Embed Size (px)
Citation preview

ZNAČILNOSTI, DIVERZITETA TER OGROŽENOST KRANJSKE ČEBELE Apis mellifera carnica Pollman 1878
- Splošni pregled znanj v domačih in tujih virih za poročilo o diverziteti Amc za projekt Amc Promo BID
dr. Gordana Glavan, Univerza v Ljubljani, Biotehniška fakulteta – Oddelek za biologijo
1. Uvod
Poglobljena razmišljanja glede ogroženosti kranjske čebele so sprožili predvsem
spomladanski pomori čebel zaradi pesticidov v nekaterih predelih Slovenije leta 2008. To
podvrsto pri nas ogroža predvsem propadanje čebeljih družin in s tem povezano siromašenje
genskega sklada (Gregori 2009). Ogroženost kranjske čebele je treba iskati v smeri njene vse
večje izpostavljenosti genski poluciji, križanju z drugimi podvrstami, kot tudi vse večji genski
homogenizaciji znotraj podvrste. Obstoj kranjske čebele ogroža tudi uporaba
fitofarmacevtskih sredstev (FFS) v kmetijstvu ter bolezni, kot je varoza.
V Sloveniji je zakonsko dovoljeno gojiti samo kranjsko čebelo (Zakon o živinoreji, 70. člen).
Kljub zakonski prepovedi prihaja do vnašanja tujih matic, ki so bile mogoče prodane kot
kranjice, a je njihov izvor nejasen. Na nevarnosti prostega pretoka blaga z vstopom v Evropo,
na dovoljene prevoze čebel in proste nakupe matic je opozarjal že Poklukar (1999). Nekateri
čebelarji, predvsem iz JZ Slovenije imajo težave z naravnim pritiskom italijanske čebele.
Vse bolj vprašljiv je čebelji križanec, buckfaška čebela, ki jo nekateri evropski čebelarji
ponekod nekontrolirano uvajajo (Gregori 2009). Isti avtor sicer navaja, da so vnosi buckfaške
čebele tudi prisotni tudi pri nas, a uradnih zapisov ali genskih analiz, ki bi to potrjevale,
nimamo. Buckfaška čebela, s katero tudi čebelarijo ponekod v Evropi, ni naravnega nastanka,
ampak jo je vzredil Adam Kehrle (ali brat Adam) iz opatije Buckfast, ko je s križanjem raznih
podvrts iskal primerno čebelo, s katero bi lahko čebelarili na Britanskem otočju, potem, ko je
pršičavost tamkajšnjo avtohtono čebelo uničila skoraj do izumrtja. Pri potomcih (križancih)
buckfaške čebele in kranjske čebele je bilo opaziti povečanje agresivnosti in zato je delo s
takimi čebelami močno oteženo (Brother Adam, 1987).
1 / 15

Do genske homogenizacije prihaja zaradi mešanja različnih ekotipov (Gregori 2009).
Omogoča jo predvsem prosta prodaja matic ter umetno narejenih čebeljih družin,
narejencev. H genski homogenizaciji prispevajo prevozi čebel na pašo v času plemenjenja
matic: te se plemenijo z lokalnimi troti, ki lahko pripadajo drugemu ekotipu. Na gensko
homogenizacijo pa pozitivno vpliva tudi testiranje matic. V okviru vzreje matic poteka
progeni test, pri katerem se matice registriranih vzrejevalcev dajo v testiranje čebelarjem v
različnih predelih Slovenije. Delež teh matic je sicer relativno majhen, saj gre letno le za nekaj
100 matic v primerjavi z vsaj 160.000 čebeljimi družinami.
Do zmanjševanja genske raznolikosti kranjske čebele prihaja tudi zaradi propadanja
čebeljih družin (Gregori 2009). To je najbolj usodna razsežnost ogroženosti kranjske čebele
pri nas, je najbolj pereč problem, ker prihaja do nepovratnih izgub in s tem siromašenja
genskega sklada. Prihaja do počasnega izumiranja edinstvenih lokalnih populacij. Genski
sklad, ki je nastajal milijone let, počasi peša, lokalne populacije izginjajo in z njimi tudi
dragoceni genski zapisi. Vzroki za to tičijo predvsem v propadanju malih čebelarstev ter
odmiranju lokalnih populacij čebel zaradi dejavnikov okolja (zime), fitofarmacevtskih sredstev
ter bolezni.
Pristopna pogodba Slovenije k EU (deklaracija št. 42) določa »ohranjanje kranjske čebele
kot avtohtone pasme čebel in avtohtone populacije s posebnimi značilnostmi na ozemlju
Republike Slovenije«. S tem prispeva k vzdrževanju biološke raznolikosti in ohranitvi
avtohtonega genskega materiala, katerega rezultat naj bi bila gensko stabilna in uravnotežena
populacija kranjske čebele. Evropska usmeritev pravi, da naj se ohranja kranjsko čebelo na
ravni krajevnih različkov ali ekotipov (De la Rua in sod., 2009), zato se tudi obravnavani vidiki
ogroženosti nanašajo v veliki meri na ekotipe. Za ohranitev lokalnih populacij, ekotipov ter
podvrste kranjske čebele bi bilo potrebno vpeljati ustrezno metodologijo za določanje
lokalnih ekotipov, ki bi temeljila na določenih morfoloških ter molekularnih znakih (Bouga s
sod., 2011). Vendar dosedanje molekularne ter morfološke raziskave niso dokazale obstoja
različnih ekotipov v Sloveniji, temveč kažejo na to, da je populacija kranjske čebele pri nas
precej homogena (Kozmus 2008; Sušnik s sod., 2004). Zaradi geografskih in podnebnih
2 / 15

značilnosti na ozemlju Slovenije bi sicer težko pričakovali izrazite meje med različnimi ekotipi.
Edina izrazitejša ovira za naravno mešanje populacij so gorski grebeni za SZ Slovenije
(Karavanke in Julijske Alpe). Tako bi lahko pričakovali le postopen prehod iz alpskega v
panonski in dinarski ekotip od zahoda proti vzhodu in juguzahodu.
2. Podvrste ter linije medonosne čebele Apis mellifera
Na osnovi morfoloških značilnosti je opisanih 28 podvrst medonosne čebele Apis
mellifera (Ruttner s sod., 1978; Ruttner 1988; Engel, 1999). Kranjska čebela oz. kranjska sivka
(Apis mellifera carnica Pollman 1879) glede na izvor uvrščajo v jugo-vzhodnoevropsko ali C
skupino podvrst medonosne čebele. Poleg omenjene skupine obstajajo še naslednje
filogenetske linije: afriška (A), severno-zahodnoevropska (M), linija O, ki naj bi bila prisotna
na Bližnjem in Srednjem vzhodu ter linija Y, ki se razprostira na območju Etiopije (Ruttner s
sod. (1978), Ruttner (1988), Arias in Sheppard (1996), Franck in sod. (2000), Palmer in sod.
(2000), Franck in sod. (2001)).
2.1. Metode za ločevanje posameznih podvrst medonosne čebele Apis mellifera
Za ločevanje podvrst čebel so najprej uporabljali barvo čebel, ki je najbolj očitna lastnost.
Kasneje se je uveljavilo merjenje določenih morfoloških lastnosti (dolžina rilčka, femurja,
tibie in metatarzusa zadnje noge; dolžina in širina sprednjih kril; dolžina dlake na 5. tergitu), s
tem pa natančnejše razvrščanje podvrst čebel (Alpatov, 1929; Rutner 1988). Analizo
ožiljenosti kril je vpeljal Goetze (1940), pristop pa je pogosto uporabljen v večini vzrejnih in
ohranitvenih programov vsake že določene podvrste (Leclercq, 1999), saj z njo enostavno
dokažemo pripadnost določeni podvrsti. Na krilih lahko merimo in izračunavamo številne
lastnosti, med katerimi so najbolj informativne in največkrat uporabljene naslednje lastnosti:
10 kotov na žilnem sistemu (A4, B4, D7, E9, G18, J10, J16, K19, L13, O26), razdalje a, b, c, in d
ter kubitalni indeks (Kauhausen-Keller in Keller, 1994; Nazzi, 1992) (Slika 1, Slika 2).
3 / 15
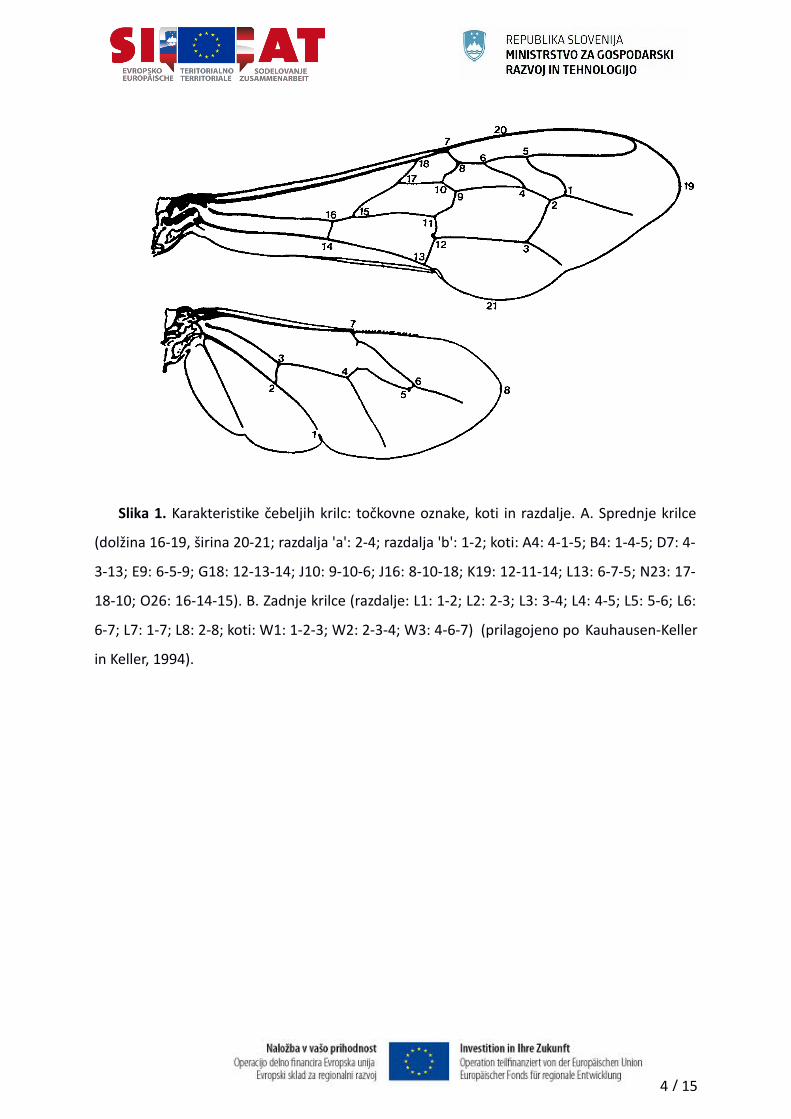
Slika 1. Karakteristike čebeljih krilc: točkovne oznake, koti in razdalje. A. Sprednje krilce
(dolžina 16-19, širina 20-21; razdalja 'a': 2-4; razdalja 'b': 1-2; koti: A4: 4-1-5; B4: 1-4-5; D7: 4-
3-13; E9: 6-5-9; G18: 12-13-14; J10: 9-10-6; J16: 8-10-18; K19: 12-11-14; L13: 6-7-5; N23: 17-
18-10; O26: 16-14-15). B. Zadnje krilce (razdalje: L1: 1-2; L2: 2-3; L3: 3-4; L4: 4-5; L5: 5-6; L6:
6-7; L7: 1-7; L8: 2-8; koti: W1: 1-2-3; W2: 2-3-4; W3: 4-6-7) (prilagojeno po Kauhausen-Keller
in Keller, 1994).
4 / 15

Slika 2. Razdalje a, b, c, in d na srednjem krilcu čebele (prilagojeno po Nazzi, 1992)
Ostali kriteriji, ki se vse bolj uveljavljajo, pa so genetski oz. molekularni. Najbolj sprejeta
je mitohondrijska analiza. Nekateri avtorji navajajo, da je le-ta zanesljivejša od morfoloških
znakov, saj se morfologija osebkov prilagaja pogojem okolja (Avise s sod., 1987; Franck s sod.,
2000; Kozmus s sod., 2007).
Mitohondrijska analiza je bila velikokrat uporabljena za biogeografske študije podvrst A.
mellifera. S to analizo so določili 5 evolucijski linij (A, C, M, O in Y) medonosne čebele z
uporabo visoko variabilnih COI-COII področij v zapisu mitohondrijske DNK (mtDNK) (Cornuet
s sod., 1991; Garnery s sod., 1992; Franck s sod., 1998, 2001). Variabilnost COI-COII področij
je rezultat sprememb v njihovi dolžini (prisotnost ter odsotnost P sekvenc, število ponovitev
Q sekvenc, manjše delecije) in substitucije nukleotidov. Najkrajši haplotip, značilen za C
5 / 15

(jugo-vzhodnoevropsko) linijo, kamor spada tudi kranjska čebela, nima P sekvence ter
vsebuje samo 1 Q sekvenco (Garnery s sod., 1998). Pri C liniji obstaja vsaj 6 haplotipov (C1
pri A. m. ligustica, C2a pri A. m. carnica, C2b pri A. m. caucasica, C2C pri A. m. carnica iz
Slovenije in Hrvaške in C2D ter C2E pri A. m. macedonica in A. m. carnica iz Srbije) in 5
polimorfnih mest (1 insercijsko/delecijsko mesto in 4 tranzicijska) ter nobenih variacij znotraj
posameznih podvrst (Franck s sod., 2000; Sušnik s sod., 2004; Kozmus s sod., 2007). V vzorcih
kranjske čebele iz Srbije so odkrili še dva haplotipa: C2I in C2J (Nedić s sod., 2009). Po drugi
strani pa je analiza mitohondrijskih ND2 sekvenc pokazala, da obstajata 2 haplotipa v
slovenski ter avstrijski populaciji A. m. carnica (Arias in Sheppard, 1996). Študije so tako
pokazale, da se na nivoju mitohondrijske DNK kranjska čebela A. m. carnica razlikuje od
ostalih podvrst. Na nivoju mitohondrijske DNK so našli še dodatno polimorfno mesto,
ugotovljeno s pomočjo restrikcijskega encima Xba1, ki loči obe podvrsti A. m. carnica ter A.
m. ligustica od ostalih podvrst A. mellifera (Meixner s sod., 1993).
Nekatere mitohondrijske haplotipe se da razlikovati preko sprememb v zapisu
mitohondrijske konzervativne tRNAleu-cox2 intergenske regije (Garneryet al., 1993, 1995,
1998; De la Rúa et al., 1998, 1999, 2000; Franck et al., 1998, 2000a, 2000b; Cánovas et al.,
2008). Ker je bilo do sedaj ugotovljenih samo nekaj mitohondrijskih haplotipov in še to na
podlagi majhnega števila polimorfizma 1 nukletida, so nekateri avtorji predlagali, (Munoz s
sod., 2009; Franck et al., 2000a, 2000b; Sušnik et al., 2004), da so tRNAleu-cox2 intergenske
regije primernejše za analizo molekularne variabilnosti znotraj C linije.
Pri razlikovanju posameznih podvrst ter ekotipov se uporablja tudi analiza mikrosatelitov
(Estoup s sod., 1995).
3. Kranjska čebela A.m.carnica3.1. Razširjenost kranjske čebele v Sloveniji ter njeni ekotipi
Izvorno območje kranjske čebele je opredeljeno severno in južno od Karavank, na obeh
straneh meje med Avstrijo in Slovenijo (Ruttner, 1988). Podvrsta se je oblikovala po koncu
zadnje ledene dobe pred približno 10.000 leti. Celotna populacija kranjske čebele je na
podlagi morfoloških znakov razdeljena na tri večje skupine (Ruttner in Hänel, 1992):
− alpska (Slovenija, Avstrija, Slovaška),
6 / 15

− panonska (Madžarska, Romunija),
− mediteranska (Hrvaška, Bosna in Hercegovina, Srbija, Črna Gora).
Kranjska čebela je danes razširjena na vseh kontinentih in je po številu družin, s katerimi
čebelarijo, takoj za italijansko. Vendar pa so te kranjske čebele v veliki meri križane z drugimi
rasami, predvsem z italijansko.
Zaradi prilagajanja krajevnim razmeram so v Sloveniji nastale edinstvene populacije
čebel, ki jih je stroka lahko prepoznavala kot posamezne krajevne različke ali ekotipe,
izkustveno pa naj bi jih prepoznavali tudi čebelarji sami, predvsem njihovo prilagojenost na
izkoriščanje lokalnih gozdnih paš. Čebelarska stroka jih je razdelila na 4 ekotipe: alpski,
dinarsko-kraški, panonski in mediteranski ekotip kranjske čebele (Rihar, 1972; Zdešar, 1999).
Poklukar (1998) pa je na podlagi morfoloških znakov (kubitalni indeks, dolžina rilčka, dolžina
goleni) preučil populacijo čebel v Sloveniji in celotno populacijo razdelil na tri večje skupine
oz. ekotipe: panonski, alpski in dinarski. Ozemlje ob italijanski meji je označil kot nedefinirano
območje zaradi mešanja kranjske čebele z italijansko čebelo.
Da bi meje med ekotipi natančneje določili so Sušnik in sod. (2004) celotno populacijo
preučili še na podlagi genetskih označevalcev, vendar razlik niso našli. Tudi analiza ožiljenosti
kril 273 vzorcev čebel iz različnih koncev Slovenije ni pokazala statističnih razlike med
posameznimi vzorci, ki bi lahko podprla obstoj različnih ekotipov kranjske čebele pri nas
(Kozmus 2008). Obe raziskavi nakazujeta, da je populacija kranjske čebele v Sloveniji
homogena. Verjetno je to posledica homogenizacije, mešanja nekoč obstoječih ekotipov.
3.2. Splošne morfološke ter biološke značilnosti kranjske čebeleV primerjavi z drugimi podvrstami je kranjska čebela precej temna in ima izrazit dolg
rilček (Ruttner 1988). Na oprsju ima rjavkaste dlačice, obročki na zadku pa so usnjeno rjave
barve z včasih nakazanimi svetlejšimi pegami na prvem in drugem obročku. Glavna odlika
kranjske čebele je njena mirnost. Med čebelarji je zelo priljubljena, saj je druga najbolj
razširjena podvrsta čebel na svetu, takoj za italijansko čebelo A. m. ligustica. Poleg tega je
znana po delavnosti, dolgoživosti, izkoriščanju paše, dobrem prezimovanju in tudi po skromni
7 / 15

porabi zimske zaloge hrane. V primerjavi z drugimi podvrstami zelo dobro izkoristi pelodno
pašo (Ruttner, 1988). Zaradi teh lastnosti so jo velikokrat uporabili pri selekciji novih linij
čebel.
3.3. Morfološke ter molekularne analize populacije kranjske čebele v Sloveniji
Poklukar (1998) je natančneje analiziral morfološke znake (kubitalni indeks, dolžina rilčka,
dolžina goleni) v populaciji čebel v Sloveniji. Analizirali so 30 vzorcev panonskega tipa, 32
alpskega, 21 notranjskega, 9 severno primorskega ter 12 južno primorskega tipa čebel. Po
uporabi multivariatne statistike pri kanonični diskriminantni analizi so na osnovi vzetih
osnovnih statistik vzorcev čebel z meritvami kubitalnega indeksa, velikosti goleni zadnje noge
in dolžine rilčka celotno populacijo razdelili v tri večje skupine oz. ekotipe: panonski, alpski in
dinarski. Povprečni kubitalni indeks, izračunan za vzorce iz cele Slovenije, je bil 2,44,
povprečna dolžina rilčka 5,57 mm ter povprečna dolžina goleni lateralno 2,39 mm. Po eni
izmed raziskav 36 čebeljih družin v Sloveniji je bil izračunan kubitalni indeks 2,69 (±0,40).
Analiza obarvanosti obročkov na zadku
V zadnjem obdobju se v populaciji kranjske čebele v Sloveniji opaža vse več čebel z
rumenimi obročki na zadku, ki za kranjsko čebelo niso značilni in so posledica križanj z
drugimi podvrstami medonosne čebele (Kozmus 2011). V primerih, da se v družini ugotovijo
čebele z rumenimi obročki na zadku, gre bodisi za uvožene italijanske čebele ali za njihove
križance (Rihar, 2003), lahko pa tudi za križance z buckfast čebelo (Borsuk in Olszewski, 2010;
Kozmus 2011). Da bi ugotovili, kolikšen del križancev-hibridov je prisotnih v populaciji
kranjske čebele v Sloveniji, je bila narejena morfološka analiza obarvanosti obročkov na
zadku čebel delavk (Kozmus 2011). Ugotovljeno je bilo, da je največji odstotek čebel z
rumenimi obročki na zadku prisoten na zahodnem delu Slovenije ob meji z Italijo (obalno
kraška in goriška regija) ter na Koroškem. Na meji z Italijo gre verjetno za mešanje z
italijansko podvrsto medonosne čebele. Avtorji navajajo dve možni razlagi, zakaj je na
območju Koroške relativno visok odstotek čebel z rumenimi obročki na zadku (od 12–16 %).
Po eni strani bi bile lahko na avstrijski strani prisotne italijanske in/ ali buckfast čebele in da
so se čebele s Koroške hibridizirale z njimi. Druga možnost nakazuje na to, da so čebelarji na
8 / 15

tem območju kupili italijanske in/ali buckfast čebele, ki so hibridizirale z avtohtono
populacijo. V povprečju je v populaciji kranjske čebele v Sloveniji 5,6 % čebel z rumenimi
obročki na zadku. Čebelarjem svetujejo, naj zamenjajo matice v družinah, v katerih so
zasledili rumeni obroček pri več kot 2% delavk (Bouga s sod., 2011).
Molekularna analiza
Leta 2004 je bila narejena mitohondrijska in nuklearna DNK analiza (analiza
mikrosatelitov) z namenom preučiti genetsko variabilnost kranjske čebele v Sloveniji (Sušnik s
sod., 2004). Osnovni cilj je bil ugotoviti genetsko 'čistost' v Sloveniji ter možnost vnosa genov
iz populacij kranjske čebele iz bližnjih držav. Rezultati analize mitohondrijev ter mikrosatelitov
so pokazali nizko gensko raznolikost med vzorci, dobljenih iz Slovenije. Pri mitohondrijski
analizi so poleg že znanega haplotipa COI-COII področja mitohondrijske DNK C2b odkrili še en
haplotip, ki so ga poimenovali C2C. Analiza vzorcev čebel A.m. carnica, dobljenih na
Hrvaškem, je pokazala podobne rezultate: vzorci so bili gensko podobni vzorcem iz Slovenije,
medtem ko pa so bili vsi omenjeni vzorci gensko zelo različni od vzorcev čebel A.m.
macedonica. Raziskovalci na osnovi genske analize trdijo, da kranjske čebele iz Slovenije
zagotavlja avtohtoni genski sklad znotraj podvrste A.m.carnica.
Vedenjske, biološke ter patološke analize čebeljih družin v Sloveniji-predlagan način
uvrščanja in odbira družin v vzrejališču čebeljih matic (Gregorc in Lokar, 2010)
V študiji, ki je obsegala 36 čebeljih družin (A.m.carnica), so čebele vrednotili glede na
mirnost, rojivost, živahnost, rasne karakteristike, kubitalni indeks (Ci), pridelavo medu, obseg
pokrite zalege, sposobnost čiščenja odmrle zalege in prisotnost spor Nosema spp.. Mirnost
čebel na satju so ocenjevali z ocenami od 1 do 4, dvakrat v letu, rojenje so ocenjevali v maju
in juniju z ocenami od 1 do 4. Živahnost so vrednotili izraženo v številu zasedenih ulic v
posamezni družini, ki so jo primerjali glede na povprečno zasedenost v vseh testiranih
družinah. Oceni 1 in 2 sta predstavljali zasedenost pod povprečjem, 3 in 4 pa zasedenost nad
povprečjem v čebelnjaku. Ocenjevali so tudi obarvanost obročkov zadka in kubitalni indeks
(Ci). Ci so izmerili na vzorcu 1749 delavk iz 36 družin. Vzorci, ki so ustrezali standardu (od 2,4
– 3,0), so dobili oceno 1, vzorci, ki standardu niso ustrezali, so dobili oceno 0 in so bili izločeni
9 / 15

iz nadaljnje obravnave. Tudi družine, pri katerih so bile ugotovljene delavke z oranžnimi
obročki zadka so bile izločene. Donos medu je bil ugotovljen s tehtanjem medenega satja
pred točenjem in po točenju medu. Razlika v teži je predstavljala količino pridelanega medu v
posamezni družini. Obseg pokrite zalege je bil izmerjen za vsako družino trikrat in sicer aprila,
maja in julija. Sposobnost čiščenja odmrle zalege so ugotavljali s pomočjo “pin-kill testa”.
Uporabili so dve polji s pokrito zalego ki sta obsegali po 100 satnih celic. V enem polju s 100
satnimi celicami so usmrtili bube, ter po 16 in 24 urah prešteli satne celice, ki so bile očiščene
odmrle zalege. Sposobnost čiščenja so izrazili v odstotku očiščenih satnih celic v časovnem
obdobju. Družine so bile glede na čistilno sposobnost razvrščene v pet skupin na osnovi
deleža očiščenih satnih celic: 5 = > 95 %; 4 = 90 – 95 %; 3 = 80 - 89 %; 2 = 70 – 79 %; in 1 = <
70 %. Vzorec delavk iz vsake družine je bil odvzet z vrha čebelje gruče v januarju, in z brade
panja v maju in septembru. Vzorci so bili preiskani na spore Nosema spp. Družine so bile
ocenjene glede na količino ugotovljenih spor: ocena 4 = 0 x 106 spor/čebelo; 3 = < 8 x 106
spor/čebelo; 2 = 8 do 30 x 106 spor/čebelo; in 1 = > 30 x 106 spor/čebelo. Vrednosti ocen za
posamezne lastnosti so sešteli v končni Selekcijski Indeks čebelje družine. Povprečna
vrednost Ci v testnih družinah je bila 2,69 (±0,40), povprečen donos medu 9,52 kg (±6,64) in
obseg pokrite zalege je v povprečju obsegal 7061 cm2 (±2813). Družine so v 24 urnem testu
odstranjevanja odmrle zalege, izvedenem v maju in juniju odstranile 83,4 % (±11,2) zalege.
Nad 90 % odmrle zalege je bilo odstranjenih v dvanajstih družinah, nad 96 % odmrle zalege
pa je bilo odstranjene v osmih družinah. Največje število spor Nosema spp. je bilo v
septembru. Na osnovi izvedenih testov, so avtorji zaključili, da je ocenjevanje in selekcija
čebeljih družin na osnovi uporabljenih karakteristik ustrezna metoda za uvrščanje družin in
odbiro v vzrejališču čebeljih matic.
Osnovni kriteriji v Sloveniji za gojenje in vzdrževanje kranjske čebeleOsnovni kriteriji za gojenje kranjske čebele v Sloveniji so naslednji (Bouga s sod., 2011,
Gregorc in Lokar 2010): - Kubitalni indeks za delavke naj bo med 2.4-3.0 ter 1.8-2.3 za trote- Barva zadka naj bo siva in ne rumena (mešanje z italijansko čebelo A.m.ligustica): če
ima rumen obroček na abdomnu več kot 2% delavk, je potrebno zamenjati matico
10 / 15

3.4. Morfološke ter molekularne analize populacije kranjske čebele iz drugih držav
Morfološka analiza vzorcev kranjske čebele iz Nemčije (Moritz 1991)
Narejena je bila primerjava ožiljenosti kril kranjske čebele vzorcev iz leta 1911 ter vzorcev
novodobne kranjske čebele iz Nemčije (Moritz 1991). Rezultati analize so pokazali, da je
novodobna kranjska čebela križanec med 'staro', avtohtono kranjsko čebelo ter podvrsto Apis
mellifera mellifera. Podobne razlike verjetno danes obstajajo med osebki kranjske čebele iz
Slovenije ter Nemčije.
Molekularna analiza vzorcev kranjske čebele iz Hrvaške
Na Hrvaškem so opisali tri ekotipe kranjske čebele: Panonski, Subalpinski in Dalmatinski
(Ruttner, 1988). Genetske študije niso našle razlik med vzorci čebel iz Hrvaške in Slovenije
(Sušnik s sod., 2004). V eni izmed študij so preučevali hrvaško obalno populacijo A. m.
carnica z uporabo mikrosatelitnih ter mitohondrijskih označevalcev (Munoz s sod., 2009).
Rezultati analize mtRNK konzervativne tRNAleu-cox2 intergenske regije so pokazali, da vzorci
čebel pripadajo Centralno-Mediteranski ter Jugovzhodni evropski evolucijski C liniji. Analiza
variabilnosti mikrosatelitov je nakazala na obstoj dveh populacij čebel znotraj istega ekotipa.
Molekularna analiza vzorcev kranjske čebele iz Srbije, Bosne in Hercegovine ter
Makedonije
Mitohondrijska analiza variabilnega COI-COII DNK področja je bila narejena na treh
ekotipih A.m.carnica: Banat (B), Syenichko-Peshterski (S) in Timok (T) (Kozmus s sod., 2007).
Rezultati so pokazali, da vzorci pripadajo C2D mitohondrijskemu haplotipu, kamor spada tudi
A.m.macedonica. Pri kranjskih čebelah iz Srbije so odkrili tudi nov haplotip, C2E, ki se od C2D
haplotipa razlikuje v dveh A-T transverzij ter eni inserciji. V vzorcih čebel ekotipa Timok so
odkrili dve dodatni polimorfni mesti: delecijo T nukleotida ter T-C tranzicijo. Ta študija kaže
na to, da je kranjska čebela iz Srbije gensko drugačna od kranjske čebele iz Slovenije.
V neki drugi raziskavi so z mitohondrijsko analizo variabilnega COI-COII DNK področja
našli še dva C2 haplotipa, C2I ter C2J (Nedić s sod., 2009). Vzorci čebel so bili nabrani na
11 / 15

različnih lokacijah v Srbiji (Vršac, Knjaževac, Kraljevo in Vranje). Analiza sekvenc iz genske
banke Gen Bank so pokazali veliko podobnost med C2I mtDNK haplotipom ter A. m. cypria
haplotipom, ki izvira iz Turčije. Določili so tudi dve novi polimorfni mesti v COI-COII mtDNK
regiji pri čebelah iz Vranja in Knjaževca. Avtorji raziskave sklepajo, da kranjske čebel v Srbiji
predstavljajo pomemben genetski sklad, ki veliko prispeva k biodiverziteti te podvrste
medonosne čebele.
Narejena je bila tudi primerjava mitohondrijske DNK podvrst Apis mellifera carnica in A.
m. macedonica, s katero so primerjali približno 1680 osebkov iz 560 čebelnjakov iz Srbije,
Bosne in Hercegovine ter Makedonije (Stevanovic s sod., 2010). V vseh vzorcih so našli C2D
mtDNK haplotip. Ti rezultati so pokazali, da imajo osebki obeh podvrst s tega področja
Balkana enak haplotip. Restrikcijski mesti za encime NcoI in SytI, ki sta značilni za podvrsto
A.m. macedonica, so pokazali za vzorce čebel iz vzhoda, juga ter jugozahoda Srbije ter za
čebel iz Makedonije, ne pa za vzorce dobljene iz severa Srbije ter Bosne in Hercegovine. Na
podlagi dobljenih rezultatov domnevajo, da je podvrsta A.m.macedonica razširjena tudi v
večjem delu Srbije, v severnem delu Srbije ter v Bosni in Hercegovini pa je verjetno prisotna
le A.m.carnica.
Na vzorcih treh ekotipov kranjske čebele iz Srbije: Banat (B), Syenichko-Peshterski (S) in
Timok (T) je bila narejena analiza kromosomov, kariotipizacija (Stanimirovic s sod., 2005). Pri
tem postopku po Giemsi barvajo kromosome, ki so v metafazi. Na obarvanih kromosomih
tako nastane tipičen vzorec črt, ki jim pravijo G-črte. Takšno barvanje je pokazalo razlike med
ekotipoma Syenichko-Peshterski (S) in Timok (T) na kromosomih 2, 4, 11 in 13. Največje
razlike v obarvanosti G-črt so ugotovili med ekotipoma Banat (B)in Syenichko-Peshterski (S)
na kromosomih 1, 2, 4, 11, 12, 13, 15 in 16. Raziskovalci ugotavljajo, da obstajajo velike
razlike med kromosomi posameznih ekotipov.
SKLEP
Tako morfološke kot molekularne analize kranjske čebele kot sosednjih populacij
medonosne čebele nakazujejo na obstoj genske pestrosti. Le ta pa je ogrožena na račun
12 / 15

velikih nihanj v populaciji čebel in gospodarjenja s čebelami. Slednje predvsem omogoča
transporte preko meja naravnega mešanj genskega sklada, bodisi na račun izkoriščanja
čebeljih paš ali z nakupom čebeljih družin in matic. Veliko nihanje v populaciji kranjske čebele
ima splošne značilnosti, ki jih poznamo tudi drugod v svetu, kjer čebelarijo z evropskimi
podvrstami čebel. Gre tako za vplive razširjanja povzročiteljev čebeljih bolezni, kakor tudi
izpostavljenost kseno biotikom. Glede na trenutno situacijo v arealu naravne razširjenosti
kranjske čebele imamo še možnost vzpostaviti trajnostni sistem za ohranjanje njene pestrosti
pa čeprav ne več v več v obsegu, kot so ga navajale starejše publikacije. Raziskava v okviru
Amc Promo BID sicer nakazuje na možnosti čezmejnega sodelovanja med čebelarji, po drugi
strani pa na vlogo večjih naravnih preprek in načinov gospodarjenja s čebelo. To so vsekakor
pomembna izhodišča za trajnostno gospodarjenje s pestrostjo genskega sklada.
LITERATURA:
Alpatov WW (1929) Biometrical studies on variation and races of the honey bee. Q Rev Biol4, 1-58.
Arias, M.C./ Sheppard, W.S. Molecular phylogenetics of honey bee subspecies (Apis mellifera L.) inferred from mitochondrial DNA sequence. Molecular phylogenetics and Evolution, 3(1996), 557–566.
Avise JC, Arnold J, Ball RM, 1987, Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics, Annu Rev of Ecol Syst, 18, 489-522.
Bouga, M., Alaux, C., Bienkowska, M., Büchler, R., Carreck, N.L., Cauia, E., Chlebo, R., Dahle, B., Dall'Olio, R., De la Rúa, P., Gregorc, A., Ivanova, E., Kence, A., Kence, M., Kezic, N., Kiprijanovska, H., Kozmus, P., Kryger, P., Le Conte, Y., Lodesani, M., Murilhas, A.M., Siceanu, A., Soland, G., Uzunov, A., Wilde, J. (2011) A review of methods for discrimination of honey bee populations as applied to European beekeeping. Journal of Apicultural Research 50, 51-84. 2011 doi: 10.3896/IBRA.1.50.1.06
Borsuk, G., Olszewski, K. 2010. Morphometric traits of buckfast and caucasian bees. Journal of Apicultural science, 54: 43–48
Brother Adam, 1987: Beekeeping at Buckfast Abbey. Northern Bee Books, Scout Bottom Farm Mytholmroy, Hebden Bridge, West Yorkshire, 122 str.
Cánovas F., De la Rúa P., Serrano J., Galián J. (2008) Geographic patterns of mitochondrial DNA variation in Apis mellifera iberiensis (Hymenoptera: Apidae), J. Zool. Syst. Evol. Res. 46, 24–30.
Cornuet J.-M., Garnery L., Solignac M. (1991) Putative origin and function of the intergenic region COI and COII of Apis mellifera: mitochondrial DNA, Genetics 1128, 393–403.
De la Rúa P., Galián J., Serrano J. (1998) Mitochondrial variability of honeybees populations from the Canary Islands, Mol. Ecol. 7, 1543–1547.
De la Rúa P., Galián J., Serrano J. (1999) Varibilidad mitocondrial en poblaciones de abejas de la miel del SurestePeninsular, Invest. Agr. Prod. Sanid. Anim. 14, 24–30.
De la Rúa P., Simon U.E., Tilde A., Mortiz R.F.A., Fuchs S. (2000) MtDNA variation in Apis cerana populations from the Philippines, Heredity 84, 124–130.
De la Rúa, P., R. Jaffé, R. Dall’Olio, I. Muñoz, J. Serrano, 2009: Biodiversity, conservation and current threats to European honeybees. Apidologie 40, str. 263-284.
13 / 15

Estoup A., Garnery L., Solignac M., Cornuet J.-M. (1995) Microsatellite variation in honey bee (Apis mellifera L.)populations: hierarchical genetic structure and test of the infinite allele and stepwise mutationmodels, Genetics 140, 679–695.
Franck P., Garnery L., Solignac M., Cornuet J.-M. (1998) The origin of West European subspecies of honeybees(Apis mellifera) new insights from microsatellite and mitochondrial data, Evolution 52, 1119–1134.
Franck, P./ Garnery, L./ Loiseau, A./ Oldroyd, P./ Helburn, H.R./ Solignac, M./ Cornuet, J.M. Genetic diversity of the honeybee in Africa: Microsatellite and mitochondrial data. Heredity, 86(2001), 420–430.
Franck P., Garnery L., Celebrano G., Solignac M., Cornuet J.-M. (2000a) Hybrid origins of honeybees from Italy (Apis mellifera ligustica) and Sicily (A. m. sicula), Mol. Ecol. 9, 907–921.
Franck P., Garnery L., Solignac M., Cornuet J.-M. (2000b) Molecular confirmation of a fourth lineage in honeybees from the Near East, Apidologie 31, 167–180.
Franck P., Garnery L., Loiseau A., Oldroyd B.P.,Hepburn H.R., Solignac M., Cornuet J.-M. (2001) Genetic diversity of the honeybee in Africa: microsatellite and mitochondrial data, Heredity 86, 420–430.
Garnery L., Cornuet J.-M., Solignac M. (1992) Evolutionary history of the honey bee Apis melliferabinferred from mitochondrial DNA analysis, Mol. Ecol. 1, 145–154.
Garnery L., Mosshine E.H., Oldroyd B.P., Cornuet J.-M. (1995) Mitochondrial DNA variation in Moroccan andSpanish honey bee populations, Mol. Ecol. 4, 465–471.
Garnery L., Solignac M., Celebrano G., Cornuet J.-M. (1993) A simple test using restricted PCR-amplified mitochondrial DNA to study the genetic structure of Apis mellifera L, Experientia 49, 1016–1021.
Garnery L. Franck P., Baudry E., Vautrin D. et al. (1998) Genetic biodiversity of the West European honeybee (Apis mellifera mellifera and Apis mellifera iberica). I. Mitochondrial DNA, Genet. Sel. Evol. 30, 31–47.
Goetze, G. Die beste Biene. Leipzig, Liedloff, Loth & Michaelis Verlag, 1940, 200 str.GREGORC, Aleš, LOKAR, Vesna. Selection criteria in an apiary of carniolan honey bee (Apis mellifera carnica)
colonies for queen rearing = Selekcijski kriteriji v čebelnjaku z družinami kranjske čebele (Apis mellifera carnica) za vzrejo matic. Journal of central european agriculture. [Online ed.], 2010, vol. 11, no. 4, str. 401-408.
Gregori J.: Kranjska čebela (Apis mellifera carnica): vidiki njene sedanje ogroženosti v Sloveniji. Acta entomologica slovenica, 17 (2), 2009
Kauhausen-Keller, D./ Keller, R. Morphometrical control of pure race breeding in the honeybee (Apis mellifera L). Apidologie, 25(1994), 133–143.
Kozmus P. Ugotavljanje prisotnosti ekotipov kranjske čebele (Apis mellifera carnica Pollman) v Sloveniji na podlagi razlik v ožiljenosti prednjih kril Carniolan bee (Apis mellifera carnica Pollman) population definition as based difference in wing venation structure. Acta agriculturae Slovenica, 92(december 2008)2, 139–149.
Kozmus P. Population of Carniolan bees (Apis mellifera carnica Pollman) in Slovenia and its hybridization with other subspecies based on colour of abdomen (Ugotavljanje stopnje hibridizacije populacije kranjske čebele (Apis mellifera carnica Pollman) v Sloveniji na podlagi obarvanosti obročkov na zadku). Acta argiculturae Slovenica, 98/2, 159–166, Ljubljana 2011
Kozmus P et al.: Analysis of mitochondrial DNA in honey bees (Apis mellifera) from Serbia Acta Veterinaria (Beograd), Vol. 57. No. 5-6, 465-476, 2007.
Leclercq, B. Biometry. Text and illustrations-extracts from CDROM “The Honeybee and man” (1999) http://www.beekeeping.com/leclercq/biometry.htm (8.oktober 2007)
Meixner M.D., Sheppard W.S., Poklukar J. (1993) Asymmetrical distribution of a mitochondrial DNA polymorphism between 2 introgressing honey bee subspecies, Apidologie 24, 147–153.
Moritz RFA (1991) The limitations of biometric control on pure race breeding in Apis mellifera. J Apic Res 30, 54-59
Muñoz I., Dall'Olio R., Lodesani M., De la Rúa P. (2009) - Population genetic structure of coastal Croatian honeybees (Apis mellifera carnica). Apidologie, 40: 617-626.
Nazzi, F. Morphometric analysis of honey bees from an area of racial hybridization in northeastern Italy. Apidologie, 23(1992), 89–96.
Nedić N, Stanisavljević LJ, Mladenović M, Stanisavljević J (2009) Molecular characterization of the honeybee Apis mellifera carnica in Serbia. Archives of Biological Science 61: 587-598.
14 / 15

Palmer, M.R./ Smith, D.R./ Kaftanoglu, O. Turkish honeybees: genetic variation and evidence for a forth lineage of Apis mellifera mtDNA. The Journal of Heredity, 91(2000), 42–46.
Poklukar, J. Zaključno poročilo o rezultatih raziskovalnega projekta v letu 1998. Kmetijski inštitut Slovenije, 1998, 12 str.
Poklukar, J., 1999: Ali bomo v Sloveniji v prihodnje še čebelarili z našo kranjsko čebelo? Slovenski čebelar, 101(3), str. 65-66.
Rihar, J., 1972: Vzrejajmo boljše čebele. Zavod za čebelarstvo Ljubljana, 160 str.Rihar, J. 2003. Vzrejajmo boljše čebele. Tretja dopolnjena izdaja. Ljubljana: 272 str.Ruttner, F (1988) Biogeography and taxonomy of honey bees. Springer-Verlag; Berlin, Germany.Ruttner, F./ Hanel, H. Active defense against Varroa mites in a Carniolan strain of honeybee (Apis mellifera
carnica Pollman). Apidologie 23(1992), 173–187.Ruttner, F./ Tassencourt, L./ Louveaux J. Biometrical-statistical analysis of the geographic variability of Apis
mellifera L., Apidologie 9(1978), 363–381.Susnik, S./ Kozmus, P./ Poklukar, J./ Meglic, V. Molecular characterization of indigenous Apis mellifera carnica in
Slovenia. Apidologie 35(2004), 623–636.Z. Stanimirovic, J. Stevanovic, M. Andjelkovic. Chromosomal diversity in Apis mellifera carnica from Serbia.
Apidologie 36 (2005) 31–42J. Stevanovic, Z. Stanimirovic, M. Radakovic, and S. R. Kovacevic. Biogeographic Study of the Honey Bee (Apis
mellifera L.) from Serbia, Bosnia and Herzegovina and Republi of Macedonia Based on Mitochondrial DNA Analyses. Russian Journal of Genetics, 2010, Vol. 46, No. 5, pp. 603–609
Zdešar, P., 1999: Pomen osnovne odbire in krajevnih različic kranjske čebele v Sloveniji. Slovenski čebelar, 101(2), str. 46-51.
15 / 15




























