Embed Size (px)
Citation preview

Univerza v Mariboru
Fakulteta za naravoslovje in matematiko
Oddelek za biologijo
MAGISTRSKO DELO
Klavdija PROT
Maribor, 2019


Univerza v Mariboru
Fakulteta za naravoslovje in matematiko
Oddelek za biologijo
Klavdija PROT
Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus
Diversity in shape of skull and molars in mouse-like hamsters, Calomyscus
MAGISTRSKO DELO
Mentor: prof. dr. Franc JANŽEKOVIČ
Somentor: prof. dr. Boris KRYŠTUFEK
Maribor, 2019


UNIVERZA V MARIBORU
FAKULTETA ZA NARAVOSLOVJE IN MATEMATIKO
IZJAVA O AVTORSTVU
Magistrsko delo je nastalo kot rezultat lastnega dela. Vsi privzeti podatki so citirani skladno z
mednarodnimi pravili o varovanju avtorskih pravic.
Podpisana Klavdija Prot izjavljam, da sem za potrebe arhiviranja oddala elektronsko verzijo
zaključnega dela v Digitalno knjižnico Univerze v Mariboru. Magistrsko delo sem izdelala
sama ob pomoči mentorja. V skladu s 1. odstavkom 21. člena Zakona o avtorskih in sorodnih
pravicah (Ur. l. RS, št. 16/07 – uradno prečiščeno besedilo, 68/08, 110/13, 56/15 in 63/16 -
ZKUASP) dovoljujem, da se zgoraj navedeno zaključno delo objavi na portalu Digitalne
knjižnice Univerze v Mariboru. Tiskana verzija magistrskega dela je istovetna elektronski
verziji, ki sem jo oddala za objavo v Digitalno knjižnico Univerze v Mariboru.
Podpisana izjavljam, da dovoljujem objavo osebnih podatkov, vezanih na zaključek študija, in
sicer za spremljanje zaposlovanja diplomantov, za potrebe Kariernega centra ter Alumni
klubov.
Maribor, ______________ Klavdija Prot

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
III
Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus.
Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko,
Oddelek za biologijo, 2019
IZVLEČEK
V sklopu magistrske naloge sem s pomočjo geometrijske morfometrije preučevala razlike v
variabilnosti lobanje ter morfometrične odnose med vrstami rodu Calomyscus. Uporabila sem
material iz različnih muzejskih zbirk. Analizirala sem okluzalno površino zgornjih meljakov
ter lobanjo iz treh perspektiv (ventralno, dorzalno in lateralno stran). Raziskava je vključevala
štirinajst lobanj, ki so pripadale osmim različnim vrstam, in sicer C. bailwardi, C. baluchi, C.
elburzensis, C. grandis, C. hotsoni, C. mystax, C. tsolovi in C. urartensis. Namen naloge je bil
ovrednotenje morfometrične pestrosti med vrstami tako v obliki kot v velikosti obravnavanih
struktur. Zanimalo me je, ali posamezni segmenti lobanje izkazujejo značilne razlike med
obravnavanimi vrstami. Analizo velikosti sem izvedla z vrednostmi centroidnih velikosti.
Izrazito sta odstopala C. tsolovi (nizke vrednosti centroidne velikosti) in C. baluchi mustersi
(visoke vrednosti centroidne velikosti). Variabilnost v obliki sem ovrednotila s prvima dvema
glavnima komponentama, dobljenima v analizi PCA. Na osnovi oblike je bila opazna
variabilnost med vrstami in znotraj njih. Izrazito je odstopal C. tsolovi, in sicer v vrednostih
dorzalne in lateralne strani lobanje. Morfometrične odnose med taksoni sem predstavila z
Evklidovimi razdaljami za vsak lobanjski segment (meljaki, ventralna, dorzalna in lateralna
stran lobanje) posebej. V obliki meljakov se je najbolj razlikoval C. hotsoni, v obliki lateralne
in dorzalne strani lobanje pa C. tsolovi. Podobnost v morfometričnih odnosih v obliki različnih
segmentov lobanje in meljakov sem ugotavljala s korelacijsko analizo med posameznimi
matrikami. Signifikantno podobnost (p < 0,05) sem potrdila le med Evklidovimi razdaljami
lateralne in dorzalne strani lobanje. V ostalih parnih primerjavah nisem ugotovila signifikantnih
korelacijskih koeficientov (p > 0,05). Dokazano so lobanjske strukture neodvisne druga od
druge. Zaključujem, da med vsemi analiziranimi vrstami najbolj izstopa C. tsolovi.
Ključne besede: Calomyscus, geometrijska morfometrija, lobanja, medvrstna variabilnost,
meljaki, morfometrični odnosi

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
IV
Prot, K. Diversity in shape of skull and molars in Mouse-like hamsters, Calomyscus.
Master of Science Thesis. University of Maribor, Faculty of Mathematics and Natural
Sciences, 2019
ABSTRACT
As part of my master thesis, I studied variances in skull variability and morphometric relations
among species of the Calomyscus genus. I worked with different material collections from
museums. I analysed the occlusal surface of the upper molar teeth as well as the skull from
three different perspectives (ventral, dorsal and lateral). The research involved fourteen skulls
from eight different species, i.e. C. bailwardi, C. baluchi, C. elburzensis, C. grandis, C. hotsoni,
C. mystax, C. tsolovi and C. urartensis. The aim of the thesis was to evaluate the morphometric
diversity among the mentioned species, in terms of the shape as well as the size of the examined
structures. I examined if separate skull segments show specific differences among the observed
species. The size analysis was conducted using values of centroid size units. A noticeable
deviation could be observed with the species C. tsolovi (low levels of centroid size) and C.
baluchi mustersi (high levels of centroid size). The variability of shape was evaluated through
the first two main components, which were gained in the Principal component analysis (PCA).
Observing the shape, a variability could be observed between and within the species. A
noticeable deviation could be observed with the species C. tsolovi, explicitly regarding the
values of the dorsal and lateral skull side. The morphometric relations among taxons have been
demonstrated using the Euclidean distance matrix for each individual skull segment separately
(molar teeth as well as the ventral, dorsal and lateral side of the skull). Regarding the shape of
the molar teeth, the most significant deviation could be observed with the species C. hotsoni.
When focusing on the shape of the lateral and dorsal skull side, the major deviation could be
observed with the C. tsolovi species. The resemblance in morphometric relations in the shape
of different skull segments and molar teeth has been examined using the correlation analysis
among individual matrices. A significant similarity (p < 0,05) could be confirmed only among
Euclidean distances in the lateral and dorsal side of the skull. In other dual comparisons no
significant correlation coefficients (p > 0,05) could be discovered. It could be proven that skull
structures do not correlate among each other and are independent. The research can be
concluded with the finding that among all of the analysed species, C. tsolovi is the most distinct
one.
Keywords: Calomyscus, geometric morphometrics, skull, variability among species, molar
teeth, morphometric relations

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
V

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
VI
Kazalo vsebine 1 Uvod ................................................................................................................................... 1
1.1 Mišji hrčki ..................................................................................................................... 1
1.2 Opis .............................................................................................................................. 2
1.3 Telesne značilnosti ....................................................................................................... 2
1.4 Razširjenost .................................................................................................................. 3
1.5 Habitat ......................................................................................................................... 4
1.6 Biologija ....................................................................................................................... 4
1.7 Morfologija lobanje in meljakov .................................................................................. 5
1.8 Geometrijska morfometrija ......................................................................................... 7
1.9 Namen dela in hipoteze ............................................................................................... 8
2 Material in metode dela ..................................................................................................... 9
2.1 Material ....................................................................................................................... 9
2.2 Oslonilne točke ............................................................................................................ 9
2.3 Prokrustova analiza ................................................................................................... 12
2.4 Analiza glavnih komponent (PCA) ............................................................................. 12
2.5 Drevo najkrajšega razvejanja (angl. Minimum spanning tree) .................................. 13
2.6 Klastrska analiza ......................................................................................................... 13
2.7 Grafikon kvantilov – Box plot.................................................................................... 13
2.8 Mantel t test .............................................................................................................. 13
3 Rezultati ............................................................................................................................ 14
3.1 Prokrustova analiza in analiza glavnih komponent ................................................... 14
3.1.1 Prokrustova analiza in analiza glavnih komponent - meljaki ............................. 14
3.1.2 Prokrustova analiza in analiza glavnih komponent - ventralna stran lobanje ... 18
3.1.3 Prokrustova analiza in analiza glavnih komponent - dorzalna stran lobanje .... 22
3.1.4 Prokrustova analiza in analiza glavnih komponent - lateralna stran lobanje .... 25
3.2 Analiza velikosti ......................................................................................................... 28
3.2.1 Analiza velikosti - meljaki ................................................................................... 28
3.2.2 Analiza velikosti - ventralna stran lobanje ......................................................... 29
3.2.3 Analiza velikosti - dorzalna stran lobanje ........................................................... 29
3.2.4 Analiza velikosti - lateralna stran lobanje .......................................................... 30
3.3 Analiza oblike ............................................................................................................. 31
3.3.1 Analiza oblike - meljaki ....................................................................................... 31

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
VII
3.3.2 Analiza oblike - ventralna stran lobanje ............................................................. 34
3.3.3 Analiza oblike - dorzalna stran lobanje .............................................................. 36
3.3.4 Analiza oblike – lateralna stran lobanje ............................................................. 39
3.4 Morfometrični odnosi med taksoni ........................................................................... 42
3.4.1 Morfometrični odnosi med taksoni - meljaki ..................................................... 42
3.4.2 Morfometrični odnosi med taksoni - ventralna stran lobanje ........................... 44
3.4.3 Morfometrični odnosi med taksoni - dorzalna stran lobanje ............................ 46
3.4.4 Morfometrični odnosi med taksoni - lateralna stran lobanje ............................ 48
3.4.5 Mantel t test ....................................................................................................... 50
4 Razprava ........................................................................................................................... 51
4.1 Analiza velikosti ......................................................................................................... 51
4.2 Analiza oblike ............................................................................................................. 51
4.3 Morfometrični odnosi med taksoni ........................................................................... 53
5 Zahvala ............................................................................................................................. 54
Literatura .................................................................................................................................. 55
6 Priloga ............................................................................................................................... 58

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
VIII
Kazalo slik
Slika 1: Mišji hrček (Calomyscus) (foto J. Vogeltanz), (Kryštufek in Vohralik, 2009). ................ 1
Slika 2: Razširjenost vrst rodu Calomyscus (povzeto po Shahabi in sod., 2013). ....................... 3
Slika 3: Lobanja in meljaki mišjih hrčkov (Calomyscus); 1. rostrum, 2. nosnica (Nasal), 3. sekalci
(Dentes incisivi) a. zgornji sekalec, b. spodnji sekalci, 4. očesne jamice (Orbita), 5. medočesni
prostor (Regio interorbital), 6. lični lok (Arcus zygomaticus), 7. maseterična plošča (Masseteric
plate - Palatina Masseterica), 8. nebni odprtini (Foramen incisivum), 9. trdo nebo (Palatum
durum), 10. prvi molar, 11. drugi molar, 12. tretji molar, 13. grebeni na molarjih, 14. kronoidni
odrastek (Processus coronoideus), 15. mandibularni kondil (Processus condylaris) (povzeto po
Shad in Darvish, 2018). ............................................................................................................... 5
Slika 4: Meljaki zgornje čeljusti (Calomyscus) s trinajstimi oslonilnimi točkami: M1 – prvi
meljak, M2 – drugi meljak, M3 – tretji meljak; A,B,C,D,E – grbice na M1; F,G,H,I – grbice na
M2. ........................................................................................................................................... 10
Slika 5: Ventralna stran lobanje (Calomyscus) z dvajsetimi oslonilnimi točkami. ................... 10
Slika 6: Dorzalna stran lobanje (Calomyscus) z enajstimi oslonilnimi točkami. ....................... 11
Slika 7: Lateralna stran lobanje (Calomyscus) s štirinajstimi oslonilnimi točkami. .................. 11
Slika 8: Shematski prikaz poteka Prokrustove analize (začetno stanje, premik v skupno
izhodišče, rotacija ter skaliranje) (povzeto po Klenovšek, 2014). ............................................ 12
Slika 9: Centroidi 13 oslonilnih točk na meljakih Calomyscus. Puščice prikazujejo vektorje
razpršenosti meritev okoli povprečja. ...................................................................................... 15
Slika 10: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
meljakih. ................................................................................................................................... 16
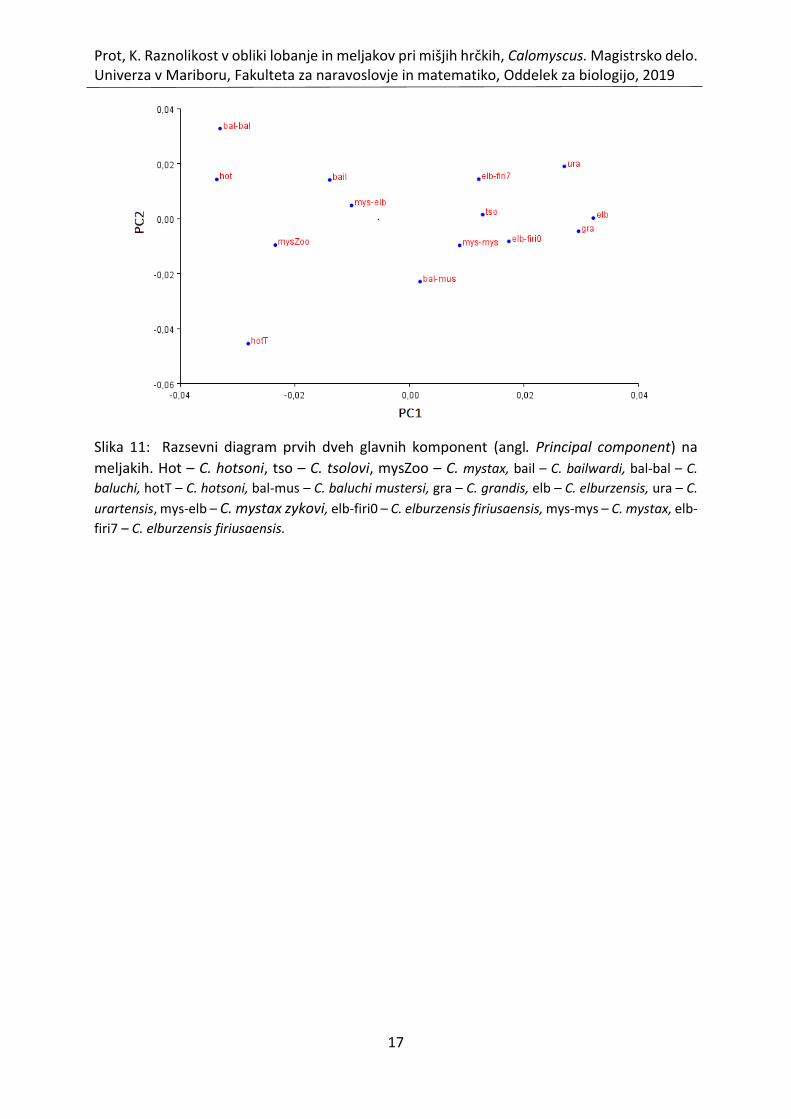
Slika 11: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
meljakih. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C. bailwardi, bal-bal – C.
baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis, elb – C. elburzensis,
ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis firiusaensis, mys-mys
– C. mystax, elb-firi7 – C. elburzensis firiusaensis. ................................................................... 17
Slika 12: Centroidi 20 oslonilnih točk na ventralni strani lobanje Calomyscus. Puščice
prikazujejo vektorje razpršenosti meritev okoli povprečja. ..................................................... 19
Slika 13: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
ventralni strani lobanje. ........................................................................................................... 20
Slika 14: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
ventralni strani lobanje. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C.
bailwardi, bal-bal – C. baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis,
elb – C. elburzensis, ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis
firiusaensis, mys-mys – C. mystax, elb-firi7 – C. elburzensis firiusaensis ................................. 21
Slika 15: Centroidi 11 oslonilnih točk na dorzalni strani lobanje pri 14 osebkih Calomyscus.
Puščice prikazujejo vektorje razpršenosti meritev okoli povprečja. ........................................ 23
Slika 16: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
dorzalni strani lobanje. ............................................................................................................. 24
Slika 17: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
dorzalni strani lobanje. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C. bailwardi,
bal-bal – C. baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis, elb – C.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
IX
elburzensis, ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis
firiusaensis, mys-mys – C. mystax, elb-firi7 – C. elburzensis firiusaensis. ................................ 24
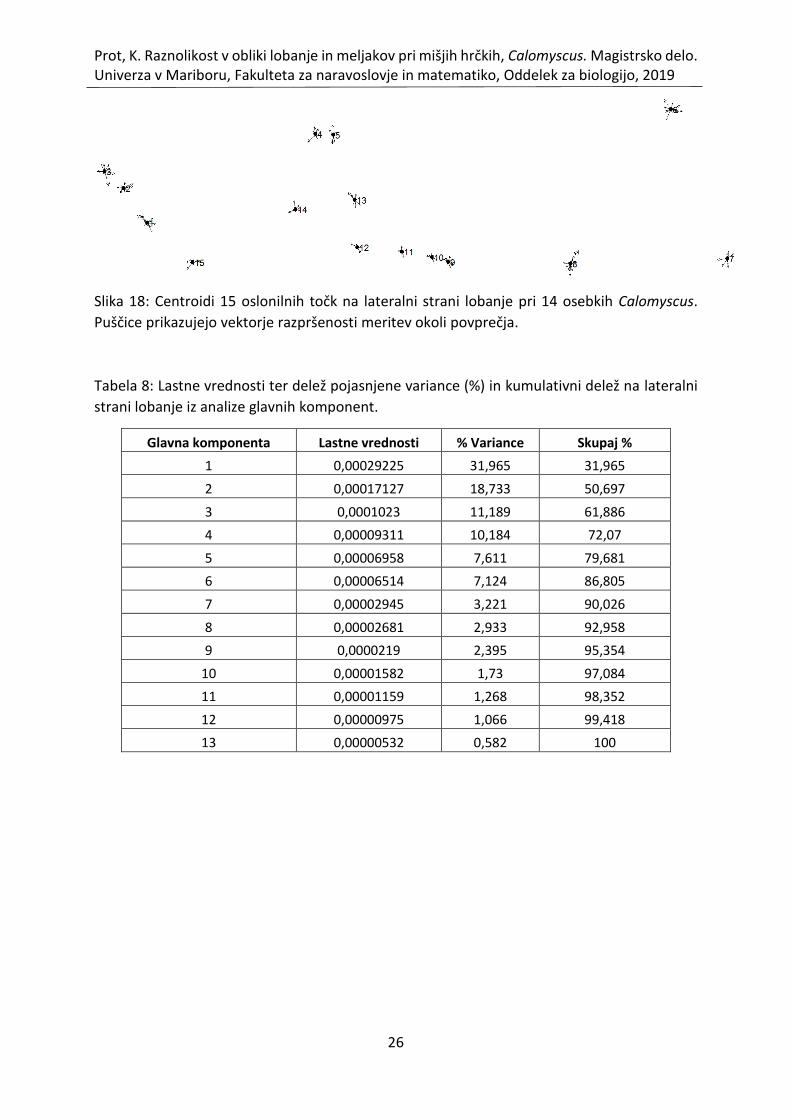
Slika 18: Centroidi 15 oslonilnih točk na lateralni strani lobanje pri 14 osebkih Calomyscus.
Puščice prikazujejo vektorje razpršenosti meritev okoli povprečja. ........................................ 26
Slika 19: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
lateralni strani lobanje. ............................................................................................................ 27
Slika 20: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
lateralni strani lobanje. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C.
bailwardi, bal-bal – C. baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis,
elb – C. elburzensis, ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis
firiusaensis, mys-mys – C. mystax, elb-firi7 – C. elburzensis firiusaensis. ................................ 27
Slika 21: Centroidna velikost meljakov pri rodu Calomyscus. Točke prikazujejo centroidno
velikost meljakov pri posamezni vrsti. ..................................................................................... 28
Slika 22: Centroidna velikost ventralne strani lobanje pri rodu Calomyscus. Posamezna točka
predstavlja centroidno velikost ventralne strani lobanje pri posamezni vrsti......................... 29
Slika 23: Centroidna velikost dorzalne strani lobanje pri rodu Calomyscus. Točke prikazujejo
centroidno velikost dorzalne strani lobanje pri posamezni vrsti. ............................................ 29
Slika 24: Centroidna velikost lateralne strani lobanje pri rodu Calomyscus. Točke prikazujejo
centroidno velikost lateralne strani lobanje pri posamezni vrsti. ............................................ 30
Slika 25: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) v morfoprostoru (na meljakih). Skupine so povezane z drevesom
najkrajšega razvejanja. ............................................................................................................. 32
Slika 26: Vizualizacija variabilnosti v obliki meljakov na podlagi prve glavne komponente (PC
1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti posamezne
anatomske strukture. ............................................................................................................... 32
Slika 27: Primerjava oblike meljakov na podlagi 1. glavne komponente. Svetlejša črta prikazuje
povprečno obliko meljakov pri PC1 = 0, medtem ko temno modra prikazuje značilnosti vrst pri
pozitivni vrednosti PC1 = 0,01 (C. urartensis, C. elburzensis, C. grandis). ................................ 33
Slika 28: Primerjava oblike meljakov na podlagi 1. glavne komponente. Svetlejša črta prikazuje
povprečno obliko meljakov pri PC1 = 0, medtem ko temno modra prikazuje značilnosti vrst pri
negativni vrednosti PC1 = -0,01 (C. baluchi, C. hotsoni). ......................................................... 33
Slika 29: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) (ventralno). Skupine so povezane z drevesom najkrajšega razvejanja.
.................................................................................................................................................. 34
Slika 30: Vizualizacija variabilnosti v obliki ventralne strani lobanje na podlagi prve glavne
komponente (PC 1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti
posamezne anatomske strukture. ............................................................................................ 35
Slika 31: Primerjava ventralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = 0,05
(C. bailwardi, C. elburzensis firiusaensis, C. hotsoni). ............................................................... 35
Slika 32: Primerjava ventralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = -
0,05 (C. elburzensis, C. grandis, C. mystax). ............................................................................. 36

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
X
Slika 33: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) (dorzalno). Skupine so povezane z drevesom najkrajšega razvejanja.
.................................................................................................................................................. 37
Slika 34: Vizualizacija variabilnosti v obliki dorzalne strani lobanje na podlagi prve glavne
komponente (PC 1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti
posamezne anatomske strukture. ............................................................................................ 37
Slika 35: Primerjava dorzalne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = 0,05
(C. bailwardi, C. elburzensis firiusaensis). ................................................................................ 38
Slika 36: Primerjava dorzalne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = -
0,05 (C. tsolovi). ........................................................................................................................ 38
Slika 37: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) (lateralno). Skupine so povezane z drevesom najkrajšega razvejanja.
.................................................................................................................................................. 40
Slika 38: Vizualizacija variabilnosti v obliki lateralne strani lobanje na podlagi prve glavne
komponente (PC 1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti
posamezne anatomske strukture. ............................................................................................ 40
Slika 39: Primerjava lateralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = 0,05
(C. tsolovi). ................................................................................................................................ 41
Slika 40: Primerjava lateralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = -
0,05 (C. elburzensis firiusaensis, C. mystax, C. hotsoni). .......................................................... 41
Slika 41: Dendrogram morfometričnih odnosov med skupinami na meljakih. ....................... 44
Slika 42: Dendrogram morfometričnih odnosov med skupinami na ventralni strani lobanje. 46
Slika 43: Dendrogram morfometričnih odnosov med skupinami na dorzalni strani lobanje. . 48
Slika 44: Dendrogram morfometričnih odnosov med skupinami na lateralni strani lobanje.. 50

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
XI
Kazalo tabel
Tabela 1: Povprečne vrednosti x in y koordinate (centroid) za 13 oslonilnih točk na meljakih
rodu Calomyscus. ..................................................................................................................... 15
Tabela 2: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na meljakih
iz analize glavnih komponent. .................................................................................................. 16
Tabela 3: Povprečne vrednosti x in y koordinate (centroid) za ventralno stran lobanje rodu
Calomyscus. .............................................................................................................................. 19
Tabela 4: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na ventralni
strani lobanje iz analize glavnih komponent. ........................................................................... 20
Tabela 5: Povprečne vrednosti x in y koordinate (centroid) za dorzalno stran lobanje rodu
Calomyscus. .............................................................................................................................. 22
Tabela 6: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na dorzalni
strani lobanje iz analize glavnih komponent. ........................................................................... 23
Tabela 7: Povprečne vrednosti x in y koordinate (centroid) za lateralno stran lobanje rodu
Calomyscus. .............................................................................................................................. 25
Tabela 8: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na lateralni
strani lobanje iz analize glavnih komponent. ........................................................................... 26
Tabela 9: Diagonalna tabela Evklidovih razdalj na meljakih. ................................................... 43
Tabela 10: Diagonalna tabela Evklidovih razdalj na ventralni strani lobanje. ......................... 45
Tabela 11: Diagonalna tabela Evklidovih razdalj na dorzalni strani lobanje. ........................... 47
Tabela 12: Diagonalna tabela Evklidovih razdalj na lateralni strani lobanje. .......................... 49
Tabela 13: Vrednosti korelacijskih koeficientov iz Mantel t testa. .......................................... 50

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
XII
Kazalo prilog
Priloga 1: Seznam proučevanih lobanj osebkov mišjih hrčkov (Calomyscus). Zbirka: SMF –
Naravoslovni muzej Senckenberg , ZFMK – Zoološki raziskovalni muzej Aleksandra Koenig , ZIN
– Zoološki inštitut Ruske akademije znanosti, Sankt Peterburg, BMNH – Prirodoslovni muzej
London. ..................................................................................................................................... 58

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
1
1 Uvod V okviru magistrske naloge sem preučevala morfometrično pestrost v rodu mišjih hrčkov
(Calomyscus) in morfometrično skladnost med posamezni segmenti lobanje. Analize sem
opravljala z metodami geometrijske morfometrije na dorzalni, ventralni in lateralni strani
lobanje ter na zgornjih meljakih.
V uvodu predstavljam biologijo mišjih hrčkov ter njihovo razširjenost. Sledi opis metodologije,
s katero sem prišla do rezultatov. Osrednji del naloge vsebuje analizo rezultatov ter njihovo
razlago v diskusiji.
1.1 Mišji hrčki
Kraljestvo: živali, Animalia
Deblo: strunarji, Chordata
Razred: sesalci, Mammalia
Red: glodavci, Rodentia
Družina: mišji hrčki, Calomyscidae (Vorontsov in Potapova, 1979)
Rod: mišji hrčki, Calomyscus (Thomas, 1905).
Slika 1: Mišji hrček (Calomyscus) (foto J. Vogeltanz), (Kryštufek in Vohralik, 2009).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
2
1.2 Opis
Mišji hrčki (Calomyscus) spadajo med glodavce. Mišje hrčke so na podlagi oblike njihovih meljakov sprva uvrščali med hrčke (družina Cricetidae). Danes jih zaradi odsotnosti obustnih mošnjičkov in bočnih žlez, daljšega repa ter lobanjskih in genetskih posebnosti uvrščamo v lastno družino Calomyscidae (Akbarirad in sod., 2015, Akbarirad in sod., 2016, Kryštufek in Vohralik, 2009).
Prvotno so vse predstavnike rodu Calomyscus prepoznali kot vrsto Calomyscus bailwardi. Z
analizami števila kromosomov, morfometrije lobanje ter drugih se je število vrst v družini
Calomyscidae povečalo na današnjih 8 vrst: C. bailwardi, C. baluchi, C. elburzensis, C. grandis,
C. hotsoni, C. mystax, C. tsolovi, in C. urartensis (Akbarirad in sod. 2016, Kryštufek in Vohralik,
2009, Wilson in sod., 2017, Shahabi, Zarei in Sahebjam, 2010).
1.3 Telesne značilnosti
Mišji hrčki spadajo med glodavce. So majhni in vitki, telo z glavo meri 60–100 mm (v povprečju 78 mm), tehtajo pa 15–30 g (v povprečju 20,4 g). Sekundarni spolni dimorfizem ni izražen. Ušesa so dolga, zaobljena in navadno gola, dolga 17–20 mm. Značilne so relativno dolge srsti, ki lahko zrastejo do 35 mm. Dlaka je na otip fina in mehka, na hrbtu v povprečju dolga 9 mm, na trebuhu pa 6 mm. Rep je daljši od glave in trupa ter meri 72–102 mm (povprečno 87 mm). Rep je dlakast, proti koncu repa postajajo dlake dolge do 12 mm in na konici oblikujejo čopek. Zadnje okončine so daljše (20,5 mm) od prednjih in imajo pet prstov. Na prednjih, krajših, nogah so le štirje prsti (Darvish in sod., 2015, Goodness, 2017, Kryštufek in Vohralik, 2009, Wilson in sod., 2017).
Barva kožuha je rumenkasto rjava, na hrbtu so prisotne temnejše dlake s črnimi konicami, medtem ko so na bokih svetlejše. Glava je svetlejša od hrbta, ušesa so svetlo rjava, z manjšimi belimi zaplatami nad bazo ušesa. Trebuh je bel vse do bokov, bela barva kožuha pa sega skoraj do oči. Zgornja stran nog je belo dlakava, s kratkimi, ostrimi kremplji. Rep je po vrhu, kot hrbet, sivo rumenkasto rjav. Proti konici repa (čopku) se dlake temnejše obarvajo, vse do črne, med njimi se pojavijo še posamezne bele dlake. Ventralna stran repa variira med belo do umazano belo obarvanostjo. Izrazito je definirana razdelitvena črta med ventralno in dorzalno stranjo osebka. Juvenilni osebki so sivi in s staranjem postopoma postajajo rjavi (Darvish in sod., 2015, Goodness, 2017, Hamidi, Darvish, Matin, 2017, Kryštufek in Vohralik, 2009, Wilson in sod., 2017).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
3
1.4 Razširjenost Vrste mišjih hrčkov so razširjenje v jugozahodni Aziji, v delu Azerbajdžana, prek večjega dela
Irana in Afganistana ter na območju severovzhodnega Pakistana. Manjše populacije se
pojavljajo tudi v Turkmenistanu in Turčiji. Mišji hrčki imajo več geografsko izoliranih populacij,
npr. na območju jugovzhodne Sirije. Glede na fosilne ostanke je rod v pliocenu in miocenu
poseljeval tudi Španijo, Francijo, otok Rodos, Kavkaz in Turčijo (Kryštufek in Vohralik, 2009,
Shahabi in sod., 2012, Shahabi in sod., 2013, Zarei in sod., 2013).
Razširjenost posameznih vrst (Kryštufek in Vohralik, 2009):
Calomyscus elburzensis Goodwin, 1938. Območje razširjenosti: gore severovzhodnega
Irana in sosednjih Turkmenistana in Afganistana.
Calomyscus grandis Schlitter in Setzer, 1973.
Območje razširjenosti: severni Iran.
Calomyscus hotsoni Thomas, 1920.
Območje razširjenosti: Balučistan v Pakistanu.
Calomyscus urartensis Vorontsov in Kartavtseva, 1979.
Območje razširjenosti: Azerbajdžan in severozahodni Iran.
Calomyscus mystax Kashkarov, 1925.
Območje razširjenosti: jugozahodni Turkmenistan.
Calomyscus tsolovi Peshev, 1991.
Območje razširjenosti: Sirija.
Calomyscus bailwardi Thomas, 1905.
Območje razširjenosti: Iran.
Calomyscus baluchi Thomas, 1920.
Območje razširjenosti: Afganistan, Pakistan.
Slika 2: Razširjenost vrst rodu Calomyscus (povzeto po Shahabi in sod., 2013).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
4
1.5 Habitat Mišji hrčki so prilagojeni na skalnate habitate. Poseljujejo predvsem suhe gorske habitate ter
nerodovitna, suha skalnata pobočja z redko vegetacijo ali brez nje, najdemo jih v suhih stepah
ter puščavskih predelih. Habitati mišjih hrčkov so porasli s skromno in redko vegetacijo,
sestavljeno iz grmičastih trav, stročnic, osatov, trnastega grmovja. Mišji hrčki so razširjeni na
nadmorskih višinah od 400 do 3500 m. Med vrstami so prisotne razlike v razširjenosti glede na
nadmorsko višino. Calomyscus bailwardi se na primer pojavlja na nadmorski višini od 760 do
2130 m, medtem ko C. baluchi najdemo na višini od 400–3500 m (Hamidi in sod., 2017,
Kryštufek in Vohralik, 2009, Wilson in Reeder, 2005).
1.6 Biologija Zaradi slabše raziskanosti mišjih hrčkov prihaja do ugibanj o obnašanju ter o njihovih bioloških
lastnostih. Mišji hrčki niso najbolj socialni, vendar si v naravi delijo zatočišča. V ujetništvu pa
se pogosteje združujejo v skupine. Osebki so majhni in agilni. Poleg tega so tudi dobri plezalci,
predvsem zaradi ostrih krempljev. Z raziskavami v ujetništvu so ugotovili občutljivost vrste na
temperaturo, ki omejuje njihovo aktivnost na popoldanske ali zgodnje jutranje ure, saj je v
okolju v tem času temperatura nekoliko nižja. Mišji hrčki ne hibernirajo, aktivni so preko celega
leta, vendar se obdobje aktivnosti skozi leto spreminja. C. mystax je spomladi in jeseni aktiven
preko dneva, poleti je aktivnost omejena na nočni čas, Calomyscus urartensis je aktiven
izključno ponoči. Brlog je ozek, horizontalen, izdelan med kamnitimi razpokami. Gnezdo je
zgrajeno iz ovčje volne in fine trave. Ta vrsta pa lahko uporablja tudi že predhodno izkopane
rove drugih vrst (Hamidi in sod., 2017, Goodness, 2017, Kryštufek in Vohralik, 2009).
Prehrana mišjih hrčkov je sestavljena predvsem iz semen, listov, popkov in cvetov.
Prehranjujejo se lahko tudi s hrano živalskega izvora, v ujetništvu pa se pojavi celo kanibalizem.
Pri analizi želodčnih vsebin so najpogosteje našli semena ter rastlinje (Goodness, 2017, Hamidi
in sod., 2017, Kryštufek in Vohralik, 2009).
Razmnoževalno obdobje mišjih hrčkov je odvisno od razpoložljivosti naravnih virov. Pri C.
mystax in C. bailwardi poteka od konca marca do začetka junija. V ujetništvu je brejost samic
29 do 31 dni. V leglu je 1–5 slepih in golih mladičev. Skoteni mladiči so veliki 37 mm ter tehtajo
med 1,53–2,95 g. Razmerje med spoloma je ob rojstvu uravnoteženo. Kožuh juvenilnih
osebkov je siv, s starostjo, v obdobju od 2–8 mesecev, obarvanje nadomestijo z rjavim
odtenkom. Mladiči sesajo do četrtega tedna ter popolnoma odrastejo po osmih mesecih
(Goodness, 2017, Kryštufek in Vohralik, 2009).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
5
Življenjska doba mišjih hrčkov je, kot ostale lastnosti, najbolj raziskana v ujetništvu in znaša 4
leta. Najvišja zabeležena starost v ujetništvu je znašala 9 let, 3 mesece in 18 dni (Goodness,
2017, Kryštufek in Vohralik, 2009).
Najpogostejši plenilci mišjih hrčkov so sove (Otus brucei), kune (Martes foina), dihurji
(Vormela peregusna) in kače (Spalerosophis atriceps, Vipera lebetina, Macrovipera lebetina)
(Goodness, 2017, Kryštufek in Vohralik, 2009).
1.7 Morfologija lobanje in meljakov Lobanja mišjih hrčkov je majhna, ozka, z ozkim in dolgim gobčnim delom (rostrum). Nosni kosti
sta ozki in segata pred sekalce, zadaj pa ne dosežeta očesnih jamic. Območje med očnicama
je široko, nekoliko konveksno, robovi dobro vidni. Na temenu terminalni grebeni niso razviti.
Lični lok je močan, s kratko maseterično ploščo, pri kateri je sprednji rob spuščen. Velikost
podočne odprtine nudi prostor, skozi katerega poteka tudi del žvekalne mišice. Nebni odprtini
sta relativno kratki in ovalni. Rob trdega neba sega do tretjega molarja. Čeljust je po obliki
podobna rodu Mus. Na spodnji čeljustnici je koronoidni odrastek relativno dolg in presega
mandibularni kondil.
Slika 3: Lobanja in meljaki mišjih hrčkov (Calomyscus); 1. rostrum, 2. nosnica (Nasal), 3. sekalci
(Dentes incisivi) a. zgornji sekalec, b. spodnji sekalci, 4. očesne jamice (Orbita), 5. medočesni
prostor (Regio interorbital), 6. lični lok (Arcus zygomaticus), 7. maseterična plošča (Masseteric
plate - Palatina Masseterica), 8. nebni odprtini (Foramen incisivum), 9. trdo nebo (Palatum
durum), 10. prvi molar, 11. drugi molar, 12. tretji molar, 13. grebeni na molarjih, 14. kronoidni
odrastek (Processus coronoideus), 15. mandibularni kondil (Processus condylaris) (povzeto po
Shad in Darvish, 2018).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
6
Za vrste Calomyscus je značilno glodavsko zobovje. V gobcu je 16 zob razporejenih po zobni
formuli 1/1, 0/0, 0/0, 3/3 = 16. Sekalci so stalno rastoči, gladki in stisnjeni, na sprednji površini
zoba je rumenorjav emajl. Molarji mišjih hrčkov so brahiodontni, kar pomeni, da imajo zobje
nizko krono. Prvi molar je največji in najdaljši, medtem ko je tretji molar reduciran. Vzorec na
molarjih pri mišjih hrčkih je podoben vzorcu pri turških predstavnikih iz rodu Cricetinae.
Molarji mišjih hrčkov so ostri in robustni. Prva dva molarja imata dve zobni grbici, obrnjeni
proti jeziku, ter tri proti ličnici. Razlika se pojavi na drugem molarju, saj se sprednja lična grbina
reducira. Tri izbokline so vidne na tretjem molarju, vendar se zaradi hitre obrabe preoblikujejo
v emajliran rob. Meljaki na mandibuli imajo tri izbokline na obeh straneh zoba, prisotna je
redukcija sprednje lične grbine na drugem meljaku. Na tretjem molarju so trije grebeni, pri
čemer ima sprednja grbica emajliran rob (Goodness, 2017, Kryštufek in Vohralik, 2009).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
7
1.8 Geometrijska morfometrija Morfometrija spada med kvantitativne statistične analize, ki temeljijo na raziskovanju
variabilnosti organizmov in oblike njihove telesne zgradbe. Je veja matematike, ki se ukvarja z
analiziranjem oblik organizmov, fosilnih ostankov, sprememb v obliki, ki so predvsem
posledica mutacije, ontogenetskih modifikacij ipd. Modifikacija določene strukture prikaže
informacijo o spremembah evolucijskega razvoja, ekoloških razmer in ontogenetskih
spremembah, kar posledično vpliva na obliko strukture. Začetki morfometrije se pojavijo v
šestdesetih letih prejšnjega stoletja in temeljijo na tradicionalni morfometriji. Ta tip
morfometrije izhaja iz multivariatnih analiz, kjer so se uporabljale meritve dolžin, razdalj ter
kotov med strukturami. Problem je neefektivnost enorazsežnih analiz (tradicionalna
morfometrija), saj ne ustvarijo popolne ureditve oslonilnih točk, pri čemer ne pridobimo
podatkov o geometrijskih odnosih med meritvami (Janžekovič, Klenovšek in Kryštufek, 2007;
Klenovšek, 2014; Slice, 2007; Zelditch, Swiderski, Sheets, 2012).
Konec osemdesetih let dvajsetega stoletja pride v analizi morfoloških struktur do nekaterih
sprememb. Razvije se geometrijska morfometrija, ki omogoči ohranitev informacij o
prostorski razporeditvi struktur ter zajemanje geometrijskih struktur skozi celoten potek
analize. Geometrijska morfometrija torej vsebuje informacije o analizi razlik in je tako
uporabnejša analizna metoda v primerjavi s tradicionalno morfometrijo.
Statistična analiza temelji na Kartezijskih koordinatah, ki so pridobljene z določanjem
oslonilnih točk. Le-te označimo po pravilih na specifičnih mestih proučevanega predmeta in z
njimi opišemo obliko (geometrijo) objektov. Razlike na objektih nastajajo zaradi sprememb v
strukturi organizmov, ki so posledica okolijskih, evolucijskih, ekoloških, ontogenetskih ter
drugih sprememb. Geometrijska morfometrija se odvija v Kendallovem geometrijskem
prostoru. Vsaka točka v prostoru predstavlja obliko konfiguracije točk v Evklidovem prostoru,
ki je neodvisna od orientacije, velikosti in položaja. V Kendallovem prostoru vsak posamezen
osebek predstavlja eno točko, z ozirom na njo pa lahko opišemo povprečno obliko in
variabilnost vzorca. Razdalje v Kendallovem prostoru ocenimo s Prokrustovimi razdaljami.
Geometrijske metode skozi celotno analizo ohranjajo vse informacije, s čimer omogočijo
določitev individualnih in skupinskih razlik, vizualizacijo vzorca itd. (Janžekovič, Klenovšek in
Kryštufek, 2007; Klenovšek, 2014; Slice, 2007; Zelditch, Swiderski, Sheets, 2012).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
8
1.9 Namen dela in hipoteze Namen magistrske naloge je predstaviti morfometrično pestrost med osmimi vrstami mišjih
hrčkov (Calomyscus). Pestrost predstavljamo ločeno za posamezni segment lobanje: ventralni
pogled, dorzalni pogled, lateralni pogled ter zgornje meljake.
Hipoteze
- A priori definirane vrste izkazujejo morfometrično variabilnost v velikosti in v obliki
lobanjskih znakov.
- Posamezni lobanjski segmenti izkazujejo različne morfometrične odnose med a priori
definiranimi vrstami.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
9
2 Material in metode dela
2.1 Material V analize sem vključila 14 lobanj, ki so pripadale 8 vrstam iz rodu Calomyscus (C. bailwardi, C.
baluchi, C. elburzensis, C. grandis, C. hotsoni, C. mystax, C. tsolovi in C. urartensis). Analizirani
material izvira iz Sirije, Irana, Afganistana in Turkmenistana. Osebki so shranjeni v sledečih
muzejskih zbirkah: Naravoslovni muzej Senckenberg (SMF), Zoološki raziskovalni muzej
Aleksandra Koenig (ZFMK), Zoološki inštitut Ruske akademije znanosti, Sankt Peterburg (ZIN),
Prirodoslovni muzej London (BMNH). Fotografije lobanj je pridobil somentor, prof. dr. B.
Kryštufek. Podroben seznam analiziranih osebkov je predstavljen v Prilogi 1.
2.2 Oslonilne točke Meritve sem izvajala na fotografijah lobanj iz treh perspektiv ter okluzalno površino zgornjih
meljakov. Ker so fotografije nastale v različnih muzejih in razdalja med objektom in lečo ni bila
stalna, sem posamezno meritev umerila s pomočjo referenčnega merila. Umeritev sem
opravila s programom tpsDig in tpsUtil.
S pomočjo programa tpsDig sem določila oslonilne točke na posamezni strani lobanje ter na
meljakih. S koordinatami oslonilnih točk opisujemo objekte in so osnova za analize morfološke
variabilnosti. Točke morajo biti na objektu nedvoumno definirane ter homologne na celotnem
materialu, saj podajo pomembne informacije o obliki objekta (Slice, 2007). S pomočjo
literature sem na meljakih izbrala in označila 13, na ventralni strani 20, na dorzalni 11 ter na
lateralni strani lobanje 15 oslonilnih točk. Analizirane strukture so simetrične, zaradi česar sem
točke določevala le na eni polovici posameznega proučevanega predmeta.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
10
Slika 4: Meljaki zgornje čeljusti (Calomyscus) s trinajstimi oslonilnimi točkami: M1 – prvi
meljak, M2 – drugi meljak, M3 – tretji meljak; A,B,C,D,E – grbice na M1; F,G,H,I – grbice na
M2.
1 – prednji rob M1
2 – iztek doline med grbicama A in C
3 – iztek doline med grbicama C in E
4 – stik med meljakoma M1 in M2
5 – iztek doline nad grbico G
6 – iztek doline med grbicama G in I
7 – stik med meljakoma M2 in M3
8 – zadnji rob M3
9 – stik med meljakoma M2 in M3
10 – iztek doline med grbicama H in F
11 – stik med meljakoma M1 in M2
12 – iztek doline med grbicama D in B
13 – iztek doline med grbicama B in A
Slika 5: Ventralna stran lobanje (Calomyscus) z dvajsetimi oslonilnimi točkami.
1 – stik sekalcev
2 – rob alveole sekalca
3 – prednji rob nebne odprtine
4 – zadnji rob nebne odprtine
5 – sutura predčeljustnice in čeljustnice
6 – prednji rob zobne jamice prvega
meljaka
7 – stik zobnih jamic prvega in drugega
meljaka
8 – stik zobnih jamic drugega in tretjega
meljaka
9 – zadnji rob zobne jamice tretjega
meljaka
10 – prehod med gobcem in ličnim lokom
11 – mesto največje izbočenosti očesne
odprtine na prednji strani
12 – mesto največje izbočenosti očesne
odprtine na zadnji strani
13 – zgornji rob slušne odprtine
14 – spodnji rob slušne odprtine
15 – proga na bobničnem mehurju
16 – zaključek bobničnega mehurja
17 – vrh zatilničnega čvršča
18 – sredina zatilnične odprtine
19 – stik med zatilnico in zagozdnico
20 – stik med levo in desno nebnico

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
11
Slika 6: Dorzalna stran lobanje (Calomyscus) z enajstimi oslonilnimi točkami.
1 – stik nosnic (distalno)
2 – stik nosnice in čeljustnice
3 – stik nosnice, čeljustnice in čelnice
4 – stik nosnic (proksimalno)
5 – krivina med gobcem in ličnim lokom
6 – stik čelnice in ličnice
7 – zadnji rob očnice
8 – dorzalni greben stika temenice in
zatilnice
9 – rob zatilnice
10 – stik med levo in desno temenico ter
zatilnico
11 – medialni stik čelnice in temenice
Slika 7: Lateralna stran lobanje (Calomyscus) s štirinajstimi oslonilnimi točkami.
1 – sprednji rob predčeljustnice medialno
(mesto največje vbočenosti med točko 1 in
3)
2 – stik predčeljustnice in nosnice
3 – vrh nosnice
4 – mesto največje vbočenosti loka očesne
odprtine
5 – prednji rob, najbolj levi del očesne
odprtine – dorzalno
6 – stik temenice in zatilnice dorzalno
(pomaknjen popolnoma na vrh lobanje)
7 – osnova zatilnega čvršča dorzalno (prvo
sedlo za zatilnim čvrščem)
8 – levi ekstrem bobničnega mehurja
9 – desni rob alveole 3. meljaka
10 – stičišče 2. in 3. meljaka
11 – stičišče 1. in 2. meljaka
12 – levi rob alveole 1. meljaka
13 – prednji rob očesne odprtine ventralno
14 – najnižja točka prednjega roba
zigomatičnega loka
15 – desni rob stičišča sekalca in zobne
jamice

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
12
2.3 Prokrustova analiza Koordinate oslonilnih točk je za nadaljevanje analiz potrebno poravnati, s čimer zmanjšamo
odstopanja, ki so nastala med fotografiranjem in digitizacijo osebkov. Koordinate sem
poravnala s Prokrustovo analizo (angl. Generalized Procrustes Analysis – GPA) v programu
MorphoJ. GPA analiza točke poravna v skupno izhodišče (premakne, zavrti ter skalira), kar
minimalizira vsoto razlik med točkami v vzorcu (slika 8). Po poteku Prokrustove analize pride
do razdelitve informacije o obliki in velikosti strukture. Oslonilne točke pridobijo nove
koordinate (Prokrustove koordinate), ki razlagajo variabilnost v obliki. Informacijo o velikosti
objekta je zajeta v centroidni velikosti. Ti informaciji omogočata ločeno obdelavo velikosti in
oblike objektov v statistiki (Klenovšek, 2014).
Slika 8: Shematski prikaz poteka Prokrustove analize (začetno stanje, premik v skupno
izhodišče, rotacija ter skaliranje) (povzeto po Klenovšek, 2014).
2.4 Analiza glavnih komponent (PCA) V analizo glavnih komponent sem vpeljala oblikovne spremenljivke, katere sem pridobila v
analizi GPA. Metoda glavnih komponent (PCA – Principal Components Analysis) je statistična
metoda, pri kateri iz množice spremenljivk pridobimo zmanjšano količino spremenljivk.
Novonastale spremenljivke imenujemo glavne komponente (PC – Principal Component), ki
ohranijo kar največ informacij in opišejo kar največ variance iz prvotnih podatkov. Glavne
komponente so linearne kombinacije prvotnih spremenljivk in med seboj niso korelirane. PC
so razporejene glede na delež pojasnjene variance, pri čemer prva glavna komponenta razloži
največji delež variance. V analizah se uporablja nekaj prvih glavnih komponent, katere skupaj
opišejo največji del skupne variance. S to metodo sem izvedla analizo variabilnosti v obliki
meljakov in lobanje, pri čemer se oblikovni prostor preoblikuje tako, da prve glavne
komponente opišejo največ variabilnosti (Klenovšek, 2014, Mitteroecker in Gunz, 2009
Zelditchin sod., 2004).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
13
2.5 Drevo najkrajšega razvejanja (angl. Minimum spanning tree) Graf je sestavljen iz poljubnega števila vozlišč, ki so lahko usmerjena ali neusmerjena. Vozlišča
so med seboj različno oddaljena in imajo neko vsoto, ki poda vrednost, koliko enot
potrebujemo za prehod iz enega vozlišča v drugo. Drevo najkrajšega razvejanja je graf, ki poda
informacijo, koliko enot je potrebnih, da pridemo iz ene točke v drugo (Algorithms, 2018,
Baloh, 2010).
2.6 Klastrska analiza Klastrska analiza (metoda razvrščanja v skupine) je multivariatna statistična metoda, ki
združuje individualna opazovanja v skupine s podobnimi lastnostmi. V skupini so združene čim
bolj podobne enote, ločene skupine pa so si med seboj čim bolj različne. Cilj analize je
zmajševanje enot in posledično njihovega števila. Rezultati so tako na pregleden način
predstavljeni v grafični obliki (Bastič, 2006, Hafner, 2013).
2.7 Grafikon kvantilov – Box plot Box plot (»škatla z brki oz. okvir z ročaji«) se uporablja za prikaz numeričnih meritev. Z grafom
nazorno prikažemo vrednosti centralne tendence ter njihovo varabilnost. Na pravokotnem
grafu je prikazanih pet vrednosti spremenljivke, in sicer: mera srednje vrednoti (Me),
maksimalna (ymax) in minimalna (ymin) vrednost spremenljivke ter prvi (Q1) in drugi (Q2) kvartil.
Z vrednostmi Ymax in ymin, ki odražata tudi varabilnost, lahko izračunamo podatek o relativni
variaciji oz. o variacijskem razmiku, medtem ko s pomočjo razlike med kvartiloma (QR = Q3 –
Q1) izračunamo kvartilni razmik, ki je absolutna mera variabilnosti (Pfajfar, 2011).
2.8 Mantel t test Mantel t test se uporablja za testiranje korelacije neodvisnih elementov med dvema
vzorcema. Test se pogosto uporablja v evolucijski biologiji in ekologiji za preverjanje
povezanosti dveh neodvisnih matrik. Mantel t test dejansko preverja odsotnost povezav med
dvema neodvisnima matrikama. Ničelna hipoteza (H0) pri t testu se glasi: »Razdalje med
objekti v vzorcu Dy niso povezane z ustreznimi razdaljami v Dx.« (Legendre, Fortin in Borcardn,
2015). Alternativna hipoteza (Ha) v tem primeru torej trdi, da so razdalje med objekti
povezane. Stopnja značilnosti je 0,05 (p < 0,05) (Legendre, Fortin in Borcard, 2015).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
14
3 Rezultati V nalogi sem analizirala 14 lobanj pri 8 vrstah iz rodu Calomyscus. Analize sem izvajala na
dorzalni, ventralni in lateralni strani lobanj ter na okuzalni površini zgornjih meljakov. Namen
analize je ovrednotenje morfometrične raznolikosti med vrstami tako glede na obliko kot na
velikost obravnavanih struktur. Rezultate sem razdelila v 3 poglavja; v prvem sem predstavila
Prokrustovo analizo in analizo glavnih komponent, v drugem analizo velikosti, v tretjem pa
analizo oblike ter morfometrične odnose med taksoni. Znotraj teh analiz sem ovrednotila
vsako izmed treh perspektiv lobanje ter okuzalno površino zgornjih meljakov.
3.1 Prokrustova analiza in analiza glavnih komponent Oslonilne točke sem s programom TpsDig določila na ventralni, lateralni, dorzalni strani
lobanje ter na zgornjih meljakih. Program TpsDig umesti oslonilne točke v koordinatni sistem.
S Prokrustovo analizo sem koordinate vseh oslonilnih točk poravnala v skupno izhodišče. S to
analizo sem korigirala različne pogoje, ki so bili posledica različne goriščne razdalje pri
fotografiranju in razlik med postavitvami objektov v optično os. V Prokrustovi analizi se
morfometrična variabilnost razdeli na informacijo o velikosti (centroidna velikost) in
informaciji o obliki (Prokrustove koordinate); posamezni vidik obravnave predstavljam v
ločenih poglavjih. Povprečne vrednosti koordinat oslonilnih točk za meljake, ventralno,
dorzalno in lateralno stran lobanje so prikazane v tabelah 1, 3, 5 in 7.
Z vektorji, ki predstavljajo variabilnost posameznih morfoloških struktur okoli centroida
oslonilne točke, sem predstavila smer in jakost variabilnost strukture. Variabilnost se pojavlja
pri vseh analiziranih straneh lobanje (dorzalno, lateralno in ventralno) ter na meljakih.
Vzroki za raznolikost v velikosti in obliki lobanje so posledica bioloških procesov, ki vplivajo na
rast in razvoj lobanje. Analiza lobanjskih struktur izkazuje opazno variabilnost med vrstami,
kar pomeni, da prihaja do morfometričnega razlikovanja vrst.
3.1.1 Prokrustova analiza in analiza glavnih komponent - meljaki Povprečne vrednosti 13 oslonilnih točk na zgornjih meljakih, ki so definirane s koordinatama
X in Y, so predstavljene v tabeli 1. Razpršenost poravnanih oslonilnih točk prikazuje slika 9, pri
čemer pika predstavlja centroid posamezne oslonilne točke, vektor pa posamezno meritev.
Vektorji, ki so krajši, torej bližje centroidu oslonilne točke, izkazujejo manjšo variabilnost
posamezne strukture ter nasprotno. Vektorji oslonilnih točk so enakomerno do eliptično
razporejeni okrog centroida, večjo variabilnost izkazuje točka 1 – prednji rob M1, 4 – stik med
meljakoma M1 in M2, 5 – iztek doline nad grbico G, 8 – zadnji rob M3, 10 – iztek doline med
grbicama H in F ter 13 – iztek doline med grbicama B in A. Najstabilnejša struktura na meljakih
je iztek doline med grbicama G in I, točka 6.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
15
Vključitev 20 oslonilnih točk na meljakih v PCA vrne 13 glavnih komponent. Prvi dve glavni
komponenti razložita 47,13 % variance v vzorcu, pri čemer prva razloži 27,94 %, druga pa 19,19
%, kar je razvidno iz tabele 2 ter slike 10. Prvih pet glavnih komponent razloži 78,57 %
variabilnosti. Slika 11 prikazuje razsevni diagram osebkov na prvi dve glavni komponenti pri
meljakih. Razvidno je, da morfoprostor ni homogen, oblikovale so se posamezne skupine, ki
se med seboj razhajajo glede na obliko meljakov. Mišji hrčki se glede na prvi dve glavni
komponenti ločijo na 4 skupine. V prvo skupino spadajo osebki z izrazito negativno vrednostjo
prve glavne komponente, in sicer osebki C. baluchi (bal-bal), C. hotsoni (hot), C. mystax
(mysZoo) ter C. hotsoni (hotT). Drugo skupino zastopata osebka C. bailwardi ter C. mystax
zykovi (mys-elb), ki imata manjše negativne vrednosti glede na prvo glavno komponento.
Tretja skupina ima pozitivne vrednosti prve glavne komponente, kamor spadajo osebki C.
baluchi mustersi (bal-mus), C. mystax (mys-mys), C. elburzensis firiusaensis (elb-firi0), C. tsolovi
(tso), C. elburzensis firiusaensi (elb-firi7). V četrto skupino pa spadajo osebki z izrazitejšo
pozitivno vrednostjo prve glavne komponente, kamor spadajo C. urartensis (ura), C.
elburzensis (elb) in C. grandis (gra).
Tabela 1: Povprečne vrednosti x in y koordinate (centroid) za 13 oslonilnih točk na meljakih
rodu Calomyscus.
Oslonilne točke meljaki X Y
1 -0,4276 0,0144
2 -0,3009 0,0870
3 -0,1543 0,0976
4 0,0063 0,0741
5 0,0394 0,0824
6 0,1768 0,0949
7 0,3175 0,0520
8 0,4645 -0,0211
9 0,2917 -0,0662
10 0,1420 -0,1085
11 -0,0197 -0,0916
12 -0,1865 -0,1347
13 -0,3493 -0,0802
Slika 9: Centroidi 13 oslonilnih točk na meljakih Calomyscus. Puščice prikazujejo vektorje
razpršenosti meritev okoli povprečja.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
16
Tabela 2: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na meljakih
iz analize glavnih komponent.
Glavna komponenta Lastne vrednosti % Variance Skupaj %
1 0,0005534 27,939 27,939
2 0,00038008 19,189 47,127
3 0,00028129 14,201 61,328
4 0,00021801 11,006 72,335
5 0,00012353 6,236 78,571
6 0,00010138 5,118 83,689
7 0,00008646 4,365 88,054
8 0,00008162 4,12 92,175
9 0,00005542 2,798 94,973
10 0,00004563 2,303 97,276
11 0,00003134 1,582 98,859
12 0,00001334 0,673 99,532
13 0,00000927 0,468 100
Slika 10: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
meljakih.
Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
17
Slika 11: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
meljakih. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C. bailwardi, bal-bal – C.
baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis, elb – C. elburzensis, ura – C.
urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis firiusaensis, mys-mys – C. mystax, elb-
firi7 – C. elburzensis firiusaensis.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
18
3.1.2 Prokrustova analiza in analiza glavnih komponent - ventralna stran
lobanje Na ventralni strani lobanje sem določila 20 oslonilnih točk. Povprečne vrednosti koordinat
oslonilnih točk, ki so definirane s koordinatami X in Y, so predstavljene v tabeli 3. Slika 12
prikazuje variabilnost ventralne strani lobanje pri analiziranih osebkih. Na ventralni strani so
vektorji enakomerno do eliptično razporejeni okrog centroida. Večja variabilnost se pojavi na
območju očesa (LM 11, 12), ušesa (LM 13, 14, 15, 16) in nevrokranija (LM 17, 18, 19, 20). Na
rostrumu je variabilnost večja le pri LM 4 in LM 10. Najstabilnejši je prednji rob nebne odprtine
LM 3.
Vključitev 20 oslonilnih točk ventralne strani lobanje v PCA vrne 13 glavnih komponent. Prvi
dve glavni komponenti skupaj razložita 53,86 % variance. Skupno prva komponenta razloži
35,43 %, druga pa 18,43 % celotne variabilnosti. Variabilnost preostalih komponent je
predstavljena v tabeli 4. Začetnih pet glavnih komponent skupaj opiše 80,41 % variabilnost
(tabela 4 in slika 13). Morfoprostor ni homogen, kar prikazuje slika 14, saj se oblikujejo 3
skupine, ki se med seboj razlikujejo glede na obliko ventralne strani lobanje.
V prvi skupini so osebki z negativnimi vrednostmi prve glavne komponente C. mystax
(mysZoo), C. mystax zykovi (mys-elb), C. tsolovi (tso), C. grandis (gra) in C. elburzensis (elb), v
drugi skupini je prisoten samo en osebek, C. urartensis (ura), ki ima pozitivno vrednost prve
glavne komponente. V tretjo skupino spadajo osebki z izrazito pozitivnimi vrednostmi prve
glavne komponente, in sicer C. elburzensis firiusaensis (elb-firi7), C. elburzensis firiusaensis
(elb-firi0), C. hotsoni (hot), C. baluchi mustersi (bal-mus), C. mystax (mys-mys), C. baluchi (bal-
bal), C. hotsoni (hotT) ter C. bailwardi (bail).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
19
Tabela 3: Povprečne vrednosti x in y koordinate (centroid) za ventralno stran lobanje rodu
Calomyscus.
Oslonilne točke ventralno X Y
1 -0,3701 -0,0214
2 -0,3624 0,0149
3 -0,2996 -0,0214
4 -0,1333 -0,0370
5 -0,2147 -0,0102
6 -0,1032 0,0175
7 -0,0510 -0,0055
8 -0,0117 -0,0094
9 0,0140 -0,0053
10 -0,1954 0,0237
11 -0,1034 0,0763
12 0,1401 0,1230
13 0,2846 0,0846
14 0,2161 0,0584
15 0,1758 0,0204
16 0,1419 -0,0395
17 0,3581 -0,0167
18 0,3047 -0,0988
19 0,1914 -0,0872
20 0,0181 -0,0665
Slika 12: Centroidi 20 oslonilnih točk na ventralni strani lobanje Calomyscus. Puščice
prikazujejo vektorje razpršenosti meritev okoli povprečja.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
20
Tabela 4: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na ventralni
strani lobanje iz analize glavnih komponent.
Glavna komponenta Lastne vrednosti % Variance Skupaj %
1 0,00033018 35,428 35,428
2 0,00017179 18,433 53,861
3 0,00011138 11,95 65,811
4 0,00007712 8,275 74,086
5 0,00005891 6,321 80,407
6 0,00004233 4,542 84,95
7 0,00003798 4,075 89,025
8 0,00002922 3,136 92,161
9 0,00001992 2,137 94,298
10 0,00001906 2,045 96,343
11 0,00001608 1,725 98,068
12 0,00001232 1,322 99,391
13 0,00000568 0,609 100
Slika 13: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
ventralni strani lobanje.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
21
Slika 14: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
ventralni strani lobanje. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C.
bailwardi, bal-bal – C. baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis,
elb – C. elburzensis, ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis
firiusaensis, mys-mys – C. mystax, elb-firi7 – C. elburzensis firiusaensis

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
22
3.1.3 Prokrustova analiza in analiza glavnih komponent - dorzalna stran
lobanje Na dorzalni strani sem določila 11 oslonilnih točk. Povprečne vrednosti koordinat oslonilnih
točk so prikazane v tabeli 5. Slika 15 prikazuje analizo variabilnosti na dorzalni strani lobanje,
kjer so vektorji enakomerno do eliptično porazdeljeni okoli centroida. Večja variabinost se
izkazuje na rostrumu (LM 1, 2, 3, 4), zadnjem robu očnice (LM 7) ter na stiku med levo in desno
temenico ter zatilnico (LM 10). Najstabilnejši je rob zatilnice (LM 9).
Vključitev 11 oslonilnih točk dorzalne strani lobanje v PCA vrne 13 glavnih komponent. Prvi
dve glavni komponenti dorzalne strani razložita 47,90 % skupne variabilnosti. Prva
komponenta razloži 28,33 %, druga 19,58 % celotne variabilnosti, variabilnost preostalih
komponent pa je predstavljena v tabeli 6. Začetnih pet glavnih komponent opiše 85,05 %
celotne variabilnosti vzorca (tabela 6, slika 16). Slika 17 prikazuje razsevni diagram osebkov na
prvi dve glavni komponenti na dorzalni strani lobanje. Morfoprostor ni homogen, saj so se
oblikovale skupine, ki se med seboj razlikujejo glede na obliko dorzalne strani lobanje.
Projekcija oblikuje štiri skupine, pri čemer se pojavi osamelec, ki odstopa do ostalih. V skupini
z izrazito negativno prvo glavno komponento je prisoten osebek C. tsolovi (tso), v drugi skupini
so osebki z negativno prvo glavno komponento C. mystax (mysZoo) in C. baluchi (bal-bal). V
tretji skupini so osebki z negativno in pozitivno vrednostjo prve glavne komponente C. grandis
(gra), C. mystax (mys-mys), C. mystax zykovi (mys-elb), C. baluchi mustersi (bal-must), C.
urartensis (ura), C. hotsoni (hotT), C. hotsoni (hot), C. elburzensis (elb) ter C. elburzensis
firiusaensis (elb-firi7). V četrti skupini sta osebka z izrazito pozitivnimi vrednostmi prve glavne
komponente C. bailwardi (bail) ter C. elburzensis firiusaensis (elb-firi0).
Tabela 5: Povprečne vrednosti x in y koordinate (centroid) za dorzalno stran lobanje rodu
Calomyscus.
Oslonilne točke dorzalno X Y
1 -0,4631 -0,0346
2 -0,3801 0,0155
3 -0,1539 -0,0275
4 -0,1432 -0,0512
5 -0,1946 0,0248
6 -0,1128 0,0458
7 0,1516 0,1511
8 0,3693 0,0975
9 0,4524 -0,0815
10 0,3190 -0,0735
11 0,1554 -0,0664

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
23
Slika 15: Centroidi 11 oslonilnih točk na dorzalni strani lobanje pri 14 osebkih Calomyscus.
Puščice prikazujejo vektorje razpršenosti meritev okoli povprečja.
Tabela 6: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na dorzalni
strani lobanje iz analize glavnih komponent.
Glavna komponenta Lastne vrednosti % Variance Skupaj %
1 0,00025136 28,328 28,328
2 0,00017371 19,577 47,904
3 0,00013806 15,56 63,464
4 0,0001319 14,865 78,329
5 0,00005966 6,723 85,052
6 0,00005201 5,861 90,913
7 0,00003185 3,59 94,503
8 0,00002237 2,521 97,024
9 0,00001381 1,556 98,58
10 0,00000541 0,61 99,19
11 0,00000338 0,381 99,571
12 0,00000283 0,319 99,89
13 0,00000098 0,11 100

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
24
Slika 16: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
dorzalni strani lobanje.
Slika 17: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
dorzalni strani lobanje. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C. bailwardi,
bal-bal – C. baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis, elb – C.
elburzensis, ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis
firiusaensis, mys-mys – C. mystax, elb-firi7 – C. elburzensis firiusaensis.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
25
3.1.4 Prokrustova analiza in analiza glavnih komponent - lateralna stran
lobanje Na lateralni strani lobanje sem določila 15 oslonilnih točk. Povprečne vrednosti koordinat
oslonilnih točk so prikazane v tabeli 7. Slika 18 prikazuje variabilnost lateralne strani lobanje
pri analiziranih osebkih. Vektorji oslonilnih točk so enakomerno do eliptično razporejeni okoli
centroida. Znotraj posameznih točk so stabilnejši predvsem meljaki (LM 9, 10, 11, 12). Močna
variabilnost je prisotna pri LM 1 (sprednji rob predčeljustnice medialno), 3 (vrh nosnice), 6
(stik temenice in zatilnice dorzalno), 7 (osnova zatilnega čvršča dorzalno), 8 (levi ekstrem
bobničnega mehurja).
Vključitev 20 oslonilnih točk na lateralni strani lobanje vrne 13 glavnih komponent. Prvi dve
glavni komponenti (slika 19) skupaj razložita 50,7 % celotne variance, pri čemer prva razloži
31,97 %, druga pa 18,73 %. Začetnih pet komponent razloži 79,68 % celotne variance.
Variabilnost preostalih komponent je predstavljena v tabeli 8. Slika 20 prikazuje razsevni
diagram osebkov na prvi dve glavni komponenti na lateralni strani lobanje. Razvidno je, da
morfoprostor ni homogen, oblikovale so se posamezne skupine, ki se razhajajo glede na obliko
lateralne strani lobanje.
Oblikujejo se štiri skupine osebkov, pri čemer ena vrsta opazno odstopa od ostalih. V prvi
skupini so živali z izrazito negativnimi vrednostmi prve glavne komponente: C. elburzensis
firiusaensis (elb-firi7), C. mystax (mys-mys) in C. hotsoni (hot). V drugi skupini so osebki z
negativnimi vrednostmi: C. elburzensis firiusaensis (elb-firi0), C. baluchi (bal-bal), C. urartensis
(ura) in C. baluchi mustersi (bal-mus). V tretji pa osebki s pozitivnimi vrednostmi prve glavne
komponente: C. mystax zykovi (mys-elb), C. bailwardi (bail), C. hotsoni (hotT), C. mystax
(mysZoo), C. grandis (gra) ter C. elburzensis (elb). Prisoten je tudi osamelec C. tsolovi (tso), ki
ima močne pozitivne vrednosti glavne komponente.
Tabela 7: Povprečne vrednosti x in y koordinate (centroid) za lateralno stran lobanje rodu
Calomyscus.
Oslonilne točke lateralno X Y
1 -0,2998 -0,0247
2 -0,3334 0,0188
3 -0,3602 0,0398
4 -0,0716 0,0983
5 -0,0469 0,0982
6 0,4166 0,1466
7 0,5008 -0,0442
8 0,2855 -0,0583
9 0,1166 -0,0615
10 0,0943 -0,0561
11 0,0524 -0,0502
12 -0,0090 -0,0468
13 -0,0144 0,0145
14 -0,0957 -0,0003
15 -0,2352 -0,0741
Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
26
Slika 18: Centroidi 15 oslonilnih točk na lateralni strani lobanje pri 14 osebkih Calomyscus.
Puščice prikazujejo vektorje razpršenosti meritev okoli povprečja.
Tabela 8: Lastne vrednosti ter delež pojasnjene variance (%) in kumulativni delež na lateralni
strani lobanje iz analize glavnih komponent.
Glavna komponenta Lastne vrednosti % Variance Skupaj %
1 0,00029225 31,965 31,965
2 0,00017127 18,733 50,697
3 0,0001023 11,189 61,886
4 0,00009311 10,184 72,07
5 0,00006958 7,611 79,681
6 0,00006514 7,124 86,805
7 0,00002945 3,221 90,026
8 0,00002681 2,933 92,958
9 0,0000219 2,395 95,354
10 0,00001582 1,73 97,084
11 0,00001159 1,268 98,352
12 0,00000975 1,066 99,418
13 0,00000532 0,582 100

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
27
Slika 19: Delež pojasnjene variabilnosti glavnih komponent (angl. Principal components) na
lateralni strani lobanje.
Slika 20: Razsevni diagram prvih dveh glavnih komponent (angl. Principal component) na
lateralni strani lobanje. Hot – C. hotsoni, tso – C. tsolovi, mysZoo – C. mystax, bail – C.
bailwardi, bal-bal – C. baluchi, hotT – C. hotsoni, bal-mus – C. baluchi mustersi, gra – C. grandis,
elb – C. elburzensis, ura – C. urartensis, mys-elb – C. mystax zykovi, elb-firi0 – C. elburzensis
firiusaensis, mys-mys – C. mystax, elb-firi7 – C. elburzensis firiusaensis.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
28
3.2 Analiza velikosti Za analizo velikosti lobanje in meljakov na posameznih osebkih rodu Calomyscus sem
uporabila vrednosti centroidnih velikosti. Na podlagi analize velikosti meljakov, lateralne,
dorzalne in ventralne strani lobanje (slike 21, 22, 23 in 24) vidimo izrazito odstopanje pri vrsti
C. tsolovi, ki je najmanjša v primerjavi z osebki ostalih vrst. Manjša dorzalna in lateralna
velikost lobanje se kaže tudi pri vrsti C. elburzensis firiusaensis. Glede na centroidno velikost
na vseh straneh lobanje vrsta C. baluchi mustersi sodi med osebke z večjimi velikostmi lobanje.
Živali, pri katerih je prisotno več osebkov iste vrste (C. hotsoni, C. mystax, C. elburzensis
firiusaensis), je prisotna variabilnost v velikosti posamezne strukture.
3.2.1 Analiza velikosti - meljaki Analiza velikosti na meljakih (slika 21) prikazuje ločitev osebkov v dve skupini glede na velikost:
manjšo in večjo. Izrazito odstopa vrsta C. tsolovi, ki je bistveno manjša od ostalih vrst. Med
večjimi osebki v velikosti meljakov prihaja do manjše variabilnosti.
Slika 21: Centroidna velikost meljakov pri rodu Calomyscus. Točke prikazujejo centroidno
velikost meljakov pri posamezni vrsti.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
29
3.2.2 Analiza velikosti - ventralna stran lobanje Osebki se glede na analizo velikosti ventralne strani lobanje ločijo na večje in manjše osebke
(slika 22). V skupino velikih osebkov spadajo vsi osebki, razen vrsta C. tsolovi, saj je le-ta
bistveno manjša od ostalih. Med večjimi vrstami prihaja do variabilnosti v velikosti lobanje, pri
čemer ima največjo lobanjo C. mystax.
Slika 22: Centroidna velikost ventralne strani lobanje pri rodu Calomyscus. Posamezna točka
predstavlja centroidno velikost ventralne strani lobanje pri posamezni vrsti.
3.2.3 Analiza velikosti - dorzalna stran lobanje Na dorzalni strani lobanje se glede na centroidno velikost lobanje formirata dve skupini
osebkov, in sicer manjša in večja skupina (slika 23). V manjšo skupino spadata vrsti C. tsolovi
ter C. elburzensis firiusaensis. Dorzalna stran lobanje je največja pri vrsti C. baluchi mustersi.
Slika 23: Centroidna velikost dorzalne strani lobanje pri rodu Calomyscus. Točke prikazujejo
centroidno velikost dorzalne strani lobanje pri posamezni vrsti.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
30
3.2.4 Analiza velikosti - lateralna stran lobanje Analiza centroidne velikosti na lateralni strani lobanje uvršča osebke v dve skupini. V skupini z
velikimi osebki so vsi osebki razen C. tsolovi ter C. elburzensis firiusaensis. Največji osebek je
C. baluchi mustersi in najmanjši C. tsolovi (slika 24).
Slika 24: Centroidna velikost lateralne strani lobanje pri rodu Calomyscus. Točke prikazujejo
centroidno velikost lateralne strani lobanje pri posamezni vrsti.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
31
3.3 Analiza oblike Analizo oblike lobanje in meljakov sem na rodu Calomyscus spremljala s pomočjo prve in druge
glavne komponente. Vrste so glede na prvi dve glavni komponenti razporejeni v morfoprostor.
Osebki so v morfoprostoru povezani z drevesom najkrajšega razvejanja, kar pomeni, da so poti
najkrajše od začetnega osebka (vozlišča) do vseh ostalih osebkov (vozlišč).
3.3.1 Analiza oblike - meljaki Med mišjimi hrčki prihaja do razlik na meljakih, kar prikazuje diagram prvih dveh glavnih
komponent (slika 25). Osebki se v morfoprostoru razporedijo v območje z negativnimi in
pozitivnimi vrednostmi prve glavne komponente. V negativno območje PC1 so uvrščene vrste:
C. baluchi, C. bailwardi ter dva predstavnika C. hotsoni in C. mystax. V pozitivno območje pa
so uvrščeni osebki: C. uratensis, C. tsolovi, C. mystax, C. elburzensis, C. grandis ter dva
predstavnika C. elburzensis firiusaensis. Vrednosti okoli nič so prisotne pri vrsti C. baluchi
mustersi. Med osebki se pojavlja medvrstna variabilnost. V razsevnem diagramu (slika 25) so
osebki povezani z drevesom najkrajšega razvejanja. Ta metoda predstavlja kontrolo za
razporeditev vrst vzdolž glavnih komponent.
Slika 26 prikazuje variabilnost posameznih oslonilnih točk na meljakih. Variabilnejše so
strukture LM 1 (prednji rob M1), LM 5 (iztek doline na grbico G), LM8 (zadnji rob M3) in LM
13 (iztek doline med grbicama B in A).
Variabilnost meljakov, katero razlaga prva glavna komponenta, sem dodatno razložila s
slikama 27 in 28. Z analizo sem spremljala oblikovne značilnosti meljakov. S spreminjanjem
PC1 med pozitivnimi (PC1 = 0,01) in negativnimi (PC1 = -0,01) vrednostmi prihaja do razlik v
obliki meljakov. Osebki s pozitivnimi PC vrednostmi imajo v primerjavi z negativnimi
vrednostmi od povprečnega vzorca manjši prvi meljak ter večji drugi in tretji meljak. Na stičišču
prvega in drugega meljaka se LM 4 (stik med meljakoma M1 in M2) in 5 (iztek doline nad grbico
G) pri pozitivnih vrednostih skoraj popolnoma približata drug drugemu. Negativna vrednost ti
dve točki loči. Razlika v variabilnosti pri M2 je v velikosti in obliki. PC1 = 0,01 imajo meljak večji
in je razširjen na območju točk 6 in 10. Na sliki 28 pri PC1 = -0,01 je M2 večji na stiku z M1 in
se do stika z M3 oži. Spreminjata se tudi velikost in oblika M3 v točki 8 – zadnji rob M3. Osebki
z negativno PC1 vrednostjo imajo v primerjavi s pozitivno točko 8 inferiorno. Meljak je pri
negativnih vrednostih ožji.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
32
Slika 25: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) v morfoprostoru (na meljakih). Skupine so povezane z drevesom
najkrajšega razvejanja.
Slika 26: Vizualizacija variabilnosti v obliki meljakov na podlagi prve glavne komponente (PC
1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti posamezne
anatomske strukture.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
33
Slika 27: Primerjava oblike meljakov na podlagi 1. glavne komponente. Svetlejša črta prikazuje
povprečno obliko meljakov pri PC1 = 0, medtem ko temno modra prikazuje značilnosti vrst pri
pozitivni vrednosti PC1 = 0,01 (C. urartensis, C. elburzensis, C. grandis).
Slika 28: Primerjava oblike meljakov na podlagi 1. glavne komponente. Svetlejša črta prikazuje
povprečno obliko meljakov pri PC1 = 0, medtem ko temno modra prikazuje značilnosti vrst pri
negativni vrednosti PC1 = -0,01 (C. baluchi, C. hotsoni).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
34
3.3.2 Analiza oblike - ventralna stran lobanje Razporeditev osebkov na prvi dve glavni komponenti ventralne strani lobanje je prikazana na
sliki 29. Osebki se v morfoprostoru razporedijo v območje z negativnimi in pozitivnimi
vrednostmi prve glavne komponente. Glede na PC1 so v negativnem območju uvrščene vrste:
C. tsolovi, C. elburzensis, C. grandis ter dva predstavnika C. mystax. Pozitivne vrednosti PC1
imajo taksoni: C. baluchi mustersi, C. mystax, C. baluchi, C. bailwardi oba predstavnika C.
elburzensis firiusaensis ter C. hotsoni. Vrednosti okoli nič se pojavljajo pri vrsti C. urartensis.
Med osebki se pojavlja medvrstna variabilnost. Na razsevnem diagramu (slika 29) so osebki
povezani z drevesom najkrajšega razvejanja, kar pomeni, da so poti najkrajše od začetnega
osebka (vozlišča) do vseh ostalih osebkov (vozlišč).
Slika 30 prikazuje variabilnost posameznih oslonilnih točk na ventralni strani lobanje.
Najstabilnejša struktura je prednji rob nebne odprtine (točka 3), medtem ko so variabilnejše
strukture na področju očesa (LM 11, 12), ušesa (LM 13, 14, 15, 16) ter nevrokranija (LM 17,
18, 19, 20).
Sliki 31 in 32 prikazujeta oblikovno značilnost ventralne strani lobanje, katero razlaga PC1. S
spreminjanjem PC1 med pozitivnimi (0,05) in negativnimi (-0,05) vrednostmi prihaja do razlike
v obliki ventralne lobanje. Osebki s pozitivno PC1 imajo krajši rostrum, prehod med rostrumom
(LM 10) in očesno odprtino (LM 11) je v primerjavi z negativnimi vrednostmi PC1 večji. Na
ventralni strani lobanje (PC = 0,05) je posteriorni del lobanje bolj zaobljen ter bolj izstopa
kavdalno v LM 18. Predel ušesa (LM 14, 15, 16) je bližje posteriornemu robu lobanje,
nasprotno je pri negativnih osebkih, kjer je ta predel pomaknjen bližje rostrumu
(anteriornemu delu lobanje).
Slika 29: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) (ventralno). Skupine so povezane z drevesom najkrajšega razvejanja.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
35
Slika 30: Vizualizacija variabilnosti v obliki ventralne strani lobanje na podlagi prve glavne
komponente (PC 1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti
posamezne anatomske strukture.
Slika 31: Primerjava ventralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = 0,05
(C. bailwardi, C. elburzensis firiusaensis, C. hotsoni).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
36
Slika 32: Primerjava ventralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = -
0,05 (C. elburzensis, C. grandis, C. mystax).
3.3.3 Analiza oblike - dorzalna stran lobanje Razporeditev osebkov na prvi dve glavni komponenti dorzalne strani lobanje je prikazana na
sliki 33. Osebki se v morfoprostoru razporedijo v območje z negativnimi in pozitivnimi
vrednostmi prve glavne komponente. V negativen morfoprostor so uvrščene vrste: C. tsolovi,
C. baluchi, C. grandis, ter C. mystax, v pozitivnega pa: C. baluchi mustersi, C. urartensis, C.
hotsoni, C. bailwardi ter predstavnika vrste C. elburzensis firiusaensis. Vrednosti okoli nič se
pojavijo pri C. hotsoni, C. elburzensis ter dveh predstavnikih C. mystax. Med osebki se pojavlja
medvrstna variabilnost. V razsevnem diagramu (slika 33) so osebki povezani z drevesom
najkrajšega razvejanja. Na dorzalni strani lobanje se C. tsolovi razporedi izrazito v negativen
morfoprostor in je edina vrsta s tako negativnimi vrednostmi.
Variabilnost posameznih oslonilnih točk na dorzalni strani lobanje prikazuje slika 34.
Variabilnejše so strukture z LM 1 – stik nosnic (distalno), 3 – stik nosnice, čeljustnice in čelnice,
4 – stik nosnic (proksimalno) ter 11 – medialni stik čelnice in temenice. Najstabilnejša je točka
7 – zadnji rob očnice.
Variabilnost dorzalne strani lobanje, katero razlaga PC1, sem dodatno razložila s slikama 35 in
36. Spremembe v obliki sem analizirala s spreminjanjem PC1 med pozitivnimi (PC1 = 0,05) in
negativnimi (PC1 = -0,05) vrednostmi. Osebki s pozitivnimi vrednostmi PC1 imajo v primerjavi
z negativnimi daljši rostrum (LM 1 in 2). Stik nosnice, čeljustnice in čelnice (LM 3) ter stik
nosnice (LM 4) je usmerjen posteriorno. Stik čelnice in ličnice (LM 6) se pri živalih s pozitivno
PC1 proksimalno vboči (proti središču lobanje). Razlika se pojavlja tudi pri LM 9 (rob zatilnice)
in LM 11 (medialni stik čelnice in temenice). LM 9 se pri pozitivnih vrednostih posteriorno
spušča, medtem ko se LM 11 premakne anteriorno na lobanjo. Pozitivne živali imajo stik
temenice in zatilnice (LM 8) dorzalno razširjen, rob zatilnice (LM 9) je posteriorno izbočen.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
37
Slika 33: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) (dorzalno). Skupine so povezane z drevesom najkrajšega razvejanja.
Slika 34: Vizualizacija variabilnosti v obliki dorzalne strani lobanje na podlagi prve glavne
komponente (PC 1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti
posamezne anatomske strukture.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
38
Slika 35: Primerjava dorzalne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = 0,05
(C. bailwardi, C. elburzensis firiusaensis).
Slika 36: Primerjava dorzalne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = -
0,05 (C. tsolovi).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
39
3.3.4 Analiza oblike – lateralna stran lobanje Razporeditev osebkov na prvi dve glavni komponenti lateralne strani lobanje je prikazana na
sliki 37. Osebki se v morfoprostoru razporedijo v območje z negativnimi in pozitivnimi
vrednostmi prve glavne komponente. Vrste z negativnimi PC1 vrednostmi so: C. hotsoni, C.
mystax, C. baluchi, C. baluchi mustersi ter C. elburzensis firiusaensis in s pozitivnimi: C.
bailwardi, C. hotsoni, C. grandis, C. elburzensis, C. tsolovi ter C. mystax. Osebki z vrednostmi
okoli nič so: C. elburzensis firiusaensis, C. mystax, in C. urartensis. Prisotna je znotrajvrstna
variabilnost. Razsevni diagram (slika 37) prikazuje povezanost osebkov z drevesom najkrajšega
razvejanja. Na lateralni strani je na območje s pozitivnimi PC1 izrazita ločitev C. tsolovi.
Variabilnosti posameznih oslonilnih točk na lateralni strani lobanje prikazuje slika 38. Velika
variabilnost je prisotna na LM 1 – sprednji rob predčeljustnice medialno (mesto največje
vbočenosti med točko 1 in 3), 3 – vrh nosnice, 5 – prednji rob, najbolj levi del očesne odprtine
– dorzalno, 6 – stik temenice in zatilnice dorzalno (pomaknjen popolnoma na vrh lobanje), 7 –
osnova zatilnega čvršča dorzalno (prvo sedlo za zatilnim čvrščem) in 8 – skrajno levi ekstrem
bobničnega mehurja.
Variabilnost lateralne strani lobanje, katero razlaga prva glavna komponenta, sem dodatno
razložila s slikama 39 in 40. Anteriorni del lobanje je pri pozitivnih vrednostih prve glavne
komponente (PC1 = 0,05) spuščen, pri čemer se stik predčeljustnice in nosnice (LM 2) ter vrh
nosnice (LM 3) približata. Sprednji rob predčeljustnice medialno (LM 1) je orientiran nižje.
Posteriorni del lobanje je večji v primerjavi z negativnimi vrednostih PC1. Superiorno so
privzdignjeni LM 4 (mesto največje vbočenosti loka očesne odprtine), LM 5 (prednji rob,
najbolj levi del očesne odprtine – dorzalno) ter LM 6 (stik temenice in zatilnice dorzalno).
Inferiorno sta spuščeni LM 7 (osnova zatilnega čvršča dorzalno) ter LM 8 (skrajno levi ekstrem
bobničnega mehurja). Meljaki so glede na pozitivne in negativne vrednosti PC1 relativno
enakomerno razdeljeni. LM 13 (prednji rob očesne odprtine ventralno) in 14 (najnižja točka
prednjega roba zigomatičnega loka) sta superiorni, prav tako je vbočen desni rob stičišča
sekalca in zobne jamice (LM 15).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
40
Slika 37: Razsevni diagram 14 osebkov rodu Calomyscus na osnovi prvih dveh glavnih
komponent (PC1 in PC 2) (lateralno). Skupine so povezane z drevesom najkrajšega razvejanja.
Slika 38: Vizualizacija variabilnosti v obliki lateralne strani lobanje na podlagi prve glavne
komponente (PC 1). Pika predstavlja povprečno obliko, vektor pa smer in jakost variabilnosti
posamezne anatomske strukture.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
41
Slika 39: Primerjava lateralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = 0,05
(C. tsolovi).
Slika 40: Primerjava lateralne strani lobanje na podlagi 1. glavne komponente. Svetlejša črta
prikazuje povprečno obliko lobanje pri PC1 = 0, medtem ko temno modra prikazuje PC1 = -
0,05 (C. elburzensis firiusaensis, C. mystax, C. hotsoni).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
42
3.4 Morfometrični odnosi med taksoni Morfometrične odnose med 14 OTU sem predstavila s pomočjo Evklidovih razdalj za vsak
segment posebej in so predstavljene v tabelah od 9 do 12. Diagonalne tabele Evklidovih razdalj
sem vključila v klastersko analizo, s katero sem odnose med posameznimi OTU-ji predstavila v
obliki dendrograma.
Izhajala sem iz domneve, da vsi lobanjski segmenti izkazujejo podobne morfološke donose. To
domnevo sem preverila s primerjanjem podobnosti med Evklidovimi razdaljami. Testiranje
sem izvedla s pomočjo Mantel t testa.
3.4.1 Morfometrični odnosi med taksoni - meljaki Morfometrični odnosi na meljakih so predstavljeni v diagonalni tabeli 9. S pomočjo Evklidovih
razdalj sem primerjala 14 OTU med seboj. Na podlagi Evklidovih razdalj meljakov se najbolj
razlikujeta vrsti C. hotsoni ter C. urartensis (razdalja 0,0895). Najmanjše Evklidove razdalje so
med vrstami C. mystax zykovi in C. mystax (razdalja 0,0402) ter C. hotsoni (hot) in C. bailwardi
(bail) (razdalja 0,0423).
Dendrogram morfometričnih odnosov na meljakih je predstavljen s sliko 41. Razdalje med OTU
so podane z Evklidskimi razdaljami. Iz dendrograma je razvidno odstopanje vrste C. hotsoni
(hotT) od ostalih vrst, ki se delijo v štiri skupine. V prvo skupino spadata sestrski vrsti C. baluchi
mustersi (bal-mu) in C. elburzensis firiusaensis (elb-firi0), od katere se je odcepil C. tsolovi (tso).
Drugo skupino sestavljata dve sestrski veji, in sicer C. grandis (gra) in C. urartensis (ura) ter C.
elburzensis (elb) in C. elburzensis firiusaensis (elb-firi7). Tretjo skupino sestavljajo vrste C.
mystax, pri čemer se od sestrske skupine C. mystax (mysZoo) in C. mystax zykovi (mys-elb)
odcepi C. mystax (mys-mys). Zadnjo skupino sestavljajo C. bailwardi (bail), C. hotsoni (hot) in
C. baluchi (bal-bal). Meljaki vrst C. mystax (mysZoo) in C. mystax elburnensis so si med seboj
najbolj podobni.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
43
Tabela 9: Diagonalna tabela Evklidovih razdalj na meljakih.
hot tso mysZoo bail bal-bal hotT bal-mus gra elb ura mys-elb elb-firi0 mys-mys elb-firi7
hot 0
tso 0,0631 0
mysZoo 0,0490 0,0612 0
bail 0,0423 0,0659 0,0579 0
bal-bal 0,0440 0,0768 0,0595 0,0548 0
hotT 0,0685 0,0829 0,0607 0,0713 0,0824 0
bal-mus 0,0607 0,0500 0,0488 0,0598 0,0756 0,0579 0
gra 0,0770 0,0573 0,0702 0,0613 0,0784 0,0751 0,0554 0
elb 0,0775 0,0662 0,0728 0,0645 0,0830 0,0816 0,0623 0,0437 0
ura 0,0676 0,0536 0,0714 0,0584 0,0704 0,0895 0,0604 0,0439 0,0493 0
mys-elb 0,0557 0,0639 0,0402 0,0600 0,0557 0,0725 0,0574 0,0658 0,0600 0,0610 0
elb-firi0 0,0688 0,0546 0,0559 0,0593 0,0754 0,0744 0,0458 0,0503 0,0579 0,0469 0,0600 0
mys-mys 0,0722 0,0716 0,0604 0,0741 0,0782 0,0793 0,0604 0,0682 0,0626 0,0678 0,0508 0,0546 0
elb-firi7 0,0596 0,0565 0,0553 0,0556 0,0575 0,0820 0,0524 0,0496 0,0435 0,0491 0,0491 0,0500 0,0515 0

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
44
Slika 41: Dendrogram morfometričnih odnosov med skupinami na meljakih.
3.4.2 Morfometrični odnosi med taksoni - ventralna stran lobanje Morfometrični odnosi na ventralni strani lobanje so predstavljeni v tabeli 10. Na ventralni
strani lobanje so v primerjavi z meljaki Evklidove razdalje krajše. Največje razdalje se pojavijo
med C. mystax (mysZoo) ter C. elburzensis firiusaensis (elb-firi0) (razdalja 0,0612), C. mystax
(mysZoo) in C. elburzensis firiusaensis (elb-firi7) (razdalja 0,0630) ter C. grandis (gra) in C.
bailwardi (bail) (razdalja 0,0567). Manjše razdalje so med vrstami C. hotsoni (hot) in C. baluchi
(bal-bal) (razdalja 0,0233), C. hotsoni (hot) in C. mystax (mys-mys) (razdalja 0,0234) ter C.
baluchi (bal-bal) in C. baluchi mustersi (bal-mus) (razdalja 0,0255).
Dendrogram morfometričnih odnosov na ventralni strani lobanje je predstavljen s sliko 42.
Razdalje med OTU so predstavljene z Evklidovimi razdaljami. Na ventralni strani lobanje se na
dendrogramu med osebki oblikujejo tri večje skupine. Najbolj podobna sta si osebka C. baluchi
(bal-bal) in C. hotsoni (hot). Na dendrogramu je razvidna relativno podobna skupina šestih
predstavnikov vrst, in sicer sestrski vrsti C. baluchi (bal-bal), C. hotsoni (hot), od katerih se
odcepita C. mystax (mys-mys), C. baluchi mustersi (bal-mus) ter sestrska skupina C. bailwardi
(bail), C. hotsoni (hotT). Osebka C. uratensis (ura) in C. elburzensis firiusaensis (elb-firi7) tvorita
sestrsko skupino, od katere se loči C. elburzensis firiusaensis (elb-firi0). Druga skupina se je
predhodno ločila od prve skupine. Na podlagi Evklidovih razdalj se je najprej ločila tretja
skupina, kamor spadajo sestrske skupine C. grandis (gra), C. elburzensis (elb) ter C. mystax
(mysZoo) in C. mystax zykovi (mys-elb), od katerih se loči C. tsolovi (tso). Glede na dendrogram
lahko opazimo, da prihaja do medvrstnih razlik med osebki C. mystax ter razlik med vrsto C.
elburzensis (elb) in njenimi podvrstami, C. elburzensis firiusaensis (elb-firi0 in elb-firi 7).
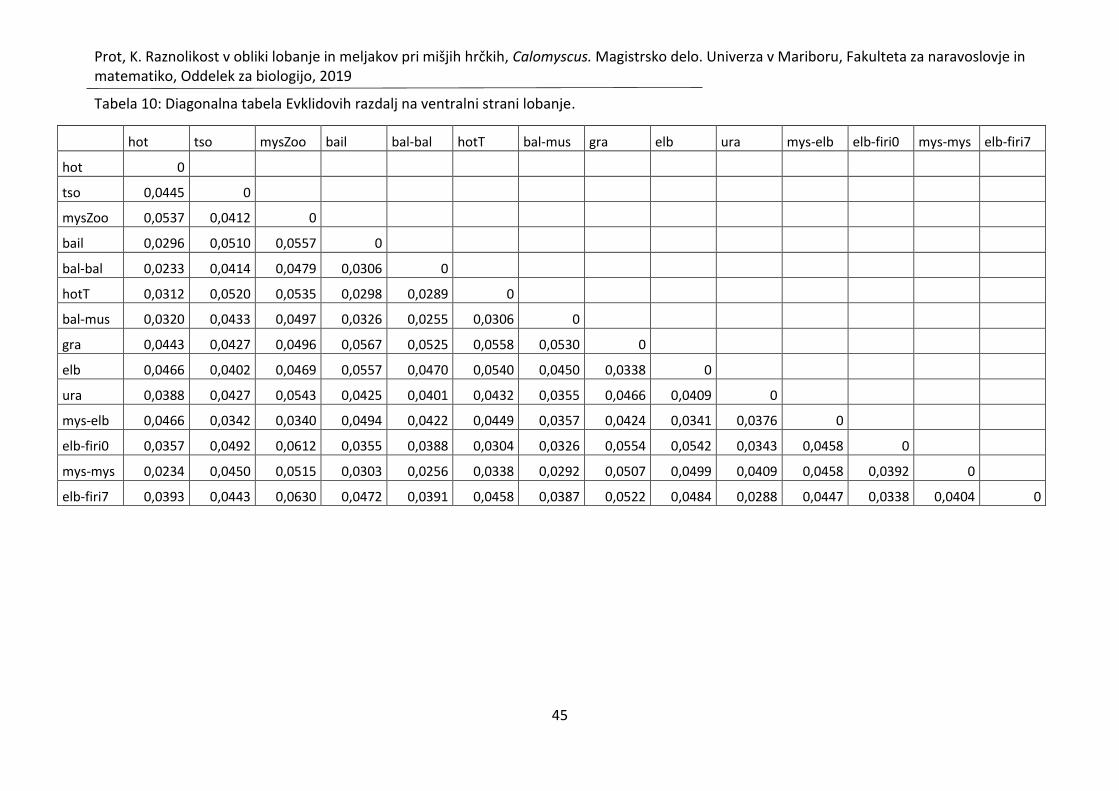
Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
45
Tabela 10: Diagonalna tabela Evklidovih razdalj na ventralni strani lobanje.
hot tso mysZoo bail bal-bal hotT bal-mus gra elb ura mys-elb elb-firi0 mys-mys elb-firi7
hot 0
tso 0,0445 0
mysZoo 0,0537 0,0412 0
bail 0,0296 0,0510 0,0557 0
bal-bal 0,0233 0,0414 0,0479 0,0306 0
hotT 0,0312 0,0520 0,0535 0,0298 0,0289 0
bal-mus 0,0320 0,0433 0,0497 0,0326 0,0255 0,0306 0
gra 0,0443 0,0427 0,0496 0,0567 0,0525 0,0558 0,0530 0
elb 0,0466 0,0402 0,0469 0,0557 0,0470 0,0540 0,0450 0,0338 0
ura 0,0388 0,0427 0,0543 0,0425 0,0401 0,0432 0,0355 0,0466 0,0409 0
mys-elb 0,0466 0,0342 0,0340 0,0494 0,0422 0,0449 0,0357 0,0424 0,0341 0,0376 0
elb-firi0 0,0357 0,0492 0,0612 0,0355 0,0388 0,0304 0,0326 0,0554 0,0542 0,0343 0,0458 0
mys-mys 0,0234 0,0450 0,0515 0,0303 0,0256 0,0338 0,0292 0,0507 0,0499 0,0409 0,0458 0,0392 0
elb-firi7 0,0393 0,0443 0,0630 0,0472 0,0391 0,0458 0,0387 0,0522 0,0484 0,0288 0,0447 0,0338 0,0404 0

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
46
Slika 42: Dendrogram morfometričnih odnosov med skupinami na ventralni strani lobanje.
3.4.3 Morfometrični odnosi med taksoni - dorzalna stran lobanje Morfometrični odnosi na dorzalni strani lobanje so predstavljeni v tabeli 11. Vrste z najdaljšimi
Evklidovimi razdaljami so C. tsolovi (tso) in C. bailwardi (bail) (evklidova razdalja 0,0689), C.
tsolovi (tso) in C. elburzensis firiusaensis (elb-firi0) (razdalja 0,0654) ter C. grandis (gra) in C.
elburzensis firiusaensis (elb-firi7) (razdalja 0,0626). Najmanjše Evklidove razdalje so med
vrstami C. mystax (mysZoo) in C. baluchi (bal-bal) (razdalja 0,0221), C. hotsoni (hot) in C.
hotsoni (hotT) (razdalja 0,0235) ter C. hotsoni (hot) in C. elburzensis firiusaensis (elb-firi7)
(razdalja 0,0251).
Dendrogram morfometričnih odnosov na dorzalni strani lobanje je predstavljen s sliko 43.
Razdalje med osebki so predstavljene z Evklidovimi razdaljami. Na dorzalni strani lobanje se
od ostalih vrst najprej ločijo C. tsolovi (tso), C. grandis (gra) in C. bailwardi (bail). Znotraj ostalih
vrst se oblikujeta dve skupini, in sicer skupina ena vsebuje sestrski skupini C. mystax (mysZoo)
in C. baluchi (bal-bal) ter C. baluchi mustersi (bal-mus) in C. mystax (mys-mys). Znotraj skupine
sta prisotna tudi najbolj podobna osebka, C. mystax (mysZoo) in C. baluchi (bal-bal). Druga
skupina je sestavljena iz sedmih osebkov: C. hotsoni (hotT), C. hotsoni (hot), C. elburzensis
firiusaensis (elb-firi7), C. elburzensis (elb), C. elburzensis firiusaensis (elb-firi0), C. uratensis
(ura) in C. mystax zykovi (mys-elb).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
47
Tabela 11: Diagonalna tabela Evklidovih razdalj na dorzalni strani lobanje.
hot tso mysZoo bail bal-bal hotT bal-must gra elb ura mys-elb elb-firi0 mys-mys elb-firi7
hot 0
tso 0,0523 0
mysZoo 0,0395 0,0393 0
bail 0,0364 0,0689 0,0437 0
bal-bal 0,0336 0,0381 0,0221 0,0423 0
hotT 0,0235 0,0447 0,0400 0,0399 0,0284 0
bal-must 0,0304 0,0588 0,0418 0,0431 0,0365 0,0359 0
gra 0,0451 0,0582 0,0394 0,0449 0,0389 0,0452 0,0481 0
elb 0,0289 0,0475 0,0372 0,0473 0,0417 0,0344 0,0376 0,0455 0
ura 0,0291 0,0538 0,0369 0,0438 0,0386 0,0350 0,0332 0,0460 0,0295 0
mys-elb 0,0351 0,0580 0,0380 0,0538 0,0416 0,0436 0,0353 0,0444 0,0335 0,0323 0
elb-firi0 0,0297 0,0654 0,0499 0,0403 0,0491 0,0367 0,0469 0,0458 0,0290 0,0354 0,0437 0
mys-mys 0,0353 0,0569 0,0353 0,0465 0,0276 0,0392 0,0283 0,0412 0,0436 0,0421 0,0342 0,0486 0
elb-firi7 0,0251 0,0570 0,0446 0,0448 0,0407 0,0342 0,0367 0,0626 0,0378 0,0429 0,0448 0,0420 0,0409 0

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
48
Slika 43: Dendrogram morfometričnih odnosov med skupinami na dorzalni strani lobanje.
3.4.4 Morfometrični odnosi med taksoni - lateralna stran lobanje
Morfometrični odnosi na lateralni strani lobanje so predstavljeni v tabeli 12. Vrste z najdaljšimi
Evklidovimi razdaljami so C. tsolovi (tso) in C. hotsoni (hot) (razdalja 0,0689), C. tsolovi (tso) in
C. mystax (mys-mys) (razdalja 0,0685) ter C. tsolovi (tso) in C. elburzensis firiusaensis (elb-firi7)
(razdalja 0,0679). Vrste z najkrajšimi razdaljami in večjo podobnostjo so C. mystax zykovi (mys-
elb) in C. elburzensis firiusaensis (elb-firi0) (razdalja 0,0251), C. grandis (gra) in C. elburzensis
(elb) (razdalja 0,0260) ter C. hotsoni (hotT) in C. grandis (gra) (razdalja 0,0277).
Dendrogram morfometričnih odnosov na lateralni strani lobanje je prikazan s sliko 44.
Razdalje med osebki so predstavljene z Evklidovimi razdaljami. Osebki formirajo 4 skupine, pri
čemer se najprej odcepi C. tsolovi (tso), osebek, ki se najbolj razlikuje od ostalih skupin. V drugi
skupini sta sestrska osebka C. mystax (mys-mys) in C. elburzensis firiusaensis (elb-firi7) ter C.
hotsoni (hot). V tretji skupini se najprej od ostalih osebkov odcepi C. bailwardi (bail), sledi
odcepitev C. urartensis (ura) ter cepitev dveh sestrskih skupin, C. baluchi (bal-bal) in C. baluchi
mustersi (bal-mus) ter C. mystax zykovi (mys-elb) in C. elburzensis firiusaensis (elb-firi0). Četrto
skupino sestavljajo najprej odcepljena C. mystax (mysZoo) in C. hotsoni (hotT) ter sestrska
skupina C. grandis (gra) in C. elburzensis (elb).

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
49
Tabela 12: Diagonalna tabela Evklidovih razdalj na lateralni strani lobanje.
hot tso mysZoo bail bal-bal hotT bal-mus gra elb ura mys-elb elb-firi0 mys-mys elb-firi7
hot 0
tso 0,0689 0
mysZoo 0,0435 0,0466 0
bail 0,0505 0,0499 0,0402 0
bal-bal 0,0445 0,0582 0,0471 0,0402 0
hotT 0,0413 0,0422 0,0381 0,0431 0,0350 0
bal-mus 0,0442 0,0537 0,0386 0,0453 0,0293 0,0377 0
gra 0,0434 0,0393 0,0291 0,0432 0,0406 0,0277 0,0365 0
elb 0,0439 0,0519 0,0352 0,0492 0,0440 0,0355 0,0402 0,0260 0
ura 0,0446 0,0524 0,0402 0,0459 0,0442 0,0450 0,0380 0,0418 0,0473 0
mys-elb 0,0482 0,0470 0,0347 0,0309 0,0301 0,0359 0,0314 0,0366 0,0428 0,0334 0
elb-firi0 0,0450 0,0494 0,0414 0,0361 0,0325 0,0377 0,0328 0,0393 0,0486 0,0312 0,0251 0
mys-mys 0,0343 0,0685 0,0373 0,0504 0,0448 0,0418 0,0409 0,0446 0,0514 0,0473 0,0422 0,0435 0
elb-firi7 0,0371 0,0679 0,0448 0,0501 0,0326 0,0461 0,0341 0,0485 0,0515 0,0370 0,0378 0,0351 0,0341 0

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
50
Slika 44: Dendrogram morfometričnih odnosov med skupinami na lateralni strani lobanje.
3.4.5 Mantel t test Z Mantel t testom sem želela preveriti domnevo, da lobanjski segmenti izkazujejo podobne
morfološke značilnosti.
Med dorzalno in lateralno stranjo lobanje je stopnja korelacije najvišja, pri čemer je korelacija
zmerna, saj je vrednost R = 0,5235, stopnja značilnosti je signifikantna (p = 0,0345). Ostale
vrednosti korelacij so nizke in ne dosegajo signifikantnosti (tabela 13). Iz tega ugotovimo, da
ni prisotne povezave med posameznimi stranmi lobanje.
Tabela 13: Vrednosti korelacijskih koeficientov iz Mantel t testa.
ventralno dorzalno lateralno
meljaki R = 0,0304 p = 0,3478
R = -0,2185 p = 0,9025
R = -0,0626 p = 0,6025
ventralno - R = 0,2065 p = 0,1151
R = 0,1433 p = 0,1716
dorzalno - - R = 0,5235 p = 0,0345

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
51
4 Razprava Namen naloge je, predstaviti morfometrično pestrost osmih vrst rodu Calomyscus, in sicer C.
bailwardi, C. baluchi, C. elburzensis, C. grandis, C. hotsoni, C. mystax, C. tsolovi in C. urartensis.
Analizirala sem okluzalno površino zgornjih meljakov ter ventralno, dorzalno in lateralno stran
lobanje s Prokrustovo analizo in analizo glavnih komponent, analizo velikosti, analizo oblike
ter analizo morfometričnih odnosov med taksoni.
4.1 Analiza velikosti Analizo velikosti sem na preiskovanih strukturah mišjih hrčkov izvedla s pomočjo vrednosti
centroidnih velikosti. Na podlagi analize zgornjih meljakov ter lobanje vidimo izrazito
odstopanje pri vrsti C. tsolovi, ki je v primerjavi z ostalimi vrstami najmanjša. Manjša dorzalna
in lateralna stran lobanje se izkazuje tudi pri vrsti C. elburzensis firiusaensis. Vrsta C. baluchi
mustersi na vseh straneh lobanje izkazuje večje velikosti glede na centroidno velikost.
Rezultati lobanje delno sovpadajo z ugotovitvami Grm (2018), kjer se je analiza velikosti
opravljala s pomočjo analize variance (ANOVA). Analiza je bila opravljena na ventralni strani
lobanje štirih vrst, in sicer C. baluchi, C. grandis, C. hotsoni ter C. sp., kamor spadajo osebki,
katerih vrsta ni znana. Telesno največje osebke v raziskavi predstavlja vrsta C. baluchi, kar
delno sovpada z ugotovitvijo v nalogi, saj je na ventralni strani lobanje v tej nalogi največji
osebek C. mystax, sledi pa ji vrsta C. baluchi mustersi. C. mystax ni vključen v analizo Grm
(2018). Peshev (1991) na podlagi zunanjih morfometričnih meritev analize velikosti ugotovi,
da so osebki C. baluchi na območju jugozahodnega Pakistana generalno večji od ostalih
zahodnih taksonov, medtem ko so C. hotsoni na splošno manjši, vendar ne tako majhni kot C.
tsolovi. Glede na drugačen tip analize pri Peshev (1991) lahko ugotovimo, da je C. tsolovi tudi
po drugih zunanjih znakih manjši med osebki v rodu Calomyscus.
4.2 Analiza oblike Analizo oblike lobanje in meljakov sem spremljala s pomočjo analize glavnih komponent. Vrste
se glede na prvo in drugo glavno komponento razdelijo v morfoprostor. Neskladja v
razporeditvi sem dodatno preverjala z metodo drevesa najkrajšega razvejanja. Osebki se v
morfoprostoru razporedijo na živali z negativnimi in pozitivnimi vrednostmi glavne
komponente, kar izkazuje razlike v obliki in velikosti posamezne strani lobanje in zgornjih
meljakov. Osebki z negativnimi vrednostmi prve glavne komponente na meljakih so: C.
baluchi, C. bailwardi ter dva predstavnika C. hotsoni in C. mystax. S pozitivnimi vrednostmi PC1
pa so: C. uratensis, C. tsolovi, C. baluchi mustersi C. mystax, C. elburzensis, C. grandis ter dva
predstavnika C. elburzensis firiusaensis. Osebki s pozitivno glavno komponento imajo manjši
M1, večji M2 in M3. Iztek doline nad grbico G se popolnoma približa stičišču M1 in M2. M2 se
na območju izteka doline med grbicama G in I ter H in F razširi v primerjavi z negativnimi
vrednostmi PC1, kjer se meljaki ožijo proti M3.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
52
Na ventralni strani lobanje so osebki s pozitivno PC1: C. urartensis, C. baluchi mustersi, C.
mystax, C. baluchi, C. bailwardi, oba predstavnika C. elburzensis firiusaensis ter C. hotsoni. Z
negativno PC1 so: C. tsolovi, C. elburzensis, C. grandis ter dva predstavnika C. mystax. Pojavijo
se spremembe med obliko in velikostjo posameznih struktur: pri pozitivnih vrednostih se
podaljša rostrum, prednji del največje izbočenosti očesne odprtine je premaknjen distalno
(stran od središča lobanje). Proksimalno (proti središču lobanje) je usmerjen zadnji del očesne
odprtine, zgornji rob slušne votline ter vrh zatilničnega čvršča. Predel ušesa je usmerjen proti
posteriornemu robu lobanje. Grm (2018) je analizo oblike testiral s pomočjo diskriminatne
analize. Analiziral je ventralno stran lobanje treh vrst mišjih hrčkov, C. baluchi, C. grandis ter
C. hotsoni, in osebke, katerih vrsta ni znana, Calomyscus sp. Vsi OTU-ji so izkazovali
signifikantne razlike med vsemi skupinami osebkov, razen C. hotsoni, ki ne izkazuje
signifikantne razlike (p > 0,05). Osebki v negativnem DF1 (C. baluchi) imajo krajši slušni
bobnični mehur, širši prednji rob orbite in daljši gobčni del lobanje. C. grandis ima v pozitivnem
območju DF1 daljši bobnični mehur, ožji prednji rob orbite in krajši gobčni del.
Na dorzalni strani so v negativen morfoprostor uvrščene vrste: C. tsolovi, C. baluchi, C. grandis,
C. hotsoni ter dva predstavnika vrste C. mystax. V pozitivnem morfoprostoru so uvrščene
vrste: C. mystax, C. baluchi mustersi, C. urartensis, C. elburzensis, C. hotsoni, C. bailwardi ter
predstavnika vrste C. elburzensis firiusaensis. Pozitivne vrednosti PC1 imajo distalno podaljšan
rostrum, stik nosnice, čeljustnice in čelnice, stik nosnic je usmerjen posteriorno. Medialni stik
čelnice in temenice je anteriorno usmerjen proti rostrumu. Stik temenice in zatilnice se razširja
dorzalno, medtem ko se rob zatilnice posteriorno izboči. Delna podobnost ugotovitev se
izkazuje v primerjavi z Akbarirad in sod. (2015), ki s kanonično variatno analizo (angl. Canonical
Variate Analysis – CVA) na dorzalni strani lobanje med seboj primerjajo osebke C. mystax, C.
eburzensis, C. hotsoni, C. grandis in osebke Calomyscus sp. Pri tem se glede na CV1 loči C.
grandis na negativno stran osi, saj ima osebek najdaljšo nosnico in medtemenico ter krajšo
temenico. Na drugi osi se ločita C. mystax in C. hotsoni, ki imata krajšo nosnico in temenico v
primerjavi s C. elburzensis. Prav tako Akbarirad in sod. (2016) analizira variabilnost v obliki med
vzorci s kanonično variatno analizo (angl. Canonical Variate Analysis – CVA), pri čemer CV1 os
loči Calomyscus sp. iz Kermanshaha v Kurdistanu in C. grandis od C. bailwardi in C. elburzensis.
Pri pozitivnih vrednostih (C. bailwardi in C. elburzensis) imajo osebki krajšo in ožjo nosnico,
širšo in daljšo temenico in medtemenico. Druga os loči Calomyscus sp. iz Kurdistana, C.
bailwardi skupino B od C. bailwardi skupine A, Calomyscus sp. iz Kermanshah, C. grandis ter
delno C. elburzensis, kjer imajo osebki s pozitivnimi vrednostmi krajšo nosnico, vendar širši
medočesni prostor ter lobanjo.
Na lateralni strani lobanje prihaja do večjih razlik v obliki in velikosti lobanje. Vrste z
negativnimi PC1 vrednostmi so: C. hotsoni, C. mystax, C. baluchi, C. baluchi mustersi ter C.
elburzensis firiusaensis, s pozitivnimi pa: C. bailwardi, C. hotsoni, C. grandis, C. elburzensis, C.
tsolovi ter C. mystax. Anteriorni del lobanje je pri pozitivnih vrednostih PC 1 spuščen in manjši,
medtem ko je posteriorni del večji. Na anteriornem delu lobanje se stik predčeljustnice in
nosnice ter vrh nosnice približata, sprednji rob predčeljustnice medialno je orientiran nižje.
Superiorno razširjeni so mesto največje vbočenosti loka očesne odprtine, prednji rob, skrajno
levi del očesne odprtine – dorzalno ter stik temenice in zatilnice dorzalno. Inferiorno spuščeni
sta osnova zatilnega čvršča dorzalno ter skrajno levi ekstrem bobničnega mehurja. Prednji rob

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
53
očesne odprtine ventralno in najnižja točka prednjega roba zigomatičnega loka sta superiorni,
desni rob stičišča sekalca in zobne jamice je vbočen. Na dorzalni in lateralni strani lobanje je
izrazita morfološka razlika med C. tsolovi in ostalimi predstavniki. Na podlagi analize v obliki in
velikosti lahko potrdim hipotezo, da a priori definirane vrste izkazujejo morfometrično
variabilnost v velikosti in v obliki lobanjskih znakov.
4.3 Morfometrični odnosi med taksoni S pomočjo Evklidovih razdalj sem predstavila morfometrične odnose med OTU, katere sem v
nadaljnji analizi vključila v klastrsko analizo ter odnose med OTU-ji predstavila v obliki
dendrograma. Na meljakih do večjih razlik med OTU-ji prihaja med vrstama C. hotsoni ter C.
urartensis, največja podobnost je med vrstama C. mystax elburnensis in C. mystax. Glede na
Evklidove razdalje najbolj odstopa C. hotsoni, saj se odcepi od štirih skupin, v katere se
oblikujejo ostali osebki. V prvi skupini so sestrski vrsti C. baluchi mustersi in C. elburzensis
firiusaensis ter C. tsolovi, v drugi skupini sta dve sestrski veji C. grandis in C. urartensis ter C.
elburzensis in C. elburzensis firiusaensis. V tretji skupini so vsi predstavniki C. mystax, v četrti
pa sestrski vrsti C. bailwardi in C. hotsoni ter odcepljena vrsta C. baluchi.
Na ventralni strani lobanje se na podlagi Evklidovih razdalj največja razlika med OTU-ji pojavi
med osebkoma C. mystax ter C. elburzensis firiusaensis, najmanjša pa med C. hotsoni in C.
baluchi. Z analizo Evklidovih razdalj se OTU-ji oblikujejo 3 skupine: v prvo skupino so uvrščeni
C. urartensis in oba osebka C. elburzensis firiusaensis, v drugi skupini so osebki C. baluchi, C.
hotsoni, C. mystax, C. baluchi mustersi, C. bailwardi ter C. hotsoni. V zadnji skupini so C.
grandis, C. elburzensis, C. mystax, C. mystax elburnensis in C. tsolovi.
Evklidove razdalje na dorzalni strani lobanje so največje med C. tsolovi in C. bailwardi, medtem
ko so najmanjše med C. mystax in C. baluchi. Morfometrični odnosi med OTU-ji na dorzalni
strani oblikujejo 3 skupine, in sicer se v prvi skupini osebki najprej ločijo glede na Evklidove
razdalje (C. tsolovi, C. grandis in C. bailwardi). Druga skupina sestoji iz: C. mystax elburnensis,
C. urartensis, oba osebka C. elburzensis firiusaensis, C. elburzensis in oba osebka C. hotsoni. V
zadnji skupini pa so C. mystax, C. baluchi mustersi, C. baluchi in C. mystax.
Evklidove razdalje na lateralni strani lobanje so najdaljše med C. tsolovi in C. hotsoni, najkrajše
med C. mystax elburnensis in C. elburzensis firiusaensis. Na lateralni strani lobanje se pojavijo
štiri skupine osebkov, pri čemer se kot prva skupina loči C. tsolovi. V drugi skupini so osebki C.
mystax C. elburzensis firiusaensis ter C. hotsoni, v tretji C. bailwardi, C. urartensis, C. baluchi,
C. baluchi mustersi, C. mystax elburnensis in C. elburzensis firiusaensis. V zadnji skupini so C.
mystax, C. hotsoni, C. grandis in C. elburzensis.
Z Mantel t testom sem ugotavljala morfološke podobnosti med lobanjskimi segmenti.
Korelacija je najvišja med lateralno in ventralno stranjo lobanje (R = 0,5235, p = 0,0345).
Nobeden izmed ostalih analiziranih lobanjskih segmentov ne izkazuje signifikantnosti, iz česar
lahko ugotovimo, da so se tekom razvoja strani lobanje razvijale neodvisno druga od druge.
Posamezni lobanjski segmenti izkazujejo različne morfometrične odnose med a priori
definiranimi vrstami, s čimer lahko tudi to hipotezo potrdim. Glede na Evklidove razdalje in
Mantel t test lahko opazimo, da posamezni lobanjski segmenti na posamezni strani lobanje
ter na meljakih izkazujejo različne povezave med posameznimi osebki.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
54
5 Zahvala
Iskreno se zahvaljujem mentorju, prof. dr. Francu Janžekoviču, za vso izkazano strokovno
pomoč, potrpljenje in dostopnost tekom nastajanja strokovnega dela.
Zahvaljujem se somentorju, prof. dr. Borisu Kryštufku, za material in ugotovitve v zvezi z
raziskovanjem mišjih hrčkov.
Zahvaljujem se družini, ki mi je omogočila študij ter me tekom le-tega tudi spodbujala in
podpirala.
Zahvala tudi Leonu in Evi, ki sta med študijem verjela vame, me spodbujala in mi stala ob
strani.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
55
Literatura
Akbarirad, S., Darvish, J., Aliabadian, M., Kilpatrick, C.W. M (2015). Biosystematic study of
Calomyscus mystax (Rodentia, Calomyscidae) from northeastern Iran. Iranian Journal of
Animal Biosystematics (IJAB). Vol.11, No.1, 65–77
Akbarirad, S., Darvish, J. in Aliabadian, M. (2015). Phylogeography of Calomyscus elburzensis
(Calomyscidae, Rodentia) around the Central Iranian Desert with Description of a New
Subspecies in Center of Iranian Plateau. Journal of Sciences, Islamic Republic of Iran 27(1):
5–21 (2016)
Akbarirad, S., Darvish, J. in Aliabadian, M. (2016). Increased species diversity of brush-tailed
mice, genus Calomyscus (Calomyscidae, Rodentia), in the Zagros Mountains, western
Iran. Mammalia, 80(5), 549–562.
Algorithms, (2018). Minimum Spanning Trees. Pridobljeno 15. 3. 2019, s strani
https://algs4.cs.princeton.edu/43mst/
Baloh, M. (2010). Realizacija porazdeljenega algoritma za izračun drevesa najkrajših poti
(Diplomsko delo). Ljubljana: Univerza v Ljubljani Fakulteta za računalništvo in informatiko.
Bastič, M., (2006). Metode raziskovanja. Maribor: Univerza v Mariboru Ekonomsko - poslovna
fakulteta Maribor.
Darvish, J., Mohammadi, Z., Mahmoudi, A., in Siahsarvie, R. (2015). Faunistic and taxonomic
study of Rodents from northwestern Iran. Iranian Journal of Animal Biosystematics, 10(2),
119–-136.
Goodness, T., (2017). Calomyscus bailwardi Mouse-like hamster (Animal Diversity Web–
ADW). Pridobljeno 7. 7. 2018, s
http://animaldiversity.org/accounts/Calomyscus_bailwardi/
Grm, B. (2018). Variabilnost lobanje mišjih hrčkov (rod Calomyscus): pristop z geometrijsko
morfometrijo (Magistrska naloga). Maribor: Univerza v Mariboru, Fakulteta za
naravoslovje in matematiko. Oddelek za biologijo.
Hafner, M. (2013). Vpliv finančnega sistema na učinke gospodarske krize (Magistrska naloga).
Maribor: Univerza v Mariboru, Ekonomsko-poslovna Fakulteta.
Hamidi, K., Darvish, J., & Matin, M. M. (2017). Ecological survey of two Calomyscidae species;
Goodwin's brush-tailed mouse and Hotson's brush-tailed mouse (Rodentia) in the eastern
parts of Iran. Acta Ecologica Sinica, 37(2), 105–114.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
56
Janžekovič, F., Klenovšek, T., Kryštufek, B. (2007). Uporaba metod geometrijske morfometrije
in multivariatne statistike v bioloških raziskavah filogenije in ontogenije. V ROSUS 2007,
Računalniška obdelava slik in njena uporaba v Sloveniji 2007: zbornik 2. Strokovne
konference, str 135–143. Maribor Fakulteta za elektroniko, računalništvo in informatiko,
Inštitut za računalništvo.
Klenovšek, T., (2014). Priročnik za uporabo geometrijske morfometrije v biologiji. Maribor:
Univerza v Mariboru, Fakulteta za naravoslovje in matematiko.
Klenovšek, T., (2014). Variabilnost lobanje dinarske voluharice Dinaromys bogdanovi
(Rodentia: Cricetidae): spolni dimorfizem, alometrija in filogenija. Doktorska disertacija,
Maribor: Univerza v Mariboru, Fakulteta za naravoslovje in matematiko.
Kryštufek, B. in Vohralik, V. (2009). Mammals of Turkey and Cyprus Rodentia II: Cricetinae,
Muridae, Spalacidae, Calomyscidae, Capromyidae, Hystricidae, Castoridae. Koper:
Univerza na Primorskem, Znanstveno-raziskovalno središče Koper, Zgodovinsko društvo
za južno Primorsko.
Legendre, P., Fortin, M. J., Borcard, D. (2015). Should the Mantel test be used in spatial
analysis?. Methods in Ecology and Evolution, 6(11), 1239–1247.
Mitteroecker, P. in Gunz, P. (2009). Advances in geometric morphometrics. Evolutionary
Biology, 36(2), 235–-247.
Norris, R. W., Woods, C. A., Kilpatrick, C. W. (2008). Morphological and molecular definition of
Calomyscus hotsoni (Rodentia: Muroidea: Calomyscidae). Journal of Mammalogy, 89(2),
306–315.
Peshev, D. (1991). On the systematic position of the mouse-like hamster Calomyscus bailwardi
Thomas, 1905 (Cricetidae, Rodentia) from the Near East and Middle Asia. Mammalia
55:107–112.
Pfajfar, L. (2011). Osnovna statistika za ekonomske in poslovne vede. Ljubljana: Univerza v
Ljubljani, Ekonomska fakulteta.
Shad, H. H., in Darvish, J. (2018). Rodents Diversity in Protected Areas of Yazd Province, Central
of Iran. Journal of Research in Biology, 1(1).
Shahabi, S., Aliabadian, M., Darvish, J., & Kilpatrick, C. W. (2013). Molecular phylogeny of
brush-tailed mice of the genus Calomyscus (Rodentia: Calomyscidae) inferred from
mitochondrial DNA sequences (Cox1 gene). Mammalia, 77(4), 425–431.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
57
Shahabi, S., Darvish, J., Aliabadian, M., Mirshamsi, O., in Mohammadi, Z. (2012). Cranial and
dental analysis of mouse-like hamsters of the genus Calomyscus (Rodentia:
Calomyscidae) from plateau of Iran. Hystrix, the Italian Journal of Mammalogy, 22(2).
Shahabi, S., Zarei, B. in Sahebjam, B. (2010). Karyologic study of three species of Calomyscus
(Rodentia: Calomyscidae) from Iran. Iranian Journal of Animal Biosystematics (IJAB),
Vol.6, No.2, 55–60, 2010.
Sherratt, E., Gower, J. G., Klingenberg, C. P. in Wilkinson, M. (2014). Evolution of Cranial Shape
in Caecilians (Amphibia: Gymnophiona). New York: Springer Science+Business Media New
York. Evol Biol (2014) 41:528–545
Slice, D. E. (2007). Geometric morphometrics. Annual Review of Anthropology, 36.
Wilson, D. E., Lacher, T. E., Mittermeier, R. A. (2017). Handbook of the Mammals of the World.
Volume 7: Rodents II. Lynx Ediciones, Barcelona.
Wilson, D. E., in Reeder, D. M. (Eds.). (2005). Mammal species of the world: a taxonomic and
geographic reference (Vol. 1). JHU Press.
Zarei, B., Darvish, J., Aliabadian, M., & Moghaddam, F. Y. (2013). Geometric morphometric
analyses of four species of brush-tailed mice, genus Calomyscus (Rodentia:
Calomyscidae), from the Iranian Plateau. Iranian Journal of Animal Biosystematics, 9(1).
Zelditch, M. L., Swiderski, D. L., Sheets, H. D. (2004). Geometric morphometrics for biologists:
A primer. Academic Press.

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
58
6 Priloga Priloga 1: Seznam proučevanih lobanj osebkov mišjih hrčkov (Calomyscus). Zbirka: SMF – Naravoslovni muzej Senckenberg , ZFMK – Zoološki
raziskovalni muzej Aleksandra Koenig , ZIN – Zoološki inštitut Ruske akademije znanosti, Sankt Peterburg, BMNH – Prirodoslovni muzej London.
inv. št. zbirka OTU Vrsta Lokaliteta Država Spol
19-11-7-64 BMNH hot C. hotsoni
83143 SMF tso C.tsolovi Thafas Syria f
87383 SMF mysZoo C.mystax Zoo Berlin m
Type_bailwardi bail C. bailwardi
Type_baluchi bal-bal C. baluchi
Type_hotsoni hotT C. hotsoni
Type_mustersi
bal-mus C. baluchi mustersi
89140 ZFMK gra C. grandis Prov. Tehran, Khojir in Sorkheh Hessar Iran
2007. 100 ZFMK elb C. elburzensis Prov. Herat, 30 km od Herat do Delaram Afganistan f
72755 ZIN ura C. urartensis Nahičevan Aterbaijan m
82802 ZIN mys-elb
C. mystax ssp. zykovi Ujetništvo Turkmenija
82850 ZIN elb-firi0
C. elburzensis firiusaensis ca. Ašhabad Turkmenistan f
82853 ZIN mys-mys C. mystax Bolšoj Balhan Turkmenistan f
87567 ZIN elb-firi7
C. elburzensis firiusaensis ca. Ašhabad Turkmenistan f

Prot, K. Raznolikost v obliki lobanje in meljakov pri mišjih hrčkih, Calomyscus. Magistrsko delo. Univerza v Mariboru, Fakulteta za naravoslovje in matematiko, Oddelek za biologijo, 2019
59





























