-1 site which are critical for binding con-tam an octameric sequence highly homologous (7 of 8nucleotides the same) to those described previously asactive in F9 (20, 22) or P19 EC cells (26). The kinetics ofbinding activity of the ECIF-1-containing complex de-creases on differentiation of the EC cells, concurrentlywith onset of IFN inducibility. Results of an in vitrotranscription assay indicate that the ECIF-1 site mediatesrepression of the IFN-/3 gene by an inhibitory activitypresent only in undifferentiated P19 cells. Using UVcross-linking, we have compared the DNA-protein com-plexes interacting with the ECIF-1 site with those bindingto other octameric sequences and observed differencesin the composition of these complexes.
Results
As is the case with the Eco-Ava fragment of the IFN-flpromoter (28), two retarded bands were seen when anuclear extract from undifferentiated P19 cells was aI-lowed to react with the radiolabeled ECIF-1 binding site(Fig. 1 , Lane 1 ). Only the upper band was seen in extractsfrom fully differentiated DMSO-treated cells (Fig. 6A,Lane 6) or from cells differentiated in the presence ofretinoic acid (data not shown).
Among the regulatory factors known to interact withthe IFN-fl promoter are IRF-1 and IRF-2, positive andnegative factors, respectively (14). These proteins havebeen shown to bind to a hexameric AA(G/T)(T/G)GAconsensus sequence (13), which is similar to the middleof the ECIF-1 site. When a concatamer of the hexamericsequence [Hex = (AAGTGA)4] which binds best to IRF-1 and IRF-2 was tested for its ability to compete with the
ECIF-1 sequence, no reduction in intensity of the re-tarded bands was seen (Fig. 1, Lane 3). When the labeledhexameric sequence was tested directly for binding ac-tivity, it was able to bind to nuclear factors present inundifferentiated cells, but the shifted bands were notcompeted away by unlabeled ECIF-1 site (Fig. 4, Lanes1 1-15). These results indicate that neither IRF-1 nor IRF-2 is directly binding with the ECIF-1 probe.
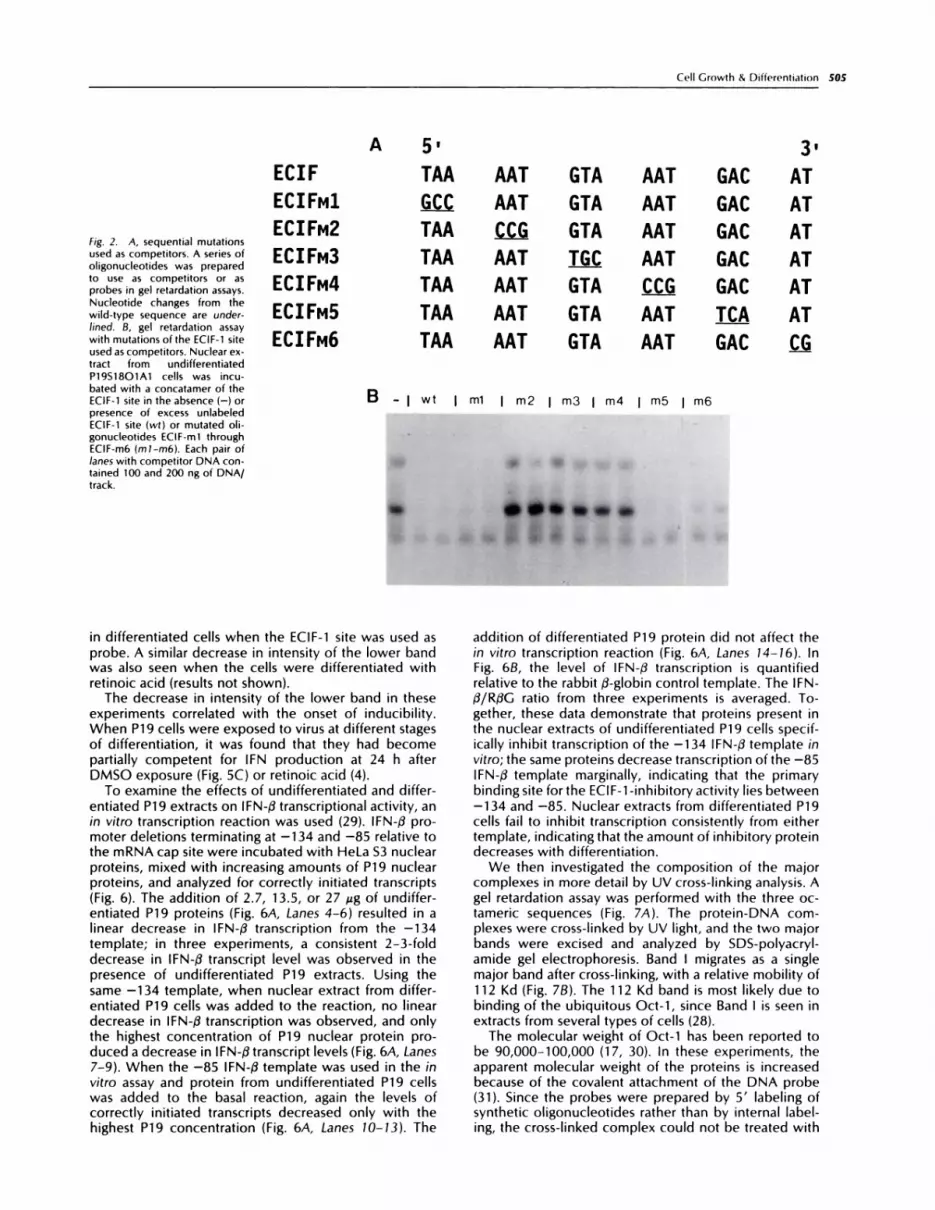
In order to delimit the nucleotides necessary for ECIF-1-binding activity in undifferentiated cells, a series ofoligonucleotides was made containing clustered pointmutations (in groups of three nucleotides) of the wild-type ECIF-1 sequence (Fig. 2A). These mutations wereused as competitors in a gel retardation assay (Fig. 28).Mutations in either the 5’ or the 3’ end of the ECIF-1site did not affect the ability of the mutant oligonucleo-tide to inhibit binding of Band I or Band II. However,changing the middle nucleotides rendered the oligonu-cleotide incapable of competing with the wild-typesequence.
Two of the mutated oligonucleotides were used di-rectly in a gel retardation assay (Fig. 3), and the resultsconfirm those of the competition study. If the 5’ end ofthe sequence was changed, then the probe still boundto both complexes. Alteration of the three middle resi-dues completely abolished binding activity.
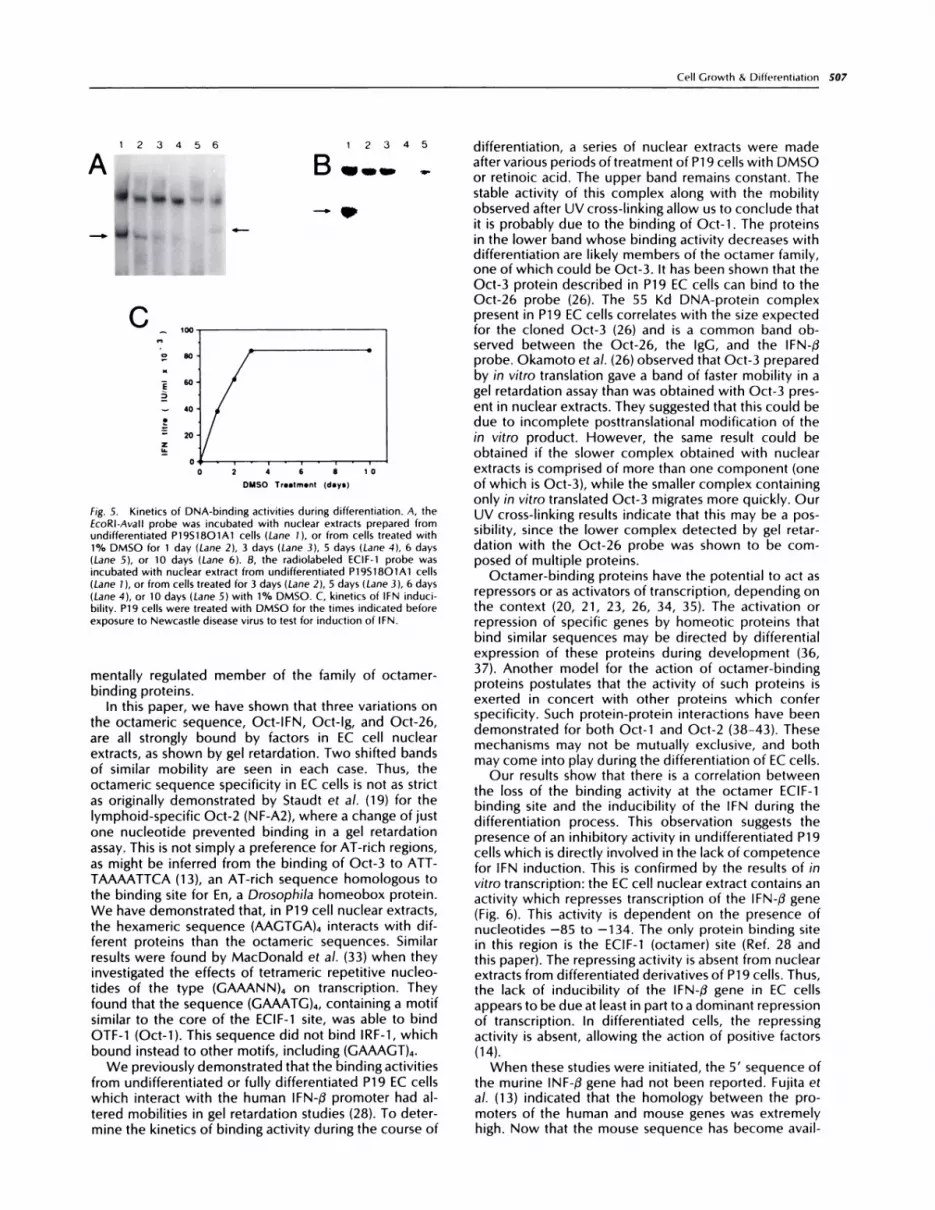
The results of these gel retardation experiments reveala core of nine nucleotides which is essential for bindingactivity for both of the complexes detected with theECIF-1 probe. This core sequence contains, in reverseorientation, an octameric sequence, ATGTAAAT, verysimilar to the ATGCAAAT octamer from the immuno-globulin enhancer, which has been shown to bind nu-clear proteins from F9 cells (20, 22), and the ATGCTAAToctamer, which is active in P19 cells (26). When oligo-nucleotides containing each of these octameric se-quences were used in a gel retardation assay with extractsfrom undifferentiated P19 cells, two similar retardedbands were seen in each case (Fig. 7,4). In addition, theECIF-1 oligonucleotide could efficiently compete withthe Oct-26 probe (Fig. 4, Lanes 6-10), and the Oct-26oligonucleotide competed with the ECIF-1 probe (datanot shown). The Oct-26 probe has the same oligonucle-otide sequence as that used by Okamoto et a!. (26), whohave shown its recognition by octamer proteins in P19cells.
If ECIF-1 is involved in the regulation of the IFN-�3gene, then one would expect a correlation between thelevels of this factor and the onset of inducibility withdifferentiation. In order to investigate this question moreclosely, a series of nuclear extracts were made from P19cells after different times of treatment with DMSO. Theactivity of these DMSO-treated extracts was examined ingel retardation experiments using the EcoRl-Avall pro-moter fragment (-281 to -39) as probe (Fig. 5A). Theupper Band I remained constant during differentiation,whereas the lower Band II decreased in intensity, startingat 1 day after the beginning of DMSO treatment. Thelower Band II was replaced at Day 10 by the slightlyhigher Band Ill characteristic of differentiated P19 cells.
When the same set of extracts was analyzed using theECIF-1 site as probe (Fig. SB), the results were similar tothose seen with the EcoRI-Avall probe. The upper BandI remained constant, whereas the lower Band II de-creased in intensity. A second lower band did not appear
A 5’
EC I FM3
ECIFM4
ECIFM5
ECIFM6
Fig. 2. A, sequential mutationsused as competitors. A series ofoligonucleotides was prepared
to use as competitors or asprobes in gel retardation assays.Nucleotide changes from thewild-type sequence are under-
lined. B, gel retardation assay
with mutations ofthe ECIF-1 siteused as competitors. Nuclear ex-tract from undifferentiatedP19S18O1A1 cells was incu-bated with a concatamer of theECIF-1 site in the absence (-) orpresence of excess unlabeledECIF-1 site (wt) or mutated oh-gonucleotides ECIF-m1 throughECIF-m6 (ml-m6). Each pair oflanes with competitor DNA con-tamed 100 and 200 ng of DNA/track.
B - I wt I ml I m2 I m3 I m4 I m5 I m6
*
,� �.IR*e4�4
Cell Growth A Differentiation 505
ECIF
EC I FM1
ECIFM2
3,TAA AAT GTA AAT GAC AT
�_c_� AAT GTA AAT GAC AT
TAA � GTA AAT GAC AT
TAA AAT � AAT GAC AT
TAA AAT GTA CC� GAC AT
TAA AAT GTA AAT J.�CA AT
TAA AAT GTA AAT GAC Ci
. ..
in differentiated cells when the ECIF-1 site was used asprobe. A similar decrease in intensity of the lower bandwas also seen when the cells were differentiated withretinoic acid (results not shown).
The decrease in intensity of the lower band in theseexperiments correlated with the onset of inducibility.When P19 cells were exposed to virus at different stagesof differentiation, it was found that they had becomepartially competent for IFN production at 24 h afterDMSO exposure (Fig. SC) or retinoic acid (4).
To examine the effects of undifferentiated and differ-entiated P19 extracts on IFN-/3 transcriptional activity, anin vitro transcription reaction was used (29). IFN-/� pro-moter deletions terminating at -134 and -85 relative tothe mRNA cap site were incubated with HeLa 53 nuclearproteins, mixed with increasing amounts of P19 nuclearproteins, and analyzed for correctly initiated transcripts(Fig. 6). The addition of 2.7, 13.5, or 27 �ag of undiffer-entiated P19 proteins (Fig. 6A, Lanes 4-6) resulted in alinear decrease in lFN-/.� transcription from the -134template; in three experiments, a consistent 2-3-folddecrease in lFN-f� transcript level was observed in thepresence of undifferentiated P19 extracts. Using thesame -134 template, when nuclear extract from differ-entiated P19 cells was added to the reaction, no lineardecrease in IFN-/3 transcription was observed, and onlythe highest concentration of P19 nuclear protein pro-duced a decrease in IFN-f3 transcript levels (Fig. 6A, Lanes7-9). When the -85 IFN-/3 template was used in the invitro assay and protein from undifferentiated P19 cellswas added to the basal reaction, again the levels ofcorrectly initiated transcripts decreased only with thehighest P19 concentration (Fig. 6A, Lanes 10-13). The
addition of differentiated P19 protein did not affect thein vitro transcription reaction (Fig. 6A, Lanes 14-16). InFig. 6B, the level of lFN-�3 transcription is quantifiedrelative to the rabbit f3-globin control template. The IFN-13/R/3G ratio from three experiments is averaged. To-gether, these data demonstrate that proteins present inthe nuclear extracts of undifferentiated P19 cells specif-ically inhibit transcription of the -134 IFN-fl template invitro; the same proteins decrease transcription of the -85lFN-� template marginally, indicating that the primarybinding site for the ECIF-1-inhibitory activity lies between-134 and -85. Nuclear extracts from differentiated P19cells fail to inhibit transcription consistently from eithertemplate, indicating that the amount of inhibitory proteindecreases with differentiation.
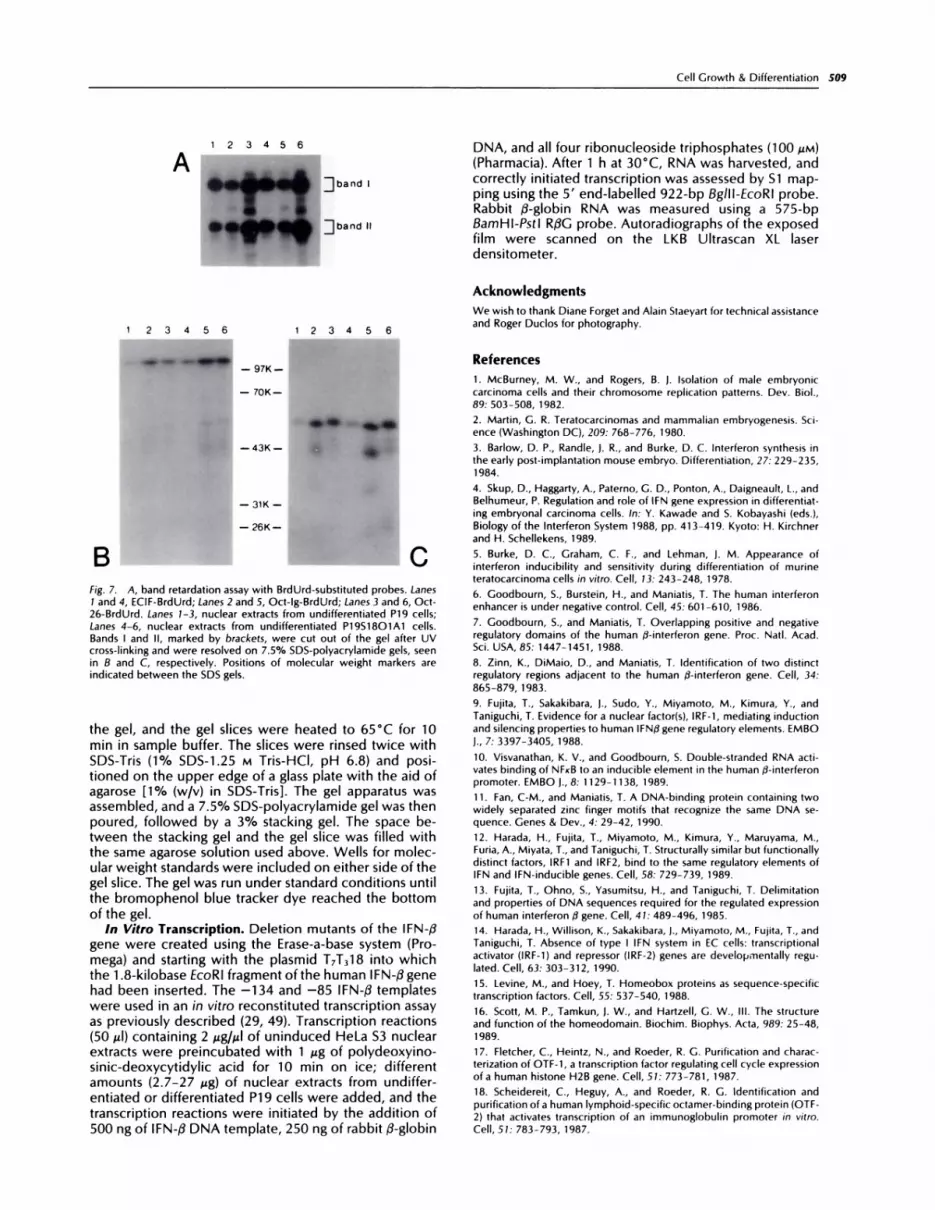
We then investigated the composition of the majorcomplexes in more detail by UV cross-linking analysis. Agel retardation assay was performed with the three oc-tameric sequences (Fig. 7A). The protein-DNA com-plexes were cross-linked by UV light, and the two majorbands were excised and analyzed by SDS-polyacryl-amide gel electrophoresis. Band I migrates as a singlemajor band after cross-linking, with a relative mobility of1 12 Kd (Fig. 7B). The 1 12 Kd band is most likely due tobinding of the ubiquitous Oct-i, since Band I is seen inextracts from several types of cells (28).
The molecular weight of Oct-i has been reported tobe 90,000-100,000 (17, 30). In these experiments, theapparent molecular weight of the proteins is increasedbecause of the covalent attachment of the DNA probe(31). Since the probes were prepared by 5’ labeling ofsynthetic oligonucleotides rather than by internal label-ing, the cross-linked complex could not be treated with
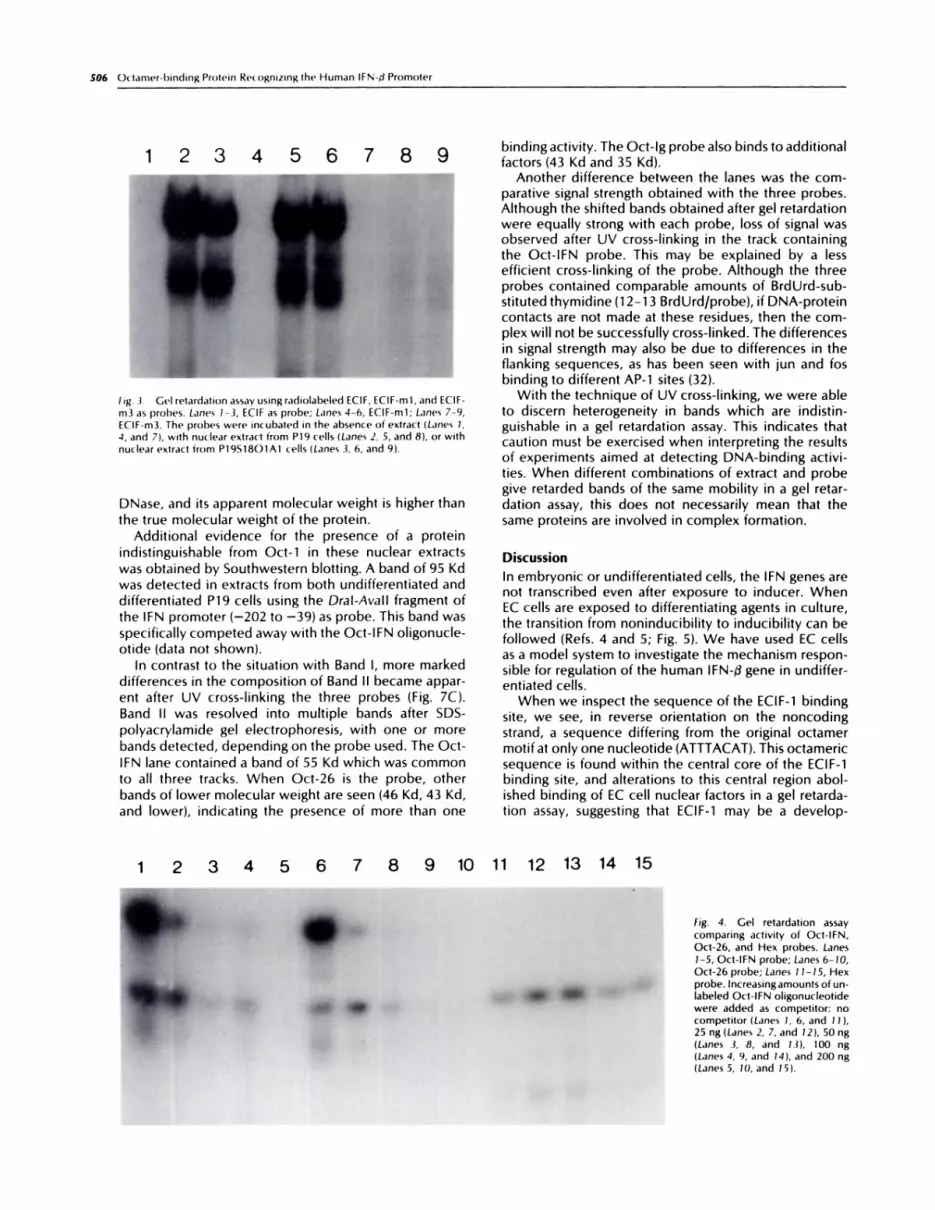
fig. .3. Ge’) retardation assay usingradiolabeled ECIF, ECIF-mi, and ECIF-
m3 as prol)es. Lanes 1-3, ECIF as probe; Lanes 4-6, ECIF-mi; Lanes 7-9,ECIF-m3. The probes were incubated in the absence of extract (Lanes 1.4, and 7), with nuclear extract from P19 cells (Lanes 2, 5, and 8), or withnuclear extract from P19S18O1A1 cells (Lanes 3, 6, and 9(.
indistinguishable from Oct-i in these nuclear extractswas obtained by Southwestern blotting. A band of 95 Kdwas detected in extracts from both undifferentiated and
differentiated P19 cells using the DraI-AvaIl fragment ofthe IFN promoter (-202 to -39) as probe. This band wasspecifically competed away with the Oct-IFN oligonucle-otide (data not shown).
In contrast to the situation with Band I, more markeddifferences in the composition of Band II became appar-ent after UV cross-linking the three probes (Fig. 7C).Band II was resolved into multiple bands after SDS-polyacrylamide gel electrophoresis, with one or morebands detected, depending on the probe used. The Oct-IFN lane contained a band of 55 Kd which was common
to all three tracks. When Oct-26 is the probe, otherbands of lower molecular weight are seen (46 Kd, 43 Kd,and lower), indicating the presence of more than one
Discussion
In embryonic or undifferentiated cells, the IFN genes arenot transcribed even after exposure to inducer. WhenEC cells are exposed to differentiating agents in culture,the transition from noninducibility to inducibility can befollowed (Refs. 4 and 5; Fig. 5). We have used EC cellsas a model system to investigate the mechanism respon-sible for regulation of the human IFN-fl gene in undiffer-entiated cells.
When we inspect the sequence of the ECIF-i bindingsite, we see, in reverse orientation on the noncodingstrand, a sequence differing from the original octamermotifat only one nucleotide (ATTTACAT). This octamericsequence is found within the central core of the ECIF-ibinding site, and alterations to this central region abol-shed binding of EC cell nuclear factors in a gel retarda-
tion assay, suggesting that ECIF-i may be a develop-
506 Octamer-binding Protein Recognizing the Human lFN�a Promoter
r) 0 5 ‘� 7 0 (� binding activity. The Oct-Ig probe also binds to additional
� � 4 U � � factors (43 Kd and 35 Kd).
Another difference between the lanes was the com-parative signal strength obtained with the three probes.Although the shifted bands obtained after gel retardationwere equally strong with each probe, loss of signal wasobserved after UV cross-linking in the track containingthe Oct-IFN probe. This may be explained by a lessefficient cross-linking of the probe. Although the threeprobes contained comparable amounts of BrdUrd-sub-stituted thymidine(12-i3 BrdUrd/probe), if DNA-proteincontacts are not made at these residues, then the com-
. plex will not be successfully cross-linked. The differences� � in signal strength may also be due to differences in the
.� .4 � flanking sequences, as has been seen with jun and fos.. binding to different AP-1 sites (32).
With the technique of UV cross-linking, we were ableto discern heterogeneity in bands which are indistin-guishable in a gel retardation assay. This indicates thatcaution must be exercised when interpreting the resultsof experiments aimed at detecting DNA-binding activi-ties. When different combinations of extract and probegive retarded bands of the same mobility in a gel retar-
DNase, and its apparent molecular weight is higher than dation assay, this does not necessarily mean that thethe true molecular weight of the protein. same proteins are involved in complex formation.
Additional evidence for the presence of a protein
12345678
*
9 10 11 12 13 14 15
.�r- -1� I, �
fig. 4. Gel retardation assay
comparing activity of Oct-IFN,
Oct-26, and Hex probes. Lanes1-5, Oct-IFN probe; Lanes 6-10,Oct-26 probe; Lanes 11-15, Hex
probe. Increasing amounts of un-labeled Oct-IFN ohigonucleotidewere added as competitor: nocompetitor (Lanes 1, 6, and 1 1),25 ng (Lanes 2, 7, and 12), 50 ng(Lanes 3, 8, and 13), 100 ng
(Lanes 4, 9, and 14), and 200 ng(Lanes 5, 10, and 15(.
C
ED
z
A _-� .�
fig. 5. Kinetics of DNA-binding activities during differentiation. A, the
EcoRl-Avall probe was incubated with nuclear extracts prepared fromundifferentiated P19S18O1A1 cells (Lane 1), or from cells treated with1% DMSO for 1 day (Lane 2), 3 days (Lane 3), 5 days (Lane 4), 6 days
(Lane 5), or 10 days (Lane 6). B, the radiolabeled ECIF-1 probe wasincubated with nuclear extract from undifferentiated P19S18O1A1 cells
(Lane 1), or from cells treated for 3 days (Lane 2), 5 days (Lane 3), 6 days(Lane 4), or 10 days (Lane 5) with 1% DMSO. C, kinetics of IFN induci-
bility. P19 cells were treated with DMSO for the times indicated beforeexposure to Newcastle disease virus to test for induction of IFN.
4 6 8
DMSO Treatment (days)
Cell Growth & Differentiation 507
1 2 3456 12345
B � � -
��ui#{149}. �.
mentally regulated member of the family of octamer-binding proteins.
In this paper, we have shown that three variations onthe octameric sequence, Oct-IFN, Oct-Ig, and Oct-26,are all strongly bound by factors in EC cell nuclearextracts, as shown by gel retardation. Two shifted bandsof similar mobility are seen in each case. Thus, theoctameric sequence specificity in EC cells is not as strictas originally demonstrated by Staudt et a!. (19) for thelymphoid-specific Oct-2 (NF-A2), where a change of justone nucleotide prevented binding in a gel retardationassay. This is not simply a preference for AT-rich regions,as might be inferred from the binding of Oct-3 to All-TAAAATTCA (13), an AT-rich sequence homologous tothe binding site for En, a Drosophila homeobox protein.We have demonstrated that, in P19 cell nuclear extracts,the hexameric sequence (AAGTGA)4 interacts with dif-ferent proteins than the octameric sequences. Similarresults were found by MacDonald et a!. (33) when theyinvestigated the effects of tetrameric repetitive nucleo-tides of the type (GAAANN)4 on transcription. Theyfound that the sequence (GAAATG)4, containing a motifsimilar to the core of the ECIF-1 site, was able to bindOTF-i (Oct-i). This sequence did not bind IRF-i, whichbound instead to other motifs, including (GAAAGT)4.
We previously demonstrated that the binding activitiesfrom undifferentiated or fully differentiated P19 EC cellswhich interact with the human IFN-f3 promoter had al-tered mobilities in gel retardation studies (28). To deter-mine the kinetics of binding activity during the course of
differentiation, a series of nuclear extracts were madeafter various periods oftreatment of P1 9 cells with DMSOor retinoic acid. The upper band remains constant. Thestable activity of this complex along with the mobilityobserved after UV cross-linking allow us to conclude thatit is probably due to the binding of Oct-i . The proteinsin the lower band whose binding activity decreases withdifferentiation are likely members of the octamer family,one of which could be Oct-3. It has been shown that theOct-3 protein described in P19 EC cells can bind to theOct-26 probe (26). The 55 Kd DNA-protein complexpresent in P19 EC cells correlates with the size expectedfor the cloned Oct-3 (26) and is a common band ob-served between the Oct-26, the IgG, and the IFN-flprobe. Okamoto et a/. (26) observed that Oct-3 preparedby in vitro translation gave a band of faster mobility in agel retardation assay than was obtained with Oct-3 pres-ent in nuclear extracts. They suggested that this could bedue to incomplete posttranslational modification of thein vitro product. However, the same result could beobtained if the slower complex obtained with nuclearextracts is comprised of more than one component (oneof which is Oct-3), while the smaller complex containingonly in vitro translated Oct-3 migrates more quickly. OurUv cross-linking results indicate that this may be a pos-sibility, since the lower complex detected by gel retar-dation with the Oct-26 probe was shown to be corn-posed of multiple proteins.
Octamer-binding proteins have the potential to act asrepressors or as activators of transcription, depending onthe context (20, 2i, 23, 26, 34, 35). The activation orrepression of specific genes by horneotic proteins thatbind similar sequences may be directed by differentialexpression of these proteins during development (36,37). Another model for the action of octamer-bindingproteins postulates that the activity of such proteins isexerted in concert with other proteins which conferspecificity. Such protein-protein interactions have beendemonstrated for both Oct-i and Oct-2 (38-43). Thesemechanisms may not be mutually exclusive, and bothmay come into play during the differentiation of EC cells.
Our results show that there is a correlation betweenthe loss of the binding activity at the octarner ECIF-ibinding site and the inducibility of the IFN during thedifferentiation process. This observation suggests thepresence of an inhibitory activity in undifferentiated P19cells which is directly involved in the lack of competencefor IFN induction. This is confirmed by the results of invitro transcription: the EC cell nuclear extract contains anactivity which represses transcription of the IFN-$ gene(Fig. 6). This activity is dependent on the presence ofnucleotides -85 to -134. The only protein binding sitein this region is the ECIF-1 (octarner) site (Ref. 28 andthis paper). The repressing activity is absent from nuclearextracts from differentiated derivatives of P1 9 cells. Thus,the lack of inducibility of the IFN-fl gene in EC cellsappears to be due at least in part to a dominant repressionof transcription. In differentiated cells, the repressingactivity is absent, allowing the action of positive factors(14).
When these studies were initiated, the 5’ sequence ofthe murine INF-/3 gene had not been reported. Fujita eta/. (13) indicated that the homology between the pro-moters of the human and mouse genes was extremelyhigh. Now that the mouse sequence has become avail-
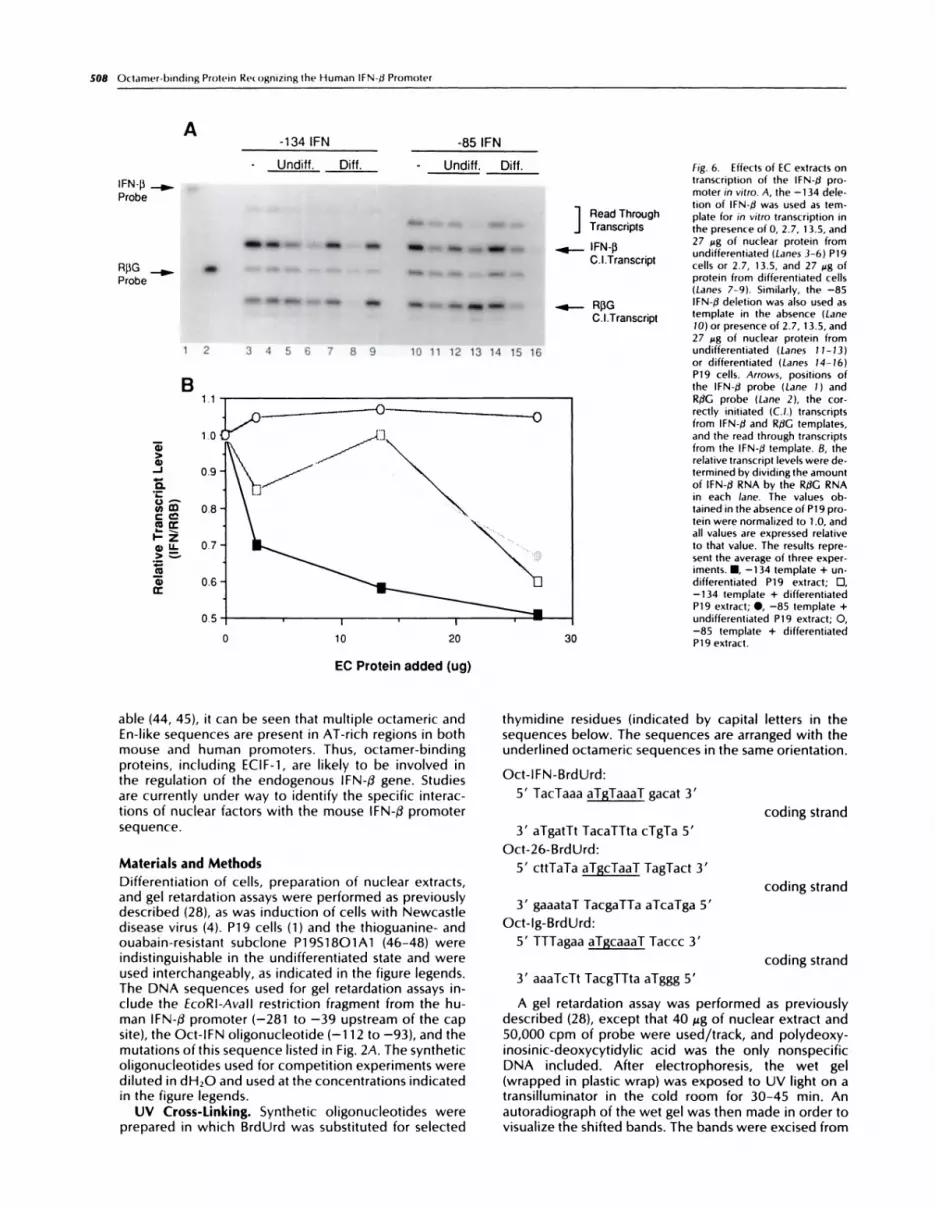
-1341FN -85IFNA
- �Undiff. Diff. - Undiff. Diff.
IFN-f� ...�..
Probe
� #{176}.. �
�- � .� � - � �
R)�G _� * .� .. � � � . �
Probe
� � � � � � � � �
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
B
6,>6)
-J
0.
U.-.
(n�
�U.
>
6)
0 10 20
508 Octamer-binding Protein Recognizing the Human IFN-d Promoter
EC Protein added (ug)
able (44, 45), it can be seen that multiple octameric andEn-like sequences are present in AT-rich regions in bothmouse and human promoters. Thus, octamer-bindingproteins, including ECIF-1, are likely to be involved inthe regulation of the endogenous IFN-fl gene. Studiesare currently under way to identify the specific interac-tions of nuclear factors with the mouse IFN-f3 promotersequence.
Materials and Methods
Differentiation of cells, preparation of nuclear extracts,and gel retardation assays were performed as previouslydescribed (28), as was induction of cells with Newcastledisease virus (4). P19 cells (1) and the thioguanine- andouabain-resistant subclone P19S18O1A1 (46-48) wereindistinguishable in the undifferentiated state and wereused interchangeably, as indicated in the figure legends.The DNA sequences used for gel retardation assays in-dude the EcoRl-Avall restriction fragment from the hu-man IFN-(3 promoter (-28i to -39 upstream of the capsite), the Oct-IFN oligonucleotide (-i12 to -93), and themutations of this sequence listed in Fig. 2A. The syntheticoligonucleotides used for competition experiments werediluted in dH2O and used at the concentrations indicatedin the figure legends.
Uv Cross-Linking. Synthetic oligonucleotides wereprepared in which BrdUrd was substituted for selected
__________ fig. 6. Effects of [C extracts ontranscription of the IFN-d pro-moter in vitro. A, the -134 dele-
tion of lFN-�J was used as tem-1 Read Through plate for in vitro transcription inJ Transcripts the presence of 0, 2.7, 13.5, and
IFN.R 27 �g of nuclear protein from.�.- P � undifferentiated (Lanes 3-6) P19
C.l.Transcript cells or 2.7, 13.5, and 27 �g of
protein from differentiated cells(Lanes 7-9). Similarly, the -85
..#{248}- R�G IFN-fl deletion was also used as
C.I.Transcript template in the absence (Lane10) or presence of 2.7, 13.5, and27 Mg of nuclear protein fromundifferentiated (Lanes 11-13)
or differentiated (Lanes 14-16)P19 cells. Arrows, positions ofthe lFN-� probe (Lane 1) andRI3G probe (Lane 2), the cor-rectly initiated (C.!.) transcriptsfrom lFN-�l and RIIG templates,
and the read through transcriptsfrom the IFN-f3 template. B, therelative transcript levels were de-termined by dividing the amount
of IFN-fl RNA by the R$G RNAin each lane. The values ob-tamed in the absence of P1 9 pro-
tein were normalized to 1 .0, and
all values are expressed relativeto that value. The results repre-sent the average of three exper-iments. #{149},-134 template + un-
differentiated P19 extract; 0,-134 template + differentiatedP19 extract; S, -85 template +
undifferentiated P19 extract; 0,-85 template + differentiated
30 P19 extract.
thymidine residues (indicated by capital letters in thesequences below. The sequences are arranged with theunderlined octameric sequences in the same orientation.
Oct-I FN-Brd Urd:
5’ TacTaaa aTgTaaal gacat 3’
3’ algatlt TacalTta clgla 5’
Oct-26-BrdUrd:
5’ cttlala algclaaT Taglact 3’
3’ gaaatal Tacgalla alcalga 5’
Oct-lg-BrdUrd:
5 , TTTagaa algcaaal Taccc 3’
3’ aaalclt Tacgllta aTggg 5’
coding strand
coding strand
coding strand
A gel retardation assay was performed as previouslydescribed (28), except that 40 �.zg of nuclear extract and50,000 cpm of probe were used/track, and polydeoxy-inosinic-deoxycytidylic acid was the only nonspecificDNA included. After electrophoresis, the wet gel(wrapped in plastic wrap) was exposed to UV light on atransilluminator in the cold room for 30-45 mm. Anautoradiograph of the wet gel was then made in order tovisualize the shifted bands. The bands were excised from
- 31K -
- 26K-
the gel, and the gel slices were heated to 65#{176}Cfor 10mm in sample buffer. The slices were rinsed twice withSDS-Tris (i% SDS-i.25 M Tris-HCI, pH 6.8) and posi-tioned on the upper edge of a glass plate with the aid ofagarose [1% (w/v) in SDS-Tris]. The gel apparatus wasassembled, and a 7.5% SDS-polyacrylamide gel was thenpoured, followed by a 3% stacking gel. The space be-tween the stacking gel and the gel slice was filled withthe same agarose solution used above. Wells for molec-ular weight standards were included on either side of thegel slice. The gel was run under standard conditions untilthe bromophenol blue tracker dye reached the bottomof the gel.
In Vitro Transcription. Deletion mutants of the IFN-f3gene were created using the Erase-a-base system (Pro-mega) and starting with the plasmid 171318 into whichthe i.8-kilobase EcoRlfragmentofthe human IFN-flgenehad been inserted. The -134 and -85 IFN-f3 templateswere used in an in vitro reconstituted transcription assayas previously described (29, 49). Transcription reactions(50 �I) containing 2 zg/jzl of uninduced HeLa 53 nuclearextracts were preincubated with 1 �zg of polydeoxyino-sinic-deoxycytidylic acid for 10 mm on ice; differentamounts (2.7-27 �zg) of nuclear extracts from undiffer-entiated or differentiated P19 cells were added, and thetranscription reactions were initiated by the addition of500 ng of IFN-f3 DNA template, 250 ng of rabbit fl-globin
Cell Growth & Differentiation 509
A123456
:i band I
:i band II
DNA, and all four ribonucleoside triphosphates (iOO zM)
(Pharmacia). After i h at 30#{176}C,RNA was harvested, andcorrectly initiated transcription was assessed by Si map-ping using the 5’ end-labelled 922-bp Bglll-EcoRI probe.Rabbit $-globin RNA was measured using a 575-bpBamHl-Pstl R/3G probe. Autoradiographs of the exposedfilm were scanned on the LKB Ultrascan XL laserdensitometer.
Acknowledgments
123456 123456
We wish to thank Diane Forget and Alain Staeyart for technical assistanceand Roger Duclos for photography.
B
.-- - 97K-
- 70K-
p. �
, -43K- �
Fig. 7. A, band retardation assay with BrdUrd-substituted probes. Lanes1 and 4, ECIF-BrdUrd; Lanes 2 and 5, Oct-lg-BrdUrd; Lanes 3 and 6, Oct-26-BrdUrd. Lanes 1-3, nuclear extracts from undifferentiated P19 cells;Lanes 4-6, nuclear extracts from undifferentiated P1951801A1 cells.Bands I and II, marked by brackets, were cut out of the gel after UV
cross-linking and were resolved on 7.5% SDS-polyacrylamide gels, seenin B and C, respectively. Positions of molecular weight markers are
indicated between the SDS gels.
References
1. McBurney, M. W., and Rogers, B. J. Isolation of male embryoniccarcinoma cells and their chromosome replication patterns. Dev. Biol.,
89: 503-508, 1982.
2. Martin, G. R. Teratocarcinomas and mammalian embryogenesis. Sci-ence (Washington DC), 209: 768-776, 1980.
3. Barlow, D. P., Randle, J. R., and Burke, D. C. Interferon synthesis in
the early post-implantation mouse embryo. Differentiation, 27: 229-235,
1984.
4. Skup, D., Haggarty, A., Paterno, G. D., Ponton, A., Daigneault, L., and
Belhumeur, P. Regulation and role of IFN gene expression in differentiat-ing embryonal carcinoma cells. In: Y. Kawade and S. Kobayashi (eds.),
Biology of the Interferon System 1988, pp. 413-419. Kyoto: H. Kirchner
and H. Schellekens, 1989.C � Burke, D. C., Graham, C. F., and Lehman, j. M. Appearance ofinterferon inducibility and sensitivity during differentiation of murine
teratocarcinoma cells in vitro. Cell, 13: 243-248, 1978.
6. Goodbourn, S., Burstein, H., and Maniatis, T. The human interferonenhancer is under negative control. Cell, 45: 601-610, 1986.
7. Goodbourn, S., and Maniatis, T. Overlapping positive and negativeregulatory domains of the human fl-interferon gene. Proc. NatI. Acad.Sci. USA, 85: 1447-1451, 1988.
8. Zinn, K., DiMaio, D., and Maniatis, T. Identification of two distinct
regulatory regions adjacent to the human �3-interferon gene. Cell, 34:
865-879, 1983.
9. Fujita, T., Sakakibara, J., Sudo, Y., Miyamoto, M., Kimura, Y., andTaniguchi, T. Evidence for a nuclear factor(s), IRF-1, mediating induction
and silencing properties to human IFNfl gene regulatory elements. EMBO
J., 7: 3397-3405, 1988.
10. Visvanathan, K. V., and Goodbourn, S. Double-stranded RNA acti-
yates binding of NFsB to an inducible element in the human fl-interferon
promoter. EMBO J., 8: 1 129-1 138, 1989.
1 1 . Fan, C-M., and Maniatis, T. A DNA-binding protein containing twowidely separated zinc finger motifs that recognize the same DNA se-quence. Genes & Dev., 4: 29-42, 1990.
12. Harada, H., Fujita, T., Miyamoto, M., Kimura, Y., Maruyama, M.,
Furia, A., Miyata, T., and Taniguchi, T. Structurally similar but functionally
distinct factors, IRF1 and IRF2, bind to the same regulatory elements of
IFN and IFN-inducible genes. Cell, 58: 729-739, 1989.
13. Fujita, T., Ohno, S., Yasumitsu, H., and Taniguchi, T. Delimitation
and properties of DNA sequences required for the regulated expression
of human interferon fi gene. Cell, 41: 489-496, 1985.
14. Harada, H., Willison, K., Sakakibara, J., Miyamoto, M., Fujita, T., and
Taniguchi, T. Absence of type I IFN system in EC cells: transcriptional
activator (IRF-1) and repressor (IRF-2) genes are developmentally regu-lated. Cell, 63: 303-312, 1990.
15. Levine, M., and Hoey, T. Homeobox proteins as sequence-specific
transcription factors. Cell, 55: 537-540, 1988.
16. Scott, M. P., Tamkun, J. W., and Hartzell, C. W., Ill. The structureand function of the homeodomain. Biochim. Biophys. Acta, 989: 25-48,
1989.
17. Fletcher, C., Heintz, N., and Roeder, R. G. Purification and charac-
terization of OTF-1, a transcription factor regulating cell cycle expressionofa human histone H2B gene. Cell, 51: 773-781, 1987.
18. Scheidereit, C., Heguy, A., and Roeder, R. G. Identification and
purification of a human lymphoid-specific octamer-binding protein (OTF-2) that activates transcription of an immunoglobuhin promoter in vitro.
Cell, 51: 783-793, 1987.

510 Octamer-binding Protein Recognizing the Human IFN-fl Promoter
19. Staudt, L. M., Singh, H., Sen. R., Wirth, T., Sharp, P. A., and Baltimore,D. A lymphoid-specific protein binding to the octamer motif of immu-noglobulin genes. Nature (Lond.), 323: 640-643, 1986.
20. Lenardo, M. J., Staudt, L., Robbins, P., Kuang, A., Mulligan, R. C.,and Baltimore, D. Repression of the IgH enhancer in teratocarcinomacells associated with a novel octamer factor. Science (Washington DC),243: 544-546, 1989.
21. Rosner, M. H., Vigano, M. A., Ozato, K., Timmons, P. M., Poirier, F.,Rigby, P. W. J., and Staudt, L. M. A POU-domain transcription factor inearly stem cells and germ cells of the mammalian embryo. Nature (Lond.),345: 686-692, 1990.
22. Sch#{246}ler,H. R., Hatzopoulos, A. K., Balling, R., Suzuki, N., and Gruss,P. A family of octamer-specific proteins present during mouse embryo-genesis: evidence for germhine-specific expression of an Oct factor.EMBO J., 8: 2543-2550, 1989.
23. Sch#{246}ler,H. R., Balling, R., Hatzopoulos, A. K., Suzuki, N., and Gruss,P. Octamer binding proteins confer transcriptional activity in nearly
mouse embryogenesis. EMBO J., 8: 2551-2557, 1989.
24. Sch#{246}ler,H. R., Dressier, G. R., Balling, R., Rohdewohld, H., andGruss, P. Oct4: a germline-specific transcription factor mapping to themouse t-complex. EMBO J., 9: 2185-2195, 1990.
25. Sch#{246}ler,H. R., Ruppert, S., Suzuki, N., dhowdhury, K., and Gruss,
P. New type of POU domain in germ line-specific protein Oct-4. Nature(lond.), 344: 435-439, 1990.
26. Okamoto, K., Okazawa, H., Okuda, A., Sakai, M., Murumatsu, M.,and Hamada, H. A novel octamer binding transcription factor is differ-entially expressed in mouse embryonic cells. Cell, 60: 461 -472, 1990.
27. Bhat, K., McBurney, M. W., and Hamada, H. Functional cloning ofmouse chromosomal loci specifically active in embryonal carcinoma stemcells. Mol. Cell. Biol., 8: 3251-3259, 1988.
28. Haggarty, A., Ponton, A., Paterno, G. D., Daigneault, L., and Skup,D. An embryonic DNA-binding protein specific for a region ofthe humanIFN�, promoter. Nucleic Acids Res., 16: 10575-10592, 1988.
29. Cohen, 1., Lacoste, J., Parniak, M., Daigneault, L., Skup, D., andHiscott, J. Stimulation of interferon fi gene transcription in vitro by purifiedNF-KB and a novel TH protein. Cell Growth & Differ., 2: 323-333, 1991.
30. Sturm, R. A., Dalton, S., and Wells, J. R. E. A 100-kD HeLa celloctamer binding protein [OBP100] interacts differently with two separateoctamer-related sequences within the SV4O enhancer. Genes & Dev., 1:
1147-1160, 1987.
31. xanthoudakis, S., and Hiscott, J. UV cross-linking of distinct proteinsto the PRDII domain of the interferon-fl promoter. Biochem. Biophys.Res. dommun., 167: 1086-1093, 1990.
32. Ryseck, R. P., and Bravo, R. c-Jun, junB and junD differ in theirbinding affinities to APi and CRE consensus sequences: effect of fosproteins. Oncogene, in press, 1991.
33. MacDonald, N. J., KuhI, D., Maguire, D., N#{228}f,D., Gallant, P., Gos-wamy, A., Hug, H., BUeler, H., Chaturvedi, M., de Ia Fuente, J., Ruffner,H., Meyer, F., and Weissmann, C. Different pathways mediate virusinducibility of the human IFN#{212}1and IFN� genes. Cell, 60: 767-779, 1990.
34. Scheuermann, R. H., and Chem, U. A developmental-specific factorbinds to suppressor sites flanking the immunoglobulin heavy-chain en-hancer. Genes & Dev., 3: 1255-1266, 1989.
35. LeBowitz, J. H., Kobayashi, T., Staudt, L., Baltimore, D., and Sharp,P. A. Octamer-binding proteins from B or HeLa cells stimulate transcrip-tion ofthe immunoglobuhin heavy-chain promoter in vitro. Genes & Dev.,2: 1227-1237, 1988.
36. Jaynes, I. B., and O’Farrell, P. H. Activation and repression of tran-scription by homeodomain-containing proteins that bind a common site.Nature (Lond.), 336: 744-749, 1988.
37. Han, K., Levine, M. S., and Manley, J. L. Synergistic activation andrepression of transcription by Drosophila homeobox proteins. Cell, 56:573-583, 1989.
38. Janson, L., and Pettersson, U. Cooperative interaction between tran-scription factors Spi and OTF-1 . Proc. NatI. Acad. Sci. USA, 87: 4732-4736, 1990.
39. Stern, S., Tanaka, M., and Herr, W. The Oct-i homeodomain directsformation of a multiprotein-DNA complex with the HSV transactivatorVP16. Nature (Lond.), 341: 624-630, 1989.
40. Kristie, T. M., LeBowitz, J. H., and Sharp, P. A. The octamer-bindingproteins form multi-protein-DNA complexes with the HSV aTIF regula-tory protein. EMBO J., 8: 4229-4238, 1989.
41 . Yoza, B. K., and Roeder, R. G. Identification of a novel factor thatinteracts with an immunoglobulin heavy chain promoter and stimulatestranscription in conjunction with the lymphoid cell-specific factor OTF2.Mol. Cell. Biol., 10: 2145-2153, 1990.
42. LeBowitz, J. H., Clerc, R. G., Brenowitz, M., and Sharp, P. A. TheOct-2 protein binds cooperatively to adjacent octamer sites. Genes &Dev., 3: 1625-1638, 1989.
43. Roebuck, K. A., Szeto, D. P., Green, K. P., Fan, Q. N., and Stumph,W. E. Octamer and SPH motifs in the Ui enhancer cooperate to activatethe Ui RNA gene expression. Mol. Cell. Biol., 10: 341-352, 1990.
44. Kuga, T. Fujita, T., and Taniguchi, T. Nucleotide sequence of themouse interferon � gene. Nucleic Acids Res., 17: 3291, 1989.
45. Dirks, W., Mittnacht, S., Rentrop, M., and Hauser, H. Isolation andfunctional characterization of the murine interferon-fl promoter. J. In-terferon Res., 9: 125-133, 1989.
46. McBurney, M. W., Jones-Villeneuve, E. M. V., Edwards, M. K. S., andAnderson, P. J. Control of muscle and neuronal differentiation in acultured embryonal carcinoma cell line. Nature (Lond.), 299: 165-167,1982.
47. Edwards, M. K. S., and McBurney, M. W. The concentration ofretinoic acid determines the differentiated cell types formed by a tera-tocarcinoma cell line. Dev. Biol., 98: 187-191, 1983.
48. Edwards, M. K. S., Harris, J. F., and McBurney, M. W. Induced muscledifferentiation in an embryonal carcinoma cell line. Mol. Cell. Biol., 3:2280-2286, 1983.
49. Hiscott, J., Alper, D., Cohen, L., Leblanc, I. F., Sportza, L., Wong, A.,and Xanthoudakis, S. Induction of human interferon gene expression isassociated with a nuclear factor that interacts with the NF-,B site of theHIV enhancer. J. Virol., 63: 2557-2566, 1989.





























