Embed Size (px)
Citation preview

NAME: MUSA MARENA
TOPIC: FILOVIRUS HAEMORRHAGIC FEVER
CONTENTS PAGES
INTRODUCTION 2
HISTORY 3-5
GENOME ABD MORPHOLOGY 6-9
PATHOGENESIS 9-13
VECTOR AND MODE OF TRANSMISSION 14-15
BINDING AND ENTRY 15-16
REPLICATION 17-19
TRANSCRIPTION AND TRANSLATION 20-24
ASSEMBLY AND BUDDING 25-26
CLINICAL FEATURES 26-27
DIAGNOSIS 27
TREATMENT 27
FILOVIRUS HAEMORRHAGIC FEVER
INTRODUCTION
Haemorrhagic fever Is an acute febrile illness characterized by malaise, myalgia, and prostration
dominated by generalized abnormalities of vascular permeability, and regulation. Bleeding
manifestations often occur, particularly in severe cases; they are usually diffuse and reflect
widespread vascular damage rather than life-threatening volume loss.
1

Filoviruses are classic emerging infections. Filoviruses have two main genera and these are Ebola
and Marburg viruses. They belong to the group of viruses called haemorrhagic fever viruses. Four
families of viruses: Arenaviruses (Junin Virus), Bunyaviruses (Nariovirus), Flaviviruses (i.e.
Yellow Fever), Filoviruses (Marburg & Ebola)they are CDC Classification: Biosafety level
(BSL)-4 Agent (cf. HIV is only 2). They are mainly RNA viruses covered in a lipid coating and
are geographically restricted to areas where host species live. Humans are not natural reservoirs
for these viruses.
They are filamentous with a linear ~13-19kb genome. They can infect mice, hamsters, guinea
pigs and monkeys, although the viral reservoir in the wild is not known. Human epidemics seem
to be related to blood-born nosocomial spread (often due to re-use of needles in hospitals;
nosocomial = hospital infection) and to close contact with infected persons (since this is a
hemorrhagic disease, this presumably would involve exposure to large amounts of blood).
Primary infections with Marburg and Ebola are 25-90% fatal. Death is thought to be due to
visceral organ necrosis (e.g., liver) due to viral infection of tissue parenchymal cells. It is
uncertain what role hemorrhage has in death. Wild-caught monkeys are now quarantined before
release to US primate centers. Cloned viruses has help in studying viral genetic and will help the
development of diagnostic serologic tests for infection, and work is also progressing to try to
develop a vaccine for Ebola virus. The first successful vaccination against this virus was reported
in 1998, by Gary Nabel's group at the University of Michigan. In this report, a DNA vaccine
encoding the Ebola virus glycoprotein was able to elicit a T-cell based immune response in
guinea pigs, which was sufficient to protect the animals against infection with a live-Ebola virus
(Xu et al. Nature Medicine 4:37, 1998). Subsequent studies in nonhuman primates have
confirmed that a DNA vaccine can represent an important component of an effective Ebola virus
vaccine. Specifically, a combination of DNA immunization and boosting with adenovirus vectors
encoding viral proteins resulted in the protection of cynomolgus macaques from an otherwise
lethal dose of highly pathogenic, wild-type Ebola Zaire virus (Sullivan et al. Nature 408:2000
HISTORY
Marburg Virus also called (African Hemorrhagic Fever, Green Monkey Disease, and Marburg
Fever). In 1967 simultaneous outbreaks of hemorrhagic fever occurred in Yugoslavia and in
Germany, in lab workers who were processing kidneys from African green monkeys. There were
31 cases and 7 deaths. The virus was first characterized in Marburg, Germany and traced to a
single shipment of Ugandan monkeys. Sporadic additional cases showed up in 1975, 1980, 1982
2

and 1987. Most recently there was the most serious outbreak of Marburg which occurred in
Angola in 2004-2005 had 252 cases with 227 deaths (90%). In 2007 there were 2 cases and 1
death (50%) in Uganda and in 2008 one case who died after 10days illness in Netherlands after
returning from a visit in Uganda
In 1976 there were epidemics of severe hemorrhagic fever in Zaire and Sudan. In Zaire, there
were approximately 300 cases with an 80% fatality rate (due to Ebola-Zaire; ZEBO). In Sudan,
there were a roughly similar number of cases, with a fatality rate of roughly 50% (due to Ebola-
Sudan; SEBO). Ebola virus was originally isolated in Zaire (now Democratic Republic of the
Congo), and it was named after a small river in N.W. Zaire. Ultra-structurally the virus resembled
Marburg virus but it was antigenically (and genetically) distinct. It now appears that at least four
and probably five EBO viruses exist -- EBO-Z (Zaire is 990-1086nm in length), EBO-S (Sudan is
974-1063nm in length), EBO-CI (Côte d’Ivoire), EBO-R (Reston is 1026-1083nm in length) and
more recently EBO-B (Bundinbugyo) from Gulu region Uganda. Figure1. The first two are
known to be highly lethal in humans and are spread via bodily fluids and by close (nonsexual)
contact. The Reston virus appears to be less lethal in humans (0 deaths in 6 cases), although it is
lethal in nonhuman primates. Outbreaks of Ebola occurred in 1995 in the Kikwit area of Zaire
(over 315 cases, with 80% fatality; due to EBO-Z) and in the Gulu region of Uganda in 2000
(over 400 cases, but with roughly 50% fatality; due to EBO-B). It is uncertain how the Kikwit
and Gulu outbreaks started. However, a smaller outbreak in 1996 in Gabon was traced to a group
of 20 young Gabonese who trapped and caught a Chimpanzee that was sick. It is believed that
exposure to Ebola occurred during the preparation of the Chimpanzee, prior to cooking and
consumption of the animal. Interestingly, Ebola was isolated only from meat-eating Chimps, and
not from strictly vegetarian members of the same troupe of animals. Outbreaks of EBO-Reston
have occurred in US primate colonies in the Washington area (Reston, 1989) and in Texas (1990,
1996). These outbreaks were contained by destruction of all animals within the affected area of
the facility. The outbreaks all appear to trace back to shipments of macaques from a single
Philippine exporter. A total of 6 humans have become infected by EBO-Reston, but none had
died. On Wednesday December 10, 2008, the Philippine Department of Agriculture and
the Philippine Department of Health announced that an Ebola virus of the Reston species
has been identified from sick pigs in three locations in the Philippines. This is the first
time globally that domestic pigs are reported to have been infected with an Ebola virus.
High rates of sickness and death in pigs and the pigs were found to be were co-infected
with Porcine Respiratory Reproductive Syndrome one (PRRS) virus and the Ebola
3
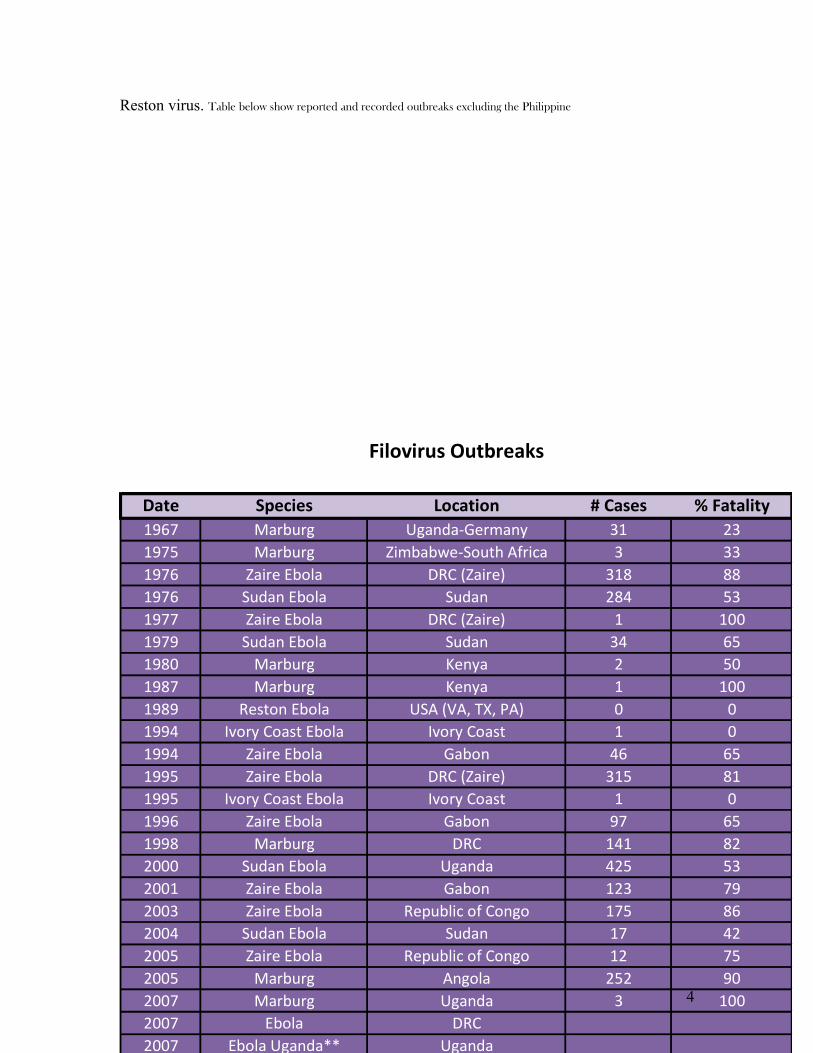
Reston virus. Table below show reported and recorded outbreaks excluding the Philippine
Date Species Location # Cases % Fatality1967 Marburg Uganda-Germany 31 231975 Marburg Zimbabwe-South Africa 3 331976 Zaire Ebola DRC (Zaire) 318 881976 Sudan Ebola Sudan 284 531977 Zaire Ebola DRC (Zaire) 1 1001979 Sudan Ebola Sudan 34 651980 Marburg Kenya 2 501987 Marburg Kenya 1 1001989 Reston Ebola USA (VA, TX, PA) 0 01994 Ivory Coast Ebola Ivory Coast 1 01994 Zaire Ebola Gabon 46 651995 Zaire Ebola DRC (Zaire) 315 811995 Ivory Coast Ebola Ivory Coast 1 01996 Zaire Ebola Gabon 97 651998 Marburg DRC 141 822000 Sudan Ebola Uganda 425 532001 Zaire Ebola Gabon 123 792003 Zaire Ebola Republic of Congo 175 862004 Sudan Ebola Sudan 17 422005 Zaire Ebola Republic of Congo 12 752005 Marburg Angola 252 902007 Marburg Uganda 3 1002007 Ebola DRC2007 Ebola Uganda** Uganda2008 Marburg Netherlands ex Uganda 1 100
Filovirus Outbreaks
4

Date Species Location # Cases % Fatality1967 Marburg Uganda-Germany 31 231975 Marburg Zimbabwe-South Africa 3 331976 Zaire Ebola DRC (Zaire) 318 881976 Sudan Ebola Sudan 284 531977 Zaire Ebola DRC (Zaire) 1 1001979 Sudan Ebola Sudan 34 651980 Marburg Kenya 2 501987 Marburg Kenya 1 1001989 Reston Ebola USA (VA, TX, PA) 0 01994 Ivory Coast Ebola Ivory Coast 1 01994 Zaire Ebola Gabon 46 651995 Zaire Ebola DRC (Zaire) 315 811995 Ivory Coast Ebola Ivory Coast 1 01996 Zaire Ebola Gabon 97 651998 Marburg DRC 141 822000 Sudan Ebola Uganda 425 532001 Zaire Ebola Gabon 123 792003 Zaire Ebola Republic of Congo 175 862004 Sudan Ebola Sudan 17 422005 Zaire Ebola Republic of Congo 12 752005 Marburg Angola 252 902007 Marburg Uganda 3 1002007 Ebola DRC2007 Ebola Uganda** Uganda2008 Marburg Netherlands ex Uganda 1 100
Filovirus Outbreaks
5
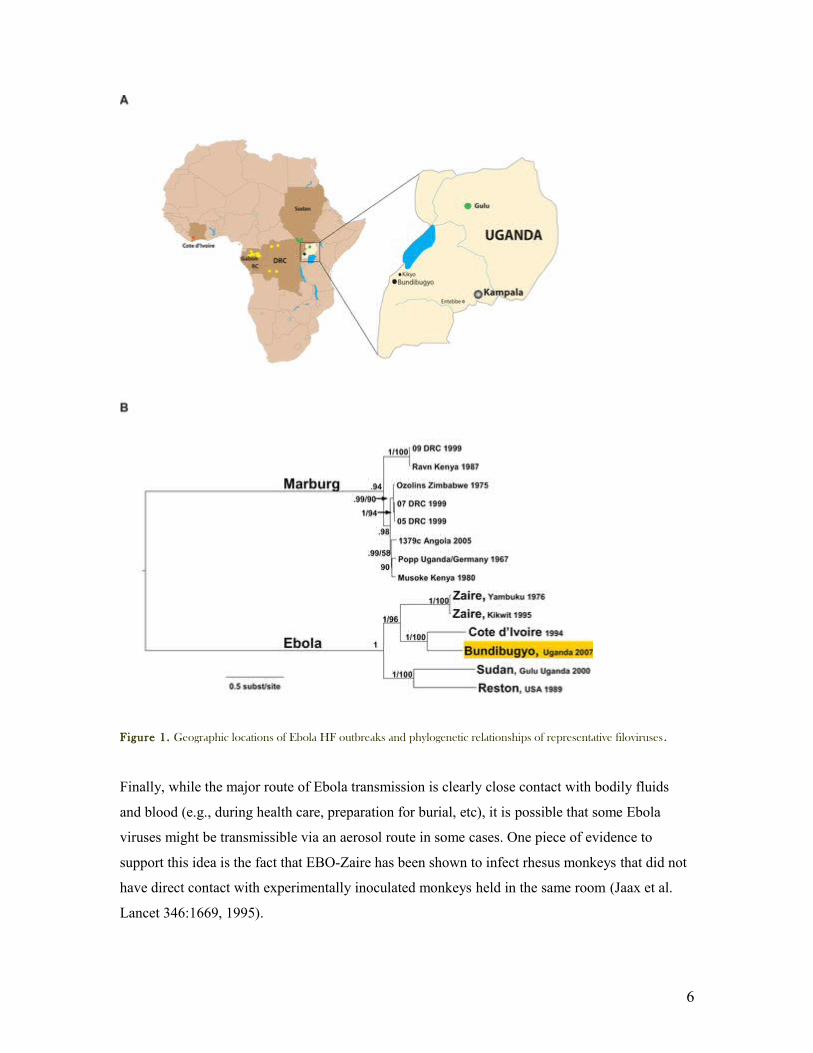
Figure 1. Geographic locations of Ebola HF outbreaks and phylogenetic relationships of representative filoviruses.
Finally, while the major route of Ebola transmission is clearly close contact with bodily fluids
and blood (e.g., during health care, preparation for burial, etc), it is possible that some Ebola
viruses might be transmissible via an aerosol route in some cases. One piece of evidence to
support this idea is the fact that EBO-Zaire has been shown to infect rhesus monkeys that did not
have direct contact with experimentally inoculated monkeys held in the same room (Jaax et al.
Lancet 346:1669, 1995).
6

GENOME AND MORPHOLOGY
Electron microscopic observation of filoviruses revealed distinctive bacilliform to filamentous
virus particles, and this highly characteristic morphology that inspired their name (Latin filium,
thread). Virion from cultured tissue are pleomorphic, appearing as either U-shaped, six-shaped
(6) or circular (torus) configuration or as elongated filamentous forms. The filamentous forms can
be seen to form branched structure. Figures2 below
The length varied 800-1080nm up to 14,000nm with Marburg virion shorter than Ebola virion
with 860nm and 12,000nm respectively. The virion have a uniform diameter of 80nm, contain a
helical ribonucleoprotein complex or nucleocapsid (NC) roughly 50nm in diameter and have a
central axial space (approx 20nm in diameter) running the length of the particle. Figures3&4. The
NC has a helical periodicity of ~5nm and is surrounded by matrix protein and a closely outer
envelope derived from the host cell plasma membrane. The virion surface is studded with
membrane anchored peplomers projecting ~10nm from the surface figure 3. The virion can appear
ragged or “moth eaten” figures 2D especially in late infection. Virion density has been determined
to be 1.14g/ml by centrifugation in potassium tartan gradient and relative molecular mass Mr 3-
6×108
The virion genome which constitute~1% of virion mass is found on single-stranded, non-
segmented, linear, negative-sense RNA (–ve ssRNA). Filoviruses genome is very similar. Their
organization generally conforms to those of paramyxoviruses and Rhabdoviruses but their
7

figure3
Complexity is more akin to those of paramyxoviruses. Filovirus genome is (~19Kb) long and
making them the largest in the mononegavirales order. They contain seven sequentially arranged
genes in the order 3’-5’ nucleoprotein (NP), viral protein (VP) 35, VP40, glycoprotein (GP),
VP30, VP24 and RNA polymerase L. The genes are delineated by conserved transcriptional
signals and begin close to the 3’ end of genome sequence with a start site and ends with a stop
(polyadenylated) site. Like in other mononegavirale these genes are usually separated by short
intergenic regions (IR) of one or more nucleotides. Unusual feature of gene overlaps are found in
the filovirus genome which have been identify in the genome of some paramyxo and
Rhabdoviruses but does not resemble those of filoviruses. Figure 3
Genes are defined by highly conserved transcriptional start signals at their 3' (3'-
CUNCNUNUAAUU-5'; consensus motif) and termination signals at their 5' ends (3'-
UAAUUCUUUUU-5') Most genes tend to possess long noncoding sequences at their 3' and/or 5'
ends, which contribute to the increased length of the genome
8

The stop site of an upstream gene overlaps the start site of a downstream gene, and overlapping
sequences are limited to the conserved transcriptional signals and are centered on 3’-UAAUU
pentanucleotide sequence common to start and stop sites. Figure4
Marburg genome have one (VP30-VP24) overlap but the characterize Ebola genome have two or
three (VP35-VP40, GP-VP30, VP24-L) overlaps. Filovirus generally have short intergenic
sequence although all have a single lengthy sequence more than 120bases separating GP and
VP30 gene in Marburg (MARV) and VP35 and VP40 on Ebola virus (EBOV). The extragenic
sequence at the 3’ end (Leader) is short in all filovirus genomes ranging in length from 50-
70bases while the length of the 5’ end (Trailer) sequence are variable. These extragenic are
thought to be templates for very small viral, nonpolyadenylated subgenomic RNAs. The extreme
3’ and 5’ ends of the genomes are conserved, show high degree of complementarity and
potentially form stem loops structure. The trailer of Zaire Ebola virus (ZEBOV) is the longest
(677bases), followed Sudan Ebola virus (SEBOV) 381bases MARV 75-95bases and Ebola
Reston REBOV 25bases
9
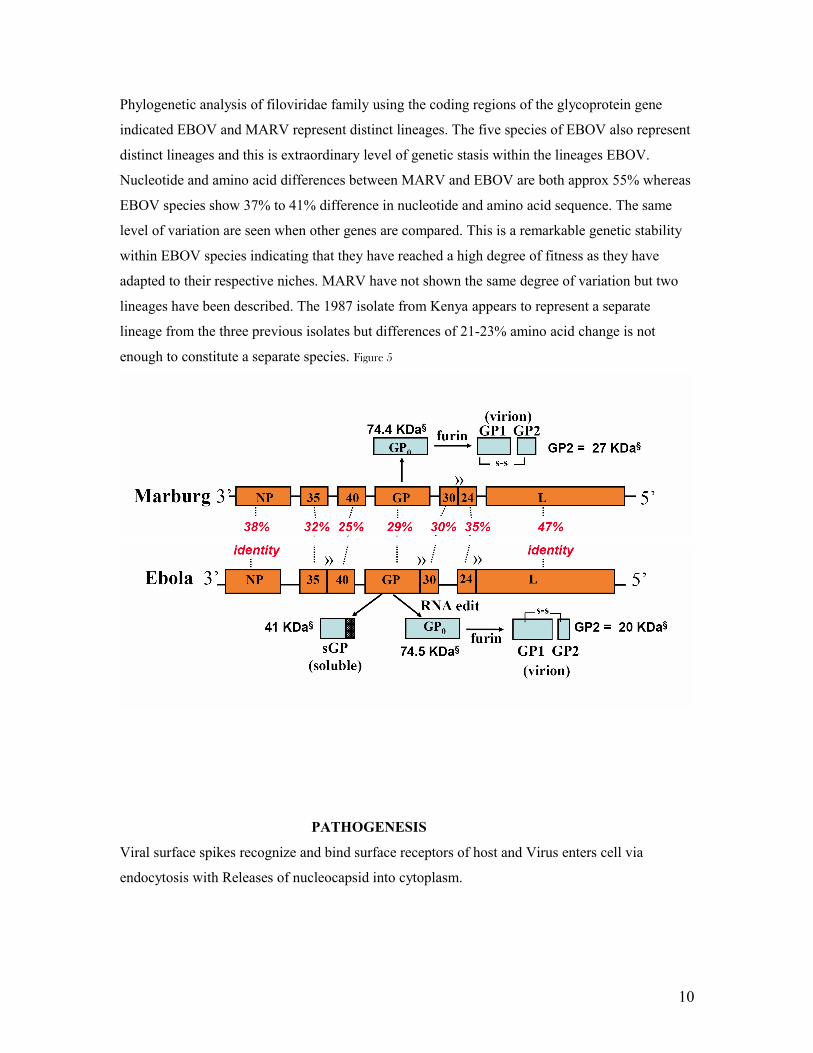
Phylogenetic analysis of filoviridae family using the coding regions of the glycoprotein gene
indicated EBOV and MARV represent distinct lineages. The five species of EBOV also represent
distinct lineages and this is extraordinary level of genetic stasis within the lineages EBOV.
Nucleotide and amino acid differences between MARV and EBOV are both approx 55% whereas
EBOV species show 37% to 41% difference in nucleotide and amino acid sequence. The same
level of variation are seen when other genes are compared. This is a remarkable genetic stability
within EBOV species indicating that they have reached a high degree of fitness as they have
adapted to their respective niches. MARV have not shown the same degree of variation but two
lineages have been described. The 1987 isolate from Kenya appears to represent a separate
lineage from the three previous isolates but differences of 21-23% amino acid change is not
enough to constitute a separate species. Figure 5
PATHOGENESIS
Viral surface spikes recognize and bind surface receptors of host and Virus enters cell via
endocytosis with Releases of nucleocapsid into cytoplasm.
10
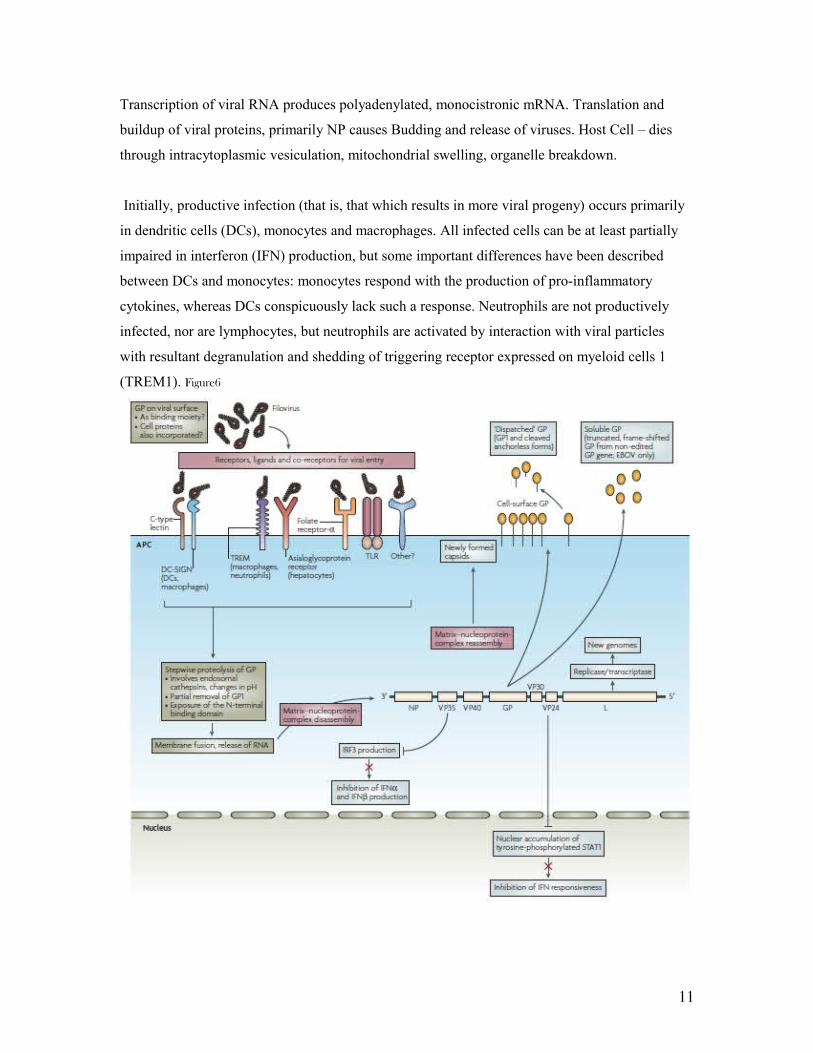
Transcription of viral RNA produces polyadenylated, monocistronic mRNA. Translation and
buildup of viral proteins, primarily NP causes Budding and release of viruses. Host Cell – dies
through intracytoplasmic vesiculation, mitochondrial swelling, organelle breakdown.
Initially, productive infection (that is, that which results in more viral progeny) occurs primarily
in dendritic cells (DCs), monocytes and macrophages. All infected cells can be at least partially
impaired in interferon (IFN) production, but some important differences have been described
between DCs and monocytes: monocytes respond with the production of pro-inflammatory
cytokines, whereas DCs conspicuously lack such a response. Neutrophils are not productively
infected, nor are lymphocytes, but neutrophils are activated by interaction with viral particles
with resultant degranulation and shedding of triggering receptor expressed on myeloid cells 1
(TREM1). Figure6
11

As viral burden increases, lymphocyte apoptosis and a generalized failure of specific immune
responsiveness are observed; we propose these to be rooted in virally induced upregulation of co-
inhibitory molecules (such as B7-H1) on DCs and monocytes, followed by interaction with
programmed death 1 (PD1) receptors on T and B cells. Infection spreads to many cells including
liver hepatocytes, and the increasing release of pro-inflammatory cytokines crosses a threshold
from beneficial to potentially harmful inflammation, also degrading vascular epithelium.
12
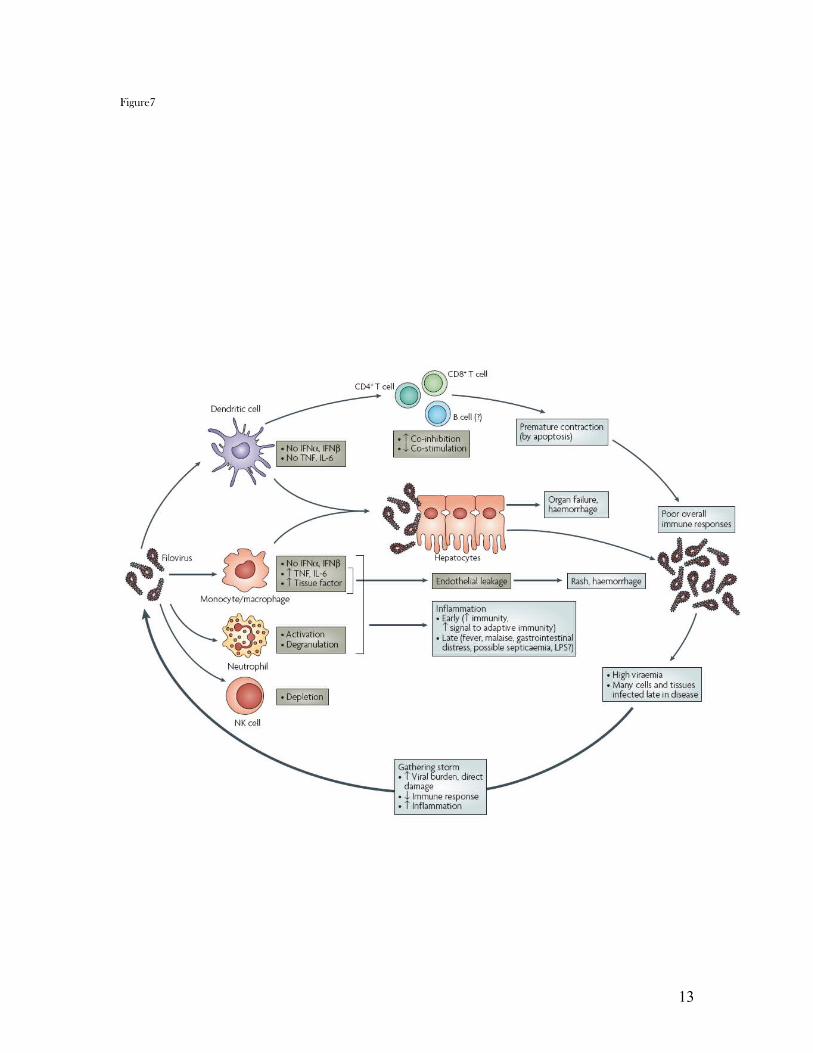
Figure7
13
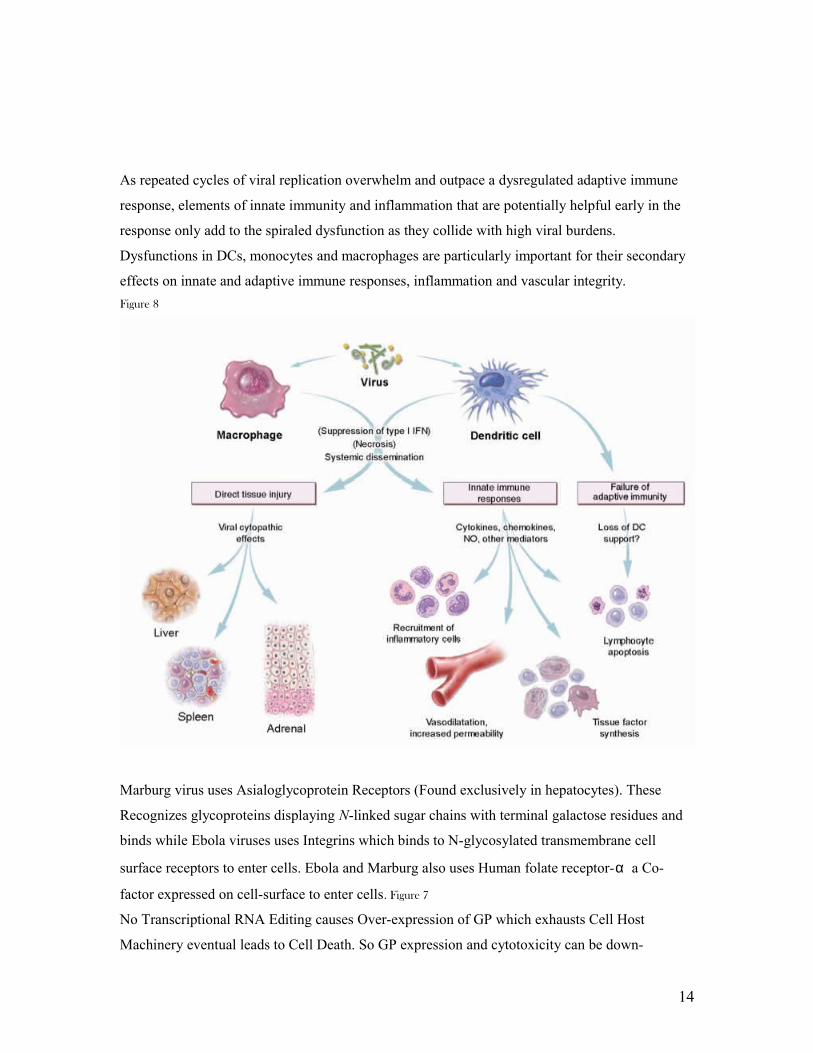
As repeated cycles of viral replication overwhelm and outpace a dysregulated adaptive immune
response, elements of innate immunity and inflammation that are potentially helpful early in the
response only add to the spiraled dysfunction as they collide with high viral burdens.
Dysfunctions in DCs, monocytes and macrophages are particularly important for their secondary
effects on innate and adaptive immune responses, inflammation and vascular integrity.
Figure 8
Marburg virus uses Asialoglycoprotein Receptors (Found exclusively in hepatocytes). These
Recognizes glycoproteins displaying N-linked sugar chains with terminal galactose residues and
binds while Ebola viruses uses Integrins which binds to N-glycosylated transmembrane cell
surface receptors to enter cells. Ebola and Marburg also uses Human folate receptor-α a Co-
factor expressed on cell-surface to enter cells. Figure 7
No Transcriptional RNA Editing causes Over-expression of GP which exhausts Cell Host
Machinery eventual leads to Cell Death. So GP expression and cytotoxicity can be down-
14

regulated by virus through transcriptional RNA editing and sGP expression. sGP inhibits early
activation of neutrophils by Binding to neutrophils via CD16b cell surface receptor. CD16b
activates neutrophils via lateral membrane interaction with CR3. sGP adsorbs neutralizing
antibodies.
Specific region of GP induces cytotoxic effects in endothelial cells. Rapid release of vasoactive
agents from infected cells Induces cell rounding and detachment from extracellular matrices this
Increases cell membrane permeability.
Proteolytic activation of GP0 precursor via cleavage of EBO-Z GP by furin is prerequisite for
fusion between viral envelope and host cell membrane. It Enables virus to replicate in host
leading to systematic infection.
Two sequences contribute to evasion of host immunity. Possible immunosuppressive sequence in
GP2 molecule and or Amino acid sequence at N-terminus suppress lymphocyte mitogen-
stimulated proliferation in vitro
Destruction of the Immune System through Infection of mononuclear phagocytes and fibroblastic
reticular system (associated with lymph nodes), Failure of early T-cell activation Disrupt antigen
trafficking and cytokine production causing Extensive apoptosis of blood leukocytes leading to
Lymphopenia (reduction in lymphocyte number) and severe damage to lymphoid tissue. Infected
Macrophages and circulating monocytes help transmit virus to other tissues
EARLY STAGE: pathology shows Striking lesions usually in liver, spleen and kidney, Necrosis
prominent in liver, lymph tissue, and spleen, little inflammatory response and viral particles
invade phagocytic cells
LATE STAGE: Liver and spleen become enlarged with excess blood, Hemorrhage in the
gastrointestinal tract, pleural, pericardial and peritoneal spaces and into the renal tubules with
deposition of fibrin, Abnormalities in coagulation parameters suggest that disseminated
intravascular coagulation is a terminal event. There is usually also profound leukopenia in
association with bacteremia.
Survived have IgG response against virus’s protein coat early on, cleared circulating antigen and
activated cytotoxic T-cells
Those who end up Dying have No IgG response and barely detectable levels of IgM
15

VECTOR AND MODE OF TRANSMISSION
Ebola Virus has No carrier state (reservoir Unknown). Researchers hypothesize that it is Zoonotic
because some outbreaks are related with contact animal like chimps and bats. Figure 9 Most
animals in Africa have positive antibodies to filoviruses and viruses have been isolated from their
serum. There is an unpopular theory that plant may be the reservoir of the virus. Human-Human
Transmission is through contact with contaminated secretions.
Ebola-Reston which Occurred in the U.S (Reston, VA) in African Green Monkeys imported from
Phillipine. Questions are asked: Why only to non-human primates? (Four scientists found to have
antibody for the disease). There is Circumstantial Evidence of airborne transmission because the
viruses spread within monkey kept within the same and between rooms (national center for
Infectious Diseases). ICEBOV was transmitted through contact with a death chimps carcass.
Figure 9
Marburg Virus maybe transmitted from animal host because fruit bats found in caves have
positive antibodies to the virus and isolates of the virus. It’s not clear whether they can transmit it
16
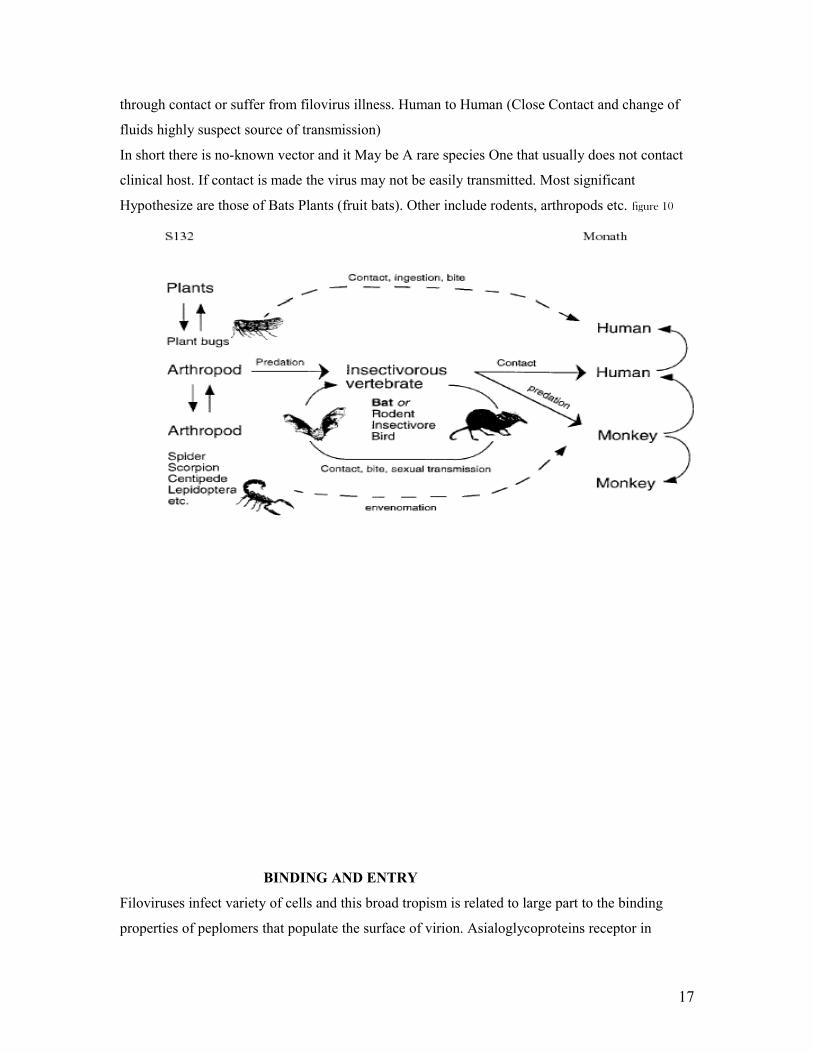
through contact or suffer from filovirus illness. Human to Human (Close Contact and change of
fluids highly suspect source of transmission)
In short there is no-known vector and it May be A rare species One that usually does not contact
clinical host. If contact is made the virus may not be easily transmitted. Most significant
Hypothesize are those of Bats Plants (fruit bats). Other include rodents, arthropods etc. figure 10
BINDING AND ENTRY
Filoviruses infect variety of cells and this broad tropism is related to large part to the binding
properties of peplomers that populate the surface of virion. Asialoglycoproteins receptor in
17

hepatocytes binds MARV but Ebola virus infects hepatocyte despite its GP lacking sialylated
glycans.
Beta 1 group of integrin are believed to interact with ZEBOV GP on cell surface and during
intracellular trafficking although cell expressing this molecules are not easily infected. Folate
receptors alpha is believe to be a co-factor in viral entry. C-type lectin present on certain
macrophages, dendritic cells and endothelium are capable of binding filovirus peplomers when
N-terminus contains mannose carbohydrate. Antibody binding to peplomers might also enhance.
Infectivity through its interaction with the Fc portion of complement protein C1q bond to the
surface of host cells. Figure 6
Figure 11
18

REPLICATION:
Filoviruses belong to the group of mononegavirale means that the viral genome is of
opposite polarity to mRNA. Thus, the viral genome cannot be used to make proteins until
it has first been transcribed. Figure 12. They include several families like Filoviridae,
Paramyxoviridae and Rhabdoviridae, plus Borna disease virus. These viruses probably
diverged from a single common ancestor as recently as the last ice age. . They are also
transcribed to produce mRNAs.
Figure 12
This has the following implications:
1. Purified virion RNA (without viral proteins) is not infectious (as noted above, it cannot
encode protein)
2. The viruses must bring their own RNA polymerase into the cell in order to make mRNA (ie,
the viral polymerase must be incorporated into the viral particle, or virion). The other key feature
of these viruses is that they make gene-unit length mRNAs (ie, each mRNA encodes only a single
protein). This is achieved by the use of transcriptional stop and start signals, which are located at
the boundaries of all of the viral genes. Stop/start transcription has two major results: Since there
is only a single promoter, located at the 3’ end of the viral genome, the polymerase can only load
onto its RNA template at one site. As it moves along the viral RNA, the polymerase encounters
19

stop/start signals at the boundaries of each of the viral genes. This results in pausing of the
enzyme, which often falls off the template. The result is that more mRNA is made from genes
that are located close to the promoter, and less mRNA is made from genes located far from the
promoter. Figure 13
Figure 13
This means that there is a polarity of transcription (see Figure below). The viruses use
this to regulate the expression of their genes, since highly expressed proteins are encoded
close to the promoter (e.g., structural proteins such as the nucleocapsid protein, N), while
proteins that are needed in only small amounts (e.g., enzymes such as the RNA
polymerase, L) are encoded far away from the promoter.
The other major consequence of stop/start transcription is that it complicates genome
replication. The only way that the complete viral RNA genome can be copied is if the
transcriptional stop/start signals can be ignored or over-ridden. This means that the
critical decision during viral RNA synthesis occurs very early on -- at the first gene
boundary (located between the leader RNA and the N gene). If the stop/start signals here
are obeyed, then only subgenomic mRNAs will be produced. However, if the stop/start
signal here is ignored or over-ridden, then a complete copy of the viral genome can be
made.
3’ end of filovirus genome has a promoter. This derives both the replication and
transcription. It derives the synthesis of full-length complementary/antigenomic RNA
from the encapsidated template. The complementary sequences and stem loop structure at
the ends of the genome are believed to be essential to filovirus replication. The initial
expression of viral genes leads to build up of viral proteins (especially NP) which are
believed to trigger the switch from transcription to replication. This switch leads to
20

synthesis and encapsidation of antigenomic RNA molecule which turn serve as a
template for genomic RNA that is also rapidly encapsidated. Depletion of capsid protein
is believe to cause a return to transcription and eventually an equilibrium is established
wherein transcription and replication are concurrent processes. As replication progress in
infected cells, NC particles containing genomic RNA accumulate and are directed to the
plasma membrane for virion assembly.
Reverse genetic system has shown that NP, VP35 and L proteins are all that is require to
transcribe and replicate MARV mingenomes but EBOV also require VP30
21

TRANSCRIPTION AND TRANSLATION
Upon entry into cell, the non segmented negative sense single stranded RNA genetic dictates that
transcription is the first (and obligatory) viral process which is similar to paramyxo and
Rhabdoviruses.
Once the nucleocapsid is released in the cytoplasm, polyadenylated, monocistronic mRNA are
synthesized from virus genes in 3’ to 5’ direction (with polar attenuated) from the encapsidated
genomic RNA template. Transcription seems to involve a process of starting and stopping as the
polymerase complex encounters conversed start (initiation) and stop
(termination/polyadenylation) sites along the genome. Transcripts are believed to be ‘capped’ at
5’ end by the L protein because it contains conserved motifs associated with this enzymatic
activity. It’s postulated that the leader sequence is transcribed while the intergenic and trailer
sequences are not transcribe. The promoters for initiating RNA synthesis are contained within
156 and 177 nucleotide regions of the genomic and antigenomic RNA 3’ termini respectively
Transcription start sites are 12 or 14 nucleotides in length and end in the consensus sequences 3’-
CUUCUAAUU EBOV and 3’-CUURUAAUU for MARV, while stop sites are 11 or 12
nucleotides long with the conserved sequence 3’-UAAUUC (U)5/6.
Polyadenylation is believed to occur by slippage or stuttering of the polymerase at the five or six
uridines ending the stop sites. Gene overlaps which are short (18-21nucleotides) does not affect
either the polyadenylation of upstream gene or initiation of transcription of downstream gene
because the transcription of VP40 and VP35 genes of ZEBOV is substantial and expression of
VP40 is very strong.
mRNAs have noncoding regions at their 3’ and /or 5’ ends which contribute to the increase length
of genome and may function in the stability of transcripts. 5’ ends of transcripts also have
potential to form stem loop structures that might affect their stability and ribosome binding
capacity/transcription
ZEBOV VP30 has a transcription activation property that is linked to a RNA secondary structure
formed at the 5’ end of the NP gene transcript as it is synthesized. The presence of VP30 is
required for transcription of down stream genes. This property is impaired by phosphorylation at
six serines and one threonine at the N-terminus and can be restored by action of cellular
22

phosphatase. In virion VP30 is partially phosphorylated and it may be the actions of phosphatase
on nucleocapsid required for before transcription proceed efficiently. Corresponding mechanism
has not be found with MARV
Filovirus genes are monocistric (each mRNA is transcribe into one protein products). The
proteins found can be subdivided into those that form nucleocapsid (NC) and those that are
associated with the envelope. The nucleocapsid-associated proteins are involved in transcription
and replication of genome whereas envelope-associated proteins are involved with either viral
assembly or virus entry.
Nucleoproteins: NP and VP30 are the major and minor nucleoprotein respectively and interact
strongly with the genomic RNA molecule to form the viral nucleocapsid. NP and VP30 are
phosphoproteins. NP has a conserved hydrophobic N-terminal half which contains all the cysteine
residues and a divergent hydrophilic C-half which contains most of the proline residues and is
extremely acidic. NP has a predicted mass of 20KDa smaller than the SDS-PAGE migration
attributed to as a result of binding to SDS molecule t negatively charge NP. The central region of
NP is highly conserved region
N-terminal third of VP30 contained a high concentration of basic amino acids (primarily arginine
residues) and may be involve in binding to virus RNA and/or acidic C-terminal of NP. VP30
contain zinc-finger motif ~70-80 residues from the N-terminus that is highly conserved in
filoviruses. ZEBOV VP30 behaves as a transcription activator that is regulated by
phosphorylation but appear absent in MARV.
POLYMERASE COMPLEX PROTEINS:
L and VP35 proteins form the polymerase complex, which acts to transcribe and replicate
filovirus genome. L provides RNA-dependent RNA polymerase activity of the complex, and
motifs linked RNA (template) binding, phosphodiester bonding and a ribonucleotide triphosphate
bonding occurs. It is the largest and least abundant protein
VP35 is a Cofactor in transcription and Replication (Cofactor in polymerase complex). VP35 is
believed to have an essential role as a cofactor that affects the mode of RNA synthesis
(transcription or replication) and acts as a linker between L and NP. VP35 has an antagonistic
effect on the interferon type1 pathway.
MATRIX PROTEIN:
23

VP40 function as matrix protein with VP24 may have a secondary/minor matrix protein function.
VP40 is most abundant protein in the virion while small amount of VP24 are incorporated into
virus particle. VP40 is a Matrix protein necessary for Virus assembly and budding. It forms
hexamers when it contacts the plasma membrane which confers added stability during assembly.
It is the most abundant protein. Both have affinity for membranes and are associated with the
virion envelope. They are easily released from virion by nonionic detergents under low salt
condition. VP40 is critical to the budding process because it initiates and drives the envelopment
of NC by the plasma membrane. VP24 precise role in replication is unclear but in ZEBOV it has
a reported in IFN type1 signaling pathway it has an antagonist activity
GLYCOPROTEIN GP
MARV GP gene encodes a single product GP, in a conventional open reading frame (ORF). It is
the fourth gene in the genome from 3’ to 5’ direction. EBOV GP gene organization and
transcription is quite unusual and distinct from MARV.
EBOV encodes their GP in two reading ORFs (0 and -1). Expression of EBOV GP requires a
transcriptional editing event. Translation of unedited transcript GP gene results in the production
of SGP, a smaller, non structural, secreted glycoprotein, the primary gene product.
24

Figure 14
In the edited transcript, the transcriptional editing event occurs at a series of seven uridines on the
genome RNA template and results in the insertion of an additional adenosine, which connects the
GP open reading frames. This may be due to stuttering of the polymerase on the poly U template
during polyadenylation. Figure 14
Approx 20-25% of the transcript are edited and this single nucleotide editing site occurs with high
degree of fidelity. The nucleotide sequence corresponding to editing site is absent in MARV GP
gene. This results in the production of both a truncated, soluble form of the Ebola virus
glycoprotein (sGP; 50-70 kD in size) and a full-length, transmembrane anchored version of the
same protein (GP; 120-150 kD in size). Ebola virus sGP and GP have different functional
properties, which may be important in disease pathogenesis. The functional subdomains of these
molecules are shown in figure 15. The soluble sGP molecule is secreted as a trimer, and is identical
at its N-terminus to the homologous region of the transmembrane glycoprotein (GP). sGP
interacts with neutrophils through CD16b, the neutrophil-specific form of the Fc © receptor III,
whereas the transmembrane glycoprotein (GP) interacts with endothelial cells but not with
neutrophils (Yang et al. Science 279:1034, 1998). It is possible that interaction of sGP with
25

neutrophils results in the blockade of early events in the activation of these cells, thereby
inhibiting inflammatory responses which might contribute to innate protection against viral
infection. sGP may also act as a "decoy" for antiviral antibodies.
GP The transmembrane glycoprotein is produced as a long precursor, which undergoes cleavage
by a cellular protease (furin), to produce GP1 and GP2. These can be viewed as being somewhat
analogous to HIV-1 gp120 and HIV-1 gp41 (which are produced by cellular proteolytic cleavage
of the gp160 precursor). Ebola virus GP2 remains in the membrane (due to its transmembrane
domain) and is responsible for mediating fusion between the virus and the plasma membrane, via
its fusion domain. The GP1 component is attached to GP2 via a non-covalent linkage, and is
thought to mediate virus attachment to its host cell(s), which include vascular endothelial cells.
Figure 11 Ebola virus GP is also cytotoxic for vascular endothelial cells in vitro, and this is thought
to contribute to the virus’ ability to trigger vascular leakage (hemorrhage) in vivo.
In the Trans-Golgi, the precursor molecule (GP0) is post-translationally cleaved by furin at Furin
cleavage site (Arg-Arg-X-Arg/LYS-Arg) yielding a heterodimer, (GP1-GP2). (Marburg, cleavage
site is more toward N-terminus). Heterodimer is linked together by one disulfide bond, a cysteine
bridge. Figure 11
Figure 16
NON STRUCTURAL GLYCOPROTEIN (sGP)
It’s expressed in EBOV viruses only. The N-terminal ~300 amino acids are identical to the
structural GP but C terminus is unique in the sequence. It is produce from a precursor molecule
cleaved by furin (or furin-like endoprotease) near the C-terminus to release a short peptide that
contain O-linked glycan and name delta peptide. Delta has no identified biological activity. sGP
is structurally distinct from GP and is secreted from infected cells as a homodimer that is likely
form in endoplasmic reticulum. It has a parallel orientation of the dimmer and held together by
disulphide bonds between N- and C terminus cysteine. figure 17 It circulate in large amount in
acutely infected human blood so is likely to contribute to disease progression
26

Figure17
ASSEMBLY AND RELEASE
When sufficient levels of negative-sense nucleocapsids and envelope-associated proteins are
reached, coalescence of these components occurs at the plasma membrane or, to a lesser extent, at
membranes forming intracellular vacuoles. Inclusion bodies forms in infected cells are induced to
form by NP but contain other proteins that form the NC. These structures are facilitated by
expression of VP35 and VP24. NC is believed to interact with VP40 molecules in the budding
process.
27
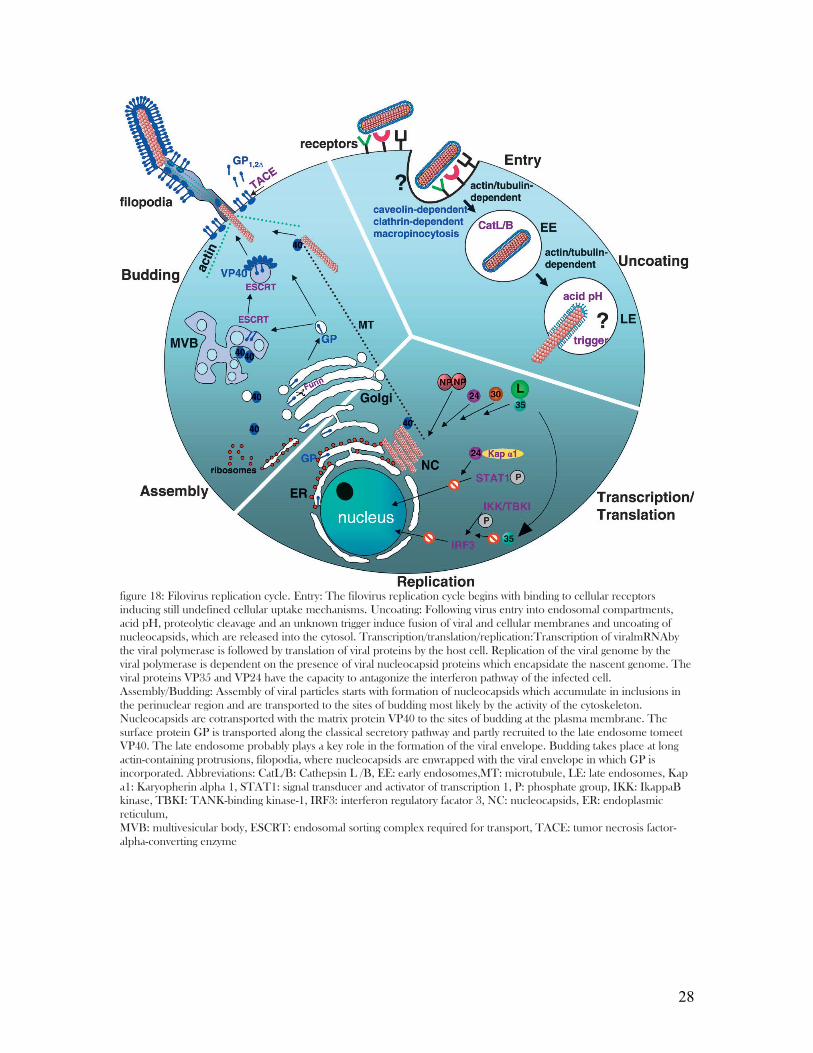
figure 18: Filovirus replication cycle. Entry: The filovirus replication cycle begins with binding to cellular receptors inducing still undefined cellular uptake mechanisms. Uncoating: Following virus entry into endosomal compartments, acid pH, proteolytic cleavage and an unknown trigger induce fusion of viral and cellular membranes and uncoating of nucleocapsids, which are released into the cytosol. Transcription/translation/replication:Transcription of viralmRNAby the viral polymerase is followed by translation of viral proteins by the host cell. Replication of the viral genome by the viral polymerase is dependent on the presence of viral nucleocapsid proteins which encapsidate the nascent genome. The viral proteins VP35 and VP24 have the capacity to antagonize the interferon pathway of the infected cell. Assembly/Budding: Assembly of viral particles starts with formation of nucleocapsids which accumulate in inclusions in the perinuclear region and are transported to the sites of budding most likely by the activity of the cytoskeleton. Nucleocapsids are cotransported with the matrix protein VP40 to the sites of budding at the plasma membrane. The surface protein GP is transported along the classical secretory pathway and partly recruited to the late endosome tomeet VP40. The late endosome probably plays a key role in the formation of the viral envelope. Budding takes place at long actin-containing protrusions, filopodia, where nucleocapsids are enwrapped with the viral envelope in which GP is incorporated. Abbreviations: CatL/B: Cathepsin L /B, EE: early endosomes,MT: microtubule, LE: late endosomes, Kap a1: Karyopherin alpha 1, STAT1: signal transducer and activator of transcription 1, P: phosphate group, IKK: IkappaB kinase, TBKI: TANK-binding kinase-1, IRF3: interferon regulatory facator 3, NC: nucleocapsids, ER: endoplasmic reticulum,MVB: multivesicular body, ESCRT: endosomal sorting complex required for transport, TACE: tumor necrosis factor-alpha-converting enzyme
28

Membrane/lipid rafts are platforms for the assembly of filovirus virion. They are rigid
microdomains (containing sphingolipids and cholesterol) present in biological membrane and are
isolated from the fluid phospholipids surround them. GP trimers conveyed to the surface
membrane have affinity for these rafts.
Posttranslational processing and intracellular trafficking of VP40 results in deposition of VP40 at
the plasma membrane via the late retrograde endosomal pathway. VP40 ZEBOV is capable of
mediating its own release from mammalian cells to form enveloped virus like particles which are
more efficiently produced what GP and NP are present. VP40 interacts with C-terminus of NP.
VP40 is bond as an oligomeric form and ubiquitinated and subsequently target to endosome by
TSG 101 and VPS 4 (components of vacuoles sorting pathway and is recruited to membrane rafts
through TSG 101 interaction with VP40 and protein raft.. raft-associated VP40 believed to
associate with NCs drawing then tightly to the membrane where they are enveloped and extruded
from host cell as infectious virions
CLINICAL FEATURES
29

Filoviruses infections are generally the most severe of viral haemorrhagic fevers. Abrupt onset
follows an incubation period of 2-21days (Ebola) and 3-9days (Marburg), averaging 4 to 10days.
It’s characterized by flue-like symptoms of fever, chills, malaise, and myalgia.
There is subsequent signs and symptoms of systemic involvement like prostration,
gastrointestinal (anorexia, nausea, vomiting, abdominal pain, diarrhea), respiratory (chest pain,
shortness of breathing, cough), vascular (conjunctival injection, postural hypotension, edema),
and neurologic (headache, confusion, coma) manifestation. Haemorrhagic manifestations develop
during the peak of the illness and include petechiae, ecchymoses, uncontrolled oozing from
venipuncture sites, mucosal hemorrhages and visceral hemorrhagic effusions. The is often a
mucopapular rash associated with varying degrees of erythema at days 5-7 of the illness, this is a
valuable differential diagnostic feature and is usually follow by desquamation in survivors.
Abdominal pain is sometimes associated with hyperamylasemia and true pancreatitis.
In late stages shock, convulsions, severe metabolic disturbances, and, in more than half the cases,
diffuse coagulopathy supervenes.
LABORATORY parameters a less characteristic
Early leucopenia with lymphopenia and subsequent neutropenia, left shift with atypical
lymphocytes, thrombocytopenia, marked elevated serum transaminase level (AST typically
exceeding ALT), hyper proteinemia and proteinuria. Prolonged prothrombin and partial
thromboplastin time, fibrin split products detectable
In later stage, secondary bacteria infection may lead to elevated white blood count. Nonfatal
cases have fever for about 5-9days, and improvement typically occurs around days 7-11days
about the time humoral antibody response. Convalescence is prolonged and sometimes
associated with myelitis, recurrent hepatitis, and psychosis, uveitis. There is an increase risk of
abortion for pregnant women and high death rate in children of infected mothers.
Fatal cases develop clinical signs early during infection and demise typically occurs between days
6 and 16 due to haemoorrhage and hypovolaemic shock
DIAGNOSIS AND PREVENTION
Diagnosis is mainly clinical with high index of suspicion. Filovirus haemorrhagic fever should be
suspected in anyone with clinical signs and symptoms with history of travel to endemic area,
jungles or caves exposure, treatment in local hospital, contact with sick person or wild/domestic
animals like monkey, ape or even pigs. Wide ranges of febrile disease most be considered and
excluded like malaria typhoid etc.
30

LABORATORY DIAGNOSIS.
Clinical microbiology and public healthy labs are ill equip to make diagnosis, so specimens
should be send to national and international reference labs capable of performing require test.
Universal precaution in patient care and sample collections to avoid direct contact with body fluid
and secretions.
Measurement of host specific immune response for detection of IgG and IgM through ELISA.
Viral particles or particle components through RT-PCR, antigens capture ELISA or immunoblot.
Electromicroscope can be use to identify virus in infected cells or through
immunohistochemistry.
Prevention is through avoidance of contact with body fluids, strick barrier nursing with use of
protective clothes and proper disposal of body secretion and fatal caderva.
Vaccine trail have prove effective in primate but it yet to be tested in human as the disease occurs
in sporadic.
TREATMENT
Is mainly supportive directed towards maintenance of effective blood volume, electrolyte
balanced. Management of shock, cerebral edema, renal failure, coagulation disorders and
secondary bacteria infection may be life threatening.
31





























