Embed Size (px)
Citation preview

Učinak genotoksičnih tvari na DNA Nastanak mutacija

GENOTOKSIČNI UČINAK NA NUKLEINSKE KISELINE
• Mutacija - naslijedna promjena vezana za genotip stanice
• Pojava mutacije podrazumijeva da je dvostruka uzvojnica DNA
u većoj ili manjoj mjeri promjenjena.
• S obzirom na te promjene, neka tvar može izazvati mutacije u
užem smislu, aneuploidiju i klastogene promjene.
• Aneuploidija podrazumjeva gubitak ili povećanje jednog
kromosoma ili 2n±1 uslijed npr. nefunkcioniranja diobenog
vretena prilikom mejotičke ili mitotičke diobe, promjenejne
strukture kinetohora i tubulina, centromera...
• Klastogeneza podrazumijeva gubitak, dodatak ili rearanžman
dijelova kromosoma.

• Mutacije mogu biti:
– spontane (tautomerne promjene baza)
– inducirane
o 1 od 109 nukleotida mutira prilikom svake mitoze
o ljudska DNA i DNA drugih sisavaca građena je od 109 nukleotida
o jedna od 109 stanica će sadržavati mutaciju
o ljudski se organizam sastoji od 1014 stanica, iz čega proizlazi da jedan čovjek nosi 105 potencijalnih stanica raka
o prema nekim istraživanjima u stanici je potrebno prisustvo mutiranih 3-20 gena uključenih u regulaciju staničnog ciklusa kako bi došlo do nastanka raka

Od mutacije do nastanka raka


Fenotipske promjene vidljive na tumorskim stanicama pokazuju da je potrebna akumulacija mnogo mutacija da bi normalna stanica postala tumorska Treba doći do mutacija u više stotina gena da bi došlo do nastanka raka Utvrđeno je da su u stanicama raka uvijek prisutne mutacije u genima koji kontroliraju stanični ciklus Stanice, ovisno o signalima iz okoline, ulaze ili ne ulaze u diobu. Signale primaju od susjednih stanica, pozitivne koji stimuliraju diobu, ili negativne koji pak sprečavaju proliferaciju Tumorske stanice same stvaraju stimulirajuće signale u procesu zvanom autokrina stimulacija ili su neosjetljive na negativne signale Kod normalnih stanica međusobnim kontaktom dolazi do inhibicije diobe stanica Tumorske stanice su izgubile svojstvo kontaktne inhibicije te rastu u više slojeva Kada nema faktora rasta ili kada su izložene toksinima ili zračenju, normalne stanice umiru Ta programirana smrt stanice, apoptoza, sprečava fiksiranje mutacija tj. proliferaciju mutiranih stanica Karakteristika tumorskih stanica je gubitak kontrolirane smrti stanice, tj. apoptoze Još jedna karakteristika tumorskih stanica je gubitak veza i komunikacija između susjednih stanica koje u normalnim stanicama služe za prijenos malih molekula važnih u kontroli staničnog rasta
Od mutacije do nastanka tumora/raka

Tumorske stanice su genetički nestabilne, tj. smanjena im je vjernost replikacije DNA i efikasnost popravka oštećenja te povećana učestalost kromosomskih aberacija i aneuploidija (promjene strukture ili broja kromosoma) Broj dioba nije ograničen kao kod normalnih stanica gdje nakon određenog broja dioba dolazi do smrti Većina ljudskih somatskih stanica ne eksprimira telomerazu. Taj je enzim odgovoran za produživanje telomera na krajevima kromosoma i eksprimira se u embriju te u stanicama raka Nije aktivan u normalnim stanicama, pa se telomera skrati do određene granice i dolazi do smrti stanice. U stanicama raka se telomeraza eksprimira, te zbog toga dolazi do nekontrolirane diobe Tumorske stanice karakterizira mogućnost stvaranja metastaza, tj. invazije tumorskih stanica u druga tkiva. Angiogeneza, tj. induciranje rasta krvnih žila još je jedna karakteristika stanica raka koja im omogućava rast i metastaziranje. To se dogodi kada se takve stanice odcijepe od primarne tumorske mase, uđu u krvotok te na taj način dođu do novog staništa unutar organizma

• Osim mutacija uzrokovanih kemijskim, fizikalnim i biološkim agensima, nastanak raka vezan je i za nestabilnost kromosoma te izostanak mehanizama popravaka
• Važna uloga onkogena i tumor supresor gena
• mutacijama protoonkogena ili tumor supresor gena može doći do nekontrolirane diobe stanica

Odnos strukture i aktivnosti kemijskih
genotoksičnih agenasa
• potencijalna elektrofilna mjesta u strukturi kemijskog spoja
koja mogu reagirati s nukleofilnim staničnim
makromolekulama
• alifatski i aromatski spojevi s nitro grupama, aromatski
spojevi s azo grupama, N-oksidi aromatskih spojeva, alkil
hidrazini, N-metiol derivati, alkil aldehidi, monohaloalkeni,
karbamati, derivati dušićnog i sumpornog iperita, N-
kloroamini, propiolaktoni, alkilhalidi aromatskih i alifatskih
spojeva, nitrozoamini, aromatski amini, oksidi i epoksidi te
svi spojevi koji nastaju kao metaboliti i imaju ovakvu
strukturu

Ciljna mjesta:

Kemijski spojevi kao mutageni
• alkilirajući agensi
• analozi dušićnih baza
• tvari koje s DNA tvore adukte
• spojevi koji izazivaju deaminaciju citozina, adenina i gvanina-deaminacija
• spojevi koji se ugrađuju između dvije baze u DNA-interkalacija
• mutageno djelovanje nekog spoja manifestira se na stanicama kćerima nastalim mitozom (mejozom)

Alkilirajući agensi
• Monoalkilirajući agensi-najčešće reagiraju s gvaninom (7N pozicija) ali i drugim pozicijama dušićnih baza
– Taj događaj ne sprečava odvajanje lanaca DNA iz strukture duple uzvojnice
– inhibicija enzima uključenih u procesiranje DNA
– neispravno odmatanje i smatanje DNA i procesiranje gena
– inhibicija staničnog rasta i apoptoza (preko p53 puta)
– nakon replikacije DNA
• u nekim stanicama indukcija mutacija i kancerogenih događaja
• dialkilirajući agensi reagiraju s dva različita 7-gvanin ostatka
• ako se taj proces događa u dva različita lanca dolazi do njihovog unakrsnog povezivanja-nemogućnost razmatanja dvostruke uzvojnice



Alkilirajući agensi
• citostatici
• spojevi koji se alkilnom (CnH2n+1) grupom vežu za DNA
• Stanice raka proliferiraju brže od normalnih stanica i sa manje kontrolnih mehanizama popravaka
• više su osjetljive na DNA oštećenja
• citotoksični i za normalne stanice (prvenstveno one koje se često dijele; GI sustav, koštana srž, testisi, ovariji)
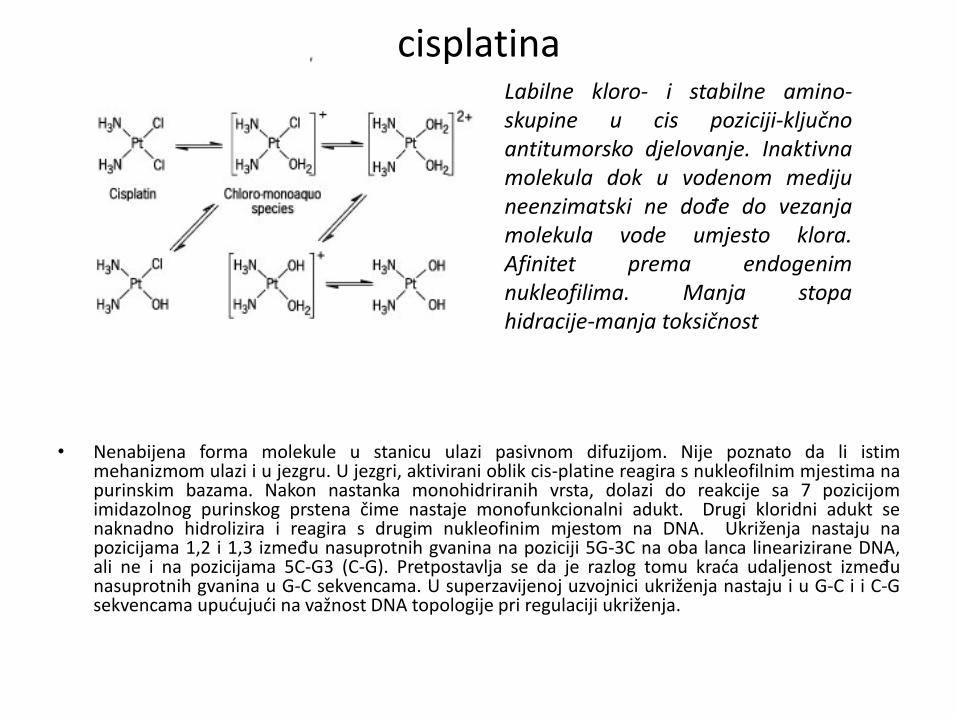
cisplatina
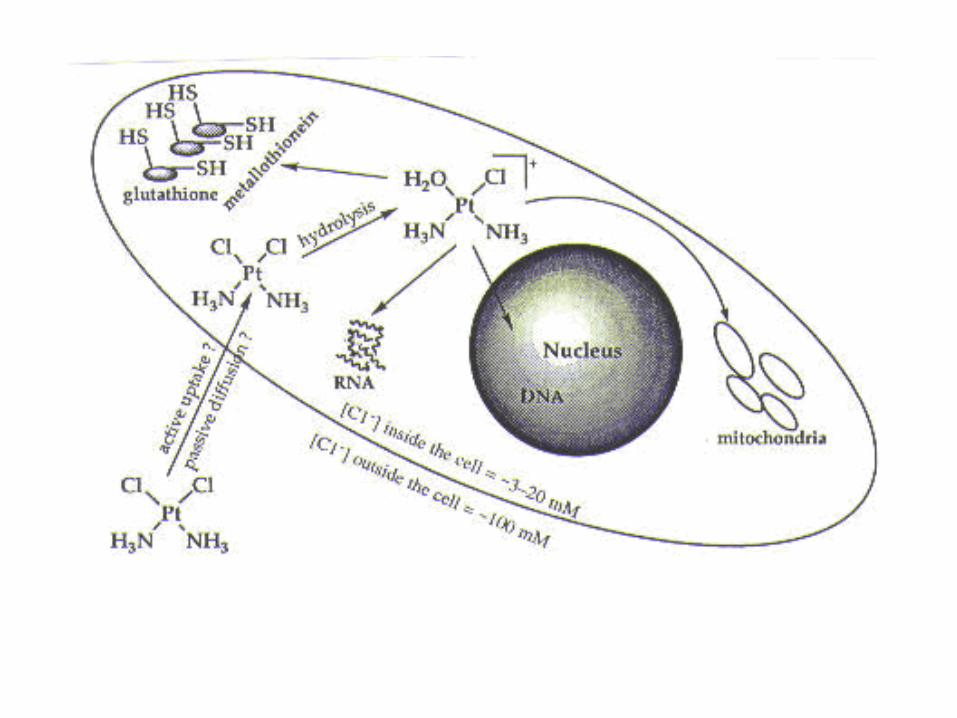
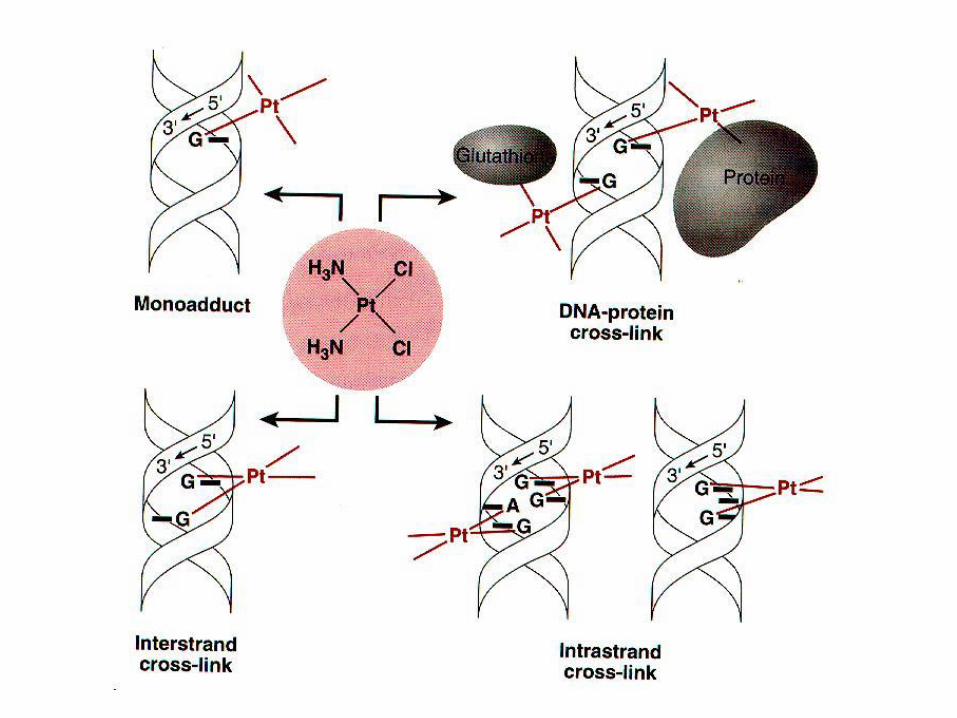
• Nenabijena forma molekule u stanicu ulazi pasivnom difuzijom. Nije poznato da li istim mehanizmom ulazi i u jezgru. U jezgri, aktivirani oblik cis-platine reagira s nukleofilnim mjestima na purinskim bazama. Nakon nastanka monohidriranih vrsta, dolazi do reakcije sa 7 pozicijom imidazolnog purinskog prstena čime nastaje monofunkcionalni adukt. Drugi kloridni adukt se naknadno hidrolizira i reagira s drugim nukleofinim mjestom na DNA. Ukriženja nastaju na pozicijama 1,2 i 1,3 između nasuprotnih gvanina na poziciji 5G-3C na oba lanca linearizirane DNA, ali ne i na pozicijama 5C-G3 (C-G). Pretpostavlja se da je razlog tomu kraća udaljenost između nasuprotnih gvanina u G-C sekvencama. U superzavijenoj uzvojnici ukriženja nastaju i u G-C i i C-G sekvencama upućujući na važnost DNA topologije pri regulaciji ukriženja.
Labilne kloro- i stabilne amino- skupine u cis poziciji-ključno antitumorsko djelovanje. Inaktivna molekula dok u vodenom mediju neenzimatski ne dođe do vezanja molekula vode umjesto klora. Afinitet prema endogenim nukleofilima. Manja stopa hidracije-manja toksičnost


Analozi dušićnih baza
• spojevi koji po svojem kemijskom karakteru mogu biti zamjenjeni s normalnom dušićnom bazom
• 5-bromouracil (5BU)
• afinitet prema adeninu
• spontanom promjenom u drugu izomernu formu-afinitet prema gvaninu
• tijekom replikacije, gvanin se insertira nasuprot BU
• u slijedećoj replikaciji gvanin će se spariti s citozinom (tranzicijska mutacija supstitucije parova baza)

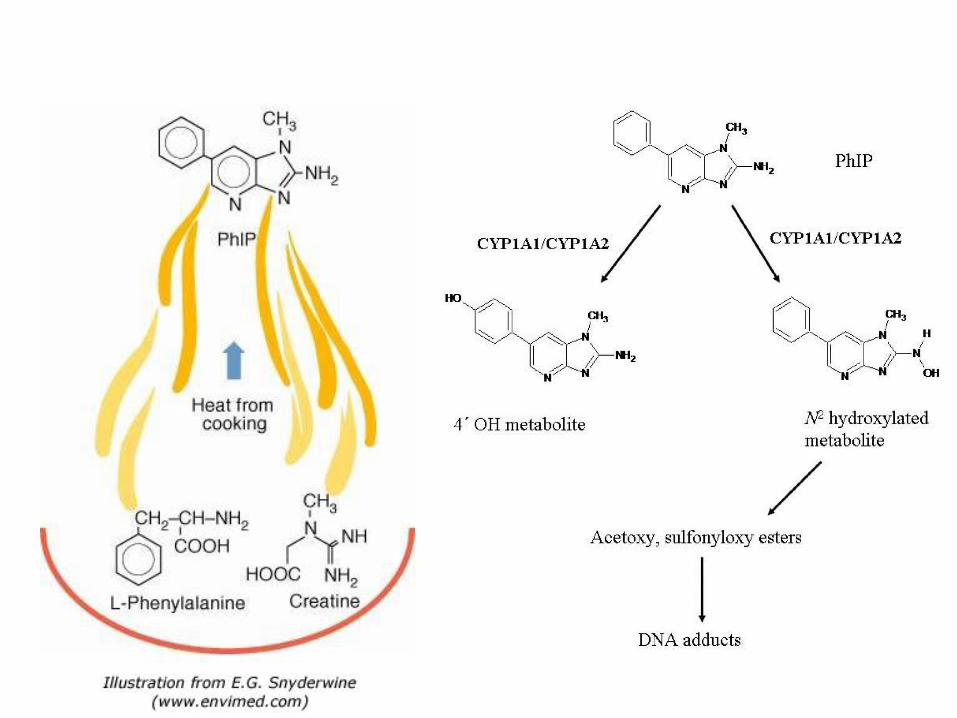
DNA adukti
• dio DNA kovalentno vezan za neki spoj
• obično inicijacija kancerogeneze
• acetaldehid, ciklički nitrozoamini, nitrozoamnini iz cigaretnog dima, benzaldehid, pirolitički produkti nastali prženjem hrane...
• mutacije koje nastaju nakon S faze
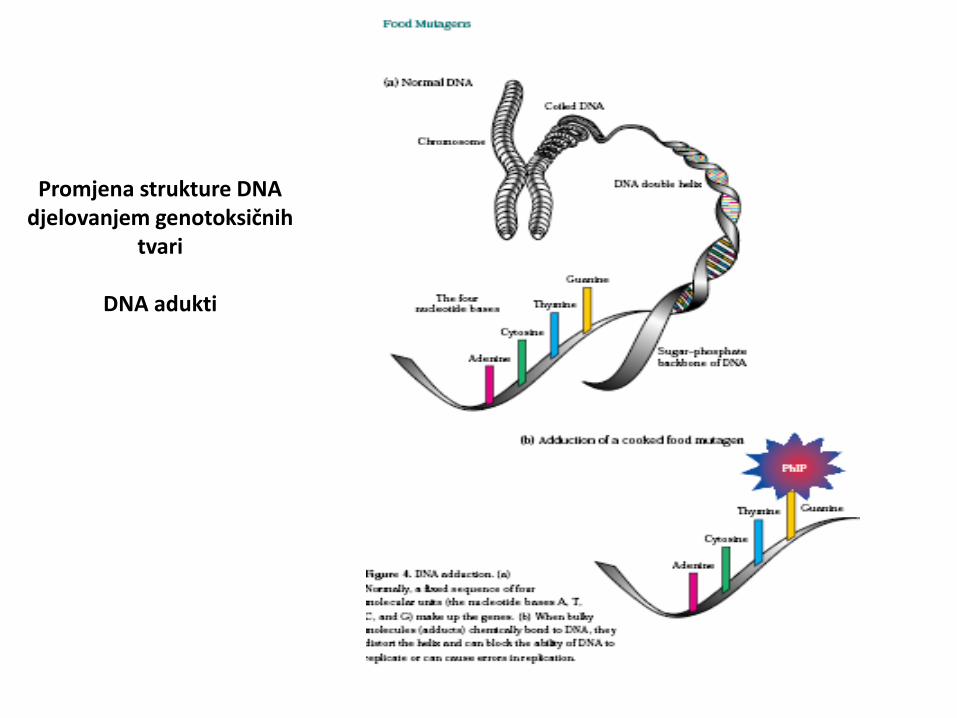
Promjena strukture DNA djelovanjem genotoksičnih
tvari
DNA adukti


Promjena strukture DNA
djelovanjem
genotoksičnih tvari
DNA adukti

• Osnovni uzrok izrazite genotoksičnosti je nastanak epoksidnih formi u fazi I
• Benz(a)piren u fazi I daje 7,8-dihidrodiol 9,10 –epoksid koji se veže za DNA.
• Diol epoksidni oblici odgovorni za izrazitu genotoksičnost PAH-ova

Deaminacija
• mutacije izazvane oksidativnom deaminacijom dušićnih baza • amino skupina dušićne baze deaminira se i zamjenjuje s
hidroksilnom skupinom • adenin kojemu se -NH2 skupina nalazi na položaju 6 deaminira se
nitritnom kiselinom i nastaje hipoksantin adenin • hipoksantin se transformira u ketotautomer koji ima afinitet
prema citozinu • rezultat pri slijedecoj replikaciji: supstitucija parova baza A : T u G:
C • Deaminacijom citozina nastaje uracil koji ima afinitet prema
adeninu-supstitucija parova baza C: G u T: A • deamniacijom gvanina nastajke ksantin koji se sa dvije vodikove
veze povezuje s citozinom • nitritna kiselina , NO, N-nitrozoindol


Deaminacija

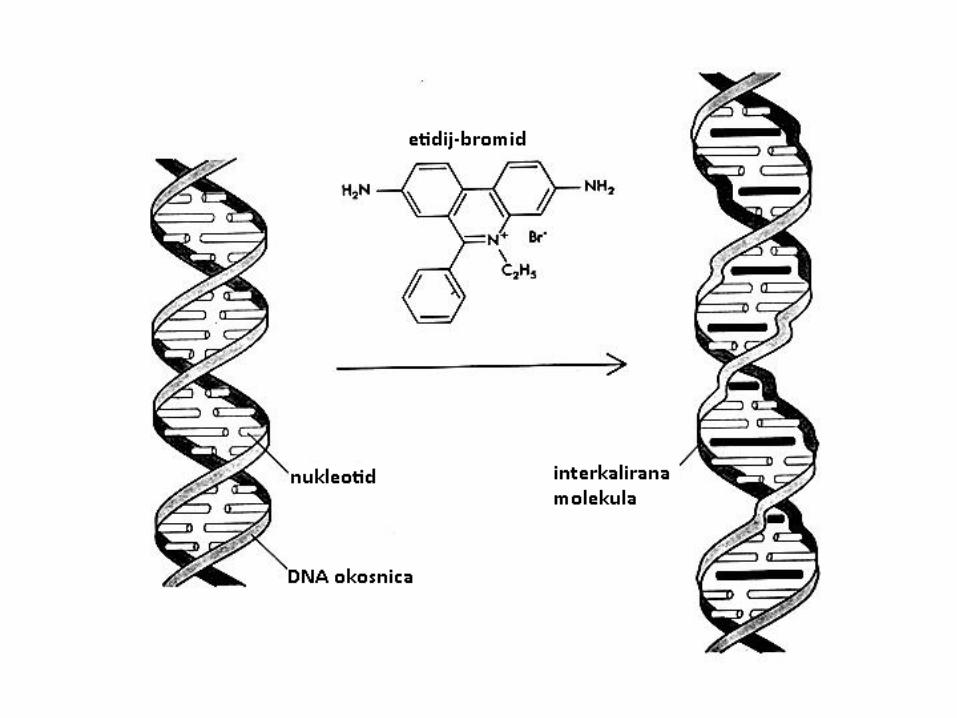
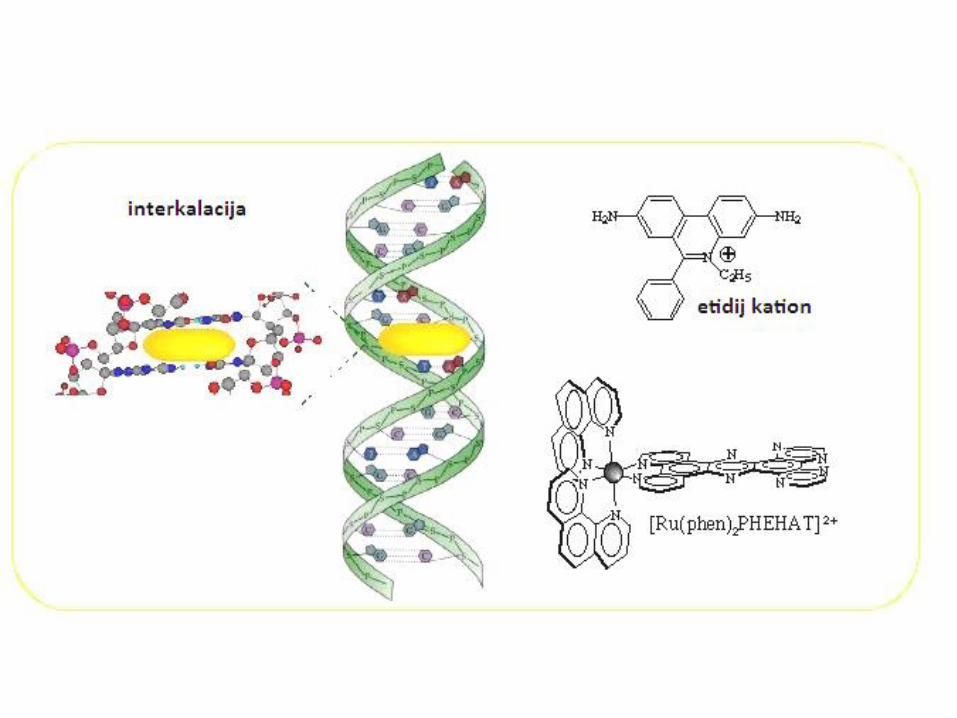
Interkalacija
• Interkalacija se javlja su slučaju kad kemikalija ima određenu strukturu i kemijsku prirodu da se može ugraditi između dviju dušićnih baza u DNA
• to su obično policikličke, aromatske i planarne strukture (berberin, etidij-bromid, proflavin, daunomicin, doksorubicin, talidomid, aflatoksin)
• da bi došlo do interkalacije, u DNA mora nastati otvor između baza-razmotavanjem
• stupanj razmatanja ovisi o interkalatoru; etidij kation (ionska forma etidij-bromida) razmata DNA pod kutem 26°, a proflavin pod kutem 17°.
• razmatanje uzrokuje razdvajanje baza-lokalne strukturne promjene DNA, izvijanje dušićnih baza
• promjena funkcije DNA, inhibicija transkripcije i replikacije te procesa popravka DNA
• mehanizam: u vodenoj otopini, kationski interkalator ima afinitet zbog elektrostatksih sila prema polianionskoj molekuli DNA. Interkalator uzrokuje izmještanje natrijevih i/ili magnezijevih iona koji uvijek okružuju DNA (zbog neutralizacije naboja) pri čemu se stvara slaba elektrostatska sila između liganda i vanjske strane DNA. Ligand ulazi u hidrofobnu unutrašnjost molekule DNA između dušićnih baza. Između baza se stvara otvor pomoću energije apsorbirane tijekom kolizije s molekulama otapala.

Točkaste mutacije
• -supstitucija parova baza
• -frameshift mutacije ili mutacije pomaka okvira čitanja
• SUPSTITUCIJA PAROVA BAZA
– -kemijska modifikacija dušićne baze. Ta se promjena može dogoditi u bilo kojoj fazi staničnog ciklusa
– inkorporacija kemijskog spoja u molekulu DNA prilikom procesa replikacije zbog sličnosti te molekule (kemijske i afiniteta enzima koji sudjeluju u replikaciji prema toj molekuli) u jedan lanac molekule DNA umjesto odgovarajuće dušićne baze

deaminacija uzrokovana nekim kemijskim sredstvom
H-hipoksantin, strukturno nalikuje gvaninu, pa afinitet prema hipoksantinu ima citozin
C≡G C≡G
A=T H=T G≡C G≡C
prva replikacija C≡G G≡C H≡C T=A G≡C C≡G PRVA STANICA KĆER DRUGA STANICA KĆER izmjenjena molekula DNA zbog neizmjenjena molekula DNA promjenjene dušićne baze (H strukturno sličan G, a ne A)
druga replikacija FIKSIRANA MUTACIJA C≡G G≡C H≡C C≡G G≡C C≡G

C≡G A=T G≡C
prva replikacija C≡G G≡C A=Bu T=A G≡Bu C≡G
druga replikacija C≡G G≡C A=T Bu=A G≡C Bu=A
treća replikacija C≡G G≡C G≡C C≡G A=T T=A T=A A=T G=C C≡G T=A A=T
Bu

Točkaste mutacije-supstitucija parova baza
• Posljedice supstitucije parova baza: – “samesense” mutacije – zbog degeneracije koda,
različiti tripleti baza kodiraju za istu aminokiselinu
– “missense” mutacije – novonastali triplet baza kodira za drugačiju aminokiselinu
– “nonsense” mutacije – mutacijom supstitucijom parova baza nastaje stop kodon (umjesto CGA za citozin)
(DNA: 5' - ATG ACT CAC TGA GCG CGA AGC TGA - 3'
3' - TAC TGA GTG ACT CGC GCT TCG ACT - 5'
mRNA: 5' - AUG ACU CAC UGA GCG CGU AGC UGA - 3'
Protein: Met Thr His Stop)

Mutacije pomaka okvira čitanja (“frameshift”) insercija ili delecija jedne ili više baza u DNA. Uslijed toga, genetički kod više nije isti
jer se pomiče okvir čitanja.
• HOW CAN THE RAT EAT (zamislite da je ovo 5 genetičkih kodova). Ako insertirate ili deletirate jednu dušićnu bazu;
• HOW CAN THE RAT EAT
• HOW CAN THR ATE AT..
• Od mjesta insercije ili delecije, kod više nema smisla. Transkripcijska informacija je potpuno izmjenjena, i jasno je da u proteinu velike molekulske mase moraju uslijed toga biti ispoljene promjene.
•
• TGT TTC TCA GGT AGT
treonin lizin serin prolin serin
• TGT TCT CAG GTA GT..
treonin lizin valin histidin
interkalacija planarnih molekula u strukturu DNA (npr. akridin)

Primjer 1: Srpasta anemija supstitucija parova baza
Mutacija u genu za β-hemoglobin, (GAG→GTG), promjena valina u glutaminsku kiselinu

Aneuploidija

Aneuploidija (2n±1) • neravnomjerna distribucija kromosoma prilikom mejoze
• neravnomjerna distribucija kromosoma prilikom mitoze
• posljedice različite: prilikom mejoze, sve stanice nastale iz aneuploidične spolne stanice sparene s drugom spolnom stanicom su aneuploidične
• prilikom mitoze, aneuploidične su stanice nastale mitozom iz jedne aneuploidične somatske stanice
• uzroci: inhibicija niti diobenog vretena, inihibicija proteina na centromeri, nepovezivanje centromera s nitima diobenog vretena


Aneuploidija u spolnim stanicama

Mejotička dioba-nastanak spolnih stanica
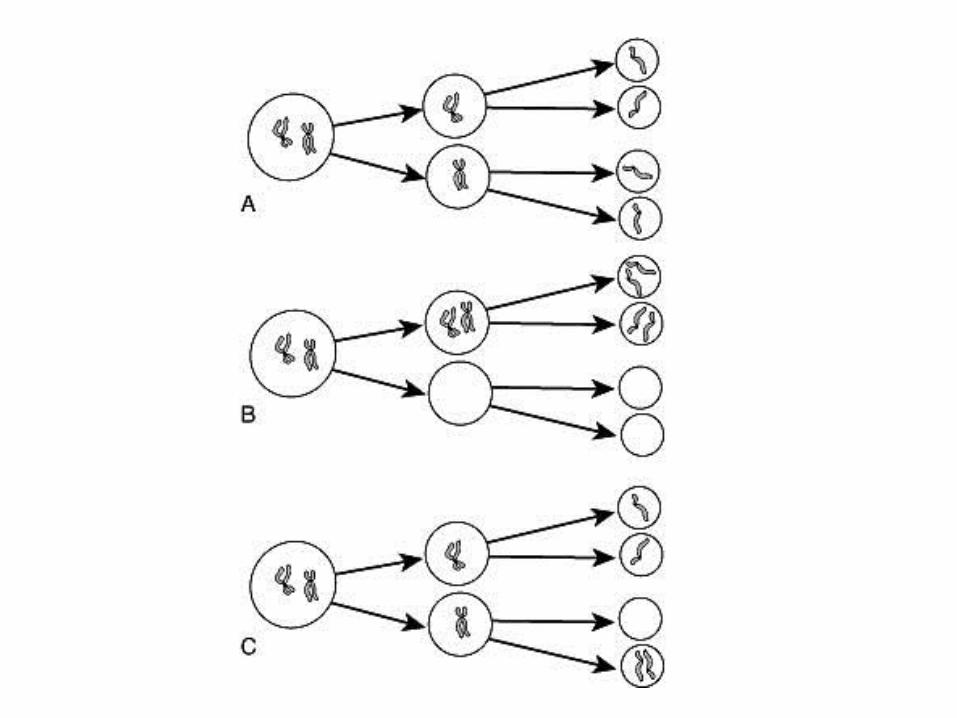

Primjer 1: Down sindrom, aneuploidija, trisomija 21-og kromosoma

Kromosomske aberacije

Kromosomski lomovi
• strukturne promjene kromosoma kao posljedica neuspješnog crossovera ili lima kromosoma uzrokovanog nekim sredstvom
• nakon loma-moguće spajanje s malom vjerojatnošću da će se spojiti isti dijelovi na istom mjestu
• ako se lom dogodi prije S faze, replicira se lom pa se govori o kromosomskom lomu, obje sestrinske kromatide imaju lom na istom mjestu
• ako se lom dogodi nakon S faze, tada lom zahvaća samo jednu kromatidu
• spojevi koji izazivaju lomove; klastogeni

Uzroci i vrste kromosomskih lomova
• klastogeni koji djeluju neovisno o S-fazi staničnog ciklusa-direktni lomovi (ionizirajuće zračenje i hidroksil radikali)
• lomovi koji nastaju uslijed procesiranja DNA koja je već prethodno oštećena nekim drugim sredstvom (ukriženja, DNA adukti, jednolančani lomovi) (zašto?)


Kromosomski lomovi


Kromatidni/kromosomski lomovi
• prelomljeni krajevi su mogu ponovno povezati bez posljedica
• ako ne dođe do povezivanja nastaju acentrični (bez centromere) ili centrični (s centromerom) fragmenti
• acentrični fragment se tijekom mitoze gubi, a njime i genetičke informacije
• dva centrična fragmenta (nastali kromosomskim lomom) se mogu povezati u novu strukturu-dicentrik
• dicentrični kromosom tijekom anafaze puca i u novonastalim stanicama kćerima jedna ima duplicirane gene a druga deletirane gene
• osim delecije i duplikacije, može doći i do inverzije
• inverzija uključuje dva loma, rotaciju za 180 stupnjeva i ponovno vezanje u strukturu kromosoma

Posljedice kromosomskih aberacija
• delecije velikih kromosomskih regija
• duplikacije velikih kromosomskih regija
• kromosomski rearanžmani:
– unutar kromosoma (inverzije)
– između različitih kromosoma (translokacije)

1. Delecija 2. Duplikacija 3. Inverzija kromosoma
Primjer 1: Delecije i duplikacije





























