Embed Size (px)
Citation preview

Transcription in Eukaryotes
by Jean-Pierre Herveg, Etienne De Plaen and a lot of friends atthe Brussels Branch of the Ludwig Institute for Cancer research (Licr) and the Christian de Duve*
Institute for cellular Patholgy (ICP).
April 2006Université Catholique de Louvain
Avenue E. Mounier, 1200 Brussels (Belgium)

Questions
1. In Prokaryotes, the sigma factor helps the RNA pol to recognize a promoter. How is this done in Eukaryotes ? 2. Describe a eukaryotic promoter.3. What are the three main postranscriptional modifications in eukaryotes ?4. What is a lariat ?5. How can the sequence of a pseudogene be recognized ?

Transcription in EukaryotesIn eukaryotes, DNA is contained within the nucleus, where DNA is transcribed into RNA. RNA must then be carried across the nuclear pores (exported) into the cytosol.
If in prokaryotes, transcription is performed by a single RNA pol, in eukaryotes, transcription is performed by 3 different RNA pols:
RNA pol I transcribes 5.8 S, 18 S, and 28 S ribosomal RNA in the nucleolus. RNA pol II transcibes m RNA and the small nuclear RNA (snRNA) RNA pol III transcribes 5 S rRNA as well as all the tRNA species.
* S mean Svedberg and is a unit of sedimentation.

"S" es la unidad Svedberg de sedimentación.
Svedberg es un físico sueco que inventó el centrífugo analítico."S" es una unidad de tiempo,1S = 10 -13 segundos16S es una sustancia que sédimente a 16S en esta máquina.16S Está también la constante de sedimentación de ARNde la pequeña unidad del ribosome en el procarioticos osmall subunit ribosomal RNA (SSU ARNr).En eucarioticos esta secuencia es 18S.Ahora, no se medie más velocidad de sedimentación de ARNpara compararlos. Se compara sus secuencias.

RNA pol I and III
RNA polymerase I (5.8 S, 18 S,and 28 S ribosomal RNA )
RNA polymerase I transcribes only the genes for ribosomal RNA, from a single type of promoter.The transcript includes the sequences of both large and small rRNAs, which are later released bycleavages and processing. There are many copies of the transcription unit,alternating with nontranscribed spacers, and organized in a cluster.
RNA polymerase III (5S rRNA and tRNA species).
The promoters fall into two general classes that are recognized in different ways by differentgroups of factors. The promoters for 5S and tRNA genes are internal;they lie downstream of the startpoint.The promoters for snRNA (small nuclear RNA) genes lie upstream of the startpoint in the moreconventional manner of other promoters. In both cases, the individual elements that arenecessary for promoter function consist exclusively of sequences recognizedby transcription factors, which in turn direct the binding of RNA polymerase.

RNA pol IIgeneral transcription factors (TF II) instead of the prokaryotic factor RNA Pol II does not contain a subunit similar to the prokaryotic factor, which can recognize the promoterand unwind the DNA double helix. In eukaryotes, these two functions are carried out by a set of proteinscalled general transcription factors. The RNA Pol II is associated with six general transcription factors,designated as TFIIA, TFIIB, TFIID, TFIIE, TFIIF and TFIIH, where "TF" stands for "transcription factor" and "II" for the RNA Pol II.
TATA-box binfing protein and TAFsTFIID consists of TBP (TATA-box binding protein) and TAFs (TBP associated factors). The role of TBPis to bind the “TATA” core promoter. TAFs may assist TBP in this process. In human cells, TAFs arformed by 12 subunits. One of them, TAF250 (with molecular weight 250 kD), has the histoneacetyltransferase activity, which can relieve the binding between DNA and histones in the nucleosome.
Pre-initation complex (PIC)The transcription factor which catalyzes DNA melting is TFIIH. However, before TFIIH can unwind DNA the RNA Pol II and at least five general transcription factors (TFIIA is not absolutely necessary)have to form a pre-initiation complex (PIC).
ElongationAfter PIC is assembled at the promoter, TFIIH, an helicase, can unwind DNA. This requires energy released from ATP hydrolysis. Then, RNA Pol II (NTPs) to synthesize a RNA transcript.During RNA elongation, TFIIF remains attached to the RNA polymerase, but all of the othertranscription factors have dissociated from PIC.The carboxyl-terminal domain (CTD) of the largest subunit of RNA Pol II is critical for elongation. In the initiation phase, CTD is unphosphorylated, but during elongation it has to be phosphorylated.
TerminationEukaryotic protein genes contain a poly-A signal located downstream of the last exon. This signal is usedto add a series of adenylate residues during RNA processing. Transcription often terminates at 0.5 - 2 kbdownstream of the poly-A signal, but the mechanism is unclear.

initiationThe promoter:Eukaryotic RNA pols lack the factor found in the prokaryotic enzyme. Instead of the Pribnow box,a TATA box, is found in most eukaryotic genes. It is located at approximately -25. Many promoters have a CAAT box and some a GC box, both at around -40 to -110 basesupstream. The location of these additional elements can vary, and they can be present oneither strand.
The basal transcription machinery: TF means trascription factor and II indicates that this TF belongsto the RNA pol II family of enzymes.TF II bind to the promoter region, guiding the polymerase to this site.They form the basal transcription machinery. The initial event in the process is the recognition ofthe TATA box by the TATA box-binding protein, a component of TFIID. This is followed by thesequential binding of other factors, including TFIIA, TFIIB, RNA polymerase II, and TFIIE.
Enhancers and silencers:Enhancer sequences, which can be located several thousand bases upstream, downstream, or in themiddle of the transcribed region, can also bind proteins which stimulate transcription.These are often tissue- and species-specific, explaining the regulation of genes in some tissues,and the host range of viruses which have usurped these sequences to stimulate transcription of theirown genes.
-------------------------------------questionDescribe a eukaryotic promoter

Acetylation: to separate DNA from the nucleosomesIn eukaryotes, the association between DNA and histones prevents access of the polymerase and generaltranscription factors to the promoter. Histone acetylation catalyzed by HATs can relieve the binding betweenDNA and histones. Although a subunit of TFIID (TAF250 in human) has the HAT activity,participation of other HATs can make transcription more efficient. The following rules apply to most (but not all) cases:
Binding of activators to the enhancer element recruits HATs to relieve association between histonesand DNA, thereby enhancing transcription.
Binding of repressors to the silencer element recruits histone deacetylases(denoted by HDs or HDACs) to tighten association between histones and DNA.
Methylation: to silence genes !Experimental evidence has shown that in certain cells there are heavily methylated genes and these genesare not expressed. On the other hand cells that have non-methylated forms of these genes are expressed.
An example of these is seen in housekeeping cells (cells that produce proteins used in "clean up" ofcellular debris and dead organelles) in which cells with non-methylated genes for these cells continuouslytranscribe or produce the materials needed to make housekeeping cells.
Also one of the X chromosomes in females is not expressed.A hypothesis for this phenomenon is linked to the heavy methylation of the inactive X chromosome.
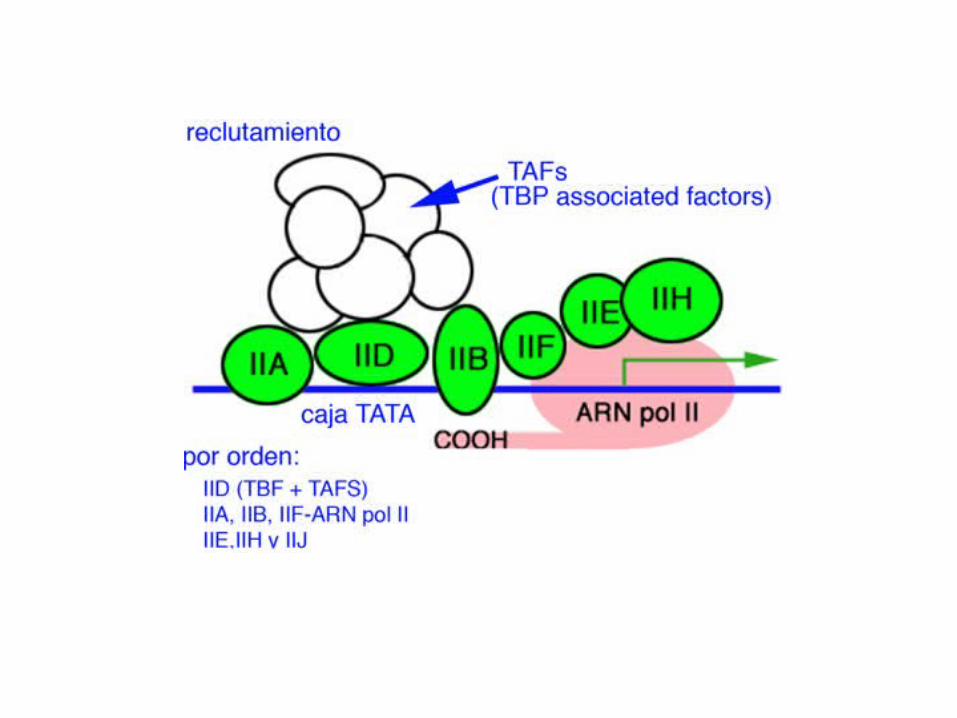

TAFs:TAFs may assist TBP in connecting the basal transciption machinery to enhancers or silencers. In human cells, TAFs are formed by 12 subunits. One of them, TAF250 (with molecular weight 250 kD), has the histone acetyltransferase activity,which can relieve the binding between DNA and histones in the nucleosome.
------------------------------------questionIn Prokaryotes, the sigma factor helps the RNA pol to recognize a promoter. How is this doneIn Eukaryotes ?

TFIIH can now use its helicase activity to unwind DNA.This requires energy released from ATP hydrolysis.
The DNA melting starts from about -10 bp. Then, RNA Pol II uses nucleoside triphosphates (NTPs)to synthesize a RNA transcript.
During RNA elongation, TFIIF remains attached to the RNA polymerase,but all of the other transcription factors have dissociated from PIC (pre-initiation complex).
The carboxyl-terminal domain (CTD) of the largest subunit of RNA Pol II is critical for elongation.In the initiation phase, CTD is unphosphorylated, but during elongation it has to be phosphorylated. This domain contains many proline, serine and threonine residues.
elongation
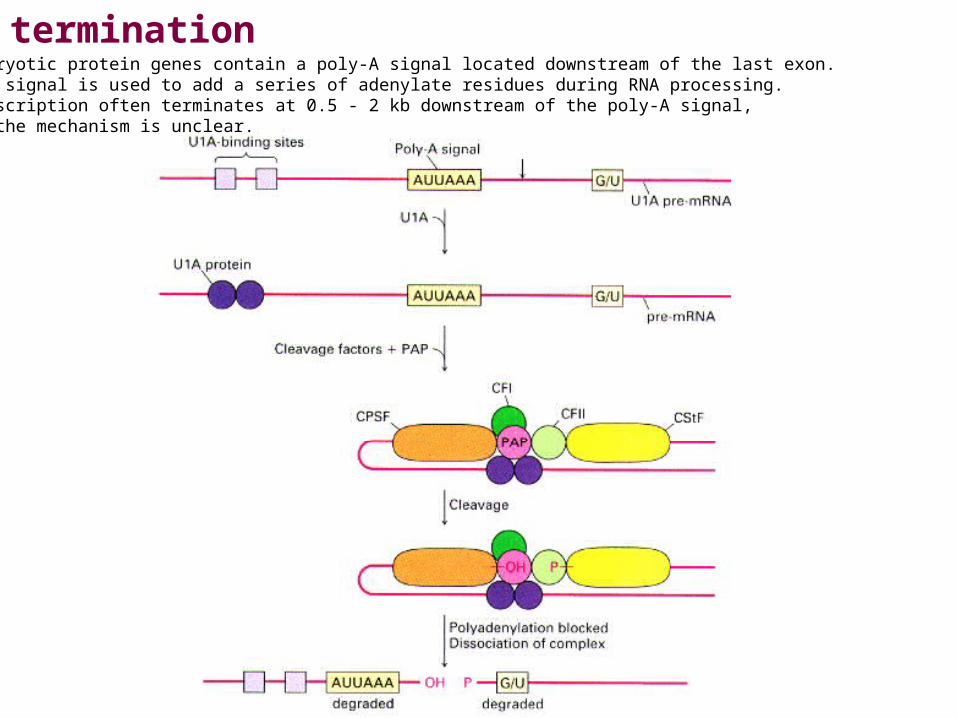
Eukaryotic protein genes contain a poly-A signal located downstream of the last exon. This signal is used to add a series of adenylate residues during RNA processing. Transcription often terminates at 0.5 - 2 kb downstream of the poly-A signal,but the mechanism is unclear.
termination

Capping
Modification of the 5'-ends of eukaryotic mRNAs is called capping.The cap consists of a methylated GTP linked to the rest of the mRNA by a 5' to 5' triphosphate "bridge”(Cap Structure). Capping occurs very early during the synthesis of eukaryotic mRNAs, even before mRNAmolecules are finished being made by RNA polymerase II. Capped mRNAs are very efficiently translatedby ribosomes to make proteins. In fact, some viruses, such as poliovirus, prevent capped cellular mRNAsfrom being translated into proteins. This enables poliovirus to take over the protein synthesizing machineryin the infected cell to make new viruses.
Polyadenylation
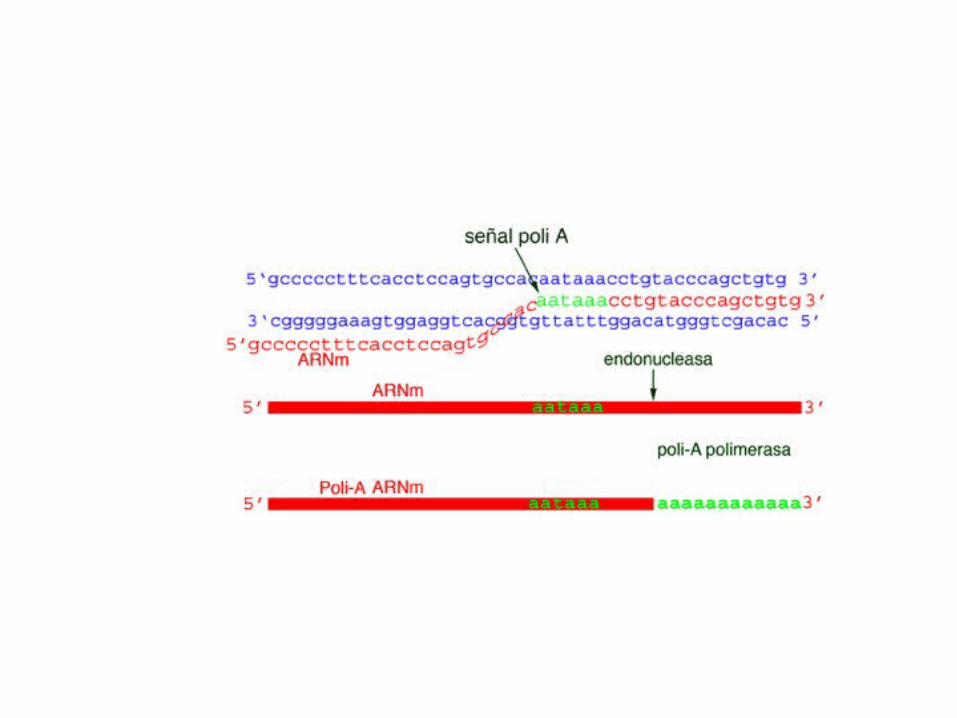
Modification of the 3'-ends of eukaryotic mRNAs is called polyadenylation (Polyadenylation Pathway).Polyadenylation is the addition of several hundred A nucleotides to the 3' ends of mRNAs.Polyadenylation signalAll eukaryotic mRNAs destined to get a poly A tail (note: most, but not all, eukaryotic mRNAs get such a tailcontain the sequence AAUAAA about 11-30 nucleotides upstream to where the tail is added.AAUAAA is recognized by an endonuclease that cuts the RNA, allowing the tail to be added by a specificenzyme: polyA polymerase.
Splicing (trans or cis):
postranscritional modifications
-------------------------QuestionWhat are the three main postranscriptional modifications in eukaryotes ?

splicing
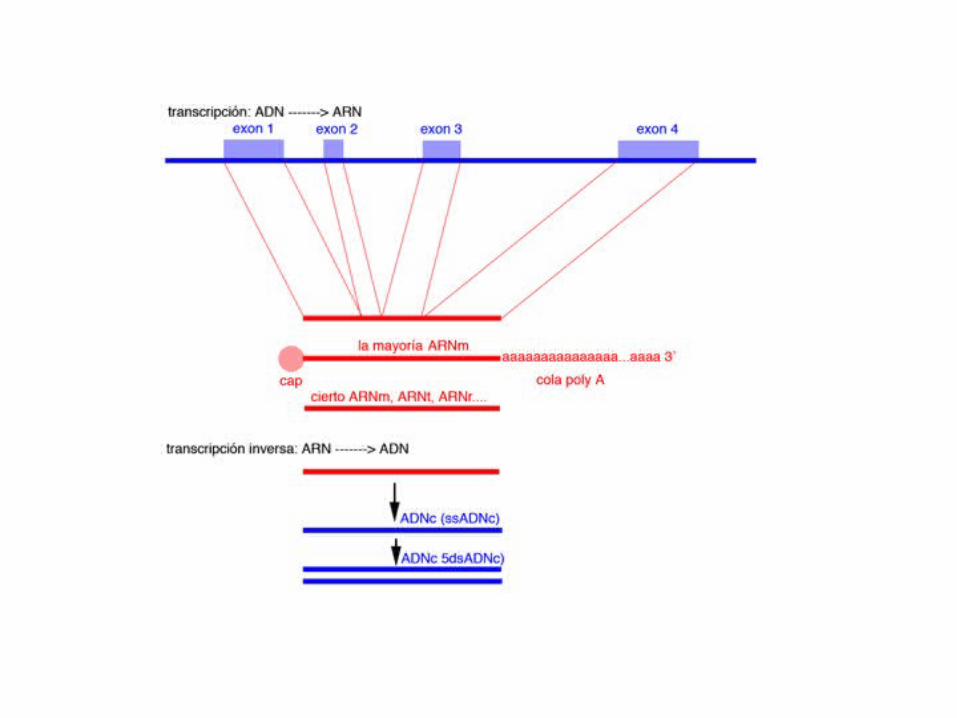
Another major difference between prokaryotic and eukaryotic mRNA is the occurence of splicingin eukaryotes. This process removes intervening sequences (introns) from the primary transcript,and precisely assembles a set of exons which form the transcript that is translated.

The fragments which are removed are not random. they have consensus sequences at the 5’and 3' splice sites.
Exonsend with the sequence AG andbegin with a G, and
Introns begin with GU andend with AG.
About 20-50 bases upstream from the 3' splice site is an adenine residue known as thebranch site:
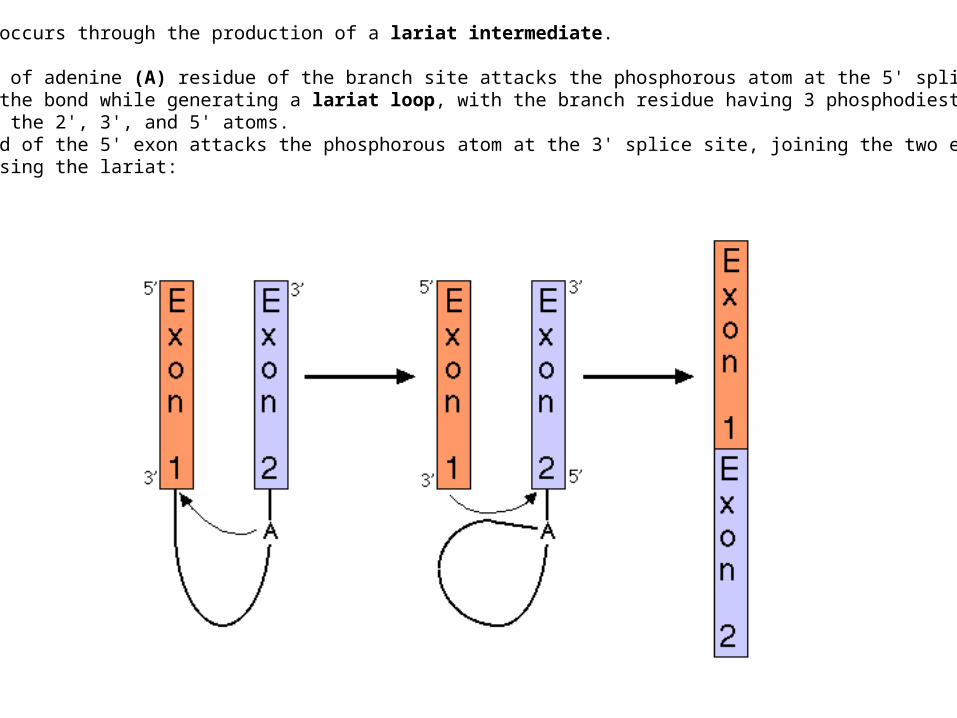
Splicing occurs through the production of a lariat intermediate.
The 2'-OH of adenine (A) residue of the branch site attacks the phosphorous atom at the 5' splice site, cleaving the bond while generating a lariat loop, with the branch residue having 3 phosphodiesterbonds, at the 2', 3', and 5' atoms.The 3' end of the 5' exon attacks the phosphorous atom at the 3' splice site, joining the two exonsand releasing the lariat:
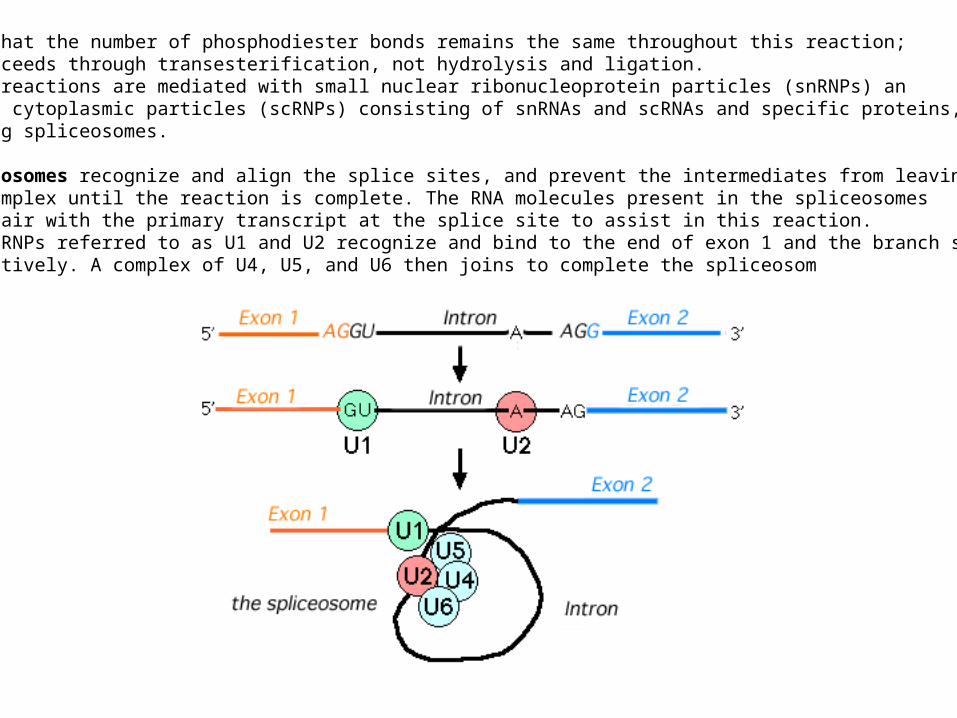
Note that the number of phosphodiester bonds remains the same throughout this reaction;it proceeds through transesterification, not hydrolysis and ligation.These reactions are mediated with small nuclear ribonucleoprotein particles (snRNPs) an small cytoplasmic particles (scRNPs) consisting of snRNAs and scRNAs and specific proteins,forming spliceosomes.
Spliceosomes recognize and align the splice sites, and prevent the intermediates from leavingthe complex until the reaction is complete. The RNA molecules present in the spliceosomesbase-pair with the primary transcript at the splice site to assist in this reaction.The snRNPs referred to as U1 and U2 recognize and bind to the end of exon 1 and the branch site,respectively. A complex of U4, U5, and U6 then joins to complete the spliceosom
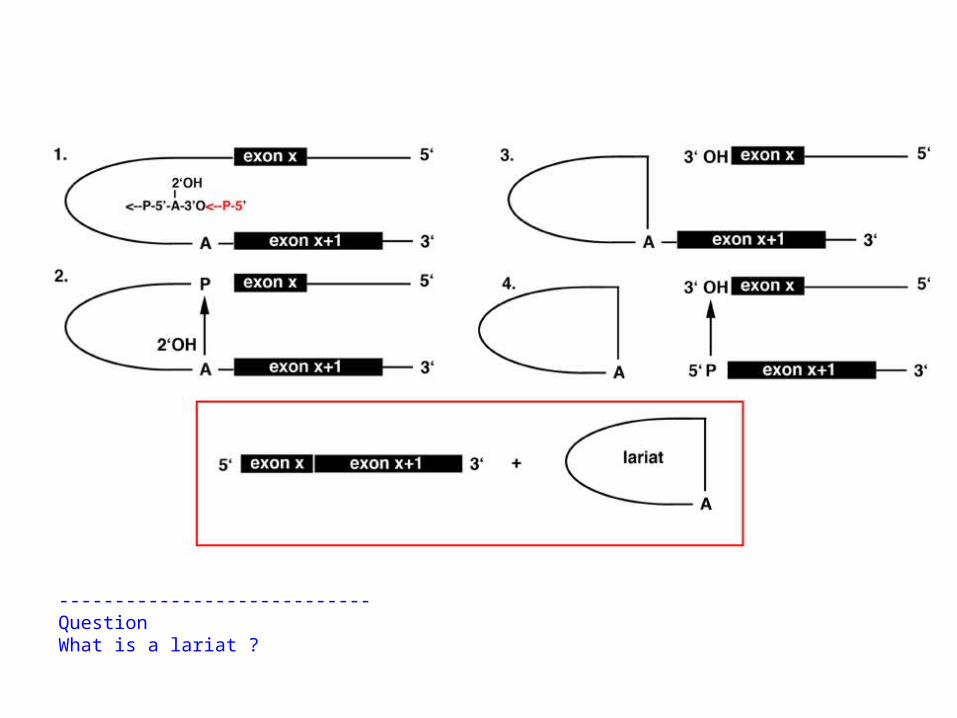
----------------------------QuestionWhat is a lariat ?
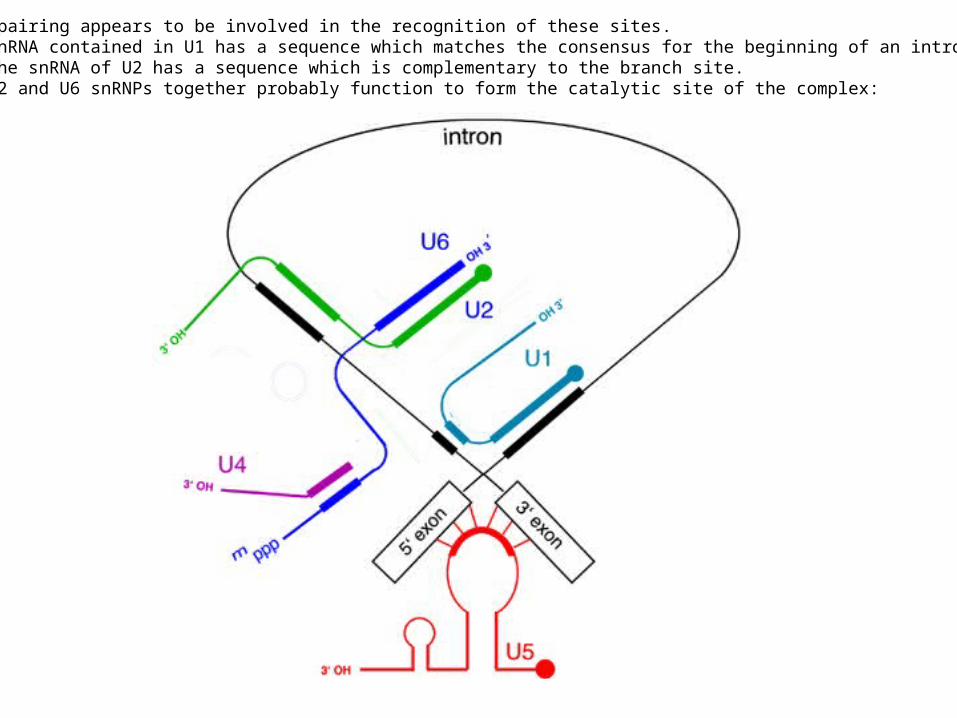
Base pairing appears to be involved in the recognition of these sites.The snRNA contained in U1 has a sequence which matches the consensus for the beginning of an intron,and the snRNA of U2 has a sequence which is complementary to the branch site.The U2 and U6 snRNPs together probably function to form the catalytic site of the complex:

alternative splicing
For those who are interested, there is also a self splicing RNA:http://138.192.68.68/bio/Courses/biochem2/RNA/SelfSplicingRNA.html

Mammalian vectors have often an intron before the polylinker,This intron increase transcription:
An intron inside a vector
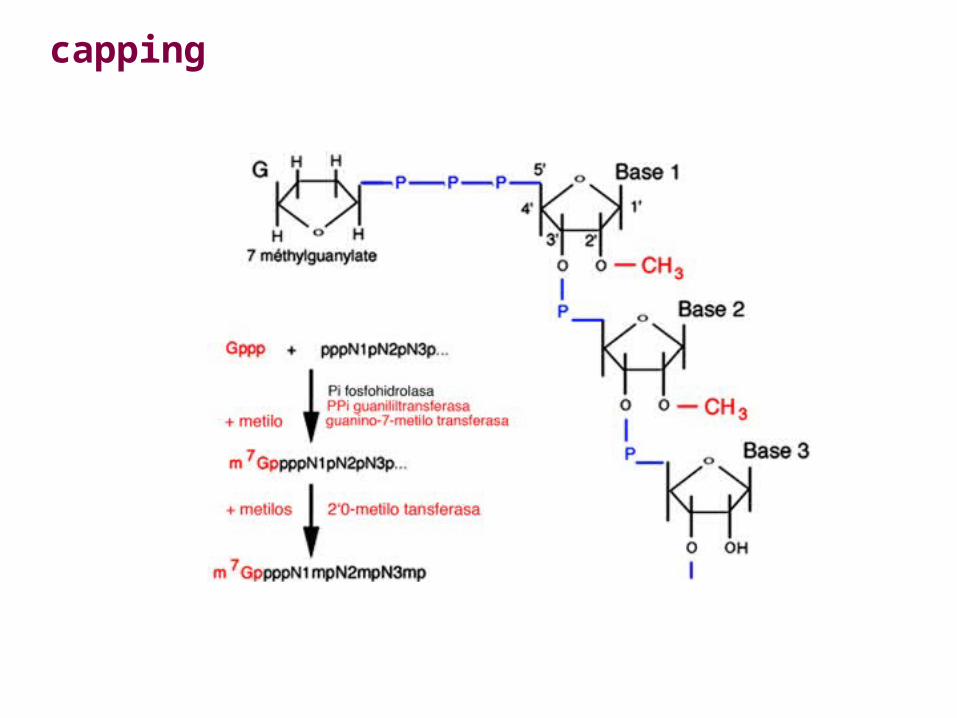
capping
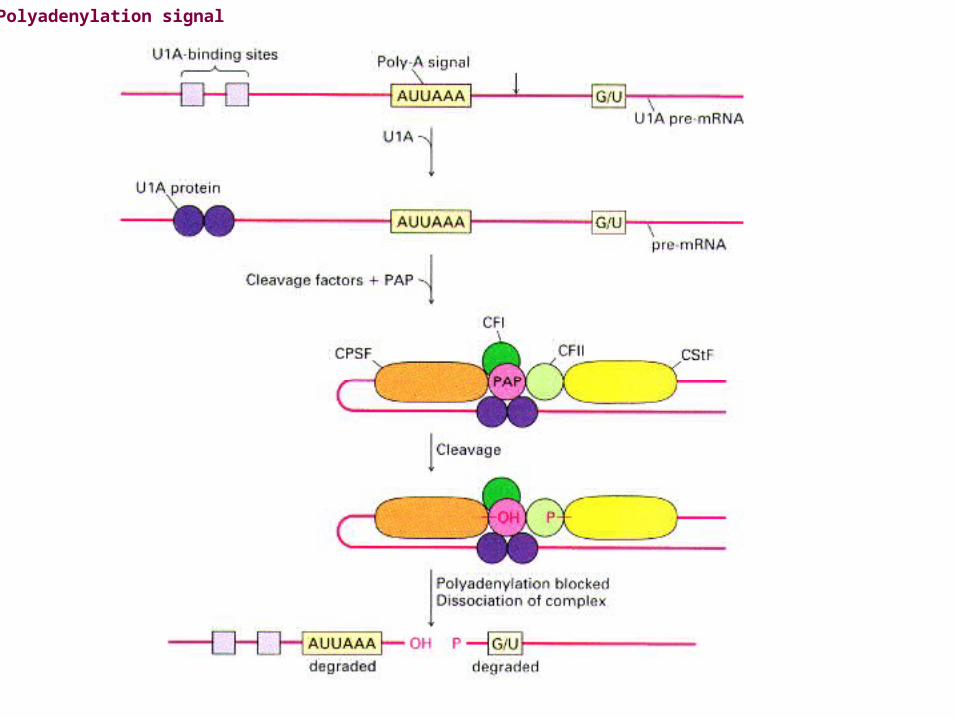
Polyadenylation signal

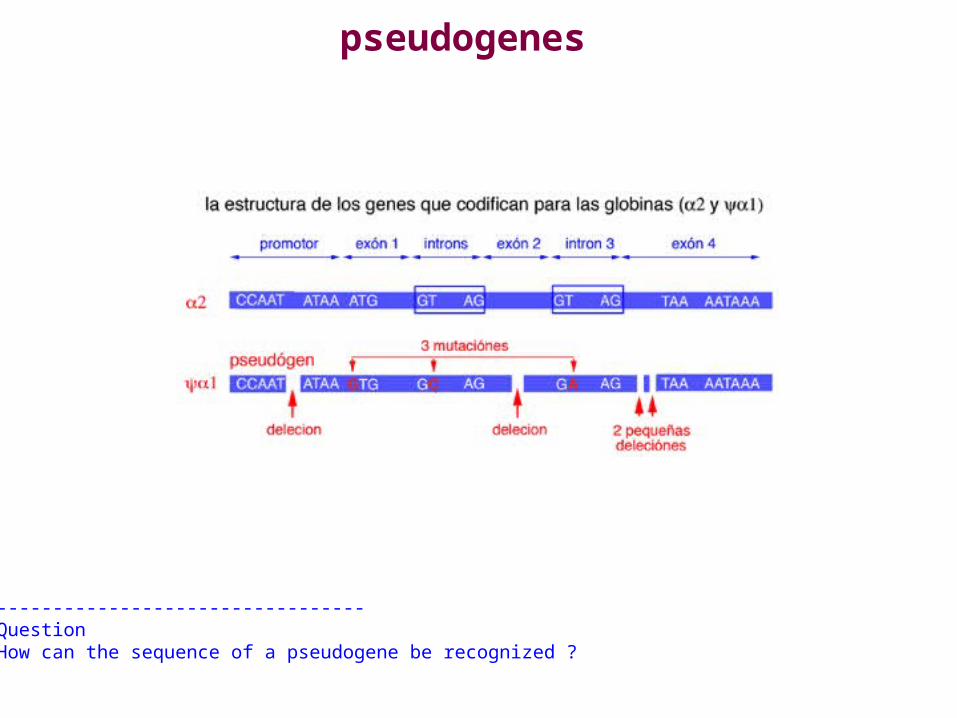
pseudogenes
---------------------------------QuestionHow can the sequence of a pseudogene be recognized ?






























