Embed Size (px)
Citation preview

Uvod u fiziologiju rastenja i razvića
biljaka

Razvojni procesi kod biljaka počinju od semena koje klija i obuhvataju formiranje biljke na kojoj sazreva novo seme.
Na njima se zasniva i na osnovu njih se realizuje ceo životni ciklus (ontogeneza) biljke. Da bi se razvijala, biljka mora da
obavlja sve svoje osnovne funkcije – usvajanje vode i mineralnih materija, fotosintezu, disanje i sintezu
organskih jedinjenja. Međutim, iako se razviće biljaka zasniva na ovim procesima, ono nije jednostavni zbir
osnovnih funkcija, već predstavlja integrisani niz morfoloških, fizioloških i biohemijskih procesa koji podležu
autonomnoj regulaciji. Ti procesi su međusobno zavisni, imaju za cilj optimalno funkcionisanje biljke u određenim
uslovima sredine. Razvojni procesi su determinisani genetičkom osnovom biljke, ali na njih utiču i faktori
spoljašnje sredine, naročito svetlost, temperatura i voda. Kod biljaka, koje su sesilni organizmi, spoljašnja sredina mnogo više utiče na razvojne procese, nego što je to slučaj
kod životinja.

Svaka biljka potiče od jedne ćelije, zigota. Zigot nastaje oplođenjem (u semenom zametku) i razvija se u embrion u semenu, a zatim, posle klijanja semena, i u višećelijski organizam. Zigot je jedina totipotentna ćelija u višećelijskom organizmu (razvićem može dati SVE tipove ćelija koji se nalaze u biljnom organizmu).
Tokom razvića biljke bitni procesi su: deoba ćelija, rastenje i diferencijacija. Rastenje je proces ireverzibilnog povećanja dimenzija organa ili biljnog organizma. U osnovi rastenja mogu biti povećanje: broja ćelija, sveže ili suve mase, zapremine, površine i/ili dimenzija (dužine, obima). NIJE svako povećanje navedenih parametara rastenje, već samo ireverzibilno povećanje. Rastenje može da se ostvari i kombinacijom povećanja nabrojanih parametara.
Diferencijacija je kvalitativna promena, proces u kome iz istih elemenata nastaju različiti. Ona podrazumeva pojavu novih tipova ćelija, tkiva ili organa i uspostavljanje višeg nivoa organizacije i složenosti organizma.

Morfogeneza je obrazovanje biljnih organa (korena, stabla, lista, cveta i dr.) iz inicijalnih meristemskih ćelija. U ovom procesu se prvo formiraju primordije (začeci) organa, a zatim se formiraju i organi. De novo morfogeneza je formiranje organa bez prethodnog formiranja primordija i na mestima gde se obično ne formiraju, u tkivima u kojima se somatske ćelije prvo dediferenciraju, a zatim rediferenciraju u određenom pravcu (kako bi dale novi organ). Ovaj proces može da se desi i spontano, ali se češće indukuje u uslovima kulture viših biljaka in vitro, jer zahteva promenu endogenih faktora, pre svega nivoa endogenih fitohormona.
Rastenje, diferencijacija i morfogeneza su međusobno povezani procesi, koji se često dešavaju uporedo, pa se ne mogu potpuno razdvojiti i ograničiti.

• Razviće se može razmatrati na nivou ćelija i na nivou celog biljnog organizma. Međutim, razviće ćelija i razviće organizma su povezani, pa se ne mogu razmatrati odvojeno.
• Ćelije prolaze kroz 3 razvojna stupnja: 1. ćelijska deoba 2. povećanje dimenzija ćelije 3. diferencijacija
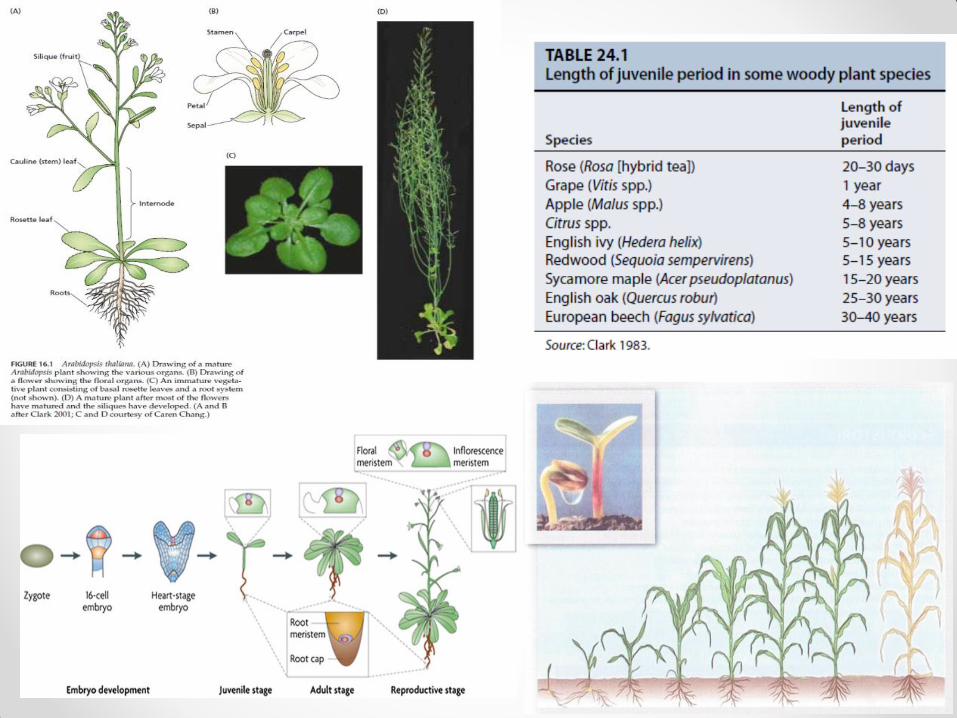
• U ontogenezi (razviće biljke od semena do semena) svakog biljnog organizma postoje 2 faze: I. vegetativna embriogeneza (embrionalni razvoj od zigota do embriona; dešava se u semenu) mirovanje semena klijanje semena razvoj mladih biljaka (juvenilna faza u kojoj se formiraju, rastu i razmnožavaju –vegetativno razmnožavanje – vegetativni organi, izdanci i koren) II. reproduktivna (period zrelosti u kojem biljke stiču sposobnost polnog razmnožavanja) indukcija cvetova razviće cvetova oprašivanje i oplođenje razviće plodova i semena
• Posle reproduktivne faze sledi faza starenja (senescencije) i smrti biljke.

Razviće biljaka je precizno vremenski usklađen i organizovan niz procesa, određenih naslednom osnovom i modifikovanih tako da usklade funkcionisanje biljaka sa uslovima sredine u kojoj živi. Zbog toga su mehanizmi kontrole razvića kod biljaka složeni. Oni se mogu ispoljiti na tri nivoa: intracelularni nivo regulacije (podrazumeva regulaciju na nivou
sinteze nukleinskih kiselina i proteina, pošto diferencijacija u suštini zavisi od aktivnosti određenih enzima)
intercelularni nivo regulacije (podrazumeva regulaciju integracije razvića na nivou organizma i odvija se na osnovu uticaja razvića jednih organa na razviće drugih – korelativni odnosi; ostvaruje se pomoću hemijskih supstanci sa signalnom funkcijom, u najvećoj meri pomoću fitohormona)
ekstracelularni nivo regulacije (podrazumeva uticaj faktora spoljašnje sredine, kao signala, na modifikaciju razvića biljke, tako da se ona funkcionalno prilagodi uslovima spoljašnje sredine)
Nivoi regulacije razvića biljke su međusobno tesno povezani. Kada se svi oni uzmu u obzir, funkcionisanje biljke se može opisati na sledeći način: biljka se prostorno i vremenski orijentiše u sredini u kojoj živi tako što prima fizičke signale iz sredine, koji se u biljci „prevode“ u hemijske faktore, najčešće fitohormone, koji na intracelularnom nivou menjaju ekspresiju specifičnih gena, sintezu i aktivnost specifičnih enzima, što menja razviće biljke tako da ona funkcioniše usklađeno sa spoljašnjom sredinom.
Ćelijski ciklus
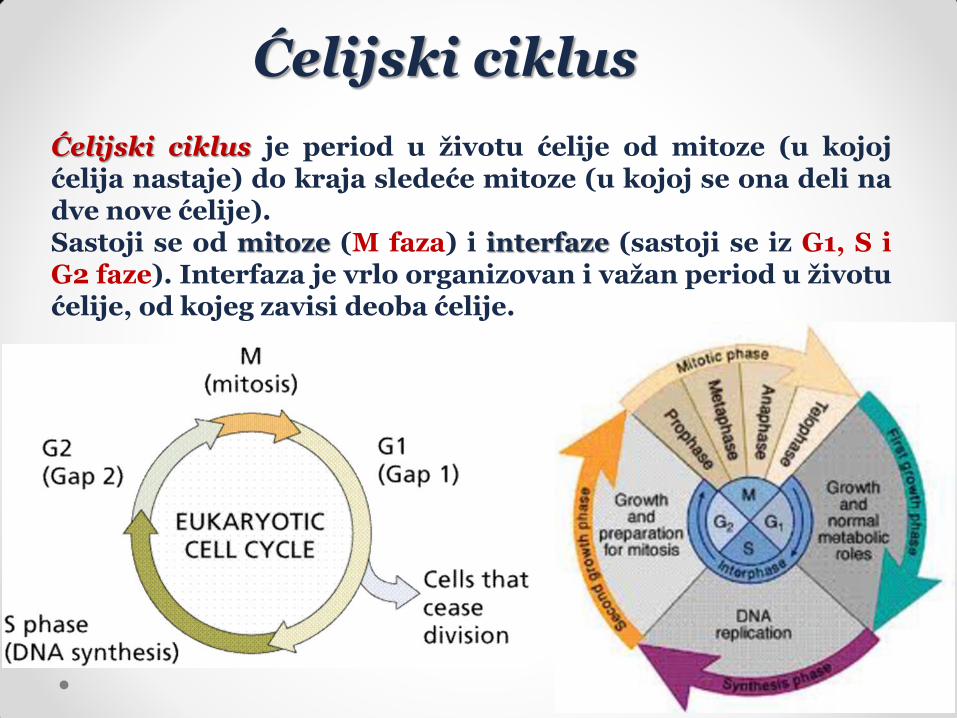
Ćelijski ciklus je period u životu ćelije od mitoze (u kojoj ćelija nastaje) do kraja sledeće mitoze (u kojoj se ona deli na dve nove ćelije). Sastoji se od mitoze (M faza) i interfaze (sastoji se iz G1, S i G2 faze). Interfaza je vrlo organizovan i važan period u životu ćelije, od kojeg zavisi deoba ćelije.

U G1 fazi se dešava priprema ćelije za S fazu, tj. sinteza nukleotida i enzima koji učestvuju u replikaciji DNK i disanju, kao i rastenje ćelije (veličina ćelije se povećava dva puta) i povećanje broja organela (proplastida, promitohondrija i membranskih struktura). U S fazi se dešava replikacija DNK i njena količina se povećava tačno dva puta. Osim toga, sintetišu se histoni i drugi nuklearni enzimi i proteini. U G2 fazi se ćelija priprema za mitozu, što podrazumeva sintezu proteina i drugih molekula koji u mitozi učestvuju. RNK se sintetiše u svim fazama ćelijskog ciklusa osim u mitozi. Tokom ćelijskog ciklusa aktivni su geni koji nisu vezani samo za neku fazu ciklusa, ali se aktiviraju i geni ćelijske deobe – cdc geni („cell division cycle genes“). Ćelije koje prestaju da se dele izlaze iz ćelijskog ciklusa tokom G1 faze (češće) ili tokom G2 faze (ređe). Kada se izlazak iz ćelijskog ciklusa desi tokom G2 faze, ćelija sadrži dvostuku količinu DNK, odnosno postaje poliploidna. Kada ćelije izađu iz ćelijskog ciklusa, ulaze u G0 fazu, koja može trajati do kraja života ćelije ili se iz nje ćelije, posle određenog vremena, mogu vratiti u fazu ciklusa u kojoj su iz njega izašle (G1 ili G2 faza) i nastaviti sa deobama. Trajanje ćelijskog ciklusa varira i zavisi od tipa ćelija. U meristemu korena ćelijski ciklus traje 20 sati, a u meristemu stabla 2-3 dana. Relativna dužina pojedinih faza ćelijskog ciklusa je uglavnom stalna.

Za ćelijski ciklus je karakteristična „vernost“ jednom planu i redosledu događaja. Ako neki proces nije kompletiran, ćelija ne ulazi u sledeću fazu ciklusa. To naročito važi za M fazu – deoba ćelije se neće desiti ako se ne kompletiraju sve faze ćelijskog ciklusa koje joj prethode. Ćelije se uvek zaustavljaju na jednom od dva mesta u ćelijakom ciklusu, koja se nazivaju kontrolnim mestima, i to su: 1. prelaz iz G1 u S fazu (G1→S) 2. prelaz iz G2 u M fazu (G2→M) Prolazi kroz kontrolna mesta su najvažniji događaji u ćelijskom ciklusu, zbog čega se regulacija ovog ciklusa obavlja baš na tim mestima. Na prvom kontrolnom mestu (nalazi se u kasnoj G1 fazi) ćelija treba da „odluči“ da li će se deliti ili će ući u G0 fazu. To „odlučivanje“ zavisi zapravo od veličine ćelije (ona treba da bude dvaput veća nego ona koju je imala posle mitoze u kojoj je nastala) i od stimulusa koji aktiviraju pripremne procese i omogućavaju da ćelija prođe kroz prvo kontrolno mesto. Ukoliko ćelija ne dostigne potrebnu veličinu i ukoliko izostanu potrebni stimulusi, ćelija ulazi u G0 fazu. Na drugom kontrolnom mestu ćelija „odlučuje“ ne da li će, nego kada će ući u mitozu. Ulazak u mitozu se zadržava na ovom mestu sve dok ne stigne signal da je replikacija DNK obavljena i da su prisutni svi činioci koji su potrebni za mitozu.

Način regulacije ćelijskog ciklusa je različit na dva kontrolna mesta: na prvom kontrolnom mestu odlučujuća je veličina ćelije i potreban je spoljašnji signal (van ćelijskog ciklusa), a na drugom kontrolnom mestu je odlučujuć signal koji potiče iz prethodno obavljene S faze i drugih procesa ćelijskog ciklusa. Faktor koji podstiče mitozu, MPF („mitosis promoting factor“) je neophodan za ćelijsku deobu (prelaz G2→M). Sastoji se iz dve komponente. Prva komponenta je protein od 34 kD (p34). Druga komponenta je protein nazvan ciklin (zato što njegova količina varira tokom ćelijskog ciklusa - nema ga odmah posle mitoze, povećava se tokom interfaze i dostiže maksimum na prelazu, da bi u anafazi deobe naglo nestao). Oba proteina kodiraju cdc geni. Kompleks p34 i ciklina (ali ne i njegove pojedinačne komponente) funkcioniše kao enzim proteinska kinaza. U tom kompleksu ciklin je regulatorna subjedinica kompleksa - određuje specifičnost za supstrat (protein koji se fosforiluje uz utrošak ATP), vreme aktivnosti i subcelularnu lokalizaciju aktivnosti, dok p34 ima kinaznu (enzimsku) aktivnost. Pošto p34 ima kinaznu aktivnost samo kad je u kompleksu sa ciklinom, on se još naziva i kinaza zavisna od ciklina, CDK („cyclin dependent kinase“). Smatra se da je kod svih eukariota kompleks ciklin/CDK regulatorni mehanizam koji kontroliše procese u ćelijskom ciklusu.

Postoji više tipova ciklina i više tipova CDK. Kod biljaka se razlikuju ciklini A, B i D (CycA, CycB i CycD), a CDK ima 5-6 tipova, od kojih je CDK A najbolje opisana i aktivna tokom S, G2 i M faze. Različiti ciklini nisu prisutni u ćeliji stalno (nestabilna su jedinjenja), već su karakteristični za za određene momente ćelijskog ciklusa – sintetišu se u specifičnim momentima, obavljaju svoju funkciju i onda brzo degraduju. Prema tome, sinteza i degradacija ciklina učestvuju u regulaciji aktivnosti CDK, ali nisu jedini regulatorni faktor. Aktivnost ciklin/CDK kompleksa zavisi od velikog broja činilaca, koji opet zavise od spoljašnjih faktora i razvojnog programa ćelije i tkiva (unutrašnji faktor). Kinaznu aktivnost CDK u toku ćelijskog ciklusa regulišu: prisustvo određenog ciklina, za koji se CDK vezuje fosforilacija i defosforilacija aminokiselina koje se nalaze na
specifičnim mestima proteina CDK prisustvo inhibitornih proteina koji se vezuju za CDK ili
inaktiviraju neke druge bitne komponente
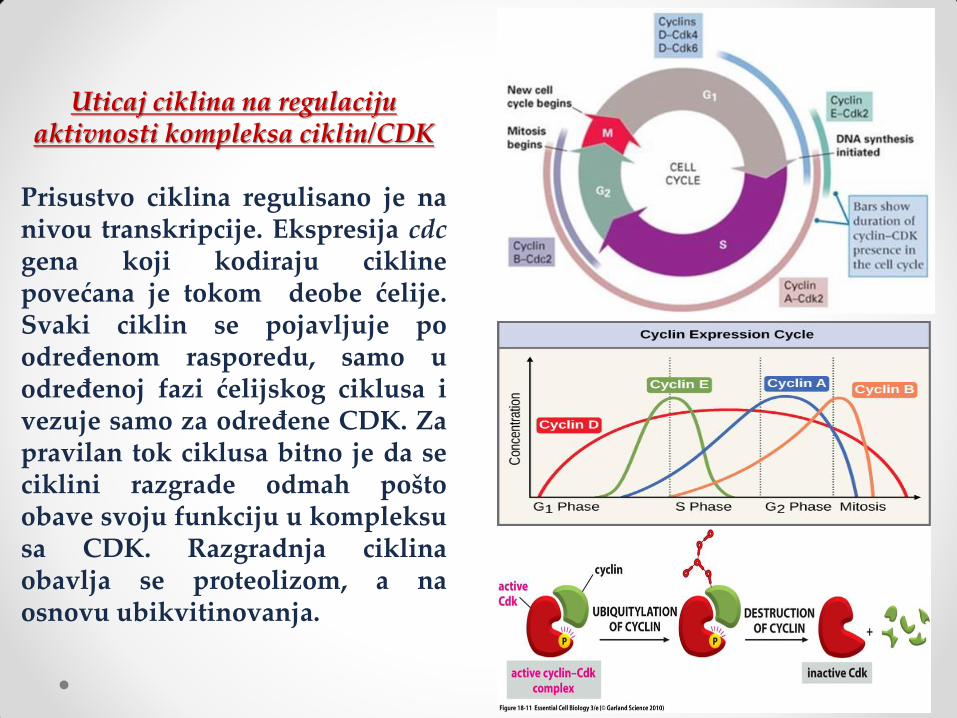
Uticaj ciklina na regulaciju aktivnosti kompleksa ciklin/CDK
Prisustvo ciklina regulisano je na nivou transkripcije. Ekspresija cdc gena koji kodiraju cikline povećana je tokom deobe ćelije. Svaki ciklin se pojavljuje po određenom rasporedu, samo u određenoj fazi ćelijskog ciklusa i vezuje samo za određene CDK. Za pravilan tok ciklusa bitno je da se ciklini razgrade odmah pošto obave svoju funkciju u kompleksu sa CDK. Razgradnja ciklina obavlja se proteolizom, a na osnovu ubikvitinovanja.

Uticaj fosforilacije i defosforilacije na aktivnost kompleksa ciklin/CDK
CDK se ubrajaju u serin/treonin kinaze i nisu aktivne odmah po formiranju kompleksa sa ciklinom. Uslov za aktivnost su fosforilacija i defosforilacija na određenim aminokiselinama CDK proteina. Za aktivnost kompleksa koji učestvuje u S/G2 fazi neophodna je fosforilacija treonina 160 (mesto pozitivne fosforilacije). Kompleks se inaktivira fosforilacijom treonina 14 i tirozina 15 (mesta negativne fosforilacije). Vezivanje fosfata za mesta negativne fosforilacije sprečava vezivanje ATP, a time i kinaznu aktivnost CDK. Fosforilaciju CDK obavljaju druge serin/treonin kinaze, koje odgovaraju na delovanje mitogena (ekstracelularni faktori koji indukuju mitozu), pa se nazivaju kinaze indukovane mitogenima, MAPK („mitogen activated protein kinases“). I MAPK se aktiviraju fosforilacijom od strane MAPKK. Između mitogena i krajnje MAPK postoji kaskada kinaza koje se uzastopno aktiviraju. MAP kinaze nisu ograničene na ćelijski ciklus, već učestvuju u transdukciji signala i u drugim važnim procesima u ćeliji. One su, međutim, te koje uvek prenose spoljašnji signal na sistem u ćeliji koji reaguje na signal. U ovoj vrsti regulacije ćelijskog ciklusa učestvuju i fosfataze, koje defosforilišu, i tako inaktiviraju, kinaze.
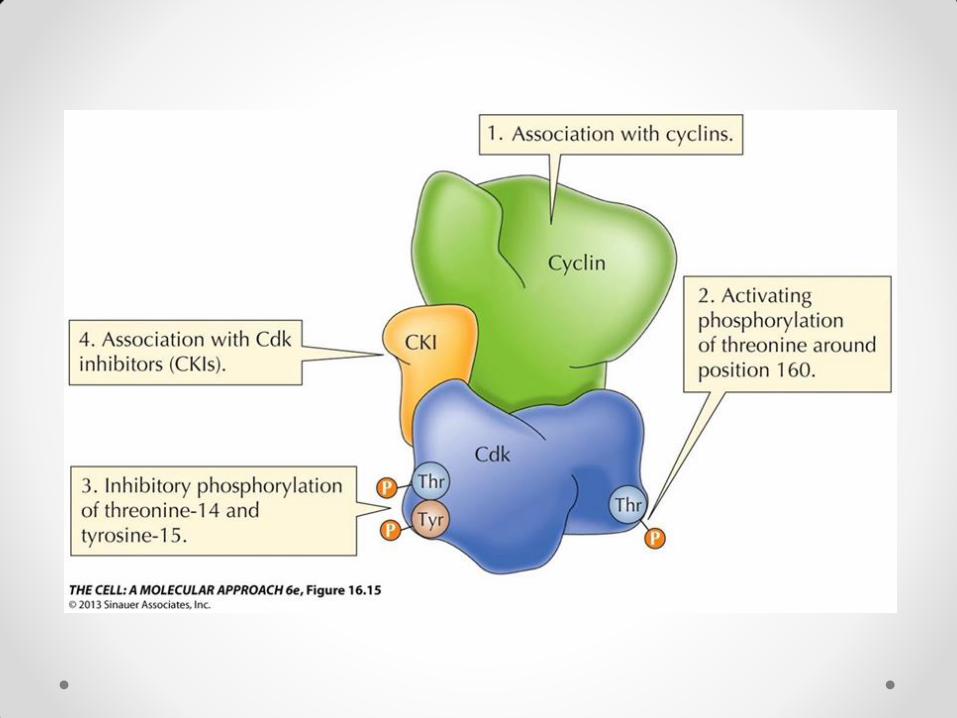

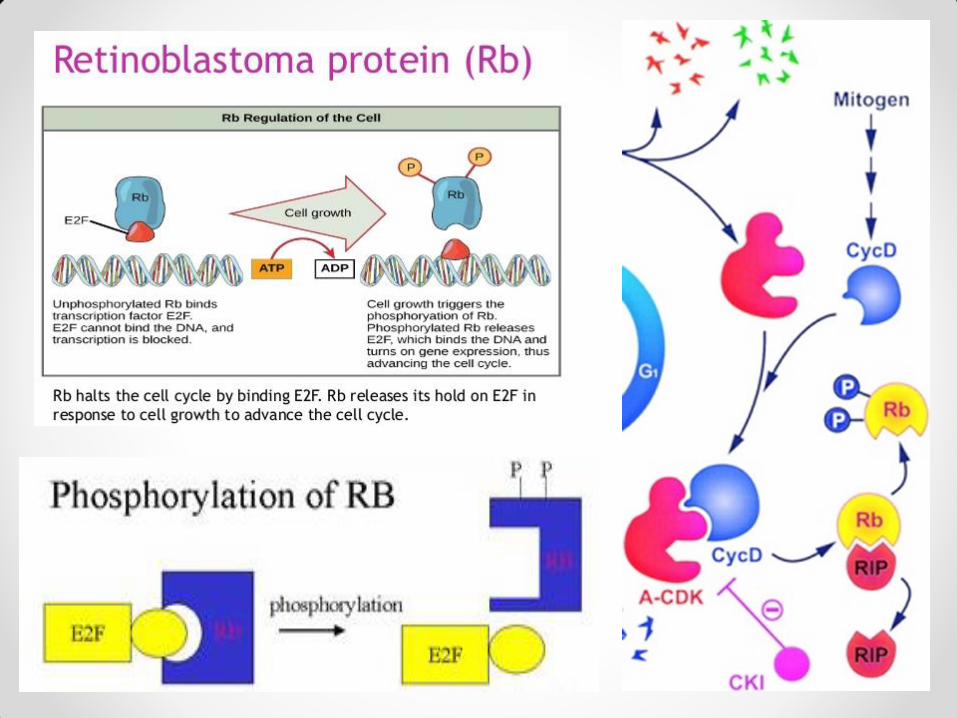
Uticaj inhibitora i represora na ćelijski ciklus Inhibitori koji se vezuju za kompleks ciklin/CDK sprečavaju dejstvo kompleksa i tako utiču na ćelijski ciklus. Biljni inhibitori aktivnosti kompleksa ciklin/CDK su ICK1 i ICK2, koji vezivanjem za kompleks sprečavaju kinaznu aktivnost. Pod dejstvom tih inhibitora dolazi do redukcije ćelijskih deoba i inhibicije rastenja kod biljaka. U represore ćelijskog ciklusa ubraja se retinoblastoma protein (Rb), koji se vezuje za transkripcioni faktor E2F i time ga inaktivira. Rezultat je izostanak replikacije DNK u S fazi i zaustavljanje ćelijskih deoba. Ciklin D3 , koji je u kompleksu sa odgovarajućom CDK, prepoznaje Rb, vezuje ga za kompleks usled čega ga CDK fosforiliše. Rb može da veže više fosfatnih grupa (hiperfosforilovano stanje) i u tom stanju otpušta E2F, koji se vezuje za DNK, dovodi do ekspresije određenih gena i omogućava sintezu DNK u S fazi, čime se ćelijski ciklus nastavlja. Rb je u G0 i G1 fazi hipofosforilovan i deluje kao represor, usled čega nema mitoze. U G1 fazi njegova fosforilacija raste i u jednom momentu Rb otpušta E2F, što dovodi do prelaska ćelije u S fazu. U diferenciranim ćelijama hipofosforilovani Rb je u velikoj količini, pa se smatra da on održava diferencirane ćelije u G0 fazi i sprečava njihovu deobu.
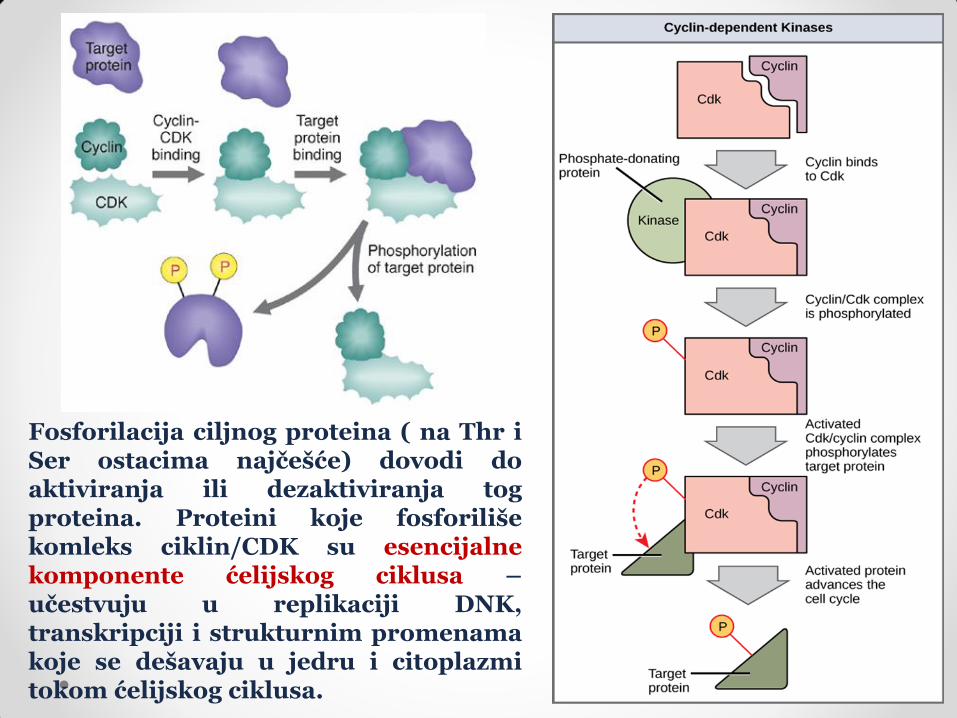
Fosforilacija ciljnog proteina ( na Thr i Ser ostacima najčešće) dovodi do aktiviranja ili dezaktiviranja tog proteina. Proteini koje fosforiliše komleks ciklin/CDK su esencijalne komponente ćelijskog ciklusa – učestvuju u replikaciji DNK, transkripciji i strukturnim promenama koje se dešavaju u jedru i citoplazmi tokom ćelijskog ciklusa.
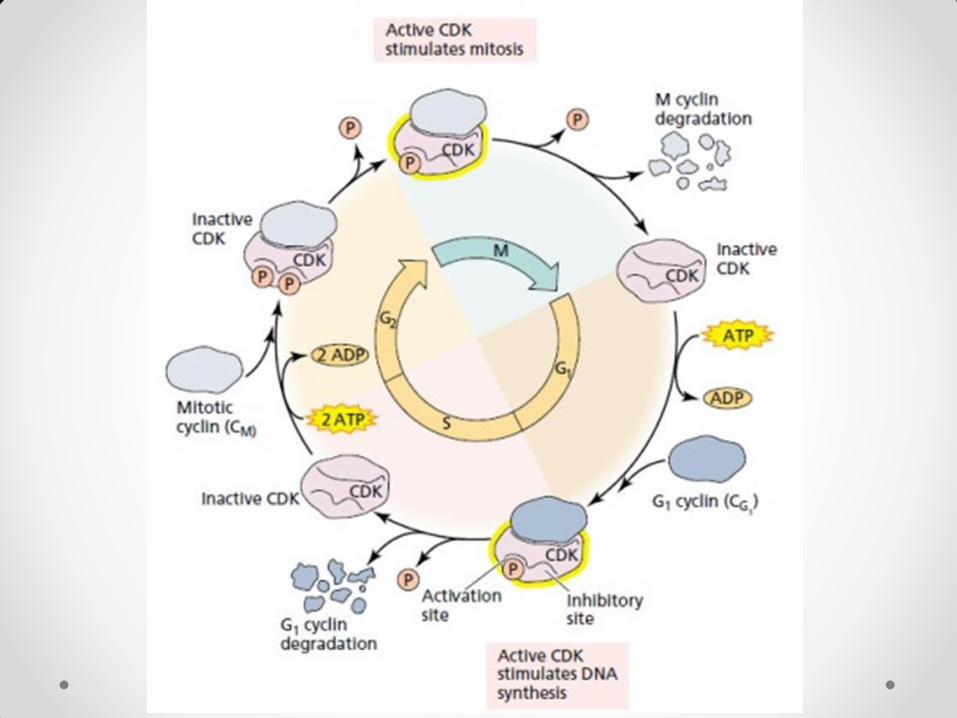

Hormonalna kontrola ćelijskog ciklusa Kod biljaka su mitogeni različiti faktori iz spoljašnje (stresni faktori, napr.) i unutrašnje sredine. Jedan od biljnih mitogena je saharoza, napr. Međutim, najbitniji mitogeni su fitohormoni. Auksini i citokinini su neophodni za indukciju ćelijskih deoba. Oni deluju u kooperaciji – ukoliko nema jednog od njih, ćelijske deobe se ne dešavaju. Auksini su neophodni na dva mesta u ćelijskom ciklusu – na kraju G1 i na kraju G2 faze, za procese u kojima učestvuje CDK, produkt cdc2 gena. Oni stimulišu sintezu CDK proteina, tako što indukuju ekspresiju gena za CDK. Citokinini deluju takođe na dva mesta u ćelijskom ciklusu. Na prelazu G2→M, gde aktiviraju neaktivnu CDK tako što indukuju sintezu fosfataza koje defosforiluju CDK (koja je već u kompleksu sa ciklinom) na mestu tirozina 15. Druga funkcija citokinina je indukcija ekspresije gena za ciklin D3 na prelazu G0→ G1, čime mirujuće ćelije vraćaju u ćelijski ciklus i vode
ih u deobu. Malo se zna o efektu drugih grupa fitohormona na ćelijski ciklus, ali je sigurno da oni nemaju ključnu ulogu u njegovoj regulaciji. Iako ostali fitohormoni evidentno utiču na ćelijski ciklus, ne zna se da li je njihov efekat direktan ili posredan. Giberelini direktno indukuju sintezu nekih ciklina u submerznim internodijama pirinča, a brasinosteroidi pojačavaju sintezu ciklina D3, a ABA stimuliše ekspresiju gena za ICK1 inhibitor.
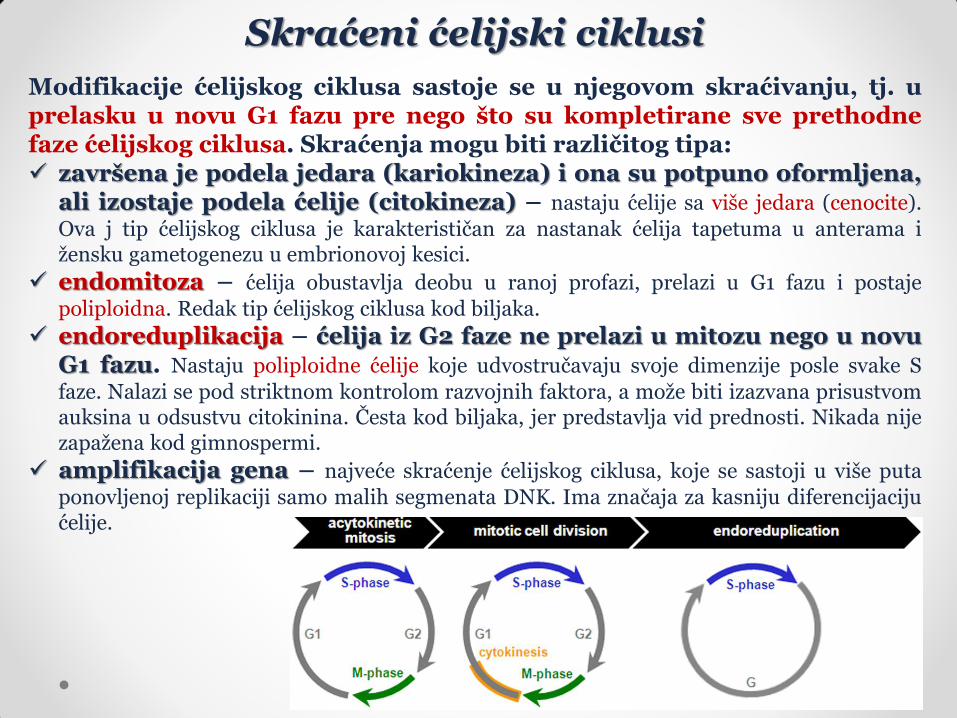
Skraćeni ćelijski ciklusi
Modifikacije ćelijskog ciklusa sastoje se u njegovom skraćivanju, tj. u prelasku u novu G1 fazu pre nego što su kompletirane sve prethodne faze ćelijskog ciklusa. Skraćenja mogu biti različitog tipa: završena je podela jedara (kariokineza) i ona su potpuno oformljena,
ali izostaje podela ćelije (citokineza) – nastaju ćelije sa više jedara (cenocite).
Ova j tip ćelijskog ciklusa je karakterističan za nastanak ćelija tapetuma u anterama i žensku gametogenezu u embrionovoj kesici.
endomitoza – ćelija obustavlja deobu u ranoj profazi, prelazi u G1 fazu i postaje
poliploidna. Redak tip ćelijskog ciklusa kod biljaka.
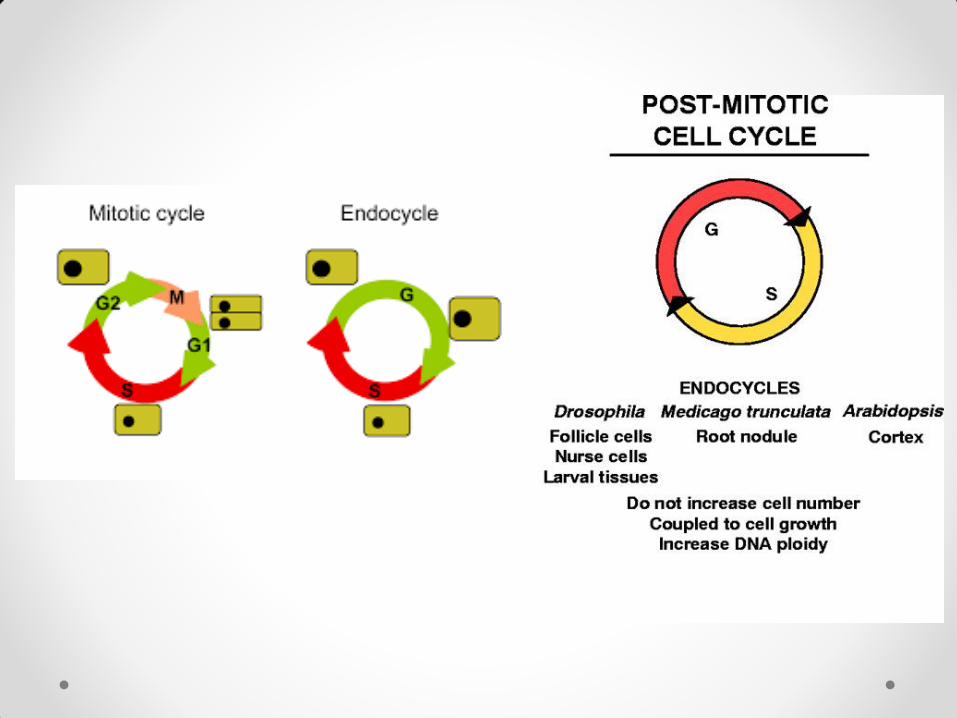
endoreduplikacija – ćelija iz G2 faze ne prelazi u mitozu nego u novu G1 fazu. Nastaju poliploidne ćelije koje udvostručavaju svoje dimenzije posle svake S
faze. Nalazi se pod striktnom kontrolom razvojnih faktora, a može biti izazvana prisustvom auksina u odsustvu citokinina. Česta kod biljaka, jer predstavlja vid prednosti. Nikada nije zapažena kod gimnospermi.
amplifikacija gena – najveće skraćenje ćelijskog ciklusa, koje se sastoji u više puta
ponovljenoj replikaciji samo malih segmenata DNK. Ima značaja za kasniju diferencijaciju ćelije.

Karakteristike rastenja biljaka
• Rastenje je proces ireverzibilnog povećanja dimenzija organa ili biljnog organizma. U osnovi rastenja mogu biti povećanje: broja ćelija, sveže ili suve mase, zapremine, površine i/ili dimenzija (dužine, obima). NIJE svako povećanje navedenih parametara rastenje, već samo ireverzibilno povećanje.
• Rastenje može da se ostvari i kombinacijom povećanja nabrojanih parametara. Deobe ćelija, na osnovu kojih se povećava broj ćelija, nisu same po sebi dovoljne da dovedu do rastenja.
• U rastenje se ne ubraja promena zapremine usled imbibicije ili osmoze. S druge strane, rastenje može da se ostvari bez vidljivog povećanja mase ili dimenzija.
• Ne doprinose podjednako sve ćelije jednog organizma njegovom rastenju.
• Rastenje biljaka je lokalizovano na meristemske zone, u kojima ćelije tokom celog života biljke zadržavaju sposobnost deobe.
• Posle deobe, ćelije-ćerke rastu i to je jedini način na koji biljke rastu, s obzirom na to da su ćelije biljnog organizma nepokretne, fiksirane apoplastom.

• Meristemi se dele na primarne i sekundarne meristeme.
• Primarni meristemi su apikalni meristemi korena i izdanka i primarni kambijum (oni se formiraju u embrionalnom razviću), a po nekim podelama i interkalarni i aksilarni meristemi („zaostali primarni meristemi“).
• Sekundarni meristemi su sekundarni kambijum, meristemi bočnih korenova i felogen (meristem kore), a po drugim podelama u sekundarne meristeme se ubrajaju i interkalarni i aksilarni meristemi. Za sve sekundarne meristeme je karakteristično da nastaju posle embrionalnog razvića, u unutrašnjim tkivima biljke, tako što se dešava dediferencijacija ćelija tih tkiva.
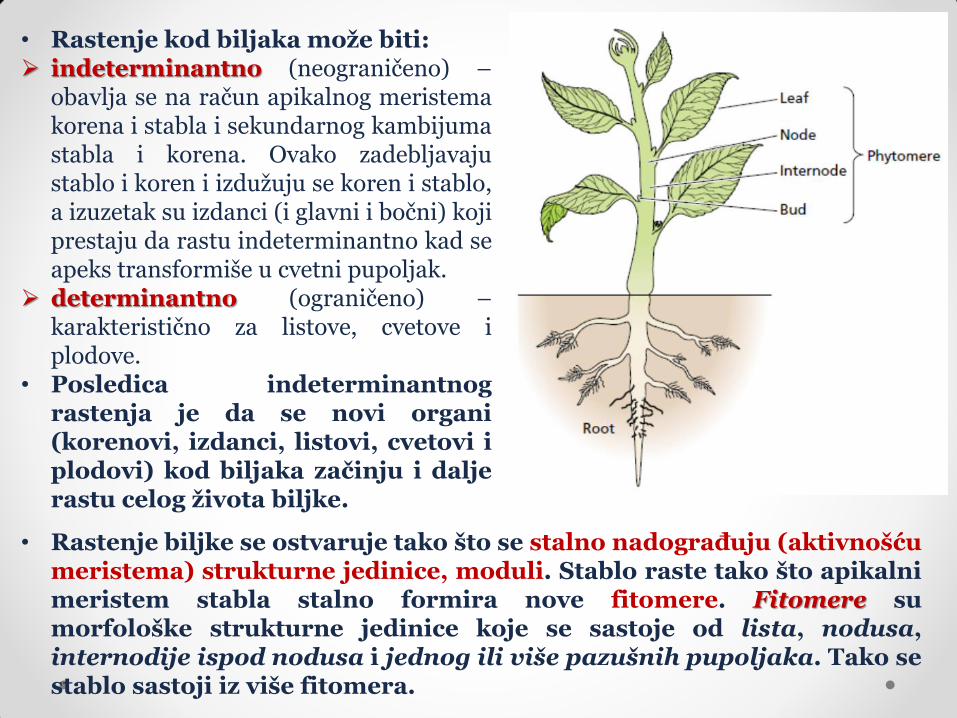
• Rastenje kod biljaka može biti: indeterminantno (neograničeno) –
obavlja se na račun apikalnog meristema korena i stabla i sekundarnog kambijuma stabla i korena. Ovako zadebljavaju stablo i koren i izdužuju se koren i stablo, a izuzetak su izdanci (i glavni i bočni) koji prestaju da rastu indeterminantno kad se apeks transformiše u cvetni pupoljak.
determinantno (ograničeno) – karakteristično za listove, cvetove i plodove.
• Posledica indeterminantnog rastenja je da se novi organi (korenovi, izdanci, listovi, cvetovi i plodovi) kod biljaka začinju i dalje rastu celog života biljke.
• Rastenje biljke se ostvaruje tako što se stalno nadograđuju (aktivnošću meristema) strukturne jedinice, moduli. Stablo raste tako što apikalni meristem stabla stalno formira nove fitomere. Fitomere su morfološke strukturne jedinice koje se sastoje od lista, nodusa, internodije ispod nodusa i jednog ili više pazušnih pupoljaka. Tako se stablo sastoji iz više fitomera.
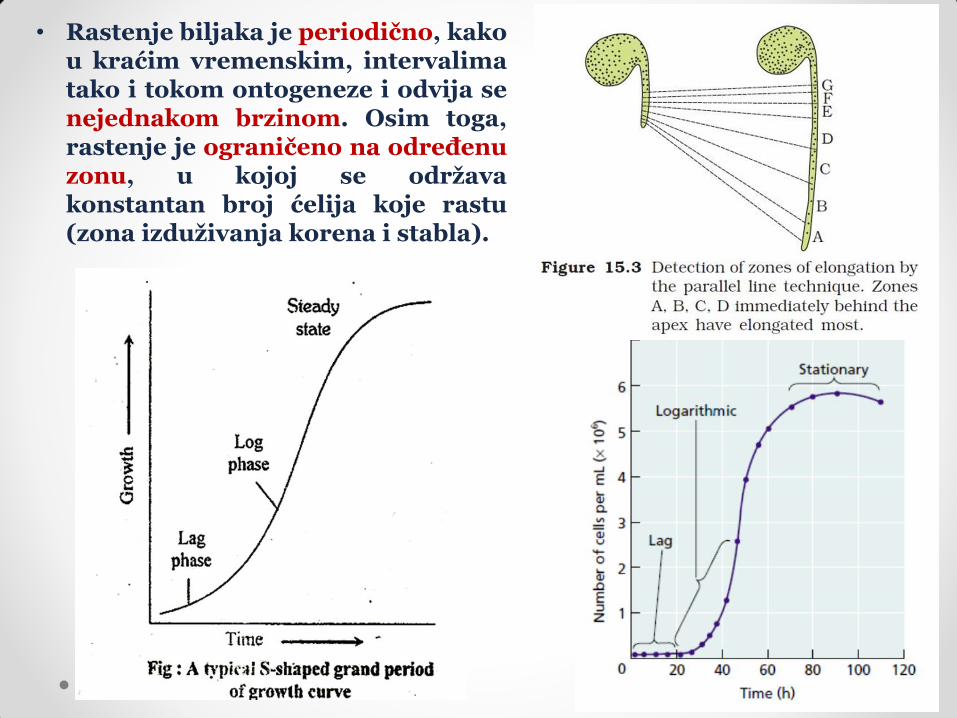
• Rastenje biljaka je periodično, kako u kraćim vremenskim, intervalima tako i tokom ontogeneze i odvija se nejednakom brzinom. Osim toga, rastenje je ograničeno na određenu zonu, u kojoj se održava konstantan broj ćelija koje rastu (zona izduživanja korena i stabla).

Brzina rastenja u funkciji udaljenosti ćelije od vrha
organa koji raste.
Prirast brzine rastenja u funkciji udaljenosti ćelije od vrha organa
koji raste.
Grafici brzine i prirasta brzine rastenja u funkciji položaja ćelije u organu daju informacije o regionima biljke u kojima je rastenje najintenzivnije.

Ćelijska osnova morfogeneze biljaka
• Na oblik i veličinu organa (morfogenezu) biljnih organizama utiču procesi rastenja i diferencijacije ćelija.
• Broj ćelija u nekom organu biljke nema presudni uticaj na veličinu i oblik organa, jer su meristemske ćelije sitne i njihov broj ne doprinosi značajno rastenju organa sve dok ćelije ne počnu da rastu.
• Međutim, na oblik organa, tj. na njegovo razviće bitno utiču položaj deobnih vretena u mitozi i pravac rastenja pojedinačnih ćelija. Osim toga, ćelije koje ulaze u sastav organa diferenciraju se u različitim pravcima.
• Pravac diferencijacije ćelija određen je još u momentu deobe ćelija i zavisi od toga da li su deobe inekvalne ili ekvalne (simetrične), zatim od položaja deobnog vretena, kao i od položaja ćelije u odnosu na susedne ćelije u tkivu.
• Pod diferencijacijom se podrazumeva postizanje kvalitativnih razlika (metaboličkih, strukturnih i funkcionalnih) među ćelijama poreklom od iste ćelije ili grupe ćelija, odnosno specijalizacija za obavljenje različitih funkcija. U užem smislu, to je razvoj morfoloških i fizioloških osobina po kojima se ćelije razlikuju. Ove, vidljive, osobine zasnivaju se na promenama na biohemijskom i molekularnom nivou, koje nisu uočljive, ali određuju dalju sudbinu ćelija.

Diferencijacija ćelija kod biljaka je često reverzibilna, naročito kada se ćelije gaje u kulturi in vitro, izolovane iz biljnog organizma. To znači da biljne ćelije mogu da se dediferenciraju (da izgube osobine nastale diferencijacijom), da ponovo steknu sposobnost deobe (koja se kod veoma diferenciranih ćelija gubi) i da se rediferenciraju u različitim pravcima (ukoliko su im dostupni određeni hormonski i nutritivni faktori), što u određenim slučajevima može da dovede do regeneracije cele biljke (procesom somatske embriogeneze, napr.). Sposobnost biljnih ćelija da se dediferenciraju ukazuje na to da diferencirane ćelije zadržavaju sve neophodne genetičke informacije neophodne za razviće cele biljke. Jedini izuzeci su ćelije biljnog organizma koje tokom razvića gube jedro (sitaste cevi u floemu) i ćelije koje na kraju razvića više nisu žive (traheje i traheide u ksilemu). Zbog opisane sposobnosti za dediferencijaciju i rediferencijaciju, biljne ćelije se smatraju totipotentnim, iako se taj termin može primeniti samo na zigot (zato što u praksi samo zigot može diferencijacijom ćelija koje nastaju njegovom deobom da dâ bilo koje biljno tkivo, što nije uvek slučaj sa somatskim ćelijama), pa se u praksi (kultura biljaka in vitro) češće koriste termini „pluripotentnost“ i „multipotentnost“. Da li će genetičke informacije koje poseduju somatske, diferencirane ćelije dovesti do regeneracije celog organizma, to zavisi od kontrole ekspresije specifičnih gena, na šta utiče veći broj spoljašnjih i unutrašnjih faktora.

Procesi u prvim fazama diferencijacije koji određuju dalju sudbinu ćelije označavaju se kao determinacija. Posle procesa determinacije ćelije nastale deobom istih meristemskih ćelija počinju uočljivo da se razlikuju. Te razlike obuhvataju: veličinu i oblik ćelija (parenhimske i prozenhimske ćelije), građu i sastav sekundarnog ćelijskog zida, broj specifičnih organela (hloroplasta napr.), sadržaj hemijskih jedinjenja (pigmenata napr.) itd.
Pravac ćelijske deobe utiče na
oblik organa. Određuju ga položaj deobnog vretena (obično je paralelno dužoj osi majke-ćelije) i ćelijske ploče (nalazi se pod pravim uglom u odnosu na osu deobnog vretena). Položaj ćelijske ploče, koja se formira u anafazi i telofazi, određen je prstenom mikrotubula po obodu ćelije – preprofazna traka.
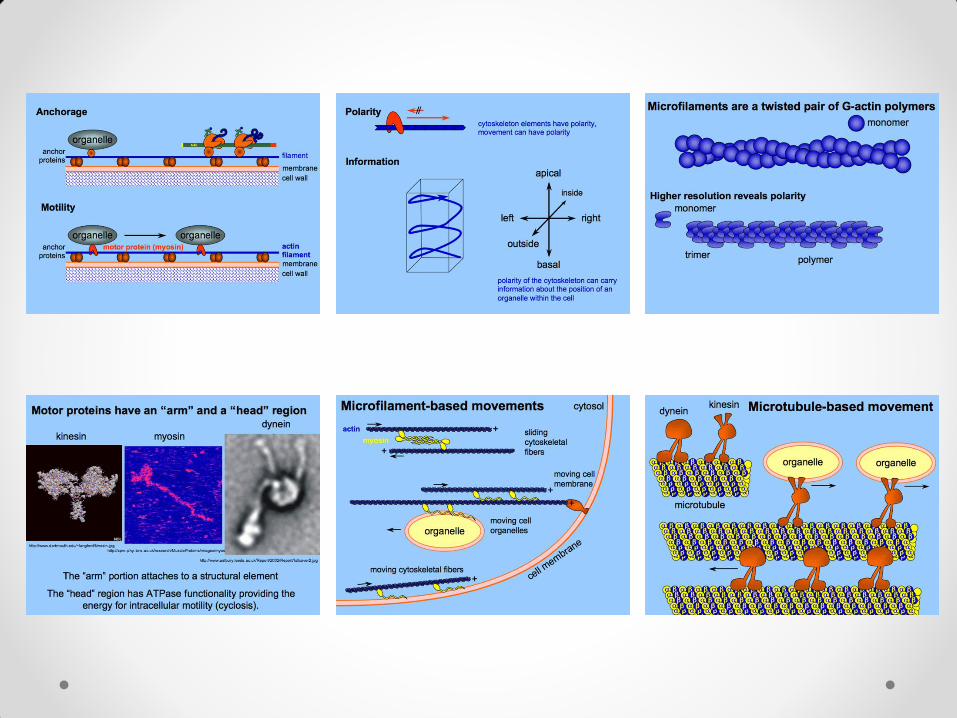
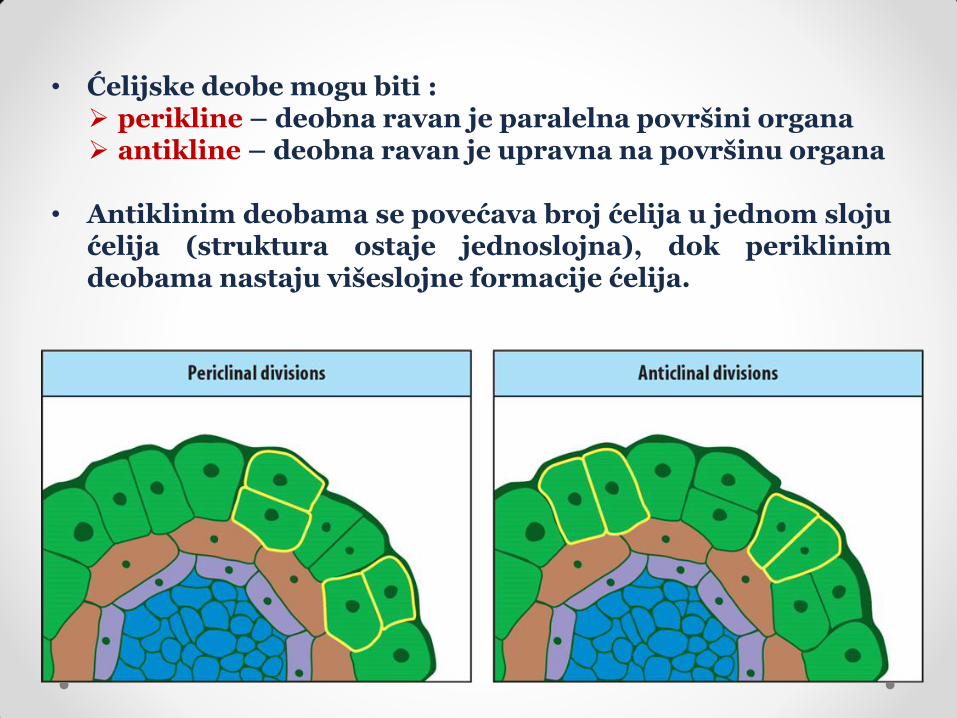
• Ćelijske deobe mogu biti : perikline – deobna ravan je paralelna površini organa antikline – deobna ravan je upravna na površinu organa
• Antiklinim deobama se povećava broj ćelija u jednom sloju
ćelija (struktura ostaje jednoslojna), dok periklinim deobama nastaju višeslojne formacije ćelija.
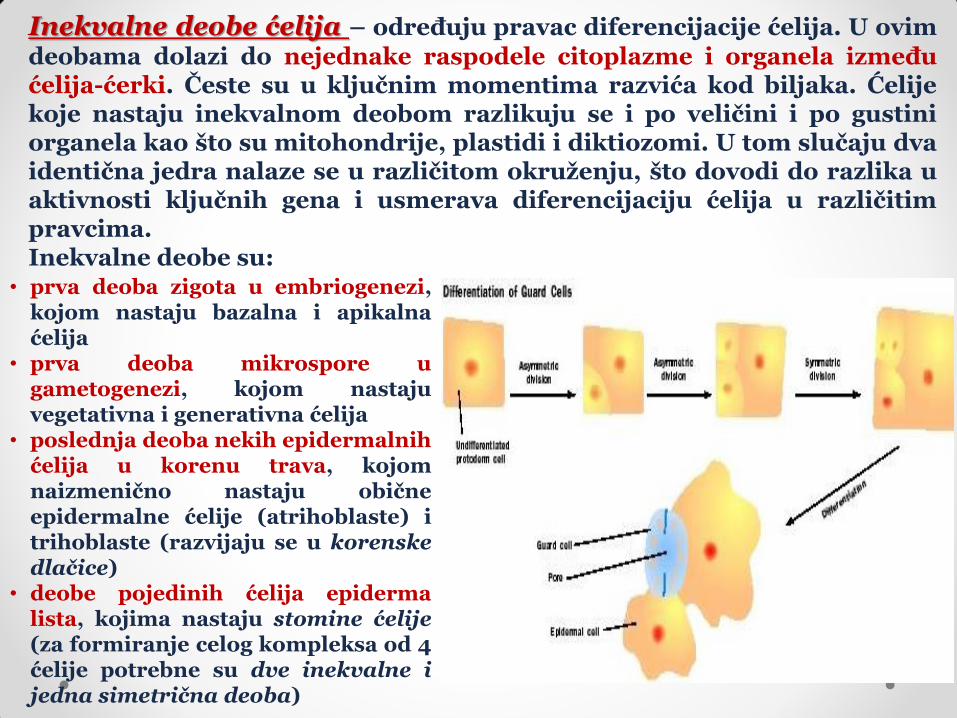
Inekvalne deobe ćelija – određuju pravac diferencijacije ćelija. U ovim
deobama dolazi do nejednake raspodele citoplazme i organela između ćelija-ćerki. Česte su u ključnim momentima razvića kod biljaka. Ćelije koje nastaju inekvalnom deobom razlikuju se i po veličini i po gustini organela kao što su mitohondrije, plastidi i diktiozomi. U tom slučaju dva identična jedra nalaze se u različitom okruženju, što dovodi do razlika u aktivnosti ključnih gena i usmerava diferencijaciju ćelija u različitim pravcima. Inekvalne deobe su:
• prva deoba zigota u embriogenezi, kojom nastaju bazalna i apikalna ćelija
• prva deoba mikrospore u gametogenezi, kojom nastaju vegetativna i generativna ćelija
• poslednja deoba nekih epidermalnih ćelija u korenu trava, kojom naizmenično nastaju obične epidermalne ćelije (atrihoblaste) i trihoblaste (razvijaju se u korenske dlačice)
• deobe pojedinih ćelija epiderma lista, kojima nastaju stomine ćelije (za formiranje celog kompleksa od 4 ćelije potrebne su dve inekvalne i jedna simetrična deoba)

Pravac rastenja ćelija – ćelije rastu u onom pravcu u kojem
ćelijski zid pruža najmanji otpor. Koji je to pravac, određuje raspored, orijentacija mikrofibrila u primarnom ćelijskom zidu. Ako su mikrofibrile orijentisane u svim pravcima ravnomerno, ćelija raste izodijametrično (povećava dimenzije u svim pravcima). Ukoliko su mikrofibrile pretežno orijentisane paralelno dužoj osi ćelija i organa (longitudinalno), dešava se zadebljavanje (povećanje obima ćelija i organa), dok pretežno transverzalna orijentacija mikrofibrila (pod uglom od 90 ° u odnosu na dužu osu ćelije ili organa) dovodi do izduživanja. Orijentacija mikrofibrila u ćelijskom zidu određena je orijentacijom mikrotubula u perifernoj citoplazmi ćelije, a na njihovu orijentaciju imaju efekta fitohormoni (giberelini i auksini u prvom redu, ali i citokinini, ABA i etilen).
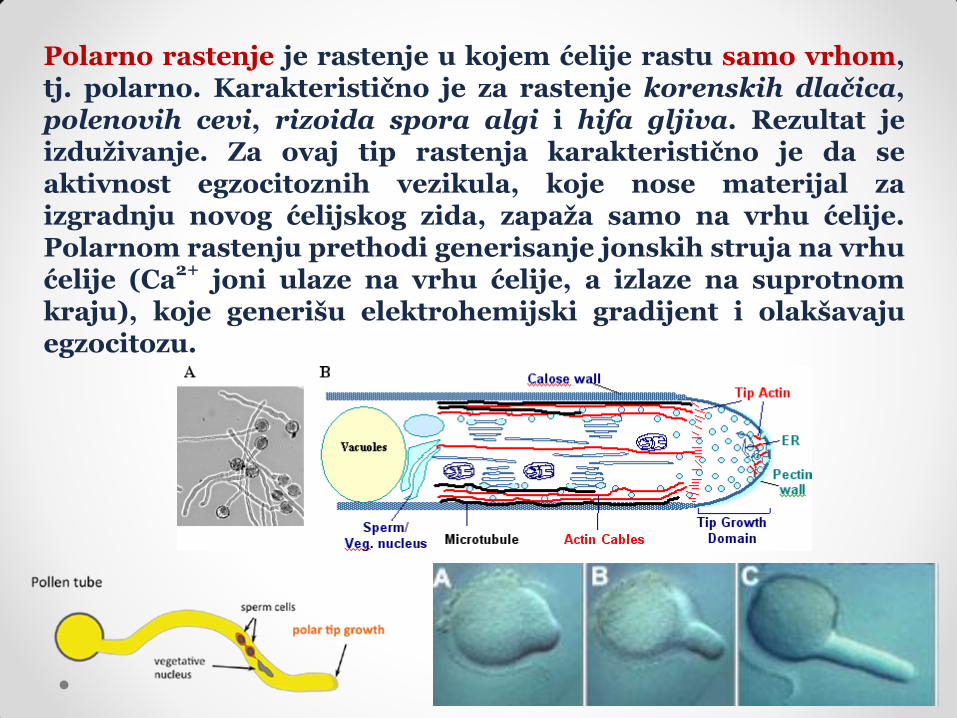
Polarno rastenje je rastenje u kojem ćelije rastu samo vrhom, tj. polarno. Karakteristično je za rastenje korenskih dlačica, polenovih cevi, rizoida spora algi i hifa gljiva. Rezultat je izduživanje. Za ovaj tip rastenja karakteristično je da se aktivnost egzocitoznih vezikula, koje nose materijal za izgradnju novog ćelijskog zida, zapaža samo na vrhu ćelije. Polarnom rastenju prethodi generisanje jonskih struja na vrhu ćelije (Ca
2+ joni ulaze na vrhu ćelije, a izlaze na suprotnom
kraju), koje generišu elektrohemijski gradijent i olakšavaju egzocitozu.

Pozicioni efekat u diferencijaciji ćelija – u toku razvića ćelije su
uglavnom predodređene svojim poreklom od određenih inicijala i položajem deobnih ravni da se diferenciraju u određenom pravcu. Međutim, u ranim fazama razvića ćelije mogu da se razvijaju u određenom pravcu ne na osnovu svog porekla, nego na osnovu svog mesta u slojevima, pa tako, ukoliko se nađu u drugom sloju od onog u kojem se obično nalaze (zbog drugačije ravni deobe), mogu da se diferenciraju drugačije nego što određuje njihovo poreklo. U tom slučaju njihovu sudbinu određuje mesto gde se nalaze, a ne njihovo poreklo. U kasnijim fazama razvića, kad ćelije dostignu zrelost, gubi se ova razvojna plastičnost.

Polarnost Polarnost je specifična morfološka i fiziološka raspodela aktivnosti na
dva kraja neke osovine (ose simetrije). Zavisno od ose simetrije, polarnost može biti: aksijalna (po uzdužnoj osi organizma) – generiše apikalno-bazalni
obrazac razvića; karakteristična je za glavni izdanak, ali i za aksilarne (bočne) organe, kao što su bočni izdanci i bočni korenovi
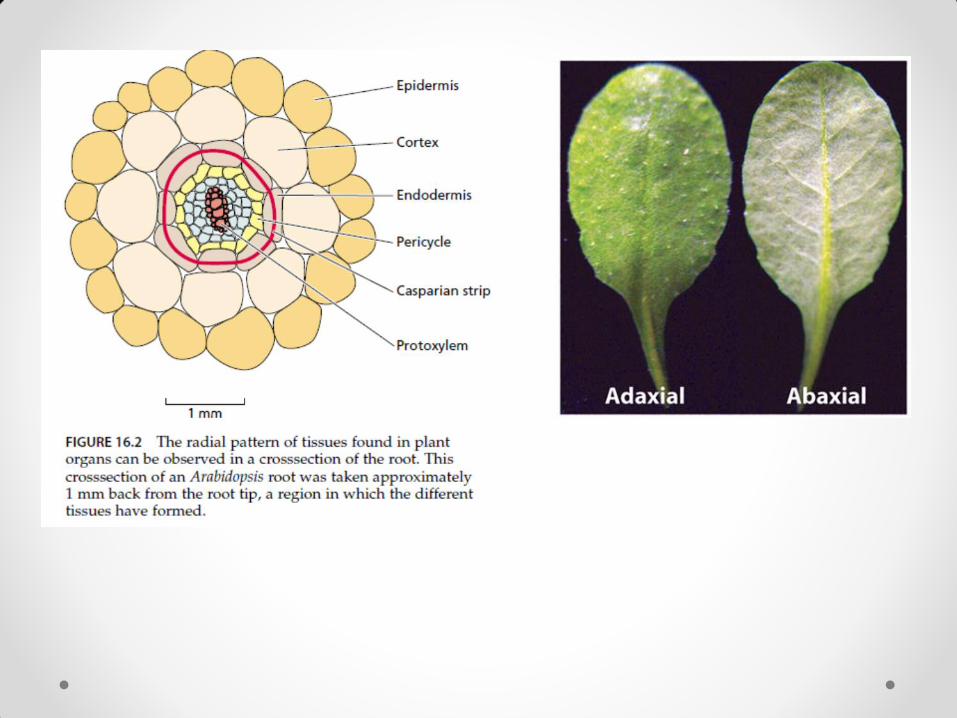
radijalna (u radijalnom pravcu) – dovodi do razlike između centra i periferije organa; karakteristična za stablo i koren
dorzoventralna – generiše razliku između gornjeg i donjeg dela organa; karakteristična za listove većine biljaka (adaksijalna i abaksijalna strana lista, tj. lice i naličje lista)
• Polarnost je fundamentalni uslov za diferencijaciju ćelija i razviće biljke. Ona određuje pravac ćelijskih deoba i rastenja.
• Oko polarne osovine obrazuju se bočni organi po principu radijalne, bilateralne ili dorzoventralne simetrije. Osim toga, duž polarne osovine kod biljaka postoji gradijent hemijskog sastava (auksina, napr.), aktivnosti i intenziteta nekih funkcija organizma.
• Polarnost indukuju spoljašnji uslovi ili okolina u kojoj se zigot nalazi, tokom ranih stadijuma razvića.
• Jednom uspostavljena polarnost ostaje nepromenjena tokom celog života biljnog organizma.

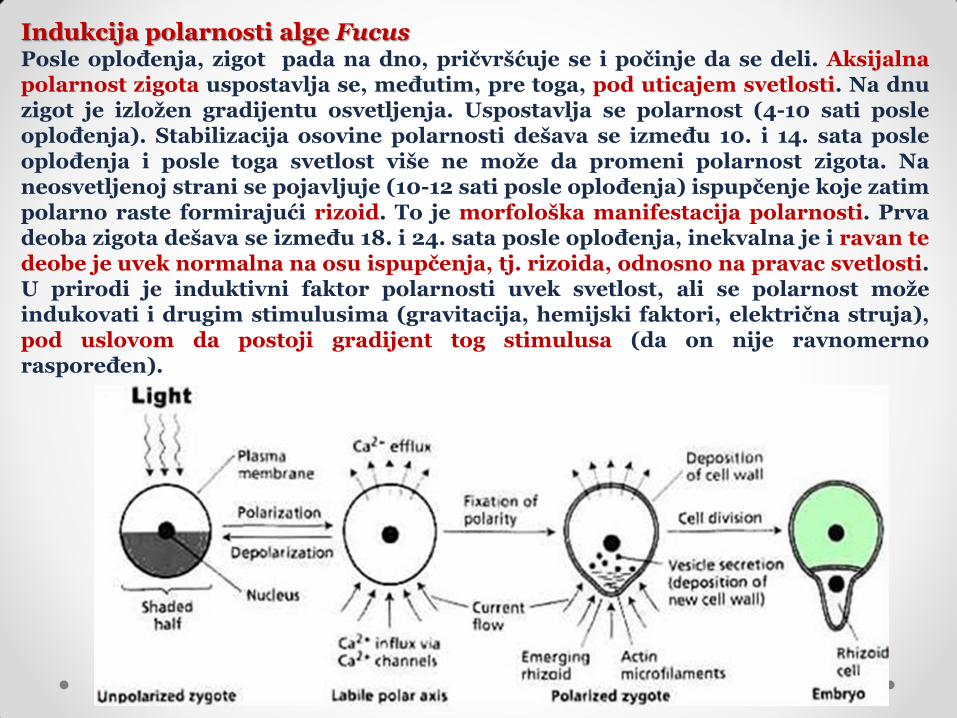
Indukcija polarnosti alge Fucus Posle oplođenja, zigot pada na dno, pričvršćuje se i počinje da se deli. Aksijalna polarnost zigota uspostavlja se, međutim, pre toga, pod uticajem svetlosti. Na dnu zigot je izložen gradijentu osvetljenja. Uspostavlja se polarnost (4-10 sati posle oplođenja). Stabilizacija osovine polarnosti dešava se između 10. i 14. sata posle oplođenja i posle toga svetlost više ne može da promeni polarnost zigota. Na neosvetljenoj strani se pojavljuje (10-12 sati posle oplođenja) ispupčenje koje zatim polarno raste formirajući rizoid. To je morfološka manifestacija polarnosti. Prva deoba zigota dešava se između 18. i 24. sata posle oplođenja, inekvalna je i ravan te deobe je uvek normalna na osu ispupčenja, tj. rizoida, odnosno na pravac svetlosti. U prirodi je induktivni faktor polarnosti uvek svetlost, ali se polarnost može indukovati i drugim stimulusima (gravitacija, hemijski faktori, električna struja), pod uslovom da postoji gradijent tog stimulusa (da on nije ravnomerno raspoređen).
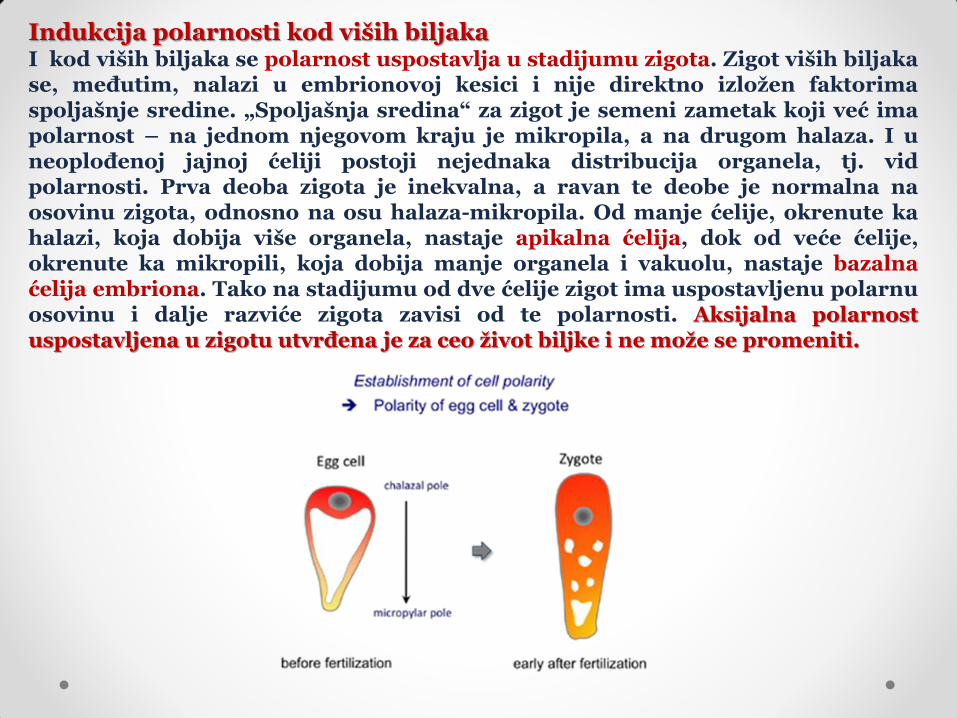
Indukcija polarnosti kod viših biljaka I kod viših biljaka se polarnost uspostavlja u stadijumu zigota. Zigot viših biljaka se, međutim, nalazi u embrionovoj kesici i nije direktno izložen faktorima spoljašnje sredine. „Spoljašnja sredina“ za zigot je semeni zametak koji već ima polarnost – na jednom njegovom kraju je mikropila, a na drugom halaza. I u neoplođenoj jajnoj ćeliji postoji nejednaka distribucija organela, tj. vid polarnosti. Prva deoba zigota je inekvalna, a ravan te deobe je normalna na osovinu zigota, odnosno na osu halaza-mikropila. Od manje ćelije, okrenute ka halazi, koja dobija više organela, nastaje apikalna ćelija, dok od veće ćelije, okrenute ka mikropili, koja dobija manje organela i vakuolu, nastaje bazalna ćelija embriona. Tako na stadijumu od dve ćelije zigot ima uspostavljenu polarnu osovinu i dalje razviće zigota zavisi od te polarnosti. Aksijalna polarnost uspostavljena u zigotu utvrđena je za ceo život biljke i ne može se promeniti.

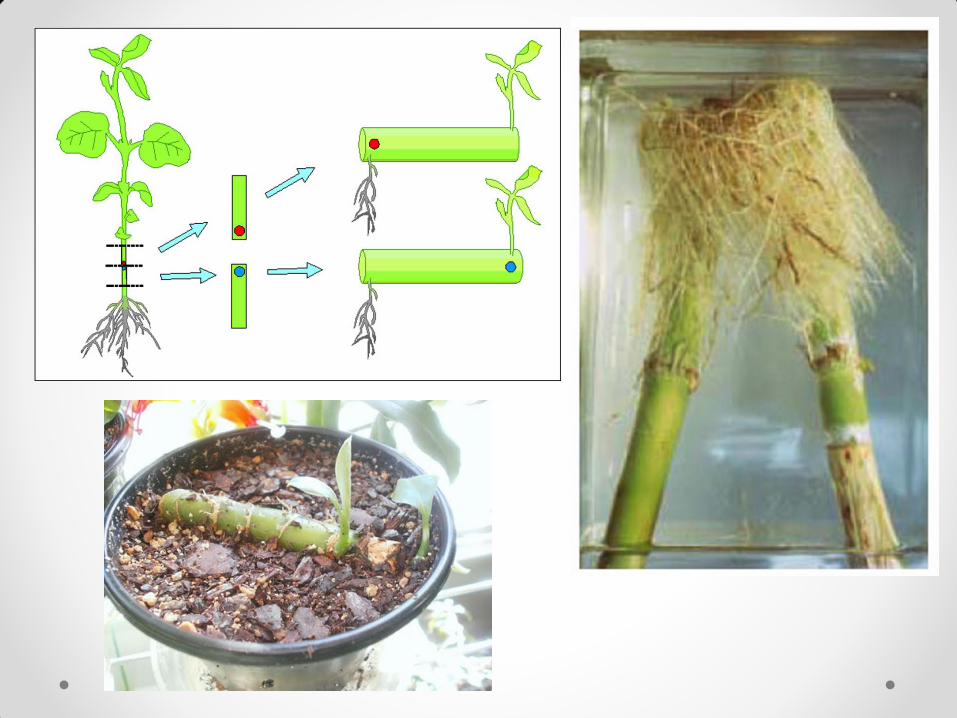
Polarnost se manifestuje u mnogim morfogenetskim i fiziološkim procesima kod biljaka. Većina ćelija se deli i raste u određenoj orijentaciji u odnosu na aksijalnu osovinu. Jedan od značajnih fizioloških činilaca polarnosti kod biljaka je polarni transport supstanci, a naročito polarni transport auksina. Polarnost se ogleda u rasporedu formiranja adventivnih korenova i pupoljaka na odsečcima grana vrbe i korena maslačka. Adventivni pupoljci se uvek formiraju na apikalnom kraju, a korenovi na bazalnom kraju odsečaka, bez obzira na to kako se orijentiše odsečak. Iz ovoga se mogu izvući dva glavna zaključka: 1. Sve ćelije imaju sposobnost da regenerišu i pupoljke i
korenove, a šta će regenerisati zavisi od toga da li se posle fragmentacije nađu na apikalnom ili na bazalnom delu odsečka.
2. Delovi biljke (tj. ćelije u njihovom sastavu), čak i u odsustvu vrha stabla i korena, pokazuju imanemtnu polarnost u skladu sa polarnom osovinom biljke od koje potiču.
U osnovi ove pojave su polarni transport auksina, koji dovodi do auksinskog gradijenta duž biljke, i odnos nivoa citokinina i auksina u pojedinim delovima biljke, tj. njihov balans.

Embriogeneza Embriogeneza je faza u razviću biljke koja počinje prvom deobom zigota, a završava se formiranjem zrelog embriona. Obuhvata niz anatomskih i morfoloških procesa i odvija se u semenom zametku (ovuli). Pre embriogeneze dešavaju se: gametogeneza, oprašivanje, dvojno oplođenje i formiranje zigota. Semeni zameci se formiraju na oplodnim listićima, karpelama, koji srastaju u plodnik (gineceum), u kojem se kasnije razvija seme kod angiospermi. Jedna ćelija nucelusa u sastavu semenog zametka se razvija kao megasporocita, deli se mejozom, daje 4 haploidne ćelije od kojih 3 degenerišu, a jedna se deli trima mitotičkim deobama i daje 8 haploidnih jedara raspoređenih u 7 ćelija koje čine embrionovu kesicu. Embrionova kesica je ženski gametofit (megagametofit).
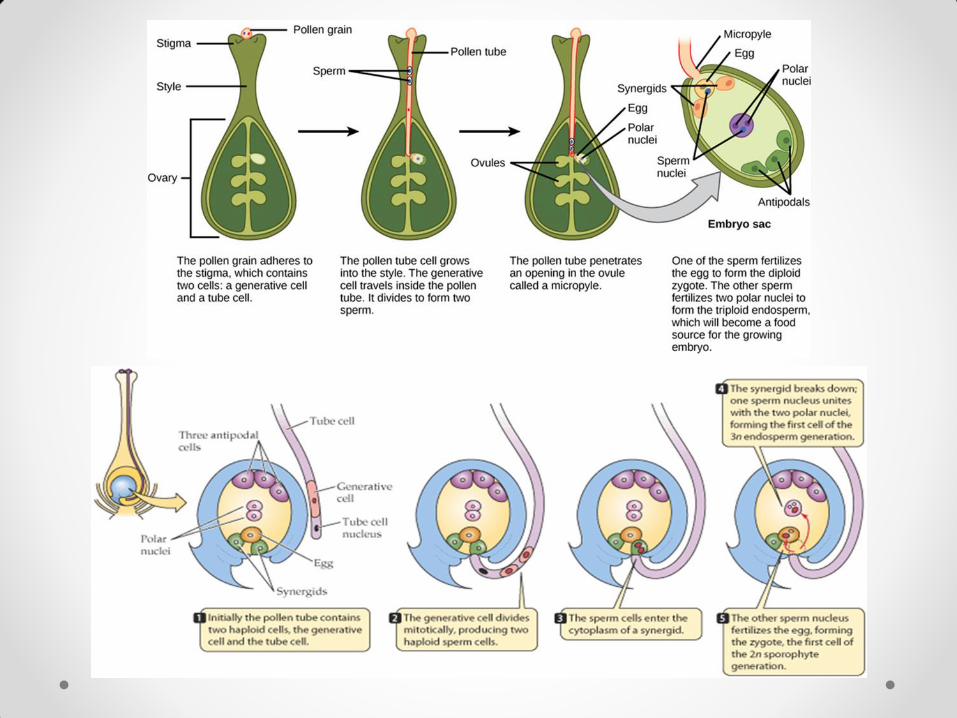

Muški gametofit, mikrogametofit, razvija se u prašnicima, koji čine andreceum. Mikrogametofit se nalazi u polenovim zrnima (mikrosporama) koja nastaju u anterama. Antere sadrže polenovu kesicu sa ćelijama koje se dele i daju ćelije inicijale tapetuma i sporogenu ćeliju. Sporogena ćelija podleže mejozi, pri čemu nastaju 4 haploidne mikrospore (tetrada).


Spajanjem generativne ćelije polena i jajne ćelije embrionove kesice nastaje zigot. Dalje razviće zigota određuje struktura jajne ćelije, u kojoj su organele neravnomerno raspoređene. U zigotu se na mikropilarnom kraju nalazi vakuola, a jedro i većina organela se nalaze na halaznom kraju zigota. Prva deoba zigota je inekvalna, a njena ravan je poprečna u odnosu na mikropila-halaza pravac. Njome počinje embriogeneza. Nastaju dve ćelije. U dvoćelijskom stadijumu zigota aksijalna polarnost je već uspostavljena. Manja, apikalna, ćelija sadrži većinu mitohondrija, proplastida i drugih organela i okrenuta je ka halazi. Ona će u daljem razviću dati većinu tkiva embriona. Veća, bazalna, ćelija sadrži vakuolu, okrenuta je ka mikropili i u daljem razviću daje suspenzor, korensku kapu i deo korena.

Prve dve deobe apikalne ćelije su uzdužne i daju kvadrant, stadijum od 4 ćelije, a ravni te dve deobe su pod pravim uglom. Sledeća deoba ćelija kvadranta je poprečna i njome nastaje oktant, stadijum od 8 ćelija. Za to vreme, poprečnim deobama bazalne ćelije formira se suspenzor. U oktantu poprečni ćelijski zidovi (O’ linija) predstavljaju granicu između budućeg apeksa sa kotiledonima i budućeg hipokotila. Ćelije oktanta dele se periklinim deobama (deobne ravni paralelne površini) i daju stadijum dermatogena (16 ćelija). Spoljašnje ćelije dermatogena se nazivaju protoderm, jednoćelijski sloj od kojeg nastaje epiderm. Sve sledeće deobe protoderma su antikline, zbog čega je epiderm na kraju jednoslojan. Unutrašnje ćelije dermatogena dele se uzdužno. Embrion ulazi u rani globularni stadijum. Istovremeno se ćelija na vrhu suspenzora podeli poprečno, a gornja ćelija se naziva hipofiza (hy). U kasnom globularnom stadijumu ćelije embriona se dele i poprečnim i uzdužnim deobama, a hipofiza samo uzdužnim deobama. Radijalna simetrija embriona se uspostavlja u stadijumu globule, a kod adultnih biljaka se zadržava u građi osovinskih organa (korena i stabla).
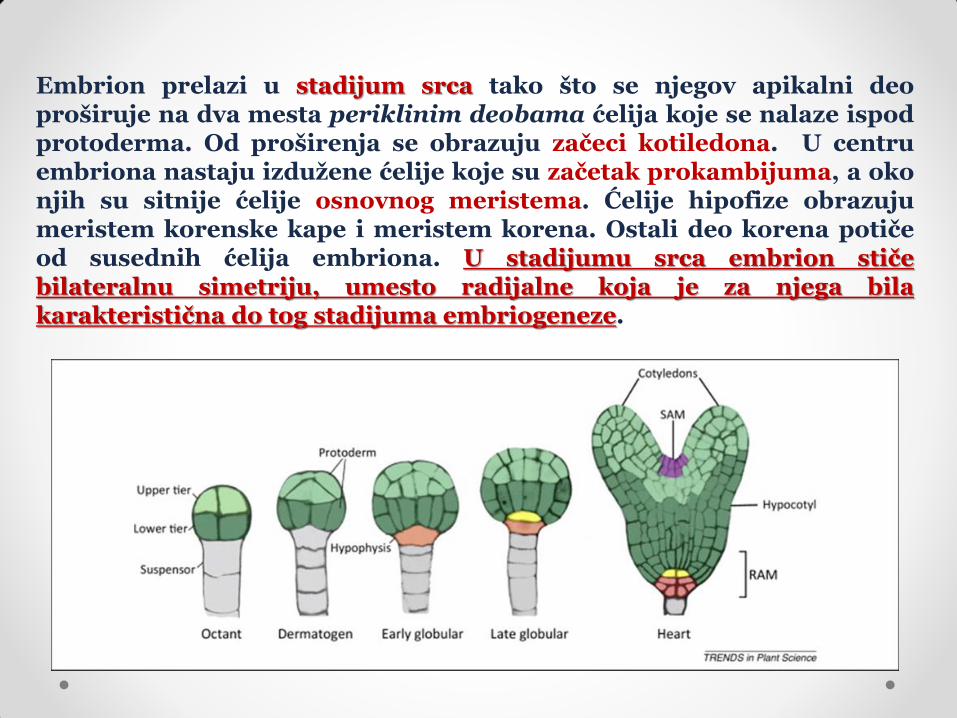
Embrion prelazi u stadijum srca tako što se njegov apikalni deo proširuje na dva mesta periklinim deobama ćelija koje se nalaze ispod protoderma. Od proširenja se obrazuju začeci kotiledona. U centru embriona nastaju izdužene ćelije koje su začetak prokambijuma, a oko njih su sitnije ćelije osnovnog meristema. Ćelije hipofize obrazuju meristem korenske kape i meristem korena. Ostali deo korena potiče od susednih ćelija embriona. U stadijumu srca embrion stiče bilateralnu simetriju, umesto radijalne koja je za njega bila karakteristična do tog stadijuma embriogeneze.
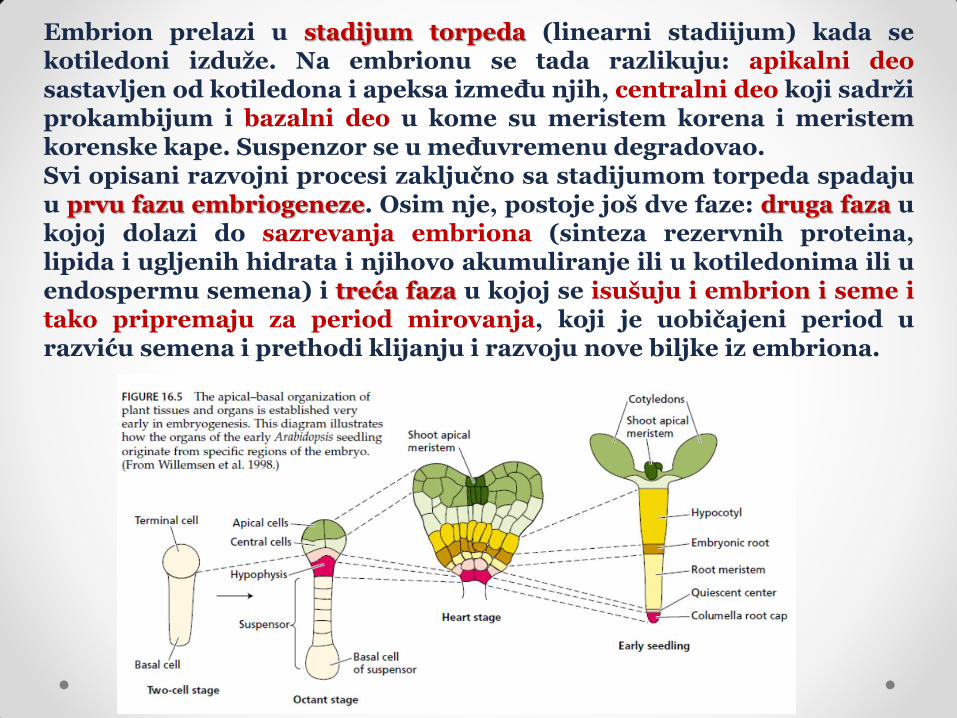
Embrion prelazi u stadijum torpeda (linearni stadiijum) kada se kotiledoni izduže. Na embrionu se tada razlikuju: apikalni deo sastavljen od kotiledona i apeksa između njih, centralni deo koji sadrži prokambijum i bazalni deo u kome su meristem korena i meristem korenske kape. Suspenzor se u međuvremenu degradovao. Svi opisani razvojni procesi zaključno sa stadijumom torpeda spadaju u prvu fazu embriogeneze. Osim nje, postoje još dve faze: druga faza u kojoj dolazi do sazrevanja embriona (sinteza rezervnih proteina, lipida i ugljenih hidrata i njihovo akumuliranje ili u kotiledonima ili u endospermu semena) i treća faza u kojoj se isušuju i embrion i seme i tako pripremaju za period mirovanja, koji je uobičajeni period u razviću semena i prethodi klijanju i razvoju nove biljke iz embriona.

Osim opisanog, kod biljaka postoji još 6 tipova embriogeneze. Za sve tipove embriogeneze je zajedničko da se tokom embriogeneze definišu posebni embrionalni domeni i da se kompetencija ćelija tih domena da se diferenciraju sve više sužava. Razvojna sudbina ćelija različitih embrionalnih domena postaje precizno određena, tako da se razviće pojedinih organa može pratiti unazad do određenog embrionalnog domena. Embriogeneza uspostavlja osnovnu organizacioni plan biljnog organizma koji se onda održava tokom ontogeneze, a to podrazumeva: bazalno-apikalni obrazac razvića (uspostavljanje
aksijalne osovine i nepromenljivo definisanje osovine stablo-koren kod adultnih biljaka)
radijalna simetrija u građi (rasporedu tkiva) osovinskih organa (korena i stabla)
uspostavljanje primarnih meristema (naročito apikalnih meristema korena i stabla), koji tokom celog života biljke omogućavaju rastenje, diferencijaciju tkiva i formiranje lateralnih organa.
Morfogeneza korena
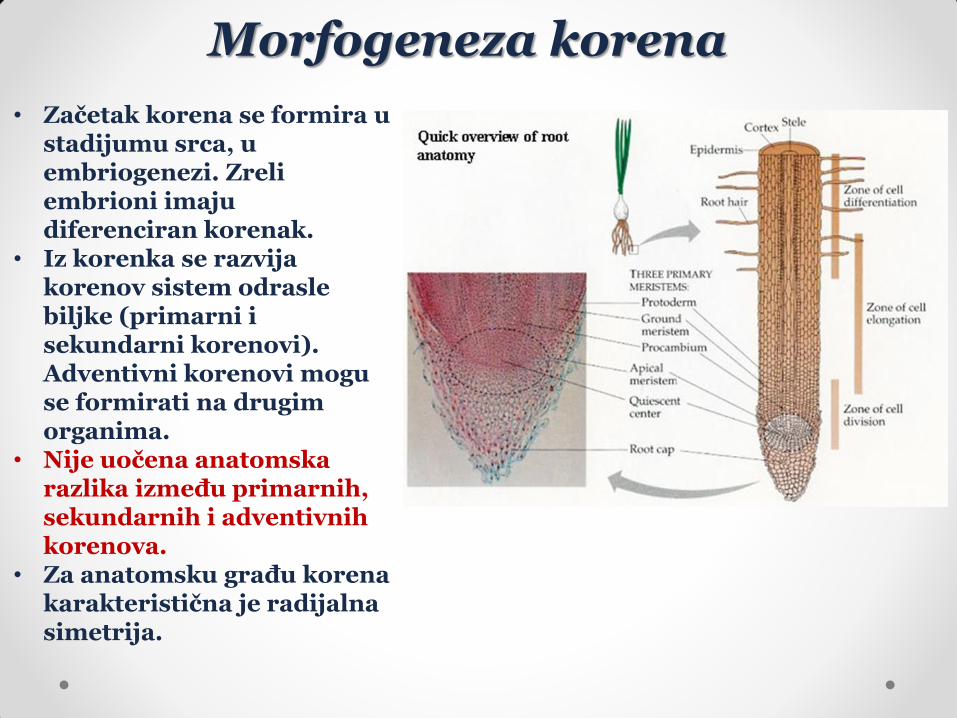
• Začetak korena se formira u stadijumu srca, u embriogenezi. Zreli embrioni imaju diferenciran korenak.
• Iz korenka se razvija korenov sistem odrasle biljke (primarni i sekundarni korenovi). Adventivni korenovi mogu se formirati na drugim organima.
• Nije uočena anatomska razlika između primarnih, sekundarnih i adventivnih korenova.
• Za anatomsku građu korena karakteristična je radijalna simetrija.
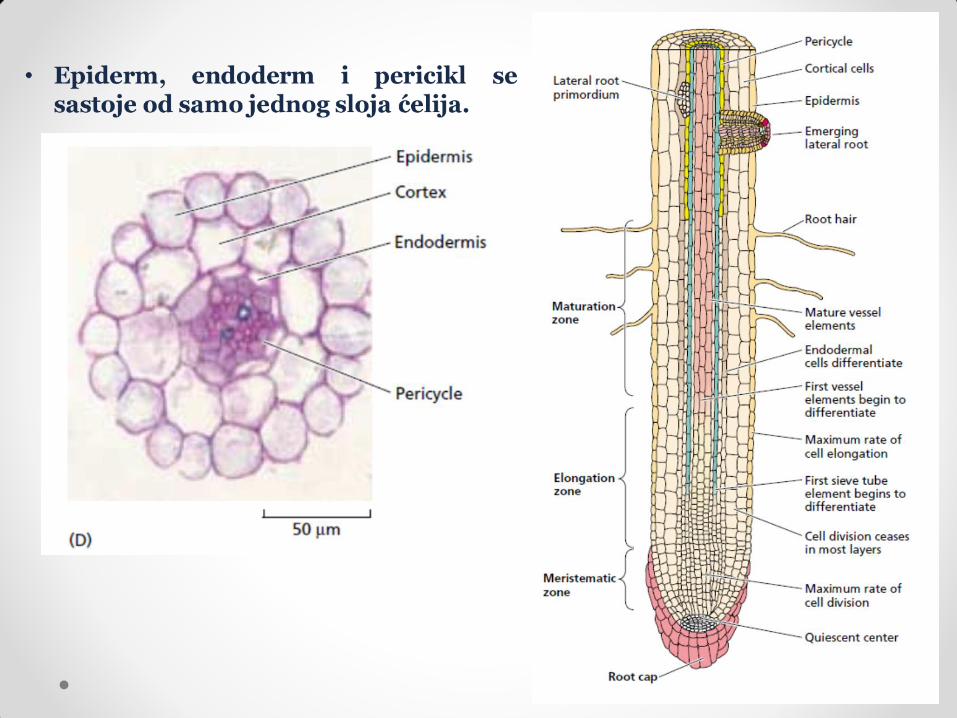

Na vrhu korena nalazi se korenska kapa. Njene funkcije su: zaštita korena od mehaničkih povreda, formiranje sluzavog omotača (ima funkciju u usvajanju nutrijenata, uspostavljanju mikorize i održavanju korisnih bakterija u rizosferi) i percepcija gravitropskog stimulusa. Nastaje od kaliptrogena (meristem) koji formira kolumelu. Ćelije kolumele su ćelije korenske kape koje prve nastaju od kaliptrogena, postavljene su centralno i glavna im je funkcija prijem gravitropskog signala (imaju amiloplaste). Kad one budu potisnute ka periferiji (aktivnošću kaliptrogena i odbacivanjem perifernih ćelija korenske kape), stiču druge funkcije.

Ispod korenske kape nalazi se primarni meristem korena (apikalni meristem korena). On se sastoji od dva kruga inicijalnih ćelija koje se stalno dele između kojih se nalaze ćelije koje se veoma retko dele. Gornji krug inicijala su ćelije od kojih se diferenciraju sva tkiva korena, donji sloj inicijala je kaliptrogen (meristem korenske kape), a ćelije koje se retko dele čine mirujući centar. Funkcija mirujućeg centra je da obezbedi rezervu meristemskih ćelija, koje se aktiviraju i zamenjuju oštećene meristemske ćelije. Inicijale gornjeg sloja meristema korena dele se poprečnim deobama. Od dve novonastale ćelije jedna ostaje meristemska, a druga raste, deli se i diferencira u određenom pravcu, dajući određeno tkivo korena. Što su dalje od vrha korena, ćelije se ređe dele, a više su diferencirane. Jedino tkivo korena čije ćelije zadržavaju sposovnost deobe je pericikl.
Formativne i proliferativne deobe: • formativnim deobama koren
se izdužuje i formiraju se nizovi ćelija (ćelijske linije ili klonovi), od kojih svaki daje jedno tkivo.
• proliferativnim deobama koren zadebljava, povećava se broj ćelija jednog tkiva u radijalnom pravcu i tkiva postaju višeslojna.

Na prelazu iz meristemske zone u zonu izduživanja korena ćelije postaju vakuolizovane, prestaju da se dele i počinju da se izdužuju. Pojavljuju se začeci protofloema, a zatim i protoksilema. U zoni diferencijacije ćelije dostižu svoju krajnju veličinu i stiču sve odlike tkiva u koje se diferenciraju. Pojedine epidermalne ćelije formiraju korenske dlačice. Provodni snopići sadrže diferencirane floemske i ksilemske elemente, a endoderm je, takođe, diferenciran. Koren postaje potpuno osposobljen za usvajanje vode i mineralnih materija.

Na rastojanju od nekoliko mm do nekoliko cm od vrha korena počinje formiranje bočnih (lateralnih) korenova. Oni se začinju u periciklu, čije ćelije zadržavaju spsobnost deobe. U periciklu se od grupe ćelija koje imaju osobine pravog meristema korena formira začetak korena (korenska primordija). Njihovim deobama i izduživanjem i diferencijacijom ćelija koje u deobama nastanu formirase koren koji se probija ka periferiji korena i na kraju izbija na površinu kroz epiderm.
Iznad zone formiranja lateralnih korenova, u korenu se dešavaju sekundarne promene – epiderm se zamenjuje peridermom (čiji su zidovi suberinizovani, pa slabo propuštaju vodu) i formira se sekundarni kambijum (čijom aktivnošću koren deblja pri bazi). Za diferencijaciju ćelija korena nije odlučujuće poreklo tih ćelija (od kojih su inicijala nastale), nego postoji poziciona kontrola (signali koji stižu iz već diferenciranog, starijeg tkiva usmeravaju diferencijaciju ćelija. Otkriveni su i geni koji specifično regulišu razviće korena.

Morfogeneza stabla Stablo je aksijalni organ radijalne simetrije, koji nosi bočne organe (listove i grane), a razvija se iz stabaoceta koje je se razvija još u embrionu. Osim apikalnog meristema (primarni meristem koji se formira u embrionu) u stablu se nalaze i aksilarni i adventivni meristemi koji nastaju kasnije u ontogenezi. Apikalni meristem stabla zaštićen je najmlađim listovima koji ga obavijaju i grade apikalni pupoljak, plumulu. Stablo dikotila raste samo vrhom, aktivnošću apikalnog meristema, dok kod monokotila deo rastenja stabla obezbeđuje interkalarni meristem u bazi organa (zaštita biljke u slučaju da vrh bude oštećen ili odstranjen – biljojedi). Na obodu apikalnog meristema obrazujju se začeci listova i bočnih izdanaka, prema rasporedu koji se naziva filotaksis (karakterističan za vrstu i razvojno određen). Na stablu se razlikuju nodusi i internodije. Bočni izdanci se formiraju u pazuhu listova i imaju istu građu kao i glavni izdanak.
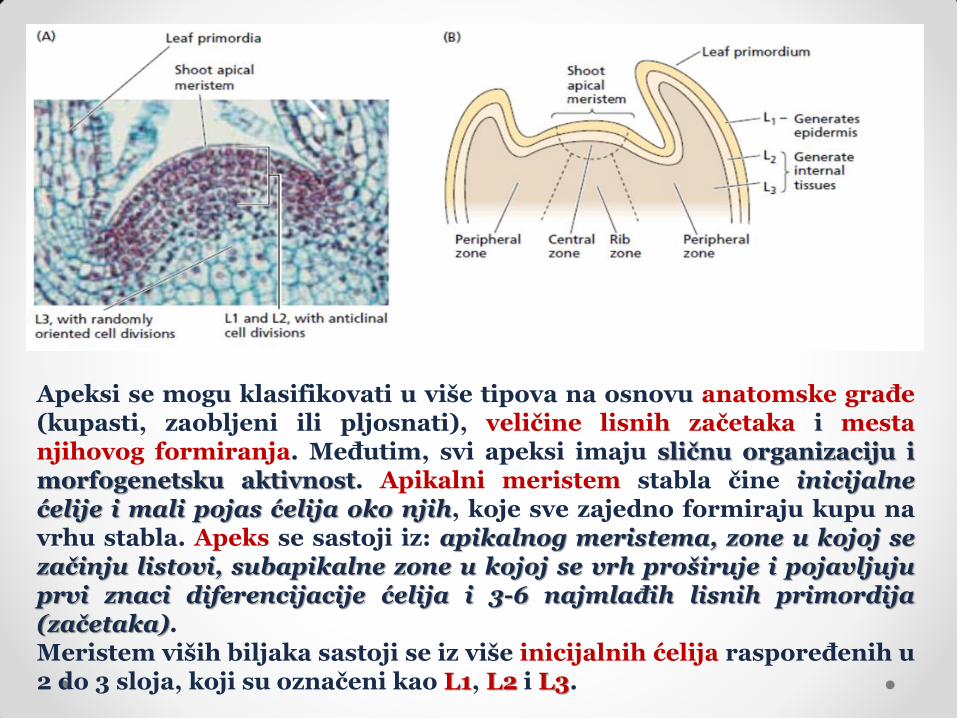
Apeksi se mogu klasifikovati u više tipova na osnovu anatomske građe (kupasti, zaobljeni ili pljosnati), veličine lisnih začetaka i mesta njihovog formiranja. Međutim, svi apeksi imaju sličnu organizaciju i morfogenetsku aktivnost. Apikalni meristem stabla čine inicijalne ćelije i mali pojas ćelija oko njih, koje sve zajedno formiraju kupu na vrhu stabla. Apeks se sastoji iz: apikalnog meristema, zone u kojoj se začinju listovi, subapikalne zone u kojoj se vrh proširuje i pojavljuju prvi znaci diferencijacije ćelija i 3-6 najmlađih lisnih primordija (začetaka). Meristem viših biljaka sastoji se iz više inicijalnih ćelija raspoređenih u 2 do 3 sloja, koji su označeni kao L1, L2 i L3.

U spoljašnjem, L1 sloju ćelije se dele isključivo antiklinim deobama i od njih nastaje jednoslojni epiderm. Ćelije srednjeg, L2 sloja takođe se dele antiklinim deobama, osim na mestima gde se začinju listovi (tamo su deobe perikline) i od njih nastaju začeci listova i bočnih izdanaka, kao i subepidermalna tkiva stabla. U unutrašnjem, L3 sloju ćelije se dele u svim pravcima, a od njih nastaju unutrašnja tkiva stabla. Ove ćelije učestvuju i u formiranju listova i bočnih izdanaka. Slojevi L1 i L2 su nezavisni jedan od drugog, ne mešaju se i svaki od njih nastaje antiklinim deobama posebnih inicijala. Prema drugoj podeli, sve ćelije koje se dele pretežno antiklino (L1 i L2 sloj) sačinjavaju tuniku, a ostale ćelije spadaju u korpus. Prema morfološkom kriterijumu, meristem stabla se može, nezavisno od postojećih slojeva, podeliti na funkcionalne zone:
centralna zona (CZ) periferijska zona (PZ) „rebrasti“ meristem

U centralnoj zoni su sve ćelije inicijale i od njih nastaju sve ostale ćelije apeksa. Oko ovih ćelija su ćelije periferijske zone među kojima su inicijale lista. Rebrasti meristem nalazi se ispod prve dve zone, a njegove ćelije daju nizove ćelija, ćelijske linije ili klonove od kojih se diferenciraju unutrašnja tkiva stabla. Apeks je organizator morfogeneze. Ćelije apeksa mogu da slede jedan od tri moguća puta diferencijacije: • da se razvijaju u listove (organi dorzoventralne simetrije i
ograničenog rastenja) • da se razvijaju u bočne pupoljke (radijalna simetrija, neograničeno
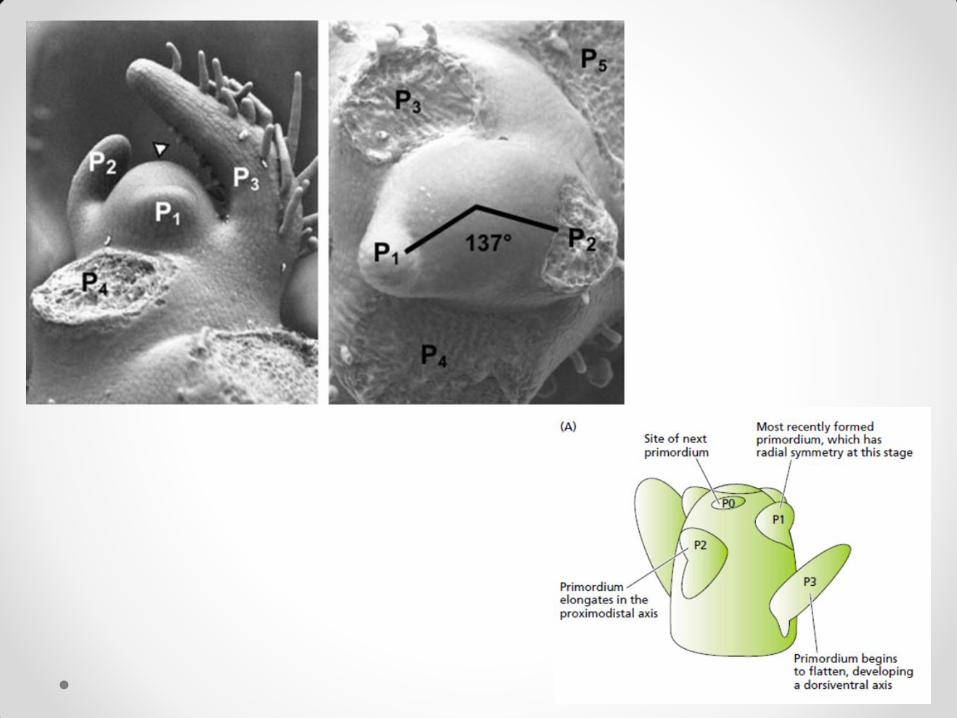
rastenje) • da ostanu ćelije stabla Ova tri razvojna puta su prostorno i vremenski precizno koordinisana. Vegetativni meristem stabla obrazuje repetitivne morfološke jedinice, fitomere. Fitomera sadrži list, nodus, internodiju ispod nodusa i jedan ili više pupoljaka u pazuhu lista. Svaka fitomera nastaje od posebne grupe meristemskih ćelija. Ćelije pojedinih funkcionalnih zona nisu ireverzibilno determinisane da prate samo jedan razvojni put. Na apeksu se mogu unapred predvideti (na osnovu geometrije rasporeda listova) mesta na kojima će se pojaviti inicijale budućih listova. Ta mesta se nazivaju centri rastenja.

Centri rastenja, iz kojih će se razviti nov bočni organ, determinisani su znatno pre pojave tog organa. U ranoj fazi razvića lisna primordija može da se pretvori u pupoljak, jer nije ireverzibilno determinisana. Apikalni meristem je glavni organizator diferencijacije lista – u njegovom odsustvu diferenciraju se pupoljci. On je organizator i formiranja provodnog tkiva (uloga auksina koji su istovremeno i ograničavajući i kontrolni faktor razvoja i ksilema i floema). Starije primordije su ireverzibilno determinisane i na njihovo razviće apikalni meristem više ne utiče. Apikalni meristem stabla je autonoman u svom razviću – izolovan iz biljke može da se razvija organizovano i da da biljku. Uslov za to je jedino da izolovani deo sadrži i lisne primordije ili da se njihovo prisustvo zameni spolja dodatim auksinom. Osnovne funkcije apikalnog meristema stabla su: da sam sebe održava i da inicira nove bočne organe tokom života biljke. Ove funkcije održavaju strukturni geni koji su pod kontrolom tri klase regulatornih („homeobox“) gena: • KN (Knotted) (ograničavaju preteranu proliferaciju meristemskih
ćelija) i STM (održavaju proliferaciju ćelija u centralnoj zoni meristema) geni
• CLAVATA (stimulišu diferencijaciju primordija lateralnih organa) i WUSCHEL (određuju diferencijaciju u ćelije stabla) geni
• geni koji regulišu razviće lisnih primordija

Razviće lista
Listovi su lateralni organi dorzoventralne simetrije. Začinju se u periferijskoj zoni apikalnog meristema stabla, periklinim deobama u L2 sloju. Obrazuje se malo ispupčenje, koje sa udaljenošću od vrha postaje sve izraženije. U pazuhu lisnih primordija formira se novo ispupčenje - to je začetak bočnog pupoljka. U razviću lista postoje tri faze: I. Formiranje ispupčenja, tj. lisne primordije na apeksu stabla II. Izduživanje ispupčenja zahvaljujući deobama ćelija na vrhu – nastaje
aksilarni organ radijalne simetrije, čije će se ćelije razviti u ćelije provodnih snopiča i lisne drške. Kad dostigne dužinu od 1 mm, deobe se proređuju na vrhu, a počinju da se dele ćelije dva naspramna bočna niza, čime se formira proširenje, a primordija dobija pljosnat oblik i dorzoventralnu simetriju – formira se lisna ploča. Meristemske ćelije ostaju aktivne samo po obodu ploče (marginalni meristem).
III.Prestaju deobe ćelija, a list raste samo na osnovu povećanja dimenzija već postojećih ćelija.
Razviće začetaka bočnog pupoljka u početku se ne razlikuje od razvića lisne primordije, međutim, pupoljak raste kao aksijalni organ radijalne simetrije, a na vrhu obrazuje apikalni meristem, isti kao onaj na vrhu stabla (aksilarni meristem).
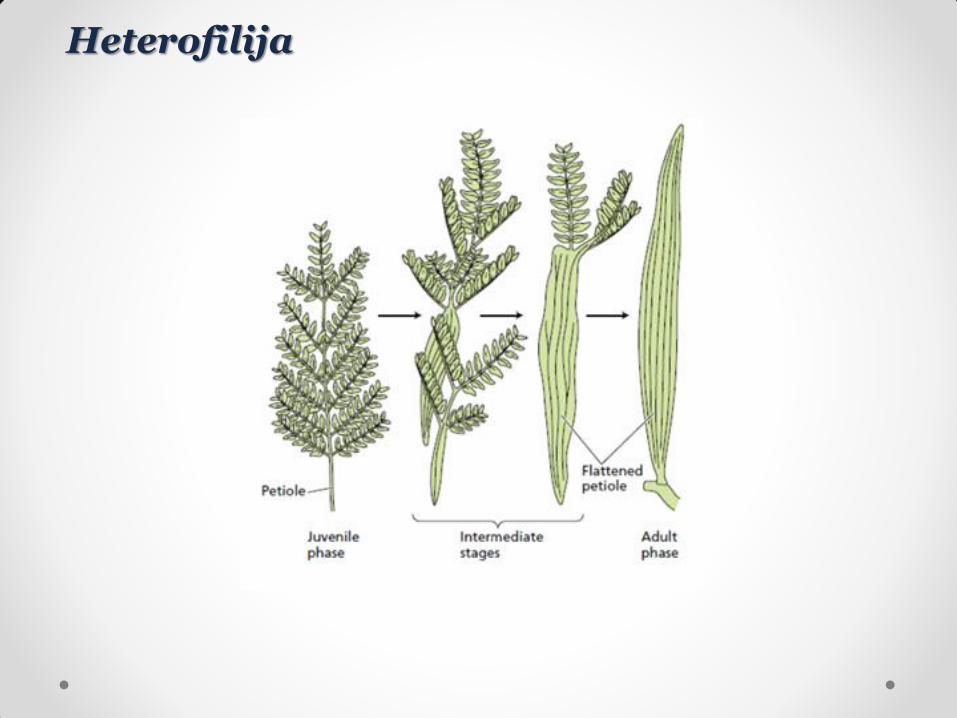
Heterofilija

Filotaksis i plastohron Filotaksis je redosled razvića listova na biljci. Pozicija listova u odnosu na meristemsku kupu je odlika vrste. Filotaksis je određen još u meristemu. Plastohron je vreme koje protekne između inicijacije dva sukcesivna lista













![UTICAJ KONCENTRACIJA Cd I FAZE RASTENJA I RAZVI]A NA …scindeks-clanci.ceon.rs/data/pdf/0351-4781/2003/0351-47810339077K.pdf · Tolerantnost biljaka prema Cd mo`e biti ste~eno svojstvo](https://img.dokumen.tips/doc/110x75/5e090c1867f0056ea75d2ced/uticaj-koncentracija-cd-i-faze-rastenja-i-razvia-na-scindeks-tolerantnost-biljaka.jpg)















