THESE DE DOCTORAT DE
L’UNIVERSITE PIERRE ET MARIE CURIE
Ecole Doctorale
Diversité du Vivant (ED 392)
Présentée par
M. Terry BRISSAC
Pour obtenir le grade de
DOCTEUR de l’UNIVERSITE PIERRE ET MARIE CURIE
Spécialité
Microbiologie et Interactions durables
Nature, Diversité et Spécificité de l’Association Lucinidae / Bactéries sulfo-oxydantes.
Soutenue publiquement le 01 octobre 2009 devant le jury composé de :
Dr. Nicole DUBILIER, Head of Symbiosis Group, Max Planck, Bremen Examinatrice
Pr. Olivier GROS, Prof. Univ. Antilles-Guyane Co-directeur
Pr. Vincent MARECHAL, Prof. Univ. Paris VI Examinateur
Dr. Hervé MERÇOT, CR, HDR, CNRS, Univ. Paris VI Directeur
Dr. Christine PAILLARD, CR, HDR, CNRS, IUEM, Brest Rapportrice
Dr. Guennadi SEZONOV, MCU, HDR, Univ. Paris VI Examinateur
Dr. Fabrice VAVRE, CR, HDR, CNRS, Univ. Lyon 1 Rapporteur
UMR 7138 UPMC - CNRS – MNHN - IRD
« Systématique, Adaptation, Evolution »
Equipe : « Génétique & Evolution » Universite Pierre et Marie Curie (Paris VI)
7, quai St Bernard
75252 PARIS CEDEX 05


REMERCIEMENTS
Je remercie le professeur Hervé Le Guyader de m’avoir accueilli dans son unité.
Je remercie Christine Paillard et Fabrice Vavre qui ont accepté d’être rapporteurs de cette thèse.
Je remercie également Nicole Dubilier, Vincent Maréchal et Guennadi Sezonov pour leur présence au sein de mon jury.
Je tiens aussi à remercier Françoise Gaill et à renouveler mes remerciements à Christine et Guennadi pour avoir accepté de faire partie de mon comité de thèse. Leurs questions et conseils lors des différentes réunion m’ont ainsi permis de prendre du recul et d’avoir une autre vision de mon sujet.
Je remercie bien sûr mes deux directeurs de thèse, Hervé et Olivier.
Hervé qui me suit depuis maintenant cinq ans, avec qui les discussions scientifiques ont quelques fois été longues et douloureuses, m’a permis de progresser et ce surtout dans la rédaction et la présentation. J’ai grâce à lui acquiq l’expérience nécessaire pour mener à bien ma quête scientifique.
Olivier qui malgré la distance géographique et le décalage horaire a toujours été disponible et de bon conseil, rendant par la même mon travail aussi plaisant à Paris qu’en Guadeloupe. Merci pour ton expertise sur les Lucinidae et de m’avoir laissé toute latitude dans l’orientation de mon travail, et le test de mes hypothèses aussi farfelues soient-elles.
Je tiens à remercier Dominique Higuet, chef d’equipe, président d’EURO-DH et tant d’autres qui malgré un emploi du temps surchargé a toujours été présent pour discuter, échanger et surtout réfléchir aux résultats et à l’avenir. J’esperes ne pas t’avoir trop fait te servir de ton tampon et suis sur que tu continueras à en faire bon usage.
Je voudrasi remercier Eric Bonnivard pour son aide et ses conseils manips ou autre ; Un grand merci pour la mèche porte-bonheur qui aurait pu débloquer SAS, MS et PCR en tout genres.
Je remercie Clémentine Vitte et Denis Poinsot, relecteurs bilingues sans qui, compte tenu de mon anglais, publier se serait révélé être une tâche nettement plus difficile.
Je remercie aussi les autres thésards côtoyés durant cette periode Anna et Yves qui dans leur rôle d’anciens m’ont beaucoup apporté par leur partage de l’expérience de la thèse (vous ne pourrez plus m’appeler « petit thésard » desormais) ; Etienne, Vincent et bien sûr Mathieu (co-fondateur d’EURO-DH) sans qui déprimer aurait surement été moins « fun », le « jukebox musical » moins

varié et flécher les pièces manips plus long. Il faut maintenant te remetre d’aplomb car il va y avoir beaucoup de choses à fêter et car tu seras maintenant seul pour « sniper » Eric & Dominique.
Je voudrais remercier Paula toujours souriante et de bonne humeur qui apporte ce brin de folie et de joie rendant les journées de travail plus légères. Merci aussi pour toute l’aide que tu fournie pour la préparation du grand jour.
Je remercie les autres membres de l’équipe « Génétique & Evolution », Micheline « ça va les jeûnes », Denis notre ACMO (Agent Chargé de la Manutention d’Objets ou déménageur), Guillaume, Isabelle, Pierre et Sophie qui même si la géographie « Paris-6-enne » (tout du moins avant ces 2 derniers mois) nous a éloignés ont bien voulu m’écouter lors de mes nombreuses répétitions et répondre à mes interrogations.
Je remercie Eric Bapteste pour les discussions sur la congruence et autres déboires phylogénétiques.
Je remercie Catherine Ozouf-Costaz et Jean-Pierre Coutanceau qui ont accompagné mes premières pérégrinations dans le monde du FISH.
Je remercie Danielle Merkiled et Phillippe Leballeur sans qui la gestion des tâches administratives (et bien plus encore) serait plus difficile, toujours disponibles et efficace spour régler tous les problèmes.
Je remercie également les Membres de l’équipe « Symbiose » Sylvie, Jean-Luc pour leur accueil lors des 2 missions scientifiques (j’insiste) que j’ai effectuées en Guadeloupe ; mais aussi les doctorants Amandine, Leslie, Melina, Nathalie, Jean-Marie, Pedro, Sophie et Yann.
Je remercie les membres de l’équipe « Répression épigénétique et éléments transposables », Anne-Laure (même si elle n’en fait plus partie), Augustin, Laure, Stéphane et Valérie, les voisins de la fameuse Tour42, pour leur présence, écoute et tou ce qui s’en suit.
Egalement un grand merci aux amis, Anna, Arnaud, Aurelia, François, Guillain (deux fois merci pour toi), Magali, Orso, Sandra, Alexandre et Nicolas (la pipoteam toujours vivante !) et ceux que j’aurais oublié (navré) pour les soirées, les pots et tous les petits moments passés à parler de tout de rien, bref à profiter de la vie.
Je tiens également à remercier le club de foot des « Suprêmes Béliers », soupapae de sécurité après les semaines « ras-le-bol ».
Je souhaiterais également et surtout remercier mes parents qui m’ont toujours soutenu et encouragé dans tous mes projets, cette réussite leur appartient tout autant.
Merci aussi à ma tante Nicole qui a permis mon adaptation rapide après mon arrivée de Guadeloupe.

Et bien sûr et surtout Clémentine, toi qui as été mon premier soutien dans les moments difficiles, c’est aussi grâce à toi que j’ai pu en arriver là. Il y a tant à dire sur ce que tu as fait (et continue à faire) que je ne te remercierais jamais assez.
Enfin merci à tous ceux qui de près ou de loin ont contribué à la réussite de cette thèse, sans oublier les palourdes qui en torturant ces pauvres bactéries ont permis l’émergence de ce manuscrit.


TABLE DES MATIERES
REMERCIEMENTS ______________________________________________ 3
TABLE DES MATIERES __________________________________________ 7
LISTE DES FIGURES ____________________________________________ 9
LISTE DES TABLEAUX _________________________________________ 11
RESUME ______________________________________________________ 15
INTRODUCTION _______________________________________________ 21
LA SYMBIOSE ____________________________________________________________ 21
LES ASSOCIATIONS CHIMIOAUTOTROPHES ______________________________________ 26
Diversité des hôtes ________________________________________________________________ 26
Diversité des symbiotes ____________________________________________________________ 27
Diversité métabolique ______________________________________________________________ 27
Diversité d’ « infection » ___________________________________________________________ 28
LES LUCINIDAE __________________________________________________________ 30
LES LUCINIDAE __________________________________________________________ 30
NATURE DE L’ASSOCIATION ___________________________________ 41
INTRODUCTION __________________________________________________________ 41
DEMARCHE EXPERIMENTALE________________________________________________ 43
RESULTATS _____________________________________________________________ 45
DISCUSSION _____________________________________________________________ 47
CONCLUSION ____________________________________________________________ 50
DIVERSITE BACTERIENNE _____________________________________ 57
DIVERSITE INTERSPECIFIQUE _______________________________________________ 57
DEMARCHE EXPERIMENTALE. _______________________________________________ 60
EVOLUTION DES COUPLES SYMBIOTIQUES ______________________________________ 64

DIVERSITE INTRA-SPECIFIQUE. ________________________________________ 71
RECHERCHE DE MARQUEURS DE DIVERSITE. ____________________________________ 73
DEMARCHE EXPERIMENTALE. _______________________________________________ 75
RESULTATS ___________________________________________________________ 82
DISCUSSION _____________________________________________________________ 91
SPECIFICITE D’ASSOCIATION _________________________________ 101
DISCUSSION GENERALE ______________________________________ 109
NATURE DE LA RELATION __________________________________________________ 111
UNE NOUVELLE VISION DE L’ASSOCIATION ____________________________________ 112
LUCINIDAE/BACTERIES SULFO-OXYDANTES. ___________________________________ 112
BIBLIOGRAPHIE _____________________________________________ 117
ANNEXE 1 : STRUCTURE DES BRANCHIES DE LUCINIDAE __________________________ 131
ANNEXE 2 : LISTE DES BACTERIES UTILISEES DANS LE CADRE DE LA RECHERCHE DE
MARQUEURS. ___________________________________________________________ 132
ANNEXE 3: PROFIL « SVARAP » DU GENE DNAE. ______________________________ 134
ANNEXE 4 : TECHNIQUES PARTICULIERES UTILISEES. ___________________________ 135
ARTICLE 1 : BRISSAC ET AL. (2009) ________________________________________ 139
ARTICLE 2 : BRISSAC ET AL. ______________________________________________ 149
ARTICLE 3 : BRISSAC ET AL. ______________________________________________ 171
DISCUSSION ____________________________________________________________ 178

LISTE DES FIGURES
Figure 1. Représentation schématique de la taxonomie des bivalves chimiosymbiotiques. ................................ 30
Figure 2 Diversité de formes chez les Lucinidae. ............................................................................................. 31
Figure 3. Habitat typique des Lucinidae. ......................................................................................................... 32
Figure 4. Mangrove de palétuviers, Rhizophora mangle. ................................................................................. 32
Figure 5. Herbier à Thalassia testudinum. ....................................................................................................... 32
Figure 6. Anatomie générale des Lucinidae. .................................................................................................... 33
Figure 7. Représentation schématique de la transmission environnementale. ................................................... 41
Figure 8. Analyse des différentes sous-populations bactériennes retrouvées dans les branchies de divers
individus de l’espêce Codakia orbicularis. ...................................................................................................... 42
Figure 9. Recherche de symbiotes re-largués. ................................................................................................. 46
Figure 10. Suivi de la population symbiotique à l’intérieur des branchies........................................................ 47
Figure 11. Représentation schématique du recrutement environnemental. ........................................................ 50
Figure 12. Carte présentant les lieux d’échantillonnage des différentes espèces de Lucinidae analysées. .......... 61
Figure 13. Phylogénie moléculaire des Lucinidae réalisée après concaténation des séquences des ARNs
ribosomaux 18S et 28S (1252 sites). ................................................................................................................ 65
Figure 14. Phylogénie moléculaire des symbiotes chimioautotrophes réalisée grâce aux séquences d’ARNr 16S
(1269 sites). .................................................................................................................................................... 67
Figure 15. Phylogénie moléculaire des symbiotes chimioautotrophes réalisée grâce aux séquences d’APS
Réductase (273 sites). ..................................................................................................................................... 68
Figure 16. Exploration des bases de données................................................................................................... 74
Figure 17. Carte des zones échantillonnées pour l’analyse de la diversité intra-spécifique. .............................. 76
Figure 18. Indice de variabilité inter-spécifique des différents gènes candidats comparés à celui de l’ARNr 16S.
....................................................................................................................................................................... 84
Figure 19. Indice de variabilité intra-spécifique des différents gènes candidats comparés à celui de l’ARNr 16S
pour différentes souches d’Escherichia coli et de Legionella pneumophila. ...................................................... 85
Figure 20. Phylogénie moléculaire des bactéries basée sur des séquences d’ARN ribosomal 16S (1354 sites). . 86
Figure 21. Phylogénies moléculaires des bactéries basées sur des séquences des différents marqueurs candidats
(dnaE : 1630 sites ; groEL : 1521 ; recA : 906 ; gyrB : 1473 ; polA : 2059 ; rpoB : 2604). .............................. 87
Figure 22. Phylogénie moléculaire des Lucinidae réalisée après concaténation des séquences des ARNs
ribosomaux 18S et 28S (1386 sites). ................................................................................................................ 89
Figure 23. Réseaux d’haplotypes reconstruits à partir des séquences obtenues pour chacun des marqueurs ..... 90
Figure 24. Représentation schématique de la distribution des hôtes et des symbiotes dans les sédiments des
herbiers à T. testudinum. ................................................................................................................................. 96
Figure 25. Représentation schématique du processus évolutif pouvant conduire à la structuration de la diversité
bactérienne observée dans les herbiers à T. testidunum de l’Îlet Cochon. ......................................................... 99
Figure 26. Représentation schématique du choix du partenaire. .................................................................... 100

Figure 27. Hypothèses de contrôle de l’acquisition du symbiote par l’hôte selon (Gros et al., 2003a). ............ 104
Figure 28. CARD-FISH réalisé à l’aide de la sonde Symco2A. ...................................................................... 106

LISTE DES TABLEAUX
Tableau A. Différents types de relations entre espèces en fonction de l’effet de l’interaction sur le succès
reproducteur des partenaires. ......................................................................................................................... 24
Tableau B. Echantillonnage de Distel et al. (1994; 1988). ............................................................................... 59
Tableau C. Echantillonnage de Durand et al. (1996) ; Durand & Gros (1996) ; Gros et al. (2000; 2003a). ...... 59
Tableau D. Echantillonnage réalisé pour cette étude de la diversité des symbiotes de Lucinidae ...................... 62
Tableau E. Amorces et conditions de PCR utilisées dans cette étude. ............................................................... 63
Tableau F. Matrice d’identité réalisée en « pairwise deletion » utilisant les séquences d’ARNr 16S. ................ 69
Tableau G. Gènes-candidats retenus à partir de la littérature. ......................................................................... 77
Tableau H. Amorces utilisées au cours de l’étude de la diversité intra-spécifique. ............................................ 81
Tableau I. Résultats de l’Au-test. ..................................................................................................................... 83
Tableau J Analyse du polymorphisme. ............................................................................................................. 91


RRREEESSSUUUMMMEEE


RESUME
15
RESUME
Les associations entre « invertébrés » marins et bactéries chimioautotrophes (i.e.
capable de fixer le carbone à partir de composés inorganiques) présentent une grande diversité
tant au niveau taxonomique, métabolique qu’écologique. Dans le cadre de cette thèse, je me
suis intéressé aux Lucinidae, famille de bivalves qui héberge dans des cellules spécialisées de
leurs branchies (les bacteriocytes), des gamma-protéobactéries endosymbiotiques,
thioautotrophes (i.e. sulfo-oxydantes). Mon travail a porté sur l’étude : (i) de la nature réelle
de cette association, (ii) de la diversité génétique des symbiotes associés et de la spécificité du
couple hôte/symbiote.
(i) Ces associations sont essentiellement décrites dans la littérature comme mutualistes
(i.e. avantageuses pour les deux partenaires). La nature des associations symbiotiques étant
souvent lié au mode de transmission du symbiote, l’hypothèse mutualiste est recevable dans
les cas ou le symbiote est transmis verticalement (e.g. Vesicomyidae). Cependant, certains
arguments peuvent conduire à une hypothèse alternative pour les modèles dits à
« transmission environnementale ». Ce mode de transmission suppose un passage du
symbiote de la génération parentale à la suivante via l’environnement. Ce schéma implique
donc la multiplication du symbiote dans l’hôte, son relarguage par ce dernier et la présence de
la forme libre du symbiote dans l’environnement. Après analyse, il apparait qu’il n’y a ni
multiplication ni relarguage du symbiote chez les Lucinidae. Cette association ne serait donc
pas mutualiste, l’entrée en symbiose constituant ainsi pour la bactérie une impasse évolutive.
(ii) L’analyse de la diversité des symbiotes indique que les couples actuels ne
proviennent pas de la co-évolution des deux partenaires à partir d’une symbiose ancestrale. En
effet, au sein de 18 espèces hôtes analysées seules 3 espèces bactériennes ont pu être décrites,
l’espèce majoritaire étant hébergée par des hôtes colonisant les herbiers et les deux autres par
des espèces de mangrove. L’association semble donc être labile et se constituer en fonction de
l’environnement. Cependant, une analyse MLST a révélé l’existence d’une diversité
symbiotique intra-spécifique. Cette diversité de souches serait structurée au sein d’une même
localité en fonction des hôtes présents. Ceci indiquerait une spécificité d’association souche
bactérienne/espèce hôte qui pourrait être due à des processus évolutifs de type « Red Queen ».
Une expérience d’infection croisée réalisée entre Codakia orbicularis et C. orbiculata atteste
de cette spécificité d’association. Il semble néanmoins qu’une certaine plasticité puisse
intervenir au cours du stade juvénile de l’hôte.
Les résultats de mon travail de thèse supportent l’hypothèse que l’association
Lucinidae/bactéries sulfo-oxydantes ne serait pas mutualiste. L’association ne se constituerait
qu’en faveur de l’hôte et selon un modèle d’évolution antagoniste. Le modèle Lucinidae
présente de nombreux points communs avec certains modèles hydrothermaux ou de
suintements froids traditionnellement considérés comme mutualistes à transmission
environnementale. Dès lors se pose naturellement la question de l’extension de nos
conclusions à ces modèles profonds.


« Tout est pour le mieux dans le
meilleur des mondes possibles »
(Dr Pangloss ; Candide, Voltaire)
Pas si sûr…


IIINNNTTTRRROOODDDUUUCCCTTTIIIOOONNN


INTRODUCTION
21
INTRODUCTION
LA SYMBIOSE
e terme même de symbiose, tel qu’il a été défini par Anton De Bary en 1879,
englobe « toute association étroite et durable entre deux organismes
hétérospécifiques ». De fait, aussi bien le mutualisme (où les bénéfices sont
réciproques pour les deux partenaires) ou à l’inverse le parasitisme (association à bénéfice
unilatéral) en passant par le commensalisme (relation bénéfique pour un partenaire et sans
incidence sur l’autre) sont des symbioses. Cependant le terme « symbiose » est souvent
assimilé de façon restrictive aux seules relations à bénéfices mutuels. Dans la suite de ce
manuscrit, le terme de symbiose sera employé suivant la définition initiale de De Bary.
Ces associations d’êtres vivants hétérospécifiques mais nécessairement compatibles
(afin que l’association puisse d’une part se constituer et d’autre part durer) constituent un des
facteurs majeurs dans l’évolution des espèces. Les associations entre organismes eucaryotes
typiquement hétérotrophes (source de carbone organique) avec des microorganismes
autotrophes (source de carbone inorganique) illustrent très bien ce dernier point. L’association
de caractéristiques métaboliques ou physiologiques différentes permet ainsi l’expression d’un
potentiel nouveau en réponse à l’environnement. Cette dernière permet à l’hôte1 de mieux
exploiter sa niche écologique ou d’en exploiter une nouvelle. Le symbiote quant à lui, trouve
dans cet hôte un nouvel environnement où son succès reproducteur est accru (e.g. protection,
meilleures conditions de reproduction, meilleure transmission).
Les modifications génotypiques, phénotypiques ou comportementales observées chez
certaines espèces sont autant de preuves qui attestent du caractère évolutif de ces associations.
On retrouve ainsi au niveau génotypique la réduction des génomes bactériens (pour revue,
voir Delmotte et al., 2006; Gomez-Valero et al., 2007; McCutcheon et al., 2009; Yu et al.,
2009) qui correspond à la perte de régions génomiques ou à la perte de gènes codants pour
des fonctions devenues inutiles ou non essentielles du fait de l’association. Autre cas, celui
1 D’une façon générale et par commodité, au sein des associations symbiotiques le plus gros partenaire est nommé hôte et le plus petit symbiote.
L

INTRODUCTION
22
des transferts de gènes entre symbiote et hôte (Blaxter, 2007). C’est l’exemple de
l’association entre le puceron du pois Acyrthosiphon pisum et la gamma-protéobactérie
Buchnera aphidicola (Gil et al., 2004; Nikoh and Nakabachi, 2009). Cette bactérie possède en
effet l’un des plus petits génomes connus (entre 420 et 650kb contenant de 400 à 600 gènes).
En dépit de la perte de gènes bactériens essentiels, cette bactérie survit grâce à l’intégration et
surtout à l’expression de ces gènes transférés au génome de l’hôte (Nikoh and Nakabachi,
2009). D’autres modifications génotypiques sont retrouvées essentiellement dans les relations
de type parasitaire. En effet, dans ce type de relations où le parasite « veut s’installer » et
l’hôte « lui échapper » nombre de modifications se produisent et sont sélectionnées. Ainsi va
la « sélection des meilleurs1 parasites par les hôtes et la sélection des meilleurs hôtes par les
parasites » (Combes, 2003). Dans ces modèles, les « gènes pour se rencontrer » chez le
parasite et les « gènes pour s’éviter » chez l’hôte évoluent de façon antagoniste tout comme
les « gènes pour tuer » et les « gènes pour survivre ». Les deux partenaires évoluent ainsi sans
cesse dans une sorte de course aux armements2 (Van Valen, 1973) où le plus important est le
succès reproducteur (fitness). Enfin, l’accélération du taux d’évolution moléculaire des endo-
symbiotes transmis verticalement constitue un autre exemple de modification génotypique
liée à la symbiose. Dans ces modèles, l’effet de la dérive génétique est amplifié par
l’existence de goulots d’étranglements au moment de la transmission des symbiotes (Peek et
al., 1998b; Woolfit and Bromham, 2003).
Au niveau phénotypique, différentes situations peuvent se présenter. Il peut s’agir de :
(i) l’apparition de cellules spécialisées tels que les bactériocytes des bivalves (Fisher, 1990)
ou des insectes (Buchner, 1965), (ii) la formation d’organes spécialisés dans la culture des
symbiotes comme l’organe lumineux du calmar Euprymna scolopes (Boettcher and Ruby,
1990; McCann et al., 2003) ou les nodules dans le cas de l’association
Rhizobium/Légumineuses (Beringer et al., 1979; Chen et al., 2007), (iii) la modification des
organes ou tissus de l’hôte telle que la réduction du tractus digestif (Fisher, 1990).
Enfin, au niveau comportemental, certains parasites sont capables de « médier » le
comportement de leur hôte de façon à faciliter leur transmission ou leur accès à différentes
ressources. Ainsi, chez la palourde Venerupis aurea, hôte intermédiaire de Meiogymnophallus
fossarum, la présence du parasite induit chez ce bivalve un changement d’orientation dans les
1 On entend ici par meilleur, le partenaire capable d’évoluer pour ne pas disparaitre. 2 Ce processus évolutif correspond à la théorie de la Reine Rouge proposée par Leigh Van Valen .

INTRODUCTION
23
sédiments. Ceci a pour effet de rendre la palourde plus accessible à son prédateur
Haematopus ostralagus qui est l’hôte final du parasite (Bartoli, 1978). Ces dernières
modifications ont été décrites sous le nom de phénotype étendu1 (Dawkins, 1982).
Enfin, l’exemple le plus remarquable de l’importance évolutive de ces associations
symbiotiques reste l’origine bactérienne de certains organites de la cellule eucaryote tels que
les mitochondries (anciennes alpha-protéobactéries) ou les chloroplastes (anciennes
cyanobactéries) (Margulis and Fester, 1991).
Entre mutualisme et parasitisme il existe différentes relations plus ou moins néfastes
ou bénéfiques pour chacun des partenaires (Tableau 1). Cependant, il existe des cas où la
nature des relations symbiotiques reste difficile à définir tant les bénéfices ou les coûts pour
les partenaires sont parfois subtils à déterminer. Ces avantages ou désavantages sont
généralement définis en fonction de l’effet de l’association sur le succès reproducteur des
deux partenaires.
La transmission du symbiote constitue également un élément essentiel à prendre en
compte. Cette dernière constitue à priori la pierre angulaire de toute association symbiotique.
En effet, le passage du symbiote d’une génération à l’autre permet la persistance de
l’association au cours du temps. Cette transmission peut être (i) verticale : passage du
symbiote de la génération parentale à la génération fille via les gamêtes (donc entre individus
directement apparentés) (ii) horizontale : échange de symbiotes (via un vecteur ou non) entre
individus contemporains apparentés ou non, (iii) environnementale. Ce dernier terme qui
correspond en tous points à une transmission horizontale est restreint aux cas où l’association
symbiotique est necessaire à l’hôte. Dans ce cadre elle définit l’infection d’une nouvelle
génération hôte par un stock de symbiotes présent dans l’environnement et provenant de la
génération parentale (Le Pennec et al., 1988).
La nature des associations symbiotiques est souvent liée au mode de transmission du
symbiote. Communément, la transmission verticale est caractéristique des associations de
type mutualiste. En effet, le devenir du symbiote dépend du devenir de son hôte et
inversement. A l’inverse, la transmission horizontale est le mode de transmission typique des
organismes pathogènes.
1 Ce concept correspond à l’expression du génotype d’un partenaire dans le phénotype de l’autre, ou comment les gènes d’un symbiote induisent des modifications morphologiques ou comportementales chez un hôte.

INTRODUCTION
24
Tableau A. Différents types de relations entre espèces en fonction de l’effet de l’interaction sur le succès
reproducteur des partenaires.
Effet sur la fitness du partenaire 1
Positif Neutre Négatif
Eff
et s
ur
la f
itn
ess
du
part
enair
e 2
Posi
tif
Mutualisme
Commensalismea
Phorésieb
Inquilinismeb
Parasitisme
Prédation N
eutr
e
Commensalisme
Phorésie
Inquilinisme
Neutralisme Amensalismec
Nég
ati
f Parasitisme
Prédation Amensalisme
Compétition
Conflit
En rouge sont notées les relations correspondant à des symbioses répondant à la définition de De BARY. a Relation où un seul partenaire tire bénéfice de la relation. L’autre partenaire n’a ni avantage ni coût à s’associer. b Ces deux relations sont des types de commensalismes particuliers. Pour la phorésie, l’un des partenaires est
transporté par l’autre (e.g. le poisson pilote [Remora sp.] des raies). Dans le cas de l’inquilinisme, un des
partenaires (généralement le plus gros) sert d’abri à l’autre (e.g. les poissons ou crustacés s’abritant dans la cavité palléale des mollusques) c Cette association correspond à l’inhibition de la croissance d’une espèce par une autre espèce (e.g. Le
champignon Penicillium sp. et sa capacité antibiotique).
Il existe toutefois des exceptions à cette règle (i.e. transmission verticale = mutualisme
et transmission horizontale = parasitisme). Tel est le cas des bactéries dites manipulatrices de
la reproduction. Ces micro-organismes ne sont pas mutualistes mais sont transmis
verticalement par voie maternelle (e.g. Cardinium, Rickettsia ou Wolbachia associées aux
arthropodes terrestres). Dans ces modèles, les symbiotes sont intégrés aux gamètes de l’hôte
(cytoplasme de l’œuf) et ont développé des mécanismes garantissant leur transmission. Cette
dernière se fait soit au détriment du sexe non transmetteur : (i) Male-killing (Hurst and
Jiggins, 2000), (ii) Féminisation (Giorgini et al., 2009; Rigaud et al., 1999) et (iii)
Parthénogénèse thélytoque (Groot and Breeuwer, 2006; Stouthamer et al., 1993), soit au
détriment de la descendance des femelles non infectées : Incompatibilité Cytoplasmique
(Gotoh et al., 2007; Hoffmann et al., 1986).

INTRODUCTION
25
Les parasites transmis par voie materno-fœtale (assimilée à la transmission verticale)
constituent une autre exception à la règle. C’est le cas par exemple du protozoaire Neospora
caninum (Cole et al., 1995). La littérature décrit également des associations où à l’évidence la
transmission du symbiote n’est pas verticale, mais où la nature de l’association symbiotique
est considérée comme mutualiste. C’est le cas de l’association Rhizobium/Légumineuses
(Beringer et al., 1979), des coraux associés aux dinoflagellés (Dubinsky et al., 1990) ou de
certaines associations (celles sans transmission verticale du symbiote) entre invertébrés
marins et bactéries chimioautotrophes. C’est d’ailleurs à ces derniers cas que le terme de
transmission environnementale a été appliqué en lieu et place de celui de transmission
horizontale.
Cependant, ces associations constituent-elles réellement une exception (i.e.
mutualisme à transmission environnementale ou horizontale) à la règle ? On peut répondre
oui, si elles sont effectivement mutualistes et qu’il y a réellement transmission du symbiote.
Mais la réponse sera non si en réalité elles correspondent à un autre paradigme, celui
d’associations non mutualistes et/ou sans transmission du symbiote. C’est à cette question que
je me suis intéressé à travers l’étude des associations entre « invertébrés » marins et bactéries
chimioautotrophes via le modèle Lucinidae/bactéries sulfo-oxydantes. D’autre part j’ai axé
mes travaux de recherche sur la diversité des bactéries sulfo-oxydantes associées aux
Lucinidae et sur l’étude de la spécificité d’association entre ces deux partenaires.

INTRODUCTION
26
LES ASSOCIATIONS CHIMIOAUTOTROPHES
e travail présenté dans ce mémoire s’inscrit dans le champ d’étude des associations
symbiotiques entre « invertébrés » marins et bactéries chimioautotrophes (i.e.
bactéries capables de fixer le carbone à partir de composés chimiques inorganiques).
Ce type d’association a été découvert au niveau des sources hydrothermales profondes
(Felbeck et al., 1981) puis a été mis en évidence au niveau des sources de fluides froids
situées sur les marges continentales (Juniper and Sibuet, 1987). Ces deux environnements
profonds se caractérisent par la présence de fortes concentrations en composés réduits tels que
le méthane ou les sulfures. Ces associations ont également été retrouvées au sein de divers
autres habitats caractérisés par la présence de composés réduits servant de source d’énergie
primaire. Ainsi les carcasses de baleines (Deming et al., 1997) ou les bois coulés (Duperron et
al., 2008b; Gros and Gaill, 2007; Gros et al., 2007) sont colonisés pas des organismes
chimiosymbiotiques exploitant les produits issus de la décomposition du substrat (e.g. bois,
lipides, etc…) par des micro-organismes. Enfin on retrouve aussi de telles associations dans
des environnements plus accessibles où les concentrations en composés réduits sont
relativement faibles tels que les herbiers à phanérogames en eau peu profondes (Berg and
Alatalo, 1984) ou les boues de mangroves (Schweimanns and Felbeck, 1985).
DIVERSITE DES ASSOCIATIONS CHIMIOAUTOTROPHES
Diversité des hôtes
Ces associations présentent une grande diversité d’hôtes. Elles ont été décrites dans de
nombreux taxons allant des mollusques aux nématodes en passant par les arthropodes, les
bryozoaires ou les annélides (pour revue, voir Dubilier et al., 2008). Ces associations peuvent
être en ecto-symbiose (ou épibiose) comme décrit chez de la crevette Rimicaris exoculata
(Polz and Cavanaugh, 1995; Zbinden et al., 2008) ou en endo-symbiose. Ce dernier type de
symbiose peut être extracellulaire, comme chez les bryozoaires du genre Watersipora
(Anderson and Haygood, 2007; Zimmer and Woollacott, 1983), les nématodes du genre
Astronema où les symbiotes sont situés dans la lumière du tube digestif (Musat et al., 2007;
Ott et al., 2004), ou les annélides des genres Inanidrilus (Blazejak et al., 2006) et Olavius
L

INTRODUCTION
27
(Woyke et al., 2006) dans lesquels les symbiotes sont localisés sous la cuticule. L’endo-
symbiose peut également être intracellulaire comme chez la plupart des Mollusques bivalves
où les symbiotes sont localisés dans des cellules spécialisées des branchies : les bactériocytes
(pour revue voir Dubilier et al., 2008; Fisher, 1990).
Diversité des symbiotes
Les symbiotes associés dans ce type de relations présentent également une certaine
diversité taxonomique. Cette dernière a essentiellement été mise en évidence grâce au
séquençage du gène de l’ARNr 16S. Sur la base des séquences de ce marqueur, les symbiotes
sulfo-oxydants par exemple formeraient au moins 9 grandes lignées distinctes (pour revue
voir Dubilier et al., 2008). La majorité des symbiotes appartient à la classe des gamma-
protéobactéries. Néanmoins, on retrouve également des alpha-protéobactéries chez les
bryozoaires du genre Watersipora (Anderson and Haygood, 2007), des delta-protéobactéries
chez les vers oligochètes du genre Olavius (Woyke et al., 2006), des epsilon-protéobactéries
chez la crevette Rimicaris exoculata (Zbinden and Cambon-Bonavita, 2003), et des
Cytophaga-Flaviobacterium-Bacterioides chez une moule du genre Idas (Duperron et al.,
2008a).
Diversité métabolique
Outre cette diversité taxonomique on trouve également chez ces symbiotes de
multiples métabolismes. Si la majorité des symbiotes possède un métabolisme de type sulfo-
oxydant, il existe également des méthanotrophes (Cavanaugh et al., 1992; Childress et al.,
1986; Duperron et al., 2005), des sulfato-réducteurs (Woyke et al., 2006), et des ferro-
oxydants (Zbinden and Cambon-Bonavita, 2003; Zbinden et al., 2004). Pour certaines
associations les métabolismes bactériens ont été inférés grâce au positionnement
phylogénétique des symbiotes (basé sur l’ARNr 16S). Par ailleurs, des analyses d’isotopes,
génomiques et/ou protéomiques des enzymes intervenant dans les différentes voies
métaboliques impliquées ont souvent confirmé les résultats obtenus à l’aide de l’ARNr 16S
(pour revue, voir Dubilier et al., 2008).

INTRODUCTION
28
Diversité d’ « infection »
Au sein de ces associations il existe aussi une diversité de « patrons d’infection ». Si
dans la majorité des cas, un seul type symbiotique (aussi bien au niveau taxonomique que
métabolique) est retrouvé, des cas de multi-infections ont été décrits. Ainsi, deux
morphotypes bactériens différents au sein des branchies de la moule Bathymodiolus
thermophilus ont été observés (Fisher et al., 1993). Ces derniers ont été caractérisés par la
détection d’activités enzymatiques confirmant ainsi la présence d’un symbiote thioautotrophe
(activité ATP sulfurylase) et d’un méthanotrophe (activité méthanol deshydrogénase) (Fisher
et al., 1993). Ceci a constitué la première observation de bi-infection au sein d’un même
individu. Plus tard, la coexistence de six symbiotes différents a été mise en évidence au sein
d’une moule du genre Idas (Duperron et al., 2008a). Ces patrons de multi-infection semblent
être courant chez les moules mais sont également retrouvés au sein d’autres types d’hôtes.
Ainsi Woyke et al. (2006) ont observé chez le ver Olavius algarvensis quatre symbiotes
différents. Deux d’entre eux sont des delta-protéobactéries sulfato-réductrices et les deux
autres des gamma-protéobactéries. Pour ces dernières le métabolisme sulfo-oxydant n’a pu
être validé que dans un seul cas. Cette association présente d’autant plus d’intérêt qu’il s’agit
de la première observation de colonisation au sein du même individu de symbiotes oxydant et
réduisant le soufre. De fait, cette symbiose présente la particularité de pouvoir fonctionner en
circuit fermé sans apport extérieur.
NATURE DES ASSOCIATIONS CHIMIOAUTOTROPHES
Au sein des associations entre « invertébrés » marins et bactéries chimioautotrophes,
la nature de l’association a été définie indépendamment du mode de transmission du
symbiote. Ces dernières sont décrites comme bénéfiques pour les deux partenaires (Stewart et
al., 2005). Du côté de l’hôte, considérant le rôle nutritionnel et adaptatif de la bactérie,
héberger un symbiote apparait essentiel (Berg and Alatalo, 1984; Fisher, 1990). En effet, la
fixation du carbone couvre une grande partie des besoins nutritionnels de l’hôte (Stewart et
al., 2005). De plus, la symbiose confère à ces derniers la capacité de fixer du carbone
indépendamment de la photosynthèse, capacité normalement absente chez les eucaryotes non
photosynthétiques. Enfin, la bactérie permettrait à l’hôte de coloniser des milieux plus
instables (e.g. sources hydrothermales, suintement froids etc…) où sont présents des

INTRODUCTION
29
dégagements de composés réduits tels que les sulfides (H2S), le soufre étant toxique pour la
cytochrome oxydase de la chaine respiratoire (Arndt et al., 2001). Du côté du symbiote,
l’intracellularité conférerait à la bactérie un environnement plus stable que les sédiments et
permettrait un meilleur accès aux substrats nécessaires à son métabolisme (e.g. CO2,
sulfides). En effet, la filtration de l’eau environnante par les branchies concentrerait ces
composés aux abords des symbiotes.
Si pour les modèles où la transmission du symbiote est verticale (e.g. Vesicomyidae,
Solemyidae) (Cary, 1994; Cary and Giovannoni, 1993; Krueger et al., 1996), l’hypothèse
mutualiste ne fait aucun doute, qu’en est-il pour les Lucinidae et les modèles sans
transmission verticale du symbiote? Ces derniers constituent-ils une exception à la règle (i.e.
mutualisme sans transmission verticale) ou l’hypothèse du mutualisme est-elle erronée? Dans
cette partie du travail je me suis intéressé au mode de transmission du symbiote chez les
Lucinidae et aux conséquences de ce dernier sur la nature de l’association afin de confirmer
ou de rejeter l’hypothèse mutualiste.

INTRODUCTION
30
LES LUCINIDAE
es Lucinidae sont une famille de bivalves appartenant à la super-famille des
Lucinoidea et à l’ordre des Veneroida (Fig. 1). Ils constituent la famille la plus
disparate et la plus riche en termes d’espèces au sein des mollusques
chimiosymbiotiques (Fig. 2 Taylor and Glover, 2005).
Figure 1. Représentation schématique de la taxonomie des bivalves chimiosymbiotiques.
L

INTRODUCTION
31
Figure 2 Diversité de formes chez les Lucinidae.
A, Plicolucina flabellata ; B, Lamellolucina trisulcata ; C, Codakia tigerina ; D, Anodontia philippiana ; E,
Miltha childrenae ; F, Eomiltha voorhoevi ; G, Austriella corrugata ; H, Ctena bella ; I, Rasta lamyi ; J,
Pompholigina gibba ; K, Myrtea spinifera.

INTRODUCTION
32
(Photo, O.Gros)
HABITATS.
Les Lucinidae s’étendent sur une large aire géographique (60°N-55°S) et sont retrouvés aussi
bien dans les zones tropicales que tempérées. Même si la majorité d’entre eux colonisent des
milieux relativement peu profonds certains
spécimens ont été décrits aux alentours des
sources de fluides froids (Duperron et al., 2007).
Ces bivalves colonisent donc une grande variété
d’habitats allant de la zone littorale (0-200m) à la
zone bathyale (200-2000m). Ils sont ainsi
retrouvés au sein des herbiers à phanérogames
(Fig. 3) (genres Zostera ou Thalassia), des boues
de mangroves de palétuviers (Fig. 4), des
sédiments réduits et d’une façon plus générale au
sein des environnements présentant des
dégagements de soufre réduit plus ou moins
importants, conséquences de la dégradation de
matière organique. Ils sont enfouis dans les
sédiments aux alentours de poches de soufre, dans des sortes de terriers communicants avec
l’extérieur (i.e. colonne d’eau) via deux conduits
permettant la circulation de l’eau (Fig. 5). Le conduit
inhalant permet l’entrée d’oxygène et de matière
organique. Il est creusé et maintenu par le pied, les
lucines n’ayant pas de siphon inhalant. Ces dernières
ne possèdent en effet qu’un siphon exhalant qui est placé
dans le conduit exhalant et permet ainsi l’évacuation des
déchets. D’autre part, des conduits plus petits qui sont
supposés avoir été creusés par l’animal à la recherche des
poches de soufre ont également été observés (Cary
et al., 1989).
(Photo, O.Gros)
P : Pied, B : Branchie, S.e. : Siphon
exhalant. (Tiré de Fisher, 1990 ;
dessiné par R.E. Kochevar)
Figure 5. Herbier à Thalassia testudinum.
Figure 4. Mangrove de palétuviers, Rhizophora
mangle.
Figure 3. Habitat typique des
Lucinidae.

INTRODUCTION
33
MORPHOLOGIE
Il a été suggéré que la symbiose constituerait le facteur le plus important dans
l’évolution de la famille des Lucinidae (Reid and Brand, 1986). Ces derniers se caractérisent
par de larges branchies hypertrophiées, l’absence de siphon inhalant, et un intestin et des
palpes labiaux réduits, toutes ces caractéristiques étant présumées dues à l’adaptation de ces
bivalves au mode de vie symbiotique.
Les branchies (Fig. 6) sont composées de deux demi-branches accolées constituées de
filaments branchiaux (Annexe 1). Ces filaments sont composés chez les adultes de trois
régions distinctes (Annexe 1): une zone ciliée courte,
une zone intermédiaire et une zone latérale (Gros et
al., 1999; Gros et al., 1997). Cette dernière
représente la majeure partie du filament branchial et
est composée de quatre types cellulaires : les
mucocytes, les cellules à grains, les cellules
intercalaires et les bactériocytes. Ces bactériocytes
constituent le type cellulaire majoritaire et abritent
les symbiotes. On trouve ces derniers dans des
vacuoles, avec généralement une seule bactérie par
vacuole. Cependant plusieurs bactéries dans une
même vacuole ont également été observées (Distel
and Felbeck, 1988).
SYMBIOSE.
Tous les Lucinidae analysés à ce jour hébergent au sein de leurs branchies des
bactéries symbiotiques. Ces dernières appartiennent à la classe des gamma-protéobactéries
(sur la base du séquençage de l’ARNr 16S) et sont sulfo-oxydantes. Le mode de transmission
du symbiote est supposé environnemental car : (i) l’acquisition du symbiote se fait au stade
juvénile après métamorphose (Gros et al., 1998b). (ii) Aucun symbiote n’a pu être détecté que
ce soit par PCR ou par microscopie électronique dans les gamètes, œufs, embryons, ou larves
de différentes lucines (Gros et al., 1996; Gros et al., 1998a). (iii) La forme libre du symbiote a
Br. Branchies, M.a. Muscles adducteurs, P.
pied, M.v. Masse viscérale. (Lucina
pectinata, Photo, O. Gros)
Figure 6. Anatomie générale des Lucinidae.

INTRODUCTION
34
été mise en évidence dans les sédiments des herbiers à Thalassia testudinum (Gros et al.,
2003b).
Cette association est considérée comme mutualiste comme toutes les symbioses entre
« invertébrés » marins et bactéries chimiotrophes. En effet, la bactérie est présumée
responsable de la détoxification du milieu (soufre). Cependant, certains auteurs ont également
incombé ce rôle à l’hémoglobine conjuguée à la présence de « sulfur-oxidizing bodies »
(Arndt et al., 2001; Liberge et al., 2001). En effet, cette dernière a été décrite dans les
branchies de diverses lucines (Ball et al., 2009; Dando et al., 1985; Frenkiel et al., 1996;
Gavira et al., 2008; Kraus and Wittenberg, 1990). Cependant l’hémoglobine a également la
capacité de fixer l’O2 en plus du H2S (Kraus, 1993). De fait, selon Dando et al. (1985) cette
dernière pourrait également agir en tant que stockeur d’oxygène durant les phases
d’enfouissement du bivalve ou protéger les symbiotes de concentrations trop élevées en O2,
les bactéries sulfo-oxydantes étant souvent micro-aérophiles (Jorgensen, 1982). Il est donc
difficile de savoir qui des symbiotes ou des hôtes détoxifie le milieu. Outre le rôle présumé de
détoxification, les symbiotes supportent une grande partie des besoins nutritionnels de l’hôte.
En effet, le Lucinidae bénéficierait du métabolisme en profitant de la fixation du carbone via
le cycle de Calvin-Benson. Du côté du symbiote, les branchies permettraient un meilleur
accès aux substrats métaboliques (CO2, H2S).
Même si cette symbiose n’est pas physiologiquement obligatoire (il est possible
d’obtenir des juvéniles asymbiotiques et de débarrasser les adultes de leurs symbiotes) cette
association apporte au bivalve un avantage sélectif certain. En effet, Gros et al. (1998b) ont
montré que les spécimens symbiotiques avaient une croissance plus importante que les non
symbiotiques. D’autre part, la prévalence de 100% dans la nature de cette association est une
observation qui va également dans ce sens.
EVOLUTION
Dans cette famille de bivalves, la capacité à établir une association symbiotique
semble être un héritage ancestral. La symbiose aurait été acquise vers -250 Millions d’années
avant la divergence des différentes familles (Thyasiridae, Lucinidae, Ungulinidae et
Fimbriidae) appartenant à la superfamille des Lucinacea (Boss, 1970; Newell, 1969). Bien
qu’elle ait été perdue totalement chez les Ungulinidae et partiellement chez les Thyasiridae

INTRODUCTION
35
(Reid and Brand, 1986), cette association a été maintenue chez les Lucinidae attestant une fois
de plus de son importantce. Cependant, les questions de l’origine et de l’histoire évolutive des
couples symbiotiques actuels ainsi que les facteurs ayant permis le maintien d’une telle
association sans transmission verticale restent posées.


NATURE DE L’ASSOCIATION
111...
NNNAAATTTUUURRREEE DDDEEE
LLL’’’AAASSSSSSOOOCCCIIIAAATTTIIIOOONNN

NATURE DE L’ASSOCIATION

NATURE DE L’ASSOCIATION
« Le rêve de toute
cellule : devenir deux
cellules »
(F. Jacob)

NATURE DE L’ASSOCIATION

NATURE DE L’ASSOCIATION
41
NATURE DE L’ASSOCIATION
INTRODUCTION
’association Lucinidae/bactéries sulfo-oxydantes est classiquement considérée
comme étant de type mutualiste. Concernant le mode de transmission du symbiote,
Gros et al. (1996; 1998a; 2003b) ayant réussi l’infection au laboratoire de juvéniles
asymbiotiques à partir de sable et ayant détecté la forme libre du symbiote dans les herbiers,
ces auteurs ont proposé l’hypothèse de transmission environnementale du symbiote.
Pour que l’association symbiotique se perpétue de générations en générations, sous les
hypothèses du mutualisme et de la transmission environnementale il faut que le processus de
transmission s’opère selon le modèle présenté Figure 7. Quant à la validité de ce modèle elle
dépend de trois paramètres : (i) la présence d’une forme libre du symbiote dans
l’environnement, (ii) la multiplication du symbiote à l’intérieur des branchies de l’hôte et (iii)
le re-largage dans l’environnement du symbiote par les adultes.
Figure 7. Représentation schématique de la transmission environnementale.
L
Les symbiotes sont acquis par les juvéniles à partir de symbiotes re-largués par les adultes de la génération
contemporaine après multiplication de ces derniers dans les branchies. A.1 : Adultes de la génération parentale,
A.2 : Adultes de la génération suivante, J.a. : Juvéniles asymbiotiques, J. : Juvéniles, ellipses : bactéries
endosymbiotiques.
NATURE DE L’ASSOCIATION
42
La forme libre du symbiote a effectivement été mise en évidence dans les sédiments
par CARD-FISH sur filtre grâce à la sonde symco2A (Gros et al., 2003b) qui est spécifique de
l’ARNr 16S de l’espèce symbiotique associée à Codakia orbicularis (Gros et al., 1996). Ces
résultats corroborent donc l’hypothèse de transmission environnementale précédemment
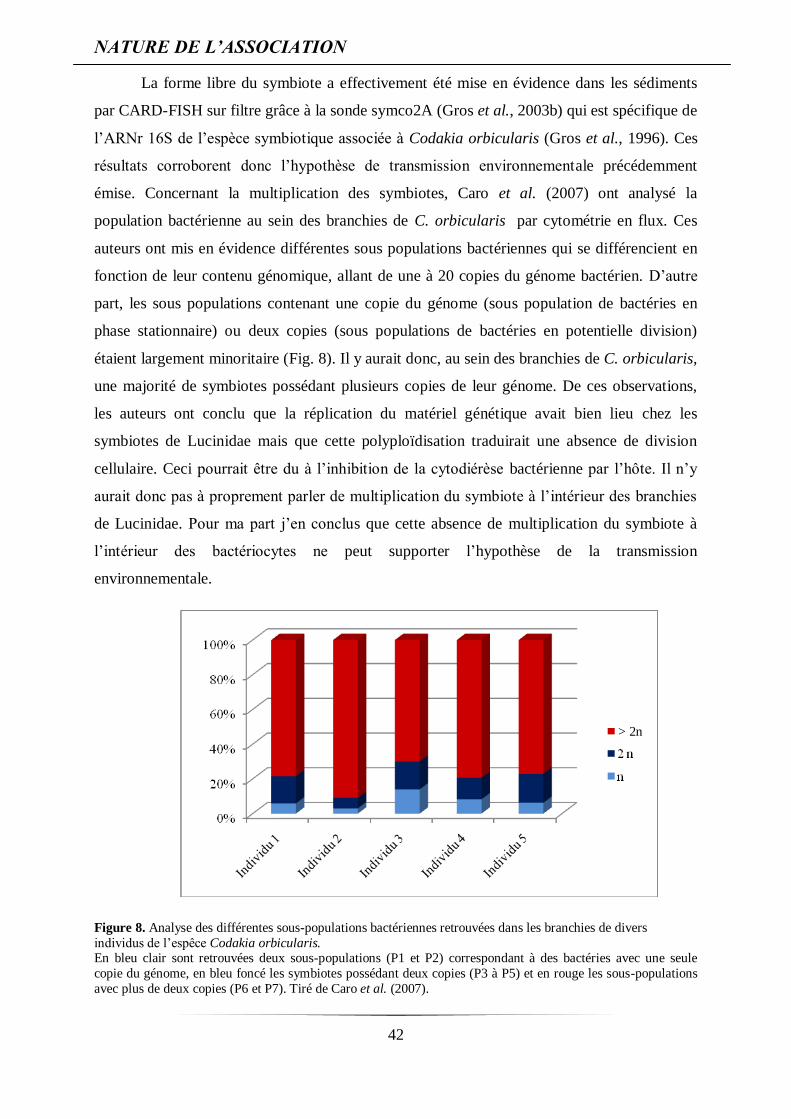
émise. Concernant la multiplication des symbiotes, Caro et al. (2007) ont analysé la
population bactérienne au sein des branchies de C. orbicularis par cytométrie en flux. Ces
auteurs ont mis en évidence différentes sous populations bactériennes qui se différencient en
fonction de leur contenu génomique, allant de une à 20 copies du génome bactérien. D’autre
part, les sous populations contenant une copie du génome (sous population de bactéries en
phase stationnaire) ou deux copies (sous populations de bactéries en potentielle division)
étaient largement minoritaire (Fig. 8). Il y aurait donc, au sein des branchies de C. orbicularis,
une majorité de symbiotes possédant plusieurs copies de leur génome. De ces observations,
les auteurs ont conclu que la réplication du matériel génétique avait bien lieu chez les
symbiotes de Lucinidae mais que cette polyploïdisation traduirait une absence de division
cellulaire. Ceci pourrait être du à l’inhibition de la cytodiérèse bactérienne par l’hôte. Il n’y
aurait donc pas à proprement parler de multiplication du symbiote à l’intérieur des branchies
de Lucinidae. Pour ma part j’en conclus que cette absence de multiplication du symbiote à
l’intérieur des bactériocytes ne peut supporter l’hypothèse de la transmission
environnementale.
Figure 8. Analyse des différentes sous-populations bactériennes retrouvées dans les branchies de divers
individus de l’espêce Codakia orbicularis. En bleu clair sont retrouvées deux sous-populations (P1 et P2) correspondant à des bactéries avec une seule
copie du génome, en bleu foncé les symbiotes possédant deux copies (P3 à P5) et en rouge les sous-populations
avec plus de deux copies (P6 et P7). Tiré de Caro et al. (2007).
> 2n

NATURE DE L’ASSOCIATION
43
Je me suis donc intéressé au troisième paramètre : le relarguage du symbiote dans
l’environnement par les adultes. Pour cela j’ai recherché, au laboratoire, à mettre en évidence
ce phénomène chez des individus de C. orbicularis et C. orbiculata en utilisant une méthode
de détection du symbiote par hybridation in situ.
DEMARCHE EXPERIMENTALE
SPECIMENS ET CONDITIONS D’ENTRETIEN
Dix individus de C. orbicularis (environ 40mm de long) et 22 individus de C.
orbiculata (environ 15mm) provenant de l’Îlet Cochon (Guadeloupe) ont été répartis au
laboratoire dans 500 ml d’eau de mer filtrée à 0,2 µm (eau de mer dépourvue de bactéries)
dans 10 cristallisoirs comme suit :
- Deux cristallisoirs contenant chacun un spécimen1 de C. orbicularis.
- Quatre cristallisoirs contenant chacun deux spécimens2 de C. orbicularis.
- Quatre cristallisoirs contenant chacun 5 ou 6 spécimens de C. orbiculata.
Afin de limiter l’éventuelle ré-acquisition de symbiotes, due à la filtration de l’eau par
le bivalve, les lucines ont été placées à mi-hauteur dans des cristallisoirs. Il n’y a eu ni ajout
de soufre, ni ajout de nourriture au cours de l’expérience. L’expérimentation a été conduite
durant un mois avec changement de l’eau tous les jours. Un contrôle positif (eau de mer
filtrée à 0,2 µm contenant un extrait de branchie de C. orbicularis) a également été réalisé afin
de s’assurer que les symbiotes pouvaient être détectés par notre technique.
DETECTION DU RELARGUAGE DES SYMBIOTES
Le seuil de détection de notre méthode a d’abord été évalué. Pour ce faire, un extrait
de branchie a été purifié sur percoll selon Caro et al. (2007) et quantifié sur cellule de
Malassez. Cet inoculum a ensuite été utilisé afin de constituer différents échantillons
1 Cette condition a été réalisée afin de limiter l’éventuelle acquisition de symbiotes relargués par un individu
hôte voisin. 2 Cette condition a été réalisée pour augmenter le relarguage s’il existe et ainsi maximiser les chances de le
détecter

NATURE DE L’ASSOCIATION
44
contenant 0, 10, 102, 10
3, 10
4, 10
5, 10
6, 10
7 bactéries/ml. Pour le relarguage, des échantillons
d’eau provenant des différents cristallisoirs ont été collectés à T0, toutes les quatre heures le
premier jour, tous les jours les cinq suivants puis pour finir tous les cinq jours. Ceci a produit
un total de 140 échantillons. Les deux types d’échantillon (seuil de détection et recherche de
symbiotes relargués) ont été fixés 4h en PFA 4% à 4°C et filtrés sur filtres GTTP (Millipore,
France). Des sections de filtre (environ ¼ de filtre) ont directement été analysés ou stockés à -
80°C avant analyse.
SUIVI DE LA POPULATION BACTERIENNE A L’INTERIEUR DES BRANCHIES PAR CARD-FISH ET MET
Afin de suivre l’évolution de la population symbiotique au sein des branchies, des
individus ont été sacrifiés à T0, T0+10 jours, T0+20 jours et T0+30 jours. Des sections de
branchies de ces individus ont été réalisées et analysées en CARD-FISH. Pour la microscopie
électronique des individus ont été sacrifiés à T0 et T0+30 jours. Après dissection, les
échantillons de branchies ont été fixés deux heures en glutaraldheyde 2,5%/Tampon
cacodylate (0,1M) à pH 7,2 (conservé iso-osmotique à l’eau de mer par addition de CaCl2)
puis lavées 3 fois dans le même tampon iso-osmotique. Les morceaux de branchies ont
ensuite été fixés plusieurs heures à température ambiante dans du tetroxide d’osmium 1%
(même tampon) et rincées en eau distillée avant post-fixation (1h en uranyle acétate aqueux
2%), déshydratation (bains croissants successifs d’éthanol et d’oxyde de propylène) puis
incluses dans un mélange Epon-Araldite. Des sections de 60nm d’épaisseur ont été réalisées,
colorées à l’uranyle d’acétate et au lead citrate avant observation par un microscope
électronique à transmission Leo912.
CARD-FISH
Les morceaux de branchies ou de filtres ont été hybridés avec les sondes EUB338, qui
s’hybride avec l’ARNr 16S de la majorité des eubactéries (Amann et al., 1990; Daims et al.,
1999) et la sonde Symco2A. L’inclusion a été réalisée selon Dubilier et al. (1995). Pour les
branchies, après inclusion, des sections de 4µm ont été réalisées et collées sur lames
SuperFrost® (Menzel-Glazer®). Les sections ont ensuite été déparaffinées dans du Toluène
100% et réhydratées (éthanol 100%, 95%, 70% suivi d’eau distillée). Les lames ont ensuite
été lavées en HCl 0,2M (12 min à RT), Tris/HCl 20mM (10 min), protéinase K (0,5 µg.mL-1
,
5 min à 37°C) puis dans du Tris/HCl 20mM (10 min).

NATURE DE L’ASSOCIATION
45
Le CARD-FISH a ensuite été réalisé selon Pernthaler et al. (2002) aussi bien pour les
sections histologiques que pour les filtres jusqu’à l’étape du tampon de lavage. Les
échantillons sont ensuite rincés deux fois en PBS 1X (15 min à RT) puis incubés dans un
tampon d’amplification (1/10 [w/v] de dextrane sulfate, PBS 1X, NaCl 2M et 1/100 [v/v] de
blocking reagent 10%), de l’eau oxygénée à 0,5% (diluée dans du PBS 1X) et 2 µL de
tyramide marquée en fluorescence pendant 15 min à 37°C dans le noir. Les sections ou filtres
sont ensuite lavés en PBS 1X, eau distillée et deux fois en éthanol absolu. Les échantillons
sont séchés, contre-colorés au DAPI (4’,6-diamino-2-phenylindole) inclus en milieu DAKO et
observés sous microscope à fluorescence epi80i (NIKON, France).
RESULTATS
RECHERCHE DE SYMBIOTES RELARGUES
Afin de s’assurer de la sensibilité de la technique utilisée, nous avons défini le nombre
minimal de bactéries nécessaires pour observer un signal dans de l’eau de mer filtrée à 0,2µm
contenant une fraction purifiée de symbiotes de C. orbicularis. Ceci a été réalisé sur des
échantillons contenant de 0 à 107
bactéries/ml. Les deux sondes utilisées, EUB338
(généraliste des eubactéries [Fig. 9c]) et Symco2A (spécifique des symbiotes de Lucinidae
[Fig. 9e et 9f]) se sont révélées suffisamment efficaces pour détecter au moins 10
bactéries/ml (densité minimale testée), le contrôle négatif (eau de mer filtrée à 0,2µm)
demeurant négatif pour les deux sondes. Concernant la détection du symbiote, sur les 140
observations réalisées (un mois d’expérimentation) aucun signal positif n’est apparu (Fig. 9g)
avec la sonde Symco2A que ce soit pour les échantillons précoces (toutes les 4h sans
changement d’eau ; de T0 à T0+20h) ou tardifs (de T0+1 à T0+30). Dans tous les cas le
contrôle positif interne d’hybridation (EUB338) a donné un signal positif (Fig. 9d). La contre-
coloration au DAPI a confirmé que les signaux d’hybridations positifs correspondaient à des
bactéries et non à des particules auto-fluorescentes du sédiment (Fig. 9a et 9b)

NATURE DE L’ASSOCIATION
46
DAPI EUB338 Symco2A
Contr
ôle
posi
tif
T0+
1 j
our
3,5.108 bactéries/mL 10 bactéries/mL
a c e f
b d g
Figure 9. Recherche de symbiotes re-largués. La rangée supérieure correspond au contrôle positif de détection ainsi qu’à l’évaluation du seuil de détection du
CARD-FISH. La rangée inférieure correspond à l’analyse de l’eau après 1 jour. (a et b) : contrôle DAPI de la
coloration des acides nucléiques. (c et d) hybridation avec EUB338 spécifique de la majorité des eubactéries. (e, f et g) hybridation avec Symco2A sonde spécifique des symbiotes de C. orbicularis et C. orbiculata sur : une
fraction purifiée de symbiotes de C. orbicularis non diluée (e) ou diluée à une densité de 10 bactéries/mL (f).
symbiotes : flèches rouges
EVOLUTION DE LA POPULATION BACTERIENNE AU SEIN DES BRANCHIES
A 0, 10, 20 et 30 jours d’expérimentation, des individus ont été sacrifiés afin de
pouvoir suivre la population bactérienne intracellulaire. Ces échantillons ont été analysés en
CARD-FISH (hybridation avec Symco2A et EUB338) et par microscopie électronique.
CARD-FISH : Dans tous les cas, les sondes s’hybrident au niveau de la zone latérale
où sont distribués les symbiotes. Aucune différence notable n’est apparue entre les sections de
branchies à T0 (Fig. 10A), T0+10 jours et T0+20 jours. A l’inverse, sur les sections provenant
d’individus sacrifiés à la fin de l’expérience (T0+30 jours) un signal d’hybridation plus faible
est observé (Fig. 10B).
TEM : Après 1 mois de jeûne on peut observer des modifications notables du filament
branchial (Fig. 10C) par rapport à l’observation d’un adulte n’ayant pas subi de jeûne. Ainsi,
on observe une diminution du nombre de bactéries et l’apparition de nombreux lysosomes
secondaires (Fig. 10D), indiquant probablement une digestion intracellulaire des symbiotes.
g

NATURE DE L’ASSOCIATION
47
Figure 10. Figure 10. Suivi de la population symbiotique à l’intérieur des branchies.
A et B : CARD-FISH réalisé à l’aide de la sonde Symco2A. A : Section de branchie de C. orbicularis. B : Section de branchie de C. orbicularis après 30 jours de jeûne. ZC, Zone Ciliée ; ZL, Zone Latérale ; échelles =20
µm.
C et D : Modifications ultra-structurales dans les branchies de C. orbicularis. C : Les bactériocytes (BC), type
cellulaire majoritaire, possèdent un noyau basal (N) et un pôle apical en contact avec l’eau de mer de la cavité
palléale (flèches blanche). Des bactéries (b) internalisées dans des vacuoles parsèment le cytoplasme. D : Après
30 jours de jeûne des bactériocytes (BC) résiduels sont observés avec peu de bactéries (b) et de larges lysosomes
secondaires (Ly). Les plus gros symbiotes ont disparu et ne demeurent que les plus petits à la périphérie de la
cellule. LS, Lacune Sanguine ; N, Noyau ; échelles = 0,5µm. (MET, Images O. Gros)
DISCUSSION
Le but de ce chapitre était de déterminer la nature exacte de la relation
Lucinidae/bactéries chimioautotrophes via l’étude du mode de transmission du symbiote. Des
études antérieures ayant montré qu’il n’y avait pas de symbiotes présents dans les gamètes,
œufs, embryons ou larves de diverses lucines ont permis de rejeter l’hypothèse de la
transmission verticale (Gros et al., 1996; Gros et al., 1998a). De plus, ces auteurs ont
également réalisé au laboratoire l’infection de juvéniles asymbiotiques de Lucinidae à partir
A ZL
ZC T0
C
b
b b
BC
BC
N
D
b
b CI
CI N
Ly
Ly
LS
B
ZC
ZL
A
ZL
ZC
T0 T0+30 jours
N
A

NATURE DE L’ASSOCIATION
48
de sable provenant des herbiers à Thalassia testudinum (Gros et al., 1998b) où la forme libre
du symbiote a été détectée (Gros et al., 2003b). De fait, ces observations ont conduit ces
auteurs à proposer l’hypothèse de la transmission environnementale i.e. infection de la
nouvelle génération à partir d’un stock de symbiotes présents dans l’environnement (Gros et
al., 1996; Gros et al., 1998a; Gros et al., 1999; Gros et al., 2003b). La transmission
environnementale étant un passage d’hôte (i.e. adultes) à hôte (i.e. juvéniles) via
l’environnement, cette dernière nécessite la présence de la forme libre dans les sédiments (qui
a été détectée), la multiplication du symbiote dans les bactériocytes (qui ne semble pas avoir
lieu) et le relarguage de ce dernier par les adultes. C’est à ce troisième paramètre non encore
étudié que je me suis intéressé. Grâce à des techniques d’hybridation in situ, j’ai recherché au
laboratoire à mettre en évidence d’éventuels symbiotes relargués.
Après 1 mois d’expérimentation, aucun signe de symbiotes relargués n’a pu être
observé. Quelles hypothèses seraient à même d’expliquer ce résultat ? (i) Les conditions
expérimentales n’étaient pas optimales pour permettre le relarguage des symbiotes. En effet,
l’expérimentation hors du milieu naturel aurait mis les différents individus en conditions de
stress induisant un comportement anormal de ceux-ci. Cependant, durant toute l’expérience il
n’y a eu aucune mortalité ni signe évident de stress due aux conditions d’entretien. (ii) La
période à laquelle a été réalisée l’expérimentation (durant le mois d’avril) ne serait pas celle
du relarguage dans la nature. En effet, la fin du développement larvaire et la métamorphose
ont lieu durant une période bien définie chez les Lucinidae (Juillet-Septembre). Ainsi sous
l’hypothèse de transmission environnementale, si le relarguage a lieu ce dernier pourrait être
restreint à cette période afin de maximiser l’infection de la nouvelle génération. L’absence de
relarguage observée pourrait simplement être due à l’absence de ce phénomène en avril. Cette
hypothèse semble peu probable car il est possible d’infecter des juvéniles durant toute l’année
et quelque soit l’âge du juvénile (O. Gros, communication personnelle). De fait, le relarguage
des symbiotes s’il a lieu ne serait pas restreint à une période particulière. (iii) Le CARD-
FISH n’est pas une technique suffisamment efficace pour détecter le relarguage. Néanmoins
le suivi de la population symbiotique (voir ci-après) ne semble pas favoriser cette hypothèse.
(iv) Le relarguage existe mais ne se produit qu’à la mort de l’individu. Cependant, seuls les
micro-organismes parasites possèdent des stratégies de transmission basées sur la mort de leur
hôte (Ewald, 1987). Or les symbiotes de Lucinidae comme les symbiotes des associations
chimioautotrophes ne sont pas considérés comme délétères (Stewart et al., 2005). Enfin (v),
ce phénomène n’a pas lieu chez les Lucinidae et par conséquent, l’hypothèse de transmission

NATURE DE L’ASSOCIATION
49
environnementale du symbiote ne peut être retenue. C’est cette dernière hypothèse que je
privilégie.
SUIVI DE LA POPULATION SYMBIOTIQUE
Le suivi de la population symbiotique à l’intérieur des branchies a révélé une
diminution du signal d’hybridation après 30 jours de jeûne. Ceci pourrait s’expliquer par un
relarguage non détecté par le CARD-FISH sur filtre. Cependant cette diminution de signal est
apparue trop importante pour correspondre à un relarguage que nous n’aurions pas été
capables de visualiser. En effet, en considérant que les branchies de bivalves hébergent
environ 3.109 bactéries et que la population bactérienne symbiotique diminue de 30% par
mois en conditions de jeûne (Caro et al., 2009), il y aurait donc eu relarguage d’environ 3.107
bactéries par bivalve par jour. Considérant notre limite de détection de 10 bactéries par ml
(i.e. signal observé à partir de 5000 bactéries par cristallisoir) ce relarguage n’aurait pu passer
inaperçu. De fait, la diminution de la population symbiotique ne saurait être due au
relarguage. Cette observation confirme donc l’inexistence de ce dernier.
Il semble que des hypothèses alternatives peuvent néanmoins expliquer la diminution
du nombre de bactéries au sein des branchies de Lucinidae. En effet, une diminution de
l’activité métabolique due aux conditions expérimentales ou la dégradation lysosomale des
symbiotes pourraient être responsables de l’évolution de la population symbiotique. La
seconde hypothèse semble à privilégier. En effet, les observations en microscopie
électronique montrent une diminution du nombre de symbiotes à l’intérieur des bactériocytes
et l’apparition de larges lysosomes secondaires (Fig. 10D) similaires à ceux décrits dans la
littérature (Frenkiel et al., 1996; Frenkiel and Mouëza, 1995; Liberge et al., 2001).
LES LUCINIDAE, EXCEPTION A LA REGLE ?
Même si la transmission environnementale n’est pas complètement rejetée par nos
expériences de relarguage en laboratoire, peu de preuves supportent cette dernière. En effet,
l’absence de transmission verticale, l’infection à partir de sable, la présence de la forme libre
du symbiote dans les sédiments, l’absence de multiplication du symbiote dans les branchies et
l’absence de re-larguage ne concourent pas globalement à valider l’hypothèse d’un passage du
symbiote d’hôte à hôte et par conséquent l’usage même du terme « transmission ». En
conséquence, l’absence de multiplication du symbiote à l’intérieur des branchies des

NATURE DE L’ASSOCIATION
50
Lucinidae (Caro et al., 2007) et de relarguage après internalisation (comme l’indique mes
résultats) me conduisent à considérer que les juvéniles de Lucinidae acquièrent leur symbiotes
uniquement à partir d’un stock de symbiotes qui ne se multiplient que dans l’environnement.
Ceci m’a donc permis de rejeter l’hypothèse de transmission environnementale du symbiote
au profit d’un strict recrutement environnemental chez les Lucinidae (Fig.11).
Dès lors, quelles peuvent être les conséquences de cette nouvelle hypothèse sur la
nature de la relation entre les Lucinidae et leurs symbiotes sulfo-oxydants ? Peut-on toujours
considérer cette association comme étant de type mutualiste ? Pour moi, non. En effet, sous
cette hypothèse de recrutement environnemental l’hôte bénéficie toujours de l’association tel
que décrit dans l’introduction (pages 26-27). Par contre, une fois internalisée, la bactérie ne
pouvant ni se multiplier ni être relarguée, l’entrée en symbiose constituerait pour cette
dernière une impasse évolutive, le succès reproducteur d’une bactérie endocytée étant alors
nul.
CONCLUSION
Dans la relation entre nature de la symbiose et mode de transmission du symbiote, il
existe face à la règle générale, des exceptions (i.e. symbiotes non mutualistes à transmission
verticale d’une part et symbiotes « mutualistes » non transmis verticalement d’autre part).
Figure 11. Représentation schématique du recrutement environnemental.
Les symbiotes sont acquis par les juvéniles à partir d’un stock de symbiotes qui se multiplient dans
l’environnement A.1 : Adultes de la génération parentale, A.2 : Adultes de la génération suivante, J.a. :
Juvéniles asymbiotiques, J. : Juvéniles, ellipses pleines : bactéries endosymbiotiques, ellipses vides : formes
libre du symbiote.

NATURE DE L’ASSOCIATION
51
C’est à cette seconde exception que semblait correspondre le cas des associations entre
Lucinidae et bactéries sulfo-oxydantes. Mes conclusions me conduisent à considérer qu’il ne
s’agit en fait pas d’une exception à la règle puisqu’il ne s’agirait pas d’une symbiose de type
mutualiste. Ceci constituerait une association symbiotique tout a fait originale puisque que la
persistance de l’association symbiotique d’une génération à l’autre se ferait sans transmission
du symbiote.


DIVERSITE BACTERIENNE
222...
DDDIIIVVVEEERRRSSSIIITTTEEE
BBBAAACCCTTTEEERRRIIIEEENNNNNNEEE
EEETTT
SSSPPPEEECCCIIIFFFIIICCCIIITTTEEE
DDD’’’AAASSSSSSOOOCCCIIIAAATTTIIIOOONNN

DIVERSITE BACTERIENNE

DIVERSITE BACTERIENNE
« Les parasites sélectionnent les
meilleurs hôtes et les hôtes les meilleurs
parasites. »
(C. Combes)

DIVERSITE BACTERIENNE

DIVERSITE BACTERIENNE
57
DIVERSITE BACTERIENNE
DIVERSITE INTERSPECIFIQUE
epuis la découverte des associations chimio-synthétiques, la connaissance de la
diversité des symbiotes chimioautotrophes a essentiellement été caractérisée via
l’étude de l’ARN ribosomal 16S (ARNr 16S). Récemment, compte tenu des
caractéristiques métaboliques de ces bactéries, d’autres marqueurs ont également été utilisés
tels que la Ribulose-1,5-bis-Phosphate Carboxylase/Oxygénase (RubisCO), caractéristique
des chimioautotrophes, l’adenosine-5’-Phosphate Réductase (APS Réductase) propre aux
sulfo-oxydantes et la méthane mono-oxygénase des méthanotrophes (Duperron et al., 2007;
Spiridonova et al., 2006; Vrijenhoek et al., 2007). Cependant, l’ARNr 16S continue à être le
marqueur le plus couramment utilisé pour caractériser les symbiotes associés dans ce type de
relations. Ce marqueur a également été utilisé dans l’étude de l’histoire évolutive de ces
symbioses, la question de l’origine des couples actuels étant posée. En effet, les deux
partenaires peuvent avoir co-évolué ou non à partir d’un couple symbiotique ancestral. La
comparaison des phylogénies moléculaires des hôtes et des symbiotes est l’un des moyens
permettant de trancher entre ces alternatives (Peek et al., 1998a). En effet, en cas de co-
évolution, les deux phylogénies doivent être congruentes attestant d’une histoire évolutive
commune des deux partenaires.
Chez les Lucinidae, des données moléculaires propres aux hôtes ou aux symbiotes ont
déjà été fournis par de précédents travaux. Cependant, aucunes données moléculaires
obtenues conjointement chez les hôtes et chez leurs symbiotes n’étaient à ce jour disponibles.
Ainsi, côté hôte, les relations entre espèces hôte appartenant aux Lucinacea (Thyasiridae,
Lucinidae et Fimbriidae etc…) ont été étudiées grâce aux ARNs ribosomaux 18S et 28S
(Williams et al., 2003). Côté symbiote, plusieurs études utilisant l’ARNr 16S ont permis la
caractérisation de la diversité symbiotique associée aux Lucinidae mais ont abouti à des
D

DIVERSITE BACTERIENNE
58
résultats contradictoires. Les premiers travaux ont été conduits par Distel et al. (1994; 1988).
Leurs analyses ont porté sur les symbiotes associés à 6 espèces hôtes réparties dans 4 genres
différents, ces espèces provenant d’habitats variés aussi bien en termes de localisation
géographique qu’en termes de biotope (Tableau B). Pour ces auteurs, les résultats obtenus
montrent que chaque espèce hôte héberge un type symbiotique qui lui est propre. Ceci leur a
suggéré une spécificité d’association entre symbiotes et hôtes les conduisant à poser
l’hypothèse « d’associations stables, espèces-spécifiques avec un seul type bactérien
dominant par espèce hôte. […] La congruence entre la phylogénie des symbiotes et la
classification des hôtes attestant d’une histoire évolutive commune » (Distel et al., 1994).
Ultérieurement, une autre série d’analyses a été réalisée au sein des Antilles Françaises
(Guadeloupe, la Désirade et Martinique) portant également sur 6 couples symbiotiques
répartis dans 4 genres d’hôtes (Tableau C). Dans ces dernières études, l’échantillonnage a été
réalisé dans un seul écosystème, les herbiers à phanérogames de Thalassia testudinum
(Durand and Gros, 1996; Durand et al., 1996; Gros et al., 2000; Gros et al., 2003a). Ces
auteurs ont mis en évidence une seule espèce bactérienne partagée par ces 6 espèces hôte de
Lucinidae. Cette dernière observation infirme donc l’hypothèse proposée précédemment (i.e.
association espèce-spécifique due à la co-évolution des deux partenaires à partir d’un couple
ancestral). En effet, selon ces auteurs, leur résultats ne « supportent pas l’hypothèse
d’associations monospécifiques entre « invertébrés » marins et bactéries chimioautotrophes »
(Gros et al., 2003a).
Ces premières études utilisant l’ARNr 16S ont abouti à des résultats contradictoires
conduisant les différents auteurs à émettre des hypothèses opposées. De plus, dans ces études,
la phylogénie moléculaire des bactéries n’a été comparée qu’à la seule classification
morphologique des hôtes. Or selon Taylor & Glover (2005), en dépit de l’intérêt porté à ce
groupe de bivalves, cette classification demeure confuse à tous les niveaux taxonomiques. Ce
n’est que plus tard, grâce à l’utilisation des outils moléculaires, qu’une grande partie de ces
incertitudes a pu être levée (Taylor et al., 2007). En conclusion, entre études moléculaires sur
les symbiotes aux conclusions contradictoires d’un côté et études moléculaires réalisées sur
les hôtes de l’autre, une comparaison entre phylogénie moléculaire des symbiotes et
phylogénie moléculaire des hôtes restait à faire. Cette dernière permettrait de mieux
comprendre l’origine et l’histoire évolutive des couples actuels.

DIVERSITE BACTERIENNE
59
Tableau B. Echantillonnage de Distel et al. (1994; 1988).
Espèces hôte Lieux de collecte Habitats Profondeursa (m)
Anodontia philipiana Ferry Reach (Bermudes) Mangrove Zone intertidale
Codakia costata Whalebone Bay (Bermudes) Herbier Zone intertidale
Codakia orbicularis Nassau(Bahamas) Herbier Zone intertidale
Lucina floridana St Joseph Bay (Floride) Herbier Zone intertidale
Lucinoma aequizonata Santa Barbara Basin (Californie) Sédiments 500
Lucinoma annulata Santa Monica Bay (Californie) Sédiments 100
a La zone intertidale correspond à la zone de balancement des marées.
Tableau C. Echantillonnage de Durand et al. (1996) ; Durand & Gros (1996) ; Gros et al.
(2000; 2003a).
Espèces hôte Lieux de collecte Habitats Profondeursa (m)
Anodontia alba Îlet Cochon (Guadeloupe) Herbier Zone intertidale
Codakia orbicularis Îlet Cochon (Guadeloupe) Herbier Zone intertidale
Codakia orbiculata Îlet Cochon (Guadeloupe) Herbier Zone intertidale
Codakia pectinella Îlet Cochon (Guadeloupe) Herbier Zone intertidale
Divaricella quadrisulcata Plage des galets (La Désirade) Herbier Zone intertidale
Linga pensylvanica Anse Macabou (Martinique) Herbier Zone intertidale
a La zone intertidale correspond à la zone de balancement des marées.
Je me suis donc intéressé à l’évolution des couples hôtes/symbiotes et à la diversité
bactérienne associée aux Lucinidae, en comparant les phylogénies moléculaires des deux
partenaires afin de déterminer si ces derniers avaient co-évolué ou non à partir d’un couple
symbiotique ancestral. J’ai réalisé ceci d’une part en reprenant les données de la littérature et
d’autre part en y ajoutant des résultats obtenus à partir de spécimens provenant de la Caraïbe
et des Philippines (Campagne PANGLAO 2005). J’ai ainsi établi la phylogénie moléculaire
des hôtes à l’aide des séquences des gènes des ARNr 18S et 28S et celle des symbiotes en
utilisant l’ARNr 16S d’une part et un gène caractéristique des bactéries sulfo-oxydantes,
l’APS Réductase d’autre part.

DIVERSITE BACTERIENNE
60
DEMARCHE EXPERIMENTALE.
ECHANTILLONNAGE.
Les différents échantillons philippins ont été collectés lors de la campagne PANGLAO
en mai 2005 (chef scientifique: P. Bouchet) dans la mer de Bohol (8,3-9,6°N et 123-124°E)
par chalutage à des profondeurs comprises entre 219 et 1775m. Les détails concernant les
différents spécimens sont présentés Tableau D. Les échantillons ont été traités comme suit
une heure après la collecte : une branchie de chaque spécimen a été stockée dans de l’éthanol
absolu pour l’analyse moléculaire et la seconde préparée pour l’étude en microscopie
électronique. Les spécimens de la Caraïbe ont été les suivants : Anodontia alba, Codakia
orbicularis et Lucina pectinata. Ces espèces hôte proviennent pour A. alba d’une mangrove
d’Haïti et d’un herbier de Guadeloupe, pour L. pectinata de mangroves d’Haïti et de
Guadeloupe et pour C. orbicularis, uniquement d’un herbier de Guadeloupe (Îlet Cochon).
EXTRACTIONS D’ADN ET AMPLIFICATIONS PAR PCR.
L’ADN matrice nécessaire à l’amplification par PCR a été extrait des branchies de
chaque individu selon la méthode CetylTrimethyl Ammonium Bromide (CTAB, Ishaq et al.,
1990). La concentration d’ADN de chaque échantillon a été contrôlée par spectrophotométrie
(BioPhotometer, eppendorf) et sur gel d’agarose 1% en comparaison avec un marqueur de
concentration (Smart Ladder, Eurogentec). Les gènes bactériens (ARNr 16S et APS
Réductase) et ceux des hôtes (ARNr 18S et 28S) ont été amplifiés à partir de l’ADN matrice
en utilisant les amorces présentées tableau E. Les amplifications ont été réalisées dans 50µL
avec 5µL de Taq buffer 10X (Invitrogen), 3mM de MgCl2 (Invitrogen), 0,4mM de chaque
dNTP, 1mM de chaque amorce, 1 unité de Taq DNA Polymérase Recombinante (Invitrogen)
et 25 à 50ng d’ADN. Les cibles sont amplifiées en 30 cycles (1 min à 94°C, 1 min au Tm
optimal [voir tableau E] et 1min 30 à 72°C) avec une dénaturation initiale de 10 min à 94°C et
une élongation finale de 15 min à 72°C. Les produits de PCR sont ensuite contrôlés sur gel
d’agarose à 1% et directement séquencés (GATC-biotech, http://www.gatc-biotech.fr).

DIVERSITE BACTERIENNE
61
Figure 12. Carte présentant les lieux d’échantillonnage des différentes espèces de Lucinidae analysées.
Distel et al. (1988 ;1994) Durand & Gros (1996) ; Durand et al. (1996)
Cette étude

DIVERSITE BACTERIENNE
62
Tableau D. Echantillonnage réalisé pour cette étude de la diversité des symbiotes de
Lucinidae
Code Espèce hôtea Lieu de collecte Habitat
b
Profondeur
(m)c
Localisation
GPS
OG18 Lucinoma sp.
Bohol Sea
Off Balicasag
Island
nd 271-318 9°29.4’N
123°43.7’E
OG57 nd Bohol Sea nd 1764-1775 9°24.3’N
124°10.7’E
OG98 Gloverina sp. Bohol/Sulu
Seas Sill nd 357-372
8°49.9’N
123°37.6’E
OG101 Cardiolucina
quadrata Bohol Sea nd 762-786
9°26.9’N
123°34.5’E
OG103 Myrtea
flabelliformis Bohol Sea nd 762-786
9°26.9’N
123°34.5’E
OG108 nd Bohol Sea nd 627 -645 9°27.4’N
123°43.1’E
ALBA Anodontia
alba
Îlet Cochon
(Guadeloupe) Herbier
Zone
intertidale
16°12'53.76''N
61°32'05.74''O
AH Anodontia
alba Haïti Mangrove
Zone
intertidale nd
CIS Codakia
orbicularis
Îlet Cochon
(Guadeloupe) Herbier
Zone
intertidale
16°12'53.76''N
61°32'05.74''O
LPEC Lucina
pectinata
La manche à eau
(Guadeloupe) Mangrove
Zone
intertidale
16°16’37’N
61°3’25.91’’O
LH Lucina
pectinata Haïti Mangrove
Zone
intertidale nd
a Concernant les échantillons des Philippines, la détermination des espèces a été réalisée par John D. Taylor
(NHM, UK) sur la base des caractères morphologiques de la coquille. b La récolte ayant été réalisée par chalutage, les biotopes précis sont inconnus pour les spécimens des
Philippines, ceci dit, compte tenu de la profondeur il est vraisemblable qu’il s’agisse de sédiments réduits. c La zone intertidale correspond à la zone de balancement des marées. d nd : non déterminé

DIVERSITE BACTERIENNE
63
ANALYSES PHYLOGENETIQUES
Les séquences obtenues ont été contrôlées par BLAST
(www.ncbi.nlm.nih.gov/blast.cgi) et alignées avec ClustalW paramétré par défaut (Thompson
et al., 1994). L’alignement a ensuite été vérifié et corrigé manuellement si nécessaire. Les
« gaps » et les positions ambigües ont été retirés de l’alignement en utilisant Gblocks 0.91b
(Castresana, 2000). L’alignement a ensuite été importé dans le logiciel TOPALi v2.5 (Milne
et al., 2004) pour définir, en utilisant ModelGenerator (Keane et al., 2006), le modèle de
substitution à utiliser pour chaque jeu de données. Le meilleur modèle est celui possédant un
score de « Bayesian Information Criterion » le plus bas (Schwarz, 1978). Les arbres ont
ensuite été reconstruits par Maximum de Vraisemblance (MV) en utilisant PhyML 2.5
(Guindon and Gascuel, 2003) et par Inférence Bayesienne (IB) en utilisant MrBayes3
(Ronquist and Huelsenbeck, 2003). Les supports pour les différents nœuds ont été évalués
utilisant les bootstrap non paramétriques (Felsenstein, 1985) obtenus pour 100 répétitions
(MV). Les probabilités postérieures de chaque nœud (IB) ont également été évaluées (runs=5,
generations=1 000 000, sampling=1 000, burn=10%).
Tableau E. Amorces et conditions de PCR utilisées dans cette étude.
Noms Séquences (5’→ 3’) Tma Cibles Ref
b
16S_UNIV-8f AGAGTTTGATCATGGCTCAG 54°C ARNr 16S [1]
16S_UNIV-1492r TACGGCTACCTTGTTACGACT
APS-FW TGGCAGATMATGATYMACGGG 58°C APS Reductase [2]
APS-RV GGGCCGTAACCGTCCTTGAA
LSU-900f CCGTCTTGAAACACGGACCAAG 52°C ARNr 28S
[3]
LSU-1600r AGCGCCATCCATTTTCAGG [4]
18S-5’f CTGGTTGATYCTGCCAGT 54°C ARNr 18S
[5]
18S-1100r CTTCGAACCTCTGACTTTGG [4]
a Température optimale pour l’amplification par PCR. b Références bibliographiques : [1] (Lane, 1991), [2] (Deplancke et al., 2000), [3] (Olson et al., 2003), [4]
(Williams et al., 2003), [5] (Winnepenninckx et al., 1998).

DIVERSITE BACTERIENNE
64
EVOLUTION DES COUPLES SYMBIOTIQUES
Que ce soit pour les gènes bactériens (ARNr 16S et APS Réductase) ou ceux des hôtes
(ARNr 18S et 28S), l’analyse des chromatogrammes a révélé une séquence unique par
échantillon analysé. Les phylogénies basées sur les séquences des ARNr 18S et 28S étant
congruentes, la phylogénie des hôtes a été réalisée après concaténation des séquences de ces
gènes (1252 sites) avec comme groupes extérieurs les Crassatellidae et les Thyasiridae,
groupes frères des Lucinidae (Fig. 12). Cette phylogénie permet de replacer nos échantillons
dans l’arbre des Lucinidae au sein des clades décrits par Williams et al. (2003). Clade
Anodontia pour OG57 et OG98, clade Myrtea pour OG108 et clade B pour OG101. L’ARNr
18S d’OG18 n’a pu être obtenu compte tenu de l’état de conservation de l’ADN. Néanmoins
la phylogénie basée sur l’ARNr 28S permet de placer ce spécimen dans le clade B. Enfin
concernant OG103, aucune séquence hôte n’a pu être obtenue, cependant la détermination de
l’espèce sur des caractères morphologiques permet de placer ce spécimen dans le clade
Myrtea (Myrtea flabelliformis, John Taylor communication personnelle). Enfin concernant les
spécimens A. alba, C. orbicularis et L. pectinata collectés aux Caraïbes (Haïti et
Guadeloupe), les séquences obtenues à partir des hôtes correspondent à celle publiées par
Williams et al. (2003).
Les Lucinidae philippins, hébergent tous des bactéries symbiotiques. En se basant sur
ces séquences d’ARNr 16S ces symbiotes appartiennent au clade monophylétique
correspondant aux symbiotes de Lucinoidea (Fig. 13). Par ailleurs, le fait qu’il s’agisse de
bactéries sulfo-oxydantes a été attesté par la détection par PCR du gène de l’APS Réductase.
L’arbre phylogénétique basé sur les séquences de ce gène (Fig. 14) indique que les symbiotes
philippins possèdent comme ceux des Lucinidae analysés à ce jour une APS Réductase
appartenant à la lignée II des APS Réductases de bactéries sulfo-oxydantes (Meyer and
Kuever, 2007)
La phylogénie des symbiotes (Fig. 13) et la matrice d’identité (Tableau F) permettent
de définir trois clades A, B et C qui correspondent à trois espèces bactériennes différentes
(pourcentage d’identité inférieur à 97%). L’espèce symbiotique « A » (Fig. 13, clade A) est
associée à l’espèce hôte Lucina pectinata. L’espèce « B » (Fig. 13, clade B) est associée aux
bivalves du genre Anodontia à l’exception d’Anodontia alba. La troisième espèce « C »
correspond aux autres symbiotrs de Lucinidae décris jusqu’alors.

DIVERSITE BACTERIENNE
65
Cet arbre a été reconstruit par Maximum de Vraisemblance (MV) et Inférence Bayesienne (IB) avec comme
modèles d’évolution le TrNef+Г (MV) et le SYM+Г (IB). Les Crassatellidae et Thyasiridae ont été choisis
comme groupes extérieurs. La robustesse des différents nœuds a été évaluée et seules les valeurs de bootstrap
(MV, audessus des branches) et les probabilités postérieures (IB, au dessous des branches) supérieures à 80%
sont affichées. Les séquences en gras correspondent aux séquences obtenues lors de cette étude et les autres sont
tirées de Williams et al. (2003). a Pour ces espèces hôte, la même séquence est retrouvée quelque soit le lieu de collecte (Haïti ou Guadeloupe). b Les 3 séquences obtenues ici correspondent à celles publiées dans Williams et al. (2003).
La phylogénie basée sur les séquences de l’ARNr 16S, montre que les symbiotes des
spécimens de lucines des Philippines appartiennent à une seule et même espèce (Fig. 13, clade
C). En effet, la matrice d’identité réalisée en « pairwise deletion » sur les séquences d’ARNr
16S (Tableau F) indique une identité moyenne entre les séquences supérieure à 97% (97,7%).
On observe de plus, trois ribotypes bactériens répartis en deux groupes. Le premier est partagé
par OG18, OG98 et OG108 (Fig. 13, groupe I). Ce groupe semble correspondre exactement à
un seul et même ribotype (identité moyenne de 99,9%). Le second groupe est représenté par
les symbiotes des spécimens OG57, OG101 et OG103 avec apparemment deux ribotypes
différents (un propre à OG57 et un partagé par OG101 et OG103 [99,8%]). Il y a donc au sein
des six espèces hôtes de Lucinidae des Philippines une seule espèce bactérienne
vraisemblablement composée de trois variants. Ces résultats recoupent ceux obtenus par
Durand & Gros (1996), Durand et al. (1996) et Gros et al. (2000; 2003a), dans lesquels
Figure 13. Phylogénie moléculaire des Lucinidae réalisée après concaténation des séquences des ARNs ribosomaux 18S et 28S (1252 sites).

DIVERSITE BACTERIENNE
66
différentes espèces hôtes hébergent la même espèce bactérienne ou le même ribotype
bactérien. Tous ces résultats infirment donc l’hypothèse d’associations espèces-spécifiques
dues à la coévolution des deux partenaires à partir d’un couple symbiotique ancestral
proposée initialement par Distel et al. (1994; 1988). J’en conclus donc que, chez les
Lucinidae, l’association serait plutôt labile et pourrait se constituer et se défaire en fonction de
l’espèce ou du type bactérien présent dans l’environnement. Qui plus est, cette hypothèse peut
également s’appliquer aux observations de Distel et al. (1994; 1988). En effet la diversité
aussi bien en terme de biotopes que de localisations géographiques de l’échantillonnage
pourrait simplement mimer une spécificité d’association liée à l’hôte. Les distances
génétiques observées entre symbiotes n’étant pas dues à une quelconque spécificité ou
coévolution mais plutôt aux distances géographiques entre les biotopes où avaient été
échantillonnées ces espèces hôte. De façon intéressante, les espèces bactériennes « A » et
« B » correspondent à des symbiotes hébergés par des hôtes retrouvés dans des
environnements où les concentrations en sulfures sont assez élevées (i.e. boues de
mangroves), comparés aux herbiers à T. testudinum (Dando et al., 1985) ou l’on retrouve
l’espèce « C ». Ces observations suggèrent donc que les espèces bactériennes se répartissent
de façon différente d’un biotope à l’autre. Ce point renforce donc l’hypothèse d’association en
fonction du type bactérien présent dans l’environnement.

DIVERSITE BACTERIENNE
67
Figure 14. Phylogénie moléculaire des symbiotes chimioautotrophes réalisée grâce aux séquences d’ARNr 16S
(1269 sites).
Cet arbre a été reconstruit par Maximum de Vraisemblance (MV) et Inférence Bayesienne (IB) avec comme
modèle d’évolution le K80+I+Г. Des bactéries marines libres ont été choisies comme groupe extérieur. La robustesse des différents nœuds a été évaluée et seules les valeurs de bootstrap (MV, 100 répétitions [au-dessus
des branches]) et les probabilités postérieures (IB, runs=5, générations=1 000 000, échantillonnage=1 000,
burn=10% [au-dessous des branches]) supérieures à 80% sont affichées. Les séquences en gras correspondent
aux séquences obtenues lors de cette étude. HAI : Haïti, GWA : Guadeloupe

DIVERSITE BACTERIENNE
68
Figure 15. Phylogénie moléculaire des symbiotes chimioautotrophes réalisée grâce aux séquences d’APS Réductase (273 sites).
Cet arbre a été reconstruit par Maximum de Vraisemblance (MV) et Inférence Bayesienne (IB) avec comme
modèles d’évolution le TrNef+Г (MV) et le K80+Г (IB). Des archae on été choisies comme groupe extérieur. La
robustesse des différents nœuds a été évaluée et seules les valeurs de bootstrap (MV, 100 répétitions [au-dessus
des branches]) et les probabilités postérieures (IB, runs=5, générations=1 000 000, echantillonnage=1 000,
burn=10% [au-dessous des branches]) supérieures à 70% sont affichées. Les séquences en gras correspondent
aux séquences obtenues lors de cette étude. HAI : Haïti, GWA : Guadeloupe.

DIVERSITE BACTERIENNE
69
Tableau F. Matrice d’identité réalisée en « pairwise deletion » utilisant les séquences d’ARNr 16S.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
1: Codakia orbicularisa ID
2: Anodontia albaa 1,000 ID
3: Lucina nassulaa 0,984 0,984 ID
4: Codakia costataa 0,975 0,975 0,979 ID
5: Lucina floridanaa 0,971 0,971 0,981 0,996 ID
6: OG57a 0,966 0,966 0,971 0,976 0,978 ID
7: OG101a 0,970 0,970 0,973 0,982 0,980 0,985 ID
8: OG103a 0,970 0,970 0,973 0,978 0,978 0,980 0,993 ID
9: Lucinoma aff. kazania 0,969 0,969 0,973 0,981 0,978 0,982 0,994 0,990 ID
10: Lucinoma aequizonataa 0,960 0,960 0,963 0,968 0,966 0,981 0,976 0,973 0,974 ID
11: OG18a 0,977 0,977 0,978 0,979 0,976 0,988 0,981 0,984 0,977 0,969 ID
12: OG98a 0,971 0,971 0,972 0,973 0,972 0,985 0,978 0,978 0,973 0,968 0,999 ID
13: OG108a 0,969 0,969 0,970 0,971 0,972 0,986 0,976 0,976 0,971 0,969 0,999 0,999 ID
14: Anodontia ovuma 0,951 0,951 0,959 0,957 0,959 0,952 0,953 0,952 0,951 0,944 0,960 0,955 0,953 ID
15: Anodontia phillipianaa 0,947 0,947 0,955 0,949 0,951 0,946 0,947 0,946 0,946 0,938 0,954 0,949 0,946 0,990 ID
16: Thyasira flexuosaa 0,942 0,942 0,944 0,945 0,942 0,941 0,941 0,939 0,940 0,933 0,950 0,944 0,942 0,947 0,945 ID
17: Lucina pectinataa 0,947 0,947 0,949 0,941 0,939 0,933 0,933 0,932 0,933 0,926 0,941 0,936 0,934 0,945 0,943 0,941 ID
a Ces séquences correspondent aux symbiotes des hôtes cités.

DIVERSITE BACTERIENNE
70
Reste néanmoins un cas particulier, celui d’Anodontia alba. Que cette espèce ait été
collectée dans les herbiers de Guadeloupe ou dans les mangroves d’Haïti, elle héberge la
même espèce bactérienne. Ces observations suggèrent donc que la forme bactérienne associée
à cette espèce hôte peut être retrouvée à la fois en mangroves et en herbiers. Des expériences
préliminaires de CARD-FISH sur filtre réalisées sur les sédiments de mangrove de
Guadeloupe grâce à une sonde spécifique du symbiote de Codakia orbicularis (Symco2A,
Gros et al., 2003b), m’ont permis de mettre en évidence que l’espèce bactérienne décrite dans
les herbiers pouvait également être présente dans ces sédiments. Il semblerait donc que cette
espèce hôte s’associe préférentiellement à la forme bactérienne d’herbiers quand les deux
formes de symbiotes cohabitent. Ainsi, en cas de coexistence de plusieurs formes bactériennes
une éventuelle spécificité d’association pourrait néanmoins exister.
Cette faible diversité observée chez les symbiotes sulfo-oxydants associés aux
Lucinidae pose d’autres questions. Comment expliquer que des hôtes si éloignés sur le plan
géographique puissent héberger la même espèce bactérienne (Fig. 13, clade C) ? En effet,
dans ce clade C, on retrouve des symbiotes hébergés par des spécimens provenant des
Philippines et d’autres comme Lucina floridana (Distel et al., 1994), Lucinoma aff. kazani
(Duperron et al., 2007), Lucinoma aequizonata (Distel et al., 1994) ou Codakia orbicularis
(Distel et al., 1988; Durand et al., 1996) provenant d’autres zones géographiques
(respectivement Floride, mer Méditerranée, Californie et Caraïbe). Deux hypothèses peuvent
expliquer le faible niveau de diversité de ces symbiotes : (i) une divergence géographique des
populations symbiotiques trop récente pour mettre en évidence une divergence génétique, (ii)
un taux d’évolution du marqueur utilisé (ARNr 16S) trop faible. La seconde hypothèse
apparaît comme plus vraisemblable car il a été démontré par Peek et al. (1998b) que les
séquences d’ARNr 16S des symbiotes acquis à partir de l’environnement (comme c’est le cas
pour les symbiotes de Lucinidae) avaient un taux d’évolution plus lent que les symbiotes à
transmission verticale et les bactéries marines libres.

DIVERSITE BACTERIENNE
71
DIVERSITE INTRA-SPECIFIQUE.
ous avons vu dans la partie précédente que la diversité des symbiotes de Lucinidae
était relativement faible. Néanmoins, cette diversité pourrait être sous estimée eu
égard au marqueur utilisé. En effet, compte tenu du taux d’évolution relativement
lent de l’ARNr 16S, il pourrait exister chez ces symbiotes une diversité que ce marqueur ne
permettrait pas de mettre en évidence. De fait, au sein des herbiers à T. testudinum des
Antilles Françaises, différentes souches appartenant à l’espèce bactérienne hébergée par les
six espèces hôte décrites ci-dessus (Tableau B) pourraient être présentes.
Ce type de patron (espèces hôte différentes partageant une même espèce bactérienne) a
déjà été décrit dans deux autres familles d’ « invertébrés » marins hébergeant également des
bactéries thioautotrophes : les Siboglinidae (Feldman et al., 1997) et les Mytilidae (Won et
al., 2003). Chez les Siboglinidae, Feldman et al. (1997) ont conclu que l’association
symbiotique semblait se constituer en fonction du biotope plutôt que de l’espèce hôte. Ainsi
trois espèces bactériennes ont été décrites. La première est hébergée par différentes espèces
hôte colonisant les sources hydrothermales basaltiques. La seconde est retrouvée chez des
espèces hôte vivant au niveau des suintements froids et la troisième est uniquement hébergée
par une espèce de ver (Escarpia spicata) colonisant les carcasses de baleines. Chez les
Mytilidae, Won et al. (2003) ont décrit deux espèces hôte (Bathymodiolus azoricus et
Bathymodiolus puteoserpentis) de la Ride Medio Atlantique hébergeant des bactéries sulfo-
oxydantes possédant le même ribotype d’ARNr 16S. Ces deux espèces hôtes sont réparties au
Nord ou au Sud de la Ride avec une zone de chevauchement au niveau du site de Broken
Spur.
Selon les auteurs, l’ARNr 16S a permis au sein de ces deux modèles de corréler la
diversité symbiotique avec des facteurs géographiques ou environnementaux. Néanmoins, la
résolution de ce marqueur ne permettait pas la visualisation de variations génétiques à
l’intérieur des « clades géographiques » ou des « clades environnementaux » définis, alors
même que les espèces hôtes y étaient différentes. D’autres analyses de diversité plus fines
utilisant d’autres marqueurs ou techniques ont donc été utilisées. Ces dernières ont permis de
mettre en évidence une diversité de souches non décelable par l’utilisation de l’ARNr 16S.
C’est ainsi que le marqueur ITS (Internal Transcribed Spacer), région transcrite de l’ARN
ribosomal située entre les ARNr 16S, 23S ou 5S a été utilisé.
N

DIVERSITE BACTERIENNE
72
Chez les Siboglinidae, l’utilisation du marqueur ITS a permis de mettre en évidence
une diversité génétique des symbiotes corrélée aux distances géographiques et non plus aux
biotopes (Di Meo et al., 2000). Ainsi chez ces vers tubiformes l’ARNr 16S révèle une
diversité répartie en fonction de l’habitat et l’ITS une diversité génétique liée à la localisation
géographique. Les auteurs en ont conclu que différents facteurs pouvaient ainsi affecter la
diversité des symbiotes de vestimentifères. Néanmoins, l’utilisation de ce marqueur ITS ne
révélait pas de diversité de souches au sein d’environnements proches ou identiques. Les
auteurs ont donc eu recours à la REP-PCR (Repetitive-Extragenic Palindrome PCR) qui a
permis de mettre en évidence une diversité de souches bactériennes influencée de façon
simultanée par trois facteurs : le genre auquel appartient l’hôte, le type d’habitat et la
localisation géographique. L’hôte semble être le premier facteur influençant la diversité des
symbiotes, ceux hébergés par des hôtes de même genre formant un clade particulier. A
l’intérieur de ces clades, les symbiotes se différencient ensuite selon le type d’habitat ou la
géographie. Dès lors, au sein des deux clades liés aux biotopes définis par l’ARNr 16S (i.e.
sources hydrothermales et suintements froids) il existerait quatre groupes de symbiotes qui se
constituent selon le genre auquel appartient l’hôte, puis selon le biotope ou la localisation
géographique.
Chez les Mytilidae, le même marqueur ITS a permis de caractériser trois souches
bactériennes différentes (sym-az, sym-pu1 et sym-pu2) réparties en deux lignées (az pour
B. azoricus et pu pour B. puteoserpentis). Ces deux lignées correspondent parfaitement pour
l’une (az) à la zone nord de la Ride Medio Atlantique où est retrouvée B. azoricus et pour
l’autre (pu) à la zone Sud où l’on rencontre plutôt B. puteoserpentis. Au niveau du site de
Broken Spur, les hôtes sont essentiellement associés avec une seule souche bactérienne (sym-
pu1) avec quelques rares cas de bi-infections (sym-az/sym-pu1). Nous retrouvons ainsi, au
niveau intra-spécifique, un patron d’association ayant une composante environnementale mais
avec également une spécificité liée à l’hôte en cas de coexistence de deux souches
symbiotiques sur le même site.
Dans ces deux modèles, l’utilisation de l’ARNr 16S dépeint donc une diversité
bactérienne relativement faible et liée à l’environnement (Nord/Sud de la Ride Medio
Atlantique pour les Mytilidae et type d’habitat pour les vestimentifères). Par contre l’emploi
de techniques plus fines met en évidence une diversité de souches influencée certes par
l’environnement mais aussi par la diversité d’hôtes hébergeant ces symbiotes.

DIVERSITE BACTERIENNE
73
Ces deux associations, même si elles sont plutôt retrouvées dans des environnements
profonds (e.g. sources hydrothermales), présentent certaines similitudes avec le modèle
Lucinidae. En effet, comme chez les lucines, le symbiote n’est pas transmis verticalement
mais est plutôt acquis à partir de l’environnement (Nussbaumer et al., 2006; Won et al.,
2003), les formes libres des symbiotes de vers tubiformes ayant même été décrites dans la
colonne d’eau à proximité des sources hydrothermales (Harmer et al., 2008). De fait, dans le
cas des Lucinidae, il pourrait exister une diversité intra-spécifique qui serait structurée en
fonction de l’environnement, du biotope ou des hôtes. C’est pourquoi j’ai analysé les
symbiotes de six espèces hôtes de Lucinidae (Anodontia alba, Divaricella quadrisulcata,
Codakia orbicularis, Codakia orbiculata, Codakia pectinella et Linga pensylvanica ; Cf. p.
33) dans le but de mettre en évidence cette éventuelle diversité intra-spécifique. Pour ce faire,
j’ai développé des marqueurs qui ont été utilisés dans une approche MLST (Multi-Locus
Sequence Typing). Cependant, au début de cette étude les bases de données étant relativement
pauvres en séquences de symbiotes de Lucinidae (Fig. 15), la première partie du travail a été
consacrée au choix de marqueurs appropriés et à l’obtention de leurs séquences.
RECHERCHE DE MARQUEURS DE DIVERSITE.
Un intérêt tout particulier a été apporté à cette partie de l’analyse, le choix des
marqueurs étant un facteur essentiel. En effet, un mauvais choix de marqueur pourrait
conduire à une mauvaise estimation de la diversité. Selon Cooper & Feil (2004), les
marqueurs utilisés couramment ou convenant bien aux analyses MLST peuvent se classer en
trois catégories : (i) Les gènes de ménage, qui sont des gènes ubiquitaires et impliqués dans
des fonctions essentielles telles que la transcription ou la traduction. En général le taux
d’évolution de ces gènes est assez bas. (ii) Les « core metabolism genes », qui sont présents
dans un groupe donné et participent essentiellement au métabolisme central du groupe que
l’on veut étudier. Ces gènes évoluent de façon plus ou moins rapide. (iii) Les hypervariables
qui peuvent ne pas être ubiquitaire dans le groupe étudié et qui évoluent très vite. Ils
correspondent souvent à des protéines de surface et peuvent intervenir dans la virulence des
micro-organismes pathogènes.

DIVERSITE BACTERIENNE
74
Figure 16. Exploration des bases de données.

DIVERSITE BACTERIENNE
75
Dans cette étude, les gènes candidats ont été choisis dans ces 3 groupes. Ainsi l’ITS1
(situé entre les ARNr 16S et 23S) a été choisi comme marqueur hypervariable. L’APS
Reductase (caractéristique des sulfo-oxydantes) et la RubisCO (propre aux
chimioautotrophes) ont été retenus comme « core housekeeping genes ». Concernant les
gènes de ménage nous avons choisi des gènes qui ont déjà été utilisés au cours d’analyses de
diversité bactérienne. Les candidats retenus à partir de la littérature sont présentés Tableau G.
Néanmoins, eu égard à leur ubiquité, ces derniers ont été soumis à une analyse critique
permettant de les valider ou non pour l’étude du groupe qui nous intéresse (i.e. les symbiotes
sulfo-oxydants). Ainsi ces marqueurs provenant de la littérature devaient répondre à 3 critères
qui nous semblent essentiels pour être qualifiés de « bon marqueur de diversité » dans le cadre
de notre étude: (i) Variabilité supérieure à l’ARNr 16S, (ii) Absence de transferts horizontaux,
(iii) Présence de séquences conservées encadrant des séquences hypervariables.
DEMARCHE EXPERIMENTALE.
ECHANTILLONNAGE.
Pour la recherche des marqueurs appropriés, des individus adultes de Codakia
orbicularis (coquille de 40mm environ) ont été collectés en Guadeloupe (Îlet Cochon
[16°12'53.76''N 61°32'05.74''O]). En ce qui concerne l’analyse de la diversité, trois adultes
d’Anodontia alba (~20mm), de C. orbicularis (~40mm), de Codakia orbiculata (~15mm) et
de Codakia pectinella (~25mm) ont été collectés à la main en Guadeloupe (Fig. 17). Les 3
spécimens de Divaricella quadrisulcata (Orbigny, 1842), ont été collectés à la Désirade
(plage des galets [16°17'46.88''N; 61°04'53.77''W]) et ceux de Linga pensylvanica (Linné,
1758) en Martinique (Anse Macabou [14°31'01.52''N; 60°49'00.86''W]). Les sub-adultes de
C.orbicularis (~10mm) ont également été échantillonnés en Guadeloupe (Îlet Cochon). Tous
les individus ont été collectés à l’intérieur (ou à proximité) des herbiers à Thalassia
testudinum. Pour chaque spécimen les branchies et les tissus non symbiotiques (i.e. pied ou
manteau) ont été conservés dans de l’éthanol absolu avant analyse.

DIVERSITE BACTERIENNE
76
Figure 17. Carte des zones échantillonnées pour l’analyse de la diversité intra-spécifique.
Ilet Cochon (Anodontia alba, Codakia orbicularis, C.orbiculata, C. pectinella)
Plage des galets (Divaricella quadrisulcata)
Anse Macabou, Martinique (Linga pensylvanica)

DIVERSITE BACTERIENNE
77
Tableau G. Gènes-candidats retenus à partir de la littérature.
Candidats Gène Fonctionsa Ref.
b
ADN Polymerase III
(sous-unité α) dnaE
Métabolisme des
purines/pyrimidines
Réplication de l’ADN
MMR
Recombinaison homologue
[1]
Chaperonine groEL
(large sous-unité de groESL) groEL
Protéine chaperonne
Repliement des protéines [2,3]
ADN Gyrase B gyrB Surenroulement de la double hélice
Recombinaison [4]
ADN Polymérase I polA
Métabolisme des
purines/pyrimidines
Réplication de l’ADN
BER/NER
Recombinaison homologue
[5]
Recombinase A recA Recombinaison homologue [1,4]
ARN Polymerase
(sous-unité β) rpoB
Métabolisme des
purines/pyrimidines
Transcription
[4]
a Fonctions associées à ces gènes selon la base de données KEGG (Kyoto Encyclopedia of Genes and Genomes,
http://www.genome.jp/kegg/genes.html). MMR : Mismatch Repair, BER :Base Excision Repair, NER :
Nucleotide Excision Repair. b [1] (Byun et al., 1999), [2] (Casiraghi et al., 2005), [3] (Tan et al., 2005), [4] (Holmes et al., 2004), [5] (Ardell
et al., 2003).
VALIDATION DES MARQUEURS DE DIVERSITE.
Nous avons testé les marqueurs choisis à partir de la littérature selon les critères
définis ci-dessus. (i) Variabilité du marqueur supérieure à celle de l’ARNr 16S : à partir d’une
matrice d’identité en « pairwise deletion » un pourcentage moyen de divergence
interspécifique (basé sur un alignement multiple de dix gamma-protéobactéries [Annexe 2]) et

DIVERSITE BACTERIENNE
78
un pourcentage moyen de divergence intra-spécifique (à partir de deux alignements multiples
de dix souches d’Escherichia coli et quatre de Legionella pneumophila [Annexe 2]) ont été
estimés. (ii) Absence de transferts horizontaux : des arbres phylogénétiques ont été
reconstruits par Maximum de Vraisemblance pour chacun des gènes en utilisant un
alignement multiple de 28 bactéries [Annexe 3]. (iii) Présence de régions conservées et de
régions variables : un alignement de dix gamma-protéobactéries (Annexe 2) a été importé
dans SVARAP 2.0 (Colson et al., 2006) afin de pouvoir visualiser la variabilité du gène le
long de sa séquence (fenêtre non glissante de 50 nucléotides). Afin de tester les transferts
horizontaux à l’intérieur du groupe des γ-protéobactéries, un Au-test (ref) a été réalisé. Ce
dernier testant la vraisemblance d’un arbre selon des séquences données. A l’aide de TREE-
PUZZLE v5.2 (option « -wsl » avec un model d’évolution HKY+Г), les topologies de nos
marqueurs ont été confrontées à un alignement de 10 séquences de γ-protéobactéries. De fait
les relations entre espèces et le positionnement de ces dernières les unes par rapport aux autres
(qui sont définis par l’ARNr 16S) ont pu être testés pour les différents marqueurs pouvant
attester de l’existence d’éventuels transferts horizontaux.
PREPARATION DE L’ADN ET CONSTRUCTION DES AMORCES SPECIFIQUES ET DEGENEREES.
L’ADN a été extrait comme décrit précédemment à partir des branchies pour les
symbiotes et à partir de tissus dépourvus de symbiotes (i.e. pied ou manteau) pour les hôtes
(Cf. page 54). Les amorces dégénérées et spécifiques utilisées pour ce travail sont détaillées
Tableau H. Les amorces dégénérées ont été déterminées en utilisant un alignement de dix
gamma-protéobactéries et le logiciel SVARAP 2.0 donnant site à site les proportions relatives
de chaque nucléotide. Les amorces spécifiques ont été déterminées en utilisant le logiciel
Primer3 ([http://frodo.wi.mit.edu/cgi-bin/primer3/primer3.cgi] Rozen and Skaletsky, 2000 ).
OBTENTION DES SEQUENCES DE SULFO-OXYDANTES POUR LES MARQUEURS VALIDES.
Nested-PCR : Une fois les marqueurs validés, le but a été d’obtenir les séquences de
ces gènes pour les symbiotes sulfo-oxydants de Lucinidae. Nous avons donc utilisé une
approche de PCR emboitée avec amorces dégénérées (Annexe 4). Cette PCR est réalisée en
deux étapes à partir de l’ADN extrait de C. orbicularis. La première étape est réalisée dans
25µL utilisant les amorces « nested » (Tableau H) et 10 à 25ng d’ADN. La seconde
amplification est réalisée dans les mêmes conditions avec les amorces « degN » au lieu des

DIVERSITE BACTERIENNE
79
« nested » et 1µL de produit de l’amplification précédente dilué 100 à 1000 fois. Les
températures optimales d’amplification pour chaque couple d’amorces sont déterminées par
PCR à gradient (Températures d’hybridation comprises entre 46 et 60°C). Le produit de PCR
obtenu est ensuite contrôlé sur gel d’agarose à 1%.
Clonage : Les fragments d’intérêt sont élués après dépôt de 100µL de produit de PCR
provenant de la seconde amplification (5x20µL). Pour ce faire, la bande d’intérêt est
découpée du gel puis un SPINX est réalisé (Spin-X®, Costar). L’ADN récupéré est ensuite
précipité sur la nuit à -20°C par ajout d’Acétate de Sodium (3M) et d’éthanol 100%. Le tout
est culotté par centrifugation (30 min à 11000 g), puis lavé 2 fois à l’éthanol 70% (15 min à
11000 g). Le culot est séché au SpeedVac concentrator (Savant) pendant 15 min et l’ADN est
resuspendu sur la nuit à +4°C dans 50 à 100µL d’eau bi-distillée « RNase/DNase free ». Le
clonage est réalisé à l’aide du kit pGEM®-T vector system (Promega) selon les
recommandations du fabricant.
Transformation : Des bactéries Escherichia coli Dh5-α compétentes sont transformées
par choc thermique (50 sec à 42°C) avec 2µL de produit de ligation. Les transformants sont
sélectionnés sur milieu LB agar + ampicilline (100mg/mL) + Xgal (25mg/mL) + IPTG
(25mg/mL). Les clones positifs (clones blancs) sont ensuite criblés par PCR pour : (i) la taille
de l’insert : une PCR avec les amorces SP6 et T7 du vecteur est réalisée, le résultat attendu
étant un fragment de la taille du fragment élué plus 130 pb (distance entre ces 2 amorces sur
le vecteur « fermé ». (ii) la séquence : deux PCR sont réalisées avec l’amorce SP6 du vecteur
et l’amorce sens ou anti-sens « degN » utilisée. Le résultat attendu étant une amplification
dans un cas et pas dans l’autre.
Séquençage : Les clones positifs sont ensuite mis en culture sous agitation sur la nuit à
37°C dans 5mL de milieu LB additionné d’ampicilline (0,1mg/mL). Les plasmides sont
ensuite extraits grâce au kit Nucléospin® Plasmid (Macherey-Nagel) et directement
séquencés (COGENICS, www.cogenics.com).
AMPLIFICATIONS PAR PCR
Après obtention des séquences des bactéries sulfo-oxydantes et validation de ces
dernières, des amorces spécifiques ont été définies et utilisées pour les PCR spécifiques. Ces
PCRs sont réalisées dans 50µL avec 5µL de Tampon Taq 10X (Invitrogen), 3mM de MgCl2

DIVERSITE BACTERIENNE
80
(Invitrogen), 0,4mM de chaque dNTP, 1mM de chaque amorce, une unité de Recombinant
Taq DNA Polymérase (Invitrogen) et 25 à 50 ng d’ADN. Les gènes des hôtes (ARNr 18S et
28S) sont amplifiés de la même façon. Tous ces produits de PCR sont directement séquencés
(GATC biotech, www.gatc-biotech.com)
ANALYSE PHYLOGENETIQUE
L’analyse phylogénétique est réalisée comme décrit lors de l’étude de la diversité
inter-spécifique (Cf. page 56).
ANALYSE DU POLYMORPHISME
Les séquences de symbiotes obtenues pour chaque marqueur ont été alignées afin de
visualiser le nombre de sites polymorphes. L’analyse des haplotypes a ensuite été réalisée
grâce au logiciel DNAsp4 (Rozas et al., 2003), les réseaux ayant été obtenus à l’aide de
NETW4510© (Fluxus Technology Ltd.) en utilisant la méthode de Median Joining (Bandelt
et al., 1999).
ANALYSE DES SEDIMENTS GRACE AU GENE DNAE
Du sable provenant de la couche supérieure des herbiers à T. testudinum de l’Îlet
Cochon a été collecté. L’ADN total a été extrait selon Zhou et al. (1996) et une PCR emboitée
en trois étapes a été réalisée à partir de 50ng d’ADN et les amorces « nested » (Tableau G).
Ce produit de PCR dilué 100 fois a servi de matrice pour la seconde amplification réalisée
avec les amorces « degN » (Tableau H) et cette procédure a été répétée une dernière fois avec
les amorces spécifiques. Le fragment d’intérêt a été élué à partir d’un gel d’agarose 1% en
utilisant le Nucleospin® Extract II kit (Macherey-Nagel) et cloné avec le kit pGEM-T® TA
cloning kit (Promega). Les vecteurs ont ensuite été utilisés pour transformer des Escherichia
coli Dh5α compétentes (Promega) et les plasmides des clones positifs ont été collectés en
utilisant le Nucleospin® plasmid kit (Macherey-Nagel). Les inserts ont ensuite été
directement séquencés (Genome express, www.gexbyweb.com).

DIVERSITE BACTERIENNE
81
Tableau H. Amorces utilisées au cours de l’étude de la diversité intra-spécifique.
Amorces Conditions
Noms Séquences (5'->3') Gène
cible Tm
b Ref
c
Hôte
s
18S-5'f CTGGTTGATYCTGCCAGT ARNr
18S
54°C 1
18S-1100r CTTCGAACCTCTGACTTTGG 2
28S_LSU-900f CCGTCTTGAAACACGGACCAAG ARNr
28S 52°C
3
28S_LSU-1600r AGCGCCATCCATTTTCAGG 2
Sym
bio
tesa
16Suniv-27f AGAGTITGATCMTGGCTCAG rrs 54°C 4 16Suniv-1492r TACGGYTACCRRGTTACGACIT
ITSdegN_1242f GCTACACACGTGCTACAATGG
ITS1
54°C 5 ITSdegN_196r TCTCGGTTGATTTCTTTTCTT
ITS_Symb-282f GGAGTGGGTTGXAAAAGAAG 58°C
ITS_Symb-954r ATCGCAAGCTACAACGTCCT
dnaEnested-1069f CCDGTDGGICCDGGICGYGGC
dnaE
46°C
5
dnaEnested-2279r TGNGATTTRTTRAANCCRTA dnaEdegN-1180f CCIGARCGKGTITCIATGCC 60°C dnaeEdegN-2076r YTGCATNACYTGYTCYTGRTA
dnaE-Symb-201f GAAGATGATCCCCTTCGAGA 64°C
dnaE-Symb-810r GTAGGAGACCTTGGCACGAC
gyrBnested-217f GHAARCGYCCDGGYATGTA
gyrB
56°C
5
gyrBnested-1809r CVACRTCVGCATCBGTCAT gyrBdegN-277f GGTDTTYGARRTDGTDGATAA
56°C gyrBdegN-1590r CHGCNGARTCMCCYTCMAC
gyrB-Symb-185f AGCCACCCGAAGAGAAGAA 58°C
gyrB-Symb-1150r TGGCAGACCGGCTATGT
CBBL-1B CACCTGGACCACVGTBTGG cbbL 60°C 6 CBBL-2C CGGTGYATGTGCAGCAGCATGCCG
aprA-Symb-23f d
GCAACGACCTGATGGGTATC aprA 58°C 5 aprA-Symb-555r
d GTCGGTACGAAGCGGTTTT
a Les amorces « nested » et « degN » correspondent aux amorces dégénérées utilisées en PCR emboitée. Les
amorces « symb » correspondent aux amorces spécifiques dessinées à partir des séquences obtenues. b Les températures d’hybridation optimales ont été obtenues par PCR à gradient c [1] (Winnepenninckx et al., 1998), [2] (Williams et al., 2003), [3] (Olson et al., 2003), [4] , [5] cette étude, [6].
d Ces amorces ont été dessinées en utilisant les séquences spécifiques de symbiotes de Lucinidae obtenues par une approche dérivée du « TE-walking » (Piednoel and Bonnivard, 2009 [Annexe 3]) à partir d’une séquence
« core » obtenue avec les amorces APS-FW/APS-RV (Deplancke et al., 2000).

DIVERSITE BACTERIENNE
82
DOUBLE CARD-FISH
Afin de tester les séquences obtenues, une sonde spécifique du marqueur dnaE
(SymdnaE-HRP414, 5’-GGCCAACGGGGAGGGATTG-3’) a été utilisée en double CARD-
FISH sur des fractions purifiées de symbiotes de C. orbicularis. Les symbiotes ont été purifiés
selon Caro et al. (2007). Une goutte de cette fraction purifiée a été déposée sur lame
Superfrost® (Menzel-Glazer®) et fixée à la chaleur. Les lames ont ensuite été traitées comme
dans le chapitre précédent (Cf. page 38). Une première hybridation a été ensuite réalisée avec
la sonde Sym_dnaE-HRP414 selon Pernthaler et al. (2002) jusqu’au Tampon de lavage. Une
seconde hybridation a ensuite été réalisée avec symco2A, sonde spécifique de l’ARNr 16S de
ces symbiotes (Gros et al., 1996). Ces lames ont ensuite été rincées 2 fois en PBS 1X (15 min
à RT) et incubées dans une solution d’Amplification Buffer (1/10 [
w/v] dextrane sulfate, 1X
PBS, 2M NaCl, 1/100 [
v/v] 10% Blocking reagent), H202 diluée à 0,5% dans du PBS 1X et 2µL
de tyramide marquée fluorescente pendant 15 min à 37°C dans l’obscurité. Les sections ont
ensuite été lavées en PBS 1X (15 min à RT), séchées, contre-colorées au DAPI et montées en
milieu DAKO avant observation avec un microscope fluorescent epi80i (NIKON, France).
RESULTATS
VALIDATION DES MARQUEURS
Variabilité des marqueurs : Dans le but de tester ce critère un indice de variabilité
inter et intra-spécifique des gènes a été évalué. Ce dernier correspond à la divergence
moyenne calculée à partir d’un alignement de 12 γ-protéobactéries (niveau inter-spécifique
[Annexe 2]). L’indice de variabilité intra-spécifique a lui été évalué sur les souches de deux γ-
protéobactéries : Escherichia coli et Legionella pneumophila (Annexe 2). On observe (Fig.
16) que la variabilité des différents marqueurs est comprise entre 30 et 40% alors que celle de
l’ARNr 16S est d’environ 15%. En ce qui concerne le niveau intra-spécifique, on observe
également que le gène de l’ARNr 16S est moins variable que ce soit pour les souches
d’Escherichia coli ou de Legionella pneumophila. Ce gène possède 0,6% de variabilité pour
les souches d’E. coli et à peine 0,1% pour les souches de L. pneumophila (Fig. 17). Dans le
même temps les marqueurs candidats varient d’environ 1% à 1,8% pour E. coli et de 1% à

DIVERSITE BACTERIENNE
83
2,5% pour L. pneumophila (Fig. 17). Tous les candidats semblent de fait correspondre aux
besoins de notre analyse.
Absence de transferts horizontaux : A partir d’un alignement multiple de 28 bactéries
(Annexe 2), une phylogénie moléculaire en Maximum de Vraisemblance a été réalisée pour
chaque marqueur candidat et comparée à la phylogénie moléculaire basée sur l’ARNr 16S
(Fig. 18). Celle-ci permet de définir les groupes bactériens et les relations entre espèces à
l’intérieur de ces groupes et servira donc de référence pour l’analyse des différents marqueurs.
On observe sur les différentes phylogénies (Fig. 19) que tous les grands groupes
monophylétiques définis par la phylogénie de l’ARNr 16S sont retrouvés pour tous les
marqueurs candidats à l’exception de groEL. En effet pour ce gène on observe par exemple
que les cyanobactéries ou les β-protéobactéries sont polyphylétiques. Suite à l’Au-test
(Tableau I, test statistique comparant les topologies de deux phylogénies ; Shimodaira, 2002),
on observe que les topologies de dnaE, gyrB, polA, et recA correspondent de façon
significative à l’ARNr 16S. On retrouve donc au sein de ces marqueurs des signatures qui
semblent être caractéristiques de l’espèce et qui permettent de replacer les espèces
bactériennes les unes par rapport aux autres.
Présence de régions conservées et de régions variables : La nécessité d’avoir des
régions conservées encadrant des régions variable a également été un critère de validation de
nos marqueurs candidats. Ainsi, un alignement de 12 gamma-protéobactéries a été importé
dans le logiciel SVARAP 2.0 afin de visualiser la variabilité des différents candidats le long
de leur séquence. On observe ainsi que chaque candidat possède ces régions conservées, qui
permettront de définir des amorces dégénérées. Ces régions conservées sont encadrées par des
régions variables qui mettront en évidence l’éventuelle diversité génétique recherchée.
Tableau I. Résultats de l’Au-test.
Topologiea ARNr 16S dnaE groEL gyrB polA recA rpoB
Classement Meilleur 5ème 6ème 2ème 4ème 3ème 7ème
Aub 0,866 0,130 0,016 0,273 0,243 0,270 0,001
a topologies comparées à celles de l’ARNr16S b p-values

DIVERSITE BACTERIENNE
84
Figure 18. Indice de variabilité inter-spécifique des différents gènes candidats comparés à celui de l’ARNr 16S.
Au vu des résultats précédents les marqueurs choisis pour l’analyse ont été : la sous
unité alpha de l’ADN polymérase III (dnaE), l’ADN Gyrase B (gyrB), l’ADN polymérase I
(polA) et la Recombinase A (recA).
OBTENTION DES SEQUENCES DES MARQUEURS RETENUS POUR L’ANALYSE
recA : Un fragment obtenu à la taille attendue a été séquencé. Cette séquence
présentait une identité nucléotidique (sur 500 pb) de 98% avec la recombinase A
d’Escherichia coli O157:H7 [TaxID : 83334]. De nouvelles amorces ont donc été dessinées.
La séquence obtenue grâce à ce nouveau couple d’amorces ne présentait aucune identité
nucléotidique avec les séquences des bases de données. Aucune amorce spécifique n’a pu
ainsi être dessinée pour ce marqueur.
polA : Un premier fragment séquencé présentait une identité nucléotidique de 79%
(76pb sur un fragment de 711) avec l’ADN Polymerase I de Bordetella pertussis Tohama I.
Un second jeu d’amorces a permis l’amplification de 800 paires de bases présentant 60 %
d’identité avec l’ADN Polymérase de Acinetobacter sp. ATCC27244 [TaxID : 525244]. Des
amorces spécifiques ont pu ainsi être déterminées à partir de cette séquence.

DIVERSITE BACTERIENNE
85
Figure 19. Indice de variabilité intra-spécifique des différents gènes candidats comparés à celui de l’ARNr 16S
pour différentes souches d’Escherichia coli et de Legionella pneumophila.
dnaE, et gyrB: Concernant ces deux gènes la première amplification PCR a permis
l’obtention de fragments à la taille attendue. Les séquences de ces derniers présentaient une
identité de séquence de 75% avec Thioalkalivibrio sp. HL-EbGR7 [TaxID : 396588] pour
dnaE, et de 85% Allochromatium vinosum DSM180 [572477] pour gyrB. Des amorces
spécifiques ont ainsi pu être déterminées pour ces différents marqueurs
VALIDATION DES SEQUENCES OBTENUES
Afin de s’assurer que les séquences obtenues à partir des branchies de C. orbicularis
correspondaient bien au symbiote de ce dernier nous avons vérifié ces séquences par analyse
phylogénétique et par CARD-FISH. Selon les différentes phylogénies nos séquences
appartiennent bien aux gamma-protéobactéries hormis pour l’ADN polymérase I dont la
séquence amplifiée appartient à un groupe non défini. De plus, les résultats d’hybridation in
situ réalisées sur des fractions purifiées de symbiotes de C. orbicularis avec une sonde
spécifique du marqueur dnaE attestent bien que les symbiotes possèdent la séquence
amplifiée. La vérification de séquence par hybridation in situ n’a pas été réalisée pour les
autres marqueurs candidats.

DIVERSITE BACTERIENNE
86
Figure 20. Phylogénie moléculaire des bactéries basée sur des séquences d’ARN ribosomal 16S (1354 sites).
Cet arbre a été reconstruit par Maximum de Vraisemblance avec comme modèle d’évolution le TrN+I+Г. Les
archées ont été utilisées comme groupe extérieur. La robustesse des différents nœuds a été évaluée par
bootstrapp et seules les valeurs supérieures à 70% sont affichées.
ANALYSE DES SEQUENCES D’ARNR 16S
Toutes les amplifications PCR réalisées à partir des branchies d’A. alba, C.
orbicularis, C. orbiculata, C. pectinella, D. quadrisulcata et L. Pensylvanica ont permis
l’obtention d’une séquence unique d’ARNr 16S après analyse des chromatographes. Ces
séquences présentent une identité nucléotidique moyenne de 99,7% (1466 pb).

DIVERSITE BACTERIENNE
87
Figure 21. Phylogénies moléculaires des bactéries basées sur des séquences des différents marqueurs candidats (dnaE : 1630 sites ; groEL : 1521 ; recA : 906 ; gyrB : 1473 ; polA : 2059 ; rpoB : 2604).
Ces arbres ont été reconstruits par Maximum de Vraisemblance avec comme modèles d’évolution SYM+I+Г
(dnaE, recA et gyrB), K80+I+Г (polA) et GTR+I+Г (groEL et rpoB) Les archées on été utilisées comme groupe
extérieur.

DIVERSITE BACTERIENNE
88
PHYLOGENIE DES HOTES
Pour toutes les amplifications des ARNr 18S et 28S, une amplification unique à la
taille attendue (respectivement ~1000 pb et ~700 pb) a été observée. Les Crassatellidae et les
Thyasiridae qui sont groupes frères des Lucinidae ont été choisis comme groupe extérieur.
Selon cet arbre phylogénétique (Fig. 20), nos échantillons sont retrouvés dans les différents
clades décrits par Williams et al. (2003). Le clade Anodontia pour A. alba, le clade A pour C.
orbicularis et C. orbiculata et le clade B pour C. pectinella, D. quadrisulcata et L.
pensylvanica. Pour A. alba et C. orbicularis les séquences obtenues ici correspondent à celles
publiées par Williams et al. (2003).
ANALYSE DU POLYMORPHISME DES SYMBIOTES.
Un total de 18 séquences par marqueur a été obtenu et le polymorphisme observé est
présenté Tableau J. Après alignement de ces séquences, 6 sites polymorphes sur 465 ont été
observés pour dnaE, 6/806 pour gyrB, 11/512 pour ITS1 8/492 pour aprA et 11/600 pour
CbbL. On note au total environ 1,5% de sites polymorphes. L’ITS1 constitue le marqueur le
plus variable (2,14%) et les gènes de métabolisme aprA et cbbL sont plus variables que les
gènes de ménage dnaE et gyrB (respectivement 1,62%, 1,83%, 1,29% et 0,74%). Selon les
réseaux d’haplotypes (Fig. 21) réalisés pour chacun des cinq marqueurs sur 18 individus (trois
par espèce hôte), différents haplotypes sont observés (entre cinq et six selon le marqueur)
révélant une diversité bactérienne intra-spécifique. De façon intéressante sauf pour ITS1
l’haplotype majoritaire ne semble pas être l’haplotype ancestral à partir duquel les autres
haplotypes seraient dérivés. De plus les différents haplotypes ne semblent pas être distribués
aléatoirement. En effet, il semblerait qu’il y ait un lien entre haplotype bactérien et espèce
hôte (Fig. 21).
Les trois sub-adultes de C. orbicularis (coquilles d’environ 5 mm de long) hébergent
le même haplotype symbiotique que les adultes. Les séquences de dnaE, gyrB et aprA
présentent 100% d’identité avec celles des adultes. Enfin le séquençage de dnaE a également
été réalisé sur la couche supérieure de sédiment comprise entre 0 et 5 cm de profondeur. Le
séquençage de cinq clones montre une séquence correspondant au symbiote de C. orbicularis
(une différence sur 465 pb). Les quatre autres clones semblent correspondre à des séquences
bactériennes différentes de celles hébergées par les lucines colonisant les herbiers à T.
testudinum de l’Îlet Cochon (identité nucléotidique moyenne de 83% avec le symbiote de C.

DIVERSITE BACTERIENNE
89
orbicularis). Enfin le séquençage de dnaE réalisé sur de C. orbicularis et C. orbiculata de
Martinique révèle que ces deux individus hébergent des symbiotes différents de ceux
retrouvés en Guadeloupe (respectivement deux et quatre différences sur 465 pb).
Figure 22. Phylogénie moléculaire des Lucinidae réalisée après concaténation des séquences des ARNs
ribosomaux 18S et 28S (1386 sites).
Cet arbre a été reconstruit par Maximum de Vraisemblance (MV) et Inférence Bayesienne (BI) avec comme
modèles d’évolution le TrNef+Г (ML) et le K80+Г (BI). Les Crassatellidae et Thyasiridae comme groupes
extérieurs. La robustesse des différents nœuds a été évaluée et seules les valeurs de bootstrap (ML, 100
répétitions [en dessous des branches]) et les probabilités postérieures (BI, runs=5, générations=1 000 000,
échantillonnage=1 000, burn=10% [au-dessus des branches]) supérieures à 95% sont affichées. Les séquences
en gras correspondent aux séquences obtenues lors de cette étude et les autres à des séquences tirées de Williams
et al. (2003).

DIVERSITE BACTERIENNE
90
Figure 23. Réseaux d’haplotypes reconstruits à partir des séquences obtenues pour chacun des marqueurs Les réseaux ont été reconstruits par la méthode de Median Joining. Les cercles correspondent aux différents haplotypes (HA à H6).La taille de ceux-ci représentent le nombre de
séquences pour chaque haplotype. La couleur des cercles représente l’espèce hôte hébergeant ces séquences bactériennes. Bleu pour C. orbicularis, vert pour L. pensylvanica,
rouge pour C. pectinella, jaune pour C. orbiculata, violet pour A. alba et noir pour D. quadrisulcata. Les barres sur les branches représentent une substitution.

DIVERSITE BACTERIENNE
91
Tableau J Analyse du polymorphisme.
Gene a
Sequences Sites
Polymorphesb
Haplotypes Nb Taille (pb)
dnaE (~3,5 kb) 18 465 6 (1.29 %) 5
gyrB (~2,4 kb) 18 806 6 (0.74 %) 5
ITS1 (~550 pb) 18 512 11 (2.14 %) 5
aprA (~1 kb) 18 492 8 (1.62 %) 6
cbbL (~1,5 kb) 18 600 11 (1.83 %) 6
TOTAL 90 2875 42 (1.46 %)
a Noms des gènes. dnaE: sous-unité alpha de l’AND Polymérase III, gyrB: AND Gyrase B, ITS1: 16S/23S
Internal Transcribed spacer, aprA: sous-unité alpha de l’Adenyly1,5-bisPhosphate Redctase, cbbL: Grande sous-
unité de la Ribulose-1,5-bisPhosphate Carboxylase/Oxygenase Large subunit. b Les pourcentages représentent la proportion de sites polymorphes par rapport à la taille du fragment considéré.
DISCUSSION
Le but de cette étude était de mettre en évidence une diversité génétique des symbiotes
sulfo-oxydants associés à six espèces de bivalve de la famille des Lucinidae (A. alba, C.
orbicularis, C. orbiculata, C., pectinella, D. quadrisulcata et L. pensylvanica) colonisant les
herbiers à T. testudinum des Antilles Françaises. En effet, ces six espèces hôte hébergent la
même espèce bactérienne sur la base du séquençage de l’ARNr 16S. Considérant ce marqueur
insuffisamment résolutif comme cela a été démontré chez les Siboglinidae (Di Meo et al.,
2000) et les Mytilidae (Won et al., 2003), j’ai cherché cette potentielle diversité chez le
Lucinidae par une approche MLST. De plus via des réseaux d’haplotypes j’ai cherché à savoir
si cette diversité bactérienne intra-spécifique était structurée ou pas en fonction de la diversité
des hôtes, de la localisation géographique ou de composantes environnementales.
CHOIX ET VALIDATION DES MARQUEURS
Les bases de données étant relativement pauvres en séquences de symbiotes de
Lucinidae, la première partie du travail a été de choisir des marqueurs convenant à notre étude
et d’obtenir les séquences de ces marqueurs. Des marqueurs correspondant aux trois
catégories de Cooper & Feil (2004): (i) gènes de ménage, (ii) gènes de « core metabolism » et

DIVERSITE BACTERIENNE
92
(iii) gènes hypervariables ont été choisis. Etant spécifiques du groupe étudié (i.e. les bactéries
sulfo-oxydantes) l’APS Reductase et la RubisCO ont été retenus comme gènes de « core
metabolism ». L’ITS correspond lui au marqueur hypervariable retenu. Concernant les gènes
de ménage, des candidats issus de la littérature ont été choisis et testés afin de vérifier leur
compatibilité avec les besoins de notre étude. Différents critères devaient donc être remplis
par ces candidats pour être ou non retenus : (i) variabilité supérieure à celle de l’ARNr 16S,
(ii) absence de transferts horizontaux et (iii) présence de régions conservées encadrant des
régions variables. Les six gènes retenus à partir de la littérature et ainsi testé ont été : dnaE,
groEL, gyrB, polA, recA et rpoB.
Variabilité supérieure à celle de l’ARNr 16S : D’une façon générale les gènes codants
pour des protéines sont attendus pour être moins variables que l’ARNr 16S ce dernier étant
contraint uniquement au niveau de sa structure alors que les séquences codant des protéines
sont contraintes non seulement pour la structure, un changement d’acide aminé pouvant
conduire à un mauvais repliement, mais aussi pour la fonction de la protéine, un changement
d’acide aminé au niveau d’un site catalytique ou de reconnaissance modifiant la fonction ou
rendant cette dernière non fonctionnelle. Néanmoins selon les indices de variabilité inter et
intra-spécifique déterminés, dnaE, groEL, gyrB, polA, recA et rpoB se sont montrés plus
variables que l’ARNr 16S.
Absence de transferts horizontaux : Ce critère est essentiel afin d’éviter de surestimer
ou sous-estimer la diversité. En effet, un gène soumis à de tels transferts (e.g. facteurs de
virulence) peut être retrouvé dans des bactéries de souches, d’espèces ou de groupes différents
et conduire à une mauvaise estimation de la diversité. Au vu des différents arbres réalisés,
dnaE, gyrB, polA, et recA ne semblent pas soumis à ce type de transferts leur phylogénie
suivant celle de l’ARNr 16S.
Présence de régions conservées et de régions variables : Ce dernier critère relève
plutôt du côté technique de la méthode utilisée ici dans la recherche de séquences. En effet, en
l’absence d’informations sur le génome des symbiotes de Lucinidae des amorces spécifiques
ne peuvent être déterminées. Par conséquent l’amplification par amorces dégénérées grâce à
des séquences conservées chez différentes espèces reste le seul moyen (hors séquençage haut
débit) pour obtenir les séquences de nos marqueurs. Il a été démontré que nos candidats
possédaient des motifs conservés qui correspondaient aux séquences codantes pour différents

DIVERSITE BACTERIENNE
93
sites catalytiques (e.g. liaison à l’ADN, hydrolyse de l’ATP …) des protéines correspondantes
(Bernad et al., 1990; Dalrymple et al., 2001; Karlin and Brocchieri, 1996; Kim et al., 1997).
Entre ces régions il existe des régions moins contraintes n’intervenant ni au niveau structural
ni au niveau fonctionnel, ces régions s’en retrouvent donc beaucoup plus variables.
Ainsi au vu de ces critères il apparait que seuls dnaE, gyrB, polA et recA peuvent être
retenus comme marqueurs de diversité dans le cas qui nous concerne.
OBTENTION ET VERIFICATION DES SEQUENCES DES MARQUEURS RETENUS.
Afin d’obtenir les séquences de nos marqueurs correspondant aux symbiotes sulfo-
oxydants de Lucinidae une stratégie de PCR emboitée avec amorces dégénérées a été utilisée.
Compte tenu de leur caractère dégénéré ces amorces sont sensées amplifier la majorité des
gamma-protéobactéries auxquelles appartiennent les symbiotes de Lucinidae. Ainsi, le risque
d’amplifier une γ-protéobactérie contaminante restait présent. Il pouvait en effet s’agir de
bactéries de l’environnement (e.g. eau de mer, sédiments, air ambiant…) ou du bivalve lui-
même (i.e. bactéries marines libres situées entre les filaments branchiaux). Cependant le
risque principal demeurait Escherichia coli, γ-protéobactérie commensale et couramment
utilisée au laboratoire. De fait la vérification par comparaison dans les banques de données,
analyse phylogénétique et hybridation in situ restait indispensable.
recA : Toutes les amplification réalisées sur ce marqueur (deux jeux d’amorces
différents) ont abouti au séquençage du gène d’Escherichia coli O157:H7. Ce résultat peut
être du à une affinité supérieure des amorces dégénérées pour les séquences d’E. coli par
rapport à la séquence de la recombinase A des symbiotes de Lucinidae.
polA : Les deux amplifications réalisées ont permis l’obtention de fragments
correspondant à polA. La première séquence ne présentait une identité nucléotidique que sur
une très faible taille (76 pb sur 611 séquencées) avec B. pertussis qui est une β-
protéobactérie. Par conséquent cette dernière a été jugée inexploitable. Concernant le second
fragment d’environ 900 pb, une identité de 60% a été retrouvée sur 914 pb. Cependant
l’analyse phylogénétique n’a pu confirmer avec certitude que ce fragment correspondait à une
γ-protéobactérie.
dnaE, et gyrB: Une bande de taille attendue a été obtenue pour toutes les
amplifications de ces marqueurs. Après séquençage, les fragments correspondaient bien au

DIVERSITE BACTERIENNE
94
gène ciblé. Les comparaisons par BLAST avec les bases de données, et les analyses
phylogénétiques ont confirmé qu’il s’agissait de γ-protéobacteries. De plus l’hybridation in
situ réalisée pour le gène dnaE a confirmé la présence de la séquence obtenue au sein des
symbiotes de Lucinidae. Par conséquent ces trois gènes, le marqueur hypervariable ITS1 et les
gènes de métabolisme cbbL et aprA ont été retenus pour l’analyse de la diversité intra-
spécifique des symbiotes associés aux Lucinidae des Antilles Françaises et les séquences de
18 individus ont été obtenues.
L’ANALYSE MLST REVELE UNE DIVERSITE GENETIQUE CHEZ LES SYMBIOTES SULFO-OXYDANTS.
Cinq marqueurs ont été utilisés appartenant aux trois catégories décrites par Cooper &
Feil (2004). Comme attendu ITS1 constitue le marqueur le plus variable (Tableau J).
Cependant ce dernier ne présente pas le plus grand nombre d’haplotypes, eu égard à
l’accumulation des sites polymorphes dans les séquences de symbiotes d’A. alba (Fig. 21).
Les gènes de ménage (dnaE et gyrB) sont les moins variables et les « core-métabolisme »
(aprA et cbbL) présentent une variabilité intermédiaire (Tableau J). Ces résultats
correspondent parfaitement à nos pré-requis (i.e. variabilité des marqueurs supérieure à celle
de l’ARNr 16S). Ainsi, notre analyse permet la visualisation d’une diversité génétique intra-
spécifique au sein des symbiotes hébergés par les six espèces de Lucinidae des Antilles
Françaises.
Le polymorphisme moyen observé est de l’ordre de 1,5% ce qui est cohérent avec les
données de la littérature (Zhang and Hewitt, 2003). Néanmoins, cette valeur peut
correspondre à une sous-estimation de la diversité eu égard à la technique employée ici (i.e.
séquençage direct des produits de PCR). En effet, seule la séquence majoritairement présente
est à chaque fois amplifiée et séquencée. De fait s’il existe au sein de ces espèces de
Lucinidae une multi-infection de souches cette dernière ne sera pas forcément détectée. Ainsi,
l’analyse attentive des différents chromatographes a permis chez un spécimen de l’espèce
Anodontia alba la visualisation de séquences minoritaires qui pourrait bien être due à la
présence de plusieurs souches bactériennes.
Se pose dès lors la question de la distribution de cette diversité. Cette dernière est-elle
répartie de façon aléatoire parmi nos différents échantillons ou structurée en fonction de la
diversité des hôtes, de la localisation géographique des échantillons ou du biotope où sont
retrouvés ces derniers ?

DIVERSITE BACTERIENNE
95
STRUCTURATION DE LA DIVERSITE BACTERIENNE
Dans le but de tester l’éventuelle structuration de la diversité des symbiotes sulfo-
oxydants associés aux lucines caribéennes des réseaux d’haplotypes ont été réalisés avec les
séquences de nos différents marqueurs (Fig. 21). Quelque soit le marqueur considéré, la
diversité bactérienne n’est pas distribuée de façon aléatoire parmi les 18 individus analysés.
De plus même si les réseaux d’haplotypes ne sont pas totalement congruents le même patron
est retrouvé : (i) les symbiotes d’A. alba sont toujours polymorphes. (ii) Les symbiotes de D.
quadrisulcata sont polymorphes ou monomorphes selon le marqueur. (iii) Les symbiotes des
4 autres espèces sont monomorphes. Cette dernière observation atteste d’une spécificité
d’association entre L. pensylvanica et son symbiote, entre C. orbicularis et le sien. De façon
surprenante il semble également exister une spécificité d’association entre une souche
particulière et deux espèces hôte (C. orbiculata et C. pectinella). Cette structuration de la
diversité semble de plus ne pas être due à un biais d’échantillonnage. En effet, la probabilité
d’obtenir au hasard une séquence bactérienne particulière pour 3 hôtes sur 18 échantillonnés,
sous l’hypothèse que la capture du symbiote est réalisée dans un unique pool bactérien est de
4,6E10-3
.
QUELS FACTEURS SONT FACTEURS RESPONSABLES DE LA STRUCTURATION DE LA DIVERSITE ?
Il existe donc chez ces symbiotes sulfo-oxydants une diversité de souches au sein de
l’espèce bactérienne partagée par les six espèces hôte des Antilles Françaises. Cette diversité
n’étant pas distribuée aléatoirement j’ai cherché les facteurs expliquant la structuration de
cette diversité. Dans un premier temps, j’ai donc considéré trois hypothèses correspondant
aux paramètres suivants : (i) la géographie (ii) l’environnement et (iii) la phylogénie des
hôtes.

DIVERSITE BACTERIENNE
96
Figure 24. Représentation schématique de la distribution des hôtes et des symbiotes dans les sédiments des
herbiers à T. testudinum.
(A) Population bactérienne structurée en fonction de la profondeur. (B) Population bactérienne non structurée.
Les cercles représentent les hôtes et les triangles les formes libres des symbiotes. Les couleurs correspondent aux
espèces (hôtes) ou aux souches mises en évidence par l’analyse MLST (symbiotes) : Jaune, C. orbiculata ;
Rouge, C. pectinella ; Violet, A. alba ; Bleu, C. orbicularis ; Bleu clair, Juvéniles ou sub-adultes de C.
orbicularis.
Influence de la géographie: Les 6 espèces hôte analysées provenant de 3 lieux de
collecte différents (La Désirade pour D. quadrisulcata, la Martinique pour L. pensylvanica et
la Guadeloupe pour A. alba, C. orbicularis, C. orbiculata et C. pectinella) la structuration
peut être due à la répartition géographique de nos échantillons. En effet, L. pensylvanica et D.
quadrisulcata ont été échantillonnés chacun dans des zones géographiques différentes et
possèdent chacun un patron d’haplotype symbiotique qui leur est propre (polymorphisme ou
non pour D. quadrisulcata et polymorphisme pour A. alba). De plus, les symbiotes des
espèces hôte échantillonnées en Martinique (C. orbicularis et C. orbiculata) présentent des
séquences différentes des symbiotes de Guadeloupe. Néanmoins, les 4 autres espèces qui ont
été échantillonnées dans le même herbier (Îlet Cochon) ne partagent pas le même haplotype.
De fait la localisation géographique ne peut être considérée comme unique composante
expliquant la structuration de la diversité observée.
Dès lors, comment expliquer la structuration de la diversité des symbiotes hébergés
par les 4 lucines de l’Îlet Cochon ? Deux autres hypothèses (écotype ou phylotype) peuvent
être envisagées.
Hypothèse de l’écotype: Sous cette hypothèse, la population bactérienne est structurée
dans l’environnement en fonction de différents paramètres environnementaux (e.g. gadients
d’H2S, d’O2 ou autre) retrouvés dans les sédiments des herbiers (Fig. 22A). De fait, la
structuration observée chez les hôtes ne serait que le reflet de la structuration de la population
bactérienne dans l’environnement. Par conséquent, des hôtes localisés à différents niveaux de
la colonne de sédiment hébergeraient des souches symbiotiques différentes. Cette hypothèse

DIVERSITE BACTERIENNE
97
est corroborée par l’observation des symbiotes de C. orbiculata et C. pectinella. En effet ces
2 espèces hôte ne sont localisées que dans la couche supérieure de sédiment eu égard à la
taille de leur siphon (Allen, 1958), et partagent quelque soit le marqueur le même haplotype
bactérien. Sous cette hypothèse, seul cet unique haplotype bactérien devrait être retrouvé dans
la couche supérieure du sédiment, dans les juvéniles (ou sub-adultes) ou dans tout Lucinidae
présent entre 0 et 5 cm de profondeur. Afin de valider cette hypothèse, les sédiments de la
couche supérieure et les sub-adultes de C. orbicularis (0,5 cm de longueur) ont été analysés.
Les résultats obtenus ne valident pas cette hypothèse. En effet, une séquence correspondant au
symbiote de C. orbicularis a été détectée dans les sédiments. De plus, les sub-adultes de cette
même espèce qui ont été échantillonnés dans la couche supérieure, hébergent la même souche
que les adultes (selon des séquences des gènes dnaE, gyrB et aprA) qui sont eux distribués
dans toute la colonne de sédiments et jusqu'à 20 cm de profondeur. Il semblerait donc qu’au
niveau de l’Îlet Cochon la population bactérienne environnementale ne soit pas structurée en
fonction de la profondeur (Fig. 22B).
Hypothèse du phylotype: Sous cette hypothèse la structuration observée serait due à la
coévolution des hôtes et des souches bactériennes à partir d’un couple symbiotique ancestral.
Ainsi les hôtes les plus proches phylogénétiquement devraient héberger les souches
bactériennes les plus proches. Afin de tester cette hypothèse, la phylogénie moléculaire des
hôtes (Fig. 20) a donc été comparée à la structuration observée (Fig. 21). On n’observe
aucune congruence entre la phylogénie des hôtes et les différents réseaux d’haplotypes. En
effet les deux hôtes hébergeant la même souche bactérienne (C. orbiculata et C. pectinella
[Fig. 21, cercle rouge et jaune]) ne sont pas phylogénétiquement les plus proches. La première
espèce hôte appartient au clade A des Lucinidae et la seconde au clade B (Fig. 20). Ainsi,
l’hypothèse du phylotype peut ici être rejetée.
Ainsi, dans le cas des six Lucinidae que j’ai étudié ici, la géographie, la diversité des
hôtes ou les conditions environnementales ne permettent pas d’expliquer la structuration de la
diversité qui existe au sein de leurs symbiotes sulfo-oxydants. En effet, même si le paramètre
« géographie » peut être invoqué dans le cas de D. quadrisulcata et de L. pensylvanica), il
semble que d’autres facteurs interviennent lorsque différentes espèces hôtes colonisent une
même localité (même lieu géographique et même biotope).Des lors, il semble que seule une
spécificité d’association hôtes/souches bactériennes due à un mécanisme sélectif puisse
conduire au schéma observé. Ceci sous l’hypothèse de l’accès à la ressource en symbiotes par

DIVERSITE BACTERIENNE
98
les hôtes. Comment envisager qu’un processus sélectif entre les hôtes et les symbiotes
puissent conduire à cette structuration de la diversité?
La figure 23 propose une hypothèse qui peut expliquer la structuration observée. La
première étape (Fig. 23[1]) correspond à l’état initial ou 4 espèces hôte exploitent le même
pool bactérien. [2] Un mutant pour les déterminants de reconnaissance apparait dans la
population bactérienne. [3] Ce dernier n’est plus reconnu par les hôtes. De fait la population
de ce nouveau mutant augmente en cas d’avantage sélectif de la « vie libre » par rapport à la
« vie associée ». Ainsi, sous l’hypothèse du recrutement environnemental, l’échappement à la
capture (et donc à l’impasse évolutive) semble expliquer l’augmentation de la proportion de
ce nouveau mutant dans l’environnement. [4] Apparition d’individus hôte capables de
reconnaitre cette nouvelle forme du symbiote. [5] Ces individus exploitant à la fois l’ancien et
le nouveau pool bactérien leur proportion augmente tout naturellement au sein de la
population hôte eu égard au bénéfice pour l’hôte (Cf chapitre I). [6] Une nouvelle association
peut se constituer si : (i) la taille de la population bactérienne environnementale est
relativement faible. En effet dans ce cas, les hôtes mutants étant les seuls à exploiter ce
nouveau pool bactérien, ces derniers ne sont pas en compétition avec d’autres espèces hôtes
pour la ressource bactérienne. (ii) La nouvelle association confère un avantage sélectif le
nouveau symbiote étant plus éfficace (e.g. fixation du carbone). Ainsi, la spécialisation pour
une souche donnée intervient via le mécanisme de choix de partenaire (Sachs et al., 2004). [7]
Des mutants pour des caractères non impliqués dans la reconnaissance peuvent également
apparaitre et continuent dans ce cas à être capturés par les hôtes. Ce cas particulier explique
de fait la présence de symbiotes polymorphes au sein de certains individus (e.g. cas d’A.
alba). En effet, nos marqueurs ne sont pas impliqués dans la reconnaissance et sont supposés
neutres, la structuration observée révèle une sélection des déterminants de reconnaissance qui
par autostop, sélectionnerait également les allèles de nos marqueurs. Ce schéma, s’il
correspond à la réalité, renforce une fois de plus l’hypothèse du recrutement environnemental
par rapport à la transmission environnementale. En effet, aucune évidence ne permet
d’observer sous l’hypothèse de la transmission les étapes [3] et [6]. Ainsi, sous cette dernière,
la bactérie qui s’échappe lors de l’étape [3] ne donnera pas de descendants sauf si elle est
capable de se multiplier dans l’environnement. Enfin lors de l’étape [6], les hôtes
« réensemençant » l’environnement via le relarguage il est peu probable que l’association
tende à se constituer avec une souche différente.

DIVERSITE BACTERIENNE
99
Figure 25. Représentation schématique du processus évolutif pouvant conduire à la structuration de la diversité
bactérienne observée dans les herbiers à T. testidunum de l’Îlet Cochon.
Hôtes
Symbiotes

DIVERSITE BACTERIENNE
100
Une hypothèse alternative à ce scénario évolutif peut également être proposée pour
expliquer le patron d’association spécifique observé au sein de l’ilet Cochon. Cette dernière
ne fait intervenir que le choix du partenaire et constituerait une réponse de l’hôte à
l’environnement (Fig. 26). Sous cette hypothèse 4 espèces hôtes s’associent à différentes
souches bactériennes présentes dans l’environnement (1). Selon les conditions
environnementales, certaines espèces hôtes étant plus adaptées à certaines souches
bactérienne (ces dernieres ne s’associent durablement qu’à la souche bactérienne qui lui
conférerait un succès reproductif accru (2).
Figure 26. Représentation schématique du choix du partenaire.
Hôtes
Symbiotes

SPECIFICITE D’ASSOCIATION
101
SPECIFICITE D’ASSOCIATION
analyse de la diversité des symbiotes associés aux Lucinidae a permis de mettre
en évidence une diversité interspécifique relativement faible. Cependant de
plus fines analyses ont révélé au sein des lucines colonisant les herbiers à
phanérogames des Antilles Françaises une diversité de souches bactériennes jusque là
insoupçonnée. Cette diversité n’est pas aléatoirement distribuée au sein des hôtes mais plutôt
structurée selon différents paramètres. Ainsi, la diversité génétique intraspécifique observée
au sein de cette espèce bactérienne semble être liée à la répartition géographique des hôtes
(e.g. Martinique, la Désirade). En effet, les souches hébergées diffèrent selon la localisation
de l’espèce hôte qui les héberge. Néanmoins, au niveau local, des hôtes différents colonisant
le même herbier n’abritent pas la même souche bactérienne. Il existe, en effet, au niveau de
l’îlet Cochon des associations spécifiques entre hôtes et souches bactériennes. Cette
spécificité d’association ne semble pas être due à une co-évolution des partenaires à partir
d’associations ancestrales. L’environnement n’influe pas non plus sur la diversité symbiotique
car au sein de ces herbiers la population bactérienne est constituée de différentes souches mais
sans structuration particulière. Dès lors, seul un processus local de co-évolution antagoniste
de type Reine-Rouge basée sur l’hypothèse de « gene-for-gene » semble pouvoir expliquer le
patron de structuration observé ici. On peut émettre l’hypothèse qu’à chaque gène de
reconnaissance de l’hôte correspondrait un gène cible chez le symbiote. L’évolution de ces
déterminant permettant tantôt au symbiote d’échapper à l’hôte, tantôt à ce dernier de capturer
ces nouveaux symbiotes. De fait, les associations spécifiques ainsi constituées devraient
pouvoir se constituer et se défaire au cours du temps. Sous un tel modèle les associations
observées aujourd’hui doivent présenter une spécificité telle qu’un hôte associé à une souche
particulière ne puisse s’associer qu’à (ou préférentiellement à) cette dernière. Il existerait alors
au sein des hôtes des mécanismes de reconnaissance et de sélection de la souche qui leur
seraient propre, et donc un contrôle au niveau intra-spécifique de l’acquisition du symbiote.
L’

SPECIFICITE D’ASSOCIATION
102
L’éventualité d’un contrôle intra-spécifique lors de l’acquisition du symbiote par les
Lucinidae est une hypothèse qui a précédemment été proposée par Gros et al. (2003a). Cette
dernière hypothèse suppose l’existence au sein de l’espèce bactérienne partagée par les six
hôtes des Antilles Française d’une diversité de souches. En effet il pourrait ne pas y avoir de
contrôle intra-spécifique mais simplement inter-spécifique, chaque hôte s’associant à toutes
les souches appartenant à l’espèce bactérienne associée à ces hôtes. Dans ce cas, la mise en
contact artificielle d’individus d’une espèce hôte avec une fraction purifiée de symbiotes
provenant des cinq autres espèces hôte aboutirait dans tous les cas à l’infection des juvéniles
(Fig. 24A). Dans le cas inverse, celui d’un contrôle intra-spécifique il ne devrait y avoir
acquisition du symbiote que si et seulement si les individus d’une espèce hôte donnée sont
mis en contact avec des symbiotes provenant de spécimens de cette même espèce hôte (Fig
24B). Ces auteurs ont donc testé ces hypothèses en réalisant des infections de juvéniles
asymbiotiques (i.e. spécimens obtenus au laboratoire et n’ayant jamais été en contact avec des
symbiotes). L’infection de juvéniles de C. orbicularis par des fractions purifiées de symbiotes
provenant des six espèces hôte citées précédemment (A. alba, C. orbicularis, C. orbiculata, C.
pectinella, D. quadrisulcata et L. pensylvanica) a donc été réalisée. Ces juvéniles ont été mis
au contact des symbiotes purifiés durant 1 mois sans changement d’eau avec inoculation à T0
et T0+15j. Les branchies des différents individus ont ensuite été disséquées et contrôlées en
Microscopie Electronique à Transmission (MET). L’expérience a montré que les juvéniles de
C. orbicularis étaient capables d’acquérir sans distinction les symbiotes des six espèces hôte
testées. Pour ces auteurs ceci suggérerait, chez les Lucinidae, une absence de discrimination
de la bactérie au niveau intra-spécifique, la discrimination n’intervenant qu’au niveau
interspécifique bactérien. Cependant compte tenu des données à la disposition de ces auteurs
(2003) ces résultats pouvaient également suggérer tout simplement une absence de diversité
de souches au sein de l’espèce symbiotique associée à ces six espèces de Lucinidae
L’analyse MLST réalisée au cours de mon travail de thèse a permis de rejeter
l’hypothèse d’une absence de diversité intra-spécifique du symbiote. Il n’y aurait donc pas de
discrimination intra-spécifique lors de l’acquisition du symbiote sur la base des résultats de
Gros et al.(2003a). Cependant, l’analyse MLST a également révélé une structuration entre
souches bactérienne/espèce hôte. C’est pourquoi j’ai donc également réalisé une expérience
d’infections sur des adultes aposymbiotiques (débarrassés de leurs symbiotes après un jeûne
de 9 mois) de C. orbiculata. Ces individus ont été mis en contact avec des fractions purifiées

SPECIFICITE D’ASSOCIATION
103
de symbiotes provenant de C. orbicularis ou C. orbiculata fraichement collectés.
L’expérimentation a été conduite durant 15 jours avec mise en contact tous les deux jours et
analyse des spécimens par hybridation in situ tous les quatre jours. Ainsi, après quatre, huit,
12 ou 15 jours, la présence de symbiotes n’est observée que dans les branchies d’adultes mis
en contact avec des fractions purifiées de symbiotes provenant de C. orbiculata (Fig. 25). Par
ailleurs les analyses moléculaires réalisées sur ces fractions purifiées révèlent uniquement la
présence de l’haplotype détecté chez C. orbiculata par l’analyse MLST. Il semble donc que
seule la souche spécifiquement associée à C. orbiculata (Fig. 21, cercles jaunes) soit capable
d’infecter les adultes de C. orbiculata. Il y aurait donc contrôle intra-spécifique lors de
l’acquisition du symbiote par les adultes.
Comment expliquer l’existence d’un contrôle intra-spécifique pour la reconnaissance
de la souche associée chez les adultes de C. orbiculata alors que ce contrôle semble ne pas
avoir lieu chez les juvéniles de C. orbicularis ? La première hypothèse serait liée à l’espèce
hôte considérée. Ainsi C. orbicularis ne contrôlerait pas au niveau intra-spécifique les
symbiotes qu’elle capture à l’inverse de C. orbiculata. Ceci semble peu vraisemblable car
tous les individus analysés (trois adultes et trois juvéniles) provenant des herbiers de l’Îlet
Cochon hébergeaient le même haplotype bactérien. Cette espèce s’associerait donc elle aussi
de façon spécifique à « sa » souche bactérienne. Une autre hypothèse serait celle d’une
plasticité d’infection différente selon le stade qui serait due à la plasticité du filament
branchial au cours du développement des hôtes et durant les phases d’infection. Sous cette
hypothèse la « capacité d’acquisition » ne serait pas la même entre les juvéniles et les adultes.
Même si tous les Lucinidae analysés à ce jour possèdent dans leurs branchies des
symbiotes sulfo-oxydants, cette association n’est pas physiologiquement obligatoire. En effet,
il est possible d’obtenir des sub-adultes non symbiotiques au laboratoire et la présence du
symbiote n’est pas obligatoire pour la métamorphose (Gros et al., 1997). Cependant, en cas de
non-infection, même si les branchies sont fonctionnelles et possèdent une zone ciliée et une
zone intermédiaire normales comme montré chez C. orbicularis (Gros et al., 1998b), la zone
latérale apparait tronquée, avec des filaments branchiaux courts. Tous les types cellulaires
(i.e. mucocytes cellules à grains et cellules intercalaires) sont présents à l’exception des
bactériocytes. Un quatrième type cellulaire, les cellules indifférenciées, est néanmoins
observé chez ces individus asymbiotiques (Gros et al., 1998b). Ces dernières cellules
correspondent aux précurseurs des bactériocytes. En effet, suite à l’endocytose des symbiotes

SPECIFICITE D’ASSOCIATION
104
au niveau du pôle apical de ces cellules indifférenciées, ces dernières se retrouvent colonisées
et se différencient en bactériocytes.
Aucun bactériocyte en division n’ayant été observé, le suivi du filament branchial
pendant les phases d’infection, de purge ou de réinfection a également été réalisé afin de
comprendre la dynamique des branchies de Lucinidae au cours ces différentes phases (N.
Elisabeth, com. Pers.). Durant les phases de purge, les bactériocytes ne disparaîtraient pas
mais réduiraient simplement de taille. Par ailleurs, durant les phases de réinfection il n’y
aurait pas de production de novo des bactériocytes mais plutôt augmentation du volume
cellulaire des bactériocytes déjà présents.
Figure 27. Hypothèses de contrôle de l’acquisition du symbiote par l’hôte selon (Gros et al., 2003a).
(A) Absence de contrôle intra-spécifique : Toutes les souches de l’espèce bactérienne à laquelle appartiennent les
symbiotes de Lucinidae sont reconnues et acquises par les hôtes. Les juvéniles d’une espèce donnée se
retrouvent infectés quelque soit la fraction purifiée mise en contact. (B) Contrôle intra-spécifique : Seule la
souche spécifiquement associée est reconnue et acquise par une espèce hôte donnée. Ainsi les juvéniles ne sont
infectés que si ils sont en contact avec une fraction purifiée provenant de la même espèce hôte. Au sein du stock
libre les formes représentent les espèces bactériennes et les couleurs les différentes souches. AE1, AE2 et AE3
représentent les adultes des espèces 1, 2 et 3. je1 : juvénile de l’espèce 1. Acquisition « naturelle » à partir de
l’environnement (flèches bleues), Infection croisée « artificielle » en laboratoire (flèches noires).

SPECIFICITE D’ASSOCIATION
105
Aussi le jeune asymbiotique ne possédant pas de bactériocytes sensu stricto mais
uniquement des cellules indifférenciées serait capable de reconnaitre et de s’associer à diverse
souches de symbiotes. Néanmoins, au vu de l’analyse des symbiotes associés aux juvéniles,
que j’ai réalisée, même si cette plasticité d’infection pourrait exister la sélection de la souche
« préférentielle » pourrait également avoir lieu. A l’inverse, chez les adultes, la différenciation
des cellules indifférenciées suite à la primo infection suivant la métamorphose pourrait
aboutir à l’apparition de bactériocytes spécialisés. Ces derniers ne disparaissant pas en cas de
purge, l’hôte ne serait capable de s’associer qu’avec cette souche particulière. Ainsi, dans
cette association, la spécificité des couples hôtes/souches bactériennes existerait, mais une
certaine plasticité des juvéniles permettrait une association plus labile qui pourrait se
constituer avec une nouvelle souche en cas d’absence (ou de disparition) de la souche
préférentiellement associée.
En conclusion, une diversité de souches au sein de l’espèce bactérienne (espèce C)
associée aux Lucinidae des Antilles Françaises a été mise en évidence. De façon remarquable,
une spécificité d’association entre espèces hôte et souches bactériennes a également été
observée. Cette spécificité d’association serait due au contrôle intra-spécifique de la souche
acquise par les hôtes. Cependant il semblerait qu’une certaine plasticité demeure chez les
juvéniles leur permettant de s’associer, si nécessaire, à différentes souches de la bactérie. Ceci
aurait pour conséquence une certaine labilité de l’association qui pourrait se faire et se défaire
en fonction de la présence ou non de la souche bactérienne préférentiellement acquise par
l’espèce hôte.

SPECIFICITE D’ASSOCIATION
106
Figure 28. CARD-FISH réalisé à l’aide de la sonde Symco2A.
(A) Section d’une branchie de C. orbiculata fraichement collecté (contrôle positif). (B) Section d’une branchie
de C. orbiculata après 9 mois de jeûne (contrôle négatif). (C et D) Sections de branchies de C. orbiculata
aposymbiotique après 4 jours (C) ou 8 jours (D) de contact avec une fraction purifiée de symbiotes provenant de C. orbicularis. (E et F) Sections de branchies de C. orbiculata aposymbiotique après 4 jours (E) ou 8 jours (F) de
contact avec une fraction purifiée de symbiotes provenant de C. orbiculata. Echelles=10 µm. Les flèches rouges
représentent les symbiotes marqués.

DDDIIISSSCCCUUUSSSSSSIIIOOONNN
GGGEEENNNEEERRRAAALLLEEE


DISCUSSION GENERALE
109
DISCUSSION GENERALE
a symbiose est considérée comme un facteur essentiel dans l’évolution des espèces
comme en témoigne par exemple les associations entre « invertébrés » marins et
bactéries chimioautotrophes. Au cours de ce travail, j’ai étudié ce type
d’associations à travers le modèle Lucinidae/bactéries sulfo-oxydantes. Ce modèle présente de
nombreux avantages tels que la facilité d’échantillonnage et les manipulations in vivo. Je me
suis donc intéressé au cours de ce travail de thèse à l’étude du mode de transmission du
symbiote, de la nature réelle de la relation symbiotique, de la diversité bactérienne associée et
enfin de la spécificité d’association entre les deux partenaires.
A l’origine l’association Lucinidae/bactéries thioautotrophes a été considérée à l’instar
de toutes les associations chimioautotrophes comme étant de type mutualiste (Stewart et al.,
2005). En effet, l’hôte bénéficie de la bactérie qui d’une part détoxifierait son environnement
(sulfides, méthane, etc.) et d’autre part supporte une grande partie de ses besoins nutritionnels
(Arndt et al., 2001). Ainsi l’hôte tire profit du métabolisme bactérien via les dérivés du cycle
de Calvin-Benson (Cavanaugh et al., 1992; Felbeck et al., 1981) et dégrade la bactérie via la
voie lysosomale (Liberge et al., 2001). De l’autre côté, l’hôte conférerait à son symbiote un
environnement protecteur et lui permettrait un meilleur accès aux ressources métaboliques
(CO2 et H2S). En l’absence de transmission verticale chez les Lucinidae, la persistance de
l’association à travers les générations, a été proposé via la transmission environnementale du
symbiote (Gros et al., 1996). En opposition à la règle généralement observée entre mode de
transmission du symbiote et nature de l’association (i.e. transmission verticale pour les
symbiotes mutualistes, transmission horizontale pour les parasites), le modèle
Lucinidae/bactérie sulfo-oxydante constituerait, parmi d’autres, une exception avec une
relation mutualiste sans transmission verticale du symbiote mais avec une transmission dite
environnementale.
Cependant l’hypothèse du couple, mutualisme/transmission environnementale, pour le
modèle Lucinidae ne me parait pas être la plus pertinente. En effet, même si la forme libre du
symbiote est présente dans l’environnement il n’y a pas multiplication des bactéries
symbiotiques dans les bactériocytes de l’hôte (Caro et al., 2007). J’ai pu montrer qu’il n’y
L

DISCUSSION GENERALE
110
aurait pas relarguage de la bactérie une fois celle-ci internalisée. Il semblerait donc qu’il n’y
aurait pas transmission du symbiote à proprement parler chez les Lucinidae. Par conséquent,
entrer en symbiose constituerait une impasse évolutive pour la bactérie, le succès reproducteur
de la bactérie internalisée étant égal à zéro. Ces considérations ne supportent donc pas
l’hypothèse mutualiste, le bénéfice de l’association (évalué au final par l’effet de l’association
sur le succès reproducteur des deux partenaires) étant unilatéral au seul avantage de l’hôte
Lucinidae. Cette association constituerait ainsi un cas tout à fait original de symbiose et
s’inscrirait donc dans un cadre opposé à celui initialement proposé. A l’idée d’une association
mutualiste à transmission environnementale se substituerait l’hypothèse d’une symbiose sans
transmission du symbiote et apparemment non mutualiste.
Dans la littérature d’autres modèles symbiotiques considérés comme mutualistes mais
sans transmission verticale du symbiote ont déjà été décrits. Tel est le cas de l’association
entre Légumineuses et Rhizobia (bactéries des genres Rhizobium, Bradyrhizobium,
Sinorhizobium, etc.). Dans cette association, la bactérie infectant la plante induit la formation
de structures spécialisées, les nodules, où ont lieu les échanges hôte/symbiotes (Oldroyd and
Downie, 2008; Soto et al., 2009). Les bénéfices étant réciproques, l’association serait donc
mutualiste. En effet le symbiote fixe l’azote atmosphérique et le transforme en ammonium qui
est directement assimilable par la plante (Oldroyd and Downie, 2008). De l’autre côté, la
plante fournit au symbiote des sucres et de l’énergie issus de la photosynthèse et nécessaires à
son métabolisme (Bethlenfalvay et al., 1978; Denison, 2000). Le symbiote peut de fait, dans
ce modèle, se multiplier et dans certains cas être relargué dans l’environnement. Autre cas
typique, l’association entre le céphalopode Euprymna scolopes et la bactérie vibrio fischeri.
Dans cette association les jeunes sont également non symbiotiques et acquièrent leurs
bactéries à partir de l’environnement (Ruby and Lee, 1998). La bactérie est localisée au
niveau des tissus de l’hôte (organe bioluminescent) où elle se multiplie avant d’être relarguée.
Par contre, le devenir des « bacterioides » (forme de la bactérie après différenciation dans les
nodules dans le cas des légumineuses) ou des symbiotes du calmar après relarguage n’étant
pas connu (i.e sont-ils perdus, se multiplient-ils dans l’environnement et/ou sont ils réacquis?)
il est de difficile de conclure quant à la réelle transmission du symbiote. On peut dès lors
également poser pour ces modèles l’hypothèse d’une absence de transmission du symbiote.
Ceci pose donc comme pour les Lucinidae la question de la réelle nature de ces associations
symbiotique.

DISCUSSION GENERALE
111
NATURE DE LA RELATION
Dans ces modèles, même s’ils sont considérés comme mutualistes, certains
mécanismes développés par les hôtes laissent à penser que la relation pourrait être unilatérale
au seul bénéfice de ces derniers. En effet, chez les légumineuses, l’évolution prédit
l’apparition de bactéries continuant à utiliser les produits de la photosynthèse mais fixant de
moins en moins d’azote. Afin d’éviter ce parasitisme des réponses de la plante qui consistent
à priver d’oxygène et d’énergie les nodules où les bactéries fixent le moins l’azote appelés
« sanctions » ont été mises en évidence (Denarié et al., 1976; Denison and Kiers, 2004). Ainsi
seules les bactéries ayant un pouvoir de fixation de l’azote important peuvent se multiplier et
être relarguées dans l’environnement. De fait, bien qu’engagée sur la voie du parasitisme,
cette symbiose semble dès lors être maintenue au seul bénéfice de l’hôte. Le même patron
d’unilatéralité est retrouvé chez le calmar. En effet, il semblerait que multiplication et
relarguage du symbiote soient intrinsèquement lié à la relation qu’entretien le calmar avec la
bactérie. Ce dernier tire profit des capacités bioluminescentes de son symbiote, celui-ci
servant à la contre-illumination1 (Jones and Nishiguchi, 2004). Or pour qu’il y ait production
de lumière une densité seuil de bactéries doit être atteinte, engendrant par quorum-sensing
l’activation de l’opéron lux (Callahan and Dunlap, 2000). Ainsi, les symbiotes acquis se
multiplient dans la journée, et émettent de la lumière durant la nuit. La partie des symbiotes
relargués par la suite permettant de diminuer la charge bactérienne afin de relancer le cycle. Il
y a donc ici une nouvelle fois une relation qui semble être au seul bénéfice d’un des deux
partenaires (hôte) bien que le symbiote puisse se multiplier et être relarguée. De fait, dans ces
modèles où la relation semble unilatérale, l’hypothèse du mutualisme ne peut être proposée
que si la valeur selective des symbiotes relargués est différente de zéro.
Chez les Lucinidae, le symbiote est acquis à partir de l’environnement de façon
continue sans multiplication dans les bactériocytes ni relarguage. Je considère que cette
relation ne peut être qualifiée de mutualiste car comme dans les modèles précédents le
bénéfice semble unilatéral (hôte) aux dépends de la bactérie. De plus chez ces bivalves le
succès reproducteur des symbiotes est nul car entrer en symbiose constitue pour la bactérie
1 Ce comportement est une technique de camouflage qui consiste à réduire sa silhouette en adoptant l’intensité et
la longueur d’onde d’une autre source de lumière.

DISCUSSION GENERALE
112
une impasse évolutive. Un tel profil s’apparente à du parasitisme. Peut-on dès lors qualifier
les Lucinidae de parasites ?
UNE NOUVELLE VISION DE L’ASSOCIATION
LUCINIDAE/BACTERIES SULFO-OXYDANTES.
Selon nos résultats, la spécificité d’association observée serait due à une évolution
antagoniste des deux partenaires. De plus, il semblerait que les juvéniles aient conservé une
certaine plasticité d’association. Ces deux caractéristiques (i.e. co-évolution antagoniste et
plasticité) sont couramment observées dans le cas des relations de type parasitaire. En effet, la
course aux armements constitue un mécanisme central dans l’évolution des couples
parasites/parasités. Le parasité évolue afin de faire tendre vers zéro le coût engendré par le
parasite (ici par évitement de la reconnaissance via l’évolution des cibles correspondantes)
alors que ce dernier tend à évoluer pour continuer à tirer bénéfice de l’association (ici par
évolution des déterminants permettant la reconnaissance de la bactérie). De plus, certains
parasites ont conservé (ou acquis) une plasticité d’infection leur permettant de « sauter »
d’une espèce partenaire à une autre afin de pouvoir élargir leur « champ d’action ». Toutes ces
observations supporteraient donc l’hypothèse du Lucinidae parasitant la bactérie sulfo-
oxydante. Cependant, au sein des relations de type parasitaire l’association au parasité est
quasi-obligatoire pour le parasite (Combes, 2003). Or, cette dernière caractéristique n’est pas
observée chez les Lucinidae bien que la prévalence de l’association soit de 100% dans la
nature. De fait, même si les Lucinidae semblent correspondre à des parasites, cette hypothèse
ne peut être totalement validée.
D’autres types d’interactions vraisemblablement à bénéfice unilatéral ont également
été décrits. Dans ces dernières l’hôte acquiert des capacités biochimiques en « s’associant » à
d’autres organismes. Ces associations sont la plupart du temps limitées dans le temps et
continuellement renouvelées. C’est l’exemple des dinoflagellés hétérotrophes Gymnodinium
acidotum qui séquestrent l’algue cryptophyte Chromonas spp. et lui « volent » ses
chloroplastes (qui demeurent fonctionnels) acquérant par la même une capacité
photosynthétique (Fields and Rhodes, 1991). Les chloroplastes demeurent actifs quelques

DISCUSSION GENERALE
113
temps puis sont dégradés par l’hôte et remplacés. Cette acquisition d’organelles à partir d’un
autre organisme a été nommée kleptoplastie1. Cette dernière est d’ailleurs supposée avoir été
une étape intermédiaire clé dans la domestication des mitochondries par la cellule eucaryote
(Cavalier-Smith, 1983).
Un tel phénomène a également été décrit chez le cilié Myrionecta rubra associé à une
autre algue cryptophyte Geminigera cryophila. Dans ce cas, en plus de la kleptoplastie
intervient la karyokleptie2 (Johnson et al., 2007). En effet, le noyau des algues est cette fois
retenu et demeure transcriptionnellement actif, permettant ainsi au cilié de contrôler les
fonctions essentielles (réplication, cytocinèse) des chloroplastes volés. De fait, l’utilisation de
ces derniers peut être plus longue que dans les cas ou intervient seulement la kleptoplastie.
Ces différentes stratégies permettent l’acquisition d’un potentiel biochimique
inexistant à partir d’un autre organisme. Les Lucinidae qui sont hétérotrophes utilisent comme
support majoritaire de leur biomasse les symbiotes sulfo-oxydants. Ceci leur confère une
capacité chimioautotrophique qu’ils ne possèdent pas naturellement. Il s’agirait donc ici
également de l’utilisation à des fins biochimiques (ici métaboliques) d’un organisme différent.
Ainsi, au vu de toutes ces observations que dire de l’association Lucinidae/bactéries
sulfo-oxydantes initialement présentée comme mutualiste à transmission environnementale.
Cette association constituerait donc une symbiose sans transmission du symbiote. Elle
constituerait un cas original dans le monde des associations chimioautotrophes. L’hôte
exploiterait son symbiote en profitant de ses capacités métaboliques. Le maintien d’une
association stable de type parasitaire serait réalisé via un mécanisme de kleptoplastie.
L’association serait donc durable avec renouvellement continu des symbiotes qui
constitueraient des pseudo-organelles permettant à l’hôte hétérotrophe de devenir autotrophe.
Au sein des associations entre « invertébrés » marins et bactéries chimioautotrophes,
certains modèles présentent de fortes similitudes avec celui des Lucinidae. En effet,
l’acquisition du symbiote à partir de l’environnement a également été décrite chez les
Mytilidae (Won et al., 2003) ou les vestimentifères (Harmer et al., 2008; Nussbaumer et al.,
1 Du grec klépto (vol)
2 Du grec karyon (noyau) et kleptie (vol)

DISCUSSION GENERALE
114
2006). De fait, ce modèle pourrait peut être s’appliquer à ces autres relations
chimiosynthétiques.

BBBIIIBBBLLLIIIOOOGGGRRRAAAPPPHHHIIIEEE


BIBLIOGRAPHIE
117
BIBLIOGRAPHIE
Allen JA (1958). On the Basic Form and Adaptations to Habitat in the Lucinacea
(Eulamellibranchia). Philosophical Transactions of the Royal Society of London.
Series B, Biological Sciences 241: 421-484.
Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, Stahl DA (1990). Combination
of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing
mixed microbial populations. Appl Environ Microbiol 56: 1919-25.
Anderson CM, Haygood MG (2007). Alpha-proteobacterial symbionts of marine bryozoans in
the genus Watersipora. Appl Environ Microbiol 73: 303-11.
Ardell DH, Lozupone CA, Landweber LF (2003). Polymorphism, recombination and
alternative unscrambling in the DNA polymerase alpha gene of the ciliate Stylonychia
lemnae (Alveolata; class Spirotrichea). Genetics 165: 1761-77.
Arndt C, Gaill F, Felbeck H (2001). Anaerobic sulfur metabolism in thiotrophic symbioses. J
Exp Biol 204: 741-50.
Ball AD, Purdy KJ, Glover EA, Taylor JD (2009). Ctenidial structure and three bacterial
symbiont morphotypes in Anodontia (Euanodontia) ovum (REEVE, 1850) from the
great barrier reef, AUSTRALIA (Bivalvia: Lucinidae). J. Mollus. Stud. (in press.):
eyp009.
Bandelt HJ, Forster P, Rohl A (1999). Median-joining networks for inferring intraspecific
phylogenies. Mol Biol Evol 16: 37-48.
Bartoli P (1978). Modification de la croissance et du comportement de Venerupis aurea
parasité par Gymnophallus fossarum (Trematoda). Haliotis 7: 23-28.
Berg CJJ, Alatalo P (1984). Potential of chemosynthesis in molluscan mariculture.
Aquaculture 39: 165-179.
Beringer JE, Brewin N, Johnston AW, Schulman HM, Hopwood DA (1979). The Rhizobium-
-legume symbiosis. Proc R Soc Lond B Biol Sci 204: 219-33.
Bernad A, Lãzaro JM, Salas M, Blanco L (1990). The highly conserved amino acid sequence
motif Tyr-Gly-Asp-Thr-Asp-Ser in alpha-like DNA polymerases is required by phage
phi 29 DNA polymerase for protein-primed initiation and polymerization.
Proceedings of the National Academy of Sciences of the United States of America 87:
4610-4614.
Bethlenfalvay GJ, Abu-Shakra SS, Phillips DA (1978). Interdependence of Nitrogen Nutrition
and Photosynthesis in Pisum sativum L: II. Host Plant Response to Nitrogen Fixation
by Rhizobium Strains. Plant Physiol. 62: 131-133.

BIBLIOGRAPHIE
118
Blaxter M (2007). Symbiont genes in host genomes: fragments with a future? Cell Host
Microbe 2: 211-3.
Blazejak A, Kuever J, Erseus C, Amann R, Dubilier N (2006). Phylogeny of 16S rRNA,
ribulose 1,5-bisphosphate carboxylase/oxygenase, and adenosine 5'-phosphosulfate
reductase genes from gamma- and alphaproteobacterial symbionts in gutless marine
worms (oligochaeta) from Bermuda and the Bahamas. Appl Environ Microbiol 72:
5527-36.
Boettcher KJ, Ruby EG (1990). Depressed light emission by symbiotic Vibrio fischeri of the
sepiolid squid Euprymna scolopes. J Bacteriol 172: 3701-6.
Boss KJ (1970). Fimbria and its lucinoid affinities (Mollusca; Bivalvia). Breviora 350: 1-16.
Buchner P (1965). Endosymbiosis of Animals with Plant Microorganisms. Interscience: New
York.
Byun R, Elbourne LD, Lan R, Reeves PR (1999). Evolutionary relationships of pathogenic
clones of Vibrio cholerae by sequence analysis of four housekeeping genes. Infect
Immun 67: 1116-24.
Callahan SM, Dunlap PV (2000). LuxR- and acyl-homoserine-lactone-controlled non-lux
genes define a quorum-sensing regulon in Vibrio fischeri. Journal of Bacteriology
182: 2811-2822.
Caro A, Got P, Bouvy M, Troussellier M, Gros O (2009). Long term starvation of a host
bivalve (Codakia orbicularis, Lucinidae) and effects on its symbiont population. Appl
Environ Microbiol.
Caro A, Gros O, Got P, De Wit R, Troussellier M (2007). Characterization of the population
of the sulfur-oxidizing symbiont of Codakia orbicularis (Bivalvia, Lucinidae) by
single-cell analyses. Appl Environ Microbiol 73: 2101-9.
Cary SC (1994). Vertical transmission of a chemoautotrophic symbiont in the protobranch
bivalve, Solemya reidi. Mol Mar Biol Biotechnol 3: 121-30.
Cary SC, Giovannoni SJ (1993). Transovarial inheritance of endosymbiotic bacteria in clams
inhabiting deep-sea hydrothermal vents and cold seeps. Proc Natl Acad Sci USA 90:
5695-9.
Cary SC, Vetter RD, Felbeck H (1989). Habitat characterization and nutritional strategies of
the endosymbiotn-bearing bivalve Lucinoma aequizonata. Mar Ecol Prog Ser 55: 31-
45.
Casiraghi M, Bordenstein SR, Baldo L, Lo N, Beninati T, Wernegreen JJ et al (2005).
Phylogeny of Wolbachia pipientis based on gltA, groEL and ftsZ gene sequences:
clustering of arthropod and nematode symbionts in the F supergroup, and evidence for
further diversity in the Wolbachia tree. Microbiology 151: 4015-22.

BIBLIOGRAPHIE
119
Castresana J (2000). Selection of conserved blocks from multiple alignments for their use in
phylogenetic analysis. Mol Biol Evol 17: 540-52.
Cavalier-Smith T (1983). Endosymbiotic origin of the mitochondrial envelope. In
Endocytobiology II. W. Schwemmler & H.E.A. Schendt: Berlin: de Gruyter. pp 265-
279.
Cavanaugh CM, Wirsen CO, Jannasch HW (1992). Evidence for Methylotrophic Symbionts
in a Hydrothermal Vent Mussel (Bivalvia: Mytilidae) from the Mid-Atlantic Ridge.
Appl Environ Microbiol 58: 3799-3803.
Chen SC, Liu HW, Lee KT, Yamakawa T (2007). High-efficiency Agrobacterium
rhizogenes-mediated transformation of heat inducible sHSP18.2-GUS in Nicotiana
tabacum. Plant Cell Rep 26: 29-37.
Childress JJ, Fisher CR, Brooks JM, Kennicutt MC, 2nd, Bidigare R, Anderson AE (1986). A
Methanotrophic Marine Molluscan (Bivalvia, Mytilidae) Symbiosis: Mussels Fueled
by Gas. Science 233: 1306-1308.
Cole RA, Lindsay DS, Blagburn BL, Sorjonen DC, Dubey JP (1995). Vertical transmission of
Neospora caninum in dogs. J Parasitol 81: 208-11.
Colson P, Tamalet C, Raoult D (2006). SVARAP and aSVARAP: simple tools for
quantitative analysis of nucleotide and amino acid variability and primer selection for
clinical microbiology. BMC Microbiol 6: 21.
Combes C (2003). L'art d'être parasite: Les associations du vivant. Flammarion: Champs,
N°545.
Cooper JE, Feil EJ (2004). Multilocus sequence typing--what is resolved? Trends Microbiol
12: 373-7.
Daims H, Bruhl A, Amann R, Schleifer KH, Wagner M (1999). The domain-specific probe
EUB338 is insufficient for the detection of all Bacteria: development and evaluation
of a more comprehensive probe set. Syst Appl Microbiol 22: 434-44.
Dalrymple BP, Kongsuwan K, Wijffels G, Dixon NE, Jennings PA (2001). A universal
protein-protein interaction motif in the eubacterial DNA replication and repair
systems. Proc Natl Acad Sci U S A 98: 11627-32.
Dando PR, Southward AJ, Southward EC, Terwilliger NB, Terwilliger RC (1985). Sulfur-
oxidizing bacteria and hemoglobin in gills of the bivalve mollusk Myrtea spinifera.
Marine Ecology-Progress Series 23: 85-98.
Dawkins R (1982). The extended phenotype. Oxford University Press: Oxford, England.
Delmotte F, Rispe C, Schaber J, Silva FJ, Moya A (2006). Tempo and mode of early gene
loss in endosymbiotic bacteria from insects. BMC Evol Biol 6: 56.

BIBLIOGRAPHIE
120
Deming JW, Reysenbach AL, Macko SA, Smith CR (1997). Evidence for the microbial basis
of a chemoautotrophic invertebrate community at a whale fall on the deep seafloor:
bone-colonizing bacteria and invertebrate endosymbionts. Microsc Res Tech 37: 162-
70.
Denarié J, Truchet G, Bergeron B (1976). Effects of some mutations on symbiotic properties
of Rhizobium. In: PS. N (ed). Symbiotic nitrogen fixation in plants. Cambridge
University Press: Cambridge. pp 47-63.
Denison RF (2000). Legume Sanctions and the Evolution of Symbiotic Cooperation by
Rhizobia. The American Naturalist 156: 567-576.
Denison RF, Kiers ET (2004). Why are most rhizobia beneficial to their plant hosts, rather
than parasitic? Microbes and Infection 6: 1235-1239.
Deplancke B, Hristova KR, Oakley HA, McCracken VJ, Aminov R, Mackie RI et al (2000).
Molecular ecological analysis of the succession and diversity of sulfate-reducing
bacteria in the mouse gastrointestinal tract. Appl Environ Microbiol 66: 2166-74.
Di Meo CA, Wilbur AE, Holben WE, Feldman RA, Vrijenhoek RC, Cary SC (2000). Genetic
variation among endosymbionts of widely distributed vestimentiferan tubeworms.
Appl Environ Microbiol 66: 651-8.
Distel DL, Felbeck H (1988). Pathways of inorganic carbon fixation in the endosymbiont-
bearing lucinid clam Lucinoma aequizonata. Part 1. Purification and characterization
of the endosymbiotic bacteria. J Exp Zool 247: 1-10.
Distel DL, Felbeck H, Cavanaugh CM (1994). Evidence for phylogenetic congruence among
sulfur-oxidizing chemoautotropihc becterial endosymbiont and their Bivalve hosts. J
Mol Evol 38: 533-542.
Distel DL, Lane DJ, Olsen GJ, Giovannoni SJ, Pace B, Pace NR et al (1988). Sulfur-
oxidizing bacterial endosymbionts: analysis of phylogeny and specificity by 16S
rRNA sequences. J Bacteriol 170: 2506-10.
Dubilier N, Bergin C, Lott C (2008). Symbiotic diversity in marine animals: the art of
harnessing chemosynthesis. Nat Rev Microbiol 6: 725-40.
Dubilier N, Giere O, Distel DL, Cavanaugh CM (1995). Characterization of
chemoautotrophic bacterial symbionts in a gutless marine worm (Oligochaeta:
Annelida) by phylogenetic 16S rRNA sequence analysis and in situ hybridization.
Appl Environ Microbiol 61: 2346-50.
Dubinsky Z, Stambler N, Benzion M, Mccloskey LR, Muscatine L, Falkowski PG (1990).
The Effect of External Nutrient Resources on the Optical-Properties and
Photosynthetic Efficiency of Stylophora-Pistillata. Proceedings of the Royal Society of
London Series B-Biological Sciences 239: 231-&.

BIBLIOGRAPHIE
121
Duperron S, Fiala-Medioni A, Caprais JC, Olu K, Sibuet M (2007). Evidence for
chemoautotrophic symbiosis in a Mediterranean cold seep clam (Bivalvia: Lucinidae):
comparative sequence analysis of bacterial 16S rRNA, APS reductase and RubisCO
genes. FEMS Microbiol Ecol 59: 64-70.
Duperron S, Halary S, Lorion J, Sibuet M, Gaill F (2008a). Unexpected co-occurrence of six
bacterial symbionts in the gills of the cold seep mussel Idas sp. (Bivalvia: Mytilidae).
Environ Microbiol 10: 433-45.
Duperron S, Laurent MC, Gaill F, Gros O (2008b). Sulphur-oxidizing extracellular bacteria in
the gills of Mytilidae associated with wood falls. FEMS Microbiol Ecol 63: 338-49.
Duperron S, Nadalig T, Caprais JC, Sibuet M, Fiala-Medioni A, Amann R et al (2005). Dual
symbiosis in a Bathymodiolus sp. mussel from a methane seep on the Gabon
continental margin (Southeast Atlantic): 16S rRNA phylogeny and distribution of the
symbionts in gills. Appl Environ Microbiol 71: 1694-700.
Durand P, Gros O (1996). Bacterial host specificity of Lucinacea endosymbionts:
interspecific variation in 16S rRNA sequences. FEMS Microbiol Lett 140: 193-8.
Durand P, Gros O, Frenkiel L, Prieur D (1996). Phylogenetic characterization of sulfur-
oxidizing bacterial endosymbionts in three tropical Lucinidae by 16S rDNA sequence
analysis. Mol Mar Biol Biotechnol 5: 37-42.
Ewald PW (1987). Transmission modes and evolution of the parasitism-mutualism
continuum. Ann N Y Acad Sci 503: 295-306.
Felbeck H, Childress JJ, Somero GN (1981). Calvin-Benson cycle and sulphide oxidation
enzymes in animals from sulphide-rich habitats. Nature 293: 291-3.
Feldman RA, Black MB, Cary CS, Lutz RA, Vrijenhoek RC (1997). Molecular phylogenetics
of bacterial endosymbionts and their vestimentiferan hosts. Mol Mar Biol Biotechnol
6: 268-77.
Felsenstein J (1985). Confidence limits on phylogenies: an approach using the bootstrap.
Evolution 39: 783-791.
Fields SD, Rhodes RG (1991). Ingestion and retention of Chroomonas spp. (Cryptophyceae)
by Gymnodinium acidotum (Dinophyceae). Journal of Phycology 27: 525-529.
Fisher CR (1990). Chemoautotrophic and methanotrophic symbioses in marine invertebrates.
Reviews in Aquatic Sciences 2: 399-436.
Fisher CR, Brooks JM, Vodenichar JS, Zande JM, Childress JJ, Burke RA (1993). The
cooccurrence of methanotrophic and chemoautotrophic sulfur-oxidizing bacterial
symbionts in a deep-sea mussel. Marine Ecology-Pubblicazioni Della Stazione
Zoologica Di Napoli I 14: 277-289.

BIBLIOGRAPHIE
122
Frenkiel L, Gros O, Mouëza M (1996). Gill structure in Lucina pectinata (Bivalvia:
Lucinidae) with reference to hemoglobin in bivalves with symbiotic sulphur-oxidizing
bacteria. Mar Biol 125: 511-524.
Frenkiel L, Mouëza M (1995). Gill ultrastructure and symbiotic bacteria in Codakia
orbicularis (Bivalvia: Lucinidae). Zoomorphology 115: 51-61.
Gavira JA, Camara-Artigas A, De Jesus-Bonilla W, Lopez-Garriga J, Lewis A, Pietri R et al
(2008). Structure and ligand selection of hemoglobin II from Lucina pectinata. J Biol
Chem 283: 9414-23.
Gil R, Latorre A, Moya A (2004). Bacterial endosymbionts of insects: insights from
comparative genomics. Environ Microbiol 6: 1109-22.
Giorgini M, Monti MM, Caprio E, Stouthamer R, Hunter MS (2009). Feminization and the
collapse of haplodiploidy in an asexual parasitoid wasp harboring the bacterial
symbiont Cardinium. Heredity 102: 365-71.
Gomez-Valero L, Silva FJ, Christophe Simon J, Latorre A (2007). Genome reduction of the
aphid endosymbiont Buchnera aphidicola in a recent evolutionary time scale. Gene
389: 87-95.
Gotoh T, Noda H, Ito S (2007). Cardinium symbionts cause cytoplasmic incompatibility in
spider mites. Heredity 98: 13-20.
Groot TV, Breeuwer JA (2006). Cardinium symbionts induce haploid thelytoky in most
clones of three closely related Brevipalpus species. Exp Appl Acarol 39: 257-71.
Gros O, Darrasse A, Durand P, Frenkiel L, Mouëza M (1996). Environmental transmission of
a sulfur-oxidizing bacterial gill endosymbiont in the tropical lucinid bivalve Codakia
orbicularis. Appl Environ Microbiol 62: 2324-30.
Gros O, De Wulf-Durand P, Frenkiel L, Mouëza M (1998a). Putative environmental
transmission of sulfur-oxidizing bacterial symbionts in tropical lucinid bivalves
inhabiting various environments. FEMS Microbiol Lett 160: 257-262.
Gros O, Duplessis MR, Felbeck H (1999). Embryonic Development and endosymbiont
transmission mode in the symbiotic clam Lucinoma aequizonata (Bivalvia:Lucinidae).
Invert Reprod Dev 36: 93-103.
Gros O, Frenkiel L, Felbeck H (2000). Sulfur-oxidizing endosymbiosis in Divaricella
quadrisulcata (Bivalvia: Lucinidae): Morphological, ultrastructural, and phylogenetic
analysis. Symbiosis 29: 293-317.
Gros O, Frenkiel L, Moueza M (1997). Embryonic, larval, and post-larval development in the
symbiotic clam Codakia orbicularis (Bivalvia: Lucinidae). Invertebrate Biology 116:
86-101.

BIBLIOGRAPHIE
123
Gros O, Frenkiel L, Moueza M (1998b). Gill filament differentiation and experimental
colonization by symbiotic bacteria in aposymbiotic juveniles of Codakia orbicularis
(Bivalvia : Lucinidae). Invertebrate Reproduction & Development 34: 219-231.
Gros O, Gaill F (2007). Extracellular bacterial association in gills of "wood mussels". Cah
Biol Mar 48: 103-109.
Gros O, Guibert J, Gaill F (2007). Gill-symbiosis in mytilidae associated with wood fall
environments. Zoomorphology 126: 163-172.
Gros O, Liberge M, Felbeck H (2003a). Interspecific infection of aposymbiotic juveniles of
Codakia orbicularis by various tropical lucinid gill-endosymbionts. Mar Biol 142: 57-
66.
Gros O, Liberge M, Heddi A, Khatchadourian C, Felbeck H (2003b). Detection of the free-
living forms of sulfide-oxidizing gill endosymbionts in the lucinid habitat (Thalassia
testudinum environment). Appl Environ Microbiol 69: 6264-7.
Guindon S, Gascuel O (2003). A simple, fast, and accurate algorithm to estimate large
phylogenies by maximum likelihood. Syst Biol 52: 696-704.
Harmer TL, Rotjan RD, Nussbaumer AD, Bright M, Ng AW, DeChaine EG et al (2008).
Free-living tube worm endosymbionts found at deep-sea vents. Appl Environ
Microbiol 74: 3895-8.
Hoffmann AA, Turelli M, Simmons GM (1986). Unidirectional Incompatibility between
Populations of Drosophila simulans. Evolution 40: 692-701.
Holmes DE, Nevin KP, Lovley DR (2004). Comparison of 16S rRNA, nifD, recA, gyrB,
rpoB and fusA genes within the family Geobacteraceae fam. nov. Int J Syst Evol
Microbiol 54: 1591-9.
Hurst GD, Jiggins FM (2000). Male-killing bacteria in insects: mechanisms, incidence, and
implications. Emerg Infect Dis 6: 329-36.
Ishaq M, Wolf B, Ritter C (1990). Large-scale isolation of plasmid DNA using
cetyltrimethylammonium bromide. Biotechniques 9: 19-20, 22, 24.
Johnson MD, Oldach D, Delwiche CF, Stoecker DK (2007). Retention of transcriptionally
active cryptophyte nuclei by the ciliate Myrionecta rubra. Nature 445: 426-8.
Jones BW, Nishiguchi MK (2004). Counterillumination in the hawaiian bobtail squid,
Euprymna scolopes Berry (Mollusca : Cephalopoda). Marine Biology 144: 1151-
1155.
Jorgensen BB (1982). Ecology of the bacteria of the sulphur cycle with special reference to
anoxic-oxic interface environments. Philos Trans R Soc Lond B Biol Sci 298: 543-61.

BIBLIOGRAPHIE
124
Juniper SK, Sibuet M (1987). Cold seep benthic communities in Japan subduction zones -
Spatial organisation - Trophic strategies and evidence for temporal evolution. Marine
Ecology-Progress Series 40: 115-126.
Karlin S, Brocchieri L (1996). Evolutionary conservation of RecA genes in relation to protein
structure and function. J Bacteriol 178: 1881-94.
Keane TM, Creevey CJ, Pentony MM, Naughton TJ, McLnerney JO (2006). Assessment of
methods for amino acid matrix selection and their use on empirical data shows that ad
hoc assumptions for choice of matrix are not justified. BMC Evol Biol 6: 29.
Kim DR, Pritchard AE, McHenry CS (1997). Localization of the active site of the alpha
subunit of the Escherichia coli DNA polymerase III holoenzyme. J Bacteriol 179:
6721-8.
Kraus DW. (1993). Symposium on Life with Sulfide, at the Annual Meeting of the American-
Society-of-Zoologists. Amer Soc Zoologists: Los Angeles, Ca, pp 112-120.
Kraus DW, Wittenberg JB (1990). Hemoglobins of the Lucina pectinata/bacteria symbiosis. I.
Molecular properties, kinetics and equilibria of reactions with ligands. J Biol Chem
265: 16043-53.
Krueger DM, Gustafson RG, Cavanaugh CM (1996). Vertical transmission of
chemoautotrophic symbionts in the bivalve Solemya velum (Bivalvia: Protobranchia).
Biol Bull 190: 195-202.
Lane DJ (1991). 16S/23S Sequencing. In: Stackebrandt E GM (ed). Nucleic acid techniques
in bacterial systematics. John Wiley and Sons.: New York. pp 115-175.
Le Pennec M, Diouris M, Herry M (1988). Endocytosis and lysis of bacteria in gill epithelium
of Bathymodiolus thermophilus, Thyasira flexuosa, and Lucinella divaricata.
(Bivalvia:Mollusca). J Shellfish Res 7: 483-489.
Liberge M, Gros O, Frenkiel L (2001). Lysosomes and sulfide-oxidizing bodies in the
bacteriocytes of Lucina pectinata, a cytochemical and microanalysis approach. Mar
Biol 139: 401-409.
Margulis L, Fester R (1991). Bellagio conference and book. Symbiosis as Source of
Evolutionary Innovation: Speciation and Morphogenesis. Conference--June 25-30,
1989, Bellagio Conference Center, Italy. Symbiosis 11: 93-101.
McCann J, Stabb EV, Millikan DS, Ruby EG (2003). Population dynamics of Vibrio fischeri
during infection of Euprymna scolopes. Appl Environ Microbiol 69: 5928-34.
McCutcheon JP, McDonald BR, Moran NA (2009). Origin of an alternative genetic code in
the extremely small and GC-rich genome of a bacterial symbiont. PLoS Genet 5:
e1000565.

BIBLIOGRAPHIE
125
Meyer B, Kuever J (2007). Phylogeny of the alpha and beta subunits of the dissimilatory
adenosine-5'-phosphosulfate (APS) reductase from sulfate-reducing prokaryotes--
origin and evolution of the dissimilatory sulfate-reduction pathway. Microbiology
153: 2026-44.
Milne I, Wright F, Rowe G, Marshall DF, Husmeier D, McGuire G (2004). TOPALi:
software for automatic identification of recombinant sequences within DNA multiple
alignments. Bioinformatics 20: 1806-7.
Musat N, Giere O, Gieseke A, Thiermann F, Amann R, Dubilier N (2007). Molecular and
morphological characterization of the association between bacterial endosymbionts
and the marine nematode Astomonema sp. from the Bahamas. Environ Microbiol 9:
1345-53.
Newell ND (1969). Classification of Bivalvia. In:Treatise on invertebrates paleontology. .
The Geological Society of America and the University of Kansas. pp N205-N224.
Nikoh N, Nakabachi A (2009). Aphids acquired symbiotic genes via lateral gene transfer.
BMC Biol 7: 12.
Nussbaumer AD, Fisher CR, Bright M (2006). Horizontal endosymbiont transmission in
hydrothermal vent tubeworms. Nature 441: 345-8.
Oldroyd GED, Downie JA (2008). Coordinating Nodule Morphogenesis with Rhizobial
Infection in Legumes. Annual Review of Plant Biology 59: 519-546.
Olson PD, Cribb TH, Tkach VV, Bray RA, Littlewood DT (2003). Phylogeny and
classification of the Digenea (Platyhelminthes: Trematoda). Int J Parasitol 33: 733-
55.
Ott J, Bright M, Bulgheresi S (2004). Symbioses between marine nematodes and sulfur-
oxidizing chemoautotrophic bacteria. Symbiosis 36: 103-126.
Peek AS, Feldman RA, Lutz RA, Vrijenhoek RC (1998a). Cospeciation of chemoautotrophic
bacteria and deep sea clams. Proc Natl Acad Sci USA 95: 9962-6.
Peek AS, Vrijenhoek RC, Gaut BS (1998b). Accelerated evolutionary rate in sulfur-oxidizing
endosymbiotic bacteria associated with the mode of symbiont transmission. Mol Biol
Evol 15: 1514-23.
Pernthaler A, Pernthaler J, Amann R (2002). Fluorescence in situ hybridization and catalyzed
reporter deposition for the identification of marine bacteria. Appl Environ Microbiol
68: 3094-101.
Piednoel M, Bonnivard E (2009). DIRS1-like retrotransposons are widely distributed among
Decapoda and are particularly present in hydrothermal vent organisms. BMC
Evolutionary Biology 9: 86.

BIBLIOGRAPHIE
126
Polz MF, Cavanaugh CM (1995). Dominance of one bacterial phylotype at a Mid-Atlantic
Ridge hydrothermal vent site. Proc Natl Acad Sci USA 92: 7232-6.
Reid RGB, Brand DG (1986). Sulfide-Oxidizing Symbiosis in Lucinaceans - Implications for
Bivalve Evolution. Veliger 29: 3-24.
Rigaud T, Moreau J, Juchault P (1999). Wolbachia infection in the terrestrial isopod oniscus
asellus: sex ratio distortion and effect on fecundity. Heredity 83: 469-75.
Ronquist F, Huelsenbeck JP (2003). MrBayes 3: Bayesian phylogenetic inference under
mixed models. Bioinformatics 19: 1572-4.
Rozas J, Sanchez-DelBarrio JC, Messeguer X, Rozas R (2003). DnaSP, DNA polymorphism
analyses by the coalescent and other methods. Bioinformatics 19: 2496-7.
Rozen S, Skaletsky H (2000). Primer3 on the WWW for general users and for biologist
programmers. Methods Mol Biol 132: 365-86.
Ruby EG, Lee KH (1998). The Vibrio fischeri-Euprymna scolopes Light Organ Association:
Current Ecological Paradigms. Appl Environ Microbiol 64: 805-812.
Sachs JL, Mueller UG, Wilcox TP, Bull JJ (2004). The evolution of cooperation. Quarterly
Review of Biology 79: 135-160.
Schwarz G (1978). Estimating the Dimension of a Model. Ann. Statist. 6: 461-464.
Schweimanns M, Felbeck H (1985). Significance of the occurrence of chemoautotrophic
bacterial endosymbionts in lucinid clams from Bermuda. Mar Ecol Prog Ser 24: 113-
120.
Shimodaira H (2002). An approximately unbiased test of phylogenetic tree selection. Syst Biol
51: 492-508.
Soto JM, Domínguez-Ferreras A, Pérez-Mendoza D, Sanjuán J, Olivares J (2009). Mutualism
versus pathogenesis: the give-and-take in plant-bacteria interactions. Cellular
Microbiology 11: 381-388.
Spiridonova EM, Kuznetsov BB, Pimenov NV, Turova TP (2006). [Phylogenetic
characterization of endosymbionts of the hydrothermal vent mussel Bathymodiolus
azoricus by analysis of the 16S rRNA, pmoL, and cbbA genes]. Mikrobiologiia 75:
798-806.
Stewart FJ, Newton IL, Cavanaugh CM (2005). Chemosynthetic endosymbioses: adaptations
to oxic-anoxic interfaces. Trends Microbiol 13: 439-48.
Stouthamer R, Breeuwert JA, Luck RF, Werren JH (1993). Molecular identification of
microorganisms associated with parthenogenesis. Nature 361: 66-8.

BIBLIOGRAPHIE
127
Tan ZJ, Xie BY, Xiao QM, Yang YH, Wan FH, Huang SW (2005). Association of different
endosymbionts with the whitefly species Bemisia tabaci and Trialeurodes
vaporariorum (Sternorrhyncha : Aleyrodidae). Entomologia Generalis 28: 173-182.
Taylor JD, Glover EA (2005). Lucinidae (Bivalvia) – the most diverse group of
chemosymbiotic molluscs. Zool J Linnean Society 148.
Taylor JD, Williams ST, Glover EA, Dyal P (2007). A molecular phylogeny of heterodont
bivalves (Mollusca : Bivalvia : Heterodonta): new analyses of 18S and 28S rRNA
genes. Zoologica Scripta 36: 587-606.
Thompson JD, Higgins DG, Gibson TJ (1994). CLUSTAL W: improving the sensitivity of
progressive multiple sequence alignment through sequence weighting, position-
specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673-80.
Van Valen L (1973). A new evolutionary law. Evol. Theory 1: 1-30.
Vrijenhoek RC, Duhaime M, Jones WJ (2007). Subtype variation among bacterial
endosymbionts of tubeworms (Annelida: Siboglinidae) from the Gulf of California.
Biol Bull 212: 180-4.
Williams ST, Taylor JD, Glover EA (2003). Molecular Phylogeny of the Lucinoidea
(Bivalvia): Non-Monophyly and Separate Acquisition of Bacterial Chemosymbiosis. J
Molluscan Stud 70: 187-202.
Winnepenninckx BM, Reid DG, Backeljau T (1998). Performance of 18S rRNA in littorinid
phylogeny (Gastropoda: Caenogastropoda). J Mol Evol 47: 586-96.
Won YJ, Hallam SJ, O'Mullan GD, Pan IL, Buck KR, Vrijenhoek RC (2003). Environmental
acquisition of thiotrophic endosymbionts by deep-sea mussels of the genus
bathymodiolus. Appl Environ Microbiol 69: 6785-92.
Woolfit M, Bromham L (2003). Increased rates of sequence evolution in endosymbiotic
bacteria and fungi with small effective population sizes. Mol Biol Evol 20: 1545-55.
Woyke T, Teeling H, Ivanova NN, Huntemann M, Richter M, Gloeckner FO et al (2006).
Symbiosis insights through metagenomic analysis of a microbial consortium. Nature
443: 950-5.
Yu XJ, Walker DH, Liu Y, Zhang L (2009). Amino acid biosynthesis deficiency in bacteria
associated with human and animal hosts. Infect Genet Evol 9: 514-7.
Zbinden M, Cambon-Bonavita MA (2003). Occurrence of Deferribacterales and
Entomoplasmatales in the deep-sea Alvinocarid shrimp Rimicaris exoculata gut.
FEMS Microbiol Ecol 46: 23-30.
Zbinden M, Le Bris N, Gaill F, Compere P (2004). Distribution of bacteria and associated
minerals in the gill chamber of the vent shrimp Rimicaris exoculata and related
biogeochemical processes. Mar Ecol Prog Ser 284: 237-251.

BIBLIOGRAPHIE
128
Zbinden M, Shillito B, Le Bris N, de Montlaur CD, Roussel E, Guyot F et al (2008). New
insigths on the metabolic diversity among the epibiotic microbial communitiy of the
hydrothermal shrimp Rimicaris exoculata. Journal of Experimental Marine Biology
and Ecology 359: 131-140.
Zhang DX, Hewitt GM (2003). Nuclear DNA analyses in genetic studies of populations:
practice, problems and prospects. Mol Ecol 12: 563-84.
Zhou J, Bruns MA, Tiedje JM (1996). DNA recovery from soils of diverse composition. Appl
Environ Microbiol 62: 316-22.
Zimmer RL, Woollacott RM (1983). Mycoplasma-Like Organisms: Occurrence with the
Larvae and Adults of a Marine Bryozoan. Science 220: 208-210.

AAANNNNNNEEEXXXEEESSS


ANNEXE 1 : STRUCTURE DES BRANCHIES DE LUCINIDAE

ANNEXE 2 : LISTE DES BACTERIES UTILISEES DANS LE CADRE DE LA
RECHERCHE DE MARQUEURS.
ETUDE DE LA VARIABILITÉ.
ESPECE (SOUCHE) BACTERIENNE TaxID
(Genbank)
Escherichia coli K12 83333
Haemophilus influenzae rd KW20 71421
Hahella chejuensis KCTC2396 349521
Legionella pneumophila str. Paris 297246
Nitrosococcus oceani ATCC19707 323261
Saccharophagus degradans 2-40 203122
Salmonella typhimurium LT2 99287
Shigella boydii Sb227 300268
Vibrio vulnificus CMCP6 216895
Yersinia pestis CO92 214092
Escherichia coli 536 362663
Escherichia coli APEC01 405955
Escherichia coli ATCC8739 481805
Escherichia coli CFT073 199310
Escherichia coli IAI39 585057
Escherichia coli K12 83333
Escherichia coli O157:H7 83334
Escherichia coli s88 585035
Escherichia coli UMN026 585056
Escherichia coli UTI89 364106
Legionella pneumophila str. Paris 297246
Legionella pneumophila str. Lens 297245
Legionella pneumophila str. Corby 400673
Legionella pneumophila str. Philadelphia 1 272624

RECHERCHE DE TRANSFERTS HORIZONTAUX
ESPECE BACTERIENNE TAXON TaxID
(Genbank)
Escherichia coli K12
γ-protéobactéria
83333
Hahella chejuensis KCTC2396 349521
Salmonella typhimurium LT2 99287
Shigella boydii Sb227 300268
Yersinia pestis CO92 214092
Bordetella pertussis Tohama I
β-protéobactéria
257313
Burkholderia pseudomallei 1710b 320372
Neisseria meningitidis Z2491 122587
Nitrosomonas europaea ATCC 19718 228410
Nitrosospira multiformis ATCC 25196 323848
Bacillus subtilis subsp. subtilis str. 168
Bacilli
224308
Lactococcus lactis subsp. cremoris MG1363 416870
Listeria monocytogenes str. 4b F2365 265669
Staphylococcus aureus subsp. aureus Mu50 158878
Streptococcus agalactiae NEM316 211110
Agrobacterium tumefaciens str. C58
α-proteobacteria
176299
Magnetospirillum magneticum AMB-1 342108
Rhizobium etli CFN 42 347834
Rhodopseudomonas palustris CGA009 258594
Mycobacterium tuberculosis H37rv Actinobacteria
83332
Streptomyces coelicolor A3(2) 100226
Chlamydia trachomatis A/HAR-13 Chlamydia
315277
Chlamydophila pneumoniae CWL029 115713
Borrelia burgdoferi B31 Spirochaetes
224326
Leptospira interrogans servoar lai str. 56601 189518
Anabaena varabbilis ATCC29413 Cyannobacteria
240292
Synechococcus elongatus PCC6301 269084

ANNEXE 3: PROFIL « SVARAP » DU GENE DNAE.
ANNEXE 4 : TECHNIQUES PARTICULIERES UTILISEES.
PCR EMBOITEE
« TE-WALKING »


AAARRRTTTIIICCCLLLEEESSS


ARTICLE 1 : BRISSAC ET AL. (2009)
FEMS Microbiol. Ecol. 67:261-267










ARTICLE 2 : BRISSAC ET AL.
Soumis à Environmental Microbiology


Unexpected structured intraspecific symbiotic diversity
within Lucinidae (Mollusca: Bivalvia).
Terry Brissac1,2,*
, Dominique Higuet1, Olivier Gros
2 and Hervé Merçot
1
1UMR 7138 UPMC-CNRS-MNHN-IRD « Systématique, Adaptation, Évolution », Equipe:
« Génétique & Evolution », Université Pierre et Marie Curie, Bat. A, 4ème
étage, 7 quai St
Bernard, 75005 Paris, France.
2 UMR 7138 UPMC-CNRS-MNHN-IRD « Systématique, Adaptation, Évolution », Equipe:
« Symbiose », Département de Biologie, Université des Antilles et de la Guyane B.P. 592,
97159 Pointe-à-Pitre Cedex, Guadeloupe (France).
Correspondence: UMR 7138 CNRS-IRD-UPMC-MNHN « Systématique, Adaptation,
Evolution », Equipe « Génétique & Evolution », Université Pierre et Marie Curie, Bat. A, 4ème
étage, 7 quai St Bernard, 75005 Paris, France. E-mail: [email protected].
Running Head: Bacterial strain diversity within Lucinidae.
Abstract
In chemoautotrophic associations, 16S rRNA usual marker was often used to characterize
symbionts diversity. Within Lucinidae, this symbiotic diversity seems no species-specific
since exactly the same sulfur-oxidizing bacterial species was described harbored by six host
species belonging to four different genus from Lesser Antilles. Nevertheless, according to the
low evolution rate of this marker, a strain diversity linked to geography, ecosystems or hosts
diversity could occur as described within Mytilidae or vestimentiferan tubeworms. In order to
highlight a potential diversity of symbionts associated with this six host species, we
developed a MLST (Multi-Locus Sequence Typing) analysis using five different markers.
According to our survey, a symbiont strain diversity of near 1.5% within Lucinidae is
obvious. Bacterial haplotype networks realized with sequences reveal whatever the marker the
same haplotype pattern. At local level (same biotope in same geographic location), this
observed diversity appears to be structured in function of host diversity. Moreover, in-situ
hybridizations realized on cross-infected individuals attest of this specificity of association.
Despite the fact that the method used here (PCR product direct sequencing) could
underestimate diversity, for a same bacterial species we observe specific associations between
hosts and bacterial strains which could be due at local level to a Red-Queen type antagonist
evolution.
Keywords: Codakia / MLST / Haplotype networks / Red-Queen evolution.

Introduction
From ecology to morphology and enzymology, many studies have permitted a better
understanding of associations between marine invertebrates and chemoautotrophic bacteria
(Dubilier et al., 2008; Felbeck et al., 1981; Fisher, 1990; Stewart et al., 2005). In these
associations, the characterization of symbionts was realized using 16S rRNA sequences
(Blazejak et al., 2006; Cary et al., 1993; Dubilier et al., 1995; Duperron et al., 2008b; Durand
et al., 1996; Polz and Cavanaugh, 1995; Polz et al., 1994). According to this maker, the
majority of symbionts involved in these interactions belong to the gamma-proteobacteria
subdivision even if alpha-, delta-, epsilon-proteobacteria and spirochaetes are also retrieved
(Dubilier et al., 2008). These associations have been described in single or multi-infections
(DeChaine and Cavanaugh, 2006; Duperron et al., 2008a; Woyke et al., 2006). 16S rRNA
sequences have also been used to study symbiont diversity compared to host diversity (Distel
et al., 1994; Distel et al., 1988; Feldman et al., 1997; Peek et al., 1998).
Commonly, species-specific associations are maintained by vertical transmission of the
symbiont (Peek et al., 1998). In fact, in cases where symbionts are vertically transmitted
(from parents to offspring via gametes), congruent evolutionary histories of hosts and
symbionts, due to co-evolution of the two partners, lead to the specific couples observed
today, like those described for example within the bivalve family of Vesicomyidae (Peek et
al., 1998). However, in cases where symbionts are not vertically acquired, symbiont diversity
does not seem related to host diversity. For example, within vestimentiferan tubeworms or
lucinid bivalves no species-specific association is apparent since different host species can be
associated to the same symbiont species (Durand and Gros, 1996; Durand et al., 1996;
Feldman et al., 1997; Gros et al., 2000; Gros et al., 2003a). In these cases the association
seems to depend on the free-living symbiont form of the bacterial species present in the
environment. Thus, host species inhabiting the same environments harbor the same (or close)
symbiont species. In the latter situation, 16S rRNA could be insufficient to highlight eventual
diversity (Di Meo et al., 2000; Fox et al., 1992; Tanabe et al., 2007; Woese, 1987). Thus in
Thalassia testudinum sea-grass beds of the Lesser Antilles, where six lucinid host species
harbor the same bacterial species (Durand and Gros, 1996; Durand et al., 1996; Gros et al.,
2000; Gros et al., 2003a), this bacterial species may present a strain diversity related to host
species (phylotype), geographic location or ecological factors (ecotype), as described for
example in vestimentiferan tubeworms (Di Meo et al., 2000) or Mytilidae (DeChaine and
Cavanaugh, 2006; Won et al., 2003). Accordingly, as in the latter case, further analyses must
be used to study the genetic variation of endosymbionts and the specificity of host/symbiont
couples within the Lucinidae/sulfur-oxidizing endosymbionts model.
To date, all lucinid genera analyzed harbor endosymbiotic thioautotrophic gamma-
proteobacteria localized in specialized cells of the gills called bacteriocytes (Fisher, 1990).
Bacterial colonization occurs at the juvenile stage after metamorphosis as previously
described in the tropical shallow-water species Codakia orbicularis (Gros et al., 1996), in the
deep-water species Lucinoma aequizonata (Gros et al., 1999) and suggested for other
Caribbean lucinids (Gros et al., 1998); symbionts being acquired from a free-living stock of
symbiosis-competent bacteria present in the environment (Gros et al., 2003b). In this paper,
we aim to analyze genetic intraspecific diversity within the symbiotic species shared by the
six lucinid host species inhabiting sea-grass beds of Guadeloupe and Martinique (Lesser
Antilles). We developped a Multi-Locus Sequence Typing (MLST) approach using five
different markers to measure strain diversity between gill-endosymbionts from different
lucinid hosts and to determine whether this diversity is structured in function of hosts,
geographic location or environmental factors.

Results
Symbionts 16S rRNA sequences analysis
All PCR amplifications realized from gills of Anodontia alba, Codakia orbicularis, Codakia
orbiculata, Codakia pectinella, Divaricella quadrisulcata and Linga pensylvanica gave an
unique 16S rRNA bacterial sequence per host without any ambiguity according to ABI
chromatograph analysis. These sequences present 99.6% mean sequence identity (1466 bp).
Host phylogeny
For all amplifications of 18S rRNA and 28S rRNA genes, a unique band of the expected size
(~1000bp and ~700bp respectively) was observed. We analyzed the six host species named
above and chose as outgroup for phylogenetic analyses, Thyasiridae and Crassatellidae, which
are sister-groups of Lucinidae (Williams et al., 2003). According to this phylogeny (Fig. 1)
the six host lucinids fall in the different clades described by Williams et al. (Williams et al.,
2003): Anodontia clade for A. alba, clade A for C. orbicularis and C. orbiculata and Clade B
for C. pectinella, D. quadrisulcata and L. pensylvanica. Concerning A. alba and C.
orbicularis, sequences obtained here correspond to those published by Williams et al.
(Williams et al., 2003).
Symbiont polymorphism analysis
Polymorphism results are presented in Table 1 and show that about 1.5% of sites are
polymorphic. ITS1 constitutes the most variable marker (2.14%), aprA and cbbL metabolism
genes are more variable than dnaE and gyrB housekeeping genes (1.62%, 1.83%, 1.29% and
0.74% respectively). According to haplotype networks (Fig. 2), realized using sequences of
five markers on 18 individuals (three per host species), we observe different haplotypes (up to
five or six according to the marker) showing an intra-specific bacterial diversity. Interestingly,
except for ITS1, the major haplotype seems not to be an ancestral haplotype from which
others are derived. Considering host species, the different haplotypes are not randomly
distributed among the 18 individuals analyzed. Thus, the observed diversity seems structured.
For C. orbicularis the three individuals present whatever the marker the same particular
haplotype. This pattern is also retrieved for individuals of L. Pensylvanica. For C. orbiculata,
C. pectinella the same haplotype is shared by the different individuals belonging to these host
species (Fig. 2). In addition, we also sequenced dnaE, gyrB, and aprA for three C. orbicularis
sub-adults (0.5 cm shell-length). According to these sequences, sub-adults harbor exactly the
same symbiont as adults (100% of sequence identity for each marker). Moreover, the layer of
sediment comprised between 0 and 5 cm was also analyzed using dnaE marker. After
sequencing of five clones we observed one clone which corresponds to C. orbicularis
symbiont (one mismatch for 465 bp). The four other clones seem to correspond to bacterial
sequences different from those harbored by host species inhabiting sea-grass beds of Îlet
Cochon (mean sequence divergence of 83% compared to C. orbicularis symbiont). Finally,
dnaE sequences for one C. orbicularis individual and one C. orbiculata individual from
Martinique were also realized. These two host species harbor bacterial sequences which are
different from those retrieved in the same host species in Guadeloupe (respectively two and
four mismatches in relation to Guadeloupe symbionts).

Cross-Infections
Slides from cross-infections were hybridized with symco2A probe (16S rRNA probe specific
of Lucinidae symbionts from Lesser Antilles (Gros et al., 2003b)). A positive hybridization in
the lateral zone where symbionts are distributed is observed throughout gill sections of freshly
collected C. orbiculata used as positive control (Fig. 3A). Gill sections from C. orbiculata
individuals kept in starvation in the laboratory during 9 months presented no hybridization
signal (Fig. 3B) suggesting that such starved individuals are devoid of endosymbionts (OG,
personal communication). Such starved individuals put in contact with C. orbicularis
bacterial gill-endosymbionts purified fraction did not present any bacterial endosymbiont
positively detected by CARD-FISH after 4 days (Fig. 3C) or 8 days (Fig. 3D) suggesting the
lack of infection by symbiosis-competent bacteria. On the other hand, when C. orbiculata
starved individuals were put in contact with gill-endosymbiont purified fraction from C.
orbiculata individuals freshly collected from the field, we observed the first step of bacterial
colonization after 4 days (Fig. 3E), a more developed infection after 8 days (Fig. 3F) up to a
complete colonization after 15 days (data not shown).
Discussion
The purpose of this study was to analyze the genetic diversity of sulfur-oxidizing
endosymbionts associated with six bivalve species of the Lucinidae family (A. alba, C.
orbicularis, C. orbiculata, C. pectinella, D. quadrisulcata and L. pensylvanica) inhabiting T.
testudinum sea-grass beds in Lesser Antilles. We sequenced 16S rRNA in symbionts of these
six host species (three individuals per host species). We found that these six host species
harbor the same bacterial species as previously described (Durand and Gros, 1996; Durand et
al., 1996; Gros et al., 2000; Gros et al., 2003a). Considering that 16S rRNA is not
discriminant enough to describe diversity at intra-specific level, we used a MLST approach to
detect a possible strain diversity within these endosymbionts, which may be structured as a
function of host diversity, geographic localization or environmental conditions.
Genetic diversity within sulfur-oxidizing symbionts displayed by MLST analysis.
We used five markers classified in three categories following Cooper and Feil (Cooper and
Feil, 2004): (i) housekeeping genes, (ii) core metabolism gene and (iii) hypervariable genes.
As expected, ITS1 is the most variable marker even if it does not present the higher number of
different haplotypes. This is due to the accumulation of polymorphic sites within symbionts of
A. alba (Fig. 2). The lowest sequence divergence is found in housekeeping genes (dnaE and
gyrB), and metabolism genes (aprA and cbbL) present an intermediary variability (Table 1).
These results are consistent with our initial requirement (i.e. greater variability than 16S
rRNA) and reveal an intra specific diversity of symbionts associated to these six Lucinidae
host species from Lesser Antilles. Indeed, as expected, we observed a mean sequence
divergence of approximately 1.5% which is consistent with the literature (Zhang and Hewitt,
2003). However, this diversity could be underestimated due to techniques employed here (i.e.
direct PCR product sequencing) which detect only the majoritary sequence of each marker.
The question is now to determine whether this bacterial diversity is randomly distributed or
structured as a function of host diversity, geographic location or environmental factors.
Structuration of symbiont diversity.
In order to test the structuration of sulfur-oxidizing symbiont diversity in Caribbean lucinids,
we realized haplotype networks using sequences of our markers. We found that bacterial
diversity is structured because whatever the marker considered haplotypes are not randomly
distributed among individuals (Fig. 2). Moreover, even if haplotype networks are not

congruent, the same patterns are retrieved for each of them: (i) polymorph for A. alba
symbionts (ii) polymorph or monomorph for D. quadrisulcata symbionts (iii) monomorph for
symbionts of the four other host species. The latter observation reveals a specificity of
association between L. pensylvanica and its symbiont, between C. orbicularis and its
symbiont, and surprisingly a specificity of association between a particular bacterial strain
and two host species (C. pectinella and C.orbiculata). In addition, this specificity is very
unlikely to result from sampling bias since the probability to obtain a particular bacterial
sequence by chance for three hosts among 18, is only 4.6 10-3
(i.e. (3/18)3) under the
hypothesis than hosts capture their symbionts randomly in the same environmental bacterial
population.
Does geography play a role in this structuration?
Our six host species have been sampled in one of three distinct geographic locations: la
Désirade (D. quadrisulcata), Martinique (L. pensylvanica) and Îlet Cochon (A. alba, C.
orbicularis, C. orbiculata and C. pectinella). Accordingly, the genetic structuration we
observed could be due to geography. This criterion could be considered for D. quadrisulcata
and L. pensylvanica. In fact, each of these two species was sampled in geographic locations
different from other host species and presents a particular symbiotic haplotype pattern. By
contrast the four other host species (A. alba, C, orbicularis, C. orbiculata and C. pectinella)
do not share the same bacterial haplotype although they have been sampled in the same sea-
grass bed (Îlet Cochon). Since geography is obviously not a pertinent explanation here, then
which factor may be involved in the structuration we observe in Îlet Cochon?
Ecotype or Phylotype?
Two hypotheses could explain the structuration of symbiotic diversity among the four
host/symbiont couples of Îlet Cochon:
(i) The ecotype hypothesis: bacterial population is structured in the environment as a function
of ecosystem parameters like sulfur and oxygen concentrations or other gradient in Thalassia
testudinum sea-grass beds sediments (Fig 4A). According to this hypothesis, the bacterial
structuration observed in hosts mimics the structuration of bacterial populations in the
environment. Consequently hosts localized at different positions of the sediment column
could harbor different bacterial strains. This possibility could be considered for C. pectinella
and C. orbiculata which are only found between 0 and 5 cm of depth due to the length of their
siphon (Allen, 1958) and which share for each marker the same bacterial haplotype (Fig. 2).
Then, only this bacterial haplotype should be retrieved in superior strata of sediment, in
juveniles (or sub-adults) or in any clam found in this superior layer. To test this hypothesis we
analyzed the sediment of this layer and also sub-adults of C. orbicularis (up to 0.5cm shell-
length) sampled in this superior layer. Our results refute the ecotype hypothesis since we
detected free-living forms of C. orbicularis symbionts in the superior layer of sediment.
Moreover, sub-adults of C. orbicularis harbor the same bacterial strain as adults (which are
distributed through the sediment layer up to 20 cm depth). It thus seems that bacterial
population is not structured as a function of sediment depth (Fig. 4B).
(ii) Phylotype hypothesis: structuration as a function of host species could be due to co-
evolution of host and bacterial strains. In this case, close host species should harbor close
symbiont species. This second hypothesis is rejected here because this pattern is not observed
when comparing host phylogeny (Fig. 1) and structured bacterial diversity (Fig.2). Indeed, the
two hosts sharing the same bacterial strain (C. orbiculata and C. pectinella [Fig.2, yellow/red
circle]) are not the closest relative at all and belong respectively to A and B Lucinidae clades
(Fig. 1). Then, within symbionts of Lucinidae inhabiting Thalassia testudinum sea-grass beds

of Îlet Cochon, neither ecotype nor phylotype hypotheses can explain the observed
structuration. This leads us to propose that the specific pattern of association between the two
partners observed here can only be explained by a local selective process between host
species and the symbiotic bacterial pool.
Local selective hypothesis.
How could a selective process between lucinid host species and their symbionts lead to
observed structuration? Figure 5 proposes a hypothesis explaining this structuration. The
proposed steps are discussed according to the environmental recruitment model (Brissac et
al., 2009). Step 1: 4 host species exploit the same bacterial pool. This step corresponds to an
initial equilibrium state. Step 2: A mutant for symbiont recognition targets appears in the
bacterial population. Step 3: This mutant is not recognized by hosts and its proportion
increase in the environment. In fact, selective valor of this new mutant is superior to the
ancient one because this symbiont escapes to internalization which constitute an evolutionary
dead-end (Brissac et al., 2009). Step 4: Apparition of mutant hosts which are able to
recognize the new symbiont. These new host variants exploit the new bacterial pool while
their conspecifics remain restricted to exploiting the ancient bacterial pool. Step 5: The
frequency of these new host individuals increases in case of a selective advantage. In fact, this
new bacterial strain could be more efficient in translation to host of fixed carbon. Moreover,
the new association could also decrease the “competitions for symbionts” because the new
host exploits a new ecological niche constituted by the mutant bacterial pool. Step 6:
Constitution of a specific association between the mutated host and the bacterial mutant. Step
7: Bacterial mutants for characters which are not implied in the recognition could appear and
continue to be captured by the host. This latter step could both explain the specificity
observed here and the presence of polymorphic symbionts (e.g. within A. alba). Our markers
are not implied in recognition and supposed to be neutral. So, the observed structuration
reveals a selection of recognition target and a selection by hitchhiking of the alleles of our
markers. This selective hypothesis reinforces the environmental recruitment model compared
with environmental transmission (Gros et al., 1996) because in this last one it is not obvious
that step 6 can occur. In fact, in this case, a new association could not be established since no
selective pressure occurs due to the reseeding of the environment by host via the release of the
symbionts (Brissac et al., 2009). Under such a model (i.e. selective hypothesis in
environmental recruitment), we must expect that in Îlet Cochon one host could not be
associated with another bacterial strain than its own. The fact that C. orbiculata did not
become infected by symbionts from C. orbicularis gill extracts agree in favor of our model.
The selective hypothesis could therefore explain the observed intra-specific structuration of
symbionts in T. testudinum sea-grass beds of the Îlet Cochon.
In conclusion the six host species inhabiting T. testudinum sea-grass beds in the Lesser
Antilles share the same bacterial species (according to 16S rRNA sequences). However our
MLST analysis reveals a pattern of intra-specific genetic diversity of symbionts. This pattern
seems both to have a geographic and also a host-species component which could be explained
by Red-Queen selective process (Van Valen, 1973).

Material & Methods
Specimen collection.
Individuals of Anodontia alba (Link, 1807), Codakia orbicularis (Linné, 1758), Codakia
orbiculata (Montagu, 1808), Codakia pectinella (Adams, 1852), were collected in
Guadeloupe (“Îlet Cochon” 16°12'53.76''N; 61°32'05.74''W). For Divaricella quadrisulcata
(Orbigny, 1842), specimens were collected in la Désirade (“plage des galets” 16°17'46.88''N;
61°04'53.77''W). Specimens of Linga pensylvanica (Linné, 1758) were collected in
Martinique (“Anse Macabou” 14°31'01.52''N; 60°49'00.86''W).
All individuals were collected by hand in (or near) Thalassia testudinum sea-grass beds. Gills
(which harbor symbionts) and other symbiont-free tissues (e.g. foot) were conserved in
absolute ethanol before investigation.
Multi-Locus Sequence Typing.
We used in this study three types of markers: (i) two housekeeping genes already used for
typing purposes in the literature: DNA Polymerase III alpha-subunit (dnaE) (Byun et al.,
1999) and DNA Gyrase subunit B (gyrB) (Holmes et al., 2004), (ii) the Internal Transcribed
Spacer (ITS1), an hypervariable region comprised between 16S and 23S rRNA genes, and (iii)
two metabolic genes already used to characterize symbiont metabolic pathway in Lucinidae:
Ribulose-1,5-bisPhosphate Carboxylase/Oxygenase (RubisCO [cbbL]) for chemoautotrophy
and Adenylyl-1,5-bisPhosphate Reductase alpha-subunit (APS Reductase [aprA]) for sulfur-
oxidizing metabolism (Duperron et al., 2007). Corresponding sequences of lucinid sulfur-
oxidizing symbionts were obtained by nested-PCR on C. orbicularis (see below) except for
RubisCO for which literature primers were used (Elsaied and Naganuma, 2001). The
sequences used for polymorphism analysis were obtained by PCR with specific primers
(Table 2).
DNA Preparation.
Template DNA for PCR amplifications was extracted from symbiont-containing gills and
symbiont-free foot tissue of each bivalve hosts according to the CTAB method (Ishaq et al.,
1990). DNA concentration was determined by spectrometry (BioPhotometer, Eppendorf) and
on a 0.8% agarose gel in comparison with a concentration marker (Smart Ladder,
Eurogentec).
Nested-PCR amplifications.
Few data for these genes within sulfur-oxidizing symbionts being available in databases, we
determined their sequences for lucinid sulfur-oxidizing symbionts. Two-step Nested-PCR
amplifications were realized with DNA extracted from C. orbicularis gills. The first step was
performed in 25µL, using degenerated “nested-primers” (Table 2) and 10-25ng of
symbiont+host template DNA mixture. The second amplification was performed in the same
conditions with “degN primers” instead of “nested-primers” and 1µL of a 100-fold to 1000-
fold dilution of the first amplification as a template. Conserved sites flanking more variable
regions were determined from aligned bacterial sequences using SVARAP 2.0 (Colson et al.,
2006). Degenerated “nested” and “degN” primers were then designed in these conserved
regions.

Specific PCR amplifications.
Specific primers were designed using Primer3 software (http://frodo.wi.mit.edu/cgi-
bin/primer3/primer3.cgi (Rozen and Skaletsky, 2000)). Bacterial sequences were amplified
from host+symbiont DNA mixture using primers presented in Table 2. Amplification was
performed in a final volume of 50µL using 10-25ng of DNA template, 1X Taq Buffer
(Invitrogen), 3mM MgCl2 (Invitrogen), 0,4mM of each dNTP, 1mM of each primer and 2
units of recombinant Taq DNA polymerase (Invitrogen). Amplicons were then directly
sequenced (GATC-biotech, www.gatc-biotech.com).
Sediment analysis.
Sediment from the supeficial layer (0-5cm) of Thalassia testudinum sea-grass beds was
collected in Îlet Cochon and total DNA was extracted following Zhou et al. (Zhou et al.,
1996). A 3-step nested PCR was realized using 50ng of DNA template and “nested” primers
(Table 2) as a first step. A 100-fold dilution of this PCR-product was then used as template
for a second amplification with “degN” primers (Table 2) and the procedure was repeated for
a last step with specific primers. Fragments of interest were then eluated from 1% agarose gel
using Nucleospin® Extract II kit (Macherey-Nagel) and cloned with pGEM-T® TA cloning
kit (Promega) according to manufacturers instructions. This vector was used to transform
Dh5α competent Escherichia coli (Promega) and plasmids from positives clones were
collected using the Nucleospin® plasmid kit (Macherey-Nagel). Inserts were then sequenced
directly using a ABI PRISM automated sequencer (Genome express, www.gexbyweb.com).
Host sequences.
DNA extracted from symbiont-free foot tissue was used in order to amplify 18S rRNA and
28S rRNA. The PCR were realized in the same conditions as symbiont-specific PCR
amplification. PCR products were then directly sequenced and phylogenetically analyzed.
Phylogenetic analysis.
Sequences were controlled by BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and aligned
with ClustalW (Thompson et al., 1994) with default parameters. The alignment was then eye-
checked and corrected if necessary. Gap positions and ambiguous sites were removed using
the online Gblocks server (Castresana, 2000). The alignment was then imported in the
TOPALi v2.5 software (Milne et al., 2004) to define the substitution model applied to each
dataset using ModelGenerator (Keane et al., 2006). The best model was chosen using the
Bayesian Information Criterion (BIC) (Schwarz, 1978). Phylogenetic analyses were then
performed with TOPALi v2.5, trees being then estimated by Maximum Likelihood (ML)
using PhyML (Guindon and Gascuel, 2003) and by Bayesian Inference (BI) using MrBayes 3
(Ronquist and Huelsenbeck, 2003). Support for individual clades was evaluated using non-
parametric bootstrapping (Felsenstein, 1985) obtained from 100 replicates ML. Posterior
probabilities for BI were also evaluated for each node.
Polymorphism analysis.
MLST symbiont sequences for each gene were analyzed using DNAsp4 (Rozas et al., 2003)
and results are presented in Table 1. Haplotype networks were realized using NETW4510©
(Fluxus Technology Ltd.) using Median Joining method (Bandelt et al., 1999).

Cross-infections experiments.
Twenty C. orbiculata individuals were purged of their symbionts by 9 month starvation in
oxygenated tanks with no sulfur added. Clams were then dispatched in four oxygenated
cristallizing dishes. Purified symbiont fractions realized according to Caro et al. (Caro et al.,
2007) from C. orbicularis or C. orbiculata gills from freshly collected animals, were added
every two days into the batches. A part of these extracts was conserved in 100% ethanol for
molecular analysis. Individuals were sacrificed every four days in order to follow a possible
infection of starved hosts. For each sacrificed individual, one gill was conserved for
molecular investigation (100% ethanol) and the other one was directly fixed (2%
paraformaldehyde in 0.2µm filtered seawater for 3h at 4°C) for CARD-FISH. Slides and
CARD-FISH experiments were realized according to Brissac et al. (2009).
Acknowledgments
We thank, Denis Poinsot and Clémentine Vitte for English revision. T.B is funded as PhD
Student by a MENRT grant (Ministère de l’Education Nationale de la Recherche et des
Technologies).
References
Allen JA (1958). On the Basic Form and Adaptations to Habitat in the Lucinacea
(Eulamellibranchia). Philosophical Transactions of the Royal Society of London. Series
B, Biological Sciences 241: 421-484.
Bandelt HJ, Forster P, Rohl A (1999). Median-joining networks for inferring intraspecific
phylogenies. Mol Biol Evol 16: 37-48.
Blazejak A, Kuever J, Erseus C, Amann R, Dubilier N (2006). Phylogeny of 16S rRNA,
ribulose 1,5-bisphosphate carboxylase/oxygenase, and adenosine 5'-phosphosulfate
reductase genes from gamma- and alphaproteobacterial symbionts in gutless marine
worms (oligochaeta) from Bermuda and the Bahamas. Appl Environ Microbiol 72:
5527-36.
Brissac T, Gros O, Mercot H (2009). Lack of endosymbiont release by two Lucinidae
(Bivalvia) of the genus Codakia: consequences for symbiotic relationships. FEMS
Microbiol Ecol 67: 261-7.
Byun R, Elbourne LD, Lan R, Reeves PR (1999). Evolutionary relationships of pathogenic
clones of Vibrio cholerae by sequence analysis of four housekeeping genes. Infect
Immun 67: 1116-24.
Caro A, Gros O, Got P, De Wit R, Troussellier M (2007). Characterization of the population
of the sulfur-oxidizing symbiont of Codakia orbicularis (Bivalvia, Lucinidae) by single-
cell analyses. Appl Environ Microbiol 73: 2101-9.
Cary SC, Warren W, Anderson E, Giovannoni SJ (1993). Identification and localization of
bacterial endosymbionts in hydrothermal vent taxa with symbiont-specific polymerase
chain reaction amplification and in situ hybridization techniques. Mol Mar Biol
Biotechnol 2: 51-62.
Castresana J (2000). Selection of conserved blocks from multiple alignments for their use in
phylogenetic analysis. Mol Biol Evol 17: 540-52.
Colson P, Tamalet C, Raoult D (2006). SVARAP and aSVARAP: simple tools for
quantitative analysis of nucleotide and amino acid variability and primer selection for
clinical microbiology. BMC Microbiol 6: 21.

Cooper JE, Feil EJ (2004). Multilocus sequence typing--what is resolved? Trends Microbiol
12: 373-7.
DeChaine EG, Cavanaugh CM (2006). Symbioses of methanotrophs and deep-sea mussels
(Mytilidae: Bathymodiolinae). Prog Mol Subcell Biol 41: 227-49.
Deplancke B, Hristova KR, Oakley HA, McCracken VJ, Aminov R, Mackie RI et al (2000).
Molecular ecological analysis of the succession and diversity of sulfate-reducing
bacteria in the mouse gastrointestinal tract. Appl Environ Microbiol 66: 2166-74.
Di Meo CA, Wilbur AE, Holben WE, Feldman RA, Vrijenhoek RC, Cary SC (2000). Genetic
variation among endosymbionts of widely distributed vestimentiferan tubeworms. Appl
Environ Microbiol 66: 651-8.
Distel DL, Felbeck H, Cavanaugh CM (1994). Evidence for phylogenetic congruence among
sulfur-oxidizing chemoautotropihc becterial endosymbiont and their Bivalve hosts. J
Mol Evol 38: 533-542.
Distel DL, Lane DJ, Olsen GJ, Giovannoni SJ, Pace B, Pace NR et al (1988). Sulfur-
oxidizing bacterial endosymbionts: analysis of phylogeny and specificity by 16S rRNA
sequences. J Bacteriol 170: 2506-10.
Dubilier N, Bergin C, Lott C (2008). Symbiotic diversity in marine animals: the art of
harnessing chemosynthesis. Nat Rev Microbiol 6: 725-40.
Dubilier N, Giere O, Distel DL, Cavanaugh CM (1995). Characterization of
chemoautotrophic bacterial symbionts in a gutless marine worm (Oligochaeta:
Annelida) by phylogenetic 16S rRNA sequence analysis and in situ hybridization. Appl
Environ Microbiol 61: 2346-50.
Duperron S, Fiala-Medioni A, Caprais JC, Olu K, Sibuet M (2007). Evidence for
chemoautotrophic symbiosis in a Mediterranean cold seep clam (Bivalvia: Lucinidae):
comparative sequence analysis of bacterial 16S rRNA, APS reductase and RubisCO
genes. FEMS Microbiol Ecol 59: 64-70.
Duperron S, Halary S, Lorion J, Sibuet M, Gaill F (2008a). Unexpected co-occurrence of six
bacterial symbionts in the gills of the cold seep mussel Idas sp. (Bivalvia: Mytilidae).
Environ Microbiol 10: 433-45.
Duperron S, Laurent MC, Gaill F, Gros O (2008b). Sulphur-oxidizing extracellular bacteria in
the gills of Mytilidae associated with wood falls. FEMS Microbiol Ecol 63: 338-49.
Durand P, Gros O (1996). Bacterial host specificity of Lucinacea endosymbionts:
interspecific variation in 16S rRNA sequences. FEMS Microbiol Lett 140: 193-8.
Durand P, Gros O, Frenkiel L, Prieur D (1996). Phylogenetic characterization of sulfur-
oxidizing bacterial endosymbionts in three tropical Lucinidae by 16S rDNA sequence
analysis. Mol Mar Biol Biotechnol 5: 37-42.
Elsaied H, Naganuma T (2001). Phylogenetic diversity of ribulose-1,5-bisphosphate
carboxylase/oxygenase large-subunit genes from deep-sea microorganisms. Appl
Environ Microbiol 67: 1751-65.
Felbeck H, Childress JJ, Somero GN (1981). Calvin-Benson cycle and sulphide oxidation
enzymes in animals from sulphide-rich habitats. Nature 293: 291-3.
Feldman RA, Black MB, Cary CS, Lutz RA, Vrijenhoek RC (1997). Molecular phylogenetics
of bacterial endosymbionts and their vestimentiferan hosts. Mol Mar Biol Biotechnol 6:
268-77.
Felsenstein J (1985). Confidence limits on phylogenies: an approach using the bootstrap.
Evolution 39: 783-791.
Fisher CR (1990). Chemoautotrophic and methanotrophic symbioses in marine invertebrates.
Rev. Aquat. Sci 2: 399-613.
Fox GE, Wisotzkey JD, Jurtshuk P, Jr. (1992). How close is close: 16S rRNA sequence
identity may not be sufficient to guarantee species identity. Int J Syst Bacteriol 42: 166-
70.

Gros O, Darrasse A, Durand P, Frenkiel L, Mouëza M (1996). Environmental transmission of
a sulfur-oxidizing bacterial gill endosymbiont in the tropical lucinid bivalve Codakia
orbicularis. Appl Environ Microbiol 62: 2324-30.
Gros O, De Wulf-Durand P, Frenkiel L, Mouëza M (1998). Putative environmental
transmission of sulfur-oxidizing bacterial symbionts in tropical lucinid bivalves
inhabiting various environments. FEMS Microbiol Lett 160: 257-262.
Gros O, Duplessis MR, Felbeck H (1999). Embryonic Development and endosymbiont
transmission mode in the symbiotic clam Lucinoma aequizonata (Bivalvia:Lucinidae).
Invert Reprod Dev 36: 93-103.
Gros O, Frenkiel L, Felbeck H (2000). Sulfur-oxidizing endosymbiosis in Divaricella
quadrisulcata (Bivalvia: Lucinidae): Morphological, ultrastructural, and phylogenetic
analysis. Symbiosis 29: 293-317.
Gros O, Liberge M, Felbeck H (2003a). Interspecific infection of aposymbiotic juveniles of
Codakia orbicularis by various tropical lucinid gill-endosymbionts. Mar Biol 142: 57-
66.
Gros O, Liberge M, Heddi A, Khatchadourian C, Felbeck H (2003b). Detection of the free-
living forms of sulfide-oxidizing gill endosymbionts in the lucinid habitat (Thalassia
testudinum environment). Appl Environ Microbiol 69: 6264-7.
Guindon S, Gascuel O (2003). A simple, fast, and accurate algorithm to estimate large
phylogenies by maximum likelihood. Syst Biol 52: 696-704.
Holmes DE, Nevin KP, Lovley DR (2004). Comparison of 16S rRNA, nifD, recA, gyrB,
rpoB and fusA genes within the family Geobacteraceae fam. nov. Int J Syst Evol
Microbiol 54: 1591-9.
Ishaq M, Wolf B, Ritter C (1990). Large-scale isolation of plasmid DNA using
cetyltrimethylammonium bromide. Biotechniques 9: 19-20, 22, 24.
Keane TM, Creevey CJ, Pentony MM, Naughton TJ, McLnerney JO (2006). Assessment of
methods for amino acid matrix selection and their use on empirical data shows that ad
hoc assumptions for choice of matrix are not justified. BMC Evol Biol 6: 29.
Lane DJ (1991). 16S/23S Sequencing. In: Stackebrandt E GM (ed). Nucleic acid techniques
in bacterial systematics. John Wiley and Sons.: New York. pp 115-175.
Milne I, Wright F, Rowe G, Marshall DF, Husmeier D, McGuire G (2004). TOPALi:
software for automatic identification of recombinant sequences within DNA multiple
alignments. Bioinformatics 20: 1806-7.
Olson PD, Cribb TH, Tkach VV, Bray RA, Littlewood DT (2003). Phylogeny and
classification of the Digenea (Platyhelminthes: Trematoda). Int J Parasitol 33: 733-55.
Peek AS, Feldman RA, Lutz RA, Vrijenhoek RC (1998). Cospeciation of chemoautotrophic
bacteria and deep sea clams. Proc Natl Acad Sci USA 95: 9962-6.
Piednoel M, Bonnivard E (2009). DIRS1-like retrotransposons are widely distributed among
Decapoda and are particularly present in hydrothermal vent organisms. BMC
Evolutionary Biology 9: 86.
Polz MF, Cavanaugh CM (1995). Dominance of one bacterial phylotype at a Mid-Atlantic
Ridge hydrothermal vent site. Proc Natl Acad Sci USA 92: 7232-6.
Polz MF, Distel DL, Zarda B, Amann R, Felbeck H, Ott JA et al (1994). Phylogenetic
analysis of a highly specific association between ectosymbiotic, sulfur-oxidizing
bacteria and a marine nematode. Appl Environ Microbiol 60: 4461-7.
Ronquist F, Huelsenbeck JP (2003). MrBayes 3: Bayesian phylogenetic inference under
mixed models. Bioinformatics 19: 1572-4.
Rozas J, Sanchez-DelBarrio JC, Messeguer X, Rozas R (2003). DnaSP, DNA polymorphism
analyses by the coalescent and other methods. Bioinformatics 19: 2496-7.
Rozen S, Skaletsky H (2000). Primer3 on the WWW for general users and for biologist
programmers. Methods Mol Biol 132: 365-86.

Schwarz G (1978). Estimating the Dimension of a Model. Ann. Statist. 6: 461-464.
Stewart FJ, Newton IL, Cavanaugh CM (2005). Chemosynthetic endosymbioses: adaptations
to oxic-anoxic interfaces. Trends Microbiol 13: 439-48.
Tanabe Y, Kasai F, Watanabe MM (2007). Multilocus sequence typing (MLST) reveals high
genetic diversity and clonal population structure of the toxic cyanobacterium
Microcystis aeruginosa. Microbiology 153: 3695-703.
Thompson JD, Higgins DG, Gibson TJ (1994). CLUSTAL W: improving the sensitivity of
progressive multiple sequence alignment through sequence weighting, position-specific
gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673-80.
Van Valen L (1973). A new evolutionary law. Evol. Theory 1: 1-30.
Williams ST, Taylor JD, Glover EA (2003). Molecular Phylogeny of the Lucinoidea
(Bivalvia): Non-Monophyly and Separate Acquisition of Bacterial Chemosymbiosis. J
Molluscan Stud 70: 187-202.
Winnepenninckx BM, Reid DG, Backeljau T (1998). Performance of 18S rRNA in littorinid
phylogeny (Gastropoda: Caenogastropoda). J Mol Evol 47: 586-96.
Woese CR (1987). Bacterial evolution. Microbiol Rev 51: 221-71.
Won YJ, Hallam SJ, O'Mullan GD, Pan IL, Buck KR, Vrijenhoek RC (2003). Environmental
acquisition of thiotrophic endosymbionts by deep-sea mussels of the genus
bathymodiolus. Appl Environ Microbiol 69: 6785-92.
Woyke T, Teeling H, Ivanova NN, Huntemann M, Richter M, Gloeckner FO et al (2006).
Symbiosis insights through metagenomic analysis of a microbial consortium. Nature
443: 950-5.
Zhang DX, Hewitt GM (2003). Nuclear DNA analyses in genetic studies of populations:
practice, problems and prospects. Mol Ecol 12: 563-84.
Zhou J, Bruns MA, Tiedje JM (1996). DNA recovery from soils of diverse composition. Appl
Environ Microbiol 62: 316-22.

Title and legend to figures
Figure Legends
Figure 1. Molecular phylogeny of Lucinidae realized using concatenated sequences from
18S and 28S rRNA genes (1391bp). Tree was reconstructed using Maximum Likelihood
(ML) and Bayesian Inference (BI) methods under TrNef+Γ (ML) and K80+ Γ (BI)
evolutionary models with Crassatellidae and Thyasiridae as outgroups. For robustness of the
clades, only bootstrap values (ML, 100 replicates [under branchs]) and Posterior Probabilities
(BI [above branchs]) up to 80% were displayed. Sequences obtained in this study are in bold
and other sequences provide from Williams et al. (2003). aOur sequences are identical to
those published by Willams et al. (2003).
Figure 2. Bacterial haplotypes networks realized using sequences from MLST genes.
Networks were reconstructed using Median Joining method. Circles represent haplotypes (H1
to H6) and size of them is proportional to number of sequences for each haplotype. Colors of
circle represent the host species harboring the bacterial sequences: blue for C. orbicularis,
green for L. pensylvanica, red for C. orbiculata, yellow for C. pectinella, purple for A. alba
and black for D. quadrisulcata. Each bar on branches represents one substitution.
Figure 3. CARD-FISH on slide with symco2A probe on Codakia orbiculata starved
individuals after addition of purified symbionts from C.orbicularis or C. orbiculata. (A)
Gill section of freshly collected C. orbiculata (CARD-FISH positive control). (B) Gill section
of C. orbiculata after 9 months of starvation (negative control). (C and D) Gill sections of
starved C. orbiculata after addition of C. orbicularis gill extract during 4 days (C) or 8 days
(D). (E and F) Gill sections of starved C. orbiculata after addition of C. orbiculata gill extract
during 4 days (E) or 8 days (F). Scale bars=10 µm. Ciliated zone (CZ). Lateral zone (LZ).
Arrows represent marked symbionts.
Figure 4. Hypothetical distributions of symbionts in Thalassia testudinum sea-grass beds
from Îlet Cochon (Guadeloupe). (A) Bacterial population structured in function of oxygen
(O2), sulfur (H2S) or other gradient. (B) Non structured bacterial population. Circles
correspond to hosts and triangle to symbionts. Colors correspond to host species (circles) or
bacterial strains (triangles): blue for C. orbicularis, Pale blue for juvenile of C. orbicularis,
yellow for C. orbiculata, red for C. pectinella and purple for A. alba.
Figure 5. Schematic representation of evolutive process which could lead to the pattern
of structuration observed in Thalassia testudinum sea-grass bed of Îlet Cochon. This
scheme shows at the final step the structured observed pattern with MLST analysis
(Association between A. alba and its strain in purple, between C. orbicularis and its one in
blue and association implying C. orbiculata and C. pectinella). Circles represent bacterial
pools. Squares, crosses, triangles and diamonds represent host species. Colors discriminates
specifically established association. Double arrows represent established association. Dotted
double arrows represent host species where individuals exploit different bacterial pools.
Dashed arrow represent apparition of bacterial mutant. Dashed/dotted arrow represent
apparition of host which are able to exploit escaped mutants.

Figures





Tables
Table 1. Polymorphism analysis data.
1 gene names. dnaE: DNA Polymerase III α-subunit, gyrB: DNA Gyrase β-subunit, ITS1: 16S/23S Internal
Transcribed spacer, aprA: Adenyly1,5-bisPhosphate Redctase α-subunit, cbbL: Ribulose-1,5-bisPhosphate
Carboxylase/Oxygenase Large subunit. 2 Percentages represent number of polymorphic sites in relation to the length of the considered fragment.
Gene 1
Sequences Polymorphic
sites 2
Haplotypes Accession
numbers Nb Length (bp)
dnaE (~3,5 kb) 18 465 6 (1.29 %) 5 To be submitted
gyrB (~2,4 kb) 18 806 6 (0.74 %) 5 To be submitted
ITS1 (~550 pb) 18 512 11 (2.14 %) 5 To be submitted
aprA (~1 kb) 18 492 8 (1.62 %) 6 To be submitted
cbbL (~1,5 kb) 18 600 11 (1.83 %) 6 To be submitted
TOTAL 90 2875 42 (1.46 %)

Table 2. Primers and PCR conditions used for each gene analyzed.
Primers PCR Conditions
Name Séquence(5'->3') Expected
size (bp)
Targeted
gene
Optimal
Tm (°C) Cycles Ref
2
Host
18S-5'f CTGGTTGATYCTGCCAGT 1000
18S
rRNA 54 30
1
18S-1100r CTTCGAACCTCTGACTTTGG 2
28S_LSU-900f CCGTCTTGAAACACGGACCAAG 700
28S
rRNA 52 40
3
28S_LSU-1600r AGCGCCATCCATTTTCAGG 2
Sym
bio
nt
1
16Suniv-27f AGAGTITGATCMTGGCTCAG 1465 rrs 54 30 4
16Suniv-1492r TACGGYTACCRRGTTACGACIT
ITS-degN-1242f GCTACACACGTGCTACAATGG
~1000
ITS1
54 35
5 ITS-degN-196r TCTCGGTTGATTTCTTTTCTT
ITS-Symb-282f GGAGTGGGTTGXAAAAGAAG 712 58 30
ITS-Symb-954r ATCGCAAGCTACAACGTCCT
dnaE-nested-1069f CCDGTDGGICCDGGICGYGGC
1171
dnaE
46 35
5
dnaE-nested-2279r TGNGATTTRTTRAANCCRTA
dnaE-degN-1180f CCIGARCGKGTITCIATGCC 857 60 35
dnaeE-degN-2076r YTGCATNACYTGYTCYTGRTA
dnaE-Symb-201f GAAGATGATCCCCTTCGAGA 667 64 30
dnaE-Symb-810r GTAGGAGACCTTGGCACGAC
gyrB-nested-217f GHAARCGYCCDGGYATGTA
1409
gyrB
56 35
5
gyrB-nested-1809r CVACRTCVGCATCBGTCAT
gyrB-degN-277f GGTDTTYGARRTDGTDGATAA 1130 56 35
gyrB-degN-1590r CHGCNGARTCMCCYTCMAC
gyrB-Symb-185f AGCCACCCGAAGAGAAGAA 932 58 30
gyrB-Symb-1150r TGGCAGACCGGCTATGT
CBBL-1B CACCTGGACCACVGTBTGG 800 cbbL 60 30 6
CBBL-2C CGGTGYATGTGCAGCAGCATGCC
G
aprA-Symb-23f 3
GCAACGACCTGATGGGTATC 532 aprA 58 33 5
aprA-Symb-555r 3
GTCGGTACGAAGCGGTTTT 1 “nested” and “degN” primers correspond to degenerated primers used in nested-PCR. “Symb” primers
correspond to specific primers designed using sequences obtained by nested-NCR. 2 References: 1 (Winnepenninckx et al., 1998), 2 (Williams et al., 2003), 3 (Olson et al., 2003), 4 (Lane, 1991),
5 this study, 6 (Elsaied and Naganuma, 2001). 3 These primers were designed using specific sequence for symbiont Lucinidae obtained using method derivate
from TE-walking (Piednoel and Bonnivard, 2009) from a “core sequence” amplified using APS-FW and APS-
RV primers (Deplancke et al., 2000).

ARTICLE 3 : BRISSAC ET AL.
Soumis à FEMS Microbiol. Ecol.


Only one majority sulfur-oxidizing symbiotic species occurs within
Lucinidae (Bivalvia).
Terry Brissac1*
, Hervé Merçot1 and Olivier Gros
2
1UMR 7138 UPMC-CNRS-MNHN-IRD « Systématique, Adaptation, Évolution », Equipe:
« Génétique & Evolution », Université Pierre et Marie Curie, Bat. A, 4ème
étage, 7 quai St
Bernard, 75005 Paris, France.
2 UMR 7138 UPMC-CNRS-MNHN-IRD « Systématique, Adaptation, Évolution », Equipe:
« Symbiose », Département de Biologie, Université des Antilles et de la Guyane B.P. 592,
97159 Pointe-à-Pitre Cedex, Guadeloupe (France).
Running title: Phylogenetic comparison of host and symbionts within Lucinidae
Keywords: Bivalvia; Comparative molecular phylogeny; EFTEM analysis; EELS; ESI;
Hemoglobin.

Introduction
Associations between chemoautotrophic bacteria and marine invertebrates were discovered in
hydrothermal vents by Felbeck et al. (1981). Later, they were described in shallow water
sediments (Berg & Alatalo, 1984; Schweimanns & Felbeck, 1985), cold seeps (Juniper &
Sibuet, 1987) whales bones (Deming et al., 1997) and sunken woods (Gros & Gaill, 2007).
This type of associations is present in several host taxa and implies various bacterial clades
and metabolisms (Dubilier et al., 2008). Among these associations, one of them concerns
Lucinidae (Bivalvia) which constitutes the most diverse bivalve family, live in a large variety
of marine habitats and are very widespread (Taylor & Glover, 2005). To date, all Lucinidae
genera studied harbor sulfur-oxidizing gamma-proteobacterial symbionts (according to 16S
rRNA sequences) localized in specialized cells, distributed throughout the lateral zone of gill
filaments, named bacteriocytes (Fisher, 1990).
Within Lucinidae, the capacity to establish symbiotic relationship seems to be an ancestral
heritage. In fact, it seems that symbiosis was acquired near -250 MY before divergence of the
different families (Thyasiridae, Lucinidae, Ungulinidae, Fimbriidae) belonging the Lucinacea
super-family (Newell, 1969; Boss, 1970). Thus, such symbiotic associations led Reid (1990)
to consider that the onset of symbiosis was the most important factor in the subsequent
evolution of the Lucinacea. Nevertheless, origin and evolutionary story of current symbiotic
couples remain obscure and ambiguous (Distel et al., 1994; Durand & Gros, 1996). In fact,
the two partners of actual couples could have coevolved or not from an ancestral symbiotic
couple. One way to distinguish between these alternatives is the comparison of host and
symbionts molecular phylogenies (Peek et al., 1998b). In fact in case of co-evolution, the two
partners must share a strong common evolutionary history and the two phylogenies must be
congruent.
Separate molecular data concerning sequences of host or symbiont has already been provided
in the literature for Lucinidae but host/symbiont sequences direct comparison has not been
realized yet for this bivalve family. On one side, molecular data concerning hosts have
already been used in order to investigate relation between Lucinoidea (Williams et al., 2003).
On the other side, 16S rRNA sequences were analyzed in order to study diversity of these
symbioses (Distel et al., 1988; Distel et al., 1994; Durand & Gros, 1996; Durand et al., 1996).
Related to hosts, Distel et al. (1988; 1994) showed specifically associated unique bacterial
ribotype per host species. Later, Durand & Gros (1996), Durand et al. (1996), Gros et al.
(2000; 2003) observed 6 host species which harbor the same symbiont species (100% of
identity on 16S rRNA sequences). If under the first observations it seems that there are
species-specific associations due to co-evolution of lucinid clams and their symbionts, ulterior
studies can not support this hypothesis. However in these studies, molecular diversity of
symbionts is compared to host morphological taxonomy and not to host molecular phylogeny.
Moreover, sampling strategies were very different, Distel et al. (1988; 1994) sampling (which
studied specimen from various ecosystem and geographic localization) being more
heterogeneous than Durand & Gros (1996), Durand et al. (1996), Gros et al. (2000; 2003)
one, which studied Lucinidae specimen from Thalassia testudinum sea-grass beds of Lesser
Antilles.
In this paper we study from a molecular point of view, Lucinidae and their bacterial
symbionts from Philippines (with additional data already published providing from various
area), in order to study evolutionary history of symbiotic current couples. Thus, we realize the

comparison of molecular phylogenies of diverse Lucinidae hosts (using 18S rRNA and
28SrRNA) and their associated symbionts (using 16S rRNA and APS Reductase [aprA]
sequences). In addition, according to the particular way of life of Lucinidae (i.e.
chemosymbiois) we conduct on Philippines samples ultrastructural (gills and symbionts) and
histochemical (occurrence of hemoglobin) analyses.
Material and Methods
Sampling
Philippians clams: Samples (Table 1) were collected during the Panglao cruise in May 2005
(chief scientist: P. Bouchet) in the Bohol sea [8,3-9,6° N and 123-124° E] using a beam trawl
at depths ranging between 219 and 1775 meters. Samples were processed on the ship within 1
hour after collection. For each lucinid specimen, the gills were dissected and treated according
to the following protocol: First gill piece was stored in 100% ethanol for molecular
investigation of bacterial diversity and host phylogeny (see below). Second gill piece was
prepared for electron microscopy. Codakia orbicularis and Lucina pectinata were sampled by
hand respectively in Thalassia testudinum sea-grass beds of “Ilet Cochon” (Guadeloupe) and
mangrove swamps of “La manche à eau” (Guadeloupe) in 2006.
Transmission Electron Microscopy (TEM) preparation
Gills were prefixed on ship for one hour at 4°C in 2.5% glutaraldehyde in 0.1M pH 7.2
caccodylate buffer, adjusted to 900 mOsM with NaCl and CaCl2 in order to improve
membrane preservation. Gills were briefly rinsed in the same buffer before to be stored in the
same buffer at 4°C until they were brought to the laboratory 2-3 weeks after. Samples were
fixed for 45 minutes at room temperature in 1% osmium tetroxide in the same buffer, then
rinsed in distilled water and post-fixed with 2% aqueous uranyl acetate for one hour before
embedding, sectioning (60nm thick), and observation with a Leo-912 microscope at 80kV.
EFTEM analysis: PEELS and ESI procedures
The EFTEM observations were performed using a LEO 912 Omega transmission electron
microscope (LEO Electron Optics GmbH, Oberkochen, Germany) at 120 kV. Acquisition was
accomplished with the ESIvision program (versions 3. 0 Soft-Imaging Software, SIS, GmbH,
D-48153 Münster). For PEELS acquisition, the primary magnification was set to a small area
delimited by the entrance aperture of the spectrometer. For ESI acquisition and to minimize
the radiation damage we used the three-window method (Jeanguillaume et al., 1978; Reimer
et al., 1991), as described in (Lechaire et al., 2008).
DNA template preparation
Template DNA for PCR amplifications was extracted from symbiont-containing gills of each
bivalve hosts according the CTAB (CetylTrimethyl Ammonium Bromide) method (Ishaq et
al., 1990). DNA concentration was determined by spectrometry (BioPhotometer, eppendorf)
and on a 1% agarose gel in comparison with a concentration marker (Smart Ladder,
Eurogentec).

PCR amplifications
Bacterial (16S rRNA and APS reductase) or host (18S rRNA and 28S rRNA) genes were
amplified from host-symbiont DNA mixture using primers presented in Table 2.
Amplification was performed in 50µL using between 25 to 50ng of DNA template, 5µL of
10X Taq buffer (Invitrogen), 3mM MgCl2 (Invitrogen), 0.4mM of each dNTP, 1mM of each
primer and 1 units of recombinant Taq DNA polymerase (Invitrogen). Target DNA were
amplified in 30 cycles (94°C for 1min, optimal Tm for 1 min [Table 2], 72°C for 1 min [1 min
30 sec for 16S rRNA]) with an initial denaturation step at 94°C for 10 min and a final
elongation step at 72°C for 10 min. PCR products were controlled on a 1% agarose gel and
directly sequenced using ABI Prism automated sequencer (GATC-biotech, http://www.gatc-
biotech.fr)
Phylogenetic analyses
Sequences were controlled by BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and aligned
with ClustalW (Thompson et al., 1994) with default parameters. The alignment was then eye-
checked and corrected if necessary. Gap positions and ambiguously sites of the alignment
were removed using Gblocks 0.91b (Castresana, 2000). Alignment was then imported in the
TOPALi v2.5 software (Milne et al., 2004) in order to define using ModelGenerator (Keane
et al., 2006) the substitution model which will be used for each dataset. The best model for
each dataset is chosen using Bayesian Information Criterion (Schwarz, 1978). Phylogenetic
analyses were then performed with TOPALi v2.5, trees being then estimated by Maximum
Likelihood (ML) using PhyML (Guindon & Gascuel, 2003) and by Bayesian Inference (BI)
using MrBayes 3 (Ronquist & Huelsenbeck, 2003). Support for individual clades was
evaluated using non-parametric bootstrapping (Felsenstein, 1985) obtain from 100 replicates
for ML. Posterior probabilities for BI were also evaluated for each node.
Results
Symbiont phylogeny
All PCR amplifications of 16S rRNA or aprA sequences gave fragments of the expected size
(~1500bp and ~300bp respectively). Direct PCR product sequencing and chromatograph
analysis indicate that for each amplification only one single detectable sequence. Symbionts
phylogenetic trees (Fig. 1 and 2) were realized using 1263 sites for 16S rRNA and 273 sites
for aprA. For 16S rRNA (Fig. 1), two free-living marine gamma-proteobacteria were used as
outgroup. We observe that 3 different bacterial species (16S rRNA identity inferior to 97%)
occur within Lucinidae (Fig. 1, clades A and B and symbiont of L.pectinata). This tree
permits to place symbionts sequences of Philippines clams in a monophyletic clade which
correspond to “Lucinoidea symbionts”. According to this tree and based on pairwise identity
matrix (Table 3), it seems that all symbionts analyzed here belong to the same bacterial
species (Fig.1, clade B) 16S rRNA identity being superior to 97% (97.7%). Moreover within
this defined species we can distinguish two different bacterial types harbored by Philippines
clams. The first one is shared by OG18, OG98 and OG108 and seem correspond exactly to
the same bacterial type (mean 16S rRNA sequence identity of 99.9%) and the second one is
present in OG57, OG101 and OG103 (98.6%). Thus, our 6 Philippines lucinid clams seem
share 2 different variants of the same bacterial species. Sulfur-oxidizing metabolism of these
symbionts harbored by Philippines clams is confirmed by the positive detection by PCR of

this gene. According to APS Reductase phylogenetic tree (Fig. 2) these symbiont sequences
fall in the lineage II of aprA in the sulfur-oxidizing bacteria clade. Unfortunately tissues
conservation not permits obtaining of aprA sequence for the specimen OG103.
Host Phylogeny
For all amplifications, only one fragment of the expected size was obtained for 18S rRNA and
28S rRNA (~1000bp and ~700bp respectively). These fragments gave unique sequences by
sample analyzed. We use as outgroup, Thyasiroidea and Crassatelloidea which were sister-
groups of Lucinidae. Host phylogeny realized with concatenation of sequences of these two
genes using 1252 sites (Fig. 3) permits to place these samples in the Lucinidae clades
described by Williams et al. (2003). Except for clade A, samples analyzed here were
distributed in all diverse described clades. So, OG108 correspond to Myrtea species, OG101
to the clade B and finally OG98 and OG57 belong to the Anodontia clade. For OG18, 18S
rRNA has not been obtained, nevertheless 28S rRNA sequence of this specimen fall in Clade
B of a 28S rRNA tree (data not shown). No sequences have been obtained for OG103. For C.
orbicularis and L. pectinata, sequences obtained here correspond to those published by
Williams et al., (2003)
Host/Symbiont phylogenies comparison
Comparing host and symbiont phylogenies they appear totally incongruent, topologies (i.e.
branching order) and time of splitting events (i.e. branch length) being different. Thus, closer
relative host species do not harbor close relative (or same) bacterial type (e.g. case of OG98
and OG57). Moreover, OG57 and OG101 which are different host species seem harboring the
same bacterial type. More interestingly, the 3 host species which harbor the same bacterial
strain (OG18, OG98 and OG108) belong to 3 different host lineages (clade B, Anodontia
clade and Myrtea clade respectively). So it appears that there is no species specific association
within Lucinidae.
Special features on ctenidial cells of the Philippines lucinids analyzed.
All analyzed individuals present ctenidial characteristics common within Lucinidae like
described by Frenkiel & Mouëza (1995). However, two individuals have displayed some
particularities. In OG18, bacteriocytes contained numerous dark granules inside the cytoplasm
(Fig.x) that probably are pigments but whose nature is unknown. In OG103, the dark aspect of
the cytoplasm with finely granular electron-dense areas suggests that intracellular hemoglobin
is present inside bacteriocytes. This was confirmed by PEELS analysis in which iron was
successfully detected in the cytoplasm of each bacteriocyte tested with a specific iron
spectrum (edge closed to 730 eV) obtained after background subtraction (Fig. xx). The ESI
method was also performed in order to image the location of the iron in the bacteriocte’s
cytoplasm. When the Fe-distribution image was superimposed onto the high-contrast image
(HCI) recorded at 250 eV (Mixmap image, Fig. xxx), iron was observed localized inside the
cytoplasm of the bacteriocyte while bacteria remain free of iron (Fig. xxx). So, iron is present
and distributed homogenously within the cytoplasm of bacteriocytes. The presence of sulfur
in the same area analyzed was tested by the PEELS method and no significant signal was
obtained (data not shown).

Ultrastructure of the bacterial gill-endosymbionts of Philippines clams.
Bacteria are individually enclosed within vacuoles. These endosymbionts are rod-shaped with
the characteristic double membrane of gram negative bacteria belonging to the γ-
proteobacteria. In OG57 and OG101 bacteria are large (4µm long x 1.5µm large), in OG18
they are longer and thinner (6µm long x 0.5µm large) and in OG98 and OG103 the two
categories are retrieved. In all cases bacteria can be big or small. Some ovoid shaped figures
can be observed but are probably due to the section orientation. Symbionts are generally
abundant per bacteriocyte excepted in OG103. Bacterial cytoplasm contains numerous non-
membrane-bound irregular inclusions (up to 50nm in diameter) appearing as black dots in
TEM sections that can be considered as polyhydroxybutyrate storage granules. Electron
lucent granules (sulfur granules) are not obvious in these gill-endosymbionts contrary to
bacterial symbionts encountered in the gills of tropical shallow water lucinids as C.
orbicularis (Lechaire et al., 2008).
Discussion
To date, diversity of host and symbiont within Lucinidae was investigated in studies with
antagonist conclusions. Thus, in Distel et al (1988; 1994) species-specific associations has
been shown being due to coevolution of the two partners. Later, associations between 6 host
species and only one bacterial species was described attesting of an association more
constituted in function of the bacteria present in the environment (Durand & Gros, 1996;
Durand et al., 1996; Gros et al., 2000; Gros et al., 2003). Nevertheless in these studies
molecular phylogeny of hosts has not been realized in comparison to symbionts one. In this
paper, we aim to study diversity of hosts and symbionts in order to bring a better
understanding of specificity of associations in this particular relationship realizing comparison
of host and symbiont molecular phylogenies.
Host/Symbiont couples relationship evolution.
We analyzed diversity of host using 18SrRNA and 28SrRNA sequences. Specimens from
Philippines analyzed here, belong to the Lucinidae monophyletic group (Fig. 3) as described
in Williams et al. (2003). Like in all lucinids described to date, Philippines clams harbor
symbiotic bacteria into their gills. According to 16S rRNA phylogeny these symbionts belong
to a well supported monophyletic clade formed by already described Lucinoidea symbionts
(Fig. 1). Moreover, PCR detection in our specimens of APS Reductase gene permits to attest
that these symbionts possess sulfur-oxidizing pathway even if sulfur or sulfur granules has not
been detected on gill sections by microscopy analyses. Concerning diversity of symbionts
analyzed in this study, it seems that they belong to the same bacterial species (Fig. 1, clade
C). This point is confirmed by sequence identity matrix showing that mean 16S rRNA
sequence identity is superior to 97% in this clade. Thus, as described in Durand & Gros
(1996), different host species could harbor the same bacteria. So, this result can not support
the hypothesis that within Lucinidae, the associations are species-specific due to coevolution
of the two partners. Thus, within Lucinidae, it seems to us that according to the acquisition
mode of the symbiont (i.e. acquisition from a free-living bacteria present in the surroundings),
the association takes place in function of the bacterial species present in the environment.
Thereby, the specificity of association described in Distel et al. (1988; 1994) could be due to
the heterogeneity of the sampling, ecosystems and geographic locations being very different

mimic specific associations. However how explain that within Lucinidae, so geographical
diverse hosts could harbor a unique bacterial species? In fact, our specimens provide from
Philippines, other like Lucinoma aff. kazanii, Lucina floridana, Lucinoma aequizonata or C.
orbicularis provide from various geographic locations (respectively Mediterranean sea,
Florida, California and Caraibe Distel et al., 1994; Durand & Gros, 1996; Duperron et al.,
2007). Two hypotheses could explain this low level of genetic diversity between these
symbionts: (i) geographic divergence of symbiotic populations too recent to permit
observation of genetic divergence, (ii) low evolution rate of the consider marker (16S rRNA).
Finally we privilege the second hypothesis because Peek et al. (1998a) described 16S rRNA
sequences of environmentally acquired symbionts (as is the case for lucinid symbionts)
evolve more slowly than vertically transmitted symbionts due to lack of bottleneck during
passage from generation to generation in associations without vertical transmitted symbionts.
How bacterial species occur within Lucinidae?
Even if, it appears that there is no species-specific association within Lucinidae, there is more
than one bacterial species associated with Lucinidae. The majority bacterial species belong to
the clade B (Fig. 1) but we can also observe 2 other species harbored respectively by Lucina
pectinata and by host species which belong to the anodontia clade except A. alba (Fig. 1,
clade A). Interestingly, these bacterial species seem correspond to symbionts present in hosts
which inhabit high sulfide environments (e.g. mangrove swamps) in comparison with shallow
water sea-grass beds or reducing sediments. This distinction between this different bacterial
species supposes that bacterial species could be different according to sulfur level of different
habitats. So this point reinforce the fact that lucind clams do not co-evolved strictly with their
symbiont but rather be associated with the symbiosis-competent bacteria present in their
environment.
Does multi-infection occur within Lucinidae?
However, in the unique bacterial species observed in Philippine clams (Fig. 1 Clade B),
according to symbiont phylogeny we can distinguish 2 different groups (Fig. 1, groups I and
II). These two groups seem correspond to the two bacterial morphotypes viewed in TEM
analysis. Thus, the large morphotype (observed in OG57 and OG101) corresponds to the
group I and the longer and thinner one (observed in OG18) corresponds to the group II.
Interestingly, for OG98 and OG103 only one bacterial sequence was detected even if the two
morphotypes are present in their gills. So, in these specimens multi-infection of two different
bacterial types could occur and could be not detected due to the technique employed here
which permit only visualization of the majority sequence (i.e. PCR direct sequencing). A
more concerted effort concerning these two lucinids (OG98 and OG103) including the
analysis of a 16S rDNA clone library issued from gill DNA and fluorescent in situ
hybridization analysis of the symbionts will be necessary to confirm the hypothesis of dual
symbiosis in such lucinid. Nevertheless, recently, a multiple symbiosis partnership was
suspected in the lucinid A. ovum according to TEM observations but the authors failed to
confirm these ultrastructural observations by molecular data as only a single 16S rDNA
sequence was obtained from 62 clones analyzed (Ball et al., 2009). However, the presence for
a same morphotype of both small and big bacterial inside a same bacteriocyte could be related
to the DNA content of new phagocytozed symbionts and old ones. Recently, Caro et al.
(2007) have shown using single-cell approaches including flow cytometry (FCM) that the
sulfur-oxidizing gill endosymbiont population of C. orbicularis was heterogeneous ranging
from one copy of their genome for the smaller cells to more than four copies for the larger
cells and even up to 20 copies when the host was kept in starvation. This multiplication of the

genomic DNA content while cell division is forbidden by the host could explain the
heterogeneous size of a single bacterial species inside a same bacteriocyte. Thus, recently
recruited symbionts (the small bacterial symbionts) are located near the apical pole and
abundantly supplied with growth substrates by the host, while their division is inhibited. As
new symbionts are recruited, the older ones migrate toward the basal pole becoming larger
with the increase of their genome.
Hemoglobin, sulfur and symbiosis
According to TEM analysis, there is no evidence of sulfur granules in gill-endosymbionts of
lucinids from Philippines conversely to those described for example in symbionts of C.
orbicularis (Lechaire et al., 2008). This is surprising because these symbionts are sulfur-
oxidizing as attest the detection of APS Reductase gene. Nevertheless, no hypothesis could
explain this particularity. Another interesting feature concerns the specimen Myrtea
flabelliformis (OG103) which present a dark cytoplasm and low number of gill-
endosymbionts. The dark aspect of bacteriocytes cytoplasm suggests the presence of
hemoglobin in gill tissues of this host which has been confirmed by PEELS and ESI analyses.
Hemoglobin, which is an oxygen or hydrogen sulfide transporter (Kraus, 1993) is scarce
within Lucinidae even if it has been described in gill tissues of some species like Anodontia
edentulata (Lebata, 2001), L. pectinata (Kraus & Wittenberg, 1990; Gavira et al., 2008) or
Myrtea spinifera (Dando et al., 1985) and suggested in Anodontia ovum (Ball et al., 2009). It
has been proposed in the literature that hemoglobin contribute to detoxification against sulfur
(which is toxic for respiratory chain and could harm tissues) in high sulfide environments as
hydrothermal vents (Arndt et al., 2001). This role of hemoglobin in detoxification has also
been suggested for the shallow water Lucinidae L. pectinata conjugated with the presence of
sulfide oxidizing bodies (Liberge et al., 2001). However, sulfur level environment of our M.
flabelliformis specimen is unknown. In addition, hemoglobin has also been described in
another species of the genus Myrtea (M. spinifera) which inhabit low sulfide environment
(Dando et al., 1985). Finally, there is no evidence of sulfur-oxidizing bodies in gill sections of
M. flabelliformis. Thus, detoxification role of hemoglobin against sulfur could not be clearly
attested for this specimen. Which other hypothesis could be posed to explain the high
hemoglobin concentration in cytoplasm of M. flabelliformis? According to Dando et al.,
(1985) high affinity of hemoglobin for oxygen could give some evidence of its role. Indeed,
hemoglobin could act as oxygen storage during burrowing or protects sulfur-oxidizing
endosymbionts against excess of oxygen, sulfur-oxidizing bacteria being frequently
mircoaerophile (Jorgensen, 1982). However, in our case there is no evidence which permit to
distinguish between these two hypotheses.
According to our results on the 6 Lucinidae species from Philippines compared to literature
data, it appears obviously that Lucinidae and their symbionts have not co-evolved from an
ancestral symbiosis. In fact, the association seems to be more constituted in function of the
bacterial type present in the environment. Moreover, according to 16S rRNA Lucinidae
appear to be associated in majority to a same bacterial sulfur-oxidizing species. Particularly
cases corresponding to bacterial species associated to hosts which inhabit high sulfur
environments (A. ovum, A. phillipiana and L. pectinata).
Acknowledgments

The material from the Philippines was obtained during the PANGLAO 2005 deep-sea cruise
aboard M/V DA-BFAR and we thank Dr P. Bouchet and L. Labe, the co-principal
investigators, for their invitation. Our participation was made possible from a MNHN grant
under the program "Status and Phylogenetic Structure of Recent and Fossil Biodiversity" led
by Dr P. Janvier. Labwork was performed within the framework of the DIWOOD project. We
thank Dominique Higuet for their fruitful comments and Denis Poinsot for English revision.
T.B. is funded as a PhD student by a MENRT (Ministère de l’Education Nationale, de la
Recherche et des Technologies) grant.
References
Arndt C, Gaill F & Felbeck H (2001) Anaerobic sulfur metabolism in thiotrophic symbioses.
J Exp Biol 204: 741-750.
Ball DJ, Purdy KJ, Glover EA & Taylor JD (2009) Ctenidial structure and three bacterial
symbiont morphotypes in Anodontia (Euanodontia) ovum (REEVE, 1850) from the
great barrier reef, AUSTRALIA (Bivalvia: Lucinidae). J molluscan Stud (in press).
Berg CJJ & Alatalo P (1984) Potential of chemosynthesis in molluscan mariculture.
Aquaculture 39: 165-179.
Boss KJ (1970) Fimbria and its lucinoid affinities (Mollusca; Bivalvia). Breviora 350: 1-16.
Caro A, Gros O, Got P, De Wit R & Troussellier M (2007) Characterization of the population
of the sulfur-oxidizing symbiont of Codakia orbicularis (Bivalvia, Lucinidae) by
single-cell analyses. Appl Environ Microbiol 73: 2101-2109.
Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in
phylogenetic analysis. Mol Biol Evol 17: 540-552.
Dando PR, Southward AJ, Southward EC, Terwilliger NB & Terwilliger RC (1985) Sulfur-
oxidizing bacteria and hemoglobin in gills of the bivalve mollusk Myrtea spinifera.
Marine Ecology-Progress Series 23: 85-98.
Deming JW, Reysenbach AL, Macko SA & Smith CR (1997) Evidence for the microbial
basis of a chemoautotrophic invertebrate community at a whale fall on the deep
seafloor: bone-colonizing bacteria and invertebrate endosymbionts. Microsc Res Tech
37: 162-170.
Deplancke B, Hristova KR, Oakley HA, McCracken VJ, Aminov R, Mackie RI & Gaskins
HR (2000) Molecular ecological analysis of the succession and diversity of sulfate-
reducing bacteria in the mouse gastrointestinal tract. Appl Environ Microbiol 66:
2166-2174.
Distel DL, Felbeck H & Cavanaugh CM (1994) Evidence for phylogenetic congruence among
sulfur-oxidizing chemoautotropihc becterial endosymbiont and their Bivalve hosts. J
Mol Evol 38: 533-542.
Distel DL, Lane DJ, Olsen GJ, Giovannoni SJ, Pace B, Pace NR, et al. (1988) Sulfur-
oxidizing bacterial endosymbionts: analysis of phylogeny and specificity by 16S
rRNA sequences. J Bacteriol 170: 2506-2510.
Dubilier N, Bergin C & Lott C (2008) Symbiotic diversity in marine animals: the art of
harnessing chemosynthesis. Nat Rev Microbiol 6: 725-740.
Duperron S, Fiala-Medioni A, Caprais JC, Olu K & Sibuet M (2007) Evidence for
chemoautotrophic symbiosis in a Mediterranean cold seep clam (Bivalvia: Lucinidae):
comparative sequence analysis of bacterial 16S rRNA, APS reductase and RubisCO
genes. FEMS Microbiol Ecol 59: 64-70.
Durand P & Gros O (1996) Bacterial host specificity of Lucinacea endosymbionts:
interspecific variation in 16S rRNA sequences. FEMS Microbiol Lett 140: 193-198.

Durand P, Gros O, Frenkiel L & Prieur D (1996) Phylogenetic characterization of sulfur-
oxidizing bacterial endosymbionts in three tropical Lucinidae by 16S rDNA sequence
analysis. Mol Mar Biol Biotechnol 5: 37-42.
Felbeck H, Childress JJ & Somero GN (1981) Calvin-Benson cycle and sulphide oxidation
enzymes in animals from sulphide-rich habitats. Nature 293: 291-293.
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap.
Evolution 39: 783-791.
Fisher CR (1990) Chemoautotrophic and methanotrophic symbioses in marine invertebrates.
Rev. Aquat. Sci 2: 399-613.
Frenkiel L & Mouëza M (1995) Gill ultrastructure and symbiotic bacteria in Codakia
orbicularis (Bivalvia: Lucinidae). Zoomorphology 115: 51-61.
Gavira JA, Camara-Artigas A, De Jesus-Bonilla W, Lopez-Garriga J, Lewis A, Pietri R, et al.
(2008) Structure and ligand selection of hemoglobin II from Lucina pectinata. J Biol
Chem 283: 9414-9423.
Gros O & Gaill F (2007) Extracellular bacterial association in gills of "wood mussels".
Cahiers De Biologie Marine 48: 103-109.
Gros O, Frenkiel L & Felbeck H (2000) Sulfur-oxidizing endosymbiosis in Divaricella
quadrisulcata (Bivalvia: Lucinidae): Morphological, ultrastructural, and phylogenetic
analysis. Symbiosis 29: 293-317.
Gros O, Liberge M & Felbeck H (2003) Interspecific infection of aposymbiotic juveniles of
Codakia orbicularis by various tropical lucinid gill-endosymbionts. Mar Biol 142: 57-
66.
Guindon S & Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large
phylogenies by maximum likelihood. Syst Biol 52: 696-704.
Ishaq M, Wolf B & Ritter C (1990) Large-scale isolation of plasmid DNA using
cetyltrimethylammonium bromide. Biotechniques 9: 19-20, 22, 24.
Jeanguillaume C, Trebbia P & Colliex C (1978) About the use of electron energy-loss
spectroscopy for chemical mapping of thin foils with high spatial resolution.
Ultramicroscopy 3: 237-242.
Jorgensen BB (1982) Ecology of the bacteria of the sulphur cycle with special reference to
anoxic-oxic interface environments. Philos Trans R Soc Lond B Biol Sci 298: 543-561.
Juniper SK & Sibuet M (1987) Cold seep benthic communities in Japan subduction zones:
spatial organization, trophic strategies and evdence for temporal evolution. Mar Ecol
Prog Ser 40: 115-126.
Keane TM, Creevey CJ, Pentony MM, Naughton TJ & McLnerney JO (2006) Assessment of
methods for amino acid matrix selection and their use on empirical data shows that ad
hoc assumptions for choice of matrix are not justified. BMC Evol Biol 6: 29.
Kraus DW (1993) Heme-proteins in sulfide-oxidizing bacteria mollusk symbioses. 112-120.
Amer Soc Z. Los Angeles, Ca.
Kraus DW & Wittenberg JB (1990) Hemoglobins of the Lucina pectinata/bacteria symbiosis.
I. Molecular properties, kinetics and equilibria of reactions with ligands. J Biol Chem
265: 16043-16053.
Lane DJ (1991) 16S/23S Sequencing. Nucleic acid techniques in bacterial systematics.John
Wiley and Sons., pp. 115-175. New York.
Lebata MJ (2001) Oxygen, sulphide and nutrient uptake of the mangrove mud clam
Anodontia edentula (Family: Lucinidae). Mar Pollut Bull 42: 1133-1138.
Lechaire J-P, Freibourg G, Gaill F & Gros O (2008) In situ characterization of sulfur in gill-
endosymbionts of the shallow water lucinid Codakia orbicularis (Linné, 1758) by
cryo-EFTEM microanalysis. Mar Biol 154: 693-700.

Liberge M, Gros O & Frenkiel L (2001) Lysosomes and sulfide-oxidizing bodies in the
bacteriocytes of Lucina pectinata, a cytochemical and microanalysis approach. Mar
Biol 139: 401-409.
Milne I, Wright F, Rowe G, Marshall DF, Husmeier D & McGuire G (2004) TOPALi:
software for automatic identification of recombinant sequences within DNA multiple
alignments. Bioinformatics 20: 1806-1807.
Newell ND (1969) Classification of Bivalvia. In:Treatise on invertebrates paleontology. The
Geological Society of America and the University of Kansas, Vol. 1 pp. N205-N224.
Olson PD, Cribb TH, Tkach VV, Bray RA & Littlewood DT (2003) Phylogeny and
classification of the Digenea (Platyhelminthes: Trematoda). Int J Parasitol 33: 733-
755.
Peek AS, Vrijenhoek RC & Gaut BS (1998a) Accelerated evolutionary rate in sulfur-
oxidizing endosymbiotic bacteria associated with the mode of symbiont transmission.
Mol Biol Evol 15: 1514-1523.
Peek AS, Feldman RA, Lutz RA & Vrijenhoek RC (1998b) Cospeciation of
chemoautotrophic bacteria and deep sea clams. Proc Natl Acad Sci USA 95: 9962-
9966.
Reid RGB (1990) Evolutionary implications of sulphide-oxidising symbioses in bivalves. In:
Morton, B. (ed.) The Bivalvia — Proceedings of a Memorial Symposium in Honour of
Sir Charles Maruice Yonge, Edinburgh, 1986 Hong Kong University Press. 127-140.
Reimer L, Rennekamp R, Fromm I & Langenfeld M (1991) Contrast in the electron
spectroscopic imaging mode of a TEM. IV. Thick specimens imaged by the most-
probable energy loss. J Microsc 162: 3-14.
Ronquist F & Huelsenbeck JP (2003) MrBayes 3: Bayesian phylogenetic inference under
mixed models. Bioinformatics 19: 1572-1574.
Schwarz G (1978) Estimating the Dimension of a Model. Ann. Statist. 6: 461-464.
Schweimanns M & Felbeck H (1985) Significance of the occurence of chemoautotrophic
bacterial endosymbionts in lucinid clams from Bermuda. Mar Ecol Prog Ser 24: 113-
120.
Taylor JD & Glover EA (2005) Lucinidae (Bivalvia) – the most diverse group of
chemosymbiotic molluscs. Zool J Linnean Society 148.
Thompson JD, Higgins DG & Gibson TJ (1994) CLUSTAL W: improving the sensitivity of
progressive multiple sequence alignment through sequence weighting, position-
specific gap penalties and weight matrix choice. Nucleic Acids Res 22: 4673-4680.
Williams ST, Taylor JD & Glover EA (2003) Molecular Phylogeny of the Lucinoidea
(Bivalvia): Non-Monophyly and Separate Acquisition of Bacterial Chemosymbiosis. J
Molluscan Stud 70: 187-202.
Winnepenninckx BM, Reid DG & Backeljau T (1998) Performance of 18S rRNA in littorinid
phylogeny (Gastropoda: Caenogastropoda). J Mol Evol 47: 586-596.

Title and legends to figures
Figure 1. Symbionts phylogenetic tree realized using 16S rRNA sequences (1269 bp).
Tree was reconstructed using Maximum Likelihood (ML) and Bayesian Inference (BI) under
K80+I+Γ evolutionary models. Robustness of clades was evaluated and only bootstrap values
(ML, 100 replicates [above branches]) and posterior probabilities (BI [under branches]) up to
80% were displayed. Taxa in bold provide from this study.
Figure 2. Symbionts phylogenetic tree realized using aprA sequences (273 bp).
Tree was reconstructed using Maximum Likelihood (ML) and Bayesian Inference (BI) under
TrNef+Γ (ML) and K80+Γ (BI) evolutionary models. Robustness of clades was evaluated and
only bootstrap values (ML, 100 replicates [above branches]) and posterior probabilities (BI
[under branches]) up to 70% were displayed. Taxa in bold provide from this study.
Figure 3. Hosts phylogenetic tree realized using contatenation of 18S rRNA and 28S
rRNA sequences (1252 bp).
Tree was reconstructed using Maximum Likelihood (ML) and Bayesian Inference (BI) under
TrNef+Γ (ML) and SYM+Γ (BI) evolutionary models. Robustness of clades was evaluated
and only bootstrap values (ML, 100 replicates [above branches]) and posterior probabilities
(BI [under branches]) up to 80% were displayed. Taxa in bold provide from this study and all
other sequences provide from (Williams et al., 2003). aSequences obtained for these two
species correspond to those published by Williams et al. (2003).
Figure 4.

Figure 1. Symbionts phylogenetic tree realized using 16S rRNA sequences (1269 bp).
Figure 2. Symbionts phylogenetic tree realized using aprA sequences (273 bp).

Figure 3. Hosts phylogenetic tree realized using contatenation of 18S rRNA and 28S
rRNA sequences (1252 bp).

Tables
Table 1. Sampling of Phillippines clams from PANGLAO 2005 cruise.
Speci
men
code
Speciesa
Samplin
g site
Locatio
n
Stati
on
Dep
th
(m)
Accession numbersb
16S
rRNA
APS
reducta
se
18S
rRNA
28S
rRNA
OG 18 -
Bohol
Sea
Off
Balicasa
g island
9°29.4’
N
123°43.
7’E
CP
2340
271
-
318
To be
submit
ted
To be
submit
ted
-
To be
submit
ted
OG 57 - Bohol
Sea
9°24.3’
N
124°10.
7’E
CP
2355
176
4 -
177
5
To be
submit
ted
To be
submit
ted
To be
submit
ted
To be
submit
ted
OG 98 Gloverin
a sp.
Bohol/S
ulu seas
sill
8°49.9’
N
123°37.
6’E
CP
2360
357
-
372
To be
submit
ted
To be
submit
ted
To be
submit
ted
To be
submit
ted
OG
101
Cardiolu
cina
quadrata
Bohol
Sea
9°26.9’
N
123°34.
5’E
CP
2388
762
-
786
To be
submit
ted
To be
submit
ted
To be
submit
ted
To be
submit
ted
OG
103
Myrtea
flabellifor
mis
Bohol
Sea
9°26.9’
N
123°34.
5’E
CP
2388
762
-
786
To be
submit
ted
- - -
OG
108c
- Bohol
Sea
9°27.4’
N
123°43.
1’E
CP
2390
627
-
645
To be
submit
ted
To be
submit
ted
To be
submit
ted
To be
submit
ted
a Species determination was realized by John D. Taylor (NHM, UK) based on morphological
characters. b Due to tissue conservation some sequences has not been obtained for some individuals.
c Microscopy analysis could not be realized for OG 108 specimen.

Table 2. Primers and PCR conditions.
Sequence (5’> 3’) Tm Name Target Reference
Sym
bio
nt
AGAGTTTGATCATGGCTCAG 54°C
16S_UNIV-8f 16S
rRNA (Lane, 1991)
TACGGCTACCTTGTTACGACT 16S_UNIV-1492r
TGGCAGATMATGATYMACGGG 58°C
APS-FW APS
reductase
(Deplancke et al.,
2000) GGGCCGTAACCGTCCTTGAA APS-RV
Host
CCGTCTTGAAACACGGACCAAG
52°C
LSU-900f 28S
rRNA
(Large
subunit)
(Olson et al., 2003)
AGCGCCATCCATTTTCAGG LSU-1600r (Williams et al., 2003)
CTGGTTGATYCTGCCAGT 54°C
18S-5’F 18S
rRNA
(Winnepenninckx et
al., 1998)
CTTCGAACCTCTGACTTTGG 18S-1100R (Williams et al., 2003)

Table 3. 16S rRNA sequences identity matrix (pairwise deletion)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
1: Codakia orbicularis symbiont ID
2: Anodontia alba symbiont 1,000 ID
3: Lucina nassula symbiont 0,984 0,984 ID
4: Codakia costata symbiont 0,975 0,975 0,979 ID
5: Lucina floridana symbiont 0,971 0,971 0,981 0,996 ID
6: OG57 symbiont 0,966 0,966 0,971 0,976 0,978 ID
7: OG101 symbiont 0,970 0,970 0,973 0,982 0,980 0,985 ID
8: OG103 symbiont 0,970 0,970 0,973 0,978 0,978 0,980 0,993 ID
9: Lucinoma aff. kazani symbiont 0,969 0,969 0,973 0,981 0,978 0,982 0,994 0,990 ID
10: Lucinoma aequizonata symbiont 0,960 0,960 0,963 0,968 0,966 0,981 0,976 0,973 0,974 ID
11: OG18 symbiont 0,977 0,977 0,978 0,979 0,976 0,988 0,981 0,984 0,977 0,969 ID
12: OG98 symbiont 0,971 0,971 0,972 0,973 0,972 0,985 0,978 0,978 0,973 0,968 0,999 ID
13: OG108 symbiont 0,969 0,969 0,970 0,971 0,972 0,986 0,976 0,976 0,971 0,969 0,999 0,999 ID
14: Anodontia ovum symbiont 0,951 0,951 0,959 0,957 0,959 0,952 0,953 0,952 0,951 0,944 0,960 0,955 0,953 ID
15: Anodontia phillipiana symbiont 0,947 0,947 0,955 0,949 0,951 0,946 0,947 0,946 0,946 0,938 0,954 0,949 0,946 0,990 ID
16: Thyasira flexuosa symbiont 0,942 0,942 0,944 0,945 0,942 0,941 0,941 0,939 0,940 0,933 0,950 0,944 0,942 0,947 0,945 ID
17: Lucina pectinata symbiont 0,947 0,947 0,949 0,941 0,939 0,933 0,933 0,932 0,933 0,926 0,941 0,936 0,934 0,945 0,943 0,941 ID
Recommended