Embed Size (px)
Citation preview

DINÁMICA POBLACIONAL DE LAS COMUNIDADES
DE BACTERIAS NITRIFICANTES EN UNA EDAR CON
SISTEMA CONVENCIONAL DE FANGOS ACTIVOS.
TRABAJO FINAL DE MÁSTER EN INGENIERÍA AMBIENTAL
Autora:
BARBARROJA ORTIZ, PAULA
Directores:
DR. ALONSO MOLINA, JOSÉ LUIS
DR. BORRÁS FALOMIR, LUIS
Valencia, Septiembre 2013


AGRADECIMIENTOS Especialmente a mis directores, José Luis y Luis, por compartir conmigo sus conocimientos, por sus útiles consejos y por su tiempo.
Al IIAMA por permitirme realizar este trabajo con ellos.
A Andrés por su paciencia y por su dedicación en este trabajo.
A mis compañeros de máster por el tiempo compartido.
A todo el personal del departamento de Química y Microbiología del Agua, por los cafés y por el apoyo recibido. Especialmente a Mariela por sus consejos y a Marta, Alba y Vero, porque sin vosotras no habría sido lo mismo.
A mi familia, especialmente a mi madre y a mi tío, por el apoyo que me han dado.
A mis amigos, por estar presentes a pesar de las distancias, especialmente a Maria y Ana por animarme tanto.
Y muy especialmente a Txema, por compartir conmigo los buenos momentos y los no tan buenos, has sido mi pilar y mi fuente de energía.


iii
RESUMEN La eliminación de nitrógeno es uno de los proceso clave en las estaciones depuradoras de aguas residuales debido a los problemas de eutrofización que el nitrógeno causa en las aguas receptoras. El sistema más comúnmente empleado para este cometido es la nitrificación-desnitrificación vía nitrato. El primer paso de este proceso es llevado a cabo por las bacterias oxidantes del amonio (BOA) y las bacterias oxidantes del nitrito (BON), las cuales presentan una alta sensibilidad a los factores ambientales y parámetros operacionales. En este trabajo se ha utilizado la técnica molecular de hibridación in situ con sondas marcadas con fluoróforos (FISH) junto con microscopía de epiflurorescencia y análisis de imagen para la identificación y cuantificación de la población de bacterias nitrificantes presentes en el licor mezcla una EDAR con sistema de aireación prolongada y eliminación biológica de nitrógeno. Las muestras fueron tomadas quincenalmente durante un año. Se han realizado estudios bivariantes y multivariantes para determinar que variables explican mejor la dinámica poblacional de las comunidades de bacterias nitrificantes en relación a las variables físico-químicas y operacionales de la EDAR. Para la identificación de las BOA se utilizaron las sondas Nso1225, Nmo218, NEU, Nse1472 y NmV, de las cuales las tres primeras dieron una abundancia significativa y establecieron a N.oligotropha como la especie dominante en la oxidación del amonio a nitrito. Para la identificación de BON se utilizaron la sonda NIT3 y Ntspa 662, siendo en este caso Nitrospira el género responsable de la oxidación de nitrito a nitrato. Tras la observación de la dinámica poblacional este estudio sugiere que la estabilidad en los rendimientos de nitrificación no está necesariamente ligada a una comunidad de bacterias nitrificantes constante. Los resultados del análisis estadístico revelaron que la dinámica poblacional estaba influenciada por las concentraciones de nitrógeno del afluente, la temperatura, la edad del fango y la carga másica.
ABSTRACT Nitrification is the key process in wastewater treatment plants (WWTPs) in order to reduce eutrophication of recipient waters. Nitrogen is commonly removed by oxidation of ammonium via nitrite. The first step in this process is carried out by the ammonia-oxidizing bacteria (AOB) and nitrite-oxidizing bacteria (NOB). Both are highly sensitive to environmental factors and operational parameters. By using fluorescence in situ hybridization (FISH) and epifluorescence microscopy, we followed biomass changes during 1 year in the mixed liquor of the WWTP with extended aeration and biological nitrogen removal system. In order to study the correlations between microbial community structures and environmental variables we used Canonical Correspondence Analysis (ACC). Ammonium oxidizing bacteria (AOB) were identified by using Nso1225, Nmo218, NEU y Nse147 and NmV probes. The first three ones

gave us an important abundance and revealed the dominance of N. oligotorpha in the ammonium oxidation. Nitrite oxidizing bacteria (NOB) were identified using NIT3 and Ntspa 662 probes, with Nitrospira as dominant NOB specie. This study suggests that stable nitrogen removal efficiency is not necessarily linked to a stable AOB community. Results reported that the dynamics of AOB community were associated with total nitrogen influent, mixed liquor temperature, sludge retention time (SRT) and Food-to-Microorganism ratio (F/M).
RESUM La eliminació de nitrogen és un dels processos clau en les estacions depuradores d’aigües residuals (EDAR) degut als problemes d’eutrofització que el nitrogen causa en les aigües receptores. El sistema emprat habitualment per a portar a terme aquest procés és la nitrificació-desnitrificació via nitrat. El primer pas d’aquest procés es portat a terme pels bacteris oxidants de l’amoni (BOA) i els bacteris oxidants del nitrit (BON), els quals presenten una elevada sensibilitat als factors ambientals i paràmetres operacionals. En aquest treball s’ha utilitzat la tècnica molecular d’hibridació in situ amb sondes marcades amb fluoròfors (FISH) junt a microscòpia de eplifluorescència i tractament d’imatge per a la identificació i quantificació de la població de bacteris nitrificants presents en el licor-mescla una EDAR amb sistema de ventilació prolongada i eliminació biològica de nitrogen en mostres preses quinzenalment durant un any. S’ha realitzat estudis de correlacions bivariants i multivariants per a determinar quines variables expliquen millor la dinàmica poblacional de les comunitats de bacteris nitrificants en relació a les variables fisicoquímiques i operacionals de la EDAR. Per a la identificació dels BOA s’utilitzaren les sondes Nso 1225, Nmo 218, NEU, Nse 1472 i NmV, de les quals les tres primeres donaren una abundància significativa i establiren a N. oligotropha com l’espècie dominant en l’oxidació d’amoni a nitrit. Per a la identificació de BON es va utilitzar la sonda NIT3 i Ntspa 662 i en aquest cas va ser Nitrospira el gènere responsable de l’oxidació de nitrit a nitrat. Després de l’observació de la dinàmica poblacional aquest estudi suggereix que l’estabilitat en els rendiments de nitrificació no està necessàriament lligada a una comunitat de bacteris nitrificants constant. Els resultats de l’anàlisi estadístic revelaren que la dinàmica poblacional estava estretament associada amb l’entrada de nitrogen en l’afluent, la temperatura, l’edat del fang i la càrrega màssica.

v
LISTADO DE ACRÓNIMOS
ACC Análisis de correspondencias canónicas
ACP Análisis de componentes principales
AM Análisis multivariante
Amo Enzima Amonio monooxigenasa
AOA Arqueas Oxidantes del Amonio
ARU Aguas Residuales Urbanas
BOA Bacterias oxidantes del amonio
BON Bacterias oxidantes del nitrito
DBO Demanda Biológica de Oxígeno
DQO Demanda Química de Oxígeno
EDAR Estación Depuradora de Aguas Residuales
FISH Fluorescense in situ hybridization
EF Edad del Fango
HAO Enzima Oxidorreductasa hidroxilamina
NKT Nitrógeno Kjeldhal
MBR Membrane bioreactor.
Nxr Enzima Nitrito oxidoreductasa
OD Oxígeno disuelto
TRH tiempo de retención hidráulico


vii
ÍNDICE
1. INTRODUCCIÓN ................................................................................... 1 1.1. EL CICLO DEL NITRÓGENO ...................................................................................... 4
1.1.1. El nitrógeno en las aguas residuales ................................................................................ 5 1.2. BIOQUÍMICA DEL PROCESO DE NITRIFICACIÓN ........................................................ 7 1.3. PRINCIPALES ORGANISMOS QUE INTERVIENEN EN EL PROCESO DE NITRIFICACIÓN .. 8
1.3.1. Bacterias Oxidantes del Amonio. .................................................................................... 9 1.3.2. Bacterias Oxidantes del Nitrito. .................................................................................... 11
1.4. TÉCNICAS MOLECULARES PARA LA IDENTIFICACIÓN DE COMUNIDADES
MICROBIANAS ................................................................................................................ 13 1.5. TÉCNICAS DE ANÁLISIS ESTADÍSTICO PARA MUESTRAS AMBIENTALES ................... 17
1.5.1. Análisis bivariante ...................................................................................................... 17 1.5.2. Análisis multivariante ................................................................................................. 19
2. OBJETIVOS ........................................................................................... 23
3. MATERIALES Y MÉTODOS .................................................................... 25 3.1. DESCRIPCIÓN DE LA EDAR .................................................................................. 25 3.2. TOMA DE MUESTRAS ............................................................................................. 26 3.3. VARIABLES FÍSICO-QUÍMICAS Y PARÁMETROS OPERACIONALES ............................. 27 3.4. TÉCNICA DE HIBRIDACIÓN IN SITU CON SONDAS MARCADAS CON FLUOROFOROS 28
3.4.1. Fijación de las muestras. .............................................................................................. 28 3.4.2. Tratamiento de los portaobjetos cubiertos con Teflón. ..................................................... 28 3.4.3. Aplicación de las muestras a los portaobjetos y permeabilización de las celulas. ................ 29 3.4.4. Hibridación in situ. ..................................................................................................... 29 3.4.5. Sondas utilizadas en el estudio ..................................................................................... 31
3.5. CUANTIFICACIÓN DE MICROORGANISMOS ............................................................ 32 3.6. ANÁLISIS ESTADÍSTICO ......................................................................................... 34
3.6.1. Tratamiento de datos realizado para determinar las correlaciones de Pearson y Spearman 36 3.6.2. Tratamiento de datos realizado en el análisis de componentes principales ......................... 36 3.6.3. Tratamiento de datos realizado en el análisis canonico de correspondencia ....................... 38
4. RESULTADOS ....................................................................................... 39 4.1. CAPTURA DE IMÁGENES DE LAS SONDAS CON SEÑAL DE HIBRIDACIÓN POSITIVA ... 39 4.2. CUANTIFICACIÓN DE LA SEÑAL DE HIBRIDACIÓN MEDIANTE TRATAMIENTO DE
IMAGEN ......................................................................................................................... 43 4.3. ANÁLISIS ESTADÍSTICO ENTRE LAS SEÑALES OBTENIDAS PARA LAS BACTERIAS BOA
Y BON Y LOS PARÁMETROS OPERACIONALES DE LA EDAR ............................................ 46 4.3.1. Variables físico-químicas y operacionales utilizadas en el análisis estadístico .................... 46 4.3.2. Análisis bivariante. Coeficientes de correlación de Pearson y Spearman ........................... 48 4.3.3. Análisis multivariante. Análisis de componentes principales ........................................... 51 4.3.4. Análisis de correspondencias canonicas ......................................................................... 53

5. DISCUSIÓN .......................................................................................... 58 5.1. DINÁMICA POBLACIONAL ..................................................................................... 58 5.1. RENDIMIENTOS DE ELIMINACIÓN .......................................................................... 61 5.2. VARIABLES DEL AFLUENTE. .................................................................................. 61 5.3. VARIABLES OPERACIONALES. ................................................................................ 62 5.4. VARIABLES DEL LICOR MEZCLA ............................................................................. 65
6. CONCLUSIONES ................................................................................... 66
7. REFERENCIAS ...................................................................................... 67

ix
ÍNDICE DE ECUACIONES
Ecuación 1. Oxidación del amoniaco a hidroxilamina. .......................................................... 7
Ecuación 2. Oxidación de la hidroxilamina a nitrito. ............................................................. 7
Ecuación 3. Formación de agua. ........................................................................................... 7
Ecuación 4. Oxidación de amonio a nitrito. .......................................................................... 7
Ecuación 5. Oxidación de la hidroxilamina a nitrito. ............................................................. 7
Ecuación 6. Formación de agua. ........................................................................................... 7
Ecuación 7. Oxidación de la hidroxilamina a nitrito. ............................................................. 8
Ecuación 8. Reacción global de nitrificación. ........................................................................ 8
Ecuación 9. Reacción de síntesis celular. ............................................................................... 8
Ecuación 10. Reacción global ............................................................................................... 8
Ecuación 11. Calculo del coeficiente de correlación lineal de Pearson .................................. 17
Ecuación 12. Calculo del coeficiente de correlación de Spearman. ....................................... 18
Ecuación 13. Vector de componentes ACP ......................................................................... 19
Ecuación 14. Modelización realizada en el ACP ................................................................. 20


xi
ÍNDICE DE FIGURAS
Figura 1. Ciclo del Nitrógeno (Metcalf &Eddy, 2000) ............................................................ 4
Figura 2. Transformaciones del nitrógeno en el proceso de nitrificación-desnitrificación vía nitrato. (Metcalf &Eddy, 2000) ...................................................................................... 6
Figura 3. Árbol filogenético de las bacterias amino oxidantes BOA basado en la secuencia del gen 16S rRNA (Seviour y Nielsen, 2010). .................................................................... 10
Figura 4. Árbol filogenético de las bacterias nitro oxidantes BON basado en la secuencia del gen 16S rRNA (Seviour y Nielsen, 2010). .................................................................... 12
Figura 5. Árbol filogenético del gen 16S rRNA basado en los linajes principales de bacterias amonio- oxidantes conocidas (Daims et al., 2009). ........................................................ 14
Figura 6. Árbol filogenético del género Nitrosomonas inferido del análisis comparativo del gen 16S rRNA (Gieseke et al., 2000). .................................................................................. 15
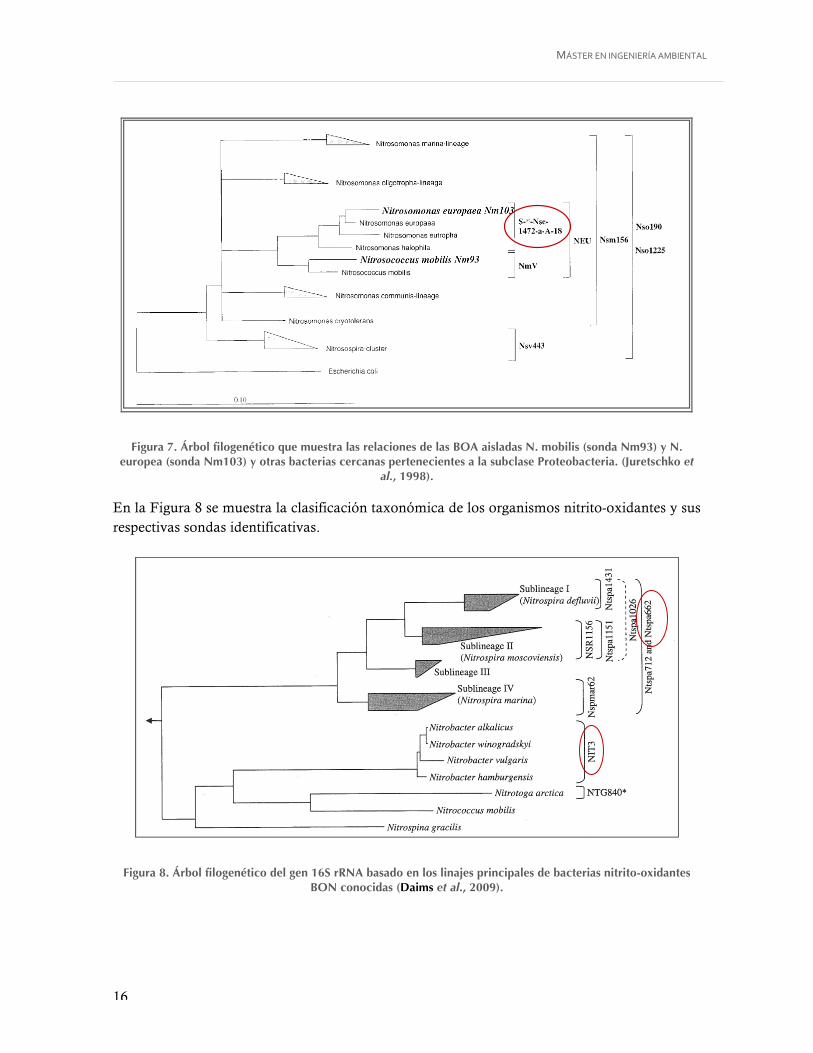
Figura 7. Árbol filogenético que muestra las relaciones de las BOA aisladas N. mobilis (sonda Nm93) y N. europea (sonda Nm103) y otras bacterias cercanas pertenecientes a la subclase Proteobacteria. (Juretschko et al., 1998). ......................................................... 16
Figura 8. Árbol filogenético del gen 16S rRNA basado en los linajes principales de bacterias nitrito-oxidantes BON conocidas (Daims et al., 2009). .................................................. 16
Figura 9. Representación esquemática de las matrices del modelo ACP. .............................. 20
Figura 10. Matriz de covarianzas ACC ............................................................................... 21
Figura 11. Esquema de depuración de la EDAR Denia-Ondara-Pedreguer (EPSAR) ............ 25
Figura 12. Sondas utilizadas para la identificación de BOA ................................................. 31
Figura 13. Sondas utilizadas para la identificación de BON ................................................. 31
Figura 14. Flujo de procesos para la cuantificación de bacterias. .......................................... 34
Figura 15. BOA. A) sonda Nso 1225, B) Mismo campo sondas EUB338mix. 600X. ............. 40
Figura 16. BOA. A) sonda NEU, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y NEU. 1000X. ............................................................................ 41
Figura 17. BOA. A) sonda Nmo 218, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y Nmo 218. 1000X. ...................................................................... 41

Figura 18. BOA. A) sonda NmV, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y NmV. 1000X. ............................................................................ 42
Figura 19. BOA. A) sonda Nse 1472, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y Nse 1472. 1000X. ...................................................................... 42
Figura 20. BOA. A) sonda Ntspa 662, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y Ntspa 662. 1000X. .......................................................... 42
Figura 21. BOA. A) sonda NIT3, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y NIT3. 1000X. ............................................................................ 43
Figura 22. Modelo de hoja Excel de resultados extraídos del software de cuantificación ....... 43
Figura 23. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables del afluente ................................................................................................... 54
Figura 24. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables del licor mezcla ............................................................................................. 55
Figura 25. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables del efluente ................................................................................................... 56
Figura 26. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables operacionales ................................................................................................ 57

xiii
ÍNDICE DE TABLAS
Tabla 1. Contaminantes de importancia en el tratamiento del agua residual ........................... 1
Tabla 2. Fraccionamiento del nitrógeno en las aguas residuales. ............................................ 5
Tabla 3. Rendimientos de eliminación en la EDAR Denia-Ondara-Pedreguer ...................... 26
Tabla 4. Esquema de la campaña de muestreo (Zornoza et al., 2010) .................................... 26
Tabla 5.Variables físico-químicos del afluente, efluente y licor mezcla en relación a la campaña de muestreo ................................................................................................................ 27
Tabla 6. Parámetros operacionales ...................................................................................... 27
Tabla 7. Solución de hibridación en función del porcentaje de formamida ............................ 29
Tabla 8. Solución de lavado en función del porcentaje de formamida ................................... 30
Tabla 9. Sondas utilizadas en el presente estudio ................................................................. 32
Tabla 10. Variables empleadas en el estudio y etiquetas ....................................................... 35
Tabla 11. Fecha de muestreos ............................................................................................. 35
Tabla 12. Variables utilizadas para realizar el análisis de componentes principales ............... 37
Tabla 13. Resumen de los resultados de las señales de hibridación ....................................... 40
Tabla 14. Promedio de las áreas en pixeles ocupadas por las BOA, desviación estándar y error de medida. .................................................................................................................. 44
Tabla 15. Promedio de las áreas en pixeles ocupadas por las BON, desviación estándar y error de medida ................................................................................................................... 45
Tabla 16. Bacterias nitrificantes expresadas como mg SSVLM / L. ...................................... 46
Tabla 17. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros físico-químicos del afluente ................................................................................................... 47
Tabla 18. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros físico-químicos del licor mezcla ............................................................................................ 47
Tabla 19. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros físico-químicos del efluente ................................................................................................... 47

Tabla 20. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros operacionales .............................................................................................................. 48
Tabla 21. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes, los rendimientos de eliminación de nitrógeno y las variables físico-químicas del afluente .... 49
Tabla 22. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes y las variables físico-químicas del licor mezcla ................................................................................... 50
Tabla 23. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes con las variables físico-químicas del efluente ............................................................................ 50
Tabla 24. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes con las variables operacionales ................................................................................................ 51
Tabla 25. Matriz de componentes rotados variables afluente ................................................ 52
Tabla 26. Matriz de componentes rotados variables licor mezcla .......................................... 52
Tabla 27. Matriz de componentes rotados variables operacionales ....................................... 53
Tabla 28. Matriz de componentes en espacio rotado variables efluente ................................. 53

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
1
1. INTRODUCCIÓN
El agua es un factor determinante para el desarrollo de la vida, además cumple importantes funciones económicas, sociales y ambientales. Si bien este no es un recurso escaso, hay que tener en cuenta que se encuentra distribuido de manera poco homogénea y solo una pequeña proporción se encuentra disponible para su uso.
El ser humano ha ido incrementando sus requerimientos de agua así como la cantidad y la carga contaminante de los vertidos a medios naturales. Esto es causa de alteraciones en la dinámica de los ecosistemas, lo que afecta a su capacidad de autodepuración y a la renovación y redistribución del agua dentro del ciclo hidrológico.
En las últimas décadas se ha observado un creciente interés por la protección de este vulnerable recurso que ha motivado el desarrollo de políticas basadas en la eficiencia, reutilización y reciclado del agua. Estas se han traducido principalmente en la aplicación de herramientas legislativas para el control de vertidos y en el desarrollo de tecnologías de tratamiento más eficientes encaminadas a la reducción de la carga contaminante de los vertidos.
Las estaciones depuradoras de aguas residuales (EDAR) son las encargadas de minimizar el impacto de estos vertidos en el medio receptor. La mayor parte de ellas emplean procesos biológicos para la degradación de la materia orgánica, eliminación de nutrientes, sólidos en suspensión y microorganismos patógenos, que son los principales compuestos contaminantes de las aguas residuales urbanas (ARU).
En la tabla que se presenta a continuación podemos ver el efecto sobre el medio ambiente de cada uno de estos contaminantes en las ARU.
Tabla 1. Contaminantes de importancia en el tratamiento del agua residual
La eliminación de nutrientes suscita un especial interés debido a los problemas de eutrofización que su vertido causa en ríos, lagos y costas. La eutrofización se puede definir como el incremento progresivo de productores primarios (algas y plantas) debido a la alteración de alguno de los factores que limitan su crecimiento: temperatura, luz y nutrientes.

MÁSTER EN INGENIERÍA AMBIENTAL
2
El crecimiento acelerado de estos organismos provoca una serie de cambios perjudiciales para los ecosistemas, los cuales se detallan a continuación (Martín, 2005):
- Aumento de la turbidez debido a mayores concentraciones de fitoplancton en el agua. Ello da lugar a una menor penetración de la luz en la columna de agua y a la posible desaparición de la vegetación de fondo por falta de radiación solar.
- Aumento de la toxicidad del agua debido a la producción de sustancias tóxicas por parte de algunas especies de fitoplancton. Esto puede tener graves consecuencias sobre la fauna piscícola, fauna terrestre que abastece de esas aguas y en los abastecimientos de poblaciones a partir de embalses eutrofizados.
- Disminución de la biodiversidad en general, tanto especies de fitoplancton como de seres superiores. Se produce una simplificación notable de los ecosistemas, haciendo a éstos más vulnerables a posibles perturbaciones futuras.
- Fluctuaciones notables en las concentraciones de oxígeno disuelto (OD) en el agua: altas durante el día y bajas por la noche debido al balance entre fotosíntesis y respiración del fitoplancton. En conjunción con épocas de alta temperatura y alta actividad de biodegradación de materia orgánica, puede dar lugar a periodos de varios días con niveles de OD extraordinariamente bajos.
- Aumento notable de pH debido a la alta tasa de consumo de CO2 por parte de fitoplancton.
El incremento de la concentración de nitrógeno en las corrientes de agua, debido al aporte derivado de la actividad antropogénica, es una de las principales causas de esta problemática en la actualidad.
Han sido varias las iniciativas legislativas comunitarias relacionadas con la protección del medio frente a la problemática de la eutrofización y la gestión más eficiente del agua:
- Directiva 2000/60/EC del Parlamento Europeo, cuyo objetivo es proteger el estado ecológico de las masas de agua continentales, aguas de transición, aguas costeras y aguas subterráneas.
- Directiva 91/271/CEE, modificada por la Directiva 98/15/CE, en la que se establecen la obligación de disponer de sistemas colectores y tratar las ARU y ARI, así como los límites de vertido de los contaminantes procedentes de EDAR. Esta directiva introduce el concepto de zona sensible a la eutrofización haciendo referencia a la importancia de eliminar nutrientes, especialmente nitrógeno y fósforo.
- Directiva 91/676/CEE para la protección de las aguas contra la contaminación producida por nitratos procedentes de fuentes agrícolas.
La Directiva 91/271/CEE fue transpuesta al ordenamiento español a través de el Real Decreto Ley 11/1995 de 20 de Diciembre, en el se definen los criterios para la clasificación de los

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
3
puntos de vertido en “zona sensible” y “zona menos sensible” de los cuales dependen los requisitos de vertido. Este Real Decreto fue desarrollado por el Real Decreto 509/1996, modificado por el Real Decreto 2116/1998, en el que se desarrolla la normativa aplicable para el control de las ARU en España.
Debido a la importancia que ha adquirido la eliminación de nitrógeno en las corrientes de agua residual, la nitrificación en las EDAR se ha convertido en uno de los procesos clave de la depuración.
Este proceso biológico, que consiste en la oxidación secuencial aerobia de amonio a nitrito y de nitrito a nitrato, depende de la presencia de poblaciones de bacterias nitrificantes quimiolitoautótrofas encargadas de catalizar dicha transformación: las bacterias oxidantes de amonio (BOA) y las bacterias oxidantes del nitrito (BON).
El lento crecimiento de las comunidades nitrificantes y la extrema sensibilidad que presentan a los factores ambientales y operacionales, entre los que se incluyen la temperatura, pH, OD, compuestos inhibidores (Posser, 1989), concentración de amonio y nitrito (Koops and Pommerening-Roser, 2001; Siripong and Rittmann, 2007), alcalinidad (Biesterfeld et al., 2003) y la disponibilidad de carbono inorgánico (Guisasola et al., 2007: Wett and Rauch 2003), hacen que el proceso de nitrificación sea un paso crítico a controlar en las estaciones depuradoras de aguas residuales.
La técnica molecular de hibridación in situ con sondas marcadas con fluoróforos (FISH) junto con la reciente aplicación de técnicas de cuantificación por tratamiento de imagen, se presentan como una de las mejores alternativas para el control y seguimiento de las poblaciones de bacterias nitrificantes debido a su rapidez y especificidad.
Aunque un gran número de estudios han investigado la dinámica poblacional de las bacterias nitrificantes en los procesos de tratamiento de aguas residuales (Daims et al., 2001a; Limpiyakorn et al., 2005; Siripong and Rittmann, 2007; Whang et al., 2009; Huang et al., 2010), la mayoría de ellos relacionan de una forma independiente la influencia de algunos de estos parámetros operacionales y físico-químicos con el rendimiento del proceso de nitrificación. Sin embargo, la eficiencia del proceso en las EDAR es el resultado de la influencia conjunta de estos parámetros, siendo los estudios que los relacionan a escala real todavía escasos (Fukushima et al., 2013; Koops et al., 2006).
Establecer las relaciones entre la comunidad de bacterias nitrificantes y las variables ambientales mediante análisis estadísticos avanzados, ayudará a esclarecer que parámetros son determinantes en el control del proceso para la mejora del rendimiento y ahorro de costes de explotación.

MÁSTER EN INGENIERÍA AMBIENTAL
4
1.1. EL CICLO DEL NITRÓGENO
La química del nitrógeno es compleja debido a los numerosos estados de oxidación en los que se puede presentar y al hecho de que el cambio en el estado de oxidación puede ser llevado a cabo por organismos vivos.
La mayor parte del nitrógeno se encuentra en la atmósfera en forma diatómica y estado gaseoso a temperatura y presión ordinaria. En su forma elemental es relativamente inerte.
El nitrógeno atmosférico es fijado a través de procesos biológicos, aunque en ocasiones también es transformado como resultado de procesos químicos espontáneos. Los organismos responsables de esta fijación son bacterias y algas (bacterias simbióticas como Rhizobium, las Cianobacterias o bien bacterias libres como Azobacter), que transforman este nitrógeno en nitrógeno inorgánico asimilable por los seres vivos (NH!!, NO!! y NO!!).
La mayor parte del nitrógeno que encontramos en el suelo está en forma orgánica debido a la descomposición y procesos metabólicos de plantas y animales. El nitrógeno orgánico pasará a nitrógeno amoniacal tras la mineralización. La materia proteica no asimilable es convertida en gran medida a amonio por la acción de las bacterias saprófitas, bajo condiciones aerobias o anaerobias.
El amonio liberado puede ser usado por las plantas para producir proteínas, el sobrante tras esta asimilación es oxidado por las bacterias nitrificantes.
Los nitratos producidos en el proceso de nitrificación pueden servir como fertilizantes para las plantas y los que se producen en exceso, pueden acabar en el agua principalmente a través de la percolación y la escorrentía.
Figura 1. Ciclo del Nitrógeno (Metcalf &Eddy, 2000)

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
5
En el medio acuático el nitrógeno se encuentra mayoritariamente en forma de nitrógeno amoniacal o nitrógeno nítrico, aunque también puede estar presente como óxido nitroso y nitrógeno orgánico incorporado a sustancias orgánicas. La cantidad de nitrógeno en las aguas ha aumentado debido al uso de fertilizantes nitrogenados empleados en la agricultura, llegando a causar la contaminación de los recursos superficiales y subterráneos. El nitrato es la forma predominante por tratarse del compuesto más estable, convirtiéndose de este modo en el contaminante más común.
Tanto en el suelo como en el agua la proporción entre estas formas del nitrógeno en sus diferentes estados de oxidación depende del proceso de nitrificación.
Bajo condiciones anóxicas los nitritos y nitratos son reducidos a través del proceso de desnitrificación a N2, que regresará a la atmósfera de este modo. En la actualidad existen otros focos de emisión de compuestos nitrogenados originados por actividades antropogénicas como la quema de combustibles fósiles y otros procesos industriales como la fabricación de abonos y amoniaco.
1.1.1. EL NITRÓGENO EN LAS AGUAS RESIDUALES
El nitrógeno en las aguas residuales procede principalmente de la metabolización de las proteínas del cuerpo humano. Fundamentalmente aparece como nitrógeno orgánico, amoniacal, nitrato y nitrito en concentraciones variables, siendo el nitrógeno amoniacal y el orgánico las formas predominantes. El sumatorio de estos cuatro compuestos da como resultado el nitrógeno total, a la suma del nitrógeno inorgánico y el amoniacal se le denomina N-Kjeldahl y a las formas de nitrito y nitrato se les denomina N-Nítrico.
Tabla 2. Fraccionamiento del nitrógeno en las aguas residuales.
El nitrógeno orgánico, que se encuentra mayoritariamente en forma de urea y proteínas, es degradado a nitrógeno amoniacal por la actividad de las bacterias amonificantes.
En disolución acuosa el nitrógeno amoniacal puede estar en forma de amoniaco (NH3) o en su forma ionizada (NH!!), siendo esta última la forma predominante en las aguas residuales debido al pH característico de las mismas.
El nitrógeno nítrico aparece mayoritariamente como nitrato debido a que el nitrito es fácilmente oxidable a nitrato y este es empleado como aceptor cuando el oxígeno es escaso, como suele ser habitual en los sistemas de alcantarillado debido a la fuerte demanda de oxígeno que ejerce la materia orgánica.

MÁSTER EN INGENIERÍA AMBIENTAL
6
En las EDAR el nitrógeno es eliminado por diferentes vías. En primer lugar el amonio es empleado como nutriente para la síntesis de material celular. Se estima entre 10% y 30% el consumo de nitrógeno para el crecimiento de la biomasa en los sistemas de fangos activos. Si existe decantación primaria esta alcanzará a eliminar entre un 5%-10% del nitrógeno en forma de nitrógeno particulado. Pero estos porcentajes de eliminación siguen siendo insuficientes para el cumplimento de los requisitos de vertido. Por ello se aplican otros procesos para aumentar el grado de eliminación.
Dentro de estos procesos, que pueden ser tanto físico-químicos, como biológicos, destaca el proceso biológico de nitrificación-desnitrificación vía nitrato, mediante el cual el amonio es oxidado a nitrato (nitrificación) y posteriormente el nitrato es reducido a nitrógeno gas y otros compuestos nitrogenados (desnitrificación).
En la Figura 2 podemos ver las transformaciones que tienen lugar en el proceso de nitrificación-desnitrificación vía nitrato en las EDAR.
Figura 2. Transformaciones del nitrógeno en el proceso de nitrificación-desnitrificación vía nitrato. (Metcalf &Eddy, 2000)

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
7
1.2. BIOQUÍMICA DEL PROCESO DE NITRIFICACIÓN
La nitrificación consiste en la oxidación secuencial del amonio a nitrato catalizada por organismos procariotas quimiolitoautótrofos (Prosser, 1989). Estos organismos emplean carbono inorgánico como fuente de carbono para la síntesis celular y nitrógeno inorgánico para la obtención de energía.
En el primer paso del proceso las BOA transforman el amonio a nitrito. Para ello son necesarias dos enzimas encargadas de catalizar el proceso; la amonio monooxigenasa (Amo) (Hollocher et al.,1981) que es la responsable de la oxidación del amonio a hidroxilamina y la oxidorreductasa hidroxilamina (HAO) (Olson and Hooper, 1983) que controla la oxidación de la hidroxilamina a nitrito.
A continuación se muestran las reacciones que tienen lugar en este proceso.
𝑁𝐻! + 𝑂! + 2𝐻! + 2𝑒!!"#
𝑁𝐻!𝑂𝐻 + 𝐻!𝑂
Ecuación 1. Oxidación del amoniaco a hidroxilamina.
𝑁𝐻!𝑂𝐻 + 𝐻!𝑂!"#
𝑁𝑂!! + 5𝐻! + 4𝑒!
Ecuación 2. Oxidación de la hidroxilamina a nitrito.
12𝑂! + 2𝐻
! + 2𝑒! → 𝐻!𝑂
Ecuación 3. Formación de agua.
La reacción global de la oxidación de amonio a nitrito puede expresarse como:
𝑁𝐻!! + 3 2𝑂! → 𝑁𝑂!! + 2𝐻! + 𝐻!𝑂
Ecuación 4. Oxidación de amonio a nitrito.
El nitrito es posteriormente oxidado a nitrato por las bacterias oxidantes del nitrito (BON) (Bock et al., 1992). Esta reacción es catalizada por la enzima nitrito oxidoreductasa (Nxr) (Bock and Wagner, 2001) para el género Nitrobacter, mientras que para el resto de BON es catalizada por la enzima denominada sistema nitrito-oxidante.
𝑁𝑂!! + 𝐻!𝑂!"#
𝑁𝑂!! + 2𝐻! + 2𝑒!
Ecuación 5. Oxidación de la hidroxilamina a nitrito.
12𝑂! + 2𝐻
! + 2𝑒! → 𝐻!𝑂
Ecuación 6. Formación de agua.
La reacción total del proceso de oxidación del nitrito a nitrato puede expresarse como:

MÁSTER EN INGENIERÍA AMBIENTAL
8
𝑁𝑂!! + 1 2𝑂! → 𝑁𝑂!!
Ecuación 7. Oxidación de la hidroxilamina a nitrito.
La reacción global del proceso de nitrificación viene dada por la expresión:
𝑁𝐻!! + 2𝑂! → 𝑁𝑂!! + 2𝐻! + 𝐻!𝑂
Ecuación 8. Reacción global de nitrificación.
En el balance total de la reacción se produce energía que es utilizada por los microorganismos para sus funciones de crecimiento y mantenimiento celular.
Como se comentaba anteriormente, una parte del nitrógeno amoniacal es empleada por los microorganismos como nutriente para la síntesis de material celular, si consideramos la fórmula química de la biomasa como C5H7NO2 esta reacción puede expresarse como:
4𝐶𝑂! + 𝐻𝐶𝑂!! + 𝑁𝐻!! + 𝐻!𝑂 → 𝐶!𝐻!𝑁𝑂! + 5𝑂!
Ecuación 9. Reacción de síntesis celular.
Con lo que la ecuación global del proceso de nitrificación quedaría de la siguiente manera:
4𝐶𝑂! + 𝐻𝐶𝑂!! + 22 𝑁𝐻!! + 37𝑂! → 𝐶!𝐻!𝑁𝑂! + 21𝑁𝑂!! + 20𝐻!𝑂 + 42𝐻!!
Ecuación 10. Reacción global
Tal y como podemos observar en la reacción global del proceso la oxidación del amonio requiere el consumo de CO2 del medio y produce H+, lo que conlleva un consumo de alcalinidad del medio.
1.3. PRINCIPALES ORGANISMOS QUE INTERVIENEN EN EL PROCESO DE
NITRIFICACIÓN
Los organismos responsables del proceso de nitrificación han sido objeto de estudio desde su descubrimiento por Winogradsky en 1890 debido a la relevancia de este proceso en el ciclo biogeoquímico del nitrógeno.
Las bacterias nitrificantes, como se ha comentado anteriormente, son organismos quimiolitoautótrofos a expensas de compuestos reducidos de nitrógeno inorgánico y respiradores aerobios por definición. Generalmente se caracterizan por ser bacterias de crecimiento lento y poseer sistemas membranosos internos complejos. Estas bacterias están ampliamente distribuidas en suelos y agua y suelen ser muy abundantes en hábitats con elevados niveles de amonio y pH alcalino. En su mayoría pertenecen al grupo de las proteobacterias.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
9
Actualmente los organismos nitrificantes conocidos se encuentran dentro de pocos linajes filogenéticos y únicamente escasas poblaciones son las que dominan habitualmente en las plantas de tratamiento de aguas residuales.
Hasta la fecha no se conoce ningún organismo capaz de realizar la oxidación completa del amonio a nitrato, aunque se ha especulado que estos organismos podrían existir (Costa et al., 2006).
1.3.1. BACTERIAS OXIDANTES DEL AMONIO.
Las BOA son las responsables de la primera etapa del proceso de nitrificación, la oxidación del amonio a nitrito.
Todos los géneros de BOA conocidos pertenecen al phylum proteobacteria, dentro del cual se distinguen dos clases, las Betaproteobacterias, cuya presencia es mayoritaria en los procesos de tratamiento de aguas residuales, y las Gammaproteobacterias, las cuales son halófilas y tienen escasa presencia en los procesos convencionales.
Las proteobacterias están ampliamente distribuidas en ambientes naturales y ambientes creados por el hombre. Inicialmente fueron subdivididas en 4 géneros Nitrosomonas, Nitrosospira, Nitrosovibro y Nitrosolobus, pero estudios posteriores basados en análisis filogenéticos del gen 16S rRNA no dieron resultados significativos que justifiquen esta diferenciación a nivel de género, por lo que se propuso incluir estos tres últimos géneros mencionados en uno mismo (Head et al., 1993).
En la mayoría de EDAR la oxidación de amonio esta asociada a Betaprotobacteria del género Nitrosomonas (Purkhold et al., 2000; Zhang et al., 2011b), aunque no se descarta que otros géneros diferentes de las Betaproteobacterias puedan contribuir al proceso de nitrificación, debido a la baja abundancia que suelen presentar (Sánchez et al., 2013).
Tal y como se observa en la Figura 3, Nitrosomonas pueden ser subdivididas en varios sublinajes filogenéticos, entre los cuales las especies más representativas en la depuración de aguas son N. europaea, N. eutropha, Nitrosococcus mobilis y N. oligotropha.
La distribución de las diferentes especies de Nitrosomonas parece depender del sistema de tratamiento y de los parámetros operacionales. Mientras que en algunos reactores domina una sola especie (Juretschko et al., 1998) otros sistemas pueden contener hasta 5 especies diferentes (Daims et al., 2001b; Gieseke et al., 2003).
La mayoría de los miembros del linaje N. europaea/N. mobilis son halotolerantes o moderadamente halófilas y tienen preferencia por las elevadas concentraciones de sustrato. Estudios en plantas piloto y a escala real sitúan a N.mobilis en reactores con elevadas concentraciones de amonio y sal (Juretschko et al., 1998), al igual que N. europaea, que aparece con altas concentraciones de amonio (Tan et al., 2008) y suelen ser dominantes en reactores que tratan aguas residuales salinas (Park et al., 2009).

MÁSTER EN INGENIERÍA AMBIENTAL
10
Figura 3. Árbol filogenético de las bacterias amino oxidantes BOA basado en la secuencia del gen 16S rRNA (Seviour y Nielsen, 2010).
Por otro lado estudios realizados en plantas piloto de cultivos puros sugieren que N. oligotropha presenta una mayor adaptación a bajas concentraciones de amonio que N. europaea. (Limpiyakorn et al., 2007; Wang et al., 2010). Recientemente, estudios en reactores MBR alimentados con concentraciones controladas de amonio han sugerido la existencia de dos poblaciones de N.oligotropha diferentes, las cuales presentan adaptaciones diferentes a la concentración de sustrato y podrían ser importantes para la nitrificación en las EDAR (Almstrand et al., 2013).
Por tanto, la concentración de amonio es un factor que repercute directamente en la composición de la comunidad de BOA, en el género Nitrosomonas la afinidad por el amonio varía entre los miembros de los diferentes linajes (Pommerening-Röser et al., 1996) pero tiende a ser relativamente similares dentro de un linaje específico.
En cultivos puros de Nitrosomonas se ha observado que crecen como células libres, regulares en forma de suspensión (Bock and Wagner 2006), mientras que en los flóculos de fangos activos y biopelículas las Nitrosomonas suelen formar agregados de células compactas casi esféricas. El tamaño de estos agregados oscila entre 10 y 50 micras. En ocasiones agregados de menor tamaño han sido observados, cuyas células presentan una organización más irregular y mayor espacio intersticial entre ellas. Normalmente no se suelen observar células individuales, aunque pueden pasar desapercibidas cuando se observan flóculos o estructuras densas.
Introducción 43
Los organismos AOB de la clase Gammaproteobacteria son halófilos que se encuentran
principalmente en ecosistemas marinos y salobres (Koop et al., 1990). No se ha observado ni
atribuido un papel significativo a estos organismos en el proceso de nitrificación que tiene lugar
en el tratamiento de aguas residuales.
Figura 1.15. Árbol filogenético hasta ahora conocido de los organismos amonioxidantes basado en la secuencia 16S rRNA (Seviour y Nielsen, 2010).
1.2.3.2 Características de los organismos nitritoxidantes (NOB)
Los organismos nitritoxidantes (NOB) son los responsables de la conversión del nitrito a nitrato,
segundo paso de la reacción de nitrificación. Esta reacción es catalizada por la enzima nitrito
oxidoreductasa (Nxr).
Desde el punto de vista filogenético los organismos NOB son un grupo más heterogéneo que las
bacterias AOB. En la Figura 1.16 se encuentra el árbol filogenético de los organismos NOB. Estos
microorganismos son procariotas quimioautótrofos, divididos en cuatro géneros Nitrobacter,
Nitrospira, Nitrococcus y Nitrospina.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
11
El género Nitrosospira, también perteneciente a clase Betaproteobacteria, es característico de la oxidación del amonio en suelos, a pesar de que también han sido detectadas en EDAR ocasionalmente. Esto puede ser debido a que Nitrosospira está mejor adaptada a bajas concentraciones de sustrato (Schramm et al., 1999), por tanto no pueden competir con Nitrosomonas en las EDAR, donde las concentraciones de amonio suelen ser más elevadas que en los suelos.
Representantes de la clase Gammaproteobacterias son Nitrosococcus halophilus y Nitrosococcus oceani que, tal y como se comentaba anteriormente, debido a sus altos requerimientos de sal son habituales en hábitats marinos junto con Nitrosomonas marina perteneciente a la clase Betaproteobacteria, a pesar de que representantes de esta especie sí han sido encontrados en ambientes salobres como en un filtro percolador de un sistema de acuicultura marina (Foesel et al., 2007).
Arqueas oxidantes del amonio
Tal y como se comentaba anteriormente, se ha sugerido que las BOA no son las únicas responsables de la oxidación del amonio en las EDAR.
Han sido numerosos los estudios acerca de la presencia de organismos del dominio Archaea los cuales son capaces de realizar la oxidación autotrófica del amonio en suelos y océanos (Schleper et al., 2005), confirmándose finalmente que estos organismos juegan un papel importante en la nitrificación en ecosistemas marinos y terrestres (Beman et al., 2008; Leininger et al., 2006; Wucher et al., 2006).
Tras este descubrimiento el punto de mira se sitúa ahora sobre el papel de las arqueas oxidantes del amonio (AOA) en los sistemas de tratamiento de aguas residuales.
AOA del phylum Crenarcheota y Thaumachaeota (Spang et al., 2010) han sido detectadas en EDAR (Parck et al., 2006; Wells et al.,2009) y en EDAR de aguas residuales industriales (Mussmann et al., 2011). A pesar de estos hallazgos todavía no está claro si estos organismos crecen exclusivamente a expensas de la oxidación autotrófica del amonio y su papel en la nitrificación en los sistemas biológicos de tratamiento de aguas residuales no ha sido esclarecido todavía ( Limpiyakorn et al., 2011; Zhang et al., 2009).
1.3.2. BACTERIAS OXIDANTES DEL NITRITO.
El grupo de BON es un grupo más diverso filogenéticamente que el de las BOA, tal y como se refleja en la Figura 4.
La mayoría de los linajes de BON son altamente versátiles y se encuentran ampliamente distribuidas en la naturaleza (ecosistemas marinos, terrestres y artificiales como las EDAR). A menudo en las EDAR, tanto en flóculos como en biopelículas, encontramos las BOA y BON

MÁSTER EN INGENIERÍA AMBIENTAL
12
en una misma área, lo que refleja la simbiosis de ambos grupos funcionales (Maixner et al., 2006).
Las especies descritas, a excepción de Nitrospira, Nitrospina y Nitrolancetus, se encuentran dentro del phylum Proteobacteria.
Figura 4. Árbol filogenético de las bacterias nitro oxidantes BON basado en la secuencia del gen 16S rRNA (Seviour y Nielsen, 2010).
Nitrobacter, dentro de las Alphaproteobacterias, ha sido detectada tanto en hábitats acuáticos
como terrestres, algunas de las subespecies descritas han sido N. winogradskyi, N. hamburgensis, N. vulgaris y N. alkalicus. Se había considerado este grupo como predominante en los sistemas de tratamiento de aguas residuales (Henze et al., 1997), hasta que estudios posteriores en plantas a escala real basados en técnicas moleculares como FISH y otras técnicas basadas en métodos no dependientes de cultivo confirmaron que Nitrospira era la especie clave en este proceso (Juretscho et al., 1998), una a excepción pueden ser reactores que tratan elevadas concentraciones de nitrito, en los que Nitrobacter puede alcanzar una abundancia relativamente alta (Daims et al., 2001a). Esto es debido a que Nitrobacter presenta mejor adaptación a altas concentraciones de NO2 mientras que Nitrospira se adapta mejor a las bajas concentraciones. (Schramm et al., 1999).
Dentro de phylum Proteobacteria aunque asociadas a ambientes marinos, encontramos los géneros Nitrococcus (N. mobilis) en la clase Gammaproteobacteria y Nitrotoga (N. artica) en la clase Betaproteobacteria (Alawi et al., 2007). El género Nitropina fue asignado provisionalmente dentro de la clase Deltaproteobacteria (Teske et al., 1994), aunque esta afiliación fue cuestionada debido a su dificultad de clasificarla únicamente con análisis filogenéticos basados en el gen 16s rRNA
44 Introducción
Figura 1.16. Árbol filogenético hasta ahora conocido de los organismos nitritoxidantes basado en la secuencia 16S rRNA (Seviour y Nielsen, 2010).
El género Nitrobacter perteneciente a la clase Alphaproteobacteria contiene las especies N.
winogradskyi, N. hamburgensis, N. vulgaris y N. alkalicus (Seviour y Nielsen, 2010). Estas especies
se han encontrado en diferentes ecosistemas acuáticos y terrestres, incluyendo sedimentos de
lagos de alta alcalinidad (Sorokin et al., 1998). Las especies Nitrosococcus mobilis
(Gammaproteobacteria) y Nitrospina gracilis (Deltaproteobacteria) se encuentran relacionadas
con ecosistemas marinos.
Por otro lado, el género Nitrospira constituye el grupo más diverso de organismos NOB conocido.
Este grupo sólo contiene dos especies descritas N. marina (Watson et al., 1986) y N. moscoviensis
(Ehrich et al., 1995). Sin embargo, se ha encontrado gran diversidad de secuencias de 16 rRNA de
organismos pertenecientes al género Nitrospira aún no caracterizada en diversas muestras de
suelo, sedimento, agua dulce y agua de mar (Daims et al., 2001a). Dentro de la familia Nitrospirae,
el género Nitrospira ha sido dividido en cuatro subgrupos filogenéticos I-IV (Daims et al., 2001a).
Las especies del género Nitrobacter pueden ser enriquecidas y desarrolladas mediante incubación
en sistemas de fangos activados y de soporte sólido. De hecho, estas especies fueron
consideradas como los organismos NOB dominantes en los sistemas de tratamiento de agua
residual. No obstante, la aplicación de técnicas más especificas para la detección microbiológica,

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
13
(Schloss and Handelsman, 2004). Estudios posteriores proponen la inclusión de este género dentro de un nuevo phylum provisionalmente denominado Nitrospinae (Lucker et al., 2013). Esta bacteria parece ser la especie dominante en la oxidación del nitrito en los ecosistemas marinos (Lucker et al., 2013). También aparece asociada a ambientes radioactivos (Weidler et al., 2007).
Tal y como se comentaba anteriormente el género Nitrospira, no está relacionado con el resto de proteobacterias, sino que se engloba dentro del phylum Nitrospirae (Daims et al., 2001a) y es clave en la oxidación del nitrito a nitrato en las EDAR. El género Nitrospira es diverso y cuenta por lo menos con cuatro sublinajes filogenéticos diferentes (Daims et al., 2001a). Han sido descritas como bacterias de difícil cultivo y únicamente pocas especies han podido ser cultivadas aisladamente. La primera especie descrita fue N. marina aislada del agua del mar (Watson et al., 1986), seguida por N. moscoviensis, la cual fue aislada a partir de un tubo de hierro de un sistema de calefacción en Moscú (Ehrich et al., 1995) y se asocia a aguas dulces.
En las EDAR Nitrospira forma agregados celulares con forma esférica o irregular, que contienen cientos o miles de células. El diámetro de estos agregados va desde 10 a 100 micras, aunque en ocasiones pueden ser encontrados agregados de mayor tamaño, los cuales habitualmente contienen estrechos "canales" y grandes cavidades (Daims, 2009)
Recientes investigaciones han descrito un nuevo género, Nitrolancetus (N. holandicus), que en contraste con el resto de BON conocidas pertenece al phylum Chloroflexi (Sorokin et al., 2012).
1.4. TÉCNICAS MOLECULARES PARA LA IDENTIFICACIÓN DE
COMUNIDADES MICROBIANAS
Los fangos activos contienen una compleja población microbiana, que en la mayoría de ocasiones no es posible caracterizar mediante métodos dependientes de cultivo (Wagner et al., 1993). Es por ello que a día de hoy se emplean gran variedad de técnicas no basadas en cultivo para el estudio de las comunidades bacterianas presentes en estos sistemas como FISH, FISH-MAR, PCR, etc.
Entre estas destaca la técnica molecular FISH la cual permite la detección e identificación de microorganismos en comunidades complejas con alto grado de especificidad y aporta información, que otros medios no pueden ofrecer, acerca de la morfología, cantidad y distribución espacial en el medio.
La técnica FISH está basada en la hibridación DNA/RNA de una región del gen 16S o 23S. Para ello emplea secuencias de oligonucleótidos que se unirán específicamente formando un hibrido con el rRNA de los organismos diana bajo condiciones específicas.
Las secuencias de DNA están marcadas con sustancias fluorescentes llamadas fluoróforos, que permiten la detección de las células diana mediante el uso de un microscopio de epifluorescencia.

MÁSTER EN INGENIERÍA AMBIENTAL
14
Las sondas FISH ideales deben ofrecer una alta sensibilidad y especificidad. La sensibilidad se refiere al nivel de fluorescencia suficiente para distinguir la célula hibridada de la fluorescencia de fondo, mientras que por especificidad se entiende la capacidad de distinguir las células diana de las que no lo son y que podrían dar una señal de hibridación falsa.
Las condiciones de hibridación son factores clave para que se mantengan los dos requisitos anteriormente mencionados. Un parámetro fundamental es la concentración de formamida, la cual permite disminuir la temperatura de desnaturalización del DNA manteniendo la morfología celular y haciendo la hibridación específica, aunque compromete la señal de la sonda de hibridación.
La sondas pueden ser diseñadas en función de la especificidad deseada, ajustándose a los diferentes niveles taxonómicos.
A continuación se presenta una relación de las sondas más comunes empleadas para la identificación de bacterias nitrificantes presentes en los sistemas de tratamiento de aguas residuales, marcadas en rojo aparecen las sondas utilizadas en el presente estudio.
Figura 5. Árbol filogenético del gen 16S rRNA basado en los linajes principales de bacterias amonio- oxidantes conocidas (Daims et al., 2009).
Liz Avendaño Universidad Politécnica de Valencia
21
protocolos. La formamida disminuye la temperatura de unión de las sondas mediante el
debilitamiento de los puentes de hidrógeno. Este compuesto disminuye la temperatura de fusión
de los híbridos DNA-RNA en 0,72 ºC por cada 1% de formamida utilizada y permite realizar la
hibridación entre los 30-50 ºC (Alonso et al, 2009).
La clasificación taxonómica de los organismos amonio-oxidantes y sus respectivas sondas
utilizadas en la técnica FISH se encuentra en la Figura 6 a continuación.
Figura 6. Árbol filogenético de 16S rARN basado en los linajes principales de bacterias amonio-
oxidantes conocidas (Daims t al, 2009).
Nmo218
Nse
1472

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
15
A continuación podemos ver los árboles filogenéticos más específicos donde se incluyen diferentes sondas empleadas en el presente estudio con sus respectivas subespecies. La sonda Nmo218 identifica las subclases que se observan en la Figura 6 que se presenta a continuación.
Figura 6. Árbol filogenético del género Nitrosomonas inferido del análisis comparativo del gen 16S rRNA (Gieseke et al., 2000).
En la Figura 7 se presentan las subespecies identificadas por la sonda Nse1472.
Liz Avendaño Universidad Politécnica de Valencia
22
Por otra parte, a continuación se presentan otros árboles filogenéticos más específicos donde se
incluyen algunas sondas con sus respectivas subespecies identificadas que fueron utilizadas en
este estudio.
La sonda Nmo218 utilizada en este estudio identifica las subclases que se observan en la Figura
7.
Figura 7. Árbol filogenético del genero Nitrosomonas inferido del análisis comparativo del gen 16S
rDNA (Gieseke et al, 2000).
Otra sonda utilizada en este estudio es la Nse1472, a continuación se presentan las subespecies
identificadas por esta sonda.
MÁSTER EN INGENIERÍA AMBIENTAL
16
Figura 7. Árbol filogenético que muestra las relaciones de las BOA aisladas N. mobilis (sonda Nm93) y N. europea (sonda Nm103) y otras bacterias cercanas pertenecientes a la subclase Proteobacteria. (Juretschko et
al., 1998).
En la Figura 8 se muestra la clasificación taxonómica de los organismos nitrito-oxidantes y sus respectivas sondas identificativas.
Figura 8. Árbol filogenético del gen 16S rRNA basado en los linajes principales de bacterias nitrito-oxidantes BON conocidas (Daims et al., 2009).
Liz Avendaño Universidad Politécnica de Valencia
23
Figura 8. Árbol filogenético que muestra las relaciones de las AOB aisladas N. mobilis Nm93 y N. europea
Nm103 y otras bacterias cercanas pertenecientes a la subclase Proteobacteria. (Juretschko et al, 1998).
La clasificación taxonómica de los organismos nitrito-oxidantes y sus respectivas sondas
identificativas correspondientes a la técnica FISH se encuentran en la Figura 9.
Figura 9. Árbol filogenético de 16S ARNr basado en los linajes principales de bacterias nitrito-oxidantes
conocidas (Daims et al, 2009).
Liz Avendaño Universidad Politécnica de Valencia
23
Figura 8. Árbol filogenético que muestra las relaciones de las AOB aisladas N. mobilis Nm93 y N. europea
Nm103 y otras bacterias cercanas pertenecientes a la subclase Proteobacteria. (Juretschko et al, 1998).
La clasificación taxonómica de los organismos nitrito-oxidantes y sus respectivas sondas
identificativas correspondientes a la técnica FISH se encuentran en la Figura 9.
Figura 9. Árbol filogenético de 16S ARNr basado en los linajes principales de bacterias nitrito-oxidantes
conocidas (Daims et al, 2009).

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
17
La técnica molecular FISH junto con las novedosas técnicas de cuantificación por tratamiento de imagen, se presentan como una de las mejores alternativas para el control y seguimiento de las poblaciones de bacterias nitrificantes debido a su rapidez y especificidad.
1.5. TÉCNICAS DE ANÁLISIS ESTADÍSTICO PARA MUESTRAS
AMBIENTALES
1.5.1. ANÁLISIS BIVARIANTE
Las técnicas de análisis bivariante expresan el grado de relación entre dos variables. Pueden considerarse, en algunos supuestos, como casos especiales o simplificados de algunas técnicas de análisis multivariante. Por ello es recomendable realizar una exploración previa mediante este tipo de análisis antes de abordar la aplicación de cualquier técnica multivariante.
Las técnicas de correlación son usadas con frecuencia para resumir la fuerza de la asociación entre dos variables métricas.
Coeficiente de correlación lineal de Pearson.
El coeficiente de correlación de Pearson es un coeficiente estadístico que se suele emplear para medir la intensidad de la relación entre dos variables linealmente relacionadas X e Y, y es calculable siempre que ambas variables se distribuyan normalmente. Se calcula con la fórmula expresada en la Ecuación 11.
r x y y x y yi
n
i
i
n
i
i
n
==
−
=
−
=∑ ∑ ∑( x)( ) / ( x) ² ( )²i i - -
1 1 1
𝑋:𝑚𝑒𝑑𝑖𝑎 𝑑𝑒 𝑋
𝑌:𝑚𝑒𝑑𝑖𝑎 𝑑𝑒 𝑌
Ecuación 11. Calculo del coeficiente de correlación lineal de Pearson
El coeficiente de correlación de Pearson está comprendido entre -1 y +1, donde los valores positivos indican una dependencia lineal creciente y los valores negativos indican una dependencia lineal decreciente. El valor absoluto del coeficiente de correlación indica el grado de la relación lineal entre las variables. Valores absolutos grandes indican las relaciones más fuertes. Cuando los valores del coeficiente sean pequeños, significativos de una pequeña dependencia lineal, se debe cambiar el modelo lineal por otro tipo de curva a fin de tener una mejor aproximación.

MÁSTER EN INGENIERÍA AMBIENTAL
18
Coeficiente de Correlación de Spearman
El coeficiente de Correlación de Spearman es una versión no paramétrica del coeficiente de correlación de Pearson. Este parámetro estadístico es adecuado para datos ordinales, o de intervalo, que no satisfagan el supuesto de normalidad. La correlación de Spearman se basa en los rangos de los datos en lugar de los valores reales, y compara dichos rangos para cada grupo de variables (Conover, 1998). El uso de este coeficiente estadístico es aconsejable cuando tenemos un número relativamente alto de categorías.
El coeficiente de correlación de Spearman requiere que las variables sean aleatorias continuas, o por lo menos una de ellas, de forma que los objetos o individuos puedan colocarse en series ordenadas (Siegel, 1983).
El coeficiente de Spearman se calcula con la fórmula expresada en la Ecuación 12.
di : la diferencia entre los rangos de X e Y, es decir di=rXi-rYi.
N: número de valores de la muestra.
Ecuación 12. Calculo del coeficiente de correlación de Spearman.
Los valores de los rangos se colocan según el orden numérico de los datos de la variable estudiada.
Su interpretación es semejante al coeficiente de correlación de Pearson. Índices cercanos a +1 indican una fuerte correlación creciente mientras que valores cercanos a -1 indican una fuerte correlación decreciente, los valores próximos a 0 indican que no hay correlación entre las variables (Siegel, 1983).
La prueba de significación asociada a ambos coeficientes únicamente prueba la hipótesis nula de que el coeficiente pueda ser igual a 0, es decir, que no exista ninguna relación lineal entre ambas variables. El nivel de significación “p” representa la probabilidad de error que se comete al aceptar el resultado observado como válido. Cuanto mayor sea este nivel de significación mayor error se comete al aceptar que las correlaciones observadas entre las variables de la muestra son indicadoras de la relación entre las respectivas variables de la población. Por ejemplo si p ≤ 0.05 existe una probabilidad del 5 % de que la relación observada entre las variables en la muestra estudiada sea casualidad. El nivel de significación 0.05 se considera un nivel de error aceptable en muchas áreas de estudio.
Liz Avendaño Universidad Politécnica de Valencia
42
= ∑ − · − ∑ − · −
: media de X.
: media de Y.
El coeficiente de correlación de Pearson está comprendido entre -1 y +1, donde los valores
próximos a +1 indican una fuerte dependencia lineal creciente y los valores de r próximos a -1
indican una dependencia lineal decreciente.
Los valores del coeficiente r pequeños indican una pequeña dependencia lineal. En estos casos se
debe cambiar el modelo lineal por otro tipo de curva a fin de tener una mejor aproximación.
3.4.2 Coeficiente de Correlación de Spearman
El coeficiente de correlación de Spearman o por rangos, también llamado rs es una medida no
paramétrica de asociación que requiere que ambas variables sean aleatorias continuas, de forma
que los objetos o individuos puedan colocarse en series ordenadas (Siegel, 1983).
El coeficiente de Spearman utiliza los números de orden (rangos) de cada grupo de variables y
compara dichos rangos (Conover, 1998). Este coeficiente es recomendable utilizarlo cuando los
datos presentan valores externos que afectan al coeficiente de Pearson o ante distribuciones no
normales.
La fórmula para calcular el Coeficiente de Spearman es la siguiente:
= 1 − 6 · ∑
−
donde:
di es la diferencia entre los rangos de X e Y, es decir di = rXi-rYi.
N es el número de valores de la muestra

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
19
1.5.2. ANÁLISIS MULTIVARIANTE
El análisis multivariante (AM) es la parte de la estadística y del análisis de datos que estudia, analiza, representa e interpreta los datos que resultan de observar más de una variable estadística sobre una muestra de individuos. Las variables observables son homogéneas y correlacionadas, sin que alguna predomine sobre las demás.
Dentro del AM se encuentran las dos técnicas que se describen a continuación:
Análisis de componentes principales
El análisis de componentes principales (ACP) es una técnica de reducción de la información disponible sobre un grupo de muestras, de los cuales se han tomado diversas variables sobre diversas características. Por ejemplo si se tienen datos de N muestras referentes a p características diferentes de éstos, es decir, X=(X1,....,Xp), el objetivo del ACP consiste en
reducir las dimensiones de la variable X creando una nueva variable U=(U1,.....,Ur), donde
r<p, cuyas componentes Ui sean combinación lineal de las Xi y que también expliquen una
proporción suficientemente grande de la dispersión total presente en los datos originales (Pérez, 1996).
Estas aproximaciones se puede orientar al estudio inicial de la variabilidad ambiental de las unidades poblacionales, y en este sentido cabe considerarla como una técnica de ordenación indirecta (Jongman et al., 1995) que permite analizar y/o modelar posteriormente la respuesta biológica de interés ante los factores extraídos por el análisis (Rottenberry and Wiens, 1980)
El proceso de cálculo del ACP es el siguiente:
El primer componente se construye buscando un vector b= (b1,...,bp) que, entre todos los que
tengan modulo unitario, atribuya a U1 la mayor varianza posible, de forma tal que este nuevo
componente constituya la mejor explicación unidimensional de la varianza total en los datos (Pérez, 1996).
Ecuación 13. Vector de componentes ACP
Procediendo con las variables sucesivas se construyen las componentes principales. Este método logra sintetizar al máximo la información con el criterio de pérdida mínima de capacidad explicativa con respecto a la varianza de las series (Pérez, 1996).
Cuando se realiza un ACP sobre una matriz de datos X, se está realizando la siguiente modelación (Aguado, 2004):
Liz Avendaño Universidad Politécnica de Valencia
43
Los valores de los rangos se colocan según el orden numérico de los datos de la variable
estudiada.
Su interpretación es semejante al coeficiente de correlación de Pearson. Para índices cercanos a
+1 la asociación es positiva, para valores cercanos a -1 la asociación es negativa y 0 significa que
no hay correlación entre las variables (Siegel, 1983).
3.4.3 Análisis de Componentes Principales (ACP)
El ACP es una técnica de reducción de la información disponible sobre un grupo de muestras, de
los cuales se han tomado diversas variables sobre diversas características. Por ejemplo si se
tienen datos de N muestras referentes a p características diferentes de éstos, es decir,
X=(X1,….,Xp), el objetivo del ACP consiste en reducir las dimensiones de la variable X creando
una nueva variable U=(U1,…..,Ur), donde r<p, cuyas componentes Ui sean combinación lineal de
las Xi y que también expliquen una proporción suficientemente grande de la dispersión total
presente en los datos originales (Pérez, 1996).
El proceso de cálculo del ACP es el siguiente:
El primer componente se construye buscando un vector b= (b1,…,bp) que, entre todos los que
tengan modulo unitario, atribuya a U1 la mayor varianza posible, de forma tal que este nuevo
componente constituya la mejor explicación unidimensional de la varianza total en los datos
(Pérez, 1996).
U1=b11·X1+b12·X2+….+b1P·Xp
Procediendo con las variables sucesivas se construyen las componentes principales. Este método
logra sintetizar al máximo la información con el criterio de pérdida mínima de capacidad
explicativa con respecto a la varianza de las series (Pérez, 1996).
Cuando se realiza un ACP sobre una matriz de datos X, se está realizando la siguiente
modelación (Aguado, 2004):
= · + = +

MÁSTER EN INGENIERÍA AMBIENTAL
20
Ecuación 14. Modelización realizada en el ACP
Donde, pa son los vectores de pesos que definen las direcciones principales de máxima
varianza en el espacio X. Estas direcciones determinan un subespacio de menor dimensión (A) que el espacio original.
La dimensión (A) se escoge de manera que no exista información importante de X en E que es
la matriz de residuos, y que, por tanto, representa el ruido. 𝑋es la estimación de X a partir de modelo con A componentes.
En la Figura 9 se muestran de forma gráfica las matrices del modelo ACP.
Figura 9. Representación esquemática de las matrices del modelo ACP.
Para una componente dada, los pesos son definidos como la dirección sobre la que se están proyectando las observaciones y están poniendo de manifiesto la contribución de las variables originales en la formación de dicha componente.
Análisis Canónico
El análisis de correspondencias canónicas (ACC) es una técnica de ordenación directa, a diferencia del ACP. El ACC asume un modelo unimodal en la respuesta de los organismos sobre combinaciones de un conjunto de variables, considerando simultáneamente la información biológica y la información ambiental. En el ACC, los ejes que explican la respuesta biológica están forzados a ser una suma ponderada de las variables estudiadas (ter Braak and Smilauer, 1998).
En los análisis canónicos se extrae toda la varianza de la matriz respuesta que esta relacionada con la matriz explicativa.
La ordenación de la matriz respuesta se precisa de forma que los vectores de ordenación resultantes son combinaciones lineales de las variables de la matriz explicativa.
El proceso del ACC es el siguiente:
Liz Avendaño Universidad Politécnica de Valencia
43
Los valores de los rangos se colocan según el orden numérico de los datos de la variable
estudiada.
Su interpretación es semejante al coeficiente de correlación de Pearson. Para índices cercanos a
+1 la asociación es positiva, para valores cercanos a -1 la asociación es negativa y 0 significa que
no hay correlación entre las variables (Siegel, 1983).
3.4.3 Análisis de Componentes Principales (ACP)
El ACP es una técnica de reducción de la información disponible sobre un grupo de muestras, de
los cuales se han tomado diversas variables sobre diversas características. Por ejemplo si se
tienen datos de N muestras referentes a p características diferentes de éstos, es decir,
X=(X1,….,Xp), el objetivo del ACP consiste en reducir las dimensiones de la variable X creando
una nueva variable U=(U1,…..,Ur), donde r<p, cuyas componentes Ui sean combinación lineal de
las Xi y que también expliquen una proporción suficientemente grande de la dispersión total
presente en los datos originales (Pérez, 1996).
El proceso de cálculo del ACP es el siguiente:
El primer componente se construye buscando un vector b= (b1,…,bp) que, entre todos los que
tengan modulo unitario, atribuya a U1 la mayor varianza posible, de forma tal que este nuevo
componente constituya la mejor explicación unidimensional de la varianza total en los datos
(Pérez, 1996).
U1=b11·X1+b12·X2+….+b1P·Xp
Procediendo con las variables sucesivas se construyen las componentes principales. Este método
logra sintetizar al máximo la información con el criterio de pérdida mínima de capacidad
explicativa con respecto a la varianza de las series (Pérez, 1996).
Cuando se realiza un ACP sobre una matriz de datos X, se está realizando la siguiente
modelación (Aguado, 2004):
= · + = +
Liz Avendaño Universidad Politécnica de Valencia
44
Donde, pa son los vectores de pesos que definen las direcciones principales de máxima varianza
en el espacio X. Estas direcciones determinan un subespacio de menor dimensión (A) que el
espacio original.
La dimensión (A) se escoge de manera que no exista información importante de X en E que es la
matriz de residuos, y que, por tanto, representa el ruido X̂ es la estimación de X a partir de
modelo con A componentes.
En la Figura 16 se muestran de forma gráfica las matrices del modelo ACP.
Figura 16. Representación esquemática de las matrices del modelo ACP.
Para una componente dada, los pesos son definidos como la dirección sobre la que se están
proyectando las observaciones y están poniendo de manifiesto la contribución de las variables
originales en la formación de dicha componente (Aguado, 2004).
3.4.4 Tratamiento por ordenador
Las correlaciones de Pearson y Spearman se obtuvieron mediante el programa Statgraphics Plus
versión 5.1. En este programa se introdujo una matriz semejante a la Figura 15 con todos los
parámetros a estudiar y se realizó un análisis estadístico correspondiente a las correlaciones de
Pearson y Spearman.
El programa genera como salida de este estudio una gran tabla con los valores de las
correlaciones y los p-valores de cada una. Estos últimos indican la bondad estadística del modelo

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
21
Sean X= (X1, …, Xp), Y=(Y1, …, Yq) dos vectores aleatorios de dimensiones p y q. Se pretende encontrar dos variables compuestas, U = Xa = a1X1+ … + apXp, V = Yb = b1Y1 + … + bqYq, siendo a = (a1, …, ap), b = (b1, …, bq)´, tales que la correlación entre ambas sea máxima. Se indicarán por S11, S22 las matrices de covarianzas (muestrales) del primer y segundo conjunto, es decir, de las variables X, Y, respectivamente, y sea S12 la matriz p x q con las covarianzas de las variables X con las variables Y. Es decir:
donde: S21 = S12
Figura 10. Matriz de covarianzas ACC
Entonces podremos suponer que var (U) = a´S11a =1 , var (V) = b´S22b = 1, reduciendo el problema a maximizar a´S12b restringido a a´S11a =1, b´S22b = 1.
Los vectores de coeficientes a, b que cumplen esta condición son los primeros vectores canónicos. La máxima correlación entre U, V es la primera correlación canónica r1.
La posibilidad de comprobar si existe una relación estadísticamente significativa entre la composición de la comunidad y una matriz de variables ambientales permite pasar de un uso meramente descriptivo de las ordenaciones a emplearlas como herramientas para el contraste de hipótesis formuladas a priori. La hipótesis nula de un ACC es que no existe relación alguna entre la matriz de variables respuesta y la matriz de variables explicativas. La significación de esta hipótesis nula se puede evaluar mediante permutaciones. En este caso, el estadístico es la suma de todos los valores propios canónicos y su significación se evalúa mediante un test de F. La hipótesis alternativa establece que la suma de todos los valores propios canónicos es mayor que la que se obtiene de matrices en las que se han permutado sus filas de datos. La suma de todos los valores propios canónicos dividida por el total de variación de la matriz Y permite obtener la proporción de la variación de Y explicada por X, que es análoga al coeficiente de determinación de las regresiones múltiples.
4.3. CORRELACI”N CAN”NICA 65
2. var(Y ) =var(bY )+var(eY ):
3. R2 =var(bY )=var(Y ):
Demost.:
1. Es consecuencia de b = : En efecto,
cov(bY ; eY ) = E(bY eY ) = E(b0X0(Y b
0X))
= b0 b
0b = 0:
2. Es consecuencia inmediata de 1).
3. De
cov(Y; bY ) = cov Y;
pX
i=1
biXi
!=
pX
i=1
bii = b0 = b
0b = var(bY );
obtenemos
R2 =cov
2(Y; bY )var(Y )var(bY )
=var(bY )var(Y )
: (4.2)
4.3. CorrelaciÛn canÛnica
Sean X = (X1; : : : ; Xp); Y = (Y1; : : : ; Yq) dos vectores aleatorios de di-mensiones p y q: Planteemos el problema de encontrar dos variables com-puestas
U = Xa = a1X1 + + apXp; V = Yb = b1Y1 + + bqYq;
siendo a = (a1; : : : ; ap)0;b = (b1; : : : ; bq)
0tales que la correlaciÛn entre ambas
cor(U; V )
sea m·xima. Indiquemos por S11;S22 las matrices de covarianzas (muestrales)del primer y segundo conjunto, es decir. de las variables X;Y; respectiva-mente, y sea S12 la matriz p q con las covarianzas de las variables X con
las variables Y: Es decir:X Y
X S11 S12Y S21 S22

MÁSTER EN INGENIERÍA AMBIENTAL
22

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
23
2. OBJETIVOS
El presente trabajo se ha desarrollado con el objetivo de estudiar las relaciones entre la población de bacterias nitrificantes presentes en el fango activo de la EDAR Denia-Ondara-Pedreguer y las variables físico-químicas y operacionales del proceso de la EDAR durante un año.
Los objetivos específicos que se pretenden alcanzar son:
- Identificar diversas subclases de BOA y BON mediante la utilización de sondas específicas con la técnica FISH.
- Cuantificar en cada muestra los grupos de bacterias nitrificantes estudiadas con respecto a la población total de bacterias.
- Estudiar las relaciones existentes entre la dinámica poblacional de las bacterias nitrificantes y su abundancia con los parámetros físico-químicos y operacionales de la EDAR.
- Estudiar la abundancia de distintas especies de BOA y BON a lo largo del tiempo y
valorar su contribución a la eliminación de amonio (N-NH4+).

MÁSTER EN INGENIERÍA AMBIENTAL
24

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
25
3. MATERIALES Y MÉTODOS
3.1. DESCRIPCIÓN DE LA EDAR
Las muestras empleadas para la realización de este proyecto fueron tomadas en la EDAR de Denia-Ondara-Pedreger ubicada en la Marina Alta (Provincia de Alicante), que trata las aguas residuales de los municipios de Denia, Ondara y Pedreguer.
Esta EDAR trata un caudal de 17.089 m3/día correspondiente a una población de 45.152 he (Datos EPSAR, 2010).
La línea de aguas cuenta con un pretratamiento compuesto por reja de gruesos, tamices, desarenador y desengrasador. El tratamiento secundario de la planta consiste en un reactor anóxico/óxico, donde el volumen total del tanque de aireación es de 12.111 m3, seguido de decantación y un tratamiento terciario de desinfección por ultravioletas. El proceso que se desarrolla es de aireación prolongada, con eliminación biológica de nitrógeno mediante nitrificación-desnitrificación vía nitrato y eliminación fisco-química de fósforo.
En la Figura 11 podemos observar el esquema de depuración.
Figura 11. Esquema de depuración de la EDAR Denia-Ondara-Pedreguer (EPSAR)
En la Tabla 3 se presentan los rendimientos de eliminación en la EDAR durante el periodo de estudio. Podemos observar los elevados rendimientos de eliminación de nitrógeno total, kjeldahl y amoniacal, lo que es significativo de un buen proceso de nitrificación.

MÁSTER EN INGENIERÍA AMBIENTAL
26
Tabla 3. Rendimientos de eliminación en la EDAR Denia-Ondara-Pedreguer
3.2. TOMA DE MUESTRAS
Las muestras utilizadas para el desarrollo de este proyecto fueron proporcionadas por el Instituto Universitario de Ingeniería del Agua y Medio Ambiente (IIAMA) de la Universidad Politécnica de Valencia (UPV).
La toma de muestras se realizó durante el periodo de diciembre de 2008 a diciembre de 2009, con una frecuencia quincenal, lo que supone un total de 23 muestras.
Las campañas de muestreo tuvieron una duración de cuatro días. Los días 1 y 2 se analizaron la DQO total, DQO soluble y DBO5 en el afluente al reactor, mientras que el día 3 se llevó a cabo un análisis completo del afluente y el efluente. El objetivo del primer análisis fue estudiar la influencia de la carga orgánica y del segundo establecer el rendimiento del proceso biológico (Zornoza et al., 2010). El cuarto día se procedió al muestreo del licor mezcla, que servirá empleado para la identificación y la cuantificación de bacterias nitrificantes.
Tanto las muestras del afluente como del efluente fueron muestras integradas, obtenidas a partir de la mezcla de muestras simples horarias en relación al caudal, mientras que las muestras del licor mezcla que se tomaron a la salida del reactor biológico fueron de tipo simple y de carácter puntual.
En la Tabla 4 se presenta un esquema de la campaña de muestreo indicándose la duración el origen y tipo de muestra.
Tabla 4. Esquema de la campaña de muestreo (Zornoza et al., 2010)

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
27
3.3. VARIABLES FÍSICO-QUÍMICAS Y PARÁMETROS OPERACIONALES
Las variables físico-químicas se determinaron mediante procedimientos normalizados de la American Public Health Association (APHA, 2005). La fracción filtrada se obtuvo a través de un filtro de lana de vidrio (Whatman GF/C) con un tamaño de poro de 1.2 µm. La fracción soluble se obtuvo a través de un filtro de 0.45 µm (Grady, 1989).
En la Tabla 5 se muestra una relación de los parámetros físico-químicos analizados que han sido usados en el presente estudio y sus respectivos días de muestreo.
Tabla 5.Variables físico-químicos del afluente, efluente y licor mezcla en relación a la campaña de muestreo
En la Tabla 6 se detalla la relación de las variables operacionales que han sido consideradas en el presente estudio.
Tabla 6. Parámetros operacionales

MÁSTER EN INGENIERÍA AMBIENTAL
28
Los valores de OD en el reactor fueron distribuidos en tres intervalos: < 0,8 OD bajo, 0,8 – 2 OD medio y > 2 ppm OD alto.
3.4. TÉCNICA DE HIBRIDACIÓN IN SITU CON SONDAS MARCADAS CON
FLUOROFOROS
A continuación se describe el protocolo empleado para la realización de la técnica FISH en las muestras empleadas en el presente estudio.
3.4.1. FIJACIÓN DE LAS MUESTRAS.
El objetivo de la fijación de las muestras es que las células mantengan su morfología e inactivar la actividad enzimática.
La fijación de las células fue realizada tras la toma de muestras siguiendo el protocolo para Gram negativas que se describe a continuación:
- Lavar 1 ml de flóculo con 500 µl de PBS, seguidamente añadir 750 µl de PFA y mantenerlo a 4ºC durante 1 a 3 horas.
- Centrifugar la muestra durante 3 minutos y eliminar el fijador.
- Lavar las celular con 500 µl de PBS 1X.
- Resuspender en 500 µl de PBS 1X y añadir 500 µl de etanol frío (4 oC).
- Guardar a -20 oC, se pueden conservar durante varios meses bajo estas condiciones..
3.4.2. TRATAMIENTO DE LOS PORTAOBJETOS CUBIERTOS CON TEFLÓN.
El objetivo del tratamiento de los portaobjetos es crear un medio adecuado en el que se mantengan las células ya fijadas tras ser aplicadas en el portaobjeto a pesar de los lavados posteriores a los que serán sometidos. Los pasos realizados se describen a continuación:
− Lavar con solución de limpieza.
− Enjuagar con agua destilada.
− Secar al aire dejándolo escurrir durante un día. Se deben proteger del polvo ambiental cubriendo con papel de aluminio.
− Cubrir con gelatina por inmersión en la solución de gelatina 0,1% con cromato sulfato potásico 0,01% preparada al momento a una temperatura de 60oC.
− Secar al aire.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
29
3.4.3. APLICACIÓN DE LAS MUESTRAS A LOS PORTAOBJETOS Y PERMEABILIZACIÓN
DE LAS CELULAS.
La metodología empleada fue la siguiente:
- Mediante uso de micropipetas se coloca en cada uno de los pocillos del portaobjetos un volumen de 4µl de muestra fijada.
− Secar al aire.
− Deshidratar en etanol 50% durante 3 minutos por inmersión.
− Deshidratar en etanol 80% durante 3 minutos por inmersión.
− Deshidratar en etanol absoluto durante 3 minutos por inmersión.
3.4.4. HIBRIDACIÓN IN SITU.
Preparación de la solución de hibridación
El primer paso de la técnica es la preparación de la solución de hibridación, cuya composición variará en función del porcentaje de formamida tal y como se indica en la Tabla 7.
Tabla 7. Solución de hibridación en función del porcentaje de formamida
- Introducir los reactivos en las cantidades establecidas en un microtubo de 2 ml.
- Añadir las sondas a la solución de hibridación según las consideraciones establecidas a continuación:
§ Por cada pocillo se necesitan 9 µl de solución de hibridación y 1 µl de sonda por tanto se prepara una solución con estas proporciones y se aplican 10 µl de esta solución a cada uno de los pocillos.
§ En estas hibridaciones se necesita utilizar más de una sonda. Se utiliza la sonda EUBMIX para identificar el total de bacterias presentes en la muestra y la sonda correspondiente a la bacteria nitrificante que se pretende identificar. En este caso se aplica las sondas de manera equimolecular, es decir en relación 1:1.

MÁSTER EN INGENIERÍA AMBIENTAL
30
§ En ocasiones para aumentar la especificidad del método es necesario emplear una sonda competidora de la bacteria nitrificante a identificar. En este caso las proporciones aplicadas de las sondas son 2:1:1, que corresponde a EUBMIX, bacteria nitrificante y competidora de la bacteria nitrificante respectivamente.
- Distribuir de manera homogénea en cada uno de los pocillos la solución de hibridación junto con las sondas.
- Colocar los portaobjetos con las muestras en un tubo falcon de 50 ml. En el tubo falcon previamente se habrá introducido papel de celulosa humedecido con 1.5 ml de la solución de hibridación. Los portaobjetos se deben mantener en posición horizontal
- Incubar las muestras durante un mínimo de 1 hora y 30 minutos a 46ºC.
Lavado de las muestras
Para eliminar el exceso de sonda que no ha hibridado y la formamida de los portaobjetos se prepara una solución de lavado con los reactivos indicados en la Tabla 8 dependiendo del porcentaje de formamida que ha sido utilizado en la solución de hibridación.
Tabla 8. Solución de lavado en función del porcentaje de formamida
El procedimiento de lavado de los portaobjetos es el siguiente:
- Sacar el portaobjetos de la incubadora e introducirlo en un tubo con la solución de lavado, mantenerlo en un baño de agua a 48 ºC durante 10 -15 minutos protegidos de la luz cubriéndolos con papel de aluminio.
- Lavar el portaobjetos con la solución de lavado y sumergirlo en agua MiliQ.
- Secar el portaobjetos a temperatura ambiente protegiéndolo de la luz.
A partir de este momento la muestra esta lista para ser visualizada, o bien se guarda a -20ºC.
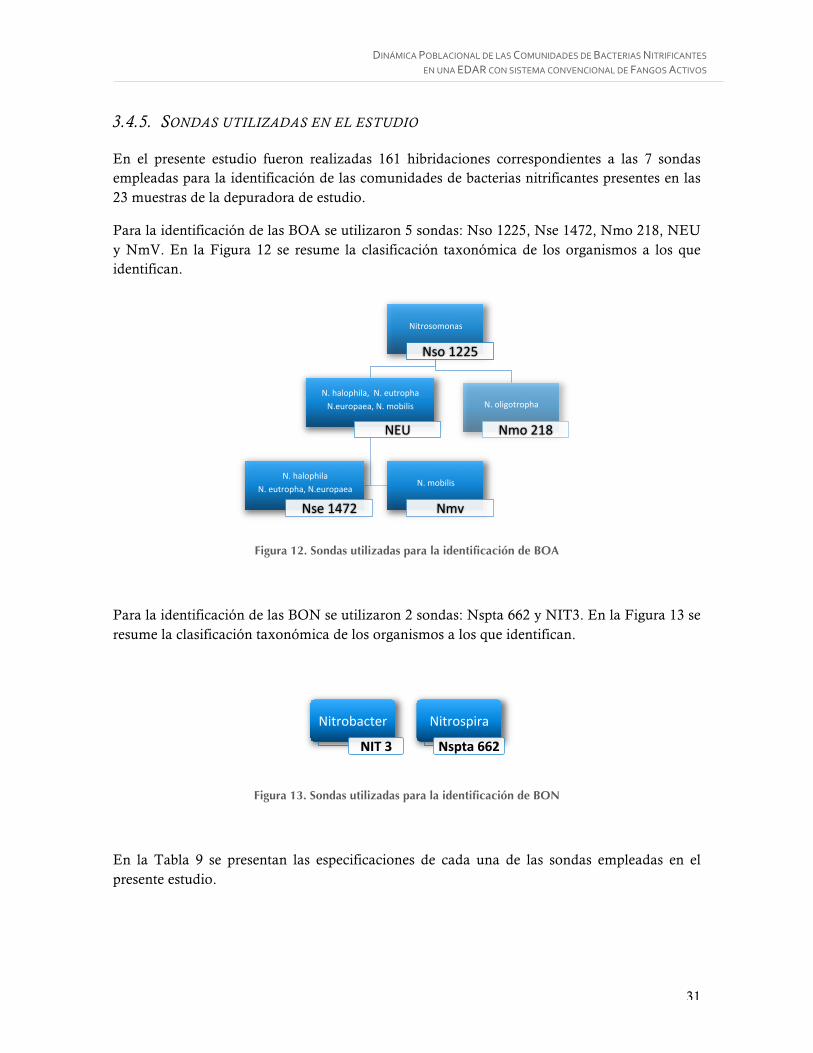
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
31
3.4.5. SONDAS UTILIZADAS EN EL ESTUDIO
En el presente estudio fueron realizadas 161 hibridaciones correspondientes a las 7 sondas empleadas para la identificación de las comunidades de bacterias nitrificantes presentes en las 23 muestras de la depuradora de estudio.
Para la identificación de las BOA se utilizaron 5 sondas: Nso 1225, Nse 1472, Nmo 218, NEU y NmV. En la Figura 12 se resume la clasificación taxonómica de los organismos a los que identifican.
Figura 12. Sondas utilizadas para la identificación de BOA
Para la identificación de las BON se utilizaron 2 sondas: Nspta 662 y NIT3. En la Figura 13 se resume la clasificación taxonómica de los organismos a los que identifican.
Figura 13. Sondas utilizadas para la identificación de BON
En la Tabla 9 se presentan las especificaciones de cada una de las sondas empleadas en el presente estudio.
Nitrosomonas
Nso 1225
N. halophila, N. eutropha N.europaea, N. mobilis
NEU
N. halophila N. eutropha, N.europaea
Nse 1472
N. mobilis
Nmv
N. oligotropha
Nmo 218
Nitrobacter
NIT 3
Nitrospira
Nspta 662

MÁSTER EN INGENIERÍA AMBIENTAL
32
Tabla 9. Sondas utilizadas en el presente estudio
3.5. CUANTIFICACIÓN DE MICROORGANISMOS
La cuantificación de microorganismos en las muestras hibridadas consiste en la observación de las muestras con microscopio de epifluorescencia y la captura de imágenes que posteriormente son procesadas por el software desarrollado por Borras (2008).
El microscopio utilizado fue un microscopio de epifluorescencia Olympus BX 50.
Previamente a la observación con el microscopio se aplicó Vectashield, un compuesto que evita la perdida de fluorescencia.
Las observaciones se realizaron a 1000 aumentos para todas las sondas excepto para la sonda Nso 1225 que se realizaron a 600 aumentos.
Por cada pocillo se fotografiaron entre 20 y 25 campos. Por cada campo se capturaron entre 2 y 3 fotografías, una primera correspondiente a las bacterias nitrificantes diana (sonda específica), la segunda de toda la comunidad bacteriana (sonda EUBmix), y en ocasiones una tercera con el filtro doble banda que ayuda a diferenciar falsas señales de la sonda de hibridación.
Las fotografías se tomaron con una cámara digital Olimpus DP 12 acoplada al microscopio con los filtros: U-MWIB (Fluoresceína, FAM) y U-MWIG (Rodamina, TAMRA).
Se realizaron aproximadamente un total de 6.600 fotografías correspondientes a las 4 sondas que dieron una señal positiva en las 22 muestras hibridadas positivamente. La observación y toma de imágenes de cada pocillo duró aproximadamente 40 minutos, lo que supuso un total de 6.240 minutos equivalentes a 104 horas empleadas en la observación y captura de imágenes de las muestras.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
33
Tras la captura de imágenes estas fueron tratadas con el programa Photoshop versión CS2 9.0, para eliminar falsas señales de la sonda de hibridación con el objetivo de que la cuantificación posterior fuera lo más precisa posible.
Como se comentaba anteriormente la cuantificación fue realizada haciendo uso del software desarrollado por Borrás. Este software desarrollado para Matlab descompone las imágenes digitales tomadas en RBG a escala binaria para realizar el conteo de píxeles a partir del cual se calcularán los porcentajes de la población de bacterias.
El algoritmo del programa permite con un simple ajuste eliminar las partes de la imagen que no son de interés mediante los parámetros Low_in y Gamma_in. Con el primero se consigue eliminar la parte de la imagen que representa la fluorescencia de fondo (background) y establece un valor de 0 a 1, por debajo del cual es un falso positivo (Borrás, 2008). El parámetro Gamma_in representa la forma de la curva que describe la relación entre los valores de intensidad de la imagen original y la nueva imagen. Cuando Gamma_in es menor a 1 la nueva imagen tendrá una intensidad más alta, es decir que será más brillante, si por el contrario este parámetro es superior a 1 la nueva imagen será más oscura (Borrás, 2008).
El programa calcula el porcentaje y la desviación estándar del área ocupada por las bacterias nitrificantes en función del total de la comunidad bacteriana presente, así como el error de la medida. Este error es calculado dividiendo la desviación estándar por la raíz cuadrada de “n”, donde n es el número de campos examinados (Borrás, 2008).
En la Figura 14 se muestra el flujo de procesos que realiza dicho software para la cuantificación.

MÁSTER EN INGENIERÍA AMBIENTAL
34
Figura 14. Flujo de procesos para la cuantificación de bacterias.
Los resultados serán expresados tanto en porcentaje (%) como en miligramos de sólidos suspendidos volátiles en el licor mezcla por litro (mgSSVLM/L).
3.6. ANÁLISIS ESTADÍSTICO
En el análisis estadístico se emplearon 35 variables (23 variables físico-químicas, 8 variables operacionales y 4 variables de bacterias nitrificantes) y 23 muestras. En la Tabla 10 se indica la nomenclatura abreviada que se ha utilizado para cada una de las variables analizadas en este estudio.
Liz Avendaño Universidad Politécnica de Valencia
39
Figura 14 Flujo de procesos para la cuantificación de bacterias.
3.4 . Análisis Estadístico.
En el análisis estadístico se emplearon 86 variables medidas (63 variables físico-químicas, 8
variables operacionales y 15 protistas) y 24 muestras, dando lugar a una matriz semejante a la que
se muestra en la Figura 15.
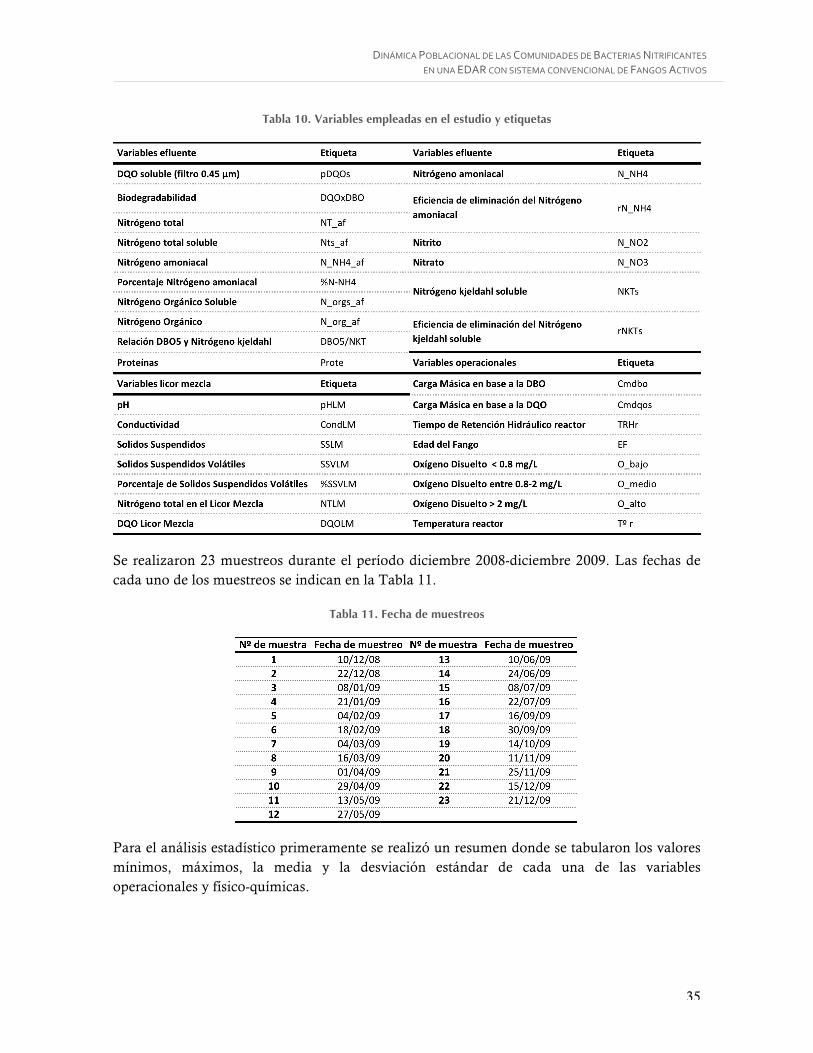
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
35
Tabla 10. Variables empleadas en el estudio y etiquetas
Se realizaron 23 muestreos durante el período diciembre 2008-diciembre 2009. Las fechas de cada uno de los muestreos se indican en la Tabla 11.
Tabla 11. Fecha de muestreos
Para el análisis estadístico primeramente se realizó un resumen donde se tabularon los valores mínimos, máximos, la media y la desviación estándar de cada una de las variables operacionales y físico-químicas.

MÁSTER EN INGENIERÍA AMBIENTAL
36
Posteriormente se emplearon diferentes técnicas estadísticas complementarias para tratar de relacionar los cambios en la composición y abundancia de las BOA y de las BON con los parámetros físico-químicos y operacionales.
Primero se realizó un análisis bivariante, que consistió en el cálculo de los coeficientes de Pearson y de Spearman.
Seguidamente se realizó un análisis multivariante mediante la aplicación de un análisis de componentes principales (ACP) y un análisis de correspondencias canónicas (ACC).
3.6.1. TRATAMIENTO DE DATOS REALIZADO PARA DETERMINAR LAS
CORRELACIONES DE PEARSON Y SPEARMAN
Previamente a la determinación de las correlaciones de Pearson y Spearman se comprobó la distribución de la normalidad de los datos obtenidos de las distintas variables mediante los test de Curtosis y Asimetría. Las variables cuyos datos no siguieron una distribución normal se transformaron a través de un cálculo logarítmico (variable = Ln [variable + 1]) (Esteban et al., 1991).
Las correlaciones de Pearson y Spearman se obtuvieron mediante el programa IBM SPSS Statistics versión 19. En este programa se introdujo una matriz con los parámetros operacionales y las especies de bacterias nitrificantes y se realizó un análisis estadístico correspondiente a las correlaciones de Pearson y Spearman.
El programa genera como salida de este estudio una tabla con los valores de las correlaciones y los p-valores de cada una. Estos últimos indican la bondad estadística del modelo ajustado. Por ejemplo p-valores iguales o inferiores a 0,05 indican una relación estadísticamente significativa al 95%.
3.6.2. TRATAMIENTO DE DATOS REALIZADO EN EL ANÁLISIS DE COMPONENTES
PRINCIPALES
Para realizar el análisis de componentes principales se utilizó el programa IBM SPSS Statistics versión 19, donde se introdujo la matriz de datos y se especificaron las variables para realizar el análisis.
La primera de las salidas del programa es la tabla que presenta la medida de adecuación muestral de Kaiser-Meyer-Olkin y la prueba de elasticidad de Bartlett.
La medida de adecuación muestral de Kaiser-Meyer-Olkin contrasta si las correlaciones parciales entre las variables son pequeñas, tomando valores entre 0 y 1, e indica que el análisis factorial es tanto más adecuado cuanto mayor sea su valor. Mientras que la prueba de esfericidad de Bartlett contrasta si la matriz de correlaciones es una matriz identidad, lo cual indicaría que el modelo factorial es inadecuado. Este estadístico se obtiene a partir de una

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
37
transformación del determinante de la matriz de correlaciones y cuanto mayor sea, y por tanto menor el nivel de significación, más improbable es que la matriz sea una matriz identidad y más adecuado resulta el análisis factorial.
Otra de las salidas del programa es la matriz anti-imagen, la diagonal de esta matriz presenta la medida de la adecuación muestral para cada una de las variables introducidas en el análisis.
En el presente estudio se han descartado todas aquellas variables cuya medida de la adecuación muestral está por debajo de 0,5, de manera que la medida de adecuación muestral de Kaiser-Meyer-Olkin y la prueba de elasticidad de Bartlett dieran valores correspondientes que establecieran la adecuación del análisis. El descarte de las variables que no se ajustaban a estos criterios dio como resultado 4 ACP con las variables indicadas en la tabla Tabla 12 .
Otra de las tablas que ayudan a establecer la adecuación del análisis es la de varianza total explicada, donde el porcentaje acumulado de la suma de las saturaciones al cuadrado de la rotación indica el porcentaje de la variabilidad de los datos que es explicada por los factores extraídos. En este estudio se han considerado adecuados los ACP con un porcentaje acumulado de varianza del 50%.
A pesar de tener en cuenta estas deferencias hay que considerar que para que el ACP resulte significativo el número de observaciones debe ser superior a 50. En este caso para que las cargas factoriales fueran significativas deberían estar por encima de 0,75. Por tanto se tendrá presente durante la evaluación de los resultados que el análisis no es riguroso, sino exploratorio y será confirmado por otros métodos avanzados de ordenación ambiental.
Tabla 12. Variables utilizadas para realizar el análisis de componentes principales
La salida empleada para la interpretación de los datos serán el diagrama de dispersión biespacial y la matriz de componentes rotados, la cual indica la carga factorial que cada una de las variables estudiadas aporta a cada uno de los componentes principales.

MÁSTER EN INGENIERÍA AMBIENTAL
38
3.6.3. TRATAMIENTO DE DATOS REALIZADO EN EL ANÁLISIS CANONICO DE
CORRESPONDENCIA
Para realizar el análisis de correspondencias canónicas se utilizó el programa XLSTAT versión 2013.4.05, donde se introdujo la matriz de datos y se especificaron las variables para realizar el análisis.
En primer lugar se comprobó si existían relaciones lineales entre las variables o bien unimodales, para ello se evaluaron los resultados de las pruebas de permutación, que indican la aceptación o rechazo de la hipótesis nula.
Como se comentaba en el apartado 1.5.2, la hipótesis nula establece que no existe relación lineal entre la matriz de variables respuesta (organismos) y la matriz de variables explicativas (variables del proceso). En los casos que se acepta la hipótesis nula no es posible llevar a cabo el ACC y se debe proceder a la realización del análisis de redundancia. En este caso todos los datos analizados dieron como resultado la aceptación de la hipótesis nula, por tanto se utilizó el ACC.
El resultado del ACC consiste principalmente en un diagrama de ordenación (biplot o triplot) formado por un sistema de ejes o dimensiones, los cuales son combinación lineal de las variables explicativas, en ellos es posible representar los sitios, las especies y las variables ambientales.
Este diagrama representa un análisis directo del gradiente. La longitud de las variables explicativas, representadas por vectores, indican su variabilidad. Cuanto menor es el ángulo entre ellas más directa es la relación entre las variables, si forman un ángulo de 90º significa que las variables no están relacionadas, y a medida que este se va a aproximando a un ángulo de 180º la relación que indica es decreciente. Cuanto más alto corte la perpendicular de la variable respuesta con la variable explicativa mayor será el grado de relación entre ambas.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
39
4. RESULTADOS
Los resultados que se obtuvieron durante la realización del proyecto corresponden, tal como se explicó en el capitulo anterior, a la realización de las hibridaciones mediante la técnica FISH de las muestras tomadas quincenalmente en el fango activo de la depuradora de Denia-Ondara-Pedreguer, durante un período de un año. Las hibridaciones se realizaron con el fin de determinar y cuantificar la cantidad de bacterias nitrificantes del grupo de las amonio-oxidantes (BOA) y nitrito-oxidantes (BON) presentes en el licor mezcla del reactor biológico.
Seguidamente se realizó un análisis cualitativo y cuantitativo de las sondas que dieron resultado positivo en las hibridaciones. Por último se realizó un análisis estadístico bivariante, un análisis de componentes principales (ACP) y un análisis de correspondencias canónicas (ACC) para determinar las relaciones entre la dinámica poblacional de las bacterias nitrificantes y las variables físico-químicas y operacionales de la EDAR.
4.1. CAPTURA DE IMÁGENES DE LAS SONDAS CON SEÑAL DE
HIBRIDACIÓN POSITIVA
En el presente estudio fueron realizadas 161 hibridaciones correspondientes a las 7 sondas empleadas para la identificación de las comunidades de bacterias nitrificantes presentes en las 23 muestras de la EDAR de estudio.
De las 23 muestras disponibles solo pudieron utilizarse 22 para la cuantificación debido a que la muestra correspondiente a la fecha 15/12/2009 no produjo hibridación para ninguna de las sondas utilizadas.
Como se comentó en el apartado de Materiales y métodos, se utilizaron 5 sondas para la identificación de las BOA, observándose una abundancia significativas en 3 de ellas, Nso 1225, NEU y Nmo 218. Por el contrario las sondas NmV y Nse 1472 no dieron señal positiva en todas las muestras hibridadas, y en los casos en que se detectó la presencia de bacterias esta fue inferior al 1%.
Para la identificación de las BON fueron utilizadas 2 sondas, la sonda NIT3 que identifica al género Nitrobacter y la sonda Ntspa 662 que identifica al género Nitrospira. La sonda Ntspa 662 mostró abundancias significativas, mientras que la sonda NIT3 únicamente dio señal positiva en algunas de las muestras analizadas, siendo todos los casos su abundancia inferior al 1%.
En la Tabla 13 se presenta una relación de las muestras hibridadas con las respectivas sondas, donde se indica por fecha de muestreo si existió presencia de bacterias (+) o si no se observaron comunidades de bacterias (-).
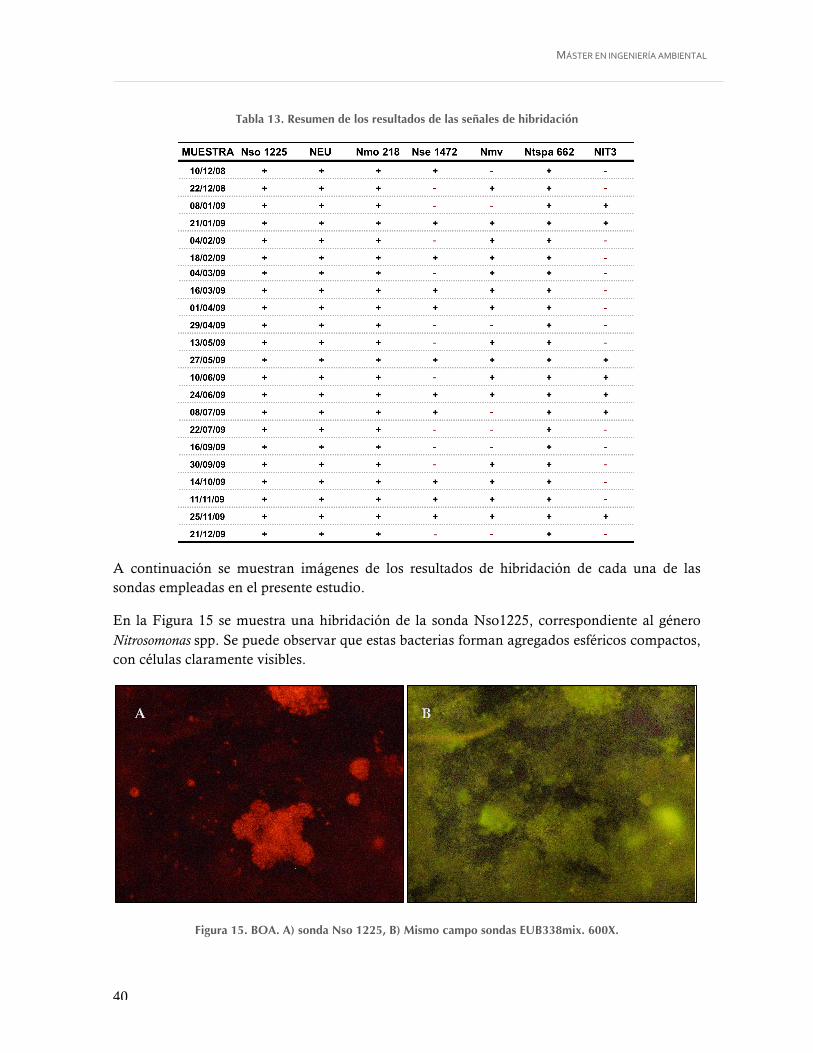
MÁSTER EN INGENIERÍA AMBIENTAL
40
Tabla 13. Resumen de los resultados de las señales de hibridación
A continuación se muestran imágenes de los resultados de hibridación de cada una de las sondas empleadas en el presente estudio.
En la Figura 15 se muestra una hibridación de la sonda Nso1225, correspondiente al género Nitrosomonas spp. Se puede observar que estas bacterias forman agregados esféricos compactos, con células claramente visibles.
Figura 15. BOA. A) sonda Nso 1225, B) Mismo campo sondas EUB338mix. 600X.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
41
En la Figura 16 se observa la hibridación de la sonda a NEU, la cual identifica a los géneros: N. halophila, N. europea, N. eutropha y Nitrosococcus mobilis. Se muestra un agregado poco compacto y algunas células libres. Los agregados presentan un tamaño mucho menor que en el caso anterior.
Figura 16. BOA. A) sonda NEU, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y NEU. 1000X.
En la Figura 17 se observa un campo correspondiente a la hibridación con la sonda Nmo218, la cual identifica las bacterias BOA del linaje N. oligotropha. Se presentan células muy visibles y compactas.
Figura 17. BOA. A) sonda Nmo 218, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y Nmo 218. 1000X.
En comparación con las especies anteriores, vemos como Nc. mobilis (sonda NmV) y las especies N. europaea y N. eutropha (Nse 1472. mobilis presentan agregados muy compactos y de tamaño mucho menor.
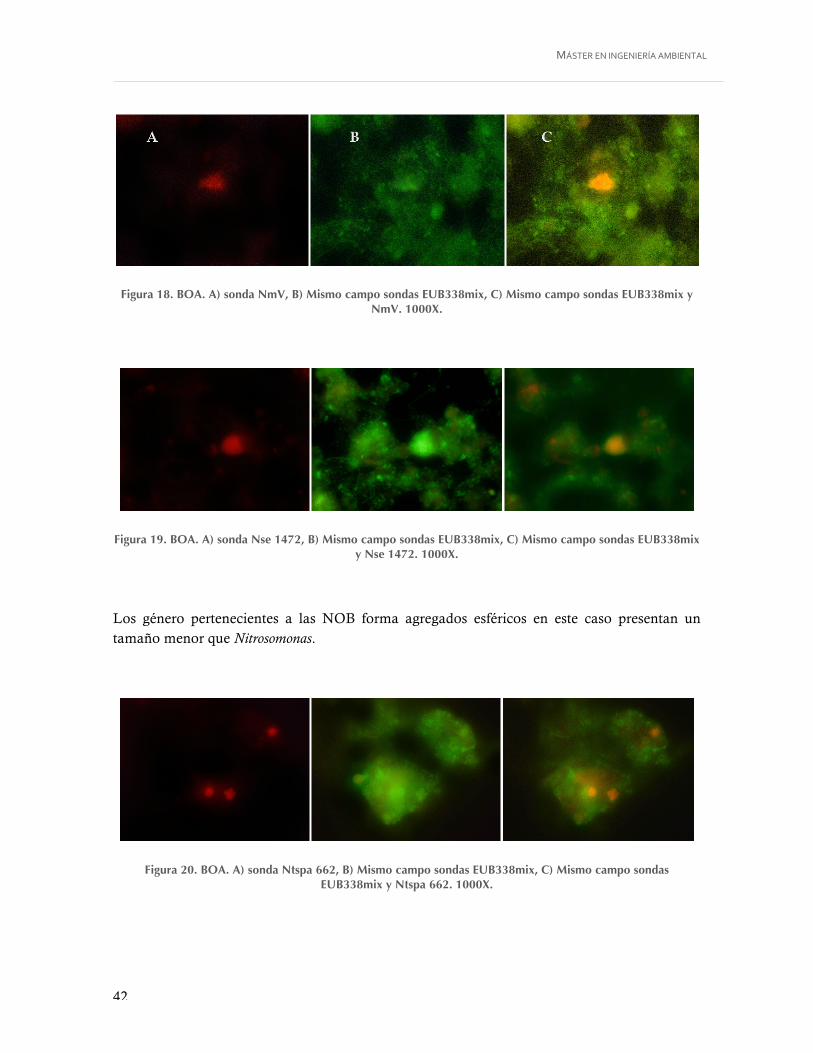
MÁSTER EN INGENIERÍA AMBIENTAL
42
Figura 18. BOA. A) sonda NmV, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y NmV. 1000X.
Figura 19. BOA. A) sonda Nse 1472, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y Nse 1472. 1000X.
Los género pertenecientes a las NOB forma agregados esféricos en este caso presentan un tamaño menor que Nitrosomonas.
Figura 20. BOA. A) sonda Ntspa 662, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y Ntspa 662. 1000X.
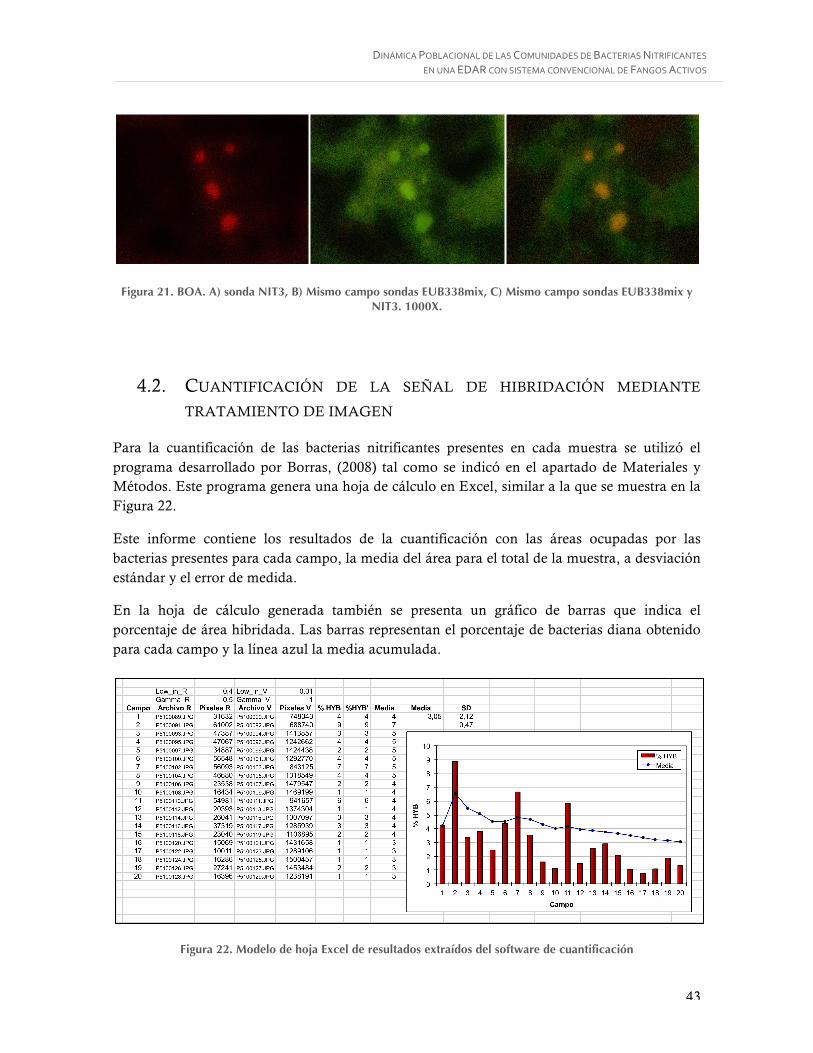
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
43
Figura 21. BOA. A) sonda NIT3, B) Mismo campo sondas EUB338mix, C) Mismo campo sondas EUB338mix y NIT3. 1000X.
4.2. CUANTIFICACIÓN DE LA SEÑAL DE HIBRIDACIÓN MEDIANTE
TRATAMIENTO DE IMAGEN
Para la cuantificación de las bacterias nitrificantes presentes en cada muestra se utilizó el programa desarrollado por Borras, (2008) tal como se indicó en el apartado de Materiales y Métodos. Este programa genera una hoja de cálculo en Excel, similar a la que se muestra en la Figura 22.
Este informe contiene los resultados de la cuantificación con las áreas ocupadas por las bacterias presentes para cada campo, la media del área para el total de la muestra, a desviación estándar y el error de medida.
En la hoja de cálculo generada también se presenta un gráfico de barras que indica el porcentaje de área hibridada. Las barras representan el porcentaje de bacterias diana obtenido para cada campo y la línea azul la media acumulada.
Figura 22. Modelo de hoja Excel de resultados extraídos del software de cuantificación
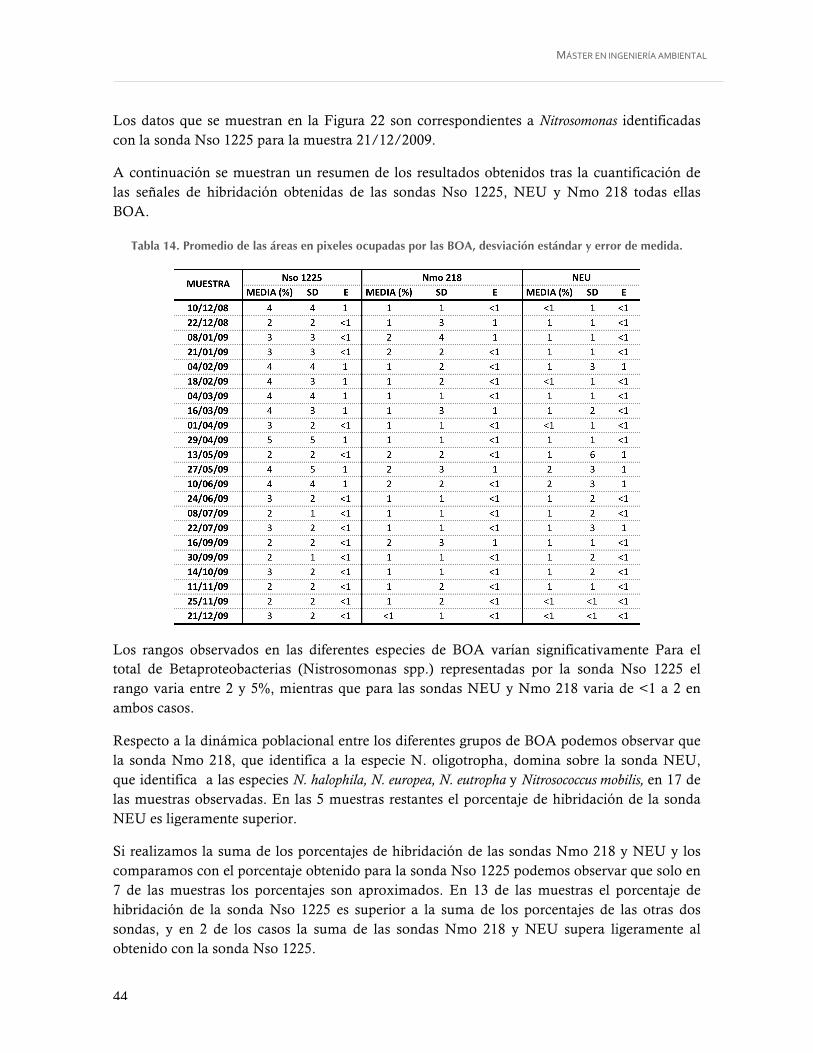
MÁSTER EN INGENIERÍA AMBIENTAL
44
Los datos que se muestran en la Figura 22 son correspondientes a Nitrosomonas identificadas con la sonda Nso 1225 para la muestra 21/12/2009.
A continuación se muestran un resumen de los resultados obtenidos tras la cuantificación de las señales de hibridación obtenidas de las sondas Nso 1225, NEU y Nmo 218 todas ellas BOA.
Tabla 14. Promedio de las áreas en pixeles ocupadas por las BOA, desviación estándar y error de medida.
Los rangos observados en las diferentes especies de BOA varían significativamente Para el total de Betaproteobacterias (Nistrosomonas spp.) representadas por la sonda Nso 1225 el rango varia entre 2 y 5%, mientras que para las sondas NEU y Nmo 218 varia de <1 a 2 en ambos casos.
Respecto a la dinámica poblacional entre los diferentes grupos de BOA podemos observar que la sonda Nmo 218, que identifica a la especie N. oligotropha, domina sobre la sonda NEU, que identifica a las especies N. halophila, N. europea, N. eutropha y Nitrosococcus mobilis, en 17 de las muestras observadas. En las 5 muestras restantes el porcentaje de hibridación de la sonda NEU es ligeramente superior.
Si realizamos la suma de los porcentajes de hibridación de las sondas Nmo 218 y NEU y los comparamos con el porcentaje obtenido para la sonda Nso 1225 podemos observar que solo en 7 de las muestras los porcentajes son aproximados. En 13 de las muestras el porcentaje de hibridación de la sonda Nso 1225 es superior a la suma de los porcentajes de las otras dos sondas, y en 2 de los casos la suma de las sondas Nmo 218 y NEU supera ligeramente al obtenido con la sonda Nso 1225.
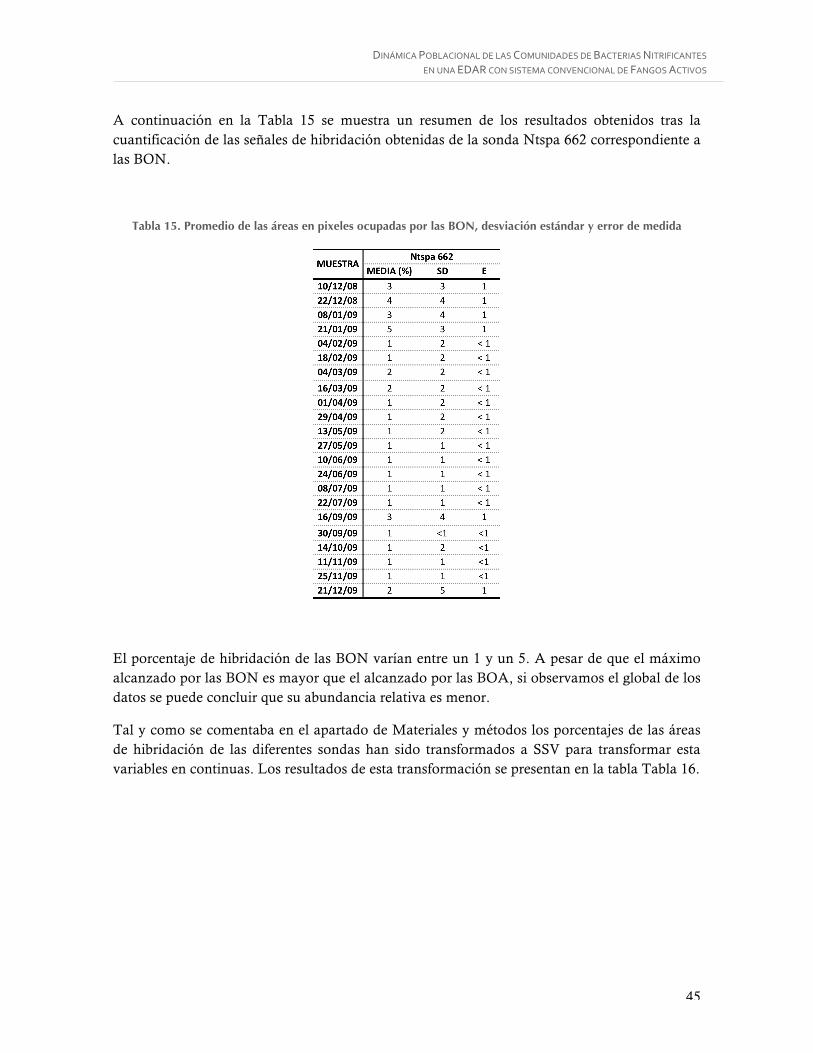
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
45
A continuación en la Tabla 15 se muestra un resumen de los resultados obtenidos tras la cuantificación de las señales de hibridación obtenidas de la sonda Ntspa 662 correspondiente a las BON.
Tabla 15. Promedio de las áreas en pixeles ocupadas por las BON, desviación estándar y error de medida
El porcentaje de hibridación de las BON varían entre un 1 y un 5. A pesar de que el máximo alcanzado por las BON es mayor que el alcanzado por las BOA, si observamos el global de los datos se puede concluir que su abundancia relativa es menor.
Tal y como se comentaba en el apartado de Materiales y métodos los porcentajes de las áreas de hibridación de las diferentes sondas han sido transformados a SSV para transformar esta variables en continuas. Los resultados de esta transformación se presentan en la tabla Tabla 16.
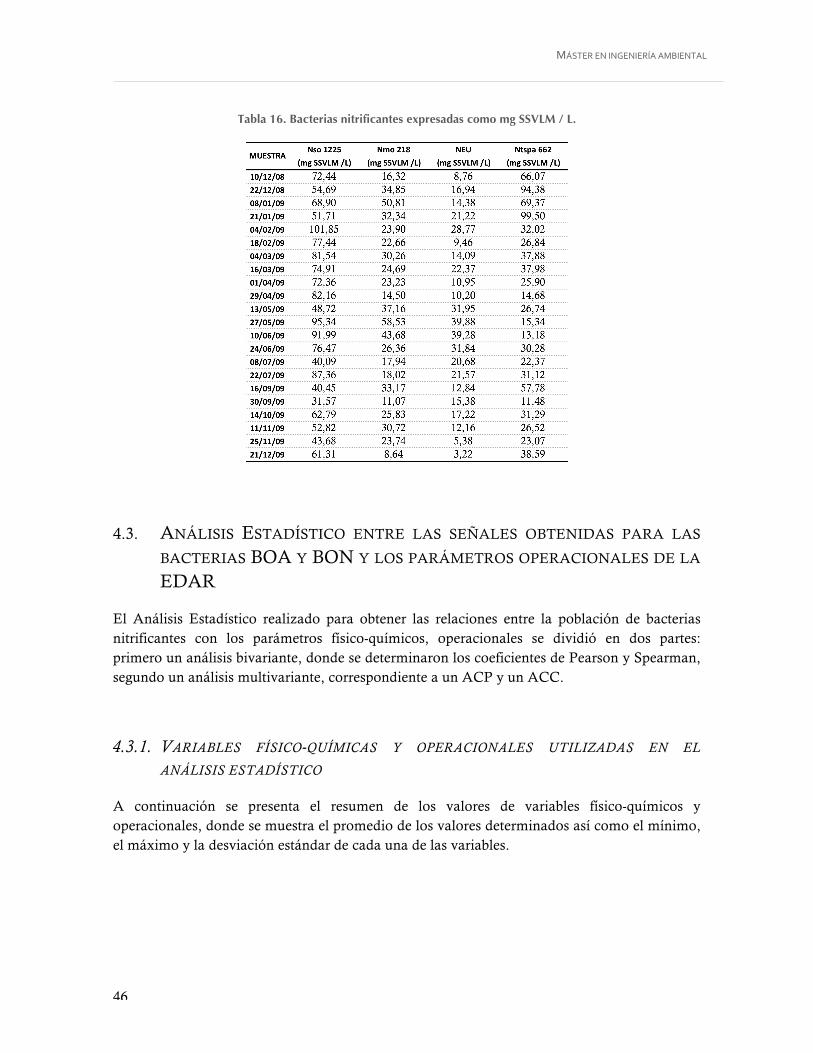
MÁSTER EN INGENIERÍA AMBIENTAL
46
Tabla 16. Bacterias nitrificantes expresadas como mg SSVLM / L.
4.3. ANÁLISIS ESTADÍSTICO ENTRE LAS SEÑALES OBTENIDAS PARA LAS
BACTERIAS BOA Y BON Y LOS PARÁMETROS OPERACIONALES DE LA
EDAR
El Análisis Estadístico realizado para obtener las relaciones entre la población de bacterias nitrificantes con los parámetros físico-químicos, operacionales se dividió en dos partes: primero un análisis bivariante, donde se determinaron los coeficientes de Pearson y Spearman, segundo un análisis multivariante, correspondiente a un ACP y un ACC.
4.3.1. VARIABLES FÍSICO-QUÍMICAS Y OPERACIONALES UTILIZADAS EN EL
ANÁLISIS ESTADÍSTICO
A continuación se presenta el resumen de los valores de variables físico-químicos y operacionales, donde se muestra el promedio de los valores determinados así como el mínimo, el máximo y la desviación estándar de cada una de las variables.
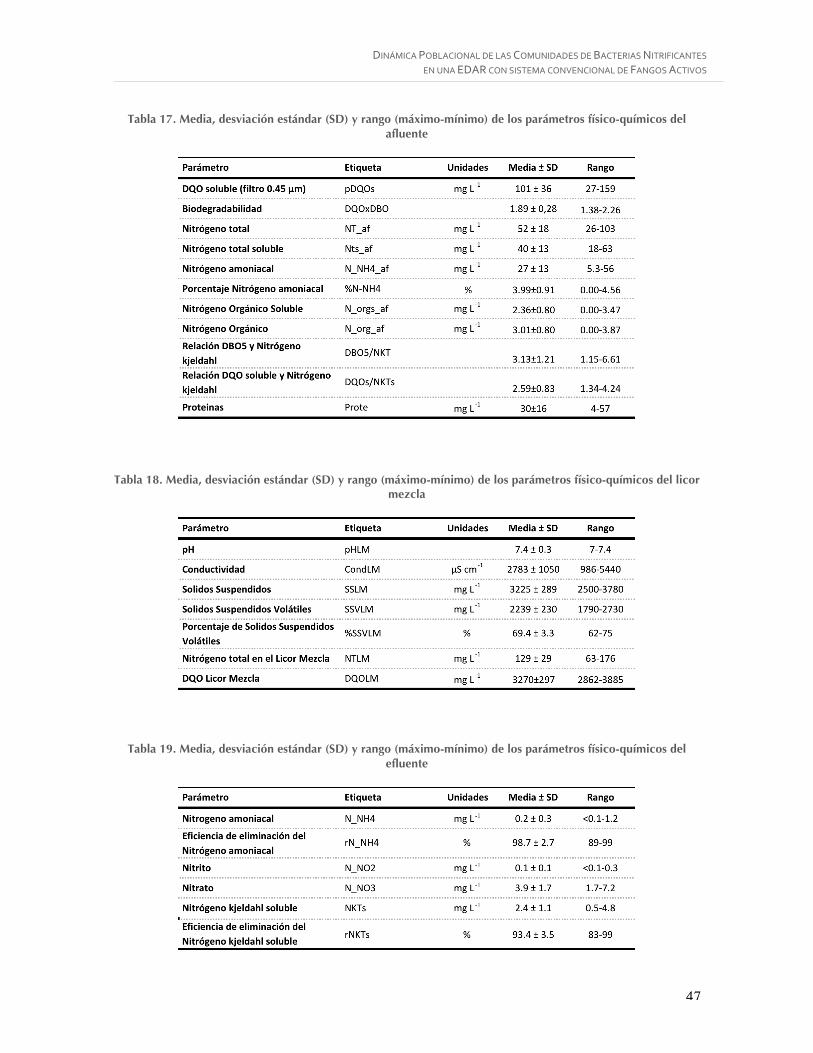
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
47
Tabla 17. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros físico-químicos del afluente
Tabla 18. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros físico-químicos del licor mezcla
Tabla 19. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros físico-químicos del efluente

MÁSTER EN INGENIERÍA AMBIENTAL
48
Tabla 20. Media, desviación estándar (SD) y rango (máximo-mínimo) de los parámetros operacionales
4.3.2. ANÁLISIS BIVARIANTE. COEFICIENTES DE CORRELACIÓN DE PEARSON Y
SPEARMAN
Los datos obtenidos tras la realización del análisis bivariante se presentan en 4 tablas diferentes que relacionan los resultados obtenidos en la cuantificación de las bacterias nitrificantes con las diferentes variables. La primera de ellas corresponde a las relaciones con las variables físico-químicas del afluente, la segunda corresponde a las variables del licor mezcla, la tercera a las del efluente y por últimos las variables operacionales. Los valores indicados en negrita y cursiva son los que presentan una significación estadística igual o superior al 95%.
En cada una de estas tablas se han correlacionado las diferentes variables de cada uno de los grupos mencionados en al párrafo anterior con el porcentaje de bacterias (correspondiente al área hibridada) las bacterias expresadas como mg de sólidos suspendidos volátiles.
El motivo de realizar las correlaciones de las diferentes variables tanto con el porcentaje de bacterias hibridadas y la cantidad de bacterias expresadas como mgSSVLM es comprobar cuál de estas dos variables correlacionan mejor con las variables físico-químicas y operacionales. Comprobándose que efectivamente correlacionan mejor los porcentajes de bacterias que sus respectivas transformaciones a mgSSVLM/L.
En la Tabla 21 se muestran las correlaciones de Pearson y Spearman entre las bacterias nitrificantes, los rendimientos de eliminación de nitrógeno y las variables físico-químicas del afluente, donde tanto la sonda NEU como la sonda Nmo 218, expresadas como porcentaje de hibridación correlacionan negativamente con la DQO soluble del afluente.
Se muestran correlaciones positivas entre el la sonda NEU expresada como mgSSVLM/L y la entrada de proteínas en el afluente.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
49
Tabla 21. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes, los rendimientos de eliminación de nitrógeno y las variables físico-químicas del afluente
En la Tabla 22 se muestran los resultados obtenidos para las correlaciones de Pearson y Spearman entre las bacterias nitrificantes, los rendimientos de eliminación de nitrógeno y las variables físico-químicas del licor mezcla.
Por lo que respecta a las especies expresada como porcentaje se observa como la sonda Nso 1225 correlaciona positivamente la cantidad de nitrógeno total del licor mezcla. Mientras que la sonda Nmo 218 presenta una correlación negativa con el pH del licor mezcla.
Las especies expresadas como mgSSVLM/L presenta correlaciones positivas para la sonda NEU y la sonda Nmo 218 con el nitrógeno total del licor mezcla.
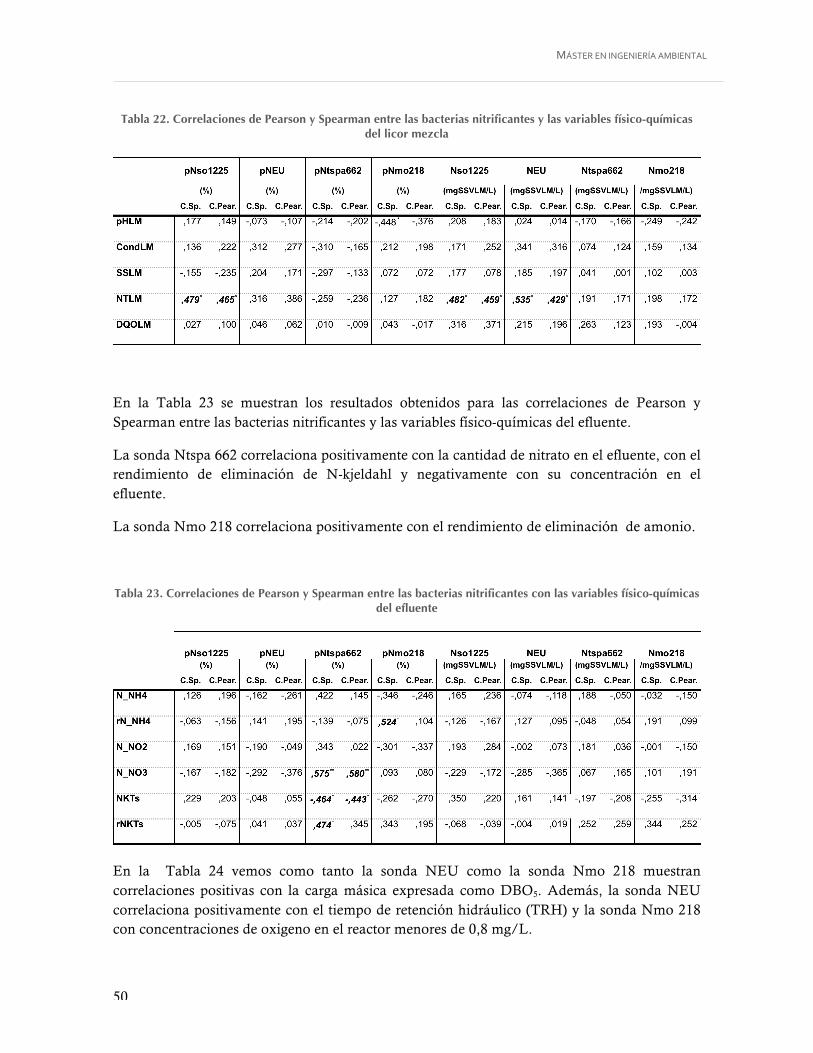
MÁSTER EN INGENIERÍA AMBIENTAL
50
Tabla 22. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes y las variables físico-químicas del licor mezcla
En la Tabla 23 se muestran los resultados obtenidos para las correlaciones de Pearson y Spearman entre las bacterias nitrificantes y las variables físico-químicas del efluente.
La sonda Ntspa 662 correlaciona positivamente con la cantidad de nitrato en el efluente, con el rendimiento de eliminación de N-kjeldahl y negativamente con su concentración en el efluente.
La sonda Nmo 218 correlaciona positivamente con el rendimiento de eliminación de amonio.
Tabla 23. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes con las variables físico-químicas del efluente
En la Tabla 24 vemos como tanto la sonda NEU como la sonda Nmo 218 muestran correlaciones positivas con la carga másica expresada como DBO5. Además, la sonda NEU correlaciona positivamente con el tiempo de retención hidráulico (TRH) y la sonda Nmo 218 con concentraciones de oxigeno en el reactor menores de 0,8 mg/L.
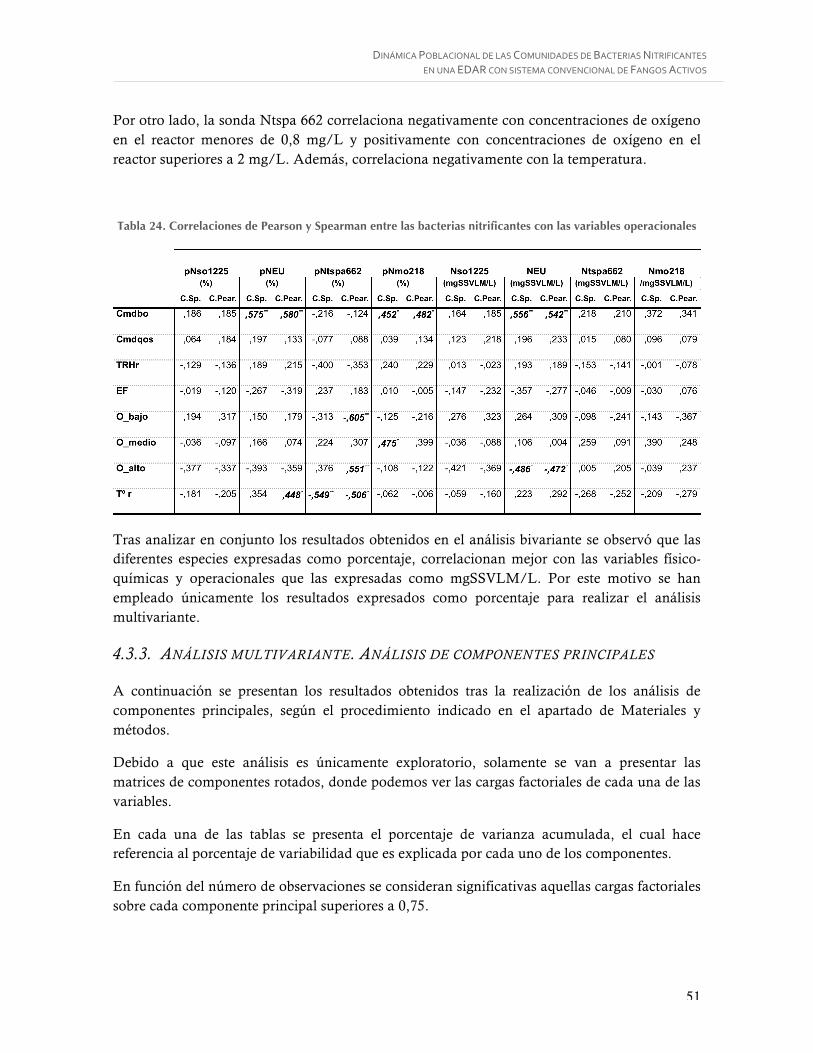
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
51
Por otro lado, la sonda Ntspa 662 correlaciona negativamente con concentraciones de oxígeno en el reactor menores de 0,8 mg/L y positivamente con concentraciones de oxígeno en el reactor superiores a 2 mg/L. Además, correlaciona negativamente con la temperatura.
Tabla 24. Correlaciones de Pearson y Spearman entre las bacterias nitrificantes con las variables operacionales
Tras analizar en conjunto los resultados obtenidos en el análisis bivariante se observó que las diferentes especies expresadas como porcentaje, correlacionan mejor con las variables físico-químicas y operacionales que las expresadas como mgSSVLM/L. Por este motivo se han empleado únicamente los resultados expresados como porcentaje para realizar el análisis multivariante.
4.3.3. ANÁLISIS MULTIVARIANTE. ANÁLISIS DE COMPONENTES PRINCIPALES
A continuación se presentan los resultados obtenidos tras la realización de los análisis de componentes principales, según el procedimiento indicado en el apartado de Materiales y métodos.
Debido a que este análisis es únicamente exploratorio, solamente se van a presentar las matrices de componentes rotados, donde podemos ver las cargas factoriales de cada una de las variables.
En cada una de las tablas se presenta el porcentaje de varianza acumulada, el cual hace referencia al porcentaje de variabilidad que es explicada por cada uno de los componentes.
En función del número de observaciones se consideran significativas aquellas cargas factoriales sobre cada componente principal superiores a 0,75.
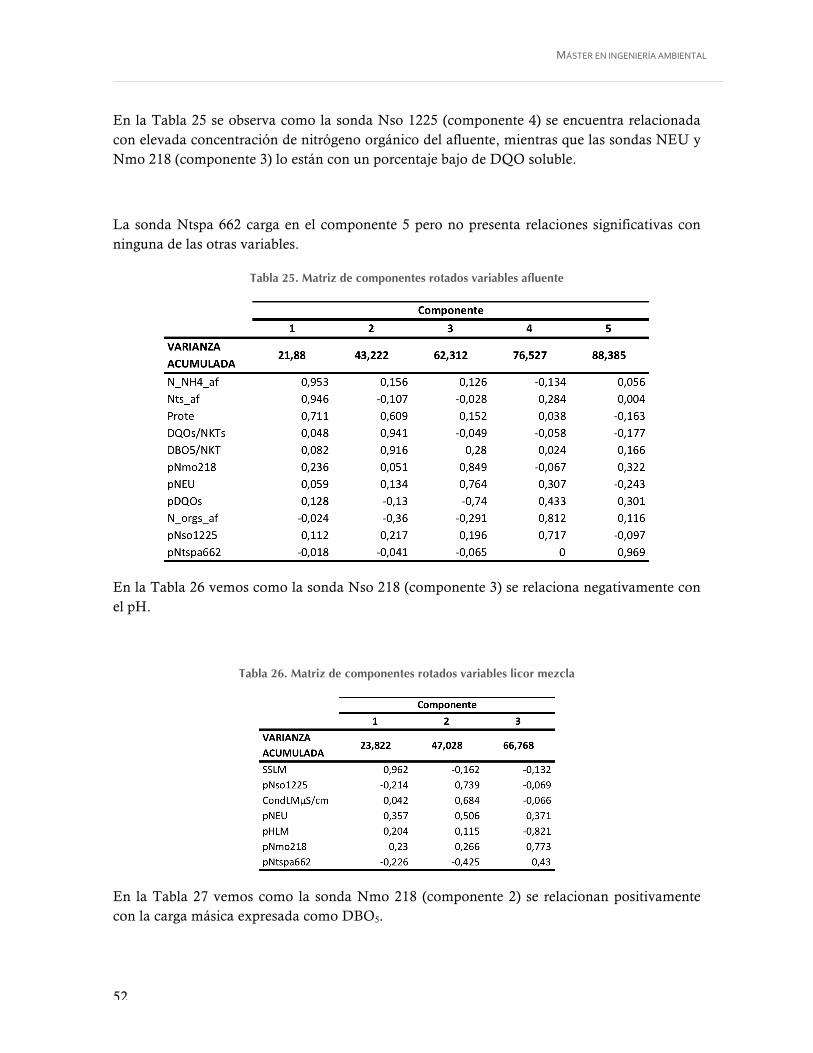
MÁSTER EN INGENIERÍA AMBIENTAL
52
En la Tabla 25 se observa como la sonda Nso 1225 (componente 4) se encuentra relacionada con elevada concentración de nitrógeno orgánico del afluente, mientras que las sondas NEU y Nmo 218 (componente 3) lo están con un porcentaje bajo de DQO soluble.
La sonda Ntspa 662 carga en el componente 5 pero no presenta relaciones significativas con ninguna de las otras variables.
Tabla 25. Matriz de componentes rotados variables afluente
En la Tabla 26 vemos como la sonda Nso 218 (componente 3) se relaciona negativamente con el pH.
Tabla 26. Matriz de componentes rotados variables licor mezcla
En la Tabla 27 vemos como la sonda Nmo 218 (componente 2) se relacionan positivamente con la carga másica expresada como DBO5.
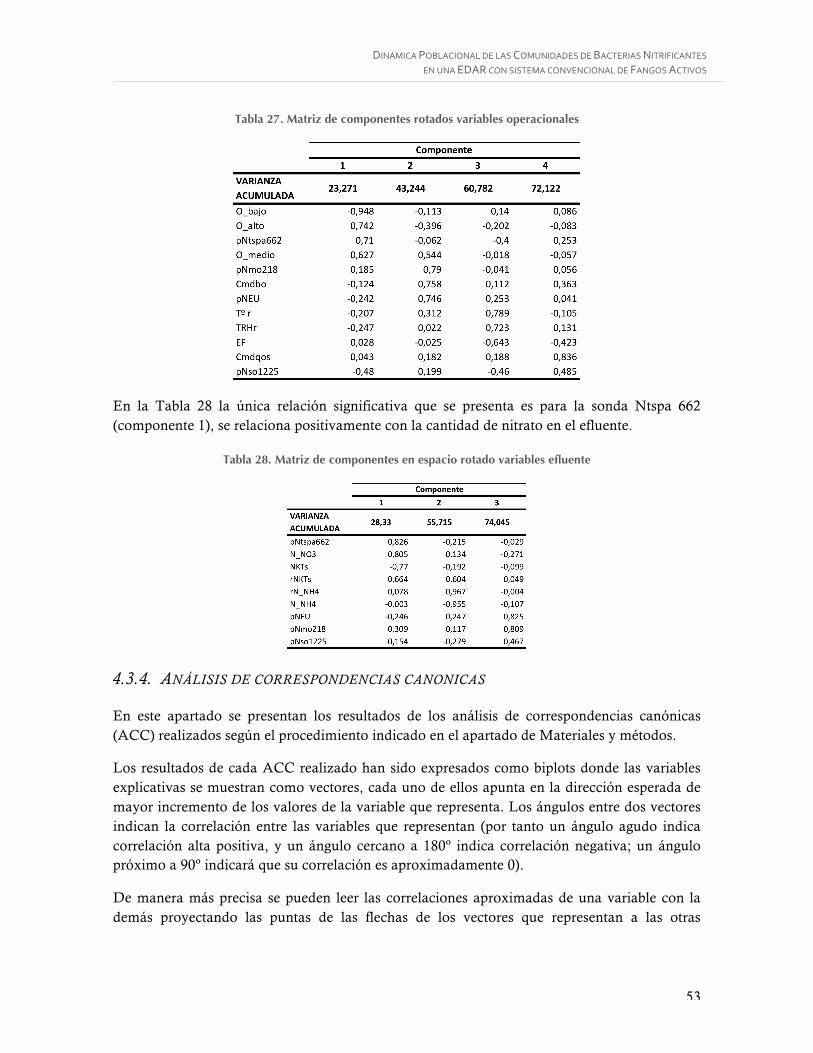
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
53
Tabla 27. Matriz de componentes rotados variables operacionales
En la Tabla 28 la única relación significativa que se presenta es para la sonda Ntspa 662 (componente 1), se relaciona positivamente con la cantidad de nitrato en el efluente.
Tabla 28. Matriz de componentes en espacio rotado variables efluente
4.3.4. ANÁLISIS DE CORRESPONDENCIAS CANONICAS
En este apartado se presentan los resultados de los análisis de correspondencias canónicas (ACC) realizados según el procedimiento indicado en el apartado de Materiales y métodos.
Los resultados de cada ACC realizado han sido expresados como biplots donde las variables explicativas se muestran como vectores, cada uno de ellos apunta en la dirección esperada de mayor incremento de los valores de la variable que representa. Los ángulos entre dos vectores indican la correlación entre las variables que representan (por tanto un ángulo agudo indica correlación alta positiva, y un ángulo cercano a 180º indica correlación negativa; un ángulo próximo a 90º indicará que su correlación es aproximadamente 0).
De manera más precisa se pueden leer las correlaciones aproximadas de una variable con la demás proyectando las puntas de las flechas de los vectores que representan a las otras
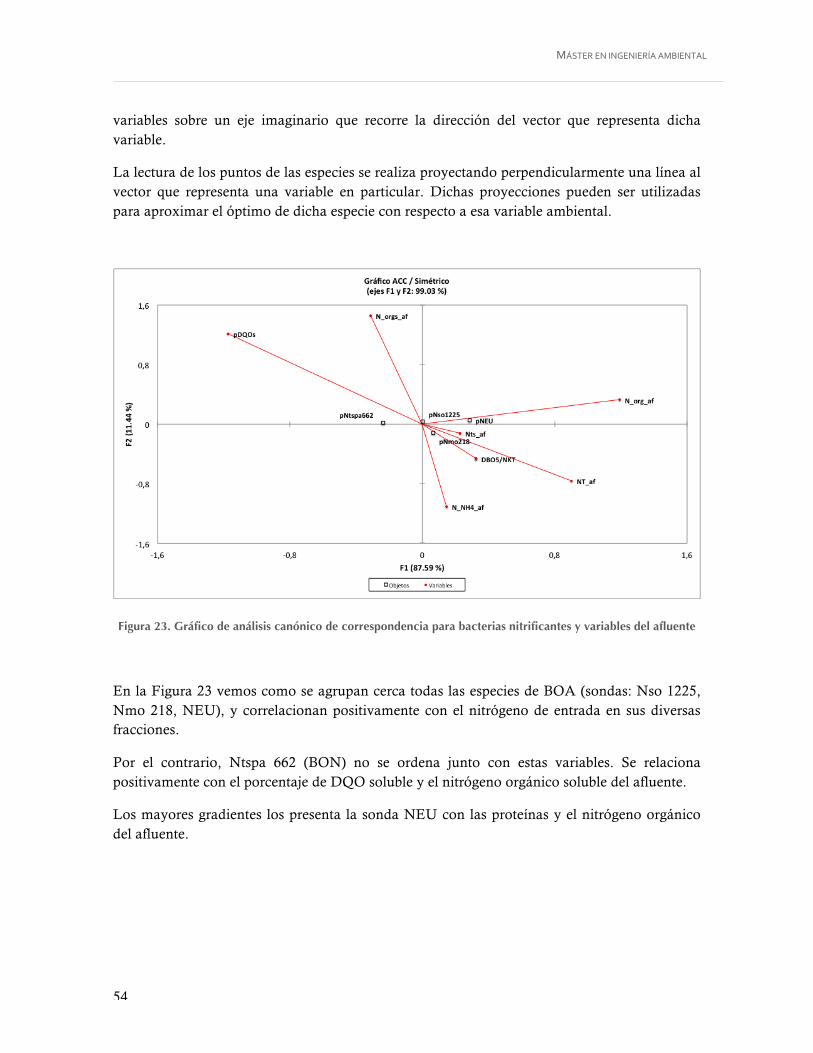
MÁSTER EN INGENIERÍA AMBIENTAL
54
variables sobre un eje imaginario que recorre la dirección del vector que representa dicha variable.
La lectura de los puntos de las especies se realiza proyectando perpendicularmente una línea al vector que representa una variable en particular. Dichas proyecciones pueden ser utilizadas para aproximar el óptimo de dicha especie con respecto a esa variable ambiental.
Figura 23. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables del afluente
En la Figura 23 vemos como se agrupan cerca todas las especies de BOA (sondas: Nso 1225, Nmo 218, NEU), y correlacionan positivamente con el nitrógeno de entrada en sus diversas fracciones.
Por el contrario, Ntspa 662 (BON) no se ordena junto con estas variables. Se relaciona positivamente con el porcentaje de DQO soluble y el nitrógeno orgánico soluble del afluente.
Los mayores gradientes los presenta la sonda NEU con las proteínas y el nitrógeno orgánico del afluente.
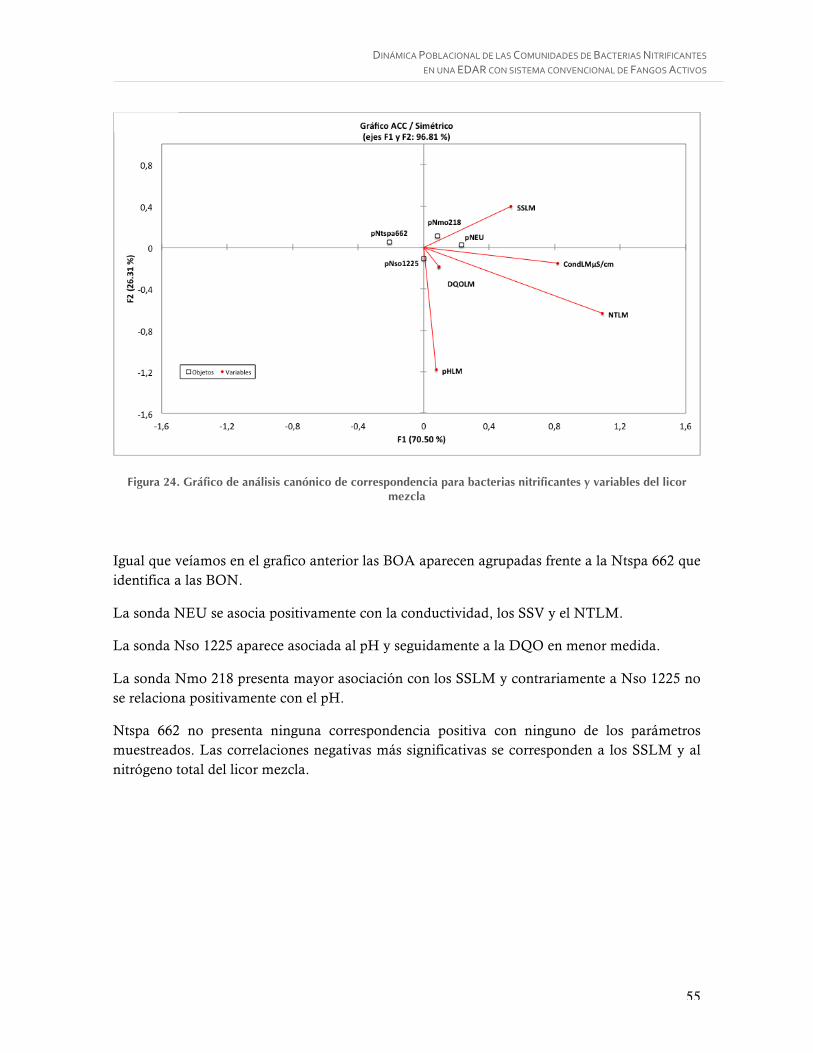
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
55
Figura 24. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables del licor mezcla
Igual que veíamos en el grafico anterior las BOA aparecen agrupadas frente a la Ntspa 662 que identifica a las BON.
La sonda NEU se asocia positivamente con la conductividad, los SSV y el NTLM.
La sonda Nso 1225 aparece asociada al pH y seguidamente a la DQO en menor medida.
La sonda Nmo 218 presenta mayor asociación con los SSLM y contrariamente a Nso 1225 no se relaciona positivamente con el pH.
Ntspa 662 no presenta ninguna correspondencia positiva con ninguno de los parámetros muestreados. Las correlaciones negativas más significativas se corresponden a los SSLM y al nitrógeno total del licor mezcla.
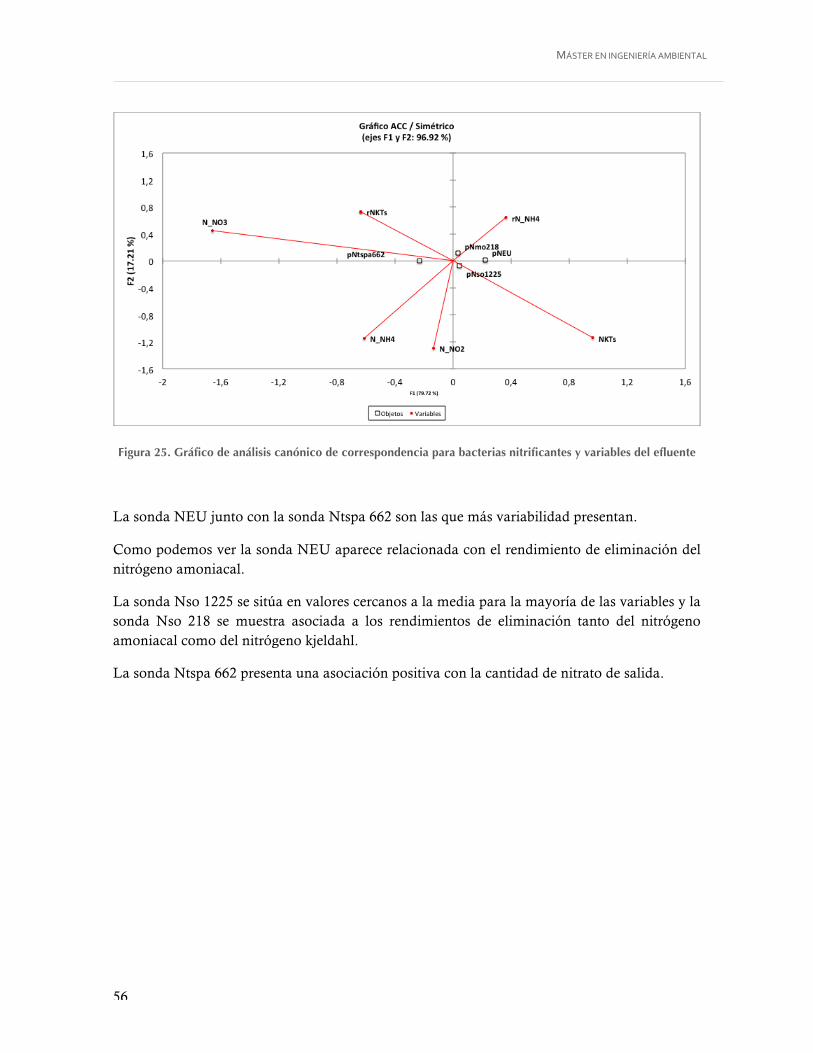
MÁSTER EN INGENIERÍA AMBIENTAL
56
Figura 25. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables del efluente
La sonda NEU junto con la sonda Ntspa 662 son las que más variabilidad presentan.
Como podemos ver la sonda NEU aparece relacionada con el rendimiento de eliminación del nitrógeno amoniacal.
La sonda Nso 1225 se sitúa en valores cercanos a la media para la mayoría de las variables y la sonda Nso 218 se muestra asociada a los rendimientos de eliminación tanto del nitrógeno amoniacal como del nitrógeno kjeldahl.
La sonda Ntspa 662 presenta una asociación positiva con la cantidad de nitrato de salida.
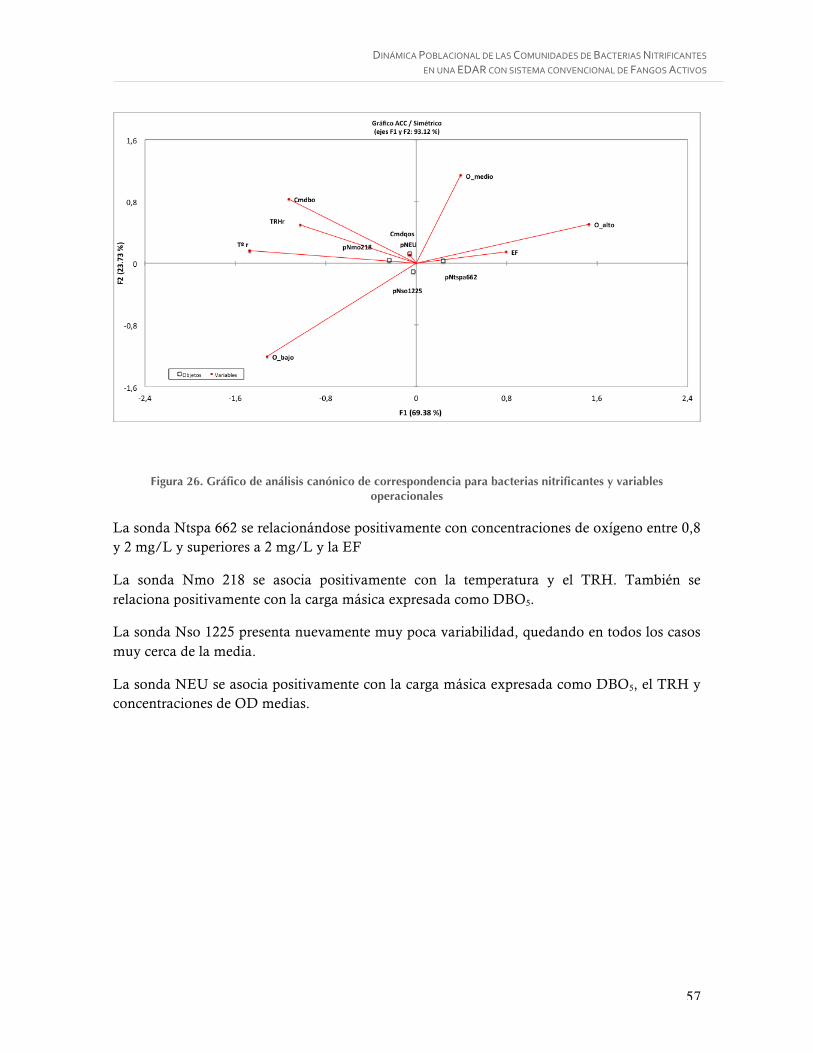
DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
57
Figura 26. Gráfico de análisis canónico de correspondencia para bacterias nitrificantes y variables operacionales
La sonda Ntspa 662 se relacionándose positivamente con concentraciones de oxígeno entre 0,8 y 2 mg/L y superiores a 2 mg/L y la EF
La sonda Nmo 218 se asocia positivamente con la temperatura y el TRH. También se relaciona positivamente con la carga másica expresada como DBO5.
La sonda Nso 1225 presenta nuevamente muy poca variabilidad, quedando en todos los casos muy cerca de la media.
La sonda NEU se asocia positivamente con la carga másica expresada como DBO5, el TRH y concentraciones de OD medias.

MÁSTER EN INGENIERÍA AMBIENTAL
58
5. DISCUSIÓN
Una de las consideraciones a tener en cuenta, previamente a esta discusión, es que la EDAR Denia-Ondara-Pedreguer durante el periodo de estudio presenta unos rendimientos de eliminación de nitrógeno muy elevados y en un rango muy estrecho de valores, por lo que el proceso de nitrificación en esta depuradora podría considerarse cercano al óptimo.
La escasa variabilidad, tanto en los rangos de los rendimientos de eliminación de nitrógeno, como en otras de las variables que intervienen en el proceso es un factor importante que afecta a los resultados obtenidos y a la interpretación de los mismos, debido a que nos movemos en un rango estrecho y siempre cercano al óptimo.
5.1. DINÁMICA POBLACIONAL
La comunidad de bacterias nitrificantes presentes en la EDAR Denia-Ondara-Pedreguer presenta una dinámica compleja y muy variable.
Según los resultados obtenidos las BOA representadas por Nitrosomonas spp. (identificada con la sonda Nso 1225) presentan una abundancia relativa entre un rango de 1-5 % del total de la población bacteriana. Las BON representadas por Nitrospira spp. (identificadas con la sonda Ntspa 662) presentan una abundancia relativa entre un rango de 1-5% del total de la población bacteriana. Se podría llegar a pensar que estos porcentajes son bajos teniendo en cuenta los elevados porcentajes de eliminación de nitrógeno amoniacal que se dan en esta depuradora (98,7%), sin embargo son acordes con otros estudios realizados. Por ejemplo Yu y Zhang (2012), caracterizaron la estructura microbiana y los genes nir expresados en la comunidad de una EDAR urbana con sistema de fangos activos en Hong Kong, encontrando que bacterias nitrificantes tales como Nitrosomonas y Nitrospira tenían una alta actividad a pesar de presentar una abundancia muy baja. También estudios realizados por Zhang et al., (2011b), detectaron una baja abundancia de BOA en fangos activos de diferentes EDAR.
En esta EDAR debido a los elevados tiempos de retención también se eliminará gran cantidad de nitrógeno por asimilación, incluso se puede llegar a producir desnitrificación en los flóculos más maduros. A pesar de ello no se descarta que otros géneros diferentes de las betaproteobacterias puedan contribuir a la actividad de nitrificación (Sánchez et al., 2013). Por ejemplo Nitrosococcus, dentro de las Gammaproteobacteria, ha sido identificado también como responsable de la nitrificación en sistemas de fangos activos (Juretschko et al., 1998; Raszka et al., 2011). También han sido identificadas como potenciales nitrificantes las AOA (ver apartado 1.3.1) o incluso organismos heterótrofos como Bacillus sp. (Kim et al., 2005), Alcaligenes faecalis (Liu et al., 2012) o Pseudomonas sp. (Su et al., 2006). A pesar de que se especula que estos organismos podrían jugar un papel importante en la nitrificación el objeto de este estudio se centra en observar la dinámica poblacional de las bacterias autótrofas nitrificantes (betaproteobacterias), consideradas como las mayores responsables de la oxidación del amonio, en relación con los parámetros operacionales.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
59
Dinámica poblacional de las BOA.
Tal y como se apuntaba en párrafos anteriores los resultados sugieren una compleja dinámica poblacional de las BOA. Solamente en 7 de las 22 muestras analizadas la suma de porcentaje de las sondas NEU (linaje N. europaea-Nc. mobilis) y la sonda Nmo 218 (N.oligotropha) dio valores similares a los porcentajes obtenidos con la sonda Nso 1225 (Nitrosomonas spp) (Tabla 14). En estos casos se podría decir que se ha conseguido caracterizar la comunidad de BOA presentes en la muestra.
En 13 de las muestras analizadas los porcentajes obtenidos con la sonda Nso 1225 fueron significativamente superiores a los obtenidos de la suma de las dos sondas anteriormente mencionadas. Esto puede ser debido a varios factores. En primer lugar apuntar al hecho de que todas las sondas vienen descritas con un porcentaje de fiabilidad, que indica qué porcentaje de individuos, pertenecientes al género o especie que se desea identificar, van a presentar señal de hibridación positiva con la sonda correspondiente. Esto significa que siempre existirá un porcentaje de individuos que no van a ser identificados. Por otra parte, algunos autores que sugieren que existen cepas de Nitrosomonas que no son detectadas por esta sonda y recomiendan el uso conjunto de la sonda Nmo 218 junto con la sonda cluster6a192. (Adamczyk, 2003; Almstrand et al., 2013) para una caracterización más completa de esta comunidad.
En las 2 muestras restantes el porcentaje obtenido por la sonda Nso 1225 fue menor que para la suma de las sondas NEU y Nmo 218. Tanto la sonda Nso 1225 como la sonda Nso 190 han sido ampliamente utilizadas para identificar la comunidad de betaproteobacterias BOA (Briesterfeld et al., 2003), no obstante en otros estudios se recomienda el uso conjunto de ambas sondas para la identificación de la comunidad de BOA (Almstrand, 2011), apuntando al hecho de que existen cepas de N. oligotropha (Hallin et al., 2005) y otras BOA (Lydmark, 2006), que no son identificadas con el uso separado de estas sondas.
Si comparamos ahora los porcentajes obtenidos con la sonda NEU y la sonda Nmo 218 se observa que N. oligotropha predomina respecto a las bacterias del linaje N. europaea/Nc. mobilis, en 17 de las 22 muestras analizadas. La dinámica observada entre estos dos géneros podría deberse a la disponibilidad de sustrato. Es bien sabido que la concentración de amonio es un factor muy importante en la composición de la comunidad de BOA, la afinidad por este sustrato varía entre miembros de los diferentes linajes de BOA pero tiende a ser estable dentro de un linaje específico (Pommerening-Röser et al., 1996). Como se observaba en la Figura 23tanto las especies englobadas en el linaje N.europaea-Nc. mobilis (sonda NEU) como N. oligotropha (sonda Nmo 218) presentan relaciones positivas con la concentración de las diferentes fracciones de nitrógeno en el afluente. Sin embargo los óptimos de N. europaea se sitúan mucho más arriba en el gradiente que los de N. oligotropha. Esto significa que N.europaea tiene mayor preferencia por concentraciones de sustrato elevadas que N. oligotropha. Estos resultados son acordes con estudios donde se ha observado, que N. oligotropha prevalece en

MÁSTER EN INGENIERÍA AMBIENTAL
60
ambientes con bajas concentraciones de sustrato (Bollmann y Laanbroek 2001; Limpiyakorn et al 2005; Otawa et al 2006; Wang et al 2010); mientras que ambientes ricos en sustrato están dominados por miembros del linaje la N. europaea - Nc. mobilis (Logemann et al 1998; Limpiyakorn et al 2005; Tan et al 2008.)
Los resultados de este estudio, sugieren que la estabilidad funcional no está necesariamente acompañada de una comunidad microbiana estable. Sin embargo, esto no significa que la dinámica de la comunidad microbiana no sea relevante para la estabilidad funcional. Gentil et al. (2007) evaluó la dinámica de la comunidad bacteriana en dos reactores de desnitrificación, encontrando durante el período de inestabilidad funcional con altas concentraciones de nitratos en el efluente, que la estructura de la comunidad cambiaba drásticamente, y este hecho correlacionaba significativamente con las variables físico-químicas del efluente. En general, dinámicas moderadas de la comunidad microbiana, necesarias para la adaptación de los organismos a las variaciones del entorno, en lugar de grandes cambios, son importantes para mantener el funcionamiento estable de los sistemas de tratamiento de aguas residuales (Miura et al., 2007). Briones et al. (2003) indican que la estabilidad funcional no se determina por la estructura de la comunidad microbiana, pero está garantizada por la presencia de un depósito de especies que pueden realizar la misma función ecológica. Como podemos observar la dinámica poblacional de este sistema es compleja y serían necesarios más estudios para desvelar cuales son las especies presentes en esta comunidad y en que porcentajes se sitúan. Estos resultados confirman observaciones realizadas por diferentes autores en sistemas de nitrificación (Daims et al., 2001; Wang et al., 2009; Wang et al., 2012; Wittebolle et al. 2008), los cuales apuntan que la existencia de una dinámica poblacional diversa, donde existen diversas especies de BOA o BON, frente a la presencia de un solo linaje de un grupo funcional, es la mejor manera de asegurar un buen sistema de nitrificación con elevados rendimientos tal y como es el caso que se presenta, ya que, a pesar de tener oscilaciones significativas en las poblaciones de BOA los rendimientos de nitrificación siguen situándose en valores óptimos.
Dinámica poblacional de las BON
En este estudio se ha identificado Nitrospira como el género dominante en la oxidación del nitrito a nitrato, tras observar que en la hibridación de las muestras con la sonda NIT3, (Nitrobacter), únicamente en 7 de las 22 muestras analizadas las hibridaciones resultaron positivas y, en todos estos la concentración de células de Nitrobacter fue muy baja. Estos mismos resultados han sido obtenidos en otras investigaciones tras la aplicación de la técnica FISH en estudios en plantas a escala real, que confirmaron que Nitrospira era la especie clave en este proceso (Juretschko et al., 1998; Schramm et al., 1998; Gieseke et al., 2003) en la mayoría de EDAR y no Nitrobacter.
Son dos los factores que parecen tener mayor influencia en la distribución de Nitrobacter y Nitrospira en las EDAR, las concentraciones de nitrito (Kim et al, 2006) y la concentración de oxígeno (Schramm et al, 1999). Se ha demostrado que Nitrobacter incrementa su abundancia

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
61
con el aumento de las concentraciones de nitrito (Vazquez-Padín et al., 2009). Huang y colaboradores (2010) realizaron estudios en un tanque aireado de fangos activos en California, donde observaron que el aumento de la concentración de OD fue adecuado para el crecimiento de Nitrobacter, mientras que Nitrospira se enriqueció de forma selectiva cuando las concentraciones de OD fueron menores de <1,0 mg /l. Estos resultados son acordes con investigaciones previas realizadas por Schramm (2003), donde se indica que la presencia de Nitrospira se ve favorecida por concentraciones de OD bajas.
5.1. RENDIMIENTOS DE ELIMINACIÓN
Se han obtenido asociaciones positivas con los rendimientos de eliminación del N-kjeldahl y N-amoniacal y la población de N. oligotropha (Figura 25), que ha sido la especie predominante durante el periodo de estudio, lo cual sería significativo de la buena señal de hibridación detectada de las sondas, así como la adecuada metodología empleada en el tratamiento y cuantificación de las imágenes (Borras, 2008).
Por el contrario las especies pertenecientes al linaje N. europaea/Nc.mobilis únicamente muestran asociación positiva con el rendimiento del nitrógeno amoniacal. La población global de Nitrosomonas no muestra ninguna de estas asociaciones, esto puede ser debido, como apuntábamos en el primer apartado de esta discusión al hecho de que existe una dinámica poblacional compleja y muy variable, además de que el estrecho rango de rendimientos de eliminación presentados limita el establecimiento de estas relaciones. Otro de los factores que pueden tener relevancia en este punto es que la elevada EF permite, que gran parte del nitrógeno sea eliminado por asimilación y a que no se puede descartar la presencia de otros organismos nitrificantes en el sistema que también contribuyen en el proceso.
El genero Nitrospira aparece asociado a la concentración de nitrato en el efluente. Aunque este parámetro dependerá del correcto funcionamiento del proceso de desnitrificación, sí tiene sentido que a mayor cantidad de BON mayor sea la cantidad de nitrato producida.
5.2. VARIABLES DEL AFLUENTE.
El estudio estadístico realizado no muestra relaciones significativas entre las variables del afluente y la dinámica poblacional de las bacterias nitrificantes de este sistema (Tabla 21, Tabla 25)
En el ACC (Figura 23) se observa como las BOA se asocian positivamente con los valores de entrada de las diferentes fracciones del nitrógeno, siendo las BOA del linaje N. europaea/Nc. mobilis, las que parecen tener una mayor preferencia por valores altos de estas variables(ver apartado 5.1 Dinámica poblacional BOA).

MÁSTER EN INGENIERÍA AMBIENTAL
62
Por otra parte la presencia de Nitrospira (NOB) parece estar más influenciada por el porcentaje de DQO soluble de entrada. En este sistema Nitrospira presenta su máxima abundancia cuando esta variable tiene valores ligeramente superiores a la media registrada en la EDAR. Teniendo en cuenta que el valor medio de la DQO soluble de entrada a la EDAR Denia-Ondara-Pedreguer durante el periodo de estudio fue de 101 ± 36 se puede afirmar que Nitrospira se verá favorecida con cargas medias-débiles de DQO. Esto es debido a que para elevados porcentajes de DQO las bacterias heterótrofas entran en competición con las autótrofas por el oxígeno, viendo estas su crecimiento limitado, no obstante el porcentaje de DQO debe ser el suficiente para que asegure el suficiente sustrato para el crecimiento de los microorganismos.
5.3. VARIABLES OPERACIONALES.
Temperatura y EF
La temperatura y la edad del fango (EF) han sido descritas como dos de las variables más influyentes en el proceso de nitrificación debido a la baja tasa de crecimiento de las bacterias nitrificantes (Gerardi, 2002). El rango de temperaturas óptimas para el proceso de nitrificación se sitúa entre los 28º y 32ºC. Para temperaturas entorno a los 16ºC se ha estimado que la velocidad del proceso de nitrificación se reduce a la mitad (Gerardi, 2002). Para valores menores la velocidad del proceso presenta una reducción drástica, siendo muy difícil que se lleve a cabo la nitrificación; en estas situaciones es necesario trabajar con EF altas para que el proceso pueda llevarse a cabo (González et al., 2010). Müller et al. (2003), indican que la EF mínima para que se produzca nitrificación es entorno a los 7 días, aunque los cambios de temperatura afectan a este parámetro. El rango de temperaturas de la EDAR durante el periodo de estudio osciló entre los 11ºC y los 26ºC, mientras que los valores de la EF se situaron entre 10 y 31 días. Tal y como podemos observar en la Figura 26 las variables EF y temperatura están enfrentadas con un ángulo de 180º lo que indica una relación inversa entre ambas, a mayor temperatura menor EF. Considerando los elevados rendimientos de eliminación de nitrógeno podríamos afirmar, que durante el periodo de estudio esta EDAR mantuvo bien compensadas estas variables operacionales y, por tanto obtener excelentes rendimientos de eliminación.
El ACC (Figura 26)presentado muestra como la población de BOA está asociada positivamente a la temperatura. Entre las especies estudiadas las pertenecientes a la especie N. oligotropha son las que mayor relación presentan con esta variable. Esto significa que la abundancia de esta especie se asocia a valores de temperatura superiores a la media registrada en la EDAR, mientras que las especies pertenecientes al linaje N. europaea/Nc. mobilis parecen ser menos dependientes. De acuerdo con estos resultados, Park et al. (2009) demostraron que la temperatura fue el factor más influyente en la comunidad de BOA presente en un reactor de tratamiento de AR salinas y que la reducción de la tasa de crecimiento no es la única repercusión de la disminución de la temperatura, esta afecta además a la actividad nitrificante y produce cambios en la composición de especies de la comunidad de BOA (Park et al., 2008).

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
63
Por el contrario Limpiyakorn et al (2005) evaluó la respuesta de la comunidad de BOA respecto a las variables estacionales en 12 EDAR, donde los resultados mostraron que la temperatura no produjo efectos detectables en las comunidades de BOA.
Por lo que respecta al género Nitrospira dentro de las BON, su presencia está más relacionada con EF elevadas. Esto puede ser debido a que Nitrospira crece más lentamente que Nitrosomonas a igual temperatura (Yu et al, 2010). Por tanto Nitrospira será más resistente frente a descensos de temperatura, que Nitrobacter, mientras se mantengan EF adecuadas. Este resultado es similar al obtenido por Gerardi , (2002), el cual sugiere que la inhibición por bajas temperaturas es mayor para Nitrobacter que para Nitrospira.
Tiempo de Retención Hidráulico
El TRH se relaciona positivamente con las especies de BOA detectadas, aunque no parece que sea factor determinante en el desarrollo de estas poblaciones.
Las asociaciones positivas podrían ser debidas al hecho de que cuando los flóculos son lo suficientemente grandes, rasgo característico de altas EF, permiten cierta retención de biomasa. Este fenómeno podría dar una ventaja competitiva a las bacterias de crecimiento lento frente a otras especies.
Carga másica
Aunque las bacterias nitrificantes sean organismos autótrofos, lo que significa que no usan materia orgánica (DBO) como fuente de carbono, la concentración de DBO juega un papel fundamental en la convivencia de las bacterias nitrificantes y las bacterias heterótrofas presentes en la comunidad, afectando a la composición de estas comunidades en el reactor. En sistemas convencionales esto es debido a que cuando se incrementa la posibilidad de acceso de la materia orgánica se favorece el crecimiento de la población heterótrofa, con el consiguiente consumo de OD asociado, lo que es un factor limitante para la población de bacterias autótrofas.
Los estudios realizados en este trabajo muestran una relación positiva entre la población de BOA y la carga másica, frente a estudios realizados por otros autores en plantas a escala real donde se han encontrado correlaciones negativas entre esta variable y la población de bacterias nitrificantes (Xia et al., 2008; Zornoza et al., 2012).
Las diferencias observadas en ambos casos son debidas al rango de valores que la carga másica toma en este sistema, el cual está comprendido entre 0,03 y 0,19 (kgDBO5/Kg SSVLM·d). Por tanto cargas másicas elevadas de este sistema se corresponde con cargas medias-bajas en sistemas convencionales. Valores más bajo de esta variable podrían convertirse en factores limitantes del crecimiento.

MÁSTER EN INGENIERÍA AMBIENTAL
64
Oxigeno
Los valores de oxígeno aportados por la EDAR están divididos en tres rangos: O2 <0,8 mg/L (OD bajo), 0,8<O2<2 mg/L (OD medio) y O2>2 mg/L (OD alto), e indican el porcentaje de tiempo en el que la concentración de oxígeno se encontró en estos valores.
En el ACC donde se muestra la relación entre el oxígeno y estos valores no se muestra relación significativa entre estas variables y, las poblaciones de bacterias nitrificantes. Se observa que tanto la sonda NEU, Nmo 218 y Ntspa se asocian positivamente con los tiempos en que el oxígeno en el reactor permaneció entre una concentración media y alta, mientras que la sonda Nso 1225 presenta una tendencia opuesta. Sin embargo no se pueden aportar datos sobre cual fue el valor promedio de la concentración de oxígeno, ni cuales fueron los máximos o mínimos alcanzados por esta variable en el periodo de estudio. Es bien sabido que el oxigeno es un parámetro clave para el funcionamiento del proceso de nitrificación. Recientes estudios llevados a cabo por Wang, et al. (2012), en los que se evaluaba la correlación de 10 variables operacionales y ambientales mediante ACC, determinaron que el OD es una variable explicativa importante que afecta a la dinámica de la comunidad BOA. Esto concuerda con los hallazgos de Wells et al. (2009) realizados en una EDAR municipal, que sugieren que el OD es una de las variables más influyentes en la dinámica de la comunidad BOA. Park et al. (2004) comparó las estructuras de la comunidad BOA en dos birreactores a escala de laboratorio operados con concentraciones altas de OD (8,5 mg / L) y bajas (0,24 y 0,12 mg / L) durante un período de 300 días. Los resultados mostraron que los miembros BOA presentes en los birreactores de OD bajo y OD alto eran en ambos casos del linaje N. europaea – Nc. mobilis, pero eran filogenéticamente diferentes, lo que indica que el OD ejerce una presión selectiva significativa en las comunidades BOA. De acuerdo con las aportaciones de Park (2004) este estudio muestra ciertas diferencias dentro de la comunidad de BOA, parece que existan especies más dependientes de esta variable que otras dentro de este mismo género, pues como vemos en el gráfico la sonda NEU presenta un gradiente ligeramente superior al mostrado por la sonda Nmo 218.
Sin embargo, el estudio de los datos estadísticos no sugiere que el oxigeno no sea uno de las variables que mejor explica la abundancia de la población de nitrificantes. De acuerdo con estos resultados Park et al. (2008) demostraron que la concentración de oxígeno no influyó en la comunidad BOA, sino más bien en su actividad. Resultados similares han sido obtenidos en varias EDAR a escala real en Tokio por Limpiyakorn et al. (2005). Por otro lado, Lydmark et al. (2007) llegó a la conclusión de que no había datos suficientes para permitir correlaciones entre las agrupaciones BOA y los niveles de oxígeno.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
65
5.4. VARIABLES DEL LICOR MEZCLA
Conductividad
Las especies pertenecientes al linaje N.europaea/Nc.mobilis parecen ser las más influenciadas por las variaciones de la conductividad, estudios anteriores (Zornoza et al., 2012) también han relacionado esta variable con la población de bacterias nitrificantes, indicando que valores muy elevados de esta variable interfieren en el crecimiento de estas especies.
pH
El pH se considera una variable que tiene influencia sobre la tasa de crecimiento de las bacterias nitrificantes. Se ha observado que la tasa máxima de nitrificación se produce entre valores de 7,2 a 9,0 aproximadamente, a valores inferiores a 6,5 la velocidad de nitrificación se reduce de forma brusca (González et al, 2010).
En este estudio se ha observado que las poblaciones más afectadas por los descensos del pH son Nitrospira y N. Oligotropha.

MÁSTER EN INGENIERÍA AMBIENTAL
66
6. CONCLUSIONES
Las principales conclusiones que se pueden extraer de este trabajo son:
- La especie N. oligotropha ha sido identificada como la especie dominante de la comunidad de BOA. La dominancia de este género se debe a las concentraciones medias y bajas de sustrato, que favorecen su crecimiento frente a las especies del linaje N. europaea/Nc. mobilis.
- El género Nitrospira ha sido el dominante dentro de la comunidad de NOB. La dominancia de esta especie se debe a las concentraciones medias y bajas de sustrato, que favorecen su crecimiento frente a las especies del linaje N. europaea/Nc. mobilis
- Los resultados del análisis estadístico muestran que la dinámica poblacional esta influenciada por las concentraciones de nitrógeno del afluente, la temperatura, la edad del fango y la carga másica.
- Los resultados de este estudio sugieren que la estabilidad funcional no está necesariamente acompañada de una comunidad microbiana estable. La existencia de una dinámica poblacional diversa, frente a la presencia de un único linaje de un grupo funcional es una buena manera de asegurar un sistema de nitrificación estable.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
67
7. REFERENCIAS
Adamczyk J, Hesselsoe M, Iversen N, Horn M, Lehner A, Nielsen PH, Schloter M, Roslev P,Wagner M (2003) The isotope array, a new tool that employs substrate-mediated labeling of rRNA for determination of microbial community structure and function. Appl Environ Microbiol 69:6875–6887.
Aguado, D. 2004. Aplicación de métodos estadísticos multivariantes para la modelación y la monitorización de un reactor discontinuo secuencial para el tratamiento de aguas residuales. Tesis Doctoral. Universidad de Valencia.
Alawi, M., Lipski, A., Sanders, T., Eva Maria, P., and Spieck, E. (2007) Cultivation of a novel cold-adapted nitrite oxidiz- ing betaproteobacterium from the Siberian Arctic. ISME J 1: 256–264.
Almstrand, R., Lydmark, P., Lindgren, P.-E., Sörensson, F., & Hermansson, M. (2013). Dynamics of specific ammonia-oxidizing bacterial populations and nitrification in response to controlled shifts of ammonium concentrations in wastewater. Applied microbiology and biotechnology, 97(5), 2183–91. doi:10.1007/s00253-012-4047-7.
Almstrand R, Lydmark P, Sörensson F, HermanssonM (2011) Nitrification potential and population dynamics of nitrifying bacterial biofilms in response to controlled shifts of ammoniumconcentrations in wastewater trickling filters. Biores Technol 201:7685–7691.
Alonso, J., Cuesta, G., Ramírez, G., Morenilla, J., Bernácer, I., Lloret, R. (2009). Manual de Técnicas Avanzadas para la Identificación y Control de Bacterias Filamentosas. P. 21-36. Ed. Epsar-Generalitat Valenciana. Valencia, España.
APHA, 2005. Standard Methods for the Examination of Water and Wastewater, 21st ed. American public health Association, Washington DC, USA.
Beman JM, Popp BN, Francis CA (2008) Molecular and biogeochemical evidence for ammonia oxidation by marine Crenarchaeota in the Gulf of California. ISME J 2: 429–441. 16.
Biesterfeld S, Farmer G, Russell P, Figueroa L (2003) Effect of alkalinity type and concentration on nitrifying biofilm activity. Water Environ Res 75(3):196–204.
Bock, E., Wagner, M. (2001) Oxidation of inorganic nitrogen compounds as an energy source. In: Dworkin H et al (eds) The prokaryotes: an evolving electronic resource for the microbiological community, release 3:7, 3rd edn. Springer-Verlag, New York.
Bock, E., Wagner, M. (2006) Oxidation of inorganic nitrogen com- pounds as energy source. In: Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E (eds) The prokaryotes. Volume 2: ecophysiology and biochemistry. Springer, New York, pp 457– 495.
Bock E., Koops H.-P., Ahlers B., Harms H. (1992) Oxidation of inorganic nitrogen compounds as energy source. in The prokaryotes, eds Balows H., Trüper H. G., Dworkin M., Harder W., Schleifer K.-H. (Springer Verlag, New York, N.Y), 2nd ed. pp 414–430.

MÁSTER EN INGENIERÍA AMBIENTAL
68
Bollmann A, Laanbroek HJ (2001) Continuous culture enrichments of ammonia-oxidizing bacteria at low ammonium concentrations. FEMS Microbiol Ecol 37:211–221.
Borrás, L. 2008. Técnicas microbiológicas aplicadas a la identificación y cuantificación de microorganismos presentes en sistemas EBPR. Tesis Doctoral. Universidad Politécnica de Valencia. Valencia. España.
Briones A, Raskin L (2003) Diversity and dynamics of microbial communities in engineered environments and their implications for process stability. Curr Opin Biotech 14: 270–276.
Conover, W. 1998. Practical nonparametric statistics. 3era. edición John Wiley & Sons. Nueva York.
Costa, E., Pérez, J., and Freft, J.U. (2006) Why is metabolic labour divided in nitrification? Trends in Microbiology 14: 213:219.
Daims, H., Maixner, F., Schmid, M.C. 2009. the Nitrifying microbes: Ammonia oxidizers, nitrite oxidizers, and anaerobic ammonium oxidizers. En: Nielsen, P., Daims, H y Lemmer, H. 2009. FISH Handbook for Biological Wastewater Treatment: Identification and quatification of microorganisms in activated sludge and biofilms by FISH. p 9-17. Ed. IWA Publishing. Londres, Reino Unido.
Daims H, Nielsen JL, Nielsen PH, Schleifer K-H, Wagner M (2001a) In situ characterization of Nitrospira-like nitriteoxidizing bacteria active in wastewater treatment plants. Appl Environ Microbiol 67:5273–5284.
Daims H, Purkhold U, Bjerrum L, Arnold E, Wilderer PA, Wagner M (2001b) Nitrification in sequencing biofilm batch reactors: lessons from molecular approaches. Water Sci Technol 43:9–18.
Ehrich, S., Behrens, D., Lebedeva, E., Ludwig,W., and Bock, E. (1995) A new obligately chemolithoautotrophic nitrite- oxidizing bacterium, Nitrospira-moscoviensis sp-nov and its phylogenetic relationship. Arch Microbiol 164: 16–23.
Esteban, G., Tellez, C., Bautista, L.M., (1991). Dynamics of ciliated protozoa communities in activated sludge process. Water Res. 25:967–972.
Foesel, B.U., Gieseke, A., Schwermer, C., Stief, P., Koch, L., Cytryn, E., et al. (2007) Nitrosomonas Nm143-like ammonia oxidizers and Nitrospira marina-like nitrite oxidiz- ers dominate the nitrifier community in a marine aquacul- ture biofilm. FEMS Microbiol Ecol 63: 192–204.
Fukushima, T, Whang L, Chiang T, Lin Y, Chevalier L, Chen M, Wu Y (2013) Nitrifying bacterial community structures and their nitrification performance under sufficient and limited inorganic carbon conditions. Appl Microbiol Biotechnol 97:6513-6523.
Gentile ME, Jessup CM, Nyman JL, Criddle CS (2007) Correlation of functional instability and community dynamics in denitrifying dispersed-growth reactors. App Env Microbiol 73: 680–690.
Gerardi, M. (2002). Nitrification and denitrification in the activated sludge process. Wiley- Interscience. Nueva York. Estados Unidos. Grady.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
69
Gieseke, A., Bjerrum, L., Wagner, M., Amman, R. (2003) Structure and activity of multiple nitrifying bacterial populations co-existing in a biofilm. Environmental Microbiology 5:355-369.
Gieseke, A., Purkhold, U., Wagner, M., Amann, R y Schramm, A. (2000). Community Structure and Activity Dynamics of Nitrifying Bacteria in a Phosphate-Removing Biofilm. Applied and Environmental Microbiology. 67: 1351-1362.
González, P., Quintans, P., Vizcaíno, M., Miguel, R., González, J., Pérez, J y García, R. (2010). Estudio de la inhibición del proceso de nitrificación como consecuencia de la acumulación de metales en el fango biológico de la EDAR de León y su alfoz. Tecnología del Agua. 322: 28-38.
Guisasola A, Petzet S, Baeza JA, Carrera J, Lafuente J (2007) Inorganic carbon limitations on nitrification: experimental assessment and modelling. Water Res 41(2):277–286.
Hallin, S., Lydmark, P., Kokalj, S., Hermansson, M., Sorensson, F., Jarvis, A., Lindgren, P.E., 2005. Community survey of ammonia-oxidizing bacteria in full-scale activated sludge processes with different solids retention time. J. Appl. Microbiol. 99 (3), 629–640.
Head, I.M., Hiorns, W.D., Embley, T.M., McCarthy, A.J. and Saunders, J.R. (1993). The phylogeny os autotrophic ammonia-oxidicing bacteria as determined by analisys of 16S ribosomal RNA gene secuences. Journal of General Microbiology 13:1147-1153.
Hence, M., Harremoës, P., la Cour Jansen, J., and Arvin, E. (1997) Wastewater Treatment. Berlin.: Springer-Verlag.
Hollocher, T.C., Tate, M.E., and Nicholas, D.J.(1981) Oxidation of ammonia by Nitrosomonas europaea. Definite 18O-tracer evidence that hydroxilamine formation involves a monooxygenase. Journal of Biological Chemistry 256: 10834-10836.
Huang Z, Gedalanga PB, Asvapathanagul P, Olson BH (2010) Influence of physicochemical and operational parameters on Nitrobacter and Nitrospira communities in an aerobic activated sludge bioreactor. Water Res 44(15):4351–4358.
Jongman, R. H. G., ter Braak, C. J. F., & van Tongeren, O. F. R. (Eds.). 1995. Data Analysis in Community and Landscape Ecology. Cambridge University Press. Cambridge.
Juretschko, S., Timmermann, G., Schmid, M., Schleifer, K. H., Pommerening-Röser, a, Koops, H. P., & Wagner, M. (1998). Combined molecular and conventional analyses of nitrifying bacterium diversity in activated sludge: Nitrosococcus mobilis and Nitrospira-like bacteria as dominant populations. Applied and environmental microbiology, 64(8), 3042–51.
Kim, D. y Kim, S. (2006). Effect of nitrite concentration on the distribution and competition of nitrite-oxidizing bacteria in nitratation reactor systems and their kinetic characteristics. Water Research. 40: 887-894.
Kim, J.K., Park, K.P., Cho, K.S., Nam, S.W., Park, T.J., and Bajpai, R. (2005). Aerobic nitrification-denitrification by het- erotrophic Bacillus strains. Bioresour Technol 96: 1897– 1906.

MÁSTER EN INGENIERÍA AMBIENTAL
70
Koops H, Purkhold U, Pommerening-R A, Timmermann G, Wagner M (2006) The lithoautotrophic ammonia-oxidizing bacteria. The Prokaryotes 5:778–811.
Koops HP, Pommerening-Roser A (2001) Distribution and ecophysiology of the nitrifying bacteria emphasizing cultured species. FEMS Microbiol Ecol 37:1–9.
Leininger S, et al. (2006) Archaea predominate among ammonia-oxidizing prokar- yotes in soils. Nature 442:806–809. 15.
Limpiyakorn, T., Fürhacker, M., Haberl, R., Chodanon, T., Srithep, P., & Sonthiphand, P. (2013). amoA-encoding archaea in wastewater treatment plants: a review. Applied microbiology and biotechnology, 97(4), 1425–39. doi:10.1007/s00253-012-4650-7.
Limpiyakorn T, Sonthiphand P, Rongsayamanont C, Polprasert C (2011) Abundance of amoA genes of ammonia-oxidizing archaea and bacteria in activated sludge of full- scale wastewater treatment plants. Bioresour Technol 102:3694–3701. 34.
Limpiyakorn T, Kurisu F, Sakamoto Y, Yagi O (2007) Effects of ammonium and nitrite on communities and populations of ammonia-oxidizing bacteria in laboratory-scale continuous-low reactors. FEMS Microbiol Ecol 60:501–512.
Limpiyakorn T, Shinihara Y, Kurisu F, Yagi O (2005) Communities of ammonia-oxidizing bacteria in activated sludge of various sewage treatment plants in Tokyo. FEMS Microbiol Ecol 54:205–117
Liu, Y., Li, Y., and Lv, Y. (2012) Isolation and characterization of a heterotrophic nitrifier from coke plant wastewater. Water Sci Technol 65: 2084–2090.
Logemann S, Schantl J, Bijvank S, van Loosdrecht MCM, Kuenen JG, Jetten M (1998) Molecular microbial diversity in a nitrifying reactor system without sludge retention. FEMS Microbiol Ecol 27:239–249
Lücker, S., Nowka, B., Rattei, T., Spieck, E., & Daims, H. (2013). The Genome of Nitrospina gracilis Illuminates the Metabolism and Evolution of the Major Marine Nitrite Oxidizer. Frontiers in microbiology, 4(February), 27. doi:10.3389/fmicb.2013.00027.
Lydmark P, Almstrand R, Samuelsson K, Mattsson A, Sorensson F, et al. (2007) Effects of environmental conditions on the nitrifying population dynamics in a pilot wastewater treatment plant. Environ Microbiol 9: 2220–2233.
Lydmark P, Lind M, Sörensson F, Hermansson M (2006) Vertical distribution of nitrifying populations in bacterial biofilms from a full-scale nitrifying trickling filter. Environ Microbiol 8:2036–2049
Maixner F, Noguera DR, Anneser B, Stoecker K, Wegl G, Wagner M et al. (2006). Nitrite concentration influ- ences the population structure of Nitrospira-like bacteria. Environ Microbiol 8: 1487–1495.
Martín, M. (2005). Técnicas e instrumentos de gestión ambiental. Editorial Nau Llibres.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
71
Metcalf & Eddy (2000). Ingeniería de aguas residuales. Tratamiento, vertido y reutilización. 3a edición. Editorial McGraw-Hill.
Miura Y, Hiraiwa MN, Ito T, Itonaga T, Watanabe Y, et al. (2007) Bacterial community structures in MBRs treating municipal wastewater: Relationship between community stability and reactor performance. Water Res 41: 627–637.
Müller, A., Wetzel, M., Loewnthal, R. y Ekama, G. (2003). Heterotrophy anoxic yield in anoxic aerobic activated sludge system treating municipal wastewater. Water Research. 37: 2435-2441.
Mussmann, M., Brito, I., Pitcher, A., Sinninghe Damsté, J. S., Hatzenpichler, R., Richter, A., … Head, I. M. (2011). Thaumarchaeotes abundant in refinery nitrifying sludges express amoA but are not obligate autotrophic ammonia oxidizers. Proceedings of the National Academy of Sciences of the United States of America, 108(40), 16771–6. doi:10.1073/pnas.1106427108.
Olson, T.C., and Hooper, A.B. (1983) Energy coupling in the bacterial oxidation of small molecules: an extracytoplasmic dehydrogenase in Nitrosomonas. FEMS Microbiology Letters 19: 47-50.
Otawa K, Asano R, Ohba Y, Sasaki T, Kawamura E, Koyama F, Nakamura S, Nakai Y (2006) Molecular analysis of ammonia- oxidizing bacteria community in intermittent aeration sequencing batch reactors used for animal wastewater treatment. Environ Microbiol 8:1985–1996.
Park, H.-D., Lee, S.-Y., and Hwang, S. (2009) Redundancy analysis demonstration of the relevance of temperature to ammonia-oxidizing bacterial community compositions in a full-scale nitrifying bioreactor treating saline wastewater. J Microbiol Biotechnol 19: 346–350.
Park JJ, Byun IG, Yu JC, Park SR, Ju DJ, et al. (2008) Analysis of nitrifying bacterial communities in aerobic biofilm reactors with different DO conditions using molecular techniques. Water Sci Technol 57: 1889–1899.
Park HD, Wells GF, Bae H, Criddle CS, Francis CA (2006) Occurrence of ammonia-ox- idizing archaea in wastewater treatment plant bioreactors. Appl Environ Microbiol 72:5643–5647.
Park HD, Noguera DR (2004) Evaluating the effect of dissolved oxygen on ammonia-oxidizing bacterial communities in activated sludge. Water Res 38: 3275–3286.
Pérez, C. 1996. Econometría y análisis estadístico multivariable con Statgraphics.Editoral RA- MA. Madrid, España.
Pommerening-Röser, A., G. Rath, and H.-P. Koops. 1996. Phylogenetic di- versity within the genus Nitrosomonas. Syst. Appl. Microbiol. 1:344–351.
Prosser J. I. (1989) Autotrophic nitrification in bacteria. Adv. Microb. Physiol. 30:125–181.
Purkhold, U., Pommerening-Röser, A., Juretschko, S., Schmid, M.C., Koops, H.P., and Wagner, M. (2000) Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA

MÁSTER EN INGENIERÍA AMBIENTAL
72
sequence analysis: implications for molecular diversity surveys. Appl Environ Microbiol 66: 5368–5382.
Sánchez, O., Ferrera, I., González, J. M., & Mas, J. (2013). Assessing bacterial diversity in a seawater-processing wastewater treatment plant by 454-pyrosequencing of the 16S rRNA and amoA genes. Microbial biotechnology. doi:10.1111/1751-7915.12052
Schleper C, Jurgens G, Jonuscheit M (2005) Genomic studies of uncultivated archaea. Nat Rev Microbiol 3:479–488.
Schloss PD, Handelsman J. (2004). Status of the microbial census. Microbiol Mol Biol Rev 68: 686–691.
Schramm, A. (2003). In Situ Analysis of Structure and Activity of Nitrifying Community in Biofilms, Aggregates, and Sediments. Geomicrobiology Journal. 20: 313-333. Serrano,
Schramm, A., D. de Beer, A., Van den Heuvel, S., Ottengraf, S., Amann, R., (1999). Microscale distribution of populations and activities of Nitrosospira and Nitrospira spp. along a macroscale gradient in a nitrifying bioreactor: quantification by in situ hybridization and the use of microsensors. Appl. Environ. Microb. 65: 3690-3696
Schramm A, De Beer D, Wagner M, Amann R (1998) Identification and activities in situ of Nitrosospira and Nitrospira spp. as dom- inant population in nitrifying fluidized bed reactor. Appl Environ Microbiol 64(9):3480–3485
Seviour, R. and Nielsen, H. (2010). Microbial ecology of activated sludge. IWA publishing, London.
Siegel, S. (1983). Estadística No Paramétrica aplicada a las ciencias de la conducta. Editoral Trillas. Mexico.
Siripong S, Rittmann B (2007) Diversity study of nitrifying bacteria in full-scale municipal wastewater treatment plants. Water Res 41(5):1110–1120.
Sorokin, D. Y., Lücker, S., Vejmelkova, D., Kostrikina, N. a, Kleerebezem, R., Rijpstra, W. I. C., Daims, H. (2012). Nitrification expanded: discovery, physiology and genomics of a nitrite-oxidizing bacterium from the phylum Chloroflexi. The ISME journal, 6(12), 2245–56.
Spang, A., Hatzenpichler, R., Brochier-Armanet, C., Rattei, T., Tischler, P., Spieck, E., Streit, W., Stahl, D.A., Wagner, M., Schleper, C., 2010. Distinct gene set in two different lineages of ammonia-oxidizing archaea supports the phylum Thaumarchaeota. Trends Microbiol. 18 (8), 331–340.
Su, J.J., Yeh, K.S., and Tseng, P.W. (2006) A strain of Pseu- domonas sp. Isolated from piggery wastewater treatment systems with heterotrophic nitrification capability in Taiwan. Curr Microbiol 53: 77–81.
Raszka, A., Surmacz-Górska, J., Zabczynski, S., and Miksch, K. (2011) The population dynamics of nitrifiers in ammonium-rich systems. Water Environ Res 83: 2159– 2169.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
73
Rotenberry, J. T. & Wiens, J. A. 1980. A synthetic approach to principal component analysis of bird/habitat relationships. En D. E. Capen (Ed.): The Use of Multivariate Statistics in Studies of Wildlife Habitat, pp. 197-208. USDA Forest Servi- ce. General Technical Report No. RM-87. Vermont.
Tan, N.C.G, Kampschreur, MJ., Wanders, W., van der Pol WLJ, van de Vossenberg J, Kleerebezem R, van Loosdrecht MCM, Jetten MSM (2008) Physiological and phylogenetic study of an ammonium-oxidizing culture at high nitrite concentrations. Syst Appl Microbiol 31:114–125
ter Braak, C. J. F. & Símilauer, P. (1998). CANOCO. Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination (Version 4). Microcomputer Power. Ithaca. New York.
Teske, A., Alm, E., Regan, J.M., Toze, S., Rittmann, B.E., and Stahl, D.A. (1994) Evolutionary relationships among ammonia-oxidizing and nitrite-oxidizing bacteria. J Bacteriol 176: 6623–6630. Third,
Vázquez-Padín, J., Figueroa, M., Mosquera-Corral, A., Campos, J.L y Méndez, R. (2009). Population dynamics of nitrite oxidizers in nitrifying granules. Water Science & Technology. 60.10: 2529-2536. Viedma,
Wagner, M., Amman, R., Lemmer, H., and Schleifer, K.-H. (1993) Probing activated sludge with oligonucleotides spe- cific for proteobacteria: inadequacy of culture-dependent methods for describing microbial community structure. Appl Environ Microbiol 59: 1520–1525. Wagner,
Wang, X., Wen, X., Xia, Y., Hu, M., Zhao, F., & Ding, K. (2012). Ammonia oxidizing bacteria community dynamics in a pilot-scale wastewater treatment plant. PloS one, 7(4), e36272. doi:10.1371/journal.pone.0036272.
Wang XH, Wen XH, Criddle C, Wells G, Zhang J, et al. (2010) Community analysis of ammonia-oxidizing bacteria in activated sludge of eight wastewater treatment systems. J Env Sci-China 22: 627–634.
Watson, S.W., Bock, E., Valois, F.W., Waterbury, J.B., and Schlosser, U. (1986) Nitrospira marina gen. nov. sp. nov.: a chemolithotrophic nitrite-oxidizing bacterium. Arch Micro- biol 144: 1–7.
Weidler, G. W., Dornmayr- Pfaffenhuemer, M., Gerbl, F. W., Heinen,W., and Stan-Lotter, H. (2007). Communities of Archaea and Bacteria in a subsurface radioactive thermal spring in the Austrian Central Alps, and evidence of ammonia-oxidizing Crenar- chaeota. Appl. Environ. Microbiol. 73, 259–270.
Wells, G.F., Park, H.-D., Yeung, C.-H., Eggleston, B., Francis, C.A., Criddle, C.S., (2009). Ammonia-oxidizing communities in a highly aerated full-scale activated sludge bioreactor: betaproteobacterial dynamics and low relative abundance of Crenarchaea. Environ. Microbiol. 11, 2310–2328.
Wett B, Rauch W (2003) The role of inorganic carbon limitation in biological nitrogen removal of extremely ammonia concentrated wastewater. Water Res 37(5):1100–1110.
Whang LM, Yang KH, Yang YF, Han YL, Chen YJ, Cheng SS (2009) Microbial ecology and performance of ammonia oxidizing bacteria (BOA) in biological processes treating petrochemical wastewater with high strength of ammonia: effect of Na2CO3 addition. Water Sci Technol 59(2):223–231.

MÁSTER EN INGENIERÍA AMBIENTAL
74
Winogradsky, S. (1890). Recherches sur les organismes de la nitrification. Ann. Inst. Pasteur 4, 213–231.
Wittebolle L, Vervaeren H, Verstraete W, Boon N (2008) Quantifying community dynamics of nitrifiers in functionally stable reactors. Appl Environ Microbiol 74:286–293.
Wuchter C, et al. (2006) Archaeal nitrification in the ocean. Proc Natl Acad Sci USA 103:12317–12322.
Xia S, Li J, Wang R (2008) Nitrogen removal performance and microbial community structure dynamics response to carbon nitrogen ratio in a compact suspended carrier biofilm reactor. Ecol Eng 32: 256–262.
Yu, K., and Zhang, T. (2012) Metagenomic and metatran- scriptomic analysis of microbial community structure and gene expression of activated sludge. PLoS ONE 7: e38183. doi:10.1371/journal.pone.0038183. Zhang,
Yu, T., Qi, R., Zhang, Y y Yang, M. (2010). Nitrifier characteristics in submerged membrane bioreactors under different sludge retention times. Water Research. 44: 2823-2830. Zornoza,
Zhang, T., Shao, M.F., and Ye, L. (2011a) 454 pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants. ISME J 6: 1137–1147.
Zhang, T., Ye, L., Tong, A.H.Y., Shao, M.-F., and Lok, S. (2011b) Ammonia-oxidizing archaea and ammonia- oxidizing bacteria in six full-scale wastewater treatment bioreactors. Appl Microbiol Biotechnol 91: 1215–1225
Zhang T, et al. (2009) Occurrence of ammonia-oxidizing Archaea in activated sludges of a laboratory scale reactor and two wastewater treatment plants. J Appl Microbiol 107:970–977.
Zornoza, A., Avendaño, L., Aguado, D., Borrás, L., & Alonso, J. L. (2012). Análisis de las correlaciones entre la abundancia de bacterias nitrificantes , parámetros operacionales y físico-químicos relacionados con el proceso biológico de nitrificación en fangos activos. VIII Jornadas de Transferencia de Tecnología sobre Microbiología del Fango Activo, Sevilla. Spain.
Zornoza A., Alonso, J., Serrano, S., Fajardo, V., Zorrilla, F., Bernácer, I., Morenilla, J. 2010b. Estudio integrado del proceso de fangos activos I. Análisis descriptivo de factores físico- químicos y biológicos implicados en su dinámica. Documentación presentada en VII Jornadas de Transferencia de Tecnología sobre Microbiología del Fango Activo. Sevilla, España.

DINÁMICA POBLACIONAL DE LAS COMUNIDADES DE BACTERIAS NITRIFICANTES
EN UNA EDAR CON SISTEMA CONVENCIONAL DE FANGOS ACTIVOS
75





























