Embed Size (px)
Citation preview

ISSN 1848‐3747
X. SIMPOZIJ PERADARSKI DANI 2013.
s međunarodnim sudjelovanjem
Hrvatska, Šibenik, 15.‐18. svibnja 2013.
X SYMPOSIUM POULTRY DAYS 2013 With International Participation Croatia, Šibenik, May 15‐18, 2013
ZBORNIK
PROCEEDINGS
Zabreb, 2013.

Izdavač / Publisher
Hrvatski veterinarski institut / Croatian veterinary institute Centar za peradarstvo / Poultry centre
Urednica / Editor Mirta Balenović
Lerkorica i prevoditeljica / Language editor Antonija Redovniković, prof.
Oblikovanje Zbornika i priprema za tisak / Proceedings design and layout Berislav Jadro
Za tisak / Of print Crescat, Zagreb
Naklada / Issue 300 primjeraka / copies
Radovi objavljeni u Zborniku recenzenti su pozitivno ocijenili.

Organizator / Organizer Hrvatski veterinarski institut
Centar za peradarstvoHeinzelova 55, Zagreb
Hrvatska
Croatian veterinary institute Poultry centre Heinzelova 55, Zagreb Croatia
Pokroviteljstvo / Under the Ausprices of Ministarstvo poljoprivrede
Republike HrvatskeMinistry of Agriculture Republic of Croatia
Ministarstvo znanosti, obrazovanja i sporta
Republike Hrvatske Ministry of Science, Education and Sport
Republic of Croatia
Svjetska udruga za znanost o peradi The World’s Poultry Science Association
Svjetska udruga veterinara u peradarstvu
World Veterinary Poultry Association
Glavni sponzor / General sponsor KOKA d.d., Varaždin / KOKA Inc., Varaždin
Predsjednik Simpozija / President of the Symposium Vladimir Savić
Tajnica Simpozija / Secretary General of the Symposium Radmila Raguž‐Đurić
Organizacijski odbor / Organising committee Mirta Balenović, Marija Berendika, Branko Bobetić, Jakov Ćorić, Zlatko Janječić,
Luka Jurinović, Fani Krstulović, Darko Majnarić, Milivoj Mikec, Radmila Raguž‐Đurić, Mirjana Runjak
Znanstveni odbor / Scientific committee Tajana Amšel Zelenika, Zdenko Biđin, Željko Cvetnić, Teufik Goletić, Boris Habrun,
Helga Medić, Suzana Milinković Tur, Mario Mitak, Stjepan Mužic, Estella Prukner‐Radovčić, Vladimir Savić, Marijana Sokolović,
Borka Šimpraga, Marina Tišljar, Olga Zorman‐Rojs, Đurđica Žutinić



Radovi objavljeni u Zborniku “Peradarski dani 2013.” biti će referirani u bazi podataka CAB International
Full of papers published in Conference Proceedings of “Poultry days 2013” will be indexed in the database CAB International

7
SADRŽAJ / CONTENTS Uvodna predavanja / Introductory lecture SVJETSKI I EU TRENDOVI U PERADARSKOJ PROIZVODNJI U RAZDOBLJU OD 2012. DO 2020. GODINE WORLD AND EU TRENDS IN POULTRY PRODUCTION IN THE PERIOD FROM 2012 TO 2020 Branko Bobetić ............................................................................................................................................... 11 CURRENT HEALTH PROBLEMS IN POULTRY PRODUCTION IN SLOVENIA AKTUALNI ZDRAVSTVENI PROBLEMI U PERADARSKOJ PROIZVODNJI U SLOVENIJI Olga Zorman Rojs, Brigita Slavec, Uroš Krapež, Rahela Juršič Cizerl, Liljana Štalcer, Marjeta Podlesnik, Tomislav Kovačič, Janez Poje, Neva Šemrov ................................................................................................ 14 Usmena izlaganja / Lectures VARIJABILNOST TROŠKOVA INKUBACIJE PRI RAZLIČITIM SANITARNIM TRETMANIMA RASPLODNIH JAJA PERADI INCUBATION COSTS VARIABILITY AT DIFFERENT SANITARY TREATMENTS OF HATCHING POULTRY EGGS Aida Kustura, Teufik Goletić, Abdulah Gagić, Emina Rešidbegović, Štefan Pintarič, Aida Kavazović, Almira Softić ............................................................................................................................................................... 18
POULTRY MEAT DECONTAMINATION WITH ELECTROLYZED OXIDIZING WATER AND CONSUMER HEALTH PROTECTION ZAŠTITA ZDRAVLJA POTROŠAČA ZBOG UPOTREBE ELEKTROOKSIDIRAJUĆE VODE KOD DEKONTAMINACIJE MESA PERADI Štefan Pintarič, Majda Biasizzo, Stanka Vadnjal............................................................................................. 23 OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA OXIDATIVE STABILITY OF COOLED AND FROZEN CHICKEN MEAT Ivan Pušić, Blanka Beer Ljubić, Zlatko Janječić, Dalibor Bedeković, Jasna Aladrović .................................. 28
CHLAMYDIA PSITTACI ASSOCIATED WITH POX VIRUS INFECTION IN LAYING HEN FLOCK CHLAMYDIA PSITTACI POVEZANA S INFEKCIJOM POX VIRUSOM U JATU KOKOŠI NESILICA Danijela Horvatek Tomić, Željko Gottstein, Vladimir Savić, Marina Tišljar, Maja Lukač, Estella Prukner-Radovčić ......................................................................................................................................................... 35 OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT POJAVNOST ESBL- I AmpC-PRODUCIRAJUĆIH SOJEVA ESCHERICHIA COLI U MESU PERADI Irena Zdovc, Majda Golob, Jerneja Ambrožič Avguštin ................................................................................. 39
PREVENTION OF MG-MS INFECTION USING LIVE VACCINES PREVENTIVA MG-MS INFEKCIJE PRIMJENOM ŽIVIH CJEPIVA László Kőrösi, János Povazsán ...................................................................................................................... 45

8
UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALUS GALUS U RADOBLJU OD 2010. DO 2012. GODINE PREVALENCE OF BACTERIA OF THE GENUS SALMOLELLA SPP. IN POULTRY SPECIES GALLUS GALLUS IN THE PERIOD BETWEEN 2010 AND 2012 Marica Lolić, Marin Periškić, Marija Agičić, Davor Balić, Mario Škrivanko, Hrvoje Krajina, Mirta Vukičević, Irena Perković, Borka Šimpraga, Kristina Matković ....................................................................................... 50
EPIDEMIOLOGICAL INVESTIGATION OF CIRCOVIRUS INFECTIONS IN PIGEONS IN SLOVENIA EPIDEMIOLOŠKO ISTRAŽIVANJE INFEKCIJE CIRCO-VIRUSIMA U GOLUBOVA U SLOVENIJI Uroš Krapež, Brigita Slavec, Adela Fratnik Steyer, Štefan Pintarič, Martin Dobeic, Olga Zorman Rojs, Alenka Dovč .................................................................................................................................................... 56
UČINAK DODATKA Ca-BUTIRATA U KRMU KOKOŠI TEŠKIH HIBRIDA NA RELATIVNU GUSTOĆU KOSTIJU – PRELIMINARNO IZVJEŠĆE THE EFFECT OF SUPPLEMENT Ca-BITURATE GIVEN IN FED OF HEAVY HYBRID BREEDING HENS ON RELATIVE BONE DENSITY - PRELIMINARY REPORT Hrvoje Mazija, Krešimir Mikulić, Željka Ervačinović ........................................................................................ 61
IMUNOGENOST CIJEPIVA PROTIV MAREKOVE BOLESTI OD SOJEVA CVI 988 I HVT FC 126 DANIH ZASEBNO ILI ISTODOBNO POSTUPKOM ZAMAGLJIVANJA TE PARENTERALNO IMMUNOGENICITY OF VACCINE AGAINST MAREK'S DISEASE STRAINS CVI 988 GIVEN SEPARATELY PARENTHERALY OR SIMULTANEOUSLY WITH STRAIN AND HVT-FC-126 GIVEN BY MEANS OF NEBULIZATION Željko Mišković, Hrvoje Mazija ........................................................................................................................ 66
PRAKTIČNA ISKUSTVA U PRIMJENI CJEPIVA OD SOLA LA SOTA PROTIV NEWCASTLESKE BOLESTI POSTUPKOM NEBULIZACIJE NETOM IZLEŽENIM TOVNIM PILIĆIMA PRACTICAL EXPIRIENCES IN APPLICATION OF VACCINE AGAINST NEWCASTLE DISEASE, STRAIN LA SOTA TO NEWLY HATCHED BROILER CHICKS BY MEANS OF NEBULISATION Željka Ervaćinović, Zvezdan Kičeec, Marica Lolić, Ivan Križek, Nikica Stevanović, Hrvoje Mazija ............... 74
TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE POULTRY RED MITE EVER ACCOMPANYING POULTRY PRODUCTION Milivoj Mikec, Tihomir Zglavnik ....................................................................................................................... 79 Poster prezentacije / Poster presentations PTIČJI HEPATITIS E - POTENCIJALNA OPASNOST ZA PERADARSKU PROIZVODNJU?! AVIAN HEPATITIS E – POTENTIAL THREAT TO THE POULTRY PRODUCTION?! Tajana Amšel Zelenika, Jelena Prpić, Marina Tišljar, Vladimir Savić, Mirta Balenović, Luka Jurinović, Lorena Jemeršić ............................................................................................................................................. 86
CRIJEVNI I IMUNOSUPRESIVNI VIRUSI POVEZANI S ETIOLOGIJOM SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA ENTERIC AND IMMUNOSSUPRESSIVE VIRUSES IN RUNTING AND STUNTING SYNDROME IN BROILER CHICKENS Marina Biđin, Darko Majnarić, Zdenko Biđin, Ivana Lojkić ............................................................................. 91

9
MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI MYCOTOXINS – CONTROL, RISK ASSESSMENT AND PREVENTIVE MEASURES IN POULTRY PRODUCTION Marijana Sokolović, Borka Šimpraga, Fani Krstulović, Marija Berendika ....................................................... 97
KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ CONTAMINATION OF POULTRY FEED WITH MYCOTOXINS DURING THE FIVE-YEAR PERIOD IN CROATIA Jelka Pleadin, Manuela Zadravec, Marijana Sokolović, Nina Perši, Ana Vulić .............................................. 105
USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS COMPARISON OF MICROBIOLOGICAL METHODS FOR DETECTION OF SALMONELLA ENTERICA SEROVAR ENTERITIDIS Marija Berendika, Borka Šimpraga, Fani Krstulović, Marijana Sokolović ....................................................... 112
PRIMJENA VIDAS TESTA ZA IZDVAJANJE BAKTERIJA RODA CAMPYLOBACTER SPP. APPLICATION OF VIDAS TEST FOR THE DETECTION OF CAMPYLOBACTER SPP. Borka Šimpraga, Fani Krstulović, Marija Berendika, Marijana Sokolović, Ivana Lohmann Janković, Ljupka Maltar .............................................................................................................................................................. 119
MONITORING THE NUMBER OF LACTOBACILLUS SPP. IN CHICKEN'S FED WITH AGARICUS BISPORUS UČINAK PLEMENITE PEČURKE AGARICUS BISPORUS NA BROJ BAKTERIJA VRSTE LACTOBACILLUS U REKTALNIM OBRISCIMA TOVNIH PILIĆA Maja Jelena Petek, Branka Gršković, Lidija Kozačinski, Danijel Špoljarić, Damir Mihelić, Ksenija Vlahović, Maja Popović, Mirta Balenović, Borka Šimpraga, Marina Tišljar, Gordan Mršić ............................................ 124 ANALIZA SOJINE SAČME NA SADRŽAJ SIROVIH BJELANČEVINA TE KONTAMINACIJU BAKTERIJAMA RODA SALMONELLA U 2012. GODINI ANALYSIS OF SOYBEAN MEAL WITH REGARD TO CRUDE PROTEIN CONTENT AND ANY CONTAMINATION OF SALMONELLA IN THE YEAR 2012 Manuela Zadravec, Jelka Pleadin, Vesna Jaki Tkalec, Darko Majnarić, Mario Mitak .................................... 129 PRERAĐENI ANIMALNI PROTEINI U HRANI ZA PERAD PROCESSED ANIMAL PROTEINS IN POULTRY FEED Manuela Zadravec, Mario Mitak, Vesna Jaki Tkalec, Darko Majnarić ............................................................ 132
ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, BATACIMA I ZABATACIMA PURANA DALMATINSKOG ZALEĐA TISSUE IN THE BREAST, THIGHS AND DRUMSTICKS OF TURKEY FROM DALMATIAN HINTERLAND Anamaria Ekert Kabalin, Sven Menčik, Mario Ostović, Hrvoje Kabalin, Mirta Balenović, Tomislav Balenović, Igor Štoković, Velimir Sušić, Kristina Starčević, Maja Maurić ....................................................... 135
PROIZVODNI REZULTATI I KVALITETA JAJA SLOBODNO DRŽANIH KOKOŠI HRVATICA PRODUCTIVITY AND EGGS QUALITY OF FREE RANGE RAISED HRVATICA HENS Zlatko Janječić, Stjepan Mužic, Dalibor Bedeković, Zoran Grgić, Gordana Duvnjak, Marina Ćurak, Tihana Bišćan ............................................................................................................................................................. 142

10
PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO BEHAVIOUR AND WELFARE ASSESSMENT OF LAYING HENS HOUSED IN CONVENTIONAL CAGES AND FREE Kristina Matković, Bara Vinković, Marija Vučemilo, Ranka Šimić, Danijel Marušić, Marica Lolić .................. 146
JAPANSKA PREPELICA (COTURNIX COTURNIX JAPONICA ) KAO MODEL U BIOMEDICINSKIM ISTRAŽIVANJIMA JAPANESE QUAIL (COTURNIX COTURNIX JAPONICA ) AS A MODEL IN BIOMEDICAL RESEARCH Gordana Gregurić Gračner, Mario Ostović, Danijela Horvatek, Damjan Gračner, Željko Pavičić ................. 154
UČINAK AKUTNE TOKSIČNOSTI L-NAME I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE (SPH) U TOVNIH PILIĆA. KRATKO PRIOPĆENJE THE IMPACT OF L-NAME AND L-ARGININE ACUTE TOXICITY INDUCED LESIONS ON ASCITES – PULMONARY HYPERTENSION SYNDROME DEVELOPMENT IN BROILER CHICKENS Marina Tišljar, Željko Grabarević, Branka Artuković, Jadranka Foršek, Petar Džaja, Marija Dinarina-Sablić, Tajana Amšel Zelenika ........................................................................................................................ 158 Okrugli stol / Round table MIKOTOKSINI U HRANI ZA ŽIVOTINJE / MYCOTOXINS IN ANIMAL FEED
MYCOTOXINS IN POULTRY: CONTAMINATION, IMPACT AND CONTROL STRATEGIES MIKOTOKSINI U PERADARSTVU: KONTAMINACIJA, UČINCI I STRATEGIJE KONTROLE RIZIKA Julia Dvorska .................................................................................................................................................. 167
MIKOTOKSINI U HRANI ZA ŽIVOTINJE – ZNAČAJ, ZAKONSKI PROPISI I PREPORUKE MYCOTOXINS IN ANIMAL FEEDS – IMPORTANCE, LEGISLATIVE CONTROLS AND RECOMMENDATIONS Marijana Sokolović .......................................................................................................................................... 172 Komercijalno predavanje / Product presentations IMPROVED GUMBORO CONTROL BY CEVAC TRANSMUNE IBD, AN IMMUNE COMPLEX VACCINE G. Bánhidi ...................................................................................................................................................... 176
SUZBIJANJE ZDRAVSTVENIH PROBLEMA U PERADARSTVU - BIOTRONIC TOP 3 Siniša Đurđević ............................................................................................................................................... 178
PERADARSKI DANI 2013. 11
SVJETSKI I EU TRENDOVI U PERADARSKOJ PROIZVODNJI U RAZDOBLJU OD 2012. DO 2020. GODINE
Branko Bobetić
Gospodarsko interesno udruženje „Croatiastočar“, Zagreb, Hrvatska
Sažetak
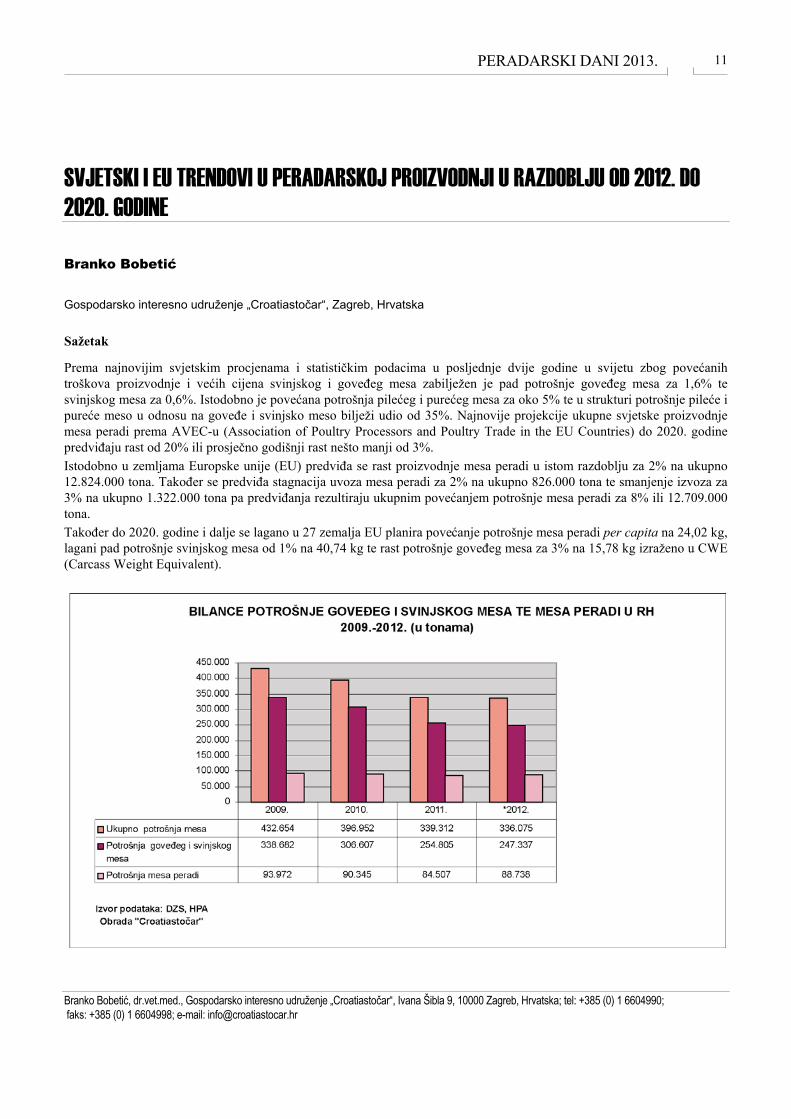
Prema najnovijim svjetskim procjenama i statističkim podacima u posljednje dvije godine u svijetu zbog povećanih troškova proizvodnje i većih cijena svinjskog i goveđeg mesa zabilježen je pad potrošnje goveđeg mesa za 1,6% te svinjskog mesa za 0,6%. Istodobno je povećana potrošnja pilećeg i purećeg mesa za oko 5% te u strukturi potrošnje pileće i pureće meso u odnosu na goveđe i svinjsko meso bilježi udio od 35%. Najnovije projekcije ukupne svjetske proizvodnje mesa peradi prema AVEC-u (Association of Poultry Processors and Poultry Trade in the EU Countries) do 2020. godine predviđaju rast od 20% ili prosječno godišnji rast nešto manji od 3%. Istodobno u zemljama Europske unije (EU) predviđa se rast proizvodnje mesa peradi u istom razdoblju za 2% na ukupno 12.824.000 tona. Također se predviđa stagnacija uvoza mesa peradi za 2% na ukupno 826.000 tona te smanjenje izvoza za 3% na ukupno 1.322.000 tona pa predviđanja rezultiraju ukupnim povećanjem potrošnje mesa peradi za 8% ili 12.709.000 tona. Također do 2020. godine i dalje se lagano u 27 zemalja EU planira povećanje potrošnje mesa peradi per capita na 24,02 kg, lagani pad potrošnje svinjskog mesa od 1% na 40,74 kg te rast potrošnje goveđeg mesa za 3% na 15,78 kg izraženo u CWE (Carcass Weight Equivalent).
Branko Bobetić, dr.vet.med., Gospodarsko interesno udruženje „Croatiastočar“, Ivana Šibla 9, 10000 Zagreb, Hrvatska; tel: +385 (0) 1 6604990; faks: +385 (0) 1 6604998; e-mail: [email protected]

SVJETSKI I EU TRENDOVI U PERADARSKOJ PROIZVODNJI U RAZDOBLJU OD 2012. DO 2020. GODINE 12
Stanje peradarske proizvodnje u RH 2011.-2012. godine
Sektor proizvodnje pilećeg i purećeg mesa
U zadnje dvije godine u reproduktivnom dijelu proizvodnje prosječno je godišnje uzgojeno oko 310.000 komada rodi-telja TL u starosti 23 tjedna. Ukupna proizvodnja ras-plodnih brojlerskih jaja procjenjuje se godišnje na oko 45.000.000 komada. Istodobno je ostvaren prosječni godiš-nji uvoz rasplodnih brojlerskih jaja od oko 20.000.000 komada te prosječni godišnji izvoz od oko 2.000.000 komada. Na osnovi podataka o inkubiranju rasplodnih brojlerskih jaja procjenjuje se prosječna godišnja proizvodnja od oko 50.000.000 jednodnevnih pilića za tov. U klaonicama pilića u RH ostvarena je ukupna proizvodnja pilećeg mesa od oko 50.000 tona, a ukupna nacionalna pro-izvodnja pilećeg mesa na bazi ukupne bilance rasplodnih jaja procjenjuje se na oko 62.000 tona. Prosječni godišnji uvoz pilećeg mesa u zadnje dvije godine iznosio je oko 13.000 tona te izvoz oko 4.000 tona, što rezultira bilancom ukupne potrošnje i prerade pilećeg mesa na godišnjoj razini od oko 70.000 tona ili oko 15 kg pilećeg mesa per capita izraženo kao CWE. Prosječni godišnji uvoz proizvoda od pilećeg mesa iznosi oko 5.640 tona, a prosječni izvoz 7.480 tona. Na osnovi bilance inkubiranja rasplodnih purećih jaja u zadnje dvije godine ostvarena je proizvodnja 1/d purića u registriranim purećim valionicama u količini od 1.465.000 komada. U klaonicama za klanje pura prosječno je godiš-nje zaklano oko 1.165.000 pura, što daje oko 13.700 tona proizvodnje purećeg mesa. Prosječni uvoz purećeg mesa od 1.615 tona te izvoz od 873 tone daje bilancu prosječne godišnje potrošnje i prerade purećeg mesa od 14.442 tone ili oko 3 kg purećeg mesa per capita CWE. Sveukupna potrošnja i prerada mesa peradi prema procjeni DZS (Državni zavod za statistiku) iznosila je u 2012. go-dini nešto više od 88.000 tona ili 19,50 kg. U 2012. godini realiziran je najveći izvoz mesa peradi i proizvoda od mesa peradi ukupne vrijednosti od oko 50.000.000 $. U strukturi uvoza mesa peradi najveći udio imaju kategorije mesa peradi za preradu. Ostvaren je ukupni izvoz proizvoda od mesa peradi u vri-jednosti od oko 37.000.000 $ te je izvoz mesnih proizvoda 3 puta veći nego uvoz. Prema metodologiji ekonomskog izračuna u poljoprivredi udio vrijednosti peradarske proizvodnje u RH u ukupnoj vrijednosti stočarske proizvodnje iznosi 17% kao i u 27 ze-malja EU.
Sektor proizvodnje konzumnih jaja
Podaci DZS pokazuju pad broja nesilica u zadnje dvije go-dine u odnosu na prethodne dvije godine za prosječno 8% godišnje.
Od ukupno 4.200.000 nesilica u 2012. godini blizu 50% smješteno je kod 149 registriranih farmi za proizvodnju konzumnih jaja, a ostatak se nalazi kod samoopskrbnih gospodarstava. Ukupna proizvodnja konzumnih jaja u RH u zadnje dvije godine procjenjuje se na oko 700.000.000 komada, od čega se kod registriranih proizvođača konzumnih jaja proizvede godišnje nešto ispod 550.000.000 komada. Procjenjuje se da je od ukupnog broja nesilica kod robnih proizvođača oko 2.000.000 komada. Udio uvoza konzumnih jaja u ukupnoj proizvodnji u 2012. godini iznosio je 1,5% ukupne proizvodnje jaja, pa je i samodostatnost u ovom sektoru kontinuirano na razini od oko 99%. Potrošnja konzumnih jaja per capita u RH procjenjuje se na oko 160 komada i značajno zaostaje u odnosu na po-trošnju u EU koja iznosi oko 225 komada. Procjena trendova peradarske proizvodnje i tržišta u RH od 2012. do 2020. godine U posljednje četiri godine u RH bilježi se kontinuirani pad ukupne nacionalne proizvodnje crvenih mesa te rast uvoza, pad izvoza te pad potrošnje. Slična je situacija i kod pilećeg i purećeg mesa, s time da je pad ukupne potrošnje i prerade mesa peradi znatno manji nego kod ostalih vrsta mesa. Samodostatnost u ukupnoj proizvodnji peradskog mesa u RH u 2012. godini, ne računajući bilance uvoza i izvoza proizvoda od mesa peradi, iznosi 85%. S obzirom na manju potrošnju mesa peradi u RH per capita u odnosu na prosjek u 27 zemalja EU realno je očekivati daljnje trendove kako u povećanju potrošnje tako i u povećanju proizvodnje svih vrsta mesa peradi. Analizirajući kretanje proizvodnje u zemljama EU od 2004. do 2011. godine evidentan je rast proizvodnje mesa peradi u 15 zemalja EU za 8%, a u 10 zemalja EU za 18%. Neke od novopridošlih članica zabilježile su rast proiz-vodnje od 8% do gotovo 80%, dok su druge zabilježile pad proizvodnje i do 20%. U uvjetima rastućeg globalnog tržišta mesa peradi u svijetu realno je očekivati u idućem sedmogodišnjem razdoblju u RH povećanje proizvodnje mesa peradi u rasponu od 20% do 30%, ovisno o vrsti mesa te kretanju potrošnje, uvoza i izvoza mesa peradi i mesnih proizvoda. U sektoru proizvodnje konzumnih jaja s obzirom na razinu sadašnje potrošnje te kontinuirani pad samoopskrbne pro-izvodnje jaja u domaćinstvima za očekivati je moguće po-većanje proizvodnje, potrošnje i prerade konzumnih jaja kod robnih proizvođača za preko 40% do 2020. godine.
Izazovi peradarske industrije
Ukupna nastojanja peradarstva RH u smislu trendova po-većanja proizvodnje i potrošnje kako mesa peradi tako i

SVJETSKI I EU TRENDOVI U PERADARSKOJ PROIZVODNJI U RAZDOBLJU OD 2012. DO 2020. GODINE 13
konzumnih jaja pretpostavljaju i čitav niz izazova od kojih će ovisiti realizacija zacrtanih programa kao što su:
- Implementacija održivosti peradarske proizvodnje u skladu s praksom višestruke sukladnosti te zaštite oko-liša i načela dobrobiti peradi u uzgoju, transportu i klao-nicama;
- Implementacija biosigurnosnih mjera i uvjeta mora osi-gurati proizvodnju zdravih, kvalitetnih i nutritivno viso-ko vrijednih proizvoda peradarske industrije. Pretpostav-ka realizacije ovih ciljeva jeste trajna i inovativna profi-laksa i preventiva u svim fazama primarne proizvodnje;
- Visoka razina sposobnosti i učinkovitosti korištenja fon-dova EU kako u segmentu realizacije budućih investicija u više od 50% postojećih kapaciteta radi dostizanja pro-izvodnih EU i svjetskih standarda te investicija u nove proizvodne kapacitete radi povećanja proizvodnje;
- Uspješnost u realizaciji zacrtanih planova u najvećoj mjeri ovisit će i o tržišnoj te proizvodnoj konkurentnosti
finalnih proizvoda, kako na domaćem tako i na ino tržištima u uvjetima potpune liberalizacije tržišta.
- Zasigurno će trebati osmisliti i novi sustav brendiranja proizvoda s osobitim naglaskom na biosigurnosne mjere, svježinu domaćih proizvoda te zdravstvenu sigurnost kroz sveobuhvatni monitoring.
- Tržišna i proizvodna globalizacija pretpostavlja i trajno restrukturiranje cjelokupnog sektora kako u proizvod-nom tako i u tržišnom smislu.
- Trajna izobrazba svih kadrova izravno ili neizravno uključenih u proizvodnom procesu, kao i sinergija prakse i istraživačke znanosti primjenom u proizvodnoj praksi trebale bi rezultirati daljnjim proizvodnim i ekonomskim razvojem ukupne peradarske proizvodnje u RH.
Izvor podataka: FAOStat, Eurostat, Godišnje izvješće AVEC-a, DZS, HPA Obrada: „Croatiastočar“

PERADARSKI DANI 2013. 14
CURRENT HEALTH PROBLEMS IN POULTRY PRODUCTION IN SLOVENIA
Olga Zorman Rojs1, Brigita Slavec1, Uroš Krapež1, Rahela Juršič Cizerl2, Liljana Štalcer2, Marjeta Podlesnik3, Tomislav Kovačič4, Janez Poje5, Neva Šemrov5
1University of Ljubljana, Veterinary Faculty, Institute of Poultry Health and Protection, Ljubljana, Slovenia 2Veterinarska ambulanta Perutnina Ptuj,Veterinarstvo d.o.o., Ptuj, Slovenia 3Veterinarska postaja Laško d.o.o., Laško, Slovenia 4PANVITA, VETERINA d.o.o., Beltinci, Slovenia 5VETAM. JATA d.o.o., Domžale, Slovenia
Summary
In the last two years, health problems in the intensive poultry production in Slovenia have been associated with different bacterial and viral diseases; some of them have not been seen in our region for years, while some were detected for the first time. Avian pathogenic Escherichia (E.) coli (APEC) and enterococci were the most prominent pathogens involved in high economic losses in broilers. APEC outbreaks led to as high as 11% mortality in some broiler flocks, and even higher were losses due to the cost of medical treatment, decreased growth rate and carcass condemnation at slaughter. According to preliminary investigations of APEC strains isolated from poultry flocks with outbreaks of colibacillosis, strains are resistant to almost all antimicrobials available for poultry treatment. In addition, extended spectrum beta-lactamase (ESBL) E. coli strains were detected. In turkeys, for the first time infection with Erysipelothrix rhusiopathiae was confirmed in a flock of 110-day old birds. High economic losses were observed in younger turkey flocks due to cardiomyopathies. The main problems in commercial layers were associated with Marek's disease. Beside this, infection with avian encephalomyelitis virus was confirmed for the first time after thirty years. Introduction
Slovenia has been and is still facing a serious economic situation, which has negative influence on all parts of economy, including agriculture. Although poultry pro-duction remains one of the most important branches in the field of farm producing animals, this economic environ-ment obviously has some negative effect. In the last two years, several health problems associated with different bacterial and viral diseases occurred. Some of them have not been seen in our region for years, while some were detected for the first time. The paper presents a short overview of health situation in poultry in the last two years.
Marek’s Disease and Avian Encephalomyelitis
Marek's disease (MD) is a neoplastic disease of poultry caused by the highly contagious alphaherpesvirus. It is the only virus-induced tumor disease for which vaccination is
available. However, vaccinated chickens can become in-fected and shed the virus into the environment (Shat and Venugopal, 2008; Baigent et al., 2006; Gimeno et al., 2012). As in other countries, all breeder and layer flocks as well as broilers raised more than 50 days should be vaccinated against MDV. A combination of ‘Rispens’ and HVT strains (CVI988 and FC-126) is used for vaccination. Such immunoprophylactic program has been working well for several years and represented the mainstay of the current control of the disease. It was also successful in multi-age farms with high standards of biosecurity practices. In the last year, MD was confirmed in five flocks, which were vaccinated against MD. The disease with high morbidity started in birds at the age of 5 weeks and lasted for more than 20 weeks. In young birds, mainly neural lesions and paralysis were seen, while in adult hens beside enlarged brachial and sciatic nerves visceral lymphomas were observed. Several outbreaks of MD confirmed that vaccination alone, without strict bio-security, is not enough to provide successful control of the
Prof. Dr. Olga Zorman Rojs, University of Ljubljana, Veterinary Faculty, Institute of Poultry Health and Protection, Ljubljana, Slovenia; Phone:+386-1-4779-242; Fax: +386 1 4779 339, e-mail:[email protected]

CURRENT HEALTH PROBLEMS IN POULTRY PRODUCTION IN SLOVENIA
15
disease. On the other hand, outbreaks could also suggest an evidence of increasing virulence of field virus (Baigent et al., 2006). Avian encephalomyelitis (AE) is one of the diseases that are expected to be under control in all countries with well-developed poultry production. Although it is well known that AE virus is very resistant in the environment, vaccination of breeders usually provides good protection of progeny (Calnek, 2008). The last AE outbreak had been confirmed before strict vaccination program was introduced in all breeder flocks. The disease was believed to be under control. In 2011, AE was observed in one smaller flock of two-week-old future layers. Day-old chicks were purchased in a big hatchery from the neighboring country. The main signs were unsteadiness and paresis. Birds were not able to move, some of them were sitting on the hocks. At necropsy, no specific lesions were seen, only anemia and dehydration were observed. Disease was confirmed by virus isolation, histologic examination and negative serologic results.
Colibacillosis and Enterococci
Escherichia coli (E. coli) and enterococci are bacterial commensals of the intestinal microflora of a variety of animals. The majority of E. coli and enterococci strains are harmless, but some are able to cause severe diseases. In the intensive poultry production, colibacilllosis is one of the most important health and economic problems. Infection with avian pathogenic E. coli (APEC) is responsible for a number of clinical conditions in birds of all ages. De-pending on the pathogenicity of the strain, host status and presence of predisposing factors, the infection manifests as colisepticemia with high mortality or localized inflam-mation in different organs. The most common conditions include peritonitis and salpingitis in broiler breeders and severe respiratory disorders in broilers. In the last years, infections with APEC in meat type birds has been associated with cellulitis, swollen head syndrome, femur necrosis and osteomyelitis. APEC outbreaks can lead to up to as high as 20% mortality, and even higher are losses due to medical treatment, decreased growth rate and carcass condemnation at slaughter (Oh et al., 2011; Shane, 2007; Kabir, 2010). Even though enterococci are considered as a normal intestinal microflora of birds and are regularly present in the poultry environment, infection with more pathogenic strains could result in acute septicemia with high mortality. In subacute or chronic form, the infection is often associated with endocarditis and leg problems, including arthritis and tenosynovitis (Oliveira et al., 2010). The most significant natural route of infection for both APEC and enterococci seem to be respiratory tract and oral infection. In addition, fecal contamination of egg can result in the E. coli or enterococci penetration through the eggshell and may spread to the chickens during hatching. The infection leads to low hatchability and very high mortality in the first week of life (Kabir, 2010).
In the last two years, infections with E. coli and enterococci caused very high economic losses in broilers as well as in broiler breeders in various forms of the disease. In some broiler flocks, APEC outbreaks caused as high as 11% mortality. Flocks were treated with anti-microbials, but very often with limited success. This situation called for special attention. Further studies were focused on detection of antimicrobial susceptibility of isolates, having in mind also the possible transition of resistance strains to humans through contaminated meat or via environmental contamination (Johnson et al., 2008). Beside this, the possible sources of infection with resistant strains were investigated. The results on E. coli and Enterococcus sp. antimicrobial resistance are shown in Figs.1 and 2.
Figure 1. Drug resistance of Escherichia coli strains isolated
from meat type poultry flocks of different ages
Figure 2. Drug resistance of Enterococcus sp. isolated from
one-day-old broiler breeder and broiler flocks
APEC isolates from poultry flocks demonstrated high rates of resistance to most of the antimicrobials available for poultry treatment. Surprisingly, higher resistance was detected in broiler breeders compared to broilers. In

CURRENT HEALTH PROBLEMS IN POULTRY PRODUCTION IN SLOVENIA 16
addition, the extended spectrum beta-lactamase (ESBL) E. coli strains were detected in some flocks of day-old breeders brought in from abroad. All enterococci isolated from day-old chicks were sus-ceptible to ampicillin, amoxicillin and florfenicol. Isolates obtained from day-old broiler flocks demonstrated high resistance rate (from 50% to 100%) to trimethoprim-sulfa, oxytetracycline and enrofloxacin. All isolates from broiler breeders were resistant to lincospectin.
Dilated Cardiomyopathy in Turkeys
In commercial turkeys, spontaneous cardiomyopathy is a disease of young birds. Some reports suggest that the syndrome could be responsible for up to 10% of mortality in young flocks. The etiology of the disease has not yet been clarified. It could be related to the hypoxia during in-cubation, poor management condition, and genetics. Tur-keys with dilated cardiomyopathy have reduced enzymes involved in energy supply, diminished concentration of ATP, and reduced concentration of fatty acids. Very similar pathologic lesions could be induced by using low doses of furazolidon, although the concentration of poly-unsaturated to fatty acids is increased in that case (Lax et al., 1994; Gyenai et al., 2012). Spontaneous cardiomyopathy occurred in two turkey flocks in a very short period. Farms were located in different areas; the origin of the day-old birds was not the same, and feed supply was also different. In both flocks, very high daily mortality was reported at the age of two weeks. The mortality was higher in males than in females. In four weeks, the mortality reached 10% in one flock and 15% in the other flock. Beside sudden death, almost no other clinical signs were observed at the beginning of the disease. On post mortem examination, a greatly enlarged heart (3 to 4 times) due to dilatation of the ventricles, congested lungs, a swollen liver, enlarged kidneys and spleen were seen. In almost all birds, ascites and hydro-pericardium were present. On histologic examination, degeneration of myofibers and myocarditis were seen in abnormal hearts. Severe microscopic lesions were also observed in livers. Routine bacteriology confirmed E. coli. Molecular investigations of different organs and intestinal contents confirmed the presence of adenovirus in one flock. In the other flock, reovirus was detected by electron microscopy. Very limited biochemical analyses of serum samples showed an increased potassium level. Analyses of the feed samples showed a slightly increased level of proteins. Conclusions Antimicrobials are still widely used in poultry industry to reduce health problems. The use of antimicrobials can lead
to the spread of resistance genes among commensal and pathogenic strains and as such, antimicrobial use should be reduced to a minimum. We have to be focused on other approaches. Strict biosecurity, effective immunoprophy-lactic programs and good management seem to be the right way to minimize the predisposing factors that can provoke health disorders in poultry flocks.
References
BAIGENT, S. J., L. P. SMITH, V. K. NAIR, R. J. CURRIE (2006): Vaccinal control of Marek disease: current challenges, and future strategies to maximize protection. Vet. Immunol. Immunopathol. 112 (1-2), 78-86.
CALNEK, B. W. (2008): Avian encephalomyelitis. In: Diseases of Poultry, 12th edn. (Y. M. Saif, A. M. Fadly, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. E. Swayne, Eds.) Black-well Publishing, Ames, Iowa, 430-441.
GIMENO, I. M., A. L. CORTES, R. L. WITTER, A. R. PANDIRI (2012): Optimization of the protocols for double vaccination against Marek’s disease by using commercially available vaccines: evaluation of protection, vaccine repli-cation, and activation of T cells. Avian Dis. 56 (2), 295-305.
GYNEAI, K., D. KAMARA, T. GENG, R. LEE-PYLE, F. PIERSON, C. LARSEN, E. SMITH (2012): An assessment of echocardiography as a diagnostic tool for dilated cardiomyopathy in Turkeys (Meleagris gallopavo). Am. J. Anim. Vet. Scie. 7(3), 120-125.
LAX, D., R. T. HOLMAN, S. B. JOHNSON, S. L. HANG, Y. LI, G. R. NOREN, N. A. STALEY, S. EINZIG (1994): Myo-cardial lipid composition in turkeys with dilated cardio-myopathy. Cardiovasc. Res. 28, 407-413.
JOHNSON, T. J., Y. WANNEMUEHLER, S. J. JOHNSON, A. L. STELL, C. DOETKOTT, J. R. JOHNSON, R. S. KIM, L. SPANJAARD, L. K. NOLAN (2008): Comparison of extraintestinal pathogenic Escherichia coli strain from human and avian sources reveals a mixed subset representing potential zoonotic pathogens. Appl. Environ. Microbiol. 74 (22), 7043-7050.
OH, J, Y., M. S. KANG, J. M. KIM, B. K. AN, E. A. SONG, J. Y. KIM, E. G. SHIN, M. J. KIM, J. K. KWON, Y. K. KWON (2011): Characterization of Escherichia coli isolates from laying hens with colibacillosis on 2 commercial egg-producing farms in Korea. Poultry Sci. 90, 1948-1954.
OLIVEIRA, M., V. SANTOS, A. FERNANDES, F. BER-NARDO, C. L. VILELA (2010): Antimicrobial resistance and in vitro biofilm-forming ability of enterococci from intensive and extensive farming broilers. Poultry Sci. 89, 1065-1069.
SHANE, S. M. (2007): E. coli continues to influence poultry health. World Poultry 23, 44-45.
SHAT, K. A., N. VENUGOPAL (2008): Marek’s disease. In: Diseases of Poultry, 12th edn. (Y. M. Saif, A. M. Fadly, J. R. Glisson, L. R. McDougald, L. K. Nolan, D. E. Swayne, Eds.) Blackwell Publishing, Ames, Iowa, 452-514.

CURRENT HEALTH PROBLEMS IN POULTRY PRODUCTION IN SLOVENIA
17
AKTUALNI ZDRAVSTVENI PROBLEMI U PERADARSKOJ PROIZVODNJI U SLOVENIJI
Sažetak
Zdravstveni problemi u intenzivnoj peradarskoj proizvodnji u Sloveniji kroz posljednje dvije godine odnose se na različite bakterijske i virusne bolesti; neke od njih nisu godinama bile zabilježene u ovom području, dok su neke otkrivene prvi put. Najizraženiji patogeni povezani s visokim gubicima kod brojlera bili su ptičja patogena Escherichia (E.) coli (APEC) i enterokoki. Epidemije APEC-a rezultirale su visokom smrtnošću od 11% u nekim jatima brojlera, dok su gubici zbog troškova za lijekove, smanjene stope rasta i povlačenja trupla nakon klanja bili još viši. Prema prvim ispitivanjima sojeva APEC izoliranih iz jata peradi s epidemijom kolibaciloze ovi sojevi su otporni na gotovo sve antimikrobne agense koji su dostupni za liječenje peradi. Uz to, otkriveni su i sojevi E. coli koji proizvode beta-laktamaze proširenog spektra (ESBL). Kod purana je infekcija bakterijom Erysipelothrix rhusiopathiae prvi put potvrđena u jatu od 110 dana starih ptica. Zabilježeni su visoki ekonomski gubici uzrokovani kardiomiopatijama u jatima mlađih ptica. Glavni problemi kod komercijalnih nesilica bili su povezani s Marekovom bolešću. Uz to, infekcija virusom ptičjeg encefalomijelitisa potvrđena je prvi put nakon trideset godina.

PERADARSKI DANI 2013. 18
VARIJABILNOST TROŠKOVA INKUBACIJE PRI RAZLIČITIM SANITARNIM TRETMANIMA RASPLODNIH JAJA PERADI
Aida Kustura1, Teufik Goletić1, Abdulah Gagić1, Emina Rešidbegović1, Štefan Pintarič2, Aida Kavazović1, Almira Softić1
1Sveučilište u Sarajevu, Veterinarski fakultet, Zavod za zootehniku i peradarstvo, Sarajevo, Bosna i Hercegovina 2Sveučilište u Ljubljani, Veterinarski fakultet, Zavod za animalnu higijenu i etologiju, Ljubljana, Republika Slovenija
Sažetak
Krajnji uspjeh rada jedne valionice ovisi, uz ostalo, od fiksnih i varijabilnih troškova inkubacije nastalih u procesu proizvodnje jednodnevnih pilića. S tim u vezi, cilj istraživanja bio je provjera primjenjivosti ekstremnih ultravioletnih zraka i negativnih iona u proizvodnoj praksi valionice brojlerskih pilića kao alternativnih sredstava sanitarnog tretmana rasplodnih jaja te usporedbom s konvencinalnim načinom sanitarnog tretmana parama formaldehida (kontrolni tretman) utvrditi njihovu ekonomsku opravdanost izraženu kroz varijabilne troškove inkubacije. Varijabilni troškovi sanitarnog tretmana 16500 rasplodnih jaja kroz četiri eksperimentalna ciklusa inkubacije parama formaldehida, ultravioletnim zrakama te kombiniranog tretmana negativnim ionima i ultravioletnim zrakama iznosili su 308,88 KM, 60,44 KM odnosno 106,80 KM. U usporedbi s referentnim tretmanom parama formaldehida udio varijabilnih troškova u cijeni jednodnevnog pileta iznosio je 18,72% kod primjene ultravioletnih zraka, te 30,76% kod kombinirane primjene ultravioletnih zraka prije početka inkubacije i primjene negativnih iona u predvalioniku, kontinuirano tokom prvih 18 dana inkubacije. Kroz dobre ekonomske pokazatelje validirani su i potvrđeni dobri higijenski te proizvodni učinci alternativnih načina sanitarnih tretmana rasplodnih jaja peradi u proizvodnim uvjetima. Kombinirana primjena ultravioletnih zraka i negativnih iona kao zamjena za fumigaciju rasplodnih jaja peradi parama formaldehida ima ekonomsku, ali i svaku drugu opravdanost. Ključne riječi: dezinfekcija, ekonomski troškovi, inkubacija, negativni ioni, UV zrake, valivost
Uvod
Jedna od osnovnih bioloških pretpostavki uspjeha in-tenzivne peradarske proizvodnje, bez obzira na vrstu peradi ili proizvodne ciljeve, je proizvodnja kvalitetnih rasplodnih jaja kao osnovnog preduvjeta potpunog oči-tovanja svih genetskih potencijala selekcioniranih jedinki uz maksimalno moguće smanjenje rizika križnih konta-minacija, vertikalnog prijenosa uzročnika specifičnih bo-lesti i ugrožavanja zdravlja peradi. Među brojne bitne negenetske čimbenike koji su pretpostavka uspjeha inten-zivnog peradarstva spadaju biosigurnosne mjere, od kojih se naročito ističu preventivni sanitarni tretmani koji pod-razumijevaju provođenje profilaktične dezinfekcije. Stoga higijena i sanitacija zauzimaju posebno mjesto u progra-mima profilakse patoloških stanja kod peradi, ali i u ostvarenju „all in – all out“ načela kao osnovnog tehno-loškog postulata ove proizvodnje. U suvremenoj praksi
peradarske proizvodnje pod pojmom 'sanitarni tretman rasplodnih jaja' podrazumijeva se samo profilaktična de-zinfekcija rasplodnih jaja koja prethodno moraju biti oda-brana i tretirana po jasno utvrđenom redoslijedu tehno-loških tretmana – od farme gdje su snešena, preko skladišta i sredstava transporta pa do ulaska u valionicu (Hristov, 2002.; Kustura, 2008.). Ukoliko tehnološki postupci kao i sam sanitarni tretman rasplodnih jaja nisu provedeni na odgovarajući način moguće je očekivati smanjenu valivost, lošu kvalitetu, slab prirast i povećanu smrtnost izleženih pilića. Cilj istraživanja bio je provjera primjenjivosti ekstremnih ultravioletnih zraka i negativnih iona u proizvodnoj praksi valionice brojlerskih pilića kao alternativnih sredstava sa-nitarnog tretmana rasplodnih jaja te usporedbom s kon-vencinalnim načinom sanitarnog tretmana parama for-maldehida (kontrolni tretman) njihova ekonomska oprav-danost izražena kroz varijabilne troškove inkubacije.
Doc. dr. Aida Kustura, Veterinarski fakultet Univerziteta u Sarajevu, Zavod za zootehniku i peradarstvo, Zmaja od Bosne br. 90, Sarajevo, Bosna i Hercegovina; Tel. i fax: 00387 33 61 88 32; E-mail: [email protected]

VARIJABILNOST TROŠKOVA INKUBACIJE PRI RAZLIČITIM SANITARNIM TRETMANIMA RASPLODNIH JAJA PERADI
19
Materijal i metode
Biološki materijal
Pokus je proveden u inkubatorskoj stanici domaćeg repro-centra teške linije, a osnovni eksperimentalni materijal predstavljala su rasplodna jaja kroz četiri inkubacijska razdoblja.
Dizajn pokusa
Dezinfekcija rasplodnih jaja parama formaldehida pro-vedena je primjenom 37%-tnog tehničkog formalina (vodeni rastvor formaldehida, HCHO). Oslobađanje para formaldehida je inicirano dodavanjem formalina u mješa-vinu destilirane vode i kalijeva permanganata (KMnO4) u odgovarajućim omjerima unutar komora predviđenih za plinjenje jaja. Sanitarni tretman rasplodnih jaja ultravioletnim zrakama prije ulaganja u predvalionike proveden je pomoću UV lampe koja emitira ultravioletne zrake valne dužine 254 nm, odnosno ultravioletne zrake skupine C koje imaju dokazana germicidna svojstva (Samek, 2000.).
U komori za sanitarni tretman rasplodnih jaja obloženoj aluminijskom folijom valionička jaja su s dva metra uda-ljenosti od UV izvora izlagana UV zrakama tri puta na dan po 15 minuta i uz ozračenost od 474 mW/ s/cm2. Kombinirani sanitarni tretman rasplodnih jaja ostvaren je primjenom UV zraka prije početka inkubacije (na ranije opisani način), a potom su jaja tijekom prvih 18 dana in-kubacije kontinuirano izlagana negativnim ionima stalnim radom ionizatora Clear Air kao izvora negativnih iona. Ovaj tip ionizatora ne zahtijeva posebnu instalaciju, a stavlja se u rad uključivanjem u električnu mrežu napona 220 V. Ionizator Clear Air proizvodi 2,5x1012 ili 2,5 trilijuna negativnih iona, odnosno 1000 negativnih iona po cm3 zraka unutar područja od 24 m2 površine.
Rezultati
Sanitarni tretman rasplodnih jaja parama formaldehida (kontrolni tretman)
Varijabilni troškovi inkubacije sanitarnog tretmana ras-plodnih jaja parama formaldehida prikazani su u tablici 1.
Tablica 1. Troškovi inkubacije rasplodnih jaja sanitarno tretiranih parama formaldehida
KM* po Pokazatelj
Pojedinačna cijena (KM*)
Cijena po tretmanu (KM*)
Cijena za sva četiri tretmana (KM*) jajetu piletu
Formalin 50,00 24,00 96,00 0,0014 0,002 KMnO4 50,00 48,00 192,00 0,0028 0,004 Destilirana voda 2,00 0,96 3,84 0,00006 0,00008 Električna energija 0,00 0,00 0,00 0,00 0,00 Radna snaga 2,84 4,26 17,04 0,0003 0,0004 Oprema 0,00 0,00 0,00 0,00 0,00 Ukupno 77,22 308,88 0,00456 0,00648
*Konvertibilna marka (1 BAM = 0,51129 €) Table 1. Incubation costs for hatching eggs sanitary treated with formaldehyde vapors
KM* Index Individual price (KM*) Price per
treatment (KM*) Price for all four
treatments (KM*) per egg per chicken Formalin 50.00 24.00 96.00 0.0014 0.002 KmNO4 50.00 48.00 192.00 0.0028 0.004 Distilled water 2.00 0.96 3.84 0.00006 0.00008 Electric power 0.00 0.00 0.00 0.00 0.00 Labor 2.84 4.26 17.04 0.0003 0.0004 Equipment 0.00 0.00 0.00 0.00 0.00 Total 77.22 308.88 0.00456 0.00648
* 1 BAM = 0,51129 €

VARIJABILNOST TROŠKOVA INKUBACIJE PRI RAZLIČITIM SANITARNIM TRETMANIMA RASPLODNIH JAJA PERADI 20
Sanitarni tretman rasplodnih jaja ultravioletnim zrakama
Varijabilni troškovi inkubacije sanitarnog tretmana rasplodnih jaja UV zrakama prikazani su u tablici 2. Tablica 2. Troškovi inkubacije rasplodnih jaja sanitarno tretiranih ultravioletnim zrakama
KM* po Pokazatelj Pojedinačna cijena (KM*)
Cijena po tretmanu (KM*)
Cijena za sva četiri tretmana (KM*) jajetu piletu
Formalin 0,00 0,00 0,00 0,00 0,00 KMnO4 0,00 0,00 0,00 0,00 0,00 Destilirana voda 0,00 0,00 0,00 0,00 0,00 Električna energija 1,00 0,20 0,80 0,0000119 0,000016 Radna snaga 2,84 14,20 56,80 0,0008452 0,0011401 Oprema 60,00 0,71 2,84 0,0000422 0,000057 Ukupno 15,11 60,44 0,0008993 0,0012131 % u odnosu na kontrolni tretman 19,58 18,72
* Konvertibilna marka (1 BAM = 0,51129 €) Table 2. Incubation costs for hatching eggs sanitary treated with ultraviolet rays
KM* Index Individual price (KM*)
Price per treatment (KM*)
Price for all four treatments (KM*) per egg per chicken
Formalin 0.00 0.00 0.00 0.00 0.00 KmNO4 0.00 0.00 0.00 0.00 0.00 Distilled water 0.00 0.00 0.00 0.00 0.00 Electric power 1.00 0.20 0.80 0.0000119 0.000016 Labor 2.84 14.20 56.80 0.0008452 0.0011401 Equipment 60.00 0.71 2.84 0.0000422 0.000057 Total 15.11 60.44 0.0008993 0.0012131 % compared to control treatment 19.58 18.72
* 1 BAM = 0,51129 €
Sanitarni tretman rasplodnih jaja negativnim ionima
Varijabilni troškovi inkubacije sanitarnog tretmana rasplodnih jaja UV zrakama u kombinaciji s negativnim ionima prikazani su u tablici 3. Tablica 3. Troškovi inkubacije rasplodnih jaja sanitarno tretiranih kombinacijom ultravioletnih zraka i negativnih iona
KM* po Pokazatelj Pojedinačna cijena (KM*)
Cijena po tretmanu (KM*)
Cijena za sva četiri tretmana (KM*) jajetu piletu
Formalin 0,00 0,00 0,00 0,00 0,00 KMnO4 0,00 0,00 0,00 0,00 0,00 Destilirana voda 0,00 0,00 0,00 0,00 0,00 Električna energija 1,00 9,04 36,16 0,000538 0,0006749 Radna snaga 2,84 14,20 56,80 0,0008452 0,0011401 Oprema 293,70 3,46 13,84 0,0002059 0,0002583 Ukupno 26,70 106,80 0,0015892 0,0019933 % u odnosu na kontrolni tretman 34,85 30,76
*Konvertibilna marka (1 BAM = 0,51129 €)

VARIJABILNOST TROŠKOVA INKUBACIJE PRI RAZLIČITIM SANITARNIM TRETMANIMA RASPLODNIH JAJA PERADI
21
Table 3. Incubation costs for hatching eggs sanitary treated a combination of negative ions and ultraviolet rays
KM* Index Individual price
(KM*) Price per treatment
(KM*) Price for all four
treatments (KM*) per egg per chicken Formalin 0.00 0.00 0.00 0.00 0.00 KmNO4 0.00 0.00 0.00 0.00 0.00 Distilled water 0.00 0.00 0.00 0.00 0.00 Electric power 1.00 9.04 36.16 0.000538 0.0006749 Labor 2.84 14.20 56.80 0.0008452 0.0011401 Equipment 293.70 3.46 13.84 0.0002059 0.0002583 Total 26.70 106.80 0.0015892 0.0019933 % compared to control treatment 34.85 30.76
* 1 BAM = 0,51129 € Rasprava i zaključci
Inkubacija oplođenih rasplodnih jaja jedna je od najosjet-ljivijih faza u reprodukciji peradi te su ostvareni rezultati valenja, iskazani kroz procenat valivosti, uvjerljiv argu-ment provodljivosti određenih inovativnih tehnoloških rje-šenja (Elbol i Brake, 2004.; Kustura i Goletić, 2004.; Hafez, 2007.). Najčešće ovakva rješenja svoju znanstvenu, stručnu i praktičnu valorizaciju dobivaju na osnovi pro-izvodnih rezultata, ali i njihovih krajnjih financijskih učinaka. Naime, uspjeh rada jedne valionice definiraju na prvom mjestu fiksni i varijabilni troškovi inkubacije nastali u pro-cesu proizvodnje jednodnevnih pilića (Sheppard, 2004.; Vučemilo, 2008.), te je uz higijenske i proizvodne učinke jedan od osnovnih ciljeva naših istraživanja bio upravo de-finiranje troškovnih pokazatelja inkubacije rasplodnih jaja sanitarno tretiranih u skladu s dizajnom pokusa. Na osnovi dobivenih rezultata utvrdili smo da je kombinirani sanitarni tretman rasplodnih jaja imao najbolju valivost (79,73%), koja je za oko 5% bila bolja u odnosu na valivost od 74,14% iz jaja tretiranih samo ultravioletnim zrakama, od-nosno za oko 9% u odnosu na kontrolni tretman jaja para-ma formaldehida. Finasijski izraženo, varijabilni troškovi sanitarnog tretmana 16500 rasplodnih jaja kroz četiri eks-perimentalna ciklusa inkubacije parama formaldehida, ultravioletnim zrakama te kombiniranog tretmana negativ-nim ionima i ultravioletnim zrakama iznosili su 308,88 KM, 60,44 KM odnosno 106,80 KM. S obzirom na ostvarene higijenske i proizvodne rezultate, kombinirani sanitarni tretman rasplodnih jaja ultraviolet-nim zrakama i negativnim ionima istodobno je imao i naj-povoljniju troškovnu strukturu cijene rasplodnog jajeta i jednodnevnog brojlerskog pileta (Kustura i sur., 2012.). Također, varijabilni troškovi sanitarnog tretmana ultravio-letnim zrakama te kombiniranog tretmana negativnim ionima i ultravioletnim zrakama bili su pet odnosno dva puta manji u usporedbi s kontrolnim sanitarnim tretmanom parama formaldehida. Imajući u vidu navedeno te činje-nicu da su i higijenski pokazatelji kombiniranog sanitarnog
tretmana bili bolji ili slični onima utvrđenim pri kontrol-nim tretmanima parama formaldehida, kombinirani sani-tarni tretman primijenjen u skladu s dizajnom ovoga istra-živanja predstavlja potpuno prihvatljivu metodu sanitacije rasplodnih jaja u inkubatorskim stanicama (Kustura i sur., 2012.). Osobita prednost ovako kombiniranog sanitarnog tretmana rasplodnih jaja ogleda se i u činjenici da nje-govom ispravnom primjenom, uz osnovne mjere zaštite, zdravlje ljudi nije ugroženo, pogotovo ne na način na koji je ono ugroženo pri, za naše prilike tradicionalno uobiča-jenom sanitarnom tretmanu rasplodnih jaja parama formal-dehida (Samek, 2000; Wakamura, 2004.). Zaključno, kroz dobre ekonomske pokazatelje sublimirani su i potvrđeni dobri higijenski te proizvodni učinci alter-nativnih načina sanitarnih tretmana rasplodnih jaja u proizvodnim uvjetima (Kustura i sar., 2012.). Naročito je važna i afirmativna činjenica da je kombinirana primjena ultravioletnih zraka i negativnih iona, kao moguća zamjena za fumigaciju jaja parama formaldehida, pokazala ne samo ekonomsku opravdanost, nego i značajno manji rizik za zdravlje ljudi te nesporne higijenske, tehnološke i eko-nomske prednosti.
Literatura
ELBOL, O., J. BRAKE (2004): Identification of critical periods for turning broiler hatching eggs during incubation. Br. Poultry Sci. 45 (5), 631-637.
KUSTURA, A., T. GOLETIĆ (2004): Tehnologija peradarske proizvodnje. Sarajevo.
HAFEZ, M.H. (2007): Breeder farms and hatchery as integrated operation. Lohmann Information, Vol. 42.
HRISTOV, S. (2002): Zoohigijena, Beograd. KUSTURA, A. (2008): Higijenski i proizvodni aspekti inkubacije
nakon različitih sanitarnih tretmana rasplodnih jaja. Disertacija. Veterinarski fakultet Univerziteta u Sarajevu, Sarajevo.
KUSTURA, A., T.GOLETIĆ, S. HADŽIABDIĆ, A. GAGIĆ, E. REŠIDBEGOVIĆ, A. GAGIĆ, Š. PINTARIČ (2012): Higijenski aspekti inkubacije nakon tri različita sanitarna

VARIJABILNOST TROŠKOVA INKUBACIJE PRI RAZLIČITIM SANITARNIM TRETMANIMA RASPLODNIH JAJA PERADI 22
tretmana rasplodnih jaja peradi, 24. Znanastveno-stručno-edukativni seminar Djelatnost dezinfekcije, dezinsekcije, deratizacije i zaštite uskladištenih poljoprivrednih proizvoda, 20.-23. ožujka 2012., Split, Hrvatska, 243-251.
SAMEK, D. (2000): Opća fizika sa osnovama biofizike. Sarajevo SHEPPARD, A. (2004): The Structure and Economics of Broiler
Production in England. Centre for Rural Research, Exeter, England.
TOMOKO WAKAMURA, MAKI SATO, A. SATO, T. DOHI, K. OKI, N. ASOU, S. HAGATA, HIROMI TOKURA (2004): A preliminary study on influence of negative air ions generated from pajamas on core, body temperature and salivary IgA during night sleep, Int. J. Occup. Med. Environ. Health, 295-298.
VUČEMILO, M. (2008): Higijena i bioekologija u peradarstvu. Zagreb.
INCUBATION COST VARIABILITY AT DIFFERENT SANITARY TREATMENTS OF HATCHING POULTRY EGGS
Summary
The ultimate success of a hatchery depends, among other factors, on the fixed and variable costs of incubation incurred in the production of day-old chickens. In this regard, the objective of the research was to check the applicability of the extreme ultraviolet rays and negative ions in the production practice of the broiler chicken hatchery, as the alternative means of the hatching egg sanitary treatment and, by comparison with the conventional sanitary treatment by formaldehyde vapors (control treatment), their economic justification expressed as variable costs of incubation. Variable costs of the sanitary treatment for 16,500 hatching eggs, through four experimental incubation cycles, with formaldehyde vapors, ultraviolet rays and the combined treatment with negative ions and ultraviolet rays were 157.93 €, 30.90 € and 54.60 €, respectively. Compared with the reference treatment by formaldehyde vapors, the share of variable costs for the experimental treatment in the price of one-day chicken was 18.72% when using ultraviolet rays and 30.76% for the combined use of ultraviolet rays before the start of incubation and application of negative ions in the incubator continuously during the first 18 days of incubation. Using positive economic indicators, good hygienic and production effects of the alternative sanitary treatments of the poultry hatching eggs were validated and confirmed in production conditions. Combined application of ultraviolet rays and negative ions as a substitute for formaldehyde vapor fumigation of the poultry hatching eggs has both economic and every other justification. Key words: disinfection, economic costs, incubation, negative ions, UV rays, hatchability

PERADARSKI DANI 2013.
23
POULTRY MEAT DECONTAMINATION WITH ELECTROLYZED OXIDIZING WATER AND CONSUMER HEALTH PROTECTION
Štefan Pintarič, Majda Biasizzo, Stanka Vadnjal
University of Ljubljana, Veterinary Faculty, Ljubljana, Slovenia
Summary
Durability of poultry meat is also dependent of microbiological contamination. During slaughtering, the surface of chicken carcasses can often be contaminated with intestinal contents. This is the main reason for contamination of poultry meat with Campylobacter spp. Neutral electrolyzed oxidizing water (NEOW) is a new generation biocide, which mainly operates on the abstraction of electrons from the environment, does not leave residuals on the surface and is not needed to rinse. We tried to find out whether it could be used for rinsing of contaminated chicken carcasses and whether this could improve the durability and safety of food. For our experiment, we took parts of chicken carcasses from one flock randomly. We selected parts of meat with skin and without it. We selected control samples and samples that were treated with 5% NEOW. All samples were then packed into controlled atmosphere (70% N2 and 30% CO2) and stored at +4 °C. Samples were tested from first to tenth day of storage for total count and presence of Campylobacter (C.) jejuni and C. coli. Since we expect a relatively low number of microorganisms on the meat surface, for evaluation of NEOW power we in parallel exposed test organisms Staphylococcus aureus on glass surfaces. On average, the total count on the meat with skin was by 80% lower after washing than without it. The results suggested a potential use of NEOW in the treatment of chicken carcasses and parts of meat with the skin. We believe that with proper use of NEOW, extended shelf life of foods can be achieved as well as reduction of contamination with Campylobacter jejuni/coli. This could improve the microbiological condition of poultry meat and so reduce the possibility of alimentary infection of consumers. In case of premises and equipment, NEOW was applied to surfaces by means of the Zeljet CS 1 mobile system (Swiss Steriplant AG). To apply it to the surfaces, we used compressed air with gas flow of 3.300 m3/min. We applied the SPN spray to the target surface at various distances with the help of the Zeljet CS1 mobile system in order to test its functionality. We also made comparison between the effectiveness of the application and other ordinary modes of biocide application. Key words: neutral electrolyzed oxidizing water, decontamination, aerosolization, Campylobacter jejuni, Campylobacter coli, poultry Introduction
Campylobacter (C.) jejuni and C. coli cause the most common gastrointestinal bacterial zoonoses. In 2009, the rate of occurrence in the EU countries was 45.6 cases per 100,000 inhabitants (EFSA, 2011). Salmonellosis is the second most frequently occurring zoonosis since its occurrence rate is 23.7 cases per 100,000 inhabitants. Campylobacteriosis usually occurs sporadically (EFSA, 2010); however, there have been cases of outbreaks. The main source of campylobacter infection is poultry. Many flocks of broiler chicken are infected by the bacteria C. jejuni and C. coli (EFSA, 2011; EFSA 2010). According to researchers, from 10.8 to 90 per cent of all poultry has
been contaminated by these organisms. In most cases, the infection happens during slaughtering. There are more critical points where contamination might happen, such as during transport, in the process of slaughtering, letting out the blood, treating carcasses with steam, feather removal, removal of internal organs and meat, and cooling (Tam et al., 2003). The most important is said to be the process of removing the offal from the carcass, which is made mechanically. During the process, many intestinal damages occur and consequently spilling of intestinal content happens, since this liquid runs down the skin and into the thoracic and abdominal cavities. There is also a possibility of cross-contamination that can happen when cutting chicken carcasses (Dickins et al., 2002). Campylobacters
Dr. Štefan Pintarič, DVM, Univerza v Ljubljani, Veterinarska fakulteta, Inštitut za higieno okolja in živali z etologijo, Gerbičeva 60, 1000 Ljubljana, Slovenija; Tel.: +38614779141; Fax.: +38614779279; E-mail: [email protected]

POULTRY MEAT DECONTAMINATION WITH ELECTROLYZED OXIDIZING WATER AND CONSUMER HEALTH PROTECTION 24
are thermolabile microorganisms, while the most common reasons for alimentary infection is either insufficient thermal treatment of meat or cross-contamination during the preparation of foodstuff. The number of campylobacter depends on many factors, such as the presence of carbon dioxide and oxygen, cooling, freezing, heating, drying, disinfection, etc., since all these processes influent the survival of microbes. In the past fifteen years, there have been many researches done on survival of Campylobacter spp. on chicken carcasses, which indicates the importance of monitoring and also points out the futility of efforts that had been done to fight campylobacterioses. The modern economic trends demand a reliable livestock production. Above all, establishment of a well-defined prevention, which also includes reliable disinfection of the livestock objects, is of vital importance, thus it is necessary to select reliable biocides. Besides their effectiveness, one expects that they will also be ecologically acceptable and their usage nontoxic to people and animals. The idea of using the electrolyzed oxidizing water (EOW) in livestock farming is gaining on its attractiveness owing to its ecological acceptability, its low toxicity and its mechanism of action (Fenner et al., 2006). When treating the surfaces of animal trunks, some biocides have also been used; with their use the level of biological contamination is trying to be decreased. The main problem is caused by residual biocide. The advantage of neutral electrolyzed oxidizing water (NEOW) is that its physical way of activity does not leave residue of any kind on the surfaces and it also offers an option of using it for animal carcass decontamination. The first methods of EOW winning were “two-chamber” systems, which resulted in the products of acid fraction (pH about 1.0; redox 1.200 mV) and alkaline phase (pH about 12-13.0; redox -800 mV). The biocide effect has acid phase, whose weak properties are corrosiveness and instability of the end product. And the product of “four-chamber” system of EOW winning is based on winning of stable EOW (fit for use for 6 months) and with pH in neutral range (pH 7.3). EOW is meta-stable and some physical factors affect its stability. In stockbreeding production, the common way of applying biocide solution to the breeding areas is by means of high-pressure device. The use of EOW in high-pressure systems seems to be unsuitable as it affects biocide properties essentially. For the purpose of disinfection with EOW, a special type of spraying nozzle for disinfection emerged on the market, which we tested in practical conditions (Fabrizzio and Cutter, 2003; Fabrizzio and Cutter, 2004; Hsu and Kao, 2004; Hsu, 2005; Kamikado et al. 2004; Liao et al., 2007; Shun-Yao, 2005; Walker et al., 2005). NEOW affects microorganisms in many ways: - Redox potential. NEOW is extremely poor in electrons. Thus it has a tendency to be in an electro-neutral environment, which can be reached by getting electrons from the environment. So, if microorganisms happen to be
in the vicinity of NEOW, it will steal electrons from the bacterial cell wall, which consequently means the collapse of the microorganism. - Oxide and superoxide ions. The emergence of these ions places the NEOW into the group of oxygen emitting biocide means. The important features of this group are rapid activity and ecological acceptance of its decom-posing products. On the other hand, there are also some disadvantages, such as turbulent reaction in the presence of organic stuff and special storage conditions (the need of special packaging with special air vents that release the pressure in the packaging), corrosiveness, irritation of respiratory organs and caustic properties. All the above disadvantages cannot be traced with the NEOW. - Chlorine, chlorides and sodium hypochloride. These compounds are formed in the process of electrolysis and are placed among the biocide compounds. However, during electrolysis they are formed in rather small amounts. - Acidity. Acidity is another important factor of disinfection. Values between 1 and 2 are distinctively acid, therefore acting as biocides to some level by themselves. The low pH values make surfaces exposed to corrosive activity. There has been an important step made forward in this field recently. By that we mean the fact that NEOW can now be obtained in physiological values (6.6 to 6.7). This means that it does not elicit corrosive effect even if concentrated.
Material and Methods
Meat treated
The experiment was conducted in the poultry slaughter-house. After cutting chicken carcasses, we put pieces of meat on trays. Pieces with the skin and without it were treated with 5 per cent NEOW solution, sprinkled with hand sprinkler. After treating the meat, we packed trays with meat on them into controlled atmosphere that is a gas mixture of 70 per cent N2 and 30 per cent CO2. The test controlling sample that was not treated with NEOW was packed exactly in the same way. The packed meat was cooled to 4 ºC, and the cool food chain was guaranteed. Samples were tested daily in the laboratory to determine total microorganism number (ISO 4833:2003), presence of the bacteria Campylobacter spp. (ISO 10272-1:2006) and to perform sensorial evaluation of meat.
Room aerosolization
The area and equipment were cleaned and prepared for disinfection. In the test, we used EOW Steriplant N manufactured by Swiss Steriplant. To carry out cold fogging (Zeljet CS 1 mobile system, Swiss Steriplant AG, Switzerland), compressed air was used that created sufficient flow over spraying nozzle. Air flow over the
POULTRY MEAT DECONTAMINATION WITH ELECTROLYZED OXIDIZING WATER AND CONSUMER HEALTH PROTECTION
25
nozzle was 3300 m3/min. The nozzle model 0/5 of the manufacturer SchlickDüsen (Germany) was used for cold fogging in both cases. To determine the presence of microorganisms on the surfaces, smears were taken from the area of 20 cm2. Fifty smears were taken. Sampling points were placed on the parts of the equipment where slaughter carcasses come in indirect contact with the equipment. Results were expres-sed in CFU/cm2. Glass surfaces with the test microorganisms Staphylo-coccus aureus ATCC 6538 were placed in parallel. The purpose of glass surfaces with the Staphylococcus aureus culture was to find out the effect of EOW without the presence of dirtiness on the surfaces and to assess the importance of cold aerosolization. The glass surfaces were placed in direct vicinity of sampling points from the equipment. The number of microorganisms on the glass surfaces was on average 5-7x106 CFU/cm2. Building 1 (evisceration) has a volume of 1100 m3. The pressure of spraying nozzle was 3.0-3.3 bar. Twelve L of undiluted biocide solution with redox value 870 mV was used for fogging, which was sprayed in 6 minutes. We spent 10 mL/m2 of undiluted biocide. Estimated volume of the room where poultry is slaughtered is 800 m3. In cold fogging, the 0/5 nozzle model was used. The pressure on the spraying nozzle was 3.0-3.3 bar. For the fogging, we used 12 L of 80% biocide solution with redox value 870 mV, which was sprayed in 6 minutes. We spent 15 mL/m2 biocide with 50% concentration.
Results and Discussion
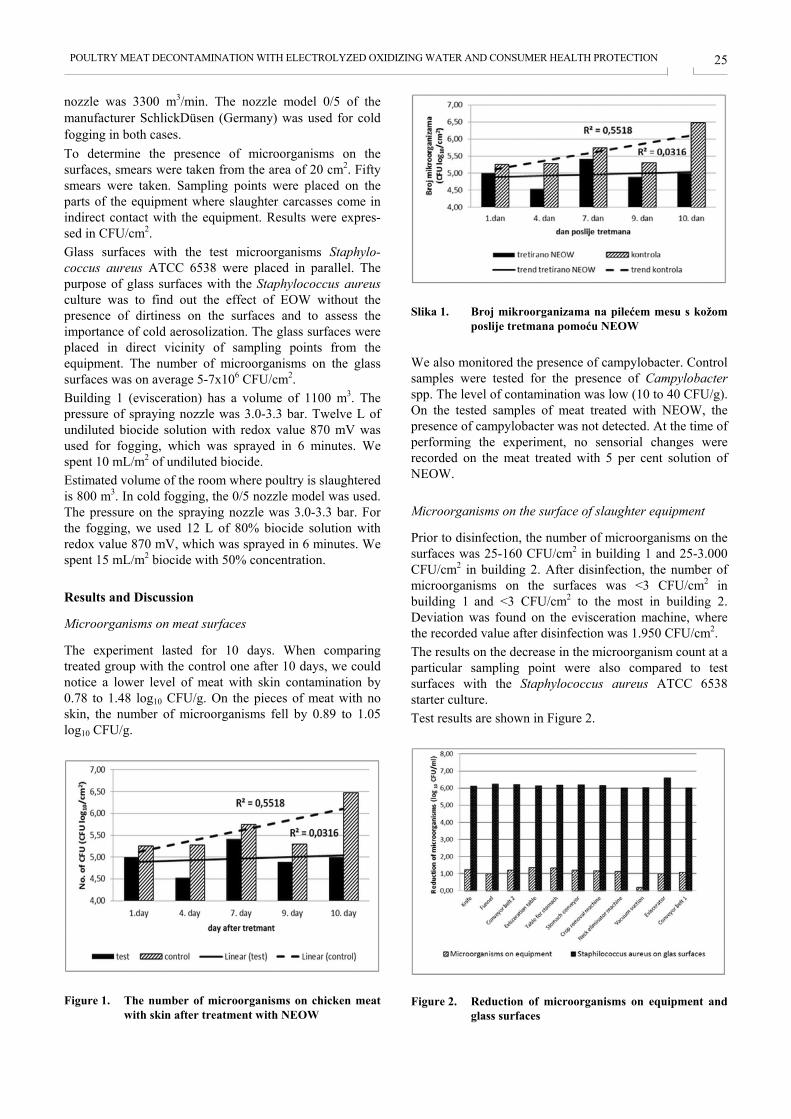
Microorganisms on meat surfaces
The experiment lasted for 10 days. When comparing treated group with the control one after 10 days, we could notice a lower level of meat with skin contamination by 0.78 to 1.48 log10 CFU/g. On the pieces of meat with no skin, the number of microorganisms fell by 0.89 to 1.05 log10 CFU/g.
Figure 1. The number of microorganisms on chicken meat
with skin after treatment with NEOW
Slika 1. Broj mikroorganizama na pilećem mesu s kožom
poslije tretmana pomoću NEOW We also monitored the presence of campylobacter. Control samples were tested for the presence of Campylobacter spp. The level of contamination was low (10 to 40 CFU/g). On the tested samples of meat treated with NEOW, the presence of campylobacter was not detected. At the time of performing the experiment, no sensorial changes were recorded on the meat treated with 5 per cent solution of NEOW.
Microorganisms on the surface of slaughter equipment
Prior to disinfection, the number of microorganisms on the surfaces was 25-160 CFU/cm2 in building 1 and 25-3.000 CFU/cm2 in building 2. After disinfection, the number of microorganisms on the surfaces was <3 CFU/cm2 in building 1 and <3 CFU/cm2 to the most in building 2. Deviation was found on the evisceration machine, where the recorded value after disinfection was 1.950 CFU/cm2. The results on the decrease in the microorganism count at a particular sampling point were also compared to test surfaces with the Staphylococcus aureus ATCC 6538 starter culture. Test results are shown in Figure 2.
Figure 2. Reduction of microorganisms on equipment and
glass surfaces

POULTRY MEAT DECONTAMINATION WITH ELECTROLYZED OXIDIZING WATER AND CONSUMER HEALTH PROTECTION 26
Slika 2. Smanjenje mikroorganizama na klaoničnoj opre-
mi i staklenim nosačima
On the evaluation of results on the surfaces of slaughter equipment, we recorded a relatively low number of micro-organisms, indicating good cleaning routine of the equipment in the slaughterhouse plant. However, it was a factor that made evaluation of the application (NEOW) with cold aerosolization difficult. Considering the guidelines of the standards for biocide testing, where there is requirement for minimal reduction of microorganisms on the surface 3 log10 CFU/cm2, it was in fact impossible to achieve in our conditions, since the highest reduction we could achieve was by 90.0% or by 1 log10 CFU/cm2. Howe-ver, the placed glass surfaces with test microorganisms S. aureus showed reduction 6 log10 CFU/cm2. In building 2, the vacuum suction machine showed deviation in particular, as in this case we achieved reduction by only 35% or 0.19 log10 CFU/cm2. The possible cause of lower reduction is the structure of the vacuum suction machine. Namely, the smear was taken from the inner side of the tube-knife, where NEOW probably did not penetrate in sufficient quantity. Because we found the highest CFU/cm2 value on this spot too, we believe this part was also less well cleaned. Most likely the present uncleanness provided protection for microorganisms and thus prevented the effect of NEOW.
Conclusion
The first results of using NEOW in the process of treating poultry meat for the purpose of diminishing the level of microorganisms are encouraging. It was found that after 10 days, the total amount of microorganisms on the meat with skin was lower by 0.78 to 1.48 log10 CFU/g, while on the meat with no skin it was lower by 0.89 to 1.05 log10 CFU/g (Fig. 1). Results show the possibility of prolonging the shelf life for at least two to three days. After 10 days, the total number of microorganisms in treated meat did not exceed 4x105 CFU/g, while the values in the control group did not exceed 3x106 CFU/g (Fig. 1). An additional proof was the fact that sensorial values of the meat were not changed even on tenth day when the experiment was
finished. According to these findings, we could say that on the one hand, with using NEOW we could extend the durability of fresh meat, while on the other hand, we could certainly acquire a safer durability in eight days, which are now being guaranteed from the producer. Our opinion is that the most critical time is summer time, when we cannot easily provide cool producer to consumer chain. This could help ensure safer food. In the evaluation of the effect of NEOW with cold fogging, we can conclude as follows: 1. The use of NEOW with tested cold fogging shows the applicability in practical conditions. 2. Results of practical testing on glass surfaces show sufficient reduction of CFU, which demonstrates the reliability of the use of NEOW with the method of application of cold fogging. 3. For disinfection of surfaces, small quantities of NEOW are required. 4. It is necessary to evaluate the effect of NEOW in the presence of dirtiness in more detail, which is the subject of further investigation. 5. It is necessary to study the effectiveness of cold aerosolization of NEOW on the slaughter equipment more closely and to study the reliability and the possibility of other ways of applying biocide solution.
Acknowledgment
We are grateful to Panvita and Obisan companies for their help in executing experiments.
References
EFSA (2011): Scientific Report of EFSA: The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2009.
EFSA (2010): The Community Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents, Antimicrobial Resistance and Foodborne Outbreaks in the European Union in 2008. The EFSA Journal: 18.
TAM, C. C., S. J. O'BRIEN, G. K. ADAK (2003): Campylo-bacter coli – an important foodborne pathogen. J Infect. 47: 28.
DICKINS, M. A., S. FRANKLIN, R. STEFANOVA, G. E. SHUTZE, I. EISENACH, V. WESLEY, D. CAVE (2002): Diversity of Campylobacter isolates from retail poultry carcasses and from humans as demonstrated by pulsed-field gel electrophoresis. J Food Prot. 65: 957-962.
FENNER, D. C., B. BURGE, H. P. KAYSER, M. M. WITTEN-BRINK (2006): The anti-microbial activity of electrolysed oxidizing water against microorganisms relevant in veteri-nary medicine. J. Vet. Med., 133-137.
FABRIZIO, K. A., C. N. CUTTER (2003): Stability of electro-lyzed oxidizing water and its efficacy against cell suspens-ions of Salmonella typhimurium and Listeria monocyto-genes. J. Food Protect. 66, 1379-1384.
FABRIZIO, K. A., C. N. CUTTER (2004): Comparison of electrolyzed oxidizing water with other antimicrobial interventions to reduce pathogens on fresh pork. Meat Sci. 68: 463-468.

POULTRY MEAT DECONTAMINATION WITH ELECTROLYZED OXIDIZING WATER AND CONSUMER HEALTH PROTECTION
27
HSU, S., H. KAO (2004): Effects of storage conditions on chemical and physical properties of electrolyzed oxidizing water. J. Food Eng. 65, 465-471.
HSU, S. (2005): Effects of flow rate, temperature and salt concentration on chemical and physical properties of electrolyzed oxidizing water. J. Food Eng. 66, 171-176.
KAMIKADO, H., A. ITO, F. AOKI, M. ENDO, A. OHTSUKA, T. MIYAMOTO, M. IIO (2004): Studies on the effect of sodium hypochlorite and electrolysed oxidizing water on the viability of Bacillus spores and the factors affecting the sporicidal action. [in Japanese]. Sci. Bull. Fac. Agric. Kyushu Univ. 59: 15-24.
LIAO, L. B., W. M. CHEN, X. M. XIAO (2007): The generation and inactivation mechanism of oxidation-reduction potential of electrolyzed oxidizing water. J. Food Eng. 78, 1326-1332.
SHUN-YAO, H. (2005): Effects of flow rate, temperature and salt concentration on chemical and physical properties of electrolyzed oxidizing water. J. Food Eng. 66, 171-176.
WALKER, S. P., A. DEMIRCI, R. E. GRAVES, S. B. SPENCER, R. F. ROBERTS (2005): Response surface modelling for cleaning and disinfecting materials used in milking systems with electrolysed oxidizing water. Int. J. Dairy Tech. 58, 65-73.
ZAŠTITA ZDRAVLJA POTROŠAČA ZBOG UPOTREBE ELEKTROOKSIDIRAJUĆE VODE KOD DEKONTAMINACIJE MESA PERADI
Sažetak
Trajnost mesa ovisi o kontaminaciji različitim mikroorganizmima. Kontaminacija pilećih trupova crijevnim sadržajem tijekom klanja veoma se često događa. Prema literaturnim izvještajima, kod 40%-85% uzoraka pilećeg mesa uzetih u trgovinama može se pronaći kontaminacija mikroorganizmima Campylobacter spp. Neutralna elektrolizirana oksidirajuća voda (NEOW) je nova generacija biocida koji uzimanjem elektrona iz okoline prelazi u stabilno stanje. Zbog toga ne ostavlja ostatke aktivnih tvari u okolini i nije ga potrebno isprati vodom kao mnoge druge dezinficijense. Zbog tih svojstava proučavali smo može li se NEOW rabiti za pranje mesa pilića kontaminiranih bakterijom Campylobacter spp. kako bi se povećala sigurnost hrane za potrošača. Zbog cjelovitog nadzora prisutnosti mikroorganizama u procesu klanja i pripreme pilećeg mesa uvrstili smo i postupak dezinfekcije opreme aerosolizacijom pomoću NEOW. U istraživanju smo ispitivali pileće meso s kožom i bez kože. Slučajno izabrani uzorci podijeljeni su na kontrolnu skupinu i skupinu uzoraka koji su prije pakiranja tretirani 5%-tnom NEOW. Obje skupine uzoraka su zatim pakirane „gastro“ postupkom zatvaranja u kontroliranu atmosferu pomoću mješavine plina (70% N2 i 30% CO2) na temperaturi +4 °C. Uzorke smo oduzimali od prvog do desetog dana skladištenja te analizirani ukupan broj mikroorganizama i prisutnost bakterija Campylobacter jejuni/coli. Usporedno smo izvodili organoleptičnu kontrolu mesa. Prostorije i opremu koja se upotrebljava u postupku klanja izložili smo djelovanju aerosola s NEOW pomoću aerosolizatora Zeljet CS1. Za aplikaciju upotrebljavali smo protok zraka 3300 m3/min i posebnu za aerosolizaciju pripremljenu dizu. U prosjeku smo utvrdili smanjenje ukupnog broja mikroorganizama na dijelovima mesa s kožom. Utvrđeno je smanjenje mikroorganizama za 77.62% do 86.17%. Na površini klaonične opreme u prosjeku smo ustanovili smanjenje ukupnog broja mikroorganizama za 1log10CFU/cm2. Smanjenje je relativno nisko, jer je bilo i malo mikroorganizma prisutnih na površini opreme. Usporedno smo kod aplikacije izložili testne površine s bakterijom Staphylococcus aureus, gdje je zabilježeno smanjenje mikroorganizama za 6log10CFU/cm2. Dobiveni rezultati ukazuju na mogućnost upotrebe NEOW na pilećem mesu za produžavanje trajanja namirnica. Isto tako, koristeći NEOW može se smanjiti prisutnost mikroorganizama Campylobacter jejuni/coli u tijeku radnog procesa. Uz primjenu NEOW u procesu klanja peradi može se ostvariti sigurnost u sprječavanju prijenosa mogućih alimentarnih infekcija i preventivno zaštititi zdravlje potrošača. Biosigurnost obrade mesa peradi može se pojačati upotrebom NEOW kod dezinfekcije prostorija aerosolizacijom. Ključne riječi: neutralna elektrolizirana oksidirajuća voda, dekontaminacija, aerosolizacija, Campylobacter jejuni, Campylobacter coli, perad

PERADARSKI DANI 2013. 28
OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA
Ivan Pušić1, Blanka Beer Ljubić,2 Zlatko Janječić3, Dalibor Bedeković3, Jasna Aladrović2
1Ministarstvo zaštite okoliša i prirode Republike Hrvatske, Zagreb, Hrvatska 2Zavod za fiziologiju i radiobiologiju, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 3Zavod za hranidbu životinja, Agronomski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska
Sažetak Najjednostavniji način očuvanja namirnica životinjskog podrijetla od oksidacijskih promjena i razvoja mikroorganizama je hlađenje ili smrzavanje. Stoga je cilj rada bio utvrditi održivost antioksidativnog sustava mesa prsnog mišićja i bataka pilića tijekom pohrane na +4 °C i -20 °C. Istraživanje je obavljeno na 84 pilića tovne pasmine Cobb 500. Prsa i bataci pilića pohranjeni su u hladnjak i na -20 °C. Uzorkovanje je obavljeno neposredno po klanju i hlađenju na +4 °C i 7. dana u uzorcima pohranjenim u hladnjaku te 30 i 90 dana nakon smrzavanja. U homogenatima tkiva određena je aktivnost superoksid dismutaze (SOD), glutation peroksidaze (GPx), katalaze te koncentracija glutationa (GSH) i lipidskih peroksida (LP). U ohlađenom prsnom mišićju aktivnost SOD i GPx značajno se smanjila pri pohrani u hladnjaku, a SOD i nakon 30 dana smrzavanja u odnosu na vrijednosti neposredno nakon hlađenja. Koncentracija GSH značajno se snizila u uzorcima zamrznutim na -20 °C nakon 90 dana u usporedbi s početnim vrijednostima. Intenzitet lipidske peroksidacije je bio značajno manji nakon sedmodnevne pohrane u hladnjaku. Aktivnost katalaze u prsnoj muskulaturi pilića nije bila podložna značajnim promjenama tijekom pohrane. U mišićima bataka aktivnosti SOD i katalaze značajno su se smanjile tijekom pohrane pilića u hladnjaku u odnosu na početnu razinu. Koncentracija GSH se smanjila 7. dana pohrane na +4 °C, kao i nakon 90 dana pohrane na -20 °C, što je pratio i porast intenziteta lipidske peroksidacije. Uočene su razlike u antioksidativnom sustavu između mišića prsa i bataka. U mišićima bataka pilića zabilježena je značajno viša aktivnost GPx na dan klanja, 7. dana pohrane u hladnjaku, te u uzorcima pohranjenim na -20 °C. Također, u mišićima bataka katalaza je bila značajno viša na početku istraživanja te nakon 30 dana od zamrzavanja. Aktivnost SOD u batacima pilića bila je značajno viša 7. dana pohrane u hladnjaku, kao i nakon 30 dana u zamrznutom stanju. Intenzitet peroksidacije bio je veći u mesu bataka 7. dana pohrane u hladnjaku. Oksidacijska stabilnost i prsnog i bedrenog mišićja pohranom se smanjuje, pri čemu je smanjenje aktivnosti SOD i katalaze kao i koncentracije GSH izraženije u mišićima bataka uz intenziviranje lipidske peroksidacije. Ključne riječi: pilići, mišići prsa, mišići bataka, oksidacijska stabilnost Uvod Kemijski sastav mesa ima značajan utjecaj na konačnu ocjenu kakvoće mesa i mesnih proizvoda. Meso peradi se po kemijskom sastavu razlikuje od mesa sisavaca. Poznato je da prsno mišićje utovljenih pilića ima više bjelančevina (21%-24%) i manje masti (2%-5%) u odnosu na mišičje bataka (18%-20% odnosno 3%-10%) (Živković i sur., 1994.). Pritom višestrukonezasićene masne kiseline čine 14% masti, dok kod sisavaca nalazimo oko 4% višestruko-nezasićenih masnih kiselina. Ove se kiseline tijekom po-hrane mesa u kontaktu s kisikom razlažu na kratkolančane spojeve poput aldehida, ketona, kiselina i alkohola. Na-vedeni se spojevi mogu nakupljati unutar mesa peradi, što može utjecati na njegova senzorna svojstva (Pearson i sur.,
1983.; Higgins i sur., 1999.; Ruíz i sur., 2001.). Uz to što dovode do stvaranja neugodnog mirisa lipidski peroksidi uzrokuju i gubitak okusa, teksture i konzistencije, što sve zajedno dovodi do smanjene prehrambene vrijednosti mesa (Gray i sur., 1996.; Valenzuela i Nieto, 1996.). Također postoje i brojni drugi čimbenici koji utječu na oksidacijske procese u mesu, kao što su izloženost mesa svjetlu, pri-sutnost kisika, temperaturni uvjeti i rast mikroorganizama (Skibsted i sur., 1998.). Stupanj oksidacije masti osim o količini masti (masnokise-linski sastav i količina nezasićenih masnih kiselina) ovisi o koncentraciji/aktivnosti spojeva s antioksidativnim djelo-vanjem, kao i o koncentraciji prooksidansa. Brojne su mo-lekule koje uklanjaju slobodne radikale i/ili oksidativno promijenjene makromolekule.
Dr. sc. Ivan Pušić, dr. med. vet., Ministarstvo zaštite okoliša i prirode, Ulica Republike Austrije 14, 10000 Zagreb, Hrvatska; E-mail: ivan.pusic@mzoip.

OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA
29
Antioksidativna obrana djeluje kao uravnotežen i koordi-niran sustav u kojem svaka molekula ovisi o djelovanju drugih molekula (Halliwell i Gutteridge, 1999.). Najvažniji enzimi su glutation peroksidaza, superoksid dismutaza i katalaza, dok od neenzimskih spojeva svakakao treba navesti glutation, albumine, vitamine E i C. Specifično, superoksid dismutaza će uklanjati superoksidni radikal prevodeći ga u manje štetan proizvod vodikov peroksid koji će se djelovanjem katalaze i glutation peroksidaze razgrađivati. Glutation se smatra najvažnijim unutarsta-ničnim antioksidansom zbog visokih koncentracija u stanici, a može sam uklanjati slobodne radikale pomoću sulfhidrilne skupine ili djelovati kao koenzim glutation peroksidaze (Kidd, 1997.). Antioksidansi se unose hranom i sintetiziraju u organizmu. Tako, primjerice, primjenom antioksidansa kao što je glutation može se usporiti oksi-dacija mioglobina u metmioglobin (Tang i sur., 2003.) i na taj način smanjiti pojava tamne boje mesa. Najjednostavniji način očuvanja namirnica životinjskog podrijetla od oksidacijskih promjena i razvoja mikroorga-nizama je hlađenje ili smrzavanje. Djelovanje antioksi-dativnog sustava odnosno oksidativna stabilnost mesa se razlikuje od stabilnosti žive stanice. Naime, pretvorba skeletne muskulature u meso rezultat je složenih bioke-mijskih reakcija gdje zbog prestanka cirkulacije i anae-robne razgradnje glikogena dolazi do povećanog nastanka mliječne kiseline uz posljedični pad vrijednosti pH koji doprinosi razgradnji bjelančevina i razvoju specifične boje, okusa, mirisa i teksture zrelog mesa. Važnu ulogu tijekom hlađenja i smrzavanja mesa ima katalaza, jer je stabilna (Pradhan i sur., 2000.), no stabilnost ovoga enzima ovisi o vrsti odnosno tipu mišićnih vlakana (Hernandez i sur., 2002.). Također, aktivnost superoksid dismutaze veća je u crvenom (oksidativnom) mišićju (Renerre i sur., 1996.). Osjetljivost masti, naročito višestrukonezasićenih masnih kiselina, na oksidaciju i promjena boje su vjerojatno glavni uzrok smanjenja kakvoće mesa tijekom pohrane. Stoga je cilj ovoga istraživanja bio utvrditi intenzitet oksidativnih promjena u mesu prsnog mišićja i bataka pilića tijekom pohrane u hladnjaku i u dubokosmrznutim uzorcima.
Materijal i metode
Istraživanje je provedeno na 84 pilića hibrida teške pasmine Cobb 500 oba spola. Pilići su tijekom pokusa podno držani, a hrana i voda su bili dostupni ad libitum. Pilići su tijekom tova hranjeni početnom smjesom za piliće prvih 21 dan, a nakon toga smjesom za rast pilića do kraja tova (tablica 1.). Po završetku tova (42 dana) pilići su transportirani na klanje. Trupovi su ohlađeni do temperature od +2 °C u tunelskom sustavu. Hlađenje je trajalo 1,5-2,0 sata, te su pohranjeni u komoru za hlađenje na 0,5 °C. Prsa i bataci pilića pohranjeni su u hladnjak i na -20 °C. Uzorkovanje je obavljeno neposredno po klanju i hlađenju na +4 °C i 7. dana u uzorcima pohranjenim u hladnjaku te 30 i 90 dana
nakon smrzavanja. Uzorci su homogenizirani u 0,14 mol/L KCl (2800 okr/min uz hlađenje tijekom 90 s; Schütt Labortechnik, Njemačka) i centrifugirani (20000xg, 30 min pri 4 °C; Sigma 3 K 15, Sigma, Njemačka). U homogenatima tkiva određena je aktivnost superoksid dismutaze (SOD), glutation peroksidaze (GPx), katalaze te koncentracija glutationa (GSH) i lipidskih peroksida (LP).
Tablica 1. Kemijska analiza početne i smjese za tov pilića
Naziv uzorka Početna smjesa
Smjesa za rast
Vlaga g/kg 113,23 112,49
Sirovi pepeo g/kg 49,75 48,15
Sirove bjelančevine g/kg 241,72 219,51
Sirova mast g/kg 41,47 41,63
Sirova vlakna g/kg 36,27 37,59
Kalcij g/kg 10,42 9,48
Fosfor g/kg 4,81 4,65
Natrij g/kg 1,73 1,82
Kalij g/kg 9,51 9,42
Selen (Na-selenit) mg/kg 0,3 0,3
Škrob g/kg 318,73 352,15
Šećer g/kg 44,48 44,89
Table 1. Diet composition according to chemical analyses
Nutrient content Starter diet Grower diet
Humidity g/kg 113.23 112.49
Ash g/kg 49.75 48.15
Protein g/kg 241.72 219.51
Fat g/kg 41.47 41.63
Raw fibre g/kg 36.27 37.59
Calcium g/kg 10.42 9.48
Phosphorus g/kg 4.81 4.65
Sodium g/kg 1.73 1.82
Potassium g/kg 9.51 9.42
Selenium (Na-selenit) mg/kg 0.30 0.30
Starch g/kg 318.73 352.15
Carbohydrate g/kg 44.48 44.89

OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA 30
Aktivnosti glutation peroksidaze i superoksid dismutaze određene su gotovim kompletima tvrtke Randox (Irska) na automatskom analizatoru SABA 18 (AMS, Italija). Aktiv-nost katalaze određena je metodom po Johanssonu i Borgu (1988.) na uređaju Thermospectronic Helios delta (Uni-cam, Cambridge, Velika Britanija). Koncentracija glutationa u tkivima određena je probom po Beutleru i sur. (1963.), dok je koncentracija lipidskih peroksida određena metodom po Ohkawi i sur. (1979.) na uređaju Thermospectronic Helios delta (Unicam, Cam-bridge, Velika Britanija). Rezultati analiza prikazani su kao srednja vrijednost i standardna devijacija ( x ± SD). Značajnost razlika među rezultatima provjerena je analizom varijance i post hoc Tukey testom ukoliko se radilo o normalnoj razdiobi, odnosno od neparametrijskih testova korištena je Kruskal-Wallisova analiza varijance i test sume rangova primjenom računalnog programa Statistica 9 (StatSoft 2005, SAD). Razlike su statistički značajne ako je p≤0,05.
Rezultati
Aktivnosti SOD i GPx u uzorcima prsne muskulature po-hranjenim na +4 °C značajno su se smanjile tijekom sedmodnevne pohrane (p<0,05) (slike 1. i 2.), a aktivnost SOD je bila značajno niža i nakon 30 dana pohrane na -20 °C (p<0,05) u usporedbi s vrijednostima nakon hlađe-nja. Koncentracija GSH značajno se smanjila u uzorcima
mišića prsa nakon pohrane 90 dana na -20 °C (p<0,05) (sli-ka 1.). Enzim katalaza u uzorcima prsne muskulature tije-kom pohrane nije pokazivao značajne promjene (slika 2.). Na slici 3. prikazan je intenzitet lipidske peroksidacije te je vidljivo da pohranom na +4 °C dolazi do značajnog smanjenja koncentracije LP (p<0,05). Aktivnosti katalaze i koncentracije GSH u mišićima bataka tijekom sedmodnevne pohrane na +4 °C kao i nakon 90 dana na -20 °C značajno su se smanjile (p<0,05) (slike 1. i 2.). Aktivnost SOD značajno se smanjila u bedrenoj muskulaturi samo pri pohrani u hladnjaku (p<0,05). Porast intenziteta lipidske peroksidacije u uzorcima bedrene muskulature uočen je 7. dana u uzorcima pohranjenim u hladnjak kao i nakon 90 dana na -20 °C (p<0,05) (slika 3.). Aktivnost GPx u mišićima bataka bila je ujednačena i stabilna neovisno o temperaturi ili duljini pohrane. Oksidativna stabilnost mišića bataka i prsa značajno se razlikovala tijekom pohrane. Aktivnost GPx bila je značajno veća u mišićima bataka u svim točkama mjerenja (p<0,05) (slika 1.). Također, aktivnost katalaze 0. i 30. dana pohrane, a SOD 7. i 30. dana pohrane bila je značajno veća u mišićima bataka (p<0,05) (slika 2.). Koncentracija GSH nakon 90 dana pohrane na -20 °C bila je značajno veća u mišićima bataka (p<0,05) (slika 1.). Početne vrijednosti LP bile su značajno veće u mišićima prsa, da bi 7. dana pohrane u hladnjaku koncentacija LP u batacima porasla na značajno veće vrijednosti u odnosu na prsno mišićje (p<0,05) (slika 3.).
Slika 1. Koncentracija GSH (lijevo) i aktivnost GPx (desno) u mišićju prsa i bataka pilića pohranjenih na +4 °C i -20 °C;
a,brazličita slova označavaju razlike tijekom pohrane u odnosu na dan 0.; A = označava razlike između tipova mišića
Figure 1. GSH concentration (left panel) and GPx activity (right panel) in breast and thigh muscles stored at +4 °C i -20 °C;
a,bdifferent superscipts refer to significant difference from day 0; A = significant differences between muscle types
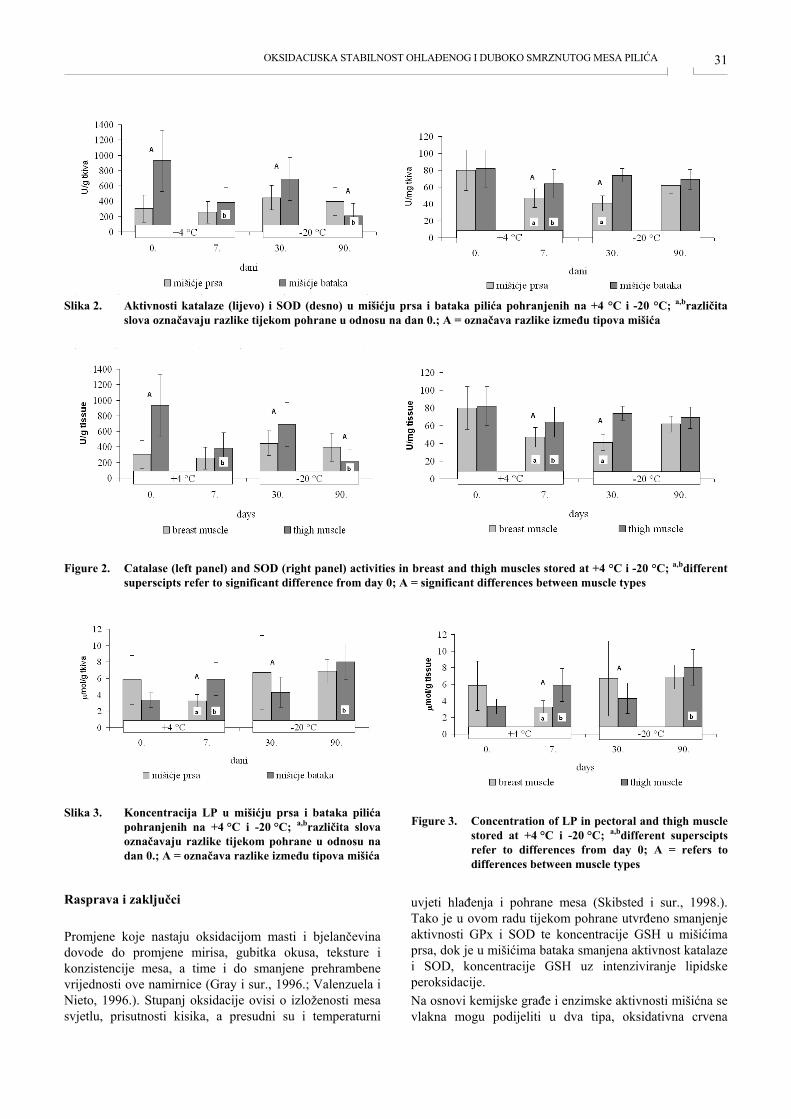
OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA
31
Slika 2. Aktivnosti katalaze (lijevo) i SOD (desno) u mišićju prsa i bataka pilića pohranjenih na +4 °C i -20 °C; a,brazličita
slova označavaju razlike tijekom pohrane u odnosu na dan 0.; A = označava razlike između tipova mišića
Figure 2. Catalase (left panel) and SOD (right panel) activities in breast and thigh muscles stored at +4 °C i -20 °C; a,bdifferent
superscipts refer to significant difference from day 0; A = significant differences between muscle types
Rasprava i zaključci Promjene koje nastaju oksidacijom masti i bjelančevina dovode do promjene mirisa, gubitka okusa, teksture i konzistencije mesa, a time i do smanjene prehrambene vrijednosti ove namirnice (Gray i sur., 1996.; Valenzuela i Nieto, 1996.). Stupanj oksidacije ovisi o izloženosti mesa svjetlu, prisutnosti kisika, a presudni su i temperaturni
uvjeti hlađenja i pohrane mesa (Skibsted i sur., 1998.). Tako je u ovom radu tijekom pohrane utvrđeno smanjenje aktivnosti GPx i SOD te koncentracije GSH u mišićima prsa, dok je u mišićima bataka smanjena aktivnost katalaze i SOD, koncentracije GSH uz intenziviranje lipidske peroksidacije. Na osnovi kemijske građe i enzimske aktivnosti mišićna se vlakna mogu podijeliti u dva tipa, oksidativna crvena
Figure 3. Concentration of LP in pectoral and thigh muscle
stored at +4 °C i -20 °C; a,bdifferent superscipts refer to differences from day 0; A = refers to differences between muscle types
Slika 3. Koncentracija LP u mišićju prsa i bataka pilića
pohranjenih na +4 °C i -20 °C; a,brazličita slovaoznačavaju razlike tijekom pohrane u odnosu nadan 0.; A = označava razlike između tipova mišića

OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA 32
vlakna i glikolitička bijela (Shephard, 1987.). Oksidativna vlakna imaju mnogo mitohondrija, veću koncentraciju mioglobina od glikolitičkih vlakana. Uglavnom koriste masne kiseline kao energetski supstrat te imaju nisku aktivnost ATP-aze i fosforilaze. Za razliku od oksidativnih vlakana, glikolitička vlakna koriste glikogen te imaju veće aktivnosti ATP-aze i fosforilaze. Također, ova se dva tipa vlakana razlikuju po razinama anitioksidativnih spojeva, kao i osjetljivosti na oksidaciju (Daun i Akesson, 2004.). Superoksid dismutaza je važna sastavnica antioksidativne zaštite stanica, jer neposredno uklanja superoksidni radikal prevodeći ga u relativno stabilan vodikov peroksid. Ovaj je enzim najvećim dijelom smješten u citoplazmi kao Zn/Cu-sadržavajući enzim, te manjim dijelom u mitohondrijima kao Mn-sadržavajući enzim, a aktivnosti mu rastu raz-mjerno izloženosti kisiku (Fridovich, 1995.). U ovom radu se aktivnost SOD manjila u obje skupine mišića tijekom pohrane u hladnjaku, a u prsnom mišićju i nakon 30 dana pohrane na -20 °C. Također, zabilježene su veće aktivnosti SOD u mišićima bataka tijekom pohrane (slika 2.). Re-zultati su u suglasju s istraživanjima Renerrea i sur. (1996.) na mišićima goveda. Kako je aktivnost SOD vezana uz broj mitohondrija, za očekivati je u oksidativnom tipu mi-šića veće aktivnosti ovoga enzima. U ovom radu u ohlađenom prsnom mišićju aktivnost GPx značajno se smanjila pri pohrani u hladnjaku, dok je u mišićima bataka bila ujednačena tijekom istraživanja (slika 1.). Također, smanjenje aktivnosti ovoga enzima utvrdili su i Hernandez i sur. (2002.) u m. longissimus dorsi kunića i svinje (Hernandez i sur., 2002.a). Oksidativni tip mišića riba (Watanabee i sur., 1996.), kao i u neki mišići goveda (Renerre i sur., 1996.) pokazuju stabilnu aktivnost tijekom pohrane kao i u ovom istraživanju. Aktivnost GPx u ovom istraživanju bila je značajno veća u mišićima bataka u kojima prevladavaju oksidativna vlakna (slika 1.), što je u skladu s rezultatima Dauna i Akessona (2004.), Renerrea i sur., (1996., 1999.), Pradhana i sur. (2000.), Hernandeza i sur. (2002.a). Kako su oksidacijski procesi naglašeniji u crvenim mišićnim vlaknima, to je veća aktivnost GPx kao kompenzatornog mehanizma za uklanjanje organskih hidroperoksida (Halliwell i Gut-teridge, 1986.). Stanice uklanjaju vodikov peroksid pomoću katalaze i glutation peroksidaze, a koji će od ova dva enzima djelovati ovisi o mjestu u stanici gdje se reakcija odvija (Chan i Decker, 1994.; Halliwell i sur., 1995.). Lokaliza-cija katalaze u mitohondrijima i peroksizomima omogu-ćuje zaštitu, osobito tijekom razdoblja pojačane mobiliza-cije makromolekula, što može stanične strukture učiniti osjetljivijima na oštećenja izazvana slobodnim radikalima (Guderley i sur., 2003.). Enzim katalaza u uzorcima prsne muskulature tijekom po-hrane nije pokazivao značajne promjene, dok se u mišići-ma bataka aktivnost smanjila (slika 2.). Lee i sur. (1996.) su uočili da se aktivnost katalaze ne mijenja u svinjskom mesu nakon 10 tjedana skladištenja na -15 °C, kao i Tere-vinto i sur. (2010.) u mesu nandua (Rhea americana) za-
mrznutom na temperaturi -20 °C tijekom 2 mjeseca. Pradhan i sur. (2000.) su utvrdili stabilnost katalaze i u prsnoj i bedrenoj muskulaturi pilića, kao i u mišićima go-veda i svinja te zaključuju da je zbog svoje stabilnosti tije-kom pohrane vrlo važan dio oksidativne stabilnosti mesa. Jednako kao i enzim GPx i aktivnost katalaze u ovom istraživanju bila je izraženija u mesu bataka, što je u skladu s rezultatima Hernandeza i sur. (2002.) te Pradhana i sur. (2000.). Oksidativni mišići skloniji su lipidskoj oksidaciji od glikolitičkih, što mora pratiti i odgovarajuća veća aktiv-nost antioksidativnih enzima pa tako i katalaze. Najveće promjene koncentracije GSH u ovom istraživanju mogu se uočiti nakon 90 dana pohrane na -20 °C, pri čemu je u mišićima bataka razina GSH veća (slika 1.). Smanjenje koncentracije GSH u zamrznutom mesu pohranjenom šest mjeseci su utvrdili i Nair i Latha (2005.), kao i Gardner i Stewart (1966.). Smanjenje koncentracije GSH ovisi o temperaturi i vremenu skladištenja (Khan i Lentz, 1977.) te pohranom njegova koncentracija postupno opada. Pretvorbom mišićja u meso poslije klanja zbivaju se pro-mjene u mesu koje dovode do poremećaja ravnoteže pro-oksidativnih i antioksidativnih čimbenika, jer se aktiv-nost/koncentracija antioksidativnih spojeva smanjuje, dok su proizvodi oksidacije makromolekula u porastu. Inten-zitet oksidacije kao i vrijednosti proizvoda ovise o duljini i temperaturi pohrane. Tako su u ovom istraživanju početne koncentracije LP veće u mišićima prsa, no tijekom pohrane oksidacija lipida u mišićima bataka je intenzivnija pa stoga raste razina LP i viša je negoli u prsnom mišićju (slika 3.). Suprotno ovome, Min i sur. (2008.) skladištenjem različitih vrsta mesa na temperaturi +4 °C tijekom sedam dana nisu zabilježili promjene koncentracije lipidskih peroksida u mesu. Za očekivati je da tkiva s visokim sadržajem antioksi-dativnih enzima budu stabilnija prema procesima lipidske peroksidacije, ali su prethodna istraživanja pokazala da su oksidativni mišići skloniji lipidskoj oksidaciji od glikoli-tičkih. Razlog ovakvog nalaza leži u činjenici da u struk-turi oksidativnih mišića ima više masti i prooksidativnih oblika željeza (Zenoble i Bowers, 1977.; Kanner i sur., 1988.; Huang i sur., 1993.; Lee i sur., 1996.). Vjerojatno bi se razgradnja oksidativnog mišićja odvijala i brže kada bi aktivnost antioksidativnih enzima bila manja. Rezultati istraživanja govore u prilog tome da oksidativna stabilnost tijekom pohrane ovisi o tipu mišićnih vlakana. Mišići bataka u kojima prevladavaju crvena mišićna vlakna podložniji su oksidaciji pa su veće aktivnosti SOD, GPx kao i katalaze kompenzatorni mehanizam za uklanjanje slobodnih radikala i lipidskih peroksida.
Literatura
BEUTLER, E., O. DURON, B. MIKUS KELLY (1963): Impro-ved method for the determination of blood glutathione. J. Lab. Clin. Med. 61, 882-886.
CHAN, K. M., E. A. DECKER (1994): Endogenous skeletal muscle antioxidants. Crit. Rev. Food Sci. Nutr. 4, 403-426.

OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA
33
DAUN, C., B. AKESSON (2004): Comparison of glutathione peroxidase activity, and total and soluble selenium content in two muscles from chicken, turkey, duck, ostrich and lamb. Food Chem. 2, 295-303.
FRIDOVICH, I. (1995): Superoxide radical and superoxide dismutases. Annu. Rev. Biochem. 64, 97-112.
GARDNER, G. A., D. J. STEWART (1966): Changes in the free amino and other nitrogen compounds in stored beef muscle. J. Sci. Food Agric. 10, 491-496.
GRAY, J. I., E. A. GOMMA, D. J. BUCKLEY (1996): Oxidative quality and shelf life of meats. Meat Sci. 1, 111-123.
GUDERLEY, H., D. LAPOINTE, M. BEDARD, J. D. DUTIL (2003): Metabolic priorities during starvation: enzyme sparing in liver and white muscle of Atlantic cod, Gadus morhua L. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2, 347-356.
HALLIWELL, B., J. M. C. GUTTERIDGE (1999): Free radicals in biology and medicine. 3rd edn. Oxford University Press, UK.
HALLIWELL, B., J. M. GUTTERIDGE (1986): Oxygen free radicals and iron in relation to biology and medicine: some problems and concepts: Arch. Biochem. Biophys. 2, 501-514.
HALLIWELL, B., M. A. MURCIA, S. CHIRICO, O. I. ARUOMA (1995): Free radicals and antioxidants in food and in vivo: what they do and how they work. Crit. Rev. Food Sci. Nutr. 1-2, 7-20.
HERNANDEZ, P., A. LOPEZ, M. MARCO, A. BLASCO (2002): Influence of muscle type, refrigeration storage and genetic line on antioxidant enzyme activity in rabbit meat. World Rabbit Sci. 4, 141-146.
HERNANDEZ, P., D. PARK, K. S. RHEE (2002a): Chloride salt type/ionic strength, muscle site and refrigeration effects on antioxidant enzymes and lipid oxidation in pork. Meat Sci., 4, 405-410.
HIGGINS, F. M., J. P. KERRY, D. J. BUCKLEY, P. A. MORRISSEY (1999): Effects of α-tocopheryl acetate supplementation and salt addition on the oxidative stability (TBARS) and warmed-over flavour (WOF) of cooked turkey meat. Br. Poult. Sci. 1, 59-64.
HUANG, C. H., H. O. HULTIN, S. S. JAFAR (1993): Some aspects of Fe2+ catalysed oxidation of fish sarcoplasmatic reticular lipid. J. Agric. Food Chem. 11, 1886-1892.
JOHANSSON, L. H., L. A. BORG (1988): A spectrophotometric method for determination of catalase activity in small tissues samples. Anal. Biochem. 174, 331-336.
KANNER, J., B. HAZAN, L. DOLL (1988): Catalytic “free” iron ions in muscle foods. J. Agric. Food Chem. 3, 412-415.
KHAN, A. W., C. P. LENTZ (1977): Effects of freezing, thawing and storage on some quality factors for portion-size beef cuts. Meat Sci. 4, 263-270.
KIDD P. M. (1997): Glutathione: systemic protectant against oxidative and free radical damage. Alter. Med. Rev. 3, 155-176.
LEE, S. K., L. MEI, E. A. DECKER (1996): Lipid oxidation in cooked turkey as affected by added antioxidant enzymes. J. Food Sci. 4, 726-728.
MIN, B., K. C. NAM, J. CORDRAY, D. U. AHN (2008): Endogenous factors affecting oxidative stability of beef loin,
pork loin and chicken breast and thigh meats. J. Food Sci. 6, 439-446.
NAIR, M. B., M. S. LATHA (2005): Glutathione antioxidant status of frozen meat samples. J. Environ. Biol. 4, 681-685.
OHKAWA, H., N. OHISHI, K. YAGI (1979): Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 95, 351-358.
PEARSON, A. M., J. I. GRAY, A. M. WOLZAK, N. A. HORESTEIN (1983): Safety implications of oxidized lipids in muscle foods. Food Technol. 37, 121-129.
PRADHAN, A. A., K. S. RHEE, P. HERNÁNDEZ (2000): Stability of catalase and its potential in lipid oxidation in meat. Meat Sci. 54, 385-390.
RENERRE, M., F. DUMONT, P. GATELLIER (1996): Anti-oxidant enzyme activities in beef in relation to oxidation of lipid and myoglobin. Meat Sci. 2, 111-121.
RENERRE, M., K. PONCET, Y. MERCIER, P. GATELLIER, B. MÉTRO (1999): Influence of dietary fat and vitamin E on antioxidant status of muscle of turkey. J. Agric. Food Chem. 1, 237-244.
RUIZ, J. A., L. GUERRERO, J. ARNAU, M. D. GUARDIA, E. ESTEVE-GARCIA (2001): Descriptive sensory analysis of meat from broilers fed diets containing vitamin E or beta-carotene as antioxidants and different supplemental fats. Poult. Sci. 7, 976-982.
SHEPHARD, R. J. (1987): Exercise physiology. B. C. Decker Inc, Toronto, Philadelphia, str. 52.
SKIBSTED, L. H., A. MIKKELSEN, G. BERTELSEN (1998): Lipid-derived off-flavors in meat. In: Flavour of Meat, Meat Products and Seafoods. Shahidi F. (Ed.) Blackie Academic & Professional, London, UK, str. 215-256.
TANG, J., C. FAUSTMAN, S. LEE, T. A. HOAGLAND (2003): Effect of glutathione on oxymyoglobin oxidation. J. Agric. Food Chem. 6, 1691-1695.
TEREVINTO, A., A. RAMOS, G. CASTROMAN, M. C. CABRERA, A. SAADOUN (2010): Oxidative status, in vitro iron-induced lipid oxidation and superoxide dismutase, catalase and glutathione peroxidase activities in rhea meat. Meat Sci. 4, 706-710.
VALENZUELA, A., S. NIETO (1996): Synthetic and natural antioxidants: food quality protectors. Grasas y Aceites 47, 186-196.
WATANABEE, F., M. GOTO, K. ABE, Y. NAKANO (1996): Glutathione peroxidase activity during storage of fish muscle. J. Food Sci. 4, 734-735, 782.
ZENOBLE, O. C., J. A. BOWERS (1977): Copper, zinc and iron content of turkey muscles. J. Food Sci. 5, 1408-1409, 1412.
ŽIVKOVIĆ, J., B. NJARI, L. KOZAČINSKI (1994): Kakvoća i higijenska ispravnost mesa u funkciji unapređivanja pera-darstva. Savjetovanje Peradarski dani. Trakošćan, str. 58-67.
Zahvala
Istraživanja je financiralo Ministarstvo znanosti, obrazo-vanja i športa RH, projekt br. 053-0531854-1866 i 053- -0531854-1853.

OKSIDACIJSKA STABILNOST OHLAĐENOG I DUBOKO SMRZNUTOG MESA PILIĆA 34
OXIDATIVE STABILITY OF COOLED AND FROZEN CHICKEN MEAT
Summary
The most effective way of protecting foodstuffs of animal origin from oxidation and microbial growth is by cooling or freezing. The aim of this study was to determine the stability of antioxidative status of chicken breasts and thigh meat during storage at +4 °C and -20 °C. The research was conducted with breast and thigh meat from 84 broilers of Cobb 500 breed. Sampling took place immediately after the slaughter and cooling down and 7 days after refrigerated storage at +4 °C in the first group. The sampling of frozen meat in the second group took place 30 and 90 days after freezing. Glutathione peroxidase (GSPx), superoxide dismutase (SOD) and catalase activites, as well as glutathione (GSH) and lipid peroxide (LP) concentrations were determined in tissue homogenates. SOD and GPx activities decreased in cooled breast meat after refrigerated storage. SOD acitvity also decreased in frozen breast meat samples 30 days after freezing. GSH concentration in breast meat significantly decreased 90 days after freezing. Lipid peroxidation was significantly lower after 7 days of refrigerated storage (+4 °C). Catalase activity was not altered in breast meat upon using different storage procedures. SOD and catalase activities in thigh meat decreased significantly after refrigerated storage. GSH concentration decreased significantly after 7 days of refrigerated storage as well as after 90 days of storage in the freezer, accompanied by elevated lipid peroxidation. We found signifiant differences in the antioxidant status between breast and thigh meat. Thigh meat had higher GPx acitvity on the day of slaughter, 7 days after refrigerated storage and after storage in the freezer. Thigh muscles had higher catalase activity in the beginning of the study as well as 30 days after freezing. SOD activity in thigh muscles was significantly higher after 7 days of refrigerated storage and 30 days after freezing. Peroxidation intensity was more pronounced in thigh than breast muscles after 7 days of refrigerated storage. Oxidative stability of both breast and thigh muscles is diminished through storage, an effect more pronounced in thigh muscles through decreased SOD and catalase activity, GSH concentration and more intensive lipid peroxidation. Key words: chicken, chicken breast meat, chicken thigh meat, oxidative stability

PERADARSKI DANI 2013.
35
CHLAMYDIA PSITTACI ASSOCIATED WITH POX VIRUS INFECTION IN LAYING HEN FLOCK
Danijela Horvatek Tomić1, Željko Gottstein1, Vladimir Savić2, Marina Tišljar2, Maja Lukač1, Estella Prukner-Radovčić1
1Department of Poultry Diseases with Clinic, Faculty of Veterinary Medicine, University of Zagreb, Zagreb, Croatia 2Poultry Center, Croatian Veterinary Institute, Zagreb, Croatia Summary In the autumn 2010, we investigated an outbreak of pox infection complicated with Chlamydia psittaci (C. psittaci) in a flock of free-ranged laying hens. Altogether 300 birds, kept as a backyard flock, started to show clinical signs of upper respiratory tract infection and severe purulent conjunctivitis was noticed in most of the birds. Mortality rate amounted up to 10% daily. Three laying hens were sacrificed and the samples were submitted to laboratory testing. Aerobic bacteriologic examination revealed the presence of Bacillus sp. and Staphylococcus sp. in conjunctival, choanal and tracheal swabs. The conjunctival and pharyngeal swabs of two sacrificed laying hens were examined by Chlamydiaceae-specific real-time PCR and C. psittaci-specific real-time PCR, and reacted positive to both methods. Samples were also examined by inoculation in chicken embryos and the presence of poxvirus in the chorioallantoic membrane was confirmed using specific PCR. The entire flock was treated with enrofloxacin (Enroxil 10%, Krka, Slovenija) via drinking water for 14 days and the symptoms resolved. The presence of poxvirus probably contributed to the severity of clinical signs caused by C. psittaci and enhanced the infection. Nevertheless, the pathogenicity of this C. psittaci strain remains indistinct, and we can only speculate that it could be higher than of other strains infecting poultry. The infection route remained unclear, but due to the free-range production, it could be possible that wild birds served as a source of infection or the pathogens persisted in the environment. The finding of C. psittaci in laying hens is of enormous importance from two standpoints: 1) significance for public health as a dangerous zoonosis, and 2) presence of C. psittaci in chickens, which are supposed to be a less prone species to this infection. Key words: Chlamydia psittaci, pox virus, laying hens Introduction Infection of birds with the obligate intracellular bacterium Chlamydia (formerly Chlamydophila) psittaci represents a significant problem in veterinary and human medicine all over the world. Avian chlamydiosis can be an inapparent subclinical infection or acute, subacute or chronic disease characterized by respiratory, digestive or systemic infection. Clinical signs vary greatly in severity and depend on the species, age of the bird, and the strain of Chlamydia involved (Vanrompay et al., 1994; Van Lock et al., 2005). Increased shedding of Chlamydia may be triggered by stress factors such as other concurrent infections or infestations, lack of food, breeding, and overcrowding. Infection in humans is also regularly reported, especially in those closely connected to infected birds (Anderson and Vanrompay, 2003; Harkinezhad et al., 2009). The illness in people is
usually respiratory and characterized by influenza-like symptoms, can lead to severe pneumonia and non-respiratory health problems, and death can result if the disease is left untreated (Crosse, 1990.). Bird breeders and persons in specific occupations (employees in poultry slaughterhouses, veterinarians, laboratory workers, zoo and pet shop workers) are more often exposed (Gosbell et al., 1999; Laroucau et al., 2009; Magnino et al., 2009). Chlamydiosis in laying hens has been previously reported (Baar et al., 1986; Malkinson et al., 1987; Arzey and Arzey, 1990), but mostly as clinically inapparent infection. Epi-demiological and laboratory examination has revealed that chickens are relatively resistant to chlamydiosis, especially older birds (Anderson and Vanrompay, 2003). On the other hand, investigation of slaughtered chickens showed that up to 85% of chicken flocks (11 of 13) tested positive to C. psittaci by PCR and culture (Dickx et al., 2010).
Dr. Danijela Horvatek Tomić, DVM; Department of Poultry Diseases with Clinic, Faculty of Veterinary Medicine, University of Zagreb, Heinzelova 55, 10000 Zagreb; Tel: 0038512390281; Fax: 0038512390135; e-mail: [email protected]

CHLAMYDIA PSITTACI ASSOCIATED WITH POX VIRUS INFECTION IN LAYING HEN FLOCK 36
Poxvirus infection is common in commercial poultry and other birds. In this regard, about 232 species in 23 orders of birds have been reported to acquire a natural poxvirus infection (Bolte et al., 1999). All avian poxviruses isolated thus far are members of the genus Avipoxvirus of the family Poxviridae (Tripathy and Reed, 2008). Avipox infection generally manifests as either cutaneous or diphtheritic form, occurring in some cases in the same bird. The most common cutaneous form develops as discrete nodular proliferative skin lesions in the non-feathered parts of the body, scab formation, and desquamation of the degenerating epi-thelium, whereas the diphtheritic form involves proliferative lesions in the mucous membrane of the mouth, pharynx, esophagus, and trachea (Tripathy and Reed, 2008). Concurrent infections with other microorganisms occur, posing difficulty in both clinical and laboratory diagnosis (Tadese et al., 2007).
Case Description
History of laying hens illness
Altogether 300 birds, kept as free-range laying hens in semi-open enclosure, started to show clinical signs of upper respiratory tract. The birds were depressed and lethargic with respiratory signs, severe purulent conjunctivitis, uni-lateral swelling of the sinuses, dyspnea, coughing, and nasal discharge. Mortality rate amounted up to 10% daily. Three laying hens were sacrificed and the samples were submitted to laboratory testing. To prevent further spread of the disease and economic losses, the flock was treated with the antimicrobial drug enrofloxacin (Enroxil 10%, Krka, Slovenija) via drinking water for 14 days, and the symptoms resolved.
Gross lesions and microbiological examination
Post mortem examination revealed an extremely high degree of cachexia in all examined hens. In one of them, the skin of the face was full of confluent focal crusts followed by the fibrin-like focal lesions widely scattered on the oral mucous membrane, and mucopurulent laryngotracheitis as well. For bacteriological examination, the conjunctival, choanal and tracheal swabs taken from sacrificed hens were plated directly on a (i) neutral agar (Difco Nutrient Agar, Becton, Dickinson and Company, Sparks, USA) and (ii) brilliant green agar (brilliant green agar-modified, Oxoid Ltd., Basingstoke, Hampshire, England) and plates were incu-bated aerobically at 37 oC for 24h. Taxonomic classification of isolated bacteria was based on colony morphology, microscopic examination and biochemical characteristics. Sabouraud dextrose agar plates (SBA; Becton Dickinson, USA) were incubated at 24 °C for 5 to 10 days for iden-tification of fungi. Fungi were identified macroscopically based on colony morphology and microscopically with lactophenol staining. Samples were also examined by inoculation into chorio-allantoic membrane of 11-day-old chicken embryos, which
were incubated for the following 5 days and the presence of fowlpox virus in the membrane was confirmed using specific PCR (Tadese et al., 2007). Because bacteriological examination did not reveal any presumptive microorganism, additional conjunctival and pharyngeal samples were taken to detect the possible presence of C. psittaci. DNA was extracted by using GenElute Mammalian Genomic DNA Miniprep Kit (Sigma-Aldrich, USA), according to the manufacturer’s instructions. Extracted DNA was examined by specific Chlamydiaceae real-time PCR, as described by Ehricht et al. (2006), targeting the 23S rRNA gene, and C. psittaci-specific real-time PCR (targeting the ompA gene), as described by Pantchev et al. (2008). PCR-amplified segments of ompA gene were sequenced by Macrogen Inc. (Korea). Nucleotide sequence has been deposited under GenBank accession numbers KC618330 for partial ompAgene sequence.
Results
The aerobic bacteriological examination revealed the pre-sence of Bacillus sp. and Staphylococcus sp. in conjunctival, choanal and tracheal swabs. Some yeasts were also found in these samples. Virologic examination in chicken embryos resulted in the formation of focal proliferative pock lesions. Fowlpox virus specific PCR gave positive result for DNA extracted from the pock lesions. The conjunctival and pharyngeal swabs of altogether 2 laying hens examined by Chlamydiaceae-specific real-time PCR and by C. psittaci-specific real-time PCR reacted positive to both methods.
Discussion
In the past, infections in poultry have been described in association with outbreaks of avian pox, since this virus can act as a predisposing factor that allows the presence of severe form of bacterial infection (Devriese et al., 1992; Van Loock et al., 2006). Findings of poxvirus in these laying hens contributed probably to the severity of clinical signs and enhanced the chlamydial infection in this case, which led to the relatively unusually high percentage of mortality. Nevertheless, the pathogenicity of this C. psittaci strain remains indistinct and we can only speculate that it could be higher than of other strains infecting poultry. Transmission of C. psittaci from birds to humans is reported regularly, particularly in high-risk individuals such as veterinarians, bird breeders or fancier. In our case, there was no clinical symptoms observed in farm workers or vete-rinarians, but on the other hand, testing was not conducted. Avian chlamydiosis has not yet been thoroughly investi-gated, probably due to the very complex diagnostic pro-cedure. Isolation and identification is the gold standard for most infectious diseases, but these techniques may be difficult to use for chlamydioses. Molecular methods are specific, sensitive and allow for rapid detection without the

CHLAMYDIA PSITTACI ASSOCIATED WITH POX VIRUS INFECTION IN LAYING HEN FLOCK
37
need of culturing of Chlamydia (Borel et al., 2008; Sachse et al., 2009). The infection routes remained unclear, but due to the free-range production at the beginning of the production cycle, it could be possible that wild birds could serve as a source of infection or the bacteria persisted in the environment. The finding of C. psittaci in laying hens is of enormous importance from two standpoints: 1) significance for public health as a dangerous zoonosis, and 2) presence of C. psittaci in chickens, which are supposed to be a less prone species to this infection.
References
ANDERSON, A. A., D. VANROMPAY (2003): Avian chlamy-diosis (psittacosis, ornithosis). In: Diseases of Poultry, 11th edn. (Y. M. Saif, Ed.), Iowa State Press, Iowa, USA, 863-879.
ARZEY, G. G., K. E. ARZEY (1990): Chlamydiosis in layer chickens. Austral. Vet. J. 67, 461.
BARR, D. A., P. C. SCOTT, M. D. O’ROURKE, R. J. COULTER (1986): Isolation of Chlamydia psittaci from commercial broiler chickens. Austral. Vet. J. 63, 377-378.
BOLTE, A. L., J. MEURER, E. F. KALETA (1999): Avian host spectrum of avipox viruses. Avian Pathol. 28, 415-432.
BOREL N., E. KEMPF, H. HOTZEL, E. SCHUBERT, P. TORGERSON, P. SLICKERS, R. EHRICHT, T. TASARA, A. POSPISCHIL, K. SACHSE (2008): Direct identification of chlamydiae from clinical samples using a DNA microarray assay: a validation study. Mol. Cell. Probes 22, 55-64.
DICKX, V., T. GEENS, T. DESCHUYFFELEER, L. TYBERG-HIEN, T. HARKINEZHAD, D. S. A. BEECKMAN, L. BRAECKMAN, D. VANROMPAY (2010): Chlamydophila psittaci zoonotic risk assessment in a chicken and turkey slaughterhouse. J. Clin. Microbiol. 48, 3244-3250.
CROSSE, B. A. (1990): Psittacosis: a clinical review, J. Infect. 21, 251-259.
EHRICHT, R., P. SLICKERS, S. GOELLNER, H. HOTZEL, K. SACHSE (2006): Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes 20, 60-63.
DeVRIESE, L. A., E. UYTTEBROEK, P. DOM, P. DEHERDT, R. DUCATELLEF, F. HAESEBROUCK (1992): Staphylococcus hyicus associated with pox in chickens and turkeys. Avian Pathol. 21, 529-533.
GOSBELL, I. B., A. D. ROSS, I. B. TURNER (1999): Chlamydia psittaci infection and reinfection in a veterinarian. Aust. Vet. J. 77, 511-513.
HARKINEZHAD, T., K. VERMINNEN, M. DE BUYZERE, E. RIETZSCHEL, S. BEKAERT, D. VANROMPAY (2009):
Prevalence of Chlamydophila psittaci infections in a human population in contact with domestic and companion birds. J. Med. Microbiol. 58, 1207-1212.
LAROUCAU, K., B. De BARBEYRAC, F. VORIMORE, M. CLERC, C. BERTIN, T. HARKINEZHAD, K. VERMIN-NEN, F. OBENICHE, I. CAPEK, C. BEBEAR, B. DURAND, G. ZANELLA, D. VANROMPAY, B. GARIN-BASTUJI, K. SACHSE (2009): Chlamydial infections in duck farms associated with human cases of psittacosis in France. Vet. Microbiol.135, 82-89.
MAGNINO, S., D. HAAG-WACKERNAGEL, I. GREIGEN-FEIND, S. HELMECKE, A. DOVČ, E. PRUKNER-RADOVČIĆ, E. REŠIDBEGOVIĆ, V. ILESKI, K. LAROUCAU, M. DONATI, S. MARTINOV, E. F. KALE-TA (2009): Chlamydial infections in feral pigeons in Europe: review of data and focus on public health implications. Vet. Microbiol.135, 54-67.
MALKINSON, M., S. MACHANY, A. ARONOVIC, K. DAVIDIV, Y. WEISMANN (1987): Mixed infection with Chlamydia psittaci, fowlpox virus and Haemophilus gallinarum in broiler breeder chicks. Vet. Rec. 120, 461-462.
PANTCHEV, A., R. STING, R. BAUERFEIND, J. TYCZKA, K. SACHSE (2008): New real-time PCR tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples. Vet. J.181, 145-150.
SACHSE, K., E. VRETOU, M. LIVINGSTONE, N. BOREL, A. POSPISCHIL, D. LONGBOTTOM (2009): Recent developments in the laboratory diagnosis of chlamydial infections. Vet. Microbiol.135, 2-21.
TADESE, T., A. E. POTTER, S. FITZGERALD, W. M. REED (2007): Concurrent infection in chickens with fowlpox virus and infectious laryngotracheitis virus as detected by immunohistochemistry and a multiplex polymerase chain reaction technique. Avian Dis. 51, 719-724.
TRIPATHY, D. N., W. M. REED (2008): Pox. In: Diseases of Poultry 11th edn. (Y. M. Saif, Ed.), Iowa State Press, Iowa, Ames, USA, 291-307.
Van LOOCK, M., T. GEENS, L. De SMIT, H. NAUWYNCK, P. Van EMPEL, C. NYLOR, H. M. HAFEZ, B. M. GOODEERIS, D. VANROMPAY (2005): Key role of Chlamydia psittaci on Belgian turkey farms in association with other respiratory pathogens. Vet. Microbiol. 107, 91-101.
Van LOOCK, M., K. LOOTS, S. Van de ZANDE, M. Van HEERDEN, H. NAUWYNCK, B. M. GODDEERIS, D. VANROMPAY (2006): Pathogenic interactions between Chlamydophila psittaci and avian pneumovirus infections in turkeys. Vet. Microbiol. 112, 53-63.
VANROMPAY, D., R. DUCATELLE, F. HAESEBROUCK (1994): Pathogenicity for turkeys of Chlamydia psittaci strains belonging to the avian serovars A, B and D. Avian Pathol. 23, 247-262.
CHLAMYDIA PSITTACI POVEZANA S INFEKCIJOM POX VIRUSOM U JATU KOKOŠI NESILICA
Sažetak
Ujesen 2010. godine istraživana je miješana infekcija virusom boginja i bakterijom Chlamydia psittaci (C. psittaci) u jatu kokoši nesilica držanih u slobodnom uzgoju. Jato od ukupno 300 kokoši počelo je pokazivati znakove infekcije gornjih dišnih puteva, uz teški gnojni konjunktivitis u većine ptica. Stopa smrtnosti iznosila je do 10% dnevno. Tri kokoši nesilice su žrtvovane, a uzorci su dostavljeni za laboratorijska ispitivanja. Aerobnom bakteriološkom pretragom dokazana je prisutnost bakterija Bacillus sp. i Staphylococcus sp. u obriscima konjunktiva, nosa i dušnika. Obrisci konjunktiva i ždrijela

CHLAMYDIA PSITTACI ASSOCIATED WITH POX VIRUS INFECTION IN LAYING HEN FLOCK 38
od ukupno dviju kokoši nesilice pregledani su i specifičnom reakcijom polimeraze u stvarnom vremenu (real-time PCR) za dokaz bakterija porodice Chlamydiaceae i za dokaz bakterije C. psittaci, te su obje reakcije bile pozitivne. Uzorci su također pregledani i virusološkom pretragom u kokošjim zamecima, te je prisutnost virusa boginja potvrđena i uporabom specifične PCR. Cijelo jato liječeno je enrofloksacinom (Enroxil 10%, Krka, Slovenija) putem vode za piće u trajanju od 14 dana, a simptomi su nestali nakon provedene terapije. Prisutnost virusa boginja vjerojatno je doprinijela težini kliničkih simptoma uzrokovanih bakterijom C. psittaci. Ipak, patogenost ovoga soja C. psittaci ostaje nejasna, kao i izvor same infekcije, no s obzirom na slobodan način uzgoja kokoši nesilica moguće je da su divlje ptice služile kao izvor infekcije ili je patogen bio trajno prisutan u okolišu životinja. Nalaz C. psittaci u nesilica od ogromnog je značenja iz dva aspekta: 1) značenja za javno zdravstvo kao opasne zoonoze i 2) općenito prisutnost C. psittaci u peradi, koja bi trebala biti manje sklona ovoj vrsti infekcije. Ključne riječi: Chlamydia psittaci, virus boginja, kokoši nesilice

PERADARSKI DANI 2013.
39
OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT
Irena Zdovc1, Majda Golob1, Tina Pirš1, Jerneja Ambrožič-Avguštin2
1University of Ljubljana, Veterinary Faculty, Institute for Microbiology and Parasitology, Ljubljana, Slovenia 2University of Ljubljana, Biotechnical Faculty, Department of Biology, Ljubljana, Slovenia
Summary The aim of our study was to investigate the prevalence of extended-spectrum β-lactamases (ESBL) and AmpC-producing Escherichia coli strains in retail chicken meat. Furthermore, associated resistance to other antimicrobials among ESBL and AmpC positive strains was also evaluated. Raw meat samples were obtained from Slovenian surveillance program on antibiotic resistance in bacteria isolated in food-producing animals in 2011 and 2012. A total of 446 broiler meat samples were examined by using non-selective and selective growth media. Isolates were phenotypically confirmed as β-lactamase producers by disk diffusion and broth microdilution method using indicator antimicrobials. Samples were heavily contaminated with E. coli producing ESBL (46.4%) and E. coli producing AmpC β-lactamases (36.9%). Among them, a high rate of isolates with associated resistance to other antimicrobials was detected. The most frequently associated resistances found in ESBL/AmpC-producing organisms were against fluoroquinolones, tetracyclines, streptomycin, trimethoprim/sulfamethoxazole and chloramphenicol. Key words: antimicrobial resistance, susceptibility testing, Escherichia coli, extended-spectrum beta-lactamases (ESBL), AmpC, poultry Introduction Escherichia coli (E. coli) normally colonizes the gastro-intestinal tract of human and warm-blooded animals and includes a variety of nonpathogenic, commensal strains, which belong to the normal gut flora. These commensal strains rarely cause diseases in contrast to pathogenic strains, which cause various types of intestinal and extra-intestinal infections in humans and animals (Kohler and Dobrindt, 2011; Kaper et al., 2004). Special types of isolates, so called avian pathogenic E. coli (APEC), are the primary causative agent of cellulitis, septicemia, and airsacculitis in poultry and they are very important bacterial pathogens in broiler chickens (Nakazato et al., 2009). Massive and long-term therapeutic use of antimicrobials may lead to selection of drug resistant strains in both commensal and pathogenic strains of E. coli. Animals are considered as potential reservoirs of multidrug-resistant (MDR) gram-negative organisms (Seiffert et al., 2013).
Bacteria acquire most resistance genes through horizontal transfer, mostly by plasmids and transposons, which are common vectors for the dissemination of antimicrobial resistance genes to other bacteria (Smith et al., 2007). During the last decade, drug resistance in Entero-bacteriaceae, obtained from human and animal sources, has increased significantly worldwide (Bailey et al., 2010; Sharma et al., 2008). The most important resistance mechanisms that reduce the efficacy of most expanded-spectrum cephalosporins and monobactams is based on the production of enzymes that inactivate the drugs by hydrolyzing their beta-lactam ring (Geser et al., 2012). Escherichia coli producing extended-spectrum beta-lactamases (ESBL) or transferrable AmpC-type beta-lactamases is a rapidly emerging public health problem. The impact of food producing animals as a possible source of resistant bacteria and their potential zoonotic trans-mission, especially along the food chain, has emerged as a major concern in human healthcare (Golob et al., 2012; Johnson et al., 2007; Leverstein-Van Hall et al., 2009; Overdevest et al., 2011). Retail meat might be an
Assist. Prof. Irena Zdovc, DVM, PhD, University of Ljubljana, Veterinary Faculty, Institute for Microbiology and Parasitology, Gerbičeva 60, Ljubljana 1000 Slovenia; tel.: 00386 (0)1 4779 158; Fax: 00386 (0)1 4779 352; e-mail: [email protected]

OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT 40
important vehicle for the community-wide dissemination of multi-resistant E. coli. (Cortes et al., 2010; Gregova et al., 2012; Kolar et al., 2010). Associated resistance to other antimicrobials such as tetracycline, fluoroquinolones, aminoglycosides and trimethoprim/sulfamethoxazole is also known (Kolar et al., 2010). Quinolone resistance is usually mediated by chromosomal mutations in the quinolone-resistant determining region (QRDR) that en-codes DNA gyrase genes. Aminoglycoside resistance is generally due to enzymatic inactivation. More recently, the appearance of carbapenem-resistant isolates in livestock has drawn even greater concerns (Seiffertet al., 2013).
Material and Methods
Sampling
The prevalence of ESBL and/or AmpC producing E. coli was estimated in retail broiler meat samples collected randomly in slaughterhouses in all regions of Slovenia. Isolation of bacteria was performed in two steps, using non-selective and selective growth media. About 25 g of each sample was enriched in 25 mL of buffered peptone water for 16-20 h at 37 °C. One loop (10 µ) of the liquid culture was streaked onto Chrom ID, ESBL agar (BioMerieux, France) containing a mixture of antibiotics, including cefpodoxime. Five typical red colonies from Chrom ID agar (Fig. 1) were subcultured on blood agar and chromogenic Uriselect agar for species confirmation.
Antibiotic susceptibility testing
The colonies that corresponded to E. coli were pheno-typically tested for their susceptibility to 18 antimicrobial agents and their combinations by the disk diffusion method (DD). All testing procedures and evaluation of the results were performed according to the Clinical and Laboratory Standards Institute (CLSI) recommendations. The anti-biotics tested were ampicillin (AM), amoxicillin/clavulanic acid (AMC), cephalothin (CF), cefpodoxime (CPD), cefotaxime (CTX), cefotaxime/clavulanic acid (CTX/C), ceftazidime (CAZ), ceftazidime/clavulanic acid (CAZ/C), cefoxitin (FOX), chloramphenicol (C), ciprofloxacin (CIP), enrofloxacin (ENR), nalidixic acid (NA), gentamicin (GM), streptomycin (S), trimethoprim-sulfamethoxazole (SXT), tetracycline (TE) and imipenem (IMP).
Phenotypic confirmation
Initial confirmatory test for ESBL was performed by the combination disk method. It compares the zones of cephalosporin disks to the ones of the same cephalosporin with clavulanic acid. The isolates, which showed at least 5-mm difference between CTX and CTX/C and/or CAZ and CAZ/C, were highly suspicious of ESBL. All isolates resistant to cefoxitin were suspicious of AmpC producing E. coli. Both types of β-lactamase producing isolates were additionally tested by the broth microdilution method
Figure 1. Types of Escherichia coli strains regarding their pathogenicity for humans and animals

OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT
41
(MIC) for phenotypic ESBL and AmpC confirmation (Fig. 2), according to the manufacturer’s instructions (Trek Diagnostic System, Sensititre).
Genotypic confirmation
Isolates from 2011 were submitted to genotype deter-mination using CTX and AmpC group specific primers (Fig. 3). The results were obtained by the multiplex PCR method, as described previously (Ambrožič-Avguštin et al., 2012). Different steps of diagnostic procedures are shown in Figures 2-4.
Results Retail poultry meat samples were heavily contaminated with E. coli producing broad-spectrum β-lactamases. Nearly half of the chicken meat samples (46.4%) contained ESBL producing E. coli (36.9%) and AmpC beta lactamases producing E. coli (see Table 1). Phenotypic confirmation using the combination disk method showed a very high degree of positive results using cefotaxime (CTX):cefotaxime/clavulanic acid (CTX/C) combination (92.9%), which indicated the presence of CTX-M resistance genes. The majority of isolates were additionally resistant to many other groups of antibiotics, especially to fluoro-quinolones. Associated resistance to other antimicrobials was tested in 140 ESBL positive and 86 AmpC positive strains. Both types of isolates were most frequently resistant to fluoroquinolones (80% of ESBL and 80.2% of AmpC isolates), tetracyclines (78.6% of ESBL and 48.8%
of AmpC isolates), and streptomycin (27.9% of ESBL and 43% of AmpC isolates). Resistance to trimethoprim/sul-famethoxazole was relatively high in ESBL (23.6%) and lower in AmpC (7%) isolates. Similar results were obtained for chloramphenicol, with higher resistance in ESBL (22.1%) and lower in AmpC (11%) isolates. However, opposite results were found for gentamicin, where high resistance was observed in 20.9% of AmpC % and only 2.1% of ESBL isolates. All isolates (100%) were susceptible to imipenem, which indicated the absence of carbapenemases. Results are shown in Table 2. Molecular typing of 61 isolates from 2011 revealed the presence of 43 isolates with bla genes belonging to CTX-M-1 group and 18 isolates with genes encoding AmpCβ -lactamases.
Table 1. Occurrence of ESBL and AmpC producing E. coli in poultry meat in Slovenia
No. of samples ESBL % AmpC % Negative %
2011 (n=80) 43 53.8 18 22.5 19 23.8
2012 (n=366 ) 164 44.8 135 36.9 67 18.3
Total (N=446) 207 46.4 153 34.3 86 19.3
Figure 2. Typical red colonies suspicious of ESBL/AmpC on
ChromID ESBL plates after 24-48 h/37 °C
Figure 3. Suspicious isolates were phenotypically confirmed
by using microdilution method
Figure 4. Molecular confirmation of resistance genes

OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT 42
Table 2. Associated resistance to other antimicrobials among ESBL and AmpC positive isolates
ESBL strains (N=140) AmpC strains (N=86) Antimicrobial tested
n R (%) n R (%)
Ampicillin (AM) 140 100 86 100
Amoxicillin/clavulanic acid (AMC) 0 0 86 100
Cephalothin (CF) 140 100 86 100
Cefpodoxime (CPD, indicator for ESBL 140 100 86 100
Cefoxitin (FOX), indicator for AmpC 0 0 86 100
Chloramphenicol (C) 31 22.1 11 12.8
Ciprofloxacin (CIP) 112 80 69 80.2
Enrofloxacin (ENR) 112 80 69 80.2
Nalidixic acid (NA) 112 80 69 80.2
Gentamicin (GM) 3 2.1 18 20.9
Streptomycin (S) 39 27.9 37 43
Trimethoprim/sulfamethoxazole (SXT) 33 23.6 6 7
Tetracycline (TE) 110 78.6 42 48.8
Imipenem (IMP) 0 0 0 0 R (%) = percentage of resistant isolates Discussion A high proportion of ESBL/AmpC producing E. coli strains in retail chicken meat samples in Slovenia was expected due to several previous reports from other EU and non EU countries (Cortes et al., 2010; Dierikx et al., 2012; Ewers et al., 2012; Geser et al., 2012; Kolar et al., 2010; Overerested et al., 2012; Seiffert et al., 2013). In general, most of the available studies are from European countries and indicate prevalence of ESBL and AmpC producing E. coli ranging between 0%-94% and 0%-13%, respectively. Data from Asia and America are limited and there is a lack of data from Africa and Australia (Seiffert et al., 2013). The associated fluoroquinolone, aminoglycoside and tri-methoprim/sulfamethoxazole resistance is quite common in ESBL strains because the genes are often transferred on the same plasmid. The high prevalence of resistant bacteria may lead to many problems in treatment, especially due to associated resis-tance to aminoglycosides and trimethoprim-sulfametho-xazole as well as a high frequency of coexistent fluoro-quinolone resistance. ESBL strains are decreasingly susceptible to 3rd and 4th generation cephalosporins in vitro, which is associated with clinically significant reductions in their efficacy. Antimicrobials containing ß-lactam/ß-lactamase inhibitor combinations are effective
against ESBL, but may not be effective against organisms that produce AmpC-type ß-lactamase. Furthermore, the majority of ESBLs do not hydrolyze cefamycins (cefoxitin and cefotetan). However, cefamycins can be hydrolyzed by AmpC-type strains. Carbapenems are normally highly stable against ß-lactamase hydrolysis and were previously agents of choice in human medicine. The presence of ESBL and AmpC producing E. coli in production animals may serve as a potential reservoir for both resistant bacteria and their resistance genes. Patho-gens can be directly transferred from animals to humans, as it is very well described in Salmonella. As observed in E. coli, animals may also harbor commensal flora, which contains resistant genes that can be transferred horizontally between bacteria by mobile genetic elements, e.g., plas-mids (Seiffert et al., 2013). At this time, the real impor-tance of different reservoirs of resistance genes is difficult to be evaluated and further studies are needed (Stefani, 2012). To access the potential public health risk of resistant bacteria in food, a more detailed comparison of ESBL and AmpC genes of E. coli isolates from animal and human origin is necessary. Recently, the emergence of carbapenemase-producing enterobacteria in people is also known and may become a potential problem in food producing animals, too. Carba-penems are not allowed for use in veterinary medicine, but control of resistant strains is highly recommended.

OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT
43
Conclusions
Findings of resistant bacteria and their potential zoonotic transmission, especially along the food chain, have emerged as a major concern in human healthcare. The retail meat might be an important vehicle for the community-wide dissemination of antimicrobial resistant E. coli. It is recommended to supplement standard surveillance methods with methods based on selective enrichment when monitoring ESBLs. Combination of chromogenic and selective culture media is very useful for rapid isolation and identification of resistant bacteria. Currently, carbapenems are in general regarded as the preferred agents for treatment of infections due to ESBL-producing organisms in humans.
Acknowledgments
This work was partly granted by the Slovenian Research Agency, Research Working Group P4-0092, research pro-ject V4-1080, and Veterinary Administration of Republic of Slovenia (Ministry of Agriculture, Forestry and Food).
References
AMBROŽIČ, J., I. ZDOVC, I. ŠTRUMBELJ (2012): Escherichia coli producing extended spectrum beta-lactamases: genotypes of isolates from diseased animals and food of animal origin compared with genotypes of human clinical isolates. 4. Baničevi dnevi, Radenci, November 2012. Zoonoze: [zbornikprispevkov], (Medicinskirazgledi, Supplement, letn. 51, 6). Ljubljana: Medicinski razgledi, 2012, 219-225.
BAILEY, J., J. L. PINYON, S. ANANTHAM, R. HALL (2010): Commensal Escherichia coli of healthy humans: a reservoir for antibiotic resistance determinants. J. Med. Microbiol. 59, 1331-1339.
Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Susceptibility Testing, Twenty-Third Informational Supplement. CLSI Document M100-S23, January 2013.
CORTES, P., V. BLANC, A. MORA, et al. (2010): Isolation and characterisation of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain. Appl. Evniron. Microbiol. 76(9): 2799-2805.
DIERIKX, M. C., E. Van DUIJKEREN, A. H. W. SCHOOR-MANS, A. Van ESSEN-ZANDBERGEN, K. VELDMAN, A. KANT, X.W. HUIJSDENS, K. Van der ZWALUW, J. A. WAGENAAR, D. J. MEVIUS (2012): Occurrence and characteristics of extended-spectrum-ß-lactamase- and AmpC-producing clinical isolates derived from companion animals and horses. J. Antimicrob. Chemother. 67(6), 1-7.
EWERS, C., A. BETHE, T. SEMMLER, S. GUENTHER, L. H. WIELER (2012): Extended-spectrum β-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin. Microbiol. Infect. 18, 646-655.
GESER, N., R. STEPHAN, H. HACHLER (2012): Occurrence and characteristics of extended-spectrum β-lactamase (ESBL) producing animals, minced and raw milk. BMC Vet. Res. 8:21.
GOLOB, M., B. KRT, I. ZDOVC (2012): Detection of ESBL- and AmpC-producing Escherichia coli isolates in poultry meat in Slovenia. V: CESAR 2012: Central European Symposium on Antimicrobial Resistance, Primošten, Croatia, 23-26 September, 2012: Book of Abstracts. Zagreb: Croatian Microbiological Society, 87.
GREGOVA, G., M. KMETOVA, V. KMET, J. VENGLOVSKY, A. FEHER (2012). Antibiotic resistance of E. coli isolated from a poultry slaughterhouse. Ann. Agric. Environ. Med. 19(1), 75-77.
JOHNSON T. J., KARIYAWASAM S., WANNEMUEHLER Y., et al. (2007): The genome sequence of avian pathogenic Escherichia coli strain O1:K1:H7 shares strong similarities with human extraintestinal pathogenic E. coli genomes. J. Bacteriol. 189(8), 3228-3236.
KOLAR, M., J. BARDON, M. CHROMA, et al. (2010): ESBL and AmpC beta-lactamase-producing Enterobacteriaceae in poultry in the Czech Republic. Vet. Med. 55(3), 119-124.
KÖHLER, C. D., U. DOBRINDT (2011): What defines extra-intestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 301, 642-647.
KAPER, J.B., J.P. NATARO, H.L.T. MOBLEY (2004): Nature Reviews/Microbiology. 2, 123-140.
LEVERSTEIN-Van HALL, MA, C. M. DIERICKX, J. COHEN STUAR, et al. (2011): Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17(6), 873-880.
NAKAZATO, G, T. AMABILE de CAMPOS, E. GUEDES STEHLING, et al.(2009): Virulence factors of avian pathogenic Escherichia coli (APEC). Pesq. Vet. Bras. 29(7), 479-486.
OVERDEVEST, I., I. WILLEMSEN, M. RIJNSBURGER, et al. (2011): Extended-spectrum β-lactamase genes of Esche-richia coli in chicken meat and humans, The Netherlands. Emerg. Infect. Dis. 17(7), 1216-1222.
SEIFFERT, N.-S., M. HILTY, V. PERRETEN, A. ENDIMIANI (2013): Extended-spectrum cephalosporin-resistant gram-negative organisms in livestock: an emerging problem for human health? Drug Resist. Update (2013), http://dx.doi.org/10.1016/j.drup.2012.12.001.
SHARMA, R., K. MUNS, T. ALEXANDER, et al. (2008): Diversity and distribution of commensal fecal Escherichia coli bacteria in beef cattle administered selected subthera-peutic antimicrobials in a feedlot setting. Appl. Environ. Microbiol. 74(20), 6178-6186.
SMITH, J. L., D. J. V. DRUM, Y. DAY et al. (2007): Impact of antimicrobial usage on antimicrobial resistance in com-mensal Escherichia coli strains colonizing broiler chickens. Appl. Environ. Microbiol. 73(5), 1404-1414.
STEFANI, S (2012): Analyzing possible intersections in the resistome among human, animal, and environmental matrices. Front. Microbiol. 3 Article 418, 1-2.

OCCURRENCE OF ESBL- AND AmpC-PRODUCING ESCHERICHIA COLI ISOLATES IN POULTRY MEAT 44
POJAVNOST ESBL- I AmpC-PRODUCIRAJUĆIH SOJEVA ESCHERICHIA COLI U MESU PERADI
Sažetak Cilj ovoga istraživanja bio je ispitati učestalost sojeva bakterije Escherichia (E.) coli koji proizvode β-laktamaze proširenog spektra (ESBL) i AmpC u mesu peradi u maloprodaji. Uz to se procjenjivala pridružena rezistencija na druge antimikrobne agense među sojevima pozitivnim na ESBL i AmpC. Uzorci sirovog mesa prikupljeni su putem slovenskog programa nadzora rezistencije na antibiotike kod bakterija izoliranih iz životinja namijenjenih za proizvodnju hrane tijekom 2011. i 2012. godine. Postupak je proveden na 446 uzoraka mesa brojlera primjenom neselektivnih i selektivnih podloga za rast. U izolatima je proizvodnja β-laktamaze fenotipski potvrđena metodom disk difuzije i mikrodilucije primjenom indikatornih antimikrobnih lijekova. Uzorci su bili teško kontaminirani bakterijom E. coli koja proizvodi ESBL (46,4%) i E. coli koja proizvodi AmpC (36,9%). Među njima otkriven je visok postotak izolata s pridruženom rezistencijom na druge antimikrobne lijekove, najčešće na fluorokinolone, tetracikline, streptomicin, trimetoprim/sulfametoksazol i kloramfenikol. Ključne riječi: antimikrobna rezistencija, ispitivanje osjetljivosti, Escherichia coli, beta-laktamaze proširenog spectra (ESBL), AmpC, perad

PERADARSKI DANI 2013.
45
PREVENTION OF MG-MS INFECTION USING LIVE VACCINES
László Kőrösi, János Povazsán
Rhone Vet Kft, Budapest, Hungary Summary Mycoplasmosis is still one of the costliest diseases for the modern poultry industry. The infected birds/flocks are carriers for life. Mycoplasma (M.) gallisepticum (MG) causes respiratory syndromes, egg-drops and increased mortality during production, and embryonic death during hatch. MG causes severe post vaccination reactions, serious respiratory syndromes, increased mortality, medication costs and increased condemnation at slaughter, with poor weight gain and feed conversion ratio in the vertically infected broilers. Mycoplasma(M.) synoviae (MS) is traditionally considered the second most important avian mycoplasma species for commercial chickens from the clinical and economic points of view. It has been associated with respiratory disease and subsequent condemnations due to airsacculitis in broilers and peritonitis and mortality in commercial layer hens, although subclinical infections of the respiratory tract seem predominant. MS is also known to cause synovitis in chickens. Significant advances in the control of mycoplasmosis have been recorded since the introduction of the third generation attenuated live M. gallisepticum (Ts11) and M. synoviae (MS-H) vaccines. These third generation mycoplasma vaccines were specifically attenuated (for temperature sensitivity (ts)) and selected to combat the effects of wild mycoplasma challenge. Key words: mycoplasmosis, mycoplasma vaccines Introduction Mycoplama (M.) gallisepticum (MG) and Mycoplasma (M.) synoviae (MS) pathogens of domestic poultry cause significant economic losses worldwide (Liers et al., 2007). The losses are due to complicated postvaccinal reactions, decreased egg production/hatchability, egg downgrading, carcass condemnation, and decreased feed efficiency due to MG and MS. Since 2000, a novel egg shell apex abnormality (EAA) has been increasingly found in table egg producing chicken flocks and broiler breeders, and MS was isolated from the oviduct of birds that produced abnormal eggs (Feberwee et al., 2009). Recent studies have shown that MS, besides causing production losses on its own, could be a significant factor in initiating Esche-richia coli peritonitis, a major cause of mortality in layers (Raviv et al., 2007).
Discussion
Control of MG and MS infection could be achieved by eradication, antibiotics and vaccines. The eradication is quite successful at elite breeding level providing free
flocks for the industry, but this eradication is by far less successful at the production level. With rapid growth of poultry production worldwide, there has been concen-tration of large numbers of birds in small areas, leading to an increased risk of exposure to pathogenic mycoplasmas. In some areas, poultry production is so concentrated that from the epidemiological point of view, it is almost like a very large multi-age farm. There has been a tendency to drift away from all-in all-out production and to concentrate production on multi-age sites, especially in case of com-mercial egg production, but in many locations multi-age management of broiler breeders or broilers may occur. Therefore, in spite of sometimes heroic efforts in bio-security and improved understanding of the survival of mycoplasmas outside the host, mycoplasma outbreaks continue to occur. Antibiotic treatments reduce mycoplasma population, clinical signs, losses, contamination of eggs and embryos, but at long-term it is associated with the development of resistance to antibiotics and residues in poultry products. Antibiotic treatment is quite costly considering the high failure rate and the industry will not accept residues in poultry products in the future.
Dr. László Kőrösi, Rhone Vet Kft, 1119 Budapest, Fehérvári út 89-95; Phone: +36309820054; fax: +3628420640; e-mail: [email protected]

PREVENTION OF MG-MS INFECTION USING LIVE VACCINES 46
Vaccination was the next attempt to control avian myco-plasmas. The first commercially available mycoplasma vaccines were oil-emulsion bacterins. Bacterins protect against egg production losses generating plenty of humoral antibodies, but provide little protection against coloni-zation by field strains of mycoplasma in respiratory tract, thus providing little value in eradication programs. Major disadvantages of bacterins are the need of two doses for optimal protection and the cost of administration. Significant advances in the control of mycoplasmosis have been achieved since the introduction of third generation attenuated live M. gallisepticum (Ts-11) and M. synoviae (MS-H) vaccines. These third generation mycoplasma vaccines were specifically attenuated, i.e. selected for temperature sensitivity (ts) and to combat the effects of wild mycoplasma challenge. Table 1 summarizes the history of avian mycoplasma vaccination. A major advantage of the ts vaccines is that attenuation makes them very safe but they still are
immunogenic (protective; not necessarily causing serum antibodies). The ts strains are temperature sensitive chemically-induced mutant strains that grow well at 33 °C, but poorly at 39.5 °C, capable of replicating in the mucosal trachea without surviving in the oviduct because of their inability to withstand temperatures higher than 39.5 °C, thereby avoiding transovarian transmission in the progeny. The useful immunity generated by live vaccines is at the mucosal surface of the respiratory tract and it is not necessarily proportional to humoral antibody. In a series of experiments, birds that were vaccinated with Ts-11 and selected with low serologic responses were taken from the field and challenged in the laboratory. They were significantly protected. Indeed, in these experiments comparison to birds vaccinated with MG bacterins with enormous amounts of humoral antibody showed them not to be protected against respiratory challenge.
Table 1. History of the development of avian mycoplasma vaccines
Vaccine strategy Examples Advantages Disadvantages
Killed bacterins 1960s
Numerous No live organisms so reversion to virulence (RTV) cannot occur
Limited duration of immunity and delay of the onset of egg production Does not stop wild type infection or vertical transmission
First generation live (vontrolled exposure) 1960s
Local wild strains and strains arriving vertically from breeding programs
Prevented egg production drops in lay by making sure infection occurred before lay
Loss of up to 20 eggs per hen and increased feed conversion ratio, etc. from subclinical effects of infection
Second generation live (mild strains) 1970s
F strain 6/85 K5831 Others
Do not have to worry about RTV issues during registration (the strain is mild)
Appears to be direct relationship between immunogenicity and residual pathogenicity
Third generation live (attenuated strains) 1980s
Ts-11 MS-H
Dissociation of the relationship between immunogenicity and pathogenicity
No transfer of protection to progeny
Subunit 1990s
MG pox vectored virus No serologic response GMO Predicted pathogen evolution will make effective life span of such vaccines short
Table 2. Summary of trials looking at humoral antibody and protection from MG challenge in birds from the field
Group Antibody at time of challenge Assessment of immunity by measurement of tracheal thickening after challenge
Non vaccinated controls No demonstrable antibody No protection against tracheal thickening with virulent MG challenge
Birds selected from vaccinated flocks with no measurable antibody No demonstrable antibody Protection against tracheal thickening
Pen trials with birds vaccinated with bacterin Nearly all birds had 4+ MG RSA antibody reactions
No protection against tracheal thickening with virulent MG challenge

PREVENTION OF MG-MS INFECTION USING LIVE VACCINES
47
Vaccinated 3w Challenged 17w
Vaccinated 3w Not challenged
Not vaccinated Challenged 17w
Figure 1. Tracheal thickening after MG challenge. All birds
had no measurable antibody at challenge We performed safety and efficacy trials with Ts-11 (Stip-kovits et al., 2003). The aims of the study were as follows: - study of the safety - study of serologic response induced by vaccine - study of efficacy of MG TS-11 vaccine on 49-day-old
MG- and MS-free chicken challenge with MG R strain (gross lesions, histology, reisolation)
- distinguishing vaccine strain from wild strains Table 3. Design of experiment
Day -14
Day old
Day 28
Day 38
Group formation 1, 2, 3 and 4 Vaccination 2 and 4 Bleeding All All All All Body weight All All All Challenge 1 and 2 Clinical observation All All All Pathology, MG reisolation All For the experiment, M. gallisepticum and M. synoviae free commercial Tetra SL layer (Bábolna Agriculture Com-pany) hens were used. The parent stock was regularly monitored serologically for the status of mycoplasma infection by SA, MG and MS blocking ELISA tests (MyGa and MySy ELISA test) with negative results. Chickens were obtained at age of one day and were reared in isolated room till the age of 49 days. From the same hatch, additional 15 one-day-old chicks were slaughtered and checked for the presence of MG antibodies as well as for the presence of mycoplasmas by culturing and PCR. At the age of 49 days (day 0 of the experiment), 60 chickens were individually labeled with colored and numbered wing tags in both wings and their body weight was measured. The birds were regrouped into four groups,
15 chickens/group, that way the average body weight of the groups did not differ from each other. Each group was placed in a separate room and different animal keepers ran them. Group 1 (NV + C) was not vaccinated but challen-ged at age of 77 days. Group 2 (V + C) was vaccinated at age of 49 days (day 0) and challenged with aerosol of 48-hour-old broth culture of M. gallisepticum R-strain (3x108
CFU/mL). (The chickens were placed in a box of 220 L volume and 10 mL of broth culture was sprayed during 100 seconds and exposure was 20 minutes). Group 3 (NV + NC) was left without vaccination and challenge as negative control. Group 4 (V + NC) was vaccinated as group 2 and left without challenge. Vaccination with Ts-11 vaccine was performed intraocularly (0.03 mL/chicken) according to the manufacturer's instructions. Chickens were clinically observed daily from day 0 to day 38. Body weight of chickens was measured before vacci-nation (day 0), for weeks post vaccination, before challenge (day 28) and on day 38. Serologic examination was performed by SA using both M. gallisepticum stained antigen (batch no. 650-1001, expiry date: 03-2003) and M. synoviae antigen (batch no. 71232A, expiry date: 07-2003) as well as by ELISA (MyGa and MySy kits (batch no. 020114, expiry date: 09-2002, batch no. 020506, expiry date: 09-2002, respectively) on day -14, day 0 (before vaccination), day 28 (before challenge) and day 38. At the end of the experimental period, chickens were euthanized and examined for lesions in thoracic air sacs and peri-toneum characteristic of M. gallisepticum infection and scored from 0 to 4 for left and right thoracic air sacs and peritoneum separately. Tracheas, lungs, spleens and livers of chickens were examined histologically. On trachea examination, attention was paid to thickening of mucosal membrane (in micrometers) infiltrated by lymphocytes. On lung examination, characteristic lesions of interstitial pneumonia, lymphohistiocytic and plasmacytic infiltrations around bronchi and bronchioli as well as the presence of catarrhal pneumonia were scored from 0 to 3. In the liver and spleen, activation of lymphofollicular foci was evaluated from 0 to 3. At the end of the experiment, an attempt was made to isolate M. gallisepticum from the trachea, lung, air sacs as well as from the liver, spleen, kidney and heart of all chickens.

PREVENTION OF MG-MS INFECTION USING LIVE VACCINES 48
After the application of TS-11 vaccine, no clinical signs were recorded during the period from day 0 to day 28 or to day 38 in any of the treated groups. After post challenge in NV + C, mild respiratory signs were recorded in 8 birds (scores 9). In contrast, in V + C only one chicken had respiratory rales. No signs were recorded in NV + NC and V + NC. No differences in body weight gains were recorded bet-ween the groups before challenge. However, a significant reduction of body weight gain was recorded in the non-vaccinated challenged group in comparison with other three groups. No difference was noticed between vacci-nated non-challenged group and non-vaccinated non-challenged group. Difference was also observed between V + C and V + NC groups. Similar results were also obtained by daily weight gain testing.
Figure 2. Air sac of a chick immunized with MG TS-11 and
challenged with MG R
Figure 3. Airsacculitis in a NV chick challenged with MG R Experimental chickens were serologically negative for antibodies against M. gallisepticum and M. synoviae on day -14 and day 0 both in SPA and ELISA. On day 28 due
to vaccination, 12 or 11 out of 15 vaccinated birds showed slight serologic response in SA to M. gallisepticum having scores 16 or 13 in SPA to M. gallisepticum antigen. Similar results were obtained on day 38. Due to challenge, all chickens reacted in both NV + C and V + C groups. In unvaccinated and challenged group, 6 animals showed sinusitis, 11 birds had tracheitis, 14 birds had airsacculitis, 7 had pneumonia, 2 had perihepatitis and 1 bird had peri-carditis. At the same time, only 1 bird showed tracheitis and 4 birds showed mild airsacculitis in V + C group. The lesion score of airsacculitis was only 4 in comparison with score 23 in the previous group. No pathologic lesions were recorded in control group and only one chicken showed mild tracheal lesions in V + NC. Due to challenge, tracheal mucosa thickened heavily in most NV + C chickens. Beside this, bronchitis, interstitial pneumonia and catarrhal pneumonia developed with rela-tive high scores of 25, 27 and 13, respectively. These para-meters were statistically significantly higher (P<0.001) than those in the group vaccinated and challenged. At the same time, these data of V + C did not differ statistically from the non-vaccinated, non-challenged or only vaccina-ted groups. Similarly, a significantly higher number of NV + C birds showed lymphohistiocytic infiltration of the liver and spleen, with significantly higher lesion scores than in V + C. There was no difference in these parameters between NV + NC and V + NC. A significant reduction of tracheal mucosa thickness (94.6 μm) was recorded in V + C chickens in comparison with that in NV + C (192.6 μm), while no difference was seen between V + NC and NV + NC (53.3 and 52.3 μm, res-pectively). However, thickness of tracheal mucosal in V + C was significantly greater than in NV + NC and V + NC. In NV + C, successful MG isolation was achieved from 30 out of 45 samples taken from respiratory tract and from 13 samples taken from other inner organs. In V + C group, mycoplasma isolation was successful from only eight samples of respiratory tract and no mycoplasma could be detected in inner organs. No mycoplasma could be detected from any chicken of the other two groups. Examination of seven different MG strains by PCR using pvpA 2R/pvpA 3F primers (Liu et al., 2001) showed no difference between strains except for MG X95 reference strain, which had amplicon of 475 bp instead of 497 bp of other mycoplasma strains. In this examination, no difference was detected between MG TS-11 vaccine strain and MG R low challenge strain. Using restriction enzyme AccI we could demonstrate difference between MG R low challenge strain showing bands of 231, 156 and 110 bp and MG TS-11 vaccine strain having two bands of 341 and 156 bp. At the same time, no difference was detected when the restriction enzyme ScrFI was used. Testing of DNA of isolates from trachea and air sacs of vaccinated as well as vaccinated and challenged chickens by using AccI enzyme demonstrated low DNA pattern characteristic of MG R, thus it was not possible to detect vaccine strains from the immunized and challenged chickens.

PREVENTION OF MG-MS INFECTION USING LIVE VACCINES
49
Using TS-11 vaccine for immunization of M. gallisepticum and M. synoviae free chickens by ocular application, no remarkable clinical signs were noticed during the obser-vation period. Application of vaccine induced slight serologic response to M. gallisepticum antigen in SPA. Vaccination of chickens did not cause decrease in weight gains. By post mortem examination of vaccinated and non-challenged group, very slight thickening of tracheal mucosa and airsacculitis were recorded in only a few birds. Vaccine strain was not isolated from inner organs. Based on these data, it is concluded that the TS-11 is safe for chickens. Due to challenge of NV + C chickens, respiratory signs, significant decrease of body weight gain, thickening of trachea, airsacculitis, interstitial and catarrhal pneumonia, bronchitis and presence of mycoplasmas in inner organs were recorded. After challenge of V chickens, no respiratory signs, interstitial or catarrhal pneumonia, bronchitis, decrease of body weight gain, or penetration of mycoplasma into the inner organs were observed. We were able to eradicate field MG positivity from a broiler breeder integration company in Hungary with long-term use of TS-11 vaccine (Kőrösi at al., 2007, Beijing Congress, oral presentation). Significant advances have been achieved with the intro-duction of live MS vaccine (MS-H) in large-scale layer farms in Japan (from 2002 to 2006). The vaccinated flocks matured earlier, had decreased mortality and increased peak and persistency production with improved eggshell quality compared with control flocks. The HH cumulative egg production was approximately 800 g more in the vaccinated group, which was equivalent to ~13 eggs. The feed conversion ratio was better in the MS live vaccine group by approximately 7%.
Conclusion
In the light of this discussion, it appears that what we need from avian mycoplasma vaccines is protection against low
intensity challenges and dampening of multiplication and transmission of field strains. Both these aims can be met with ts vaccines and many producers around the world are reaping benefits of these vaccines with none of the downsides of other older vaccines. Do we need serologic responses to monitor vaccination or field challenges? – who cares as long as the birds are protected. We have now developed sophisticated methods of strain identification using genomic tests to analyze field problems. Field experiences utilizing live vaccines have been very favorable in commercial layers, and field experiences in multi-age broiler breeders are also favorable. These experiences suggest that live vaccines may be viable tools for the eradication of mycoplasma infection on multi-age commercial poultry farms.
References
FEBERWEE, A., J. de WITT, W. LANDMAN (2007): Myco-plasma synoviae-associated apex abnormalities. XV WVPA Congress, Beijing (oral presentation).
KÖRÖSI, L., J. POVAZSÁN, C. NEUBAUER, M. HESS, P. KISZEL (2007): Field experiences concerning the longterm use of live, apathogenic, thermosensitive MG vaccine (TS 11) in a broiler PS farm in Hungary. XV WVPA Congress, Beijing (oral presentation).
MORROW, C., K. WHITHEAR (2012): Mycoplasma ts vaccines – 20 years of field experience, pen trials and myths. Int. Hatchery Pract., 25 (5) 13-15.
ZIV RAVIV, N. FERGUSSON-NOEL, V. LAIDINIS, R. WOOTEN, S. H. KLEVEN (2007): Role of Mycoplasma synoviae in commercial layer Escherichia coli peritonitis syndrome. Avian Dis., 51, 685-690.
STIPKOVITS L., J. BÍRÓ, J. POVAZSÁN, L. KŐRÖSI, R. GLÁVITS (2003): Study of safety and efficacy of Mycoplasma gallisepticum TS-11 vaccine. XIII WVPA Congress, Denver (oral presentation).
Ts-11(live MG) vaccine and MS-H (live MS) vaccine manufactured by Bioproperties, Australia under the trademark Vaxsafe MG and Vaxsafe MS.
PREVENTIVA MG-MS INFEKCIJE PRIMJENOM ŽIVIH CJEPIVA
Sažetak
Mikoplazmoza je još uvijek jedna od najskupljih bolesti u suvremenoj peradarskoj industriji. Zaražene ptice/jata doživotno ostaju nositeljima. Mycoplasma (M.) gallisepticum (MG) uzrokuje dišni sindrom, pad nesivosti i povećanu smrtnost tijekom proizvodnog ciklusa, te smrtnost embrija tijekom leženja. MG uzrokuje teške reakcije nakon cijepljenja, ozbiljan dišni sindrom, povećanu smrtnost, troškove za lijekove te povećanu stopu zabrane stavljanja na tržište kod klanja, uza slab porast težine i omjer konverzije hrane kod vertikalno zaraženih brojlera. Mycoplasma (M.) synoviae (MS) se tradicionalno smatra drugom najvažnijom vrstom ptičje mikoplazme za komercijalne kokoši s kliničkog i ekonomskog stajališta. Povezuje se s dišnom bolešću i naknadnom neupotrebljivošću zbog upale zračnih vrećica kod brojlera te zbog peritonitisa i smrtnosti kod komercijalnih nesilica, iako se čini da prevladavaju subkliničke infekcije respiracijskog sustava. Također je poznato da MS uzrokuje sinovitis kod kokoši. Značajan napredak u kontroli mikoplazmoze bilježi se od uvođenja treće generacije oslabljenih živih cjepiva M. gallisepticum (Ts11) i M. synoviae (MS-H). Ova treća generacija cjepiva mikoplazme specifično je oslabljena (prema temperaturnoj osjetljivosti (ts)) i odabrana da se odupre prirodnim učincima mikoplazme. Ključne riječi: mikoplazmoza, cjepiva mikoplazme

PERADARSKI DANI 2013. 50
UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALLUS GALLUS U RAZDOBLJU OD 2010. DO 2012. GODINE
Marica Lolić1, Marin Periškić1, Marija Agičić1, Davor Balić1, Mario Škrivanko1, Hrvoje Krajina1, Mirta Vukičević1, Irena Perković1, Borka Šimpraga2, Kristina Matković3
1Veterinarski zavod Vinkovci, Hrvatski veterinarski institut, Vinkovci, Hrvatska 2Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska 3Zavod za higijenu, ponašanje i dobrobit životinja, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska Sažetak U Veterinarskom zavodu Vinkovci tijekom 2010. do 2012. godine ukupno je pretraženo 4886 uzoraka (1559 uzoraka navlaka za obuću iz objekata za tov pilića, 1646 uzoraka izmeta konzumnih nesilica, 503 uzoraka izmeta rasplodnih nesilica, 217 uzoraka konzumnih jaja i 961 uzorak neizleženih jaja) na prisutnost bakterija iz roda Salmonella. Izdvajanje i identifikacija salmonela iz jaja i izmeta obavljeni su standardnim metodama (HRN EN 6579: 2003; ISO 6579: 2007. Amandman 1: Dodatak D). Pripadnost serovara određena je serološkom tipizacijom polivalentnim i monovalentnim antiserumima za pojedine antigene salmonela. Ukupni broj izdvojenih sojeva salmonela iz pretraživanih uzoraka podrijetlom iz jata peradi vrste Gallus gallus u 2010. bio je 81 ili 5,02%, u 2011. godini 22 ili 1,32%, a u 2012. godini 40 ili 2,50%. Najčešće izdvojeni serovar u svim pretraženim uzorcima bio je Salmonella (S.) Enteritidis. Serovar S. Enteritidis izdvojen je u slijedećim uzorcima: 20 navlaka za obuću iz objekata za tov pilića, 46 izmeta konzumnih nesilica i četiri izmeta rasplodnih nesilica. Serovar Salmonella Typhimurium izdvojen je u dva uzorka izmeta tovnih pilića. Od ostalih serovarova najčešće su izdvojeni S. Mbandaka i S. Fillmore, dok su ostali serovarovi izdvojeni u znatno manjem broju. U navedenom razdoblju ukupno je pretraženo 947 jata peradi Gallus gallus pričem su u njih 56 utvrđene bakterije roda Salmonella. Serovar S. Enteritidis utvrđen je u četiri jata tovnih pilića, 16 jata konzumnih nesilica i dva jata rasplodnih nesilica. Serovar Salmonella Typhimurium utvrđen je u jednom jatu tovnih pilića. Ključne riječi: salmonele, tovni pilići, konzumne nesilice, rasplodne nesilice, jaja, izmet Uvod
Salmoneloze su značajan javnozdravstveni problem koji je povezan s potrošnjom hrane životinjskog podrijetla. Prema izvješću Europske agencije za sigurnost hrane u 2010. godini nastavio se trend smanjenja broja dokazanih slu-čajeva salmonelnih infekcija u ljudi kao i proteklih šest godina. Prema navedenom izvješću u 2010. godini broj oboljelih ljudi salmonelnim infekcijama pao je za 8,8% u odnosu na 2009. godinu (EFSA, 2012.). Mnogi stručnjaci iz područja epidemiologije objavili su razne putove kojima se salmonele mogu širiti unutar proizvođača koji se bave peradarskom proizvodnjom u Europi (Heyndrickx i sur., 2002., Davies i Breslin, 2003., Namata sur, 2008.). Infek-
cije salmonelama i salmoneloze u peradi proširene su di-ljem svijeta i uzrokom su velikih ekonomskih gubitaka prouzročenih uginućima, smanjenom proizvodnjom jaja, lošom kvalitetom pilića, visokim troškovima liječenja, visokim troškom nadzora i suzbijanja (Hafez, 2005.). Do-maće i divlje životinje, kao i čovjek, rezervoari su salmo-nela u prirodi. Uz njih, izvor infekcije mogu biti voda, hrana, glodavci, insekti, stelja i zaražena perad. Prijenos i kontaminacija jaja salmonelama mogući su dvojako: transovarijski (vertikalni prijenos) pričem se žumančana membrana ili bjelanjak inficiraju salmonelama putem infi-ciranih reproduktivnih organa (Okamura, 2001.). Uvjet za vertikalni prijenos je infekcija prije stvaranja ljuske u jajovodu; drugi način je prolazak kroz ljusku (horizontalni
Dr. sc. Marica Lolić, Veterinarski zavod Vinkovci, Hrvatski veterinarski institut, Josipa Kozarca 24, 32100 Vinkovci, Hrvatska; Tel: ++ 385 (0) 32 331 288; E-mail: lolic@veinst.

UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALLUS GALLUS U RAZDOBLJU OD 2010. DO 2012. GODINE
51
prijenos) pričem salmonele iz okoliša prodiru kroz ljusku jaja. Prolaskom jaja kroz kloaku ljuska jaja se može onečistiti izmetom kokoši u kojem je prisutna salmonela, pa može doći do njenog prodora kroz ljusku (Messens i sur., 2005., De Reu i sur., 2006.). Jata jednodnevnih pilića mogu se zaraziti salmonelama vertikalnim prijenosom iz zaraženih rasplodnih jata ili horizontalno prijenosom tije-kom leženja, utovara ili tijekom prijevoza do farme (Cox i sur., 1990., Cason i sur., 1994., Chriél i sur., 1999.), a na farmi od peradarske nastambe (Rojas i sur, 2002., Davies i Wray, 1996.). Kao potencijalni izvor infekcija spominju se i kukci (Davies i Wray, 1994.). Neprikladno čišćenje i dezinfekcija važni su čimbenici prisutnosti salmonela u peradarskim objektima (Davies i Breslin, 2003., Rose i sur., 2003., Gradel i sur., 2005., Marin i sur., 2009.). Dezinsekcija i deratizacija tijekom odmora peradarske na-stambe važni su čimbenici kontrole salmoneloza (Davies i Breslin, 2003., Carrique-Mas i sur., 2008.). Zaražene ži-votinje, osobito glodavci, mogu sudjelovati u ponovnoj kontaminaciji peradarskog objekata salmonelama nakon čišćenja i dezinfekcije. Svojim izmetom glodavci mogu onečistiti uskladištenu hranu koja se često spominje kao jedan od najznačajnijih izvora infekcije salmonelama pera-di. Heyndrickx i sur. (2002.) smatraju da hrana i voda u peradarskoj nastambi imaju značajan utjecaj na zdravstve-no stanje jata. Iako je u gotovim smjesama i sirovinama, pogotovo životinjskog podrijetla, teško uvijek dokazati pri-sutnost salmonela, često se isti serotip nađe u hrani i peradi koja je tu hranu konzumirala. Smanjenje rizika od infekcije i redukcija salmonela koje se prenose hranom postiže se primjenom probiotika i cijepljenjem peradi (Hafez, 2008.). Preventivna mjera zaštitite od salmoneloznih infekcija rasplodnih i konzumnih nesilica je cijepljenje živim ili mrtvim cjepivima. Cijepljenje tovnih pilića nije opravdano zbog njihovog kratkog životnog vijeka koji nije dovoljan da pilići steknu imunitet. U Republici Hrvatskoj kontrola infekcije peradi vrste Gallus gallus salmonelama provodi se u skladu sa zahtjevima nacionalnih programa kontrole salmoneloza (Nacionalni program kontrole salmoneloze u tovnih pilića vrste Gallus gallus u Republici Hrvatskoj, 2010., 2011., 2012.; Nacionalni program kontrole salmone-loze u odraslih rasplodnih jata (rasplodna jata u proiz-vodnji) vrste Gallus gallus u Republici Hrvatskoj, 2010., 2011., 2012.; Nacionalni program kontrole salmoneloze u jatima konzumnih nesilica vrste Gallus gallus u Republici Hrvatskoj, 2010., 2011., 2012.). Svrha ovoga rada je prikazati pojavnost bakterija roda Salmonella u izmetu, neizleženim i konzumnim jajima peradi vrste Gallus gallus pretraženim u Veterinarskom zavodu Vinkovci tijekom razdoblja od 2010. do 2012. godine.
Materijali i metode
Od 2010. do 2012. godine u Laboratoriju za dijagnostiku i Laboratoriju za mikrobiologiju hrane za ljude i hrane za životinje u Veterinarskom zavodu Vinkovci ukupno je pre-traženo 4886 uzoraka podrijetlom iz jata peradi Gallus gallus na prisutnost bakterija iz roda Salmonella. Pre-
traženo je 1559 uzoraka navlaka za obuću iz objekata za tov pilića, 1646 uzoraka izmeta konzumnih nesilica, 503 uzorka izmeta rasplodnih nesilica, 254 uzorka konzumnih jaja i 924 uzorka neizleženih jaja. Uzorke su dostavile ovlaštene veterinarske organizacije u skladu s donesenim Naredbama o mjerama zaštite životinja od zaraznih i na-metničkih bolesti i njihovom financiranju za odgovarajuću godinu (NN 7/2010., NN 1/2011., NN 17/2012.). Izdva-janje i identifikacija salmonela iz konzumnih i rasplodnih jaja i izmeta obavljena je standardnom metodom HRN EN ISO 6579:2003, a iz uzoraka navlaka za obuću i izmeta standardnom metodom ISO 6579:2007., Amandman 1: Dodatak D. Od svakog uzorka navlaka za obuću ili izmeta peradi Gallus gallus odvagano je 25 g i dodano u 225 mL pufe-rirane peptonske vode (BPW, Buffered Peptone Water, Biolife, Italija) te inkubirano pri 37±1 °C, 18±2 sata. Na-kon inkubacije uzorci izmeta precijepljeni su na modifici-ranu polumeku podlogu Rappaport Vassiliadis (MSRV, Rappaport-Vassiliadis Semisolid Medium Modified, Bio-life, Italija) na način da je sadržaj nacijepljene peptonske vode nacijepljen ubodom u podlogu na tri mjesta u količini od 0,1 mL te inkubiran pri 41,5±1 °C, 24±3 sata. Nakon 24 sata bakterije su s podloge MSRV u području prisutnosti zone migracije mikrobiološkom ušicom precijepljene na selektivne hranjive podloge XLD (Xylose Lysine Deoxy-cholate Agar, Biolife, Italija) i CSA (Chromogenic Salmonella Agar Base, Biolife, Italija) te inkubirane pri 37±1 °C, 24±3 sata. Od uzorka konzumnih i neizleženih jaja napravljena je ho-mogena smjesa (žumanjak i bjelanjak 10 jaja). Odvagano je 50 g homogenog uzorka i dodano u 450 mL puferirane peptonske vode te inkubirano pri 37±1 °C, 18±2 sata. Na-kon inkubacije je 0,1 mL inkubiranog medija precijepljeno u epruvetu s 10 mL Rappaport Vassiliadis bujona (RVS, Rappaport Vassiliadis Soy Broth, Biolife, Italija) i 1 mL u epruvetu s 10 mL Muller-Kauffmann tetrationat/novobio-cin bujona (MKTT, Muller Kauffmann Tetrathionate Broth Base Iso Formulation, Biolife, Italija + Novobiocin MKTT Selective Supplement, Biolife, Italija). Epruvete s RVS bujonom inkubirane su pri 41,5 °C±1 °C, 24±3 sata, a epruvete s MKTT bujonom pri 37 °C±1 °C, 24±3 sata. Nakon inkubacije iz svakog bujona mikrobiološkom uši-com nacijepljeno je oko 10 μL na čvrste selektivne podloge XLD i CSA koje su inkubirane pri 37 °C±1 °C, 24±3 sata. Nakon pregleda XLD i CSA ploča sumnjive kolonije porasle na čvrstim hranjivim podlogama (četiri do pet), precijepljene su na trostruki šećer agar sa željezom (TSI, Triple Sugar Iron Agar USP, Biolife Italija) i LIA (Lysine Iron Agar, Biolife, Italija). Epruvete s TSI agarom i LIA agarom inkubirane su pri 37 °C±1 °C, 24±3 sata. Za biokemijsku identifikaciju salmonela korišteni su testo-vi BBL Crystal (Becton Dickinson, SAD) i Vitek 2 Compact 15, (bioMérieux, Francuska), a za serološku tipi-zaciju polivalentni i monovalentni antiserumi (Bio-rad, Francuska). Aglutinacijom na predmetnom stakalcu potvr-

UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALLUS GALLUS U RAZDOBLJU OD 2010. DO 2012. GODINE
52
đena je pripadnost bakterija rodu Salmonella. Serovar salmonela određen je prema shemi Kauffmann White (Gri-mont i Weill, 2007.). Izdvojeni bakterijski sojevi Salmo-nella potvrđeni su u Nacionalnom referentnom laboratoriju Hrvatskoga veterinarskog instituta, Centra za peradarstvo, Zagreb.
Rezultati i rasprava
U Veterinarskom zavodu Vinkovci od ukupno 4886 pre-traženih uzoraka podrijetlom iz jata peradi Gallus gallus u 143 izdvojene su bakterije roda Salmonella. Pritom je u 2010. godini izdvojen 81 soj ili 5,02%, u 2011. godini 22 soja ili 1,32%, a u 2012. godini 40 sojeva ili 2,50%. Najčešći izdvojeni serovar u svim pretraženim uzorcima bio je S. Enteritidis. Serovar S. Enteritidis izdvojen je iz slijedećih uzoraka: 20 navlaka za obuću iz objekata za tov pilića, 46 izmeta konzumnih nesilica i četiri izmeta rasplodnih nesilica. Serovar S. Typhimurium izdvojen je iz dva uzorka navlaka za obuću. Od ostalih serovarova najčešće su izdvojeni serovar S. Mbandaka iz 30 uzoraka navlaka za obuću i pet uzoraka neizleženih jaja, serovar S. Fillmore iz osam uzoraka navlaka za obuću iz objekata za
tov pilića, dva uzorka izmeta rasplodnih nesilica i tri uzorka neizleženih jaja i serovar S. Tennessee iz sedam uzoraka izmeta konzumnih nesilica, dok su serovarovi S. Mowanjum, S. Agona, S. Montevideo, S. Coeln, S. Stanleyville i S. Senftenberg izdvojeni u znatno manjem broju (tablica 1.). Na temelju dobivenih rezultata u razdoblju od 2010. do 2012. godine utvrđeno je da je S. Enteritidis bila dominantan izolirani serovar. Takve nalaze potvrđuju i izvješća drugih autora (Imberechts i Dierick, 2004., Krstulović i sur. 2007., De Reu i sur. 2008., Marin i sur. 2011.). U 2010. godini serovar S. Enteritidis izdvojen je iz 14 uzoraka navlaka za obuću iz objekata za tov pilića, 27 uzoraka izmeta konzumnih nesilica i dva uzorka izmeta rasplodnih nesilica. Ostali serovarovi Salmonella izolirani su iz 26 uzoraka navlaka za obuću iz objekata za tov pilića, dva uzorka izmeta konzumnih nesilica, četiri uzorka izmeta rasplodnih nesilica i osam uzoraka neizleženih jaja. U 2011. godini serovar S. Enteritidis izdvojen je iz četiri uzorka navlaka za obuću iz objekata za tov pilića i osam uzoraka izmeta konzumnih nesilica, dok su ostali serovarovi salmonela izdvojeni iz četiri uzorka navlaka za obuću i šest uzoraka izmeta konzumnih nesilica.
Tablica 1. Bakterije roda Salmonella izdvojene iz pretraživanih uzoraka podrijetlom iz jata peradi vrste Gallus gallus u
razdoblju od 2010. do 2012. godine u Veterinarskom zavodu Vinkovci
2010. 2011. 2012.
Izmet tovnih pilića
Izmet konzum. nesilica
Izmet rasplod. nesilica
Neizležena jaja
Konzum. jaja
Izmet tovnih pilića
Izmet konzum. nesilica
Izmet rasplod. nesilica
Neizležena jaja
Konzum. jaja
Izmet tovnih pilića
Izmet konzum. nesilica
Izmet rasplod. nesilica
Neizležena jaja
Konzum. jaja
Bakterija roda Salmonella
Pretraženo/pozitivno/ (%) Pretraženo/pozitivno/ (%) Pretraženo/pozitivno/ (%)
S. Enteritidis 514/14 (2,72)
481/27 (5,61)
257/2 (0,77)
310/0 (0)
52/0 (0)
485/4 (0,82)
590/8 (1,36)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0,36)
575/11 (1,91)
36/2 (5,56)
328/0 (0)
101/0 (0)
S. Typhimurium 514/0
(0) 481/0
(0) 257/0
(0) 310/0
(0) 52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0,36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Mbdaka 514/14 (2,72)
481/0 (0)
257/0 (0)
310/5 (1,61)
52/0 (0)
485/4 (0,82)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/12 (2,14)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Fillmore 514/8 (1,56)
481/0 (0)
257/2 (0,77)
310/3 (0,97)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Mowanjum 514/2 (0,39)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Agona 514/2 (0,39)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Montevideo 514/0
(0) 481/0
(0) 257/0
(0) 310/0
(0) 52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Norwich 514/0
(0) 481/2 (0,42)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Tennessee 514/0
(0) 481/0
(0) 257/0
(0) 310/0
(0) 52/0 (0)
485/0 (0)
592/2 (0,34)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/5 (0,87)
36/0 (0)
328/0 (0)
101/0 (0)
S. Coeln 514/0
(0) 481/0
(0) 257/0
(0) 310/0
(0) 52/0 (0)
485/0 (0)
590/2 (0,34)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0,36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Stanleyville 514/0
(0) 481/0
(0) 257/0
(0) 310/0
(0) 52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0,36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Senftenberg 514/0
(0) 481/0
(0) 257/0
(0) 310/0
(0) 52/0 (0)
485/0 (0)
590/2 (0,34)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0,36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
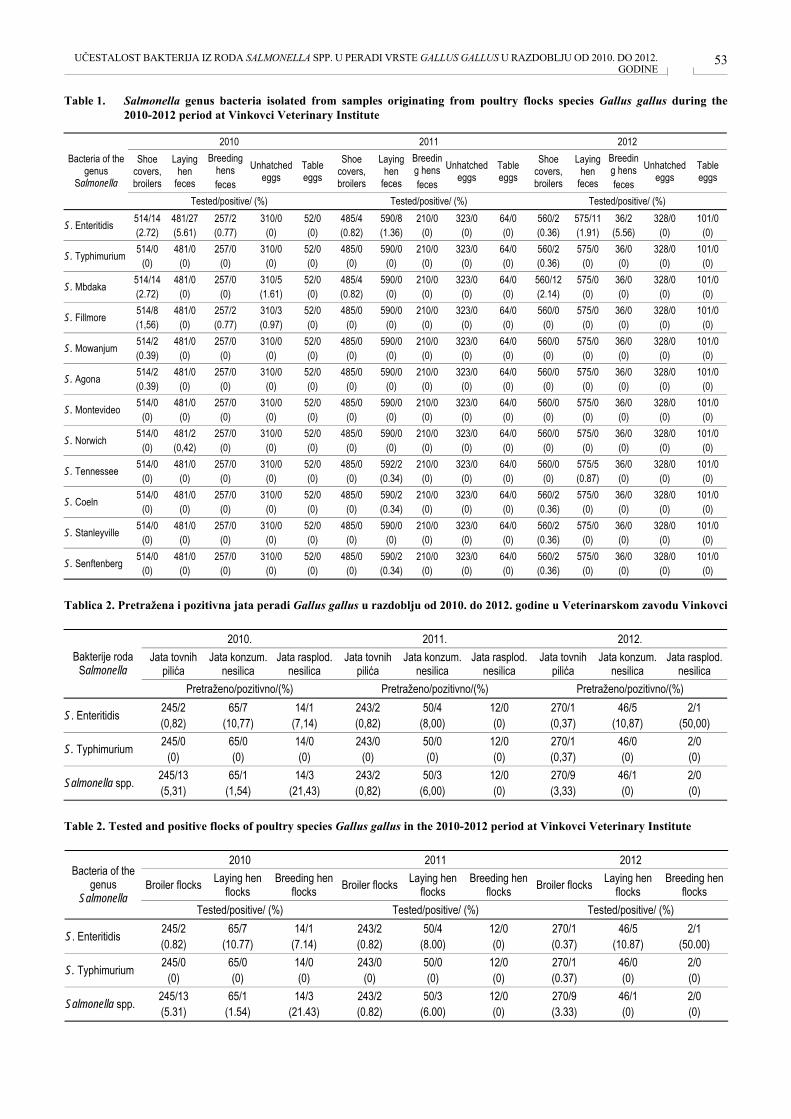
UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALLUS GALLUS U RAZDOBLJU OD 2010. DO 2012. GODINE
53
Table 1. Salmonella genus bacteria isolated from samples originating from poultry flocks species Gallus gallus during the 2010-2012 period at Vinkovci Veterinary Institute
2010 2011 2012 Shoe
covers, broilers
Laying hen
feces
Breeding hens feces
Unhatched eggs
Table eggs
Shoe covers, broilers
Laying hen
feces
Breeding hens feces
Unhatched eggs
Table eggs
Shoe covers, broilers
Laying hen
feces
Breeding hens feces
Unhatched eggs
Table eggs
Bacteria of the genus
Salmonella Tested/positive/ (%) Tested/positive/ (%) Tested/positive/ (%)
S. Enteritidis 514/14 (2.72)
481/27 (5.61)
257/2 (0.77)
310/0 (0)
52/0 (0)
485/4 (0.82)
590/8 (1.36)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0.36)
575/11 (1.91)
36/2 (5.56)
328/0 (0)
101/0 (0)
S. Typhimurium 514/0 (0)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0.36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Mbdaka 514/14 (2.72)
481/0 (0)
257/0 (0)
310/5 (1.61)
52/0 (0)
485/4 (0.82)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/12 (2.14)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Fillmore 514/8 (1,56)
481/0 (0)
257/2 (0.77)
310/3 (0.97)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Mowanjum 514/2 (0.39)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Agona 514/2 (0.39)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Montevideo 514/0 (0)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Norwich 514/0 (0)
481/2 (0,42)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Tennessee 514/0 (0)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
592/2 (0.34)
210/0 (0)
323/0 (0)
64/0 (0)
560/0 (0)
575/5 (0.87)
36/0 (0)
328/0 (0)
101/0 (0)
S. Coeln 514/0 (0)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/2 (0.34)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0.36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Stanleyville 514/0 (0)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/0 (0)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0.36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
S. Senftenberg 514/0 (0)
481/0 (0)
257/0 (0)
310/0 (0)
52/0 (0)
485/0 (0)
590/2 (0.34)
210/0 (0)
323/0 (0)
64/0 (0)
560/2 (0.36)
575/0 (0)
36/0 (0)
328/0 (0)
101/0 (0)
Tablica 2. Pretražena i pozitivna jata peradi Gallus gallus u razdoblju od 2010. do 2012. godine u Veterinarskom zavodu Vinkovci
2010. 2011. 2012. Jata tovnih
pilića Jata konzum.
nesilica Jata rasplod.
nesilica Jata tovnih
pilića Jata konzum.
nesilica Jata rasplod.
nesilica Jata tovnih
pilića Jata konzum.
nesilica Jata rasplod.
nesilica Bakterije roda
Salmonella Pretraženo/pozitivno/(%) Pretraženo/pozitivno/(%) Pretraženo/pozitivno/(%)
S. Enteritidis 245/2 (0,82)
65/7 (10,77)
14/1 (7,14)
243/2 (0,82)
50/4 (8,00)
12/0 (0)
270/1 (0,37)
46/5 (10,87)
2/1 (50,00)
S. Typhimurium 245/0 (0)
65/0 (0)
14/0 (0)
243/0 (0)
50/0 (0)
12/0 (0)
270/1 (0,37)
46/0 (0)
2/0 (0)
Salmonella spp. 245/13 (5,31)
65/1 (1,54)
14/3 (21,43)
243/2 (0,82)
50/3 (6,00)
12/0 (0)
270/9 (3,33)
46/1 (0)
2/0 (0)
Table 2. Tested and positive flocks of poultry species Gallus gallus in the 2010-2012 period at Vinkovci Veterinary Institute
2010 2011 2012
Broiler flocks Laying hen flocks
Breeding hen flocks Broiler flocks Laying hen
flocks Breeding hen
flocks Broiler flocks Laying hen flocks
Breeding hen flocks
Bacteria of the genus
Salmonella Tested/positive/ (%) Tested/positive/ (%) Tested/positive/ (%)
S. Enteritidis 245/2 (0.82)
65/7 (10.77)
14/1 (7.14)
243/2 (0.82)
50/4 (8.00)
12/0 (0)
270/1 (0.37)
46/5 (10.87)
2/1 (50.00)
S. Typhimurium 245/0 (0)
65/0 (0)
14/0 (0)
243/0 (0)
50/0 (0)
12/0 (0)
270/1 (0.37)
46/0 (0)
2/0 (0)
Salmonella spp. 245/13 (5.31)
65/1 (1.54)
14/3 (21.43)
243/2 (0.82)
50/3 (6.00)
12/0 (0)
270/9 (3.33)
46/1 (0)
2/0 (0)

UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALLUS GALLUS U RAZDOBLJU OD 2010. DO 2012. GODINE
54
U 2012. godini serovar S. Enteritidis izdvojen je iz dva uzorka izmeta tovnih pilića, 11 uzoraka izmeta konzumnih nesilica i dva uzorka izmeta rasplodnih nesilica, dok su ostali serovarovi salmonela izdvojeni iz 18 uzoraka na-vlaka za obuću iz objekata za tov pilića i pet uzoraka izmeta konzumnih nesilica (tablica 1.). U 2012. godini ukupni broj izdvojenih salmonela ponovno je porastao, što može biti posljedica nedovoljnih službenih kontrola siro-vina ili gotovih smjesa za perad. To potvrđuje tvrdnju Jonesa (2011.) da je nalaz salmonela u hrani sigurno prijetnja kako za zdravlje i proizvodnost peradi tako i za zdravlje ljudi. Analizirani uzorci bili su podrijetlom iz 947 jata peradi vrste Gallus gallus i to 758 jata tovnih pilića, 161 jata konzumnih nesilica te 28 jata rasplodnih nesilica. Serovar S. Enteritidis utvrđen je u 2010. godini u dva jata tovnih pilića, odnosno 0,82%, sedam jata konzumnih nesilica, odnosno 10,77% i jednog jata rasplodnih nesilica, odnosno 7,14%. U 2011. godini isti serovar izdvojen je iz dva jata tovnih pilića odnosno 0,82% i četiri jata konzumnih ne-silica odnosno 8,00%. U 2012. godini utvrđen je u jednom jatu tovnih pilića odnosno 0,37%, pet jata konzumnih nesilica odnosno 10,87% i jednom jatu rasplodnih nesilica odnosno 50,00%. Serovar S. Typhimurium utvrđen je u jednom jatu tovnih pilića odnosno 0,37% (tablica 2.). Prema izvješću Europske agencije za sigurnost hrane u većini europskih zemalja učestalost pojava salmoneloza u peradi pokazuje trend opadanja (EFSA, 2012.).
Zaključci
Na temelju dobivenih rezultata može se zaključiti da je najmanji broj izdvojenih salmonela, kao i broj pozitivnih jata bio u 2011. godini. S. Enteritidis bila je najčešći izdvojeni serovar salmonela iz uzoraka podrijetlom iz jata peradi Gallus gallus. Serovar S. Typhymurium izoliran je iz samo jednog jata tovnih pilića 2012. godine. Od ostalih serovarova najčešće su izdvajani S. Mbandaka i S. Fillmore, dok je ostalih serovarova izdvojeno 0,47%. Pro-vođenjem Nacionalnih programa kontrola salmoneloza u peradi vrste Gallus gallus broj pozitivnih jata zaraženih bakterijom S. Enteritidis se smanjuje, iako je ona i dalje prisutna u peradarskoj proizvodnji. Da bi se smanjio rizik zaražavanja peradi, a time i zaražavanja ljudi salmonelama potrebno je, uz redovitu kontrolu izmeta i proizvoda od peradi kontinuirano kontrolirati sirovine i gotove smjese za hranidbu peradi, te provoditi trajno obrazovanje osoba koje se bave peradarskom proizvodnjom.
Literatura
CASON, J., N. COX, J. BAILEY (1994): Transmission of Salmonella typhimurium during hatching of broiler chicks. Avian Dis. 38, 583-588.
CHRIÉL, M., H. STRYHN, G. DAUPHIN (1999): Generalized linear mixed models analysis of risk factors for contami-
nation of Danish broiler flocks with Salmonella typhi-murium. Prev. Vet. Med. 40, 1-17.
COX, N., J. BAILEY, L. BLANKENSHIP, R. MEINERS-MANN, N. STERN, F. McHAN (1990): Fifty percent colonization dose for Salmonella typhimurium administered orally and intracloacally to young broiler chicks. Poultry Sci. 69, 1809-1812.
DAVIES, R.H., M. BRESLIN (2003): Observations on Salmo-nella contamination of commercial laying farms before and after cleaning and disinfection. Vet. Rec. 152, 283-287.
DAVIES, R.H., C. WRAY (1995): Mice as carriers of Salmonella enteritidis on persistently infected poultry units. Vet. Rec. 137, 337-341.
DAVIES, R.H., C. WRAY (1996): Persistence of Salmonella enteritidis in poultry units and poultry food. Br. Poult. Sci. 37, 589-596.
De REU, K., K. GRIJSPEERDT, W. MESSENS, M. HEYNDRICKW, M.UYTTENDAELE, J. DEBEVERE, L. HERMAN (2006): Eggshell factors influencing eggshell penetration and whole egg contamination by different bacteria, including Salmonella enteritidis. Int. J. Food Microbiol. 112, 253-260.
De REU, K., W. MESSENS, M. HEYNDRICKW, T. B. RO-DENBURG, M. UYTTENDAELE, L. HERMAN (2008): Bacterial contamination of table eggs and the influence of housing system. World Poult. Sci. J. 64, 5-19.
EFSA (2012): The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2010. EFSA Journal, 10 (3): 2597:442.
GRADEL, K.O., L. RANDALL, A. SAYERS, R. H. DAVIES (2005): Possible associations between Salmonella persis-tence in poultry houses and resistance to commonly used disinfectants and a putative role of mar. Vet. Microbiol. 107, 127-138.
GRIMONTH, P. A.D., F.-X. WEILL (2007): Antigenic formulae of the Salmonella serovars.
WHO Collaborating Centre for Reference and Research on Salmonella. Institut Pasteur, 1-166.
HAFEZ, H. M. (2005): Governmental regulations and concept behind eradication and control of some important poultry diseases. World Poult. Sci. J. 61, 569-581.
HAFEZ, H. M. (2008): European perspective on the control and eradication of some poultry diseases. 1st Mediterranean Summit of WPSA, Porto Carras, Chalkidiki, Greece, 7-10 May 2008, Book of Proceedings, pp 62-72.
HEYNDRICKX, M., D. VANDEKERCHOVE, L. HERMAN, I. ROLLIER, K. GRIJSPEERDT, L. ZUTTER (2002): Routes for Salmonella contamination of poultry meat: epidemio-logical study from hatchery to slaughterhouse. Epidemiol. Infect. 129, 253-265.
IMBERECHTS, H., K. DIERICK (2004): Report on zoonotic agents in Belgium in 2002, pp. 65. Veterinary and Agrochemical Research Centre-Scientific Institute of Public Health.
JONES, F. T. (2011): A review of practical Salmonella control measures in animal feed. J. Appl. Poult. Res.20, 102-113.
KRSTULOVIĆ, F., B. ŠIMPRAGA, M. SOKOLOVIĆ, T. ZGLAVNIK (2007): Bakterije roda Salmonella u kokoši konzumnih jaja. Praxis Veterinaria 55, 41-46.

UČESTALOST BAKTERIJA IZ RODA SALMONELLA SPP. U PERADI VRSTE GALLUS GALLUS U RAZDOBLJU OD 2010. DO 2012. GODINE
55
MARIN, C., S. BALASCH, S. VEGA, M. LAINEZ (2011): Sources of Salmonella contamination during broiler pro-duction in Eastern Spain. Prev. Vet. Med. 98, 39-45.
MARIN, C., A. HERNANDIZ, M. LAINEZ (2009): Biofilm development capacity Salmonella strains isolated in poultry risk factors and their resistance against disinfectants. Poultry Sci. 88, 424-431.
MESSENS, W., K. GRIJSPEERDT, L. HERMAN (2005): Eggshell penetration by Salmonella: a review. World Poult. Sci J 61, 71-85.
NAMATA, H., E. MÉROC, M. AERTS, C. FAES, J. CORI˜NAS-ABRAHANTES, H. IMBERECHTS, K. MINTIENS (2008): Salmonella in Belgian laying hens: an identification of risk factors. Prev. Vet. Med. 83, 323-336.
Naredba o mjerama zaštite životinja od zaraznih i nametničkih bolesti i njihovom financiranju u 2010. godini, NN RH 7/2010.
Naredba o mjerama zaštite životinja od zaraznih i nametničkih bolesti i njihovom financiranju u 2011. godini, NN RH 1/2011.
Naredba o mjerama zaštite životinja od zaraznih i nametničkih bolesti i njihovom financiranju u 2012. godini, NN RH 17/2012
OKAMURA M, Y. KAMIJIMA, T. MIYAMOTO, H. TANI, K. SASAI, E. BABA (2001): Differences among six Salmonella serovars in abilities to colonize reproductive organs and to contaminate eggs in laying hens. Avian Dis. 45, 61-69.
ROJAS, M.J., M. GARCÍA, V. MASDEU (2002): Resultados del análisis microbiológico de yacijas de paja de arroz utilizadas en la avicultura. Revista Cubana de Ciencia Avícola 26, 121-123.
ROSE, N., J. P. MARIANI, P. DROUIN, J. Y. TOUX, V. ROSE, P. COLIN (2003): A decision-support system for Salmonella in broiler-chicken flocks. Prev. Vet. Med. 59, 27-42.
PREVALENCE OF BACTERIA FROM THE GENUS SALMONELLA SPP. IN POULTRY SPECIES GALLUS GALLUS IN THE 2010-2012 PERIOD
Summary At the Croatian Veterinary Institute in Vinkovci, a total of 4886 samples were tested for Salmonella spp. (1559 samples of shoe covers from broiler objects, 1646 samples of laying hen feces, 503 samples of breeding hen feces, 217 samples of table eggs and 961 samples of unhatched eggs) during the 2010-2012 period. Isolation and identification of Salmonella species from eggs and feces was performed by standard methods (HRN EN 6579: 2003; ISO 6579: 2007, Amendment 1: Annex D). Serovar was determined by serotyping with polyvalent and monovalent antisera corresponding to specific Salmonella antigens. The number of Salmonella strains isolated from the samples originating from poultry flocks species Gallus gallus was 81 (5.02%) in 2010, 22 (1.32%) in 2011 and 40 (2.50%) in 2012. The most commonly isolated serovar from all samples was Salmonella (S.) Enteritidis. The S. Enteritidis serovar was isolated from the following samples: 20 shoe covers from broiler objects, 46 samples of laying hen feces and 4 samples of breeding hen feces. The Salmonella Typhimurium serovar was isolated from 2 samples of broiler feces. Among other serovars, most commonly isolated were S. Mbandaka and S. Fillmore, while other serovars were rare. In the study period, a total of 947 flocks of poultry species Gallus gallus were tested and Salmonella was present in 56 of them. The Salmonella Enteritidis serovar was present in 4 samples of broiler flocks, 16 laying hen flocks and 2 breeding hen flocks. The S. Typhimurium serovar was detected in one sample of broiler flock. Key words: Salmonella, broilers, laying hens, breeding hens, eggs, feces

PERADARSKI DANI 2013. 56
EPIDEMIOLOGICAL INVESTIGATION OF CIRCOVIRUS INFECTIONS IN PIGEONS IN SLOVENIA
Uroš Krapež1, Brigita Slavec1, Adela Fratnik Steyer2, Štefan Pintarič3, Martin Dobeic3, Olga Zorman Rojs1, Alenka Dovč1
1Institute of Poultry Health, Veterinary Faculty, University of Ljubljana, Ljubljana, Slovenia 2Institute of Microbiology and Immunology, Faculty of Medicine, University of Ljubljana, Ljubljana, Slovenia 3Institute of Animal and Environmental Hygiene with Ethology, Ljubljana, Slovenia
Summary
Pigeon circovirus (PiCV) infections were investigated in feral and racing pigeons (Columba livia var. domestica) by the real-time polymerase chain reaction (PCR) method. Pigeon circovirus was detected by real-time PCR in cloacal swabs and pharyngeal swabs taken from 74 feral pigeons that were caught at various locations in the city of Ljubljana, Slovenia. PiCV infections were detected in the majority of the tested birds. Cloacal swab and pharyngeal swab samples were also collected from 15 flocks of racing pigeons in Slovenia. Bio-aerosol samples were taken in each of 15 investigated flocks using a Coriolis delta Air sampler. PiCV infections were detected in all 15 flocks. A higher viral load was observed in cloacal swab and pharyngeal swab samples taken from racing pigeons younger than 6 months compared to the values detected in racing pigeons older than 1 year. PiCV was also detected by real-time PCR in bio-aerosol samples obtained from each of the investigated flocks. The results showed a high prevalence of PiCV infections in feral and racing pigeons. Key words: pigeon circovirus, feral pigeons, racing pigeons, real-time PCR, prevalence Introduction Pigeon circovirus (PiCV) has been associated with a range of clinical signs, including lethargy, weight loss, ruffled feathers, respiratory distress, diarrhea, regurgitation, fluid-filled crop and increased mortality in young racing, orna-mental and meat pigeons (Woods et al., 1994; Abadie et al., 2001; Raue et al., 2005; Todd et al., 2006). Circovirus infection of pigeons was first recognized and characterized in the United States in 1990 (Woods et al., 1993). Since then, PiCV has been detected in pigeons from South Africa (Gardens, 1993), Australia, Canada (Woods et al., 1994) and Europe (Smyth and Carroll, 1995; Soike, 1997; Coletti et al., 2000; Abadie et al., 2001). The PiCV genome is a circular, single-stranded DNA, ranging from 2032 to 2040 nucleotides (Todd et al., 2008). Based on genomic similarities, PiCV is classified together with psittacine beak and feather disease virus, porcine circo-viruses types 1 and 2, goose circovirus, canary circovirus, duck circovirus and swan circovirus, as member of the
Circovirus genus in the family Circoviridae (Todd et al., 2005; Carstens, 2010). In the absence of a cell-culture virus propagation system and virus-specific antibodies for detecting virus antigen, several polymerase chain reaction (PCR) methods able to detect small amounts of viral DNA have been developed in recent years (Soike et al., 2001; Todd et al., 2002; Duchatel et al., 2005; Franciosini et al., 2005; Duchatel et al., 2009). Viral DNA has been detected in cloacal swabs, pharynx swabs, trachea, blood, sera, different organs, ovary, testes, semen and embryos by various PCR methods (Hattermann et al., 2002; Todd et al., 2002; Roy et al., 2003; Duchatel et al., 2005; Duchatel et al., 2006; Todd et al., 2006; Duchatel et al., 2009). PiCV infections have been detected in young birds with clinical signs (Soike et al., 2001; Todd et al., 2006; Duchatel et al., 2009) and in apparently healthy young birds (Duchatel et al., 2006; Todd et al., 2006; Duchatel et al., 2009) as well as in apparently healthy older birds, aged from 1 to 9 years (Duchatel et al., 2005; Duchatel et al., 2006; Duchatel et al., 2009).
Uroš Krapež, Institute of Poultry Health, Veterinary Faculty, Cesta v Mestni log 47, 1115 Ljubljana, Slovenia; tel.: +386 1 4779 247; fax. +386 1 4779 339; e-mail address: [email protected]

EPIDEMIOLOGICAL INVESTIGATION OF CIRCOVIRUS INFECTIONS IN PIGEONS IN SLOVENIA
57
Although much knowledge has been gathered about the prevalence of the infection and its relevance to pigeon health in racing, ornamental and meat pigeons, little is known about the prevalence of infection in feral pigeons. To our knowledge, only Hattermann (Hattermann et al., 2002) have tested 9 blood samples from feral pigeons, which were negative for viral DNA by the PCR method and one genome sequence of PiCV from a feral pigeon has been published in GenBank (accession no. DQ915962). The purpose of the present study was to investigate the prevalence of PiCV in a population of feral pigeons in Ljubljana and in a population of racing pigeons in Slovenia. Cloacal swabs and pharyngeal swabs taken from feral and racing pigeons and bio-aerosol samples collected in pigeon houses were investigated for the presence of viral DNA by real-time PCR.
Materials and Methods
Sampling methods
Seventy-four apparently healthy feral pigeons (Columba livia var. domestica) were caught by traps at six different lo-cations in Ljubljana city as part of monitoring of the popu-lation of feral pigeons. Eleven out of the 74 pigeons were younger than 1 year. The birds were ringed and released after sampling. None of the birds was harmed during the procedure. Air distances between sampling sites ranged from 1140 m to 4570 m, with an average distance of 2670 m. Based on the knowledge gathered from monitoring and ringing of feral pigeons in the city of Ljubljana for several years, it is believed that the captured pigeons belong to six flocks, corresponding to the locations, although some mixing of pigeons between flocks cannot be excluded. Pharyngeal swabs and cloacal swabs were taken from each bird individually. Cloacal swabs and pharyngeal swabs were stored at -70 oC until DNA isolation. Pharyngeal swabs and cloacal swabs were taken from pigeons of 15 racing pigeon flocks in Slovenia. Five pigeons younger than 6 months and 5 pigeons older than 1 year per each flock were individually sampled. Bio-aerosol samples were taken in each of 15 investigated flocks using a Coriolis delta Air sampler (Coriolis, France). Bio-aerosol sampling was performed as described by Jereb et al. (2010). Two bio-aerosol samples per flock were taken. One sample was taken in the compartment of the pigeon house where the young pigeons (younger than 6 months that have not been racing yet) were kept and the second sample was taken in the compartment of the pigeon house where older racing birds were kept. Samples of the cloacal swabs, pharyngeal swabs and bio-aerosol samples were stored at -70 oC until DNA isolation.
DNA extraction and real-time PCR
Prior to DNA isolation, 2 mL of sterile phosphate-buffered saline was added to cloacal and pharyngeal swabs taken from feral pigeons. DNA was extracted from each sample.
Cloacal and pharyngeal swabs taken from racing pigeons were pooled. Pools were formed, in 2 mL of sterile phosphate-buffered saline, from five samples of pharyngeal or cloacal swabs taken from pigeons younger than 6 months and from pharyngeal or cloacal swabs taken from pigeons older than 1 year. Pigeons were from the same flock. As a result, two samples (one from young and one from old pigeons) of pharyngeal and two samples (one from young and one from old pigeons) of cloacal swabs per flock were obtained. DNA was extracted by QIAamp DNA Mini Kit (Qiagen, Washington, USA). The input volume for all types of samples was 200 µL. Extracted DNA samples were eluted in 100 µL buffer. Detection of viral DNA in pharyngeal swabs and cloacal swabs from 74 feral pigeons as well as in pooled pharyngeal swab samples, pooled cloacal swab samples and in bio-aerosol samples from racing pigeons was performed by SYBRGreen real-time PCR developed by Duchatel et al. (2009). Briefly, a region of 139 bp of PiCV genome was amplified by oligonucleotide primer YP09 5’-GGT ACC CGC ATA AGG TGC CCG T-3’ and primer YP10 5’-TTG ATC CGC CGG AAGAGC GCC T-3’. PCR reactions were performed with the following program: denaturation step at 95 °C for 15 min, followed by 45 cycles of denaturation at 95 °C for 30 sec, annealing at 64.3 °C for 30 sec and extension at 72 °C for 30 sec. MaximaTMSYBR Green/Rox APCR Master Mix (2X) (Fermentas, Vilnius, Lithuania) was used for amplification reactions.
Results and Discussion
The real-time PCR test showed that 55 of 74 (74.3%) cloacal swabs and 40 of 74 (54.1%) pharyngeal swabs from feral pigeons of Ljubljana, Slovenia, were positive for PiCV (Table 1). The detection rate of viral DNA in cloacal swabs (74.3%) and pharyngeal swabs (54.1%) taken from feral pigeons in the city of Ljubljana was higher than the detection rate reported by Duchatel et al. (2006) for cloacal swabs (15%) and pharyngeal swabs (25%) of adult racing pigeons. The results of investigation in racing pigeons in Slovenia showed that PiCV infection was detected in all 15 flocks (Table 2). A similar high prevalence (93.7%) of PiCV infection in racing pigeons flocks has also been reported for pigeon flocks in Hungary (Csagola et al., 2011). The results of our study showed that the virus persisted in the majority of feral pigeons as well as in all tested racing pigeon flocks. Based on these results, it could be concluded that the PiCV is widespread both in the population of feral pigeons and in the population of racing pigeons in Slovenia. A higher prevalence of PiCV infection was detected in samples from 11 feral pigeons younger than 1 year, where all birds were positive for PiCV in pharyngeal swab samples and 10 were positive in the cloacal swab samples. Similar results were observed in racing pigeon flocks.

EPIDEMIOLOGICAL INVESTIGATION OF CIRCOVIRUS INFECTIONS IN PIGEONS IN SLOVENIA 58
Table 1. Detection of pigeon circovirus (PiCV) in cloacal swabs and pharyngeal swabs taken from feral pigeons by real-time PCR with average threshold cycle (Ct) and 95% confidential interval (CI) calculated for positive samples
Cloaca Pharynx
Location No. of pigeons
No. of young pigeons PiCV positive Average Ct 95% CI PiCV positive Average Ct 95% CI
A 20 7 (35%) 16 (80%) 27.02 ± 5.05 24.55-29.49 11 (55 %) 24.28 ± 6.87 20.22–28.34 B 34 2 (5.8%) 25 (73.5%) 31.53 ± 2.17 30.68-32.38 19 (55.9%) 32.29 ± 3.46 30.73-33.85 C 12 1 (8.3%) 10 (83.3%) 32.46 ± 1.19 31.72-33.2 4 (33%) 30.1 ± 4.12 26.06-34.14 D 3 1 (33.3%) 1 (33.3%) N.D.A N.D. 3 (100%) 29.46 ± 5.49 23.25-35.67 E 3 0 (0%) 3 (100%) 31.66 ± 1.38 30.1-33.22 2 (66.6%) 29.9 ± 1.87 27.31-32.49 F 2 0 (0%) 0 (0%) N.D. N.D. 1 (50%) N.D. N.D. TOTAL 74 11 (14.8%) 55 (74.3%) 30.38 ± 3.96 29.33–31.43 40 (54.1%) 29.52 ± 5.68 27.76–31.28 AN/D = Not done.
Table 2. Detection of pigeon circovirus (PiCV) in cloacal swabs and pharyngeal swabs taken from racing pigeons and in bio-aerosol samples collected in pigeon houses by real-time PCR with average threshold cycle (Ct) and 95% confidential interval (CI) calculated for positive samples
Type of sample PiCV positive Average Ct 95% CI Clocal swab (<6mB) 15 (100%) 20.64 ± 5.65 17.79 – 23.49 Clocal swab (>1yC) 13 (86.6% 31.2 ± 5.02 28.66 – 33.74 Pharyngeal swab (<6m) 15 (100%) 21.55 ± 4.17 19.44 – 23.66 Pharyngeal swab (>1y) 14 (93.3%) 30.29 ± 2.58 27.47 – 33.11 Bio-aersol (<6m) 15 (100%) 26.97 ± 2.95 25.49 – 28.46 Bio-aersol (>1y) 15 (100%) 27.23 ± 2.48 25,97 – 28.49 B Pigeons younger than 6 months C Pigeons older than 1 year Viral DNA was detected in all tested samples (cloacal and pharyngeal swabs and bio-aerosol samples) taken from pigeons younger than 6 months. In racing pigeons older than 1 year, PiCV was detected in 100% of bio-aerosol samples, 93.3% of pharyngeal swab samples and 86.6% of cloacal swab samples (Table 2). For feral pigeons, positive cloacal swab samples presented a mean cycle threshold (Ct) value of 30.38±3.96 (95% confidence interval (CI) 29.33-31.43)) and positive pharyngeal swab samples presented a mean Ct value of 29.52±5.68 (95% CI 27.76-31.28) (Table 1). Positive cloacal swab samples and pharyngeal swab samples from 11 feral pigeons younger than 1 year showed lower mean Ct values of 24.78±4.75 (95% CI 21.84-27.72) and 24.15±6.41 (95% CI 20.36-27.94), respectively. Similar, although lower mean Ct values were detected for cloacal swabs 20.64±5.65 (95% CI 17.79-23.49) and also for pharyngeal swabs 21.55±4.17 (95% CI 19.44-23.66) of racing pigeons younger than 6 months (Table 2). The mean Ct values for cloacal and pharyngeal swabs of racing pigeons older than one year (Table 2) were similar to those obtained for feral pigeons. The lower Ct values detected in young feral and racing pigeons suggested higher viral
loads in cloacal swabs and pharyngeal swabs compared to the amounts of excreted virus through cloaca and trachea in older feral and racing pigeons. These results are similar to those published by Duchatel et al. (2006) and are also consistent with the proposal of the same authors (Duchatel et al., 2006) that low amount of detected viral DNA in older birds is probably not the result of recent infections, but rather that the virus DNA has persisted from infections that occurred when the pigeons were young. Comparison of the results between feral pigeons caught at the six different locations showed no major deviations (Table 1). Similar to feral pigeons, comparable results were found between 15 investigated racing pigeon flocks, except for flock 2, where young pigeons excreted more virus, and flocks 12 and 13, where young pigeons excreted less virus than young pigeons in other flocks, as well as for flock 6, where older pigeons excreted less virus through cloaca than older pigeons in other flocks. The concentration of virus in the air of pigeon houses as measured by bio-aerosol sampling and detection of the virus by real-time PCR was relatively high. For example, more viral DNA was detected in the air of young and old

EPIDEMIOLOGICAL INVESTIGATION OF CIRCOVIRUS INFECTIONS IN PIGEONS IN SLOVENIA
59
pigeon compartment of pigeon house (mean Ct values of 26.97±2.95 and 27.23±2.48, respectively) than in cloacal and pharyngeal swabs taken from pigeons older than 1 year (Table 2). Detection of rather high concentrations of PiCV in bio-aerosol samples indicates the most common way of the virus transmission that is through inhalation of virus bound to the feather dust, which can be found in large amounts in the air of pigeon houses. Daily exposure to PiCV viral particles in the air of pigeon houses, as it is common between pigeon’s breeders, could have some implications on human health. For example, Bougiouklis hypothesized in a recently published study (Bougiouklis, 2007) that avian circoviruses could be one of the potential triggers of the pigeon breeder’s lung, a common extrinsic allergic alveolitis or hypersensitivity pneumonitis. The results of our study showed a rather high concentration of the virus in the air of pigeon houses. To our knowledge, this is the first report on the presence of PiCV in the air of pigeon houses. Further investigation, concerning human and animal health, should be performed to confirm or abandon the hypothesis on PiCV as one of the potential triggers of pigeon’s breeder lung disease.
Acknowledgments
The authors would like to thank Tea Poljanec and Darja Krelj for technical assistance.
References
ABADIE, J., F. NGUYEN, C. GROIZELEAU, N. AMENNA, B. FERNANDEZ, C. GUEREAUD, L. GUIGAND, P. ROBART, B. LEFEBVRE, M.WYERS (2001): Pigeon circovirus infection: pathological observations and suggested pathogenesis. Avian Pathol. 30, 149-158.
BOUGIOUKLIS, P. A. (2007): Avian circoviruses of the genus Circovirus: a potential trigger in pigeon breeder's lung (PBL)/bird fancier's lung (BFL). Med. Hypotheses 682, 320-323.
CADAR D., A. CSÁGOLA, M. LORINCZ, K. TOMBÁCZ, M. SPÎNU, T. TUBOLY (2011). Genetic diversity of pigeon circovirus in Hungary. Arch.Virol. 156, 2233-2239.
CARSTENS, E. B. (2010): Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2009). Arch. Virol. 155, 133-146.
COLETTI, M., M. P. FRANCIOSINI, G. ASDRUBALI, F. PASSAMONTI (2000): Atrophy of the primary lymphoid organs of meat pigeons in Italy associated with circovirus like particles in the bursa of Fabricius. Avian Dis. 44, 454-459.
DUCHATEL, J. P., D. TODD, A. CURRY, J. A. SMYTH, J. C. BUSTIN, H. VINDEVOGEL (2005): New data on the transmission of pigeon circovirus. Vet. Rec. 157, 413-415.
DUCHATEL, J. P., D. TODD, J. A. SMYTH, J. C. BUSTIN, H. VINDEVOGEL (2006): Observations on detection, excretion and transmission of pigeon circovirus in adult, young and embryonic pigeons. Avian Pathol. 35, 30-34.
DUCHATEL, J. P., D. TODD, C. WILLEMAN, B. LOSSON (2009): Quantification of pigeon circovirus in serum, blood, semen and different tissues of naturally infected pigeons using a real-time polymerase chain reaction. Avian Pathol. 38, 143-148.
FRANCIOSINI, M. P., E. FRINGUELLI, O. TARHUNI, G. GUELFI, D. TODD, P. CASAGRANDE PROIETTI, N. FALOCCI, G. ASDRUBALI (2005): Development of a polymerase chain reaction-based in vivo method in the diagnosis of subclinical pigeon circovirus infection. Avian Dis. 49, 340-343.
GARDENS, G. H. (1993): Two very small viruses – a pres-umptive identification. J. S. Afr. Vet. Assoc. 64, 2.
HATTERMANN, K., D. SOIKE, C. GRUND, A. MANKERTZ (2002): A method to diagnose pigeon circovirus infection in vivo. J. Virol. Methods 104, 55-58.
JEREB, G., Š. PINTARIČ, J. MICUNOVIČ, U. KRAPEŽ, B. SLAVEC, B. POLJŠAK, A. DOVČ (2010). Air borne microorganisms in pigeons colonized attics. International Aerosol Conference, August 29-September 3, Helsinki, Finland.
RAUE, R., V. SCHMIDT, M. FREICK, B. REINHARDT, R. JOHNE, L. KAMPHAUSEN, E. F. KALETA, H. MÜLLER, M. E. KRAUTWALD-JUNGHANNS (2005): A disease complex associated with pigeon circovirus infection, young pigeon disease syndrome. Avian Pathol. 34, 418-425.
ROY, P., A. S. DHILLON, L. LAUERMAN, H. L. SHIVAPRASAD (2003): Detection of pigeon circovirus by polymerase chain reaction. Avian Dis. 47, 218-222.
SMYTH, J. A., B. P. CARROLL (1995): Circovirus infection in European racing pigeons. Vet. Rec. 136, 173-174.
SOIKE, D. (1997): Circovirus infections in pigeons. Tierarztl. Prax. 25, 52-54.
SOIKE, D., K. HATTERMANN, K. ALBRECHT, J. SEGALES, M. DOMINGO, C. SCHMITT, A. MANKERTZ (2001): A diagnostic study on columbid circovirus infection. Avian Pathol. 30, 605-611.
TODD, D., J. P. DUCHATEL, J. H. WESTON, N. W. BALL, B. J. BORGHMANS, D. A. MOFFETT, J. A. SMYTH (2002): Evaluation of polymerase chain reaction and dot blot hybridisation tests in the diagnosis of pigeon circovirus infections. Vet. Microbiol. 89, 1-16.
TODD, D., J. P. DUCHATEL, J. C. BUSTIN, F. T. SCULLION, M. G. SCULLION, A. N. SCOTT, A. CURRY, N. W. BALL, J. A. SMYTH (2006): Detection of pigeon circovirus in cloacal swabs: implications for diagnosis, epidemiology and control. Vet. Rec. 159, 314-317.
TODD, D., M. BENDINELLI, P. BIAGINI, S. HINO, A. MANKERTZ, S. MISHIRO, C. NEIL, H. OKAMOTO, S. RAIDAL, B. W. RITCHIE, G.C. TEO (2005): Circoviridae. In: Virus Taxonomy, VIIIth Report of the International Committee for the Taxonomy of Viruses. C. M. Fauquet, M. A. Mayo, J. Maniloff, U. Desselberger, L. A. Ball, eds. Elsevier/Acadenic Press, London. pp. 327-334.
TODD, D., E. FRINGUELLI, A. N. SCOTT, B. J. BORGH-MANS, J. P. DUCHATEL, H. L. SHIVAPRASAD, S. R. RAIDAL, J. X. ABADIE, M. P. FRANCIOSINI, J. A. SMYTH (2008): Sequence comparison of pigeon circo-viruses. Res. Vet. Sci. 84, 311-319.
WOODS, L. W., K. S. LATIMER, B. C. BARR, F. D. NIAGRO, R. P. CAMPAGNOLI, R. W. NORDHAUSEN, A. E. CASTRO (1993): Circovirus-like infection in a pigeon. J. Vet. Diagn. Invest. 5, 609-612.
WOODS, L. W., K. S. LATIMER, F. D. NIAGRO, C. RIDDELL, A. M. CROWLEY, M. L. ANDERSON, B. M. DAFT, J. D. MOORE, R. P. CAMPAGNOLI, R. W. NORDHAUSEN (1994): A retrospective study of circovirus infection in pigeons: nine cases (1986-1993). J. Vet. Diagn. Invest. 6, 156-164.

EPIDEMIOLOGICAL INVESTIGATION OF CIRCOVIRUS INFECTIONS IN PIGEONS IN SLOVENIA 60
EPIDEMIOLOŠKO ISPITIVANJE INFEKCIJE GOLUBOVA CIRKOVIRUSOM U SLOVENIJI Sažetak Infekcije golubinjim cirkovirusom (PiCV) ispitane su kod slobodno živućih i uzgojenih golubova (Columba livia var. domestica) metodom lančane reakcije polimeraze (PCR) u stvarnom vremenu. Golubinji cirkovirus otkriven je ovom metodom u obriscima kloake i obriscima ždrijela uzetih od 74 divlja goluba uhvaćenih na raznim mjestima u gradu Ljubljani, Slovenija. Infekcija PiCV otkrivena je u većine ispitanih ptica. Uzorci obrisaka kloake i ždrijela prikupljeni su i u 15 jata uzgojenih golubova u Sloveniji. Uz to, uzorci bioaerosola prikupljeni su u 15 jata uzgojenih golubova pomoću uređaja Coriolis delta Air sampler. Infekcija PiCV otkrivena je u svih 15 jata. U uzorcima obrisaka kloake i obrisaka ždrijela uzgojenih golubova mlađih od 6 mjeseci utvrđeno je veće virusno opterećenje u usporedbi s vrijednostima zabilježenim kod uzgojenih golubova starijih od 1 godine. PiCV je isto tako otkriven pomoću navedene metode u uzorcima bioaerosola iz svakog od ispitivanjih jata. Rezultati su pokazali visoku učestalost infekcije PiCV kod slobodno živućih i uzgojenih golubova. Ključne riječi: golubinji cirkovirus; slobodno živući golubovi; uzgojeni golubovi; lančana reakcija polimeraze u stvarnom vremenu; učestalost

PERADARSKI DANI 2013.
61
UČINAK DODATKA Ca-BUTIRATA U KRMU KOKOŠI TEŠKIH HIBRIDA NA RELATIVNU GUSTOĆU KOSTIJU – PRELIMINARNO IZVJEŠĆE
Hrvoje Mazija1, Tihomir Dragojlov2, Željka Ervačinović3
1Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 2 Veterinarska ambulanta za kućne ljubimce, Zagreb, Hrvatska 3 “Galis” d.o.o., Slavonski Brod, Hrvatska
Sažetak
Kokoši teških hibrida hranjene su dodatkom pripravka koji je sadržavao 21% inkapsuliranog Ca-butirata (CB) u matriksu tri specifična nosača (E-55, E-327 i E-558) te dodatak Ca-soli jestive masne kiseline (E-470). Kokošima dviju skupina (A i B) je od 30. do 51. tjedna života u krmu dodavano 1‰ (1000 g/t) hrane CB, što se u usporedbi s kontrolnim skupinama nesilica (C i D) koje nisu jele spomenuti dodatak općenito povoljno odrazilo na njihovo zdravlje i proizvodna svojstva. Napose je povećana nesivost, masa jaja, kakvoća ljuske jaja i leživost pilića. U dobi od 51 tjedna po četiri kokoši svake skupine je žrtvovano te im je određena gustoća bedrene i podkoljeničnih kostiju (femur i tibija). Primijenjen je postupak denzitometrije pomoću uređaja za kompjutoriziranu tomografiju (CT), a rezultati su statistički izračunati. Pokazalo se da dodatak 1‰ CB u krmu nesilica značajno povoljno djeluje na gustoću kostiju, što je potvrđeno CT postupkom. S obzirom na postignuti uspjeh istraživanja očekuje se i povoljan utjecaj CB na tamno crvenu diskoloraciju kostiju i okolnog tkiva u tovnih pilića, opisanu kao sindrom crnih kostiju.
Ključne riječi: kokoši nesilice konzumnih jaja, bedrena kost, podlaktična kosti, gustoća kostiju
Uvod
Intenzivna proizvodnja peradi, napose tov pilića, usmjereni su prema poboljšanju proizvodnih svojstava, prvenstveno na rast mišićne mase uz istodobno optimalno iskorišćivanje hrane. Ubrzanje rasta i poboljšana iskoristivost hrane doveli su do pojave niza bolesti koštanog sustava koji se morao prilagoditi većem opterećenju i potrebi bržeg sa-zrijevanja koštanog tkiva. Genetskim odabirom u većine hibridnih linija pilića poboljšana je čvrstoća kostiju, ali je istodobno poboljšan i pristup očuvanju zdravlja lokomo-tornog sustava pilića uvođenjem kvalitetnijih tehnoloških rješenja uzgoja. Ta je genetska osnova nužno zahtijevala drukčiji pristup u hranidbi peradi. Ipak, sve veća tjelesna masa postizana u sve kraćem razdoblju ima za posljedicu nove zdravstvene poteškoće vezane uz lokomotorni sustav. Kosti rastu u dva smjera, uzdužno kod dugih kostiju pro-cesom enhondralnog rasta koji se temelji na epifizealnom rastu gdje se hondrociti diferenciraju u proliferativne hon-drocite. Razmnožavanjem ovih stanica nastaju plosnate stanice koje se diferenciraju i hipertrofiraju, zatim izdužuju te stanu lučiti novu komponentu koštanog matriksa, ko-lagen tipa X (Howlet, 1980., Almedia Paz i sur., 2004.).
Tvorba kostiju odvija se u donjoj hipertrofičnoj zoni izgradnjom mreže trabekula. Produženjem kosti mreža trabekula postupno se resorbira te ustupa mjesto koštanoj srži. U relativno složenom procesu u šupljini kosti osteo-klasti resorbiraju kost proširujući tako, uz istodobni rast, tvorbu kosti s vanjske strane. U ranoj fazi rasta ovaj se proces odvija brzo pa se šupljine kosti ne ispunjavaju u cijelosti koštanom masom prije početka endostealne re-sorpcije, ali s usporenjem rasta stupanj tog ispunjenja se povećava. Zbog toga kost postaje porozna, a postoje i dokazi da ta poroznost raste u većine današnjih hibrida tovnih pilića (Julian, 1998.). Brojni su poremećaji u oblikovanju kostiju, od kojih je najrasprostranjeniji nekroza glave femura. Radi se zapravo o osteomijelitisu povezanom s bakterijskom infekcijom uglavnom bakterijom Staphylococcus aureus ili rjeđe Escherichia coli. Ove se bakterije zadrže u krvnim žilama ploče rasta kosti, osobito femura, gdje je opskrba krvlju slaba, ali se isto tako može raditi i o hranidbenom poblemu ako se ove bakterije zaustave u oštećenom hrskavičnom tkivu kod rahitisa ili tibijalne diskondroplazije (Reece, 1992.). Također je moguć i rahitis kao posljedica nedo-statka kalcija, fosfora ili D vitamina u mladih ptica (Dinev,
Hrvoje Mazija, profesor emeritus, Veterinarski fakultet Sveučilišta u Zagrebu, Heinzelova 55, 10000 Zagreb, Hrvatska; e-mail: [email protected]

UČINAK DODATKA Ca-BUTIRATA U KRMU KOKOŠI TEŠKIH HIBRIDA NA RELATIVNU GUSTOĆU KOSTIJU – PRELIMINARNO IZVJEŠĆE
62
2011.). Napose je to učestala pojava zbog neravnoteže kalcija i fosfora, odnosno zbog malapsorpcije, infekcije koja općenito remeti apsorpciju nutrijenata (Rath i sur., 2000.). Tibijalna diskondroplazija obilježena je nakupljanjem ili predhipertrofijom hondrocita i zbog toga zadebljanjem sloja hrskavice između proliferativne i hipertrofične zone (Edwards, 1984.). Bolest je opisana u brzo rastućih pilića, pataka i purana svugdje u svijetu. Radi se o narušavanju procesa diferencijacije kojim se proliferativni hondrociti mijenjaju u hipertrofične. Zona rasta hrskavice akumulira se u području tibiotarzisa, međutim, ne postiže očekivanu veličinu u hipertrofičnoj zoni te podliježe prijevremenoj nekrozi (Orth i Cook, 1994.). Ipak, mehanizam ove bolesti nije u cijelosti razjašnjen (Leach i Monsonego-Ornan, 2007.). Osobito je značajna pojava tzv. sindroma crnih kostiju (SCK) u tovnih pilića i purića. Radi se o tamnjenju kostiju nogu i okolnog mišićnog tkiva, naročito nakon kuhanja. Pojava je učestalo opisivana posljednjih godina te značajno proširenija negoli se vjerovalo. Zbog toga ima štetne posljedice kako na kakvoću mesa utovljenih pilića tako i na dobrobit životinja. Po svemu sudeći, SCK je povezan s cijeđenjem krvi iz koštane srži kroz strukturu kosti, što se osobito zapaža u pilića već na liniji klanja, a zahvaćeni su femur i tibija. Krv se cijedi na proksimalnom dijelu tibije, neposredno ispod ploče rasta, te difundira uz kost ispod periosta. Smrzavanjem se istiskuje još više krvi iz koštane srži te ona ulazi u okolno tkivo (Lyon i Lyon, 2002.). Također i kuhanjem se ovaj proces naglašava, a tamna boja mesa odaje odbojan izgled konzumentima. Istražujući po-javu SCK u kuhanim proizvodima od pilećeg mesa uzetih s polica veletrgovina, Smith i Northeut (2003.) su našli 11% uzoraka na koje su kupci imali primjedbe ili ih odbili zbog diskoloracije. Diskoloracija pilećeg mesa smatra se po-greškom u kakvoći i dovodi do uklanjanja s linije klanja. U smrznutim proizvodima SCK čini se nije povezan s tipom proizvoda. Histološka pretraga dokazuje općenito vrlo tanko područje mineralizacije i zato slabo međusobno povezivanje struk-tura kosti (Whitehead i Flaming, 2009.). Očevidno postoje putevi u toj strukturi kojima otječe krv te se nakuplja ispod periostealnog sloja. Načini na koje se nastoji ublažiti SCK povezani su s po-boljšanom hranidbom, zapravo pokušajima da se ubrza mi-neralizacija kostiju i tako ojača njihova struktura, napose gustoća cjevastih kostiju nogu. Jedan od postupaka je i davanje dodatnih količina vitamina D (25-hydroxyvitamin D). Poboljšanje, smanjeno crnjenje cjevastih kostiju (tibija) statistički je značajno (P=0,043) (Ledwaba i Roberson, 2003., Saunders-Blades i Korver, 2006.). Učinak dodavanja mineralnih pripravaka u krmu tovnih pi-lića radi suzbijanja SCK nije opisan. Zato nalaz povećanja gustoće cjevastih kostiju (femur i tibija) u kokoši kojima je u krmnu smjesu dodavan mineralni pripravak kalcijev butirat (Ca-butirat) zaslužuje pozornost. Kalcijevu butiratu empirijske formule (CH3-CH2-CH2-COO)2Ca pripisuju se brojna korisna svojstva. Sporo se otapa u probavnom sustavu, nakon što lipaza i soli žuči razgrade trigliceridski omotač. Djeluje na metabolizam stanica izazivajući njiho-
vu apoptozu. Djeluje zaštitno na sluznice probavnog susta-va, ima i antibakterijsko djelovanje te inhibira rast E. coli, salmonela, kampilobaktera, klostridija i drugih, dok isto-dobno potiče korisne bakterije. Pokaže li se učinkovitim u suzbijanju SCK mogao bi se koristiti u tovu brzorastućih pilića, a određene pretpostavke upućuju na mogućnost da bi Ca-butirat mogao povoljno djelovati i u sprječavanju (ublažavanju) osteoporoze u sisavaca. Više je proizvođača pripravaka koji sadrže Ca-butirat. Komercijalni pripravak Ca-butirat što smo ga mi koristili, prema deklaraciji, inkap-suliran je u matriksu tri nosača i to E-55, E-327 i E-558, te sadrži približno 21,0% aniona butirata. Kalcij je organskog podrijetla, a u korištenom pripravku dolazi u obliku kalci-jeve soli jestive masne kiseline E-470. Pripravak se prepo-ruča kao dodatak krmnoj smjesi za tovne piliće (po toni: 500 g u početnoj, 750 g u završnoj I. i 500-750 g u za-vršnoj II. smjesi), te za kokoši nesilice bilo rasplodnih ili konzumnih jaja (1,0-1.5 kg/tona). Najnovijom uredbom Europske komisije, Food Standard Agency (www.good.gov.uk) od 6. siječnja 2013. zabranjena je upotreba tri aditiva što sadrže aluminij, a među njima je i bentonit (E-558). Prema tome, upotreba pripravka što smo ga mi koristili, u obliku kakvom se danas stavlja na tržište je zabranjeno. Kako se radi tek o jednom od aditiva, biti će ga lako zamijeniti nekim od škriljevaca bez aluminija. Zato ovaj rad i ima smisla.
Materijali i metode Pripravak Ca-biturata je korišten u jatu nesilica hibrida teških linija kokoši u razdoblju od 30. do 51. tjedna života. U odnosu na kontrolnu skupinu nesilica koje nisu dobivale pripravak pokazao je povoljan učinak općenito na pro-izvodna svojstva nesilica. U dobi od 51 tjedna po dvije kokoši kontrolne i pokusne skupine žrtvovane su dislo-kacijom vrata. Noge su odvojene od trupa, skinuta je koža i nakon odsjecanja tarzalne kosti kuhane su u vodi dva sata kako bi se kosti lakše odvojile od mekog tkiva, mišićja i tetiva. Kosti, bedrena i podkoljenična, su zatim isušene pri temperaturi 105 ºC (24 h) te je njihova gustoća određena denzitometrijskim postupkom kompjutorizirane tomogra-fije (CT). U tu svrhu korišten je uređaj MSCT Toshiba Aquilion® (Toshiba Medical, Japan). Dijagnostička medi-cinska radna stanica VEPRO MEDIMAGE® (VEPRO AG, Njemačka) poslužila je za analitičke postupke denzito-metrije i to: - Histogramsku analizu CT podataka, segmentaciju, gra-fičko predstavljanje područja visokih gustoća (VOI LUT), prijetvorba i prikaz područja visokih gustoća te histogram korigiranih vrijednosti s obzirom na razlike u površinama slojeva. - Prihvat 16-bitnih CT podataka pretvaran je u 8-bitni format za naknadnu obradu. - Širina CT sloja iznosila je 1 mm. Uzorci 1 i 2 potječu od kokoši dobi 51 tjedan koje su tijekom 21 tjedna jele u krmu dodani pripravak Ca-butirata, dok su uzorci 3 i 4 kosti kontrolne skupine koja nije dobivala pripravak. Za četiri kontrolne kosti – lijeva i desna bedrena i potko-ljenična kost (ossa femur et tibia) skupina 1 i 4 analizarane su CT snimke.
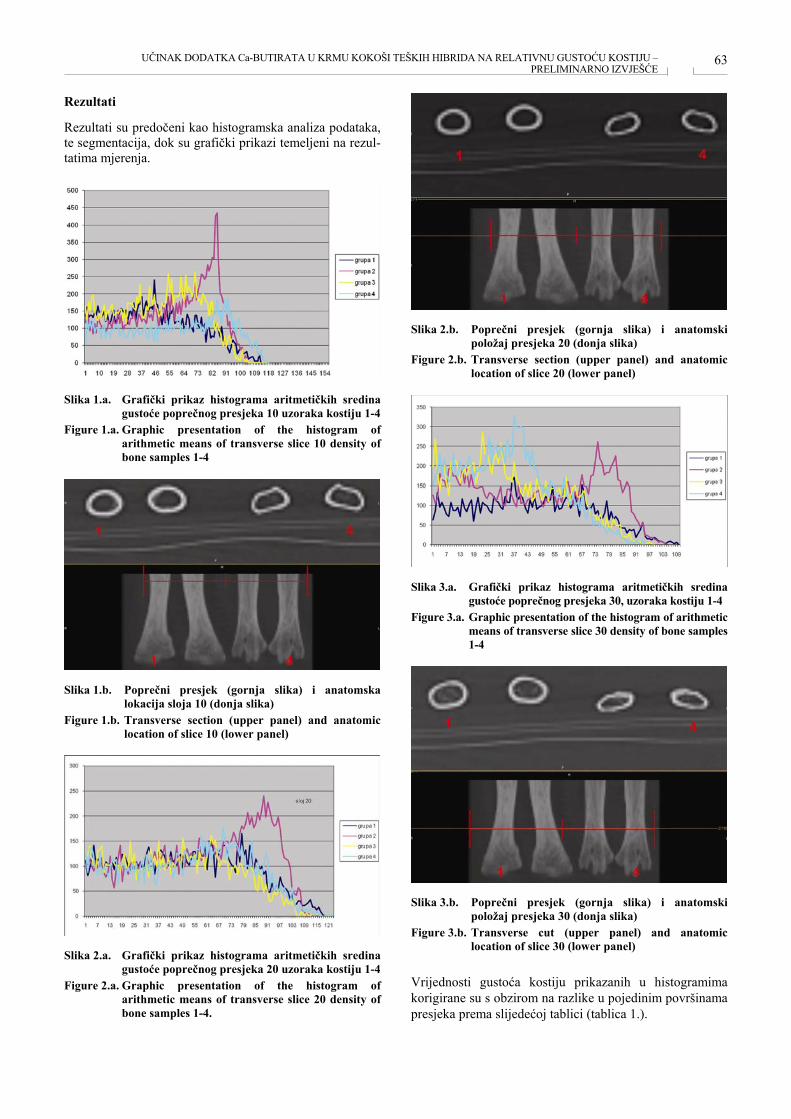
UČINAK DODATKA Ca-BUTIRATA U KRMU KOKOŠI TEŠKIH HIBRIDA NA RELATIVNU GUSTOĆU KOSTIJU – PRELIMINARNO IZVJEŠĆE
63
Rezultati
Rezultati su predočeni kao histogramska analiza podataka, te segmentacija, dok su grafički prikazi temeljeni na rezul-tatima mjerenja.
Slika 1.a. Grafički prikaz histograma aritmetičkih sredina gustoće poprečnog presjeka 10 uzoraka kostiju 1-4
Figure 1.a. Graphic presentation of the histogram of arithmetic means of transverse slice 10 density of bone samples 1-4
Slika 1.b. Poprečni presjek (gornja slika) i anatomska lokacija sloja 10 (donja slika)
Figure 1.b. Transverse section (upper panel) and anatomic location of slice 10 (lower panel)
Slika 2.a. Grafički prikaz histograma aritmetičkih sredina gustoće poprečnog presjeka 20 uzoraka kostiju 1-4
Figure 2.a. Graphic presentation of the histogram of arithmetic means of transverse slice 20 density of bone samples 1-4.
Slika 2.b. Poprečni presjek (gornja slika) i anatomski položaj presjeka 20 (donja slika)
Figure 2.b. Transverse section (upper panel) and anatomic location of slice 20 (lower panel)
Slika 3.a. Grafički prikaz histograma aritmetičkih sredina gustoće poprečnog presjeka 30, uzoraka kostiju 1-4
Figure 3.a. Graphic presentation of the histogram of arithmetic means of transverse slice 30 density of bone samples 1-4
Slika 3.b. Poprečni presjek (gornja slika) i anatomski položaj presjeka 30 (donja slika)
Figure 3.b. Transverse cut (upper panel) and anatomic location of slice 30 (lower panel)
Vrijednosti gustoća kostiju prikazanih u histogramima korigirane su s obzirom na razlike u pojedinim površinama presjeka prema slijedećoj tablici (tablica 1.).

UČINAK DODATKA Ca-BUTIRATA U KRMU KOKOŠI TEŠKIH HIBRIDA NA RELATIVNU GUSTOĆU KOSTIJU – PRELIMINARNO IZVJEŠĆE
64
Tablica 1. Korigirane vrijednosti gustoća bedrene (kosti 1 i 3) i potkoljenične (kosti 2 i 4) kosti pokusne i kontrolne skupine kokoši
Table 1. Corrected values of density of thigh (bones 1 and 3) and drumstick (bones 2 and 4)
g1 g2 g3 g4 Kost 1** Bone 1 908 833 933 926 Kost 2* Bone 2 976 856 950 948 Kost 3** Bone 3 975 880 1009 997 Kost 4* Bone 4 993 888 1009 1022 Ukupno/Total 3852 3457 3901 3893 Faktor korekcije/Factor of correction 103,8% 115,7% 102,5% 102,7%
*dodan Ca-butirat/Ca-butyrate added **bez Ca-butirata/without Ca-butyrate Naglašena je gustoća kostiju kokoši nesilica što su jele Ca-butirat primiješan u krmnu smjesu.
Rasprava i zaključak
Postignuti uspjeh primjene komercijalnog pripravka Ca-butirata u kokoši nesilica hibrida teških linija bio je osnova promišljanju o mogućnosti njegova djelovanja općenito na koštane strukture, bilo domaće peradi ili sisavaca. Kao pre-liminarno istraživanje, zbog sličnosti u procesu sazrijeva-nja kostiju ptica i sisavaca, naročito u sadržaju i tvorbi ko-lagenskih niti i načinu oblikovanja lamelarnog tipa kostiju (Boskey i sur., 1999.), moguć je sličan ili čak istovjetan učinak Ca-butirata u sisavaca. Mogao bi se tako primijeniti kao pomoćno sredstvo u suzbijanju osteoporoze. Pripravak Ca-butirata što smo ga mi koristili značajno je poboljšao gustoću bedrene i potkoljenične kosti kokoši. Međutim, treba uzeti u obzir i trajanje primjene pripravka u nesilica (21 tjedan) te činjenicu da se radilo o nesilicama teških hibrida koje su intenzivno nesle jaja i tu trošile kalcij i fosfor radi izgradnje jajne ljuske. Neosporno, fizio-loški proces nešenja nije bio narušen u nesilica koje su jele dodani Ca-butirat, dok je kod kontrolne skupine bez tog pripravka gustoća kostiju bila manja. Temeljem postignutog uspjeha istraživanja nemoguće je tvrditi da Ca-butirat može spriječiti pojavu sindroma crnih kostiju u pilića, jer je i vrijeme njihova tova u odnosu na 21 tjedan primjene u kokoši neusporedivo kraće. Ostaje otvoreno pitanje bi li kratkotrajnom primjenom u krmi brzorastućih tovnih pilića (35-45 dana) pozitivni učinak na sindrom crnih kostiju bio i ostvaren ili je za to potrebno dulje vrijeme. Postignuti povoljni učinak, poboljšanje proizvodnih svoj-stava kokoši nesilica od kojih je korišten uzorak kostiju ukazuje na općenito pozitivno djelovanje Ca-butirata na zdravlje peradi i osobito na zdravlje koštanog sustava. Zaključno treba navesti da istraživanja valja nastaviti u najmanje dva smjera. Najprije bi trebalo zamijeniti zabra-njeni (Anonimno, 2012.) bentonit nekim škriljcem koji ne sadrži aluminij, primjerice, multikomponentnim holocen-
skim mineralom koji ne sadrži aluminij i za koji je doka-zano da ne djeluje štetno ili uopće ne utječe na kakvoću mesa utovljenih pilića (Medić i sur., 2010.). Istodobno bi trebalo učinak pripravka Ca-butirata istražiti na animalnom modelu sisavaca s induciranom osteoporozom, kakav su primjerice ovariektomizirane ovce (Turner, 2001.).
Literatura
ALMEDIA P.Z, I. C. L., A. A. MENDES, T. S. TAKITA, L. VULKANO, P. C. GUERRA, R. G. GARCIA (2005): Comparison of techniques for tibial dyschondroplasia assessment in broiler chickens. Braz. J. Poult. Sci. 7, 27-32.
Anonimno (2012): Food Standard Agency. Restrictions on the use of additives containing aluminium (www.good.gov.uk).
BOSKEY, A. L., T. M. WRIGHT, R. D. BLANK (1999): Colla-gen and bone strength. J. Bone Miner. Res. 14, 330-335.
DINEV, I. (2011): Comparative pathomorphological study of rickets types in broiler chickens. Iran. J. Vet. Sci. Technol. 3(1), 1-10.
EDWARDS, H. Jr. (1984): Studies on the etiology of tibial dyschondroplasia in chickens. J. Nutr. 114, 1001-1013.
HOWLET, C. R. (1980): The fine structure of the proximal growth plate metaphysis of the avian tibia: endochondral osteogenesis. J. Anat. 130, 745-768.
JULIAN, R. J. (1998): Rapid growth problems: ascites and skeletal deformities in broilers. Poult. Sci. 77, 1773-1780.
LEACH, R. M. Jr., E. MONSONEGO-ORNAN (2007): Tibial dyschondroplasia 40 years later. Poult. Sci. 86, 2053-2058.
LEDWABA, M. F., K. D. ROBERSON (2003): Effectiveness of twenty-five-hydroxycholecalciferol in the prevention of tibial dyschondroplasia in Ross cockorels depends on dietary calcium. Poult. Sci. 82, 1769-1777.
LYON, B. G., C. E. LYON (2002): Color of uncooked and cooked broiler leg quarters associated with chilling temperature and holding time. Poult. Sci. 81, 1916-1920.
MEDIĆ, H., S. VIDAČEK, N. MARUŠIĆ, A. JURIĆ, T. PETRAK, H. MAZIJA (2010): Multicomponent Holocene mineral compound given in feed does not influence quality of chicken meat. XII European Poultry Conference, Tours, Francuska, 23-27.08.2010.

UČINAK DODATKA Ca-BUTIRATA U KRMU KOKOŠI TEŠKIH HIBRIDA NA RELATIVNU GUSTOĆU KOSTIJU – PRELIMINARNO IZVJEŠĆE
65
ORTH, M. W., M. E. COOK (1994): Avian tibial dyschondro-plasia: a morphological and biochemical review of the growth plate lesions and its cause. Vet. Pathol. 31, 403-414.
RATH, N. C., G. R. HUFF, W. E. HUFF, J. M. BALOG (2000): Factors regulating bone maturity and strength in poultry. Poult. Sci. 79, 1024-1032.
REECE, R. L. (1992): The role of in fectious agents in leg abnormalities in growing birds. U Bone Biology and Skeletal Disorders in Poultry. pp. 231-263. C. C. Whitehead, ed. Carfax Publishers, Abingdon, UK.
SAUNDER-BLADES, J., D. KORVER (2006): HyD and Poultry: Bones and Beyond. DSM
Nutritional products Satellite Symposium. XII European Poultry Conference, Verona, 10-14 September 2006.
SMITH, P. D., J. K. NORTHCUT (2003): Red discoloration of fully cooked chicken products. J. Appl. Poult. Res. 12, 515-521.
TURNER, A. S. (2001): Animal models of osteoporosis – necessity and limitations. Eur. Cell Mater. 1,66-81.
WHITEHEAD, C., B. FLAMING (2009): The black bone syndrome in broilers – a challenge for breeders? Int. Hatch. Pract. 23(8), 7-9.
THE EFFECT OF Ca-BUTYRATE SUPPLEMENT ADDED TO FEED OF HEAVY HYBRID BREEDING HENS ON RELATIVE BONE DENSITY – PRELIMINARY REPORT Summary Heavy hybrid breeding hens were fed Ca-butyrate (CB) supplement containing 21% of encapsulated calcium butyrate in a matrix of three specific carriers (E-55, E-327 and E-558), and Ca-salt of edible fatty acid E-470. Hens of two groups were given Ca-butyrate from 21 to 51 weeks of life (1‰ (1000 g/t), which resulted in generally better production yield and health, compared to the control group of hens that did not receive the supplement. In particular, this supplementation increased the number of eggs, egg weight, quality of egg shell, and hatchability. At the age of 51 weeks, 4 hens from each group were sacrificed and bone density of the femur and tibia was determined using computerized tomography (CT) and statistical data processing. It was shown that the addition of 1‰ CB significantly improved bone density. Considering the favorable results achieved, positive influence of CB on the black bone syndrome in chickens described as dark red discoloration of bones and surrounding muscle tissue, can also be expected. Key words: heavy laying hens, femur, tibia, bone density

PERADARSKI DANI 2013. 66
IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
Željko Mišković1, Hrvoje Garay2, Hrvoje Mazija3
1"Posavina koka", Orašje, Bosna i Hercegovina 2Veterinarska stanica Orašje, Orašje, Bosna i Hercegovina 3Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska
Sažetak
Istražena je imunogenost cjepiva protiv Marekove bolesti od stanično pridruženog virusa CVI 988 danog parenteralno intramuskularno odnosno istodobno s herpesvirusom purana HVT-FC-126 danog postupkom nebulizacije netom izleženim pilenkama. Tijekom dvogodišnjeg razdoblja cjepivo CVI 988 dano je ukupno 243.122 pilenki u 40 jata, a 307.955 pilenki u 50 jata cijepljeno je istodobno postupkom zamagljivanja (HVT-FC-126) i parenteralno intramuskularno (CVI 988). Bez obzira na način davanja postupci cijepljenja nisu izazvali štetnih kliničkih reakcija. Tijekom uzgojnog razdoblja od 18 tjedana od Marekove bolesti uginulo je 4,98% pilenki cijepljenih jedino sojem CVI 988, te njih 0,23% u jatima cijepljenim istodobno spomenutim načinima s oba soja virusa. Nove spoznaje o značenju ranog postizanja specifične otpornosti protiv Marekove bolesti kakva se ostvaruje postupkom nebulizacije aktualiziraju primjenu cjepiva od soja HVT-FC-126 zamagljivanjem. Ključne riječi: Marekova bolest, cijepljenje, soj HVT-FC-126, nebulizacija, soj CVI 988, kokoši nesilice Uvod Marekova bolest (MB) najznačajnija je neoplastična zaraza kokoši. Uzročnik, stanično pridruženi (Calnek i Hitchner, 1969.) alfa-herpesvirus (Buckmaster i sur., 1988.) pripada serotipu 1 te je srodan neonkogenim herpesvirusima kokoši serotipa 2 i herpesvirusu purana označenog kao herpesvirus 3. S obzirom na patotip sojevi virusa MB (VMB) svrstani su bilo u blage (medium, m), virulentne (virulent, v), vrlo viru-lentne (very virulent, vv) ili u novije vrijeme vrlo virulentne + (very virulent plus, vv+) sojeve (Witter, 1998.). Gospo-darske štete koje je MB izazivala rasle su sukladno razvoju peradarstva, a temeljene na izazvanom uginuću i smanjenoj nesivosti u kokoši, te uginuću i klaoničkom odbacivanju morfološki promijenjenih trupova utovljenih pilića. Samo zbog uginuća peradi u SAD 1984. godine neposredne štete iznosile su 12 milijuna USD, s ostalim štetama i cijenom cijepljenja ukupno 169 milijuna USD, dok je u svijetu ukupna šteta iznosila 943 milijuna USD. Danas štete od MB
u svijetu iznose između 1 i 2 milijarde USD (Morrow i Fehler, 2004.). Porast virulencije VMB u proteklih 60-ak godina je vrlo značajan, najprije zbog pojave vv VMB 1987.-ih godina, a zatim i vv+VMB ranih 1990.-ih godina (Witter, 1998.). Anketni upitnik kojim se nastojalo saznati koja je proširenost MB u SAD i u svijetu (Dunn i Gimeno, 2013.) distribuiran je u 116 zemalja svijeta, a obuhvatio je članice WVPA, proizvodne tvrtke u peradarsvu, znanstvene institucije te proizvođače specifičnih cjepiva. U gotovo 50% zemalja porasla je učestalost MB u proteklih 10 godina, navodeći kao glavni razlog tome slabljenje imunog sustava ili u manjoj mjeri (16% ispitanika) povećanu virulentnost VMB. Opsežna primjena cjepiva od soja Rispens razlogom je što je učestalost MB opala u 42% anketiranih zemalja. Prva cjepiva protiv MB izrađena su svega nekoliko godina nakon što je dokazana njena etiologija (Churchill i sur., 1969., Okazaki i sur., 1970., Rispens i sur., 1972.). Cjepivo od herpesvirusa purana (serotip 3) (HVT) koji su prvi izdvojili Kawamura i sur. (1969.) izradili su Okazaki i sur.
Željko Mišković, "Posavina koka", Ulica XIV 74, 76270 Orašje, Bosna i Hercegovina; telefon 0038731712368; e-mail: [email protected]

IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
67
(1970.). Za razliku od cjepnih sojeva VMB koje je, kao stanično pridružene, nemoguće liofilizirati, soj HVT-FC-126 dolazi na tržište liofiliziran, što je bila osnova njegove ma-sovne primjene. Ipak, prednost se daje stanično pridruženim cjepivima (Witter, 1987.) zbog solidnijeg imuniteta koji po-tiču, odnosno kombinaciji oba tipa cjepnih sojeva (stanično pridruženih i HVT-FC-126). Uz sojeve od atenuiranih VMB serotipa 1 (CVI 988 Rispens) kao cjepivo koriste se i pri-rodno apatogeni sojevi VMB serotipa 2 (SB1), a u slučaju pojave vv+VMB često i kombinacija sva tri soja (Anon., 2010.). Način primjene specifičnih cjepiva protiv MB je gotovo isključivo parenteralni, bilo subkutani, intramuskularni ili in ovo kokošjim zamecima u dobi od 18 dana (Johnston i sur., 1997.). Već g. 1995. u SAD cijepljeno je in ovo gotovo 55% kokoši (tovni pilići i buduće nesilice), odnosno godišnje preko 5 milijardi rasplodnih jaja (Johnston i sur., 1997.). Koristilo se je dvovaljano cjepivo, kombinacija sojeva HVT i SB-1 (Sarma i sur., 1995.). Danas se protiv MB cijepi gotovo 100% tovnih pilića u SAD i značajan broj pilića u europskim zemljama. Osobito je zanimljiv rezultat istraživanja brzine nastupa vi-remije u kokošjih zametaka cijepljenih in ovo 17. dana inku-biranja virusom CVI988 (Zhang i Sharma, 2001.). Dokazali su da viremija nastupa 2-3 dana nakon leženja pilića, znatno prije negoli kod parenteralno cijepljenih pilića neposredno nakon leženja te da su bolje od njih specifično zaštićeni. Prvi pokušaj raspršivanja cjepiva protiv MB od HVT u obli-ku spreja datira iz 1975. godine (Kleven i Edison, 1975.). Uspjeh je bio tek djelomičan, jer je za postizanje uspjeha trebala enormna količina cjepiva, pa postupak nije prihvaćen u praksi. Vrlo sitnu česticu suspenzije cjepiva od soja HVT-FC-126 pomoću ultrazvučnog raspršivača tovnim su pilići-ma primijenili Pridybailo i sur. (1986.), no korišteno cjepivo bilo je neprikladno za nebulizaciju pa niti ovaj pokušaj nije uspio. Pravi uspjeh primjene sitne čestice aerosola promjera 3-5 mikrona uslijedio je tek nakon što su Čajavec i sur. (1992.) izradili liofiliziorano cjepivo od soja herpesvirusa purana (MARIKAL® HVP-FC-126).) koje se je moglo radi primje-ne otopiti u običnoj destiliranoj vodi. Iako namijenjeno za parenteralnu primjenu, to su cjepivo Mazija i Čajavec (1993.) i Mazija i sur. (1994.), koristivši ultrazvučni raspršivač (Mazija i Štimac, 1993.) dali uspješno netom izle-ženim pilićima te dokazali virusni antigen u dišnom sustavu cijepljenih pilića. Stančić (1996.) je cijepila tovne piliće pro-tiv MB postupkom nebulizacije primjenom cjepiva MARI-KAL® HVP-FC-126 i dokazala neštetnost postupka, a Kokić (1998.) je postigla izniman uspjeh u uzgoju pilenki budućih nesilica konzumnih jaja. Viremija izazvana cjepivom od soja HVT- FC-126 (TAD MAREK® VAC LYO) danim netom izleženim pilićima postupkom nebulizacije nastupila je već nakon 48 sati i u odnosu na parenteralno dano cjepivo bila dvostruko odnosno čak trostruko veća šestog te dvanaestog dana (Voss i Mazija, 2000. neobjavljeno). Zbog sve slabijeg učinka cjepiva od soja FC 126 ono je postupno zamijenjeno kombinacijom sa
stanično pridruženim cjepnim virusima (HVT-FC-126 + Rispens CVI 9898 ili Sb1), najčešće u odnosu 3 + 1 te se da-nas takva uglavnom koriste svugdje u svijetu. Razlike u učestalosti pojave MB između SAD gdje bolest stagnira proteklih godina s najnižom učestalosti g. 2007. (0,0008% uginuća) i zemalja Sjeverne Afrike, Istočne Euro-pe, Istočne Azije i Južne Amerike, gdje je MB u porastu Dunn i Gimeno (2013.) pripisuju doduše imunosupresivnim bolestima i povećanoj patogenosti VMB kojim je izazvana, ali i primjeni nespecifičnih mjera preventive koje su u SAD značajno bolje negoli u drugim zemljama. Dunn i sur. (2010.) dokazali su da izazivačka infekcija patogenim VMB nakon koje ubrzo (za 24 sata) slijedi superinfekcija VMB ima daleko pogubniji učinak negoli superinfekcija nakon 13 dana (patogeni virus dokazan u 55% prema 5% pilića). Ovime je samo potvrđena davna spoznaja da rano izlaganje patogenom VMB već cijepljenih pilića u cijelosti obez-vrjeđuje učinak danog cjepiva (Zanella, 1982.), odnosno da je stupanj specifične zaštite potaknut cijepljenjem to slabiji što je razmak do izlaganja patogenom virusu kraći (Okazaki i sur., 1970.). Viremiju od HVT kao mjeru osjetljivosti prema patogenom VMB istražili su Cho i sur. (1976.) i našli da je specifična zaštita to jača što je viremija izraženija. Istraživanja brzine nastupa viremije od HVT u pilića nakon davanja specifičnog cjepiva postupkom nebulizacije (Mazija i Voss, 2000., neobjavljeno) potakla su istraživanja distribu-cije u tkivima DNK genoma HVT danog bilo postupkom nebulizacije (Mazija i Čajavec, 1993.) ili usporedno pot-kožno (Gottstein i sur., 2007.). U uzorcima tkiva pluća genom HVT dokazan je nakon 24, 48 odnosno 96 sati, dok su uzorci pluća pilića cijepljenih potkožno istovjetnim viru-som bili negativni. Gimeno i sur. (2008.) ustvrdili su da viremija s HVT nije u korelaciji sa specifičnom zaštitom, no Holland i Silva (1999.) su dokazali da je tkivo slezene, a ne periferna krv mjesto nakupljanja herpesvirusa purana. Zbog navedenog razloga, a i zbog nastojanja da se speci-fična refraktornost prema ranoj infekciji patogenim sojevima VMB što je moguće prije postigne istražen je imuni učinak cjepnog soja VMB, CVI 988, danog parenteralno intramu-skularno bilo samostalno ili u kombinaciji sa sojem HVT- -FC-126 danim postupkom nebulizacije. U tom smislu ovaj rad predstavlja dopunjenu interpretaciju istraživanja što ih opisuje Mišković (2004.) i aktualnih znanstvenih spoznaja o imunoprofilaksi Marekove bolesti.
Materijal i metode
Pokusna perad
Anamnestički podaci Istraživanje je provedeno u području izrazito opsežne pro-izvodnje kokoši, jednako tovnih pilića i nesilica konzumnih jaja. Zbog male međusobne udaljenosti nastambi s peradi učestale su naročito virusne zaraze koje je bilo nemoguće spriječiti uobičajenim postupcima specifične imunoprofi-lakse. Odnosi se to osobito na zaraznu bolest burze (Pav-lović, 1998.) te Marekovu bolest (Mišković, 2004., neob-javljeno). Prema epizootiološkim podacima MB je u pilenki,

IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
68
budućih nesilica jaja za konzum (hibrid Lohmann Brown) cijepljenih jedino sojem CVI 988, tijekom uzgojnog raz-doblja od prosječno 18 tjedana izazvala uginuće značajno veće od dopustivog (Mišković, 2002., neobjavljeno). Jata netom izleženih ženskih pilića od vlastitih roditeljskih jata hibrida lakih linija Lohmann Brown ili rjeđe iz uvezenih jaja hibrida Hy Line Brown (Mađarska) odnosno uvezenih rasplodnih jaja iz Njemačke (Lohmann Brown) cijepljena su protiv MB intramuskularno cjepivom od soja CVI 988 (NOBILIS® RISMAVAC) prema preporuci proizvođača. Nakon toga isporučeni su u nastambe za uzgoj podnim
načinom. Nastambe su bile prosječnih dimenzija 30x12 m (360 m2), u manjem broju dvoetažne (720 m2), često vrlo derutne. Za dezinfekciju prostora nakon mehaničkog čišće-nja korištena je otopina Na OH, te pripravci Virkon S i HP-3 metodom zamagljivanja. Uzgojno razdoblje različito je kod pojedinih jata, najmanje 81, a najviše 151 dana ili prosječno 124,37 dana, što je nešto manje od tehnološki poželjnih 18 tjedana. Međuturnusna stanka očito nije planirana, jer je iznosila od najmanje pet ili sedam dana pa sve do osam mjeseci. Tijekom uzgojnog razdoblja pilenke su osim protiv MB cijepljene prema slijedećem programu:
Tablica 1. Program imunoprofilakse* bez uključene Marekove bolesti
Dob prilikom cijepljenja Bolest Korišteno cjepivo (soj virusa) Postupak primjene 9. dan Newcastleska bolest La Sota Pitkom vodom 14. dan Zarazna bolest burze Intermedijarni Pitkom vodom 19. dan Zarazni bronhitis H 120 Pitkom vodom 28. dan Newcastleska bolest La Sota Pitkom vodom 32. dan Zarazna bolest burze Intermedijarni Pitkom vodom 37. dan Zarazni bronhitis H 120 Pitkom vodom 8. tjedan Newcastleska bolest La Sota Pitkom vodom 9. tjedan Boginje peradi Kokošji atenuirani Ubod u krilnu opnu 11.-12. tjedan Zarazni bronhitis H 120 Pitkom vodom 12.-13. tjedan Newcastleska bolest La Sota Pitkom vodom 16.-18. tjedan NB + ZB + EDS NB + ZB + EDS** Intramuskularno
40. dan i 10. tjedan = kontrola imunosti 9. tjedan = skraćivanje kljunova *Opisani program imunoprofilakse ne preporuča se u Hrvatskoj **inaktivirano uljno cjepivo Table 1. Program of immunoprophylaxis* without Marek's disease included
Age at vaccination Disease Vaccine (strain) used Mode of application Day 9 Newcastle disease La Sota Drinking water Day 14 Infectious bursal dis. Intermediate Drinking water Day 19 Infectious bronchitis H 120 Drinking water Day 28 Newcastle disease La Sota Drinking water Day 32 Infectious bursal dis. Intermediate Drinking water Day 37 Infectious bronchitis H 120 Drinking water Week 8 Newcastle disease La Sota Drinking water Week 9 Chicken pox Atenuated chicken Wing web Week 11-12 Infectious bronchitis H 120 Drinking water Week 12-13 Newcastle disease La Sota Drinking water Week 16-18 ND + IB + EDS'76 ND + IB + EDS'76** Intramuscularly
Day 40 and week 10 = immunity control; week 9 = debeaking **Inactivated oil vaccine *This program is not recommended in Croatia

IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
69
Sukladno tehnološkim prilikama i kojiput gotovo izostaloj međuturnusnoj stanci, uginuća pilenki su se tijekom uzgoj-nog razdoblja također kretala u širokom rasponu, od najma-nje 1,02% te povećana do najviše 47,74% bilo zbog pojave zarazne bolesti burze ili ugušenja, odnosno stalno prisutne kokcidioze i oportunističkih bakterijskih bolesti. Marekova bolest početkom opisivanog razdoblja (2001.-2003. godine) nije predstavljala uzgojni problem. Pojavljivala se najprije tek nakon spolne zrelosti kokoši i postupno kompromitirala tvrtku isporučitelja uzgojenih jata. U uzgoju pojavljuje se sredinom 2001. godine u jatima s relativno malim ukupnim uginućem (3,47% i 3,48%) te dosiže udjel od najviše 1,79%. Zatim naglo porasta oscilirajući oko prosječnih 5,85%, no u rasponu od najmanje 0,70% pa do najviših 18,89% (u jatu s ukupnim uginućem 47,74%). Pojava uginuća od MB već u uzgojnom razdoblju neosporno dokazuje ovu zarazu koja puni intenzitet očituje početkom razdoblja nešenja, što je uvjetovano nizom stresora poput cijepljenja, uznemiravanja te aktivacijom hormonskog sustava reprodukcije, na što uka-zuje veća učestalost MB u ženki (Payne i Venugopal, 2000.). Zbog opisane pojave MB koju nije bilo moguće spri-ječiti u opisanoj proizvodnoj praksi klasičnom primjenom stanično pridruženog cjepiva od soja CVI 988 istražen je učinak simultanog davanja cjepiva od soja CVI 988 (NOBI-LIS® RISMAVAC) netom izleženim pilićima parenteralno intramuskularno, a zatim i cjepiva od soja herpesvirusa pura-na FC 126 (MARIKAL ® HVP-FC-126) postupkom nebuli-zacije (Mazija i Čajavec, 1993.). Iako ovakav postupak nije prethodno dovoljno istražen, epizootiološke prilike koje su prijetile potpunim prestankom proizvodnje kokoši nesilica u opisivanom području, te iskustvo s primjenom cjepiva TAD MAREK® VAC LYO (Voss i Mazija, 2000., neobjavljeno) bili su dovoljnim razlogom za njegovu opsežnu primjenu. Ukupno je istraživanjem obuhvaćeno 551.077 pilenki izleže-nih u lokalnoj valionici proizvodne tvrtke koja ih je uzgojila bilo u kooperantskim odnosima ili u vlastitoj proizvodnji. Tijekom razdoblja istraživanja (2001.-2003. godine) bilo je to 90 jata prosječne veličine 6.018 pilenki s najmanje 2.502, a najviše 12.580 komada. Način uzgoja pilenki bio je podni s prosječno 13 pilenki po m2, a tehnološki uzgojni uvjeti
jedino unutar nastambe zadovoljeni. Međusobna udaljenost pojedinih nastambi, međutim, bila je nedovoljna da se spri-ječi širenje uzročnika bolesti zrakom, a razlike u dobi poje-dinih jata nisu omogućile primjenu temeljnog tehnološkog postulata zaštite zdravlja peradi (ista dobna kategorija).
Korištena cjepiva i način cijepljenja
Korištena su dva različita cjepiva i to MARIKAL® SPF (Pliva, Zagreb) koje sadrži soj virusa MB HVT-FC-126, te cjepivo NOBILIS® RISMAVAC (Nizozemska) od stanično pridruženog soja Rispens CVI 988. Od spomenutog broja pilenki njih 243.122 cijepljeno je cjepivom NOBILIS® RISMAVAC (skupina A), a glavnina od 307.955 pilenki (skupina B) istodobno s oba cjepiva. Skupini A cjepivo je dano parenteralno intramuskularno, prema preporuci proiz-vođača, a pilenke skupine B najprije su cijepljene parente-ralno cjepivom NOBILIS® RISMAVAC, a zatim još i cjepi-vom MARIKAL® SPF postupkom nebulizacije (Mazija i sur., 1994.). Cijepljeni pilići, bez obzira na način cijepljenja, još su 24 sata zadržani u prostoru valionice i tek tada ispo-ručeni u uzgojne nastambe. Učinjeno je to kako bi se omo-gućio razvoj viremije i potakla specifična imunost protiv MB (Cho i sur., 1976., Mazija i Voss, 2000., neobjavljeno).
Rezultati
Zdravlje pilenki u pokusu
Cijepljenje bez obzira na način nije izazvalo nikakvih klinič-kih očitovanja koja bi se mogla povezati s primijenjenim postupcima ili različitim cjepivima. U tablici 2. prikazan je uspjeh uzgoja pilenki. Prosječno uginuće pilenki do dobi od 18 tjedana tijekom razdoblja uzgoja uz primjenu jedino cjepiva soja Rispens CVI 98 (devet i pol mjeseci) za 1,94% je više negoli u pilenki cijepljenih s oba cjepiva (Rispens CVI 988 + HVT FC 126). Međutim, struktura uzroka uginuća temeljem razudbe zna-čajno se razlikuje među skupinama pilenki, a prikazana je tablično za obje (A i B) (tablica 3.).
Tablica 2. Uspjeh uzgoja pilenki do dobi od 18 tjedana različito cijepljenih protiv Marekove bolesti
Pokusna skupina Broj pilenki Korišteno cjepivo* Postupak cijepljenja Uginuće tijekom 18 tj. n (%)
Uginuće od MB n (%)
A 40 jata 243.122 CVI 988 i/m 20.533 (8,45) 12.097 (4,98) B 50 jata 307.955 CVI 988 +HVT FC 126 i/m +nebulizacija 20.046 (6,51) 708 (0,23) *NOBILIS® RISMAVAC (Rispens CVI 988) *MARIKAL® SPF (HVT FC 126) Table 2. Success of replacement chickens rearing up to age of 18 weeks, vaccinated against Marek's disease by different means
Experimental group
Number of chickens Vaccine* used Mode of vaccination Mortality rate during 18 w.
n (%) Mortality of MB
n (%) A 40 flocks 243,122 CVI 988 i.m. 20,533 (8.45) 12,097 (4.98) B 50 flocks 307,955 CVI 988 + HVT FC 126 i.m. + nebulization 20,046 (6.51) 708 (0.23) *NOBILIS® RISMAVAC (Rispens CVI 988) *MARIKAL® SPF (HVT FC 126)
IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
70
Tablica 3. Uzroci uginuća pilenki tijekom uzgoja s obzirom na način zaštite od Marekove bolesti
A. Ulazni broj pilenki: 243.122 B. Ulazni broj pilenki: 307.955 Korišteno cjepivo: CVI 988 i/m Korišteno cjepivo: CVI 988 + HVT FC 126 (i/m + nebulizacija) Uzrok uginuća
Uginuće % % od uginuća Uginuće % % od uginuća
E. coli infekcija 1.321 0,54 6,43 3.030 0,98 15,12 Kokcidioza 323 0,13 1,57 1.174 0,38 5,86 Ugušenje 451 0,19 2,20 14.184 4,61 70,76 Iskrvarenje 84 0,03 0,41 242 0,08 1,21 Trauma 165 0,07 0,80 397 0,13 1,98 Marekova bolest* 12.097 4,98 58,91 708 0,23 3,53 Kronična dišna b. 255 0,10 1,24 156 0,05 0,77 Zarazna b. burze 5.812 2,39 28,31 0 0,00 0,00 Kanibalizam 25 0,01 0,12 155 0,05 0,77 Ukupno uginuće 20.533 8,45 20.046 6,51
*Uginuću pribrojen „škart“ s očitovanim znakovima Marekove bolesti Table 3. Causes of death in replacement chickens during rearing period according to vaccination against Marek's disease
A. No. of chickens housed: 243,122 B. No. of chickens housed: 307,955 Vaccine used: CVI 988 i.m. Vaccine used: CVI 988 + HVT FC 126 (i.m. + nebulization) Cause of death
Mortality rate % % of mortality Mortality rate % % of mortality
E. coli infection 1,321 0.54 6.43 3,030 0.98 15.12 Coccidiosis 323 0.13 1.57 1,174 0.38 5.86 Suffocation 451 0.19 2.20 14,184 4.61 70.76 Bleeding 84 0.03 0.41 242 0.08 1.21 Trauma 165 0.07 0.80 397 0.13 1.98 Marek's disease* 12,097 4.98 58.91 708 0.23 3.53 C. R. D. 255 0.10 1.24 156 0.05 0.77 Inf. bursal dis. 5,812 2.39 28.31 0 0.00 0.00 Cannibalism 25 0.01 0.12 155 0.05 0.77 Total mortality 20,533 8.45 20,046 6.51
*"runts" with clinical symptoms of Marek's disease Udio uginuća od MB je 21,65 puta manji u pilenki ci-jepljenih parenteralno (Rispens CVI 988) i nebulizacijom (HVT FC 126).
Rasprava i zaključci
Specifično cijepljenje protiv MB čini se jedinim uspješnim postupkom njena suzbijanja bez kojega bi peradarstvo trpilo enormne štete. Od početka sustavne primjene cjepiva g. 1968. (Churchill i sur., 1969.) istodobno jača virulencija uzročnika MB. Tijekom niza godina korištena cjepiva prila-
gođavana su epizootiološkim prilikama, naročito porastu patogenih svojstava uzročnog virusa početkom 1990.-ih godina (Witter, 1998.) te njegovim također promijenjenim genomskim svojstvima, iako se vjeruje da uspješnost cijep-ljenja ovisi o nizu epigenetskih čimbenika poput imuno-supresije (Mitra i sur., 2012.). Rana infekcija patogenim VMB izazvat će tako ranu citolitičku infekciju te imati za posljedicu smanjenu imunokompetenciju. Imunosupresija uključuje jednako humoralnu i staničnu imunost, što je po-vezano s limfopenijom zbog citolize B i T limfocita. Limfo-penija najčešće je posljedica zaražavanja vrlo virulentnim

IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
71
sojevima virusa MB (Calnek i sur., 1998.). Odatle proizlazi i potreba zaštite netom izleženih pilića od VMB koji se nalazi u kontaminiranom okolišu. Na ovu činjenicu upućuje i pojavnost MB u SAD gdje je ona postala gotovo zanemariva (0,0008%), ponajprije zbog sprječavanja rane infekcije pato-genim VMB iz okoliša, što se postiže primjenom nespeci-fičnih mjera profilakse, ali i gotovo dvostruka njena učesta-lost u zemljama Afrike, Istočne Europe, Istočne Azije i Južne Amerike zbog imunosupresivnih bolesti (Dunn i Gimeno, 2011.). Do pojave epizootije MB koju opisujemo došlo je očevidno zbog izostale mogućnosti primjene nespecifičnih mjera pre-ventive u području prenapučenom nastambama s peradi, međusobno udaljenim tek nekoliko desetaka metara. Tako-đer, zbog te male udaljenosti nije bilo moguće spriječiti niti opsežno zaražavanje netom cijepljenih i u nastambe uselje-nih pilenki virusom iz okoliša. Kompromitiranje učinka inače ispravno obavljenog cijepljenja i inače učinkovitog cjepiva (soj Rispens CVI 988 ) bilo je tako za očekivati (Zanella, 1982.). Tako je značenje rane infekcije patogenim VMB iz okoliša i njen pogubni utjecaj na zdravlje već cijepljenih pilića što je opisuju Dunn i sur. (2010.) dokazano u proizvodnoj praksi. Pilenke skupine A, njih 243.122 ko-mada, cijepljene protiv MB jedino sojem Rispens CVI 988, ugibale su od MB 21,65 puta učestalije negoli pilenke cijepljene najprije ovim sojem intramuskularno, a zatim so-jem HVT-FC-126 postupkom nebulizacije. Marekova bolest je u toj skupini osnova za 58,91% ukupnog uginuća, dok su ostale bolesti zastupljene u različitom stupnju i bez pravil-nosti. Tako na zaraznu bolest burze otpada 28,3% uginuća, ali se zapravo radi o tek dvije enzootije s visokim pomorom. Sva ostala uginuća tehnološke su naravi i povezana sa stup-njem primjene nespecifičnih mjera preventive, najučestalije kolibaciloze (6,3% uginuća) i ugušenja (2,20%) ili pogre-šaka u tehnologiji poput očitovane kronične dišne bolesti (1,24%). Pilenke skupine B (ukupno useljeno 307.955) cijepljene su najprije intramuskularno cjepnim sojem CVI 988 (NOBILIS® RISMAVAC), a zatim postupkom nebulizacije sojem HVT-FC-126 (MARIKAL® SPF). Od MB uginulo je ukupno 0,2% pilenki, odnosno od ukupnog uginuća tek 0,23%. Najviše pilenki uginulo je od nespecifičnih uzroka, ugušenja (70,76% od uginulih) ili ko-libaciloze, što je posljedica izostale sanitarne obrade jaja prije ulaganja u ležionike (15,12%). Ostala uginuća bezna-čajna su, s rijetkom pojavom kokcidioze (5,86% od ugi-nulih), što ne treba nužno tumačiti kao nedostatak, nego više kao poticanje imunosti prema toj protozoarnoj invaziji. Uspjeh istodobne primjene dvaju cjepiva od čega jednog postupkom inhalacije (HVT-FC-126, MARIKAL® SPF) treba pripisati upravo činjenici da je netom izleženo pile došlo najprije u dodir s imunogenom unešenim u dišni su-stav, što je i prirodni način ulaska virusa MB u organizam, najčešće patogenog (Calnek i sur., 1970.). Parenteralna pri-mjena cjepiva protiv MB ne spriječava inhalaciju patogenog
virusa iz okoliša te tako omogućuje nesmetano zaražavanje pilića, sa svim negativnim posljedicama, naročito u slučaju stresa (Zanella, 1982.). U usporedbi s cijepljenjem in ovo kokošjih zametaka dobi 17-18 dana kad viremija, iako nije mjera specifične imunosti (Gimeno i sur., 2008.), no u korelaciji je s količinom anti-gena virusa MB u tkivu slezene (Holland i Silva, 1999.) nastupa za 2-3 dana, postupak nebulizacije virusa HVT-FC-126 (MARIKAL® SPF) predstavlja značajnu prednost. Na-laz HVT-FC-126 u leukocitima već 24 sata nakon primjene postupkom nebulizacije u usporedbi s parenteralno danim virusom, koji je dokazan tek sedmog dana i to u gotovo dvo-struko nižem titru (broj plakova 4,0 prema 2,5), u cijelosti tumači prednosti postupka nebulizacije (Mazija i Voss, 2000., neobjavljeno) i osnovu neposredne zaštite pilića. Pa-renteralno cjepivo (CVI 988) potaći će nastavak već zapo-čete imunosti i tako spriječiti pojavu Marekove bolesti. To neposredno dokazuje nagli prestanak uginuća od MB u jati-ma pilenki koje su se nakon uvođenja postupka nebulizacije cjepnog virusa HVT FC 126 (MARIKAL® SPF) uz intra-muskularno davanje soja CVI 988 (NOBILIS® RISMA-VAC) našle u istovjetnim nastambama i istovjetnom konta-miniranom okolišu, znači i izvoru zaražavanja patogenim virusom MB, kao i prethodne generacije. Zbog navedenog postupak nebulizacije cjepiva od virusa HVT-FC-126 (MARIKAL® SPF) treba ponovno nastaviti koristiti istodobno uz parenteralnu primjenu stanično pridru-ženih cjepnih sojeva virusa MB, kakav je primjerice CVI 988. To bi se naročito moglo preporučiti u zemljama Sje-verne Afrike, Istočne Europe, Istočne Azije i Južne Amerike u kojima je MB u značajnom porastu zbog uglavnom izo-stalih mogućnosti primjene nespecifičnih postupaka zaštite od bolesti (Dunn i Gimeno, 2011.). Kako je cjepivo (MA-RIKAL® SPF) jedinstveno u svijetu s obzirom na to da se je-dino može primijeniti resuspendirano u običnoj destiliranoj vodi te zato i koristiti u postupku nebulizacije, moguće ga je predstaviti bez rezerva i u tom smislu.
Literatura
Anonimno (2010): Marek's disease. U: OIE Terrestrial Manual 2010. Chapter 2.1.13.
BUCKMASTER, A. E., S. D. SCOTT, M. J. SANDERSON, M. E. G. BOURSNELL, N. L. J. ROSS, M. M. BINNS (1988): Gene sequence and mapping data from Marek's disease virus and herpesvirus of turkeys: implications for herpesvirus classification. J. Gen. Virol. 69, 2033-2042.
CALNEK, B. W., S. B. HITCHNER (1969): Localization of viral antigen in chickens infected with Marek's disease herpes virus. J. Natl. Cancer Inst. 43, 935-949.
CALNEK, B. W. H. K. ADLDINGER, D. E. Kahn (1970): Feather follicle epithelium: a source of enveloped and infectious cell-free herpesvirus from Marek's disease. Avian Dis. 14, 219-233.
CALNEK, B. W., R. W. HARRIS, C. BUSCAGLIA, K. A. SCHAT, B. LUCIO (1998): Relationship between the

IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
72
immunosuppressive potential and the pathotype of Marek’s disease virus isolates. Avian Dis. 42, 124-132.
CHO, B. R., R. K. BALCH, R. W. HILL (1976): Marek's disease vaccine breaks: differences in viremia of vaccinated chickens between those with and without Marek's disease. Avian Dis. 20 (3), 496-503.
CHURCHILL, A. E., R. C. CHUBB, W. BAXENDALE (1969): The attenuation, with loss of oncogenicity of the herpes-type virus of Marek's disease (strain HPRS-16) on passage in cell culture. J. Gen. Virol. 4, 322-333.
ČAJAVEC, S., Đ. NEMARNIK, H. MAZIJA, I. REŽIĆ (1992): Stabilnost i imunogenost liofilizirane vakcine Marekove bo-lesti MARIKAL® (HVP-FC-126). XIV. međunarodno savjeto-vanje PLIVA i stočarska proizvodnja. Poreč, 5.-6. listopada 1992. Zbornik sažetaka. str. 10.
DUNN, J. R., I. GIMENO (2013): Current status of Marek's disease in the United States and worldwide based on a questionnaire survey. Avian Dis. (prihvaćeno za tisak prosinac 2012). Print ISSN: 0005-2086.
DUNN, J. R., R. L. WITTER, R. F. SILVA, L. F. LEE, J. FINLAY, B. A. MARKER, J. B. KANEENE, R. M. FULTON, S. D. FITZGERALD (2010): The effect of the time interval between exposure on the susceptibility of chickens to superinfection with Marek's disease virus. Avian Dis. 54, 1838-1049.
GIMENO, I. M., A. L. CORTES, R. F. SILVA (2008): Load of challenge Marek's disease virus DNA in blood as criterion for early diagnosis of Marek's disease tumors. Avian Dis. 52, 203-208.
GOTTSTEIN, Ž., I. CIGLAR GROZDANIĆ, Ž. CVETIĆ, H. MAZIJA (2007): Detection of HVT FC 126 in lung tissue of chickens vaccinated by means of nebulization against Marek's disease. 15th World Veterinary Poultry Congress, Abstract Book. Beijing, China: WVPA, 2007., str. 442.
HOLLAND, M. S., R. F. SILVA (1999): Tissue-specific restriction of latent HVT
transcription. Acta Virol. 43, 148-151. JOHNSTON, P. A., H. LIU, T. O’CONNELL, P. PHELPS, M.
BLAND, J. TYCZKOWSKI, A. KEMPER, T. HARDING, A. AAVAKIAN, E. HADAD, C. WHITFILL, R. GILDERS-LEEVE, C. A. RICKS (1997): Applications in in ovo technology. Poult. Sci. 76:165-178.
KAWAMURA, H., D. J. KING, D. P. ANDERSON (1969): A herpesvirus isolated from kidney cell culture of normal turkeys. Avian Dis. 13, 853-863.
KLEVEN, S. H., C. S. EIDSON (1975): Aerosol administration of turkey herpesvirus vaccine. Proc. 24th Western Poult. Dis. Conf. and 9th Poult. Health Symp. March 18-20, 1975. str. 21.
KOKIĆ, A. (1998): Učinci cjepiva protiv Marekove bolesti pri-pravljenog od soja FC-126 primijenjenog pilenkama lakih hibrida kokoši postupkom raspršivanja odnosno parenteralno. Magistarska rasprava. Veterinarski fakultet Sveučilišta u Zagrebu. Zagreb, 1998.
MORROW, C., F. FEHLER (2004): Marek's disease: a worldwide problem. U: Marek's Disease. An Evolving Problem (F. Davison, V. Nair, ur., str. 49-61. London: Elsevier Academic Press.
MAZIJA, H., S. ČAJAVEC (1993): The distribution of HVT FC 126 antigen in respiratory system of chickens after aerosol vaccination with Marek's disease vaccine. Proc Xth Int. Congr. of the WVPA. Sydney, Australia, 16-19 August 1993. Ref. 150.
MAZIJA, H., S. ČAJAVEC, Đ. VRANEŠIĆ, Đ. NEMARNIK, E. PRUKNER-RADOVČIĆ (1994): Immunogenicity of Marek's disease vaccine HVT FC-126 applied by aerosol of fine particles. Proc. 9th Eur. Poult. Conf. Glasgow UK. August 7-12, 1994. str. 195-196.
MAZIJA, H., T. ŠTIMAC (1993): Patentna prijava uređaja SONOVAC 095. Državni zavod za intelektualno vlasništvo. Patentna prijava: P950425A.
MIŠKOVIĆ, Ž. (2004): Imunogenost cjepiva protiv Marekove bolesti od sojeva CVI 988 ili HVT-FC-126 u pilenki cijep-ljenih zasebno parenteralno ili usporedno parenteralno i za-magljivanjem. Diplomski rad. Veterinarski fakultet Sveučilišta u Zagrebu. Zagreb, 2004.
MITRA, A., J. LOU, H. HUANMING, K. CUI, K. ZHAO, J. SONG (2012): Marek’s disease virus infection induces widespread differential chromatin marks in inbred chicken lines. BMC Genomics 13, 557.
OKAZAKI, W., H. G. PURCHASE, B. R. BURMESTER (1970): Protection against Marek's disease by vaccination with a herpesvirus of turkey. Avian Dis. 14, 413-429.
PAVLOVIĆ, S. (1998): Značajke enzootije zarazne bolesti burze konzumnih nesilica u Bosanskoj Posavini. Magistarski rad. Veterinarski fakultet Univerziteta u Sarajevu. Sarajevo, 1998.
PAYNE, L. N., K. VENUGOPAL (2000): Neoplastic diseases: Marek's disease, avian leukosis and reticuloendotheliosis. Rev. Sci. Tech. Off. Int. Epiz. 19 (2), 544-564.
PRIDYBAILO, N. D., R. N. KORVIN, V. D. SOKOLOV, V. A. LOKH, A. I. SEDUNOVA, T. B. SLEPENKO, V. I. SMOLENSKII, V. I. BOGOLEPOV, S. V. SAFRANOV, R. M. VORONTSOV, E. V. LOMAKINA (1986): Efficacy of aerosol vaccination against Marek's disease. Veterinarya Moscow USSR No. 10, 31-32.
RISPENS, B. H., H. van VLOTEN, N. MASTENBROEK, H. J. L. MAAS (1972): Control of Marek's disease in Netherlands. I Isolation of an avirulent Marek's disease virus (strain CVI 988) and its use in laboratory vaccination trials. Avian Dis. 16, 108-125.
SARMA G., W. GREER, R. P. GILDERSLEEVE, D. L. MURRAY, A. M. MILES (1995): Field safety and efficacy of in ovo administration of HVT + SB-1 bivalent Marek’s disease vaccine in commercial broilers. Avian Dis. 39:211-217.
WITTER, R. L. (1987): New serotype 2 and attenuated serotype 1 Marek's disease vaccine virus: comparative efficacy. Avian Dis. 31, 752-765.
WITTER, R. L. (1998): Control strategies for Marek's disease: a perspective for the future. Poult. Sci. 77, 1197-1203.
ZANELLA, A. (1982): Marek's disease – survey on vaccination failure. Den. Biol. Stand. 52, 29-37.
ZHANG, Y., J. M. SHARMA (2001): Early posthatch protection against Marek's disease in chickens vaccinated in ovo with a CVI988 serotype 1 vaccine. Avian Dis. 45, 639-945.

IMUNOGENOST CJEPIVA PROTIV MAREKOVE BOLESTI OD SOJA CVI 988 DANOG PARENTERALNO ZASEBNO ILI ISTODOBNO SA SOJEM HVT-FC-126 DANOG POSTUPKOM ZAMAGLJIVANJA
73
IMMUNOGENICITY OF THE CVI 988 STRAIN VACCINE AGAINST MAREK'S DISEASE ADMINISTERED SEPARATELY PARENTERALLY OR SIMULTANEOUSLY WITH HVT-FC-126 STRAIN BY NEBULIZATION Summary Immunogenicity of the vaccine against Marek's disease cell associated CVI 988 strain given alone parenterally and simultaneously with HVT-FC-126 turkey herpes virus by nebulization to newly hatched chickens was investigated. During a two-year period, the CVI 988 vaccine was administered to a total of 243,122 chickens in 40 flocks, while 307,955 chickens in 50 flocks were vaccinated simultaneously by nebulization (HVT-FC-126) and intramuscularly (CVI 988). Regardless of the mode of vaccination or the vaccine used, chickens did not show any adverse clinical reaction. During the growing period, 4.98% of hens vaccinated only with CVI 988 strain died from Marek's disease compared to only 0.23% mortality rate in the flocks vaccinated simultaneously parenterally with CVI 988 and with HVT-FC-126 by nebulization. The new knowledge about the significance of early achievement of specific immunity against Marek's disease that is generated by HVT-FC-126 nebulization actualizes its implementation in practice. Key words: Marek's disease, vaccination, HVT-FC-126 strain, nebulization, CVI 988 strain, laying hens

PERADARSKI DANI 2013. 74
PRAKTIČNA ISKUSTVA U PRIMJENI CJEPIVA OD SOJA LA SOTA PROTIV NEWCASTLESKE BOLESTI POSTUPKOM NEBULIZACIJE NETOM IZLEŽENIM TOVNIM PILIĆIMA
Željka Ervaćinović1, Zvezdan Kičeec1, Marica Lolić2, Ivan Križek3, Nikica Stevanović4, Hrvoje Mazija5
1"Gallis" d.o.o., Slavonski Brod, Hrvatska 2Veterinarski zavod Vinkovci, Hrvatski veterinarski institut, Vinkovci, Hrvatska 3Phoenix Farmacija d.d., Osijek, Hrvatska 4Veto d.o.o., Slavonski Brod, Hrvatska 5Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska Sažetak
Istražena je uspješnost cijepljenja postupkom nebulizacije netom izleženih tovnih pilića hibridne linije ROSS 308 protiv newcastleske bolesti cjepivom PESTIKAL® La Sota SPF, Genera, d.d., Kalinovica. Štetna cijepna reakcija se nije očitovala, a pilići su tvorili specifična heminhibicijska protutijela u relativno visokom titru. Pokusnim zaražavanjem pilića sojem Herts 33 virusa pilića u dobi od 42 dana dokazana je specifična imunost pilića jednako pokusne i kontrolne skupine, što se tumači horizontalnim širenjem cijepnog virusa i dovoljno dugim razdobljem za postizanje zadovoljavajuće imunosti. Uspjeh tova pilića prosuđen je temeljem postignute tjelesne mase, uginuća, učinkovitosti korištene krmne smjese te je izračunat indeks proizvodnje (EPEF) koji je za 10,273 indeksnih jedinica bio viši u cijepljenih pilića negoli u necijepljenih pilića. Ključne riječi: newcastleska bolest, PESTIKAL® La Sota SPF, nebulizacija, tovni pilići, specifična imunost, proizvodni uspjeh Uvod
Newcastleska bolest (NB) je virusna, izrazito kontagiozna zaraza domaće peradi i drugih ptica koju uzrokuje virus reda Mononegavirales, obitelji Paramyxoviridae, roda Avulavirus. Sojevi virusa NB (VNB) međusobno se razli-kuju patotipom i tropizmom. Svojstva različitih patotipova VNB opisali su Alexander i Allan (1974.). Newcastleska bolest uzrokuje velike gospodarske štete zbog neposredno izazvanog uginuća, troškova mjera sprječavanja, suzbijanja i njena iskorjenjivanja. Ova se bolest nalazi na listi A Me-đunarodnog ureda za epizootije (OIE) pa se i u Republici Hrvatskoj suzbija temeljem zakonskih odrednica (Ano-nimno, 2012.). Tim se mjerama prvenstveno naređuje spe-cifično cijepljenje sve primljive peradi postupcima koji uključuju i raspršivanje, dakle i nebulizaciju (Mazija i Štimac, 1999.).
Uobičajeni postupak cijepljenja je raspršivanje vodene suspenzije cjepiva promjera čestica 120-150 μm odnosno u odrasle peradi 30 μm. Cjepivo glavninom dospijeva u prednje dijelove dišnog sustava. Netom izleženi pilići ne cijepe se ovim sojem, jer često dovodi do neželjenih reakcija u dišnom sustavu. Međutim, primijeni li se postupkom nebulizacije izaziva dugotrajnu imunost koja traje najmanje 49, a moguće i više dana. Pritom ne oštećuje dišni sustav i ne izaziva štetne cijepne reakcije (Mazija i sur., 2000., Mazija i sur., 2009.). Ne-bulizacija je relativno nov postupak davanja cjepiva netom izleženoj peradi. Posebnim uređajem koji proizvodi aerosol vodene suspenzije cijepnog virusa, čestice promjera 2-5 μm dospijevaju i u najdublje dijelove dišnog sustava pa se uz pluća apsorpcija virusa odvija i u zračnim vrećicama te u pneumatiziranim kostima (Mazija i Štimac, 1999.). Ultrazvuk ne oštećuje cijepni virus, nego mu, naprotiv,
Dr. sc. Željka Ervaćinović, dr.med.vet., Gallis, d.o.o., Vjenceslava Novaka 40, Slavonski Brod; telefon: 035/448-845, fax: 035/448-399, e-mail: [email protected]

PRAKTIČNA ISKUSTVA U PRIMJENI CJEPIVA OD SOJA LA SOTA PROTIV NEWCASTLESKE BOLESTI POSTUPKOM NEBULIZACIJE NETOM IZLEŽENIM TOVNIM PILIĆIMA
75
tijekom kratkog izlaganja povisuje titar zbog razbijanja aglomerata virusnih čestica (Bašić, 2001.) Okulonazalno davanje cjepiva vrlo je učinkovit, ali u indu-strijskoj proizvodnoj peradi zbog pojedinačne primjene neprihvatljiv način. Davanje cjepiva pitkom vodom se ne preporuča, jer soj La Sota ne očituje tropizam prema probavnom sustavu te ga je potrebno dati i 20.000 puta više radi postizanja istovjetnog uspjeha kao i postupkom raspršivanja (Giambrone, 1985.). Uz to, učinak danog cjepiva može biti ometan onečiš-ćenjem pitke vode otopljenim mineralima (soli Cl, Fe, Zn, Cu) ili pak deterdžentima i dezinficijensima koji mogu inaktivirati cijepni virus (Cazaban, 2004.). Primjena cje-piva u pitkoj vodi je neprihvatljiv način osobito u epizo-otiološki ugroženom području, jer imunost nastupa tek za 12-15 dana. Usporede li se navedeni načini cijepljenja, značajnu pred-nost ima raspršivanje, jer se refraktarnost prema patoge-nom virusu NB očituje već za 60 sati, dok još nije došlo do tvorbe specifičnih protutijela. Okulonazalni način primjene cjepiva potiče tvorbu protutijela približno sedmog dana kad nastupa otpornost prema infekciji. Cijepljenje protiv NB cjepivom od soja La Sota, bez obzira na način pri-mjene, izaziva supresiju rasta i nepovoljno utječe na konverziju hrane (Abbas i sur., 2006.).
Materijal i metode
Pokusna perad
Korišteni su jednodnevni tovni pilići oba spola, potomci nesilica hibrida teške linije ROSS 308, podrijetlom iz tvrtke Valionica, d.o.o., Slavonski Brod. Njihovo roditelj-sko jato je tijekom razdoblja uzgoja bilo višekratno cijep-ljeno protiv newcastleske bolesti. Pokusom su obuhvaćena dva jata pilića razdvojena žičanom mrežom, no smještena u istu nastambu korisne površine 130 m2.
Pilići jata 1 su cijepljeni u valionici cjepivom PESTIKAL®
La Sota SPF (Genera d.o.o., Kalinovica) postupkom nebulizacije, dok pilići jata 2 nisu cijepljeni te su poslužili kao kontrolna skupina. Uobičajeno, nastamba je prethodno sanitarno obrađena i pripremljena za prijam pilića. Broj useljenih jednodnevnih pilića i trajanje tova prikazani su u tablici 1. Pilići su primljeni na papir koji je petog dana tova uklonjen te su preostalo razdoblje boravili na stelji od drv-nih strugotina. Krmna smjesa je u prvih pet dana ponuđena u podnim hranilicama koje su ručno punjene, a od šestog dana iz visećih hranilica. Oba jata jela su od 1. do 21. dana početnu smjesu s 22% bjelančevina, završnu smjesu I. s 20% bjelančevina od 22. do 40. dana te završnu smjesu II. s 18% bjelančevina od 41. do 47. dana tova. Na kraju tova izračunat je utrošak i iskoristivost (konverzija) hrane. Pilići su napajani ad libitum iz sustava siski ("nipple") uz dodane zvonolike pojilice. Tijekom prva tri dana pitkom vodom dobivali su multivitaminski pripravak Muvisel prašak (Genera, d.d. Kalinovica) u količini od 100 g na 200 litara vode. Tjelesna masa useljenih jednodnevnih pilića bila je 44 grama. Mikroklimatski uvjeti tijekom trajanja tova usklađivani su prema preporukama proizvođača hibrida (Ross 308 Broiler Management, 2009).
Cjepivo Korišteno je cjepivo PESTIKAL® La Sota SPF (Genera d.d., Kalinovica), namijenjeno cijepljenju pilića, kokoši i purana protiv newcastleske bolesti. Sadržava lentogeni živi VNB soj La Sota umnožen u kokošjim SPF zamecima. Jedna doza sadržava najmanje 106 EID50 spomenutog vi-rusa. Cjepivo je liofilizirano i uz dodatak posebnog stabili-zatora (PPGF). Način primjene cjepiva Netom izleženi tovni pilići jata 1 cijepljeni su protiv NB postupkom nebulizacije. Za tu svrhu korišten je ultrazvučni raspršivač SONOVAC® 2006. Liofilizirano cjepivo je re-suspendirano u destiliranoj vodi tako da se je na svako pile odnosila po jedna doza. Sukladno tome određeno je i vrijeme izlaganja pilića cjepivu kroz 60 sekunda. Kontrola zdravlja pilića Pilići oba jata nadzirani su svaki drugi dan tijekom trajanja tova kad su izvagani (10% pilića) te je određena potrošnja krmne smjese i vode, zabilježeno uginuće, izdvajani su kržljavci (škartiranje), ocijenjeno opće zdravlje te mikro-klimatski uvjeti u nastambama. Pripravak Muvisel prašak primijenjen je radi sprječavanja stresa prije cijepljenja pro-tiv gumborske bolesti i imunosupresije zbog zamjene po-četne krmne smjese završnom smjesom I. (Findrik i sur., 1980.).
Serološke probe Za ocjenu uspješnosti primjene cjepiva punkcijom krilne vene (v. cutanea ulnaris) izvađena je krv od dvadeset na-sumce odabranih pilića i to neposredno nakon leženja te u dobi od 14 i 42 dana, kako bi se odredila najprije količina materalnih serumskih protutijela, vrijeme njihova izlučiva-nja odnosno mogućeg učinka na dano cjepivo te na visinu titra protutijela tijekom trajanja tova pilića do dobi od 47 dana. Za pretraživanje krvi primijenjen je uobičajeni beta postupak IH virusom NB (Allan i sur., 1978.). Na kraju tova pilića, izračunat je europski indeks učinko-vitosti proizvodnje (European Production Efficacy Factor, EPEF) prema jednadžbi:
U postotak preživjelih nisu uračunati izlučeni (škartirani) pilići.
Izazivačka infekcija
Koristili smo dvije skupine po 10 pilića iz svakog jata izdvojenih u dobi od 42 dana, te ih okulonazalno zarazili s po 106 EID50 velogenog virusa NB soj Herts 33. Pilići su tijekom pokusa, idućih sedam dana od zaražavanja, bili smješteni u odgovarajućem prostoru (kavezi) na Veterinar-skom fakultetu u Zagrebu te svakodnevno tijekom sedam dana promatrani radi uočavanja moguće pojave kliničkih znakova bolesti.

PRAKTIČNA ISKUSTVA U PRIMJENI CJEPIVA OD SOJA LA SOTA PROTIV NEWCASTLESKE BOLESTI POSTUPKOM NEBULIZACIJE NETOM IZLEŽENIM TOVNIM PILIĆIMA
76
Tablica 1. Osnovni podaci o tovnim pilićima korištenim u istraživanju
Table 1. Basic data on fattening chickens used in the study
Jato / Flock 1 Jato / Flock 2 Način cijepljenja Mode of vaccination nebulizacija
nebulization Necijepljeni
not vaccinated Podrijetlo pilića Origin of the chickens Valionica d.o.o. Valionica d.o.o.
Broj useljenih pilića Number of chicks housed 1000 400
Trajanje tova (dani) Fattening period (days) 47 47
Rezultati Klinički, cijepljeni (jato 1) i necijepljeni (jato 2) pilići nisu se međusobno razlikovali. Useljeni u nastambe približno 8-10 sati nakon cijepljenja uobičajeno su jeli i pili te se dobro snalazili u prostoru. Tijekom 47 dana tova nisu primijećene cijepne reakcije niti je bilo zdravstvenih poteškoća, a utov-ljeni pilići postigli su zadovoljavajuću tjelesnu masu i visok stupanj preživljavanja (tablica 2.). Tablica 2. Uspjeh tova pilića u pokusu
Tehnološki pokazatelji Jato 1 Jato 2 Broj useljenih pilića 1000 400 Mortalitet 15 (1,5%) 12 (3,00%) Izdvojeni pilići 4 (0,40%) 4 (1,00%) Prosječna tjelesna masa (g) 3.442 3.415 Konverzija hrane (kg/kg) 1.96 1.97 Table 2. Fattening success rate of chickens used in the
study
Technological parameter Flock 1 Flock 2 No. of chickens housed 1000 400 Mortality rate 15 (1.5%) 12 (3.00%) Cull chickens 4 (0.40%) 4 (1.00%) Mean body weight (g) 3.442 3.415 Feed conversion (kg/kg) 1.96 1.97 Oba jata uspješno su utovljena. Pilići jata 1 su postigli povoljnu prosječnu tjelesnu masu od 3,442 kg s također povoljnom konverzijom hrane od 1,96 kg u 47 dana. Jato 2 postiglo je prosječnu masu od 3,415 kg s konverzijom od 1,97 kg/kg. Nadalje, stupanj preživljavanja u jatu 1 vrlo je visok te iznosi 98,08%, a patoanatomskom pretragom uginulih pilića nisu utvrđeni znaci koji bi upućivali na pojavu bolesti (tehnološko uginuće). Zamjetno je da je jato 1 koje je cijepljeno protiv NB postiglo bolje rezultate tova
uz nešto manju konverziju hrane te manju smrtnost. To se očituje i u vrijednosti EPEF koja za cijepljeno jato (jato 1) iznosi 368,038, a za necijepljeno (jato 2) 357,765, što čini razliku od 10,273 indeksnih jedinica u korist cijepljenih pilića. S obzirom na gustoću populacije (13 pilića/m2) na kraju tova se po m2 našlo prosječno 10,5 pilića odnosno 36 kg pilića žive vage. To ponešto (za 4 kg/m2) prelazi standarde (32 kg/m2), no nije se štetno odrazilo na zdravlje pilića. Iz rezultata seroloških pretraga (tablica 3.) u dobi pilića od 1, 14 i 42 dana uočljiv je visok titar pasivno stečenih HI protutijela u jednodnevnih pilića. Titri HI protutijela za VNB prikazani su u tablici 3. i slici 1. Tablica 3. Titar HI protutijela za VNB u krvnom serumu
tovnih pilića (log2)
Jato 1. dan 14. dan 42. dan 1 4,9 2,55 1,95 2 4,9 1,45 1,44
Table 3. Finding of specific HI antibodies for NDV in
blood sera of chickens in the course of trial (log2)
Flock Day 1 Day 14 Day 42 1 4.9 2.55 1.95 2 4.9 1.45 1.44
Slika 1. Nalaz specifičnih HI protutijela za VNB u
krvnom serumu pilića tijekom pokusa (%)
Figure 1. Finding of specific HI antibodies for NDV in blood
sera of chickens in the study (%)

PRAKTIČNA ISKUSTVA U PRIMJENI CJEPIVA OD SOJA LA SOTA PROTIV NEWCASTLESKE BOLESTI POSTUPKOM NEBULIZACIJE NETOM IZLEŽENIM TOVNIM PILIĆIMA
77
Izazivačka infekcija je u obje skupine pilića (cijepljeni i necijepljeni) polučila istovjetni rezultat, tj. preživljavanje svih pilića bez očitovanja znakova bolesti. S obzirom na ovaj ishod pilići nisu žrtvovani.
Rasprava
Cijepljenje netom izleženih tovnih pilića postupkom nebu-lizacije cjepivom od La Sota soja VNB nije izazvalo štetnu kliničku reakciju. Postupak je siguran i učinkovit u poti-canju dugotrajne specifične imunosti, što u cijelosti potvr-đuje rezultate preliminarnih istraživanja (Mazija i sur., 2009.). Specifična materalna protutijela VMB nisu omela učinak danog cjepiva, no nalaz protutijela u 35% jednako cijepljenih i necijepljenih pilića u dobi od 42 dana ukazuje na horizontalni prijenos cijepnog virusa (Abbas i sur., 2006.). Postignuti proizvodni uspjeh utovljenih pilića ocijenjen europskim indeksom učinkovitosti proizvodnje bolji je u cijepljenih pilića za 10,273 indeksnih jedinica, što je iznenađujuće s obzirom na uvriježeno vjerovanje da svako cijepljenje izaziva stres. To se osobito odnosi na cijepni soj La Sota VNB koji se zato ne primjenjuje netom izleženim pilićima (Abbas i sur., 2006.). Ipak, u slučaju cijepljenja iz nužde soj La Sota moguće je koristiti za cijepljenje i klasičnim postupkom raspršivanja, a očeki-vanu cijepnu reakciju zbog djelovanja oportunističkih bak-terija može se spriječiti primjenom antibiotika 3-4 dana nakon toga (Mazija i sur., 1981.). Visoki titar specifičnih HI protutijela što su ih pilići stekli od višekratno cijepljenih nesilica dokazan je u svih pilića (4,9 log2) i nije omeo učinak cijepnog virusa. Ova se činjenica može pripisati enormnoj količini cijepnog virusa koji je primijenjenim postupkom nebulizacije unesen do najdubljih dijelova dišnog sustava te nije zaustavljen u nje-govim prednjim dijelovima, što se događa kod uobičajenog postupka raspršivanja cjepiva (Hayter i Besch, 1974.). Treba spomenuti i malobrojnost sesilnih makrofaga u dišnom sustavu ptica (Toth i Siegel, 1986.), što ne sprje-čava ulazak cijepnog virusa u epitelne stanice dišnog susta-va koji tako interferira s mogućim ulaskom patogenog virusa. Izostalu cijepnu reakciju na primijenjeno cjepivo protiv NB od soja La Sota treba pripisati i homogenom aerosolu čestica suspenzije cijepnog virusa. Cijepljeni pilići, ali isto tako i necijepljeni, podnijeli su izazivačku infekciju patogenim Herts 33 sojem VNB bez obzira na niski titar specifičnih HI protutijela dokazanih 42. dana života u cijepljenih (1,95 log2 ) odnosno necijep-ljenih (1,44 log2) pilića. Otpornost prema patogenom virusu valja pripisati prvenstveno staničnoj, a ne humoral-noj imunosti (Al Garib i sur., 2003.). Ovakav rezultat izazivačke infekcije razumljiv je s obzirom na činjenicu da su obje skupine pilića boravile u istom prostoru te je omo-gućeno horizontalno širenje cijepnog virusa. Ovo je prvi opis opsežnije primjene cijepnog soja La Sota postupkom nebulizacije u gotovo terenskim uvjetima te je bio osnova njegova prihvaćanja u proizvodnoj praksi. Bez obzira na uspješnost opisanog postupka postoji objektivna
mogućnost da se pojedina cjepiva nominalno istovjetnog sadržaja (La Sota soj VNB) međusobno razlikuju. Osnova toga je mješovita populacija cijepnih VNB sadržanih u različitim cjepivima, na što upućuju Wakamatsu i sur. (2006.). S obzirom na ovu činjenicu naglašavamo da se opisani rezultat istraživanja odnosi na soj (ili mješavinu klonova) La Sota sadržan u cjepivu PESTIKAL® La Sota SPF (Genera d.o.o., Kalinovica). Zaključno, temeljem rezultata preliminarnih (Mazija i sur., 2000.) i opisanih istraživanja može se ustvrditi da je cijepljenje netom iz-leženih pilića protiv NB još u valionici, postupkom ne-bulizacije zbog povoljnijeg proizvodnog uspjeha cijep-ljenih pilića, izostale cijepne reakcije i dugotrajne imu-nosti, u cijelosti zadovoljavajući postupak zaštite peradi od newcastleske bolesti.
Literatura
ABBAS, T., M. A. MUNNER, M. D. AHMED, M. A. KHAN, M. YOUNUS, I. KHAN (2006): Comparative efficacy of five different brands of com-mercial Newcastle disease Lasota virus vaccines in broilers. Pakistan Vet. J. 26(2), 55-58.
ALEXANDER, D. J., W. H. ALLAN (1974): Newcastle disease virus pathotypes. Avian Pathol. 3(4), 269-278.
Al-GARIB, S. O., A. L. J. GIELKENS, E. GRUYS, G. KOCH (2003): Review of Newcastle disease virus with particular reference to immunity and vaccination. World Poult. Sci. J. 59, 185-200.
ALLAN, W. H., J. E. LANCASTER, B. TOTH (1978): Newcastle disease vaccines – their production and use. FAO Animal Production Series. No. 10 Rome: FAO.
ALLAN, W. H., R. E. GOUGH (1974): Standard haemaggluti-nation inhibition test for Newcastle disease. A comparison of macro and micro methods. Vet. Rec. 95, 120-123.
Anonimno (2012): Naredba o mjerama zaštite životinja od zaraz-nih i nametničkih bolesti i njihovom financiranju u 2012. godini. Narodne novine broj 17/12 i 100/12.
BAŠIĆ, I. (2001): Osjetljivost prema ultrazvuku lentogenih so-jeva virusa newcastleske bolesti različitog tropizma. Diplom-ski rad. Veterinarski fakultet Sveučilišta u Zagrebu. str. 1-30.
BEARD, C. W., R. P. HANSON (1984): Newcastle disease. U: Diseases of Poultry, 8. izd. (M. S. Hofstad, H. J. Barnes, B. W. Calnek, W. M. Reid, H. W. Yoder, ur.). Iowa State University Press. Ames IA. str. 452-470.
CAZABAN, C. (2004): Water quality determines the success of oral vaccination. World Poultry 20, 32-34.
FINDRIK, M., V. ŠERMAN, H. MAZIJA (1980): Istraživanje utjecaja hranidbe na solidnost stečenog imuniteta protiv atipične kuge peradi. I. Utjecaj zamjene hrane na uspjeh istodobne vakcinacije protiv atipične kuge peradi i prirast tjelesne težine pilića. Vet. Arhiv 50, 115-119.
GIAMBRONE, J. J. (1985): Laboratory evaluation of Newcastle disease vaccination programs for broiler chickens. Avian Dis.29(2), 479-487.
HAYTER, R. B., E. L. BESCH (1974): Airborne-particle de-position in the respiratory tract of chickens. Poult. Sci. 53, 1507-1511.
MAZIJA, H., S. ČAJAVEC, Đ. NEMARNIK, E. PRUKNER-RADOVČIĆ, N. ERGOTIĆ (2000a): Day old commercial

PRAKTIČNA ISKUSTVA U PRIMJENI CJEPIVA OD SOJA LA SOTA PROTIV NEWCASTLESKE BOLESTI POSTUPKOM NEBULIZACIJE NETOM IZLEŽENIM TOVNIM PILIĆIMA
78
chickens given La Sota virus by ultrasonic nebulizer resist challenge with Herts 33 strain at day 42. Abstracts & Pro-ceedings, XXI World's Poultry Congress, Montreal, 33-34.
MAZIJA, H., S. ČAJAVEC, E. PRUKNER-RADOVČIĆ, N. ERGOTIĆ, I. CIGLAR GROZDANIĆ, Ž. GOTTSTEIN, A. KOKIĆ, W. L. RAGLAND (2009): Immunogenicity and safety of La Sota strain of Newcastle disease virus adminis-tered to newly hatched chicks by nebulization. Acta Vet. Brno 78, 137-144.
MAZIJA, H., T. ŠTIMAC (1999): Ultrazvučni raspršivač vakcina protiv Marekove i drugih bolesti peradi. Hrvatski patentni glaasnik 4, 877. Patent broj: A 61 D 7/00.
MAZIJA, H., Z. BIĐIN, A. ŠTAJER, M. MIKEC (1981): Vakci-nacija iz nužde kokoši protiv atipične kuge peradi (Emer-gency vaccination against Newcastle disease). Praxis Vet. 29: 45-52.
TOTH, T. E, P. B. SIEGEL (1986): Cellular defense of the avian respiratory tract: paucity of free-residing macrophages in the normal chicken. Avian Dis. 30(1), 67-75.
WAKAMATSU, N., D. J. KING, B. S. SEAL, B. P. H. PEETERS, C. C. BROWN (2006) The effect on patho-genesis of Newcastle disease virus LaSota strain from a mutation of the fusion cleavage site to a virulent sequence. Avian Dis. 50(4), 483-488.
PRACTICAL EXPERIENCES WITH THE APPLICATION OF VACCINE AGAINST NEWCASTLE DISEASE LA SOTA STRAIN TO NEWLY HATCHED BROILER CHICKS BY NEBULIZATION Summary The effect of vaccination of newly hatched broiler chickens hybrid ROSS 308 against Newcastle disease with the PESTIKAL® La Sota SPF vaccine (Genera, d.d., Kalinovica) by nebulization was investigated. All broiler flocks were placed in the same poultry houses under the same technological conditions. There was no vaccination reaction and rather high antibody titer values were recorded in broiler chicks. Upon experimental infection at the age of 42 days, specific immunity was proven by serologic tests and hemagglutination inhibition for Newcastle disease. During fattening period, the chicks were clinically monitored, production data (body weight, mortality and feed conversion) were recorded, and productive index was calculated (EPEF). In vaccinated chickens, it was by 10.273 index units higher compared to non-vaccinated chickens. Key words: Newcastle disease, Pestikal® La Sota SPF, nebulization, broiler chicks, specific immunity, production results

PERADARSKI DANI 2013.
79
TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE
Milivoj Mikec, Tihomir Zglavnik
Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak
U ovom radu opisane su dosadašnje spoznaje i postupci suzbijanja tekuti u peradi te istraživanja (biopredatori tekuti, vakcinacija, entomopatogenične gljive itd.) na rezultatima kojih će se u skoroj budućnosti temeljiti program njihove eradikacije. Invazija peradarskih nastambi s tekuti ima za posljedicu zdravstvene i proizvodne probleme kokoši, koji se na kraju proizvodnog ciklusa očituju kao velika gospodarska šteta. Zasad još uvijek ne znamo dovoljno biologiju grinja, što je i razlog naše neučinkovitosti u borbi protiv njih. Trenutno, peradari se najčešće koriste kemijskim akaricidima jer drže da će se njihovom uporabom najbrže riješiti tekuti, no uglavnom njihove želje ostaju neispunjene budući da korišteni pripravci teško dospijevaju do grinja jer su one u skrovitim mjestima ili na domaćinima duboko ispod perja. Poradi toga, nužno je koristiti kombinaciju više uobičajenih i alternativnih metoda suzbijanja da se ostvari neki uspjeh i broj tekuti održi na podnošljivoj razini jer će tada šteta biti manja. Naime, dok je hematopoetski sustav životinje u stanju kompenizirati manjak krvi koju grinje posišu, do tada i nema neke veće štete, no kada broj tekuti dosegne kritičnu masu nastaju zdravstveni i proizvodni problemi. Zbog toga je važno provoditi kontrolu tijekom proizvodnje da bi se na vrijeme reagiralo. Ključne riječi: perad, tekut, preventiva, terapija Uvod Dermanyssus gallinae ili tekut uzrok su velikih problema u peradarskoj proizvodnji koji se na završetku očituju kao sla-bija nesivost i povećano uginuće, a visina štete zavisi o stupnju invadiranosti kokoši (Mul i sur., 2009.). Suzbijanje ektoparazitoza u peradarskoj proizvodnji, poglavito ako se radi o grinjama, sizifovski je posao. Naime, može se reći da je tekut gotovo najveći zdravstveni i proizvodni problem u peradarstvu otkad ono postoji (Maurer i sur.,1988; Hoglund i sur., 1995.; Kilpinen i Steenberg, 2009.). Jednom kada tekut uđe u nastambu gotovo je nemoguće riješiti se je, bu-dući da se radi o parazitu koji izvan domaćina može pre-živjeti devet mjeseci u temperaturnim uvjetima od 9 ºC do 25 ºC (Nordenfors i sur., 1999.). Ako tome pridodamo da se ona zavlači u najskrovitija i teško dostupna mjesta perad-njaka i boravi na kokošima ispod perja, onda se može shvatiti zašto je dermanisoza takva napast za peradarsku proizvodnju. Unatoč silnim naporima svih sudionika u pro-cesu proizvodnje te pratećih kemofarmacijskih i znanstvenih institucija još uvijek nemamo pravi odgovor za tu ekto-parazitozu. Spoznaje se svakodnevno proširuju i povećava se broj korištenih akaricida, no riješiti se tekuti gotovo je
nemoguće. Prema posljednjim obavljenim rezultatima istra-živanja oko 70% svjetskog uzgoja infestirano je tekutima, a neke zemlje s razvijenom peradarskom proizvodnjom, pri-mjerice Nizozemska, trpi godišnju štetu zbog tekuti od 11 milijuna eura (Emous i sur., 2005.), a Europska unija oko 130 milijuna eura, što je gotovo pola eura po kokoši.
Životni ciklus i stanište tekuti
Ektoparaziti peradi su artropodi ili člankonošci koji žive na koži ili perju domaćina, koji se u uzgoje šire putem tehno-loške opreme, odjeće radnika, jaja te divljih ptica i glodava-ca. No najčešći put širenja je sama perad preko koje se invadiraju čiste farme, a što se događa tijekom useljenja novih jata. Dvije su skupine ektoparazita zanimljivih za pe-radarstvo, a to su pauši koje se hrane mrtvim stanicama kože i perjem, te grinje ili tekut koje se hrane krvlju i tema su ovoga rada. Kokošja tekut ili crvena grinja hematofagni su ektoparaziti prošireni diljem svijeta koji uz kokoši invadiraju još i pura-ne, golubove, kanarince te druge divlje ptice (oko 30 vrsta). Prvi opis Dermanyssus gallinae dao je De Geer (1778.). Uza spomenute ptice tekuti napadaju i čovjeka, poglavito za
Dr. sc. Milivoj Mikec, Hrvatski veterinarski institut, Centar za peradarstvo, Hienzelova 55, 10000 Zagreb, Hrvatska; [email protected]

TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE 80
vrijeme iseljavanja kokoši iz nastambi kada im on ostaje jedini izbor (Sikes and Chamberlain, 1954.). Grinje peradi su iz porodice Dermanyssidae i obuhvaćaju kokošju tekut te sjevernu i tropsku kokošju tekut (Biđin, 2008.). Kokošja tekut ili Dermanyssus galline proširena je diljem svijeta i najčešće je nalazimo u toplim krajevima, u nastambama s konzumnim i rasplodnim nesilicama. Brojlerska jata veoma su rijetko invadirana, dok su uzgojna nešto češće, ali ne toliko kao nesilice. Sjeverna kokošja tekut ili Ornithonyssus sylviarum je ektoparazit u svim područjima s uzgojem pe-radi, divljih ptica pa i čovjeka. Ona se često zamijeni kokošjom tekuti, ali za razliku od nje ima drukčiji izgled dorzalne i analne ploče. Uz to, ona se na ptici zadržava tije-kom dana i noći, a na njoj obavlja svoj razvojni ciklus. Kokošja tekut nakon hranjenja napušta domaćina i zavlači se u procijepe i pukotine u kojima odlaže jaja, a gotovo uvijek se može naći na gnoju i stelji (Maurer,1993.). Samo mali broj tekuti zadržava se na domaćinu tijekom dana i obično ga u jutarnjim satima napuštaju (Wood, 1917.). Treba reći da se sve više može čuti kako opisano ponašanje kokošje tekuti u intenzivnoj proizvodnji baš i nije takvo i da se više odnosi na dvorišne kokoši. Naime, tekuti smeta dnevno svjetlo pa zato napušta perad tijekom izlaska iz kokošinjca i zavlači se na tamna mjesta unutar njega. Budući da su su-vremeni peradnjaci bez dnevnog i s umjetnim svjetlom slabog intenziteta tekut nema razloga bježati s domaćina i može se reći da se u velikom broju zadržava na domaćinu cijelo vrijeme, što još više otežava borbu protiv nje. Tropska kokošja tekut vezana je isključivo uz tropske krajeve i vrlo je slična sjevernoj kokošjoj tekuti od koje se razlikuje izgle-dom dorzalne ploče. Za naše peradarstvo nema nekakvog značenja. Odrasla ženka veličine je od 0,7 do 1,1 mm i različitih je boja, od sive do tamno crvene, što ovisi o količini posisane krvi. Jaja odlaže u okoliš kokoši 12 do 14 sati nakon hranjenja i ovisno o temperaturi okoliša larve se izlegu za 48 do 72 sata nakon polaganja jaja. Za jedan do dva dana presvuku se u protonimfu (0,35 do 0,60 mm) i nakon toga u deutonimfu (0,55 do 0,80 mm), koja se nakon sisanja krvi pretvara u odraslu tekut (Maurer i sur., 1988.; Axtell i Arends, 1990.). Cijeli razvojni ciklus (slika 1.) traje od 5,5 do 7 dana, što ovisi o ambijentalnim uvjetima (Kirkwood, 1968.; Nordenfors i sur., 1999.). Tekut se razmnožava samo u prisutnosti njihova domaćina, a krv sišu samo nimfe i odrasle ženke, dok mužjaci to tek povremeno rade. Hranje-nje traje do 60 minuta, nakon čega tekut napušta domaćina (Maurer i sur. 1988.), ali ostaje uz njegovo ležište. Parenje se odvija oko 30 minuta, a ženke polažu jaja oko tri dana nakon svakog hranjenja. Polaganje jaja obnaša se pri temperaturi od 5 do 45 ºC, a najveći broj jajašaca odlaže se pri temperaturi od 20 ºC do 25 ºC i relativnoj vlazi od 70 (Maurer i Baumgartner, 1992.; Nordenfors i sur., 1999.). Ženka nese jaja u razdoblju od oko 8 tjedana, a kada temperatura u peradnjaku padne ispod 9 ºC nema razvoja niti razmnožavanja. Isto se događa kod niske vlage i tem-perature iznad 40 ºC, a ugibanje tekuti nastupa kod tempe-rature iznad 55 ºC. Dosta nejasnoća u razumijevanju biolo-gije tekuti unosi činjenica da je u laboratorijskim uvjetima
spolni odnos mužjaka i ženki nakon leženja 50:50, dok u prirodnom okruženju jedan mužjak dolazi na nekoliko sto-tina ženki (Oliver, 1965.; Maurer i Baumgartner, 1992.).
Slika 1. Razvojni ciklus Dermanyssus gallinae (V. Maurer,
FiBL). Sezonska pojavnost tekuti uglavnom je potaknuta tempera-turom (Kirkwood, 1968.), no primijećeno je da ona ovisi više o brzini porasta temperature negoli o stalnosti tempera-ture zraka (Kilpinen i Steenberg, 2009.). Aktivnost tekuti najveća je po noći kada je i najlakše uočimo (Kirkwood, 1968.; Guy i sur., 2004.). Naime, nakon zamračenja tekut napušta svoja skrovišta i odlazi na perad penjući se uz rešetke kaveza i po njenim nogama, a odabire mjesta na tijelu s najmanje perja i gdje je koža tanja. Uglavnom su to vrat, leđa i područje ramenog zgloba (Maurer, 1993.), jer na tim mjestima ih kokoš teško može dohvatiti i pojesti. Brzina kretanja tekuti je otprilike 10 sekundi za 1 cm. Pronalazak domaćina je složen i drži se da u tome vjerojatnu ulogu ima ugljični dioksid (Kilpinen, 2005.), toplina i vibracije (Kilpinen i Mullens, 2004.). Neka istraživanja ukazuju na to da bi u pronalaženju domaćina ulogu mogli imati lipidi kože i feromoni (Enterkin i Oliver, 1982.; Zeman, 1988.). Nakon nekoliko dana gladovanja tekuti su jako agresivne i ne prave razliku hoće li napasti kokoš, čovjeka ili druge toplokrvne kralježnjake, mada treba reći da preferiraju ptice. Pri nor-malnom intenzitetu svjetla kretanje tekuti je slabo ili ni-kakvo, što je vjerojatno obrambeni mehanizam da je kokoš ne vidi i pojede, no zato se po mraku kreće veoma dobro, jer tada nema opasnosti od domaćina koji ih tada ne vidi. Ponekad se kokošja tekut stalno zadržava na kokošima i na njima se razmnožava, što je vjerojatno povezano sa stanjem mikroklime i intenzitetom svjetla u nastambama te s jačinom invazije. Odrasla tekut bez krvnog obroka može preživjeti do dva tjedna na temperaturi od 20 ºC do 25 ºC, ali na temperaturi od 5 ºC do 10 ºC i do devet mjeseci, a nimfe do pet mjeseci. U intenzivnoj kaveznoj i podnoj peradarskoj proizvodnji jaja obilje je skrovitih mjesta u koja se tekut zavlači (traka za skupljanje jaja, gnojna mjesta, ventilacijski kanali, ispod ko-rita za hranu, okovi i brave za zatvaranje kaveza, gnijezda, prečke itd.). Tako je utvrđeno da se tekut voli razmnožavati i

TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE
81
skrivati u pukotinama manjim od 2 mm i to u materijalima od papira, plastike i drveta, a ne voli aluminij i staklo (Chirico, 2009.). No, uglavnom se može reći da je jačina invazije slabija u kavezno držanih kokoši negoli u onih u podnom ili slobodnom držanju. Istraživanja su pokazala da se populacija tekuti najviše razvila 30 dana nakon početka invazije, ali valja napomenuti da je gustoća naseljenosti tekuti farmska specifičnost. Optimalna temperatura za život i razvoj grinja je 25 ºC do 30 ºC, pa su najveće invazije u ljetnim mjesecima (Maurer i Baumgartner, 1992.).
Problemi u proizvodnji
Tekut je uzrok stalne neugode i nemira kokoši, poremećaja spavanja, čupkanja perja i kanibalizma (Chauve,1998.), gu-bitka težine, anemije i slabije nesivosti (Nordenfors i sur., 1999.; Kilpinen, 2001.). Uočljiva anemija s blijedom krije-stom i podbradnjacima te ružičast kljun (Kilpinen i sur., 2005.), kao i ekstremno uginuće (do 50%) posljedica je vrlo jake invazije, no to se nasreću ipak rijetko događa. Takva stanja moguća su kada se na životinji nađe od 150 do 200 000 grinja. Uz trajno nizak broj tekuti kokoši kompenziraju gubitak krvi svojim hematopoetskim resursom, no regenera-tivna anemija posljedica je veoma jakih invazija. Kokoš invadirana s 20 000 tekuti tijekom noći izgubi do 4 grama krvi, jer adult tekut posiše u obroku oko 2 µL krvi (Sikes i Chamberlain, 1954.), što je oko 3% za kokoš tešku 1,8 kg. Tekutima napadnuta perad u stalnom je kroničnom stresu i kao takva podložna nizu drugih bolesti zbog pada imuniteta, pa je pojava primjerice kolibaciloze u takvim jatima veoma česta. Istraživanja na terenu govore o padu nesivosti za 10% do 20%, a zabilježeno je i do 90% (Axtell i Arends, 1990.; Arkle, 2007.). Kod podno držanih nesilica (matična jata) česta je pojava podnih jaja, jer kokoši bježe iz gnijezda zbog uznemiravanja. Uz pad nesivosti opada i kvaliteta jaja zbog prisutnosti krvnih mrlja na ljusci (od zdrobljene tekuti na traci za jaja) i ekskremenata tekuti (crvenkasto crne točkice), tako da se broj nekomercijalnih ili jaja druge klase kod sred-nje jake invazije poveća za do 15% (Chauve, 1998.). Treba napomenuti da uz opisane štete tekut može prenositi niz bakterijskih i virusnih bolesti ljudi i životinja (Nordenfors i sur., 1999.; Kilpinen, 2001.). Tako je utvrđen prijenos mikroorganizama Salmonella Enteritidis, Escherichia coli, Shigella spp, Staphylococcus spp. (Moro i sur., 2009.), Cardinium spp., Spiroplasma spp., Rickettsiella spp., Schi-neria spp. (De Luna, 2009.), a još i P. multocida, Chlamydia spp., Borrelia anserina, Erysipelothrix rhusiopathiae, Liste-ria monocytogenes, Coxiella burnetii i više virusnih bolesti (Toga virus, Saint Louis encefalitis, boginje peradi, ptičju leukozu, newcastlesku bolest). Kao što je prije rečeno, tekuti znaju napadati i ljude. Ugrizi su bolni i variraju od blage iritacije do teške urtikarije s kožnim ekcemom (papulovezikularni dermatitis). Osip koji se sastoji od malenih crvenkastih papula može se najviše naći na rukama, prsima i leđima (Sahibi i sur., 2008.; Potenza i sur., 2008.). Problemi najčešće nestaju bez po-sebne terapije. No, ima još veći problem, a to je negativan učinak korištenih kemikalija u svrhu uništavanja tekuti.
Naime, one ne samo da imaju izravan štetan učinak na zdravlje djelatnika koji rade u peradnjacima, već i neizravan na konzumente kroz prehranu jajima iz takve proizvodnje (Hamscher i sur., 2003.). Istraživači se slažu da postoje indicije koje upućuju na prisutnost tekuti u jatu i da bi trebali temeljito pregledati unutrašnjost nastambe i perad, a to su: - pojačano žeđanje - slabija nesivost - pojačano uzimanje hrane uz lošu konverziju - imunosupresija - izbjegavanje invadiranih mjesta u nastambi - pojava bolesti u nesilica koje tekuti prenose - slaba kakvoća perja
Monitoring, prevencija i suzbijanje
Rano otkrivanje tekuti vrlo je važno za očuvanje zdravlja i proizvodnost nesilica. Potencijalna skrovišta u nastambama su stranice kaveza, valovi, prečke, izmet, pukotine itd. Kada radnici u farmi primijete jasne znakove invazije (feces tekuti i njihove nakupine na opremi, krvave mrlje na jajima, ugrizi po rukama), tada je već invazija rasprostanjena i jaka. Zbog toga je dobro koristiti zamke za tekut koje su razradili i vrednovali Nordenfors i Chirico (2001.), ali i neke već prije korištene zamke i metode otkrivanja. Dobrim se pokazalo lupanje po kavezu i postavljanje bijele plahte ili papira ispod njega, kao i brisanje kaveza bijelom krpom na kojoj će u slučaju prisutnosti tekuti ostati krvav trag. Na tim se mjestima mogu vidjeti i ekskrementi tekuti. Za nadziranje se također koriste prije spomenute zamke. One su od rebrastog kartona ili plastike, jer će se tekut skrivati u tamne prostore nabora rebrastog kartona, zatim tuba zamke, sjedala zamke i tzv. monitoring zamke. Sve se one postavljaju u neposrednu blizinu kokoši, uz kaveze ili gnijezda, a dobro ih je tretirati nekim akaricidom, no u tom slučaju moraju se udaljiti da nisu dostupne kokošima. Nadalje, možemo i sami napraviti zamke od papirnatih ručnika koje zavežemo u čvorove. Ovakve zamke se objese na nekoliko mjesta u kavezu i tekuti se, ako ih ima, skupljaju unutar čvorova trake. Novija razmišljanja idu u smjeru korištenja elektroničkih senzora u smislu ranog otkrivanja tekuti, ali za to još treba pričekati neko vrijeme. Iz jedne u drugu farmu tekuti se uglavnom prenose preko kutija za jaja i druge transportne opreme, preko ljudi (rad-nika, vakcinatora, posjetitelja), divljih ptica i glodavaca. Naročito su opasni vrapci koji gnijezda rade od kokošjeg perja. Dakle, veoma je važno provoditi optimalne tehno-loške i biosigurnosne mjere (kontrola nove peradi i opreme, dezinfekcija, dezinsekcija, deratizacija farmi), što je najbolja prevencija. Osobitu pozornost treba posvetiti sanitarnom po-stupku u nastambama, što podrazumijeva korištenje visoko-tlačnih prskalica, a u slučaju jake invazije u prijašnjem proizvodnom ciklusu i nekog valjanog akaricida. U suzbijanju tekuti koristimo se konvencionalnim i alterna-tivnim metodama. Prve imaju za cilj ubiti tekut ili spriječiti invaziju, dok alternativne metode uključuju korištenje modi-

TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE 82
ficirane osvijetljenosti i temperature, mirisa, predatora teku-ti, entomopatogenične gljive i cjepiva u svrhu ograničenja i širenja invazije.
Konvencionalne metode
Prva od konvencionalnih metoda koju ću opisati je dobro čišćenje i pranje nastambe te opreme. Premda se misli da tim postupkom ne pridonosimo u većoj mjeri ograničenju broja tekuti, istina je sasvim suprotna. Naime, ukoliko tu sanitarnu mjeru obavimo na ispravan način (pranje pod tlakom i toplom vodom uz korištenje nekog detergenta ili primjena vakum čiščenja)smanjenje broja tekuti i njihovih jaja biti će veoma visoko (Nordenfors i Hoglund, 2000.). Također je dobro tijekom sanitarnog postupka kao i u vrijeme pri-sutnosti peradi u nastambi koristiti neki akaricid, ali uza sve mjere opreza (korištenje zaštitne odjeće i maske) No, s njima ne treba pretjerivati zbog moguće opasnosti od konta-minacije jaja te rizika od otrovanja za kokoši i ljude. Uz to, dokazan je brzi razvoj rezistencije na ta sredstva kao što je opisano u Italiji (Marangi i sur., 2009.), Švedskoj (Norden-fors i sur., 2001.), Velikoj Britaniji (Thind i Ford, 2007.) i Francuskoj (Beugnets i sur., 1997.). S obzirom na broj ko-rištenih kemijskih sredstava u borbi protiv tekuti i nezadovo-ljavajuće rezultate očito je da ona baš i nisu najbolje rješenje za taj problem. Zadovoljni bismo bili nekim kemijskim pripravkom koji bi imao slijedeće osobine: - brzo i učinkovito djelovanje - ne stvara rezistenciju i - nije toksičan za ljude i životinje Nažalost, zasad nemamo takav kemijski akaricid. Lepeza korištenih akaricidnih pripravaka prilično je široka, a oni se uglavnom temelje na: - organofosfornim spojevima (inhibitori acetilkolinesteraze
u tekuti) - kloriranim ugljikovodicima (inhibitori acetilkolinesteraze
u tekuti) - karbamatima (inhibitori acetilkolinesteraze u tekuti) - piretroidima (neurotoksini) Njihova je učinkovitost na tekut neupitna, no prigovori se odnose na očuvanje sigurnosti hrane i ekološku sferu te tok-sičnost za ljude i životinje. Međutim, slabiji rezultati njihove primjene u naseljenom peradnjaku posljedica su teške dostupnosti djelatnih tvari u akaricidnim sredstvima za te-kut, jer je ona dobro zaštićena u svojim skrovištima i ispod perja kokoši. Organofosforni spojevi, klorirani ugljikovodici i karbamati sprječavaju sintezu acetilkolina, a piretroidi djeluju na živčani sustav artropoda. Kao što je prije rečeno, akaricidna sredstava veoma lagano razviju rezistenciju i zbog toga treba raditi rotaciju pri-pravaka. Zasad se dobrim pokazalo korištenje po jednog pripravka iz svake skupine. Primjerice, jednom tretiramo organofosfornim pripravkom, zatim karbamatnim i nakon
njega nekim piretroidom. Treba naglasiti da se špricanje akaricida u tijeku proizvodnje obnaša u nastambi u kojoj nema jaja i hrane za nesilice poradi njihove moguće konta-minacije. U posljednje vrijeme sve se više u suzbijanju tekuti koristi dijatomejska zemlja ili kako je još zovu inertna prašina (amorfni silicijski dioksid). Naime, radi se o geološkom depozitu nastalom taloženjem jednostaničnih biljnih organi-zama u slatkoj ili slanoj vodi, koji se usitnjava u prašak i kao takav prodaje. On se raspršuje na opremu i životinje te lijepi na insekte. Pritom uništava im vanjski sloj abrazijom i adsorpcijom, pa tako načet insekt dehidrira (Kilpinen i Steenberg, 2009.). Isti autori utvrdili su da korištenjem inert-ne prašine 50% tekuti ugine u prva 24 sata. Dijatomejska zemlja potpuno je neotrovna za ljude i životinje, a zasad rezistencija nije primijećena. Pripravci u obliku pudera mo-gu iritirati kožu, no to nije slučaj ako se koristi gel ili tekućina. Učinkovitost postupka ovisna je o kakvoći dijato-mejske zemlje, okolišnim čimbenicima i količini pripravka na tretiranoj površini. Suzbijanje tekuti korištenjem visoke temperature (iznad 45 ºC kroz pet dana ili 52 ºC kroz dva dana) pokazalo se vrlo učinkovitim. To je postupak kojim se koriste u Nizozemskoj i Norveškoj, a primjenjuje se u nastambama za vrijeme me-đuturnusne stanke. Prema njihovim iskustvima nakon tako provedenog tretmana u novom proizvodnom ciklusu nisu imali problema s tekuti. Naročito dobri rezultati postižu se ako se uz visoku temperaturu koristi neki akaricid. Tako primjerice u Norveškoj primjenjuju toplinu i akaricidno sredstvo, pa nisu imali reinfekcija u tretiranim nastambama. Nizozemska koja primjenjuje samo toplinski tretman u više je navrata imala ponovne invazije tekuti u tijeku proiz-vodnje. No, ovo treba uzeti s rezervom, jer uzrok tome može biti u nemogućnosti postizanja visoke temperature u cijeloj nastambi te velikoj gustoći i blizini ostalih peradnjaka. Inače valja reći da je korištenje visokih temperatura (toplinski tretman) dosta skupo i nosi sa sobom rizik oštećenja opreme. Zaključak stručnjaka iz Nizozemske ipak je da bi bilo dobro uz toplinu koristiti i akaricide, budući da uvijek postoji mo-gućnost preživljavanja tekuti kod samo toplinskog tretmana. Naime, teško je unutar cijele nastambe postići željenu temperaturu koja će ubiti tekuti. Može se također spomenuti i svrsishodnost korištenja niskih temperatura u suzbijanju tekuti. Radi se o temperaturama ispod 10 ºC, no treba reći da rezultati tog postupka nisu dobri kao pri korištenju visokih temperatura. Također je utvrđeno da su mala i visoka rela-tivna vlažnost zraka ograničavajući čimbenik jačeg širenja tekuti (Migeon i sur., 2009.), no to nema neke važnosti za peradarsku praksu, jer je to samo povremeni vanjski čim-benik koji je dobrodošao u rješavanju tog problema. Ponekad je dobro, kao što rade u nekim zemljama, dizaj-nirati unutrašnjost nastambi tako da na utvrđene migracijske puteve tekuti (od skrovišta do kokoši) postavimo prikladne barijere na koje se nanese akaricid, esencijalno ulje ili sili-katna zemlja, što će spriječiti tekut da dođe do kokoši tijekom noći.

TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE
83
Alternativne metode
Dobra alternativna metoda za suzbijanje i širenje tekuti unu-tar nastambe je primjena modificiranog programa osvijetlje-nosti. Istraživači u Belgiji, koristeći program u kojem je bilo 15 minuta svjetla i 45 minuta mraka, uspjeli su znatno smanjiti populaciju tekuti u peradnjacima (Zoons, 2004.). Takav učinak ostvaren je u još nekim zemljama, mada su neki izvijestili da on nestaje kroz neko vrijeme nakon nje-gove primjene. Ostaje nejasno na koji način ovakav program osvijetljenosti djeluje na tekut. Moguće je da tako određen ritam osvjetljenja inhibira aktivnost tekuti, te ona ima premalo vremena za doći do kokoši ili naći skrovište i tada postaje laka meta za kokoš da je pojede. No, s ovakvim programom svjetla ima jedan problem. Naime, direktiva EU nalaže da kokoši nesilice moraju imati konstantan mrak od osam sati, a neke druge mogućnosti s primjenom svjetla još nisu istražene. Druga alternativna metoda odnosi se na korištenje sredstava koja odbijaju tekuti (repelenti). Francuski istraživači izvješ-ćuju da rezultati primjene tih sredstava nisu uvijek predvid-ljivi, što je zbunjujuće. Zapravo, ponkad su rezultati dobri, a ponekad ne. Sigurno je da mirisi mogu ostvariti neki povo-ljan učinak u borbi protiv tekuti, ali treba pronaći ispravan koncept u njihovoj primjeni, a on bi se trebao temeljiti na iskustvu oko primjene pripravka te poznavanju biologije štetnika. Dosad su u uporabi repelenti na bazi češnjaka, korijandera, timijana, cimeta, spearminta, hrena, klinčića itd. Pripravci na toj osnovi daju se kroz hranu ili se nanose na opremu u okolišu nesilica. Prirodni akaricidi uključuju biljne akaricide, to su esenci-jalna ulja, trave i biljni ekstrakti, a oni sadrže kemijske kom-ponente koje ubijaju tekut (George i sur., 2008a., 2008b.; Maurer i sur., 2009.). No, budući da koncentracija otrova u njima može biti različita s obzirom na uvjete rasta i pod-neblja, treba biti vrlo pažljiv u izradi djelatne otopine, jer ne treba zaboraviti na moguće opasno djelovanje iste na ljude i životinje te na okoliš. Drugi su bioakaricidi i zapravo radi se o bakterijama i njihovim toksinima koje ubojito djeluju na insekte. To su primjerice Bacillus thuringiensis (Hassanain i sur., 1997.), Saccharopolyspora spinosa (Anastas i sur., 1999.) i Boophilus microplus (Davey i sur., 2001.). Posebice se od njih kao vrlo učinkoviti ističu spinosadi, produkti fermentacije Saccharopolyspora spinosa, za koje drže da su nova generacija bioakaricida (Harrington i sur., 2011.). Predatori grinja (Lealapidae, Cheyletidae) slijedeća su alter-nativna opcija koja daje ohrabrujuće rezultate. Zapravo ovi predatori (Hypoaspis aculeifer, Androlaelaps casalis i Chey-letus eruditus) okupiraju ista skrovišta kao i tekuti. Pritom ih proždiru i poremećuju njihovo prirodno grupiranje te dina-miku reprodukcije. U izboru predatora mora se paziti da nemaju utjecaja na zdravlje ljudi i životinja i da nanose najviše šteta tekuti (Maurer, 1993.; Lesna i sur., 2009.). Ova su istraživanja još u tijeku i najviše se rade u Nizozemskoj. Za razliku od istraživača u Nizozemskoj, oni u Danskoj i Velikoj Britaniji istražuju entomopatogenične gljive koje su u stanju zaraziti i ubiti grinje (Beauveria bassiana, Metar-
hizium anisopliae, Paecilomyces fumosoroseus). Spore ovih gljiva ulaze preko kutikule domaćina u njihovo tijelo i ubi-jaju ga, a iz njega se spore umnažaju i šire na ostale grinje (Steenberg i sur., 2006.; Tavassoli i sur., 2008.). Vrlo je važno da one nisu opasne za ljude, životinje i jaja te da imaju zadovoljavajući učinak u primjeni (Vestergaard i sur., 2003.). Odabrane gljive moraju preživjeti mikroklimu u pe-radnjaku, jer se pokazalo da visoka razina amonijaka kao i metalna podloga može inhibirati njihov rast (Hong i sur., 2005.). Zasad se još radi na odabiru najboljih vrsta gljiva za tu namjenu. Dosta velika očekivanja imamo od cijepljenja protiv tekuti. Istraživanja su još u tijeku (Arkle i sur., 2008.), a radi se na odabiru najboljeg antigena iz proteinskog sastava tekuti koji će u cijepljenih kokoši izazvati najbolju tvorbu protutijela za uništavanje tekuti nakon sisanja krvi. Zasad je utvrđeno uništenje do 30% populacije tekuti, no istraživači vjeruju da će se izborom najboljeg antigena taj učinak bitno popraviti.
Zaključak
Možemo zaključiti da ima dosta toga nepoznatog iz života i reprodukcije tekuti, što je i glavni razlog zbog čega ju ne možemo iskorijeniti (Mul i sur., 2009.). No, postoje rješenja kojima možemo održavati populaciju grinja na podnošljivoj razini. Težište borbe treba biti na preventivnim mjerama, provedbi biosigurnosti u farmama i dobro tempiranom mo-nitoringu, a u zahvaćenim jatima valja koristiti kombinaciju više konvencionalnih i alternativnih metoda suzbijanja (Harrington i sur., 2011.). Zasad najbolji učinak ima uporaba visokih temperatura tijekom sanacije nastambi u kombinaciji s kemijskim akaricidom, a u skoro vrijeme treba računati s uporabom entomopatogeničnih gljiva, bioakaricida, preda-tora tekuti i cijepljenjem (Mul i sur., 2009.).
Literatura
ANASTAS, P., M. KIRCHCHOFF, T. WILLIAMSON (1999): Spinosad, a new natural product for insect control. Green Chemistry, August 1999:G88.
ARKLE, S. (2007): Development of a vaccine against the poultry red mite (Dermanyssus gallinae), PhD Thesis, Newcastle University, UK.
ARKLE, S., D. HARRINGTON, C. De LUNA, D. GEORGE, J. GUY, A. E. SPARAGANO (2008): Immunological control of poultry red mite: the use of whole mite antigens as a candidate vaccine. Ann. N. Y. Acad. Sci. 1149, 36-40.
AXTELL, R.C., J. J. ARENDS (1990): Ecology and management of arthropod pests of poultry. Annu. Rev. Entomol. 35, 101-126.
BEUGNETS, F., C. CHAUVE, M. GAUTHEY, L. BEERT (1997): Resistance of the poultry mite to pyrethroids in France. Vet. Rec. 140, 577-579.
BIĐIN, Z. (2008): Bolesti peradi. Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, 2008.
CHAUVE, C. (1998): The poultry red mite Dermanyssus gallinae (De Geer, 1778): current situation and future prospects for control. Vet. Parasitol. 76, 239-245.

TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE 84
CHIRICO, J., R. TAUSON (2002): Traps containing acaricides for the control of Dermanyssus gallinae. Vet. Parasitol. 110, 109-116.
DAVEY, R. B., J. E. GEORGE, D. E. SNYDER (2001): Efficacy of a single whole body spray treatment of spinosad against Boophilus microplus on cattle. Vet. Parasitol. 99, 41-52.
De LUNA, C., C. V. MORO, J. GUY, L. ZENNER, O. SPARAGANO (2009): Endosymbiotic bacteria living inside the poultry red mite. Exp. Appl. Acarol. 48, 91-99.
EMOUS, R. A., T. G. NIEKERK, M. F. MUL (2005): 11 milion € damage for the sector: enquiry into the cost of mite to the poultry industry in Dutch. De pluimveehouderij, 35, 8-9.
ENTREKIN, D. L., J. H. OLIVER (1982): Aggregation of the chicken mite, Dermanyssus gallinae. J. Med. Entomol. 19, 671-678.
GEORGE, D., K. CALAGHAN, J. H. GUY, O. A. E. SPARAGANO (2008a): Lack of prolonged activity of lavander essential oils as acaricides against the poultry red mite under laboratory conditions. Res. Vet. Sci. 85, 540-542.
GEORGE, D., T. J. SMITH, O. A. E. SPARAGANO, J. H. GUY (2008b): The influence of time since last blood meal on the toxicity of essential oils to the poultry red mite. Vet. Parasitol. 155, 333-335.
GUY, J. H., M. KHAJAVI, M. M. HLALELE, O. SPARAGANO (2004): Red mite prevalence in laying units in Northern England. Br. Poult. Sci. 45, S15-S16.
HAMSCHER, G., B. PRIESS, J. HARTUNG, M. I. NOGOSSEK, G. GLUENDER, H. NAU (2003): Determination of propoxur residues in eggs by liquid chromatography diode array detection after treatment of stocked housing facilities for the poultry red mite. Anal. Chim. Acta 483, 19-26.
HARRINGTON, D. W. J., D. R. GEORGE, J. H. GUY, O. A. E. SPARAGANO (2011): Opportunities for integrated pest management to control the poultry red mite. World Poult. Sci. J. 67, 83-92.
HASSANAIN, M. A., M. F. E. GARHY, F. A. ABDEL-GHAFFAR, A. El SHARABY, K. N. A. MEGEED (1997): Biological control studies of soft and hard ticks in Egypt. Parasitol. Res. 83, 209-213.
HOGLUND, J., H. NORDENFORS, A. UGGLA (1995): Preva-lence of the poultry red mite in different types of production system for egg layers in Sweden. Poultry Sci. 74, 1793-1798.
HONG, T. D., S. EDGINGTON, R. H. ELLIS, M. AQUINO de MURO, D. MOORE (2005): Saturated salt solutions for humi-dity control and the survival of dry powder and oil formula-tions of Beauveria bassiana conidia. J. Invertebr. Pathol. 89, 136-143.
KILPINEN, O. (2001): Activation of the poultry red mite, by increasing temperatures. Exp. Appl. Acarol. 25, 859-867.
KILPINEN, O., B. A. MULENS (2004): Effect of food deprivation on response of the mite Dermanyssus gallinae, to heat. Med. Vet. Entomol. 18, 368-371
KILPINEN, O. (2005): How to obtain a blood meal without being eaten by the host: the case of the poultry red mite. Physiol. Entomol. 30, 232-240.
KILPINEN, O., A. ROEPSTORFF, A. PERMIN, G. NORGAARD NIELSEN, L. G. LAWSON, H. B. SIMONSEN (2005): Influence of Dermanyssus gallinae and Ascaridia galli infections on behaviour and health of laying hens. Br. Poult. Sci. 46, 2634-2638.
KILPINEN, O., T. STEENBERG (2009): Inert dusts and their effects on the poultry red mite. Exp. Appl. Acarol. 48, 51-62.
KIRKWOOD, A. C. (1968): Some observations on the feeding habits of the poultry mites Dermanyssus gallinae and Liponyssus sylviarum. Entomol. Exp. Appl. 11, 315-320.
LESNA, I., P. WOLFS, F. FARANJI, L. ROY, J. KOMDEUR, M. W. SABELIS (2009): Candidate predators for biological control of the poultry red mite. Exp. Appl. Acarol.48, 63-80.
MARANGI, M. A.CAFIERO, G. CAPELLI, A. CAMARDA, O. A. E. SPARAGANO, A. GIANGASPERO (2009): Evaluation of poultry red mite susceptibility to some acaricides in a field population from Italy. Exp. Appl. Acarol.48, 11-18.
MAURER, V.(1993): The dynamics of Dermanyssus gallinae populations interacting with laying hens and the predatory mite Cheyletus eruditus (Acari: Cheyletidae). Dissertation ETHZ No. 10330, 104.
MAURER, V., J. BAUMGARTNER (1992): Temperature influence on life table statistics of the chicken mite. Exp. Appl. Acarol.15, 27-40.
MAURER, V., M. BIERI, D. W. FOLSCH (1988): Host finding of Dermanyssus gallinae in poultry houses. Arch. Geflugelkd. 52, 209-215.
MAURER, V., E. PERLER, F. HECKENDORN (2009): In vitro efficacies of oils, silicas and plant preparations against the poultry red mite. Exp. Appl. Acarol.48, 31-41.
MIGEON, A., F. FERRAGUT, K. FIABOE, M. KNAPP, M. NAVAJAS (2009): Modelling the potential distribution of the invasive tomato red spider mite, Tetranycus evansi. Exp. Appl. Acarol.48, 199-212.
MORO, C. V., C. De LUNA, T. ALEXANDER, J. GUY, A. E. OLIVER (2009): The poultry red mite: potential vector of pathogenic agents. Exp. Appl. Acarol. 48,93-104.
MUL,M., T. VANNIEKERK, J. CHIRICO, V. MAURER, O. KILPINEN, O. SPARAGANO, B. THIND, J. ZOONS, D. MOORE, B. BELL, A. GJEVRE, C. CHAUVE (2009): Control method for Dermanyssus gallinae in system for laying hens: results o fan international seminar. World Poult. Sci. J. 65, 589-600.
NORDENFORS, H., J. HOGLUND (2000): Long term dynamics of Dermanyssus gallinae in relation to mite control measures in aviary system for layers. Br. Poult. Sci. 41, 533-540.
NORDENFORS, H., J. CHIRICO (2001): Evaluation of a sampling trap for Dermanyssus gallinae. J. Econ. Entomol. 94, 1617-1621.
NORDENFORS, H., J. HOGLUND, A. UGGLA (1999): Effects of temperature and humidity on oviposition, molting and longe-vity of Dermanyssus gallinae. J. Med. Entomol. 36, 68-72.
NORDENFORS, V., J. HOGLUND, R. TAUSON, J. CHIRICO (2001): Effect of permethrin impregnated plastic strips on Dermanyssus gallinae in loose housing systems for laying hens. Vet. Parasitol. 102, 121-131.
OLIVER, J. H. Jr.(1965): Karyotypes and sex determination in some Dermanyssid mites. Ann. Entomol. Soc. Am. 58, 567-573.
POTENZA.L., M. ASSUNTACAFIERO, L. CUCCHIARINI, G. LASALANDRA, A. GIANGASPERO, O. A. E. SPA-RAGANO, A. CAMARDA (2008): Dermanyssus gallinae mites collected from pigeon nests and laying hens: a molecular study based on the ITS region. BSP Spring. Trypanosomi-asis/Leishmaniasis and Malaria Meetings. March 30 to April 2, Newcastle upon Tyne, UK. 169.

TEKUT, VJEČNI PRATITELJ PERADARSKE PROIZVODNJE
85
SAHIBI, H., O. A. E. SPARAGANO, A. RHALEM (2008): Dermanyssus gallinae: Acari parasite highly aggressive but still ignored in Marocco. BSP Spring. Trypanosomiasis/Leish-maniasis and Malaria Meetings. March 30 to April 2, Newcastle upon Tyne, UK.,173.
SIKES, R. K., R. W. CHAMBERLAIN (1954): Laboratory obser-vations on three species of bird mites. J. Parasitol. 40, 691-697.
STEENBERG, T., O. KLIPINEN, J. B. JESPERSEN, M. D. SOLERCRUZ, C. VEGAROBLES, M. BIRKETT, S. DEWHIRST, J. PICKETT (2005): Advances in the develop-ment of novel control methods against chicken mites. Bulletin-OILB/SROP 28:89-91.
TAVASSOLI, M., A. OWNAG, S. H. POURSEYED, K. MARDANI (2008): Laboratory evaluation of three strains of the entomopathogenic fungus Metarhizium anisopliae for controlling Dermanyssus gallinae. Avian Pathol. 37, 259-263.
THIND, B. B., H. L. FORD (2007): Assessment of susceptibility of the poultry red mite to some acarides using an adapted filter paper based bioassay. Vet. Parasitol. 144, 344-348.
VESTERGAARD, S., A. CHERRY, S. KELLER, M. GOETTEL (2003): Safety of Hyphomycete fungi as microbial control agents in: Environmental Impacts of Microbial Insecticides (H. M. T. Hokkanen, A. E. Hajek, ur.) 32-62.
WOOD, H. P. (1917): The chicken mite: its life history and habits. United States Department of Agriculture, Bulletin 553.
ZEMAN, P.(1988): Surface skin lipids of birds, a proper host kairomone and feeding inducer in the poultry red mite. Exp. Appl. Acarol. 5, 163-173.
ZOONS, J. (2004): The effects of light programs on red mite in battery cage housing. in: Welfare of the Laying Hen (G. C. Perry, Ed.), 416.
POULTRY RED MITE EVER ACCOMPANYING POULTRY PRODUCTION Summary The state-of-the-art and procedures used in poultry red mite control, as well as the respective studies (red mite biopredators, vaccination, entomopathogenic fungi, etc.) the results of which will provide a basis for developing a program of their eradication in the near future, are presented. Red mite invading poultry housings results in health and productivity problems in chicken, which manifest as great economic losses at the end of the production cycle. The biology of mites is not yet fully understood, which is the cause of inefficient mite control. Currently, poultry keepers mostly use chemical acaricides, since they believe it to be the most efficient method of poultry mite control; however, it usually proves unsuccessful because these products can hardly reach mites, which slip to hidden sites or deep under the host's feathering. It is therefore necessary to use a combination of several conventional and alternative methods of mite control to achieve some success and to reduce mite count to an acceptable level, thus also reducing the ensuing losses. Namely, as long as the hematopoietic system of the animal can compensate for the blood loss caused by mite blood sucking, the damage remains within acceptable limits; however, when the mite count increases drastically, then both health and production problems occur. Thus, strict control should be performed throughout the production cycle to enable timely intervention if needed. Key words: poultry red mite, prevention, therapy

PERADARSKI DANI 2013. 86
PTIČJI HEPATITIS E – POTENCIJALNA OPASNOST ZA PERADARSKU PROIZVODNJU?!
Tajana Amšel Zelenika1, Jelena Prpić2, Marina Tišljar1, Vladimir Savić1, Mirta Balenović1, Luka Jurinović1, Lorena Jemeršić2
1Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska 2Odjel za virologiju, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak
Gubitci u peradarskoj proizvodnji često su uzrokovani padom nesivosti, kasnijim spolnim dozrijevanjem kokoši, lošom kvalitetom ljuske jaja itd. Jedan od mogućih, ali vrlo rijetko spominjanih uzroka može biti i hepatitis E. Zbog sličnosti sa simptomima zaraznog bronhitisa, sindroma pada nesivosti, tehnološkim pogreškama u odgoju pilenki i sl. bolest može biti zanemarena i neuspješno liječena simptomatski. Stoga je cilj ovoga rada prikazati osnovne značajke bolesti. U radu su prikazani i rezultati preliminarnog istraživanja pojavnosti virusa u organima kokoši koje su patološko-anatomskom pretragom pokazivale simptome hepatitisa E. Ključne riječi: hepatitis E, hepevirus, kokoš Uvod Bolesti jetre i slezene često prate intenzivnu peradarsku proizvodnju. No, osim u slučajevima velikog pomora kao što je kod Marekove bolesti, limfoidne leukoze ili bakte-rijskih septikemija koje se mogu dokazati na osnovi spe-cifične patohistološke slike, uzročnik takvih bolesti naj-češće ostane neistražen. Najnovija istraživanja u svijetu ukazuju na to da se u mno-gim slučajevima bolesti kod kojih je uočena punokrvnost i nekroza jetre i slezene, tzv. HSS (engl. hepatitis-spleno-megaly syndrome) ili kod povećanja jetre i slezene, BLSD (engl. big liver and spleen disease) zapravo radi o vi-rusnom hepatitisu E (Haqshenas i sur., 2001.; Agunos i sur., 2006.; Payn i sur.,1999.). Bolest je raširena diljem svijeta. Nakon prvih opisa bolesti u Australiji i Americi infekcija hepatitisa E virusom (HEV) dokazana je i u Italiji (Massi i sur., 2005.), Kanadi (Agunos i sur., 2006.), Mađarskoj (Morrow i sur., 2008.), Kini (Zhang i sur., 2008.), Španjolskoj (Peralta i sur., 2009.) i Rusiji (Irza i Sprygin , 2012.). Uzročnik bolesti je RNK virus iz roda Hepevirus (HEV), porodice Hepeviridae (Emerson i sur., 2004.).
Slika 1. Ptičji hepatitis E virus 1.012x1325 pix (Izvor:
http://3dciencia.com/blog/wp-content/uploads/ /2011/02/HEV-hepatitis-E-virus-hepevirus-capsid-small-non-enveloped-particles.jpg , 16.01.2013.)
Dr. sc. Tajana Amšel Zelenika, Hrvatski veterinarski institut, Podružnica Centar za peradarstvo, Heinzelova 55, Zagreb, Hrvatska. Tel: 385(0)1 2441-394. Fax: 385(0)1 2441-396. E-mail: [email protected].

PTIČJI HEPATITIS E – POTENCIJALNA OPASNOST ZA PERADARSKU PROIZVODNJU?!
87
Virusni hepatitis E opisan je u ljudi te mnogih vrsta do-maćih i divljih životinja te peradi. Poznato je 5 genetskih skupina virusa. Genotipovi 1 i 2 izdvojeni su iz ljudi, dok su genotipovi 3 i 4 izdvojeni iz ljudi i raznih vrsta sisavaca. Ptičji HEV svrstan je u zasebnu 5. genotipsku skupinu i samo se u 50% nukleotida podudara s ostalim genotipo-vima HEV-a (Huang i sur., 2004.; Huang i sur., 2005.; Meng, 2010.). Sekvenciranjem genoma i filogenetskom analizom utvrđena je heterogenost unutar ptičjeg soja HEV (Sun i sur., 2004b.). Za razliku od svinjskih izolata ptičji HEV ne posjeduje zoonotski potencijal i ne može inficirati druge primate (Meng i sur., 1998.a,b; Meng, 2000.a,b; Tei i sur., 2003.; Huang i sur., 2004.). Kokoš je jedini prirodni domaćin ptičjeg HEV-a, iako su pokusno zaraženi purani u dobi od 8 tjedana razvili vire-miju i pokazivali znakove virusnog proljeva. Isto tako virus je dokazan i u intravenski zaraženih kokošjih embrija (Billiam i sur., 2005.; Huang i sur., 2005.; Sun i sur., 2004.; Payne i sur., 1993.).
Slika 2. Subkapsularna krvarenja jetre u pokusno zaražene
kokoši. (Izvor: Journal of Virology, jvi.asm.org/ /content/79/6/3429.full, 16.01.2013).
Glavni izvor infekcije je feces zaraženih kokoši. Bolest se prenosi oralno fekalnim putem, dok vertikalni prijenos nije zabilježen (Huang i sur., 2002.; Guo i sur., 2007.). Pilići su uspješno zaraženi i pokusnom okulonazalnom infekcijom (Billiam i sur., 2005.). Inkubacija bolesti traje od 1 do 3 tjedna, no bolest često prolazi bez vidljivih simptoma, a uginuća su relativno rijetka (Huang i sur., 2002.). U pojedinim jatima nesilica u kojih se očituje sindrom HS zabilježen je pad nesivosti do 20% i to najčešće u jatima dobi od 30-72 tjedna. U mlađe peradi bolest u pravilu ima subklinički tijek (Sun i sur., 2004.a,b). Sve dobne skupine kokoši osjetljive su na BLS, ali klinički simptomi vidljivi su pretežno u nesilica starih 24 tjedna.
Uginuća nisu znatna, no pad nesivosti može doseći do 20%. Obično je nagli pad nesivosti i prvi znak infekcije HEV-om. Infekcija pilenki najčešće rezultira kasnijim spolnim dozrijevanjem, nesilice ne dosežu očekivanu pro-izvodnju jaja, jaja su mala s tankom i slabo pigmentiranom ljuskom. U starijih nesilica pad nesivosti traje oko 3-6 tjedana i još je toliko vremena potrebno za oporavak. Ponekad se javlja letargija, anoreksija i gubitak perja (McFerran, 1994.).
Slika 3. Infiltracija limfocita i segmentni flebitis u portal-nom traktu. (Izvor: http://www.worldpoultry.net/ /Broilers/Health/2013/1/Avian-hepatitis-E-virus-in-fection-in-chickens-Part-2-1137296W/, 16.01.2013.)
U peradi koja je uginula nakon infekcije ptičjim HEV-om često se primijete atrofični jajnici, krvarenja u abdomenu te povećanje jetre koja biva prekrivena krvarenjima s mogu-ćom pojavom subkapsularnih hematoma, dok je slezena blago do umjereno povećana.
Slika 4. Mikroskopske lezije u slezeni. Limfoidna hiperplazija
u pokusno inficirane kokoši. (Izvor: Journal of Viro-logy, jvi.asm.org/content/79/6/3429.full, 16.01.2013.)
Mikroskopski, lezije na jetri variraju od multifokalnih krvarenja do opsežnih područja nekroze, te infiltracije

PTIČJI HEPATITIS E – POTENCIJALNA OPASNOST ZA PERADARSKU PROIZVODNJU?! 88
heterofila i mononuklearnih upalnih stanica oko portalnog trakta (Meng i sur., 2008.). Na jetri se često javljaju gra-nulomi, a povremeno i tromboza portalne vene. U pokusno inficirane peradi patološko anatomski uočene su i promje-ne na timusu u vidu blage kortikalne hipoplazije timusa, te blaga upala bubrega i pluća (Billiam i sur., 2005.). Česta pojava su i peritonitis i kronični bakterijski hepatitis, što upućuje na moguće imunosupresivno djelovanje virusa HE (Morrow i sur., 2008.). Sumnja na pojavu hepatitisa E (HE) postavlja se temeljem kliničkih simptoma i patološko histološkog nalaza. Virus ptičjeg HE vrlo je teško izolirati. Elektronskim mikro-skopom moguće je dokazati virusnu česticu veličine 30-50 nm u žuči inficiranih ptica, što može potvrditi sumnju na infekciju HEV. Trenutno se dijagnoza postavlja na temelju dokaza virusne RNA metodom lančane reakcije polimera-zom uz prethodnu reverznu transkripciju (RT-PCR, engl. reverse transcription polymerase chain reaction) ili do-kazom protutitjela imunoenzimskom probom (ELISA, engl. enzyme-linked immunosorbent assay). Međutim, osjetljivost i specifičnost ovih metoda još je neistražena. Naime, još ne postoje komercijalni kitovi za dokaz pro-tutijela postupkom ELISA. Serološki moguć je dokaz i imunodifuzijom u gelu upotrebom pročišćenog antigena dobivenog iz slezene i jetre bolesnih pilića (Payne i sur. 1999.; Todd i sur., 1993.). Međutim, sam serološki dokaz protutijela nije dovoljan za dokaz infekcije, jer se u akutno inficiranih pilića viremija i širenje virusa fekalnim putem javlja znatno ranije nego organizam odgovori stvaranjem IgG protutijela. Istraživanjem serološki pozitivnih jata u SAD-u i Španjolskoj utvrđeno je da se virus može dokazati u 15%-100% pilića (Huang i sur. 2002.; Peralta i sur., 2009.). Zbog svega navedenog dijagnoza se danas temelji na me-todi RT-PCR, premda nisu utvrđene univerzalne PCR početnice koje bi prepoznale sve genetske varijacije unutar sojeva ptičjeg HEV (Huang i sur., 2002.; Billam i sur., 2005.). Diferencijalno dijagnostički HE treba razlikovati od bolesti masne jetre (patoanatomski je vidljivo nakupljanje krvi u abdominalnoj šupljini, kao i krvarenja na jetri), trovanja rodenticidima (antikoagulansima), traume i sl. Nešto je lakše HE razlučiti od Marekove bolesti, limfodine leukoze ili bakterijske septikemije koje imaju karakteristične histo-loške promjene. Ne postoji učinkovit lijek za hepatitis E. Prevencija se sastoji od pojačanih mjera biosigurnosti na farmama. Istra-žuje se mogućnost preventivnog cijepljenja, no do danas na tržištu nema komercijalnog cjepiva. Cilj našega istraživanja i izlaganja je upozoriti držaoce pe-radi na bolest o kojoj se vrlo malo govori, a potencijalna je opasnost za značajne gubitke u proizvodnji jaja. S obzirom na to da Hrvatski veterinarski institut, Laboratorij za dijag-nostiku klasične svinjske kuge, molekularnu virologiju i genetiku od 2009. godine istražuje učestalost virusa HE u domaćih i divljih životinja (Jemeršić i sur., 2010.; Jemeršić i sur., 2011.; Prpić i sur., 2012.), uspostavljena je suradnja i započeto je istraživanje učestalosti virusa HE u peradi. U
istraživanje su se uključili Laboratorij za patologiju i La-boratorij za virusologiju i serologiju Centra za peradarstvo. U ovom radu ćemo prikazati rezultate dosadašnjeg istra-živanja.
Materijali i metode
Tijekom 2011. i 2012. godine prikupljane su jetre i slezene kokoši različite dobi i načina držanja. Svi organi prikup-ljeni su od životinja u kojih su patološko-anatomskom pretragom dokazane promjene u jetri i slezeni.
RT-PCR
Uzorke organa pretražili smo metodom RT-PCR. Virusna RNK izolirana je prema protokolu kompleta QIAamp® Viral RNA Kit (Qiagen, Njemačka). Postupak reverzne transkripcije obavljen je kompletom za reverznu transkripciju SuperScriptTM III First-Strand Syn-thesis System for RT-PCR (Invitrogen, SAD) pomoću uređaja Gene Amp PCR System 9700 (Applied Bio-systems, SAD). Odsječci gena za kapsidni protein i gena za helikazu umnoženi su lančanom reakcijom polimeraze. Korištena su dva para početnica, jedan specifičan za odsje-čak gena za kapsidni protein, drugi specifičan za odsječak gena za helikazu (Huang i sur., 2002.; Bilic i sur., 2009.). Tablica 1. Reakcijski ciklusi umnožavanja odsječka gena za
kapsidni protein i gena za helikazu
Naziv koraka Vrijeme/temperatura Početna denaturacija 3 min/94 °C Denaturacija 30 s/94 °C Sparivanje početnica i kalupa (annealing) 30 s/57 °C
Produljenje lanca
35 ciklusa umnožavanja
1 min/72 °C Završno produljenje lanca 10 min/72 °C Elektroforeza je izvedena u 1,5%-tnom agaroznom gelu veličine 100x115x40 mm (širina x duljina x visina) prire-đenom u 1xTBE-puferu. Elektroforeza je provedena tije-kom 60 minuta pri sobnoj temperaturi uz primjenu napona od 70 V.
Rezultati
Tablica 2. Patološko anatomskom pretragom dokazano je:
Patološko anatomska dijagnoza Broj lešina Ascites 16 Hepatomegalija 13 Supkapsularna krvarenja u slezeni 1 Hiperplastični tumor slezene 7 Nekrotični hepatitis 2

PTIČJI HEPATITIS E – POTENCIJALNA OPASNOST ZA PERADARSKU PROIZVODNJU?!
89
Rezultati pretraživanja na prisutnost RNK virusa hepatitisa E u svim pretraženim organima dali su negativni rezultat.
Rasprava
Na farmama nesilica diljem Hrvatske česta je pojava pada nesivosti, kasnije spolno dozrijevanje kokoši, loša kvaliteta ljuske jaja itd. S obzirom na veliku sličnost sa simptomima zaraznog bronhitisa, sindromom pada nesivosti, tehno-loškim pogreškama u odgoju pilenki i sl. često se bolest zanemari i liječi simptomatski. No, takova slika bolesti velikim dijelom može biti uzrokovana i virusom hepatitisa E, tim više što je HE dokazan u mnogim zemljama uklju-čujući i Mađarsku i Italiju s kojima graničimo. Morrow i sur. (2008.) opisuju dokaz HEV u farmama nesilica u Mađarskoj. Jata u kojima je dokazan HEV pokazivala su simptome pada nesivosti nakon postizanja vrha proiz-vodnje, depresiju, djelomično mitarenje, povećanu jetru i slezenu, blago povišeno uginuće i krvarenja u abdomenu. Česta je pojava peritonitisa, kroničnog bakterijskog hepa-titisa i ascitesa u prsnoj šupljini. Isti simptomi opisani su i u Sjevernoj Americi i Italiji (Huang i sur., 2002.; Massi i sur., 2005.). Jata nesilica čija je tjelesna težina odgovarala uzgojnim standardima vratila su proizvodnju za otprilike mjesec dana, no jata s nesilicama nešto veće tjelesne težine imala su dugotrajniji pad nesivosti. U Australiji bolest je prouzročila veće ekonomske gubitke u teških nesilica gdje je gubitak iznosio oko 8 jaja po kokoši u 50% jata. Gubitak je procijenjen na 2,8 milijuna dolara (Payne i sur., 1999.). Negativan nalaz našega istraživanja ni u kojem slučaju ne može dovesti do zaključka da hepatitis E nije prisutan u kokoši u Hrvatskoj. Naime, loša gospodarska situacija i teške prilike u kojima se našlo hrvatsko peradarstvo rezul-tirale su sve manjim istraživanjem pojava bolesti na farma-ma. Za primjer ćemo reći da je u 2005. godini u Labora-toriju za patologiju Centra za peradarstvo pretraženo ukupno 547 nesilica (390 lakih i 157 teške pasmine), a tijekom 2011. godine samo 95 nesilica (21 laka i 74 teške pasmine). Dijagnostika se ograničila na minimum pretraga koje su nužne prema zakonu tj. Naredbi. Uzrok sve ma-njem broju patološko anatomski pretraženih lešina u Cen-tru za peradarstvo ne leži u sve zdravijoj i otpornijoj peradi. Veliki troškovi uzgoja peradi i održavanja farmi, nažalost, natjerali su mnoge peradare da svoje troškove svedu samo na one neophodno potrebne, pa se mnoštvo problema na farmama uzrokovanih bolestima liječe simp-tomatski ili se u potpunosti previde. Tako i organi koje smo uzorkovali, iako su pokazivali patološko anatomsku sliku koja ukazuje na bolesti jetre i slezene koje mogu biti uzrokovane HEV, ne predstavljaju reprezentativni uzorak. Zato je i cilj našeg istraživanja i izlaganja bio upozoriti držaoce peradi na bolest o kojoj se mnogo ne govori, a potencijalna je opasnost za značajne gubitke u proizvodnji jaja.
Zahvala
Istraživanje je financiralo Ministarstvo znanosti, obrazo-vanja i športa Republike Hrvatske, znanstveni projekt
"Primjena genetičkih analiza u veterinarskoj medicini" br. 048-0481186-1183.
Literatura
AGUNOS, A. C., D. YOO, S. A. YOUSSEF, D. RAN, B. BINNINGHTON, D. B. HUNTER (2006): Avian hepatitis E virus in an outbreak of hepatitis-splenomegaly syndrome and fatty liver haemorrhage syndrome in two flaxseed-fed layer in Ontario. Avian Pathol. 35, 404-412.
BILIC, I., B. JASKULASKA, A. BASIC, C. J. MORROW, M. HESS (2009) Sequence analysis and comparison of avian hepatitis E virus from Australia and Europe indicate the existence of different genotypes. J. Gen. Virol. 90, 863-873.
BILLAM, P., F. HUANG, Z. F. SUN, F. W. PIERSON, R. B. DUNCAN, F. ELVINGER, D. K. GUENETTE, T. E. TOTH, X. J. MENG (2005): Systematic pathogenesis and replication of avian hepatitis E virus in specific – pathogen-free adult chickens. J. Virol. 79, 3429-3437.
EMERSON, S. U., D. ANDERSON, A. ARANKALLE, X. J. MENG, M. PURDY, G. G. SCHLAUDER, S. A. TSAREV (2004): Hepevirus. U Virus Taxonomy, VIIIth report of the ICTV. Fauqet C.M., M.A. Mayo, J. Maniloff, U. Dessel-berger, L.A. Ball (ur.), 851-855. Elsevier/Academic Press, London.
GUO, H., E. M. ZHOU, Z. F. SUN, X. J. MENG (2007): Egg whites from eggs of chickens infected experimentally with avian hepatitis E virus contain infectious virus, but evidence of complete vertical transmission is lacking. J. Gen. Virol. 88, 1532-1537.
HAQSHENAS, G., H. L. SHIVAPRASAD, P. R. WOOLCOCK, D. H. READ, X. J. MENG (2001): Genetic identification and characterization of a novel virus related to human hepatitis E virus from chickens with hepatitis-splenomegaly syndrome in the United states. J. Gen. Virol. 82, 2449-2462.
HUANG, F. F., G. HAQSHENAS, H. L. SHIVAPRASAD, D. K. GUENETTE, P. R. WOOLCOCK, C. T. LARSEN, F. W. PIERSON, F. ELVINGER, T. E. TOTH, X. J. MENG (2002): Heterogeneity and seroprevalence of a newly identified avian hepatitis E virus from chickens in the United States. J. Clin. Microbiol. 40, 4197-4202.
HUANG, F. F., Z. F. SUN, S. U. EMERSON, R. H. PURCELL, H. L. SHIVAPRASAD, F. W. PIERSON, T. E. TOTH, X. J. MENG (2004): Determination and analysis of the complete genomic sequence of avian hepatitis E virus (avian HEV) and attempts to infect rhesus monkeys with avian HEV. J. Gen. Virol. 85, 1609-1618.
HUANG, F. F., F. W. PIERSON, T. E., TOTH, X. J. MENG (2005): Construction and characterization of infectious cDNA clones of chicken strain hepatitis E virus (HEV), avian HEV. J. Gen. Virol. 86, 2585-2593.
IRZA, V., A. SPRYNG (2012): Big liver and spleen syndrome is a new viral disease in Russian chicken flocks. J. Agric. Biol. 4, 73-77.
JEMERŠIĆ, L., B. ROIĆ, J. BALATINEC, T. KEROS (2010): Hepatitis E – Da li smo ugroženi? (Hepatitis E – Are we threatened?). Veterinarska stanica, 41(5), 383-397.
JEMERŠIĆ, L., J. BALATINEC, B. ROIĆ, T. KEROS (2011): Detection of hepatitis E virus in domestic pigs and wild boars in Croatia. EcoHealth/Wilcox, Bruce A. (ur.). USA: Springer Science and Business Media, 2011. 144-144. 1st International One World One Health Congress. Melbourne, Australia, February 14-16, 2011.

PTIČJI HEPATITIS E – POTENCIJALNA OPASNOST ZA PERADARSKU PROIZVODNJU?! 90
MASSI, P., G. TOSI, D. GELMETTI, A. LQAVAZZA, G. LOMBARDI, G. TORCOLI (2005): Big liver and spleen disease in broiler breeders in Italy. It. J. Anim. Sci. 4, 303-305.
McFERRAN, J. B. (1994): Big liver and spleen disease. In M. S. McNulty, McFerran, J. B. (ur). New and Evolving Virus Disease in Poultry. Commission of the European Com-munities, Brussels, 299-304.
MENG, X. J., P. G. HALBUR, J. S. HAYNES, T. S. TSAREVA, J. D. BRUNA, R. L. ROYER, R. H. PURCELL, S. U. EMERSON (1998a): Experimental infection of pigs with the newly identified swine hepatitis E virus (swine HEV), but not with human strains of HEV. Arch. Virol. 143, 1405-1415.
MENG, X. J., P. G. HALBUR, M. S. SHAPIRO, S. GOVINDARAJAN, J. D. BRUNA, I. K. MUSHAWAR, R. H. PURCELL, S. U. EMERSON (1998b): Genetic and experimental evidence for cross-species infection by swine hepatitis E virus. J. Virol. 72, 9714-9721.
MENG, X. J. (2000a): Novel strains of hepatitis E virus identified from humans and other animal species: is hepatitis E a zoonosis? J. Hepatol. 33, 842-845.
MENG, X. J. (2000b): Zoonotic and xenozoonotic risks of hepatitis E virus. Infect. Dis. Rev. 2, 35-41.
MENG, X. J., H. L. SHIVAPRASAD, C. PAYNE (2008): Hepatitis E virus infections. U: Saif M. i sur. (ur.) Diseases of Poultry. 12. izd. Blackwell Publishing Press, 443-452.
MENG, X. J. (2010): Hepatitis E virus: animal reservoirs and zoonotic risk. Vet. Microbiol. 140, 256-265.
MORROW, C., S. GYÖZÖ, M. ESZTER, K. AKOS, A. WOOD, S. RICHTER, B. JASKULSKA, M. HESS (2008): Avian hepatitis E virus infection and possible associated clinical disease in broiler breeder flocks in Hungary. Avian Pathol. 37, 527-535.
PAYNE, C. J, S. L. PLANT, T. M. ELLIS, P. V. HILLER, W. HOPKINSON (1993): The detection of big liver and spleen agent in infected tissues via intravenous chick embryo inoculation. Avian Pathol. 22, 245-256.
PAYNE, C. J, T. M. ELLIS, S. L. PLANT, A. R. GREGORY, G. E. WILCOX (1999): Sequence data suggest big liver and
spleen disease virus (BLSV) is genetically related to hepatitis E virus. Vet. Microbiol. 68, 119-125.
PERALTA, B., M. BIARNÉS, G. ORDÓŇEZ, R. PORTA, M. MARTΊN, E. MATEU, S. PINA, X. J. MENG (2009): Evidence of widespread infection of avian hepatitis E virus (avian HEV) in chickens from Spain. Vet. Microbiol. (2009), doi:10.1016/j.vetmic.2008.12.010.
PRPIĆ, J., L. JEMERŠIĆ, S. ČERNI, T. KEROS, D. BRNIĆ, D. ŠKORIĆ (2012): Detection and phylogenetic analysis of hepatitis E virus in Croatian wild boar serum, liver and spleen samples. 5th Croatian Congress of Microbiology with International Participation. p. 54. Primošten, Croatia, 26-30 September 2012.
SUN, Z. F., C. T. LARSEN, F. F. HUANG, P. BILLIAM, F. W. PIERSON, T. E. TOTH, X. J. MANG (2004a): Generation and infectivity titration o fan infectious stock of avian hepatitis E virus (HEV) in chickens and cross-species infection of turkey with avian HEV. J. Clin. Microbiol. 42, 2658-2662.
SUN, Z. F., C. T. LARSEN, A. DUNLOP, F. F. HUANG, F. W. PIERSON, T. E. TOTH, X. J. MENG (2004b): Genetic identification of avian hepatitis E virus (HEV) from healthy chicken flocks and characterization of the capsid gene of 14 avian HEV isolates from chickens with hepatitis-spleno-megaly syndrome in different geographical regions of the United States. J. Gen. Virol. 85, 693-700.
TEI, S., N. KITAJIMA, K. TAKAHASHI, S. MISHIRO (2003): Zoonotic transmission of hepatitis E virus from deer to human beings. Lancet 362, 371-373.
TODD, D. K. A., A. MAWHINNEY, V. A. Mc ALINDEN, A. J. DOUGLAS (1993): Development of an enzyme-linked immunosorbent assay for the serological diagnosis of big liver and spleen disease. Avian Dis. 37, 811-816.
ZHANG, L., E. M. ZHOU, Z. Z. CUI, S. W. DONG, S. T. JING, J. PENG (2008) Serological investigation of avian hepatitis E virus infection and its relationship with chicken hepatitis-splenomegaly syndrome [in Chinese]. China Poult. 30, 22-24.
AVIAN HEPATITIS E – A POTENTIAL RISK FOR POULTRY PRODUCTION?!
Summary
Losses in poultry production are frequently caused by a decrease in hatchability, delayed sexual maturation of chickens, lower quality of eggshell, etc. Hepatitis E might be one of the possible but very rarely mentioned causes of these events. The disease may be neglected and unsuccessfully treated symptomatically due to overlapping of its symptoms with those associated with infectious bronchitis, syndrome of hatchability decline, technological errors in chicken breeding, etc. Therefore, the aim of this paper is to present the basic characteristics of the disease, along with the results of a preliminary study of the hepatitis E virus prevalence in the organs of chickens in which pathoanatomical examination revealed hepatitis E symptoms. Key words: hepatitis E, hepevirus, chicken

PERADARSKI DANI 2013.
91
CRIJEVNI I IMUNOSUPRESIVNI VIRUSI U ETIOLOGIJI SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA
Marina Biđin1, Darko Majnarić2, Zdenko Biđin1, Ivana Lojkić3
1Zavod za bolesti peradi s klinikom, Veterinarski fakultet Sveučilišta u Zagrebu, Hrvatska 2Veterinarski zavod Križevci, Hrvatski veterinarski institut, Križevci, Hrvatska 3Odjel za virologiju, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak
Opisano je istraživanje proširenosti nekih crijevnih i imunosupresivnih virusa u jatima intenzivno uzgajanih tovnih pilića na području Republike Hrvatske. Pretraženo je ukupno 28 jata različitog zdravstvenog stanja i dobi. Istraživanjem su obuhvaćena jata pilića zahvaćena blagim do srednje jakim proljevom i kržljavošću, kao i klinički zdravi pilići, čiji su organi uzorkovani nakon klanja. Skupni uzorci crijeva i slezena pilića su pretraženi lančanom reakcijom polimeraze na virus ptičjeg nefritisa (ANV), kokošji astrovirus (CAstV), kokošji parvovirus (ChPV), virus zarazne anemije pilića (CAV) i kokošji adenovirus (FAV). Pretragama je dokazana prisutnost jednog ili više virusa u ukupno 21 jatu, od čega je ANV bio najčešće dokazan virus u bolesnih i klinički zdravih ptica. Molekularna i filogenetska analiza hrvatskih CAV sekvenci temeljena na odsječku kapsidnog proteina je pokazala da su domaći sojevi najsličniji kineskom izolatu CH SCK/05-01/GD/664. Nalaz pretraživanih virusa u jatima pilića zahvaćenim crijevnom bolešću doprinosi poznavanju virusne etiologije sindroma zarazne kržljavosti, kao i epizootioloških osobitosti ovoga područja. Ključne riječi: sindrom zarazne kržljavosti, crijevni virusi, imunosupresivni virusi
Uvod
Sindrom zarazne kržljavosti tovnih pilića je jedan od go-spodarski najznačajnijih pojava u brzorastućih pilića di-ljem svijeta. Etiologija sindroma u kojem prevladavaju enterovirusi je složena i nije konačno riješena, a gubici rastu zbog povećanog uginuća, troškova liječenja i veće količine klaoničkog konfiskata (Biđin, 2008.). Crijevni je trakt ptica jako specijaliziran sustav s brojnim posebnim funkcijama vezanim za probavu i apsorpciju hra-njivih tvari, a ujedno i je i glavna razdjelnica između vanj-skog okoliša i unutarnjosti i mjesto prodora brojnih uzroč-nika, poglavito različitih crijevnih virusa koji se umnažaju u crijevnom epitelu i pridruženom limfoidnom tkivu, na-pose Fabricijeve burze. Zato su crijevne bolesti važna kategorija zaraznih bolesti peradi (Schat i Myers, 1991.) To značenje potječe iz činjenice da današnji moderni tovni pilići mogu ostvariti svoja genima određena proizvodna svojstva ako su opskrbljeni optimalno sastavljenom hra-nom i ako nije došlo do njene poremećene resorpcije (ma-
lapsorpcije). Uspješna će proizvodnja tovnih pilića biti tek ako je svaka hraniva komponenta u optimalnoj količini u smjesi i ako je takva smjesa optimalno pasirala probavnim traktom. Poremećen unos esencijalnih hraniva u organizam ptice remeti i normalni razvoj jedinstvenog imunog aparata vezanog za sluznicu čija je glavna komponenta limfoidno tkivo vezano za crijevo. Taj sustav sadrži limfoidne tvorbe kao što je Fabricijeva burza i drugo limfodino tkivo u lamini propriji crijeva (Kelsall i Strober, 1999.). Maldigestija i malapsorpcija se najjednostavnije određuju kao nemogućnost crijeva da probavi i resorbira sastojke iz hrane. Posljedično nastale promjene mogu biti vezane za kržljav rast, proljev, a ostali se simptomi mogu očitovati sekundarnim nestašicama vitamina, minerala, proteina i masti te posljedično rahitisom, kržljavošću, živčanim simptomima, slabim opernaćenjem, anemijom (Schleifer, 1995.). Različito kliničko očitovanje i nedovoljno poznata etiologija sindroma kržljavosti i zaostalosti razlog su broj-nim opisima te pojave u svijetu kao "sindrom kržljavosti i slabosti nogu", "helikopterska bolest", "bolest žutih glava",
Dr. sc. Marina Biđin, dr. med. vet., Zavod za bolesti peradi s klinikom, Veterinarski fakultet Sveučilišta u Zagrebu, Heinzelova 55, Zagreb, Hrvatska, tel: 01 2390-281; e-mail: [email protected]

CRIJEVNI I IMUNOSUPRESIVNI VIRUSI U ETIOLOGIJI SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA 92
"malapsorpcijski sindrom" ili "sindrom proljeva i kržlja-vosti", "zarazni proventrikulitis", "sindrom blijedih ptica", dok se u nas on obično opisuje kao "sindrom zarazne kržljavosti" (Kouwenhoven i sur., 1978.; 1983.; Good, 1982.; Schleifer, 1995.; Shane, 2002.; Biđin, 2008.). Poremećenu probavu i resorpciju hrane mogu uzrokovati zarazni ili nezarazni uzročnici te fizički čimbenici. Od za-raznih uzročnika prevladavaju virusi, bakterije i protozoa (Schleifer, 1995.). Dosadašnja iskustva na osnovi terenskih nalaza i pokusnih rezultata ukazuju da je etiologija sindroma pretežno zara-zna. Iz pilića s različito teškim proljevom, ali i klinički potpuno zdravih ptica su najčešće izdvojeni različiti ente-rovirusi (rota-, astro-, parvo-, entero-, toga- i kalicivirusi) te adeno-, reovirusi (za njih se drži da su „okidači“ u etiologiji sindroma) (Kisary i sur., 1984.; McNulty i sur., 1984.; Decaesstecker i sur. 1986.; Frazier i sur., 1986.; Connor i sur., 1987.; Yason i Schat, 1987.; Goodwin i sur.; 1996.; Todd, 2009; de Witt i sur., 2011.). Crijevnim se virozama peradi u nas kao zasebnom nozo-loškom entitetu donedavno nije posvećivala osobita pozor-nost. Zbog opće poznatih problema umnažanja nekih od spomenutih enterovirusa dijagnostika se u početku temelji-la uglavnom na serološkom dokazu protutijela, pričem se u pravilu dokazivao grupno specifičan antigen za pojedini virus. Tek razvojem molekularnih dijagnostičkih metoda počela se slagati epizootiološka slika proširenosti infekcije u domaće peradi određenim enterovirusom. Uza specifične enteroviruse na zdravlje probavnog sustava značajno djeluje imuni sustav domaćina. Jaka atrofija Fa-bricijeve burze te sekundarno timusa i slezene, uzrokovana virusom zarazne bolesti burze, u pilića u prvim danima života omogućuje sinergetsko djelovanje i drugih imuno-supresivnih virusa poput virusa zarazne anemije pilića i drugim manje patogenim enterovirusima. Infekcija jako vi-rulentnim virusom zarazne bolesti burze može produžiti zadržavanje virusa zarazne anemije pilića i poremetiti na-stanak dobne otpornosti za taj virus koja je potpuna u imuno zrelih pilića u dobi 3 tjedna, čak i prije (Rosen-berger i Cloud, 1989.; Ismail i Saif, 1991; Imai i sur., 1999; Todd, 2000.; Jackwood i Sommer-Wagner, 2005.; Durairaj i sur., 2011.). U radu su opisani rezultati ciljanog istraživanja prošireno-sti nekih enterovirusa (virus ptičjeg nefritisa, ANV; ko-košji astrovirus, CastV; virus zarazne anemije pilića, CAV; kokošji parvovirus, ChPV; i kokošji adenovirus, FAV) u jatima tovnih pilića koji su odavali znakove sindroma zarazne kržljavosti u farmama intenzivne proizvodnje u Hrvatskoj.
Materijali i metode
Pilići Dijagnostičko istraživanje proširenosti nekih imunosupre-sivnih i crijevnih virusa (ANV, CAstV, CAV, ChPV i FAV) je provedeno na 28 jata tovnih pilića svrstanih u tri dobne skupine; 1-7 dana (7 jata), 7- 21 dan (13 jata) i od
21. dana pa sve do završetka tova (8 jata). Za potrebe istraživanja pretraživana su jata koja su pokazivala zna-kove enteritisa očitovanog u rasponu od blagog do srednje jakog proljeva i kržljavosti. Unutar najstarije dobne sku-pine su uz uginule pretražena i 4 jata (jato 25, 26, 27 i 28 u tablici 2.) klinički zdravih pilića čiji su organi prikupljeni nakon klanja. Od svakog je jata uzorkovano 10-15 pilića. Pripremanje uzoraka i pročišćavanje virusnih nukleinskih kiselina (NK) Za potrebe istraživanja su od svih pilića uzorkovani crijevo i slezena. Uzorci su do pročišćavanja zamrznuti pri -20 oC. Nakon odmrzavanja su od pojedinačnih napravljeni skupni uzorci crijeva, odnosno slezene, tako da je svaki uzorak sadržavao otprilike 5 organa po jatu. U uzorke je dodano 750 μL puferirane fosfatne otopine (PBS), a za pročišća-vanje virusnih NK iz dobivenih homogenata je rabljen High Pure Viral Nucleic Acid Kit (Roche Diagnostics, Mannheim, Njemačka) prema uputama proizvođača. Viru-sne NK pročišćene su u ukupnom volumenu od 50 μL. Reverzna transkripcija (RT) i lančana reakcija polimeraze (PCR) Od pročišćenih je uzoraka odpipetirano 5 μL RNK koja je poslužila za reakciju reverzne transkripcije. Za sintezu cDNK je rabljena reverzna transkriptaza SuperScript TM III reverse transcriptase (Invitrogen, Carlsbad, Kalifornija, SAD) i ''random hexamers'' početnica. Reakcija je izve-dena u uređaju GeneAmp PCR System 2400 (Applied Biosystems, Foster City, Kalifornija, SAD) pri 42 oC tije-kom 50 minuta, a zatim je inaktivirana tijekom 15 minuta pri 70 oC. Uzorci su metodom PCR pretraženi na ANV, CAstV, CAV, ChPV i FAV. Svaka reakcija PCR je provjerena pozitivnom (prethodno dokazan i sekvenciran virus) i negativnom kontrolom (destilirana voda). Reakcija PCR (u ukupnom volumenu od 50 μL) je sadržavala 5 μL cDNK ili DNK, 25 μL Go Taq® Green Master Mix (Pro-mega, Madison, SAD) i 0,20 μM uzvodne i nizvodne početnice. Početnice za svaki pojedini virus i uvjeti reak-cije PCR su prilagođeni za svaku pretragu prema referen-cama prikazanim u tablici 1. Proizvodi reakcije PCR su analizirani elektroforezom u 2%-tnom agaroznom gelu obojanom etidij-bromidom. Sekvenciranje i analiza sekvenca Svaki uzorak pozitivan na jedan ili više virusa je poslan u tvrtku Macrogen Inc. (Seoul, Koreja), gdje je sekvenciran u oba smjera. Sekvence su analizirane pomoću programa BLAST (http://blast.ncbi.nlm.nih.gov./Blast.cgi), BioEdit (verzija 7.0.1.) (Hall, 1999.) i Mega 5 programa (Tamura i sur., 2011.) koji je korišten za filogenetsku analizu. U radu su opisane sekvence dobivene iz uzoraka pozitivnih na CAV, koje su upisane u banku gena (GenBank, http://www.ncbi.nlm.nih.gov/genbank/ pod pristupnim bro-jevima KC602112, KC602113, KC602114 i KC602115.

CRIJEVNI I IMUNOSUPRESIVNI VIRUSI U ETIOLOGIJI SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA
93
Rezultati
PCR Rezultati PCR pretrage 28 jata pilića na ANV, CAstV, CAV, ChPV i FAV su prikazani u Tablici 2. Tablica 1. Početnice za dokaz virusa pretraživanih u ovom
radu Table 1. Primers used for detection of viruses tested in this
study
Virus Virus
Ciljni gen Target gene
Veličina proizvoda PCR Size of PCR product
Referenca Reference
ANV ORF 1b 255 bp Biđin i sur., 2011. CAstV ORF 1b 362 bp Day i sur., 2007. CAV VP 1 675 bp Todd i sur., 1992. ChPV NS 561 bp Zsak i sur., 2009. FAV Hexon 1319 bp Hess i sur., 1999. Molekularna i filogenetska analiza CAV sekvenca Jata pozitivna na CAV, kao i nazivi sekvenca su prikazani u tablici 2. Domaće sekvence međusobno dijele 96% do 99,3% nukleotidne i 99,5%-100% sličnosti u uspoređi-vanom odsječku kapsidnog proteina. Slika 1. prikazuje dendrogram dobiven filogenetskom analizom hrvatskih sekvenca i 10 CAV sekvenca dokazanih na različitim zemljopisnim područjima. Domaće sekvence su pokazale nešto veću sličnost sa stranima na aminokiselinskoj (98,6%-100%) nego na nukleotidnoj razini (96,1%-99%).
Rasprava i zaključak
Istraživanje proširenosti nekih crijevnih i imunosupresiv-nih virusa povezanih s etiologijom sindroma zarazne kržljavosti pilića je pokazalo da je infekcija virusom ANV najčešća u sve tri pretraživane dobne skupine tovnih pilića (tablica 2.). To ne iznenađuje budući da je poznato da se ANV širi i vertikalnim i horizontalnim putem, a jednodnevni pilići se smatraju najosjetljivijim na infekciju (Imada i sur., 1981). S obzirom na to da istraživanjem nisu praćena ista jata u različitim vremenskim razmacima i zbog nepoznavanja nji-hovog zdravstvenog stanja u prvom tjednu života možemo pretpostaviti, ali ne i egzaktno ustvrditi da je upravo hori-zontalno širenje virusa odgovorno za povećanje broja ANV pozitivnih jata u dobi od 7 dana pa sve do kraja tova. Ipak, longitudinalno istraživanje proširenosti astrovirusa u jatima pilića koje su opisali Smyth i sur. (2009.) naglašava važnost i udio horizontalnog prijenosa CAstV i ANV u prvim da-nima života pilića u odnosu na vertikalno inficirane piliće. Osim u jatima s proljevom, nalaz ANV u klinički zdravog jata upućuje na subkliničku infekciju, a poznato je da se ANV može naći i u zdravih ptica (Pantin-Jackwood i sur., 2008.; Zhao i sur., 2011.). Uz ANV po zastupljenosti pozi-tivnih jata slijedi ChPV koji je na području Republike Hrvatske već opisan kao uzročnik prisutan u jatima pilića i purića zahvaćenih crijevnom bolešću (Biđin i sur., 2011.).
Parvovirus je poznat kao uzročnik enteropatije i jedan je od virusa što se povezuju sa sindromom zarazne kržljavosti (Guy, 1998.; Zsak i sur., 2009.). Infekcija adenovirusom je dokazana u 5 jata dobi 7-21 dana i najčešće je bila u koinfekciji s jednim ili više pretraživanih virusa (tablica 2.). Takav nalaz mješovite infekcije crijevnim i imunosupresiv-nim virusima objašnjava kliničke simptome (proljev, zaosta-janje u rastu, raslojenost jata) koji se očituju kao sindrom zarazne kržljavosti. Tablica 2. Rezultati PCR pretrage tovnih pilića na neke
crijevne i imunosupresivne viruse Table 2. Results of PCR examination for some enteric and
immunosuppressive viruses in broiler chickens
Dob Age
Jato Flock ANV CAstV CAV ChPV FAV
1 - - - - - 2 - - - - - 3 - - - + - 4 + - - - - 5 + - - - - 6 - - - - -
1-7 dana/days
7 + - +a - - 8 + - - + - 9 + - - + -
10 + - - - - 11 + - - - - 12 + - - + - 13 + - +b - + 14 - - - - + 15 + + - + - 16 - + - - - 17 + - +c - + 18 - - - - + 19 + - - - +
7-21 dana/days
20 + - - + - 21 + - - + - 22 - - - + - 23 - - - + - 24 + - - - - 25 + - +d - - 26 - - - - - 27 - - - - -
21 dan/day -
End of fattening period
28 - - - - - asekvenca M12-738 (KC602112), bsekvenca M12-740 (KC602113), csekvenca M12-962 (KC602114), dsekvenca M12-964 (KC602115) asequence M12-738 (KC602112), bsequence M12-740 (KC602113), csequence M12-962 (KC602114), dsequence M12-964 (KC602115)

CRIJEVNI I IMUNOSUPRESIVNI VIRUSI U ETIOLOGIJI SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA 94
Uza spomenute viruse prirodna infekcija virsuom CAV je dokazana u 4 jata različite dobi. CAV je najranije bio dokazan u pilića dobi do 7 dana, ali je također nađen i u pilića koji za života nisu pokazivali nikakve znakove bolesti. Budući da je dokazan u jatu pilića starijem od tri tjedna možemo pretpostaviti da nije došlo do nastanka bolesti, jer su pilići dotad već razvili dobnu rezistenciju na virus. Osim što pilići nisu bili cijepljeni protiv CAV, molekularnom i filogenetskom analizom odsječka kapsidnog proteina je potvrđeno da dokazani virus pripada u terenske, a ne cijepne sojeve. Filogenetski, hrvatski su sojevi najsličniji s CAV izdvojenim u Kini, dok su evolucijski najviše udaljeni od izolata podrijetlom iz SAD-a (slika 1.). Zanimljivo je i da su sve hrvatske sekvence osim M12-964 aminokiselinskim sastavom identične sekvenci CL37 iz Čilea, koja pripada u skupinu girovirusa dokazanih u ljudi (Phan i sur., 2012.). Sindrom zarazne kržljavosti nastaje međusobnim djelo-vanjem više zaraznih i nezaraznih čimbenika, od kojih su u ovom radu obrađeni neki najčešći crijevni i imunosupresivni virusi. Iz ovog se istraživanja može zaključiti da je infekcija virusom ANV puno češća u odnosu na ostale viruse, pričem je zanimljivo da je CAstV, iako pripadnik iste porodice virusa kao i ANV, bio najrjeđe zastupljen virus u tovnih pilića. Istraživanja provedena u drugim dijelovima svijeta pokazuju suprotne rezultate, pričem su oba astrovirusa go-tovo jednako prisutna u komercijalno uzgajanih pilića i često dolaze u koinfekciji (Pantin-Jackwood i sur., 2006.; Smyth i sur., 2009.). Iako je za potpunu sliku etiologije sindroma zarazne kržljavosti svakako potrebno uzeti u obzir i druge
čimbenike, dokaz miješane virusne infekcije ukazuje na si-nergetsku ulogu virusa u nastanku sindroma zarazne kržlja-vosti. Suprotno tome, pretraživanje klinički zdravih jata je dalo očekivano negativni nalaz pretrage na sve viruse u tri od ukupno četiri jata. Zaključno, istraživanje virusa što cirkuliraju jatima tovnih pilića pomaže poznavanju etiologije sindroma zarazne kržljavosti i postavljanju dijagnoze oboljelih jata.
Literatura
BIĐIN, Z. (2008): Sindrom zarazne kržljavosti. U Bolesti peradi. Veterinarski fakultet Sveučilišta u Zagrebu, 201-206.
BIĐIN, M., I. LOJKIĆ, Z. BIĐIN, M. TIŠLJAR, D. MAJNARIĆ, M. MIKEC (2011): Detection and characterization of avian nephritis virus in ducklings. Avian Pathol. 40, 173-177.
CONNOR, T. J., F. McNEILLY, J. B. McFERRAN, M. S. McNULTY (1987): A survey of avian sera from Northern Ireland for antibody to avian nephritis virus. Avian Pathol. 16, 15-20.
DAY, J. M., E. SPACKMAN, M. J. PANTIN-JACKWOOD (2007): A multiplex RT-PCR test for differential identification of turkey astrovirus type 1, turkey astrovirus type 2, chicken astrovirus, avian nephritis virus, and avian rotavirus. Avian Dis. 51, 681-684.
DECAESSTECKER, M., G. CHARLIER, G. MEULEMANS (1986): Significance of parvoviruses, entero-like viruses and reoviruses in aetiology of the chicken malabsorption syndrome. Avian Pathol. 15, 676-782.
Slika 1. Filogenetsko Neighbour-joining stablo dobiveno analizom 219 aminokiselina izvedenih iz nukleotidnih sekvenca
odsječka VP 1 gena od 14 CAV izolata. Ljestvica na dnu slike označava evolucijsku udaljenost izračunatuPoissonovim korekcijskim modelom, uz 1000 ,"bootstrap" replikacija
Figure 1. Phylogenetic Neighbour-joining tree based on the analysis of 219 amino acids deduced from partial VP1 gene
nucleotide sequences of 14 CAV isolates. The scale in the bottom indicates evolutionary distance computed using thePoisson correction model with 1000 bootstrap replicates

CRIJEVNI I IMUNOSUPRESIVNI VIRUSI U ETIOLOGIJI SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA
95
DURAIRAJ, V., HOLLY S. SELLERS, E. G. LINNEMANN, A. H. ICARD, E. MUNDT (2011): Investigation of the antigenic evolution of filed isolates using the reverse genetics system of infectious bursal disease virus (IBDV). Arch. Virol. 156, 1717-1728.
FRAZIER, J. A., H. FARMER, M. F. MARTLAND (1986): A togavirus-like agent in the pancreatic duct of chickens with infectious stunting syndrome. Vet. Rec. 119, 208-209.
GOOD, R. E. (1982): The pale bird syndrome. Poult. Dig. 41, 298. GOODWIN, M. A., S. HAFNER, D. I. BOUNOUS, K. S.
LATIMER, E. C. PLAYER, F. D. NIAGRO, R. P. CAMPAGNOLI, J. BROWN (1996): Viral proventriculitis in chickens. Avian Pathol. 25, 369-379.
GUY, J. S. (1998): Virus infections of the gastrointestinal tract of poultry. Poultry Sci. 77, 1166-1175.
HALL, T. A. (1999): BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95.98.
HESS, M., R. RAUE, H. M. HAFEZ (1999): PCR for specific detection of haemorrhagic enteritis virus of turkeys, an avian adenovirus. J. Virol. Methods 81, 199-203.
IMAI, K., M. MASE, K. TSUKAMOTO, H. HIHARA, N. YUASA (1999): Persistent infection with chicken anaemia virus and some effect of highly virulent infectious bursal disease virus infection in its persistency. Res. Vet. Sci. 67, 233-238.
IMADA, T., T. TANIGUCHI, S. YAMAGUCHI, T. MINETOMA, M. MAEDA, H. KAWAMURA (1981): Susceptibility of chickens to avian nephritis virus at various inoculation routes and ages. Avian Dis. 25, 294-302.
ISMAIL, N. M, Y. M. SAIF (1991): Immunogenicity of infectious bursal disease viruses in chickens. Avian Dis. 35, 460-469.
JACKWOOD, D. J., S. E. SOMMER-WAGNER (2005): Mole-cular epidemiology of infectious bursal disease viruses: distribution and genetic analysis of newly emerging viruses in the United States. Avian Dis. 49, 220-226.
KELSALL, B., W. STROBER (1999): Gut associated lymphoid tissue, antigen handling and T-lymphocyte responses. U: Mucosal Immunology. Ogra, P. L., J. Mestecky, M. E. Lamm, W. Strober, J. Binenstock, J. R. McGhee (ur.). New York. Academic Press. pp. 293-317.
KISARY, J., B. NAGY, Z. BITAY (1984): Presence of parvoviruses in the intestine of chickens showing stunting syndrome. Avian Pathol. 13, 339-343.
KOUWENHOVEN, B., M. VERTOMMEN, E. GOREN (1983): Runting and stunting syndrome in broiler: the disease with many names and faces. Int. Un. Immunol. Proc. No. 66, 73-96.
KOUWENHOVEN, B., M. VERTOMMEN, J. H. H. van ECK (1978): Runting and leg weakness in broilers: involvement of infectious factors. Vet. Sci. Commun. 2, 253-259.
McNULTY, M. A., G. M. ALLAN, T. J. CONNOR, J. B. McFERRAN, R. M. McCRACKEN (1984): An entero-like virus associated with the runting syndrome in broiler chickens. Avian Pathol. 13, 429-439.
PANTIN-JACKWOOD, M. J., E. SPACKMAN, P. R. WOOLCOCK (2006): Molecular characterization and typing of chicken and turkey astrovirus circulating in the United States: implication for diagnostics. Avian Dis. 50, 397-404.
PANTIN-JACKWOOD, M. J., J. M. DAY, M. W. JACKWOOD, E. SPACKMAN (2008): Enteric viruses detected by molecular methods in commercial turkey flocks in the United States between 2005-2006. Avian Dis. 52, 235-244.
PHAN, T. G., L. LIU, M.G. O'RYAN, H. CORTES, N. MAMANI, I. J. BOKOUNGOU, C. WANG, C. M. LEUTENEGGER, E. DELWART (2012): A third gyrovirus species in human faeces. J. Gen. Virol. 93, 1356-1361.
ROSENBERGER, J. K., S. S. CLOUD (1989): The isolation and characterization of chicken anemia agent (CAA) from broilers in the United States. Avian Dis. 33, 707-713.
SCHAT, K. A., T. J. MYERS (1991): Avian intestinal immunity. Crit. Rev. Poultry Biol. 3, 19-34.
SCHLEIFER, J. (1995): Malabsorption in broilers. Poultry Int. 34, 80-84.
SHANE, S. (2002): Runting and stunting syndrome. Poultry Int. 41, 38-41.
SMYTH, V. J., H. L. JEWHURST, B. M. ADAIR, D. TODD (2009): Detection of chicken astrovirus by reverse transcrip-tase-polymerase chain reaction. Avian Pathol. 38, 293-299.
TAMURA, K., D. PETERSON, N. PETERSON, G. STECHER, M. NEI, S. KUMAR (2011): MEGA5: Molecular Evo-lutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 28, 2731-2739.
TODD, D., K. A. MAWHINNEY, M. S. McNULTY (1992): Detection and differentiation of chicken anemia virus isolates by using the polymerase chain reaction. J. Clin. Microbiol. 30, 1661-1666.
TODD, D. (2000): Circoviruses: immunosuppressive threats to avian species: review. Avian Pathol. 29, 373-394.
TODD, D. (2009): Identification of chicken enterovirus-like viruses, duck hepatitis virus type 2 and duck hepatitis virus type 3 as astroviruses. Avian Pathol. 38, 21-30.
de WITT, J. J., G. B. DAM, J. M. van de LAAR, Y. BIERMANN, I. VERSTEGEN, F. EDENS, C. C. SCHREIER (2011): Detection and characterization of a new astrovirus in chicken and turkeys with enteric and locomotion disorders. Avian Pathol. 40, 453-461.
YASON, C. V., K. A. SCHAT (1987): Pathogenesis of rotavirus infection in various age groups of chickens and turkeys. Clinical signs and virology. Am. J. Vet. Res. 48, 977-983.
ZHAO, W., X. G. HUA, C. L. YUAN, L. CUI, T. L. SHAN, X. Q. DAI, A. L. ZHU, Y. YU, C. X. ZHU, Z. B. YANG (2011): Sequence analyses of the representative Chinese-prevalent strain of avian nephritis virus in healthy chicken flocks. Avian Dis., 55, 65-69.
ZSAK, L., K. O. STROTHER, M. J. DAY (2009): Development of a polymerase chain reaction procedure for detection of chicken and turkey parvoviruses. Avian Dis. 53, 83-88.

CRIJEVNI I IMUNOSUPRESIVNI VIRUSI U ETIOLOGIJI SINDROMA ZARAZNE KRŽLJAVOSTI TOVNIH PILIĆA 96
ENTERIC AND IMMUNOSUPPRESSIVE VIRUSES IN RUNTING AND STUNTING SYNDROME IN BROILER CHICKENS Summary A survey of the prevalence of some enteric and immunosuppressive viruses in commercially reared broiler flocks in Croatia is described. A total of 28 flocks of different health status and ages were tested. Flocks that suffered from mild to severe diarrhea and stunting were covered with this research. The organs of clinically healthy broilers were also tested for the virus presence. Pooled intestinal and spleen samples were tested by polymerase chain reaction for avian nephritis virus (ANV), chicken astrovirus (CAstV), chicken parvovirus (ChPV), chicken anemia virus (CAV) and fowl adenovirus (FAV). The presence of one or more viruses was detected in 21 flocks, with ANV being the most common virus found in tested flocks. Molecular and phylogenetic analysis of Croatian CAV sequences based on the fragment of the capsid protein showed their high similarity with the Chinese isolate CH SCK/05-01/GD/664. This investigation contributes by revealing the viral etiology of runting and stunting syndrome, as well as epizootiologic characteristics of this area. Key words: runting and stunting syndrome, enteric viruses, immunosuppressive viruses

PERADARSKI DANI 2013.
97
MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI
Marijana Sokolović, Borka Šimpraga, Fani Krstulović, Marija Berendika
Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak
Mikotoksini su sekundarni metaboliti brojnih vrsta plijesni koji mogu uzrokovati poremećaje i bolesti u ljudi i životinja. Ovaj rad prikazuje najčešće mikotoksine značajne za uzgoj peradi s obzirom na njihov nastanak, toksičnost, štetne učinke u peradi i rizik za zdravlje. Također daje osvrt na postupke kontrole te zakonske i preventivne mjere koje se primjenjuju u Republici Hrvatskoj. Ključne riječi: mikotoksini, perad, mikotoksikoze, plijesni Uvod Mikotoksini su raznolika skupina sekundarnih metabolita različitih vrsta plijesni koje rastu na odgovarajućim supstra-tima. Svojim rastom plijesni kao primarni ili znatno češće kao oportunistički patogeni (najčešće respiratornim putem) mogu uzrokovati nastanak bolesti koje se nazivaju mikoze (npr. aspergiloza). Mikotoksikoze, bolesti uzrokovane miko-toksinima, najčešće nastaju trovanjem (hranom, rjeđe zra-kom ili putem kože). Ovisno o vrsti mikotoksina, količini i trajanju izloženosti toksinu, dobi, spolu i zdravstvenom sta-tusu životinje, uzrokuju različite simptome. Pojedini miko-toksini mogu sinergistički djelovati s drugim toksinima i/ili mikotoksinima te tako smanjiti otpornost životinje na druge štetne tvari i uzročnike infekcije. Ako dođe do pojave miko-toksikoze, liječenja u pravilu nema, već se primjenjuju ne-specifične mjere (promjena hrane, poboljšani higijenski uvjeti i sl.) a u posljednje vrijeme i sve je češća primjena različitih veterinarskih pripravaka koji u određenoj mjeri mogu vezati toksine i tako smanjiti njihov štetni učinak (Henry i sur., 2002.). U literaturi mikotoksini se klasificiraju na razne načine. Primjerice, mogu se podijeliti prema organu na koji primar-no djeluju (hepatotoksini, nefrotoksini, neurotoksini, imuno-toksini i dr.), prema kemijskoj strukturi i biosintetskom podrijetlu (kumarini, poliketidi i dr.), prema specifičnom učinku na razini stanice (mutageni, karcinogeni i dr.), prema bolestima koje uzrokuju (asperigiloza i dr.) te prema pli-jesnima koje ih proizvode (Aspergillus toksini, Fusarium
toksini, Penicillium toksini i dr.). Međutim, niti jedna od ovih podjela nije u potpunosti točna. Mikotoksini su prirodne molekule male molekularne težine, različitih kemijskih struktura, podrijetla, bioloških i toksič-nih učinaka, a naziv su dobili davne 1962. godine kada je u Velikoj Britaniji došlo do uginuća više od 100.000 puran-skih pilića te oboljenja i drugih vrsta životinja. Kemijskim i mikrobiološkim analizama brašna od kikirikija podrijetlom iz Brazila kojim su hranjene te životinje pronađeni su fluo-rescirajući sekundarni metaboliti plijesni Aspergillus flavus po kojoj su dobili i ime (Blount, 1961.). Danas se smatra da postoji više od 400 različitih mikotoksina, ali je za svega manji broj evidentirana uzročno-posljedična veza s nastan-kom poremećaja i bolesti u ljudi i životinja. Rastu plijesni i proizvodnji mikotoksina pogoduju različiti okolišni uvjeti (vremenske prilike, širok raspon temperatura, visoka vlaga zraka i visoka relativna vlaga hrane). Mikotok-sini se proizvode na različitim supstratima uključujući žita-rice, uljarice, orašaste plodove, hranu i hranu za životinje. Prema Europskoj Direktivi 2002/32//EC hrana za koju po-stoji izrazita opasnost kontaminacije aflatoksinima uključuje kikiriki, kokos, palmine koštice, sjeme pamuka, tvrdi orah babasu, kukuruz i proizvode od navedene hrane. Do konta-minacije mikotoksinima može doći prije žetve (kao poslje-dica stresa uzrokovanog sušom) i nakon žetve (prilikom pre-rade, transporta i neodgovarajuće pohrane žitarica). Primje-rice, optimalnim uvjetima za proizvodnju aflatoksina smatra se visoka temperatura zraka (24 °C do 35 °C, visoka vlaga zraka (85%), visoka vlaga hrane (17%), a pogoduju ošteće-
Dr. sc. Marijana Sokolović, dr.med.vet, Centar za peradarstvo, Hrvatski veterinarski institut, Henizelova 55, 10000 Zagreb, RH; Tel.: ++385(0)12441-394; Fax: ++385(0)12441-396; E-mail: [email protected]

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI 98
nja uzrokovana insektima i glodavcima te prisutnost drugih plijesni i/ili mikroorganizama (WHO, 1998.; EFSA, 2004.). Bolesti uzrokovane mikotoksinima, mikotoksikoze, mogu biti akutne i kronične. Akutnu toksičnost obilježava nagli nastanak i karakteristična reakcija otrovanja, kao što je pri-mjerice gore opisani slučaj uginuća velikog broja puranskih pilića koji je nazvan „X bolest purana“ (Blount, 1961.; WHO, 1998.). Kronični oblik bolesti nastaje nakon dugo-trajnog izlaganja (najčešće hranom) malim dozama toksina, što posljedično rezultira nastankom uglavnom ireverzibilnih promjena (npr. tumora). Budući da specifičnog liječenja mikotoksikoza nema, preventiva nastanka jedina je prava mjera za sprječavanje štetnog učinka mikotoksina. Ona uključuje odgovarajuće mjere dobre proizvođačke prakse, higijene i kontrole u svim fazama proizvodnje, pohrane i korištenja hrane. Cilj ovoga rada je dati osvrt na mikotoksine koji se najčešće susreću u hrani za perad te prema dosadašnjim saznanjima imaju štetne učinke za zdravlje i proizvodnost peradi, a mogu predstavljati i prijetnju za zdravlje ljudi.
Aflatoksini
Jedni od prvih izdvojenih mikotoksina su aflatoksini. Otkri-veni su i identificirani nakon gore opisanog akutnog otro-vanja 100.000 puranskih pilića u Velikoj Britaniji. Pritom su oboljele i druge vrste životinja (pačići, fazani, goveda, svinje i ovce). Poveznica smrtnosti ovih životinja bila je hrana, odnosno brašno od kikirikija podrijetlom iz Brazila koje je bilo kontaminirano navedenim mikotoksinima. U brašnu su otkrivene četiri molekule derivata difuranokumarina (B1, B2, G1, G2) koje su fluorescirale plavom ili zelenom bojom kada su bile izložene UV svjetlu. Količina tih tvari određena je tankoslojnom kromatografijom i toksini su dobili naziv po plijesni Aspergillus flavus koja ih proizvodi. Proizvodnja aflatoksina, iako rjeđe, zabilježena je i u drugim vrstama plijesni roda Aspergillus (A. bombycis, A. ochraceoroseus, A. nomius i A. pseudotamari). Uz navedena četiri toksina 50% sojeva plijesni Aspergillus flavus i Aspergillus parasi-ticus mogu proizvoditi čak 18 različitih toksina (npr. P1, Q1, aflatoksikol), od kojih je aflatoksin B1 najtoksičniji i naj-češći u prirodi. Metaboliti aflatoksina M1 i M2 izdvojeni su iz mlijeka krava u laktaciji te su prema tome dobili i ime. Uz mlijeko, njihova prisutnost dokazana je u jajima i drugim proizvodima životinjskog podrijetla, koji tako predstavljaju neizravnu alimentarnu prijetnju za zdravlje ljudi (Goto i sur., 1996.; Peterson i sur., 2001.; EFSA, 2004.). Toksičnost Za aflatoksine, točnije aflatoksin B1, je dokazan toksični i karcinogeni učinak na ljude i životinje. Pritom aflatoksin B1 je i do danas jedini mikotoksin za koji je dokazana karcino-genost u ljudi te je prema izvješću Internacionalne agencije za istraživanje raka (International Agency for Research on Cancer) i svrstan u I. skupinu karcinogena (Shank i sur., 1972.; Squire, 1981.; IARC, 1982.). LD50 vrijednost aflatoksina B1 (tablica 1.) u većine životinjskih vrsta kreće
se od 1 do 50 mg/kg, dok je najtoksičniji za pastrve, mačke i pačiće (LD50 <1 mg/kg) (Leeson i sur., 1995.; Rawal i sur., 2010.). Simptomi u peradi Akutna aflatoksikoza rezultira smrću i različita je ovisno o vrsti, spolu, dobi, zdravstvenom stanju životinje te načinu unosa toksina u organizam. Simptomi uključuju anoreksiju, gubitak tjelesne težine, depresiju, ataksiju i uginuće unutar jednog do dva tjedna sa znakovima opistotonusa i nogu ispruženih prema natrag (Hamilton i sur., 1972.). Kronična aflatoksikoza rezultira nastankom tumora, imunosupresije i sličnih kroničnih poremećaja. Primarni ciljni organ djelo-vanja toksina je jetra, a oštećenja (različitog stupnja ovisno o vrsti) su uz perad dokazana i u riba, glodavaca i primata. U kroničnim slučajevima primjećuje se smanjeni prirast, pove-ćani slezena i gušterača te regresija Fabricijeve burse (Ha-milton, 1992.). Sami simptomi ovise o dozi i dužini izlože-nosti toksinu. Primjerice, doza od 0,5 mg/kg aflatoksina B1 u hrani tovnih pilića u trajanju od četiri tjedna uzrokovati će značajan pad tjelesne težine i unosa hrane, dok kratkotrajna izloženost tovnih pilića količini od 1,0 mg/kg u hrani neće uzrokovati značajnije poremećaje proizvodnih karakteri-stika. Ipak, prisutnost mikotoksina u hrani remeti i pravilan unos drugih esencijalnih tvari u hrani (npr. kokcidiostatika, vitamina i/ili aminokiselina) (Jewers, 1990.). U kokoši nesilica dolazi do smanjene proizvodnje jaja i težine jaja, nastanka masne jetre te promjene nekih bioke-mijskih parametara kada je aflatoksin B1 prisutan u hrani u količini od 1 mg/kg ili više u trajanju od četiri tjedna ili više. Iako se aflatoksin eliminira vrlo brzo iz tkiva nakon ukla-njanja kontaminirane hrane (72-96 sati) neki su autori za-bilježili i postojanje rezidua aflatoksina M2 u tkivu bubrega i jajima i nakon sedam dana (Trucksess i sur., 1983.; Leeson i sur, 1995.; EFSA, 2004.).
Fumonizini
Fumonizini su prvi puta opisani i karakterizirani 1988. go-dine. Iako je identificirano šest različitih fumonizina (A1, A2, B1, B2, B3 i B4) najčešći predstavnik ove skupine toksina je fumonizin B1. Proizvode ga brojne vrste plijesni roda Fusarium i to osobito Fusarium verticillioides (nazi-vana i Fusarium moniliforme-Gibberella fukokuroi), Fusa-rium proliferatum i Fusarium nygamai te jedna vrsta plijesni Alternaria alternata (Bezuidenhout i sur., 1988.). Od svih navedenih plijesni, plijesan F. verticillioides najčešće je povezana s proizvodnjom ovoga mikotoksina na kukuruzu (u vegetativnom i reproduktivnom tkivu), pričem najčešće uopće ne izaziva bolest biljke. Iako je ova plijesan prisutna u gotovo svim uzorcima kukuruza, svaki kukuruz nije ujedno i kontaminiran fumonizinom (Plumlee i Galey, 1994.). Toksičnost Prema Internacionalnoj agenciji za istraživanje raka fumo-nizini su svrstani u skupinu 2B (potencijalni karcinogeni) (Rheeder i sur., 2002.). Prema rezultatima istraživanja

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI
99
Weibkinga i sur. (1993.) minimalna učinkovita doza koja će uzrokovati poremećaj zdravlja i proizvodnosti u pilića iznosi 75 mg/kg. S obzirom na to da je količina ovoga toksina u hrani za perad znatno manja, u pravilu se smatra da fumo-nizini ne predstavljaju značajniju prijetnju za perad. Ipak, kako je hrana često kontaminirana različitim mikotoksina, ne može se zanemariti njegov potencijalni sinergistički učinak s primjerice aflatoksinom i/ili trikotecenima (SCF, 2000.). Simptomi u peradi Fumonizini uzrokuju poremećaje prvenstveno u drugih vrsta životinja, jer interferiraju sa sfingolipidnim metabolizmom te dolazi do leukoencefalomalacije u konja i kunića (Ma-rasas i sur., 1988.; Bucci i sur., 1996.), edema pluća i hidro-toraksa u svinja (Harrison i sur., 1990.), hepatotoksičnih i karcinogenih učinaka i apoptoze u jetri štakora (Gelderblom i sur., 1988.). U peradi su potrebne znatno veće doze fumo-nizina kroz dulje razdoblje za nastanak kliničkih simptoma, smanjenja tjelesne težine i smrtnosti (125-274 mg/kg) i smatra se da alimentarna doza fumonizina od 80 mg/kg u pravilu ne utječe na proizvodne karakteristike u pilića (Javed i sur., 1993.; Henry i sur., 2000.; SCF, 2000.).
Ohratoksin
Ohratoksini predstavljaju skupinu od sedam metabolita pli-jesni. Prvi puta su izdvojeni 1965. godine iz plijesni Asper-gillus ochraceus (Shotwell i sur., 1969.). Toksine iz ove sku-pine proizvode brojne vrste plijesni roda Aspergillus (A.) alliaceus, A. auricomus, A. carbonarium, A. glaucus, A. melleus i A. niger (Bayman i sur., 2002.). Uz rod Aspergillus smatra se da samo jedna vrsta plijesni roda Penicillium (P. verrucosum) može proizvoditi ohratoksin (Pitt, 1987.). U prirodi je redovito prisutan samo ohratoksin A i to u žita-ricama poput ječma, pšenice, zobi, raži i kukuruza. Može se proizvoditi u na temperaturi okoliša od 4 °C u žitaricama koje imaju sadržaj vlage od 18,5% do 40,4% (Leeson i sur., 1995.). Uz navedeno, otkriven je i u krvi i različitim tkivima životinja te u mlijeku i mesu (Marquardt i Frohlich, 1992.). Toksičnost Ohratoksin A je najtoksičniji predstavnik svoje vrste i pri-marno oštećuje bubrege, dakle djeluje nefrotoksično. Smatra se najtoksičnijim za domaću perad te ima nižu LD50 vri-jednost i od aflatoksina i T-2 toksina (tablica 1.). Također postoje podaci da djeluje i hepatotoksično, imunosupre-sivno, a potencijalni je teratogen i karcinogen (Leeson i sur., 1995.). Simptomi u peradi Iako se ohratoksin redovito povezuje s uzgojem svinja na koje naročito štetno djeluje uzrokujući nefropatiju, utvrđeno je da može uzrokovati pobol i smrtnost u peradi (Hamilton i sur., 1982.; Burns i Dwivedi, 1986.). Internacionalna agen-cija za istraživanje raka svrstala je ohratoksin u skupinu mogućih karcinogena za ljude (skupina 2B) (IARC, 1982.).
U tovnih pilića koji su kontaminirani visokim dozama ohra-toksina A (16 mg/kg) dolazi do dijareje, tremora i drugih neuralnih poremećaja i uginuća unutar 24 sata. Manje vrijednosti ohratoksina u hrani (3 mg/kg) uzrokuju dijareju, dehidraciju i povećanu smrtnost. Od ostalih simptoma može se uočiti smanjeni rast, povećana konzumacija vode i vodenasti feces (Huff i sur., 1974.). U kokoši nesilica dolazi do smanjenog unosa hrane i pro-izvodnje jaja, smanjene mase jaja te učestalije pojave pro-mjena na ljuski jajeta. Smatra se da je minimalna količina ohratoksina A u hrani za perad potrebna za nastanak pore-mećaja u proizvodnji jaja 0,5 mg/kg (Leeson i sur., 1995.).
Trikoteceni
Trikoteceni su skupina od više od 190 mikotoksina koje proizvode prvenstveno plijesni roda Fusarium, ali i plijesni drugih rodova kao što su Myrothecium, Phomopsis, Stachybotrys, Trichoderma, Trichothecium i druge. Ime su dobili po "trichothecinu", prvom izoliranom članu ove sku-pine toksina. Obilježava ih posjedovanje 12,13-epoksitriko-tecenske strukture i olefinske veze. Mogu se podijeliti na markocikličke i nemakrocikličke toksine, ovisno o tome po-sjeduju li makrociklički ester ili ester-ester vezu između atoma ugljika na položajima C-4 i C-15 (Krska i sur., 2001.; Sokolovic i sur., 2008.). Makrocikličke trikotecene proiz-vode plijesni rodova Myrothecium, Stachybotrys i Tricho-thecium, a najpoznatiji makrociklički trikoteceni su satratok-sini, roridini i verukarini (Leeson i sur., 1995.). Nemakro-ciklički trikoteceni se dalje mogu podijeliti po strukturnim karakteristikama u skupinu A (T-2 toksin, neosolaniol, diacetoksiscirpenol) i skupinu B (fusarenon-x, nivalenol, deoksinivalenol). Najznačajniji predstavnici ove skupine toksina s obzirom na učestalost i štetne učinke u peradi su T-2 toksin, deoksi-nivalenol i diacetoksiscirpenol, pričem je deoksinivalenol (vomitoksin) najčešće prisutan u žitaricama u suptropskim područjima. Vrućine i visoka vlaga zraka predstavljaju čim-benike koji pogoduju infekciji biljaka plijesnima roda Fusarium. Uz odgovarajuće loše uvjete skladištenja dolazi do proizvodnje ovih trikotecena. Primjerice, T-2 toksin na-staje na oštećenim žitaricama koje sadrže vlagu u koncen-traciji od 13% do 22%. Nastaje u širokom rasponu tem-peratura (0-32 °C), s time da su optimalne temperature za proizvodnju toksina ispod 15 °C (Bergers i sur., 1985.). Od žitarica, najčešće su prisutni u kukuruzu, pšenici, ječmu, zobi i raži. Toksičnost Oni su izraziti inhibitori sinteze proteina u eukariota, a sama toksičnost pripisuje se posjedovanju različitih funkcionalnih skupina. Primjerice, 12,13-epoksidni prsten T-2 toksina od-govoran je za njegovu toksičnost, a proces de-epoksidacije rezultira gubitkom štetnog učinka. Toksičnost se smanjuje i procesom deacetilacije tijekom kojega nastaje prvi metabolit HT-2 toksin (nešto slabije toksičan od izvorne molekule) te krajnji proizvod T-2 tetraol čiji je štetni učinak gotovo

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI 100
sedam puta slabiji od izvorne molekule. Toksičnost, kao i kod drugih toksina, ovisi o načinu ulaska u organizam, vre-menu izlaganja toksinu, dozi te dobi, spolu i općem zdravstvenom stanju životinje. Najtoksičniji predstavnik ove skupine toksina je T-2 toksin koji djeluje citotoksično, imunosupresivno, štetno djeluje na probavni i neurološki sustav te kožu (tablica 1.) (Leeson i sur., 1995.; JECFA, 2001.; Sokolovic i sur., 2008.). Deoksinivalenol je u peradi znatno manje toksičan (tablica 1.) i u pravilu služi kao bio-marker onečišćenja drugim fuzarijskim mikotoksina. Ipak, procjenjuje se da ukupna količina svih trikotecenskih miko-toksina u uzorku hrane za životinje ne bi smjela biti veća od 0,5 mg/kg. Simptomi u peradi Konzumacija trikotecena može uzrokovati nastanak ošte-ćenja sluznice probavnog sustava, hemoragije, proljev i povraćanje, dok izravni kontakt može uzrokovati dermatitis. Pri nižim dozama toksin uzrokuje smanjeni prirast, smanjeni unos hrane, zaostajanje u rastu, gubitak tjelesne težine i odbijanje hrane u peradi kao i u drugih životinja. U svakom slučaju, kada se uoči smanjeni unos hrane, inhibicija rasta sa slabijim prirastom, nastanak lezija u sluznici usne šupljine, depigmentacije kože nogu, smanjena kvaliteta perja, neuro-loški poremećaji treba posumnjati na moguću kontaminaciju trikotecenskim mikotoksina, a naročito T-2 i HT-2 toksi-nom. U kokoši nesilica može se uočiti i smanjena proiz-vodnja jaja koja u pravilu imaju tanju ljusku, te smanjena valivost, cijanoze i leukopenija (Hoerr, 2003.). Zearalnon
Zearalenon (ZEA) je sekundarni metabolit plijesni Fusarium graminearum (telomorf Gibberella zeae) i dobio je ime po G. zeae, jer je po kemijskoj strukturi lakton fenolne resor-
ciklične kiseline. Prije se je često koristio i naziv F-2. Iako se svrstava u mikotoksine s obzirom na to da on ima izrazitu biološku aktivnost (može izazvati znakove estrusa u krmača kojima su uklonjeni ovariji, dok količina od 1-5 mg/kg u hrani može uzrokovati vulvovaginitis u mladih svinja), zapravo uopće nije toksičan. Strukturno i biološki nalikuje 17 β-estradiolu, hormonu ovarija koji regulira vezanje za estrogenske receptore na odgovarajućim stanicama. U hrani se često nalazi u koncentracijama manjim od 1 mg/kg, ali je nađen i u količini većoj od 30 mg/kg (Leeson i sur., 1995.). Proizvode ga plijesni Fusarium graminearum, Fusarium culmorum, Fusarium equiseti i Fusarium crookwellense. Uz zearalenon u ovu skupinu pripadaju i zearalenol (molekula slabije estrogene aktivnost) te zeranol koji je svojedobno ko-rišten kao anabolički agens za krave i goveda. I sam zearalenon je prije korišten za liječenje nekih stanja u ljudi (postmenopauzalni simptomi i kao oralno kontracepcijsko sredstvo) (Hiddy i sur., 1977.; Kuiper-Goodman i sur., 1987.). Toksičnost Toksičnost toksina iz skupine zearalenona je relativno mala, ali već male količine ovoga toksina (reda veličine µg/kg) mogu izazvati već spomenute vidljive promjene funkcije re-produktivnog sustava. Koncentracije mikotoksina koje su prisutne u hrani za životinje najopasnije su za svinje, dok u peradi uglavnom nisu uočeni znakovi poremećaja zdravlja i proizvodnosti. Dokazana je povezanost konzumacije konta-minirane hrane u svinja i hiperestrogenizam i to kod količina od svega 1,0 mg/kg. Veće doze toksina uzrokuju neuspješnu oplodnju, pobačaje i druge smetnje reproduktivnog sustava. Slični reproduktivni simptomi uočeni su i u krava i ovaca (El-Nezami i sur., 2002.). Internacionalna agencija za istra-živanje raka procijenila je da nema dovoljno dokaza za procjenu njegove karcinogenosti (IARC, 1993.).
Tablica 1. LD50 vrijednosti mikotoksina u pojedinih kategorija i vrsta peradi
Mikotoksin Vrsta životinje LD50* Literaturna referenca Pačići 0,34-0,56 mg/kg Patterson, 1973.
Aflatoksin B1 Pilići 6,5-16,5 mg/kg Patterson, 1973.
Fumonizin Pileći embrij 18,73 µg/jaje Henry i Wyatt, 2001. Pačići 0,5 mg/kg van der Merwe i sur., 1965. Tovni pilići 2,14-3,6 mg/kg Huff i sur., 1974. Ohratoksin A Purani 7,84 mg/kg Chang i sur., 1981.
T-2 toksin Pilići dobi 1 dan 4,97 mg/kg Hoerr i sur., 1982. Pilići dobi 7 dana 6,8 mg/kg Hoerr i sur., 1982. HT-2 toksin Pilići dobi 7 dana 7,22 mg/kg Hoerr i sur., 1982. Deoksinivalenol Pilići dobi 7 dana 140 mg/kg Hoerr i sur., 1982. Diacetoksiscirpenol Pilići dobi 7 dana 2,0 mg/kg Hoerr i sur., 1982. Zearalenone Pilići >15.000 mg/kg Scudamore, 1998.
*LD50 po kilogramu tjelesne težine

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI
101
Simptomi u peradi Prema dosadašnjim istraživanjima, alimentarna izloženost peradi različitim količinama mikotoksina (od 30 do 800 mg/kg) nije rezultirala promjenama proizvodnih parametara kao niti nastankom simptoma koji bi mogli ukazivati na moguće otrovanje. S druge strane, neki autori navode pad nesivosti i depresiju rasta peradi i pri koncentraciji od svega 0,5 mg/kg (Chi i sur., 1980.; Allen i sur., 1981.; Leeson i sur., 1995.). Kao i u slučaju deoksinivalenola, prisutnost zearalenona u hrani za životinje može se i redovito se koristi kao "bio-marker" prisutnosti nekih drugih možda štetnijih fuzarijskih mikotoksina (Romer, 1990.). Uz navedeno, kao i u slučaju drugih mikotoksina ne smije se zanemariti njegov potenci-jalni sinergistički učinak.
Ostali mikotoksini
Uz navedene, postoji velik broj toksina za koje nema dovoljno podataka o štetnim učincima za zdravlje ljudi i/ili životinja, iako postoje podaci u literaturi koji na njihov štetni učinak ukazuju. Nadalje, današnje metode otkrivanja mikotoksina su skupe i usredotočene su na manji broj tok-sina, pričem se "zanemaruje" mogući štetni učinak pojedi-nih tvari. Prema istraživanjima nekih autora, trebalo bi procijeniti potencijalno štetni učinak koji bi mogli imati mikotoksini poput patulina, citrinina, penicilinske kiseline, ergot alkaloida i drugih metabolita plijesni. Isto je moguće tek procjenom njihove učestalost preciznim i pouzdanim analitičkim metodama, ali i dokazivanjem uzročno-poslje-dične veze između prisutnosti mikotoksina u hrani i na-stanka poremećaja ili bolesti u primjerice peradi (Leeson i sur., 1995.; Hoerr, 2003.).
Sigurnost hrane, procjena rizika, kontrole i zakonski propisi
Zbog procjene da je u prosjeku svake godine gotovo 25% hrane u cijelom svijetu kontaminirano mikotoksinima, kao i da su toksikogene plijesni redovito prisutne u prirodi po-
trebne su odgovarajuće mjere kontrole i procjene učesta-losti i rizika za zdravlje ljudi i životinja. Mikotoksini se, prema nekim autorima, smatraju jednim od najznačajnijih kroničnih alimentarnih rizičnih čimbenika, značajnijim i od pesticida, aditiva hrani i drugih kontami-nanata (Kuiper-Goodman, 1995). Uz navedeno, mikotok-sini su "odgovorni" i za ekonomske gubitke budući da se značajne količine hrane moraju uništiti zbog kontamina-cije. Ponekad kontaminirana hrana završi kao hrana za životinje te nastaju problemi u uzgoju životinja (smanjeni prirast, bolesti i uginuća), a mogu rezultirati i prisutnošću rezidua u mlijeku, jajima i mesu životinja. Kao što je već navedeno, aflatoksini u hrani za krave i kokoši se bio-transformiraju u aflatoksin M1 koji se izlučuje mlijekom i jajima. Ohratoksin u hrani za svinje nakuplja se u tkivima te je moguć nalaz rezidua u mesu. Budući da je nemoguće izbjeći prisutnost plijesni i pro-izvodnju mikotoksina, uspostavljene su mjere kontrole i monitoringa učestalosti mikotoksina u hrani i hrani za ži-votinje. Maksimalne količine mikotoksina nisu jednoliko propisane u svim zemljama svijeta, a u većini zemalja uglavnom su propisane mjere za nekoliko toksina u hrani za ljude te najčešće samo za aflatoksin u hrani za životinje. U Republici Hrvatskoj primjenjuje se Pravilnik o najvećim dopuštenim količinama određenih kontaminanata u hrani (NN 146/2012.) koji je usvojio zahtjeve Europske Direk-tive 1881/2006. Za hranu za životinje u Republici Hrvat-skoj također su prihvaćene Uredbe Europske komisije o nepoželjnim tvarima u hrani za životinje (EC Direktiva 2002/32/EZ) te su Pravilnikom propisane samo maksi-malne dozvoljene količine aflatoksina B1 (tablica 2.) (NN 124/2012.). Procjena rizika mikotoksina za zdravlje u idealnim bi slu-čajevima trebala uključivati potpunu toksikološku i epide-miološku procjenu (učestalost toksina i procjenu dnevnog unosa u organizam, usporedbu toksičnih učinaka u različi-tih vrsta životinja i dr.) kao i procjenu izloženosti. Ali, u menadžmentu mikotoksina redovito je potrebo poduzeti odgovarajuće mjere prije nego svi takvi podaci uopće budu dostupni. Za životinje, kao i za ljude, određuje se odgova-
Tablica 2. Najveće dopuštene količine nepoželjnih tvari u hrani za životinje (NN 124/2012.)
Nepoželjne tvari Proizvodi namijenjeni za hranu za životinje Najveća dopuštena količina u mg/kg
(ppm), kada udio vlage u hrani za životinje iznosi 12%
Krmiva 0,02 Dopunske i potpune krmne smjese 0,01 osim: – krmnih smjesa za mliječna goveda i telad, mliječne ovce i janjad, mliječne koze i jarad, prasad i mladu perad 0,005 1. Aflatoksin B1
– krmnih smjesa za goveda (osim mliječnih goveda i teladi), ovce (osim mliječnih ovaca i janjadi), koze (osim mliječnih koza i jaradi), svinje (osim prasadi) i perad (osim mlade peradi)
0,02
2. Glavnica raži (Claviceps purpurea) Krmiva i krmne smjese koje se sastoje od nemljevenih žitarica 1000

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI 102
rajuća „sigurna doza“ čijim unosom ne bi trebalo doći do nastanka štetnih učinaka za zdravlje (Kuiper-Goodman, 1995.). Za postizanje što točnijih procjena i podaci na kojima se temelji procjena rizika trebaju biti točni, pouzdani i pro-vjerljivi. Poznato je da mikotoksini nisu jednoliko raspro-stranjeni u hrani, te je stoga pravilno uzorkovanje hrane ključno za procjenu kontaminiranosti. Ako uzorak nije pra-vilno uzorkovan može doći do pogrešne procjene kvalitete neke hrane (redovito većeg lota hrane) te tako dovesti i do neželjenih zdravstvenih i/ili ekonomskih posljedica. Kako bi se spriječile takve pogreške izrađeni su planovi uzorko-vanja temeljeni na nasumičnom uzorkovanju, koji s do-voljnom sigurnošću omogućuju pravilnu procjenu većeg lota hrane na temelju analize manjeg laboratorijskog uzorka (Dickens i sur., 1979.; Bennett i Klich, 2003.). Nakon osiguranja reprezentativnog uzorka isti je potrebno pripremiti za analizu (homogenizirati, ekstrahirati) te u ko-načnici detektirati odgovarajućim metodama. Količina mi-kotoksina može se otkriti različitim metodama uključujući tankoslojnu kromatografiju, visokoučinkovitu tekućinsku kromatografiju, masenu spektrometriju ili imunoenzimnim testovima. Iako se količina mikotoksina u velikom lotu hrane ne može utvrditi sa stopostotnom sigurnošću, postu-pak uzorkovanja redovito se smatra najvećim izvorom nesigurnosti dobivenih analitičkih rezultata.
Preventivne mjere
Mjere koje sprječavaju kontaminaciju žitarica prije i nakon žetve uključuju agrotehničke mjere, odabir sorta žitarica te primjenu preporučenih postupaka pri proizvodnji žitarica, rana sjetva, navodnjavanje pri vrućim i suhim klimatskim uvjetima kako bi se izbjegao učinak suše, smanjenje ošte-ćenja uzrokovanih insektima, rana žetva, smanjenje ošteće-nja zrna tijekom žetve. Zatim je potrebno osigurati pra-vilno sušenje i pohranu žitarica, održavati i kontrolirati čistoću prostorija za pohranu. Dekontaminacija hrane kontaminirane mikotoksinima uklju-čuje fizikalne (npr. razdvajanje oštećenih zrna), kemijske i biološke metode koje u stanovitoj mjeri mogu poboljšati kvalitetu same hrane. U današnje vrijeme poseban naglasak je na primjeni adsorbensa (organskih i neorganskih) koji mogu smanjiti negativni utjecaj mikotoksina na životinje.
Zaključak
Mikotoksini predstavljaju opasnost za zdravlje i proizvod-nost životinja te ujedno predstavljaju i javnozdravstveni problem (izravna ili neizravna izloženost ljudi putem proizvoda životinjskog podrijetla). Uz to uzrokuju i zna-čajne ekonomske gubitke za proizvođače peradi. Redovi-tim kontrolama (unutarnjim ili zakonski propisanim) u svim fazama proizvodnje hrane i hrane za životinje mogu se smanjiti neželjeni učinci mikotoksina na zdravlje ljudi i/ili životinja. Pritom osobitu pozornost treba posvetiti uzorkovanju reprezentativnog uzorka na temelju kojega
ćemo procijeniti veći lot hrane. Dostupne analitičke me-tode osiguravaju precizne i pouzdane rezultate. Ipak, tre-nutna analitika usredotočena je na mali broj mikotoksina, dok bi za stvarnu procjenu rizika trebalo analizirati veći broj uzoraka hrane i hrane za životinje na široki spektar mikotoksina. Usporedbom takvih podataka s toksikološkim podacima omogućiti će se pravodobna reakcija na pojavu kontaminacije te time osiguranje kvalitetne hrane i hrane za životinje te u konačnici zaštita zdravlja ljudi i životinja.
Literatura
ALLEN, N. K., C. J. MIROCHA, G. WEAVER, S. AAKHUS-ALLEN, F. BATES (1981): Effects of dietary zearalenone on finishing broiler chickens and young turkey poults. Poult. Sci. 60(1), 124-131.
BAYMAN, P., J. L. BAKER, M. A. DOSTER, T. J. MICHAILIDES, N. E. MAHONEY (2002): Ochratoxin production by the Aspergillus ochraceus group and Asper-gillus alliaceus. Appl. Environ. Microbiol. 68(5), 2326-2329.
BENNETT, J. W., M. KLICH (2003): Mycotoxins. Clin. Microbiol. Rev. 16(3), 497-516.
BERGERS, W. W. A., J. G. M. M. van der STAP, C. E. KIENTZ (1985): Trichothecene production in liquid stationary cultures of Fusarium tricinctum NRRL 3299 (Synonym: F. sporotrichioides): comparison of quantitative brine shrimp assay with physicochemical analysis. Appl. Environ. Microbiol. 50(3), 656-62.
BEZUIDENHOUT, S. C., W. C. A. GELDERBLOM, C. P. GORST-ALLMAN, R. M. HORAK, W. F. O. MARASAS; G. SPITELLER, R. VLEGGAAR (1988): Structure eluci-dation of the fumonisins, mycotoxins from Fusarium moniliforme. J. Chem. Soc. Chem. Commun. 11:743-745.
BLOUNT, W. P. (1961): Turkey “X” disease. Turkeys. 77(9), 52-61. BUCCI, T., D. K. HANSEN, J. B. LABORDE (1996):
Leukoencephalomalacia and hemorrhage in the brain of rabbits gavaged with mycotoxin fumonisin B1. Nat. Toxins 4(1), 51-52.
BURNS, R. P., I. P. DWIVEDI (1986): The natural occurrence of ochratoxin A and its effects in poultry. A review. Part II. Pathology and immunology. World Poult. Sci. 42, 48-62.
CHANG, C. F., J. A. DOERR, P. B. HAMILTON (1981): Experimental ochratoxicosis in turkey poultry. Poult. Sci. 60(1), 114-119.
CHI, M. S., C. J. MIROCHA, G. A. WEAVER, H. J. KURTZ (1980): Effect of zearalenone on female White Leghorn chickens. Appl. Environ. Microbiol. 39(5), 1026-1030.
Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs (Text with EEA Relevance).
DICKENS, J. W., T. B. WHITAKER, R. J. MONROE, J. N. WEAVER (1979): Accuracy of subsampling mills for granular material. J. Am. Oil Chem. 56(9), 842-844.
Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed.
EFSA (2004): Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to Aflatoxin B1 as undesirable substance in animal feed. EFSA Journal 39, 1-27.

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI
103
El-NEZAMI, H., N. POLYCHRONAKI, S. SALMINEN, H. MYKKANEN (2002): Binding rather than metabolism may explain the interaction of two food-grade Lactobacillus strains with zearalenone and its derivative (')alpha-earalenol.. Appl. Environ. Microbiol. 68(7), 3545-3549.
GELDERBLOM, W. C. A., K. JASKIEWICZ, W. F. O. MARASAS, P. G. THIEL, R. M. HORAK, R. VLEGGAAR, N. P. J. KRIEK (1988): Fumonisins – novel mycotoxins with cancer-promoting activity produced by Fusarium monili-forme. Appl. Environ. Microbiol. 54(7), 1806-1811.
GOTO, T., D. T. WICKLOW, Y. ITO (1996): Aflatoxin and cyclopiazonic acid production by a sclerotium-producing Aspergillus tamarii strain. Appl. Environ. Microbiol. 62(11), 4036-4038.
HAMILTON, P. B., W. E. HUFF, J. R. HARRIS, R. D. WYATT (1982): Natural occurrences of ochratoxicosis in poultry. Poult. Sci. 61(9), 1832-1836.
HAMILTON, P. B., H. T. TUNG, J. R. HARRIS, J. H. GAINER, W. E. DONALDSON (1972): The effect of dietary fat on aflatoxicosis in turkeys. Poult. Sci. 51(1), 165-170.
HAMILTON, P. B. (1992): Justification for mycotoxin research. Current Contents 23(46), 10.
HARRISON, L. R., B. M. COLVIN, J. T. GREENE, L. E. NEWMAN, J. R. COLE, Jr. (1990): Pulmonary edema and hydrothorax in swine produced by fumonisin B1, a toxic metabolite of Fusarium moniliforme. J. Vet. Diagn. Invest. 2(3), 217-221.
HENRY, M. H., R. D. WAYTT, O. J. FLETCHERT (2000): The toxicity of purified fumonisin B1 in broiler chicks. Poult. Sci. 79(10), 1378-1384.
HENRY, M. H., R. D. WYATT (2001): The toxicity of fumonisin B1, B2 and B3, individually and in combination, in chicken embryos. Poult. Sci. 80(4), 401-407.
HENRY, S. H., F. X. BOSCH, J. C. BOWERS (2002): Aflatoxin, hepatitis and worldwide liver cancer risks, p. 229-320. U: J. W. DeVries, M. W. Trucksess, L. S. Jackson (ur.). Mycotoxins and Food Safety. Kluwer Academic/Plenum Publications, New York, N.Y.
HIDY, P. H. R. S. BALDWIN, R. L. GREASHAM, C. L. KEITH, J. R. Mc MULLEN (1977): Zearalenone and some derivatives: production and biological activities. Adv. Appl. Microbiol. 22, 59-82.
HOERR F. J. (2003): Mycotoxicoses. U: Saif Y.M. (ur.) Diseases of Poultry. 11th ed., Ames, Iowa, USA: Iowa State Uni-versity Press, str. 1103-1132.
HOERR FJ, CARLTON WW, YAGEN B, JOFFE AZ (1982): Mycotoxicosis caused by either T-2 toxin or dia-cetoxyscirpenol in the diet of broiler chickens. Fundam. Appl. Toxicol. 2(3), 121-124.
HUFF, E. W., R. D. WYATT, T. L. TUCKER, P. B. HAMILTON (1974): Ochratoxicosis in the broiler chicken. Poult. Sci. 53(4), 1585-1591.
IARC, International Agency for Research on Cancer (1993): Monographs on the Evaluation of the Carcinogenic Risk of Chemicals to Humans. Some Naturally Occurring Subs-tances: Food Items and Constituents, Heterocyclic Amines and Mycotoxins. Lyon: IARC Press.
IARC, International Agency for Research on Cancer (1982): The evaluation of the carcinogenic risk of chemicals to humans. IARC Monograph Supplement 4. International Agency for Research on Cancer, Lyon, France.
JAVED, T, G. A. BENNET, J. L. RICHARD, M. A. DOMBRINKKURTZMAN, L. M. COTE, W. B. BUCK (1993): Mortality in broiler chicks on feed amended with Fusarium proliferatum culture material or with purified fumonisin B1 and moniliformin. Mycopathologia 123(4), 171-184.
JECFA, Joint Expert Committee on Food Additives (2001): T-2 and HT-2 toxins. Monographs & Evaluations No. 47, 2001. http://www.inchem.org/documents/jecfa/jecmono/ v47je06.htm [14.03.2014].
JEWERS, K. (1990): Mycotoxins and their effect on poultry production. Options Mediterraneennes, Ser. A. No. 7. L'aviculture en Mediterranee.
KRSKA, R., S. BAUMGARTNER, R. JOSEPHS (2001): The state-of-the-art in the analysis of type-A and type-B tricothecene mycotoxins in cereals. Fresenius J. Anal. Chem. 371(3), 285-299.
KUIPER-GOODMAN, T., P. M. SCOTT, H. WATANABE (1987): Risk assessment of the mycotoxin zearalenone. Regul. Toxicol. Pharmacol. 7(3), 253-306.
KUPIER-GOODMAN, T. (1995): Mycotoxins: risk assessment and legislation. Toxicol. Lett. 82-83, 853-859.
LEESON, S., G. DIAZ, J. D. SUMMERS (1995): Poultry Meta-bolic Disorders and Mycotoxins. Ontario, Canada: Uni-versity Books.
MARASAS, W. F. O., T. S. KELLERMAN, W. C. A. GELDERBLOM, J. A. W. COETZER, P. G. THIEL, J. J. van der LUGT (1988): Leukoencephalomalacia in horse induced by fumonisin B1 isolated from Fusarium moniliforme. Onderstepoort J. Vet. Res. 55(4), 197-203.
MARQUARDT, R. R., I A. A FROHLICH (1992): A review of recent advances in understanding ochratoxicosis. J. Anim. Sci. 70(12), 3968-3988.
PATTERSON, D. S. P. (1973): Metabolism as a factor in determining the toxic action of the aflatoxins in different animal species. Food Cosmet. Toxicol. 11(2), 287-294.
PETERSON, S. W., Y. ITO, B. W. HORN, T. GOTO (2001): Aspergillus bombycis, a new aflatoxigenic species and ge-netic variation in its sibling species, A. nomius. Mycologia 93(4), 689-703.
PITT, J. I. (1987): Penicillium viridicatum, Penicillium verru-cosum, and production of ochratoxin A. Appl. Environ. Microbiol. 53(2), 266-269.
PLUMLEE, K. H., F. D. GALEY (1994): Neurotoxic myco-toxins: a review of fungal toxins that cause neurological disease in large animals. J. Vet. Intern. Med. 8(1), 49-54.
Pravilnik o izmjenama Pravilnika o nepoželjnim tvarima u hrani za životinje kojim se provode Uredbe Europske komisije o izmjenama Direktive 2002/32/EZ Europskog parlamenta i Vijeća od 7. svibnja 2002. o nepoželjnim tvarima u hrani za životinje (124/2012).
Pravilnik o najvećim dopuštenim količinama određenih kontami-nanata u hrani (NN 146/2012).
RAWAL, S, J. E. KIM, R. Jr. COULOUMBE (2010): Aflatoxin B1 in poultry: toxicology, metabolism and prevention. Res. Vet. Sci. 89(3), 325-331.
RHEEDER, J. P., W. F. MARASAS, H. F. VISMER (2002): Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 68(5), 2102-2105.
ROMER, T. (1990): Advances in mycotoxin analysis: marking fusarium. Feed Manag. 41, 30-32.

MIKOTOKSINI – KONTROLA, PROCJENA RIZIKA I PREPORUKE U UZGOJU PERADI 104
SCF, Scientific Committee on Food (2000): Opinion of the Scientific Committee on Food on Fusarium toxins. Part 3: Fumonisin B1 (FB1). SCF/CS/CNTM/MYC/24FINAL. http://ec.europa.eu/food/fs/sc/scf/out73_en.pdf [14.03.2013.]
SCUDAMORE, K.A., NAWAZ, S., HETMANSKI, M.T. (1998): Mycotoxins in ingredients of animal feeding stuffs: II Determination of mycotoxins in maize and maize products. Food Addit. Contam. 15(1), 30-55.
SHANK, R. C., N. BHAMARAPRAVATI, J. E. GORDON, G. N. WOGAN (1972): Dietary aflatoxins and human liver cancer. IV Incidence of primary liver cancer in two municipal populations in Thailand. Food Cosmet. Toxicol. 10(2), 171-179.
SHOTWELL, L. L., C. W. HESSELTINE, M. L. GOULDEN (1969): Ochratoxin A: occurrence as natural contaminant of a corn sample. Appl. Microbiol. 17(5), 765-766.
SOKOLOVIC M., V. GARAJ-VRHOVAC, B. ŠIMPRAGA (2008): T-2 toxin: incidence and toxicity in poultry. Arh. Hig. Rada. Toksikol. 59(1), 43-52.
SQUIRE, R. A. (1981): Ranking animal carcinogens: a proposed regulatory approach. Science 214(4523), 877-880.
TRUCKSESS, M.W., L. STOLOFF, K. YOUNG, R. D. WYATT, B. L. MILLER (1983): Aflatoxicol and aflatoxins B1 and M1 in eggs and tissues of laying hens consuming aflatoxin-contaminated feed. Poult. Sci. 62(11), 2176-2182.
van der MERWE, K. J., P. S. STEYN, L. FOURIE, D. B. SCOTT, J. J. THERON (1965): Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 205(976), 1112-1113.
WEIBKIG, T. S., D. R. LEDOUX, D. R. BROWN, G. E. ROTTINGHAUS (1993): Effects of feeding Fusarium moniliforme culture material, containing known levels of fumonisin B1, on the young broiler chick. Poult. Sci. 72(3), 456-466.
WHO, World Health Organisation (1998): Safety evaluation of certain food additives and contaminants. WHO Food Additives Series 40. http://www.inchem.org/documents/ /jecfa/ jecmono/v040je16.htm [14.03.2013.]
MYCOTOXINS – CONTROL, RISK ASSESSMENT AND PREVENTIVE MEASURES IN POULTRY PRODUCTION Summary Mycotoxins are secondary metabolites of diverse mould species that can cause illness in humans and animals. This paper gives an overview of the most important mycotoxins in intensive poultry production. Mentioned will be their production, toxicity, toxic effects in poultry and risk assessment. The paper includes an overview of the current legal and preventive control measures applied in the Republic of Croatia. Key words: mycotoxins, poultry, mycotoxicosis, moulds

PERADARSKI DANI 2013.
105
KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ
Jelka Pleadin1, Manuela Zadravec1, Marijana Sokolović2, Nina Perši1, Ana Vulić1
1 Odjel za veterinarsko javno zdravstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska
2 Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak
Mikotoksini kao sekundarni toksični metaboliti plijesni različitih rodova čest su uzrok kontaminacije hrane za životinje, uključujući i hrane za perad. Cilj istraživanja bio je odrediti razinu kontaminacije hrane za perad mikotoksinima u Hrvatskoj tijekom petogodišnjeg razdoblja uzorkovanja. Ukupno je analizirano 255 uzoraka dostavljenih u razdoblju od 2007. do 2012. godine iz različitih tvornica stočne hrane iz svih hrvatskih županija. Primjenom validiranih imunoenzimskih metoda ELISA kvantitativno su određene koncentracije aflatoksina B1 (AFB1), okratoksina A (OTA), zearalenona (ZEA), deoksinivalenola (DON), fumonizina (FUM) i T-2 toksina (T-2). Dobiveni rezultati ukazali su na najveću razinu i učestalost kontaminacije hrane za perad mikotoksinom DON (1126±825 μg/kg), te FUM (586±448 μg/kg) i ZEA (165±189 μg/kg), a određene su značajno niže koncentracije T-2 (13,5±11,6 μg/kg), AFB1 (3,02±2,45 μg/kg) i OTA (1,78±1,43 μg/kg). Iako su u pojedinim uzorcima utvrđene povišene razine fuzarijskih mikotoksina, naročito u uzorcima iz sjeverozapadnog dijela zemlje nakon izrazito hladne i vlažne 2010. godine, njihove koncentracije bile su u skladu s dopuštenim ili preporučenim vrijednostima u Republici Hrvatskoj kao i Europskoj uniji. Ključne riječi: hrana za perad, mikotoksini, Hrvatska, ELISA Uvod Mikotoksini su proizvodi toksikogenih plijesni, najčešće iz rodova Aspergillus, Penicillium i Fusarium, a čine vrlo česte kontaminante hrane i hrane za životinje. Među najzastup-ljenijim mikotoksinima u hrani za životinje su aflatoksin B1, okratoksin A, zearalenon, T-2 toksin, deoksinivalenol i fumonizin, a uzrokuju značajne gubitke tijekom procesa proizvodnje i skladištenja u stočarskoj industriji. Čimbenici koji utječu na nastanak plijesni te posljedičnu tvorbu mikotoksina odnose se na sadržaj vlage, temperaturu, razinu aeracije, prisutnost gljivičnih spora, insekata i meha-ničkih oštećenja zrna (CAST, 2003.; Sforza i sur., 2006.). Problem mikotoksina naročito je izražen tijekom kišovitih godina s izraženim temperaturnim promjenama (Pleadin i sur., 2012.a; Pleadin i sur., 2012.b), kada značajno poraste kontaminacija plijesnima uzrokujući pojačanu tvorbu miko-toksina (Pepeljnjak i sur., 1979.; Pepeljnjak i sur., 2008.; Pleadin i sur., 2013.). Osobito značenje pridaje se i isto-dobnom pojavljivanju nekoliko fuzarijskih mikotoksina u istom uzorku žitarice ili hrane za životinje, često uz prisut-
nost aflatoksina B1 (Placinta i sur., 1999.). Iako su u pret-hodnom razdoblju razvijene brojne specifične i osjetljive analitičke metode za određivanje količine mikotoksina u hrani za životinje, i nadalje je najšira primjena imunoloških kvantitativnih ili polukvantitativnih metoda (Krska i sur., 2007.). Istraživanja su pokazala da mikotoksini uzrokuju bolesti na-zvane mikotoksikozama te da imaju genotoksično, nefro-toksično, citotoksično, estrogeno i teratogeno djelovanje kod ljudi i životinja (Morgavi i Riley, 2007.). Istodobno mogu djelovati na različite načine i na više ciljnih tkiva u orga-nizmu, što ovisi o toksičnosti pojedinog mikotoksina te dozi i vremenu izloženosti. Izravne posljedice ishrane životinja kontaminiranom hranom za životinje odnose se na sma-njenje odnosno odbijanje unosa hrane i njenu lošu konver-ziju, smanjenu tjelesnu težinu životinja, povećanu inciden-ciju bolesti i smanjene reproduktivne sposobnosti (Binder i sur., 2007.). Među navedenim mikotoksinima aflatoksin B1 je najjači hepatokarcinogen u sisavaca te je prema Inter-national Agency of Research on Cancer klasificiran kao karcinogen 1. skupine (IARC, 1993.).
Doc. dr. sc. Jelka Pleadin; Hrvatski veterinarski institut, Savska cesta 143, 10000 Zagreb; Tel.: 01/6123-626; Fax: 01/6123-670; E-mail: [email protected]

KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ 106
Višegodišnja istraživanja u Republici Hrvatskoj govore o učestaloj kontaminaciji hrane (žitarica) i hrane za životinje mikotoksinima te upućuju na nužnost sustavne kontrole u cilju prevencije negativnih učinaka na zdravlje ljudi i ži-votinja, kao i ekonomskih gubitaka u stočarskoj industriji. Vrlo malo podataka u Europi, kao i u Hrvatskoj, objavljeno je o kontaminaciji hrane za perad (Zinedine i Mañes, 2009.; Pleadin i sur., 2012.c). Stoga je cilj ovoga istraživanja bio utvrditi razinu kontaminacije hrane za perad mikotoksinima u uzorcima iz tvornica i mješaona stočne hrane iz različitih hrvatskih županija tijekom posljednjeg petogodišnjeg razdoblja.
Materijal i metode
Uzorci za analizu Istraživanje je provedeno na ukupno 255 uzoraka hrane za perad dostavljenih na analizu u razdoblju od 2007. do 2012. godine. Uzorci su uzorkovani u tvornicama i mješaonama stočne hrane sukladno normama ISO 6497:2002 i ISO 6498:1998 iz različitih županija Republike Hrvatske podije-ljenih u četiri regije (sjeverozapadna, središnja, istočna i juž-na). Usitnjavanje uzorka u fini prah provedeno je uporabom analitičkog mlina (Cylotec 1093, Tecator, Švedska). Uzorci su čuvani na tamnom i hladnom mjestu do provedbe analiza.
Kemikalije Imunoenzimski (ELISA) kitovi nabavljeni su od R-Bio-pharm (Darmstadt, Njemačka). Kitovi sadržavaju mikrotitra-cijsku ploču s 96 jažica, 6 standardnih otopina različitih koncentracija mikotoksina (ng/mL), enzimski konjugat, anti-tijelo, otopinu supstrata i kromogena, sol za pripremu pufera za ispiranje (10 mM fosfatni pufer, pH=7,4) i stop otopinu (1 N sulfatna kiselina). Standardi mikotoksina za validaciju ispitnih metoda nabavljeni su od Sigma-Aldrich Chemie GmbH (Steinheim, Njemačka). Sve ostale kemikalije ko-rištene u analizama bile su analitičke čistoće.
Ekstrakcija mikotoksina U analizama AFB1, FUM, ZEA i T-2 ekstrakcija je pro-vedena pomoću 25 mL 70%-tnog metanola. U ekstrakciji OTA korišteno je 100 mL 0,13 M natrij-hidrogen karbo-natnog pufera (pH 8,1), a pri određivanju DON 25 mL destilirane vode. Uzorci su određeno vrijeme ostavljeni
mućkati na tresilici pri brzini od 300 o/min (HS 260 Control, Ika). Sadržaj je potom profiltriran (crna vrpca), izdvojeni supernatant razrijeđen redestiliranom vodom, te je 50 µL otopine korišteno u testu ELISA.
Metoda ELISA Određivanje količine mikotoksina provedeno je prema upu-tama proizvođača ELISA kita uz uporabu automatiziranog kemijskog analizatora ChemWell 2910 (Awareness Techno-logy, USA). Nakon dodatka svih komponenti iz kita reakcija je zaustavljena dodatkom stop otopine te je apsorbancija izmjerena pri valnoj duljini od 450 nm. Nakon konstruiranja baždarne krivulje određena je koncentracija mikotoksina u otopini (μg/L) te je uzimajući u obzir faktor razrijeđenja uzorka utvrđena koncentracija u hrani za perad (μg/kg). Do-bivene koncentracije korigirane su za vrijednost iskorištenja analitičke metode.
Statistička obrada podataka Statistička analiza provedena je korištenjem programa Sta-tistica Ver. 7 software (StatSoft Inc. Tulsa, OK, 1984-2004, USA). Za određivanje razlike u kvantitativnim vrijedno-stima mikotoksina primijenjen je t-test, a statistički značajne razlike izražavane su na razini vjerojatnosti 0,05.
Rezultati
Vrijednosti validacijskih parametara primijenjenih analitič-kih metoda za pojedini mikotoksin prikazane su u tablici 1. Iskorištenje analitičkih metoda kretalo se u rasponu od 78,9% do 98,5%, a ponovljivost određivanja u rasponu od 76,5% do 96,3%, s koeficijentima varijabilnosti (CV) ma-njim od 10%. Koncentracija mikotoksina određena u hrani za perad, broj uzoraka u kojima je otkriven pojedini mikotoksin u pro-matranom razdoblju (% pozitivnih uzoraka) te srednje (mean), minimalne (min) i maksimalne (max) vrijednosti koncentracija, kao i najveće dopuštene količine (NDK) ili najveće preporučene količine (NPK) prikazani su u tablici 2. Tablica 3. prikazuje koncentracije mikotoksina po regijama Hrvatske određene u cjelovitom promatranom razdoblju Na slici 1. prikazane su srednje vrijednosti (±SD) koncen-tracije analiziranih mikotoksina u hrani za perad po go-dinama od 2007. do 2012.
Tablica 1. Vrijednosti validacijskih parametara metoda ELISA Table 1. Values of validation parameters for ELISA methods
Parametar / Parameter LOD (μg/kg)
Iskorištenje / Recovery (%)
CV (%)
Ponovljivost / Repeatability (%)
CV (%)
AFB1 1 82,6 5,3 79,8 9,2 OTA 1,3 93,7 6,4 92,8 8,9 ZEA 1,8 93,2 4,8 90,1 6,9 DON 150 98,5 5,1 96,3 7,8 FUM 25 78,9 6,3 76,5 8,2 T-2 4 94,5 4,3 93,1 6,5
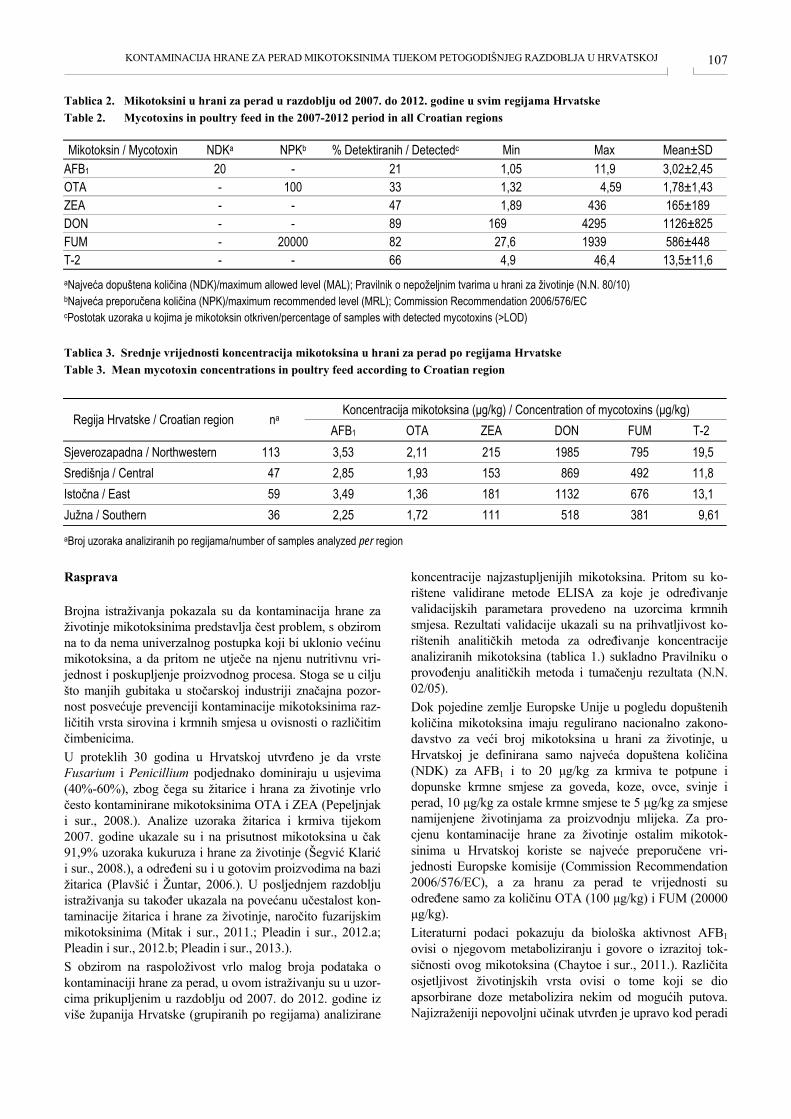
KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ
107
Tablica 2. Mikotoksini u hrani za perad u razdoblju od 2007. do 2012. godine u svim regijama Hrvatske Table 2. Mycotoxins in poultry feed in the 2007-2012 period in all Croatian regions
Mikotoksin / Mycotoxin NDKa NPKb % Detektiranih / Detectedc Min Max Mean±SD AFB1 20 - 21 1,05 11,9 3,02±2,45 OTA - 100 33 1,32 4,59 1,78±1,43 ZEA - - 47 1,89 436 165±189 DON - - 89 169 4295 1126±825 FUM - 20000 82 27,6 1939 586±448 T-2 - - 66 4,9 46,4 13,5±11,6 aNajveća dopuštena količina (NDK)/maximum allowed level (MAL); Pravilnik o nepoželjnim tvarima u hrani za životinje (N.N. 80/10) bNajveća preporučena količina (NPK)/maximum recommended level (MRL); Commission Recommendation 2006/576/EC cPostotak uzoraka u kojima je mikotoksin otkriven/percentage of samples with detected mycotoxins (>LOD) Tablica 3. Srednje vrijednosti koncentracija mikotoksina u hrani za perad po regijama Hrvatske Table 3. Mean mycotoxin concentrations in poultry feed according to Croatian region
Koncentracija mikotoksina (μg/kg) / Concentration of mycotoxins (μg/kg) Regija Hrvatske / Croatian region na
AFB1 OTA ZEA DON FUM T-2 Sjeverozapadna / Northwestern 113 3,53 2,11 215 1985 795 19,5 Središnja / Central 47 2,85 1,93 153 869 492 11,8 Istočna / East 59 3,49 1,36 181 1132 676 13,1 Južna / Southern 36 2,25 1,72 111 518 381 9,61 aBroj uzoraka analiziranih po regijama/number of samples analyzed per region Rasprava
Brojna istraživanja pokazala su da kontaminacija hrane za životinje mikotoksinima predstavlja čest problem, s obzirom na to da nema univerzalnog postupka koji bi uklonio većinu mikotoksina, a da pritom ne utječe na njenu nutritivnu vri-jednost i poskupljenje proizvodnog procesa. Stoga se u cilju što manjih gubitaka u stočarskoj industriji značajna pozor-nost posvećuje prevenciji kontaminacije mikotoksinima raz-ličitih vrsta sirovina i krmnih smjesa u ovisnosti o različitim čimbenicima. U proteklih 30 godina u Hrvatskoj utvrđeno je da vrste Fusarium i Penicillium podjednako dominiraju u usjevima (40%-60%), zbog čega su žitarice i hrana za životinje vrlo često kontaminirane mikotoksinima OTA i ZEA (Pepeljnjak i sur., 2008.). Analize uzoraka žitarica i krmiva tijekom 2007. godine ukazale su i na prisutnost mikotoksina u čak 91,9% uzoraka kukuruza i hrane za životinje (Šegvić Klarić i sur., 2008.), a određeni su i u gotovim proizvodima na bazi žitarica (Plavšić i Žuntar, 2006.). U posljednjem razdoblju istraživanja su također ukazala na povećanu učestalost kon-taminacije žitarica i hrane za životinje, naročito fuzarijskim mikotoksinima (Mitak i sur., 2011.; Pleadin i sur., 2012.a; Pleadin i sur., 2012.b; Pleadin i sur., 2013.). S obzirom na raspoloživost vrlo malog broja podataka o kontaminaciji hrane za perad, u ovom istraživanju su u uzor-cima prikupljenim u razdoblju od 2007. do 2012. godine iz više županija Hrvatske (grupiranih po regijama) analizirane
koncentracije najzastupljenijih mikotoksina. Pritom su ko-rištene validirane metode ELISA za koje je određivanje validacijskih parametara provedeno na uzorcima krmnih smjesa. Rezultati validacije ukazali su na prihvatljivost ko-rištenih analitičkih metoda za određivanje koncentracije analiziranih mikotoksina (tablica 1.) sukladno Pravilniku o provođenju analitičkih metoda i tumačenju rezultata (N.N. 02/05). Dok pojedine zemlje Europske Unije u pogledu dopuštenih količina mikotoksina imaju regulirano nacionalno zakono-davstvo za veći broj mikotoksina u hrani za životinje, u Hrvatskoj je definirana samo najveća dopuštena količina (NDK) za AFB1 i to 20 μg/kg za krmiva te potpune i dopunske krmne smjese za goveda, koze, ovce, svinje i perad, 10 μg/kg za ostale krmne smjese te 5 μg/kg za smjese namijenjene životinjama za proizvodnju mlijeka. Za pro-cjenu kontaminacije hrane za životinje ostalim mikotok-sinima u Hrvatskoj koriste se najveće preporučene vri-jednosti Europske komisije (Commission Recommendation 2006/576/EC), a za hranu za perad te vrijednosti su određene samo za količinu OTA (100 μg/kg) i FUM (20000 μg/kg). Literaturni podaci pokazuju da biološka aktivnost AFB1 ovisi o njegovom metaboliziranju i govore o izrazitoj tok-sičnosti ovog mikotoksina (Chaytoe i sur., 2011.). Različita osjetljivost životinjskih vrsta ovisi o tome koji se dio apsorbirane doze metabolizira nekim od mogućih putova. Najizraženiji nepovoljni učinak utvrđen je upravo kod peradi

KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ 108
(patke > brojleri > nesilice), a potom kod svinja (Guen-gerich, 2003.). Ranija istraživanja govore o razini konta-minacije hrane za perad mikotoksinom AFB1 u rasponu od 20 do 200 µg/kg, a u četiri uzorka određena je razina kontaminacije u rasponu od 2000 do 5625 µg/kg, nadalje povezane s kliničkom aflatoksikozom brojlera (Kichou i Walser, 1993.). U ovom istaraživanju utvrđena je općenito niska razina kontaminacije hrane za perad mikotoksinom AFB1 (tablica 2.), s maksimalnom koncentracijom (11,9 µg/kg) određenom u istočnom dijelu Hrvatske, što je ipak značajno manja vrijednost od propisane NDK (20 µg/kg). Značajno veće razine mikotoksina određene u hrani za perad odnose se na one roda Fusarium. Podatke o kontaminaciji različitih žitarica i proizvoda na bazi žitarica ovim miko-toksinima objavili su i drugi europski i hrvatski autori (Tanaka i sur., 1990., Placinta i sur., 1999.; Domijan i sur., 2005.; Sokolović i Šimpraga, 2006.; Mitak i sur., 2011.). Također, Schothorst i Egmond (2004.) dali su pregled razina fuzarijskih mikotoksina određenih u usjevima 12 europskih zemalja te utvrdili da je DON najzastupljeniji trihotecen u žitaricama bez obzira na zemlju u kojoj je istraživanje provedeno. Binder i sur. su 2007. objavili rezultate istra-živanja o pojavljivosti fuzarijskih mikotoksina u Europi s obzirom na geografsko podrijetlo. Najveće koncentracije određene su za DON i iznosile su 5510 µg/kg u Sjevernoj Europi, 8020 µg/kg u Središnjoj Europi te 3036 µg/kg u regiji Južna Europa s Mediteranom. Najveće koncentracije ZEA i FUM određene su u Južnoj Europi s Mediteranom i iznosile su 2348 µg/kg za ZEA i 3120 µg/kg za FUM (Binder i sur., 2007.). Istraživanje Mankevičiene i sur. (2007.) provedeno na uzorcima žitarica koje su se koristile za proizvodnju hrane za životinje (2004. i 2005.) također je rezultiralo visokom kontaminacijom mikotoksinom DON, određenom u 62,5%-100% u količini do 1121 µg/kg. U hrani za perad iz ovoga istraživanja određena je također najveća prosječna količina za DON (1126±825 µg/kg) s maksimalnom vrijednošću 4295 µg/kg (tablica 2.). Desetogodišnja istraživanja kontaminacije mikotoksinom ZEA s područja Republike Hrvatske u kukuruzu i krmnim smjesama ukazala su na kontaminaciju 21,19% uzoraka ku-kuruza i 14,74% krmnih smjesa i najvećom koncentracijom od 1200 µg/kg (Mitak i sur., 2001.). Daljnja istraživanja također su ukazala na kontaminaciju mikotoksinom ZEA oko 20%-25% uzoraka kukuruza (Mitak i sur., 2011.). Re-zultati ovoga istraživanja ukazali su na prosječnu razinu kontaminacije ovim mikotoksinom od 165±189 µg/kg, s maksimalnom koncentracijom 436 µg/kg i većim udjelom uzoraka u kojima je otkriven (47%). Međutim, pritom je važno istaknuti i da se posljednjih godina u analitici miko-toksina koriste značajno osjetljivije metode ELISA s nižim granicama detekcije, što zasigurno rezultira i većim brojem uzoraka u kojima su mikotoksini otkriveni u odnosu na ranije razdoblje. Domijan i sur. (2005.) analizirali su uzorke kukuruza iz raz-ličitih dijelova Hrvatske prikupljene nakon jesenske žetve. Utvrđene su visoke koncentracije fumonizina B1 u rasponu
od 196,8 µg/kg do 1377,6 µg/kg uz njegovo otkrivanje u svim analiziranim uzorcima. Pleadin i sur. (2012.c) odredili su u hrani za perad prosječnu količinu FUM od 643±768 µg/kg. Kontaminacija FUM u ovom istraživanju određena je u 82% uzoraka, s prosječnom količinom 586±448 µg/kg i rasponom od 27,6 µg/kg do 1939 µg/kg. Svi analizirani uzorci udovoljavali su najvećim preporučenim količinama ovog mikotoksina u hrani za perad (2006/576/EC). U Europi je provedeno vrlo malo istraživanja o pojavljivosti T-2, a imajući u vidu utvrđenu toksičnost ovog mikotoksina potrebno je daljnje prikupljanje podataka i definiranje zakonskih vrijednosti najvećih dozvoljenih količina (Vulić i sur., 2011.). Objavljeni podaci o zastupljenosti T-2 toksina u Hrvatskoj, određeni u različitim vrstama krmnih smjesa tijekom razdoblja od 1998. do 2004. godine, govore o vri-jednostima od 100 do 500 µg/kg (Sokolović i Šimpraga, 2006.). U istraživanju Binder i sur. (2007.), najveće koncentracije T-2 toksina od 1776 µg/kg određene su u području Sjeverne Europe. U ovom istraživanju određena je prosječna količina T-2 od 13,5±11,6 µg/kg, podjednaka re-zultatima dobivenim u hrani za perad iz istraživanja Pleadin i sur.(2012.c). Tablica 3. prikazuje koncentracije mikotoksina po regijama Hrvatske određene u cjelovitom promatranom razdoblju, ukazujući na najveću razinu kontaminacije hrane za perad iz sjeverozapadne regije za sve analizirane mikotoksine. Me-đutim, u ostalim regijama je uzorkovan i značajno manji broj uzorka. Veći broj uzoraka iz drugih regija (središnja, istočna i južna) dao bi zasigurno utemeljeniju spoznaju i bo-lju usporedivost rezultata s obzirom na razinu kontaminacije mikotoksinima, ali ujedno u sjeverozapadnom dijelu Hrvat-ske nalazi se najveći broj mješaona hrane za perad u udnosu na ostale regije. Uspoređujući koncentracije određivanih mikotoksina u hrani za perad po godinama utvrđena je statistički značajno veća (p<0,05) koncentracija mikotoksina u 2010. godini u odnosu na ostale godine u promatranom razdoblju (slika 1.). Tije-kom 2010. godine utvrđene su značajno veće razine konta-minacije fuzarijskim mikotoksinima u odnosu na ostale godine, što se može obijasniti izrazito kišovitom i vlažnom 2010. godinom (podaci Državnog hidrometerološkog zavo-da). Klimatske prilike kao značajan čimbenik za tvorbu plijesni te posljedično tvorbu mikotoksina zasigurno su mogle uzrokovati povišenu kontaminaciju hrane za perad. Ranije provedena istraživanja u Hrvatskoj tijekom vlažnih godina, za vrijeme produljenih zima i temperaturnih osci-lacija, također su ukazala na povećane koncentracije miko-toksina. Tako su tijekom izuzetno vlažne 1978. godine maksimalne koncentracije u žitaricama i krmivima za OTA iznosile 68000,9 µg/kg, a za ZEA 275000,8 µg/kg. Godine 1980. određena je količina OTA od 4000,7 µg/kg, a izrazito hladne i vlažne 2004. godine određen je ZEA u koncentraciji od 29000,43 µg/kg (Pepeljnjak i sur., 2008.). U ovom istra-živanju određene su značajno manje količine ovih miko-toksina u odnosu na navedene, a količina OTA ujedno je bila značajno manja i u odnosu na NPK u hrani za perad (100 µg/kg). Istraživanje je pokazalo općenito najveću za-

KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ
109
stupljenost DON, koji je otkriven u 89% analiziranih uzo-raka s najvećom razinom 2010. godine (2675±3115 µg/kg), značajno većom u odnosu na ostale godine.
Zaključak
Dobiveni rezultati ukazali su na najveću učestalost kontami-nacije hrane za perad deoksinivalenolom, potom fumonizi-nom i zearalenonom, dok su razine T-2 toksina, aflatoksina B1 i okratoksina A značajno manje. Iako su u pojedinim uzorcima utvrđene povišene razine fuzarijskih mikotoksina, naročito u uzorcima iz sjeverozapadnog dijela zemlje tije-kom izrazito hladne i vlažne 2010. godine, njihove kon-centracije bile su u skladu s dopuštenim ili preporučenim
vrijednostima u Republici Hrvatskoj, kao i u Europskoj uniji. U cilju zaštite zdravlja ljudi i životinja nužna je pro-vedba sustavnog nadzora kontaminacije mikotoksinima, općenito hrane za životinje, a time i hrane za perad.
Literatura
BINDER, E. M., L. M. TAN, L. J. CHIN, J. HANDL, J. RICHARD (2007): Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 137, 265-282.
CHAYTOR A. C., J. A. HANSEN, E. van HEUGTEN, M. TODD SEE, S. W. KIM (2011): Occurrence and decontamination of mycotoxins in swine feed. Asian Aust. J. Anim. Sci. 24, 723-738.
Slika 1. Srednje vrijednosti koncentracije mikotoksina (±SD) u Hrvatskoj po godinama od 2007. do 2012. Figure 1. Mean mycotoxin concentrations (±SD) in Croatia according to year from 2007 to 2012.

KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ 110
DOMIJAN, A. M., M. PERAICA, B. CVJETKOVIĆ, S. TURČIN, Ž. JURJEVIĆ, D. IVIĆ (2005): Mould contamination and co-occurrence of mycotoxins in maize grain in Croatia. Acta. Pharm. 55, 349-356.
EC (2006): Commission Recommendation 2006/576/EC of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Official Journal of the European Union L 229/7.
GUENGERICH, F. P. (2003): Cytochrome P450 oxidations in the generation of reactive electrophiles: epoxidation and related reactions. Arch. Biochem. Biophys. 409, 59-71.
IARC (International Agency for Research on Cancer) (1993): Some Naturally Occurring Substances: Food Items and Constituents, Heterocyclic Aromatic Amines and Mycotoxins. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans, Vol. 56. IARC, Lyon.
KICHOU, F., M. M. WALSER (1993): The natural occurrence of aflatoxin B1 in Moroccan poultry feeds. Vet. Hum. Toxicol. 35, 105-108.
KRSKA, R., E. WELZIG, H. BOUDRA (2007): Analysis of Fusa-rium toxins in feed. Anim. Feed Sci. Technol. 137, 241-264.
MANKEVIČIENE A, B. BUTKUTĖ, Z. DABKEVIČIUS, S. SUPRONIENĖ (2007): Fusarium mycotoxins in Lithuanian cereals from the 2004-2005 harvests. Ann. Agric. Environ Med. 14, 103-107.
MITAK, M., T. GOJMERAC, P. PAVIČIĆ, S. TOPOLKO (2001): Kontaminacija krmiva i krmnih smjesa za svinje zearale-nonom u razdoblju od 1990-1999. godine. Veterinarska stanica 32, 205-209.
MITAK, M., J. PLEADIN, N. PERŠI, A. VULIĆ, M. ZADRA-VEC (2011): Mycotoxins in feed commodities and mixtures during 2009 and 2010. Veterinarska stanica 42, 139-145.
MORGAVI, D.P., R.T. RILEY (2007): A historical overview of field disease outbreaks known or suspected to be caused by consumption of feeds contaminated with Fusarium toxins. Anim. Feed Sci. Technol. 137, 201-212.
PEPELJNJAK, S., S. ČUTURIĆ, S. TOPOLKO, M. MUNK (1979): Faktori koji utječu na stvaranje mikotoksina kod kukuruza. Krmiva 5, 109-112.
PEPELJNJAK, S., Z. CVETNIĆ, M. ŠEGVIĆ-KLARIĆ (2008): Okratoksin i zearalenon: kontaminacija žitarica i krmiva u Hrvatskoj (1977-2007) i utjecaj na zdravlje životinja i ljudi. Krmiva 3, 147-159.
PLACINTA, C. M., J. P. F. D'MELLO, A. M. C. MacDONALD (1999): A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 78, 21-37.
PLAVŠIĆ, F., I. ŽUNTAR (2006): Uvod u analitičku toksiko-logiju, Zagreb: Školska knjiga.
PLEADIN, J., N. PERŠI, M. MITAK, M. ZADRAVEC, M. SOKOLOVIĆ, A. VULIĆ, V. JAKI, M. BRSTILO (2012a): The natural occurrence of T-2 toxin and fumonisins in maize samples in Croatia. Bull. Environ. Contam. Toxicol. 88, 863-866.
PLEADIN, J., M. SOKOLOVIĆ, N. PERŠI, M. ZADRAVEC, V. JAKI, A. VULIĆ (2012b): Contamination of maize with deoxynivalenol and zearalenone in Croatia. Food Control 28, 94-98.
PLEADIN, J., N. PERŠI, A. VULIĆ, M. ZADRAVEC (2012c): Survey of mycotoxin feed contamination in Croatia. Bio-technol. Anim. Husban. 28(2), 167-177.
PLEADIN, J., N. VAHČIĆ, N. PERŠI, D. ŠEVELJ, K. MARKOV, J. FRECE (2013): Fusarium mycotoxins' occurrence in cereals harvested from Croatian fields. Food Control 32, 49-54.
Narodne novine (02/05): Pravilnik o provođenju analitičkih metoda i tumačenju rezultata. Narodne novine.
Narodne novine (78/11): Pravilnik o izmjenama i dopunama pra-vilnika o najvećim dopuštenim količinama određenih konta-minanata u hrani. Narodne novine.
SCHOTHORST, R. C., H. P. EGMOND (2004): Report from SCOOP task 3.2.10. Collection of occurrence data of Fusarium toxins in food and assessment of dietary intake by the population of EU member states, subtask: trichothecenes. Toxicol. Lett. 153, 133-143.
SFORZA, S., C. Dall' ASTRA, R. MARCHELLI (2006): Recent advances in mycotoxin determination in food and feed by hyphenated chromatographic techniques/mass spectrometry. Mass Spectrom. Rev. 25, 54-76.
SOKOLOVIĆ, M., B. ŠIMPRAGA (2006): Survey of tricho-thecene mycotoxins in grains and animal feed in Croatia by thin layer chromatography. Food Control 17, 733-740.
ŠEGVIĆ KLARIĆ, M., S. PEPELJNJAK, Z. CVETNIĆ, I. KO-SALEC (2008): Komparacija ELISA i TLC/HPLC metoda za određivanje zearalenona i okratoksina u žitaricama i krmi. Krmiva 50, 235-244.
TANAKA, T., S. YAMAMOTO, A. HASEGAWA, N. AOKI, J.R. BESLING, Y. SUGUIRA, Y. UENO (1990): A survey of the natural occurrence of Fusarium mycotoxins, deoxynivalenon, nivalenon and zearalenone, in cereals harvested in the Netherlands. Mycopathologia 110, 19-22.
CAST (2003): Mycotoxins: risks in plant, animal and human systems. Council for Agricultural Science and Technology. Task Force Report No. 139. Ames, IA.
VULIĆ, A., J. PLEADIN, N. PERŠI (2011): Determination of T-2 and HT-2 toxins in commodities and feed in Croatia. Bull. Environ. Contam. Toxicol. 86, 294-297.
ZINEDINE, A., J. MAÑES (2009): Occurrence and legislation of mycotoxins in food and feed from Morocco. Food Control 20, 334-344.
CONTAMINATION OF POULTRY FEED WITH MYCOTOXINS DURING A FIVE-YEAR PERIOD IN CROATIA Summary Mycotoxins as secondary metabolites of toxic moulds from different genera are frequent causes of feed contamination, including poultry feed. The aim of this study was to determine the level of contamination of poultry feed with mycotoxins in Croatia during a five-year sampling period. A total of 255 samples submitted during the 2007-2012 period from different

KONTAMINACIJA HRANE ZA PERAD MIKOTOKSINIMA TIJEKOM PETOGODIŠNJEG RAZDOBLJA U HRVATSKOJ
111
feed factories in all Croatian regions were analyzed by use of validated immunoenzymatic ELISA methods. The concentrations of aflatoxin B1 (AFB1), ochratoxin A (OTA), zearalenone (ZEA), deoxynivalenol (DON), fumonisins (FUM) and T-2 toxin (T-2) were quantitatively determined. The results showed the highest level and frequency of contamination of poultry feed with DON (1126±825 mg/kg), FUM (586±448 mg/kg) and ZEA (165±189 mg/kg), and significantly lower concentrations of T-2 (13.5±11.6 mg / kg), AFB1 (3.02±2.45 mg/kg) and OTA (1.78±1.43 mg/kg). Although elevated levels of Fusarium mycotoxins were found in some of the samples, especially those from the northwestern part of the country after a very cold and wet year 2010, their concentrations were in accordance with the permitted or recommended values in the Republic of Croatia and in the European Union. Key words: poultry feed, mycotoxins, Croatia, ELISA

PERADARSKI DANI 2013. 112
USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS
Marija Berendika, Marijana Sokolović, Borka Šimpraga, Fani Krstulović
Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak Bakterije roda Salmonella jedne su od najčešćih uzročnika alimentarnih infekcija u ljudi. Pritom se hrana životinjskog podrijetla (meso, jaja, mlijeko i mliječni proizvodi) smatra glavnim izvorom infekcije u ljudi. U primarnoj peradarskoj proizvodnji potrebno je redovito kontrolirati moguće izvore infekcije uključujući i hranu za životinje. U ovom radu procijenjena je uspješnost izdvajanja bakterije Salmonella Enteritidis iz uzoraka hrane za životinje pomoću standardne referentne horizotalne metode HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008, te alternativnih brzih metoda VIDAS® UP Salmonella (SPT), RapidChek® SELECTTM Salmonella i RapidChek® SELECTTM Salmonella Enteritidis. Korišteni su uzorci hrane za životinje umjetno kontaminirani različitim brojem bakterijskih kolonija (dvije razine kontaminacije) te smo usporedili specifičnost, osjetljivost i točnost navedenih metoda. Na temelju dobivenih rezultata može se zaključiti da su sve četiri metode bile jednako uspješne u izdvajanju bakterije Salmonella Enteritidis iz uzoraka hrane za životinje te da alternativne metode s odgovarajućom specifičnošću, osjetljivošću i točnošću u značajno kraćem roku od referentne metode omogućavaju otkrivanje salmonela u uzorcima hrane za životinje.
Ključne riječi: Salmonella Enteritidis, hrana za životinje, ISO, VIDAS, RapidChek
UVOD Bakterije roda Salmonella rasprostranjene su u čitavom svijetu i čest su uzročnik bolesti u ljudi i životinja. Prema trenutnoj klasifikaciji rod Salmonella podijeljen je u dvije vrste: Salmonella enterica i Salmonella bongori (Grimont i Weill, 2007.). S. enterica podijeljena je u šest podvrsta (enterica, salamae, arizonae, diarizonae, houtenae i indica). Dosad je izdvojeno više od 2.500 serotipova bakterije od kojih većina pripada podvrsti Salmonella enterica. Bakterije roda Salmonella koje nisu specifične za pojedinog domaćina mogu inficirati različite vrste životinja te ljude. Ti serotipovi su uzročnici bolesti u ljudi uzroko-vanih hranom, a hrana životinjskog podrijetla (meso, jaja, mlijeko i mliječni proizvodi) smatra se glavnim izvorom infekcije u ljudi. U posljednjih nekoliko godina serotip Salmonella Enteritidis najčešći je uzročnik obolijevanja u ljudi. Od svih zabilježenih slučajeva obolijevanja u ljudi na području zemalja Europske Unije ovaj serotip uzrokovao je infekcije u 52,3% slučajeva tijekom 2009. godine, odnosno 45% slučajeva tijekom 2010. godine. Prisutnost serotipa S. Enteritidis dokazana je u uzorcima jaja i mesu peradi (EFSA, 2012.). Do kontaminacije mesa i jaja namijenjenih
prehrani ljudi može doći u klaonicama, tijekom prerade i pripreme hrane ili križnom kontaminacijom. U klaonicama dovoljan je mali broj zaraženih ptica za kontaminaciju čitave proizvodne linije. U primarnoj proizvodnji do pri-jenosa bakterija roda Salmonella u ptica može doći verti-kalnim putem (putem jaja) ili horizontalno (konzumira-njem kontaminirane vode, hrane, putem fecesa, prašine i dr.) (Cox i sur., 2000.; Foley i sur., 2011.; Howard i sur., 2012.). Bakterije iz roda Salmonella smatraju se izrazito otpornima na različite uvjete okoliša i mogu preživjeti tjednima i mjesecima u okolišu, a smatra se da mogu preživjeti i 26 mjeseci u hrani za životinje (D'Aoust i Sewell, 1986.; Davies i Wray, 1996.). Zbog navedenog hrana za perad jedan je od mogućih izvora infekcije salmonelama u peradi, a neizravno i infekcija u ljudi. Ipak, prema podacima iz literature izravnu povezanost prisut-nosti bakterija u hrani za životinje s obolijevanjem živo-tinja i obolijevanjem u ljudi nije uvijek moguće dokazati (Jones, 2011.; EFSA, 2012.). U svakom slučaju, u cilju sprječavanja nastanka infekcija bakterijama iz roda Salmonella u uzgoju peradi neophodno je kontrolirati sve potencijalne izvore infekcije, pa tako i hranu za životinje. Hrana za životinje uobičajeno se pretražuje referentnom
Marija Berendika, dipl.ing.preh.teh., Centar za peradarstvo, Hrvatski veterinarski institut, Henizelova 55, 10000 Zagreb, Hrvatska; Tel.: +385 (0)1 2441-394; Fax: +385 (0)1 2441-396; E-mail: [email protected]

USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS
113
horizontalnom metodom za dokazivanje Salmonella spp. u hrani i hrani za životinje (HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008) koja uključuje postupak pred-obogaćivanja u neselektivnom hranjivom bujonu, zatim obogaćivanje u selektivnom hranjivom mediju i na-cjepljivanje na selektivne hranjive podloge. Sojevi bakte-rija porasli nakon takvog izdvajanja zatim se identificiraju i potvrđuju biokemijskim testovima i serološkom tipiza-cijom. Sam postupak traje tri do pet dana za negativni rezultat, odnosno i do sedam dana za potvrdu pozitivnog rezultata. Zbog dugotrajnosti ove metode, a u cilju bržeg dobivanja rezultata razvijen je veći broj alternativnih metoda koje su se pokazale uspješne u probiru uzoraka hrane i uzoraka iz okoliša. Neke od tih metoda su i uspješno validirane od strane nezavisnih tijela (AOAC, AFNOR, MicroVal, ISO 16040). Tako postoje metode koje koriste nove postupke uzgoja bakterija, imunomagnetsku separaciju, imunoen-zimski testovi te različite molekularne metode. Takvi testovi redovito omogućuju dobivanje prvih rezultata za manje od dva dana. Test VIDAS® UP Salmonella (SPT) je brza metoda otkrivanja salmonela u hrani i u uzorcima iz okoliša koju obilježava jednostavna priprema i rezultati za manje od 19 sati. Metoda se temelji na tehnologiji proteina rekombi-nantnih bakteriofaga i imunoenzimskom fluorescentnom testu (Enzyme Linked Fluorescent Assay, ELFA) koji se analizira pomoću automatiziranih instrumenata VIDAS. Rezultat se tumači kao pozitivan ili negativan usporedbom dobivene granične vrijednosti fluorescencije (engl. thres-hold value, TV) s internim referentnim vrijednostima TV. Testom je moguće otkriti prisutnost i vrlo malog broja (ne)pokretnih bakterija roda Salmonella u ispitivanom uzorku (jedna bakterija). Metoda je validirana od strane nezavisnih tijela (AOAC i AFNOR) za otkrivanje sal-monela iz različitih uzoraka hrane i hrane za životinje (bioMérieux, 2012.; Eurofins, 2012.). Test RapidChek® SELECTTM Salmonella Enteritidis (SE) je brzi imunoenzimski test (tzv. „sendvič“ test) u kojem se koriste dvije vrste specifičnih antitijela (speci-fičnih za salmonele skupine D1 uključujući i serotip S. Enteritidis). Prva skupina specifičnih antitijela vezana je na površinu membrane kao „kontrolna linija“. Druga skupina specifičnih antitijela (označenih koloidnim zlatom) smještena je na površini membrane trakice reagensa kao „testna linija“ uzvodno od „kontrolne linije“. Uzorak kapilarno „putuje“ iz filtar područja trakice reagensa do područja sa „zlatom označenim specifičnim antitijelima“ pričem se prisutne bakterije serotipa S. Enteritidis specifično vežu na površinu membrane u tzv. „sendvič“ imunoenzimski kompleks i nastaje specifična reakcija (nastanak crvene testne linije). Višak zlatom označenih specifičnih antitijela veže se u području membrane kon-trolne linije i nastaje crvena kontrolna linija koja je ujedno i provjera ispravnosti testa. Kompleks antigen-antitijelo se ne stvara u odsutnosti salmonela skupine D1 te pritom
dolazi do nastanaka samo crvene kontrolne linije, ali ne i crvene testne linije. Metoda je validirana od strane ne-zavisnog tijela (AOAC) za otkrivanje salmonela skupine D1 za različite uzorke (obrisci nastamba, uzorci jaja i otopine za ispiranje trupova). Testom je moguće detektirati prisutnost bakterija serotipa S. Enteritidis za 32 do 48 sati, a test je prikladan za uzorke kontaminirane vrlom malim brojem bakterija (jedna bakterija) (Sdix, 2011.). Test RapidChek® SELECTTM Salmonella je brzi test koji koristi prednost visoko selektivnog postupka oboga-ćivanja i imunoenzimske detekcije salmonela. Imunoen-zimski test također se temelji na načelu „sendviča“. Koriste se dvije skupine specifičnih antitijela (specifičnih za bak-terije roda Salmonella spp.) vezanih na površinu trakice reagensa kao i u prethodno opisanom testu za salmonele skupine D1. Testovi su također jednostavni za primjenu, a rezultati se dobiju za 24 do 48 sati. Test je validiran od strane nezavisnog tijela (AFNOR) za različite uzorke hrane, hrane za životinje i uzorke iz okoliša. Pozitivne rezultate testa potrebno je potvrditi kao i kod ostalih alternativnih brzih testova nacijepljivanjem na minimalno dva selektivna agara te biokemijskim testovima i sero-loškom tipizacijom (Sdix, 2010.). Cilj ovoga rada bio je usporedba standardne referentne horizontalne metode za dokazivanje prisutnosti Salmonella spp. (HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008) u hrani za životinje s alternativnim brzim metodama određivanja prisutnosti bakterija iz roda Salmonella i to: VIDAS® UP Salmonella (SPT), RapidChek® SELECTTM Salmonella Enteritidis (SE) i RapidChek® SELECTTM Salmonella.
Materijal i metode
Uzorci hrane U radu je korišten uzorak hrane za perad. Uzorak je prije testiranja ispitan na prisutnost patogenih bakterija (uklju-čujući i bakterije iz roda Salmonella) te je utvrđeno da je isti mikrobiološki ispravan. Kako bi se eliminirala poten-cijalna bakterijska flora uzorak je steriliziran pri 121 °C u trajanju od 15 minuta. Uzorak je podijeljen u 60 manjih uzoraka (25,00 g) koji su korišteni u testovima. Uzorak hrane je umjetno kotaminiran bakterijom S. Enteritidis s dvije ciljane razine kontaminacije (1 CFU/25 g i 10 CFU/25 g). Za svaku ispitivanu metodu ispitano je pet uzoraka za svaku razinu kotaminacije i pet uzoraka negativne kontrole (isti uzorak hrane u koji je dodana samo otopina za pripremu decimalnog razrjeđenja bez bakterije). Bakterijska kultura U radu je korištena 24-satna kultura referentnog soja bakterije S. Enteritidis (ATCC 13076). Iz razrjeđenja od 0,5 po McFarlandu pripremljena su decimalna razrjeđenja do 10-7 te su za kontaminaciju uzorka hrane korištena razrjeđenja od 10-6 i 10-7. Istodobno je određen i točan broj

USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS 114
bakterija u pripremljenim razrjeđenjima nacijepljivanjem na hranjivi agar kako bi se provjerile ciljane razine kontaminacije. Referentna metoda HRN EN ISO 6579 Dokazivanje prisutnosti bakterija roda Salmonella prove-deno je prema uputama standardne referentne metode HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008. Ukratko, uzorak hrane za životinje homogeniziran je u omjeru 1:9 s puferiranom peptonskom vodom (engl. Buffered Peptone Water (BPW), bioMérieux, Francuska) i inkubiran pri temperaturi od 37 ºC±1 ºC u trajanju od 18±2 sata. U postupku obogaćivanja uzorka korištene su dvije tekuće selektivne hranjive podloge na način da je 0,1 mL pred-obogaćenog uzorka prenesen pipetom u epruvetu s 10 mL selektivnog Rappaport-Vassiliadis bujona (engl. Rap-paport Vassiliadis Soya broth (RVS), bioMérieux, Fran-cuska) i inkubiran pri 41,5 ºC±1 ºC kroz 24±3 sata. Istodobno je 1,0 mL pred-obogaćenog uzorka prenesen u epruvetu s 10 mL selektivnog Mueller-Kauffmannovog tetrationat/novobiocin bujona (engl. Mueller-Kauffmann tetrathionate/novobiocin broth (MKTTn), bioMérieux, Francuska) i inkubiran pri 37 ºC±1 ºC kroz 24±3 sata. Nakon toga su iz RVS bujona i MKTTn bujona uzorci nacijepljeni mikrobiološkom ušicom (10 µL) na gotove čvrste selektivne hranjive podloge, XLD agar (engl. xylose lysine deoxycholate agar (XLD), bioMérieux, Francuska) i SM2 agar (engl. chromIDTM Salmonella Agar (SM2), bioMérieux, Francuska) te su ploče inkubirane pri 37 ºC±1 ºC kroz 24±3 sata. Test VIDAS® UP Salmonella (SPT) Dokazivanje prisutnosti bakterija roda Salmonella testo-vima VIDAS® UP Salmonella (SPT) (bioMérieux, Fran-cuska) provedeno je prema uputi proizvođača. Ukratko, uzorak hrane za životinje homogeniziran je u omjeru 1:9 s puferiranom peptonskom vodom (bioMérieux, Francuska) i 1 mL Salmonella saplementa (bioMérieux, Francuska) te inkubiran pri 41,5 ºC±1 ºC kroz 21±3 sata. Iz tako pripremljenog uzorka mikrobiološkom ušicom su nacijepljene gotove čvrste selektivne hranjive podloge XLD agar (bioMérieux, Francuska) i SM2 agar (bio-Mérieux, Francuska) i inkubirane pri 37 ºC±1 ºC kroz 24±3 sata. Istodobno je 2-3 mL preneseno pipetom u epruvetu s poklopcem te zagrijano u vodenoj kupelji na 95-100 ºC kroz 5 minuta. Ohlađeni uzorak prenesen je pipetom u količini od 0,5 mL u jažice testa VIDAS® UP Salmonella (SPT). Testovi su analizirani pomoću instru-menta mini VIDAS® te je rezultat očitan nakon 48 minuta usporedbom vrijednosti TV (engl. test value threshold). Vrijednost TV manja od 0,25 u analiziranom uzorku označavala je odsutnost salmonela u uzorku (TV negative), dok je vrijednost TV veća ili jednaka 0,25 očitana kao pozitivan uzorak, odnosno u uzorku je bila dokazana prisutnost salmonela (TV positive). Rezultati su uspoređeni
s poraslim bakterijama na čvrstim selektivnim hranjivim podlogama. Test RapidChek® SELECTTM Salmonella Test RapidChek® SELECTTM Salmonella (SDIX, USA) sastoji se od primarnog medija RapidChek SELECT, sekundarnog medija RapidChek SELECT, saplementa RapidChek SELECT i trakica RapidChek SELECT Salmo-nella. Test je napravljen prema preporuci proizvođača (Sdix, 2010.). Ukratko, uzorak hrane za životinje homo-geniziran je u omjeru 1:9 s primarnim medijem sa sam-plementom (SDIX, USA) (predhodno zagrijanim na 41,5 ºC) u trajanju od 30 sekundi. Sadržaj je inkubiran pri 41,5 ºC kroz 19±3 sata. Nakon inkubacije 0,1 mL obo-gaćenog uzorka preneseno je pipetom u epruvetu s 1,0 mL sekundarnog medija (SDIX, USA) prethodno zagrijanog na 41,5 ºC. Epruvete su inkubirane pri 41,5 ºC kroz 7±1 sat. Istodobno je mikrobiološkom ušicom uzorak hrane u primarnom mediju nacijepljen na čvrste selektivne hranjive podloge XLD agar (bioMérieux, Francuska) i SM2 agar (bioMérieux, Francuska) koje su inkubirane pri 37 ºC±1 ºC kroz 24±3 sata. Nakon inkubacije uzorka u sekundarnom mediju u epru-vete su umetnute trakice za dokazivanje prisutosti salmo-nela, tako da je označeni kraj trakice bio na dnu epruvete (onaj na kojem je nacrtana strelica). Nakon 10 minuta očitane su promjene na trakicama. Pojava jedne crvene kontrolne linije na trakici ukazivala je na negativan rezultat (kontrolna crvena linija ujedno je bila i dokaz da je korištena trakica ispravna). Pojava dvije crvene linije označavala je pozitivan uzorak, odnosno uzorak u kojem je dokazana prisutnost bakterija iz roda Salmonella. Test RapidChek® SELECTTM Salmonella Enteritidis (SE) Test RapidChek® SELECTTM Salmonella Enteritidis (SE) (SDIX, USA) sastoji se od primarnog medija RapidChek SELECT, sekundarnog medija RapidChek SELECT, sa-plementa RapidChek SELECT i trakica RapidChek SELECT Salmonella. Test je napravljen prema preporuci proizvođača (Sdix, 2011.). Ukratko, uzorak hrane je homogeniziran u omjeru 1:9 s primarnim medijem (SDIX, USA) (prethodno zagrijanog sa saplementom na 42 ºC±2 ºC) u trajanju od 30 sekundi. Sadržaj je inkubiran pri 42 ºC±2 ºC kroz 19±3 sata. Nakon inkubacije 0,2 mL obogaćenog uzorka preneseno je pipetom u epruvetu s 2,0 mL sekundarnog medija (SDIX, USA) prethodno zagrija-nog na 42 ºC. Epruvete su inkubirane pri 42 ºC kroz 7±1 sat. Istodobno je mikrobiološkom ušicom uzorak hrane u primarnom mediju nacijepljen na čvrste selektivne hranjive podloge XLD agar (bioMérieux, Francuska) i SM2 agar (bioMérieux, Francuska) koje su inkubirane pri 37 ºC±1 ºC kroz 24±3 sata. Nakon inkubacije uzorka u sekundarnom mediju uzorci u epruvetama su ispitani trakicama za dokazivanje prisutnosti salmonela istim postupkom koji je prethodno opisan za test RapidChek® Salmonella.
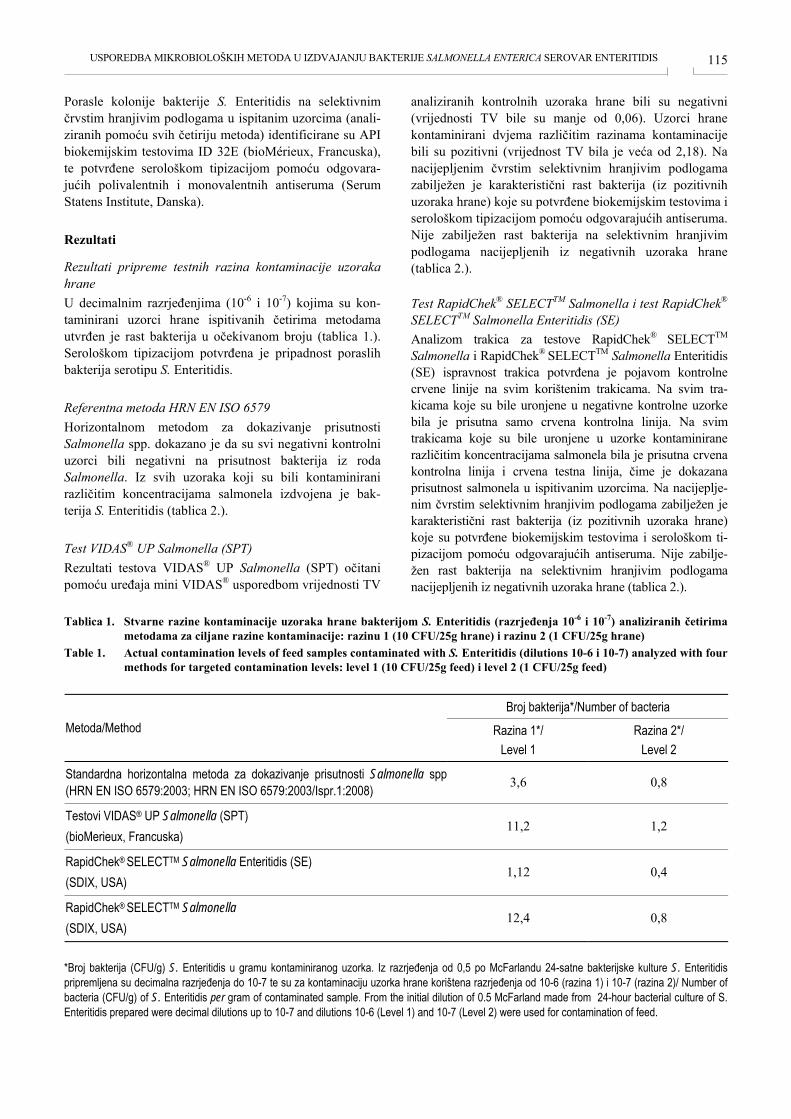
USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS
115
Porasle kolonije bakterije S. Enteritidis na selektivnim črvstim hranjivim podlogama u ispitanim uzorcima (anali-ziranih pomoću svih četiriju metoda) identificirane su API biokemijskim testovima ID 32E (bioMérieux, Francuska), te potvrđene serološkom tipizacijom pomoću odgovara-jućih polivalentnih i monovalentnih antiseruma (Serum Statens Institute, Danska).
Rezultati
Rezultati pripreme testnih razina kontaminacije uzoraka hrane U decimalnim razrjeđenjima (10-6 i 10-7) kojima su kon-taminirani uzorci hrane ispitivanih četirima metodama utvrđen je rast bakterija u očekivanom broju (tablica 1.). Serološkom tipizacijom potvrđena je pripadnost poraslih bakterija serotipu S. Enteritidis.
Referentna metoda HRN EN ISO 6579 Horizontalnom metodom za dokazivanje prisutnosti Salmonella spp. dokazano je da su svi negativni kontrolni uzorci bili negativni na prisutnost bakterija iz roda Salmonella. Iz svih uzoraka koji su bili kontaminirani različitim koncentracijama salmonela izdvojena je bak-terija S. Enteritidis (tablica 2.).
Test VIDAS® UP Salmonella (SPT) Rezultati testova VIDAS® UP Salmonella (SPT) očitani pomoću uređaja mini VIDAS® usporedbom vrijednosti TV
analiziranih kontrolnih uzoraka hrane bili su negativni (vrijednosti TV bile su manje od 0,06). Uzorci hrane kontaminirani dvjema različitim razinama kontaminacije bili su pozitivni (vrijednost TV bila je veća od 2,18). Na nacijepljenim čvrstim selektivnim hranjivim podlogama zabilježen je karakteristični rast bakterija (iz pozitivnih uzoraka hrane) koje su potvrđene biokemijskim testovima i serološkom tipizacijom pomoću odgovarajućih antiseruma. Nije zabilježen rast bakterija na selektivnim hranjivim podlogama nacijepljenih iz negativnih uzoraka hrane (tablica 2.).
Test RapidChek® SELECTTM Salmonella i test RapidChek® SELECTTM Salmonella Enteritidis (SE) Analizom trakica za testove RapidChek® SELECTTM Salmonella i RapidChek® SELECTTM Salmonella Enteritidis (SE) ispravnost trakica potvrđena je pojavom kontrolne crvene linije na svim korištenim trakicama. Na svim tra-kicama koje su bile uronjene u negativne kontrolne uzorke bila je prisutna samo crvena kontrolna linija. Na svim trakicama koje su bile uronjene u uzorke kontaminirane različitim koncentracijama salmonela bila je prisutna crvena kontrolna linija i crvena testna linija, čime je dokazana prisutnost salmonela u ispitivanim uzorcima. Na nacijeplje-nim čvrstim selektivnim hranjivim podlogama zabilježen je karakteristični rast bakterija (iz pozitivnih uzoraka hrane) koje su potvrđene biokemijskim testovima i serološkom ti-pizacijom pomoću odgovarajućih antiseruma. Nije zabilje-žen rast bakterija na selektivnim hranjivim podlogama nacijepljenih iz negativnih uzoraka hrane (tablica 2.).
Tablica 1. Stvarne razine kontaminacije uzoraka hrane bakterijom S. Enteritidis (razrjeđenja 10-6 i 10-7) analiziranih četirima
metodama za ciljane razine kontaminacije: razinu 1 (10 CFU/25g hrane) i razinu 2 (1 CFU/25g hrane) Table 1. Actual contamination levels of feed samples contaminated with S. Enteritidis (dilutions 10-6 i 10-7) analyzed with four
methods for targeted contamination levels: level 1 (10 CFU/25g feed) i level 2 (1 CFU/25g feed)
Broj bakterija*/Number of bacteria
Metoda/Method Razina 1*/ Level 1
Razina 2*/ Level 2
Standardna horizontalna metoda za dokazivanje prisutnosti Salmonella spp (HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008) 3,6 0,8
Testovi VIDAS® UP Salmonella (SPT) (bioMerieux, Francuska)
11,2 1,2
RapidChek® SELECTTM Salmonella Enteritidis (SE) (SDIX, USA)
1,12 0,4
RapidChek® SELECTTM Salmonella (SDIX, USA)
12,4 0,8
*Broj bakterija (CFU/g) S. Enteritidis u gramu kontaminiranog uzorka. Iz razrjeđenja od 0,5 po McFarlandu 24-satne bakterijske kulture S. Enteritidis pripremljena su decimalna razrjeđenja do 10-7 te su za kontaminaciju uzorka hrane korištena razrjeđenja od 10-6 (razina 1) i 10-7 (razina 2)/ Number of bacteria (CFU/g) of S. Enteritidis per gram of contaminated sample. From the initial dilution of 0.5 McFarland made from 24-hour bacterial culture of S. Enteritidis prepared were decimal dilutions up to 10-7 and dilutions 10-6 (Level 1) and 10-7 (Level 2) were used for contamination of feed.

USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS 116
Tablica 2. Rezultati usporedne analize uzoraka hrane na prisutnost bakterije S. Enteritidis četirima metodama Table 2. Results of comparison of four methods for isolation of S. Enteritidis bacteria from feed
Vrsta uzorka*/Sample category Metoda/Method Kontrola/
Control Razina 1*/
Level 1 Razina 2*/
Level 2
Standardna horizontalna metoda za dokazivanje prisutnosti Salmonella spp. (HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008) 0/5 5/5 5/5
Testovi VIDAS® UP Salmonella (SPT) (bioMerieux, Francuska) 0/5 5/5 5/5
RapidChek® SELECTTM Salmonella Enteritidis (SE) (SDIX, USA)
0/5 5/5 5/5
RapidChek® SELECTTM Salmonella (SDIX, USA)
0/5 5/5 5/5
*Kontrola = negativni kontrolni uzorak hrane koji nije bio kontaminiran bakterijskom kulturom S. Enteritidis; razina 1 i razina 2 označavaju razinu kontaminacije uzorka hrane bakterijama S. Enteritidis. Iz razrjeđenja od 0,5 po McFarlandu 24-satne bakterijske kulture S. Enteritidis pripremljena su decimalna razrjeđenja do 10-7 te su za kontaminaciju uzorka hrane korištena razrjeđenja od 10-6 (razina 1) i 10-7 (razina 2)/ Control = negative control feed sample that was not contaminated with S. Enteritidis; Level 1 and Level 2 correspond to the level of feed contamination with S.Enteritidis. From the initial dilution of 0.5 McFarland made from 24-hour bacterial culture of S. Enteritidis prepared were decimal dilutions up to 10-7 and dilutions 10-6 (Level 1) and 10-7 (Level 2) were used for contamination of feed. Rasprava
Iz prikazanih rezultata izdvajanja bakterije S. Enteritidis iz kontrolnih (negativnih) i kontaminiranih (pozitivnih) uzo-raka hrane za životinje četirima uspoređenim metodama može se zaključiti da su pomoću svih metoda jednakom točnošću negativni uzorci identificirani kao negativni, a da je u pozitivnim uzorcima (u obje razine kontaminacije) dokazana prisutnost bakterije S. Enteritidis. Vrijednosti specifičnosti, osjetljivosti i točnosti svih ispitivanih alter-nativnih metoda odgovaraju vrijednostima referentne me-tode. S obzirom na to da su sve testirane metode validirane od strane nezavisnih tijela očekivana je uspješnost izdva-janja salmonela koja je dokazana i u ovom radu. Slični rezultati dobiveni su i u istraživanjima drugih autora (Muldoon i sur., 2009.; Brenneman i sur., 2012.; Muldoon i sur., 2011.; Torlak i sur., 2012.). Testovi VIDAS® UP Salmonella (SPT) nedavno su razvi-jeni i validirani od strane nezavisnih tijela u skladu s međunarodnim standardom EN ISO 16140:20003 (AOAC i AFNOR) (Eurofins, 2012). Prema navedenom izvješću osjetljivost ove alternativne metode iznosi 97,4%, a gra-nica otkrivanja je od 0,3 do 1,3 stanica bakterija u uzorku od 25 grama (ili mililitra), što je odgovaralo rezultatima referentne metode za otkrivanje salmonela. Među-laborato-rijska usporedba metode (opisana u istom izvješću) prove-dena u 16 laboratorija uključivala je tri razine kontami-nacije (0, 1,16 i 10,9 CFU/25g hrane) i osam ponavljanja te usporedbu s referentnom metodom. Svih 16 laboratorija uspješno je identificiralo negativne uzorke kao i pozitivne uzorke u kojima je korištena viša razina kontaminacije. U uzorcima kontaminiram nižom razinom kontaminacije 55 uzoraka bilo je pozitivno metodom VIDAS® UP Salmo-
nella (SPT), a 62 referentnom metodom. Time je dokazana specifičnost metode od 100,0%, osjetljivost metode od 74,6% i relativna točnost od 89,0%. Prema dostupnim podacima u literaturi nije objavljen rad o usporedbi navedenog testa s drugim brzim testovima za otkrivanje bakterije S. Enteritidis u uzorcima hrane za životinje. Uspješnost tehnologije VIDAS u otkrivanju salmonela zabilježena je za druge testove (npr. VIDAS Salmonella (SLM)) koji se temelje na načelima testova ELFA (McMahon i sur., 2004.; Johnson i sur., 2009.; Crowley i sur., 2011.). Pritom su metode VIDAS bile jednako uspješne kao i referentna metoda za otkrivanje bakterija iz roda Salmonella u različitim uzorcima hrane. Ovim istraživanjem uspješno je dokazana primjena testa na uzorcima hrane za životinje umjetno kontaminiranim razli-čitim razinama kontaminacije bakterijom S. Enteritidis. Uz preduvjet da laboratorij posjeduje instrument VIDAS, na-vedena metoda mogla bi se koristiti za probir uzoraka hra-ne za životinje na prisutnost bakterija iz roda Salmonella uz obveznu potvrdu pozitivnih uzoraka nacjepljivanjem na selektivne hranjive podloge za rast salmonela, biokemij-skim testovima i serološkom tipizacijom. Prema uputi proizvođača, testovi RapidChek® SELECTTM Salmonella Enteritidis (SE) su predviđeni za otkrivanje bakterije iz roda Salmonella skupine D1 (S. Enteritidis) u uzorcima iz okoliša (obrisci iz peradarskih nastamba) i hrane (jaja i meso peradi). Testovi omogućuju otkrivanje prisutnosti S. Enteritidis unutar 32-48 sata, ovisno o vrsti uzorka. Svi sumnjivi uzorci na prisutnost bakterija iz roda Salmonella također se moraju dodatno potvrditi nacjeplji-vanjem uzorka na selektivne hranjive podloge. Testovi RapidChek® SELECTTM Salmonella, prema uputi proiz-vođača, predviđeni su za otkrivanje bakterija iz roda

USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS
117
Salmonella spp. u uzorcima hrane (mljeveno goveđe meso, sirovo mljeveno meso peradi, jajima, otopina za ispiranje trupova peradi, kuhano pureće meso) te u uzorcima iz okoliša (spužvice i brisevi). Testovi omogućuju otkrivanje prisutnosti bakterija Salmonella unutar 24-44 sata, ovisno o vrsti uzorka. U uputi je također navedeno ograničenje ovoga testa prema kojem se njime ne mogu otkriti bakterije roda Salmonella spp. koje pripadaju skupini O:18 (K prema staroj klasifikaciji). Uspješnost ovih alternativnih metoda s obzirom na spe-cifičnost, osjetljivost i relativnu točnost te usporedbu s referentnom metodom za otkrivanje bakterija iz roda Salmonella spp. ispitivali su i drugi autori (Muldoon i sur., 2009.; Brenneman i sur., 2012.; Muldoon i sur., 2011.; Torlak i sur., 2012.). Prema rezultatima Muldoona i sur. (2011.) metoda RapidChek SELECTTM Salmonella Enteri-tidis (SE) ispitana s 82 soja bakterije iz roda Salmonella skupine D1 (na uzorcima iz okoliša, uzorcima jaja i vode za ispiranje trupova peradi) pokazala je osjetljivost i speci-fičnost od 100% te točnost veću od referentne metode (137%), čime su autori dokazali izrazitu robusnost ovoga brzog testa. Istraživanje ovih autora napravljeno je s istim razinama kontaminacije kao i naše istraživanje te možemo zaključiti da naši rezultati potvrđuju robusnost navedenoga testa na uzorcima hrane za perad. Isti autori dobili su slične rezultate i za test RapidChek®
SELECTTM Salmonella u otkrivanju bakterije S. Typhi-murium u uzorcima maslaca od kikirikija (Muldoon i sur., 2009.). Slične rezultate opisali su i Brenneman i sur. (2012.) koji su ispitivali uspješnost testa RapidChek® SELECTTM Salmonella u izdvajanju vakcinalnih i divljih sojeva bakte-rije Salmonella enterica serovar Typhi u uzorcima stolice ljudi. Test je pokazao gotovo jednake rezultate kao i refe-rentna metoda za vakcinalne sojeve i nešto slabiju osjet-ljivost od referentne metode za divlje sojeve bakterije. Usporedbom rezultata naših istraživanja, upute proizvo-đača i literaturnih podataka možemo zaključiti da naši re-zultati potvrđuju robusnot testova RapidChek® SELECTTM Salmonella i RapidChek® SELECTTM Salmonella Enteri-tidis u uzorcima hrane za životinje kontaminiranim niskom razinom (1 CFU/25 g) i višom razinom (10 CFU/25 g) bakterije S. Enteritidis. S obzirom na postupak pripreme uzorka pretpostavljamo da u hrani nisu bile prisutne druge bakterije i/ili plijesni koje bi evenutalno mogle utjecati na rezultate istraživanja. Nadalje, u istraživanju je korišten referentni ATCC soj bakterije S. Enteritidis te bi bilo korisno slično istraživanje ponoviti na većem broju prirodno kontaminiranih uzoraka hrane za životinje.
Zaključci
Na tržištu se nudi velik broj različitih dijagnostičkih te-stova za brzo otkrivanje prisutnosti patogenih bakterija, osobito bakterija iz roda Salmonella. Cilj ovih testova je brzo otkrivanje izvora kontaminacije kako bi se spriječila
infekcija u ljudi i životinja, odnosno kako bi se osigurala zdravstveno ispravna hrana za ljude i životinje. Standardna referentna metoda za otkrivanje salmonela traje minimalno tri do pet dana za negativan rezultat, odnosno i do sedam dana za potvrdu pozitivnog rezultata. Primjena alternativnog testa VIDAS® UP Salmonella (SPT) omogućuje dobivanje rezultata unutar 19 sati upotrebom instrumenta VIDAS sa zadovoljavajućom specifičnošću, osjetljivošću i točnošću. Uz preduvjet da laboratorij posje-duje potrebni instrument, navedeni testovi mogli bi se ko-ristiti za probir potencijalnih izvora infekcije, jer značajno ubrzavaju proces dobivanja pouzdanih rezultata. Testovi RapidChek® SELECTTM Salmonella i RapidChek®
SELECTTM Salmonella Enteritidis također su se pokazali robusnim i korisnim za probir potencijalnih izvora infek-cije. S obzirom na jednostavnost primjene imaju prednost u odnosu na referentnu metodu i test VIDAS®, jer se mogu upotrijebiti i u slabije opremljenim laboratorijima, a osigu-ravaju osjetljive, precizne i točne rezultate. Uz navedeno, RapidChek® SELECTTM Salmonella Enteritidis je speci-fičan za otkrivanje salmonela skupine D1, dok test RapidChek® SELECTTM Salmonella otkriva i ostale sku-pine (osim bakterija skupine O:18). Kod svih alternativnih metoda neophodno je provjeriti po-zitivne rezultate izolacijom bakterija na selektivnim hranji-vim podlogama te identificirati bakterije biokemijskim testovima i serološkom tipizacijom.
Literatura
bioMÈRIEUX (2012): VIDAS® UP Salmonella (SPT), REF 30707, package insert version 9300912 D-EN-2012/04.
COX, N. A., M. E. BERRANG, J. A. CASON (2000): Salmonella penetration of egg shells and proliferation in broiler hatching eggs – a review. Poult.Sci. 79(11):1571-1574.
CROWLEY, E., P. BIRD, K. FISHER, K. GOETZ, M. J. BENZINGER JR, J. AGIN, D. GOINS, R. L. JOHNSON (2011): Evaluation of VIDAS Salmonella (SLM) easy Salmonella method for the detection of Salmonella in a variety of foods: collaborative study. J. AOAC Int. 94(6):1821-1834.
D'AOUST, J. Y., A. M. SEWELL (1986): Slow rehydration for detection of Salmonella spp. in feeds and feed ingredients. Appl. Environ. Microbiol. 51(6):1220-1223.
DAVIES, R. H., C. WRAY (1996): Persistence of Salmonella Enteritidis in poultry units and poultry food. Br. Poult. Sci. 37(3):589-596.
EUROFINS (2012): NF Validation certification of VIDAS® UP Salmonella method (VIDAS® SPT – ref. 30707). Summary Report. Link: http://www.afnor-validation.com/rapports-synthese/BIOMERIEUX/Synt-BIO-12-32-10-11.pdf [02.03.2013.]
European Food Safety Authority (EFSA) (2012): The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2010. EFSA Journal 10(3):2597.
FOLEY, S. L., R. NAYAK, I. B. HANNING, T. J. JOHNSON, J. HAN, S. C. RICKE (2011): Population dynamics of Salmonella enterica serotypes in commerical egg and poultry production. Appl. Environ. Microbiol. 77(13):4273-4279.

USPOREDBA MIKROBIOLOŠKIH METODA U IZDVAJANJU BAKTERIJE SALMONELLA ENTERICA SEROVAR ENTERITIDIS 118
GRIMONT, P. A. D., F. X. WEILL (2007): Antigenic formulae of the Salmonella serovars. WHO Collaboratoing Centre for Reference and Research on Salmonella – Institut Pasteur, 9. izdanje. Link: http://www.pasteur.fr/ip/portal/action/Web-driveAction Event/ oid/01s-000036-089 [02.03.2013.]
HOWARD, Z. R., C. A. O'BRYAN, P. G. CRANDALL, S. C. RICKE (2011): Salmonella Enteritidis in shell eggs: current issues and prospects for control. Food. Res. Int. 45(2):755-764.
HRN EN ISO 6579:2003 Mikrobiologija hrane i stočne hrane – Horizontalna metoda za otkrivanje Salmonella spp. (ISO 6579:2002; EN ISO 6579:2002); Microbiology of food and animal feeding stuffs – horizotal method for the detection of Salmonella spp. (ISO 6579:2002; EN ISO 6579:2002)
HRN EN ISO 6579:2003/Ispr.1:2008 Mikrobiologija hrane i hrane za životinje – Horizontalna metoda za dokazivanje prisutnosti Salmonella spp. (ISO 6579:2002/Cor 1:2004; EN ISO 6579:2002/AC:2006); Microbiology of food and animal feeding stuffs – horizontal method for the detection of Salmonella spp. (ISO 6579:2002/Cor 1:2004; EN ISO 6579:2002/AC:2006)
ISHA (2010): AFNOR Certification Validation Study RapidChekTM SELECTTM Salmonella test system for the detection of Salmonella spp. in food, feed and environmental samples. Link: http://www.afnor-validation.com/rapports-synthese/SDIX/SyntSDI 34-01 04-10 (en).pdf [02.03.2013.]
JOHNSON, R., J. MILLS, J. COLON-REVELES (2009): VIDAS Salmonella (SLM) assay method EasySLM with ChromID
Salmonella (SM2) Agar. Performance Tested Method 020901. J. AOAC Int. 92(6):1861-1864.
JONES, F. T. (2011): A review of practical Salmonella control measures in animal feed. J. Appl. Poult. Res. 20(1):102-113.
McMAHON, W. A., A. M. SCHULTZ, R. L. JOHNSON (2004): Evaluation of VIDAS Salmonella (SLM) immunoassay method with Rappaport-Vassiliadis (RV) medium for detection of Salmonella in foods: collaborative study. J. AOAC Int. 87(4):867-883.
MULDOON, M. T., J. LI, M. SUTZKO, A. C. OLSSON-ALLEN, G. TEANEY, V. GONZALEZ (2009): RapidChek SELECT Salmonella. Performance Tested Method 080601. J. AOAC Int. 92(6):1890-1894.
MULDOON, M. T., V. GONZALES, M. I. SUTZKO, A. C. OLSSON ALLEN, S. CREAMER, D. V. ONISK, K. LINDPAINTNER (2011): RapidChek SELECT Salmonella Enteritidis test system for the detection of Salmonella Enteritidis in poultry house drag swabs, shell egg pools, and chicken carcass rinsates. J. AOAC Int. 94(4):1138-1153.
SDIX (2010): RapidChek® SELECTTM Salmonella Sample Pack, Part Numbers: 7000190S and 7000194S, User Guide version 03/23/10.
SDIX (2011): RapidChek® SELECTTM Salmonella Enteritidis (SE) Sample Pack (5-Strips), Part Number: 7000220S, User Guide version 04/07/11.
TORLAK, E., I. M. AKAN, M. INAL (2012): Evaluation of RapidChek Select for the screening of Salmonella in meat and meat products. J. Microbiol. Meth. 90(3):217-219.
COMPARISON OF MICROBIOLOGICAL METHODS FOR DETECTION OF SALMONELLA ENTERICA SEROVAR ENTERITIDIS Summary Bacteria of the genus Salmonella are one of the most common causes of food-borne infection in humans. Foods of animal origin (meat, eggs, milk and milk products) are considered as the primary source of infection in humans. In primary poultry production, regular control of the potential source of infection is necessary and it also includes animal feed. The efficacy of the standard reference horizontal method HRN EN ISO 6579:2003; HRN EN ISO 6579:2003/Ispr.1:2008 and alternative fast methods VIDAS® UP Salmonella (SPT), RapidChek® SELECTTM Salmonella and RapidChek® SELECTTM Salmonella Enteritidis for isolation of Salmonella Enteritidis from animal feed samples was evaluated. Samples of artificially contaminated animal feed with different number of bacterial colonies (two contamination levels) were used for comparison of specificity, sensitivity and accuracy rates of the above mentioned methods. Based on the study results, it is concluded that all four methods were equally successful in isolation of Salmonella Enteritidis from animal feed samples. Alternative methods detected Salmonella bacteria in animal feed samples with appropriate specificity, sensitivity and accuracy and in a significantly shorter time than the reference method. Key words: Salmonella Enteritidis, animal feed, ISO, VIDAS, RapidChek

PERADARSKI DANI 2013.
119
PRIMJENA TESTA VIDAS ZA IZDVAJANJE BAKTERIJE RODA CAMPYLOBACTER SPP.
Borka Šimpraga1, Fani Krstulović1, Marijana Sokolović1, Marija Berendika1, Ivana Lohman-Janković2, Ljupka Maltar2
1Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska 2Uprava za veterinarstvo, Ministarstvo poljoprivrede, ribarstva i ruralnog razvoja Republike Hrvatske, Zagreb, Hrvatska Sažetak Tijekom veljače 2013. godine u Laboratoriju za bakteriologiju Centra za peradarstvo pretražena su 43 skupna uzorka sadržaja slijepih crijeva na prisutnost bakterija roda Campylobacter spp. Izdvajanje je obavljeno prema uputama standardne metode HRN EN ISO 10272:2008-1 i VIDAS® Campylobacter automatskog kvalitativnog imunoenzimskog fluorescentnog testa za otkrivanje kampilobaktera iz hrane za ljude i životinje, uzorcima iz okoliša proizvodnje i tekućine za ispiranje trupova peradi. Iz 27,9% skupnih uzoraka sadržaja slijepih crijeva i jednom i drugom korištenom metodom dokazane su bakterije roda Campylobacter spp. Na temelju dobivenih rezultata može se zaključiti da su se obje korištene metode pokazale jednako uspješnim za otkrivanje bakterije roda Campylobacter spp. u uzorcima slijepih crijeva. Prednost metode VIDAS® Campylobacter (CAM) je dobivanje negativnog nalaza u znatno kraćem razdoblju u odnosu na standardnu metodu. Ključne riječi: Campylobacter, perad, VIDAS, ISO
Uvod
Prema posljednjim izvješćima Europske agencije za sigur-nost hrane (EFSA) i Europskog centra za prevenciju i kontrolu bolesti (ECDC) objavljenim u ožujku 2012. godine kampilobakterioza je u 2010. godini bila najčešća zoonoza (EFSA, 2012.). Prema broju prijavljenih i potvrđenih sluča-jeva infekcija termotolerantnim bakterijama roda Campylo-bacter (C.) spp. u ljudi, od kojih su najučestaliji C. jejuni, C. coli i C. lari u 2010. godini prijavljeno je i potvrđeno 218.963 slučaja (48,6 slučajeva na 100.000 stanovnika), što je bilo povećanje za 6,7% u odnosu na 2009. godinu. Izvor infekcije u ljudi mogu biti različiti postupci u pripremi hrani u domaćinstvima, osobito priprema i konzumacija pilećeg mesa, ali i druge vrste hrane (mliječni proizvodi, juhe, sosovi, preljevi, začini, slatkiši te pekarski proizvodi), vode, kao i prijenos s osobe na osobu (EFSA, 2012.). Iste izvore infekcije uočili su i drugi autori (Adak i sur., 2005.; Wingstrand i sur.; 2006.; Humphrey i sur., 2007.; Heuvelink i sur., 2009.), što ukazuje na mogući rizik pri konzumaciji onečišćenog pilećeg mesa, ali i rizik konzumacije nepasteri-ziranog mlijeka. Infekcije bakterijama roda Campylobacter spp. u ljudi zabilježene su najčešće kao sporadični, pojedinačni slučajevi
ili rijetko obiteljska izbijanja. Kampilobakterioza je izrazito sezonskog karaktera i najveći broj obolijevanja javlja se ljeti. U svih vrsta ptica i divljih ptica termotolerantni kampilobakteri su učestali komenzali u probavnom sustavu i ptice se smatraju i njihovim glavnim rezervoarom (Achen i sur., 1998.; Corry i Alabay, 2001.; Waldenstrom i sur., 2002.). Prisutnost ovih bakterija dokazana je i u probavnim sustavima drugih životinja (Sandberg i sur., 2002.; Stanley i Jones, 2003.) te se kao važni rezervoari kampilobaktera također spominju životinje koje se uzgajaju na farmama uključujući goveda, svinje, ovce, koze, kao i glodavce, pse i mačke (Oporto i sur., 2007.). U peradi do infekcije najčešće dolazi u dobi od tri tjedna te u vrlo kratkom razdoblju cijelo jato bude inficirano, pričem nema kliničkih znakova bolesti, ali velika količina bakterije izlučuje se putem fecesa te dolazi do širenja bolesti koprofagijom, onečišćenom hranom i vodom (van Gerwe i sur., 2005.; Bull i sur., 2006.; Sparks, 2009.; Pérez-Boto i sur., 2010.). Najveća koncentracija kampilobaktera nalazi se u sadržaju slijepih crijeva (Wallace i sur., 1997.). Od ostalih čimbenika u nastanku i širenju infekcije u peradi navode se depopulacija jata, velik broj životinja, osoblje, prisutnost drugih životinja i također je dokazano da bakterije iz okoliša u nastambe za perad prenose muhe (Hald i sur., 2004.; Adkin i sur., 2006.).
Dr. sc. Borka Šimpraga, Hrvatski veterinarski institut, Centar za peradarstvo, Heinzelova 55, 10000 Zagreb, Hrvatska; Tel. +385 (0) 1 2441394; faks. +385 (0) 1 2441396; e-mail: [email protected]

PRIMJENA TESTA VIDAS ZA IZDVAJANJE BAKTERIJE RODA CAMPYLOBACTER SPP. 120
Kampilobakterioza je zoonoza i važan je javnozdravstveni problem u mnogim zemljama svijeta. U cilju poboljšanja kvalitete zdravlja i sprječavanja nastanka infekcija kampilo-bakterima u ljudi potrebno je primijeniti odgovarajuće bio-sigurnosne mjere u primarnoj proizvodnji te uz redovite kontrole smanjiti učestalost spomenutih bakterija. Strategija kontrole morala bi obuhvatiti preventivne mjere sadržane u provođenju niza različitih metoda. U nizu metoda vrlo važna je i primjena modificirane prehrane uz dodatke koji sma-njuju kolonizaciju kampilobaktera u jatu, kloriranje vode za piće, cijepljenje, primjena probiotika, prebiotika, primjena bakteriofaga te bakteriocina (Rosenquist i sur., 2009.; de Zoete i sur., 2007.). Primjena mjera za smanjenje kolonizacije probavnog susta-va peradi kampilobakterima tijekom primarne proizvodnja vrlo je važan preduvjet smanjenja onečišćenja pilećeg mesa i proizvoda od pilećeg mesa te smanjenja pojave obolijevanja ljudi. Zaključno, primjena probiotika i prebiotika u kombi-naciji sa strogim biosigurnosnim mjerama i kontrolom muha najvažnije su mjere stretegije smanjenja kampilobaktera u jatima peradi (Pasquali i sur., 2011.; Ghareeb i sur., 2012.). Kako nema jednostavnog "zlatnog standarda" u laboratorij-skoj dijagnostici i izdvajanju bakterija roda Campylobacter spp. (dehidracija, oksigenacija, smrzavanje i visoke tempe-rature smanjuju njihovu pokretljivost), nužno je uzorke što brže i na odgovarajući način dopremiti u laboratorij. U Laboratoriju za bakteriologiju Centra za peradarstvo počeo su uza standardnu metodu HRN EN ISO 10272:2008-1 za izdvajanje kampilobaktera koristiti automatski kvalitativni test VIDAS® koji se analizira pomoću instrumenata iz obitelji VIDAS®. Metoda je validirana od strane nezavisnih tijela (AOC i AFNOR) prema normi EN ISO 16040. Cilj ovoga rada bio je usporediti uspješnost izdvajanja i do-kazivanja bakterija roda Campylobacter spp. standardnom metodom i testom VIDAS® Campylobacter. Prikazani rezultati rada proizišli su iz Programa praćenja bakterija roda Campylobacter spp. i njihove otpornosti na antimikrobne pripravke u jatima i trupovima tovnih pilića u Republici Hrvatskoj u 2013. godini Ministarstva poljoprivrede Re-publike Hrvatske.
Materijal i metode
Izdvajanje termotolerantnih bakterija roda Campylobacter spp. standardnom metodom Izdvajanje termotolerantnih bakterija roda Campylobacter spp. obavljeno je prema uputama standardne metode HRN EN ISO 10272:2008-1, kao što je prethodno opisano (Šimpraga i sur., 2011.). Ukratko, sadržaj 10 slijepih crijeva istisnut je u sterilnu plastičnu vrećicu, homogeniziran i izvagan. Tako pripremljeni uzorak razrijeđen je u volumnom omjeru 1:9 Bolton bujonom za obogaćivanje (Bolton broth, Oxoid, Engleska). Nakon inkubacije u mirkoaerofilnim uvjetima uzorak je precijepljen na dvije selektivne podloge: modificirani cefoperazon deoksikolat agar s ugljenom (mCCD agar, Oxoid, Engleska) i Campy Food ID agar (CFA, bioMérieux, Francuska) te inkubiran također u mikroaerofilnim uvjetima.
Nakon inkubacije sa selektivnih agara tipične ili sumnjive kolonije precijepljene su na ploče kolumbija krvnog agara (kolumbija agar s dodatkom 5%-10% ovčje krvi) koje su inkubirane u mikroaerofilnim uvjetima. Čista kultura izrasla na kolumbija krvnom agaru (Columbia agar, bioMérieux, Francuska; ovčja krv, Biognost, USA) rabila se za fiziološku i biokemijsku diferencijaciju. Fizološka diferencijacija Za fiziološku diferencijaciju izrasla bakterijska kultura ispitana je na pokretljivost, morfologiju, mikroaerofilni rast pri 25 °C i aerobni rast pri 41,5 °C. Za ispitivanje morfologije napravljeni su mikroskopski pripravci, a za daljnja ispitivanja rabile su se kulture u kojima su mikroskopski ustanovljene zakrivljene ili spiralne štapićaste bakterije. Za ispitivanje pokretljivosti jedna kolonija s kolumbija krvnog agara suspendirana je u jedan mililitar Brucela bujona te je pregledana na pokretljivost pomoću mikroskopa. Bakterije roda Campylobacter spp. odlikuju se pokretljivošću poput vadičepa. Za ispitivanje mikroaerofilnog rasta pri 25 °C i za ispitivanje aerobnog rasta pri 41,5 °C odabrane kolonije nacijepljene su na kolumbija krvni agar, inkubirane u mikroaerofilnim uvje-tima pri 25 °C kroz 44±4 sata i u aerobnim uvjetima pri 41.5 °C kroz 44±4 sata te su nakon inkubacije ploče pregledane na vidljivi rast kolonija kampilobaktera. Biokemijska diferencijacija Za biokemijsku diferencijaciju napravljeni su testovi na oksidazu, katalazu, hidrolizu hipurata te API Campy (bioMérieux, Francuska), kao što je prethodno opisano (Šimpraga i sur., 2011.). API Campy je standardizirani sustav za identifikaciju kampilobaktera temeljen na mini-jaturnim testovima i specijalno prilagođen bazi podataka koji se izvodi prema uputi proizvođača. Reakcije se oči-tavaju pomoću vrijednosti navedenih u tablici očitavanja, a identifikacija na osnovi liste profila (bioMérieux, Fran-cuska). Otkrivanje termotolerantnih bakterija roda Campylobacter spp. VIDAS® testom Usporedno s izdvajanjem bakterija standardnom metodom ISO paralelni skupni uzorci sadržaja slijepih crijeva anali-zirani su testom VIDAS® testom prema uputi proizvođača (bioMérieux, Francuska). Ukratko, homogenat skupnog uzorka sadržaja slijepih crijeva nacijepljen je na CampyFood bujon u omjeru 1:9 i inkubiran u mikroaerofilnim uvjetima pri 41,5 °C kroz 48±4 sata. Nakon inkubacije uzorci su precijepljeni na selektivne čvrste hranjive podloge po-stupkom opisanim za standardnu metodu, a nekoliko mililitara bujona pipetom je prebačeno u epruvetu te držano pri 95-100 °C pet minuta. Tako zagrijani bujon je ohlađen, homogeniziran te korišten u testu VIDAS®. Prije ispitivanja uzoraka na uređaju miniVIDAS® ispitani su standardi kojima se uređaj umjerava i na temelju kojih se dobivaju interne referentne vrijednosti fluorescencije koje služe za

PRIMJENA TESTA VIDAS ZA IZDVAJANJE BAKTERIJE RODA CAMPYLOBACTER SPP.
121
usporedbu dobivenih vrijednosti fluorescencije uzoraka te procjenu pozitivne/negativne reakcije.
Rezultati
Metodom izdvajanja termotolerantnih bakterija roda Cam-pylobacter spp. standardnom metodom od ukupno 43 skupna uzorka slijepih crijeva bakterije karakterističnog rasta izdvojene su iz 12 uzoraka (27,9%) te je fiziološkom diferencijacijom dokazana pripadnost rodu Campylobacter. Biokemijskom diferencijacijom bakterija roda Campylobac-ter dokazano je da šest sojeva pripada vrsti C. jejuni, a šest vrsti C. coli. Metodom otkrivanja termotolerantnih bakterija roda Cam-pylobacter spp. testom VIDAS® u istovjetnim uzorcima dobiven je isti rezultat kao i u prethodnoj metodi. Dodatno nacjepljivanje svih uzoraka na selektivne čvrste hranjive podloge potvrdilo je rezultate testa VIDAS®. Od ukupno 43 skupna uzorka slijepih crijeva pretražena na prisutnost bakterija roda Campylobacter spp. prema upu-tama standardne metode (HRN EN ISO 10272:2008-1) i testa VIDAS kampilobakter, u 12 skupnih uzoraka slijepih crijeva s obje korištene metode dokazane su bakterije roda Campylobacter spp.
Rasprava i zaključci
Zbog velike učestalost bakterija roda Campylobacter i nji-hovog značenja potrebno je odrediti učinkovite kontrolne mjere u primarnoj peradarskoj proizvodnji kako bi se spriječile pojave obolijevanja u ljudi. Zbog navedenog potrebno je primjenjivati odgovarajuće osjetljive i pouzdane postupke otkrivanja i identifikacije spomenutih bakterija. Iako se za izdvajanje kampilobaktera redovito koristi stan-dardna metoda ISO, zbog karakterističnog rasta i osjet-ljivosti ovih bakterija postoji niz istraživanja kojima se još uvijek nastoje prilagoditi pojedini postupci za njihovo uspje-šnije i brže izdvajanje iz različitih uzoraka. Uobičajene mi-krobiološke metode otkrivanja bakterija roda Campylobac-ter spp. u peradi u pravilu uključuju postupke obogaćivanja selektivnim bujonima te nacjepljivanje na čvrste selektivne podloge, a ponekad i izravno nacjepljivanje na čvrste selektivne podloge. Prema standardnoj horizontalnoj metodi za dokazivanje pri-sutnosti Campylobacter spp. (HRN EN ISO 10272:2008-1) bujon izbora je Bolton bujon, a čvrsta podloga je mCCDA te druga čvrsta selektivna podloga po izboru za rast kampi-lobaktera. Uza spomenute podloge u različitim istraživa-njima korištene su i druge kombinacije selektivnih podloga kao što su Preston bujon, čvrsta selektivna podloga po Skirowu i druge, kao i različite modifikacije koje se razlikuju po količini i vrsti antibiotika, dodavanju krvi, ugljena i slično. U pravilu postoje dvije osnovne skupine podloga i to podloge s dodatkom krvi i podloge s dodatkom ugljena i one omogućuju selektivno izdvajanje bakterija kampilobaktera, dok se daljnja diferencijacija do vrste mora napraviti biokemijskim testovima budući da još uvijek nema odgovarajućih selektivnih podloga za tu namjenu. Oboga-
ćivanje se smatra korisnim u slučaju da se u uzorku nalazi mali broj kampilobaktera ili su oni oštećeni pod utjecajem različitih stresnih čimbenika okoliša (npr. promjene tempe-rature, dehidracija, oksigenacija i dr.) (Corry i sur., 1995.). Dodavanje antibiotika pomaže u inhibiciji rasta ostalih bakterija koje se nalaze u uzorku. Dodavanje krvi i ugljena pomaže u uklanjanju toksičnih derivata kisika kao što su peroksidi i superoksidni ioni. Ti produkti toksično djeluju na kampilobaktere, jer oni ne posjeduju enzime superoksid dizmutazu i peroksidazu (Corry i sur., 1995.). Osim spe-cifičnih sastava selektivnih bujona i čvrstih podloga, inku-bacija pri 42 °C također djeluje selektivno na rast ovih termotolerantnih bakterija (Corry i sur., 1995.; Paulsen i sur., 2005.). Uz mikrobiološke metode u posljednje vrijeme razvijene su i alternativne metode koje su se pokazale uspješne u doka-zivanju kampilobaktera iz nekih vrsta uzoraka. Alternativne metode uključuju različite enzimatske imunotestove (Endtz i sur., 2000.), molekularne metode (Rodgers i sur., 2012.) te enzimatske fluorescentne testove (Liu i sur., 2009.; bioMerieux, 2012.). Metoda VIDAS Campylobacter je brzi imunoenzimski test koji omogućuje otkrivanje kampilobaktera pomoću postupka ELFA (enzimski vezani fluorescentni test). Test radi na načelu otkrivanja specifičnih antigena koji je u interlabora-torijskom istraživanju provedenom sa 17 laboratorija na uzorcima kontaminiranim s tri razine kontaminacije (0 CF/25 g, 3 CFU/25 g i 30 CFU/25 g) pokazao relativnu točnost od 55,7%, specifičnost od 94,3% i osjetljivost od 92,6% u odnosu na referentnu metodu ISO za otkrivanje kampilobaktera. Time je dokazano da postoje stanovita odstupanja od referentne metode, s time da je ova alter-nativna metoda dala bolje rezultate osjetljivosti od refe-rentne metode koja je imala osjetljivost od samo 43,1%, dok je u prethodnim istraživanjima osjetljivost referentne metode bila veća. Kao razlog navedenom odstupanju navodi se i postojanje druge mikroflore u uzorku koja bi mogla utjecati na rast osjetljivih bakterija kao što su kampilobakteri, dok primjena načela specifičnih antigena i antitijela može biti nešto preciznija u otkrivanju bakterija u uzorcima u kojima je moguća i veća razina drugih bakterija. S obzirom na to da se kampilobakter u primarnoj proizvodnji okriva iz skupnih uzoraka sadržaja slijepih crijeva, ne može se zanemariti potencijalni inhibicijski učinak kompetitivne mikroflore, iako bi kampilobaktera u slijepim crijevima ipak trebalo biti najviše. Prema podacima iz dostupne literature nismo pronašli ra-dove koji su uspoređivali test VIDAS s referentnom meto-dom u skupnim uzorcima sadržaja slijepih crijeva peradi. U usporedbi metode na drugim vrstama uzoraka (npr. mesa) Liu i sur. (2009.) su dobili slične rezultate točnosti otkri-vanja kampilobaktera s mini VIDAS-om, molekularnim me-todama i standardnim mikrobiološkim metodama. Slične rezultate dobili su i drugi autori (Paulsen i sur., 2005.). U našem istraživanju od ukupno 43 skupna uzorka slijepih crijeva pretražena na prisutnost bakterija roda Campylo-bacter spp. rezultati referentne metode i testa VIDAS® Campylobacter bili su isti. U cilju daljnje provjere testa

PRIMJENA TESTA VIDAS ZA IZDVAJANJE BAKTERIJE RODA CAMPYLOBACTER SPP. 122
VIDAS i negativne uzorke smo provjerili nacjepljivanjem na selektivne čvrste podloge. Rezultati ovih provjera također su bili identični kao i prethodna dva testa. Test VIDAS ima prednost u brzini identifikacije negativnih uzoraka budući da test može biti gotov za 48 sati u odnosu na četiri dana koja su potrebna za otkrivanje negativnih uzoraka referentnom metodom. U slučaju pozitivnih uzoraka rezultat dobiven testom VIDAS treba provjeriti nacjepljiva-njem na selektivne čvrste podloge te potvrditi izolirane sojeve fiziološkom i biokemijskom karakterizacijom, čime se pozitivan nalaz dobiva nakon pet do sedam dana. Kod referentne metode potrebno je šest do osam dana do krajnje identifikacije kampilobaktera do vrste. Nedostaci testa VIDAS su u nemogućnosti diferencijacije vrsta, potrebni instrumenti i relativna cijena testa. Ipak, zbog brzine bi u analizi velikog broja uzoraka i u cilju što bržeg otkrivanja negativnih uzoraka svakako bilo korisno primijeniti ove testove VIDAS. Nalaz kampilobaktera u 12 skupnih uzoraka slijepih crijeva (od ukupno 43 pretražena) ukazuje na očekivanu učestalost bakterija u slijepim crijevima, što je u skladu s prijašnjim rezultatima (Šimpraga i sur., 2012.). Uočili smo tek manju razliku u učestalosti vrsta, jer su u ovom istraživanju bili jednako zastupljeni i C. jejuni i C. coli, dok je u prethodnim istraživanjima C. coli bila izdvajana u većem postotku. Naše istraživanje daje prikaz uspješnosti dviju metoda (referentne i alternativne metode VIDAS) korištenih za otkrivanje bakterija roda Campylobacter u sadržaju slijepih crijeva peradi. Na temelju rezultata može se zaključiti da je dokazana jednaka uspješnost obiju metoda. Međutim, potrebno je analizirati veći broj uzoraka kao bi se moglo doći do relevantnijih podataka.
Literatura
ACHEN, M., T. Y. MORISHITA, E. C. LEY (1998): Shedding and colonization of Campylobacter jejuni in broilers from day of hatch to slaughter age. Avian Dis. 42, 732-737.
ADAK, G. K., S. M. MEAKINS, H. YIP, B. A. LOPMAN, S. J. O'BRIAN (2005): Disease risks from foods, England and Wales, 1996-2000. Emerg. Infect. Dis. 11, 365-372.
ADKIN, A., E. HARTNETT, L. JORDAN, D. NEWELL, H. DAVISON (2006): Use of a systematic review to assist the development of Campylobacter control strategies in broilers. J. Appl. Microbiol. 100, 306-315.
bioMérieux, Francuska (2012): VIDAS Campylobacter (CAM), REF 30111. 07999 M.
BULL, S. A., V. M. ALLEN, G. DOMINGUE, F. JORGENSE, J. A. FROST, R. URE, R. WHYTE, D. TINKER, J. E. CORRY, J. GILLARD-KING, T. J. HUMPHREY (2006): Sources of Campylobacter colonising housed broiler flocks during rearing. Appl. Environ. Microbiol. 72, 645-652.
CORRY, J. E. L., H. I. ATABAY (2001): Poultry as a source of Campylobacter and related organisms. J. Appl. Microbiol. 90(S6), 96S-114S.
CORRY, J. E. L., D. E. POST, P. COLIN, M. J. LAISNEY (1995): Culture media for the isolation of Campylobacters. Int. J. Food Microbiol 26, 43-76.
De ZOETE, M. R. J. P. van Putten, J. A. Wagenaar (2007): Vaccination of chickens against Campylobacter. Vaccine 25(39), 5548-5557.
European Food Safety Authority (EFSA) (2012): The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2010. EFSA Journal 10(3), 2597.
ENDTZ, H. P., C. W. ANG, V. D. N. BRAAK, B. DUIM, A. RIGTER, L. J. PRICE, D. L WOODWARD, F. G. RODGERS, W. M. JOHNSON, J. A. WAGENAAR, B. C. JACOBS, H. A. VERBRUGH, V. A. BELKUM (2000): Molecular characterization of Campylobacter jejuni from patients with Guillan-Barre and Miller Fisher syndromes. J. Clin. Microbiol. 38(6), 2297-2301.
GHAREEB, K. K., W. W. W. A. AWAS, M. M. MOHNL, R. R. PORTA, M. M. BIARNES, J. J. BOHM, G. G. SCHATZMAYR (201S.): Evaluating the efficacy o fan avian-specific probiotic to reduce the colonization of Campylobacter jejuni in broiler chickens. Poult. Sci. 91(8), 1825-1832.
HALD, B., H. SKOVGARD, D. D. BANG, K. PEDERSEN, J. DYBDAHL, J. B. JESPERSEN, M. MADSEN (2004): Flies and Campylobacter infection of broiler flocks. Emerg. Infect. Dis. www.cdc.gov/eid, Vol (10), 1490-1492.
HAUVELINK, A. E., C. van HEERWAARDEN, A. ZWARTKRUIS, J. J. H. C. TIL BURG, M. H. BOS, F. G. C. HEILMAN, A. HOFHUIS, T. HOEKSTRA, E. de BOER (2009): Two outbreaks of campylobacteriosis associated with the consumption of raw cow's milk. Int. J. Food Microbiol. 134, 70-74.
HRN EN ISO 10272 : 2008 – 1. Mikrobiologija hrane i hrane za životinje – Horizontalna metoda za dokazivanje prisutnosti i brojenje Campylobacter spp. 1.dio: Metoda dokazivanja.
HUMPHREY, T., S. O'BRIEN, M. MADSEN (2007): Campylo-bacters as zoonotic pathogens: a food production perspective. Int. J. Food Microbiol. 117, 237-257.
LIU, L., S. K. HUSSAIN, R. S. MILLER, O. A. OYARZABAL (2009): Efficacy of Mini VIDAS for the detection of Campylobacter spp. from retail broiler meat enriched in Bolton broth, with or without the supplementation of blood. J. Food Protect. 72(11), 2428-2432.
OPORTO, B. J. I. ESTEBAN, G. ADURIZ, R. A. JUSTE, A. HURTADO (2007): Prevalence and strain diversity of thermophilic campylobacters in cattle, sheep and swine farms. J. Appl. Microbiol. 103(4), 977-984.
PASQUAKI, F., A. De CESARE, G. MANFREDA, A. FRANCHINI (2011): Campylobacter control strategies in European poultry production. World Poult. Sci. J. 67, 5-18.
PAULSEN, P. P. KANZLER, F. HILBERT, S. MAYRHOFER, S. BAUMGARTNER, F. J. M. SMULDERS (2005): Comparison of three methods for detecting Campylobacter spp. in chilled or frozen meat. Int. J. Food Microbiol. 103, 229-233.
PÉREZ-BOTO, D, GARCΊA-PEÑA J. FRANCISCO, JUAN C. ABAD-MORENO, M. DOLORES HURTADO-PIZARRO, IRATXE PÉREZ-COBO, M. AURORA ECHEITA (2010): Drinking water as the source of Campylobacter coli infection in grandparent heavy breeders. Avian Pathol. 39, 483-487.
RODGERS, J. D., J. R. LAWES, A. B. VIDAL. J. ELLIS-IVERSEN, A. RIDLEY, E. J. PLEYDELL, L. F. POWELL, M. TOSZEGHY, K. STAPLETON, F. A. CLIFTON-HADLEY (2012): Characteristics and comparative performance of direct culture, direct PCR and enumeration methods for detection and quantification of Campylobacter spp. in broiler caeca. 159, 390-396.

PRIMJENA TESTA VIDAS ZA IZDVAJANJE BAKTERIJE RODA CAMPYLOBACTER SPP.
123
ROSENQUIST, H., L. BOYSEN, C. GALLIANO, S. NORDEN-TOFT, S. ETHELBERG, B. BORCK (2009): Danish strategies to control Campylobacter in broilers and broiler meat: fact and effects. Epidemiol. Infect. 137, 1742-1750.
SANDBERG, M., B. BERGSJØ, M. HOFSHAGEN, E. SKJERVE, H. KRUSE (2002): Risk factors for Campylobacter infection in Norwegian cats and dogs. Prev. Vet. Med. 55, 241-253.
ŠIMPRAGA, B., I. LOHMAN JANKOVIĆ, F. KRSTULOVIĆ, M. SOKOLOVIĆ, A. LABROVIĆ (2011): Rezultati ispitiva-nja antimikrobne osjetljivosti bakterija Campylobacter jejuni i Campylobacter coli izdvojenih iz tovnih pilića. Zbornik Peradarski dani, Šibenik, 11.-14.05.2011., str. 108-113.
SPARKS, N. H. C. (2009): The role of water supply system in the infection and control of Campylobacter in chicken. World Poult. Sci. J. 65, 459-473.
STANLEY, K., K. JONES (2003): Cattle and sheep farms as reservoirs of Campylobacter. J. Appl. Microbiol. 94, 104S-113S.
van GERWE, T. J. W. M., A. BOUMA, W. F. JACOBS-REITSMA, J. van den BROEK, D. KLINKENBERG, J. A. STEGEMAN, J. A. P. HEESTERBEEK (2005): Quantifying transmission of Campylobacter spp. among broilers. Appl. Environ. Microbiol. 71, 5765-5770.
WALDENSTRÖM, J., T. BROMAN, I. CARLSSON, D. HASSELQUIST, R. P. ACHTERBERG, J. A. WAGENAAR, B. OLSEN (2002): Prevalence of Campylobacter jejuni, Campylobacter lari, and Campylobacter coli in different ecological guilds and taxa of migrating birds. Appl. Environ. Microbiol. 68, 5911-5917.
WALLACE, J. S., K. N. STANLEY, J. E. CURRIE, P. J. DIGGLE, K. JONES (1997): Seasonality of thermophilic Campylobacter populations in chickens. J. Appl. Microbiol. 82, 219-224.
WIGSTRAND A., J. NEIMAN, J. ENGBERG, E. MOLLER NIELSEN, P. GERNER-SMIDT, H. C. WEGWNER, K. MOLBAK (2006): Fresh chicken as main risk factor for campylobacteriosis, Denmark. Emerg. Infect. Dis.12(2), 280-284.
APPLICATION OF VIDAS TEST FOR ISOLATION OF CAMPYLOBACTER SPP.
Summary During February 2013, in the Poultry Center Laboratory of Bacteriology, a total of 43 group samples of appendices were tested for the presence of Campylobacter spp. bacteria. Isolation was performed according to the directions in the standard method HRN EN ISO 10272:2008-1 and with VIDAS® Campylobycter qualitative enzyme-linked fluorescent imuunoassay for detection of Campylobacter from food and feed, environmental samples and carcass rinsing. Campylobacter was detected in 27.9% of group samples of appendices with both methods. According to the presented data, we can conclude that both methods proved to be equaly successful for detection of these bacteria from group samples of appendices. The benefit of VIDAS® Campylobacter (CAM) methods is achievement of negative results in a shorter period than with the reference method. Key words: Campylobacter, poultry, ISO, VIDAS

PERADARSKI DANI 2013. 124
MONITORING THE NUMBER OF LACTOBACILLUS SPP. IN CHICKENS FED WITH AGARICUS BISPORUS
Maja Jelena Petek1, Branka Gršković1,2, Maja Popović3, Igor Špoljarić1, Borka Šimpraga4, Marijana Sokolović4, Mirta Balenović4, Lidija Kozačinski3, Daniel Špoljarić3, Damir Mihelić3, Ksenija Vlahović3, Gordan Mršić1,2
1 Ivan Vučetić Forensic Science Center, Ministry of Interior, Zagreb, Croatia 2 University Center for Forensic Sciences, University of Split, Split, Croatia 3 Faculty of Veterinary Medicine, University of Zagreb, Zagreb, Croatia 4 Poultry Center, Croatian Veterinary Institute, Zagreb, Croatia Summary Enteric infectious diseases are the most common cause of losses in intensive production of chickens. Antibiotic growth promoters (AGP) have been added to chicken feed for many years to prevent such diseases as well as to promote their growth. Nowadays, when nonclinical use of AGP has been gradually abandoned, there is an intensive search for alternative strategies directed towards control and prevention of losses in poultry. These strategies include numerous biological and synthetic compounds that could decrease the negative effects of stress and protect poultry from infection. It is already known that mushrooms have numerous biologically active compounds (such as β-glucans) with definite antibacterial, antiviral, antitumoral and immunostimulatory effects. In this study, we replaced AGP with Agaricus bisporus (white button mushroom) and monitored the number of Lactobacillus spp. bacteria in rectal swabs of chickens (aged from 1 day to 38 days) fed commercial feed supplemented with mushrooms. The results showed the number of Lactobacillus spp. to be by 14.6% higher in chickens fed mushrooms compared to control group. Key words: antibiotic growth promoter replacement, poultry feed, white button, rectal swab Introduction
Clinical and subclinical doses of antibiotics have been used for decades as treatment of infectious diseases as well as growth promoters in intensive poultry production. However, the frequent use of antibiotic growth promoters (AGP) gradually led to the loss of their efficiency. New bacterial strains resistant to antibiotics have been emerging, posing threat to human health. Therefore, EU countries have placed a ban on adding AGP to animal feed (Regulation EC no. 1831/2003). Consequently, there is intensive search for alternative strategies in the control and prevention of poultry losses. The strategies include numerous biological and synthetic preparations that can decrease the negative effects of stress and protect poultry
from infections. The most efficient in protection and at the same time harmless for consumers and environment are bioactive components of feed (nutraceutics) and substances that anticipate the function of the immune system (immunomodulators) (Gallois et al., 2009). Such substances are natural additives, like bees' products, plant substances, mushrooms and natural minerals, which can almost completely recoup all nutritive ingredients that the animals would consume in natural pasture as well as improve their immune system and prevent ailments. Mushrooms are the natural source of many biologically active macromolecules with curative properties, from which some, including glucans, have been used in Asian and Mediterranean countries in veterinary medicine for centuries, both as prevention and therapy (Breen, 1990; Chang and Buswell, 1996).
Assist. Prof. Gordan Mršić, PhD, Ministry of Interior, Ivan Vučetić Forensic Science Center, Ilica 335, 10000 Zagreb, Croatia; Tel: +385 1 3788 231; Fax: +385 1 3788 051; e-mail: [email protected]

MONITORING THE NUMBER OF LACTOBACILLUS SPP. IN CHICKENS FED WITH AGARICUS BISPORUS
125
The white button mushroom Agaricus bisporus is one of the most frequently bred species of mushrooms in the world. Good nutritive properties of white button, with low fat and high protein and carbohydrate content, make it a very acceptable food not only for humans, but also for domestic animals intended for human consumption. It was shown that β-glucans isolated from white button, besides antitumoral effect, also act as immunostimulants to systemic and local (gut) immunity of some species of farm animals (Barbison et al., 2002; Brown et al., 2003; Shen et al., 2007). It has been documented that dried white button preparations have beneficial effect on gut histomorphology and population of commensal microbiota in fattening chickens (Giannenas et al., 2010a; Giannenas et al., 2010b). Enteric infectious diseases are the most common cause of losses in intensive production of chickens. Diarrhea is frequently associated with enteropathogenic Escherichia coli. It was shown that Lactobacillus spp. effectively reduced the level of Escherichia coli in the small intestine in pigs (Barrow et al., 1980). Also, other authors have reported that Lactobacillus spp. have antagonistic effects on viability and growth of Escherichia coli (Fernandes et al., 1987). Consequently, in this study, we monitored the effect of Agaricus bisporus, which we added to the standard chicken feed as a replacement for AGP, on the growth of Lactobacillus spp. in rectal swabs of chickens fed the supplement of mushrooms in comparison to control group.
Material and Methods
Agaricus bisporus The Agaricus bisporus (Fig. 1) mushrooms were obtained from commercial cultivation (Šampiko, Budačka Rijeka, Croatia). Fresh mushrooms were dried in driers for six hours at 42 °C. Dried mushrooms were milled to powder.
Chicken feed In this study, three types of powdered commercial chicken feed (PIPO, Čakovec, Croatia) were used, as follows: 1) starter mixture for chickens aged 1 to 14 days, 2) finisher I mixture for chickens aged 14-28 days, and 3) finisher II mixture for chickens aged 28-38 days. Dried powdered mushrooms were added into all three types of standard fattening chicken feed in a concentration of 20 g/kg. Experimental animals In this study, 60 fattening chickens (ROSS 308, 30 male, 30 female) from commercial flock (PIPO, Čakovec, Croatia) were included (Fig. 3). Chickens were housed for 38 days in separate stainless floor pens under controlled temperature and light conditions (Department for Animal Nutrition, Faculty of Veterinary Medicine, Zagreb, Croatia). They were divided into two experimental groups (C and M), each containing 30 randomly selected chickens (15 female and 15 male). Each group was housed in separate pens using wood shavings as litter according to technological conditions of intensive breeding (Vučemilo, 2008). Control (C) group of chickens was fed commercial feed as follows: age 1 to 14 days with starter, age 14 to 28 days with finisher I, and age 28 to 38 days with finisher II. Chickens from group M were fed in the same manner with the commercial feed (starter, finisher I and finisher II) in which the preparation of powdered white button was added in a concentration of 20 g/kg. During the experiment, access to water and feed was ad libitum. Experimental animals were treated according to the Croatian legislation on animal protection. Rectal swabs Rectal mucosa swabs were taken from seven randomly chosen animals of each group on days 0, 14, 28 and 38 of the experiment. The number of Lactobacillus spp. We used the method on MRS agar in microaerophilic conditions for detection and enumeration of Lactobacillus spp. (Biolife Manual, 3rd edn., 101-102, 2003). The correlations between days of feeding (0, 14, 28, 38) and number of Lactobacillus spp. bacteria from rectal mucosa swabs in the control (C = not treated chickens) group and group fed with Agaricus bisporus (M=20 g/kg) were calculated using the Kendall’s τ rank correlation as a method of nonparametric statistics using Statistica 8.0 (StatSoft Inc., 2008).
Results and Discussion
Figure 2 shows Lactobacillus spp. from rectal mucosa swabs of experimental chickens.
Figure 1. Structure of powdered dried biomass of Agaricus
bisporus visualized by scanning electron micro-scope TESCAN MIRA3 LMH
Slika 1. Struktura praškaste biomase plemenite pečurkeAgaricus bisporus vizualizirane skenirajućimelektronskim mikroskopom TESCAN MIRA3LMH

MONITORING THE NUMBER OF LACTOBACILLUS SPP. IN CHICKENS FED WITH AGARICUS BISPORUS 126
On day 38 of the experiment, the value of Kendall τ rank correlation coefficient was significant for M group of chickens (p<0.05) as compared with control group (Table 1), i.e. the Agaricus bisporus supplement raised the number of Lactobacillus spp. by 14.6% in comparison to control chickens. Table 1. Number of Lactobacillus spp. from rectal mucosa
swabs taken from experimental chickens on days 0, 14, 28 and 38 of the experiment
Tablica 1. Broj bakterija vrste Lactobacillus iz rektalnih obrisaka uzetih od pokusnih pilića 0., 14. 28. i 38. dana pokusa
No. of Lactobacillus spp. bacteria (cfu/mL) on days of experiment
Kendall τ rank correlation coefficient
Group (n=7)
0 14 28 38 τ C 3.9x107 3.9x107 4.4x107 4.7x107 0.91 M 4.2x107 4.9x107 5.0x107 5.8x107 0.99*
*p<0.05
Considerable efforts have been focused lately on the understanding of enteric infections, their diagnosis, in-cluding biology of pathogens, host resistance and therapy in intensive large-scale production of food animals (chickens and pigs). On the other hand, little is known about the prevention of such diseases with immuno-modulatory and dietary strategies. Until recently, these problems have been overcome by adding the sub-therapeutic doses of AGP in feed in poultry/swine industry
to enhance the production efficiency (Yang et al., 2009). AGPs were used not only to improve growth, but also to control enteric infections. Today, when nonclinical use of antibiotics has been gradually abandoned, there is intensive search for alternative strategies directed towards control and prevention of losses in poultry. It was shown that a promising replacement for AGP could be the poly-saccharides such as β-glucans from different sorts of mushrooms, especially Shiitake and Maitake, which are assigned as having curative properties (Aida et al., 2009). The effects of β-glucans (isolated from baker's yeast Saccharomyces cerevisiae) on pig growth, digestion and immunity is a very important issue in veterinary medicine (Li et al., 2006; Price et al., 2009). Furthermore, the preparations of β-glucans from baker's yeast S. cerevisiae and Sclerotium rolfsii mushroom showed their ability to protect pigs from infection by enterotoxic Escherichia coli strains (Soltanian et al., 2009). Also, β-glucans from barley, oat and brown sea algae Laminaria spp. have shown beneficial effects on productivity and immunity of fattening animals (O'Shea et al., 2010). Colibacillosis in poultry is typically a localized or systemic disease that occurs as a consequence of imbalance between Escherichia coli and host resistance, caused by increased number of bacteria. It has been reported that Lactobacillus spp. of pig gastric origin inhibits growth of a broad spectrum of enteropathogens including Escherichia coli (Conway and Kjelleberg, 1989). Also, Oztürk et al. describe a strong antibacterial effect of the Agaricus mushroom preparations on gram-positive bacteria, especially on Micrococcus luteus, Micrococcus flavus and Bacillus subtilis (Oztürk et al., 2011). Furthermore, Giannenas et al. showed that Agaricus bisporus, added to a
Figure 2. Lactobacillus spp. from rectal mucosa swabs taken from experimental chickens visualized by scanning electron
microscope TESCAN MIRA3 LMH Slika 2. Bakterije vrste Lactobacillus iz rektalnih obrisaka uzetih od pokusnih pilića, vizualizirane skenirajućim elektronskim
mikroskopom TESCAN MIRA3 LMH

MONITORING THE NUMBER OF LACTOBACILLUS SPP. IN CHICKENS FED WITH AGARICUS BISPORUS
127
commercial feed in a concentration of 20 g/kg, raised the number of Lactobacillus spp. in chicken cecum (Giannenas et al., 2010b), which is in compliance with this study where Agaricus bisporus raised the number of Lactobacillus spp. by 14.6% in comparison to control group (Table 1).
Conclusion
Our results showed the ability of Agaricus bisporus to raise the number of Lactobacillus spp. in chicken intestine, thus contributing to current attempts at establishing the mushrooms of the genus Agaricus as good replacement for AGP, capable of reducing enteric diseases.
Acknowledgment
The current study was supported by grants from the Ministry of Agriculture (VIP/2012) and Ministry of Science, Education and Sports of Republic of Croatia (053-0532265-2255).
References
AIDA, F. M. N. A., M. SHUHAIMI, A. G. YAZID (2009): Mushroom as a potential source of prebiotics: a review. Trends Food. Sci. Tech. 20, 567-575.
BARBISON, L. F., M. MIYAMOTO, C. SCOLASTICI, D. M. F. SALVADORI, L. R. RIBEIRO, A. F. EIRA (2002): Influence of aqueous extract of Agaricus blazei on rat liver toxicity induced by different doses of diethylnitrosamine. J. Ethnopharmacol. 83, 25-32.
BARROW, P. A., B. E. BROOKER, R. FULLER, M. J. NEWPORT (1980): The attachment of bacteria to the gastric epithelium of the pig and its importance in the microecology of the intestine. J. Appl. Microbiol. 48, 147-154.
BREENE, W. M. (1990): Nutritional and medicinal value of specialty mushrooms. J. Food Prot. 53, 883-894.
BROWN, G. D., S. GORDON (2003): Fungal β-glucans and mammalian immunity. Immunity 19, 311-315.
CHANG, S. T., J. A. BUSWELL (1996): Mushroom nutraceuticals. World J. Microbiol. Biotechnol. 12, 473-476.
CONWAY, P. L., S. KJELLEBERG (1989): Protein mediated adhesion of Lactobacillus fermentum strain 737 to mouse stomach epithelium. J. Gen. Microbiol. 135, 1175-1186.
FERNANDES, C. F., K. M. SHAHANI, M. A. AMER (1987): Therapeutic role of dietary lactobacilli and lactobacillic fermented dairy products. FEMS Microbiol. Rev. 46, 343-356.
GALLOIS, M., H. J. ROTHKǑTTER, M. BAILEY, C. R. STOKES, I. P. OSWALD (2009): Natural alternatives to in-feed antibiotics in pig production: can immunomodulators play a role? Animal 3, 1644-1661.
GIANNENAS, I., D. TONTIS, E. ISHAKE (2010a): Influence of dietary mushroom Agaricus bisporus on intestinal morphology and microflora composition in broiler chickens. Res. Vet. Sci., 89, 21-28.
GIANNENAS, I., I. S. PAPPAS, S. MAVRIDIS, G. KONTOPIDIS, J. SKOUFOS, I. KYRIAZAKIS (2010b): Performance and antioxidant status of broiler chickens supplemented with dried mushroom (Agaricus bisporus) in their diet. Poult. Sci. 89, 303-311.
LI, J., D. F. LI, J. J. ZING, Z. B. CHENG, C. H. LAI (2006): Effects of β-glucan extracted from Saccharomyces cerevisiae on growth performance, and immunological and somatotropic responses of pigs challenged with Escherichia coli lipopolysaccharide. J. Anim. Sci. 84, 2374-2381.
O'SHEA, C. J., T. SWEENEY, M. B. LYNCH, D. A. GOHAN, J. J. CALLAN, J. V. O'DOHERTY (2010): Effect of β-glucans contained in barley- and oats-based diets and exogenous enzyme supplementation on gastrointestinal fermentation of finisher pigs and subsequent manure odor and ammonia emission. J. Anim. Sci. 88, 1411-1420.
OZTÜRK, M., M. E. DURU, S. KIVRAK, N. MERCAN-DOĞAN, A. TÜRKOGLU, M. A. OZLER (2011): In vitro antioxidant, anticholinesterase and antimicrobial activity studies on three Agaricus species with fatty acid compositions and iron contents: a comparative study on the three most edible mushrooms. Food Chem. Toxicol. 49, 1353-1360.
PRICE, K. L., H. R. TOTTY, H. B. LEE, M. D. UTT, G. E. FRITZNER, I. YOON, M. A. PONDER, J. ESCOBAR (2010): Use of Saccharomyes cerevisiae fermentation product on growth performance and microbiota of weaned pigs during Salmonella infection. J. Anim. Sci. 88, 3896-3908.
SHEN, J., H. REN, C. TOMIYAMA-MIYAJI, Y. SUGA, T. SUGA, Y. KUWANO, T. IIAI, K. HATAKEYAMA, T. ABO (2007): Potentiation of intestinal immunity by micellary mushroom extracts. Biomed. Res. 28, 71-77.
SOLTANIAN, S., E. STUYVENT, E. COX, P. SORGELOOS, P. BOSSIER (2009): Beta-glucans as immunostimulants in vertebrates and invertebrates. Crit. Rev. Microbiol. 35, 109-138.
VUČEMILO, M. (2008): Hygiene and bioecology in poultry. Faculty of Veterinary Medicine, University of Zagreb, Zagreb.
YANG, Y., P. A. IJI, M. CHOCT (2009): Dietary modulation of gut microflora in broiler chickens: a review of the role of six kinds of alternatives to in-feed antibiotics. World Poult. Sci. J. 65, 97-114.
UČINAK PLEMENITE PEČURKE AGARICUS BISPORUS NA BROJ BAKTERIJA VRSTE LACTOBACILLUS U REKTALNIM OBRISCIMA TOVNIH PILIĆA
Sažetak
Infekcijske crijevne bolesti su najčešći uzrok gubitaka u proizvodnosti pilića u intenzivnom uzgoju. Kako bi se spriječile takve infekcije, kao i radi poboljšanja rasta, u hranu za piliće godinama su se dodavali antibiotski promotori rasta (APR). U

MONITORING THE NUMBER OF LACTOBACILLUS SPP. IN CHICKENS FED WITH AGARICUS BISPORUS 128
današnje vrijeme kada se neklinička uporaba APR-a polako napušta provode se brojna istraživanja alternativnih strategija usmjerenih prema kontroli i prevenciji gubitaka u uzgoju peradi. Strategije uključuju niz prirodnih i sintetskih spojeva koji bi mogli smanjiti negativne utjecaje stresa i zaštititi perad od infekcije. Poznato je da gljive sadrže mnoge biološki aktivne spojeve (kao što su β-glukani) s utvrđenim antibakterijskim, antivirusnim, antitumorskim i imunostimulacijskim svojstvima. U ovom radu standardnu hranu za tovne piliće obogatili smo plemenitom pečurkom Agaricus bisporus kao zamjenom za APR te smo odredili broj bakterija vrste Lactobacillus u rektalnim obriscima plića (u dobi od 1 do 38 dana) hranjenih obogaćenom hranom. Na kraju pokusa broj bakterija vrste Lactobacillus u pilića hranjenih plemenitom pečurkom bio je za 14,6% veći u usporedbi s kontrolnim pilićima. Ključne riječi: prehrana, perad, gljiva, rektalni obrisak

PERADARSKI DANI 2013.
129
ANALIZA SOJINE SAČME NA SADRŽAJ SIROVIH BJELANČEVINA TE KONTAMINACIJU BAKTERIJAMA RODA SALMONELLA U 2012. GODINI
Manuela Zadravec1, Jelka Pleadin1, Vesna Jaki Tkalec2, Darko Majnarić2, Mario Mitak1
1 Hrvatski veterinarski institut, Zagreb, Hrvatska 2 Vaterinarski zavod Križevci, Hrvatski veterinarski institut, Križevci, Hrvatska Sažetak Soja kao najvažniji nositelj komponente bjelančevina u smjesama za perad dodaje se u smjese za perad u količini od 18% do 22%, ovisno o kategoriji životinja i sadržaju sirovih bjelančevina. Zbog tako visokog udjela dodatka soje vrlo je važna njena higijenska ispravnost u pogledu moguće kontaminacije bakterijama roda Salmonella. Salmonele u organizam ulaze alimentarnim putem uzrokujući najčešće subkliničke oblike salmoneloze, a oboljele životinje mogu širiti uzročnika izmetom (horizontalni put) i jajima (vertikalni put). U radu su prikazani rezultati analiza 36 uzoraka sojine sačme uzorkovane u 2012. godini na sadržaj sirovih bjelančevina i prisutnost bakterija roda Salmonella. Udio sirovih bjelančevina kretao se u rasponu od 41,26% do 48,40%, sa srednjom vrijednošću od 45,11±1,76%. Od 36 analiziranih uzoraka sojine sačme četiri (11%) uzorka bilo je kontaminirano bakterijama roda Salmonella. Iz dobivenih rezultata može se zaključiti da je tijekom 2012. godine sojina sačma u pogledu sadržaja sirovih bjelančevina zadovoljila kriterij kakvoće, dok je s obzirom na kontaminaciju bakterijama roda Salmonella predstavljala ozbiljan epizotiološki čimbenik. Ključne riječi: sojina sačma, sirove bjelančevine, Salmonella spp. Uvod Zbog visokog sadržaja sirovih bjelančevina i velikog udjela u krmnim smjesama sojina sačma ima veliku ulogu u hranidbi peradi. Međutim, zbog česte kontaminiranosti bakterijama roda Salmonella ima značajnu ulogu u epi-zootiologiji salmoneloza u peradi. Sojina sačma je proizvod visoke hranjive i biološke vrijednosti dobiven suvremenim procesom prerade kvalitetnog oljuštenog sojinog zrna. Sojina sačma predstavlja najznačajniji izvor bjelančevina (44%-46%) i zaštitnih hranjivih sastojaka, minerala i vitamina u hranidbi peradi svih uzrasta i kategorija (Dei, 2011.). Odgovarajućom termičkom obradom inaktiviraju se antinutritivni čimbenici, čime se postiže maksimalna isko-ristivost bjelančevina. Sojina sačma je bogata amino kiseli-nama (lizin, triptofan), koje su inače deficitarne u obrocima baziranim na proteinima žitarica, kao što su kukuruz ili ječam (Swick, 1999.; Britzman, 2001.). Udio sojine sačme u krmnim smjesama za perad kreće se od 18% do 30% ovisno o kategoriji životinja i sadržaju bjelančevina same sojine sačme. Zbog tako velikog udjela sojina sačma ima veliku ulogu i u higijenskoj ispravnosti krmnih smjesa za perad.
Soja se može kontaminirati bakterijama iz roda Salmonella tijekom berbe, prerade i skladištenja. Rizik je to veći što je sojina sačma izloženija pticama i glodavcima (Maciorowski i sur., 2006.). Kontaminirana soja dugo ostaje izvor zaraze zbog toga što salmonele tjednima mogu živjeti izvan orga-nizma nositelja preživljavajući smrzavanje, ali i temperature okoliša iznad 35 °C (Petkar i sur. 2011.). S obzirom na opa-snost koju predstavlja pojava salmoneloze (kontaminirano meso peradi i jaja) često se krmne smjese za hranidbu peradi peletiraju, što smanjuje sadržaj bakterija roda Salmonella u krmnim smjesama, ali ni podvrgavanje krmnih smjesa tem-peraturi peletiranja ne uništava salmonele u potpunosti (Jones, 2011.). Hranidbom peradi hranom kontaminiranom salmonelama perad se može zaraziti i oboljeti od salmone-loze (Li i sur., 2012.). Salmoneloze peradi su zoonoze od kojih obolijevaju sve vrste paradi te se pojavljuju u čitavom svijetu. Salmoneloza peradi može imati klinički oblik, pri-čem oboljele životinje pokazuju znakove bolesti, i sub-klinički oblik koji je mnogo značajniji zato što životinje izmetom izlučuju uzročnika bolesti bez vidljivih znakova bolesti. Uz izmetom (horizontalni način širenja) oboljele životinje mogu izlučivati salmonele i putem jaja (vertikalni
Manuela Zadravec dr. med. vet., Hrvatski veterinarski institut, Savska c. 143, 10000 Zagreb, Republika Hrvatska, tel: +385 1 6123 617, fax: +385 1 6123 670, e-mail: [email protected]

ANALIZA SOJINE SAČME NA SADRŽAJ SIROVIH BJELANČEVINA TE KONTAMINACIJU BAKTERIJAMA RODA SALMONELLA U 2012. GODINI
130
način širenja). Na taj način zaraženo meso i jaja peradi postaju izvori zaraze i za ljude (Kralj i Herceg, 1996.). Zbog uvida u epizootiološku situaciju salmoneloza peradi i kakvoću sojine sačme analizirano je 36 uzoraka sojine sač-me namijenjene umješavanju u hranu za perad s područja sjeverozapadnog, istočnog i središnjeg dijela Republike Hrvatske.
Materijali i metode
Trideset i šest uzoraka sojine sačme s različitih područja Republike Hrvatske (sjeverozapadna, istočna i središnja re-gija) namijenjenih za proizvodnju krmnih smjesa za perad pretraženo je na prisutnost bakterija roda Salmonella. U istim uzorcima sojine sačme određen je udio sirovih bje-lančevina.
Bakteriološka pretraga
Akreditiranom metodom HRN ISO 6579:2003 (Horizon-talna metoda za otkrivanje Salmonella spp.) analizirani su uzorci sojine sačme na prisutnost bakterija roda Salmonella. Inokulirano je 25 g homogenizirane sojine sačme u 225 mL puferirane peptonske vode (Merck, Njemačka) pri 37±1 ºC. Nakon 16-18 h 1,0 mL inokuliranog uzorka prenesen je u MKTTn bujon (Merck, Njemačka) i 0,1 mL u RVS bujon (Biokar Diagnostic, Francuska). MKTTn bujon inkubiran je 24 h pri 37±1 ºC, a RVS bujon pri 40,5-42 ºC. Nakon 24 h ezom zapremine 10 µL nacijepljen je sadržaj bujona na dvije ploče XLD agara (Bio Rad, SAD) i Rambach agara (Merck, Njemačka). Nakon inkubacije u trajanju od 24 h pri 37±1 ºC karakteristične kolonije roda Salmonella presađene su na TSI agar (Merck, Njemačka). U slučaju karakteristične re-akcije za salmonele, alkalična (crvena) kosina i kiselo (žuto) dno agara uza stvaranje plina (mjehurići) i vodikovog sul-fida (zacrnjenje agara), porasle kolonije su aglutinirane poli-valentnim serumom Omni-O (Bio Rad, SAD) i dodatno pro-vjerene bikemijskim nizom BBL CRYSTAL (BD, SAD).
Određivanje udjela bjelančevina
Za određivanje udjela sirovih bjelančevina uzorci sojine sač-me homogenizirani su pomoću analitičkog mlina (Cylotec 1093, Tecator). Udio (%, w/w) je određivan primjenom akreditirane metode (HRN EN ISO 5983-1:2008 i HRN EN ISO 5983-2:2010) po Kjeldahlu. Testni dio uzorka (0,7 g) razgrađen je pri temperaturi od 420 ºC u trajanju od 1 h, uz dodatak 12 mL koncentrirane sulfatne kiseline i katalizatora kalijevog sulfata i bakrenog (II)-sulfata pentahidrata uz upo-rabu bloka za razaranje (Unit 8 Basic, Foss). Nakon hlađenja razorenih uzoraka (~ 40 ºC) provedena je destilacija uz dodatak otopine natrijevog hidroksida (50 mL) i vode (80 mL) u kivetu te otopine borne kiseline (30 mL) u titrator. Titracijski postupak proveden je pomoću 0,1 N kloridne kiseline do prelaska boje otopine iz zelenkaste u ružičastu uz dodatak indikatora. Cjeloviti postupak destilacije i titracije proveden je korištenjem automatiziranog uređaja (Kjeltec
8400, Foss). Računskim postupkom određen je udio dušika, a potom i udio sirovih bjelančevina množenjem udjela dušika s faktorom konverzije 6,25.
Statistička obrada podataka
Statistička analiza provedena je korištenjem statističkog pa-keta Statistica for Windows (10,0 Statsoft Inc., 2011.). Za određivanje razlike u kvantitativnim vrijednostima udjela sirovih bjelančevina primijenjen je t-test, a statistički zna-čajne razlike izražavane su na razini vjerojatnosti 0,05.
Rezultati i rasprava
Od 36 analiziranih uzoraka sojine sačme bakteriološkom pretragom u četiri uzorka izdvojene su bakterije iz roda Sal-monella, što prestavlja značajnih 11% pozitivnih uzoraka.
Slika 1. Kontaminacija sojine sačme bakterijama roda
Salmonella u 2012. godini Figure 1. Contamination of soybean meal with Salmonella
in 2012. Tablica 1. Udio sirovih bjelančevina u uzorcima sojine
sačme po razredima kakvoće Table 1. The share of crude protein in samples of soybean
meal according to quality grade
Udio sirovih bjelančevina (%) Razred kakvoće
Min po Pravilniku*
(%)
Broj uzoraka Min Max Mean±SD
I. 46 11 46,02 48,40 46,99±0,95 II. 44 16 44,16 45,99 45,09±0,63 III. 40 9 41,16 43,88 42,83±1,01
*minimalni udio sirovih bjelančevina u sojinoj sačmi definiran Pravilnikom o izmjeni i dopuni pravilnika o kakvoći stočne hrane (N.N. 22/2006) *minimum share of crude protein in soybean meal defined by Ordinance amending of the Ordinance on the Quality of Animal Feed (NN 22/2006) U ispitivanim uzorcima utvrđena je srednja vrijednost udjela sirovih bjelančevina od 45,11±1,76%, s najmanjom vri-jednošću od 41,16% i najvećom od 48,40%. Pravilnikom o izmjeni i dopuni pravilnika o kakvoći stočne hrane (N.N. 22/2006) definiran je po razredima kakvoće najmanji udio sirovih bjelančevina koji soja treba sadržavati (prvi, drugi i treći razred kakvoće).

ANALIZA SOJINE SAČME NA SADRŽAJ SIROVIH BJELANČEVINA TE KONTAMINACIJU BAKTERIJAMA RODA SALMONELLA U 2012. GODINI
131
U tablici 1. prikazane su minimalne, maksimalne te srednje (±SD) vrijednosti udjela sirovih bjelančevina po razredima kakvoće sojine sačme. Najveći broj uzoraka bio je drugog razreda (n=16), zatim prvog (n=11) te trećeg razreda kakvoće (n=9). Ranijim istraživanjima (Jones i Richardson, 2004.; Hacking i sur., 1978.; MacKenzie i Bains, 1976.) utvrđeno je da su uljarice najčešće kontaminirani sastojci stočne hrane. S obzi-rom na to da se sojina sačma umiješava u vrlo visokom postotku (18%-30%) u hranu za perad, postoji vrlo velika vjerojatnost da se perad zarazi bakterijama roda Salmonella (Maciorowski i sur., 2006.). Kako udio sojine sačme u krmnim smjesama raste s obzirom na njen sadržaj sirovih bjelančevina, smanjenjem udjela sojine sačme, a to znači i boljom kakvoćom sojine sačme s obzirom na udio sirovih bjelančevina u krmnoj smjesi smanjuje se i rizik od pojave salmoneloze (Jones, 2011.). U našem istraživanju većina analizirane sojine sačme spada u drugi razred kakvoće te se stoga može zaključiti da je pojavnost salmoneloze uzroko-vane kontaminiranom sojinom sačmom razmjerno mala. Međutim, 11% analiziranih uzoraka kontaminirano je sal-monelama te se stoga rizik od pojave salmoneloze povećava. Iz dobivenih rezultata može se zaključiti da je tijekom 2012. godine sojina sačma u pogledu sadržaja sirovih bjelančevina zadovoljila kriterij kakvoće, dok je s obzirom na konta-minaciju bakterijama roda Salmonella predstavljala ozbiljan epizotiološki čimbenik.
Literatura
BRITZMAN, D.G. (2001): Soybean meal, an excellent protein source for poultry feeds. ASA Technical Bulletin. Link: http://www.asaim-europe.org/Backup/pdf/Britzman.pdf [04.03.2013.]
DEI, H.K (2011): Soybean as a feed ingredient for livestock and poultry. Recent trends for enhancing the diversity and quality of soybean products. Link: http://cdn.intechopen.com/pdfs/ /22604/InTech-Soybean_as_a_feed_ingredient_for_lives-tock_and_poultry.pdf [04.03.2013.]
HACKING, W.C., W.R. MITCHELL, H.C. CARLSON (1978): Salmonella investigation in an Ontario feed meal. Can. J. Comp. Med. 42, 400-406.
JONES, F. T., K. E. RICHARDSON (2004): Salmonella in commercially manufactured feeds. Poultry Sci. 83, 384-391.
JONES, F. T. (2011): A review of practical Salmonella control measures in animal feed. J. Appl. Poult. Res. 20, 102-113.
KRALJ M, M. HERCEG (1996): Bolesti peradi. Veterinarski priručnik, V. izdanje. Medicinska naklada, Zagreb.
LI, X., L. A. BETHUNE, Y. JIA, R. A. LOVELL, T. A. PROESCHOLDT, S. A. BENZ, T. C. SCHELL, G. KAPLAN, D. G. McCHESNEY (2012): Surveillance of Salmonella prevalence in animal feeds and characterization of the Salmonella isolates by serotyping and antimicrobial susceptibility. Foodborne Pathog. Dis. 9, 1-6.
MACIOROWSKI, K. G., P. HERRERA, M. M. KUNDINGER, S. C. RICKE (2006): Animal feed production and contamination by foodborne Salmonella. J. Verbr. Lebensm.1, 197-209.
Mac KENZIE, M. A., B. S. BAINS (1976): Dissemination of Salmonella serotypes from raw ingredients to chicken carcasses. Poultry Sci. 55, 957-960.
Narodne novine (22/2006): Pravilnik o izmjeni i dopuni pravilnika o kakvoći stočne hrane.
PETKAR, A., W. Q. ALALI, M. A. HARRISON, L. R. BEUCHAT (2011): Survival of Salmonella in organic and conventional broiler feed as affected by temperature and water activity. Agric. Food Anal. Bacteriol. 1, 175-185.
SWICK, R. A. (1999): Considerations in using protein meals for poultry and swine. ASA Technical Bulletin AN21. Link: http://www.asaimsea.com/index.php?language=en&screenname=__docs_Technical%20Bulletins|Animal%20Nutrition [04.03.2013.]
ANALYSIS OF SOYBEAN MEAL WITH REGARD TO CRUDE PROTEIN CONTENT AND CONTAMINATION WITH SALMONELLA IN 2012 Summary Soybean meal, because of its high crude protein content, is added to the feed mixture for poultry in the amount of 20%-35%, depending on the category of animals and the content of crude protein. Because of such a large share of soy additive, the hygienic status regarding any possible contamination with Salmonella is very important. Salmonellosis is a food borne infection causing mostly subclinical forms of the disease. An infected animal spreads pathogens via its feces (horizontal path) and eggs (vertical path). This paper presents the results of an analysis of 36 samples of soybean meal sampled in 2012 with regards to the crude protein content and the presence of salmonella. The proportion of crude protein ranged from 41.26% to 48.40%, mean 45.11±1.76%. Out of 36 samples of soybean meal analyzed, 4 (11%) samples were contaminated with Salmonella. These results suggest that in the year 2012, the soybean meal generally met the quality criteria in terms of crude protein content; however, concerning contamination by the bacteria of the genus Salmonella, it is a serious epizootic factor. Key words: soybean meal, crude protein, Salmonella spp.

PERADARSKI DANI 2013. 132
NALAZ PRERAĐENIH ANIMALNIH PROTEINA U HRANI ZA PERAD
Manuela Zadravec1, Mario Mitak1, Vesna Jaki Tkalec2, Darko Majnarić2
1 Hrvatski veterinarski institut, Zagreb, Hrvatska 2 Veterinarski zavod Križevci, Hrvatski veterinarski institut, Križevci, Hrvatska
Sažetak
Prerađeni animalni proteini (PAP) dozvoljeni su za hranidbu peradi ako su podrijetlom od ribe. Međutim, zbog neželjenih organoleptičkih svojstava (mirisa i okusa) u finalnim proizvodima peradarske proizvodnje, mesu i jajima, upotreba ribljeg brašna dozvoljena je samo za mlađe kategorije na početku tova. S obzirom na to da je upotreba PAP-a pod strogim nadzorom zbog epizootiološkog značenja za pojavnost spongiformnih encefalopatija, nedeklarirana upotreba ribljeg brašna predstavlja značajan epizootiološki propust. Tijekom 2011. i 2012. godine mikroskopski je pretraženo 30 smjesa za perad. Četiri krmne smjese za hranidbu nesilica sadržavale su nedeklarirano riblje brašno. Mogućnosti za dospijeće nedeklariranog ribljeg brašna u hranu za perad su nedovoljno očišćene miješalice u mješaonama stočne hrane ili namjerno dodavanje kako bi se uravnotežio sadržaj sirovih bjelančevina. Ključne riječi: prerađeni animalni proteini, hrana za perad, sirovi protein, spongiformne encefalopatije Uvod Prerađeni animalni proteini (PAP) podrijetlom od ribe (riblje brašno) koriste se u hrani za perad kao izvor proteina. Riblje brašno sadrži veće količine aminokiselina lizina, metionina i cisteina (Swick, 1999.) od sojine sačme koja se također koristi u hranidbi peradi kao izvor proteina. Uz dobar aminokiselinski sastav riblje brašno sadrži Ca i P (Hendriks i sur., 2002.) u povoljnom omjeru, a zbog saržaja masti od 2%-12% (Windstor, 2001.) dobro je i za pokrivanje energetskih potreba životinja. Međutim, hranidba peradi ribljim brašnom, pogotovo kozumnih nesilica i tovne peradi, ima za rezultat neugodan miris i okus jaja i mesa po ribi (Varga i sur., 2005.). Stoga je hranidba ribljim brašnom ograničena samo na mlađe kategorije životinja. U novije vrijeme, zbog pojave transmisivne spongiformne encefalopatije (Budka i sur., 2008.) zabranjena je upotreba PAP-a podrijetlom od drugih vrsta životinja osim ribe za hranidbu farmskih životinja (EC 999/2001.), pozanata kao "Feed ban". "Feed ban" je osnovna preventivna mjera koja se provodi u svrhu sprječavanja širenja transmisivnih ence-falopatija, a sastoji se od zabrane uporabe PAP-a u hrani za farmske životinje. Uporaba ribljeg brašna, isključivo pod strogim nadzorom, dozvoljena je u hranidbi svinja i peradi te
mladih preživača u fazi dojenja. Stoga često dolazi do nenamjerne kontaminacije u kafilerijama ili miješalicama u mješaonama stočne hrane koje nemaju odvojene linije za proizvodnju krmnih smjesa za preživače i nepreživače. Međutim, zbog nemogućnosti korištenja PAP-a na druge načine dolazi i do namjernog patvorenja ribljeg brašna PAP-om podrijetlom od drugih životinjskih vrsta (Murray, 2004.). S obzirom na to da je upotreba PAP-a pod strogim nad-zorom zbog epizootiološkog značenja za pojavnost spongi-formnih encefalopatija nedeklarirana upotreba ribljeg brašna predstavlja značajan epozootiološki propust. Kako bismo dobili uvid u korištenje ribljeg brašna u hrani za perad mikroskopskom pretragom analizirali smo smjese za perad s područja Republike Hrvatske. Mikroskopska pre-traga službena je pretraga u Europskoj Uniji, propisana Direktivom EC 51/2013. Ovom metodom može se otkriti sadržaj PAP-a u ribljem brašnu i krmnim smjesama u koncentraciji od 0,1% dodanih prerađenih animalnih proteina.
Materijali i metode
Mikroskopskom pretragom tijekom 2011. i 2012. godine pretraženo je 30 smjesa za perad, od toga 15 smjesa za
Manuela Zadravec dr.med.vet., Hrvatski veterinarski institut, Savska c. 143, 10000 Zagreb, Republika Hrvatska, tel: + 385 1 6123 617, fax: + 385 1 6123 670, e-mail: [email protected]

NALAZ PRERAĐENIH ANIMALNIH PROTEINA U HRANI ZA PERAD
133
kokoši neselice, pet dopunskih krmnih smjesa za tov pilića, sedam smjesa za tov pilića i tri smjese za tov purića podrijetlom iz mješaona stočne hrane s cijelog područja Republike Hrvatske. Glavnina smjesa (60%) podrijetlom je iz sjeverozapadnog dijela Hrvatske gdje se i nalazi većina proizvodnih kapaciteta stočne hrane u Republici Hrvatskoj.
Slika 1. Pretražene krmne smjese Figure 1. Feeds analyzed Peletirane smjese samljevene su u laboratorijskom mlinu. Uzorak je zatim prosijan kroz sito s promjerom otvora 0,5 mm. Ta prosijana frakcija promatrana je svjetlosnim mikro-skopom pod različitim povećanjima, a tražile su se dlake, perje i mišićni fragmenti. Kao pomoć u pretraživanju uzorak je obojen Fehlingovim reagensom (mišićno tkivo obojilo se ljubičasto) ili cistin reagensom (životinjski dijelovi koji sadrže cistin obojeni su tamno smeđe do crno). Frakcija koja je veća od 0,5 mm promatrana je stereo-mikroskopom. Svjetlosnim mikroskopom pregledana je barem jedna povr-šina pokrovnice 18 mm x 18 mm prosijane frakcije (engl. slide). Za dobivanje koncentriranog sedimenta najmanje 10 g samljevenog uzorka sedimentirano je u separacijskom lijevku s oko 100 mL tetrakloretilena. Nakon protresivanja i sedimentacije u trajanju od najmanje tri minute odvojen je sediment. Mješavina je potom još jednom promiješana i ostavljena sedimentirati najmanje tri minute. Tada se odvo-jio sediment i ostavio sušiti. Ako je sediment sadržavao velike dijelove prosijan je kroz sito veličine otvora 0,5 mm. Sediment je promatran pod stereo-mikroskopom i svjetlo-snim mikroskopom (40-400x). Za lakše promatranje pod svjetlosnim mikroskopom upotrebljen je glicerol kao uklop-no sredstvo. Za lakšu identifikaciju djelića kostiju sediment je obojen alizarinskim crvenilom. Kao pomoć u identifikaciji struktura animalnih tkiva i ostalih čestica koristili smo program ARIES, Atlas of Feed Microscopy III (Buki, 2005.) i referalne uzorke.
Rezultati i rasprava
Mikroskopskom pretragom smo uz očekivane biljne kompo-nente u četiri smjese za nesilice utvrdili mišićna vlakna, škrge, ljuske i dijelove kostiju podrijetlom od ribe. Od pre-gledanih 30 uzoraka hrane za životinje 12% nije udovolja-
valo deklaraciji, što ukazuje na značajan propust u kontroli tržišta hrane za perad, a to povlači za sobom i smanjenu kakvoću proizvedenog mesa i jaja.
Slika 2. Prerađeni animalni proteini (PAP) u hrani za
perad Figure 2. Processed animal proteins (PAP) in poultry feed Riblje brašno u hrani za nesilice nije ništa neobično s obzi-rom na to da je perad dozvoljeno hraniti ribljim brašnom. Riblje brašno kao nedeklarirana komponenta moglo je do-spjeti u hranu zbog nedovoljno očišćenih miješalica (nedo-voljno čišćenje) pričem je došlo do unakrsne kontaminacije. Druga mogućnost je da je riblje brašno dodano s namjerom poboljšanja aminokiselinske komponente i sadržaja Ca i P, ali zbog zakonske regulative (NN 26/1998.) nije dozvoljeno dodavati ga u hranu za nesilice pa tako ni prikazati na deklaraciji. Takav postupak čini hranu patvorenom (NN 46/2007.). I jedan i drugi način predstavljaju značajan epi-zootiološki propust s obzirom na pojavnost spongiformnih encefalopatija i zabranu hranjenja životinja PAP-om. Riblje brašno može biti patvoreno PAP-om podrijetlom od drugih vrsta životinja zbog poboljšanja svojstva ili zbog iskorišta-vanja zabranjenih PAP-a podrijetlom od kopnenih sisavaca i ptica (Campagnoli i sur., 2004.). Bellagamba i suradnici (2003.) pišu o kontaminaciji/patvorenju ribljeg brašna PAP-om različitih kopnenih životinja na talijanskom tržištu u relativno visokom postotku od 0,25% do 0,5% dodanog PAP-a. U Češkoj (Krčmar i Renčova, 2001.), Poljskoj (Weiner i sur., 2012.) i Velikoj Britaniji (Cawthraw i sur., 2009.) analizirane su krmne smjese i riblja brašna na pri-sutnost PAP-a podrijetlom od kopnenih sisavaca i ptica te se došlo do spoznaja da je 10%-20% ispitivanih smjesa sadrža-valo nedozvoljene PAP. Sustavnim praćenjem pojavnosti spongiformnih encefalopatija od strane DG SANCO (Direc-torate General For Health and Consumers – European Com-mission) došlo se do važne spoznaje da je pojavnost spongiformnih encefalopatija značajno pala od uvođenja potpune zabrane hranjenja farmskih životinja (Feed Ban) 2001. godine. Prema izvještaju DG SANCO za 2011. go-dinu, u Europi broj oboljelih životinja od transmisivnih encefalopatija 2001. godine iznosio je 2167, dok je 2010. taj broj bio 45 (DG SANCO, 2011.), što znači smanjenje pedeset puta! Zbog ove činjenice vrlo je važno da je riblje brašno na tržištu strogo kontrolirano s obzirom na kontami-

NALAZ PRERAĐENIH ANIMALNIH PROTEINA U HRANI ZA PERAD 134
naciju/patvorenje zbog poboljšavanja svojstva ili iskoristi-vost PAP-a od drugih vrsta životinja osim riba. Uz to, proizvodi životinja koje su nepropisno hranjene ribljim brašnom nisu tražene tržišne kakvoće u pogledu mirisa i okusa.
Literatura
ARIES (2004): Animal remains identification and evaluation system, Version 1.0. In STRATFEED Consortium, RIKILT. Wegeningen, The Netherlands.
BELLAGAMBA, F., F. VALFRE, S. PANSERI, V.M. MORETTI (2003): Polymerasae chain reaction-based analysis to detect terrestrial animal protein in fish meal. J. Food Protect. 66, 682-685.
BUDKA, H., B. GOOSSENS, G. RU (2008): BSE and TSEs: past, present and future. Trends Food Sci. Technol. 19, 34-39.
BUKI, R. E. (2005): Atlas of microscopy. Animal originated components. Garadi Zita. Budapest, Hungary.
CAMPAGNOLI, A., L. PINOTTI, G. TOGNON, F. CHELI, A. BALDI, V. Dell'ORTO (2004): Potential application of electric nose in processed animal proteins (PAP) detection in feedstuffs. Biotechnol. Agron. Soc. Environ. 8, 253-255.
CAWTHRAW, S., G. C. SAUNDERS, T. C. MARTIN, J. SAWYER, O. WINDL, S. D. REANELY (2009): Real-time PCR detection and identification of prohibited mammalian and avian material in animal feeds. J. Food Protct. 72, 1055-1062.
European Commission, Directorate General for Health and Consumers (2011): Report on the monitoring and testing of ruminants and the presence of TSE in the EU in 2010. Link: http://ec.europa.eu/food/food/biosafety/tse_bse/docs/annual_report_tse2010_en.pdf
European Union (2001): Regulation (EC) 999/2001 laying down rules for the prevention, control and eradication of certain transmissible spongiform encefalopathy.
European Union (2013): Amending Regulation (EC) 51/2013 as regards the methods of determination of constituents of animal origin for the official control of feed.
HENDRIKS, W. H., C. A. BUTTS, V. D. THOMAS, K. A. C. JAMES, P. C. A. MOREL, M. W. A. VERSTEGEN (2002): Nutritional quality and variation of meal and bone meal. Asian Aust. J. Anim. Sci. 15, 1507-1516.
KRČMAR, P., E. RENČOVA (2001): Identification of bovine-specific DNA in feedstuffs. J. Food Protect. 64, 117-119.
MURRAY, I., K. J. KAFFKA, Z. SEREGELY (2004): PQS detects adulteration of fishmeal with meat and bone meal – a contribution to help eradicate "mad cow disease". Near infrared spectroscopy. Proceedings of the 11th International Conference. Cordoba, Spain.
Narodne novine (26/1998): Pravilnik o kakvoći stočne hrane. Narodne novine.
Narodne novine (46/2007): Zakon o hrani. Narodne novine. SWICK, A. R. (1999): Consideration in using protein meals for
poultry and swine. ASA Techn. Bull. 21, 1-11. VARGA, C., V. STRELEC, M. VOLK (2005): Proizvodni rezultati
i kakvoća okusa mesa pilića koji u prehrani nisu dobivali životinjske bjelančevine. Meso 7, 30-33.
WINDSTOR, M. L. (2001): Fish meal. Torry Research Station. Link: http://www.fao.org/wairdocs/tan/x5926e/ /x5926e00.htm#Accompanying%20Notes
WEINER, A., A GOLEBIOVSKA, I. PAPROCKA, K. KWIATEK (2012): Detection and identification of banned processed animal protein in feeding stuffs by microscopic and PCR methods. PJV Sci 15, 163-164.
DETECTION OF PROCESSED ANIMAL PROTEIN IN POULTRY FEED
Summary
Processed animal proteins (PAP) are allowed to be fed to poultry if originating from fish. However, due to adverse organoleptic properties (odor and flavor) in meat and eggs as final products of poultry production, the use of fishmeal is allowed only for younger categories at the beginning of fattening period. As the use of PAP is under strict supervision because of its epidemiological role in the etiology of spongiform encephalopathy, any undeclared use of fishmeal is a major epizootiologic omission. In 2011 and 2012, 30 samples of poultry feed were tested microscopically for the presence of PAP. Four samples of feed for laying hen contained undeclared fishmeal. Undeclared fishmeal can reach poultry feed through inappropriately cleaned mixers in feed factory or intentionally being added to balance the content of crude protein. Key words: processed animal protein, poultry feed, crude protein, spongiform encephalopathy

PERADARSKI DANI 2013.
135
ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA
Anamaria Ekert Kabalin1, Sven Menčik1, Mario Ostović2, Hrvoje Kabalin3, Mirta Balenović4, Tomislav Balenović1, Igor Štoković1, Velimir Sušić1, Kristina Starčević1, Maja Maurić1
1 Zavod za stočarstvo, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 2 Zavod za higijenu, ponašanje i dobrobit životinja, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 3 Veterinarska stanica Jastrebarsko, Ambulanta Pisarovina, Pisarovina, Hrvatska 4 Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska
Sažetak
Ekstenzivan tov purana dalmatinskog zaleđa („dalmatinskih tuka“) na obiteljskim gospodarstvima usmjeren je na sezonsku potrošnju purećeg mesa, uglavnom zimi. Način uzgoja, uza specifičnost genotipa, dijelom su razlog karakterističnoj aromi i cijenjenosti njihova mesa. Istraživanjem je obuhvaćeno 20 tuka (po 10 svakog spola) u dobi od 7±0,5 mjeseci, uzgajanih u ujednačenim uvjetima. Da bismo pobliže okarakterizirali kakvoću mesa najznačajnijih dijelova trupa provedena je disekcija prsa, zabataka i bataka te utvrđena zastupljenost pojedinih tkiva (kože i potkožnog masnog tkiva, mišićnog tkiva, tetiva i ovojnica, masnog tkiva te kosti) kod utovljenih jedinki. Prosječni randman u purana iznosio je 70,41±4,91%, a u pura 70,83±3,24%. Iako su apsolutne vrijednosti tjelesne mase živih jedinki, kao i mase prsa, zabataka i bataka bile značajno veće u muških jedinki (p<0,001 za sve), njihov udio u tjelesnoj masi živih jedinki bio je ujednačen po spolu. Nadalje, nisu zabilježene značajne razlike između udjela pojedinih tkiva u prsima purana i pura. U zabatacima purana utvrđen je značajno veći udio kože i potkožnog masnog tkiva (p<0,01), kosti i tetiva (p<0,05) te manji udio mišićnog tkiva (p<0,05). Također, u batacima muških jedinki udio tetiva i ovojnica bio je značajno veći (p<0,001), a udio mišićnog tkiva manji (p<0,01). Ključne riječi: pureće meso, ekstenzivan uzgoj, udio tkiva Uvod Na prinos i kakvoću mesa purana utječu mnogi čimbenici među kojima su način uzgoja, sastav hrane, završna tjelesna masa (tržišna težina), dob, spol, kao i genotip jedinki (Halvorson i sur., 1991.; Brake i sur., 1994.; Waldroup i sur., 1997.; Roberson i sur., 2003.; Bogosavljević-Bošković i sur., 2005.; Nestor i sur., 2005.; Hulet, 2006.; Laudadio i sur., 2009.; Medić i sur., 2009.; Sogut i sur., 2010.). Werner i sur. (2008.) navode da uslijed povećane potrošnje purećeg mesa dolazi do novog izazova u proizvodnji s obzirom na to da ubrzana „industrijska“ proizvodnja, kao i sve intenzivniji rast pojedinih sojeva purana utječu na kakvoću mesa. Pritom se kakvoća purećeg mesa može procjenjivati s različitih stajališta. Sa stajališta proizvođača procjenjuje se na osnovi randmana, većeg udjela najvažnijih dijelova trupa I. i II. kategorije, poželjne konformacije, te optimalnog rasporeda
masnog tkiva i neoštećene kože. Potrošač danas, u cilju očuvanja vlastitog zdravlja, pridaje veću pozornost speci-fičnosti i ukusnosti mesa, poznatom podrijetlu te načinu uzgoja. Takav stav krajnjeg kupca čini jednim od glavnih čimbenika razvoja ekološko-organskog uzgoja. Ekstenzivan način uzgoja smanjuje troškove prehrane ispašom purana, a time i ukupne proizvodnje purećeg mesa (Alders i Pym, 2009.; Sogut i sur., 2010.). Takav način uzgoja pogoduje i osiguranju dobrobiti purana (Platz i sur., 2003.). Podjela mesa peradi na svijetlo i tamno meso zasniva se na njegovoj boji koja je posljedica relativnog odnosa bijelih i crvenih vlakana u mišićima (Medić i sur., 2009.). Pritom pod svijetlim mesom podrazumijevamo prsne mišiće broj-lera i purana, a pod tamnim mišiće nogu. Pureće meso ima visoku nutritivnu vrijednost, prije svega visok sadržaj bjelančevina, a nizak sadržaj masti (osobito prsni mišići), te se ubraja u dijetetske proizvode, a na tržištu uglavnom
Prof. dr. sc. Anamaria Ekert Kabalin, Veterinarski fakultet Sveučilišta u Zagrebu, Zavod za stočarstvo, Heinzelova 55, 10000 Zagreb; tel/fax: +385 1 2390138; e-mail: [email protected]

ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA 136
postiže optimalan omjer cijene i kakvoće (Roberson i sur., 2003.). Bitan čimbenik koji pritom određuje kakvoću mesa je zastupljenost pojedinih tkiva u važnijim osnovnim dijelovima trupa (Bogosavljević-Bošković i sur., 2005.). Tradicionalni uzgoj arhaične forme purana na širem po-dručju dalmatinskog zaleđa tek je krajem prvog desetljeća ovoga stoljeća privukao pozornost znanstvenika i stručnjaka, kada je započeta provedba projekta „Dalmatinska tuka – fenotipske odlike i ekološke odrednice areala“. U okviru istraživanja nastojalo se što detaljnije okarakterizirati jedin-ke, analizirati njihova morfološka svojstva te utvrditi geo-grafsku rasprostranjenost (Ekert Kabalin i sur., 2009.a,b,c, 2011., 2012.; Menčik i sur., 2009.). Na brojnim seoskim domaćinstvima dalmatinskog zaleđa puran ima važnu ulogu u prehrani lokalnog stanovništva i njegova proizvodnja usmjerena je na sezonsku potrošnju, pričem se većina utov-ljenih jedinki prodaje tijekom zime, uglavnom kao blagdan-ski specijalitet. Meso dalmatinskih tuka, zbog specifičnosti uzgoja, poznati je specijalitet tijekom turističke sezone. Pri ekstenzivnom uzgoju tuke se drže u skromnim životnim uvjetima, uglavnom na otvorenom, izuzev prva dva mjeseca života (topla faza uzgoja), noću te tijekom zime. Životinje na zelenim površinama pronalaze hranu (trava, djetelina, skakavci, masline, aromatično bilje), a po potrebi im se obrok nadopunjuje kukuruzom, mekinjama, manjom količi-nom prekrupe, salatom, lupinama luka, kupusom i slično (tj. svim biljnim ostacima iz kuhinje). Ekološko-organska pe-radarska proizvodnja zadnjih godina zauzima sve značajnije mjesto u uzgoju i proizvodnji domaćih životinja u zemljama Europske Unije i drugim visokorazvijenim zemljama svijeta (Mužic i sur., 2008.). Nadalje, zabilježen je trend povećanja interesa za očuvanje ukupne bioraznolikosti samoodrživim, nekonvencionalnim, ekološko-organskim uzgojem domaćih životinja (Rizzi i sur., 2009.). Cilj istraživanja prikazanog u radu bio je procijeniti kakvoću mesa kod autohtone forme purana tradicionalno uzgajanog u ekstenzivnim uvjetima na širem području dalmatinskog zaleđa. Pritom su, kao pokazatelj kakvoće, promatrani udjeli osnovnih tkiva u vrijednijim dijelovima trupa (prsima, zabatacima i batacima).
Materijali i metode
Istraživanjem je obuhvaćeno dvadeset jedinki u dobi od 7±0,5 mjeseci (10 purana i 10 pura) uzgajanih ekstenzivno u ujednačenim uvjetima. Trupovi tuka obrađeni su prema
metodi koju su opisali Hahn i Spindler (2002.). Uz tjelesnu masu živih jedinki prikazan je prosječni randman te masa i udio prsa, zabataka i bataka u ukupnoj živoj masi jedinki. Randman je računat kao udio očišćenog, eviscerirenog, obrađenog i ohlađenog trupa u tjelesnoj masi živih jedinki prije klanja. Prosječni udio kao i najmanja i najveća utvr-đena masa pojedinih tkiva (kože i potkožnog masnog tkiva, mišića, tetiva i ovojnica, masnog tkiva i kosti) utvrđeni su disekcijom prsa (cijelih), zabataka i bataka (lijevih) prema metodi koju je opisao Jensen (1983.). Obrada i analiza rezultata provedena je pomoću referentnog programa Statistica v.10 (StatSoft Inc., 2011.). Značajnost razlika između utvrđene mase te udjela pojedinih dijelova trupova i tkiva u purana i pura testirana je Studentovim T-testom te Mann-Whitneyevim U testom.
Rezultati i rasprava
Prosječna tjelesna masa živih jedinki, randman, kao i udio prsa, zabataka i bataka u tjelesnoj masi živih jedinki prikazani su u tablicama 1. i 2. Iz prikazanih tablica 1. i 2. uočljivo je da su prosječna tjelesna masa kao i masa prsa, zabataka i bataka u muških jedinki bile značajno veće u odnosu na vrijednosti utvrđene u ženki (p<0,001). No, ukupna iskoristivost (randman) trupa kao i udio pojedinih važnijih dijelova u ukupnoj masi živih jedinki bili su ujednačeni u oba spola (p>0,05). Iako purani dalmatinskog zaleđa morfološki uvelike nalikuju divljem izvornom obliku broncastog purana, a njihovi uvjeti držanja staništu divljeg purana (Ekert Kabalin i sur., 2009.a,b,c, 2011., 2012.), iskoristivost trupova i udio najvrijednijih dijelova tijela su im nešto veći. Večerek i sur. (2008.) su utvrdili da prosječni randman divljih purana iznosi 67,99%, s prosječnom živom tjelesnom masom je-dinki oba spola od 3932,00±915,31 g. U istraživanju Hu-leta (2006.) prosječna masa pura tri komercijalna hibrida (BUTA, Hybrid i Nicholas) u dobi od 15 tjedana kretala se od 8895,4 do 9823,6 g, dok je randman iznosio od 78,80% do 79,78%. Herendy i sur. (2003.) su uspoređivali proiz-vodne značajke te iskoristivost broncastog purana (nalik izvornom tipu) u ekstenzivnom uzgoju i komercijalnog hibrida BUT. Pritom su ustanovili da se randman bronca-stog purana kretao u rasponu od 62,9% do 65,3%, a komercijalnog BUT hibrida od 76,1% do 83,4%, tj. bio je bolji za oko 13%-18%.
Tablica 1. Utvrđena tjelesna masa te randman po spolu Table 1. Body weight and yield of carcasses according to sex
Purani / Turkey-cocks Pure / Turkey-hens
arit.sred. / mean ± std.dev. / SD arit.sred.
/ mean ± std.dev. / SD p-vrijednost /
p-value
Tjelesna masa (g) / Body weight (g) 5848,00 ± 488,28 3880,00 ± 312,98 <0,001
Randman (%) / Yield (%) 70,41 ± 4,91 70,83 ± 3,24 >0,05

ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA
137
Tablica 2. Prosječna masa te udio prsa, zabataka i bataka u tjelesnoj masi živih jedinki Table 2. Mean mass and share of breast, thighs and drumsticks in live body weight
Purani / Turkey-cocks Pure / Turkey-hens arit. sred.
mean ± std. dev. SD
arit. sred. mean ± std. dev.
SD
p-vrijednost p-value
masa (g) weight (g) 1253,40 ± 267,83 749,20 ± 103,67 <0,001 Prsa s kožom, potkožnim
masnim tkivom i prsnom kosti Breast with skin, subcutaneous
fat and chest bone udio / share (%) 21,30 ± 3,54 19,25 ± 1,46 >0,05
masa / weight (g) 822,40 ± 140,81 534,20 ± 77,87 <0,001 Prsni mišić bez kosti Breast muscle without bone udio / share (%) 13,98 ± 1,52 13,74 ± 1,33 >0,05
masa / weight (g) 572,20 ± 51,92 399,80 ± 43,02 <0,001 Zabataci / Thighs
udio / share (%) 9,80 ± 0,70 10,29 ± 0,48 >0,05
masa / weight (g) 541,40 ± 52,07 349,00 ± 24,41 <0,001 Bataci / Drumsticks
udio / share (%) 9,28 ± 0,75 9,06 ± 0,79 >0,05
Sogut i sur. (2010.) navode kako su pojedini istraživači utvrdili da način uzgoja tovnih pasmina i hibrida purana s pristupom ispaši ili bez toga nije značajno utjecao na masu trupa, prsa ili nogu (bataka i zabataka), što ukazuje na značajan utjecaj genotipa jedinki na navedene pokazatelje. Nadalje, značajan je utjecaj stope rasta, poglavito nakon 16. tjedna, na prinos prsnog mišićja, pričem teži, ranozre-liji sojevi imaju veći udio prsnog mišićja (www.hybrid-turkeys.com). Rezultati istraživanja Herendya i sur. (2003.) pokazali su da komercijalni BUT hibrid ima prosječno 10% veći udio prsnog mišićja od ekstenzivno uzgajanog broncastog purana. Prosječna masa prsa u istraživanju Večereka i sur. (2008.) u divljih purana oba spola iznosila je 518,24±164,72 g, uz randman u odnosu na živu masu tijela od 12,92%. Prosječni randman prsa (bez kosti) u našem istraživanju bio je nešto veći i iznosio 13,98% u
purana te 13,74% u pura. Prosječna masa prsa s kosti u pura tri komercijalna hibrida obuhvaćenih istraživanjem Huleta (2006.) kretala se od 2662,6 do 2841,6 g, pričem je randman u živoj masi iznosio 28,96% do 29,94%. Prosječ-ni randman u ovom istraživanju bio je 21,30% u purana te 19,25% u pura. Nadalje, masa bataka s kožom u istraživanju Večeraka i sur. (2008.) iznosila je 778,22±167,68 g, a randman 19,92%. Prema Huletu (2006.) ukupna masa nogu pura tri komercijalna hibrida iznosila je 2116,5-2299,2 g, uz rand-man od 23,38%-24,25% u ukupnoj tjelesnoj masi živih je-dinki. Prosječna masa bataka i zabataka u našem istraži-vanju bila je 19,08% žive mase u purana te 19,35% u pura. U tablicama 3. do 5. te na slikama 1. do 3. prikazani su rezultati disekcije prsa, zabataka i bataka u jedinki oba spola.
Tablica 3. Rezultati disekcije prsa po spolu Table 3. Dissection of breast according to sex
Purani / Turkey-cocks Pure / Turkey-hens prosječni udio u prsima (%) average share in breast (%)
prosječni udio u prsima (%) average share in breast (%) Dijelovi prsa / Breast parts
arit. sred. mean ± std. dev.
SD arit. sred.
mean ± std. dev. SD
p-vrijednost p-value
Koža i potkožno masno tkivo Skin and subcutaneous fat 18,04 ± 7,99 13,23 ± 3,86 >0,05
Kost / Bone 15,59 ± 2,56 15,83 ± 2,68 >0,05
Mišićno tkivo / Muscle tissue 63,69 ± 6,16 68,85 ± 3,81 >0,05
Tetive i ovojnice / Tendons and sheaths 1,52 ± 0,81 1,00 ± 0,49 >0,05
Masno tkivo / Adipose tissue 1,16 ± 0,48 1,09 ± 0,62 >0,05
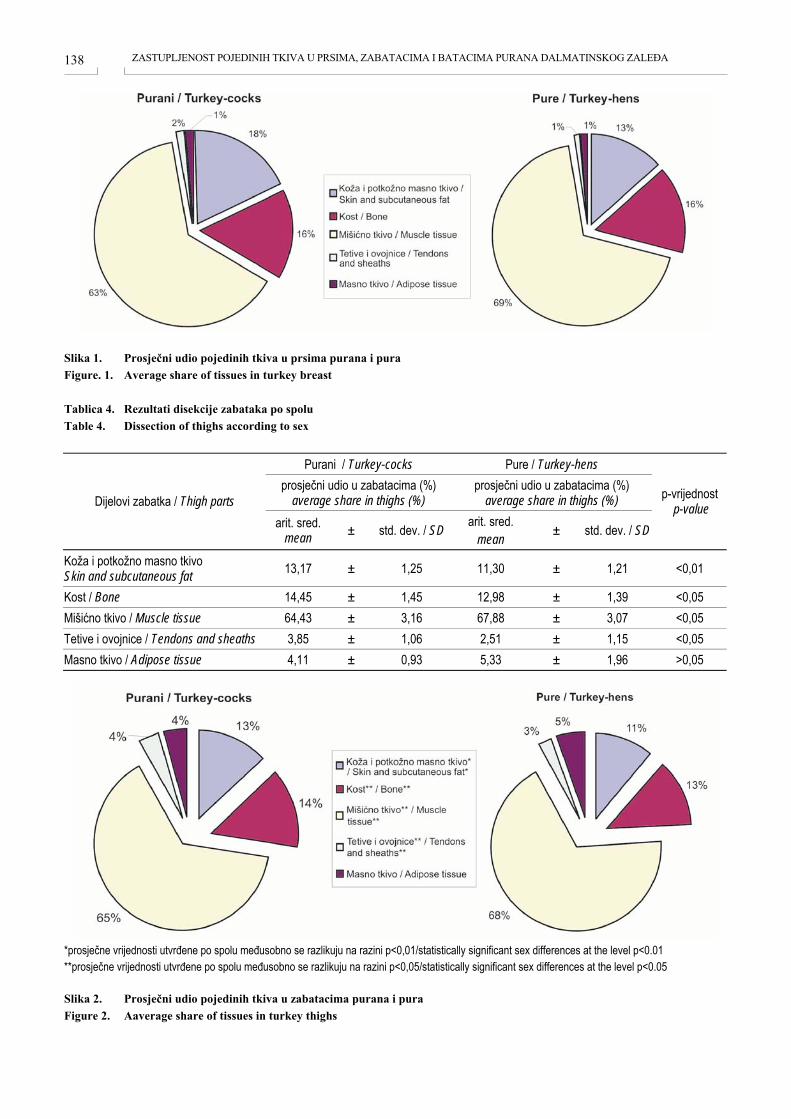
ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA 138
Slika 1. Prosječni udio pojedinih tkiva u prsima purana i pura Figure. 1. Average share of tissues in turkey breast Tablica 4. Rezultati disekcije zabataka po spolu Table 4. Dissection of thighs according to sex
Purani / Turkey-cocks Pure / Turkey-hens prosječni udio u zabatacima (%)
average share in thighs (%) prosječni udio u zabatacima (%)
average share in thighs (%) Dijelovi zabatka / Thigh parts arit. sred.
mean ± std. dev. / SD arit. sred. mean ± std. dev. / SD
p-vrijednost p-value
Koža i potkožno masno tkivo Skin and subcutaneous fat 13,17 ± 1,25 11,30 ± 1,21 <0,01
Kost / Bone 14,45 ± 1,45 12,98 ± 1,39 <0,05 Mišićno tkivo / Muscle tissue 64,43 ± 3,16 67,88 ± 3,07 <0,05 Tetive i ovojnice / Tendons and sheaths 3,85 ± 1,06 2,51 ± 1,15 <0,05 Masno tkivo / Adipose tissue 4,11 ± 0,93 5,33 ± 1,96 >0,05
*prosječne vrijednosti utvrđene po spolu međusobno se razlikuju na razini p<0,01/statistically significant sex differences at the level p<0.01 **prosječne vrijednosti utvrđene po spolu međusobno se razlikuju na razini p<0,05/statistically significant sex differences at the level p<0.05 Slika 2. Prosječni udio pojedinih tkiva u zabatacima purana i pura Figure 2. Aaverage share of tissues in turkey thighs
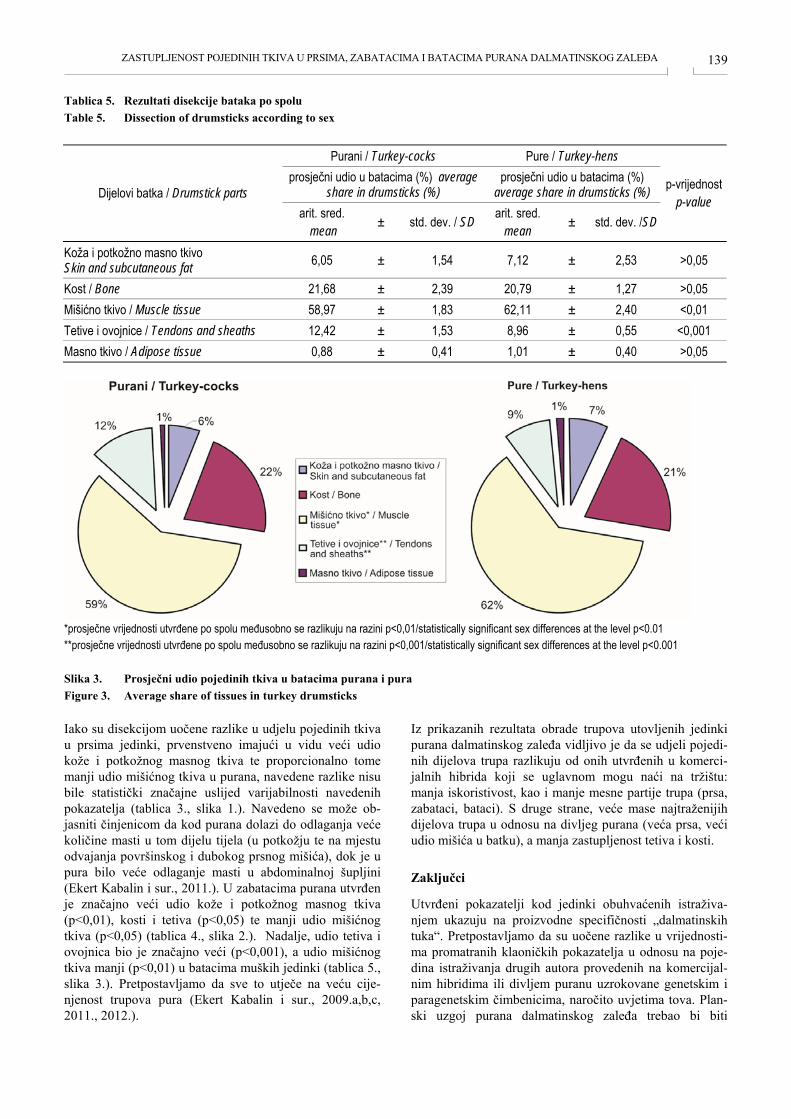
ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA
139
Tablica 5. Rezultati disekcije bataka po spolu Table 5. Dissection of drumsticks according to sex
Purani / Turkey-cocks Pure / Turkey-hens prosječni udio u batacima (%) average
share in drumsticks (%) prosječni udio u batacima (%)
average share in drumsticks (%) Dijelovi batka / Drumstick parts arit. sred.
mean ± std. dev. / SD arit. sred. mean ± std. dev. /SD
p-vrijednost p-value
Koža i potkožno masno tkivo Skin and subcutaneous fat 6,05 ± 1,54 7,12 ± 2,53 >0,05
Kost / Bone 21,68 ± 2,39 20,79 ± 1,27 >0,05 Mišićno tkivo / Muscle tissue 58,97 ± 1,83 62,11 ± 2,40 <0,01 Tetive i ovojnice / Tendons and sheaths 12,42 ± 1,53 8,96 ± 0,55 <0,001 Masno tkivo / Adipose tissue 0,88 ± 0,41 1,01 ± 0,40 >0,05
*prosječne vrijednosti utvrđene po spolu međusobno se razlikuju na razini p<0,01/statistically significant sex differences at the level p<0.01 **prosječne vrijednosti utvrđene po spolu međusobno se razlikuju na razini p<0,001/statistically significant sex differences at the level p<0.001 Slika 3. Prosječni udio pojedinih tkiva u batacima purana i pura Figure 3. Average share of tissues in turkey drumsticks
Iako su disekcijom uočene razlike u udjelu pojedinih tkiva u prsima jedinki, prvenstveno imajući u vidu veći udio kože i potkožnog masnog tkiva te proporcionalno tome manji udio mišićnog tkiva u purana, navedene razlike nisu bile statistički značajne uslijed varijabilnosti navedenih pokazatelja (tablica 3., slika 1.). Navedeno se može ob-jasniti činjenicom da kod purana dolazi do odlaganja veće količine masti u tom dijelu tijela (u potkožju te na mjestu odvajanja površinskog i dubokog prsnog mišića), dok je u pura bilo veće odlaganje masti u abdominalnoj šupljini (Ekert Kabalin i sur., 2011.). U zabatacima purana utvrđen je značajno veći udio kože i potkožnog masnog tkiva (p<0,01), kosti i tetiva (p<0,05) te manji udio mišićnog tkiva (p<0,05) (tablica 4., slika 2.). Nadalje, udio tetiva i ovojnica bio je značajno veći (p<0,001), a udio mišićnog tkiva manji (p<0,01) u batacima muških jedinki (tablica 5., slika 3.). Pretpostavljamo da sve to utječe na veću cije-njenost trupova pura (Ekert Kabalin i sur., 2009.a,b,c, 2011., 2012.).
Iz prikazanih rezultata obrade trupova utovljenih jedinki purana dalmatinskog zaleđa vidljivo je da se udjeli pojedi-nih dijelova trupa razlikuju od onih utvrđenih u komerci-jalnih hibrida koji se uglavnom mogu naći na tržištu: manja iskoristivost, kao i manje mesne partije trupa (prsa, zabataci, bataci). S druge strane, veće mase najtraženijih dijelova trupa u odnosu na divljeg purana (veća prsa, veći udio mišića u batku), a manja zastupljenost tetiva i kosti.
Zaključci
Utvrđeni pokazatelji kod jedinki obuhvaćenih istraživa-njem ukazuju na proizvodne specifičnosti „dalmatinskih tuka“. Pretpostavljamo da su uočene razlike u vrijednosti-ma promatranih klaoničkih pokazatelja u odnosu na poje-dina istraživanja drugih autora provedenih na komercijal-nim hibridima ili divljem puranu uzrokovane genetskim i paragenetskim čimbenicima, naročito uvjetima tova. Plan-ski uzgoj purana dalmatinskog zaleđa trebao bi biti

ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA 140
usmjeren na razvijanje strategije koja s jedne strane omo-gućuje optimalno iskorištavanje dostupnih resursa, a s dru-ge strane istodobno opskrbljuje tržište proizvodom visoke kakvoće. S obzirom na povećano zanimanje za očuvanje bioraznolikosti samoodrživim, ekološko-organskim uzgo-jem vjerujemo da uzgoj tuka može pomoći regionalnom razvoju, a specifičan gastronomski proizvod doprinijeti prepoznatljivosti dalmatinskog zaleđa, pa i Hrvatske na međunarodnom tržištu.
Literatura
ALDERS, R. G., R. A. E. PYM (2009): Village poultry: still important to millions, eight thousand years after domesti-cation. World Poult. Sci. J. 65, 2, 181-190.
BOGOSAVLJEVIĆ-BOŠKOVIĆ, S., S. MITROVIĆ, V. RADO-VIĆ, M. PETROVIĆ (2005): Odabrani parametri kvalitete mesa brojlerskih pilića iz različitih sistema gajenja. Biotech. Anim. Husbandry 21, 3-4, 133-140.
BRAKE, J., G. B. HAVENSTEIN, P. R. FERKET, D. V. RIVES, F. G. GICSBRECHT (1994): Relationship of sex, strain and body weights to carcass yield and offal production in turkeys. Poult. Sci. 74, 1, 161-168.
EKERT KABALIN, A., S. MENČIK, I. ŠTOKOVIĆ, Š. HORVATH, A. GRGAS, T. BALENOVIĆ, V. SUŠIĆ, I. KARADJOLE, M. OSTOVIĆ, Ž. PAVIČIĆ, D. MARKO-VIĆ, D. MARGUŠ (2009a): Future of Dalmatian turkey – traditional local form of poultry in Croatia. 60th Annual Meeting of the European Association for Animal Production (EAAP 2009), Barcelona, Spain (Local breeds: what future? 1. Selection), 16-17.
EKERT KABALIN, A., S. MENČIK, M. OSTOVIĆ, I. ŠTO-KOVIĆ, A. GRGAS, Š. HORVATH, T. BALENOVIĆ, V. SUŠIĆ, I. KARADJOLE, Ž. PAVIČIĆ (2012): Morpho-logical characteristics of Dalmatian turkey: preliminary results. Maced. J. Anim. Sci. 2, 3, 277-280.
EKERT KABALIN, A., S. MENČIK, M. OSTOVIĆ, M. BALENOVIĆ, I. ŠTOKOVIĆ, V. SUŠIĆ, T. BALENOVIĆ, Ž. PAVIČIĆ (2011): Fenotipske odlike purana dalmatinskog zaleđa. Stočarstvo 65, 2, 139-145.
EKERT KABALIN, A., Š. HORVATH, D. MARKOVIĆ, D. MARGUŠ, V. SUŠIĆ, T. BALENOVIĆ, I. KARADJOLE, I. ŠTOKOVIĆ, S. MENČIK, Ž. PAVIČIĆ, A. GRGAS, M. BALENOVIĆ, M. OSTOVIĆ (2009b): Identification and characterisation of Dalmatian turkey. 1st Conference of the Balkan Network for the Animal Reproduction & Bio-technology. Sofia, Bulgaria, 16-17.
EKERT KABALIN, A., Š. HORVATH, V. SUŠIĆ, T. BALE-NOVIĆ, I. KARADJOLE, M. BALENOVIĆ, D. MARGUŠ, D. MARKOVIĆ, A. GRGAS, Ž. PAVIČIĆ, I. ŠTOKOVIĆ, S. MENČIK, M. OSTOVIĆ (2009c): Dalmatinska tuka – arhaična forma peradi. Stočarstvo 63, 1, 49-55.
HAHN, G., M. SPINDLER (2002): Methods of dissection of turkey carcasses. World Poult. Sci. J. 58, 2, 179-197.
HALVORSON, J. C., P. E. WAIBEL, E. M. OJU, S. L. NEOL, M. E. El HALAWANI (1991): Effect of diet and population density on male turkeys under various environmental con-ditions. 2: Body composition and meat yield. Poult. Sci. 70, 4, 935-940.
HERENDY, V., Z. SÜTŐ, P. HORN (2003): Characteristics of improvement in the turkey production in the last 30 years. Agric. Conspec. Sci. 68, 2, 127-131.
HULET, R. M. (2006): Comparative meat yield of modern com-mercial turkeys. [http://www.zootecnicainternational.com/ /article-archive/processing/903-comparative-meat-yield-of-modern-commercial-turkeys.html (10.11.2011.)]
JENSEN, J. F. (1983): Method of dissection of broiler carcasses and description of parts. World's Poultry Science Association European Federation Working Group V, Copenhagen, Den-mark. Papworth's Pendragon Press, Cambridge, UK, p 32.
LAUDADIO V., V. TUFARELLI, M. DARIO, F. P. D'EMIL-LIO, A. VICENTI (2009): Growth performance and carcass characteristics of female turkeys as affected by feeding programs. Poult. Sci. 88, 4, 805-810.
MEDIĆ, H., S. BIDAČEK, K. SEDLAR, V. ŠATOVIĆ, T. PETRAK (2009): Utjecaj vrste i spola peradi te tehnološkog procesa hlađenja na kvalitetu mesa. Meso 11, 4, 222-231.
MENČIK, S., A. EKERT KABALIN, T. BALENOVIĆ, Ž. PA-VIČIĆ, I. ŠTOKOVIĆ, M. OSTOVIĆ, A. GRGAS (2009): Utvrđivanje morfoloških obilježja dalmatinskih tuka. Zbor-nik sažetaka. Znanstveno-stručni sastanak Veterinarska zna-nost i struka. Zagreb, 96-97.
MUŽIC, S. G. KRALIK, R. RAGUŽ-ĐURIĆ, Z. JANJEČIĆ, B. BOBETIĆ (2008): Peradarska proizvodnja u Republici Hrvatskoj. Krmiva 50, 6, 353-358.
NESTOR, K. E., J. W. ANDERSON, D. HARTZLER, S. G. VELLEMAN (2005): Genetic variation in pure line and crosses of large-bodied turkeys. 4: Body shape and carcass traits. Poult. Sci. 84, 12, 1825-1834.
PLATZ, S., J. BERGER, F. AHRENS, U. WEHR, W. RAMBECK, W. AMSELGRUBER, M. H. ERHARD (2003): Health, productivity and behaviour of conventional turkey breeds under ecological outdoor rearing conditions. Proceedings of the ХІ International Congress on Animal Hygiene, Mexico, pp 259-264.
RIZZI, C., M. BARUCHELLO, G. M. CHIERICATO (2009): Slaughter performance and meat quality of three Italian chicken breeds. Ital. J. Anim. Sci. 8 (Suppl. 3), 228-230.
ROBERSON, K. D., A. P. RAHN, R. J. BALANDER, M. W. ORTH, D. M SMITH, B. L. BOOREN, A. M. BOOREN, W. N. OSBURN, R. M. FULTON (2003): Evaluation of the growth potential, carcass compounds and meat quality characteristics of three commercial strains of tom turkeys. J. Appl. Poult. Res. 12, 2, 229-236.
SOGUT, B., O. F. KURBAL, H. DOMIRULUS, H. INCI (2010): Growth performance of Big-6 Broiler White turkeys in the different rearing conditions. J. Anim. Vet. Adv. 9, 7, 1156-1159.
VEČEREK. V., V. ŠERMAN, F. VITULA, E. STRAKOVÁ, P- SUCHÝ, N. MAS, Z. LUKAC (2008): Klaonička vrijednost odabranih vrsta pernate divljači. Krmiva 50, 6, 335-344.
WALDROUP, P. W., J. A. ENGLAND, A. L. WALDROUP, N. B. ANTHONY (1997): Response of two strains of Large White male turkeys to amino acid levels when diets are changed at three- or four-week intervals. Poult. Sci. 76, 11, 1543-1555.
WERNER, C., J. RIEGEL, M. WICKE (2008): Slaughter performance of four different turkey strains, with special focus on the muscle fiber structure and the meat quality of the breast muscle. Poult. Sci. 87, 9, 1849-1859.
www.hybridturkeys.com – Info Sheet Hybrid, A Hendrix Genetics Company: Breast Meat Yield in Commercial Turkeys: Utilizing the Genetic Resource [pristupano 10.11.2011.]

ZASTUPLJENOST POJEDINIH TKIVA U PRSIMA, ZABATACIMA I BATACIMA PURANA DALMATINSKOG ZALEĐA
141
TISSUE SHARE IN THE BREAST, THIGHS AND DRUMSTICKS OF TURKEYS FROM DALMATIAN HINTERLAND
Summary
Extensive fattening of turkeys at family households in Dalmatian hinterland (so called "Dalmatian tuka") is focused on seasonal consumption of turkey meat, mostly during winter. Breeding as well as the specificity of the genotype are some of the reasons for the characteristic aroma and reputability of their meat. This survey was performed on 20 turkeys (10 of either sex), mean age 7±0.5 months, bred under uniform conditions. Carcass dissection was performed to characterize the quality of the most important parts of meat (breast, thighs and drumsticks) in which the share of tissues was determined (skin and subcutaneous adipose tissue, muscle tissue, tendons and sheaths, adipose tissue and bone). The mean yield in turkeys-cocks was 70.41±4.91% and in turkey-hens 70.83±3.24%. The absolute values of body weight of live animals and the mass of breast, thighs and drumsticks were significantly higher in males (p<0.001 all). However, their share in the live body weight was uniform. We found no significant difference between tissue shares in the breast. Turkey-cocks had a considerably higher share of skin and subcutaneous adipose tissue (p<0.01), bones and tendons (p<0.05) and lower share of muscle tissue (p<0.05) in thighs. Also, the share of tendons and sheaths was significantly higher (p<0.001) in drumsticks of males, while the share of muscle tissue was lower (p<0.01). Key words: turkey meat, extensive breeding, tissue share

PERADARSKI DANI 2013. 142
PROIZVODNI REZULTATI I KVALITETA JAJA SLOBODNO DRŽANIH KOKOŠI HRVATICA
Zlatko Janječić1, Stjepan Mužic1, Dalibor Bedeković1, Zoran Grgić1, Gordana Duvnjak2, Marina Ćurak1, Tihana Bišćan3
1Agronomski fakultet Sveučilišta u Zagrebu, Hrvatska 2Hrvatska poljoprivredna agencija, Zagreb, Hrvatska 3Poljoprivredna savjetodavna služba, Zagreb, Hrvatska Sažetak Slobodan način držanja kokoši nesilica jedan je od propisanih alternativnih načina proizvodnje kokošjih jaja u Europi, a proizvodnja konzumnih jaja na obiteljskim gospodarstvima u Hrvatskoj uglavnom se odvija na taj način. Cilj istraživanja bio je utvrditi proizvodnost hrvatske autohtone pasmine kokoši hrvatice u uvjetima slobodnog načina držanja. Kokoši u dobi od 20 tjedana useljene su na 11 obiteljskih gospodarstava, pričem su na svakom bila dva jata po 10 kokoši. Praćenje proizvodnih rezultata trajalo je 32 tjedna. Sve kokoši su hranjene po volji kompletnom krmnom smjesom koja je sadržavala 15% sirovih bjelančevina i 11,28 MJ/ME, a na raspolaganju su imale i zatravnjeni ispust po 10 m2 po kokoši. Praćen je broj i masa jaja te konzumacija krmne smjese. Za utvrđivanje čvrstoće ljuske, boje žutanjaka i vrijednosti Haughove jedinice iz svakog je jata u 10. tjednu istraživanja prikupljeno 10 jaja. Analizom isplativosti (cost-benefit) izračunati su ekonomski pokazatelji proizvodnje. Prosječan broj jaja po nesilici tijekom istraživanja iznosio je 130,98 jaja, konverzija krmne smjese 3,46/kg jajčane mase, a prosječna masa jaja iznosila je 51,4 grama. Čvrstoća ljuske bila je 3,10 kp/cm2, boja žutanjka 11 prema ljestvici La Roche, a vrijednost Haughovih jedinica 66,02. Ukupni prihodi koji su uključivali iznose dobivene prodajom jaja i državna poticajna sredstva bili su 57.742,50 kn, dok su ukupni varijabilni troškovi bili 21.666,60 kn. Pokriće varijabilnih troškova za svih 11 obiteljskih gospodarstava iznosio je 36.075,90 kn odnosno 3279,63 kn za jedno gospodarstvo. Prema navedenom zaključujemo da se kokoš hrvatica može u potpunosti preporučiti za proizvodnju jaja slobodnim načinom držanja na malim obiteljskim gospodarstvima u Hrvatskoj. Ključne riječi: proizvodni rezultati, kvaliteta jaja, slobodno držanje, kokoš hrvatica Uvod Dosadašnja se industrijska proizvodnja konzumnih ko-košjih jaja u gotovo svim zemljama svijeta odvijala u zatvorenim objektima, kaveznim načinom držanja pričem su uglavnom korišteni visoko selekcionirani hibridi kokoši nesilica. U posljednja tri desetljeća svjedoci smo sve većeg društvenog zanimanja za dobrobit životinja (Levy, 2004.), koji je za posljedicu imao i odredbu Komisije EU (1996.) kojom se od 1. siječnja 2012. proizvodnja konzumnih jaja u zemljama članicama mora odvijati u alternativnim susta-vima držanja konzumnih nesilica koji najčešće uključuju okruženja (prostor sa životinjama) bez kaveza, s moguć-nošću pristupa peradi prostoru izvan peradnjaka. Držanje i smještaj konzumnih nesilica u alternativnim sustavima
proizvodnje je jedna od najkontroverznijih i najdrastičnijih mjera koje su propisane u stočarskoj proizvodnji glede dobrobiti životinja. Slobodno držanje kokoši nesilica na zatravnjenim ispusti-ma jedan je od dozvoljenih alternativnih načina proizvod-nje konzumnih jaja koji ima neke elemente tradicionalnog držanja peradi (Savory, 2004.). Njime se kokošima nesili-cama želi osigurati veći prostor za kretanje i omogućiti prirodnije ponašanje (Appleby and Hughes, 1991.). U proizvodnji konzumnih jaja slobodnim načinom držanja mogu se koristiti hibridne kokoši nesilice stvorene za taj način proizvodnje ili čiste pasmine kokoši specifične za uzgoj u određenom podneblju. U Hrvatskoj postoji izvorna pasmina kokoši pod imenom hrvatica koja je stvorena na području Međimurja i Podra-
Prof. dr. sc. Zlatko Janječić, Sveučilište u Zagrebu, Agronomski fakultet Zagreb, Zavod za hranidbu životinja, Svetošimunska 25, 10000 Zagreb, Republika Hrvatska. Tel.: ++385 (0) 1 239 3951, fax: ++385 (0) 1 239 3932, e-mail: [email protected]
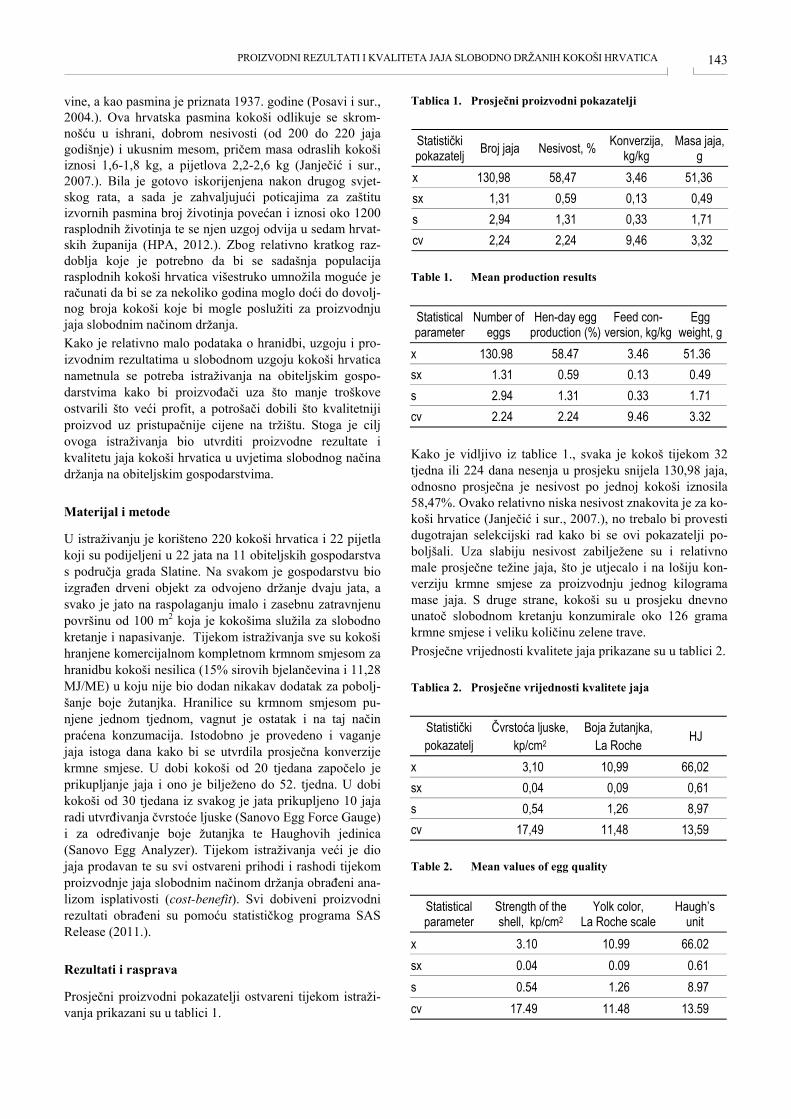
PROIZVODNI REZULTATI I KVALITETA JAJA SLOBODNO DRŽANIH KOKOŠI HRVATICA
143
vine, a kao pasmina je priznata 1937. godine (Posavi i sur., 2004.). Ova hrvatska pasmina kokoši odlikuje se skrom-nošću u ishrani, dobrom nesivosti (od 200 do 220 jaja godišnje) i ukusnim mesom, pričem masa odraslih kokoši iznosi 1,6-1,8 kg, a pijetlova 2,2-2,6 kg (Janječić i sur., 2007.). Bila je gotovo iskorijenjena nakon drugog svjet-skog rata, a sada je zahvaljujući poticajima za zaštitu izvornih pasmina broj životinja povećan i iznosi oko 1200 rasplodnih životinja te se njen uzgoj odvija u sedam hrvat-skih županija (HPA, 2012.). Zbog relativno kratkog raz-doblja koje je potrebno da bi se sadašnja populacija rasplodnih kokoši hrvatica višestruko umnožila moguće je računati da bi se za nekoliko godina moglo doći do dovolj-nog broja kokoši koje bi mogle poslužiti za proizvodnju jaja slobodnim načinom držanja. Kako je relativno malo podataka o hranidbi, uzgoju i pro-izvodnim rezultatima u slobodnom uzgoju kokoši hrvatica nametnula se potreba istraživanja na obiteljskim gospo-darstvima kako bi proizvođači uza što manje troškove ostvarili što veći profit, a potrošači dobili što kvalitetniji proizvod uz pristupačnije cijene na tržištu. Stoga je cilj ovoga istraživanja bio utvrditi proizvodne rezultate i kvalitetu jaja kokoši hrvatica u uvjetima slobodnog načina držanja na obiteljskim gospodarstvima.
Materijal i metode
U istraživanju je korišteno 220 kokoši hrvatica i 22 pijetla koji su podijeljeni u 22 jata na 11 obiteljskih gospodarstva s područja grada Slatine. Na svakom je gospodarstvu bio izgrađen drveni objekt za odvojeno držanje dvaju jata, a svako je jato na raspolaganju imalo i zasebnu zatravnjenu površinu od 100 m2 koja je kokošima služila za slobodno kretanje i napasivanje. Tijekom istraživanja sve su kokoši hranjene komercijalnom kompletnom krmnom smjesom za hranidbu kokoši nesilica (15% sirovih bjelančevina i 11,28 MJ/ME) u koju nije bio dodan nikakav dodatak za pobolj-šanje boje žutanjka. Hranilice su krmnom smjesom pu-njene jednom tjednom, vagnut je ostatak i na taj način praćena konzumacija. Istodobno je provedeno i vaganje jaja istoga dana kako bi se utvrdila prosječna konverzije krmne smjese. U dobi kokoši od 20 tjedana započelo je prikupljanje jaja i ono je bilježeno do 52. tjedna. U dobi kokoši od 30 tjedana iz svakog je jata prikupljeno 10 jaja radi utvrđivanja čvrstoće ljuske (Sanovo Egg Force Gauge) i za određivanje boje žutanjka te Haughovih jedinica (Sanovo Egg Analyzer). Tijekom istraživanja veći je dio jaja prodavan te su svi ostvareni prihodi i rashodi tijekom proizvodnje jaja slobodnim načinom držanja obrađeni ana-lizom isplativosti (cost-benefit). Svi dobiveni proizvodni rezultati obrađeni su pomoću statističkog programa SAS Release (2011.).
Rezultati i rasprava
Prosječni proizvodni pokazatelji ostvareni tijekom istraži-vanja prikazani su u tablici 1.
Tablica 1. Prosječni proizvodni pokazatelji
Statistički pokazatelj Broj jaja Nesivost, % Konverzija,
kg/kg Masa jaja,
g x 130,98 58,47 3,46 51,36 sx 1,31 0,59 0,13 0,49 s 2,94 1,31 0,33 1,71 cv 2,24 2,24 9,46 3,32 Table 1. Mean production results
Statistical parameter
Number of eggs
Hen-day egg production (%)
Feed con-version, kg/kg
Egg weight, g
x 130.98 58.47 3.46 51.36 sx 1.31 0.59 0.13 0.49 s 2.94 1.31 0.33 1.71 cv 2.24 2.24 9.46 3.32 Kako je vidljivo iz tablice 1., svaka je kokoš tijekom 32 tjedna ili 224 dana nesenja u prosjeku snijela 130,98 jaja, odnosno prosječna je nesivost po jednoj kokoši iznosila 58,47%. Ovako relativno niska nesivost znakovita je za ko-koši hrvatice (Janječić i sur., 2007.), no trebalo bi provesti dugotrajan selekcijski rad kako bi se ovi pokazatelji po-boljšali. Uza slabiju nesivost zabilježene su i relativno male prosječne težine jaja, što je utjecalo i na lošiju kon-verziju krmne smjese za proizvodnju jednog kilograma mase jaja. S druge strane, kokoši su u prosjeku dnevno unatoč slobodnom kretanju konzumirale oko 126 grama krmne smjese i veliku količinu zelene trave. Prosječne vrijednosti kvalitete jaja prikazane su u tablici 2. Tablica 2. Prosječne vrijednosti kvalitete jaja
Statistički pokazatelj
Čvrstoća ljuske, kp/cm2
Boja žutanjka, La Roche HJ
x 3,10 10,99 66,02 sx 0,04 0,09 0,61 s 0,54 1,26 8,97 cv 17,49 11,48 13,59 Table 2. Mean values of egg quality
Statistical parameter
Strength of the shell, kp/cm2
Yolk color, La Roche scale
Haugh’s unit
x 3.10 10.99 66.02 sx 0.04 0.09 0.61 s 0.54 1.26 8.97 cv 17.49 11.48 13.59
PROIZVODNI REZULTATI I KVALITETA JAJA SLOBODNO DRŽANIH KOKOŠI HRVATICA 144
Kao što se vidi u tablici 2., prosječna čvrstoća ljuske kod analiziranih jaja bila je zadovoljavajuća, jer su kokoši dio gradivnih elemenata za tvorbu ljuske zasigurno podmi-rivale i hranjivima pronađenim u ispustima. U prilog tome govori i boja žutanjka koja je bila intenzivno žuta do narančasta i rezultat je apsorpcije i deponiranja karotenoida koje su kokoši pohranile iz konzumirane trave u žutanjak jajeta. Nešto lošije vrijednosti Haughovih jedinica (A) mo-gu se pripisati visokim dnevnim temperaturama (iznad 30 ºC) tijekom mjeseca srpnja i nepravilnog dnevnog sa-kupljanja jaja. Analiza isplativosti (cost-benefit) s ukupnim prihodima i rashodima te pokrićem varijabilnih troškova prikazana je u tablici 3. U tablici 3. prikazani su ukupni prihodi koji su uključivali iznose dobivene prodajom jaja i državna poticajna sredstva i iznosili su 57.742,50 kn, dok su ukupni varijabilni troškovi bili 21.666,60 kn. Pokriće varijabilnih troškova za svih 11 obiteljskih gospodarstava iznosio je 36.075,90 kn, odnosno 3279,63 kn za jedno gospodarstvo. Kada bi se u
prihode uračunala i dobit od eventualne prodaje jedno-dnevnih pilića, tada bi i pokriće varijabilnih troškova za svako gospodarstvo bilo znatno veće. Ovime je vidljivo da je određeni dohodak tijekom godine na jednom obitelj-skom gospodarstvu moguće ostvariti držanjem nekoliko rasplodnih jata kokoši hrvatica, čime ženska populacija koja je uglavnom bila uključena u provedbu istraživanja ima i veće značenje unutar porodice, što se u potpunosti uklapa u prioritete Vlade RH o samozapošljavanju žena u ruralnim sredinama i sprječavanju napuštanja istih.
Zaključak
Provedeno istraživanje doprinijelo je popularizaciji hrvat-ske izvorne pasmine kokoši hrvatice i njezinom širenju u druge dijelove Hrvatske te se zaključuje da se njihovo korištenje u proizvodnji konzumnih jaja slobodnim na-činom držanja na obiteljskim gospodarstvima može prepo-ručiti, poglavito ako se za rasplodna jata nastave isplaćivati državni poticaji.
Tablica 3. Analiza isplativosti (cost-benefit)
Količina Mjerna jedinica Cijena, kn Ukupno, kn Varijabilni troškovi Kokoši i pijetlovi 242 Kljun 25,00 6.050,00 Krmna smjesa 6200 Kg 2,50 15.500,00 Veterinarski troškovi 242 Kljun 0,48 116,60 Ukupni varijabilni troškovi 21.666,60 Prihodi Jaja 28815 Komad 1,50 43.222,50 Državni poticaji 242 Kljun 60,00 14.520,00 Ukupni prihodi 57.742,50 Pokriće varijabilnih troškova 36.075,90 Table 3. Economic analysis
Quantity Measure unit Price per unit, € Total, € Variable costs Hens and rosters 242 Piece 3.20 774.40 Feed 6200 Kg 0.32 1 984.00 Veterinarian costs 245 Piece 0.06 14.70 Total variable costs 2 773.10 Income Eggs 28.815 Piece 0.20 5 763.00 Government incentives 242 Piece 7.90 1 911.80 Total income 7 674.80 Variable costs covered 4 901.70

PROIZVODNI REZULTATI I KVALITETA JAJA SLOBODNO DRŽANIH KOKOŠI HRVATICA
145
Zahvala
Prikazani rezultati proizišli su iz VIP-projekta ”Alterna-tivna proizvodnja jaja na obiteljskim gospodarstvima”, provedenog uz potporu Ministarstva poljoprivrede, ribar-stva i ruralnog razvoja Republike Hrvatske i grada Slatine.
Literatura
APPLEBY, M. C., B. O. HUGHES (1991): Welfare of laying hens in cages and alternative systems; environmental, physi-cal and behavioural aspects. World's Poultry Science Jour-nal, 47:109-128.
European Commission (1996) Report on the Welfare of Laying Hens. Brussels: Scientific Veterinary Committee, Animal Welfare Section, 30 October 1996.
HPA (2012): Godišnje izvješće 2012. HPA, Križevci. JANJEČIĆ, Z., S. MUŽIC, VLASTA HERAK-PERKOVIĆ
(2007): Proizvodnost kokoši Hrvatica. Praxis Veterinaria 3:117-124.
LEVY, N. (2004): What Makes Us Moral? Crossing the Boundaries of Biology. Oneworld, Oxford, UK.
POSAVI, M., M. ERNOIĆ, R. OZIMEC, F. POLJAK (2002): Hrvatske pasmine domaćih životinja. Ministarstvo zaštite okoliša i prostornog uređenja, Zagreb.
SAS Release (2011): SAS/STAT 9.3. User's Guide. SAS Institute Inc., Cary, NC.
SAVORY, C. J. (2004): Laying hen welfare standards: a classic case of ”power to the people”. Animal Welfare 13:S153-S158.
PRODUCTIVITY AND EGG QUALITY OF FREE RANGE RAISED HRVATICA HENS
Summary The aim of this study was to determine the productivity of the Croatian breed of hens hrvatica under free-range raising. Hens aged 20 weeks were housed in 11 family farms, two flocks of 10 hens each. Production results were monitored for 32 weeks. All hens were fed complete feed mixtures (15% protein and 11.28 MJ/ME), and they had access to the grass discharge (10 m2/hen). Data on the number and weight of eggs and feed consumption were collected. The strength of the shell, yolk color and Hough’s unit values of 10 eggs of each flock were determined in week 10 of investigation. Cost benefit analysis was done to calculate economic indicators of the production. The mean number of eggs per hen was 130.98 eggs, feed conversion 3.46 kg/kg of egg mass and mean egg weight 51.4 grams. The strength of the shell was 3.10 kp/cm2, yolk color at La Roche scale was 11 and the value of Haugh’s unit was 66.02. Total income included amounts received by selling eggs and government incentives amounted to 7 674.80 €, while the variable costs were 2 773.10 €. Contribution of cover for all 11 family farms was 4 901.70 € or 445.61 € /farm. According to the above, we conclude that the hen hrvatica can be fully recommended for free-range raising on small family farms in Croatia while there are government incentives. Key words: productivity, egg quality, free range egg production, hrvatica hen

PERADARSKI DANI 2013. 146
PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO
Kristina Matković1, Bara Vinković2, Marija Vučemilo1, Ranka Šimić3, Danijel Marušić4, Marica Lolić5
1 Zavod za higijenu, ponašanje i dobrobit životinja, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 2 Plotnikova 12, Zagreb, Hrvatska 3 Ministarstvo poljoprivrede Republike Hrvatske, Zagreb, Hrvatska 4 Brodsko-posavska županija, Slavonski Brod, Hrvatska 5 Veterinarski zavod Vinkovci, Hrvatski veterinarski institut, Vinkovci, Hrvatska
Sažetak
Sredinom prošloga stoljeća većina tradicionalne animalne proizvodnje značajno se promijenila. To je naročito izraženo kod držanja kokoši nesilica. U nekoliko godina mali uzgoji slobodno držanih nesilica zamijenjeni su intenzivnim baterijskim sustavima. To je sve više poticalo rasprave o fiziološkim potrebama, osjećajima i pravima životinja. Pitanje je kako procijeniti normalno ponašanje, što na njega ukazuje, te koliki je njegov udjel u ocjeni ukupnog stanja dobrobiti. Neki autori predlažu za polaznu točku ponašanje što ga kokoši prakticiraju u prirodnom okolišu. Stoga kod procjene dobrobiti treba temeljito razmotriti, a potom zaključiti koliko se pojedina kokoš u modernom uzgoju može ponašati u skladu s prirodnim ponašanjem. Isto tako nužno je pronaći odgovor na pitanje koliko uopće mogućnost manifestiranja prirodnog ponašanja osigurava visoku razinu dobrobiti. Uz to, kokoši u modernim uvjetima držanja mogu razviti drugačije obrasce ponašanja pa time omogućiti zadovoljavajuću dobrobit. Na odabranim gospodarstvima mjereni su pokazatelji mikroklimatskog kompleksa u zatečenom stanju objekata, te su prikupljani vrijednosni pokazatelji za procjenu dobrobiti nesilica, kao što su dostupnost hrani, vodi, tehničko stanje kaveza, uređaja za prozračivanje objekta, tehnike za izgnojavanje i drugo. Potom su analizirani elementi ponašanja i kondicije nesilica, također nužnih za procjenu njihove dobrobiti. Prema dobivenim rezultatima provedenog istraživanja može se zaključiti da smještaj nesilica u kaveze nije odgovarajući s aspekta dobrobiti zbog nemogućnosti kretanja, čeprkanja, kupanja u prašini, šepurenja i zadovoljenja drugih etoloških potreba, ali je ostvaren zadovoljavajući higijenski standard, laka je kontrola nesilica, uz vrlo mali broj oštećenih i prljavih jaja. Hranjenje i napajanje bilo je zadovoljavajuće prema tehnološkim zahtjevima proizvodnje, kao i zdravstveno stanje jata. Ključne riječi: ponašanje, dobrobit, kokoši nesilice, kavezno držanje, slobodno držanje
Uvod U prošlosti veza između ljudi i životinja bila je nepo-sredna. Tradiciju njegovanja takvih veza nalagala su pra-vila o postupcima sa životinjama što su ih na razne načine propisivale i kontrolirale društvene zajednice. Razvitkom ljudskog društva ne samo u ekonomskom, već i u filozof-skom i etičkom smislu, propitivana su znanja o postupcima sa životinjama, naročito životinja držanih na farmama. Dobrobit životinja kao znanstvena disciplina nastaje iz-među 50.-ih i 80.-ih godina prošloga stoljeća i od tada se razvija velikom brzinom. Puno komponenata ove disci-
pline i osnova bazirana na etici postojale su puno prije. Bolest i zdravlje, hranidba, smještaj životinja, njega, i u taj kontekst još dodani stres, fiziologija i ponašanje, dobije se zaokružena cjelina – dobrobit. Najpoznatija definicija kaže da je dobrobit stanje u kojem se jedinka pokušava nositi sa svojim okolišem (Broom, 1991.). Sve je počelo 1960.-ih godina knjigom Ruth Harrison "Animal Machines", kojom se i javnost zainteresirala za dobrobit životinja. Ljudi su počeli sve više javno dizati svoj glas o tome da sa životinjama treba postupati humano, javlja se briga za okoliš. Počinje se razvijati koncept odr-živosti.
Doc. dr. sc. Kristina Matković, dr.vet.med., Veterinarski fakultet Sveučilišta u Zagrebu, Zavod za higijenu, ponašanje i dobrobit životinja, Heinzelova 55, Zagreb, Hrvatska; e-mail: [email protected]

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO
147
Glede dobrobiti, s jedne strane su proizvođači koji žele životinje koje proizvode i osiguravaju im zaradu, a s druge strane sve jača svijest o dobrobiti životinja, sve glasnija društva za zaštitu životinja (Ladewig, 2008.). Najteže je pronaći ravnotežu imeđu te dvije strane, a i prava istina je negdje između. U intenzivnoj, mnoštvenoj proizvodnji jednostavno se životinje mora lišiti nekih slo-boda u zamjenu za redovite obroke, osiguran zaklon i njegu. Zanimanje znanosti za pitanja dobrobiti životinja potak-nulo je 1965. godine istraživanje o uvjetima u kojima uistinu žive farmske životinje. Istraživanje je provedeno, a rezultati su poznati kao Brambellov izvještaj. U tom do-kumentu je istaknuto pet odrednica slobode životinja i danas najčešće korištenih za definiciju dobrobiti. To su sloboda od: • gladi i žeđi • stresa • bolova, ozljeda i bolesti • nemogućnosti izražavanja normalnog ponašanja • straha i tjeskobe. Na tragu spomenutih istraživanja donesen je niz zakona o zaštiti i dobrobiti životinja. Stavljanje brige o životinjama u zakonske okvire potaknuli su dodatno sami potrošači traženjem odgovornije skrbi za domaće životinje. Ti su zahtjevi prepoznati pod pojmom dobrobit, pojmu koji bilo u užem ili širem smislu te riječi nije jednostavno definirati. Prema Šuleku (1990.) dobrobit znači dobar bitak, dobra bit. Anić (1998.) isti pojam objašnjava kao stanje dobrog zdravlja, sigurnosti, sreće i prosperiteta pojedinca ili skupine, a Hornby (1986.) pod pojmom welfare smatra uvjete za dobro zdravlje, udobnu radnu i životnu sredinu. Iz ovoga se može zaključiti da dobrobit životinja (engl. animal welfare) predstavlja uvjete u kojima životinja može ostvariti svoje biološke i fiziološke, vrsno specifične po-trebe, kao što su hranidba i napajanje, zatim imati primje-ren zaklon, preciznije prostor za smještaj u kojem može ostvariti fizičku, psihičku i termičku udobnost, potom socijalni kontakt sa životinjama iste vrste, manifestirati osnovne oblike ponašanja i biti slobodna od neugoda što ih nanose patnja, stres, bolesti i povrede. Uza spomenuto ovaj pojam označava brigu za životinje da one pod ljudskom skrbi mogu živjeti bez patnji i boli, što znači da im je čovjek izgradio siguran smještaj, osigurao dobru, u skladu s njihovom prirodom, hranidbu, osigurao zaštitu od bolesti, da im je osigurao pomoć veterinara ako su i kada su bolesne, te im pružio potrebnu ljubav i pažnju. Visoko specijalizirana peradarska proizvodnja dovela je kokoši do iscrpljujućih proizvodnih mogućnosti. Zapravo, kokoši su prve domaće životinje koje su prevedene iz prirodnog tradicionalnog načina držanja u strogo kontro-lirane uvjete, a peradarstvo je postalo ne samo velik biznis, već i obrazac za intenzivno, gotovo industrijsko iskorišta-vanje domaćih životinja. Iako je suvremeno peradarstvo nalik tvornici kojoj je na prvom mjestu profit, ipak se ozbiljno propituju etička načela takvog načina proizvodnje, naročito proizvodnje
jaja za ljudsku hranu. Javnost je počela negodovati na činjenicu da se nesilice drže u prenapučenim objektima, natiskane u tijesnim kavezima, prisiljene jesti i piti što im je ponuđeno, jedva se kretati i samo proizvoditi jaja. Slijedom navedenog ovim istraživanjem željela se procije-niti dobrobit i ponašanje kokoši nesilica smještenih u kon-vecionalne kaveze i slobodno.
Materijali i metode
Procjena dobrobiti nesilica obavljana je promatranjem, mjerenjem i opisivanjem stanja kao što su dostupnost hrani i vodi, izloženost uvjetima mikroklime, potom fizičko stanje nesilica, te njihovo ponašanje. Svi pokazatelji odre-đivani su u peradnjacima šest obiteljskih poljoprivrednih gospodarstava (OPG) Brodsko-posavske županije, u pet se kokoši nesilice drže konvencionalno, a u jednom slobodno. Mjerenja su obavljana deset puta u proljeće i ljeto 2011. godine. Temperatura zraka, relativna vlaga zraka i brzina strujanja zraka mjereni su uređajem Testo 400 (Testo Inc., Lenz-kirch, Njemačka), osvijetljenost je određivana luksmetrom Testo 0500 (Testo Inc., Lenzkirch, Njemačka), sadržaj ugljikovog dioksida i amonijaka uređajem Dräger-Multi-warn II (Dräger Njemačka). Broj bakterija i gljivica mjeren je uređajem Merck Mas 100 (Merck KgaA, Darmstadt, Njemačka). Procjena dobrobiti i ponašanje procijenjeni su prema pri-ručniku Welfare Quality – Assessment Protocol for Poultry, Bristol, Wageningen.
Rezultati i rasprava
Dostupnost hrani i vodi
Hranidba kokoši nesilica jaja za jelo od velike je važnosti i sudjeluje u do 70% svih troškova proizvodnje (Vučemilo, 2008.). Budući da se koriste selekcionirani linijski hibridi koji su velikog genetskog potencijala važno je hranu pra-vilno izbalansirati i osigurati optimalne količine i omjere. S obzirom na intenzivan metabolizam peradi, fiziološki vi-soku tjelesnu temperaturu i relativno kratak probavni sustav njihove potrebe za energijom su velike te hrana mora biti lako probavljiva i koncentrirana, a to su visoko-energetska koncentrirana krmiva (kukuruz, pšenica, ječam, raž i druge žitarice). Proizvođači iz analiziranih gospodar-stava većinom su sami miješali hranu uz dodatak premiksa. U konvencionalnim načinima držanja nesilica one se hrane iz žljebastih hranilica koje su smještene s vanjske strane svakog reda kaveza. Rapodjela hrane u hranilice je pomoću kolica ili lancem, što ovisi o tipu kaveza (Vučemilo, 2008.). Za napajanje peradi koristi se automatski nipl sustav koji se sastoji od četvrtaste plastične cijevi smještene duž gor-njeg dijela svakog reda kaveza u koju su učvršćeni nipli. Dnevno je nesilici potrebno osigurati 250 mL vode. Uko-liko je temperatura u objektu povišena kokoši popiju i do pet puta više vode nego što konzumiraju hrane.

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO 148
Tablica 1. Utvrđeni pokazatelji tehničke naravi s učincima na dobrobit nesilica (proljeće 2011. godine)
Pokazatelji OPG
Dostupnost hrani Dostupnost vodi Raspoloživo prostora/cm2/nesilica
Nesilice ubrzano dišu Nesilice drhte
I. Da/hranilica duž cijelog kaveza 2 nipl pojilice/kavezu 270 ne Ne II. Da/hranilica duž cijelog kaveza 2 nipl pojilice/kavezu 270 ne Ne III. Da/hranilica duž cijelog kaveza 2 nipl pojilice/kavezu 270 ne Ne IV. Da/hranilica duž cijelog kaveza 2 nipl pojilice/kavezu 270 ne Ne V. Da/hranilica duž cijelog kaveza 2 nipl pojilice/kavezu 270 ne Ne
OPG = obiteljsko poljoprivredno gospodarstvo; svih pet promatranih sustava bili su kavezni Table 1. Identified technical indicators, with effects on the welfare of laying hens (spring, 2011)
Indicators FAH
Food availability Water availability Available space/cm2/hen
Hens breathing fast Hens tremble
I Yes/feeder along the cage 2 nipple drinker/cage 270 No No II Yes/feeder along the cage 2 nipple drinker/cage 270 No No III Yes/feeder along the cage 2 nipple drinker/cage 270 No No IV Yes/feeder along the cage 2 nipple drinker/cage 270 No No V Yes/feeder along the cage 2 nipple drinker/cage 270 No No
FAH = family agricultural husbandry; all five observed systems were cages Izloženost osnovnim pokazateljima mikroklime U objektu je nužno održavati temperaturno vlažne odnose koji zadovoljavaju linijski hibrid tijekom cijele proizvodnje. Optimalna temperatura je između 18 i 22 °C (Vučemilo, 2008.). Tijekom zime temperatura ne bi smjela biti niža od 15 °C, a ljeti viša od 26 °C, jer se tada značajno smanjuje nesivost. Visoke temperature opasnije su od niskih, a ako se kroz duže vrijeme zadrže visoke temperature nesivost opada za 15% do 20%, jaja imaju tanju ljusku. Zbog toga je jako važan sustav za prozračivanje koji tada mora raditi mak-simalnom jačinom, uz otvaranje svih otvora.
Ovaj sustav je od vitalne važnosti za uspješnost peradnjaka. U svakom satu po jednom kilogramu žive mase nesilica u peradnjak treba ući 4 do 6 m3 svježeg zraka. Brzina strujanja treba biti od 0,25 do 0,30 m/s, ljeti ne veća od 0,50 m/s, bez propuha. Poželjna optimalna vlaga je između 65% i 75%. Tijekom zime ona može jako porasti, djelomice i stoga što nesilica tijekom 24 sata disanjem izluči 250 cm3 vode. Svjetlosni režim jedan je od najvažnijih čimbenika u proiz-vodnji jaja, jer utječe na spolno sazrijevanje i samu nesivost, stoga ga je puno lakše provoditi u objektima bez prozora.
Tablica 2. Srednje vrijednosti izmjerenih pokazatelja mikroklime u peradnjacima OPG-a Brodsko-posavske županije
(proljeće/ljeto 2011. godine)
Pokazatelji Koncentracija OPG Temperatura
zraka (°C) Vlaga zraka
(%) Brzina strujanja
zraka (m/s) Amonijaka (NH3 ppm)
Ugljičnog dioksida (CO2 vol%)
Osvijetljenost (lx)
Ukupno bakterija (CFU/m3 )
Ukupno gljivica
(CFU/m3)
I. 22 42 0,11 11 0,05 31 1,3x104 4,4x104
II. 25 59 0,10 3 0,04 33 0,7x104 0,1x104
III. 25 65 0,20 8 0,03 24 1,5x104 1,6x104
IV. 29 61 0,15 6 0,04 22 1,2x104 0,7x104
V. 24 54 0,06 5 0,03 24 1,3x104 0,1x104
OPG = obiteljsko poljoprivredno gospodarstvo; svih pet promatranih sustava bili su kavezni

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO
149
Table 2. Mean values of the measured microclimate parameters in poultry houses at FAHs in Brod-Posavina County (spring/summer 2011)
Indicators Concentration FAH Air
temperature (°C)
Relative humidity (%)
Air velocity (m/s) Ammonia
(NH3 ppm) Carbon dioxide
(CO2 vol%) Light (lx) Total bacteria
(CFU/m3 ) Total fungi (CFU/m3)
I 22 42 0.11 11 0.05 31 1.3x104 4.4x104
II 25 59 0.10 3 0.04 33 0.7x104 0.1x104
III 25 65 0.20 8 0.03 24 1.5x104 1.6x104
IV 29 61 0.15 6 0.04 22 1.2x104 0.7x104
V 24 54 0.06 5 0.03 24 1.3x104 0.1x104
FAH =family agricultural husbandry; all five observed systems were cages Fizičko stanje kokoši nesilica Tablica 3. Ocjena fizičkog stanja nesilica na promatranim OPG-ima Brodsko-posavske županije (jesen 2011./proljeće 2012.
godine)
Obiteljsko poljoprivredno gospodarstvo Fizička obilježja promatrano po 10 nesilica I. II. III. IV. V.
Tjelesna masa (g), izražene srednje vrijednosti 2000 2100 1900 2100 1800 Zadebljanje mekuši 4 3 4 4 4 Čistoća nogu 4 2 3 4 4
vrat 4 2 2 4 4 prsa 3 2 3 4 4 leđa 4 4 4 4 4 krila 3 1 2 4 3
Oštećenje perja
rep 4 2 4 4 4 glava 4 3 4 4 4 trtica 4 4 4 4 4 Oštećenje kože kloaka 3 2 3 4 3
Svih pet promatranih sustava bili su kavezni Table 3. Physical condition rating of hens at family agricultural husbandries in Brod-Posavina County (autumn 2011/spring
2012)
Family agricultural husbandry Physical characteristics, observed 10 hens I II III IV V
Body weight (g), expressed as mean values 2000 2100 1900 2100 1800 Thickening of the paws 4 3 4 4 4 Leg cleanliness 4 2 3 4 4
Neck 4 2 2 4 4 Chest 3 2 3 4 4 Beck 4 4 4 4 4 Wings 3 1 2 4 3
Feather damage
Tail 4 2 4 4 4 Head 4 3 4 4 4 Tail bone 4 4 4 4 4 Skin damage Cloaca 3 2 3 4 3
All five observed systems were cages

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO 150
Vrijednosni bodovi za ocjenu fizičkog stanja:
Fizičko stanje Broj bodova Bez oštećenja 4 Neznatno oštećenje 3 Umjereno oštećenje 2 Ozbiljno oštećenje 1
The value points for evaluation of the physical condition:
Physical condition Score No damage 4 Mild damage 3 Moderate damage 2 Serious damage 1
Ako više od 3 nesilice od 10 promatranih imaju oštećenja označena s 1 ili 2 boda treba pratiti ostale nesilice u jatu radi utvrđivanja uzroka oštećenja.
Slike 1. do 6. koje slijede odabrani su primjeri za ilu-straciju fizičkog stanja koja su poslužila za procjenu dobrobiti, prikazano u tablici 3.
Slika 1. Noga nesilice - nema oštećenja (4 boda) Figure 1. Hen leg - no damage (4 points)
Slika 3. Krilo nesilice - nema oštećenja (4 boda) Figure 3. Hen wing - no damage (4 points)
Slika 5. Umjerena oštećenja perja na vratu i prsima (2 boda) Figure 5. Moderate damage to the feathers on the neck and
chest (2 points)
Slika 2. Noga nesilice - umjereno oštećenje (2 boda) Figure 2. Hen leg - Moderate damage (2 points)
Slika 4. Krilo nesilice - ozbiljna oštećenja (1 bod) Figure 4. Hen wing - serious damage (1 point)
Slika 6. Umjerena oštećenja perja repa i područja kloake (2 boda)
Figure 6. Moderate damage to the tail feathers and cloaca area (2 points)

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO
151
Ponašanje
Odstupanje od ponašanja kao kriterij stanja dobrobiti ži-votinje zahtijeva definiciju "normalnog" ponašanja. S obzi-rom na to da se ponašanje kao fiziološka reakcija smatra sredstvom adaptacije, mijenja se ovisno o promjeni uvjeta okoliša. Različita ponašanja nastala kao adaptivne reakcije na promjene u okolišu treba smatrati "normalnim" (Hughes i Duncan, 1988.). Dobrobit je narušena kada su prijeđene granice, npr. izazov je izvan raspona reakcijske sposob-
nosti ptice. Bihevioralna zapažanja traju određeno vrijeme i zahtijevaju posebnu obuku osoblja. Velika je raznolikost u promjenama ponašanja ovisno o dobu dana, godišnjem dobu i nepredvidljivim podražajima koji se razlikuju među proizvodnim sustavima (Keeling, 1995.). Stoga je teško odrediti kvantitativni raspon "nor-malnog" ponašanja. Promatranje životinja u proizvodnim sustavima ne može pružiti pouzdane informacije koje se mogu primijeniti na svim postojećim sličnim sustavima.
Tablica 4. Kvantitativna procjena ponašanja kokoši nesilica smještenih kavezno na OPG-ima Brodsko-posavske županije (ljeto
2012. godine)
Ljestvica procijenjenih vrijednosti Oblici ponašanja nesilica Minimalna MaksimalnaAktivne Opuštene Plašljive Uzrujane Depresivne Mirne Zadovoljne Napete Nesigurne Energične Frustrirane Dosađuju se Prijateljski raspoložene Zaokupljene Nervozne Sretne Tužne Table 4. Quantitative evaluation of the behavior of laying hens housed in cages at family agricultural husbandries in Brod-
Posavina County (summer, 2012)
Scale of estimated values Hen behavior Minimum MaximumActive Relaxed Fearful Upset Depressed Calm Satisfied Strained Unsafe Vigorous Frustrated Bored Friendly Busy Nervous Happy Sad

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO 152
Kokoši nesilice smještene kavezno, na osnovi subjektivne procjene, tijekom promatranja iskazivale su slabu ak-tivnost, nisu bile energične niti zaokupljene, većinom su iskazale obrasce ponašanja plašljivosti i depresije. Vidljivo su se dosađivale i nisu bile prijateljski raspoložene niti
znatiželjne. Tijekom promatranja, uprkos neodgovarajućim uvjetima okoliša, nisu primijećena abnormalna ponašanja kao što su kljucanje, kanibalizam i agresija. Vučemilo i sur. (2008.) zabilježili su slične obrasce ponašanja kokoši nesilica.
Tablica 5. Procjena dobrobiti kokoši nesilica temeljem osnovnih načela na OPG-ima Brodsko-posavske županije (ljeto 2012.
godine)
Obiteljsko poljoprivredno gospodarstvo I. II. III. IV. V. VI. Načela Kriteriji
Da Ne Da Ne Da Ne Da Ne Da Ne Da Ne Odsustvo gladi x x x x x x
Hranidba Izostanak žeđi x x x x x x Komfor x x x x x x Ugodna toplina x x x x x x x x x x x
Držanje Mogućnost kretanja x x x x x x Izostanak ozljeda x x x x x x x x Izostanak bolesti x x x x x x x
Fizičko stanje Odsustvo boli x x x x x x x x x
Izrazi društvenog ponašanja x x x x x x Vrsti primjereno ponašanje x x x x x x Dobri odnosi čovjek-životinja x x x x x x x Ponašanje Pozitivno emocionalno ponašanje x x x x x x
I., II., III., IV., V. = kavezni način držanja; VI. = slobodni način držanja Table 5. Laying hen welfare assessment based on the basic principles at family agricultural husbandries in Brod-Posavina
County (summer, 2012)
Family agricultural husbandry I II III IV V VI Principle Criterion
Yes No Yes No Yes No Yes No Yes No Yes No Absence of hunger x x x x x x
Feeding Absence of thirst x x x x x x Comfort x x x x x x Pleasant warmth x x x x x x x x x x x
Housing Possibility of movement x x x x x x Absence of injury x x x x x x x x Absence of disease x x x x x x x
Physical condition Absence of pain x x x x x x x x x
Expressions of social behavior x x x x x x Species appropriate behavior x x x x x x Good man animal relations x x x x x x x Behavior Positive emotional behavior x x x x x x
I, II, III, IV, V = cage housing; VI = free housing

PROCJENA PONAŠANJA I DOBROBITI KOKOŠI NESILICA DRŽANIH U KONVENCIONALNIM KAVEZIMA I SLOBODNO
153
U kavezno držanih nesilica, kao i u slobodno držanih nesi-lica, hranidba i napajanje bili su zadovoljavajući te su nesilice stoga bile slobodne od gladi i žeđi (tablice 1. i 5.). Glede načina držanja komfor je bio zadovoljavajući kod slobodno držanih nesilica, dok je ugodna temperatura izo-stala kod oba načina držanja, ali su u vrijeme istraživanja (ljeto) vladale visoke vanjske temperature koje je u zatvo-renim sustavima bilo teško spustiti, čak i maksimalnim pro-zračivanjem (tablica 2.). Mogućnost kretanja bila je moguća samo kod slobodnog sustava. Slične vrijednosti zabilježili su Matković i sur. (2009.), Vučemilo i sur. (2010.). Ozljede i bol su izostale u većini promatranih jata osim kod nekolicine kavezno držanih jedinki u zadnjim danim iskorištavanja, koje su imale ozljede na vratu i prsima (tablica 3.). Slobodno držane jedinke bile su izložene stalnim napadima grabežlji-vaca. Ponašanje svojstveno vrsti iskazivale su samo slobod-no držane nesilice.
Zaključak
Prema dobivenim rezultatima provedenog istražavanja može se utvrditi da smještaj nesilica u kaveze nije odgovarajući s aspekta dobrobiti zbog nemogućnosti kretanja, čeprkanja, kupanja u prašini, šepurenja i zadovoljenja drugih etoloških potreba, ali je ostvaren zadovoljavajući higijenski standard, laka je kontrola nesilica, uz vrlo mali broj oštećenih i prlja-vih jaja. Hranjenje i napajanje bilo je zadovoljavajuće prema tehnološkim zahtjevima proizvodnje, kao i zdravstveno sta-nje jata.
Literatura
ANIĆ, V. (1998): Rječnik hrvatskog jezika. Novi Liber. Zagreb. BROOM, D. M. (1991): Animal welfare: concepts and measure-
ment. J. Anim. Sci. 16, 4167-4175. HORNBY, A. S. (1986): Oxford Advanced Learnerly Dictionary of
Current English. Oxford University Press i Cankarjeva založba. Ljubljana.
HUGHES, B. O., I. J. H. DUNCAN (1988): The notion of ethological ‘need’, models of motivation and animal welfare. Anim. Behav. 36, 1696-1707.
KEELING, L. J. (1995): Spacing behaviour and an ethological approach to assessing optimum space allocations for groups of laying hens. Appl. Anim. Behav. Sci. 44, 171-186.
LADEWIG, J. (2008): The role of veterinarian in animal welfare. Acta Vet. Scand. 50 (Suppl 1): S5.
MATKOVIĆ, K., M. VUČEMILO, B. VINKOVIĆ, M. BENIĆ (2009): Air quality and welfare of cage housed laying hens. Proceedings of the ISAH Congress 2009, 897-900.
ŠULEK, B. (1990): Hrvatsko-njemačko-talijanski rječnik znanstve-nog nazivlja. Reprint izd. 1874.-1875. Globus. Zagreb.
VUČEMILO, M. (2008): Higijena i bioekologija u peradarstvu. Veterinarski fakultet, Zagreb.
VUČEMILO, M., K. MATKOVIĆ, B. VINKOVIĆ, S. RADOVIĆ, M. BENIĆ (2008): Higijena, dobrobit i ponašanje nesilica smještenih u klasičnim kavezima i alternativnim sustavima držanja. Stočarstvo 6, 495-501.
VUČEMILO, M., B. VINKOVIĆ, K. MATKOVIĆ, I. ŠTO-KOVIĆ, S. JAKŠIĆ, S. RADOVIĆ, K. GRANIĆ, Đ. STUBIČAN (2010): The influence of housing systems on the air quality and bacterial eggshell contamination of table eggs. Czech J. Anim. Sci. 6, 243-249.
BEHAVIOR AND WELFARE ASSESSMENT OF LAYING HENS HOUSED IN CONVENTIONAL CAGES AND FREE
Summary
Most of the traditional animal production has changed significantly in the middle of the last century. This is particularly pronounced for laying hen housing. Small freely housed hen farming was replaced by battery-intensive systems in a few years. All of this encouraged discussion on physiological needs, feelings and rights of animals. The question is how to estimate normal behavior, how it is manifested and what is the share of behavior in assessing the overall state of welfare. As a starting point, some authors suggest it to be behavior that hens practice in natural environment. Therefore, when assessing their welfare, the extent should be thoroughly considered and established to which an individual hen in modern farming can act in accordance with its natural behavior. In addition, it is necessary to find answer to the question how the ability to manifest natural behavior ensures a high level of welfare. Besides, hens in modern housing systems can develop different patterns of behavior and thus provide sufficient welfare. Indicators of microclimate complex in objects were measured in selected husbandries and valuable data collected for assessing the welfare of laying hens, such as availability of food and water, technical conditions of the cages, ventilation facilities, techniques for slurry operations, etc. Then the elements of behavior and fitness of hens necessary to assess their welfare were analyzed. Results of the study revealed that accommodation of hens in cages is not appropriate in terms of welfare due to the impossibility of movement, tinkering, bathing in the dust, prancing and other ethologic needs, while achieving a satisfactory hygienic level, easy control of the hens, with a very small number of damaged and dirty eggs. Feeding and watering were satisfactory according to technological requirements of production, as well as the health status of the flock. Key words: behavior, welfare, laying hens, cage housing, free housing

PERADARSKI DANI 2013. 154
JAPANSKA PREPELICA (Coturnix coturnix japonica ) KAO MODEL U BIOMEDICINSKIM ISTRAŽIVANJIMA
Gordana Gregurić Gračner1, Mario Ostović1, Danijela Horvatek2, Damjan Gračner3, Željko Pavičić1
1Zavod za higijenu, ponašanje i dobrobit životinja,Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 2Zavod za bolesti peradi s klinikom, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 3Klinika za unutarnje bolesti, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska Sažetak
Prepelice se koriste kao modeli u brojnim područjima biomedicinskih istraživanja, pričem najranija izvješća o korištenju japanske prepelice kao laboratorijske životinje potječu iz druge polovice 20. stoljeća. Prema Pravilniku o zaštiti životinja koje se koriste u pokusima ili u druge znanstvene svrhe (NN RH br. 47/2011.) prepelica je također jedina ptica koja se navodi u popisu životinja koje se uzgajaju za korištenje u pokusima. U radu stoga predstavljamo biološke osobitosti prepelice, navodimo smjernice za smještaj i držanje te neke primjere njezina korištenja kao modela u biomedicinskim istraživanjima. Ključne riječi: japanska prepelica, laboratorijska životinja
Taksonomija i biološke osobitosti japanske prepelice
Prepelice (Coturnix) su rod ptica iz reda Galliformes (ko-koši), porodice Phasianidae. Rasprostranjene su širom svije-ta. Neke od danas živućih vrsta su europska obična prepelica (Coturnix coturnix), kineska prepelica (Coturnix chinensis), afričke prepelice (Coturnix adansonii i Coturnix dele-gorguei), australska prepelica (Coturnix pectoralis), austra-loazijska prepelica (Coturnix ypsilophora) te kišna prepelica (Coturnix coromandelica). Većina navedenih vrsta dijeli se u podvrste (Anonimno, 2013.). Tako je japanska prepelica (Coturnix coturnix japonica) jedna od podvrsta europske obične prepelice koja se spominje još u Starom zavjetu te prikazuje na staroegipatskim reljefima (Reese i Reese, 1962.). Japanska prepelica udomaćena je u Japanu potkraj 11. stoljeća, međutim, tek od 14. stoljeća počinje njezin uz-goj i to, unatoč grubom pjevu, kao ptice pjevice (Nemanič i Berić, 1995.). Nakon prvog svjetskog rata u japanskoj pokrajini Airhi počela je selekcija prepelica na nesivost, a danas u privredi Japana zauzimaju značajno mjesto, odmah iza uzgoja kokoši, i to zbog velikih proizvodnih mogućnosti i skromnih zahtjeva za smještajem i hranom. Sve se više uzgajaju i u SAD-u i Europi, pa tako i u Hrvatskoj.
Japanska prepelica spada među najmanje pripadnice reda kokoši. Mjereno od vrha kljuna do kraja repa dugačka je od 17 do 20 cm, od toga 3 cm otpada na zaobljeni rep. Raspon krila joj je oko 34 cm. Ima mali, slab kljun i kratke noge. Gornji dio tijela je smeđ, a sredina trbuha žućkasto bijela (Senčić, 1994.). Spolni dimorfizam uočava se u dobi od 3 tjedna i to na osnovi boje perja i uzorka na njima (Hawkins i sur., 2001.). Tjelesna masa odraslih mužjaka je oko 140 g, a ženki 135 g (Supić i sur., 2000.). Pilići su nakon valjenja teški 5 do 10 g (Senčić, 1994.). Spolnu zrelost dostiže s 5 do 6 tjedana, ovisno o trajanju svjetla. Uzgoj je najuspješniji kada skupinu sačinjavaju mužjak i dvije ženke (Hawkins i sur., 2001.). Prosječno godišnje daje 260 do 300 jaja prosječne mase 10,6 g. Inkubacija jaja traje 16-17 dana. Japanska prepelica izgubila je instinkt kvocanja pa se jaja inkubiraju u inku-batorima. U povoljnim uvjetima inkubiranja postotak va-ljenja prepeličjih pilića vrlo je visok, 87% do 95% (Senčić, 1994.). Za prepelicu je znakovit vrlo brzi rast. Već u trećem tjednu života poveća tjelesnu masu osam do deset puta. Kao najmanja kokoš po svojim je proizvodnim sposobnostima biološki fenomen (Nemanič i Berić, 1995.). Tijekom godine
Dr. sc. Gordana Gregurić Gračner, Zavod za higijenu, ponašanje i dobrobit životinja, Veterinarski fakultet Sveučilišta u Zagrebu, Heinzelova 55, 10 000 Zagreb, Hrvatska; Telefon: +385 1 2390 293; Fax: +385 1 2441 390; E-mail: [email protected]

JAPANSKA PREPELICA (Coturnix coturnix japonica) KAO MODEL U BIOMEDICINSKIM ISTRAŽIVANJIMA
155
dana može proizvesti 6 do 7 generacija, što ukazuje na nje-zinu veliku nesivost, plodnost i brzu reprodukcijsku moć (Vučemilo, 2008.). Ženke žive 2,5 do 3 godine, a mužjaci i do 5 godina (Ottinger, 1992.).
Smjernice za smještaj i držanje japanskih prepelica koje se koriste u pokusima ili u druge znanstvene svrhe
U Republici Hrvatskoj na snazi je Pravilnik o zaštiti živo-tinja koje se koriste u pokusima ili u druge znanstvene svrhe (NN RH br. 47/2011.). U Dodatku I. Pravilnika popisane su životinje koje se koriste u pokusima iz članka 21. Pravilnika. Jedina ptica u navedenom popisu upravo je prepelica. Prema članku 21. Pravilnika, životinje navedene u tome popisu namijenjene korištenju u pokusima moraju biti uzgojene životinje, osim u slučajevima kada Uprava za veterinarstvo odobri opće ili posebno izuzeće na način određen spo-menutim Pravilnikom. Prema Pravilniku, životinje koje se koriste za pokuse ili u druge znanstvene svrhe moraju biti primjereno njegovane, ne smije ih se nepotrebno izlagati boli, patnji, iscrpljenosti ili trajnom ozljeđivanju, a kad je to neizbježno, isto se mora svesti na najmanju moguću mjeru. Također, na najmanju moguću mjeru treba smanjiti bilo koje ograničenje njihovih fizioloških i etoloških potreba. Tako se i prepelicama mora osigurati primjeren smještaj, okoliš, mo-gućnost barem minimalnog kretanja, hrana, voda i njega odgovarajuća njihovom zdravlju i dobrobiti. Time se osi-gurava nesmetan tijek opita, odnosno izbjegavaju se mogući negativni utjecaji na njegove konačne rezultate. Skupine prepelica trebalo bi formirati prije postizanja spolne zrelosti kako bi se izbjegla pojava agresije. Tek bi se u iz-nimnim slučajevima prepelice trebale držati pojedinačno. Ptice treba sustavno nadgledati kako bi se na vrijeme uočila pojava socijalnog stresa te izbjegle daljnje posljedice istoga (Hawkins i sur., 2001.). Preporuča se držanje prepelica na otvorenom, u aviarijima, no pritom im treba osigurati zaklon od nepovoljnih vre-menskih prilika ili pak dodatni izvor topline tijekom hlad-nijeg razdoblja. Okoliš ptica u takvim nastambama mora biti što sličniji prirodnom, a preporuča se držanje osam ptica na 19 m2 avijarija. Kako je u takvim uvjetima zbog prisutnosti fecesa i vlažne zemlje rizik od obolijevanja ptica veći, nužan je pojačan nadzor nad njihovim zdravstvenim stanjem. Osobe koje skrbe o prepelicama moraju biti vrlo pažljive i nadasve vješte kako bi se, naročito prilikom hvatanja, iz-bjeglo ozljeđivanje ptica. Ako se pak prepelice za pokuse drže u zatvorenim nastambama preporuča se držanje do najviše 50 jedinki na m2 (Hawkins i sur., 2001.). Isto tako, prilikom smještaja prepelica u kaveze, unatoč ograničenom prostoru, treba voditi računa o mogućnosti zadovoljenja njihovih vrsno specifičnih potreba, pričem im je moguće obogatiti okoliš različitim predmetima poput kuglica, cijevi, boca, kocaka, ogledala i sl., a svakako im treba osigurati i materijal za „kupanje“ kao što su pijesak ili piljevina (Hawkins i sur., 2001.). Međutim, ne smije se koristiti piljevina od drveta obrađenog kemikalijama (NN RH br. 47/2011.).
Prepelicama je, također, potrebno osigurati mjesta za odmor, kutije za gniježđenje i materijale za izgradnju gnijezda. Nai-me, prepelice najradije nesu jaja na zaklonjenom mjestu, stoga im se u nastambama trebaju osigurati predmeti u svrhu zaklona kakvu im u prirodi pružaju biljke (Hawkins i sur., 2001.). Pri kaveznom držanju, prema Pravilniku, za prepelice mase 120 do 140 g najmanja podna površina iznosi 350 cm2 ako se u kavezu drži jedna ptica, 250 cm2 za dvije ptice, 200 cm2 za tri ili više ptica, uz 4 cm hranilice po ptici. Visina kaveza predstavlja vrijednost u cm okomitog razmaka između naj-više točke na podu i najniže točke na gornjem dijelu kaveza. Pravilnik propisuje najmanju visinu kaveza od 15 cm. Me-đutim, kako prepelice u prirodnom okolišu uzlijeću kratko i brzo, gotovo okomito, krov kaveza ne bi smio biti niži od 30 cm upravo zbog takvog karakterističnog uzlijetanja, jer u slučaju nižeg kaveza dolazi do ozljeđivanja glave koje nali-kuje ozljedama nastalim uslijed kanibalizma. Poklopci ka-veza moraju ptici pružati osjećaj zaštite, no isto tako i omo-gućavati dostatnu razinu osvjetljenja i u kavezima smješte-nim na nižim razinama u baterijama. U tu svrhu najbolje je stoga koristiti poklopce od fleksibilne plastične mreže (12x16 mm), jer ne uzrokuju ozljeđivanje glave, omogućuju dobru osvijetljenost donjih kaveza i bolje strujanje zraka u bateriji (Hawkins i sur., 2001.). Kavezi moraju biti izvedeni tako da se životinje ne mogu ozlijediti te izrađeni od materijala koji nisu štetni za njihovo zdravlje. Ako kavezi nisu za jednokratnu uporabu materijali moraju biti prihvatljivi s obzirom na održavanje higijenskih prilika (NN RH br. 47/2011.). Prepelicama treba osigurati smještaj u izoliranim, dobro pro-zračivanim prostorima, bez propuha i naglih promjena tem-perature. Prepelice su plašljive životinje (Vučemilo, 2008.), stoga nastambe i laboratoriji u kojima se provode pokusi moraju biti izolirani od buke. Prestrašene prepelice obično nesu mala jaja (Vučemilo, 2008.), što može poslužiti kao pokazatelj povišene razine stresa. Optimalni raspon vrijednosti temperature za prostorije u ko-jima se drže prepelice, prema Pravilniku, treba biti između 20 i 24 ºC, te mora biti osigurana precizna kontrola tempe-rature. Preporučena relativna vlaga zraka u nastambama za prepelice u laboratorijskim uvjetima je između 45% i 70% (Hinshaw i sur., 1969.). Sustav za prozračivanje treba osigurati 15 do 20 promjena zraka u jednome satu, međutim, ako se u nekom prostoru nalazi manji broj jedinki, može biti dovoljno i 8 do 10 promjena zraka, pričem je zabranjeno kruženje neobnovljenog zraka. Također, sustav za prozrači-vanje trebao bi biti dizajniran tako da, prema potrebi, može osigurati i grijanje i hlađenje zraka (NN RH br. 47/2011.). Prema Nemanič i Berić (1995.) za prepelice u uzgoju prepo-ručuje se 23 sata svjetla u prva dva tjedna, a zatim 14-18 sati, jačine 1 W/m2. Japanske prepelice su prilično otporne na bolesti (Supić i sur., 2000.), no novonabavljene životinje moraju se staviti u karantenu kako ne bi ugrozile životinje koje se već nalaze u nastambi, dok bolesne ili ozlijeđene životinje moraju biti smještene odvojeno uz prikladan smještaj, hranu i vodu za napajanje (NN RH br. 47/2011.).

JAPANSKA PREPELICA (Coturnix coturnix japonica) KAO MODEL U BIOMEDICINSKIM ISTRAŽIVANJIMA 156
Hrana za prepelice treba odgovarati hranidbenim potrebama svakog životnog stadija, a bilo bi dobro da je raznovrsna i ponuđena na različite načine, tako da zadovolji njihovu potrebu za traženjem hrane. To znači da bi hranu trebalo obogatiti voćem, sjemenkama, zrnjem, kukcima i sl. Također im na raspolaganju treba biti vapnenac ili ljušture kamenice. Voda im mora biti dostupna ad libitum (Hawkins i sur., 2001.) u posebnim kapljičnim pojilicama ili šalicama (Supić i sur., 2000.). Pritom Pravilnik propisuje da u slučaju kapljičnih pojilica ili šalica trebaju biti predviđene po dvije za pticu. Automatske sustave za napajanje treba redovito provjeravati, servisirati i ispirati kako bi se izbjegle nezgode i širenje zaraza, a za praćenje kakvoće vode potrebno je redovito ispitivanje sustava na bakteriološko zagađenje. Pri odabiru, proizvodnji i pripremi hrane za životinje treba izbjeći svako kemijsko, fizičko i mikrobiološko onečišćenje. Svi dozatori hrane za životinje ili posude u kojima je hrana ponuđena moraju biti redovito čišćeni i prema potrebi sterilizirani (NN RH br. 47/2011.).
Neki primjeri korištenja japanske prepelice kao modela u biomedicinskim istraživanjima
Znanstvenici su još godine 1962. ustanovili da je japansku prepelicu, za razliku od mnogih drugih laboratorijskih ži-votinja, moguće koristiti u istraživanjima u uvjetima koji su daleko od optimalnih, da su osobito pogodne za istraživanja u eksperimentalnoj psihologiji, da je nije teško pripitomiti, kao i da je nezajažljivog apetita (Reese i Reese, 1962.). Istraživanje Stattelmana i sur. (1975.) pokazalo je da je prepelice moguće dresirati nagrađujući ih brašnastim crvima kao hranom, no samo u slučaju ako su na njih navikle još kao pilići. Godine 1985. Ratnamohan je objavio pregledni rad o kori-štenju japanske prepelice u virusološkim istraživanjima. Takatsuji i sur. (1986.) su pak koristili albino prepelice kao životinjske modele u istraživanju etiologije i patogeneze glaukoma. U ženki japanskih prepelica, unatoč primjeni visokih doza testosterona, nije moguće potaknuti karakteristike spolnog ponašanja mužjaka. Uzrok tome je potpuno različita građa medijalne preoptičke jezgre koja ima značajnu ulogu u izra-žavanju spolnog ponašanja mužjaka, a navedene spoznaje primjenjive su u istraživanju centra u mozgu odgovornog za spolno ponašanje (Panzica i sur., 1998.; 2001.). U japanskih prepelica, isto tako, vrlo je definiran proces starenja s pri-mjetnim smanjenjem funkcije reproduktivnog, metaboličkog i osjetilnog sustava. Stoga se, primjerice, koriste kao model u istraživanju neuroendokrinih promjena vezanih uza sma-njenje funkcije reproduktivnog sustava, uključujući endo-krine i ponašajne komponente, s naročitim naglaskom na istraživanje u mužjaka (Ottinger, 2001.). Huss i sur. (2008.) navode da su japanske prepelice iznimno pogodne za istraživanja u razvojnoj biologiji, kao i da su zbog svoje fiziologije usporedive s ljudskom iznimno pogodne kao mo-del u istraživanjima procesa starenja te raznih bolesti. U istraživanjima u području molekularne biologije japanske prepelice predstavljaju idealan životinjski model za pro-
izvodnju transgeničnih ptica korištenjem lentiviralnih vek-tora (Poynter i sur., 2009.). Seredynski i sur. (2011.) su prepelice koristili u istraživanju specifičnih uloga dva podtipa estrogenog receptora bitnih u aktivaciji spolnog ponašanja mužjaka, jer je prethodno ustanovljeno da te receptore imaju mnogi kralježnjaci, uključujući i čovjeka. Korištene su, također, u komparativnoj studiji sposobnosti intuitivnog učenja (Zucca i sur., 2005.), a u istraživanju sposobnosti orijentacije u prostoru korištenjem prepoznat-ljivih obilježja postigle su isto tako odlične rezultate (Ruploh i sur., 2011.).
Umjesto zaključka
Na osnovi svega navedenog, razvidno je da je japanska prepelica, ptica skromnih životnih zahtjeva i iznimno prila-godljiva laboratorijskim uvjetima, fiziologije u mnogome kompatibilne ljudskoj, postala standardni i u mnogim bio-loškim i medicinskim istraživanjima nezamjenjivi životinj-ski model velikog potencijala.
Literatura
Anonimno (2013): Avibase – the world bird database. Link: http://avibase.bsc-eoc.org/species.jsp?lang=HR&avibaseid= =110CF4251A857B0D [01.02.2013]
HAWKINS, P., D. B. MORTON, D. CAMERON, I. CUTHILL, R. FRANCIS, R. FREIRE, A. GOSLER, S. HEALY, A. HUDSON, I. INGLIS, A. JONES, J. KIRKWOOD, M. LAWTON, P. MONAGHAN, C. SHERWIN, P. TOWN-SEND (2001): Laboratory birds: refinements in husbandry and procedures. Fifth report of BVAAWF/FRAME/RSPCA/ /UFAW Joint Working Group on Refinement. Lab. Anim. 35, 1-163.
HINSHAW, W. R., B. R. BURMESTER, A. A. CREAMER, C. W. HESS, J. R. HOWES, W. M. INSKO, W. O. WILSON (1969): Coturnix (Coturnix coturnix japonica): standards and guidelines for the breeding, care, and management of laboratory animals: a report. Subcommittee on Avian Standards. Institute of Laboratory Animal Resources. National Academy of Sciences, Washington, D. C. Link: http://books.google.hr/books?hl=hr&id=zmsrAAAAYAAJ&q= [13.01.2013]
HUSS, D., G. POYNTER, R. LANSFORD (2008): Japanese quail (Coturnix japonica) as a laboratory animal model. Lab. Anim. (NY) 37, 513-519.
Narodne novine (47/11): Pravilnik o zaštiti životinja koje se koriste u pokusima ili u druge znanstvene svrhe. Narodne novine.
NEMANIČ, J., Ž. BERIĆ (1995): Peradarstvo. Nakladni zavod Globus, Zagreb, Hrvatska.
OTTINGER, M. A. (1992): Neuroendocrine mechanisms regu-lating reproductive endocrinology and behavior during aging. Poult. Sci. Rev. 4, 235-248.
OTTINGER, M. A. (2001): Quail and other short-lived birds. Exp. Gerontol. 36, 859-868.
PANZICA, G. C., C. CASTAGNA, C. VIGLIETTI-PANZICA, C. RUSSO, O. TLEMÇANI, J. BALTHAZART (1998): Organi-zational effects of estrogens on brain vasotocin and sexual behavior in quail. J. Neurobiol. 37, 684-699.

JAPANSKA PREPELICA (Coturnix coturnix japonica) KAO MODEL U BIOMEDICINSKIM ISTRAŽIVANJIMA
157
PANZICA, G., C. VIGLIETTI-PANZICA, J. BALTHAZART (2001): Sexual dimorphism in the neuronal circuits of the quail preoptic and limbic regions. Microsc. Res. Tech. 15, 364-374.
POYNTER, G., D. HUSS, R. LANSFORD (2009): Japanese Quail: An Efficient Animal Model for the Production of Transgenic Avians. Cold Spring Harb. Protoc., doi:10.1101/pdb.emo112
RATNAMOHAN, N. (1985): The management of Japanese quail and their use in virological research. Vet. Res. Commun. 9, 1-14.
REESE, E. P., T. W. REESE (1962): The quail, Coturnix coturnix, as a laboratory animal. J. Exp. Anal. Behav. 5, 265-270.
RUPLOH, T., A. KAZEK, H. J. BISCHOF (2011): Spatial orientation in Japanese quails (Coturnix coturnix japonica). Link: http://www.ncbi.nlm.nih.gov/pmc/articles/ /PMC3233574/ [13.01.2013]
SENČIĆ, Đ. (1994): Peradarstvo. Gospodarski list d.d., Zagreb, Hrvatska.
SEREDYNSKI, A. L., G. F. BALL, J. BALTHAZART, T. D. CHARLIER (2011): Specific activation of estrogen receptor alpha and beta enhances male sexual behavior and neuro-
plasticity in male Japanese quail. Link: http://www.plos-one.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0018627 [31.01.2013]
STATTELMAN, A. J., R. T. TALBOT, D. B. COULTER (1975): Olfactory thresholds of pigeons (Columba livia), quail (Colinus virginianus) and chickens (Gallus gallus). Comp. Biochem. Phys. A 50, 807-809.
SUPIĆ, B., N. MILOŠEVIĆ, T. ČOBIĆ (2000): Živinarstvo. Poljoprivredni fakultet Univerziteta u Novom Sadu, Novi Sad, Srbija.
TAKATSUJI, K., Y. SATO, S. IIZUKA, H. NAKATANI, A. NAKAMURA (1986): Animal model of closed angle glaucoma in albino mutant quails. Invest. Ophthalmol. Vis. Sci. 27, 396-400.
VUČEMILO, M. (2008): Higijena i bioekologija u peradarstvu. Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska.
ZUCCA, P., F. ANTONELLI, G. VALLORTIGARA (2005): Detour behaviour in three species of birds: quails (Coturnix sp.), herring gulls (Larus cachinnans) and canaries (Serinus canaria). Anim. Cogn. 8, 122-128.
JAPANESE QUAIL (Coturnix coturnix japonica) AS A MODEL IN BIOMEDICAL RESEARCH Summary Quail has been used as a model in many areas of biomedical research, with the earliest reports on using Japanese quail as laboratory animals dating from the second half of the 20th century. According to the Regulations on the Protection of Animals Used for Experimental and Other Scientific Purposes (Official Gazette No. 47/2011), quail is the only bird included in the list of animals reared for use in experiments. Therefore, biological properties of Japanese quail, guidelines for quail housing and keeping, and examples of its use as a model in biomedical research are presented. Key words: Japanese quail, laboratory animal

PERADARSKI DANI 2013. 158
UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
Marina Tišljar1, Željko Grabarević2, Branka Artuković2, Jadranka Foršek3, Petar Džaja4, Marija Dinarina-Sablić5, Tajana Amšel Zelenika1
1Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska 2Zavod za veterinarsku patologiju, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 3Klinika za unutarnje bolesti, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 4Zavod za sudsko i upravno veterinarstvo, Veterinarski fakultet Sveučilišta u Zagrebu, Zagreb, Hrvatska 5Ireks Aroma, d.o.o., Zagreb, Hrvatska Sažetak Na temelju rezultata kliničke, patomorfološke i hematološke pretrage te indeksa mase organa određivan je učinak akutne toksičnosti N-ω-nitro-L-arginin-metil-estera (L-NAME), neselektivnog inhibitora i L-arginina, prekursora dušik (II) oksida (DO) na pojavu ascitesa – sindroma plućne hipertenzije u 25 tovnih pilića muškoga spola (AVIAN 34; dob: 30 dana) nakon jednokratne intraperitonejske primjene 50 mg/kg tj. m., 100 mg/kg tj. m. i 150 mg/kg tj. m. L-NAME-a (LN50, LN100 i LN150), 150 mg/kg tj. m. L-NAME-a i 100 mg/kg tj. m. L-arginina (LN150/LA) i 0,5 mL/kg tj.m. fiziološke otopine (0,9% NaCl) (K, kontrolna skupina). Prvi klinički znakovi karakteristični za hipoksiju očitovali su se šest sati nakon primjene najveće koncentracije L-NAME (150 mg/kg tj. m.) i kao slabije izraženi znakovi hipoksije u skupinama tretiranim manjim koncentracijama L-NAME (50 mg/kg tj. m. i 100 mg/kg tj. m.). Makroskopskom i mikroskopskom patomorfološkom pretragom utvrđene su promjene karakteristične za predascitični stadij. Ključne riječi: dušik (II) oksid, L-arginin, L-NAME, tovni pilići, ascites – sindrom plućne hipertenzije Uvod
Ascites, sindrom plućne hipertenzije (SPH), prepoznat kao sindrom plućne arterijske hipertenzije (PAH) (Wideman i sur., 2013.) te izrazito nalik idiopatskoj arterijskoj hiper-tenziji u ljudi (idiopathic pulmonary arterial hypertension, IPAH), donedavno poznatoj pod nazivom primarne plućne hipertenzije (PPH) (Wideman i Hamal, 2011.), metaboličko je stanje brzorastućih tovnih pilića koje nastaje postupnim poremećajem rada srčanožilnog sustava s posljedičnim pa-tološkim promjenama tipičnima za hipoksični organizam. U brojnim su znanstvenim preglednim radovima obrađeni pri-kupljeni literaturni podatci o utjecajima tehnologije hranidbe i uzgoja, okoliša i nasljeđa na patogenezu PAH, s posebnim naglaskom na hipoksiju kao neposredan uzrok ove bolesti (Lopez-Coello i sur., 1986.; Hernandez, 1987.; Bottje and Wideman, 1995; Dewill i sur., 1996.; Grabarević i sur.,
1997.; Shlosberg et al., 1998.; Currie, 1999; Wideman, 2000., 2001.; Julian, 2000., 2007.; DeCuypere i sur., 2000; Tankson i sur., 2001, 2002; Tišljar, 2004; Tišljar i sur., 2005, 2011; Wideman i sur., 2004, 2007; Olkowski, 2007.; Pavlidis i sur., 2007.; Baghbanzadeh and DeCuypere, 2008.). Najučestalija u tovnih pilića u dobi od 4-5 tjedana, ova se bolest očituje s prosječnom stopom smrtnosti od 5%-8% (sporadično i do 30%) (Balog, 2003.; De Smit i sur., 2005.; Julian, 2005.; Pavlidis i sur., 2007.). Već 4%-tna smrtnost tovnih pilića uzrokovana ascitesom-SPH donosi godišnji gubitak od milijun dolara u svjetskoj peradarskoj pro-izvodnji (Maxwell i Robertson, 1997.). Uz pojačanu transudaciju u tjelesne šupljine s krajnjim ishodom nastanka ascitesa i uginućem (Broz i Ward, 2007.) SPH se u pre-živjelih pilića očituje znatnim smanjenjem tjelesne mišićne mase, što predstavlja izrazito velik ekonomski problem za
Dr. sc. Marina Tišljar, dr. vet. med., Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska; tel.: ++385 (0)1 2441 392; fax: ++385 (0)1 2441 396, e-mail: [email protected]

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
159
svjetsku peradarsku proizvodnju (Rajani i sur., 2011.; Veldkamp i sur., 2000.). U razdoblju od 1991. do 2012. godine u prikazu strukture smrtnosti zaraznih i nezaraznih bolesti domaće peradi intenzivnog uzgoja u Republici Hrvatskoj (neobjavljeni podatci, Tišljar, 2013.) prosječna smrtnost tovnih pilića uginulih od posljedica ascitesa-SPH iznosi 13%. Dosadašnji pokušaji razjašnjavanja povećane rezistencije u krvnim žilama pluća tovnih pilića sada se uglavnom usre-dotočuju na određivanje uloge kemijskih medijatora vazo-konstrikcije i vazodilatacije, kao i na patološke (strukturne) promjene unutar plućnih aretriola (primjerice, patologija i remodeliranje krvnih žila) tijekom nastanka plućne arterijske hipertenzije (PAH) (Wideman i sur., 2013.). Naime, u brzorastućih tovnih pilića sve su raspoložive krvne žile pre-težito krutih, nedovoljno rastezljivih stijenki, smanjene spo-sobnosti za preuzimanje dodatnog volumena krvi i najčešće krcate krvlju tako da plućno krvožilje ima smanjen kapacitet pohrane krvi čak i u besprijekornim uvjetima (posljedica otpora na protok krvi kroz pluća koji je prisutan već u u predkapilarnim arteriolama) (Wideman i Kirby, 1995.; Wideman i sur., 2007.; Martinez-Lemus i sur., 1999.; Wideman, 2000., 2001.; Chapman and Wideman, 2001.; Odom i sur., 2004.; Khajali i sur., 2011.). Jedan od izrazito jakih i učinkovitih plućnih vazodilatatora, dušik (II) oksid (DO), sintetiziran je iz L-arginina posre-dovanjem sintaza DO-a (SOD) (Bowen i sur., 2006.). Ovaj spoj smanjuje otpor protoku krvi u plućima sisavaca opu-štanjem glatkog mišićja krvnih žila, oblikovanjem otpušta-nja vazokonstriktora pomoću aktiviranih leukocita, desen-zibilizacijom endotelnog odgovora na vazokonstriktore i inhibiranjem proliferacije glatkog mišićja (muskularizacije) u predkapilarnim arteriolama (Gaston and Stamler, 1997.; Arnal i sur., 1999.; Albrecht i sur., 2003.). Na isti način djeluje i u plućima kokoši (pilića): smanjuje otpor u krvnim žilama dilatacijom stijenke i njihovu osjetljivost na vazo-konstriktorski učinak spojeva kao što su, primjerice, endo-telin-1 i tromboksan A2 (Grabarević i sur.,1997.; Wideman i sur., 1998, 2004., 2005.; Martinez-Lemus i sur., 2003.; Villamor i sur., 2002.; Wang i sur., 2002.; Weidong i sur., 2002.; Moreno de Sandino and Hernandez, 2003.; Odom i sur., 2004.; Tišljar, 2004.; Tišljar i sur., 2005., 2011.). Vazokonstrikciju pobuđenu hipoksijom i pojavu PH u tovnih pilića izloženih tijekom duljeg vremena hladnoći mo-guće je uspješno spriječiti dodavanjem L-arginina (prekur-sora DO-a i inhibitora SPH) u hranu (Wilson i sur., 1988.; Khajali i Wideman, 2010.). U potpunosti je suprotan učinak dugotrajnog pokusnog tretiranja tovnih pilića neselektivnim inhibitorom DO-a L-NAME (ponovljeno intraperitonejsko injiciranje tijekom 5 tjedana; 10 mg/kg tj.m.) (Grabarević i sur. 1997.; Tišljar, 2004.; Tišljar i sur., 2005., 2011.) ili dodavanja L-NAME-a (100 ppm tijekom 4 tjedna) u hranu pilića (Wang i sur., 2002.), uslijed čega postupno dolazi do pojave predascitičnog stadija, ascitesa-SPH i uginuća. U dosadašnjim su brojnim radovima o ulozi DO-a u na-stanku ascitesa-SPH u tovnih pilića poglavito obrađivani
rezultati istraživanja učinka njegovih prekursora, L-arginina i/ili vazokonstriktora i neselektivnog inhibitora DO-a L-NAME kao i enzima koji nadziru njegovu tvorbu, sintaza DO-a (SOD), tijekom duljeg razdoblja njihova djelovanja u organizmu pilića. Budući da L-NAME u manjim koncentracijama (npr. 10 mg/kg tj. m.) primjenjivanima tijekom duljeg razdoblja dokazano uzrokuje nastanak ascitesa-SPH, dok istodobno prekursor DO-a L-arginin smanjuje pojavnost ove nezarazne metaboličke bolesti srčanožilnoga sustava i/ili ljekovito djeluje na organizam već zahvaćen bolešću (istraživanje učinka kronične toksičnosti L-arginina i L-NAME na pojavu ascitesa-SPH u tovnih pilića (Grabarević i sur., 1997.; Wang i sur., 2002.; Wideman i sur., 2004.; Odom i sur., 2004.; Tišljar, 2004.; Tišljar i sur., 2005., 2011.; Tan i sur., 2007.), predmetno se je istraživanje usmjerilo na istraživanje učinka akutne toksičnosti jednokratnih većih doza L-NAME-a (50 mg/kg tj. m.; 100 mg/kg tj.m. i 150 mg/kg tj.m.) i L-arginina (100 mg/kg tj. m.) na pojavu ascitesa-SPH u tovnih pilića.
Materijal i metode
Dvadeset i pet tridesetdnevnih pilića muškoga spola (AVIAN 34) odabrano je za jednokratni jednodnevni pokus u kojemu je intraperitonejski (i.p.) apliciran L-NAME u različitim koncentracijma (50 mg/kg tj. m. (LN50), 100 mg/kg tj. m. (LN100) i 150 mg/kg tj. m. (LN150)); istodobno je aplicirano 100 mg/kg tj. m. L-arginina (LA) i 150 mg/kg tj. m. (LN150) (LA/LN150) te fiziološka otopina (f.o.) 0,9% NaCl (0,5 mL/kg tj. m.) (K)). Rabljene kemikalije
U svim su pokusima rabljene slijedeće kemikalije: 1. L-arginin (prekursor DO-a) (ICN Biomedicals, Cleveland, SAD); 2. L-NAME (N-ω-nitro-L-arginin metil-ester HCl/neselek-tivni inhibitor DO-a/) (ICN Biomedicals, Cleveland, SAD) 3. Fiziološka otopina (f.o.), 0,9% NaCl. Protokol pokusa Skupine od pet tovnih pilića muškoga spola (AVIAN 34) (dob: 30 dana) jednokratno su dobile L-NAME u dozama od 50 mg/kg tj. m. (LN50), 100 mg/kg tj. m. (LN100) i 150 mg/kg tj. m. (LN150) te istodobno 150 mg/kg tj. m. (LN150) i 100 mg/kg tj. m. L-arginina (LA) (LA/LN150) i fiziološku otopinu 0,9% NaCl (0,5 mL/kg tj. m.). Šest sati nakon primjene pilići su žrtvovani cervikalnom dislokacijom nakon omamljivanja kloroformom. K (5 pilića) – pilići primili 0,5 mL/kg tj m. f.o. (0,9% NaCl) L-NAME (LN50) (5 pilića) – pilići primili L-NAME u dozi 50 mg/kg tj. m. L-NAME (LN100) (5 pilića) – pilići primili L-NAME u dozi 100 mg/kg tj. m.

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
160
L-NAME (LN150) (5 pilića) – pilići primili L-NAME u dozi 150 mg/kg tj m. L-NAME (LN150) i L-arginin (LN150/LA) – pilići primili L-NAME u dozi 150 mg/kg tj. m. i L-arginin u dozi 100 mg/kg tj. m. Nakon šestosatnog promatranja vladanja pilića tijekom ko-jega su im uskraćene hrana i voda za piće pilići su izvagani i žrtvovani. Dobiveni uzorci krvi proslijeđeni su na hema-tološku pretragu. Tijekom razudbe izvagani su parenhimski (srce i jetra) i imunokompetentni organi (Fabricijeva burza, timus i slezena) te pluća, nakon čega je određivan indeks njihovih masa. Navedeni su organi (srce, pluća, slezena i jetra) fiksirani u 10%-tnom neutralnom formalinu te pod-vrgnuti patohistološkoj pretrazi. Patohistološka pretraga Nakon višednevne fiksacije organa u 10%-tnom neutralnom formalinu uzorci tkiva su uklopljeni u parafin i rezani na kliznom mikrotomu (MICROM HM 325, Zeiss, Austrija) na rezove debljine 4 μm. Deparafinirani su preparati bojani hematoksilinom i eozinom (HE) i pretraženi svjetlosnim mikroskopom (LEICA DMLB, Njemačka) te slikani digitalnim fotoaparatom PIXERA Pro 150ES. Hematološka pretraga Ukupan broj eritrocita i leukocita te vrijednosti hemoglobina i hematokrita određeni su na aparatu COULTER Counter (Engleska), dok je diferencijalna krvna slika određena iz krvnih razmaza bojanih po May-Grünwald-Giemsi (MGG). Indeks mase organa Indeksi mase jetre, srca, slezene i pluća utvrđeni su izraču-navanjem odnosa mase organa i tjelesne mase te množenjem dobivenog rezultata s tisuću.
Statistička obrada rezultata Rezultati hematološke pretrage te indeks mase organa obrađivani su po Mann-Whitney-U testu za dvije nezavisne skupine (Ivanković i sur., 1988.).
Rezultati
Klinička slika
Prvi klinički znakovi hipoksije utvrđeni su u pokusnih pilića šest sati nakon početka aplikacije. Najveći stupanj cijanoze sluznice, kože i mišićja glave i vrata te kože i mišićja prsnoga područja bio je vidljiv u skupini tretiranoj najvećom koncentracijom L-NAME (150 mg/kg tj. m.) (LN150) i u skupini pilića kojima je istodobno aplicirano 150 mg/kg tj. m. L-NAME i 100 mg/kg tj. m. L-arginina (LA/LN150). Slabije izražena cijanoza kože i mišićja prsnoga područja uočena je u pilića ostalih pokusnih skupina (LN100 i LN50). U pojedinih je pilića zapaženo i otežano disanje (LN150/LA; LN150).
Patoanatomski nalaz
Najveći stupanj cijanoze mekih tkiva (sluznice, kože i mišićja) utvrđen je u području glave, vrata te u koži i mišićju prsnoga područja pilića iz skupina LN150 i LA/LN150. Slabije izražen serozni edem potkožja u prsnome području, hidroperikard, umjerena dilatacija desne klijetke srca i povećana transudacija peritonejske tekućine primijećeni su u pilića skupine LN150, LN150/LA i LN100. U istih je skupina uočen plućni edem i punokrvnost jetre. U pilića kontrolne skupine nije bilo vidljivih patoanatomskih promjena.
Patohistološki nalaz
Izrazite su promjene zastupljene u srcima pilića koji su primili najveću koncentraciju L-NAME (tablica 1.).
Tablica 1. Patohistološke promjene organa pilića koji su jednokratno dobili fiziološku otopinu (0,9% NaCl - 0,5 mg/kg tj. m.;
skupina K); 50 mg/kg tj. m. L-NAME, 100 mg/kg tj. m. L-NAME i 150 mg/kg tj. m. L-NAME (LN50, LN100 i LN150) te istodobno 150 mg/kg tj. m. L-NAME i 100 mg/kg tj. m. L-arginin (LN150/LA)
Skupina Slezena Jetra Srce Pluća
K B. o. B. o. Uočljiv p.v. edem i umjerena punokrvnost Umjerena punokrvnost
LN50 Nekroza stanica tipa limfocita
Uočljiva punokrvnost Hiperplazija mononuklearnih stanica i umjeren p.v. edem
Umjereni p.v. edem, ostalo kao u skupini K
LN100 Nekroza stanica tipa limfocita
Punkrvnost i uočljiv intersticijski edem
Izrazita punokrvnost Izrazit p.v. i subendokardijski edem Miokardioliza
Vidljiv p.v. edem, ostalo kao u skupini K
LN150 Punokrvnost i nekroza stanica tipa limfocita Uočljiva punokrvnost P.v. i i.f. edem Miokardioliza Izrazit p.v. edem, ostalo kao u
skupini K
LA/LN150 B.o. Uočljiv intersticijski edem Nalaz kao u LN100 (bez nalaza miokardiolize)
Izrazito krvarenje i p.v. edem, ostalo kao u skupini K
p.v. = perivaskularni; i.f. = interfibrilarni

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
161
Table 1. Histopathologic changes in broilers intraperitoneally (ip) treated with a single dose of saline (0.90 % w/v NaCl; 0,5 mL/kg BW – control group /K/; L-NAME (doses 50, 100, 150 mg/kg BW /LN50-, LN100- and LN150- groups) and L-arginine (100 mg/kg BW) in combination with L-NAME (150 mg/kg BW) (LN150/LA- group)
Group Spleen Liver Heart Lungs
K No changes No changes Noticeable pv edema and hyperemia Noticeable hyperemia
LN50 Necrosis of lymphoid cells Noticeable hyperemia Mononuclear cell hyperplasia
Noticeable pv edema Noticeable pv edema and hyperemia
LN100 Necrosis of lymphoid cells
Noticeable hyperemia and edema
Distinct hyperemia and pv and subendocardial edema Myocardiolysis
Distinct pv edema Noticeable hyperemia
LN150 Hyperemia and necrosis of lymphoid cells Noticeable hyperemia Pv and if edema Myocardiolysis Distinct pv edema Noticeable
hyperemia
LA/LN150 No changes Noticeable interstitial edema Pv and if edema Distinct hemorrhages and pv
edema Noticeable hyperemia
pv = perivascular; if = interfibrillar Patohistološki nalaz u kontrolnoj skupini pilića ukazuje na peri-vaskularni edem i umjerenu punokrvnost u srcu i plućima. U sle-zeni pilića iz skupina koje su primile 50 mg/kg tj. m., 100 mg/kg tj. m. i 150 mg/kg tj. m. L-NAME prevladava nekroza stanica tipa limfocita u tipu apoptoze. Prevladavajuća je pojava punokrvnosti i edema u jetri pilića tretiranih s L-NAME. U srcu pilića svih pokusnih skupina očit je perivaskularni i/ili interfibrilarni edem, a u skupinama koje su dobile 100 mg/kg tj. m. L-NAME i 150 mg/kg tj. m. L-NAME utvrđena je i miokardioliza.
Rezultati hematološke pretrage
Statistički znakovito odstupanje od prosječne sred-nje vrijednosti hematoloških parametara u pokusu istraživanja akutne toksičnosti agonista i antagoni-sta DO-a (P<0,05; Mann-Whitney-U test) utvrđeno je za vrijednost broja leukocita (slika 1.) i hemo-globina (slika 2.).
Slika 1. Srednje vrijednosti broja leu-kocita u pokusu istraživanjaučinka akutne toksičnosti L-ar-ginina i L-NAME na pojavuascitesa-SPH u tovnih pilića
Figure 1. Mean values of total leukocytecount in the experiment inves-tigating the impact of L-NAME and L-arginine acute toxicity induced lesions on ascites-PHS development in broiler chickens

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
162
Slika 2. Srednje vrijednosti hemo-globina u pokusu istraživa-nja učinka akutne toksično-sti L-arginina i L-NAME na pojavu ascitesa-SPH u tovnih pilića
Figure 2. Mean hemoglobin values inthe experiment investigatingthe impact of L-NAME and L-arginine acute toxicity in-duced lesions on ascites-PHS development in broilerchickens
Slika 3. Prosječne srednje vrijednostiindeksa mase jetre u pokusuistraživanja učinka akutnetosičnosti L-NAME i L-argi-nina na pojavu ascitesa –SPH u tovnih pilića
Figure 3. Mean mass index zvalues ofthe liver in the experiment ofthe investigation of the im-pact of L-NAME and L-argi-nine acute toxicity inducedlesions on ascites – PHS development in broilerchickens

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
163
Znatna su odstupanja u vrijednostima broja leukocita u sku-pini LN100 (32,67x109/L) i LN150/LA (24,99x109/L) u usporedbi s kontrolnom skupinom (44,56x109/L): u objema su pokusnim skupinama znatno manje vrijednosti od onih u kontrolnoj. Znatno veće vrijednosti utvrđene su u skupini LN150 (43,57x109/L) u usporedbi sa skupinom LA/LN150. Prosječne vrijednosti broja leukocita veće su u skupinama LN100 (32,67x109/L) i LN50 (35,81x109/L) od istih u skupini LA/LN150. Vrijednosti broja leukocita znatno su veće u skupini LN150 od onih u skupini pilića tretiranih s LN100. Znatno su manje vrijednosti hemoglobina u skupinama LA/LN150 (94x10g/L), LN100 (90x10g/L) i LN50 (90x10g/L) u usporedbi s kontrolnom skupinom (106x10g/L). Najveće vrijednosti pokazuje kontrolna skupina, dakako veće i od skupine LN150 (98x10g/L), iako ova razlika nije statistički značajna. Veće su vrijednosti u skupini LN150 od onih u skupinama koje su dobile 50 i 100 mg/kg tj. m. L-NAME i skupini LA/LN150. Ni u ovom slučaju razlika između srednjih vrijednosti nije statistički značajna. Indeks mase organa Znatno su manje vrijednosti u skupini LN150/LA (26,90) u usporedbi s kontrolnom skupinom (30,81). Kontrolna sku-pina pokazuje daleko najveću vrijednost indeksa mase jetre od svih skupina u ovome pokusu, no razlike između kon-trolne i preostalih skupina (osim LN150/LA) nisu statistički značajne.
Rasprava
Istraživanje djelovanja različitih koncentracija neselektivnog inhibitora DO-a - L-NAME (50 mg/kg tj. m., 100 mg/kg tj. m. i 150 mg/kg tj. m.) i prekursora DO-a L-arginina (100 mg/kg tj.m.) jednokratno apliciranih pilićima u dobi od 30 dana provedeno je sa svrhom utvrđivanja učinka akutne tok-sičnosti ovih spojeva na pojavu ascitesa-SPH u tovnih pilića. Prvi su rezultati kliničkog promatranja polučeni šest sati nakon primjene. U pilića kojima je injicirano 150 mg/kg tj.m. LN (LN150) i u kombinaciji iste koncentracije L-NAME s L-argininom (100 mg/kg tj.m.) primijećen je najveći stupanj cijanoze sluznica glave i vrata te potkožja i kože glave, vrata i prsnog područja. Manji stupanj navedenih promjena uočen je u skupini koja je dobila 100 mg/kg tj.m LN (LN100) i 50 mg/kg tj.m. LN (LN50). Razudbom je utvrđena serozna provlaženost potkožja u prsnome području, hidroperikard i povećan stupanj provla-ženosti prsnotrbušne šupljine (LN150, LN100). Smanjenje kon-centracije DO-a u krvnoj plazmi s posljedičnim porastom plućnog arterijskog tlaka i nastankom SPH utvrđeni su već jedan sat nakon intravenske primjene L-NAME tovnim pilićima (Wang i sur., 2002.). U pilića u predascitičnom stanju uočljiva je cijanoza, dje-lomice anemija prsnoga mišićja, pojačana provlaženost prsnotrbušne šupljine, umjerena dilatacija desne klijetke srca ponekad praćena hidroperikardom, umjereni edem pluća te hiperplazija jetre (Crespo i Shivaprasad, 2003.). Hidro-perikard, kao i punokrvnost i edem pluća i jetre utvrđeni su
u većine pokusnih pilića šest sati nakon primjene L-NAME (v. Klinička slika i Patološkoanatomski nalaz). U prilog rezultatima našega kliničkog pregleda, a djelomice i razud-benom nalazu dobivenom u izrazito kratkom razdoblju od početka primjene istraživanih tvari govore i podatci Juliana i sur. (1987.; 1989. a, b) i Juliana (1990.) glede pojave uginuća pilića oboljelih od PH bez prethodno vidljivih kliničkih znakova bolesti (ascitesa). Većina istraživača ascitesa-SPH naglašava sličnost patomorfološkog nalaza pilića zahvaćenih hipoksijom s nalazom u ascitičnih pilića (Hall i Machicao, 1968.; Maxwell i sur., 1986. a, b). U pilića tretiranih s L-NAME utvrđena je u slezeni nekroza stanica tipa limfocita u tipu apoptoze. Ovaj se nalaz može pripisati tropizmu L-NAME limfoidnom tkivu slezene ili pak akutnom toksičnom učinku velikih koncentracija L-NAME-a (Grabarević i sur., 1997.; Tišljar, 2004.; Tišljar i sur., 2005., 2011.). Sličan je nalaz u slezeni i tijekom infekcije virusom zarazne bolesti Fabricijeve burze. No, budući da patomorfološkom i patohistološkom pretragom nisu utvrđene promjene u burzi, a ni klinički znakovi nisu ukazivali na bolesno stanje životinja prije aplikacije, virus nije izdvajan. U jetri je prevladavala primjetna punokrvnost u pilića skupina LN50, LN100 i LN150. U skupinama LN100 i LN150/LA utvrđen je i intersticijski edem. Očito je da jednokratna primjena L-NAME-a u najvećoj koncentraciji (150 mg/kg tj. m.) izaziva opsežne promjene u stijenci krvnih žila tako što sprječava proizvodnju DO-a i izaziva povećanu propusnost krvnih žila s pratećim edemom. Opsežna krvarenja i izrazit perivaskularni edem utvrđeni su u plućima pileta koje je istodobno dobilo L-arginin i LN150. Sličan je nalaz utvrđen u plućima pilića koji su L-arginin primili neposredno pred kraj pokusnoga razdoblja (Graba-rević i sur., 1997.; Tišljar, 2004.; Tišljar i sur., 2005.; 2011., istraživanje učinka kronične toksičnosti L-NAME-a i L-arginina na pojavu ascitesa-SPH u tovnih pilića /ponavljano intraperitonejsko injiciranje L-NAME-a 10 mg/kg tj. m. i L-arginina 100 mg/kg tj. m. tijekom prvih pet tjedana života). Nalaz perivaskularnog edema i miokardiolize u pilića koji su primili 100 mg/kg tj. m. L-NAME-a odgovarao bi nalazu u srcu pilića koji L-NAME primaju svaki drugi dan tijekom četiri pokusna tjedna (10 mg/kg tj. m.) (Grabarević i sur., 1997.; Tišljar, 2004.; Tišljar i sur., 2005.; 2011.). Izraziti se kardiotropizam u pilića tretiranih neselektivnim inhibitorom DO-a L-NAME-om očitovao nekrozom miofibrila i poja-vom subendokardijskoga edema četiri tjedna nakon sustavne primjene L-NAME u koncentraciji od 10 mg/kg tj. m. (Grabarević i sur., 1997.; Tišljar, 2004.; Tišljar i sur., 2005.; 2011.) i šest sati nakon primjene daleko većih koncentracija L-NAME-a (100 mg/kg tj. m. i 150 mg/kg tj. m. istodobno s 100 mg/kg tj. m. L-arginina). Ponovno, ali ovoga puta na temelju rezultata istraživanja učinka akutne toksičnosti L-NAME-a i L-arginina, dokazan je i sustavni toksični učinak L-NAME-a. Dok je u pokusu istraživanja kronične toksičnosti taj učinak L-NAME-a ovisio o duljini razdoblja tijekom kojega je spoj apliciran pilićima (10 mg/kg tj. m. svaki drugi dan tijekom prvih pet tjedana života) (Grabarević i sur., 1997.; Tišljar, 2004.;

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
164
Tišljar i sur., 2005.; 2011.), u ovome je istraživanju izravno ovisio o koncentraciji L-NAME-a. Statistički značajno manji broj leukocita u skupini pilića tretiranoj L-argininom u usporedbi s brojem leukocita u kontrolnoj i u skupinama koje su dobivale isključivo L-NAME (50, 100 i 150 mg/kg tj. m.) može upućivati na "zaštitni" učinak jednokratno primijenjenog prekursora DO-a glede nastupa hipoksije, odnosno predascitičnog stanja s posljedičnim povećanjem broja leukocita u skupinama tretiranim neselektivnim inhibitorom DO-a L-NAME (slika 1). Dušik (II) oksid, prepoznat kao ključni zaštitni modulator tijekom oštećenja koje nastaje posredstvom leukocita i tromboze u plućnom krvotoku sisavaca, vjerojatno je moć-niji primarni intrapulmonarni modulator štetnog učinka tromboksana (TxA2) i PAF-a (engl. platelet activating factor) od prostaciklina (PGI2) (Grabarević i sur., 1997.). Oportuni nalaz istodobno statistički značajno manjih koncentracija hemoglobina u pokusnim skupinama (LN50, LN100 i LA/LN150) od one u kontrolnoj i u skupini LN150 (iako navedene vrijednosti nisu statistički značajno manje od one u LN150 skupini) (slika 2.) ostaje predmetom daljnjih istraživanja s obzirom na dosadašnje podatke o vrijedno-stima hemoglobina u zdravih i ascitičnih pilića (Maxwell i sur., 1986.a) te na oskudan broj istraživačkih radova o učinku akutne toksičnosti DO-a na pojavu ascitesa-SPH u tovnih pilića. Od izračunatih indeksa mase organa bitne su razlike utvr-đene samo u vrijednostima indeksa mase jetre (slika 3.). Statistički znatno manja vrijednost od kontrolne skupine (30,81) nalazi se u skupini LN150/LA (26,90). Autori pret-postavljaju kako je prekratko razdoblje proteklo od primjene tvari da bi došlo do bitnih promjena u masi jetre, no svakako je znakovito da su najveće vrijednosti zabilježene u kontrol-noj skupini, dok su u tretiranim skupinama vrijednosti bile daleko niže.
Zahvala
Istraživanje je financiralo Ministarstvo znanosti, obrazova-nja i športa RH (projekti br. 0053019 i 048-0531863-0464).
Literatura
ARNAL, J.-F., A. T. DINH-XUAN, M. PUEYO, B. DARBLADE, J. RAMI (1999): Endothelium-derived nitric oxide and vascular physiology and pathology. Cell. Mol. Life Sci. 55, 1078-1087.
ALBRECHT, E. W., C. A. STEGEMAN, P. HEERINGA, R. H. HENNING, H. van GOOR (2003): Protective role of endothelial nitric oxide synthase. J. Pathol. 199, 8-17.
BAGHBANZADEH, A., E. DeCUYPERE (2008): Ascites synd-rome in broilers: physiological and nutritional perspectives. Avian Pathol. 37, 117-126.
BALOG, J. M. (2003): Ascites syndrome (pulmonary hypertension syndrome) in broiler chickens: are we seeing the light at the end of the tunnel? Avian Poult. Biol. Rev. 14, 99-126.
BOTTJE, W., R. F. WIDEMAN, Jr. (1995): Potential role of free radicals in the pathogenesis of pulmonary hypertension syndrome. Poult. Avian Biol. Rev. 6, 211-231.
BOWEN, O. T., G. F. ERF, N. B. ANTHONY, R. F. WIDEMAN (2006): Pulmonary hypertension triggered by lipopoly-saccharide in ascites-susceptible and resistant broilers is not amplified by aminoguanidine, a specific inhibitor of inducible nitric oxide synthase. Poult. Sci. 85, 528-536.
BROZ, J., N. E. WARD (2007): The role of vitamins and feed enzymes in combating metabolic challenges and disorders. J. Appl. Poult. Res. 16, 150-159.
CHAPMAN, M. E., R. F. WIDEMAN (2001): Pulmonary wedge pressures confirm pulmonary hypertension in broilers initiated by an excessive pulmonary arterial (precapillary) resistance. Poult. Sci. 80:468-473.
CRESPO, R., H. L. SHIVAPRASAD (2003): Developmental, metabolic, and other noninfectious disorders, Ch. 31. U: Diseases of Poultry, 11. izd. (Saif Y. M., ur.) Iowa State University Press, Ames, Iowa, USA, 1055-1103.
CURRIE, R. J. (1999): Ascites in poultry: recent investigations. Avian Pathol. 28, 313-326.
DeSMIT, L., K. TONA, V. BRUGGEMAN, O. ONAGBESAN, M. HASSANZADEH, L. ARCKENS, E. DeCUYPERE (2005): Comparison of three lines of broilers differing in ascites susceptibility or growth rate. 2. Egg weight loss, gas pressures, embryonic heat production, and physiological hormone levels. Poult. Sci. 84, 1446-1452.
DeCUYPERE, E., J. BUYSE, N. BUYS (2000): Ascites in broiler chickens: exogenous and endogenous structural and functional causal factors. World Poult. Sci. J. 56, 367-377.
DeWILL, E., N. BUYS, G. A. ALBERS, E. DeCUYPERÉ (1996): Different characteristics in chick embryos of two broiler lines differing in susceptibility to ascites. Br. Poult. Sci. Fed. Proceed. 22, 182.
GASTON, B., J. S. STAMLER (1997): Nitrogen oxides. In: The Lung, 2. izd. R. G. Crystal, J. B. West, E. R. Weibel, P. J. Barnes, ur. Lippincott-Raven, New York, NY, 239-253.
GRABAREVIĆ, Ž., M. TIŠLJAR, P. DŽAJA, B. ARTUKOVIĆ, S. SEIWERTH, P. SIKIRIĆ (1997): The influence of BPC 157 on nitric oxide agonist and antagonist induced lesions in broiler chicks. J. Physiol. (Paris) 91, 139-149.
HALL, S. A., N. MACHICAO (1968): Myocarditis in broiler chickens reared at high altitude. Avian Dis. 12, 75-84.
HERNANDEZ, A. (1987): Hypoxic ascites in broilers: a review of several studies done in Colombia. Avian Dis. 31, 658-661.
IVANKOVIĆ, D. (1988.): Osnove statističke analize za medici-nare. Medicinski fakultet Sveučilišta u Zagrebu, Zagreb.
JULIAN, R. J., G. W. FAIRS, H. FRENCH, M. QUINTON (1987): The effect of increased sodium in the drinking water on right ventricular hypertrophy, right ventricular failure, and ascites to weight gain in broiler and roaster chickens. Avian Dis. 31, 130-135.
JULIAN, R. J., J. A. FRAZIER, M. GORYO (1989a): Right ventricular hypertrophy, right ventricular failure and ascites in broiler chickens caused by amiodarone-induced lung pathology. Avian Pathol. 18, 161-174.
JULIAN, R. J., I. McMILLAN, M. QUINTON (1989b): The effect of cold and dietary energy on right ventricular hypertrophy, right ventricular failure and ascites in meat-type chickens. Avian. Pathol. 18, 675-684.
JULIAN, R. J. (1990): Cardiovascular disease. U: Poultry Diseases, 3. izd. (Jordan, T. W. ur.), Baillière Tindall, London, 330-353.

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
165
JULIAN, R. J. (2000): Physiological, management and environ-mental triggers of the ascites syndrome: a review. Avian Pathol. 29, 519-527.
JULIAN, R. J. (2005): Production and growth related disorders and other metabolic diseases of poultry – a review. Vet. J. 169, 350-369.
JULIAN, R. J. (2007): The response of the heart and pulmonary arteries to hypoxia, pressure, and volume. A short review. Poult. Sci. 86, 1006-1011.
KHAJALI, F., R. F. WIDEMAN (2010): Dietary arginine: metabolic, environmental, immunological and physiological interrelationships. World Poult. Sci. J. 66, 751-766.
KHAJALI, F., M. TAHMASEBI, H. HASSANPOUR, M. R. AKBARI, D. QUJEQ, R. F. WIDEMAN (2011): Effects of supplementation of canola meal-based diets with arginine on performance, plasma nitric oxide and carcass characteristics of broiler chickens grown at high altitude. Poult. Sci. 90, 2287-2294.
LOPEZ-COELLO, C., T. W. ODOM, R. F. WIDEMAN (1986): Un importante factor en la mortalidad de asaderos. Industria Avicola 33, 12-17.
MARTINEZ-LEMUS, L. A., R. K. HESTER, E. J. BECKER, J. S. JEFFREY, T. W. ODOM (1999): Pulmonary artery endo-thelium-dependent vasodilation is impaired in a chicken model of pulmonary hypertension. Am. J. Physiol. 277:R190-197.
MARTINEZ-LEMUS, L. A., R. K. HESTER, E. J. BECKER, G. A. RAMIREZ, T. W. ODOM (2003): Pulmonary artery vasoactivity in broiler and leghorn chickens: an age profile. Poult. Sci. 82,1857-1964.
MAXWELL, M. H., G. W. ROBERTSON (1997): World broiler ascites survey 1996. Poult. Int. 36, 16-30.
MAXWELL, M. H., G. W. ROBERTSON, S. SPENCE (1986a): Studies on an ascitic syndrome in young broilers. I Haematology and pathology. Avian Pathol. 15, 511-524.
MAXWELL, M. H., G. W. ROBERTSON, S. SPENCE (1986b): Studies on ascitic syndrome in young broilers. II Ultra-structure. Avian Pathol. 15, 525-538.
MORENO de SANDINO, M., A. HERNANDEZ (2003): Nitric oxide synthase expression in the endothelium of pulmonary arterioles in normal and pulmonary hypertension chicks subjected to chronic hypoxia. Avian Dis. 47, 1291-1297.
ODOM , T. W., L. A. MARTINEZ-LEMUS, R. K. HESTER, J. BECKER, S. JEFFREY, G. A. MEININGER, G. A. RAMIREZ (2004): In vitro hypoxia differentially affects constriction and relaxation responses of isolated pulmonary arteries from broiler and leghorn chickens. Poult. Sci. 83, 835-841.
OLKOWSKI, A. A. (2007): Pathophysiology of heart failure in broiler chickens: Structural, biochemical and molecular characteristics. Poult. Sci. 86, 999-1005.
PAVLIDIS, H. O., J. M. BALOG, L. K. STAMPS, J. D. HUGHES, Jr., W. E. HUFF, N. B. ANTHONY (2007): Divergent selection for ascites incidence in chickens. Poult. Sci. 86, 2517-2529.
RAJANI, J., M. A. KARIMI TORSHIZI, S. RAHIMI (2011): Control of ascites mortality and improved performance and meat shelf-life in broilers using feed adjuncts with presumed antioxidant activity. Anim. Feed. Sci. Technol., 170, 239-245.
SHLOSBERG, A., M. BELLAICHE, E. BERMAN, A.B. DAVID, N. DEEB, A. CAHANER (1998): Comparative effects of added sodium chloride, ammonium chloride, or potassium bicarbonate in the drinking water of broilers, and feed
restriction, on the development of the ascites syndrome. Poult. Sci. 77, 1287-1296.
TAN X., W. D. SUN, J. C. LI, J. Q. PAN, Y. J. LIU, J. Y. WANG, X. L. WANG (2007): L-arginine prevents reduced expression of endothelial nitric oxide synthase (NOS) in pulmonary arterioles of broilers exposed to cool temperatures. Vet. J. 173, 151-157.
TANKSON, J. D., J. P. THAXTON, Y. VIZZIER-THAXTON (2001): Pulmonary hypertension syndrome in broilers caused by Enterococcus faecalis. Infect. Immun. 69, 6318-6322.
TANKSON, J. D., J. P. TANKSON, Y. VIZZIER-THAXTON (2002): Morphological changes in heart and lungs of broilers experiencing pulmonary hypertension syndrome caused by Enterococcus faecalis. Poult. Sci. 81, 365-370.
TIŠLJAR, M. (2004): Učinak antagonista i agonista dušik (II) oksida na pojavu ascitesa – sindroma plućne hipertenzije u tovnih pilića. Doktorska disertacija. Veterinarski fakultet Sve-učilišta u Zagrebu. Zagreb.
TIŠLJAR, M., Ž. GRABAREVIĆ, B. ARTUKOVIĆ, P. DŽAJA, J. FORŠEK, R. BARIĆ RAFAJ, Lj. BEDRICA, V. HERAK-PERKOVIĆ, B. ŠIMPRAGA, H. MAZIJA (2005): Istraži-vanje kronične toksičnosti agonista i antagonista dušik (II) oksida i njezin učinak na pojavu ascitesa – sindroma plućne hipertenzije u tovnih pilića. Zbornik, VI. simpozij "Peradarski dani 2005." s međunarodnim sudjelovanjem / Balenović, M. (ur.).Zagreb: Centar za peradarstvo, 2005;123-137.
TIŠLJAR, M., Ž. GRABAREVIĆ, B. ARTUKOVIĆ, P. DŽAJA, S. ĆENAN, T. AMŠEL ZELENIKA, R. G. COOPER, M. DINARINA-SABLIĆ (2011): The impact of L-NAME and L-arginine chronic toxicity induced lesions on ascites – pulmonary hypertension syndrome development in broiler chickens. Coll. Antropol. 35, 547-556.
TIŠLJAR, M. (2013): neobjavljeni podatci. VELDKAMP, T., R. P. KWAKKEL, P. C. M. FERKET, J.
SIMONS, J. P. NOORDHUIZEN, A. PIJPERS (2000): Effects of ambient temperature, arginine-to-lysine ratio and electrolyte balance on performance, carcass and blood para-meters in commercial male turkeys. Poult. Sci. 79, 1608-1616.
VILLAMOR, E., K. RUIJTENBEEK, V. PULGAR, J. G. R. De MAY, C. BLANCO (2002): Vascular reactivity in intra-pulmonary arteries of chicken embryos during transition to ex ovo life. Am. J. Physiol. Regul. Integr. Comp. Physiol. 282, R917-R927.
WANG, W., G. F. ERF, R. F. WIDEMAN, Jr. (2002): Effect of cage vs. floor litter environments on the pulmonary hyper-tensive response to intravenous endotoxin and on blood-gas values in broilers. Poult. Sci. 8, 1728-1737.
WEIDONG, S., W. XIALONG, W. JINYONG, J. RUIPING (2002): Pulmonary arterial pressure and electrocardiograms in broiler chickens infused intravenously with L-NAME, an inhibitor of nitric oxide synthase, or sodium nitroprusside (SNP), a nitric oxide donor. Br. Poult. Sci. 43, 306-312.
WIDEMAN, R. F., M. F. FORMAN, J. D. HUGHES, Y. K. KIRBY, N. MARSON, N. B. ANTHONY (1998): Flow-dependent pulmonary vasodilation during acute unilateral pulmonary artery occlusion in jungle fowl. Poult. Sci. 77, 615-626.
WIDEMAN, R. F., M. E. CHAPMAN, W. WANG, G. F. ERF (2004): Immune modulation of the pulmonary hypertensive response to bacterial lipopolysaccharide (endotoxin) in broilers. Poult. Sci. 83, 624-637.
WIDEMAN, R. F., G. F. ERF, M. E. CHAPMAN (2005): Nω-nitrol-arginine methyl ester (l-NAME) amplifies the pulmo-

UČINAK AKUTNE TOKSIČNOSTI L-NAME-A I L-ARGININA NA POJAVU ASCITESA – SINDROMA PLUĆNE HIPERTENZIJE U TOVNIH PILIĆA. KRATKO PRIOPĆENJE
166
nary hypertensive response to microparticle injections in broilers. Poult. Sci. 84, 1077-1091.
WIDEMAN, R. F., M. E. CHAPMAN, K. R. HAMAL, O. T. BOWEN, A. G. LORENZONI, G. F. ERF (2007): An inadequate pulmonary vascular capacity and susceptibility to pulmonary arterial hypertension in broilers. Poult. Sci. 86, 984-998.
WIDEMAN, R. F., D. D. RHOADS, G. F. ERF, N. B. ANTHONY (2013): Pulmonary arterial hypertension (ascites syndrome) in broilers: a review. Poult. Sci. 92, 64-83.
WIDEMAN, R. F., Y. K. KIRBY (1995): Evidence of a ventilation perfusion mismatch during acute unilateral pulmonary artery occlusion in broilers. Poult. Sci. 74, 1209-1217.
WIDEMAN, R. F., K. R. HAMAL (2011): Idiopathic pulmonary arterial hypertension: an avian model for plexogenic arterio-pathy and serotonergic vasoconstriction. J. Pharmacol. Toxicol. Methods 63, 283-295.
WIDEMAN, R. F. (2000): Cardio-pulmonary hemodynamics and ascites in broiler chickens. Poult. Av. Biol., R. R. Dietert, M. A. Ottinger, ur. 11:21-43.
WIDEMAN, R. F. (2001): Pathophysiology of heart/lung dis-orders: pulmonary hypertension syndrome in broiler chickens. World Poult. Sci. J. 57, 289-307.
WILSON, J. B., R. J. JULIAN, K. BARKER (1988): Lesions of right heart failure and ascites in broiler chickens. Avian Dis. 32, 246-261.
THE IMPACT OF L-NAME AND L-ARGININE ACUTE TOXICITY INDUCED LESIONS ON ASCITES – PULMONARY HYPERTENSION SYNDROME DEVELOPMENT IN BROILER CHICKENS Summary The impact of L-arginine (LA), a precursor for the synthesis of nitric oxide (NO), and N-ω-nitro-L-arginine methyl ester (L-NAME, LN), a non-selective inhibitor of the enzyme producing nitric oxide (nitric oxide synthase; NOS) acute toxicity induced lesions on ascites – pulmonary hypertension syndrome development was investigated in 25 thirty-day-old male broiler chickens (AVIAN 34). The effect of intraperitoneal (ip) single dose application of saline, 0.90% w/v NaCl (1 mL/kg BW), L-NAME (doses 50, 100, 150 mg/kg BW) and L-arginine (100 mg/kg BW) in combination with L-NAME (150 mg/kg BW) was evaluated 6 hours after the application according to clinical signs, gross pathomorphological findings and histopathologic findings in the spleen, liver, heart, lungs, bursa of Fabricius and thymus as well as hematologic analysis results and organ mass index values of the liver, spleen, heart, thymus and bursa of Fabricius. Histopathologic examination revealed that L-NAME caused severe tissue damage (myocardial necrosis and necrosis of lymphoid cells in the spleen). This effect of L-NAME was successfully inhibited by the application of L-arginine. Hematologic analysis showed significant hemoglobin and leukocyte count decrease in all treated groups of chicks (L-NAME- and L-NAME/L-arginine treated groups). Key words: nitric oxide, L-arginine, L-NAME, broilers, ascites – pulmonary hypertension syndrome

PERADARSKI DANI 2013.
167
MYCOTOXINS IN POULTRY: CONTAMINATION, IMPACT AND CONTROL STRATEGIES
Julia Dvorska
Sumy National Agrarian University, Sumy, Ukraine Summary The level of the mycotoxin contamination of broiler breeder feeds was determined using Alltech 37+ Program (UPLC-MS/MS). Feed contained low levels of ochratoxins, Type B trichothecenes (DON group) and fumonisins. Type A trichothecenes (T-2 group) were detected in moderate concentrations. Slight reduction in feed intake and egg production, oral lesions, comb necrosis, tissue necrosis, lymphoid, hematopoietic and oral mucosa lesions, occasionally abnormal feathering and thinning of the eggshells with an increase in the percentage of embryonic mortality and decreased hatchability were observed on the farm. At necropsy, the lesions were found mainly in the liver, which was reddened due to necrosis and congestion, or yellow due to lipid accumulation. Use of adsorbents on a base of glucomannans was effective in preventing the negative effect of mycotoxins on broiler breeders. Key words: mycotoxins, occurrence, poultry, control strategies, glucomannans Introduction Mycotoxins are secondary metabolites produced by fungi, especially those belonging to the genera Aspergillus, Penicillium and Fusarium. Mycotoxin contamination can occur in all agricultural commodities in the field and/or during storage, if conditions are favorable to fungal growth. More than 500 mycotoxins have been identified in feeds and feed ingredients to date. However, only a few have been tested in the field. Although the type of mycotoxin tested depends on the toxin prevalence, aflatoxins are analyzed around the world because of their carcinogenic properties. South Asian feed mills sometimes are also tested for ochratoxin A and T-2 toxin if the standards are available. In North America, deoxynivalenol (vomitoxin or DON) is widely tested, with occasional testing for T-2 toxin, zearalenone and fumonisins. In Europe, five mycotoxins (aflatoxins, deoxynivalenol, zearalenone, fumonisins and ochratoxin A) are covered by EU legislation (regulation or recommendation). Trans-gressions of these limits are rarely observed in official monitoring programs. However, low-level contamination by Fusarium toxins is very common (e.g., deoxynivalenol (DON) is typically found in more than 50% of the samples) and co-contamination is frequently observed. Multi-myco-toxin studies have reported 75%-100% of the samples to
contain more than one mycotoxin, which could impact animal health already at low doses. Co-occurrence of mycotoxins is likely to arise for at least three different reasons: (i) most fungi are able to simultaneously produce a number of mycotoxins, (ii) commodities can be conta-minated by several fungi, and (iii) completed feed is made from various commodities (Streit et al., 2012). Monitoring of mycotoxin contamination of grain and feed samples from various regions of Ukraine and two regions of Russia was conducted during the 2005-2010 period. Zeara-lenone, deoxynivalenol and fumonisins were determined by ELISA, and T-2 toxin and HT-2 toxin by bioautographic method. Deoxynivalenol, T-2 toxin, HT-2 toxin, zearalenone and fumonisins were detected in 72%, 33%, 5%, 51% and 74% of the samples at a mean concentration of 60, 44, 131, 25 and 734 μg/kg, respectively. In 0.8%-3.3% of the samples, mycotoxin concentrations exceeded the maximum permissible levels established in Ukraine and other count-ries. The data indicate the widespread occurrence of mycotoxins as contaminants of grain and feed in Ukraine (Trufanov, 2011).
Material and Methods
Feed samples were collected from one of the broiler farms in the east of Ukraine, where some clinical signs of mycotoxin problem were observed.
Julia Dvorska, Assoc. Prof., Sumy National Agrarian University, Sumy, Ukraine; Tel: +380677355506; E-mail: [email protected]

MYCOTOXINS IN POULTRY: CONTAMINATION, IMPACT AND CONTROL STRATEGIES 168
Alltech has developed the 37+ Program to assist in the detection of more than 37 mycotoxins using a holistic approach. The objective of the program is to evaluate feedstuffs for multiple mycotoxins using ultra-performance liquid chromatography coupled to tandem mass spectrometry (UPLC-MS/MS) methodology developed at the Alltech Global Research Headquarters in Kentucky. The mass spectrometry technique, LC-MS2, utilizes the high sensitivity and selectivity of liquid chromatography to dramatically enhance the definition and quantification of mycotoxins in complex matrices such as feedstuffs. This technique combines separation capabilities (LC) with the accurate detection of mass particles (MS). MS/MS is a mass spectrometer capable of doing several rounds of mass spectrometry separated by mass fragmentation. This approach allows for recording of the analyte atomic mass or fragments and its unequivocal assignment, with its fragmentation enabling its specific fingerprinting. The
successful addition of the LC-MS2 analytical technique to Alltech core competencies allowed for development of analytical methods investigating more than 30 different mycotoxins quantitatively, and more than 50 others qualitatively in less than 15 minutes per sample analyzed with limits of detection in the ppt range. Patterns of contamination are now investigated in feed matrices to better capture mycotoxin profiles and the associated risks for the animal consuming contaminated feed material. Feed samples were sent to the Alltech Global Research Headquarters in Kentucky for 37+ mycotoxin analysis. Clinical signs and pathomorphological changes were also studied in broilers fed analyzed feed on the farm.
Results
Level of contamination in one of the feed samples is presented in the table below.
Sample: 006, Poultry Feed Species: Broiler breeders
Toxins Amount, ppb Low risk Medium risk Low risk Aflatoxin (B1) 0 <5 10 20
Aflatoxins (B1+B2+G1+G2) 0 <10 25 50
Ochratoxins (A+B) 5.528 <10 25 50
Type B Trichothecenes* 263.955 <500 1000 1500
Type A Trichothecenes** 43 <25 50 100
Fumonisins (B1+B2+B3) 1325.764 <1000 2500 5000
Zearalenone group 0 <1000 1750 2500
Other Penicillium mycotoxins*** 0 <25 50 100
Other Aspergillus mycotoxins**** 0 <25 50 100
Ergot toxins 0 <250 500 1000
* Type B Tricho = deoxynivalenol (DON) + 15-acetyl DON + 3-acetyl DON + fusarenon X + nivalenol + DON-3-glucoside ** Type A Tricho = T-2 + HT-2 + diacetoxyscirpenol (DAS) + neosolaniol *** Penicillium mycotoxins = patulin + penicillic acid + roquefortine C + mycophenolic acid + wortmannin **** Aspergillus mycotoxins = gliotoxin + patulin + sterigmatocystin + verruculogen
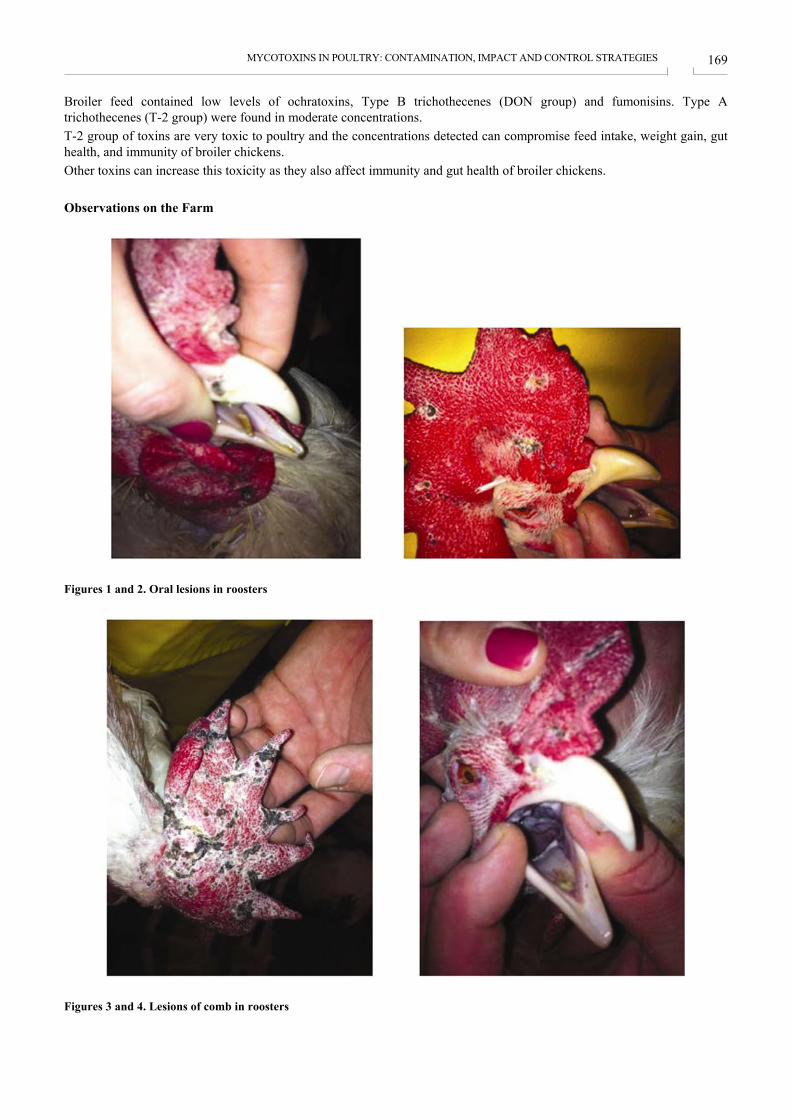
MYCOTOXINS IN POULTRY: CONTAMINATION, IMPACT AND CONTROL STRATEGIES
169
Broiler feed contained low levels of ochratoxins, Type B trichothecenes (DON group) and fumonisins. Type A trichothecenes (T-2 group) were found in moderate concentrations. T-2 group of toxins are very toxic to poultry and the concentrations detected can compromise feed intake, weight gain, gut health, and immunity of broiler chickens. Other toxins can increase this toxicity as they also affect immunity and gut health of broiler chickens. Observations on the Farm
Figures 1 and 2. Oral lesions in roosters
Figures 3 and 4. Lesions of comb in roosters

MYCOTOXINS IN POULTRY: CONTAMINATION, IMPACT AND CONTROL STRATEGIES 170
Figures 5 and 6. Liver lesions
Figure 7. Egg abnormalities
On the farm, we observed slight reduction in feed intake and egg production, oral lesions (Figs. 3 and 4), tissue necrosis, lymphoid, hematopoietic and oral mucosa lesions, some-times abnormal feathering, and thinning of the eggshells with an increase in the percentage of embryonic mortality and decreased hatchability (Fig. 7). Necrotic lesions were also found in rooster combs; almost 50% of them were affected. We found that oral lesions in broiler breeder occurred in approximately 60% of the lots where these birds were fed a diet contaminated with mycotoxins. At necropsy, the lesions were mainly found in the liver, which was reddened due to necrosis and congestion, or yellow due to lipid accumulation. Hemorrhages were also observed.
Upon the receipt of feed analysis, we suggested the farmer to use effective adsorbent, esterified glucomannan, a yeast cell wall derivative of Saccharomyces cerevisiae, to prevent the negative affect of mycotoxins.
Discussion and Conclusion
The presence of mycotoxins in poultry diets can determine the losses in poultry production system. From the harvest of grain, the effects of fungal and mycotoxicologic conta-mination can be perceived, and its effects can reflect on the production of chickens (Binder, 2007). The control depends on the implementation of appropriate policies within the agricultural management, production systems and storage, which are the root of the problem. Research in these areas should be developed, reflecting better results in economic and productive livestock, but also for the improvement of healthy food for human consumption. Use of Alltech 37+ Program provided a glimpse of the mycotoxin profile of feedstuffs. Further data will improve the understanding of mycotoxin prevalence. The presence of multiple mycotoxins is a common phenomenon in the field, and this can lead to mycotoxin interactions and, ultimately, to an increased risk to animal health and performance. The implementation of a mycotoxin control program based on hazard analysis and critical control point principles for farms and feed mills represents an integrated approach to control mycotoxin challenges. There are various approaches to control or combat mycotoxin problems. The simplest strategy is based on the

MYCOTOXINS IN POULTRY: CONTAMINATION, IMPACT AND CONTROL STRATEGIES
171
prevention of the formation of mycotoxins in feeds by special management programs including storage at low humidity levels and prevention of grain damage during processing. However, modern agronomic technology is not able to eliminate pre-harvest infection of susceptible crops by field fungi. Therefore, this strategy can only partially be effective; and in countries with warm and humid conditions, this strategy could be quite costly. Esterified glucomannan, a yeast cell wall derivative of Saccharomyces cerevisiae, effectively prevent the negative effect of mycotoxins in poultry through the binding and reduction in their bioavailability (Dvorska, 2011).
References
BINDER, E. M. (2007): Managing the risk of mycotoxins in mo-dern feed production. Anim. Feed Sci. Technol., 133, 149-166.
DVORSKA, J. (2011): Mycosorb in prevention of fusariotoxicosis in poultry. Effective Feed Industry, 5 (77) 33-35.
FUJIMOTO, H. (2011): Yeasts and Molds: Mycotoxins: Classifi-cation, Occurrence and Determination. In: Encyclopedia of Dairy Sciences. 2nd edn. (J. W. Fuquay, ed.) Academic Press, San Diego, CA, USA, 792-800.
SULYOK, M., R. KRSKA, R. SCHUHMACHER (2007): A liquid chromatography/tandem mass spectrometric multi-mycotoxin method for the quantification of 87 analytes and its application to semi-quantitative screening of moldy food samples. Anal. Bioanal. Chem., 389, 1505-1523. [doi: 10.1007/s00216-007-1542-2.]
SWEENEY, M. J., A. D. W. DOBSON (1998): Mycotoxin pro-duction by Aspergillus, Fusarium and Penicillium species. Int. J. Food Microbiol., 43, 141-158. [doi: 10.1016/S0168-1605(98)00112-3.]
STREIT, E., G. SCHATZMAYR, P. TASSIS, E. TZIKA, D. MARIN, I. TARANU, C. TABUC, A. NICOLAU, I. APRODU, O. PUEL, I. OSWALD (2012): Current situation of mycotoxin contamination and co-occurrence in animal feed – focus on Europe. Toxins (Basel), 4 (10), 788-809.
TRUFANOV, O. (2011): Monitoring of mycotoxin occurrence in feed and commodities in Ukraine. Modern Problems Toxicol. 1 (2), 35-39.
MIKOTOKSINI U PERADARSTVU: KONTAMINACIJA, UČINCI I STRATEGIJE KONTROLE RIZIKA Sažetak Razina kontaminacije mikotoksinima hrane za piliće brojlere određivala se pomoću Alltech37+ Programa (UPLC-MS/MS). Hrana je sadržavala niske razine okratoksina, trihotecena tipa B (skupina DON) i fumonizina. Trihoteceni tipa A (skupina T-2) otkriveni su u umjerenim koncentracijama. Na gospodarstvu je zabilježeno manje smanjenje unosa hrane i proizvodnje jaja, oralne promjene, nekroza kreste, nekroza tkiva, limfoidne i hematopoetske promjene, promjene oralne sluznice, ponegdje slabija opernaćenost i tanja ljuska jajeta, uz porast postotka smrtnosti embrija i smanjenje broja izlegnutih pilića. Pri obdukciji promjene su uglavnom utvrđene na jetri, koja je bila crvena zbog nekroze i kongestije, ili pak žuta zbog nakupljanja masti. Primjena adsorbenata na bazi glukomanana pokazala se učinkovitom u sprječavanju negativnih učinaka mikotoksina na piliće brojlere. Ključne riječi: mikotoksini, pojavnost, perad, strategije kontrole, glukomanani

PERADARSKI DANI 2013. 172
MIKOTOKSINI U HRANI ZA ŽIVOTINJE – ZNAČENJE, ZAKONSKI PROPISI I PREPORUKE
Marijana Sokolović
Centar za peradarstvo, Hrvatski veterinarski institut, Zagreb, Hrvatska SAŽETAK Mikotoksini, značajni u peradarstvu, uglavnom su sekundarni proizvodi metabolizma plijesni rodova Aspergillus, Fusarium i Penicillium. Plijesni često kontaminiraju hranu i hranu za životinje te u odgovarajućim klimatskim i okolišnim uvjetima proizvode mikotoksine u svim fazama proizvodnje i uporabe hrane: prije žetve, tijekom žetve ili tijekom transporta i pohrane. Za određene mikotoksine utvrđen je štetni učinak za zdravlje ljudi i životinja, a u proizvodnji hrane i u uzgoju životinja predstavljaju i značajne ekonomske gubitke. Zbog nezaobilazne prisutnosti plijesni i toksina u hrani istraživani su brojni postupci smanjivanja njihovih štetnih učinaka. Različiti postupci dekontaminacije i inaktivacije toksina imaju ograničene učinke i redovito nisu niti primjenjivi niti ekonomski isplativi u praksi. S obzirom na učestalost toksina, njihove potencijalne štetne učinke i ograničene mogućnosti njihove inaktivacije jedino odgovarajući preventivni postupci u proizvodnji, preradi i pohrani hrane mogu smanjiti potencijalne ekonomske gubitke i štetne učinke za zdravlje ljudi i životinja.
Ključne riječi: mikotoksini, hrana za životinje, preventivne mjere
UVOD Hrana kontaminirana plijesnima i mikotoksinima uzrokuje širok raspon učinaka, od gotovo neprimjetnih do značajnih zdravstvenih, ali i ekonomskih gubitaka koji nastaju kao posljedica smanjene proizvodnje jaja te slabijih proizvodnih rezultata peradi u tovu. Uz negativne učinke plijesni na hrandibeni sastav i organoleptičke osobine hrane, hrana može sadržavati i sekundarne metabolite, mikotoksine. Bu-dući da su u hrani za životinje često prisutne različite vrste plijesni koje mogu proizvoditi različite mikotoksine, hrana je u praksi često istodobno kontaminirana različitim miko-toksinima. Samim time su i negativni učinci veći i složeniji nego što ih može uzrokovati samo jedna vrsta toksina. U peradi se često mogu uočiti simptomi karakteristični za mikotoksikoze, iako se analizom hrane utvrde tek niske razine pojedinih mikotoksina. Takvi simptomi su tada naj-češće rezultat interakcije različitih mikotoksina (sinergi-stički, aditivni ili antagonistički učinak), pričem je obično izražen jedan od simptoma toksičnosti (Manning i sur., 1985.; CAST, 2003.; Devegowda i Murthy, 2005.). Iako postoji velik broj poznatih plijesni (oko 100.000 vrsta) mikotoksine najčešće proizvode plijesni rodova Aspergillus, Fusarium i Penicillium (tablica 1.), dok je toksikogenost
dokazana i u rodova Alternaria, Cephalosporium, Myro-thecium, Stachybotrys, Trichoderma, Trichotecium, Verti-cillium i drugima (Pitt, 2000.; CAST, 2003.). Mikotoksini su heterogena skupina tvari različitih struktura štetni učinci kojih ovise o vrsti toksina, količini i dužini unosa toksina, vrsti životinje, spolu, dobi, zdravstvenom statusu životinje, uvjetima u kojima se životinje drže te prisutnosti drugih štetnih tvari i mikotoksina u hrani. Iako postoji više od 400 poznatih mikotoksina, zbog njihove učestalosti u hrani kao i dosad evidentiranih štetnih učinaka u ljudi i životinja naj-značajnijim se smatraju aflatoksini, ohratoksin A, trikoteceni tipa A (T-2/HT-2 toksin, diacetoksiscirpenol), trikoteceni tipa B (deoksinivalenol), fumonizini i zearalenon. Na štetne učinke mikotoksina može se posumnjati ako se u životinja uoče: odbijanje i smanjeni unos hrane, slabija kon-verzija hrane, pad tjelesne težine, smanjena proizvodnost i povećana učestalost bolesti koje nastaju u životinja slabijeg imunog sustava. Za većinu mikotoksina dokazani su različiti toksični učinci koji mogu biti akutni (hepatotoksični, nefro-toksični učinak i poremećaj sinteze proteina), kronični (kar-cinogeni, mutageni ili teratogeni učinak) te redovito imuno-supresivni učinak. Najčešći simptomi trovanja mikotoksi-nima u peradi prikazani su u tablici 2. (Leeson i sur., 1995.; Pitt, 2000.; CAST, 2003.; Binder i sur., 2007.)
Dr. sc. Marijana Sokolović, dr.med.vet, Centar za peradarstvo, Hrvatski veterinarski institut, Henizelova 55, 10000 Zagreb, Hrvatska; Tel.: ++385(0)12441-394; Fax: ++385(0)12441-396; E-mail: [email protected]

MIKOTOKSINI U HRANI ZA ŽIVOTINJE – ZNAČENJE, ZAKONSKI PROPISI I PREPORUKE
173
Tablica 1. Plijesni, mikotoksini i učinci u peradi (CAST, 2003.)
Mikotoksin Plijesni Aflatoksin (B1, B2, G1, G2) A. flavus; A. parasiticus; A. nomius; A. pseudotamarii Ohratoksin A A. ochraceus; P.verrucosum Trikoteceni: T-2 i HT-2 toksin Diacetoksiscirpenol (DAS) Deoksinivalenol (DON)
Fusarium, Myrothecium, Phomopsis, Stachybotrys, Trichoderma, Trichothecium i druge
Fumonizin (B1, B2, B3) F. verticillioides (sin. moniliforme); F. proliferatum; F. nygamai Zearalenon F. graminearum; F. culmorum, Fusaium equiseti, Fusarium crookwellense
Table 1. Moulds, mycotoxins and effects in poultry (CAST, 2003)
Mycotoxin Moulds Aflatoxin (B1, B2, G1, G2) A. flavus; A. parasiticus; A. nomius; A. pseudotamarii Ochratoxin A A. ochraceus; P.verrucosum Trichothecenes: T-2 and HT-2 toxin; Diacetoxyscirpenol (DAS); Deoxynivalenol (DON)
Fusarium, Myrothecium, Phomopsis, Stachybotrys, Trichoderma, Trichothecium i druge
Fumonisin (B1, B2, B3) F. verticillioides (sin. moniliforme); F. proliferatum; F. nygamai Zearalenone F. graminearum; F. culmorum, Fusaium equiseti, Fusarium crookwellense
Tablica 2. Štetni učinci mikotoksina u peradi (Leeson i sur., 1995.; Pitt, 2000.; CAST, 2003.; Binder i sur., 2007.)
Mikotoksin Simptomi Aflatoksin (B1, B2, G1, G2)
Akutno: gubitak tjelesne težine, depresija, ataksija i uginuće unutar 1-2 tjedna Kronično: imunosupresija, nastanak tumora, oštećenje jetre, slezene, gušterače i Fabricijeve burze, slabija proizvodnost (slabiji prirast, pad proizvodnje i težine jaja)
Ohratoksin A Akutno: dijareja, tremor, neuralni poremećaji i uginuće unutar 24 sata Kronično: dijareja, dehidracija, povećani mortalitet, smanjeni prirast, povećana konzumacija vode i vodenasti feces, smanjena proizvodnja jaja, smanjena masa jaja i promjene na ljuski jajeta
Trikoteceni: T-2 i HT-2 toksin Diacetoksiscirpenol (DAS) Deoksinivalenol (DON)
Akutno i kronično: oštećenje sluznice probavnog sustava, hemoragije, proljev i povraćanje, dermatitis, depigmentacije kože nogu, smanjena kvaliteta perja, neurološki poremećaji, smanjeni unos hrane, smanjeni prirast, zaostajanje u rastu, gubitak tjelesne težine, odbijanje hrane, smanjena proizvodnja jaja (tanka ljuska), smanjena valivost, cijanoze i leukopenija
Fumonizin (B1, B2, B3) Kronično: smanjenje tjelesne težine i povećana smrtnost Zearalenon Kronično: pad nesivosti i depresija rasta peradi
Table 2. Toxic effects of mycotoxins in poultry (Leeson i sur., 1995; Pitt, 2000; CAST, 2003; Binder i sur., 2007)
Mycotoxin Symptoms Aflatoxin (B1, B2, G1, G2)
Acute: weight loss, depression, ataxia, death within 1-2 weeks Chronic: immunosuppression, tumors, lesions of liver, spleen, pancreas and bursa of Fabricius, lower productivity (reduced weight gain, low egg production and egg weight)
Ochratoxin A Acute: diarrhea, tremor, neurologic disorders and death within 24 hours Chronic: diarrhea, dehydration, increased mortality, reduced weight gain, increased water consumption and watery feces, low egg production, reduced egg weight and egg shell changes
Trichothecenes: T-2 and HT-2 toxin; Diacetoxyscirpenol (DAS); Deoxynivalenol (DON)
Lesions of mucous membranes, hemorrhage, diarrhea and vomiting, dermatitis, depigmentation of the skin of the legs, low feather quality, neurologic disorders, reduced feed intake and growth, weight loss, feed refusal, low egg production (thin shell), reduced hatchability, cyanosis and leukopoenia
Fumonisin (B1, B2, B3) Reduced weight gain and increased mortality Zearalenone Low egg production and reduced weight gain

MIKOTOKSINI U HRANI ZA ŽIVOTINJE – ZNAČENJE, ZAKONSKI PROPISI I PREPORUKE 174
Učestalost u hrani. U objavljenoj literaturi velik je broj izvješća o nalazu toksigenih plijesni i mikotoksina u hrani i hrani za životinje. Pritom je u značajnom broju dokazana povezanost pristunosti mikotoksina u hrani i štetnog učinka u ljudi i životinja. S obzirom na to da se procjenjuje kako je godišnje četvrtina žitarica u cijelom svijetu kontaminirana mikotoksinima, mala je vjerojatnost da će životinje tijekom svog proizvodnog ciklusa biti hranjene hranom u kojoj mikotoksina nema. Zbog određenih klimatskih promjena kao i globalizacije trgovine nije više moguće govoriti o zemljopisnim područjima u kojima se neki mikotoksini na-laze, već je stvarna opasnost od prisutnosti pojedinih mi-kotoksina podjednaka u čitavom svijetu. Budući da rast plijesni i proizvodnja mikotoksina ovise o uvjetima okoliša, uz pravilne postupke u uzgoju, kontrolu proizvodnje i uvoza količina mikotoksina u hrani ovisi i o uvjetima transporta i skladištenja (Leeson i sur., 1995.; CAST, 2003.; EFSA, 2004.). Aflatoksini su u hrani prisutni često u vrlo niskim koncen-tracijama (nekoliko μg/kg), ali mogu biti prisutni u rasponu i većem od 0,1 mg/kg. Najčešće su prisutni u kukuruzu, sjemenu pamuka, kikirikiju, kokosu, palminim košticama, začinima, smokvama, brazilskim orasima, pistacijama i drugim orašastim plodovima, dok su nešto rjeđe prisutni u drugim vrstama hrane. Učestalost aflatoksina u svijetu kreće se u rasponu od 6% do 88%, odnosno 19%-51% na području Europe. Ohratoksin A pronađen je u kukuruzu, pšenici, soji, zobi, riži, vinu, pivu i kavi u koncentracijama uglavnom manjim od 0,5 mg/kg. Učestalost u žitaricama i hrani za životinje iznosi 11%-55%, a prisutan je u koncentracijama do 1 mg/kg i tek rjeđe su zabilježene koncentracije veće od 5 mg/kg. Trikoteceni su česti u žitaricama i hrani za životinje. Najčešći mikotoksin koji se može smatrati i biomarkerom prisutnosti drugih mikotoksina je deoksinivalenol. Manji je broj izvješća o prisutnosti drugih trikotecenskih mikotoksina uključujući T-2 toksin, HT-2 toksin i diacetoksiscirpenol. Deoksinivalenol je pronađen u 17%-71% uzoraka hrane u rasponu koncentracija do 8 mg/kg, a rjeđe u većim koncen-tracijama. Fumonizini su u pravilu redovito prisutni u ku-kuruzu u čitavom svijetu i to u koncentracijama manjim od 1 mg/kg, iako su zabilježene i koncentracije veće od 1 mg/kg. Fumonizini su prisutni u hrani u rasponu od 12% do 76%. Zearalenon je otkriven u kukuruzu, pšenici, ječmu i soji i to najčešće u koncentracijama manjim od 1 mg/kg. Učestalost zearalenona u hrani u svijetu kreće se u rasponu od 8% do 63% (CAST, 2003.; Sokolovic i Simpraga, 2006.; Binder i sur., 2007.). Zakonski propisi. Zbog izrazito štetnog učinka pojedinih mikotoksina dokazanog istraživanjima u uvjetima in vivo i in vitro, kao i iz epidemioloških istraživanja te nezaobilazne prisutnosti mikotoksina u hrani uvedeni su propisi kojima se nastoji smanjiti izloženost ljudi i životinja navedenim toksi-nima. Propisi su uvedeni temeljem procjene rizika i pro-pisuju najveće dopuštene količine pojedinih toksina u hrani i hrani za životinje. Primjerice, u Republici Hrvatskoj pri-mjenjuju se Pravilnik o najvećim dopuštenim količinama određenih kontaminanata u hrani (NN 146/2012.) i Pravilnik o izmjenama Pravilnika o nepoželjnim tvarima u hrani za
životinje kojim se provode Uredbe Europske komisije o izmjenama Direktive 2002/32/EZ Europskog parlamenta i Vijeća od 7. svibnja 2002. o nepoželjnim tvarima u hrani za životinje (NN 124/2012.). Uza zakonske propise postoje i odgovarajuće smjernice i preporuke s obzirom na količine pojedinih toksina u hrani i hrani za životinje. Preporuke. Mikotoksini su u pravilu vrlo otporni na ke-mijske i toplinske postupke koji učinkovito uništavaju spore plijesni. Najučinkovitija mjera sprječavanja izloženosti ljudi i životinja mikotoksinima je uklanjanje i uništavanje konta-minirane hrane. Međutim, navedena mjera zbog ekonomskih razloga u pravilu nije primjenjiva u praksi. Stoga, za sprječavanje štetnih učinaka mikotoksina redovito postoje dvije mogućnosti: sprječavanje rasta plijesni i proizvodnje mikotoksina te dekontaminacija kontaminirane hrane. U cilju sprječavanja rasta plijesni i proizvodnje mikotoksina preporuča se uzgoj biljaka otpornih na infestaciju plijesni-ma, postupci žetve i pohrane koji minimaliziraju kontami-naciju uključujući rotaciju žitarica i pravodobnu primjenu fungicida. Ipak, sprječavanje rasta plijesni prije žetve re-lativno je ograničeno zbog nemogućnosti utjecaja na klimatske promjene koje pogoduju nastanku plijesni kao što su nedostatak ili preobilne kiše tijekom uzgoja. Skladištenje žitarica od osobite je važnosti u prevenciji rasta plijesni i proizvodnji mikotoksina, jer pritom kvaliteta žitarica ovisi o čimbenicima makro- i mikro okoliša te prisutnosti različitih organizama (mikroorganizama, insekata, glodavaca i ptica). Stoga, održavanje odgovarajuće vlage žitarica (<13%) i drugih higijenskih uvjeta pohrane u pravilu će spriječiti neželjeni rast plijesni te posljedičnu proizvodnju toksina (Leeson i sur., 1995.; CAST, 2003.). Dekontaminacija kontaminirane hrane uključuje fizikalne, kemijske i biološke metode. Fizikalna dekontaminacija uključuje mehaničko razdvajanje oštećenih zrna, izlaganje toplini, zračenje i ekstrakciju s odgovarajućim otapalima (otopine etanola, acetona, izopropanola, metanola i dr.). Iako je navedenim metodama dokazana sposobnost smanjenja koncentracije mikotoksina u određenoj mjeri, one redovito nisu isplative, sigurne i primjenjive u praksi. Tvari koje mogu kemijski detoksificirati i/ili inaktivirati mikotoksine su različite kiseline, baze, aldehidi, bisulfidi, oksidirajući agen-si i pojedini plinovi. Međutim, kao i u slučaju fizikalne dekontaminacije, većina ovih postupaka nije primjenjiva u praksi, ne djeluju na sve vrste toksina, a vrlo često navedene kemijske tvari nisu dovoljno sigurne. Biološka dekontami-nacija podrazumijeva primjenu pojedinih mikroorganizama kojima se sprječava proizvodnja ili dolazi do inaktivacije toksina. Takav učinak dokazan je za pojedine netoksigene sojeve plijesni (A. flavus i A. parasiticus), kvasce i bakterije (Flavobacterium aurantiacum i Eubacterium spp.) i enzime. Sličan učinak dokazan je i primjenom pojedinih hranidbenih modifikacija (povećane razine metionina, selena i vitamina te dodatak nekih biljnih tvari). Jedna od učinkovitijih mjera sprječavanja štetnih učinaka toksina je neutralizacija već prisutnih mikotoksina uporabom inertnih adsorbensa koji priječe apsorpciju toksina u probavnom sustavu. U tu svrhu istraživana je učinkovitost različitih glina u inaktivaciji mi-kotoksina. Tako su ispitani bentoniti, zeoliti i alumino-

MIKOTOKSINI U HRANI ZA ŽIVOTINJE – ZNAČENJE, ZAKONSKI PROPISI I PREPORUKE
175
silikati. Uspješnost adsorpcije navedenih glina ovisi o strukturi i vrsti toksina i kreće se u širokom rasponu od 0 do 87%. Mineralne gline su se pokazale učinkovite u adsorpciji aflatoksina, dok su vrlo mali učinak pokazale u zaštiti od zearalenona, T-2 toksina, diacetoksiscirpenola i ohratoksina. Uz relativno malu sposobnost adsorpcije različitih toksina problem kod mineralnih glina je i visok stupanj inkluzije. Njihovom primjenom dokazano je slabije iskorištenje mine-rala bakra, cinka, klora, magnezija, mangana i natrija iz hrane. Uz mineralne gline ispitani su i drugi adsorbensi (or-ganski, neorganski i mješoviti) kao što su granulirani akti-virani ugljik, kolestiramin, hidratizirani kalcij-aluminosili-kati i glukomanan. Prema rezultatima istraživanja ovi dodaci hrani za životinje uz uspješno vezanje mikotoksina ujedno i povoljno djeluju na proizvodne rezultate u peradi u uzgoju (bolji prirast, očuvanje nesivosti, veća otpornost životinja na druge infekcije i sl.) (Volkl i Karlovsky, 1998.: Schiedeler, 1993.; CAST, 2003.; Devegowda i Murthy, 2005.).
Zaključci
Hrana za životinje često je kontaminirana mikotoksinima i to najčešće manjim koncentracijama razlitičih toksina. Una-toč mjerama prisutnost mikotoksina je neizbježna i speci-fičnog liječenja u slučaju intoksikacije nema. Primjena određenih adsorbenata može povoljno djelovati na zdravlje i proizvodnost životinja, ali apsolutna sigurnost nikada se ne postiže. Sprječavanje štetnih učinaka toksina temelji se na preventivi koja uključuje sprječavanje rasta plijesni na ži-taricama u svim fazama proizvodnje, prerade i pohrane. Uporaba zdravih i kvalitetnih sirovina u proizvodnji hrane te kontinuirana provjera kvalitete sirovina i gotovih proizvoda omogućuju unapređenje proizvodnje, smanjenje ekonomske štete i zaštitu zdravlja ljudi i životinja.
Literatura
BINDER, E. M., M. TAN, L. J. CHIN, J. HANDL, J. RICHARD (2007): Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 137:265-282.
CAST Report (2003): Mycotoxins: risks in plant, animal, and human systems. U: J. L. Richard, G. A. Payne (ur.), Council for Agricultural Science and Technology Task Force Report No. 139, Ames, Iowa, USA. ISBN 1-887383-22-0.
DEVEGOWDA, G., T. N. K. MURTHY (2005): Mycotoxins: effects on poultry performance and health. U: D. E. Diaz (ur.), The Mycotoxin Blue Book. Nottingham University Press, Nottingham, Velika Britanija. pp. 25-56.
EFSA (2004): Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to aflatoxin B1 as undesirable substance in animal feed. EFSA Journal 39:1-27.
LEESON, S., G. DIAZ, J. D. SUMMERS (1995): Poultry Meta-bolic Disorders and Mycotoxins. Ontario, Canada: University Books.
MANNING, R. O., T. P. BROWN, R. D. WYATT, O. J. FLETCHER (1985): The individual and combined effects of citrinin and ochratoxin A in broiler chicks. Avian Dis. 29:986-997.
PITT, J. I. (2000): Toxigenic fungi and mycotoxins. Br. Med. Bull. 56(1):184-192.
SCHIEDELER, S. E. (1993): Effects of various types of alumino-silicates and aflatoxin B1 on aflatoxin toxicity, chick perfor-mance and mineral status. Poult. Sci. 72:282-288.
SOKOLOVIC, M., B. SIMPRAGA (2006): Survey of trichothecene mycotoxins in grains and animal feed in Croatia by thin layer chromatography. Food Control 17(9):733-740.
VOLKL, A., P. KARLOVSKY (1998): Biological detoxification of fungal toxins and its use in plant breeding, feed, and food production. Natural Toxins 7:1-23.
MYCOTOXINS IN ANIMAL FEEDS – IMPORTANCE, LEGISLATIVE AND RECOMMENDATIONS Summary Mycotoxins, which are important in poultry production, are mostly secondary metabolites of moulds belonging to the genera Aspergillus, Fusarium and Penicillium. Moulds are often contaminants of food and feed and in favorable climate and environmental conditions produce mycotoxins in all phases of food production and manipulation, i.e. pre-harvest, during harvest, or on transport and storage. Toxic effects on human and animal health and significant economic losses in the production of food, feed and animals have been documented for some mycotoxins. Since the presence of moulds and mycotoxins in food is unavoidable, a lot of research has been done to find procedures reducing these toxic effects. Different toxin decontamination and inactivation procedures have limited efficiency and are usually not applicable or economically cost effective in everyday work. Due to the incidence of toxins, their potential toxic effects and limited possibilities of inactivation, only appropriate preventive procedures in the production, processing and storage of food and feed can reduce the potential economic losses and toxic effects on human and animal health. Key words: mycotoxins, animal feed, preventive measures

PERADARSKI DANI 2013. 176
IMPROVED GUMBORO CONTROL BY CEVAC TRANSMUNE IBD, AN IMMUNE COMPLEX VACCINE
G. Bánhidi
Technical manager / poultry CEVA SA Eastern Europe / Central Europe Gumboro disease virus, being a quite resistant pathogen, is capable to cause a persistent or recurrent infection of poultry farms. However, it is vital to prevent the infection of flocks since it can cause sub-clinical infection and significant economic losses even without being recognized and diagnosed. The prevention basically stands on two legs: biosecurity and vaccination, each of them equally important to secure profitable broiler production.
Vaccination against Gumboro disease is not easy for a couple of reasons. Each chick within a flock posses a specific amount of circulating maternal antibodies against Gumboro virus that can protect them for a while but are also able to neutralize vaccine viruses. Furthermore, the level of virus pressure (quantity and type of field viruses challenging the flock) is often not known. Therefore, veterinarians face the following crucial questions when thinking about Gumboro vaccination: - which vaccine strain to choose? - what is the best date to vaccinate the flock? - how many vaccinations should be given for sufficient protection? - how to administer efficiently the vaccine? Unfortunately, even little mistakes in vaccination timing or vaccine administration may lead to sub-optimal flock protection, latent infection and loss of performance. In case of vvIBD all that may be topped with significant mortality. Thanks to Cevac Transmune IBD, representing the innovative immune complex technology, the difficulties of IBD vaccination can now be overcome. This vaccine is made of antigen antibody complexes. The vaccine virus strain Winterfield 2512 is bound to specific IBD antibodies in an antigen-antibody complex. With Cevac Transmune IBD, one single dose administered in the hatchery either in ovo or subcutaneously is enough to protect the broilers throughout its life. How can this vaccine eliminate the above mentioned problems of traditional vaccination?
While the vaccine virus of a traditional vaccine can easily be eliminated even by low level of maternal antibodies, in Cevac Transmune IBD the vaccine virus is protected by the complex formed with specific antibodies. After injection the antigen-antibody complexes enter the bird’s bood circulation and upon reaching the spleen and thymus these complexes are captured by specific immune cells (follicular dendritic cells) present in those organs. The antigen-antibody complexes bound to the follicular dendritic cells are not permanently stable. With time, the specific antibodies will detach from the complex and the viral part is then released in the blood circulation. If the levels of circulating maternal antibodies are still high enough, the vaccine virus will be rapidly identified and neutralised by the immune system. As
long as the maternally derived antibodies remain higher than the vaccine “breakthrough” level, the vaccine viruses released from the spleen will always be neutralized. This process continues in time and after a while the level of MDA will not be enough for neutralising the released vaccine virus. In that moment the W2512 vaccine strain will be able to reach the bursa, replicate there and induce active immunity. It usually occurs around 14 to 28 days of age.

IMPROVED GUMBORO CONTROL BY CEVAC TRANSMUNE IBD, AN IMMUNE COMPLEX VACCINE
177
The immune system of chicks vaccinated with Cevac Transmune IBD will be constantly challenged by the vaccine and will respond the earliest possible time when the declining level of maternal antibodies allow it. In contrast to the drinking water vaccination there will be no birds that are already sensitive to infection before the vaccination, and also there will be no birds receiving the vaccine too early and missing immune response because of vaccine virus neutralization by MDA. Further to that, within-flock heterogeneity is not a problem anymore: one single vaccination at hatchery can solve the vaccination of the whole flock regardless the differences between birds. It is also known that the mistakes of drinking water vaccination frequently result in sub-optimal protection and decreased performance due to sub-clinical infections. Cevac Transmune IBD can fully eliminate all the potential weaknesses of drinking water method, thus ensuring constant, high-level protection to all flocks all along the year. Optimal flock performance needs freedom from sub-clinical infections
In summary, with the use of Cevac Transmune IBD all the broilers will be: - individually and surely vaccinated already in the hatchery (no question about the quality of on-farm vaccination) - protected against each forms of Gumboro disease (no question about vaccine choice) - immunized at the right time according to their level of MDA (no question about vaccination timing and number of vaccinations) Thanks to its mechanism of action, Cevac Transmune IBD goes further than conventional drinking water vaccination. It will protect 100% of the broilers and allow a full expression of their genetic ability for your own benefit.

PERADARSKI DANI 2013. 178
SUZBIJANJE ZDRAVSTVENIH PROBLEMA U PERADARSTVU - BIOTRONIC TOP 3
Siniša Đurđević, dipl. ing.
BIOMIN Hrvatska d. o. o.
Zakiseljivači se u intenzivnoj peradarskoj proizvodnji koriste više od 25 godina. Već samo dodavanje mravlje kiseline u obliku soli (formiata) značajno je snizilo mortalitet i konverziju hrane. Nakon toga došli su zakiseljivači druge generacije koji su sadržavali mješavinu kiselina i njihovih soli. Treća generacija zakiseljivača dovela nas je do zakiseljivača sa sekvencijalnim otpuštanjem aktivnih tvari uz dodatak EC (E. Coli kontrolera). Kontrola štete uz pomoć dobro proučenog mehanizma djelovanja organskih kiselina i njihovih soli, ustaljena je praksa proteklih nekoliko desetljeća. Uobičajeni konvencionalni zakiseljivači dosegnuli su maksimum učinkovitosti. Daljnje usavršavanje proizvoda postalo je neizvjesno i neperspektivno. Danas se u BIOMIN-u možemo pohvaliti četvrtom generacijom zakiseljivača, koja uz mješavinu kiselina i soli ima ugrađenu fitokemijsku supstancu uz patentirani permeabilizator. Uz doziranje od 1 – 1,5 kg / t gotove smjese, najnovija generacija zakiseljivača ne samo da je izrazito učinkovita već je i financijski isplativija. Bakterioze izazvane E. coli i Salmonella spp. u suvremenom stočarstvu izazivaju značajne gubitke, usprkos neprekidnom usavršavanju nutricionizma i zdravstvene zaštite životinja. Najmodernija kontrola štete izazvana gram-negativnim bakterijama odvija se uz pomoć permeabilizatora, kemijskih tvari koje pomažu dezintegraciju vanjske membrane mikroorganizma i olakšavaju drugim antimikrobnim tvarima ulaz u stanicu pojačavajući djelovanje aktivne tvari. U prisustvu permeabilizatora maksimizira se učinkovitost konvencionalnih kiselina koje je moguće dozirati u smanjenim koncentracijama. Permeabilizatori pomažu učinkovitost fitogenih kemijskih tvari. Fitokemikalije su prirodne tvari koje proizvode biljke i imaju zaštitnu, antimikrobnu i/ili antioksidativnu funkciju. Cimet aldehid je prirodno fitokemijsko sredstvo koje sprječava dijeljenje bakterijske stanice, odnosno onemogućava diobu i razmnožavanje bakterija. Fitokemikalije su prirodni kontrolor diobe bakterijske stanice. Biotronic TOP 3 sadrži permeabilizator, fitokemikalije i organske kiseline na sporo otpuštajućem mediju (SRM). Potvrđen je u praksi u brojnim znanstvenim i komercijalnim pokusima diljem svijeta. Bez negativnih učinaka i kontraindikacija otvara novo poglavlje u kontroli štete izazvane gram–negativnim bakterijama.





























